Embed Size (px)
Citation preview

ESCUELA TÉCNICA SUPERIOR DE INGENIEROS AGRÓNOMOS UNIVERSIDAD POLITÉCNICA DE MADRID
Departamento de Biotecnología
PROTEÍNAS DE DEFENSA COMO POTENCIALES PANALERGENOS VEGETALES: QUITINASAS DE CLASE I Y
PROTEÍNAS DE TRANSFERENCIA DE LÍPIDOS.
Tesis Doctoral ARACELI DÍAZ PERALES
2000

ESCUELA TÉCNICA SUPERIOR DE INGENIEROS AGRÓNOMOS UNIVERSIDAD POLITÉCNICA DE MADRID
Departamento de Biotecnología
Proteínas de defensa como potenciales panalergenos vegetales: Quitinasas de clase I y Proteínas de Transferencia
de Lípidos.
Memoria presentada por Dña. Araceli Díaz Perales para optar al grado de Doctor.
Madrid, 1 de Septiembre de 2000
ú^
V°B° El Director de la Tesis
Fdo. Gabriel Salcedo Duran Profesor Titular de Bioquímica y Biología Molecular
Universidad Politécnica de Madrid

TRIBUNAL
PRESIDENTE :
VOCAL:
VOCAL:
VOCAL:
VOCAL SECRETARIO:
CALIFICACIÓN:

Reconocimientos
El presente trabajo ha sido realizado en la Unidad de Bioquímica,
Departamento de Biotecnología, de la E.T.S. Ingenieros Agrónomos,
Universidad Politécnica, Madrid, en el marco de los proyectos PB95-0035 y
PB 98-0735 del plan sectorial de PGC (MEC). Araceli Díaz Perales ha sido
becaria del programa FPI (MEC) durantes los años 1997-2000.
La consecución de muchos de los resultados recogidos en esta
memoria no hubiera sido posible sin la colaboración de los siguientes
investigadores:
Dra. R. Sánchez-Monge (Dpto. Biotecnología, E.T.S. Ingenieros
Agrónomos, UPM) cuya participación ha sido esencial en todas las etapas
de este trabajo, tanto en su diseño como en su realización experimental.
Dr. C. Blanco (Sección de Alergia, Hospital de Gran Canaria Dr.
Negrín, Las Palmas de Gran Canaria) ha aportado ideas en todo lo referente
a la caracterización de alérgenos implicados en el síndrome látex-frutas, así
como, las muestras de sueros de pacientes alérgicos. También realizó las
pruebas cutáneas y los ensayos de inhibición con CAP comerciales
correspondientes.
Drs. C. Aragoncillo y C. Collada (Dpto. Biotecnología, E.T.S.
Ingenieros de Montes, UPM) colaboraron activamente en la purificación y
caracterización de las quitinasas como panalergenos y cedieron los
anticuerpos policlonales monoespecíficos frente a las mismas. Los Drs. C.
Aragoncillo y L. Gómez (Dpto. Biotecnología, E.T.S. Ingenieros de Montes,
UPM) suministraron desinteresadamente el clon ADNc de la quitinasa de
clase I de castaña.
Drs. D. Barber y M. Lombardero (ALK-Abelló, Madrid) sugirieron la
identificación inicial de los alérgenos de bajo peso molecular de manzana y
melocotón, y han participado en la caracterización de las LTPs alergénicas.
También aportaron parte del material vegetal utilizado en ese trabajo.
Drs. F. J. García-Sellés (Dpto. de Alergología, Hospital Virgen de la
Arrixaca, Murcia) y M. Fernández-Rivas (Sección de Alergia, Fundación

Alcorcón, Madrid) suministraron los sueros de pacientes alérgicos a frutas de
la familia Rosaceae y realizaron las pruebas clínicas en los estudios de
caracterización de las LTPs como alérgenos.
Dra. M. Pernas (Dpto. Biotecnología, E.T.S. Ingenieros Agrónomos,
UPM; actualmente en CNB, CSIC, Madrid) participó en la purificación de la
LTP de castaña y en las etapas iniciales del aislameinto de clones de ADNc
de las de manzana y melocotón.
Dra. M.T. Villalba (Dpto. Bioquímica y Biología Molecular, Facultad de
Ciencias Químicas, Universidad Complutense, Madrid) cedió
desinteresadamente el vector pPIC 9, así como la cepa de levadura de
Pichia pastoris GS115. Además aportó ideas para la expresión de las
proteínas recombinantes.
J. Várela (CIB, CSIC, Madrid) determinó las secuencias
aminoterminales de las distintas proteínas, así como la composición de
aminoácidos de la quitinasas de clase II de aguacate.
A. Rubio (Dpto. Biotecnología, E.T.S. Ingenieros Agrónomos, UPM;
actualmente en CNIO, CSIC, Madrid) realizó las secuencias de nucleótidos
de las distintas contrucciones de ADN presentadas en este trabajo.
Drs. T. Carrillo, N. Ortega y M. Álvarez (Sección de Alergia, Hospital
de Gran Canaria Dr. Negrín, Las Palmas de Gran Canaria) participaron en la
selección de pacientes con el síndrome látex-frutas.
Dr. M.J. Chrispeeis (Dept. Biología, Universidad de California, San
Diego, USA) cedió el suero específico frente a N-oligosacáridos complejos
de plantas.
Dr. Z. Chen (Instituto de Investigación de Medicina Ocupacional,
BGFA, Bochum, Alemania) de cuyo laboratorio proviene la heveína de látex
purificada.
J. García (Dpto. Biotecnología, E.T.S. Ingenieros Agrónomos, UPM)
colaboró en todos los montajes fotográficos presentados en este trabajo, D.
Lamoneda (del mismo Dpto.) en la elaboración de algunas figuras, y F.
García, A. García y C. Rojas ( mismo Dpto.) prestaron su asistencia técnica.

Agradecimientos
Llegado este momento, siempre queda lo más difícil. No hay duda
que habrá muchas personas que no estén en estos agradecimientos, aunque
deberían haber estado.
En primer lugar, agradecer el cariño, la paciencia y el tesón de mis
jefes, Gabi y Rosa, que han hecho que este trabajo sea más sencillo y
divertido en todos los sentidos.
A todos mis compañeros de laboratorio que me han ayudado, y que
siempre escucharon mis quejas. En especial, a Mónica y Gloria, mis
"jefecillas" que nunca dejaron que decayera mi ánimo. Sin ellas, todo
hubiera sido más difícil. A Arantxa, Emilia y Marta, tanto por su apoyo
como por su interés, y sobretodo, por el último cigarro de la tarde. A
"Rosita" que aunque ha llegado tarde, me ha ayudado como la que más.
A Nines, Gema y Zamira porque siempre hubo un abrazo y un
consuelo. Por su amistad y su alegría.
Al resto de mis compañeros, que están o han estado, por sus risas.
A los chicos de Montes, que están cuando se les necesita, pero,
especialmente, a "Alvarito" porque siempre se ha prestado a ayudarme, y
hacerme reir.
A la gente de Abelló, que me hicieron pasar muy buenos ratos
mientras estuve allí.
A los técnicos de la unidad; a Joaquín, por su paciencia; a Feli, por su
inestimable ayuda; A Lola, por su eficiencia; a Carlos, por su interés; y a
Ángel, por sus "pastillas" contra el dolor de cabeza.

A todos en general por haberme regalado cuatro años maravillosos,
incluyendo los malos ratos.
Me gustaría hacer una mención especial a mis amigos que
aguantaron mis desplantes, mis cortes por teléfono, y mi mal humor. A
Mari Carmen y Miguelón, a Chus y Jesús, a Queli, David y Juanchi. A
vosotros porque siempre habéis y estáis ahí.
A mi familia, sin la cual no lo hubiera conseguido. A los "asociados",
por tratarme como una hermana. A mis hermanas. Minerva y Rebeca, por
su amistad, y porque sois las mejores. Pero, sobretodo, esta tesis es de
Dani, mi saco de boxeo y mi fiel compañero.
Y, todo esto, ha sido gracias a mis padres, que siempre me han
apoyado en "mis locuras" y gracias a cuyo esfuerzo soy lo que soy. Para
ellos, no hay palabras suficientes de gratitud. Simplemente, gracias.

A mis padres y mis hermanas,
quienes nunca aceptaron que abandonase.
A todas aquellas personas que estuvieron
conmigo a pesar de los malos tiempos,
Chus, Jesús, Queli, David y Juanchi.
A nuestro castillo y a las estrellas
que vemos desde él, siempre presentes.

índice

índice
I.- INTRODUCCIÓN.
1.1.- HIPERSENSIBILIDAD TIPO I, ALÉRGENOS E IgE. 2
I.2.- ALÉRGENOS DE ALIMENTOS VEGETALES Y PROTEÍNAS
DE DEFENSA. 4
I.2.1.- Alérgeno de alimentos vegetales y reacciones
cruzadas. 4
1.2.2.- Proteínas de defensa y panalergenos vegetales. 5
I.3.- SÍNDROME LÁTEX-FRUTAS. 8
1.3.1.- Alergia al látex. Alérgenos caracterizados. 8
1.3.2.- Reacciones cruzadas de látex y alimentos:
síndrome látex-frutas. 10
1.3.3.- Heveína y proteínas con dominio heveína. 11
I.4.- ALERGIA A FRUTOS DE LA FAMILIA Rosaceae Y
PROTEÍNAS DE TRANSFERENCIA DE LÍPIDOS (LTPs). 15
1.4.1- Síndrome de polen de abedul-frutas-hortalizas:
B e t v l 15
I.4.2.- Alergia a rosáceas en el área mediterránea: LTPs. 16
I.5.- OBJETIVOS DE LA TESIS 19
II.- MATERIAL Y MÉTODOS.
11.1.-MATERIAL. 22
11.1.1.-Material vegetal. 22
II. 1.2.-Sueros de pacientes. 23

índice
.2.- MÉTODOS DE PURIFICACIÓN Y CARACTERIZACIÓN
DE PROTEÍNAS. 26
11.2.1.- Extracción de proteínas. 26
11.2.1.1.- Extracto crudo de proteínas solubles
(PBS). 26
II.2.1.2.- Preparaciones enriquecidas en quitlnasas. 26
11.2.1.3.- Preparaciones enriquecidas en LTPs. 27
11.2.2.- Cuantificación de proteínas. 27
11.2.3.- Métodos cromatográficos. 27
11.2.3.1.- Cromatografía de filtración molecular. 27
11.2.3.2.- Cromatografía de afinidad. 28
11.2.3.3.- Cromatografía de intercambio catiónico. 28
11.2.3.4.- Cromatografía líquida de alta presión
(HPLC). 29
11.2.4.- Métodos electroforéticos. 30
11.2.5.- Ensayos de actividad quitinasa. 30
11.2.6.- Determinación de la composición de aminoácidos. 31
11.2.7.- Determinación de secuencias aminoterminales. 31
11.2.8.- Determinación de pesos moleculares por
espectrometría de masas. 31
.3.- MÉTODOS INMUNOQUÍMICOS. 32
11.3.1.- Purificación de anticuerpos policlonales. 32
11.3.2.- Inmunodetección de proteínas transferidas a
membranas. 32
11.3.2.1.- Electrotransferencia a membranas de
nitrocelulosa o difluoruro de polivinilideno
(PVDF). 32
11.3.2.2.- Inmunodetección frente a anticuerpos
policlonales purificados. 33
11.3.2.3.- Detección de glicoproteínas. 34

índice
II.3.2.3.1.- Detección de glicoproteínas por
inmunoensayo con digoxigenina. 34
II.3.2.3.2.- Inmunodetección con anticuerpos
específicos frente a p (1-2) xilosa y
a(1-3)fucosa. 34
11.3.2.4.- Inmunodetección frente a sueros de
pacientes alérgicos. 34
11.3.2.4.1.- Inmunodetección con fosfatasa
alcalina. 35
11.3.2.4.2.- Inmunodetección con
Quimioluminiscencia. 35
II.3.2.4.3.- Ensayos de inmunoinhibición en
membrana. 35
II.3.3.- Enzimoinmunoensayo (ELISA). 36
II.3.3.1.- Determinación de IgE específica. 36
11.3.3.2.- Ensayos de inhibición. 36
II.3.4.- Pruebas de inhibición por radioinmunoensayo
(RAST). 37
11.3.5.- Pruebas cutáneas (SPT). 37
.4.- MÉTODOS DE ÁCIDOS NUCLEICOS. 38
11.4.1.- Extracción del ARN total de piel de manzana y
melocotón. 38
11.4.2.- Obtención de los clones de ADNc correspondientes
a las LTPs de melocotón y manzana. 39
11.4.3.- Subclonaje en el vector de expresión pPIC 9 de
la levadura Pichia pastoris. 40
11.4.3.1.- Obtención del ADNc del dominio heveína,
catalítico y de la proteína madura Cas s 5
(quitlnasa de clase I de castaña). 41
li.4.3.2.- Subclonaje en el vector de expresión
pPIC 9. 43

índice
11.4.4.- Transformación de la cepa de P. pastoris GS115. 44
11.4.5.- Expresión de proteínas en P. pastoris. 45
III.- RESULTADOS.
III.1.- QUITINASAS DE CLASE I Y SÍNDROME LÁTEX-FRUTAS 47
111.1.1.- Caracterización y purificación de los alérgenos
mayoritarios de castaña y aguacate. 47
III.1.2.- Purificación y caracterización de los alérgenos
mayoritarios de plátano. 56
III.1.3.- Reacciones cruzadas en el síndrome látex-frutas:
quitinasas de clase I como potenciales
panalergenos. 60
III. 1.4.- Los panalergenos implicados en el síndrome látex-
frutas son inducibles por etileno e inactivados por
tratamiento térmico. 65
III.1.5.- Producción de Cas s 5 y su dominio catalítico en
Pichia pastoris: papel central del dominio heveína
en la alergenicidad de las quitinasas de clase I. 71
III.2.- LTPs COMO POTENCIALES PANALERGENOS VEGETALES
III.2.1.- Purificación de las LTPs de polen de Artemisia y
semilla de castaño: reactividad cruzada con los
alérgenos de manzana y melocotón. 78
III.2.2.- Clonaje de los ADNc correspondientes a los
alérgenos Pru p 3 y Mal d 3. 85
IV.- DISCUSIÓN.
IV.1.- PROTEÍNAS DE DEFENSA Y ALERGIA. 89

índice
IV.2.-ALÉRGENOS IMPLICADOS EN EL SÍNDROME
LÁTEX-FRUTAS. 90
IV.2.1.- Las quitlnasas de clase I son alérgenos
mayoritarios en castaña, aguacate, y
plátano. 90
IV.2.2.- Las quitlnasas de clase I como potenciales
panalergenos vegetales. 93
IV.2.3.- Los panalergenos del síndrome látex-frutas
son termolábiles e inducibles por etileno. 97
IV.2.4.- El dominio heveína, y no el catalítico, es
responsable de la alergenicidad de las
quitlnasas de clase I. 99
IV.3.- LTPs: UNA FAMILIA DE PANALERGENOS CUYOS
MIEMBROS PRESENTAN DIFERENTE ACTIVIDAD
ALERGÉNICA. 103
V.- RESUMEN Y CONCLUSIONES. 110
VI.-SUMMARY. 114
Vil.-BIBLIOGRAFÍA. 117

Abreviaturas
Abreviaturas
AOX Gen que codifica para la aicoiiol oxidasa de Pichia pastoris
Art V 3 Alérgeno de A. vulgaris (LTP)
BSA Albúmina de Suero Bovino
BCIP/NBT 5-bromo-4-cloro-3-indolil fosfato / azul de nitrotretrazolium
BMGY Buffered glycerol-complex medio
BMMY Buffered methanol-complex medio
BMG Buffered minimal glicerol med io
BMM Buffered minimal methanol medio
CAP Capsulated hidrophylic carrier polymer
Csl 3 Quitinasa de clase I de castaña
Csll 1 Quitinasa de clase II de castaña
Cas s 3 Alérgeno de castaña (LTP)
Cas s 5 Alérgeno de castaña (Quitinasa)
EDTA Ácido etilendiaminotetracético
ELISA Enzyme linked immunosorbent assay
ES Error estándar
HIS4 Gen que codifica para la histidil deshidrogenasa de P. pastoris
FPLC Fast-Performance Liquid Chromatography
HPLC l-liglit-Performance Liquid Chromatography
IgE Inmunoglobulina E
LB Medio de Luria-Bertani
LTP(s) Proteínas de Transferencia de Lípidos
MALDI Matrix-assisted láser desorption/ionization
Mal d 3 Alérgeno de manzana (LTP)
M D Minimal dextrose m ed io
MM Minimal methanol medio
Mus a 1.1 Alérgeno de plátano (Quitinasa)
Mus a 1.2 Alérgeno de plátano (Quitinasa)
MOPS Ácido 3-(N-morfolinolpropanosulfónico)
OD Densidad Óptica

Abreviaturas
PAGE Electroforesis en geles de poliacrilamida
PBS Tampón fosfato salino
Par j 1 Alérgeno de polen de Paríetaría judaica (LTP)
PR Pathogenesis related
Prs a 1 Alérgeno de aguacate (Quitinasa)
Pru p 3 Alérgeno de melocotón (LTP)
PvChl Quitinasa de clase I de judía verde
PVDF Difluoruro de polivinilideno
PvPhy Fitohematoglutinina de judía blanca
RAST Radioallergosorbent test
rCas s 5 Alérgeno de castaña recombinante
rCAT Proteína recombinante del domino catalítico de Cas s 5
SAO Síndrome de Alergia Oral
SPT Skin prick test
TBS Tampón tris salino
WGA Aglutinina de germen de trigo
YNB Yeast nitrogen base

Introducción

Introducción
1.1.- HIPERSENSIBILIDAD TIPO I, ALÉRGENOS E IgE
La alergia (hipersensibilidad tipo I) afecta a un 20% de la población
mundial, y su desarrollo depende tanto de factores genéticos como ambientales
(Kay, 1997). La característica fisiopatológica central de esta enfermedad es la
producción de anticuerpos tipo inmunogiobulina E (IgE) frente a antígenos
(alérgenos) que son inocuos en personas sanas (Ishizaka et al., 1966;
Johansson y Bennich, 1967). Los síntomas inmediatos de la alergia (rinitis,
dermatitis, asma, anafilaxia) están producidos por la interacción entre la IgE y el
alérgeno, que induce una respuesta inmune, cuyo resultado final es la
liberación de mediadores inflamatorios (histamina, leucotrienos, etc.)
responsables de dichos síntomas clínicos.
Alérgeno Mastocíto/Basófilo
"f)V^ \yfvy
IgE - •
^ IL4, IL13
Cg) Péptldo IL5
derivado \ del alérgeno O-
Eosinófilo
o ,0 r.„^
Hipersensibilidad inmedita
""iRespuesta tardía
Figura 1: Esquema de la respuesta inmunológica en alergia.
La alergia se debe, al menos en parte, a una desviación del sistema
inmune hacia células T colaboradoras de tipo TH2 (van Neerven, 1999; Corry y
Kheradmand, 1999). En el proceso de sensibilización al alérgeno (Fig. 1), éste
entra en contacto con Células Presentadoras de Antígeno (APC) que lo
degradan a péptidos de 15-20 aminoácidos. Los péptidos son expuestos en la
membrana de éstas células, asociados a moléculas del Complejo Mayor de
Histocompatibilidad II (MHC-HLA II), donde son reconocidos por los linfocitos T,
que se diferencian en células TH2, y segregan, entre otras citoquinas,
interleuquinas 4, 5, y 13 (IL-4, -5, -13). IL-4 e IL-13 son requeridas para la
diferenciación y síntesis de IgE por células B e IL-5 para el desarrollo y

Introducción
diferenciación de eosinófiios (implicados en respuestas tardías). El complejo
IgE-alergeno se une a receptores de alta afinidad (FcsRI) de mastocitos y
basófilos, y su entrecruzamiento induce la desgranulación, liberándose
mediadores inflamatorios como histamina, leucotrienos, etc. La presencia de
IgE unida a receptores facilita la reacción a nuevas exposiciones.
El balance TH1/TH2 (TH1, células T colaboradoras tipo 1, que secretan
citoquinas diferentes a las de TH2, como IL-2 e interferón y (IFN-y), y que
conjuntamente movilizan mecanismos de defensa contra patógenos
intracelulares y antagonizan respuestas IgE) también ha sido invocado para
explicar el notable aumento de enfermedades alérgicas en países desarrollados
(Romagnani, 1998; Cookson, 1999). El cambio en el patrón de enfermedades
infecciosas, y en general en el contacto con organismos patógenos, debido a
las mejores condiciones higiénicas, programas de vacunación y mejores
hábitos nutricionales, podría haber provocado una falta importante de señales
para el desarrollo de líneas TH1 en los primeros años de vida.
De lo expuesto anteriormente, puede deducirse que alérgenos e IgE son
moléculas centrales en la reacción de hipersensibilidad tipo I. El diagnóstico de
esta enfermedad se realiza por la detección de IgE específicas en sueros de
pacientes, y su relevancia clínica puede confirmarse por pruebas cutáneas y de
provocación (Fuchs, 1979). La vía específica de tratamiento (inmunoterapia)
implica la administración de dosis crecientes del alérgeno para inducir
respuestas inmunitarias no reactivas al mismo (Bousquet et al., 1998). La
mejora de ambos aspectos clínicos (diagnosis y terapia) conlleva la
identificación y caracterización molecular de los antígenos responsables, su
producción en cantidades utilizables para el diagnóstico, y su manipulación
estructural para generar especies moleculares hipoalergénicas (Valenta et al.,
1999 a, b).

Introducción
I.2.- ALÉRGENOS DE ALIMENTOS VEGETALES Y PROTEÍNAS DE
DEFENSA.
Las alergias con mayor significación poblacional están producidas por
aeroaiergenos que toman contacto con el organismo por inhalación (proteínas
de pólenes, ácaros, epitelio de animales, etc.; Kay, 1997). Sin embargo, la
"percepción" y sensibilización a alimentos se está incrementando en los países
occidentales (Stanley y Bannon, 1999). En Estados Unidos, encuestas a
consumidores arrojan cifras de hasta un 30% de personas alérgicas (no
confirmado clínicamente) a algún producto alimentario (Sloan y Powers, 1986).
Se estima que la población europea afectada es de un 1-2%, siendo más
frecuente en niños (hasta un 8%; Brujinzeel-Koomen eí a/., 1995). En países
anglosajones, leche, huevo y cacahuete son los alimentos más implicados en
reacciones alérgicas durante los primeros años de vida, mientras cacahuete y
otros frutos secos, pescado y marisco son ios más comunes en adultos
(Stanley y Bannon, 1999). Estos perfiles pueden cambiar según los hábitos
alimentarios de los diferentes países. Así, por ejemplo, y en el caso de
alimentos de origen vegetal, la alergia a lentejas y garbanzos en niños (Crespo
et a/., 1995) y a frutas de la familia Rosaceae en adultos (Fernández-Rivas et
al., 1997) se encuentran entre las más frecuentes en España.
i.2.1.- Alérgenos de alimentos vegetales y reacciones cruzadas.
El número de alérgenos aislados y caracterizados de diferentes
organismos se ha incrementado de forma importante en la última década. Aún
así, el listado de Allergen Nomenclature (International Union of Immunological
Societies, Allergen Nomenclature Sub-Commitee, Marzo 2000) sólo incluye 40
alérgenos de alimentos de origen vegetal. Un análisis de sus propiedades
bioquímicas e inmunológicas no permite deducir ningún rasgo común asociado
a la inducción o reconocimiento de IgE. En el caso de alérgenos alimentarios
(Stanley y Bannon, 1999), se han sugerido como características generales, el

Introducción
ser proteínas con pesos moleculares menores de 70 kDa, abundantes en
alimentos, y resistentes a proteasas y tratamientos térmicos.
Un hecho frecuente en pacientes alérgicos a un determinado agente
(alimentos, pólenes, etc.) es su sensibilización a otras fuentes de alérgenos
vegetales. Estas reacciones cruzadas se han descrito entre agentes similares
de especies muy relacionadas, como pólenes de abedul y avellano (Valenta et
al., 1991) o frutas de la familia Rosaceae (Pastorello et al., 1994), y también
entre diferentes agentes de especies filogenéticamente distantes, como polen
de abedul y frutas de manzano (Ebner et al., 1991) o látex y diversas frutas
(aguacate, plátano, etc.; Blanco etal., 1994).
La reactividad cruzada está originada por la presencia en los agentes
implicados de alérgenos homólogos que comparten epítopos comunes (Vieths,
1997). La distribución de miembros de la familia Bet v 1 (el alérgeno principal de
polen de abedul) en diferentes pólenes, frutas y hortalizas (ver epígrafe 1.2.2 de
la Introducción), es uno de los ejemplos mejor documentados.
Un segundo tipo de epítopos que parece responsable de reacciones
cruzadas en agentes de origen vegetal son N-oligosacáridos complejos unidos
a residuos de asparagina, que se han detectado en muchas glicoproteínas de
plantas (Aalberse y van Ree, 1997). Los residuos de p-(1,2)-xilosa y/o a-(1,3)-
fucosa que forman parte de estos N-oligosacáridos contribuyen
significativamente a la unión de IgE específica y a las reacciones cruzadas
entre glicoproteínas vegetales, y entre éstas y las presentes en algunos
insectos (al menos, in vitro; García-Casado etal., 1996a; Batanero etal., 1999;
van Ree eí a/., 2000a).
I.2.2.- Proteínas de defensa y panalergenos vegetales.
Varias de las nuevas familias de alérgenos vegetales (principalmente de
frutas, hortalizas y pólenes) descritas en los últimos años pertenecen a las
denominadas proteínas de defensa o PRs (Pathogenesis Related; Salcedo eí
al., 1999). Estas proteínas están implicadas en los mecanismos de defensa
(inducidos o constitutivos) que poseen las plantas frente al ataque de plagas y

Introducción
patógenos. Su actividad antifúngica, antibacteriana o insecticida en ensayos in
vitro, su inducción in planta por la interacción con estos organismos, y/o una
mayor resistencia de plantas transgénicas con niveles altos de expresión del
transgen, han sustentando su potencial papel en protección vegetal (Shewry y
Lucas, 1997; IVIelchers y Striver, 2000). Los ejemplos más significativos de
proteínas de defensa relacionados con alergias producidas por alimentos
vegetales se resumen en la Tabla 1.
Tabla 1: Ejemplos de proteínas de defensa relacionadas con la alergia a alimentos vegetales.
Familia de proteínas/ Especie
Familia de Bet v 1 Manzana Pera Albaricoque Cereza Avellana Apio Zanatioria
Taumatina Manzana Cereza Pimienta
Proteínas de transferencia de lípidos Manzana Melocotón Albaricoque Ciruela Maiz
Heveina-Proheveina Nabo
Nombre
Mal d i P y r d Pruarl Pruavl Cora l Ap ig l D a u d
Mald2 Pru av 2 Capa 1
Mald3 Prup3 Pru ar 3 Prud3 Zea m 14
Brar2
Referencia
Vieths and Schoning, 1996 No. Accesión AF05730 No. Accesión U93165 Scheurereía/., 1997 Breitenederefa/., 1993 Breitenederefa/., 1995 Hoffmann-Sommergruberefa/., 1999
Hsielieía/., 1995 Inschlagefa/., 1998 EbnerCefa/., 1998
Sánchiez-Mongeefa/., 1999 Sánciiez-Monge etal., 1999 Pastorelloefa/.,2000 Pastorelloefa/., 1994 No. Accesión P 19656
Hanninenefa/., 1999
La caracterización de Bet v 1, el alérgeno principal de polen de abedul
(Breiteneder et al., 1989), permitió establecer su homología con proteínas de
defensa de guisante (familia PR-10). Posteriormente, se han identificado
proteínas de 16-18 kDa homologas a Bet v 1 en varios pólenes de árboles de la
familia Fagaceae (aliso, avellano, carpe; Valenta et al., 1991), y en varias frutas
de la familia Rosaceae (ver Tabla 1), así como en apio (Breiteneder et al., 1995)
y en zanahoria (Hoffmann-Sommergruber et al., 1999). Se ha demostrado la
presencia de epítopos comunes en Bet v 1 y sus homólogos de alimentos
(Frisch et al., 1997; Kazemi-Shirazi et al., 2000), lo que explicaría la

Introducción
cosensibilización a frutas (principalmente manzana) y hortalizas (apio,
zanahoria) que muestran pacientes polínicos alérgicos a abedul en el área
centroeuropea.
Un segundo grupo de proteínas de defensa (familia PR-5) implicadas en
alergia son las taumatinas. Hasta el momento, se han localizado miembros de
esta familia de 23-30 kDa con capacidad de ligar IgE específica en frutas
(manzana, cereza) y especias (pimienta) (ver Tabla 1). En este grupo se
requieren estudios adicionales que completen la caracterización bioquímica e
inmunológica de los potenciales alérgenos, así como las posibles reacciones
cruzadas entre los mismos.
Proteínas de Transferencia de Lípidos (LTPs) han sido identificadas
como alérgenos principales de frutas de la familia Rosaceae (Sánchez-Monge
et al., 1999; Pastorello et al., 1999 a, b y 2000) en poblaciones mediterráneas
no expuestas a polen de abedul (ver epígrafe 1.4.2 de esta Introducción).
Evidencia preliminar sugiere que las LTPs pueden ser también alérgenos
importantes en maíz (EA Pastorello, Comunication Oral, 7*' International
Symposium on Immunological, Chemical and Clinical Problems of Food Allergy,
Giardini-Naxo, Italia, 1998) y cerveza (Curioni etal., 1999).
Además de las tres familias de panalergenos citadas anteriormente,
otros tipos de proteínas de defensa han sido implicados en reacciones de
hipersensibilidad a alimentos. Tal es el caso de proheveína/heveína en nabo y
látex (ver apartado 1.3 de esta Introducción) y de inhibidores de a-amilasas
heterólogas presentes en harinas de cereales (García-Casado et al., 1996b;
James et al., 1997; Kusaba-Nakayama et al., 2000). En el caso de látex, un
candidato propuesto para explicar la cosensibilización con algunos alimentos es
el alérgeno Hev b 2, identificado como una p-1,3-glucanasa (familia PR-2;
Breiteneder y Scheiner, 1998).
La relación entre alérgenos de alimentos vegetales y proteínas de
defensa debe sustentarse con trabajos adicionales tanto a nivel estructural
como epidemiológico. Sin embargo, los datos ya disponibles permiten
vislumbrar la incidencia de dicha relación en aspectos tales como:

Introducción
a.- Sensibilización cruzada de pacientes alérgicos a diferentes
alimentos y/o pólenes. La mayoría de las familias de proteínas de
defensa tienen una distribución ubicua entre las diferentes especies de
plantas cultivadas. En este contexto, será esencial comparar la
reactividad de distintos miembros de una misma familia de proteínas en
diferentes especies y/o tejidos de la planta.
b.- Los cambios en el nivel de expresión de algunas proteínas de
defensa con estrés biótico o abiótico pueden suponer variaciones
significativas de la cantidad de alérgeno en función de las condiciones de
recolección y almacenamiento, así como de los tratamientos para la
elaboración del producto final.
c- Por último, las propiedades alergénicas de algunas proteínas
de defensa deben considerarse en los programas de obtención de
plantas transgénicas resistentes a determinadas plagas o patógenos.
I.3.- SÍNDROME LÁTEX-FRUTAS
1.3.1.- Alergia al látex. Alérgenos caracterizados.
El látex obtenido del árbol del caucho {Hevea brasiliensis) es
ampliamente utilizado en la producción de material sanitario (guantes,
catéteres, etc.), así como en muchos de los productos de uso cotidiano
(balones, globos, chupetes, etc.). Su implantación en nuestra sociedad ha ido
acompañada de un aumento de la alergia a este producto, pasando a ser una
de las alergias ocupacionales más importantes (Niggeman y Breiteneder,
2000). Esta alergia afecta, principalmente, a personal de los centros de la salud,
o trabajadores de invernaderos, así como también a pacientes que requieren
operaciones reiteradas o que pasan largas temporadas en los hospitales. Se
estima que más del 18% de los trabajadores de la salud, así como el 50% de
los niños con espina bífida, sufren procesos adversos provocados por látex
(Posch et al., 1998). En general, la prevalencia de esta alergia es del 4,2% en
Introducción
Alemania, y cerca del 4,5% en Canadá por prueba cutánea, aunque muchos
Individuos no presentan síntomas clínicos (Fuchs etal., 1998).
La vía de sensibilización al látex puede ser cutánea, a través de
mucosas o heridas, o aérea, inhalando el polvo de los guantes que puede
adsorber proteínas de éstos (Posch et al., 1998). Los síntomas clínicos de esta
alergia van desde urticaria localizada y rinoconjuntivitis hasta asma, y más
raramente anafilaxia (Smedley, 2000).
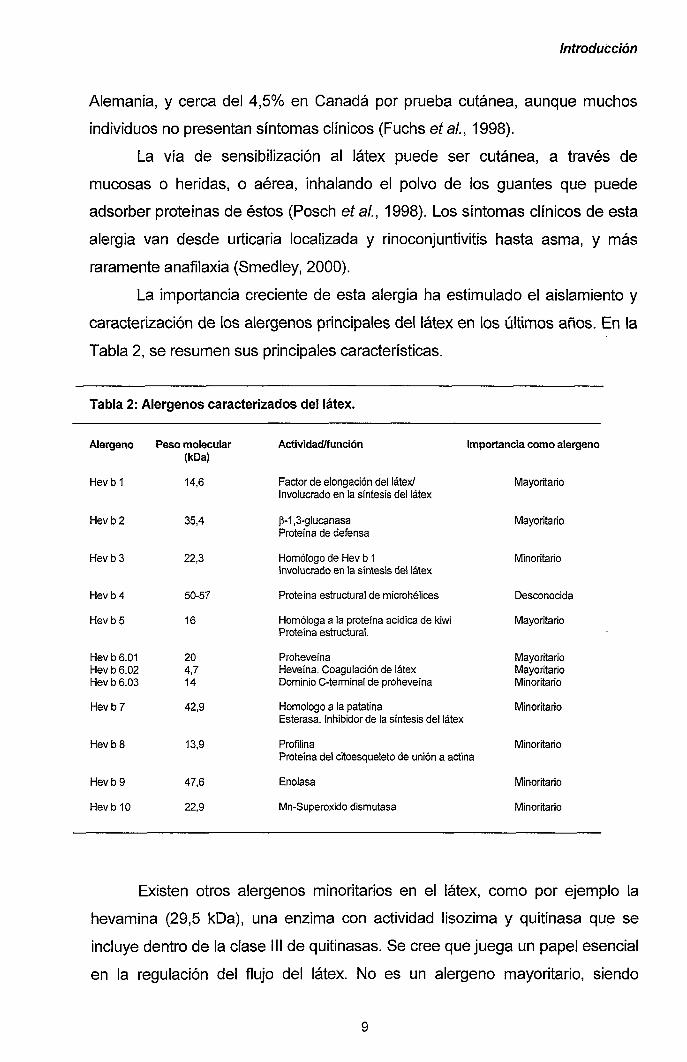
La importancia creciente de esta alergia ha estimulado el aislamiento y
caracterización de los alérgenos principales del látex en los últimos años. En la
Tabla 2, se resumen sus principales características.
Tabla 2:
Alérgeno
Hevb1
Hevb2
HevbS
Hevb4
HevbS
HevbS.OI Hev b 6.02 Hev b 6.03
Hevb7
HevbS
Hevb9
Hev b 10
Alérgenos caracterizados del látex.
Peso molecular (kDa)
14,6
35,4
22,3
50-57
16
20 4,7 14
42,9
13,9
47,6
22,9
Actividad/función Importancia como alérgeno
Factor de elongación del látex/ Involucrado en la síntesis del látex
p-1,3-glucanasa Proteína de defensa
Homólogo de Hev b 1 Involucrado en la síntesis del látex
Proteína estructural de microhélices
Homologa a la proteína acídica de kiwi Proteína estmctural.
Proheveína Heveína. Coagulación de látex Dominio C-terminal de proheveína
Homologo a la palatina Esterasa. Inhibidor de la síntesis del látex
Profilina Proteína del citoesqueleto de unión a actina
Enolasa
Mn-Superoxido dismutasa
Mayoritario
Mayoritario
IVIinoritario
Desconocida
Mayoritario
Mayoritario Mayoritario Minoritario
Minoritario
Minoritario
Minoritario
Minoritario
Existen otros alérgenos minoritarios en el látex, como por ejemplo la
hevamina (29,5 kDa), una enzima con actividad lisozima y quitinasa que se
incluye dentro de la clase III de quitinasas. Se cree que juega un papel esencial
en la regulación del flujo del látex. No es un alérgeno mayoritario, siendo

Introducción
reconocido únicamente por el 3,4% de los pacientes alérgicos (Alenius eí al.,
1995). También se han descrito como alérgenos minoritarios otras qultinasas
de látex (Breiteneder y Scheiner, 1998).
Un aspecto a destacar en la alergia al látex, es el patrón diferencial de
alérgenos principales a los que están sensibilizados los distintos grupos de
población afectados. Así, por ejemplo, en pacientes que han sido sometidos a
reiteradas operaciones (p.e. niños con espina bífida), Hev b 1 y Hev b 3 son los
alérgenos más importantes. Ambos comparten un 73% de identidad de
secuencia. Hev b 1 es reconocido (prueba cutánea) por un 80% de los
pacientes de este grupo sensibilizados al látex (Chen et al., 1997a). Por el
contrario, en personal sanitario afectado por látex, la proheveína (Hev b 6.01) y
su domino N-terminal, denominado heveína (Hev b 6.02) son los mayoritarios.
Más del 80% de estos profesionales reconocen éstos alérgenos, mientras que
sólo lo hacen un 27% de pacientes alérgicos con espina bífida (Alenius eí al.,
1995).
El reconocimiento diferencial de alérgenos entre los distintos grupos que
padecen alergia al látex puede explicarse por factores como la edad cuando se
produce la sensibilización, y, sobre todo, la vía por la cual se induce ésta. El
personal sanitario suele tomar contacto con los alérgenos del látex vía
respiratoria o por la piel, mientras que es muy probable que los enfermos de
espina bífida se sensibilicen por las mucosas o por contacto directo entre los
alérgenos y sangre (Posch etal., 1998).
I.3.2.- Reacciones cruzadas de látex y alimentos: Síndrome látex-
frutas.
La alergia al látex ha sido asociada a otras alergias, como polinosis a
Artemisia y/o Ambrosia (Fuchs eí al., 1998; Levy eí ai, 2000). Sin embargo, la
asociación más importante y más ampliamente documentada es entre el látex y
ciertas frutas. Se estima que más del 50% de los pacientes alérgicos al látex
están sensibilizados a una o más frutas, siendo un número alto de los casos
descritos como reacciones de anafilaxia. Dada la importancia de esta
10

Introducción
asociación, en 1994, se propuso llamarla síndrome látex-frutas (Blanco et al.,
1994).
Las frutas más claramente asociadas al síndrome son aguacate,
castaña, plátano y kiwi. También se han descrito casos de reacciones cruzadas
entre el látex y otras frutas, como melocotón, pina, fruta de la pasión, uva,
guayaba, chirimoya, papaya, mango, melón y frutos secos (almendra, piñones y
cacahuete; Brehiereía/., 1997).
Como se puede observar, en este síndrome se asocian especies muy
distantes filogenéticamente, lo cual implica la presencia de antígenos comunes
en todos ellas. Sin embargo, al inicio de esta Tesis, no se habían identificado
los alérgenos de las reacciones cruzadas entre látex y frutas. Los datos
preliminares más fiables estaban basados en inmunodetecciones de extractos
crudos de frutas usando sueros de pacientes alérgicos a látex y alimentos.
Wadee et al., (1990) detectaron una banda reactiva de 30 kDa en melocotón,
mandarina, cereza, guayaba, plátano y látex. Lavaud et al., (1995) identificaron
un potencial alérgeno mayoritario de 30 kDa en látex, aguacate y plátano. En
esta última fruta, otros grupos han detectado también posibles alérgenos de 30-
37 kDa que cruzaban con látex (Alenius etal., 1996b; Delbourg etal., 1996). En
ninguno de los casos citados fueron purificados y caracterizados los
componentes reactivos de 30 kDa. Las propuestas de algunos tipos de
proteínas, que incluían lisozimas o proteasas del tipo papaína, como
responsables de la cosensibilización con látex (Brehier et al., 1997) estaban
basadas en la presencia de las mismas en algunos frutos asociados al
síndrome, y/o en látex, sin una evidencia bioquímica, inmunológica y clínica
complementaria.
I.3.3.- Heveína y proteínas con dominio heveína.
La proheveína, como ya se ha comentado, constituye uno de los
alérgenos más importantes del látex (Alenius et al., 1995, 1996a; Chen et al.,
1997b). Se sintetiza como un precursor de 204 aminoácidos, siendo los 17
primeros del N-terminal el péptido señal. El producto que incluye los 187
11

Introducción
restantes, pierde 14 aminoácidos del C-termina! dando lugar a la proheveína de
20kDa, que es reconocida por los pacientes alérgicos. Sin embargo, esta
proteína puede sufrir una escisión entre los residuos 49 y 50, dando lugar a dos
péptidos, el N-terminal de 4,7 kDa y el dominio C-terminal de 14 kDa. A su vez,
el primero pierde de 4 a 6 aminoácidos de su C-terminal convirtiéndose en la
heveína. El dominio C-terminal puede hidrolizarse tras la separación. Se cree
que su función es servir como señal de localización vacuolar, lo cual sería
coherente con que la heveína se encuentre principalmente en lutoides
derivados de vacuolas (Soedjanaatmadja et al., 1995).
Tanto la heveína como el domino C-terminal de la proheveína han sido
identificados también como alérgenos, denominándose Hev b 6.02 y Hev b
6.03, respectivamente. Sin embargo, parece que es en la primera donde se
localiza la mayor alergenicidad (Alenius et al., 1996a; Chen et al., 1997b), ya
que sólo del 15 al 21% de los pacientes sensibilizados a la proheveína
reconocen el dominio C terminal por inmunodetección y ELISA (Alenius eí al.,
1996a). De hecho, de los 6 epítopos secuenciales mapeados en la proheveína,
tres se localizan en el domino N-terminal, los cuales además son aquellos que
fijan más IgE de pacientes alérgicos al látex (Beezhold etal., 1997).
La heveína es un pequeño péptido de 43-45 aminoácidos, entre los
cuales se incluyen 8 cisteínas que forman 4 puentes disulfuro. Su estructura
tridimensional, determinada por NMR, consiste en una triple lámina p con una
pequeña a-hélice conectando la tercera de estas láminas (Asensio et al., 1995;
1997). Se incluye dentro de la familia de proteínas de unión a quitina. La quitina
es el componente principal de la pared celular de los hongos, así como del
exoesqueleto de los artrópodos. De hecho, se ha descrito que la heveína puede
inhibir el crecimiento de hongos (van Parijs et al., 1991), y que su síntesis es
inducible por ataque de patógenos, además de por herida y etileno (Broekaert
et al., 1997). Estas características han conducido a proponer su función como
proteína de defensa, aunque también se ha sugerido su papel en la
coagulación del látex (Gidrol etal., 1994).
Se han descrito proteínas homologas a la heveína (o proheveína) en
algunas especies vegetales como Arabidopsis thaliana (Potter et al., 1993),
12

Introducción
nabo {Brassica napis; Hanninen et al., 1999) y saúco {Sambucus nigra; Van
Damme et al., 1999). En este último caso, el precursor de la heveína tiene una
extensión C-terminal homologa al dominio catalítico de las quitinasas de clase
V, aunque no se ha demostrado su actividad enzimática. De los tres ejemplos
citados anteriormente, la proheveína de nabo es el único estudiado en el
contexto de alergia. Esta proteína se induce por herida y/o etileno, y es
reconocida por sueros de pacientes alérgicos al látex en extractos de plantas
tratadas por estos agentes, mientras que en muestras de plantas no tratadas,
no hay reconocimiento de ningún componente (Hanninen et al., 1999). Estos
datos sugieren que algunos homólogos a la heveína son putativos alérgenos, y
que su expresión puede favorecerse con tratamientos hormonales o estrés.
Como ya se ha mencionado, la heveína y sus homólogos pertenecen a
la familia de proteínas de unión a quitina. Dentro de esta familia se incluyen
lectinas de diferentes tipos: las merolectinas, o proteínas con un solo dominio
heveína o dominio heveína truncado; y las halolectinas, que están compuestas
por 2, 3, 4, ó 7 dominios homólogos a la heveína. Este es el caso de la
aglutinina de germen de trigo que está formada por dos monómeros, cada uno
de los cuales tiene 4 repeticiones en tándem de un domino homólogo a la
heveína. Por otra parte, las lectinas de las solanáceas están compuestas por un
domino N-terminal de unión a quitina, formado por tres repeticiones de heveína,
ligado a una extensión rica en 0-glicosilaciones (Van Damme etal., 1999).
En otras proteínas, el domino heveína aparece ligado a un dominio con
función enzimática. Este es el caso de las endoquitinasas de clase I, IV y V.
Las endoquitinasas son enzimas ampliamente distribuidas en la naturaleza.
Catalizan la hidrólisis al azar de enlaces internos (3-1,4 glicosídicos de los
polímeros de quitina. En algunas quitinasas se ha demostrado su actividad
inhibitoria frente al crecimiento de hongos (Schiumbaum et al., 1986; Broekaert
etal., 1986; Mauch etal., 1988), del mismo modo, que se ha demostrado que al
ser sobreexpresadas en plantas transgénicas aumentan la resistencia de éstas
a hongos patógenos (Broglie et al., 1991; Jach et al., 1995; Grisson et al.,
1996). Actualmente, las quitinasas están clasificadas en las familias PR-3, PR-
4, PR-8 y PR-11 de proteínas de defensa (Van Loon et al., 1999).
13

Introducción
Heveína ^ ^ ^ ^
Clase I \/ff^
Clase II
Clase III
Clase IV
0'\/\j>\j^/-^,r\j^ / v > v V \ .A. / \ / \ ^ ^ ^ / \
^
«-ev ^ 3 ^ Figura 2: Esquema de las clases de quitinasas. Los recuadros con entramado de líneas simbolizan el dominio heveína; los recuadros blancos son dominios catalíticos homólogos; el recuadro con entramado ondulante simboliza el dominio catalítico característico de las quitinasas de clase III.
Basándose en su estructura primaria, se pueden agrupar las quitinasas
en cinco clases (Fig. 2; Collinge etal., 1993; Meins etai, 1994). Las clases i, II,
y IV se engloban dentro de la familia 19 de glicosil-hidrolasas. Tienen dominios
catalíticos muy similares entre sí, aunque en las de tipo IV, el dominio catalítico
ha sufrido deleciones. Los dominios catalíticos de las quitinasas de clase I y II
pueden llegar a tener hasta un 80% de identidad. Por otra parte, las clase I y IV
tienen una extensión N-terminal homologa a la heveína de látex .
La clase III de quitinasas agrupa a enzimas de hongos, bacterias,
plantas y animales, siendo incluidas en la familia 18 de las glicosil-hidrolasas.
Las quitinasas de clase V tienen un dominio catalítico homólogo a las de
tipo I y II, aunque en su extensión N-terminal aparecen dos repeticiones
homologas a la heveína (Meins etal., 1994; Neuhaus etal., 1996).
Dentro de las clases de quitinasas con dominio heveína, el de las de
clase I tiene un alto grado de identidad con la heveína del látex. El tamaño
molecular de estas enzimas oscila entre 30 y 35 kDa. Teniendo en cuenta que
en extractos de aguacate, plátano y otras frutas era reconocida una banda de
30 kDa por sueros de pacientes alérgicos a látex y frutas, y dada la homología
de secuencia entre la heveína y el dominio N-terminal de las quitinasas de
clase I, estas proteínas eran candidatos plausibles para explicar las reacciones
cruzadas en el síndrome látex-frutas.
14

Introducción
I.4.- ALERGIA A FRUTOS DE LA FAMILIA Rosaceae Y PROTEÍNAS DE
TRANSFERENCIA DE LÍPIDOS (LTPs).
Los frutos de especies de la familia Rosaceae son los alimentos que
más frecuentemente se han implicado en alergias de adultos producidas por
vegetales. Estas frutas se han asociado a tres modelos de hipersensibilidad:
reacciones cruzadas con pólenes de fagáceas, donde los alérgenos
responsables son proteínas de la familia de Bet v 1 (Fritsch eí al., 1997);
reacciones cruzadas a pólenes de gramíneas, siendo la familia de las profilinas
presuntamente responsables de dichas reacciones (van Ree et al., 1995); y
alergia a frutas con o sin polinosis asociada, donde las proteínas de
transferencia de lípidos (LTPs) son alérgenos principales (Sánchez-Monge et
al., 1999).
Los pólenes implicados en la asociación de la polinosis y frutas de
rosáceas dependen del área geográfica. En centro y norte de Europa, esta
asociación es principalmente entre polen de abedul (y/u otras fagáceas),
mientras que en el sur de Europa, la alergia a rosáceas se asocia
principalmente a gramíneas (Van Ree etal., 1995; Cuesta-Herranz etal., 1999).
1.4.1.- Síndrome de polen de abedul-frutas-hortalízas: Bet v 1.
Como ya se ha comentado, en el centro y norte de Europa, la reacción
cruzada más importante es el síndrome polen de abedul, frutas (principalmente
manzana) y otros alimentos de origen vegetal. Prácticamente el 70% de los
pacientes que sufren polinosis a abedul también están sensibilizados a frutos
de rosáceas. Los síntomas clínicos de esta alergia, en más del 78% de los
casos, se engloban dentro de lo que se conoce como síndrome de alergia oral
(SAO), y suele producirse a los pocos minutos de la ingestión del alimento; Por
regla general, el SAO va precedido, temporalmente, de polinosis, por lo que se
piensa que la sensibilización ai polen de abedul trae como consecuencia la
sensibilización a las frutas (Ortolani etal., 1988).
15

Introducción
El alérgeno mayoritario del polen de abedul es Bet v 1 (Breiteneder et al.,
1989), siendo reconocido por el 95% de los pacientes que sufren alergia a este
polen. Se han descrito reacciones cruzadas con otros pólenes, generalmente
de especies de la familia de las fagáceas (Ipsen y Hansen, 1991; Valenta et al.,
1991), y también, a frutos de rosáceas y otros alimentos (Breiteneder et al.,
1995; Vieths et al., 1995; Vieths y Schoning, 1996). En todas las especies
implicadas en la reacción cruzada, se han encontrado proteínas homologas a
Bet V 1, lo que explicaría por qué la sensibilización a estos pólenes puede
inducir la sensibilización a determinados alimentos (Deviller y Pauli, 1997).
Un segundo alérgeno del polen del abedul, que no tiene tanta relevancia
en la polinosis, es Bet v 2, reconocido por el 20% de los pacientes (Valenta et
al., 1992). Bet v 2 se engloba dentro de la familia de las profilinas, proteínas de
13-14 kDa, altamente similares, cuya función biológica es formar complejos con
la actina, regulando su polimerización, y participando así en la forma y
movimientos celulares. Miembros de esta familia han sido identificados en
pólenes de gramíneas y Artemisia, así como en muchos alimentos. Estas
proteínas podrían ser las responsables de las asociaciones entre alergia a
polen de abedul y a pólenes de gramíneas y/o Artemisia, ya que las profilinas
de polen de estas especies comparten hasta un 80% de identidad de
secuencia. También, han sido identificadas profilinas en manzana y otras
rosáceas, así como en avellana, apio y zanahoria, que podrían estar implicadas
en la asociación entre alergias a pólenes, frutas y hortalizas (van Ree eí al.,
1995).
I.4.2.- Alergia a rosáceas en el área mediterránea: LTPs.
La alergia alimentaria a rosáceas es una de las más frecuentes en
población adulta en España e Italia, suponiendo el 2% de los individuos que
tienen algún tipo de alergia a alimentos (Pastorello et al., 1997). La mayoría de
los pacientes con este tipo de alergia también están sensibilizados a pólenes.
Sin embargo, un 15-21% de alérgicos a frutos de rosáceas no presenta
polinosis asociada (Ortolani et al., 1988).
16

Introducción
Los síntomas clínicos son distintos para aquellos pacientes que tienen
alergia a rosáceas y pólenes, de los que sólo están sensibilizados a frutas. En
el primer caso, las manifestaciones suelen estar englobadas dentro del
síndrome de alergia oral; en cambio, los alérgicos sin polinosis presentan una
entidad clínica severa con un alto porcentaje de anafilaxia (Fernández-Rivas et
al., 1997). Por otro lado, los pacientes alérgicos a rosáceas presentan una
tolerancia distinta si la misma fruta es ingerida pelada o sin pelar. El 44% de los
pacientes alérgicos a manzana y el 41% de los pacientes alérgicos a pera
presentaban síntomas cuando ingerían dichas frutas con piel, pero las
toleraban cuando lo hacían peladas (Fernández-Rivas y Cuevas, 1999).
En el área mediterránea, libre de polen de abedul y otras fágales en la
mayoría de sus regiones, la sensibilización a Bet v 1 (o sus homólogos) es
mínima en pacientes alérgicos a frutas de la familia Rosaceae (porcentaje de
reconocimiento de un 6% en Madrid; Cuesta-Herranz et al., 1999). Los
alérgenos principales en estas poblaciones son LTPs de las rosáceas, en
pacientes con o sin polinosis asociada. En pacientes con polinosis asociada, se
ha propuesto un papel adicional de las profilinas (reconocidas en un 44% de los
casos en pruebas cutáneas; Van Ree etal., 1995).
Las LTPs se han identificado como los alérgenos más importantes en
melocotón y manzana (Sánchez-Monge et al., 1999; Pastorello et al., 1999 a y
b) y albaricoque (Pastorello et al., 2000), aunque es muy posible, que en otras
especies de la familia Rosaceae, existan proteínas de esta familia implicadas
en reacciones alérgicas (Pastorello etal., 1994; Fernández-Rivas etal., 1997).
Las LTPs son una familia de proteínas de plantas de 7 a 9 kDa. En su
secuencia, se encuentran 8 cisteínas que forman 4 puentes disulfuro, lo que les
confiere gran estabilidad. Esto podría explicar su resistencia al calor, ya que las
LTPs persisten en zumos, néctares y otros derivados de frutas de rosáceas
(Brenna et al., 2000). Este grupo de proteínas es muy heterogéneo,
presentando una identidad de secuencia de menos del 30%. Solamente se
conservan las ocho cisteínas y 12 posiciones, las ocupadas por residuos
hidrofóbicos o aromáticos. Sin embargo, la estructura terciaria, constituida por
17

Introducción
cuatro a-hélices unidas por brazos flexibles, está muy conservada (Kader,
1996).
El papel de las LTPs de plantas ha sido muy controvertido. Aunque
inicialmente se propuso que participaban en la transferencia intracelular de
lípidos, su localización ha hecho dudar de esta función. Actualmente, se supone
que actúan como proteínas de defensa, y, tal vez, en las síntesis de cutina en la
superficie de algunos órganos (Kader, 1996; García-Olmedo et al., 1995).
Estas proteínas se expresan como preproteína, siendo el N-terminal un
péptido señal de transporte a membrana. Se han descrito LTPs prácticamente
en todos los tejidos, aunque los niveles más altos se suelen asociar a las capas
epidérmicas y periféricas de los órganos, lo que puede estar asociado a su
capacidad de inhibir el crecimiento de algunos microorganismos o su relación
con respuesta frente a estrés osmótico o de sequía. En algunos miembros de
esta familia se ha comprobado su actividad antimicrobiana, aunque LTPs de
distintas especies vegetales se pueden comportar de forma diferente (Molina et
al., 1993 a, b). Del mismo modo, se ha demostrado que los genes que codifican
LTPs son inducibles por distintos factores ambientales como temperaturas
extremas, estrés salino o sequía (Kader, 1996).
Por tanto, las LTPs son un grupo de proteínas ampliamente distribuidas
en el mundo vegetal, y si bien no mantienen una alta identidad de secuencia
entre especies de distintas familias, si conservan básicamente su estructura
tridimensional. Estas características, junto a su gran resistencia térmica,
sustentan la importancia de las LTPs como potenciales panalergenos (Asero,
1999).
18

Introducción
I.5.-OBJETIVOS DE LA TESIS
Si bien la alergia alimentaria está incrementando su importancia clínica,
el número de alérgenos aislados y caracterizados de fuentes vegetales es aún
muy limitado. La identificación de los componentes alergénicos de un alimento
puede ayudar a mejorar tanto el diagnóstico como la terapia utilizados, y
contribuir a entender mejor los casos de cosensibilización con otros alimentos
y/o con pólenes.
Entre los alérgenos vegetales identificados en la última década, más de
la mitad están incluidos en alguna de las familias de proteínas de defensa. Este
hecho ha llevado a considerar nuevos aspectos relacionados con la alergia
alimentaria, como son los posibles cambios en los niveles de alérgenos en
respuesta a estrés bióticos, tratamientos hormonales, condiciones de
almacenamiento, etc., y a prevenir posibles efectos deletéreos en el consumo
humano de productos transgénicos que sobreexpresen este tipo de proteínas.
En este contexto, el objetivo general de esta Tesis ha sido estudiar las
implicaciones en reacciones alérgicas por ingestión de alimentos vegetales, de
dos familias de proteínas de defensa, quitinasas de clase I y proteínas de
transferencia de lípidos (LTPs). La primera se ha analizado con relación al
síndrome látex-frutas, y la segunda en grupos de pacientes alérgicos a frutas de
rosáceas. Este objetivo general se ha abordado tratando de cubrir los
siguientes objetivos específicos:
1.- Purificar y caracterizar a los alérgenos mayoritarios de castaña,
plátano y aguacate en el contexto del síndrome látex-frutas.
2.- Determinar la alergenidad de las quitinasas de clase I frente a las de
clase II, tanto en pruebas in vitro como in vivo.
3.- Estudiar la implicación de las quitinasas de clase I en la
sensibilización a otros frutos asociados con la alergia a látex.
Determinar, también, el posible papel de los N-oligosacáridos complejos
en estas reacciones cruzadas.
19

Introducción
4- Comparar el patrón de reconocimiento de sueros de pacientes
alérgicos a látex pero no frutas, frente al patrón de sueros de pacientes
con síndrome látex-frutas.
5.- Estudiar el comportamiento de las quitinasas alergénicas frente a
tratamientos térmicos, y su posible inducción con etileno.
6.- Evaluar el papel de los dominios heveína y catalítico de las quitinasas
de clase I en sus propiedades alergénicas. Para ello se obtendrán las
proteínas recombinantes correspondientes a la enzima completa y a sus
dos dominios, expresando los clones de cada uno de estos productos en
la levadura Pichia pastorís.
1 - Purificar LTPs de especies alejadas filogenéticamente de la familia
Rosaceae, como son el polen de Artemisia vulgaris y la semilla de
Castanea sativa (ambas relacionadas en distinta medida con la alergia a
melocotón). Comparar la capacidad alergénica de estas proteínas, con
las que exhiben los homólogos de manzana y melocotón, en ensayos in
vitro.
8.- Estudiar la reactividad in vivo (pruebas cutáneas) de las cuatro LTPs
mencionadas, analizando un grupo amplio de pacientes alérgicos a
melocotón.
9.- Aislar y secuenciar los clones de ADNc de las LTPs de manzana y
melocotón.
20

Matertaíy Métodos

Material y Métodos
11.1.- MATERIAL BIOLÓGICO
11.1.1.- Material vegetal.
En este trabajo se han utilizado semillas de castaño {Castanea sativa
Mili.), y frutos de aguacate {Persea americana L.), y plátano canario {Musa x
paradisiaca L., híbrido de M. acuminata Colla x M. balbisiana Colla) para la
purificación de quitinasas de clase I y II.
Para determinar la implicación de estas enzimas en el síndrome látex-
frutas, se empleó extractos de chirimoya {Annona clierimola Mill.j, fruta de la
pasión {Passiflora edulis Sims.), kiwi {Actinidia ctiinensis Planch.), pina
{Annanas comosus (Stickm.) Merr.), papaya {Carica papaya L.), mango
{Mangifera indica L.), melocotón {Prunus pérsica (L.) Batsch.), pera común
{Pyrus communis Bunga), níspero {Mespilus germánica L.), melón {Cucumis
meló L.), uva {Vitis vinifera L.), tomate {Lycopersicon esculentum Mil!.),
patata {Solanum tuberosum L), pistacho {Pistacia vera Mili.), almendra
{Prunus dulcis L.), cacahuete {Aractiis liypogaea L.), nuez {Juglans regia L.),
piñón {Pinus pinea L.), látex del árbol del caucho {Hevea brasiliensis (H.B.K.)
Muell. Arg.) y trigo {Triticum aestivum (L.) Thell.).
Para el estudio del efecto de los tratamientos con etileno y calor de las
quitinasas de clase I, fueron utilizados frutos de judía Pliaseolus vulgaris L.,
cultivar lluro, recogidas a las 8 semanas (vainas; judía verde) o a las 14
semanas (semillas maduras; judía blanca). El tratamiento con etileno se
realizó pulverizando sobre las judías verdes una solución de etephon (ácido
2-cloroetil-fosfónico; Sigma;1mg/ml) o agua (control), y manteniendo las
vainas tratadas en bolsas de celofán selladas. Las muestras se congelaron y
liofilizaron a las 24 horas (etileno 24 h), 48 horas (etileno 48 h) y 72 horas
(etileno 72 h). El tratamiento térmico se realizó mediante cocción en agua
hirviendo de las muestras de judía verde durante 15 minutos.
Para purificar LTPs, se partió de piel liofilizada de melocotón y de
manzana {Maius domestica Borkh.), semillas de castaño y polen de
artemisia {Artemisia vulgaris L.).
22
Material y Métodos
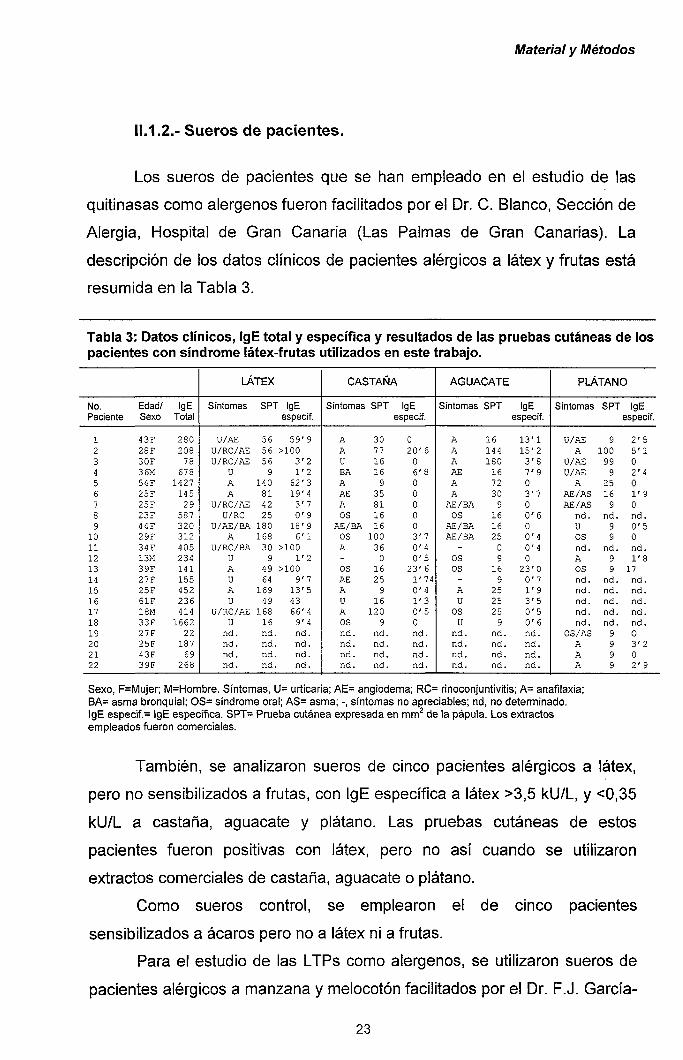
11.1.2.- Sueros de pacientes.
Los sueros de pacientes que se han empleado en el estudio de las
quitinasas como alérgenos fueron facilitados por el Dr. C. Blanco, Sección de
Alergia, Hospital de Gran Canaria (Las Palmas de Gran Canarias). La
descripción de los datos clínicos de pacientes alérgicos a látex y frutas está
resumida en la Tabla 3.
Tabla 3: Datos clínicos, IgE total y específica y resultados de las pruebas cutáneas de los pacientes con síndrome látex-frutas utilizados en este trabajo.
No. Paciente
1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18 19 20 21 22
Edad/ Sexo
43F 28F 30F 36M 54F 25F 25F 23F 44F 29F 34F 13M 39F 27F 25F 61F 18M 33F 27F 25F 43F 39F
IgE Total
280 208
78 678
1427 145
29 587 320 312 405 234 141 155 452 236 414
1662 22
187 69
268
LÁTEX
Síntomas
U/AE U/RC/AE U/RC/AE
U A A
U/RC/AE U/RC
U/AE/BA A
0/RC/BA U A U A U
U/RC/AE U
nd . nd . nd . nd .
SPT
56 56 56
9 140
81 42 25
180 158
30 9
49 64
169 49
168 16 nd . nd . nd . nd .
IgE específ.
5 9 ' 9 >100
3 ' 2 1'2
5 2 ' 3 1 9 ' 4
3 ' 7 0 ' 9
1 8 ' 9 5 ' 1
>100 1'2
>100 9 ' 7
1 3 ' 5 43 6 6 ' 4
9 '4 nd . nd . nd . nd .
CASTAÑA
Síntomas
A A U BA A AE A OS
AE/BA OS A
-OS AE A U A OS nd . nd . nd . nd .
SPT
30 77 16 16
9 35 81 16 16
100 36
0 16 25
9 15
120 9
nd . nd . nd . nd .
IgE específ.
0 2 0 ' 5
0 6 ' 8 0 0 0 Q 0 3 ' 7 0 ' 4 0 ' 5
2 3 ' 6 1 '74 0 ' 4 1 '3 0 ' 5 0 nd . nd . n d . nd .
AGUACATE
Síntomas SPT
A A A AE A A
AE/BA OS
AE/BA AE/BA
-OS OS
-A U
OS U
nd . nd . nd . nd .
16 144 180
16 72 30
9 16 16 25
0 9
16 9
25 25 25
9 n d . nd . n d . nd .
IgE específ.
1 3 ' 1 1 5 ' 2
3 ' 8 7-9 0 3 ' 7 0 0 ' 6 0 0 ' 4 0 ' 4 0
2 3 ' 0 0 ' 7 1 '9 3 ' 5 0 ' 5 0 ' 6 nd . nd . nd . nd .
PLÁTANO
Síntomas
U/AE A
U/AE U/AE
A AE/AS AE/AS
nd. U OS nd . A OS nd . nd . nd . nd . nd .
OS/AS A A A
SPT
9 100
99 9
25 16
9 nd .
9 9
nd . 9 9
nd . nd . nd . nd . nd .
9 9 9 9
IgE específ.
2 ' 5 5 ' 1 0 2 ' 4 0 1 '9 0 nd . 0 ' 5 0 nd . 1'8
17 nd . nd . nd . nd . nd . 0 3 ' 2 0 2 ' 9
Sexo, F=Mujer; M=Hombre. Síntomas, U= urticaria; AE= angiodema; RG= rinoconjuntivitis; A= anafilaxia; BA= asma bronquial; 0S= síndrome orai; AS= asma; -, síntomas no apreciables; nd, no determinado. IgE especif.= IgE específica. SPT= Prueba cutánea expresada en mm^ de la pápula. Los extractos empleados fueron comerciales.
También, se analizaron sueros de cinco pacientes alérgicos a látex,
pero no sensibilizados a frutas, con IgE específica a látex >3,5 kU/L, y <0,35
kU/L a castaña, aguacate y plátano. Las pruebas cutáneas de estos
pacientes fueron positivas con látex, pero no así cuando se utilizaron
extractos comerciales de castaña, aguacate o plátano.
Como sueros control, se emplearon el de cinco pacientes
sensibilizados a ácaros pero no a látex ni a frutas.
Para el estudio de las LTPs como alérgenos, se utilizaron sueros de
pacientes alérgicos a manzana y melocotón facilitados por el Dr. F.J. García-
23

Material y Métodos
Selles (Departamento de Alergología, Hospital Virgen de la Arrixaca, Murcia)
y la Dra. M. Fernández-Rivas (Sección de Alergia, Fundación Alcorcen,
Madrid). Los datos clínicos de los pacientes alérgicos a frutos de rosáceas
se resumen en la Tabla 4. Como sueros negativos fueron empleados los de
cinco pacientes con IgE específica y prueba cutánea negativa a frutas de
rosáceas.
24

Tabla 4: Datos clínicos, IgE total y específica y resultados de las pruebas cutáneas de los pacientes con alergia a manzana y melocotón analizados en este trabajo.
Melocotón Manzana Castaña Artemisia
P a c i e n t e
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 35 37 38 39 40 41 42 43 44 45 46 47
E d a d / Sexo
14F 12F 17F 13F 12M 28M 27F 29F 30F 15F 20M 26M 25F 14F 6F
21M 13M 6M 17M 24F 21F 12M 11M 25F 23F 37F 31M 29M 26M 19F 29M 25F 20F 30F 25F 29F 30F 30F 33F 15M 11F 24M 19F 26M 31F 17M 20F
IgE to ta l (ku/1)
63.6 35.6 58.8 46.4 583 222 221 396 158
1340 127 394 85.3 475 44.1 nd 100 141 13.4 454 24.9 834 179 71.7 66 151 454 970 441 294 471 19.2
>2000 89.9 566 406 152 693 1165 412 199 146 356 326 313 475 235
P o l i n o s i s
No No No No Si Si Si Si Si Si Si Si Si Si No Si Si Si Si Si No No Si No Si Si Si SI Si Si SI Si Si Si Si Si Si Si Si Si Si Si Si Si Si Si Si
s í n t o m a s
OAS U U A U U
OAS OAS
U U U A U U
OAS CU U A U U A
CU CU A U U U U A U A
CU A A U A U A
OAS U U U
CU OAS
U U A
SPT
63.9 46.4 15.5
107.9 20.4 142
-27.9 64.2
-82.5 51.6 70.1 73.4 27.3 81
38.2 43.6 87.7 34
123.8 17.1 35.9 104.5 96.4 18.9
127.6 28.1 36.2 26.8 67.8 35.1 41.9 16.1 68.5 90.7 80.7 36.9 51.1 69.3 19.5 24.4 41.9 17.9 43.9 74.2 32
I g E E s p e c í f .
1.85 0.99 13.9 1.24 6.50 29.90 0.74 1.27 9.61 1.19 3.67 13.60 0.85 4.41 2.59 2.90 1.79 8.19 7.16 9.78 2.44 33.20 2.28 2.81 4.56 14.90 12.0
31.90 16.0 5.20 8.52 6.81 3.13 3.53 15.80 3.52 2.35 8.56 4.03 30.30 2.76 6.45 36.20 10.90 3.38
36.30 9.27
Síntomas
.
. u
-7 OAS
-OAS
A A
U U
U
U U A
-OAS
--. u A U A
OAS U
CU U
-OAS U U A
SPT
33.3
-10.1 37.8
-59.2
28.7
33.4 17.6 22.1
25.6 9.3
20.5 35
31.5 10.5 7.3
20.1 31
50.4 23.9 68.3 20.8 15.9 17.4 70.2 15.5 44.8 7.4
62.8 24.1 15.6 34.5 22.3 62.9 20.1 20.4 24.8 24.9 33.9 61.7 27.4
I g E E s p e c i f .
0.53 0.51 12.60 1.19 6.36 12.20 1.47 1.31 5.64 2.45 3.36 9.50
<0.35 5.09 2.26 1.05 2.49 7.35 6.35 10.0 0.40
43.50 2.04 2.52 3.65 15.0 7.31
27.10 14.30 2.06 11.10 6.86 1.51 3.41 7.45 0.81 0.66 7.01 4.27 14.40 1.67 4.14 24.30 8.74 2.01 25.60 5.57
Síntomas
.
. ?
--?
?
?
s u
OAS U u A
-OAS OAS
---.
OAS
-A
SPT
-. ---9.2
-10.7 16.1
. 18.1 9.5
59.5 14.3 15.5 18.3 24.8 20.3
-9.7 78
----
. 8.9
8.4 26.4
--10.8 16
I g E E s p e c i f .
<0.35 <0.35 5.26
<0.35 1.24 2.46 0.97 0.66
6.11 1.26
<0.35 0.38 1.47
<0.35 <0.35
0.58 1.92 1.30
<0.35
0.49 1.02 2.52 2.91 1.60 11.60 6.66 1.23 6.05 1.83 0.38 1.86 0.39
<0.35 <0.35 <0.35 1.13 3.51
<0.35 0.46 7.83 0.69 0.61 3.47 2.10
SPT
--
10.2 14.9
-8
7.9
7
20.9 37.5
8.7 11.2 31.5 48.9 42.9 44.9 35.7 36.9
10 45.4
. 25.6
. ---11.1 8.6
-27 45.4 29.6 48.7 106.4 13.3
I g E Especif.
<0.35 <0.35 4.47
<0.35 2.83 2.33 3.62 0.68 1.72 10.60 0.81 2.12
<0.35 4.05
<0.35 <0.35 1.81 0.62 0.68 6.89
<0.35 12.10 0.68
2.20 4.37 2.64 11.30 3.54 1.15
<0.35 1.73
<0.35 3.43 0.78
<0.35 <0.35 <0.35 4.49 1.09
<0.35 0.74 4.34 1.79 7.80 8.47 1.72
Sexo, F= Mujer; M=Hombre. Síntomas, OAS= Síndrome de Alergia Oral; U=Urt¡car¡a; A=Anaf¡laxia; CU=Urt¡car¡a de contacto; S=Shock anafiláctico; -, tolera la fruta; ?=desconoce su tolerancia a la fruta. SPT, prueba cutánea expresada en mm^ de pápula; valores inferiores a 7 mm^ se consideraron negativos (-). nd= no determinado.

Material y Métodos
11.2.- MÉTODOS DE PURIFICACIÓN Y CARACTERIZACIÓN DE
PROTEÍNAS.
11.2.1.- Extracción de proteínas.
11.2.1.1.- Extracto crudo de proteínas solubles (PBS).
La piel o los frutos completos fueron liofillzados y posteriormente
pulverizados con N2 líquido. El pulverizado se desiipidó con acetona (3x 1:5
p/v; 1 h a 4°C). En algunos casos, se realizó una segunda deslipidación con
una mezcla de etenetanol (3:1 v/v; 1:5 p/v; 1 h a 4°C).
Los residuos deslipidados fueron extraídos en tampón PBS (fosfato
sódico 100 mM, pH 7, NaCI 0,15 M; 1 h; 4°C). Tras centrifugar (12.000 rpm;
30 min; 4°C), el sobrenadante se dializó frente a agua destilada (4°C; 48h)
y se liofilizó.
El extracto de látex se obtuvo a partir de látex natural con bajo
contenido en amoniaco (Cauchos Guzmán S.A.; Valencia). La preparación
fue diluida en PBS (1:5 p/v), centrifugada (2 x 12.000 rpm; 30 min; 4°C) y el
sobrenadante fue dializado frente a agua destilada y liofilizado.
11.2.1.2.- Preparaciones enriquecidas en quitinasas.
Los extractos PBS fueron precipitados con (NH4)2S04 al 80% de
saturación. Tras centrifugarse (12.000 rpm; 30 min; 4°C), los precipitados
fueron resuspendidos en tampón PBS, y se acidificaron hasta un pH de 3,5
con HCI 0,1M. Después de centrifugar en las mismas condiciones, los
sobrenadantes fueron dializados frente a agua destilada y liofillzados.
26

Material y Métodos
11.2.1.3.- Preparaciones enriquecidas en LTPs.
Las proteínas del polen de artemisia y de piel de melocotón y
manzana deslipidados fueron extraídas con tampón Tris-HCI 0,1 M, EDTA 10
mM, pH7,5 (1:5 p/v; 1h; 4°C). Después de centrifugar (12.000 rpm; 30 min;
4°C), el sobrenadante fue dializado y liofilizado (Extracto Tris-HCI).
El precipitado de la centrifugación se resuspendió en agua destilada y
se centrifugó (15.000 rpm; 15 min; 4°C) y el nuevo precipitado se extrajo con
LiCI 1,5 M (1 h; 4°C). Se volvió a centrifugar (15.000 rpm; 20 min; 4°C) y el
sobrenadante fue dializado frente a agua destilada y liofilizado (Preparación
enriquecida en LTPs).
11.2.2.- Cuantifícación de proteínas.
Los extractos de proteínas se cuantificaron según el método de
Bradford(1976).
Las proteínas purificadas fueron cuantificadas por el método del ácido
bicinconinico (Smitli etai, 1985).
11.2.3.- Métodos cromatográfícos.
11.2.3.1.- Cromatografía de filtración molecular.
Para la purificación de la LTP de castaña, inicialmente se fraccionó el
extracto PBS (30 mg de proteína) por cromatografía de filtración molecular
utilizando Sephacryl S-200 HR en una columna de 90 x 2,5 cm, con un flujo
medio de 85 ml/h, usando como tampón de elución acetato amónico 0,1M,
pH 6,8.
Para la purificación de las proteínas recombinantes rCas s 5 y rCAT,
se utilizó una columna preempaquetada Superdex 75 HR 16/26 (Pharmacia
Diagnostics, Uppsala, Suecia), con un flujo de 60 ml/h, usando como tampón
de elución acetato amónico 0,2M, pH 6,8. El sobrenadante del cultivo de las
27

Material y Métodos
levaduras transformadas (2-4 mg de proteínas) fue resuspendido en el
mismo tampón y aplicado a la columna.
11.2.3.2.- Cromatografía de afinidad.
Como primer paso de purificación de las quitinasas nativas de
castaña, aguacate y plátano se realizó una cromatografía de afinidad con
quitina regenerada según el método de Molano eí al. (1970). Para ello, 2
gramos de Chitosan (Sigma) se trituraron con N2 líquido y se resuspendieron
en 40 mi de ácido acético 10% (v/v), dejándolos toda la noche a temperatura
ambiente. Posteriormente, la mezcla se diluyó con 180 mi de metanol y se
filtró al vacío con lana de vidrio. Después se añadieron 2 mi de anhídrido
acético en agitación y se dejó 30 minutos a temperatura ambiente, hasta
conseguir una textura de gel. Se homogeneizó en un omnimixer (Omni
International, CT, Estados Unidos) hasta conseguir el tamaño de partícula
deseada para la cromatografía. Tras ello, se montó una columna de 45x2,5
cm.
Las preparaciones enriquecidas en quitinasas (30mg de proteína)
fueron disueltas en tampón Tris-HCI 20mM, pH 8, y se cargó la columna de
quitina regenerada equilibrada con el mismo tampón. La elución fue llevada
a cabo con 300 mi (3 horas) de tampón acetato sódico 20mM, pH 5,5
seguido de 400 mi (3 horas) de ácido acético 20mM, pH 3,2.
11.2.3.3.- Cromatografía de intercambio catiónico.
Para purificar las quitinasas, las fracciones retenidas en la columna de
afinidad de quitina fueron sometidas a una cromatografía de intercambio
catiónico en una columna Mono S HR 5/5 preempaquetada de Pharmacia
(1ml de volumen), acoplada a un sistema de FPLC (Pharmacia Diagnostic,
Uppsala, Suecia). La columna fue equilibrada con tampón acetato sódico
10mM, pH 5,3 (tampón A) y eluido con el mismo tampón con NaCI 0,5M
(tampón B). El gradiente que se utilizó fue 0% a 50% de tampón B en 40
28

Material y Métodos
minutos, y de 50% a 100% de tampón B en 20 minutos, con un flujo de
1ml/min. Las fracciones que contenían las distintas clases de quitinasas
fueron desalinizadas en centricones de 10 kDa frente a agua y concentradas
(Amicon Separations, Bedford, MA, Estados Unidos).
Para purificar las LTPs de manzana y melocotón, se utilizaron
cartuchos de intercambio catiónico Sep-Pak Accell Plus CM de 500mg
(Waters, Mildford, MA, Estados Unidos). Se aplicaron preparaciones
enriquecidas en LTPs (5mg de proteína), resuspendidas en tampón ácido
fórmico-NaOH 20mM, pH 4. La muestra se eluyó con el mismo tampón con
NaCI 0,75M, con un flujo de 1 ml/min.
11.2.3.4.- Cromatografía líquida de alta presión (HPLC).
El extracto Tris-HCI de polen de artemisa fue fraccionado por RP-
HPLC en una columna preparativa Vydac-C4 (22x250 mm; tamaño de
partícula 10|am; The Separations Group, Hesperia, CA, Estados Unidos). La
elución fue llevada a cabo en un gradiente lineal de 2-propanol en 0,1% de
ácido trifluoroacético (0% a 30% en 120 min; 30% a 50% en 30 min; y 80 min
a 50%; flujo 1 ml/min). Las fracciones enriquecidas en la LTP de artemisa
fueron repurificadas en un segundo sistema de RP-HPLC en una columna
de Nucleosil 300-C4 (8 x 250 mm; tamaño de partícula 5 |am; Sugelabor,
Madrid), eluyendo con un gradiente lineal de acetonitrilo en 0,1% de ácido
trifluoroacético (0% a 80% en 150 min; 0,5 ml/min).
Las fracciones de castaña enriquecidas en LTP, procedentes de la
columna de Sephacril S-200, fueron separadas por RP-HPLC en una
columna preparativa Vydac-C4 (22x250 mm; tamaño de poro 10 |j,m), en un
gradiente lineal de acetonitrilo en 0,1% de ácido trifluoroacético (10%
durante 45 min; 10% a 35% en 140 min; 35% a 50% en 100 min y 50% a
85% en 15 min; 1 ml/min).
29

Material y Métodos
11.2.4.- Métodos electroforéticos.
Para la electroforesis de proteínas se utilizaron geles de poliacrilamida
en presencia de dodecil sulfato sódico (SDS-PAGE) en condiciones no
reductoras. Se siguió básicamente el método de Laemmii (1970). Para el gel
separador se empleó una solución de poliacrilamida al 15% (p/v) en Tris-HCI
0,125M, pH 8,8, 0,1% de SDS (p/v), y para el gel espaciador una solución de
poliacrilamida al 4% (p/v) en Tris-HCI 0,125M, pH 6,8, 0,1% de SDS (p/v).
Las muestras se disolvieron en Tris-HCI 0,0625M, pH 6,8, 2% de SDS (p/v),
urea 8M, 0,001% de azul de bromofenol (p/v).
Para la detección de las LTPs, se emplearon geles de gradiente de
poliacrilamida (4-20%) prefabricados de Bio-Rad (Tris-Glicina; BioRad
Laboratories, Hercules, CA, Estados Unidos), o bien geles de gradiente de
poliacrilamida (10-20%) prefabricados de Novex (Tris-Tricina; Novex, San
Diego, CA, Estados Unidos). Las muestras se prepararon de la misma forma
que se ha descrito anteriormente.
Las electroforesis se llevaron a cabo en el sistema de minigeles de
Bio-Rad (Bio-Rad Miniprotean II system) o de Novex (X Cell™ Mini-Cell)
durante 1 hora a 20 mA/gel, utilizando como tampón de electroforesis Tris
0,025M, glicina 0,192M, pH 8,3, 0,1% de SDS (p/v).
La tinción de proteínas se realizó con una mezcla de Coomassie
Brillant Blue R-250 al 2% y Bismark Brown R al 0,05% (7,5:10 v/v), en
metanol 40% (v/v), ácido acético 7% (v/v), según el método de Choi et al.
(1996).
M.2.5.- Ensayo de actividad quitínasa.
La actividad quitinasa fue determinada por un microensayo
colorimétrico con carboximetil-quitin-Remazol Brillant Violet 5R como
sustrato (Wirth y Wolf,1990). La muestra se disolvió en 300 |iil de tampón
acetato sódico 0,2M, pH 5,5, y se añadió 100 |J.I del sustrato (2mg/ml) . Se
30

Material y Métodos
incubó 30 min a 37°C y posteriormente se paró la reacción con 100 pl de HCI
2N, manteniéndose 10 min en hielo. Tras centrifugarse (10 min a 10.000
rpm) se determinó la absorbancia a 550 nm.
11.2.6.- Determinación de la composición de aminoácidos.
El análisis de aminoácidos fue realizado tras oxidación perfórmica e
hidrólisis acida de las proteínas. Las muestras se disolvieron en 20}il de
ácido fórmico al 97% (v/v) y 30^1 de ácido perfórmico, obtenido a partir de
9,7 mi de ácido fórmico al 97% y 0,5 mi de H2O2 al 30% (v/v). La oxidación
transcurrió en un baño con hielo durante 3 horas. Finalmente, se diluyeron
con agua destilada y se liofilizaron, (Hirs, 1967). Las muestras oxidadas se
hidrolizaron durante 24 horas a 110°C con HCI 5,7M, fenol 5mM. Los
hidrolizados fueron fraccionados en un analizador Pharmacia Biochrom 20
(J. Várela, CIB, CSIC, Madrid).
11.2.7.- Determinación de secuencias aminoterminales.
Las secuencias N-terminales se determinaron siguiendo el método
estándar con un secuenciador de fase gaseosa Applied Biosystems 477 A
(J.Varela, CIB, CSIC, Madrid).
11.2.8.- Determinación de pesos moleculares por espectrometría
de masas.
Los pesos moleculares de proteínas se determinaron por
espectrometría de masas (MALDI-TOF) usando un espectrómetro Kompact
MALDI-2 con extracción retardada (Kratos Analytical, Manchester, Gran
Bretaña).
31

Material y Métodos
11.3.- MÉTODOS INMUNOQUÍMICOS.
11.3.1.- Purificación de anticuerpos poíiclonaíes.
Los anticuerpos poíiclonaíes de conejo frente a qultinasa de clase II
de castaña (CslM) fueron cedidos por los Drs. C. Aragoncillo y C. Collada
(Dept. Biotecnología, ETS Ingenieros de Montes, UPM).
La purificación de los anticuerpos poíiclonaíes monoespecíficos se
realizó según el método descrito por Olmsted (1986). Se insertó 100 |ig de la
proteína purificada en un gel de poliacrilamida de un solo pocilio continuo.
Una vez transferida la proteína a membrana de nitrocelulosa, ésta se tiñó
con una solución de rojo Ponceau 0,2% (p/v) disuelto en 3% de ácido acético
(v/v) (Boheringer-Manheim, Alemania), se cortó la banda que contenía la
proteína y se destiñó con agua destilada. La banda fue bloqueada con TBS
(Tris-HCI 20mM, NaCI 150mM, pH 8,3), 2% (p/v) de leche desnatada en
polvo, durante 2 horas a temperatura ambiente. Posteriormente, se incubó
con el suero de anticuerpos poíiclonaíes en TBS, 0,2% (p/v) de leche
desnatada en polvo (dilución 1:15). Tras lavarse con el mismo tampón 5
minutos, se eluyó el anticuerpo con tampón Glicina-HCI 0,1 M, pH 2,5,
durante 1,5 horas a temperatura ambiente. El eluido se neutralizó con Tris
0,5M, pH 11, se dializó frente a agua destilada 24 horas y se liofilizó.
Posteriormente, se redisolvió en TBS y se tituló.
11.3.2.- ínmunodetección de proteínas transferidas a membrana.
11.3.2.1- Electrotransferencia a membranas de nitrocelulosa o
de difluoruro de polivinilideno (PVDF).
Las proteínas separadas por electroforesis se electrotransfirieron a
membranas de nitrocelulosa (tamaño de poro 0,45 pim, Millipore) o de PVDF
(Immobilon P, tamaño de poro 0,45 ¡im, Millipore), según el método de
Gültekin y Heerman (1988).
32

Material y Métodos
Las membranas de PVDF se activaron previamente en metanol
durante 15 segundos, y posteriormente fueron equilibradas en tampón de
transferencia (Tris-Bórico 0,05IV1, pH 8,3). La electrotransferencia fue llevada
a cabo en un sistema de Bio-Rad (Bio-Rad Mini Trans-Blot Cell), con el
mismo tampón, durante 1 hora e intensidad constante de 80 mA.
Las membranas de nitroceiulosa fueron electrotransferidas utilizando
tampón Towbin (Tris 0,025M, Glicina 0,2I\/I, metanol 20% (v/v); Towbin et al.,
1974). La transferencia fue llevada a cabo en el sistema de de Bio-Rad
durante 1 hora a 80 mA utilizando el mismo tampón.
11.3.2.2.- Inmundetección frente a anticuerpos policlonales
purificados.
Las membranas, con las proteínas transferidas, fueron bloqueadas
con tampón TBS con 2% (p/v) de leche desnatada en polvo durante toda la
noche a 4°C. Tras ello, se incubaron con el anticuerpo correspondiente
diluido en tampón TBS con 1% (v/v) de suero de cabra y 0,2% (p/v) de leche
desnatada en polvo durante 2 horas a temperatura ambiente. Los
anticuerpos policlonales monoespecíficos frente a CslH fueron utilizados a
una dilución 1:250.
Tras la incubación con los anticuerpos, las membranas fueron lavadas
con el tampón TBS con 0,2% (p/v) leche desnatada en polvo (3x5 min).
Posteriormente, se incubaron con el anticuerpo secundario antl-lgG de
conejo conjugado a fosfatasa alcalina (dilución 1:500; Sigma) en TBS 0,2%
leche desnatada en polvo con gelatina 0,4% (p/v), durante 2 horas a
temperatura ambiente. Después, se volvieron a lavar las membranas con
TBS 0,2% leche desnatada en polvo (3x5 min), y se revelaron con la
solución del sustrato de la fosfatasa alcalina (Sigma Fast^'^ fosfato 5-bromo-
4-cloro-3-indolil / azul de nitrotetrazolium; BCIP/NBT).
33

Material y Métodos
II.3.2.3.- Detección de glicoproteínas.
II.3.2.3.1.-Detección de glicoproteínas por inmunoensayo con
digoxigenina.
Las glicoproteínas fueron identificadas por el sistema de detección de
azúcares de Roche (Roche Molecular Biochemicals, Mannhein, Alemania),
siguiendo el protocolo indicado por la casa comercial. Las proteínas fueron
oxidadas y conjugadas con digoxigenina antes de ser separadas por SDS-
PAGE y eletrotransferidas a membranas de nitrocelulosa. La digoxigenina
incorporada fue detectada en un inmunoensayo enzimático utilizando un
anticuerpo conjugado a fosfatasa alcalina.
ll.3.2.3.2.-lnmunodetección con anticuerpos específicos frente
a p (1-2) xilosa y a (1-3) fucosa.
Para detectar oligosacáridos complejos de plantas unidos a
asparagina, se utilizó un suero específico frente a P(1-2) xilosa y a(1-3)
fucosa, cedido por el Dr. M.J. Chrispeeis (Dept. Biología, Universidad de
California, San Diego, USA; Lauriere et al., 1989). Las membranas con las
proteínas transferidas fueron tratadas según se describe en el punto 11.3.2.2
de este apartado, utilizando como anticuerpo primario el específico frente a
oligosacáridos complejos (dilución 1: 6000).
11.3.2.4.- Inmunodetección frente a sueros de pacientes
alérgicos.
11.3.2.4.1.- Inmunodetección con fosfatasa alcalina.
Las membranas con las proteínas transferidas fueron bloqueadas con
tampón PBS (Fosfato sódico 10mM, NaCI 15mM, pH 7, 0,01% de
Thimerosal) con 2% de leche desnatada en polvo durante 2 horas a
34

Material y Métodos
temperatura ambiente. A continuación, fueron incubadas con una mezcla de
sueros de pacientes alérgicos diluida (1:3) con tampón PBS, 0,2% de leche
desnatada en polvo, durante toda la noche a temperatura ambiente.
Después de lavar con PBS (3x5 min) y equilibrar con TBS, 0,2% leche
desnatada (3x10 min), las membranas fueron incubadas durante 2 horas a
temperatura ambiente, con un anticuerpo monoclonal anti-lgE humana ligado
a fosfatasa alcalina (clon GE-1, Sigma; dilución 1:500) diluido en tampón
TBS, 0,2% leche desnatada con 0,4% de gelatina. Se lavaron de nuevo con
TBS, 0,2% leche desnatada y se revelaron con el sustrato de la fosfatasa
alcalina (BCIP/NBT; Sigma).
11.3.2.4.2.- Inmunodetección por quimioluminiscencia.
Las membranas con las proteínas transferidas fueron bloqueadas con
tampón PBS y albúmina de suero bovino (BSA) al 5%, durante 2 horas a
temperatura ambiente. Tras el bloqueo, se incubaron con la mezcla de los
sueros de los pacientes diluida (1:10) con tampón PBS, 0,5% BSA, 0,05%
Tween-20, durante toda la noche. Después de lavar con PBS, 0,1% Tween-
20 (4x5 min), se incubaron 2 horas a temperatura ambiente con líquido
ascítico de ratón (monoclonal HE-2 anti-lgE humana; dilución 1:3000).
Posteriormente, se lavaron y se incubaron con anticuerpos de conejo anti-
IgG de ratón ligados a peroxidasa (DAKO A/S, Dinamarca) durante 1 hora a
temperatura ambiente. La detección de los componentes que ligaban IgE fue
llevada a cabo por quimioluminiscencia, siguiendo las instrucciones del
fabricante (ECL-Amersham).
11.3.2.4.3.- Ensayos de inmunoinhibición en membrana.
Los ensayos se realizaron incubando la mezcla de sueros de
pacientes sin diluir con el inhibidor correspondiente durante 3 horas a
temperatura ambiente, y posteriormente, llevando a cabo la inmunodetección
según se ha descrito.
35

Material y Métodos
La heveína purificada de látex, empleada en algunos de estos
ensayados, proviene del laboratorio del Dr. Z. Chen (Instituto de
Investigación de Medicina Ocupacional, BGFA, Bochum, Alemania).
11,3.3.- Enzimoinmunoensayo (ELISA).
11.3.3.1- Determinación de la IgE específica.
Sobre placas de poliestireno de 96 pocilios (Costar 3590, Corning,
NY, Estados Unidos), se aplicaron 50{a.l de una solución de 20fj,g/ml de LTP
purificada por pocilio, y se fijó durante 1 hora a 37°C. Después de bloquear
con tampón PBS con 1 % BSA, los pocilios fueron incubados con 50 ,1 de la
mezcla de sueros de pacientes (dilución 1:6) durante 3 horas a temperatura
ambiente. Tras lavarse con tampón PBS, 0,1% Tween 20, se incubó con
50|j,l de anti-lgE humana ligada a peroxidasa disuelta en tampón PBS, 0,1%
Tween-20 (dilución 1:500; DAKO A/S, Dinamarca), durante 1 hora a
temperatura ambiente. Se volvió a lavar con el mismo tampón y se añadió el
sustrato de la peroxidasa (0,012% de H2O2, 0,66mg/ml o-fenilenediamina,
OPD; DAKO). La reacción se paró después de 30 minutos con 50|j,l de
H2SO4 4N. Se midió la absorbancia a 492nm. Cada ensayo se realizó por
duplicado y como control negativo se utilizaron pocilios sin fijar LTP. Los
resultados se expresaron en unidades de RAST (RU/ml).
11.3.3.2.- Ensayos de inhibición.
Los ensayos de inhibición por ELISA se realizaron incubando la
mezcla de sueros sin diluir con el inhibidor correspondiente durante 3 horas
a temperatura ambiente. Posteriormente, se siguió con lo descrito en el
apartado 11.3.3.1.
El porcentaje de inhibición de fijación de IgE fue determinado como:
Inhibición (%)= 100x(1-[(OD-ODioo%)/(ODoo/o-ODioo%])
36

Material y Métodos
Donde OD es la absorbancia de cada pocilio tras ser Incubado con la
mezcla de suero más el Inhibidor; ODo% es la absorbancia obtenida al
Incubar con la mezcla de sueros sin inhibidor; y ODioo% es la absorbancia en
ausencia de la mezcla de sueros.
11.3.4.- Pruebas de inhibición por radioinmunoensayo (RAST).
Los ensayos de inhibición por RAST se realizaron utilizando el
sistema de CAP-Inmunogenic de Pharmacia. La mezcla de sueros se incubó
con el inhibidor correspondiente a una determinada concentración, a partir
de la cual se realizaron 5 diluciones seriadas con tampón PBS 0,05% BSA.
Tras 1 hora de incubación a temperatura ambiente en una microplaca, se
añadió el CAP comercial. La cuantificación de los niveles de IgE específica
fue llevada a cabo siguiendo el manual de la casa comercial (CAP System,
Pharmacia & Upjohn Diagnostics).
11.3.5.- Pruebas cutáneas (SPT, Skin Príck Test).
Las pruebas cutáneas con extractos y proteínas purificadas fueron
llevadas a cabo siguiendo un método convencional (SPT) usando una
lanceta DOME (Hollister-Stier) Bayer de Morrow-Brown (1 mm de incisión;
Bayer S.A, Madrid). Todas las muestras fueron redisueltas en glicerina al
50% (v/v), NaCI 0,9% (p/v), realizándose dos punciones en el antebrazo de
los pacientes. Las pápulas desarrolladas tras 15 minutos desde la aplicación
de la muestra fueron transferidas desde la piel hasta un papel adhesivo
mediante una técnica de "trazado", y su área fue medida por planimetría
(HAFF planimeter 317). No se consideraron como positivas las áreas
<7mm^. Fosfato de histamina (10mg/ml) y tampón PBS fueron usados como
controles positivos y negativos, respectivamente.
37

Material y Métodos
HA- MÉTODOS DE ÁCIDOS NUCLEICOS.
11.4.1.- Extracción del ARN total de piel de manzana y melocotón.
La extracción del ARN total se realizó según el método de Lagrimlni et
al. (1987). Se pulverizó tejido en N2 líquido, y se añadió 1 volumen de
tampón de extracción (ácido p-aminosalicílico al 4% y ácido 1,5-
naftalendisulfónico al 1%; 400|Lil/500pg de tejido), 1 volumen de fenol
equilibrado con Tris-HCI 50mM, pH 8, y 1 volumen de cloroformo: alcohol
isoamílico (24:1, v/v). Se centrifugó a 8.000 rpm durante 10 min a 4°C, y se
recogió la fase acuosa. Dicha fase fue precipitada con acetato sódico 3M, pH
5,3 (10% del volumen total) y etanol absoluto (2 volúmenes), manteniéndose
en hielo 30 min. Tras centrifugarse (8000 rpm, 10 min a 4°C), se lavó el
precipitado con etanol al 80% (v/v). El precipitado se dejó secar a
temperatura ambiente, y posteriormente se resuspendió en agua destilada.
Tras ello, se añadió 2/3 del volumen de LiCI 8M y se incubó toda la noche a
4°C. Se centrifugó y el precipitado fue lavado con etanol 80% (v/v). Por
último el precipitado fue resuspendido en agua destilada.
La calidad del ARN extraído fue comprobada en geles
desnaturalizantes de agarosa al 1,2% (p/v) en presencia de formaldehído al
6% (v/v). El tampón de electroforesis contenía MOPS 20 mM, acetato sódico
5mM, pH 7, EDTA 1 mM. Las muestras se aplicaron disueltas en tampón de
electroforesis con formamida 45% (v/v), formaldehído al 16% (p/v), glicerol
5% (v/v), azul de bromofenol 0,02% (p/v) y bromuro de etidio (0,1mg/ml).
11.4.2.- Obtención de los clones ADNc correspondientes a las
LTPs de melocotón y manzana.
El ARN total de piel de manzana y melocotón sirvió como molde para
obtener la primera cadena del ADN complementario utilizando el sistema
"First Strand cDNA" de Pharmacia, siguiendo el protocolo que recomienda la
38

Material y Métodos
casa comercial y utilizando como oligonucleótido cebador la secuencia:
TCGACTCTAGAGGATCCGAATTCAAGC(T)i5.
Una vez obtenido el ADN complementario, se realizó una primera
Reacción en Cadena de la Polimerasa (PCR) con 4^] de la primera hebra
del ADN complementario, siguiendo el programa: 7 min de desnaturalización
a 96°C; 30 ciclos de 1 min de desnaturalización a 96°C, 1 min de
anillamiento a 55°C, 1 min de polimerización a 72°C; y 12 min de
polimerización terminal a 72°C. Posteriormente, se realizó una
reamplificación con 2JJ,I del producto obtenido, siguiendo el mismo programa,
pero con una temperatura de anillamiento de 65°C. Para las amplificaciones,
se utilizó la polimerasa Taq-Gold de Perkin-Elmer, y la mezcla de reacción
se preparó siguiendo las recomendaciones de la casa comercial. Las
secuencias de los oligonucleótidos que se emplearon para ambas PCR
fueron: para el extremo 3', CTGCAGGTCGACTCTAGAGGATCC; para el
extremo 5' de la LTP de melocotón, TAACATGTGGCCAAGTGTCCAGCGC
ACTTGCACCATGC; y para el extremo 5' de la LTP de manzana, TAACAT
GTGGCCAAGTGACCAGCAGCCTTGCACCATGC. Los oligonucleótidos se
dedujeron a partir de la secuencia aminoterminal de las proteínas purificadas
y de la secuencia de nucleotidos de la LTP de almendra disponible en el
banco de datos. La LTP de almendra tiene un 95% de identidad de
secuencia de aminoácidos con la de melocotón.
Los productos de PCR fueron separados por electroforesis en geles
de agarosa (1,2% p/v) redisuelta en tampón TAE (Tris-acetato 40mM, EDTA
2mM, pH 8) con bromuro de etidio (0,1mg/ml). Para aplicar las muestras en
los geles, se les añadió 2|il de azul de bromofenol 0,02% (p/v). Las
electroforesis se desarrollaron a 90 mV en tampón TAE.
Cuando se obtuvieron bandas del tamaño esperado, éstas fueron
purificadas de agarosa por el sistema de QUIAEX II (Quiagen, Alemania) y
posteriormente fueron clonadas en el vector pGEM T Easy de Promega
(Madison, Estados Unidos), siguiendo el protocolo de la casa comercial. El
vector con el inserto fue introducido en Escherichia coli DH5a, y la selección
de clones positivos se realizó por resistencia a ampicilina (0,1mg/ml) en
39

Material y Métodos
medio LB en placa ( 0,5M NaCI, 1% triptona, 0,5% extracto de levadura, 15g
de agar). Estos clones fueron crecidos toda la noche a 37°C en medio LB
líquido (igual que el de placa, pero sin agar), y el ADN plasmídico fue aislado
por el sistema Wizard Plus Minipreps (Promega).
El ADN fue secuenciado usando el ABI PRISM Dye Terminator Kit,
con un secuenciador automático ABI 377 (Applied Biosystem, Perkin-Elmer),
utilizando la ADN polimerasa Ampli Taq FS. La mezcla de reacción se
preparó siguiendo el sistema Big Dye™ Terminator RR Mix (Perkin-Elmer
Biosystem).
11.4.3.- Subclonaje en el vector de expresión pPIC 9 de la levadura
Pichia pastoris.
Pichia pastoris es una levadura metalotrófica que es capaz de utilizar
el metanol como fuente de carbono, cuando no existe otra. En su genoma,
están presentes dos genes que codifican para sendas proteínas con
actividad alcohol oxidasa, A0X1 y A0X2. En presencia de metanol, como
única fuente de carbono, la expresión de A0X1 supone un 80 % del total.
Aprovechando esta característica, el sistema de expresión de P. pastoris
consiste básicamente en sustituir por recombinación el gen A0X1 por el que
interesa expresar. Asimismo, dicha levadura expresa muy pocas proteínas
extracelulares. Por otra parte, la cepa que se ha empleado de levadura,
GS115, es deficitaria en crecimiento en medio sin histidina.
Para la expresión en levadura, se ha utilizado el vector de expresión
pPIC 9 (Fig. 3), que incluye el péptido señal de secreción extracelular del
factor a de Sacctiaromyces cerevisiae previo al sitio de clonaje múltiple en
fase, donde se introducirá el inserto que interesa expresar. Precediendo al
péptido señal y posterior al sitio de multiclonaje, se incluyen los extremos 5'
y 3', respectivamente, del gen A0X1, para dirigir la recombinación. Además,
en este vector se ha incluido un origen de replicación en bacterias, el gen de
la resistencia a ampicilina (para la selección de transformantes en bacteria) y
el gen I-IIS4, de la histidol deshidrogenasa que le permite crecer a la
40

Material y Métodos
levadura en medio sin histidina (para la selección de transformantes en
levadura).
3'AOX1(TT)
& l t Fragmento del promotor 5' AOXI: 1-948 Péptido señal de secreción (S): 949-1218 Sitio de clonaje múltiple: 1192-1241 Fragmento de Z'AOXI, terminación de la transcripción (TT): 1253-1586 Gen H/S4; 4514-1980 Origen de replicación en bacteria (ColEI): 6708-6034 Gen de resistencia a ampicilina: 7713-6853
Figura 3: Esquema del vector de expresión pPIC 9 de Pichia pastoris.
II.4.3.1.- Obtención del ADN complementario de los dominios
heveína y catalítico, y de la proteína madura Cas s 5 (quitinasa
clase I de castaña).
En la Figura 4, se representa la secuencia de nucleótidos y
aminoácidos de Cas s 5 (sin péptido señal), la quitinasa de clase I alergénica
de semilla de castaña. El clon de ADNc (Aliona et al., 1996) fue cedido por
los Drs. C. Aragoncillo y L. Gómez (Dept. Biotecnología, ETS Ingenieros de
Montes, UPM). Se han señalado los fragmentos de ADN que sirvieron para
subclonar el dominio heveína, el dominio catalítico, y la proteína completa.
El subclonaje de estos fragmentos se realizó por PCR mediante
oligonucleótidos específicos utilizando la TaqGold polimerasa de Perkin-
Elmer. Para la amplificación se utilizó un programa convencional: 4 min de
desnaturalización a 94°C; 25 ciclos de 1 min a 94°C, 1 min a 55°C y 1 min a
72°C; y 12 min de finalización a 72°C. Los cebadores fueron diseñados a
partir de la secuencia del ADN complementario y añadiendo los sitios de
restricción Xho I/Eco Rl, que iban a servir para clonar los fragmentos en el
vector de expresión pPIC 9. Por otra parte, al oligonucleótido diseñado para
amplificar el extremo 3' del dominio heveína se incorporó un codon de
parada de la traducción, así como, en el cebador del extremo 5' del
41

Material y Métodos
'GAA CAÁ TGT GGC AGA CAÁ GCT GGG GGT GCT GCG TGT GCG AAC 42 E Q C G R Q A G G A A C A N AAC TTA TGT TGT AGC CAG TTT GGA TGG TGT GGC AAC ACÁ GCT 84 N L C C S Q F G W C G N T A GAG TAC TGT GGA GCT GGT TGC CAG AGC CAG TGT TCT TCT CCT 12 6 E Y C G A G C Q S Q C S S P
HD CD
ACA^ACT ACT ACT TCA TCT CCT AC#GCT AGT AGT GGT GGT GGT 168 T / \ T T T S S P T / \ A S S G G G ( t a a ) (E) GGC GAT GTT GGC AGC CTT ATC AGT GCG TCT CTT TTT GAT CAÁ 210 G D V G S L I S A S L F D Q ATG CTT AAA TAT AGG AAC GAT CCA AGA TGT AAA AGT AAT GGA 252 M L K Y R N D P R C K S N G TTC TAC ACT TAC AAT GCT TTC ATT GCT GCT GCT CGA TCT TTC 2 94 F Y T Y N A F I A A A R S F AAT GGC TTT GGC ACÁ ACT GGT GAT GTT ACT ACÁ CGT AAA AGA 336 N G F G T T G D V T T R K R GAG CTT GCG GCT TTC TTA GCT CAÁ ACC TCT CAT GAG ACC ACÁ 378 E L A A F L A Q T S H E T T GGA GGG TGG GCA ACT GCA CCA GAT GGC CCA TAT GCA TGG GGA 420 G G W A T A P D G P Y A W G TAT TGT TTT GTT ATG GAA AAT AAC AAG CAÁ ACC TAT TGT ACC 4 62 Y C F V M E N N K Q T Y C T TCA AAA TCT TGG CCA TGT GTT TTT GGA AAA CAÁ TAT TAT GGT 504 S K S W P C V F G K Q Y Y G CGG GGA CCT ATC CAÁ CTC ACT CAC AAC TAC AAC TAT GGG CAÁ 54 6 R G P I Q L T H N Y N Y G Q GCA GGT AAA GCC ATT GGA GCT GAT CTC ATA AAC AAT CCA GAT 588 A G K A I G A D L I N N P D CTT GTA GCC ACÁ AAC CCC ACC ATA TCG TTT AAG ACÁ GCC ATA 630 L V A T N P T I S F K T A I TGG TTT TGG ATG ACÁ CCG CAÁ GCA AAC AAG CCA TCT AGC CAC 672 W F W M T P Q A N K P S S H GAT GTG ATC ATT GGA AAT TGG AGA CCC TCT GCT GCT GAC ACÁ 714 D V I I G N W R P S A A D T TCA GCT GGT CGA GTT CCA AGC TAT GGT GTA ATC ACC AAC ATT 75 6 S A G R V P S Y G V I T N I ATC AAT GGT GGC CTT GAA TGT GGC CAT GGC TCT GAT GAT AGG 7 98 I N G G L E C G H G S D D R GTG GCT AAT AGG ATT GGG TTT TAT AAG AGG TAC TGT GAC ACÁ 84 0 V A N R I G F Y K R Y C D T TTG GGA GTA AGC TAT GGG AAC AAC TTA GAT TGC TAT AAT CAÁ 882 L G V S Y G N N L D C Y N Q
CD
AAG CCC TTT G C C • t a a 8 94 K P F A
Figura 4: ADNc de la proteína madura de Cas s 5. Debajo de la secuencia de nucleótidos se tía añadido la secuencia de aminoácidos. El inicio y el final de las secuencia empleadas para subclonar el dominio heveína (HD) y el dominio catalítico (CD) se han indicado por flechas. Del mismo modo se ha señalizado el lugar donde se añadió el codon de parada (taa) en la secuencia del dominio heveína, y del glutámico (E) en la secuencia del domino catalítico. Por otra parte, se han subrayado los aminoácidos que corresponden a una diana de N-glicosílación
42

Material y Métodos
domino catalítico, se añadió un codon de ácido glutámico (E) con el fin de
facilitar el corte del péptido de localización extracelular. Las secuencias de
las parejas de los cebadores diseñados fueron:
<CCGCTCGAGAAAAGAGAACAATGTGGCA>/<CCTCGAATTCATGTAGGA
GAAGAACACTGGC> para los extremos 573' del dominio heveína;
<CCGCTCGAGAAAAGAGAAGCTAGTAGTGGTGG>/<TGGCCTTAGCTATT
TAGGCAAAGGGC> para los extremos 573' del dominio catalítico;
<CCGCTCGAGAAAAGAGAACAATGTGGCA>/<TGGCCTTAGCTATTTAGG
CAAAGGGC> para los extremos 5'/3' de la proteína completa.
11.4.3.2.- Subclonaje en el vector de expresión pPIC 9.
Una vez obtenidos los productos de PCR de las 3 construcciones,
éstos fueron aislados de geles de agarosa, clonados en el vector pGEM
TEasy e introducidos en E.coli. La selección de transformantes se realizó por
crecimiento selectivo en medio LB con ampicilina (ver apartado 11.4.2 del
Material y Métodos). Se aisló, a su vez, el ADN plasmídico de los clones
positivos que se digirió con las enzimas de restricción Xho l/EcoR I. Los
fragmentos digeridos se clonaron en pPIC 9 digerido con las mismas
enzimas, y se introdujeron en E.coli. La selección de los clones fue, de
nuevo, por resistencia a ampicilina. De los clones positivos, el ADN
plasmídico (pPIC 9 con el inserto) fue linearizado con Bgl II (en el caso del
inserto con el domino heveína) o con Sac I (en el caso del inserto con el
dominio catalítico y la quitinasa de clase I de castaña). El vector linearizado
fue empleado para la transformación de la levadura.
11,4.4.- Transformación de la cepa de P. pastoris GS115.
Una colonia de P. pastoris GS115 fue crecida en medio líquido YPD
(1% extracto de levadura, 2% peptona bacteriológica, 2% glucosa) a 30°C
(300 rpm en un incubador con agitación), hasta llegar a una densidad de
cultivo de 5-10 ODeoo (ODeoo- 5x10'' células/ml). Tras ello, el cultivo fue
43

Material y Métodos
alicuoteado añadiendo glicerol al 15%, y congelado con N2 líquido para
posteriormente almacenarlo a -80°C.
Una alícuota fue crecida en 500 mi de medio YPD durante toda la
noche hasta una ODeoo de 1,3-1,5. Posteriormente, fue centrifugada a 2500
rpm 10 minutos a 4°C. El precipitado fue resuspendido en 500 mi de agua
destilada, y se volvió a centrifugar en las condiciones antes mencionadas. El
nuevo precipitado se resuspendió en 250 mi de agua destilada, y se repitió la
centrifugación. El tercer precipitado se resuspendió en 20 mi de sorbitol 1M,
y se volvió a centrifugar. El precipitado final de células se resuspendió en 1-2
mi de sorbitol 1M, obteniéndose las células electrocompetentes.
Dichas células electrocompetentes fueron incubadas en hielo 15
minutos, con 10 |ag de ADN del vector pPIC 9 linearizado con Bgl II o Sac I.
Transcurrido ese tiempo, las células con el ADN fueron introducidas en la
cubeta de electroporación P/N 620 (2 mm de ancho; BTX Inc., San Diego,
Estados Unidos). Para la electroporación, se empleó un Electro Cell
Manipulator 600 BTX con unas condiciones de 2,5 kV de resistancia de alto
voltaje, sin capacitancia, 129 ohm de resistencia, una carga de 1,5 kV y un
campo de fuerza estimado de 7,5 kV/cm en 5 mseg. Tras el pulso, se añadió
1 mi de sorbitol 1M, y las células se recuperaron plaqueando 200-600|_il de la
mezcla final en placas con medio MD (1,34% YNB, 4x 10'^% biotina, 2%
glucosa y 15g agar). Se dejaron crecer a 30°C durante 3-4 días. La selección
de transformantes se realizó por crecimiento diferencial en ausencia de
histidina.
Una vez que crecieron los transformantes, estos fueron seleccionados
en placa con medio con glucosa (MD) y placas con medio con metanol (MM;
MD con 0,5% de metanol y sin glucosa). Para ello, se realizaron réplicas de
los transformantes en ambos medios y se dejaron crecer 4 días a 30°C.
Aquellos transformantes que eran capaces de crecer en MD, pero tenían
crecimiento retardado en MM, eran candidatos a que se hubiera sustituido el
gen A0X1 por el subclon de Cas s 5 correspondiente.
44

Material y Métodos
11.4.5.- Expresión de proteínas en P. pastorís.
Los transformantes con crecimiento retardado en placa con medio MM
fueron crecidos en medio líquido BMGY (1% extracto de levadura, 2%
peptona bacteriológica, tampón fosfato potásico lOOmM, pH 6, 1,34% de
YNB, 4 X 10"^% biotina y 1% glicerol) o BMG (tampón fosfato potásico
lOOmM, pH 6, 1,34% YNB, 4 x 10 "^% biotina y 1% glicerol), durante 4 días a
30°C (300 rpm hasta una ODeoo de 2-5). Posteriormente, se indujo la
expresión con metano!, centrifugando las células y cambiando el medio de
cultivo a BMMY (igual que el medio BMGY, pero sin glicerol y con 0,5% de
metanol), o a BMM (igual que el medio BMG, pero con 0,5% de metanol y
sin glicerol) a 30°C y 300 rpm. En el nuevo medio, no hay crecimiento de los
transformantes, pero sí inducción de la expresión. El máximo de ésta se
consiguió en todos los casos a los 4 días de inducir con metanol. Una vez
llegado al máximo de expresión, los cultivos fueron centrifugados, y el
sobrenadante fue dializado frente a acetato amónico 0,2M (48 horas a 4°C) y
liofilizado.
Para expresar el dominio heveína, los transformantes
correspondientes fueron crecidos en medio BMGY e inducidos en BMMY.
Sin embargo, para la expresión del dominio catalítico y de la quitinasa de
clase I de castaña, fue empleado el medio BMG e inducido en medio BMM.
La cuantificación de proteínas del sobrenadante se realizó por el
método de Bradford (1976).
45

Jlesuítadbs
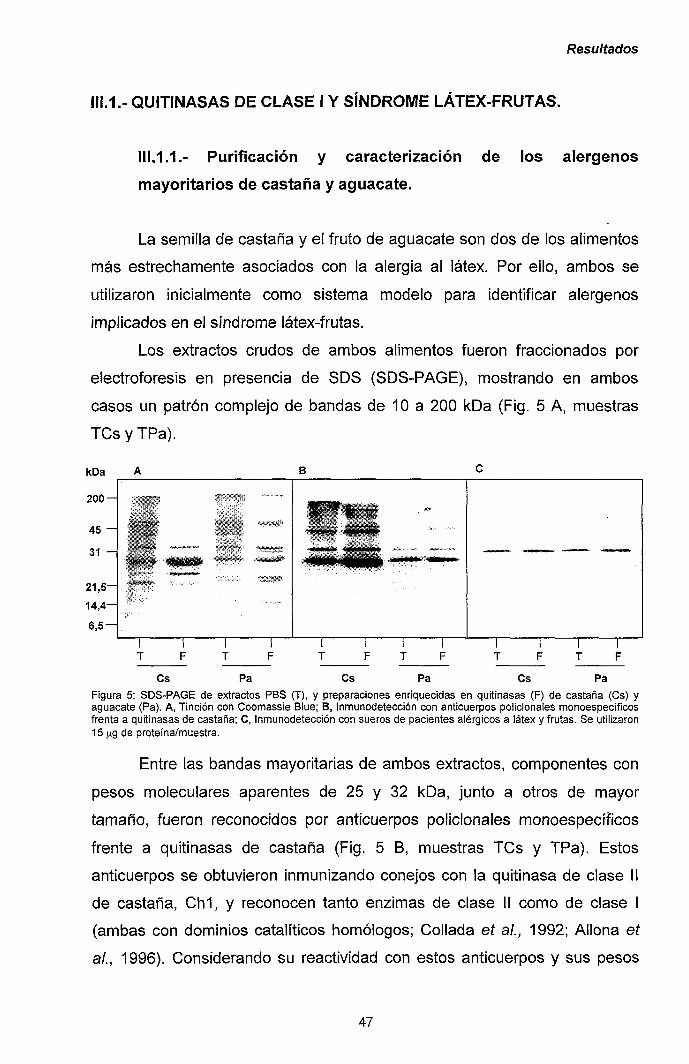
Resultados
III.1.- QUITINASAS DE CLASE I Y SÍNDROME LATEX-FRUTAS.
III.1.1.- Purificación y caracterización de los alérgenos
mayoritarios de castaña y aguacate.
La semilla de castaña y el fruto de aguacate son dos de los alimentos
más estrechamente asociados con la alergia al látex. Por ello, ambos se
utilizaron inicialmente como sistema modelo para identificar alérgenos
implicados en el síndrome látex-frutas.
Los extractos crudos de ambos alimentos fueron fraccionados por
electroforesis en presencia de SDS (SDS-PAGE), mostrando en ambos
casos un patrón complejo de bandas de 10 a 200 kDa (Fig. 5 A, muestras
TCs y TPa).
2 0 0 -
Cs Pa Os Pa Os Pa
Figura 5: SDS-PAGE de extractos PBS (T), y preparaciones enriquecidas en quitinasas (F) de castaña (Os) y aguacate (Pa). A, Tinción con Coomassie Blue; B, Inmunodetección con anticuerpos policlonales monoespecíficos frenta a quitinasas de castaña; C, Inmunodetección con sueros de pacientes alérgicos a látex y frutas. Se utilizaron 15 |.ig de proteína/muestra.
Entre las bandas mayoritarias de ambos extractos, componentes con
pesos moleculares aparentes de 25 y 32 kDa, junto a otros de mayor
tamaño, fueron reconocidos por anticuerpos policlonales monoespecíficos
frente a quitinasas de castaña (Fig. 5 B, muestras TCs y TPa). Estos
anticuerpos se obtuvieron inmunizando conejos con la quitinasa de clase II
de castaña, Chl, y reconocen tanto enzimas de clase II como de clase í
(ambas con dominios catalíticos homólogos; Collada eí al., 1992; Aliona et
al., 1996). Considerando su reactividad con estos anticuerpos y sus pesos
47

Resultados
moleculares, las bandas de 25 y 32 kDa corresponderían potencialmente a
quitinasas de clase II y I, respectivamente.
La detección de proteínas que fijaban IgE específica se realizó con
una mezcla de cuatro sueros de pacientes alérgicos a látex y frutas
(correspondientes a los pacientes 1, 2, 3, y 6 de la Tabla 3 de Material y
Métodos). Tanto la mezcla como los cuatro sueros individuales (datos no
mostrados), reconocieron una única banda de 32 kDa en los extractos PBS
de castaña y aguacate (Fig. 5 C, muestras TCs y TPa). Los sueros utilizados
como control negativo no reaccionaron con ningún componente de ambos
extractos.
En función de los datos anteriores, se obtuvieron preparaciones de
castaña y aguacate enriquecidas en quitinasas por precipitación selectiva a
pH 3,5 de los extractos PBS (ver apartado 11.2.1.2 de Material y Métodos).
Estas preparaciones mostraron un incremento en las bandas de 25 y 32
kDa, y un reconocimiento de componentes por sueros antiquitinasa y de los
pacientes alérgicos similar a la de los extractos crudos (Fig. 5, muestras F).
Los resultados descritos sugieren que algunas quitinasas de castaña
y de aguacate podrían ser alérgenos importantes en estas especies. Por
ello, se procedió a purificar miembros representativos de las clases I y II de
estas enzimas partiendo de las preparaciones enriquecidas en los mismos
(Fig. 6). Estas preparaciones fueron inicialmente fraccionadas mediante
cromatografía de afinidad en columnas de quitina regenerada (Fig. 6 A, para
castaña, y 6 D, para aguacate). Este método permitió obtener fracciones
retenidas que incluían las bandas de 25 kDa (pico 1 de Figs. 6 A y 6 D) y 32
kDa (pico 2 de Figs. 6 A y 6 D) como componentes mayoritarios. Cada una
de estas fracciones fueron separadas por un segundo sistema
cromatográfico en columnas de intercambio catiónico (Fig. 6 B,C, E, y F).
En el caso de castaña, se purificaron dos proteínas denominadas Csll
1 y Csl 3. Su correspondencia con las quitinasas Ch1 (clase II) y Ch3 (clase
I), previamente identificadas (Collada et al., 1992), se confirmó por coelución
del correspondiente enzima con los picos señalados en la figura 6B y 6C.
48
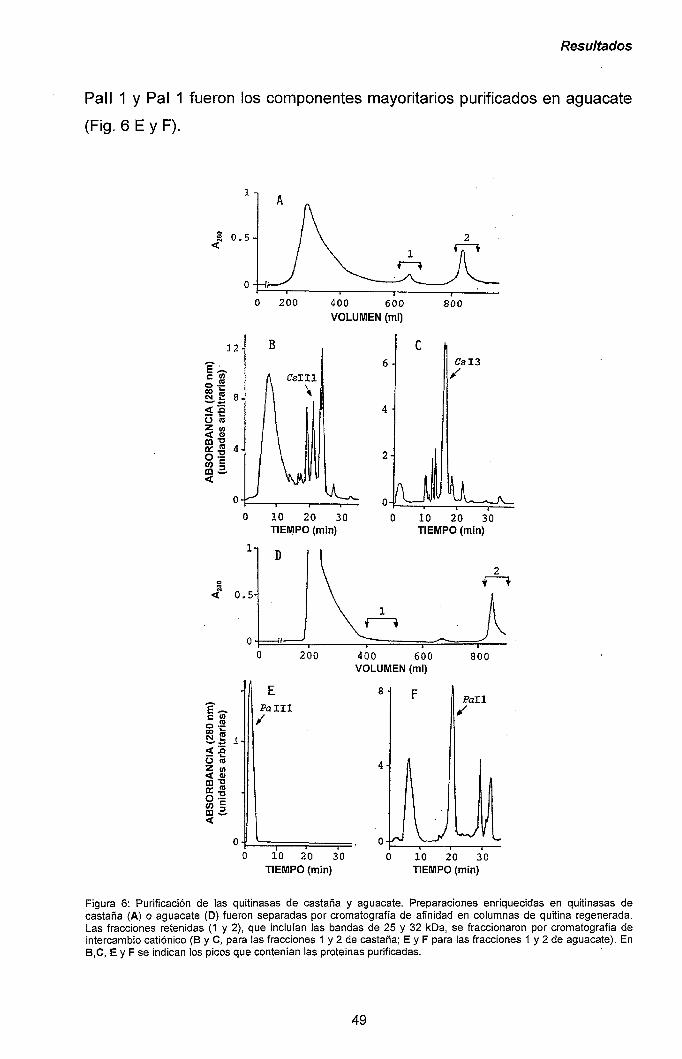
Resultados
Pal I 1 y Pal 1 fueron los componentes mayoritarios purificados en aguacate
(Fig. 6 E y F).
0.5
200 400 600 VOLUMEN (mi)
800
c 6-
4 •
2-
„.AJI
Csl3 /
ii_^ O 10 20 30
TIEMPO (mln) O 10 20 30
TIEMPO (mln)
TIEMPO (mln) 10 20 30 TIEMPO (mln)
Figura 6: Purificación de las quitinasas de castaña y aguacate. Preparaciones enriquecidas en quitinasas de castaña (A) o aguacate (D) fueron separadas por cromatografía de afinidad en columnas de quitina regenerada. Las fracciones retenidas (1 y 2), que incluían las bandas de 25 y 32 kDa, se fraccionaron por cromatografía de intercambio catiónico (B y C, para las fracciones 1 y 2 de castaña; E y F para las fracciones 1 y 2 de aguacate). En B,C, E y F se indican los picos que contenían las proteínas purificadas.
49
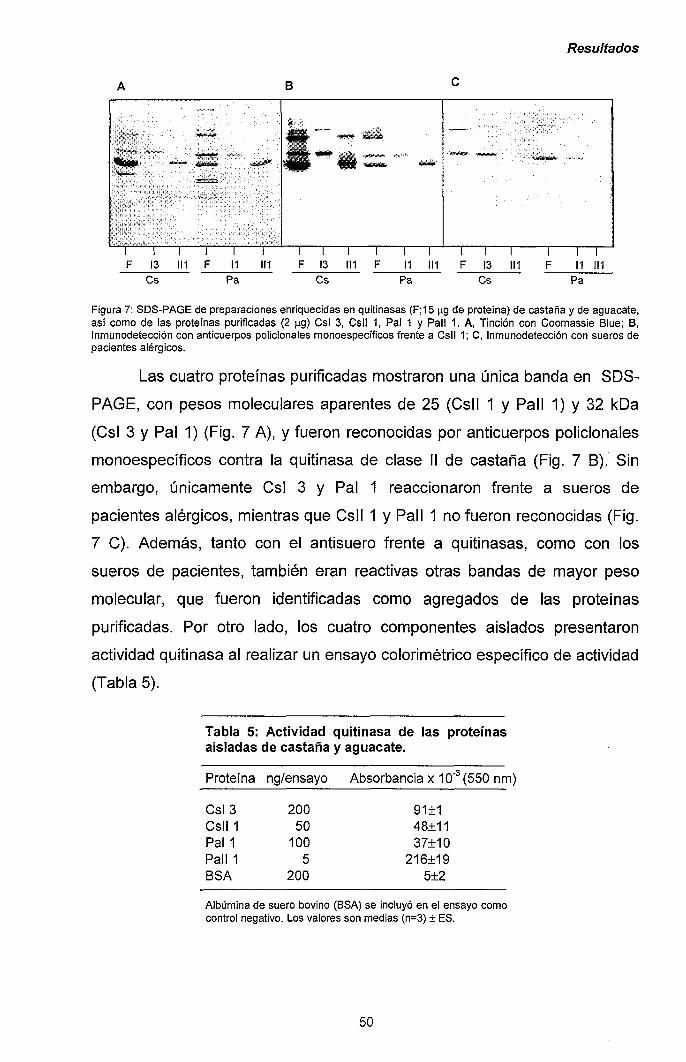
Resultados
B
1 — I — \ — I — r F 13 111 F 11
j B p ^ ^
1 — \ — I — r F 13 111 F
1 1 f-11 111 F 13 111 111
1 I 11 111
Cs Pa Cs Pa Cs Pa
Figura 7: SDS-PAGE de preparaciones enriquecidas en quitinasas (F;15 \ig de proteína) de castaña y de aguacate, así como de las proteínas purificadas (2 \IQ) Csl 3, Csll 1, Pal 1 y Pall 1. A, Tinción con Coomassie Blue; B, Inmunodetección con anticuerpos policlonales monoespecíficos frente a Csll 1; C, Inmunodetección con sueros de pacientes alérgicos.
Las cuatro proteínas purificadas mostraron una única banda en SDS-
PAGE, con pesos moleculares aparentes de 25 (Csll 1 y Pall 1) y 32 kDa
(Csl 3 y Pal 1) (Fig. 7 A), y fueron reconocidas por anticuerpos policlonales
monoespecíficos contra la quitinasa de clase II de castaña (Fig. 7 B). Sin
embargo, únicamente Csl 3 y Pal 1 reaccionaron frente a sueros de
pacientes alérgicos, mientras que Csll 1 y Pall 1 no fueron reconocidas (Fig.
7 C). Además, tanto con el antisuero frente a quitinasas, como con los
sueros de pacientes, también eran reactivas otras bandas de mayor peso
molecular, que fueron Identificadas como agregados de las proteínas
purificadas. Por otro lado, los cuatro componentes aislados presentaron
actividad quitinasa al realizar un ensayo colorimétrico específico de actividad
(Tabla 5).
Tabla 5: Actividad quitinasa de las proteínas aisladas de castaña y aguacate.
Proteína ng/ensayo Absorbanciax 10'^(550 nm)
Csl 3 CslM Pan Pall 1 BSA
200 50
100 5
200
91±1 48±11 37±10
216±19 5±2
Albúmina de suero bovino (BSA) se incluyó en el ensayo como control negativo. Los valores son medias (n=3) + ES.
50

Resultados
Un dato adicional en la caracterización de las proteínas fue la
obtención de sus secuencias N-terminales. Mientras en el caso de Csll 1 y
Csl 3 había determinaciones previas de sus estructuras primarias (Collada et
al., 1992; Aliona et al., 1996), no ocurría lo mismo con las proteínas de
aguacate. La secuenciación del extremo N-terminal de Csl 3 y Pal 1 (Fig. 8)
permitió mostrar el alto grado de similitud de ambas proteínas, que
compartían 22 de los 27 residuos secuenciados. Además, ambas tenían más
del 80% de identidad con la correspondiente secuencia de la heveína de
látex. La secuencia de Csl 3 era idéntica a la previamente deducida a partir
de su correspondiente clon de ADNc (Aliona et al., 1996).
Alérgeno Secuencia aminoterminal Identidad (%) 1 10 20 Pal 1 Csl 3 Hev.
Pal 1 EQCGRQAGGALCPGGLCCSQFGWCGST — 82 85 Csl 3 EQCGRQAGGAACANNLCCSQFGWCGNT — 82 Heveína EQCGRQAGGKLCPNNLCCSQWGWCGST
Figura 8; Secuencia N-terminal de la quitinasa de clase I de aguacate (Pal 1) y de castaña (Csl 3), y de la heveína de Hevea brasHiensis. En negrita, se han señalado los residuos diferentes de Csl 3 y de la heveína con respecto a Pal 1.
El análisis de dos preparaciones distintas de Pal I 1 indicó que esta
quitinasa tenía probablemente bloqueado su extremo N-terminal. Se
determinó su composición de aminoácidos, con el fin de compararla con las
de Csll 1 y Csl 3 previamente descritas (Collada et al., 1992; Tabla 6). El
perfil de aminoácidos de Pall 1 era más similar al de Csll 1 que al de Csl 3,
especialmente en el contenido en cisteína, que diferencia a las enzimas de
clase II, sin dominio heveína, de las de clase I que tienen dicho dominio, rico
en el citado aminoácido.
El conjunto de resultados expuestos permite definir a Pal 1 y Csl 3
como quitinasas de clase I con un dominio heveína N-terminal, y a Pal! 1 y
Csll 1 como sus homólogos de clase II carentes de dicho dominio. Los
resultados de inmunodetección con sueros de pacientes alérgicos (Fig. 7 C)
indicaban que únicamente Pal 1 y Csl 3 poseían capacidad de ligar IgE
específica. Este hecho fue plenamente confirmado por ensayos de
inmunoinhibición (Fig. 9). Las quitinasas de clase I inhibieron completamente
51
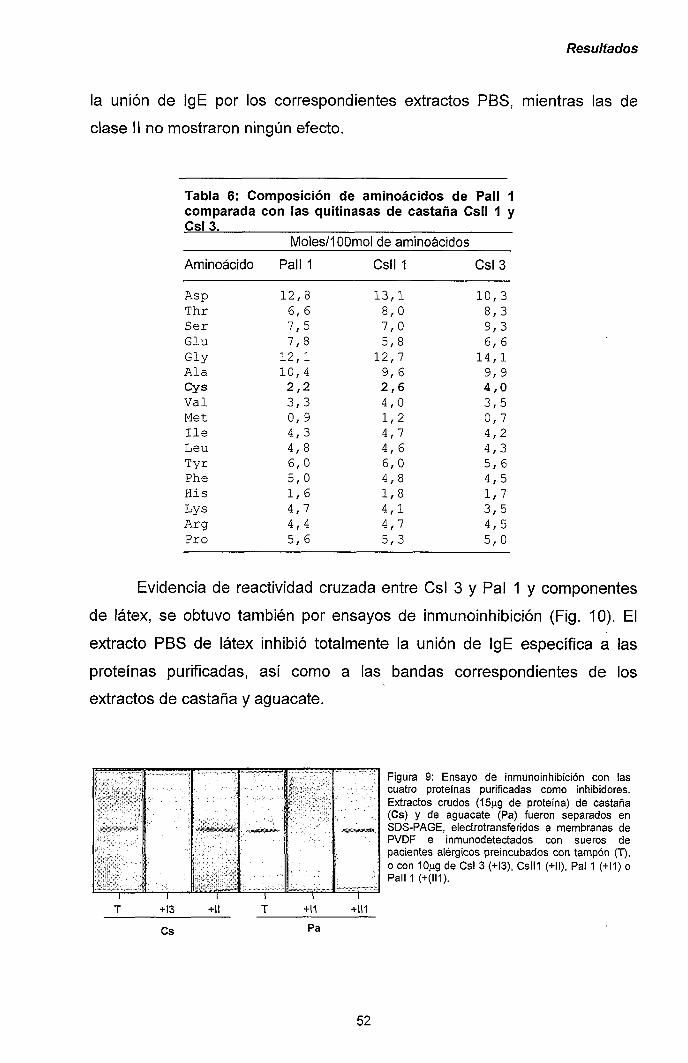
Resultados
la unión de IgE por los correspondientes extractos PBS, mientras las de
clase II no mostraron ningún efecto.
Tabla 6: Composición de aminoácidos de Pall 1 comparada con las quitinasas de castaña Csll 1 y Csl3.
Moies/100mol de aminoácidos Aminoácido Pall 1 CslM Csl3
Asp Thr Ser Glu Gly Ala Cys Val Met H e Leu Tyr Phe His Lys Arg Pro
12,8 6,6 7,5 7,8
12, 1 10,4 2,2 3,3 0,9 4,3 4,8 6,0 5,0 1,6 4,7 4,4 5, 6
13,1 8,0 7,0 5,8
12,7 9,6 2,6 4,0 1,2 4,7 4,6 6,0 4,8 1,8 4,1 4,7 5,3
10,3 8,3 9,3 6,6
14,1 9,9 4,0 3,5 0,7 4,2 4,3 5, 6 4,5 1,7 3,5 4,5 5,0
Evidencia de reactividad cruzada entre Csl 3 y PaM y componentes
de látex, se obtuvo también por ensayos de inmunoinhibición (Fig. 10). El
extracto PBS de látex inhibió totalmente la unión de IgE específica a las
proteínas purificadas, así como a las bandas correspondientes de los
extractos de castaña y aguacate.
1 ' [
j 1
^™"r"
.,j;!=ssí
1
í
1
JlíaíV***'!*»^
„.. ^.
1 +13
Os
+11 +11
Pa
+111
Figura 9: Ensayo de inmunoinhibición con las cuatro proteínas purificadas como inhibidores. Extractos crudos (15p.g de proteina) de castaña (Os) y de aguacate (Pa) fueron separados en SDS-PAGE, electrotransferidos a membranas de PVDF e inmunodetectados con sueros de pacientes alérgicos preincubados con tampón (T), o con lOng de Csl 3 (+13), Cslll (+11), Pal 1 (+11) o Pall 1 (+(111).
52
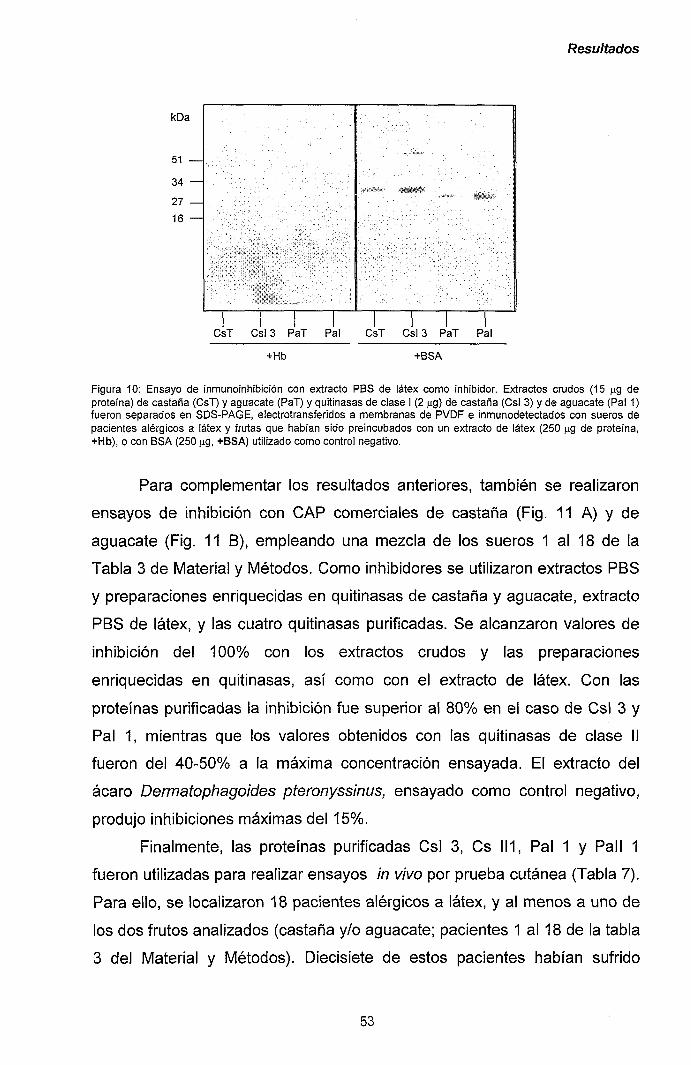
Resultados
kDa
CsT Csl3 PaT Pal
+Hb
CsT Csl3 PaT Pal
+BSA
Figura 10: Ensayo de Inmunoinhibiclón con extracto PBS de látex como inhibidor. Extractos crudos (15 i g de proteína) de castaña (CsT) y aguacate (PaT) y quitinasas de clase I (2 ng) de castaña (Csl 3) y de aguacate (Pal 1) fueron separados en SDS-PAGE, electrotransferidos a membranas de PVDF e inmunodetectados con sueros de pacientes alérgicos a látex y frutas que habían sido preincubados con un extracto de látex (250 ^g de proteína, +Hb), o con BSA (250 \ig, +BSA) utilizado como control negativo.
Para complementar los resultados anteriores, también se realizaron
ensayos de inhibición con CAP comerciales de castaña (Fig. 11 A) y de
aguacate (Fig. 11 B), empleando una mezcla de los sueros 1 al 18 de la
Tabla 3 de Material y Métodos. Como inhibidores se utilizaron extractos PBS
y preparaciones enriquecidas en quitinasas de castaña y aguacate, extracto
PBS de látex, y las cuatro quitinasas purificadas. Se alcanzaron valores de
inhibición del 100% con los extractos crudos y las preparaciones
enriquecidas en quitinasas, así como con el extracto de látex. Con las
proteínas purificadas la inhibición fue superior al 80% en el caso de Csl 3 y
Pal 1, mientras que los valores obtenidos con las quitinasas de clase II
fueron del 40-50% a la máxima concentración ensayada. El extracto del
acaro Dermatophagoides pteronyssinus, ensayado como control negativo,
produjo inhibiciones máximas del 15%.
Finalmente, las proteínas purificadas Csl 3, Cs III , Pal 1 y Pall 1
fueron utilizadas para realizar ensayos in vivo por prueba cutánea (Tabla 7).
Para ello, se localizaron 18 pacientes alérgicos a látex, y al menos a uno de
los dos frutos analizados (castaña y/o aguacate; pacientes 1 al 18 de la tabla
3 del Material y Métodos). Diecisiete de estos pacientes habían sufrido
53
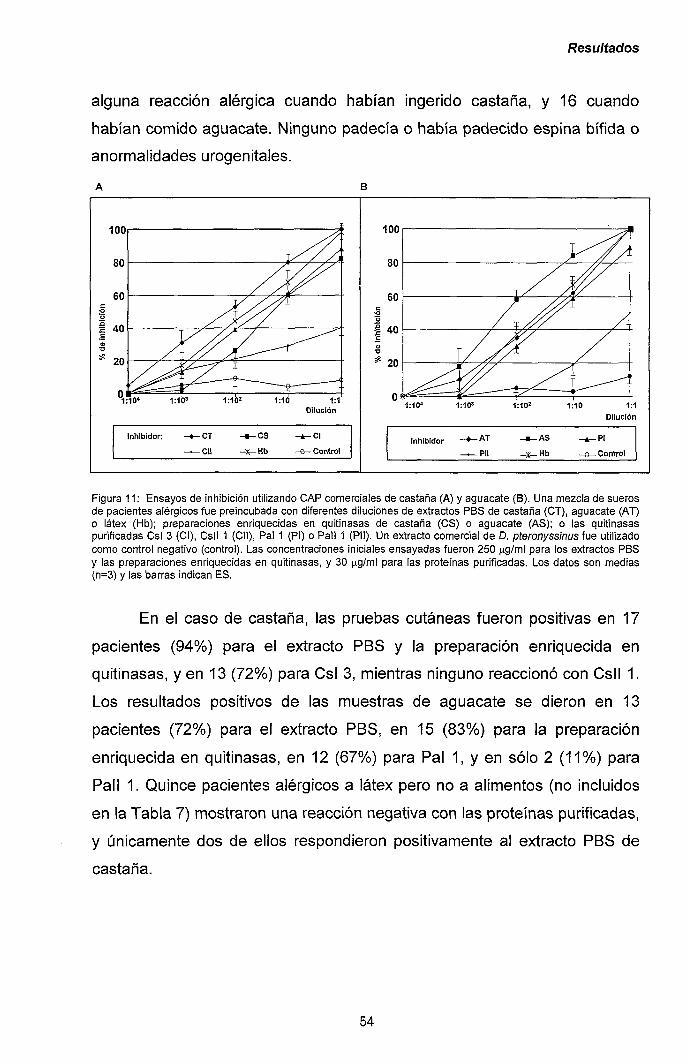
Resultados
alguna reacción alérgica cuando habían ingerido castaña, y 16 cuando
habían comido aguacate. Ninguno padecía o había padecido espina bífida o
anormalidades urogenitales.
100
1:10« 1:10' 1:10^ 1:10 1:1 Dilución
Inhibidor: -CT
-CU
-es -Hb
-Cl
- Control
100
§ 40
inhibidor -AT
-Pll
-AS
-Hb
-Pl
- Control
Figura 11: Ensayos de inhibición utilizando CAP comerciales de castaña (A) y aguacate (B). Una mezcla de sueros de pacientes alérgicos fue preincubada con diferentes diluciones de extractos PBS de castaña (CT), aguacate (AT) o látex (Hb); preparaciones enriquecidas en quitinasas de castaña (OS) o aguacate (AS); o las quitinasas purificadas Csl 3 (01), Gsll 1 (CU), Pal 1 (Pl) o Pall 1 (Pll). Un extracto comercial de D. pteronyssinus fue utilizado como control negativo (control). Las concentraciones Iniciales ensayadas fueron 250 |xg/ml para los extractos PBS y las preparaciones enriquecidas en quitinasas, y 30 ng/ml para las proteínas purificadas. Los datos son medias (n=3) y las barras indican ES.
En el caso de castaña, las pruebas cutáneas fueron positivas en 17
pacientes (94%) para el extracto PBS y la preparación enriquecida en
quitinasas, y en 13 (72%) para Csl 3, mientras ninguno reaccionó con Csll 1.
Los resultados positivos de las muestras de aguacate se dieron en 13
pacientes (72%) para el extracto PBS, en 15 (83%) para la preparación
enriquecida en quitinasas, en 12 (67%) para Pal 1, y en sólo 2 (11%) para
Pall 1. Quince pacientes alérgicos a látex pero no a alimentos (no incluidos
en la Tabla 7) mostraron una reacción negativa con las proteínas purificadas,
y únicamente dos de ellos respondieron positivamente al extracto PBS de
castaña.
54
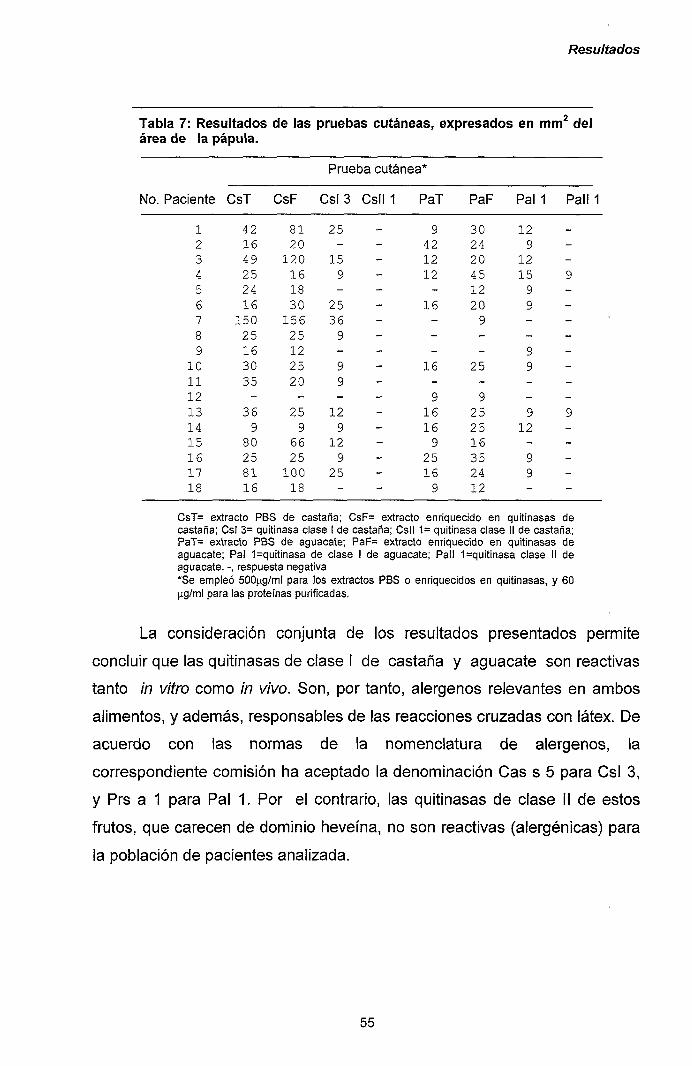
Resultados
Tabla 7: Resultados de las pruebas cutáneas, expresados en mm^ del área de la pápula.
Prueba cutánea*
No. Paciente CsT CsF Csl 3 Csll 1 PaT PaF Pal 1 Pall 1
1 2 3 4 5 6 7 8 9
10 11 12 13 14 15 16 17 18
42 16 49 25 24 16 150 25 16 30 35 -36 9
80 25 81 16
81 20 120 16 18 30 156 25 12 25 20 -25 9
66 25 100 18
25 -15 9 -
25 36 9 -9 9 -12 9
12 9
25 -
9 42 12 12 -16 -
-
16
9 16 16 9
25 16 9
30 24 20 45 12 20 9
_
25
9 25 25 16 35 24 12
12 9
12 15 9 9 -
9 9
-
9 12 -9 9 -
CsT= extracto PBS de castaña; CsF= extracto enriquecido en quitinasas de castaña; Csl 3= quitinasa clase I de castaña; Csll 1= quitinasa clase II de castaña; PaT= extracto PBS de aguacate; PaF= extracto enriquecido en quitinasas de aguacate; Pal 1=quitinasa de clase I de aguacate; Pall 1=quitinasa clase II de aguacate. -, respuesta negativa *Se empleó SOO^g/ml para los extractos PBS o enriquecidos en quitinasas, y 60 |.ig/ml para las proteínas purificadas.
La consideración conjunta de los resultados presentados permite
concluir que las quitinasas de clase I de castaña y aguacate son reactivas
tanto in vitro como in vivo. Son, por tanto, alérgenos relevantes en ambos
alimentos, y además, responsables de las reacciones cruzadas con látex. De
acuerdo con las normas de la nomenclatura de alérgenos, la
correspondiente comisión ha aceptado la denominación Cas s 5 para Csl 3,
y Prs a 1 para Pal 1. Por el contrario, las quitinasas de clase II de estos
frutos, que carecen de dominio heveína, no son reactivas (alergénicas) para
la población de pacientes analizada.
55
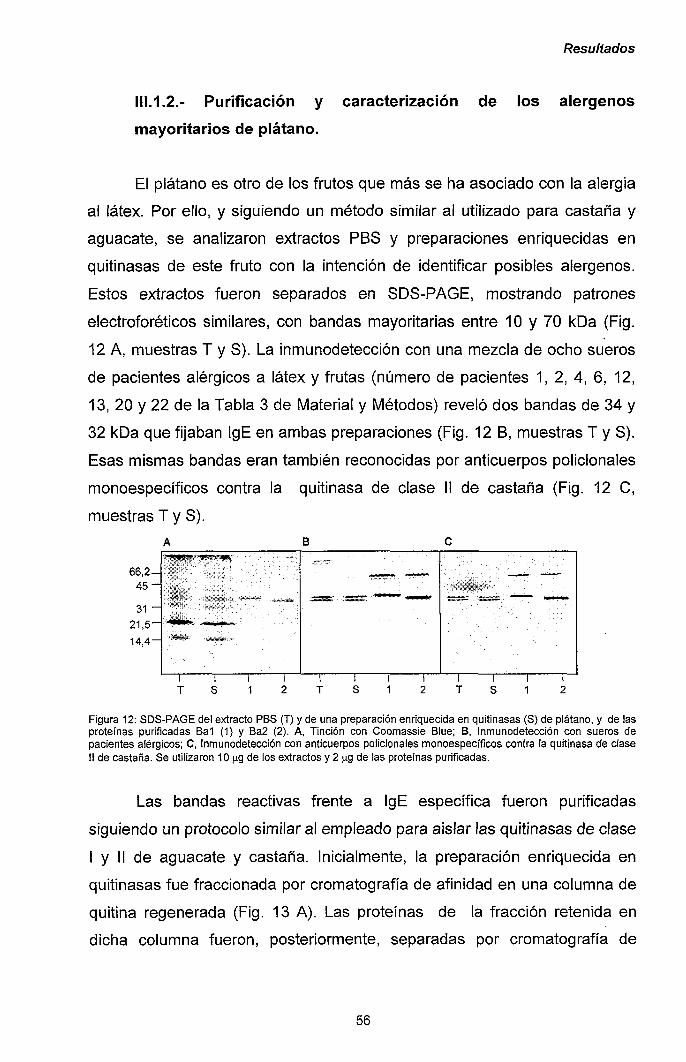
Resultados
III.1.2.- Purificación y caracterización de los alérgenos
mayoritarios de plátano.
El plátano es otro de los frutos que más se ha asociado con la alergia
al látex. Por ello, y siguiendo un método similar al utilizado para castaña y
aguacate, se analizaron extractos PBS y preparaciones enriquecidas en
quitinasas de este fruto con la intención de identificar posibles alérgenos.
Estos extractos fueron separados en SDS-PAGE, mostrando patrones
electroforéticos similares, con bandas mayoritarias entre 10 y 70 kDa (Fig.
12 A, muestras T y S). La inmunodetección con una mezcla de ocho sueros
de pacientes alérgicos a látex y frutas (número de pacientes 1, 2, 4, 6, 12,
13, 20 y 22 de la Tabla 3 de Material y Métodos) reveló dos bandas de 34 y
32 kDa que fijaban IgE en ambas preparaciones (Fig. 12 B, muestras T y S).
Esas mismas bandas eran también reconocidas por anticuerpos policlonales
monoespecíficos contra la quitinasa de clase II de castaña (Fig. 12 C,
muestras T y S).
A B C
66,2-4 5 -
31 -21,5-14,4-
'**"*iSSw^"*s*Kmiiv
<<•. ...^..„.
-mmm- —.•.....
l i l i
, .-
•"""'i'i'' •— "'
1 1
• , , , , . , —
• •mmmm
1 1
- • ' . • . * • - ,
iriísr: '•^^'-'^^
1 1
——
1» lili
1 1 1 1 1
Figura 12: SDS-PAGE del extracto PBS (T) y de una preparación enriquecida en quitinasas (S) de plátano, y de las proteínas purificadas Bal (1) y Ba2 (2). A, Tinción con Coomassie Blue; B, Inmunodetección con sueros de pacientes alérgicos; O, Inmunodetección con anticuerpos policlonales monoespecíficos contra la quitinasa de clase II de castaña. Se utilizaron 10 jxg de los extractos y 2 fig de las proteínas purificadas.
Las bandas reactivas frente a IgE específica fueron purificadas
siguiendo un protocolo similar al empleado para aislar las quitinasas de clase
I y II de aguacate y castaña. Inicialmente, la preparación enriquecida en
quitinasas fue fraccionada por cromatografía de afinidad en una columna de
quitina regenerada (Fig. 13 A). Las proteínas de la fracción retenida en
dicha columna fueron, posteriormente, separadas por cromatografía de
56
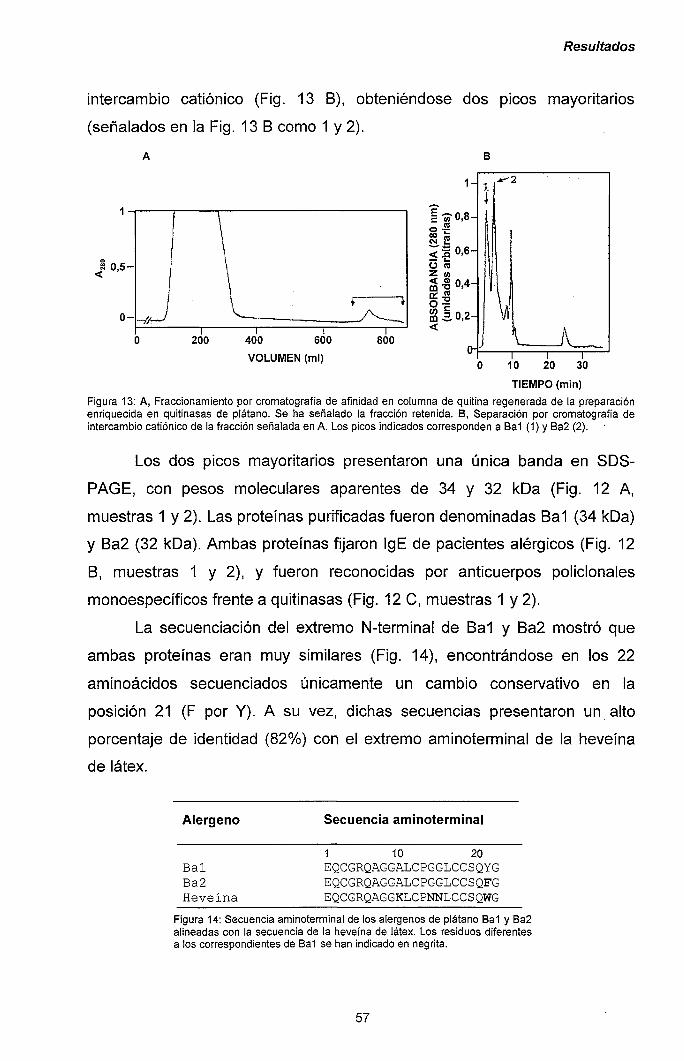
Resultados
intercambio catiónico (Fig. 13 B), obteniéndose dos picos mayoritarios
(señalados en la Fig. 13 B como 1 y 2).
0,5-
400 600 VOLUMEN (mi) „ ^„ „ „ , „
^ ' O 10 20 30 TIEMPO (min)
Figura 13: A, Fraccionamiento por cromatografía de afinidad en columna de quitina regenerada de la preparación enriquecida en quitinasas de plátano. Se ha señalado la fracción retenida. B, Separación por cromatografía de intercambio catiónico de la fracción señalada en A. Los picos indicados corresponden a Ba1 (1) y Ba2 (2).
Los dos picos mayoritarios presentaron una única banda en SDS-
PAGE, con pesos moleculares aparentes de 34 y 32 kDa (Fig. 12 A,
muestras 1 y 2). Las proteínas purificadas fueron denominadas Bal (34 kDa)
y Ba2 (32 kDa). Ambas proteínas fijaron IgE de pacientes alérgicos (Fig. 12
B, muestras 1 y 2), y fueron reconocidas por anticuerpos policlonales
monoespecíficos frente a quitinasas (Fig. 12 C, muestras 1 y 2).
La secuenciación del extremo N-terminal de Bal y Ba2 mostró que
ambas proteínas eran muy similares (Fig. 14), encontrándose en los 22
aminoácidos secuenciados únicamente un cambio conservativo en la
posición 21 (F por Y). A su vez, dichas secuencias presentaron un alto
porcentaje de identidad (82%) con el extremo aminoterminal de la heveína
de látex.
Alérgeno Secuencia aminoterminal
Bal Ba2 Heveína
1 10 20 EQCGRQAGGALCPGGLCCSQYG EQCGRQAGGALCPGGLCCSQFG EQCGRQAGGKLCPNNLCCSQWG
Figura 14: Secuencia aminoterminal de los alérgenos de plátano Bal y Ba2 alineadas con la secuencia de la heveína de látex. Los residuos diferentes a los correspondientes de Bal se han indicado en negrita.
57
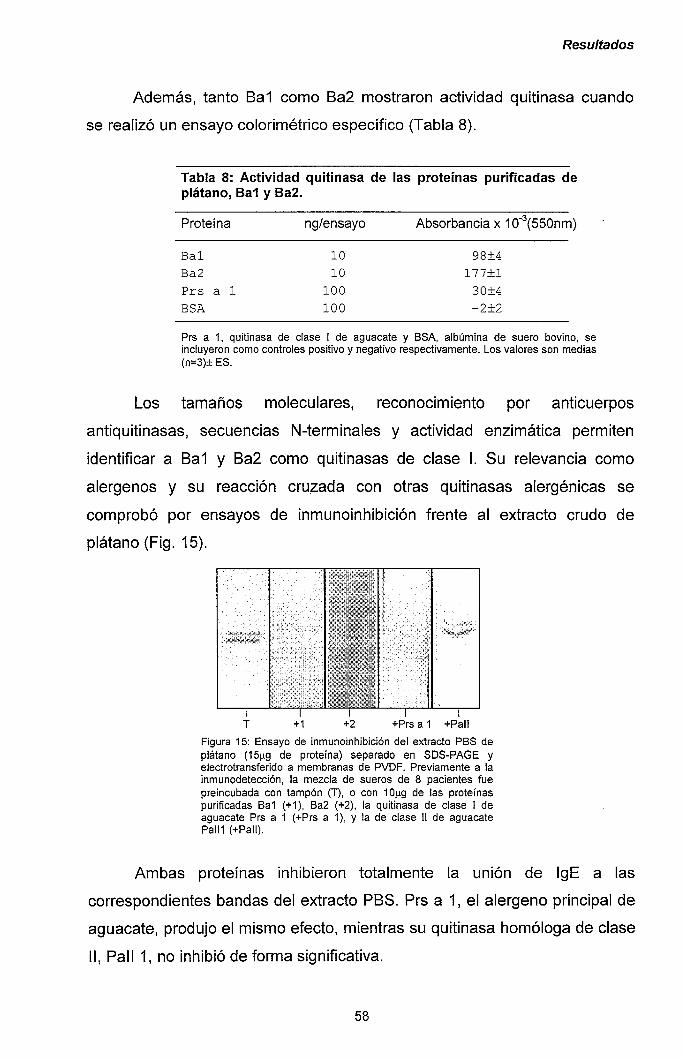
Resultados
Además, tanto Ba1 como Ba2 mostraron actividad quitinasa cuando
se realizó un ensayo colorimétrico específico (Tabla 8).
Tabla 8: Actividad quitinasa de las proteínas purificadas de plátano, Ba1 y Ba2.
Proteína ng/ensayo Absorbancia x 10" (550nm)
Bal Ba2
Prs a 1
ESA
10 10
100
100
98+4
177±1
30±4
-2±2
Prs a 1, quitinasa de clase I de aguacate y BSA, albúmina de suero bovino, se incluyeron como controles positivo y negativo respectivamente. Los valores son medias (n=3)± ES.
Los tamaños moleculares, reconocimiento por anticuerpos
antiqultinasas, secuencias N-terminales y actividad enzimática permiten
identificar a Ba1 y Ba2 como quitinasas de clase I. Su relevancia como
alérgenos y su reacción cruzada con otras quitinasas alergénicas se
comprobó por ensayos de inmunoinhibición frente al extracto crudo de
plátano (Fig. 15).
T +1 +2 —r
+Prs a 1 +Pall Figura 15: Ensayo de inmunoinhibición del extracto POS de plátano (15)19 de proteina) separado en SDS-PAGE y electrotransferido a membranas de PVDF. Previamente a la inmunodetección, la mezcla de sueros de 8 pacientes fue preincubada con tampón (T), o con lO^g de las proteínas purificadas Bal (+1), Ba2 (+2), la quitinasa de clase I de aguacate Prs a 1 (+Prs a 1), y la de clase II de aguacate Pain (+Pall).
Ambas proteínas inhibieron totalmente la unión de IgE a las
correspondientes bandas del extracto PBS. Prs a 1, el alérgeno principal de
aguacate, produjo el mismo efecto, mientras su quitinasa homologa de clase
II, Rail 1, no inhibió de forma significativa.
58

Resultados
1:10' 1:10' 1:10' 1:1 1:1
Dilución
Inhibidores: 1-^-2-x-C:
Figura 16: Ensayos de inhibición utilizando CAP comercial de plátano. Como inhibidores se emplearon extracto PBS (T) y una preparación enriquecida en quitinasas (S) de plátano, así como las proteínas purificadas Bal (1) y Ba2 (2). El extracto comercial de D. pteronyssinus fue usado como control negativo (C). La mezcla de 15 sueros fue preincubada con diluciones crecientes de una concentración inicial de inhibidores de 250 jig/ml para los extractos y 30 ng/ml para las proteínas purificadas.
Estos resultados se confirmaron por ensayos de inhibición en CAP
(Fig. 16). Para dichos ensayos se empleó un CAP comercial de plátano que
fue incubado con una mezcla de sueros de 15 pacientes (número de
pacientes 1, 2, 3, 4, 5, 6, 7, 9, 10, 12, 13, 19, 20, 21, y 22 de la Tabla 3 de
Material y Métodos). Los valores de inhibición que se obtuvieron fueron del
90% para ambas proteínas purificadas (Fig. 16, muestras 1 y 2), siendo más
efectivas que el extracto crudo de plátano (T) o el enriquecido en quitinasas
(S), cuando se consideran cantidades equivalentes de proteína en el ensayo.
Por último, se realizaron ensayos in vivo por prueba cutánea,
utilizando extracto PBS y una preparación enriquecida en quitinasas de
plátano, así como las proteínas purificadas Bal y Ba2, en el grupo de 15
pacientes con alergia a látex y plátano (señalado anteriormente). En la Tabla
9, se resumen los resultados de este ensayo. El 93% de los individuos
tuvieron una reacción positiva con el extracto PBS de plátano, y el 87% con
el enriquecido en quitinasas. Bal fue reconocida por el 53% de los
pacientes, mientras que Ba2 lo fue por el 60% de los mismos. Puede, por
tanto, concluirse que las quitinasas de clase I también son alérgenos
59
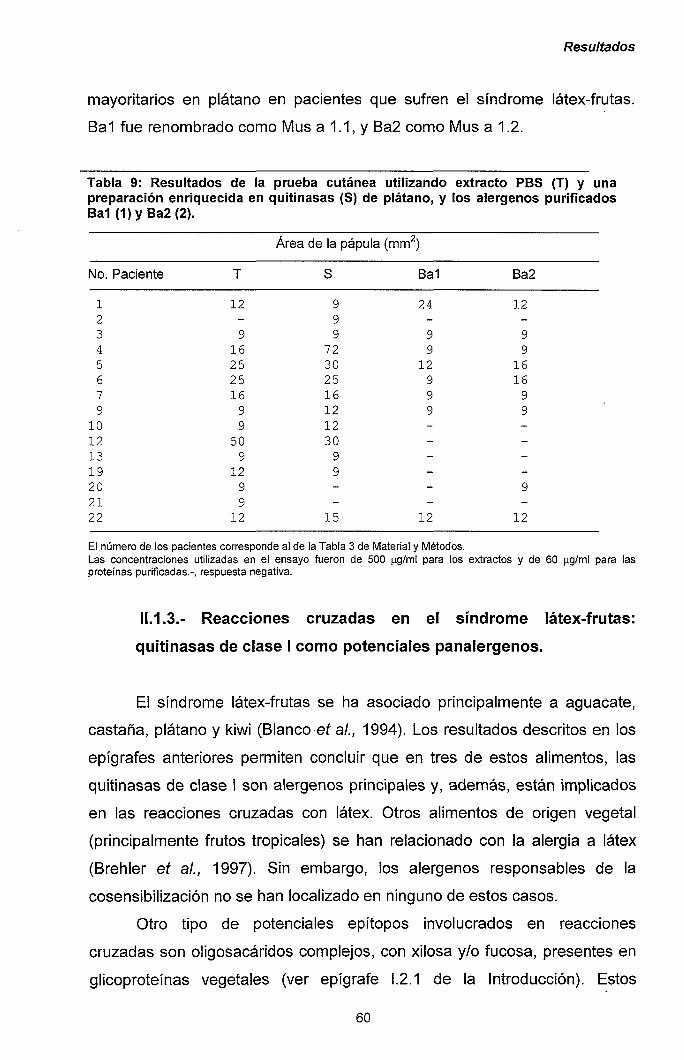
Resultados
mayoritarios en plátano en pacientes que sufren el síndrome látex-frutas.
Ba1 fue renombrado como Mus a 1.1, y Ba2 como Mus a 1.2.
Tabla 9: Resultados de la prueba cutánea utilizando extracto PBS (T) y una preparación enriquecida en quitinasas (S) de plátano, y los alérgenos purificados Ba1 (1)yBa2(2).
No. Paciente
1 2 3 4 5 6 7 9 10 12 13 19 20 21 22
T
12 -9 16 25 25 16 9 9 50 9 12 9 9 12
Área de la pápula
S
9 9 9 72 30 25 16 12 12 30 9 9 --15
(mm^)
Ba1
24 -9 9 12 9 9 9 ------12
Ba2
12 -9 9 16 16 9 9 ----9 -12
El número de los pacientes corresponde al de la Tabla 3 de Material y Métodos. Las concentraciones utilizadas en el ensayo fueron de 500 ^g/ml para los extractos y de 60 ng/ml para las proteínas purificadas.-, respuesta negativa.
11.1.3.- Reacciones cruzadas en el síndrome látex-frutas:
quitinasas de clase I como potenciales panalergenos.
El síndrome látex-frutas se ha asociado principalmente a aguacate,
castaña, plátano y kiwi (Blanco ef al., 1994). Los resultados descritos en los
epígrafes anteriores permiten concluir que en tres de estos alimentos, las
quitinasas de clase I son alérgenos principales y, además, están implicados
en las reacciones cruzadas con látex. Otros alimentos de origen vegetal
(principalmente frutos tropicales) se han relacionado con la alergia a látex
(Brehier et al., 1997). Sin embargo, los alérgenos responsables de la
cosensibilización no se han localizado en ninguno de estos casos.
Otro tipo de potenciales epítopos involucrados en reacciones
cruzadas son oligosacáridos complejos, con xilosa y/o fucosa, presentes en
glicoproteínas vegetales (ver epígrafe 1.2.1 de la Introducción). Estos
60

Resultados
oügosacáridos se han descrito en la cosensibilización entre pólenes y látex
(Fuchsefa/., 1998).
Para evaluar cual es el posible papel de las quitinasas y de los
oligosacáridos complejos en el síndrome látex-frutas, fueron analizados
extractos de 20 alimentos vegetales con diferente grado de asociación al
síndrome, así como extracto de látex y leche en polvo comercial (ver
Material y Métodos). Un patrón complejo, con bandas entre 6 y 100 kDa, se
obtuvo en la mayoría de las muestras, aunque hubo extractos que mostraron
un número muy limitado de bandas (p.e. pina; Fig. 17 A).
Para determinar el papel de los N-oligosacáridos complejos, se realizó
una inmunodetección usando un suero específico frente a los mismos
(Fig. 17 B). Se observaron varias bandas reactivas, de entre 30 y 100 kDa,
en casi todos los extractos. La leche desnatada, que se utilizó como control
negativo (Fig. 17 B, muestra M), no mostró ningún componente reactivo
frente a este suero, como era esperado.
Las putativas quitinasas presentes en los distintos extractos se
localizaron mediante una inmunodetección con anticuerpos policlonales
monoespecíficos frente a la quitinasa de clase 11 de castaña (Fig. 17 C).
Además de en castaña, donde se detectaron las bandas de 25 (clase II) y 32
kDa (clase I) anteriormente descritas, fueron localizadas potenciales
quitinasas (de 1 a 6 bandas, con tamaños moleculares de 20 a 75 kDa,
dependiendo de la muestra) en 14 alimentos analizados. Algunas de estas
bandas reactivas también fijaban IgE específica cuando los extractos fueron
inmunodetectados con una mezcla de sueros de pacientes alérgicos (Fig. 17
D; pacientes 1, 2, 4, 6, 12, 13, 20, y 22 de la Tabla 3 de Material y Métodos).
Estos componentes reconocidos por ambos sueros se localizaron en
chirimoya (46 kDa), fruta de la pasión (35 kDa), kiwi (41 kDa), látex (35-45
kDa), papaya (30-35 kDa), mango (46 kDa), tomate (31 kDa) y trigo (33
kDa). Bandas adicionales que unían IgE específica, pero que no
61
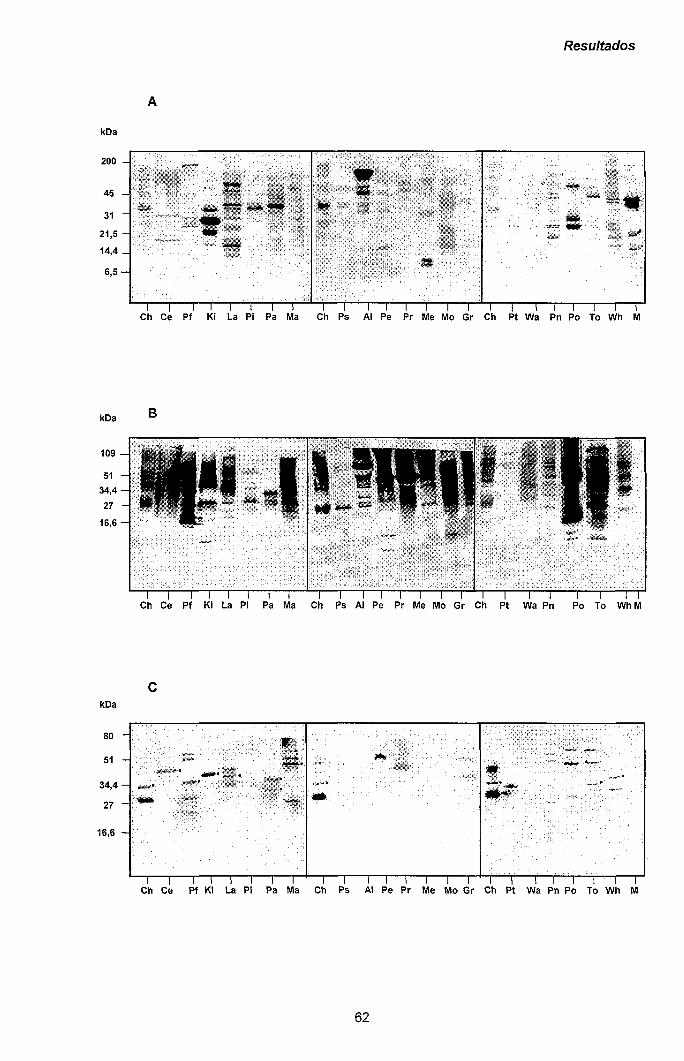
Resultados
kDa
200
45
31
21,5
14,4
6,5-
-
^
1
' ,tf^^, ^^a^ íi^fünif *iéii0
l i l i l í
—
m
1
W 1 ^
- "?.;;
1 1 1
•-•,, , -
1 9
l i l i
'•' <mm
1 I l i l i
.-
1 1 Ch Ce Pf Ki La Pi Pa Ma Ch Ps Al Pe Pr Me Mo Gr Ch Pt Wa Pn Po To Wh M
kDa B
109 -
1 I r Ch Ce Pf Ki La Pi Pa Ma Ch
1—I—I—I—I—I—m-Ps Al Pe Pr Me Mo Gr Ch
1—r Wa Pn
1 TT Po To Wh M
kDa
80
51
34,4-
27 •
16,6 -
,
1
r tíií^í^^*
1 1 1
íKww^ a
t ^ ' '
1 1 I 1
. ^ * r
1 1
^»*
l i l i l í 1 1 1
•' «*» * "
, — ••
1 1 1 1 1 La Pi Pa Ma Ch Ps
62
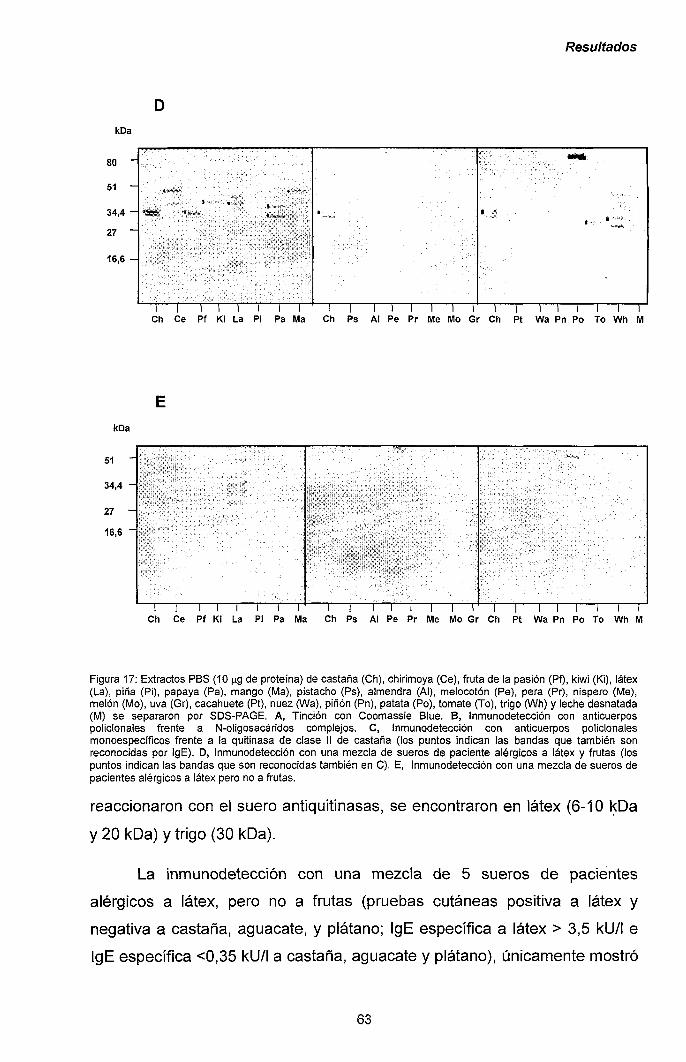
Resultados
kDa
T — I — 1 — I — 1 — 1 — r " — I — i — I — 1 — 1 — I — I — r — I — i — I — \ — 1 — I — I — r Ch Ce Pf Ki La P¡ Pa Ma Ch Ps Al Pe Pr Me Mo Gr Ch Pt Wa Pn Po To Wh M
kDa
51 -
34,4 -
27
16,6 -
I I l i l i l í I I l i l i l í
^.
1 1 l i l i l í Ch Ps Al Pe Pr Me Mo Gr Ch Pt Wa Pn Po To Wh M
Figura 17: Extractos PBS (10 ng de proteína) de castaña (Ch), chirimoya (Ce), fruta de la pasión (Pf), kiwi (Ki), látex (La), pina (P¡), papaya (Pa), mango (Ma), pistacho (Ps), almendra (Al), melocotón (Pe), pera (Pr), níspero (Me), melón (Mo), uva (Gr), cacahuete (Pt), nuez (Wa), piñón (Pn), patata (Po), tomate (To), trigo (Wh) y leche desnatada (M) se separaron por SDS-PAGE. A, Tinción con Coomassie Blue. B, Inmunodetección con anticuerpos policlonales frente a N-oligosacárídos complejos. C, Inmunodetección con anticuerpos policlonales monoespecíficos frente a la quitinasa de clase II de castaña (los puntos indican las bandas que también son reconocidas por IgE). D, Inmunodetección con una mezcla de sueros de paciente alérgicos a látex y frutas (los puntos indican las bandas que son reconocidas también en C). E, Inmunodetección con una mezcla de sueros de pacientes alérgicos a látex pero no a frutas.
reaccionaron con el suero antiquitinasas, se encontraron en látex (6-10 kDa
y 20 kDa) y trigo (30 kDa).
La inmunodetección con una mezcla de 5 sueros de pacientes
alérgicos a látex, pero no a frutas (pruebas cutáneas positiva a látex y
negativa a castaña, aguacate, y plátano; IgE específica a látex > 3,5 kU/l e
IgE específica <0,35 kU/l a castaña, aguacate y plátano), únicamente mostró
63
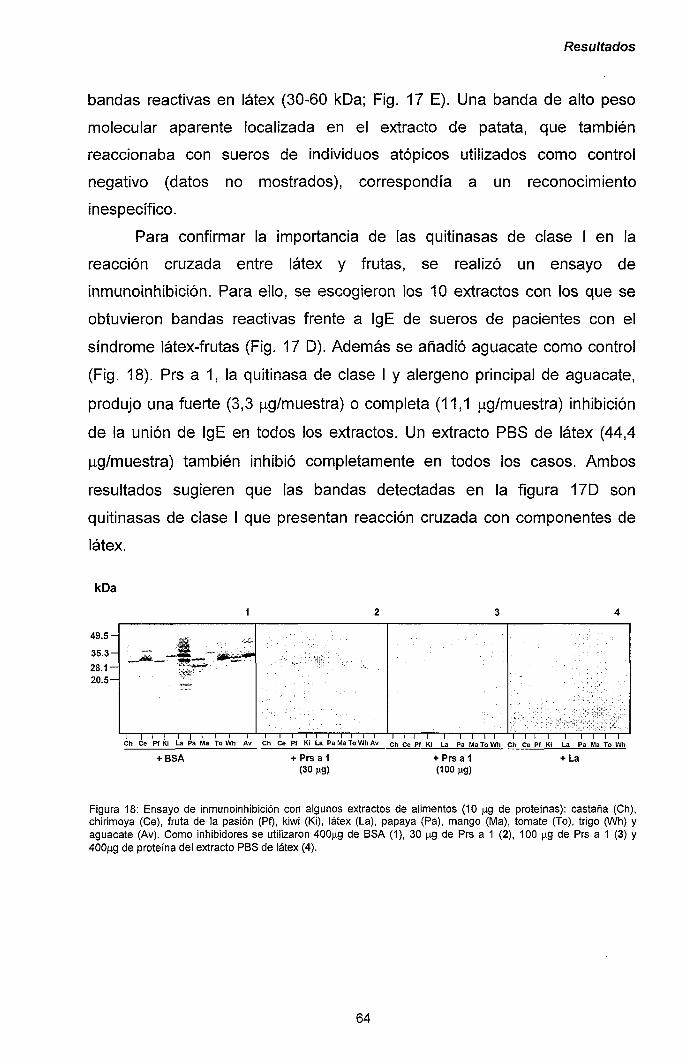
Resultados
bandas reactivas en látex (30-60 kDa; Fig. 17 E). Una banda de alto peso
molecular aparente localizada en el extracto de patata, que también
reaccionaba con sueros de individuos atópicos utilizados como control
negativo (datos no mostrados), correspondía a un reconocimiento
inespecífico.
Para confirmar la importancia de las quitinasas de clase I en la
reacción cruzada entre látex y frutas, se realizó un ensayo de
inmunoinhibición. Para ello, se escogieron los 10 extractos con los que se
obtuvieron bandas reactivas frente a IgE de sueros de pacientes con el
síndrome látex-frutas (Fig. 17 D). Además se añadió aguacate como control
(Fig. 18). Prs a 1, la quitinasa de clase I y alérgeno principal de aguacate,
produjo una fuerte (3,3 |j,g/muestra) o completa (11,1 )j,g/muestra) inhibición
de la unión de IgE en todos los extractos. Un extracto PBS de látex (44,4
pg/muestra) también inhibió completamente en todos los casos. Ambos
resultados sugieren que las bandas detectadas en la figura 17D son
quitinasas de clase I que presentan reacción cruzada con componentes de
látex.
kDa
49.5-35.3-28.1-20.5-
T—|—I—i—I—I—I—I—I—r^—1—I—I—n—I—i—1—n—n—r-i—i—i—i—i—n—n—n—i—i—i—i—i—r-Ch Ce Pf K¡ La Pa Ma To Wh Av Ch Ce Pf Kl U PaMaToWhAv ch Ce Pf Ki La Pa MaToWh Ch Ce Pf Ki La Pa Ma To Wh
+ BSA + Prs a 1 (30 lg)
+ Prsa1 (100 ng)
+ La
Figura 18: Ensayo de inmunoinhibición con algunos extractos de alinnentos (10 ng de proteínas): castaña (Ch), chirimoya (Ce), fruta de la pasión (Pf), kiwi (Ki), látex (La), papaya (Pa), mango (Ma), tomate (To), trigo (Wh) y aguacate (Av). Como inhibidores se utilizaron 400ng de BSA (1), 30 ^g de Prs a 1 (2), 100 \ig de Prs a 1 (3) y 400ng de proteína del extracto PBS de látex (4).
64
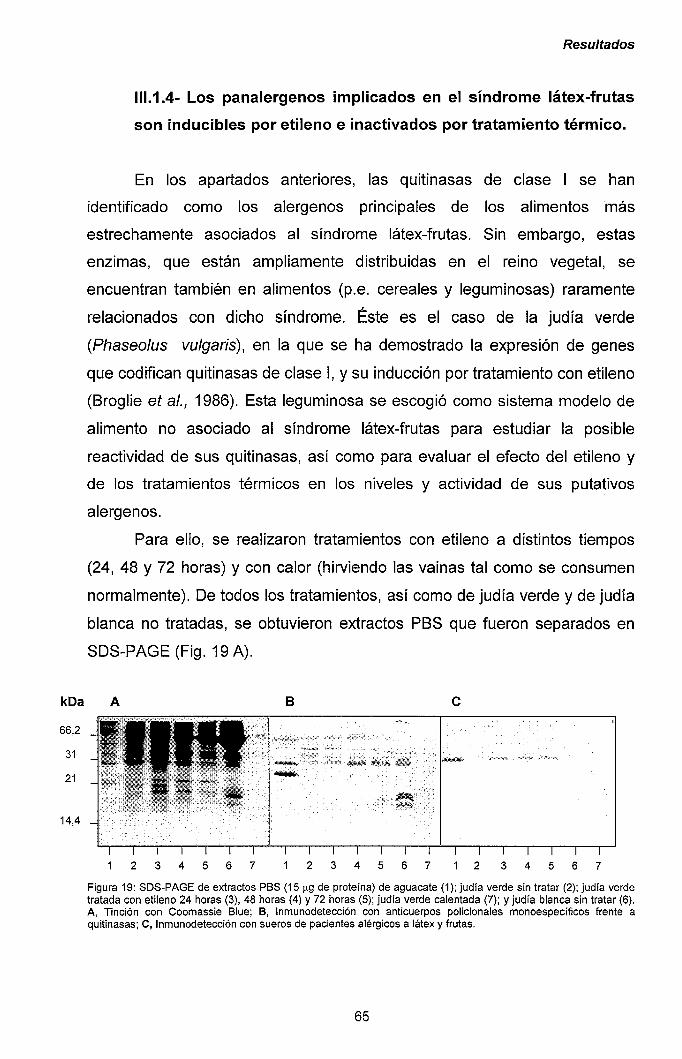
Resultados
ill.1.4- Los panalergenos implicados en el síndrome látex-frutas
son inducibles por etileno e inactivados por tratamiento térmico.
En los apartados anteriores, las quitinasas de clase I se han
Identificado como los alérgenos principales de los alimentos más
estrechamente asociados al síndrome látex-frutas. Sin embargo, estas
enzimas, que están ampliamente distribuidas en el reino vegetal, se
encuentran también en alimentos (p.e. cereales y leguminosas) raramente
relacionados con dicho síndrome. Éste es el caso de la judía verde
{Phaseolus vulgaris), en la que se ha demostrado la expresión de genes
que codifican quitinasas de clase I, y su inducción por tratamiento con etileno
(Broglie eí al., 1986). Esta leguminosa se escogió como sistema modelo de
alimento no asociado al síndrome látex-frutas para estudiar la posible
reactividad de sus quitinasas, así como para evaluar el efecto del etileno y
de los tratamientos térmicos en los niveles y actividad de sus putativos
alérgenos.
Para ello, se realizaron tratamientos con etileno a distintos tiempos
(24, 48 y 72 horas) y con calor (hirviendo las vainas tal como se consumen
normalmente). De todos los tratamientos, así como de judía verde y de judía
blanca no tratadas, se obtuvieron extractos PBS que fueron separados en
SDS-PAGE (Fig. 19 A).
kDa A B C
Figura 19: SDS-PAGE de extractos PBS (15 ng de proteína) de aguacate (1); judía verde sin tratar (2); judía verde tratada con etileno 24 horas (3), 48 tioras (4) y 72 horas (5); judia verde calentada (7); y judía blanca sin tratar (6). A, Tinción con Coomassie Blue; B, Inmunodetección con anticuerpos policlonales monoespecíficos frente a quitinasas; C, Inmunodetección con sueros de pacientes alérgicos a látex y frutas.
65

Resultados
En todas las muestras se detectó un modelo electroforétlco complejo,
con bandas entre 20 y 70 kDa. Los extractos de judías verdes que habían
sufrido un tratamiento térmico constituyeron una excepción. En este caso, no
se reveló ninguna banda, aun cuando la cantidad de proteína insertada en el
gel (cuantificada por el método de Bradford) fue la misma que en el resto de
las muestras (Fig. 19, muestra 7). Este mismo resultado se obtuvo al
analizar extractos de judías verdes comerciales enlatadas (datos no
mostrados).
La inmunodetección con anticuerpos policlonales monoespecíficos
frente a quitinasas mostró las dos bandas ya descritas de 25 (clase II) y 32
kDa (clase I; Prs a 1) en un extracto de aguacate utilizado como control (Fig.
19 B). Una putativa quitinasa de clase I de 32 kDa también fue reconocida
en todas las muestras de judía, excepto en la sometida a tratamiento
térmico. Se observó un incremento de la banda correspondiente en los
extractos de judías verdes tratadas con etileno (máximo en la muestra de 72
horas; Fig. 19, muestras 2-5).
Esos mismos extractos fueron utilizados para realizar una
inmunodetección con una mezcla de 8 sueros de pacientes alérgicos a látex
y frutas (pacientes número 1, 2, 4, 6, 12, 13, 20, y 22 de la Tabla 3 de
Material y Métodos). La banda de 32 kDa fue reactiva tanto en el extracto de
aguacate como en los de judía verde (Fig.19 C). Sin embargo, no se detectó
ninguna banda en la muestra de judía verde que había sido calentada, ni en
el extracto de judía blanca, aunque sí hubo una banda de 32 kDa reconocida
por el antisuero frente a quitinasas en este último caso.
Con el fin de purificar putativas quitinasas de clase I, los extractos de
judía verde sin tratar y tratadas 72 horas con etileno, así como el de las
semillas maduras (judía blanca), fueron separados por cromatografía de
intercambio catiónico (Fig. 20).
Proteínas de 32 kDa estaban incluidas en los dos picos señalados
(Fig. 21, muestras 3 y 5). La comparación de los perfiles correspondientes a
judía verde sin tratar y tratada (Fig. 20 A y B) indicó que el tratamiento con
etileno provocaba un fuerte incremento del componente de 32 kDa.
66
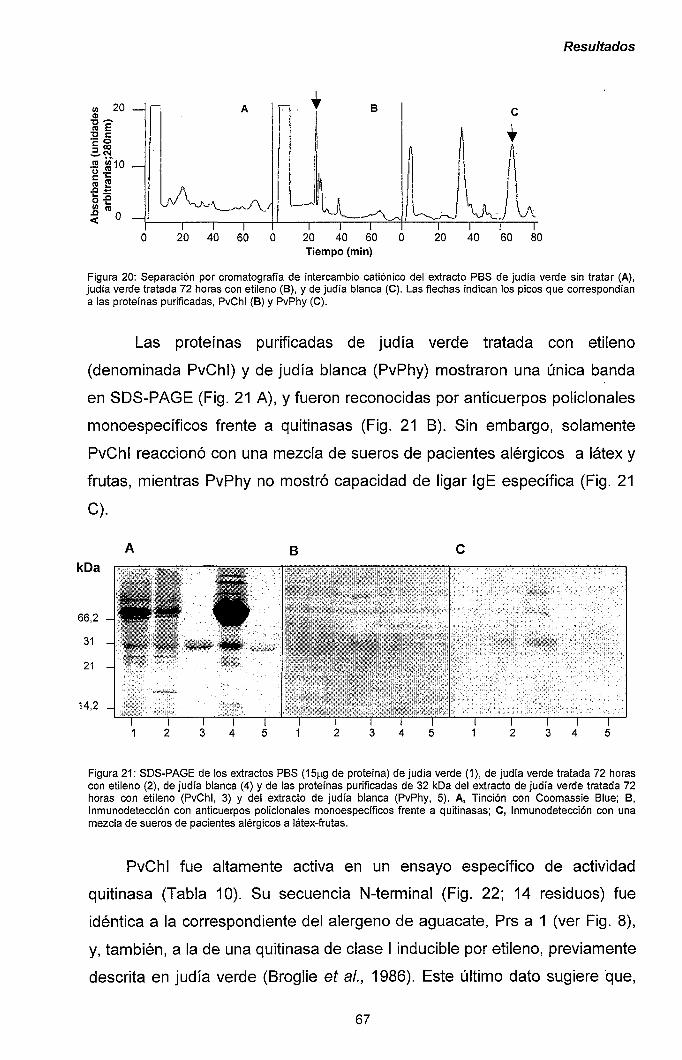
Resultados
20 ta
o •s? •n c 'E ° § 00
.2 ig10 ü .— II o 5 <n ra < O
U/' A •-x/vy^.
n 1 1 — 20 40 60 O
n 1 T" 20 40 60 Tiempo (min)
C
! u ^ \ii^/ L' M.
1 1 r 20 40 60 80
Figura 20: Separación por cromatografía de intercambio catiónico del extracto PBS de judía verde sin tratar (A), judía verde tratada 72 horas con etlleno (B), y de judia blanca (C). Las flechas indican los picos que correspondían a las proteínas purificadas, PvChl (B) y PvPhy (C).
Las proteínas purificadas de judía verde tratada con etileno
(denominada PvCiil) y de judía blanca (PvPliy) mostraron una única banda
en SDS-PAGE (Fig. 21 A), y fueron reconocidas por anticuerpos policlonales
monoespecíficos frente a quitinasas (Fig. 21 B). Sin embargo, solamente
PvCiil reaccionó con una mezcla de sueros de pacientes alérgicos a látex y
frutas, mientras PvPhy no mostró capacidad de ligar IgE específica (Fig. 21
C).
B kDa
Figura 21: SDS-PAGE de los extractos PBS (15ng de proteína) de judía verde (1), de judía verde tratada 72 horas con etileno (2), de judía blanca (4) y de las proteínas purificadas de 32 l<Da del extracto de judía verde tratada 72 horas con etileno (PvChl, 3) y del extracto de judía blanca (PvPhy, 5). A, Tinción con Coomassie Blue; B, Inmunodetección con anticuerpos policlonales monoespecíficos frente a quitinasas; C, Inmunodetección con una mezcla de sueros de pacientes alérgicos a látex-frutas.
PvChl fue altamente activa en un ensayo específico de actividad
qultinasa (Tabla 10). Su secuencia N-terminal (Fig. 22; 14 residuos) fue
idéntica a la correspondiente del alérgeno de aguacate, Prs a 1 (ver Fig. 8),
y, también, a la de una quitinasa de clase I inducible por etileno, previamente
descrita en judía verde (Broglie eí al., 1986). Este último dato sugiere que.
67

Resultados
probablemente, PvChl y la quitinasa identificada sean la misma proteína. Si
éste es el caso, su relación con Prs a 1 se ve sustentada por el alto grado de
identidad de secuencia entre este alérgeno y la quitinasa de judía (74%),
basada en las estructuras primarias deducidas de los correspondientes
clones de ADNc (Broglie et al., 1986; Sowka eí al., 1998a).
Tabla 10: Actividad quitinasa de ios extractos PBS de judía verde sin tratar, judía verde tratada 72 horas con etileno, judía blanca, y de las proteínas purificadas PvChI y PvPhy.
Muestra ng/ensayo Absorbancia x 10"^(550nnn)
14 5±5
622±2
118±2
636±4
207±8
25±2
GbE, judía verde; GbE72, judía verde tratada 72 horas con etileno; WbE, judía blanca. Los valores son medias (n=3)± ES .
La relevancia de PvCiní como potencial alérgeno de judía, así como su
reactividad cruzada con Prs a 1, se confirmó por ensayos de
inmunoinhibición (Fig. 23). En ambos casos, las proteínas purificadas
inhibieron completamente la unión de IgE por la banda del extracto
relacionado (PvChl/ aguacate; Prs a 1/judía tratada con etileno).
GbE
GbE72
WbE
P v C h l
PvPhy
500
500
500
100
10
500
Proteína
P v C h l
PvPhy
PHA-L
Secuencia N-terminal
EQCGRQAGGALCPG
SNDIYFNFQRFXET
SNDIYFNFQRFNET
Figura 22: Secuencia de aminoácidos N-terminal de las proteínas de 32 kDa purificadas de judía verde (PvChl), y de judía blanca (PvPhy). Esta última se ha comparado con la secuencia de la fltohemaglutinina de judía PHA-L, previamente descrita (Hoffman y Donaldson, 1985).
68
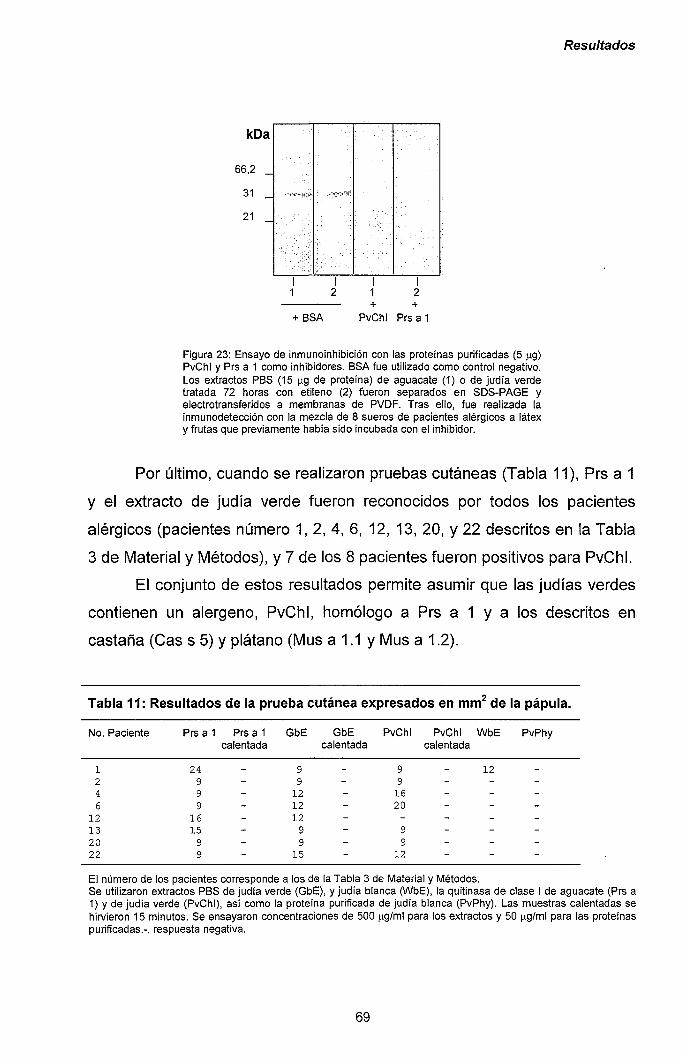
Resultados
kDa
66,2
31
21
+ BSA
I I 1 2 + +
PvChl Prs a 1
Figura 23: Ensayo de inmunoinhibición con las proteínas purificadas (5 ng) PvChl y Prs a 1 como inhibidores. BSA fue utilizado como control negativo. Los extractos PBS (15 ng de proteína) de aguacate (1) o de judía verde tratada 72 horas con etileno (2) fueron separados en SDS-PAGE y electrotransferidos a membranas de PVDF. Tras ello, fue realizada la inmunodetección con la mezcla de 8 sueros de pacientes alérgicos a látex y frutas que previamente había sido incubada con el inhibidor.
Por último, cuando se realizaron pruebas cutáneas (Tabla 11), Prs a 1
y el extracto de judía verde fueron reconocidos por todos los pacientes
alérgicos (pacientes número 1, 2, 4, 6, 12, 13, 20, y 22 descritos en la Tabla
3 de Material y Métodos), y 7 de los 8 pacientes fueron positivos para PvChl.
El conjunto de estos resultados permite asumir que las judías verdes
contienen un alérgeno, PvChl, homólogo a Prs a 1 y a los descritos en
castaña (Cas s 5) y plátano (Mus a 1.1 y Mus a 1.2).
Tabla 11: Resultados de la prueba cutánea expresados en mm^ de la pápula.
No. Paciente Prs a l Prs a 1 calentada
GbE GbE calentada
PvChl PvChl WbE calentada
PvPhy
1 2 4 6
12 13 20 22
24 9 9 9
16 15 9 9
--------
9 9 12 12 12 9 9
15
--------
9 9 16 20
-9 9
12
12
El número de ios pacientes corresponde a los de la Tabla 3 de Material y Métodos. Se utilizaron extractos PBS de judía verde (GbE), y Judía blanca (WbE), la quitinasa de clase I de aguacate (Prs a 1) y de judía verde (PvChl), así como la proteína purificada de judía blanca (PvPhy). Las muestras calentadas se hirvieron 15 minutos. Se ensayaron concentraciones de 500 (j.g/ml para los extractos y 50 ng/ml para las proteínas purificadas.-, respuesta negativa.
69

Resultados
Por lo que se refiere a PvPhy, la proteína de 32 kDa purificada de
judía blanca, además de no ser reconocida por la mezcla de sueros de
pacientes alérgicos (Fig. 21 C), no mostró reactividad en ensayos in vivo
(Tabla 11), ni actividad quitinasa a niveles significativos (Tabla 10). Todos
estos datos son coherentes con la falta de reactividad del extracto de judía
blanca frente a sueros de pacientes alérgicos (Fig. 19, muestra 6). La
determinación de la secuencia N-terminal de PvPhy (Fig. 22) permitió su
identificación como PHA-L, una fitohemaglutinina de judía previamente
descrita (Hoffman y Donaldson, 1985). Su reconocimiento por anticuerpos
policlonales monoespecíficos frente a quitinasas de plantas puede explicarse
por la homología entre PHA-L y el dominio catalítico de las mismas (como
las de castaña y cebada).
Para estudiar el efecto del calor en las propiedades alergénicas de las
quitinasas de clase I se utilizaron muestras sin tratar y tratadas térmicamente
(100°C durante 15 minutos) de aguacate y judía verde. El tratamiento
provocó una pérdida total de la capacidad de unir IgE por la banda de 32
kDa (Prs a 1), en preparaciones de aguacate enriquecidas en quitinasas
(Fig. 24, muestras AvF). Estos resultados se confirmaron analizando los
alérgenos purificados Pv Chl y Prs a 1. La muestra de PvChl tratada fue
indetectable después de SDS-PAGE y no fue reconocida ni por anticuerpos
policlonales monoespecíficos frente a quitinasas, ni por una mezcla de
sueros de pacientes alérgicos (Fig. 24, muestras PvChI). Idénticos
resultados se obtuvieron al analizar Prs a 1. Además, el tratamiento térmico
conllevaba la pérdida de actividad enzimática de ambos alérgenos, así como
cambios en su perfil cromatográfico de HPLC (datos no mostrados). Dos
proteínas no relacionadas con las quitinasas, albúmina de suero bovino y un
alérgeno de melocotón (BSA y LTP; Fig. 24 D), no se vieron afectadas, al
menos en su movilidad y detección después de SDS-PAGE, por el
tratamiento térmico.
La inactivación por calor de los alérgenos Prs a 1 y PvChI se vio
plenamente confirmada al realizar pruebas cutáneas in vivo (Tabla 11). En
70
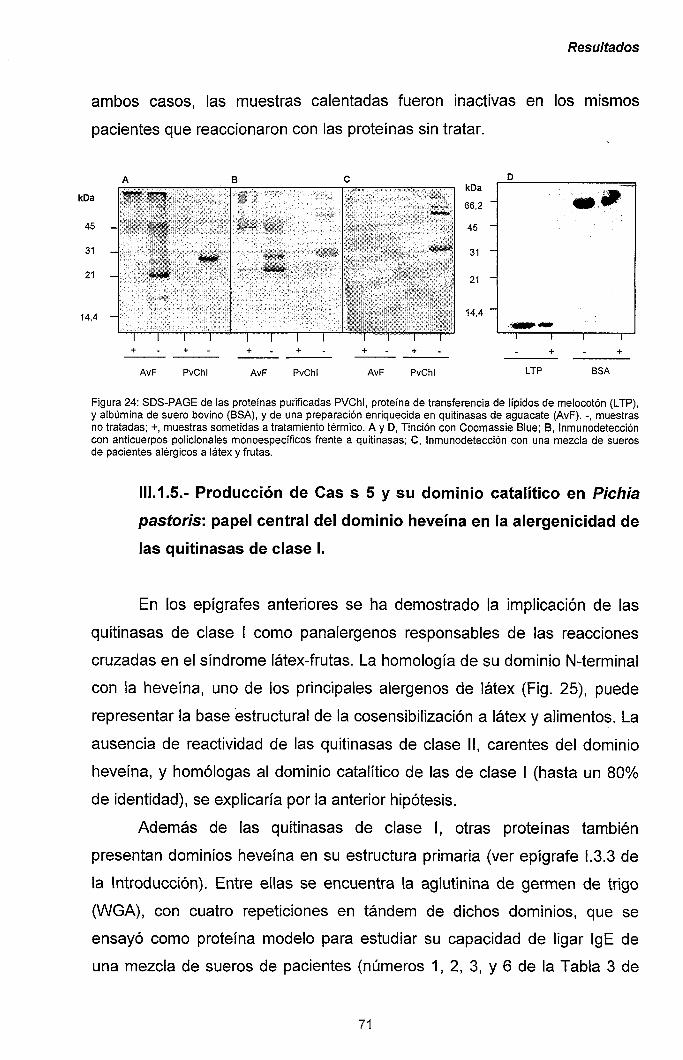
Resultados
ambos casos, las muestras calentadas fueron Inactivas en los mismos
pacientes que reaccionaron con las proteínas sin tratar.
kDa
45
31
21
14,4
V^^^ W^^
'•í
^'^
<i
1 1
mm
1
f" •
i^^is ;;•'•;•-
l i l i
—. ii«í¿
••¡m»^
l i l i
AvF PvChl AvF PvChI AvF PvChI LTP BSA
Figura 24: SDS-PAGE de las proteínas purificadas PVChl, proteína de transferencia de lípidos de melocotón (LTP), y albúmina de suero bovino (BSA), y de una preparación enriquecida en quitinasas de aguacate (AvF). -, muestras no tratadas; +, muestras sometidas a tratamiento térmico. A y D, Tinción con Coomassie Blue; B, Inmunodetección con anticuerpos policlonales monoespecíficos frente a quitinasas; C, Inmunodetección con una mezcla de sueros de pacientes alérgicos a látex y frutas.
III.1.5.- Producción de Cas s 5 y su dominio catalítico en Pichia
pastorís: papel central del dominio heveína en la alergenicidad de
las quitinasas de clase I.
En los epígrafes anteriores se ha demostrado la implicación de las
quitinasas de clase I como panalergenos responsables de las reacciones
cruzadas en el síndrome látex-frutas. La homología de su dominio N-terminal
con la heveína, uno de los principales alérgenos de látex (Fig. 25), puede
representar la base estructural de la cosensibilización a látex y alimentos. La
ausencia de reactividad de las quitinasas de clase II, carentes del dominio
heveína, y homologas al dominio catalítico de las de clase I (hasta un 80%
de identidad), se explicaría por la anterior hipótesis.
Además de las quitinasas de clase I, otras proteínas también
presentan dominios heveína en su estructura primaria (ver epígrafe 1.3.3 de
la Introducción). Entre ellas se encuentra la aglutinina de germen de trigo
(WGA), con cuatro repeticiones en tándem de dichos dominios, que se
ensayó como proteína modelo para estudiar su capacidad de ligar IgE de
una mezcla de sueros de pacientes (números 1, 2, 3, y 6 de la Tabla 3 de
71
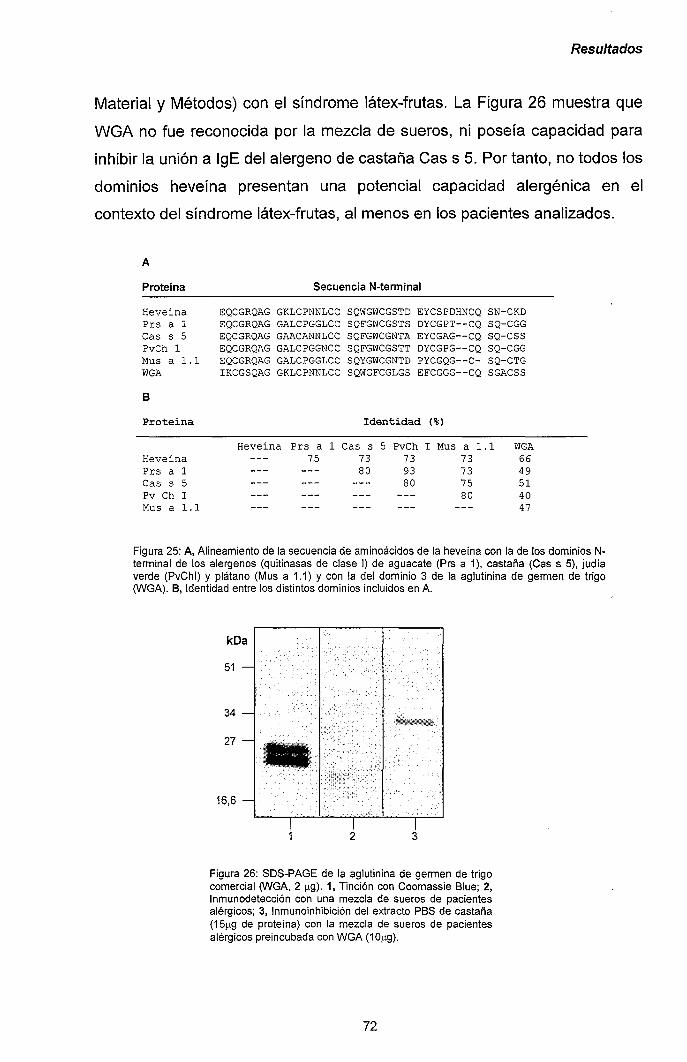
Resultados
Material y Métodos) con el síndrome látex-frutas. La Figura 26 muestra que
WGA no fue reconocida por la mezcla de sueros, ni poseía capacidad para
Inhibir la unión a IgE del alérgeno de castaña Cas s 5. Por tanto, no todos los
dominios heveína presentan una potencial capacidad alergénica en el
contexto del síndrome látex-frutas, al menos en los pacientes analizados.
A
Proteína
Heve ina P r s a 1 Cas s 5 Pv Ch I Mus a 1.1
Secuencia N-terminal
Heveína Prs a 1 Cas s 5 PvCh 1 Mus a 1.1 WGA
B
Proteína
EQCGRQñG EQCGRQAG EQCGRQAG EQCGRQAG EQCGRQAG IKCGSQAG
GKLCPNNLCC GALCPGGLCC GAACANNLCC GALCPGGNCC GALCPGGLCC GKLCPNNLCC
SQWGWCGSTD EYCSPDHNCQ SQFGWCGSTS DYCGPT—CQ SQFGWCGNTA EYCGAG—CQ SQFGWCGSTT DYCGPG—CQ SQYGWCGNTD PYCGQG—C-SQWGFCGLGS EFCGGG—CQ
Identidad (%)
SN-CKD SQ-CGG SQ-CSS SQ-CGG SQ-CTG SGACSS
Heveína Prs a 1 Cas s 75 73
80
PvCh I Mus a 1.1 73 73 93 73 80 75
80
WGA 66 49 51 40 47
Figura 25: A, Alineamiento de la secuencia de aminoácidos de la heveína con la de los dominios N-terminal de los alérgenos (quitinasas de clase I) de aguacate (Prs a 1), castaña (Cas s 5), judia verde (PvChl) y plátano (Mus a 1.1) y con la del dominio 3 de la aglutinina de germen de trigo (WGA). B, Identidad entre los distintos dominios incluidos en A.
kDa
51 -
34 —I
27
16,6
Figura 26: SDS-PAGE de la aglutinina de gennen de trigo comercial (WGA, 2 |.ig). 1, Tinción con Coomassie Blue; 2, Inmunodetección con una mezcla de sueros de pacientes alérgicos; 3, Inmunoinhibición del extracto PBS de castaña (15(ig de proteina) con la mezcla de sueros de pacientes alérgicos preincubada con WGA (10(xg).
72

Resultados
47 Cas S 5 EQCGRQAGGA ACANNLCCSQ FGWCGNTAEY CGAGCQSQCS SPTTTTSSPT Prs a 1 EQCGRQAGGA LCPGGLCCSQ FGWCGSTSDY CGPTCQSQCG GVTPSP Mus a 1.1 EQCGRQAGGA LCPGGLCCSQ YGWCGNTDPY CGQGC-SQCT GSTPSPSTPS PvChl EQCGRQAGGA LCPGGNCCSQ FGWCGSTTDY CGPGCQSQCG GPSPAPT
90 Cas s 5 ASSGGGGDVG SLISASLFDQ MLKYRNDPRC KSNGFYTYNA FIAAARSFNG Prs a 1 GGGVA SLISQSVFNQ MLKHRNDAAC QAKGFYTYNA FIAAANSFNG Mus a 1.1 GGGSVG SIISSSLF-E MLKHRNDAAC PGKGFYTYNA FIAAANSFSG PvChl DLS ALISRSTFDQ MLKHRNDGAC PAKGFYTYDA FIAAAKAYPS
139 Cas S 5 FGTTGDVTTR KRELAAFLAQ TSHETTGGWA TAPDGPYAWG YCFVM-ENNK Prs a 1 FASVGDTATR KREIAAFLAQ TSHETTGGWA TAPDGPYAWG YCFLKEQGNP Mus a 1.1 FGTTGD-ATR E PvChl FGNTGDTATR KREIAAFLGQ TSHETTGGWA TAPDGPYAWG YCFVR-ERNP
189 Cas S 5 QTYCTSKS-W PCVFGKQYYG RGPIQLTHNY NYGQAGKAIG ADLINNPDLV Prs a 1 PDYCVPTAQW PCAPGKKYYG RGPIQISYNY NYGPAGRAIG YDLINNPDAV PvChl STYCSATPQP PCAPGQQYYG RGPIQISWNY NYGQCGRAIG VDLLNKPDLV
239 Cas S 5 ATNPTISFKT AIWFWMTPQA NKPSSHDVII GNWRPSAADT SAGRVPSYGV Prs a 1 ATDPVISFKT ALWFWMTPQS PKPSCHNVIT GRWTPSAADR AAGRLPGYGV PvChl ATDSVISFKS ALWFWMTAQS PKPSSHDVIT SRWTPSSADV AARRLPGYGT
289 Cas S 5 ITNIINGGLE CGHGSDDRVA NRIGFYKRYC DTLGVSYGNN LDCYNQKPFA Prs a 1 VTNIINGGLE CGRGQDSRVQ DRIGFFKRYC DLLGVGYGNN LDCYSQTPFG PvChl ITNIINGGIE CGKGFNDKVA DRIGFYKRYC DLLGVSYGSN LDCYNQRSFG
301 Prs a 1 PvChl
VSTNPLAASS — NSLLLSDLVT SQ
B
Protexna Identidad (%)
Cas s 5 Prs a 1 Mus a 1.1 PvChl Cas s 5 - 71 65,4 67,4 Prs a 1 - - 72,5 75,8 Mus al.l - - - 64,2
Figura 27: A, Alineamiento de la secuencia de aminoácidos de las quitinasas alergénicas Cas s 5 (No. Accesión U48687), Prs a 1 (No. Accesión Z78202), Mus a 1.1 (Clon parcial; No. Accesión AF001524) y • PvChl (No. Accesión P06215). B, Porcentaje de identidad entre las secuencias.
La comparación de las secuencias completas de aminoácidos de las
cuatro quitinasas alergénicas identificadas en este trabajo (Fig. 27) reveló la
alta similitud que hay entre las mismas, no sólo en sus dominios heveína,
sino también en los catalíticos.
La disponibilidad de un clon ADNc de Cas s 5 (cedido por los Drs. C.
Aragoncillo y L. Gómez, E.T.S. Ingenieros de Montes, UPM), condicionó la
elección de esta quitinasa como modelo para estudiar el papel de los
73

Resultados
dominios lieveína y catalítico en la alergenicidad. Partiendo del clon
completo, se eliminó la secuencia que codificaba el péptido señal, y
utilizando cebadores específicos se subclonaron los fragmentos
correspondientes a la proteína madura completa (rCas s 5), al dominio
catalítico (rCAT) y al dominio heveína (ver Fig. 4 de Material y Métodos).
Cada uno de estos subclones se insertó en el vector de expresión
extracelular pPIC 9 (en fase con un péptido señal específico; Fig.3 del
Material y Métodos), con el que se transformó la cepa GS115 de la levadura
Pichia pastoris.
Las células transformadas con cada una de las tres construcciones, y
con el vector sin inserto como control, se incubaron hasta alcanzar una
ODeoo de 2-2,5 (3 días), y se indujeron con metanol (4 días). Las proteínas
recombinantes se purificaron a partir de los sobrenadantes de los
correspondientes cultivos. rCas s 5 y el producto truncado sin el dominio
heveína, rCAT, se obtuvieron en cantidad apreciable (aproximadamente
30mg/l de cultivo) siguiendo este protocolo. Sin embargo, el dominio heveína
recombinante presentó serias dificultades en su expresión y purificación.
Probablemente debido a su bajo tamaño molecular (4,2 kDa), su detección
en geles resulta difícil, aun utilizando tinción de plata y cantidades de
proteína de hasta 10 [ig. Además, los picos de HPLC donde se detectó el
péptido recombinante, mostraron una fuerte contaminación con putativos
fragmentos (de 400 a 900 aminoácidos, determinados por MALDI) de
proteínas de pared de P. pastoris (con hidroxiprolina en posición 3 de su
secuencia), al secuenciar su extremo N-terminal. Considerando estos
resultados, el dominio heveína recombinante no se incluyó en los ensayos
que se describen a continuación.
Los sobrenadantes de los cultivos que expresaban rCas s 5 y rCAT
mostraron una banda mayoritaria de 35 y 25 kDa, respectivamente (Fig. 28
A). Las proteínas recombinantes se purificaron por filtración molecular (Fig.
28, B y C) usando una columna de Superdex 75. Las fracciones obtenidas
se sometieron a un paso final de repurificación por cromatografía de
intercambio catiónico, donde se obtuvo un único pico en ambos casos.
74
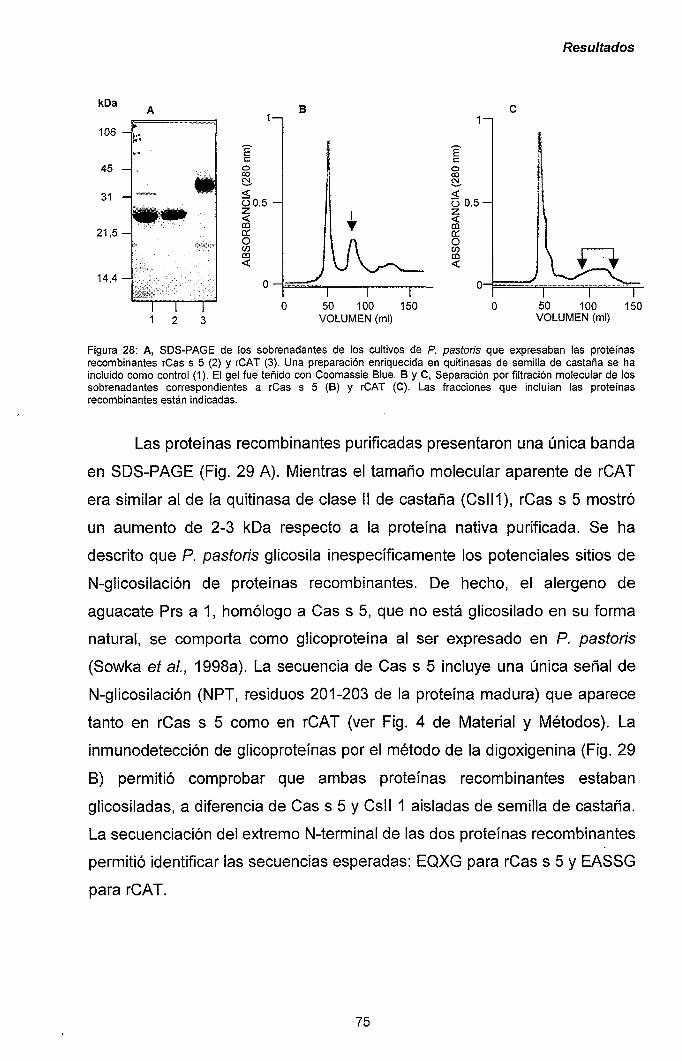
Resultados
kDa
106 -
45 -
31 -
21,5 -
1 4 , 4 -
A
^ ™ ^
j | | j |k o oo
< ü0,5 z < m D; O w CQ <
O -
I
1 ^ r 1 2 3
50 100 150 VOLUMEN (mi)
50 100 150 VOLUMEN (mi)
Figura 28: A, SDS-PAGE de los sobrenadantes de los cultivos de P. pastoris que expresaban las proteínas recombinantes rOas s 5 (2) y rCAT (3). Una preparación enriquecida en quitinasas de semilla de castaña se ha incluido como control (1). El gel fue teñido con Coomassie Blue. B y C, Separación por filtración molecular de los sobrenadantes correspondientes a rCas s 5 (B) y rCAT (C). Las fracciones que incluían las proteínas recombinantes están indicadas.
Las proteínas recombinantes purificadas presentaron una única banda
en SDS-PAGE (Fig. 29 A). IVIientras el tamaño molecular aparente de rCAT
era similar al de la quitinasa de clase II de castaña (CslH), rCas s 5 mostró
un aumento de 2-3 kDa respecto a la proteína nativa purificada. Se ha
descrito que P. pastoris glicosila inespecíficamente los potenciales sitios de
N-glicosilación de proteínas recombinantes. De hecho, el alérgeno de
aguacate Prs a 1, homólogo a Cas s 5, que no está glicosilado en su forma
natural, se comporta como glicoproteína al ser expresado en P. pastoris
(Sowka et al., 1998a). La secuencia de Cas s 5 incluye una única señal de
N-glicosilación (NPT, residuos 201-203 de la proteína madura) que aparece
tanto en rCas s 5 como en rCAT (ver Fig. 4 de Material y Métodos). La
inmunodetección de glicoproteínas por el método de la digoxigenina (Fig. 29
B) permitió comprobar que ambas proteínas recombinantes estaban
glicosiladas, a diferencia de Cas s 5 y Csll 1 aisladas de semilla de castaña.
La secuenciación del extremo N-terminal de las dos proteínas recombinantes
permitió identificar las secuencias esperadas: EQXG para rCas s 5 y EASSG
para rCAT.
75
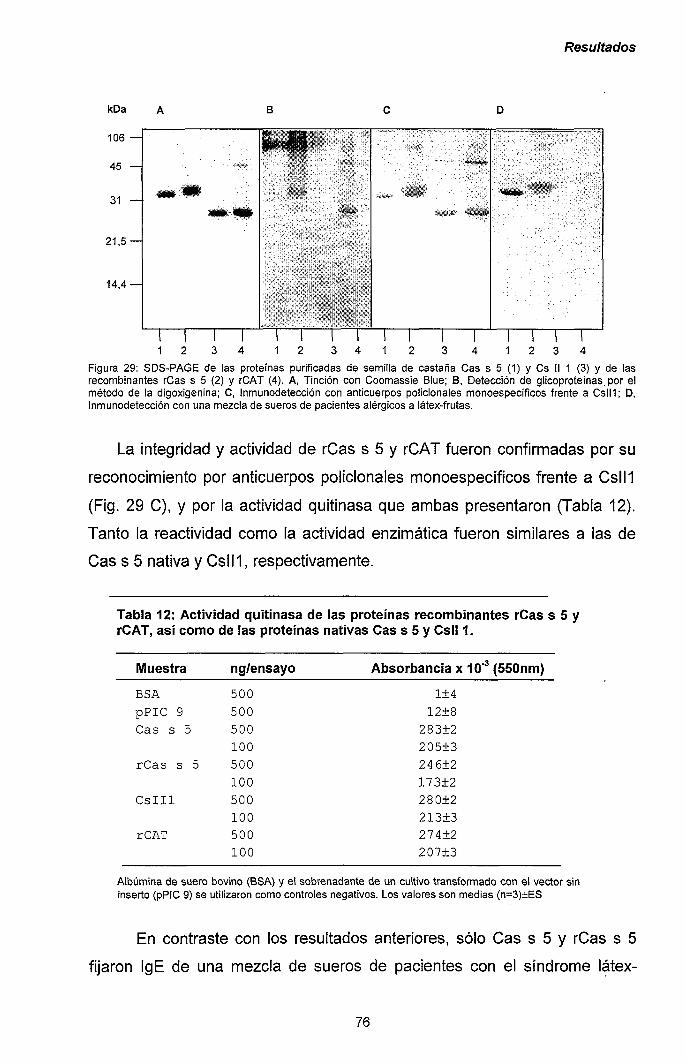
Resultados
kDa
106 —
45
31 —I
21,5-
14,4-
•¿»'
'^^¿^¿^
1 1 1 1 Figura 29: SDS-PAGE de las proteínas purificadas de semilla de castaña Cas s 5 (1) y Cs II 1 (3) y de las recombinantes rCas s 5 (2) y rCAT (4). A, Tinción con Coomassie Blue; B, Detección de glicoproteínas.por el método de la dlgoxigenina; C, Inmunodetección con anticuerpos policlonales monoespecíflcos frente a Csll l ; D, Inmunodetección con una mezcla de sueros de pacientes alérgicos a látex-frutas.
La integridad y actividad de rCas s 5 y rCAT fueron confirmadas por su
reconocimiento por anticuerpos policlonales monoespecíficos frente a CslH
(Fig. 29 C), y por la actividad quitinasa que ambas presentaron (Tabla 12).
Tanto la reactividad como la actividad enzimática fueron similares a las de
Cas s 5 nativa y CslH, respectivamente.
Tabla 12: Actividad quitinasa de las proteínas recombinantes rCas s 5 y rCAT, así como de las proteínas nativas Cas s 5 y Csll 1.
Muestra ng/ensayo Absorbancia x 10 (550nm)
BSA pPIC 9
Cas s 5
rCas s 5
CsIIl
rCAT
500 500
500
100
500
100
500
100 500
100
1±4
12±8
283±2
205±3
24 6±2
173+2
280±2
213±3
274±2
207±3
Albúmina de suero bovino (BSA) y el sobrenadante de un cultivo transformado con el vector sin inserto (pPIC 9) se utilizaron como controles negativos. Los valores son medias (n=3)+ES
En contraste con los resultados anteriores, sólo Cas s 5 y rCas s 5
fijaron IgE de una mezcla de sueros de pacientes con el síndrome látex-
76

Resultados
frutas (pacientes 1, 2, 4, 6, 12, 13 y 20 de la tabla 3 de Material y Métodos).
La proteína recombinante sin el dominio heveína, rCAT, así como Cslíl, no
mostraron reactividad. Estos datos se confirmaron plenamente por ensayos
de inmunoinhibición (Fig. 30). Mientras Cas s 5 o su recombinante
provocaron una inhibición completa, la preincubación de la mezcla de sueros
con rCAT o Cslll no ejerció afecto alguno. La heveína de látex purificada
produjo una fuerte disminución de la unión de IgE a Cas s 5 y rCas s 5.
Puede, por tanto, concluirse que la deleción del dominio heveína de Cas s 5
conlleva una pérdida completa de sus propiedades alergénicas (al menos, in
vitro).
1 1 , ""
•
1 :
í 1 ; E
^ : ^ } l i
\ \ í : ' 1 •
;' !
i
< !
i (
, • . j ;
i ! í
1
•
! •"
j ; \ - ^
1
1-1 2 3 4 5 6 1 2 3 4 5 6
CassS rCas s 5
Figura 30: Ensayo de inmunoinhibición utilizando las proteínas purificadas Cas s 5 y rCas s 5 (Ijag). La mezcla de sueros de pacientes alérgicos fue preincubada con 6 ng de albúmina de suero bovino (1), Cas s 5 (2), Csll I (3), rCas s 5 (4), rCAT (5) y heveína de látex (6).
III.2.- LTPs COMO POTENCIALES PANALERGENOS VEGETALES.
Las proteínas de transporte de lípidos (LTPs) han sido identificadas
como alérgenos principales de manzana y melocotón en la alergia a frutas
de rosáceas no relacionada con polinosis a Bet v 1 (ver epígrafe 1.4.2 de la
Introducción; Sánchez-Monge eta\., 1999; Pastorello eí a/,, 1999a y b). Esta
familia de proteínas está ampliamente distribuida en alimentos vegetales y
77

Resultados
pólenes, lo que sugiere que podría estar implicada en otros tipos de alergia,
y representar un nuevo tipo de panalergenos vegetales.
III.2.1.- Purificación de las LTPs de polen de Artemisia y semilla
de castaño: reactividad cruzada con los alérgenos de manzana y
melocotón.
Algunos de los pacientes sensibilizados a frutas de rosáceas
presentan, en ensayos in vitro, reactividad cruzada con polen de Artemisia,
pero no con polen de abedul, lo que podría indicar, al menos en España, que
existe una cosensibilización al polen de esta planta herbácea y a las citadas
frutas.
El castaño es una especie de la familia Fagaceae, muy distanciada
filogenéticamente de las rosáceas. Sin embargo, se observó que algunos
pacientes alérgicos a manzana y melocotón, también presentaban
sensibilización a castaña, pero no a polen de ésta ni al de otros miembros de
su familia. Esto descartaba, en principio, la implicación de homólogos de Bet
V 1, agente principal de la cosensibilización a polen de abedul y frutas en el
centro y norte de Europa.
Con el fin de estudiar la posible implicación de las LTPs en las
reacciones cruzadas entre frutas de rosáceas, polen de artemisa y semilla
de castaña, se aislaron los miembros mayoritarios de esta familia de
proteínas en cada uno de los cuatro materiales vegetales.
Los alérgenos (LTPs) de manzana y melocotón fueron aislados a
partir de extractos de LiCI de la piel de estas frutas, previamente extraídos
con tampón Tris-HCI. Los extractos se fraccionaron por cromatografía de
intercambio catiónico (cart:uchos Sep-Pak Accell Plus CM, Waters, Irianda;
resultados no mostrados).
El posible polipéptido homólogo de polen de artemisa fue purificado a
partir del extracto Tris-HCI fraccionado por RP-HPLC en una columna Vydac
C4 (Fig. 31 A). Las fracciones que contenían la putativa LTP, junto con otras
proteínas, fueron separadas en un segundo sistema de RP-HPLC utilizando
78
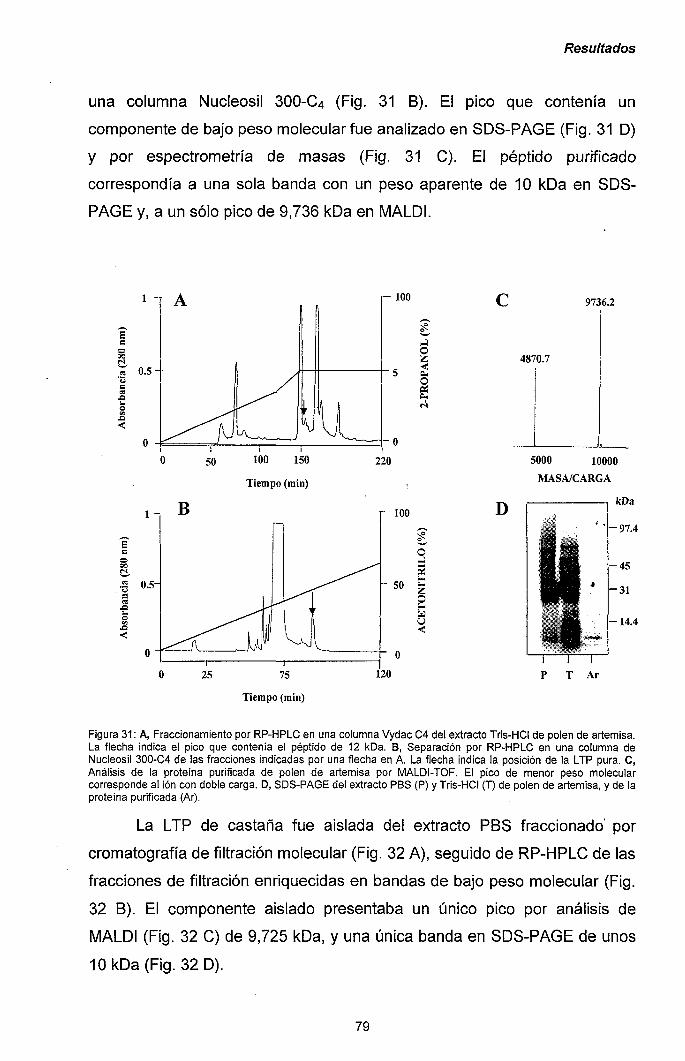
Resultados
una columna Nucleosil 3OO-C4 (Fig. 31 B). El pico que contenía un
componente de bajo peso molecular fue analizado en SDS-PAGE (Fig. 31 D)
y por espectrometría de masas (Fig. 31 C). El péptido purificado
correspondía a una sola banda con un peso aparente de 10 kDa en SDS-
PAGE y, a un sólo pico de 9,736 kDa en MALDI.
9736.2
4870.7
Tiempo (min)
5000 10000
MASA/CARGA
.2 0.5-
£1
<
B
u 1
0 25 i
75
1 12
r 100
o 2
50 I O H u <
D kDa
-97.4
14.4
Tiempo (min)
Figura 31: A, Fraccionamiento por RP-HPLC en una columna Vydac 04 del extracto Tris-HCi de polen de artemisa. La flecha indica el pico que contenía el péptido de 12 kDa. B, Separación por RP-HPLC en una columna de Nucleosil 300-04 de las fracciones indicadas por una flecha en A. La flecha indica la posición de la LTP pura. C, Análisis de la proteina purificada de polen de artemisa por MALDI-TOF. El pico de menor peso molecular corresponde al ion con doble carga. D, SDS-PAGE del extracto PBS (P) y Tris-HC! (T) de polen de artemisa, y de la proteína purificada (Ar).
La LTP de castaña fue aislada del extracto PBS fraccionado' por
cromatografía de filtración molecular (Fig. 32 A), seguido de RP-HPLC de las
fracciones de filtración enriquecidas en bandas de bajo peso molecular (Fig.
32 B). El componente aislado presentaba un único pico por análisis de
MALDI (Fig. 32 C) de 9,725 kDa, y una única banda en SDS-PAGE de unos
10 kDa (Fig. 32 D).
79
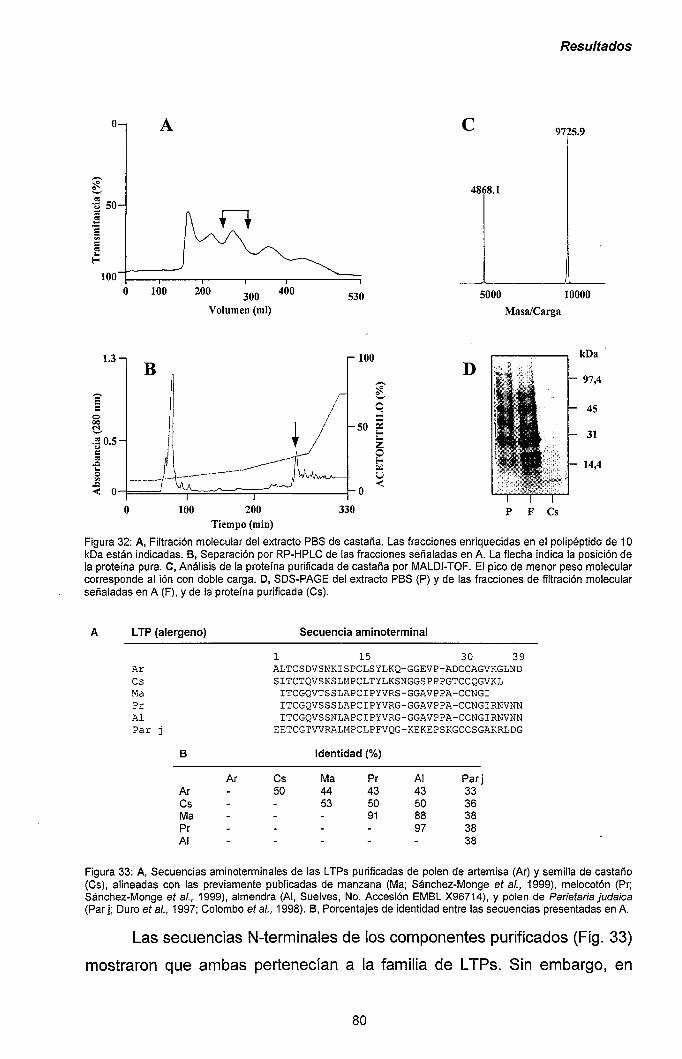
Resultados
9725.9
4868.1
300 Volumen (mi)
5000 10000 Masa/Carga
1.3 n
.2 0.5
rlOO D
-50
O ¡¿ e¿ H Z O H tá U <
330 P F Cs Tiempo (min)
Figura 32: A, Filtración molecular del extracto PBS de castaña. Las fracciones enriquecidas en el polipéptido de 10 kDa están indicadas. B, Separación por RP-HPLC de las fracciones señaladas en A. La flecha indica la posición de la proteína pura. C, Análisis de la proteína purificada de castaña por MALDI-TOF. El pico de menor peso molecular corresponde al ion con doble carga. D, SDS-PAGE del extracto PBS (P) y de las fracciones de filtración molecular señaladas en A (F), y de la proteína purificada (Cs).
LTP (alérgeno)
Ar Cs Ma Pr Al Par j
B
Ar Ar Cs Ma Pr Al
Secuencia aminoterr
1 15 ALTCSDVSNKISPCLSYLKQ-
nina!
30 39 -GGEVP-ADCCAGVKGLND
SITCTQVSKSLMPCLTYLKSNGGSPPPGTCCQGVKL ITCGQVTSSLAPCIPYVRS-ITCGQVSSSLAPCIPYVRG-ITCGQVSSNLAPCIPYVRG-
EETCGTVVRALMPCLPFVQG-
Identidad (%)
Cs Ma Pr 50 44 43
53 50 91
.
.
-GGAVPPA-CCMGI -GGAVPPA-CCNGIRNVNN -GGAVPPA-CCNGIRNVNN -KEKEPSKGCCSGAKRLDG
Al Parj 43 33 50 36 88 38 97 38
38
Figura 33: A, Secuencias aminoterminales de las LTPs purificadas de polen de artemisa (Ar) y semilla de castaño (Cs), alineadas con las previamente publicadas de manzana (Ma; Sánchez-Monge ef al., 1999), melocotón (Pr; Sánchez-Monge ef al., 1999), almendra (Al, Suelves, No. Accesión EMBL X96714), y polen de Paríetaria judaica (Parj; Duro etal., 1997; Colombo etal., 1998). B, Porcentajes de identidad entre las secuencias presentadas en A.
Las secuencias N-terminaies de los componentes purificados (Fig. 33)
mostraron que ambas pertenecían a la familia de LTPs. Sin embargo, en
80
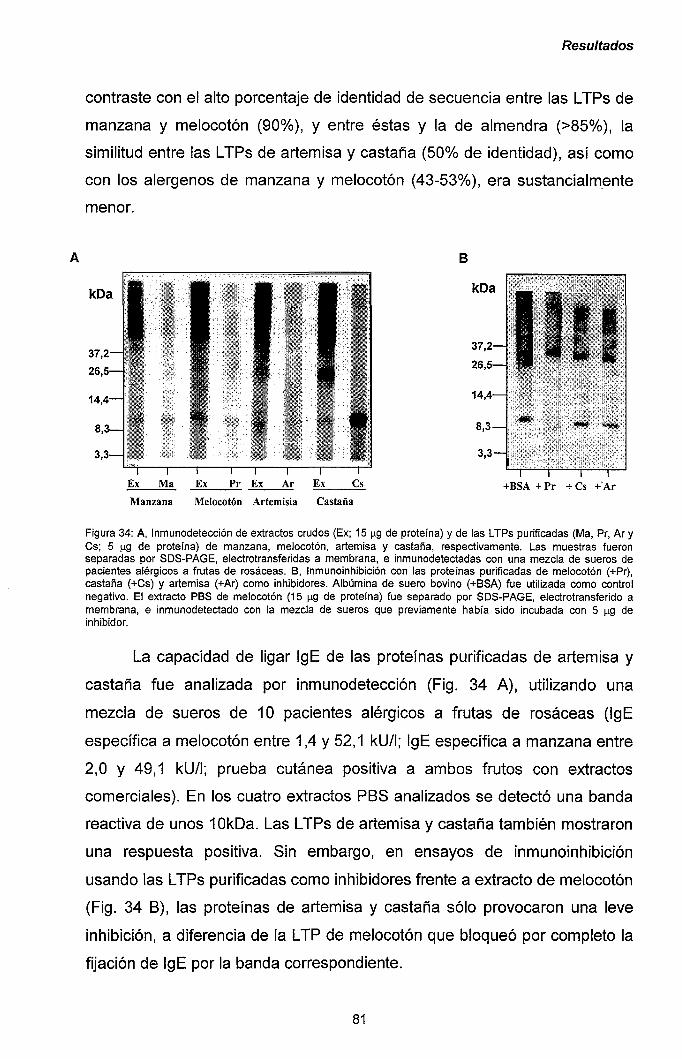
Resultados
contraste con el alto porcentaje de identidad de secuencia entre las LTPs de
manzana y melocotón (90%), y entre éstas y la de almendra (>85%), la
similitud entre las LTPs de artemisa y castaña (50% de identidad), así como
con los alérgenos de manzana y melocotón (43-53%), era sustancialmente
menor.
B
1 r +BSA + Pr + Cs +'Ar
Manzana Melocotón Artemisia Castaña
Figura 34: A, Inmunodetección de extractos crudos (Ex; 15 ^g de proteína) y de las LTPs purificadas (Ma, Pr, Ar y Cs; 5 ¡j.g de proteína) de manzana, melocotón, artemisa y castaña, respectivamente. Las muestras fueron separadas por SDS-PAGE, electrotransferidas a membrana, e inmunodetectadas con una mezcla de sueros de pacientes alérgicos a frutas de rosáceas. B, Inmunoinhibición con las proteínas purificadas de melocotón (+Pr), castaña (+Cs) y artemisa (+Ar) como inhibidores. Albúmina de suero bovino (+BSA) fue utilizada como control negativo. El extracto PBS de melocotón (15 ^g de proteina) fue separado por SDS-PAGE, electrotransferido a membrana, e inmunodetectado con la mezcla de sueros que previamente había sido incubada con 5 ng de inhibidor.
La capacidad de ligar IgE de las proteínas purificadas de artemisa y
castaña fue analizada por inmunodetección (Fig. 34 A), utilizando una
mezcla de sueros de 10 pacientes alérgicos a frutas de rosáceas (IgE
específica a melocotón entre 1,4 y 52,1 kU/l; IgE específica a manzana entre
2,0 y 49,1 kU/l; prueba cutánea positiva a ambos frutos con extractos
comerciales). En los cuatro extractos PBS analizados se detectó una banda
reactiva de unos lOkDa. Las LTPs de artemisa y castaña también mostraron
una respuesta positiva. Sin embargo, en ensayos de inmunoinhibición
usando las LTPs purificadas como inhibidores frente a extracto de melocotón
(Fig. 34 B), las proteínas de artemisa y castaña sólo provocaron una leve
inhibición, a diferencia de la LTP de melocotón que bloqueó por completo la
fijación de IgE por la banda correspondiente.
81
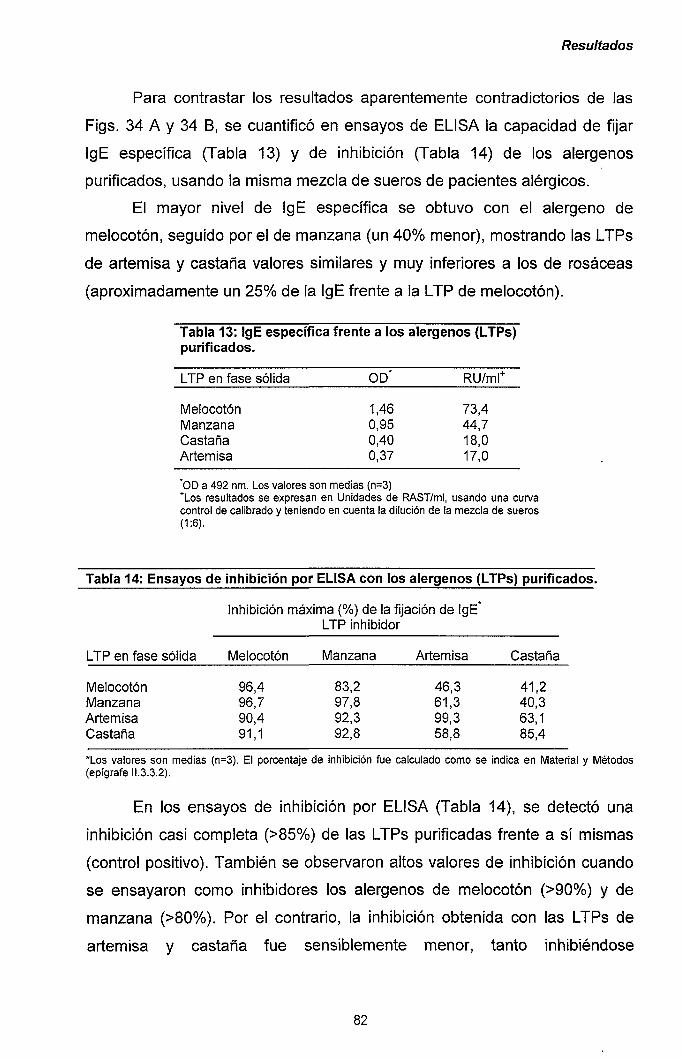
Resultados
Para contrastar los resultados aparentemente contradictorios de las
Figs. 34 A y 34 B, se cuantificó en ensayos de ELISA la capacidad de fijar
IgE específica (Tabla 13) y de inhibición (Tabla 14) de los alérgenos
purificados, usando la misma mezcla de sueros de pacientes alérgicos.
El mayor nivel de IgE específica se obtuvo con el alérgeno de
melocotón, seguido por el de manzana (un 40% menor), mostrando las LTPs
de artemisa y castaña valores similares y muy inferiores a los de rosáceas
(aproximadamente un 25% de la IgE frente a la LTP de melocotón).
1,46 0,95 0,40 0,37
73,4 44,7 18,0 17,0
Tabla 13: IgE específ ica frente a los a lérgenos (LTPs) puri f icados.
LTP en fase sól ida OD ^ RU/mr
Melocotón Manzana Castaña Artemisa
'OD a 492 nm. Los valores son medias (n=3) *Los resultados se expresan en Unidades de RAST/ml, usando una curva control de calibrado y teniendo en cuenta la dilución de la mezcla de sueros (1:6).
Tabla 14: Ensayos de Inhibición por ELISA con los alérgenos (LTPs) purificados.
Inhibición máxima (%) de la fijación de IgE* LTP inhibidor
LTP en fase sólida Melocotón Manzana Artemisa Castaña
Melocotón 96,4 83,2 46,3 41,2 Manzana 96,7 97,8 61,3 40,3 Artemisa 90,4 92,3 99,3 63,1 Castaña 91,1 92,8 58,8 85,4
*Los valores son medias (n=3). El porcentaje de inhibición fue calculado como se indica en Material y Métodos (epígrafe 11.3.3.2).
En los ensayos de inhibición por ELISA (Tabla 14), se detectó una
inhibición casi completa (>85%) de las LTPs purificadas frente a sí mismas
(control positivo). También se observaron altos valores de inhibición cuando
se ensayaron como inhibidores los alérgenos de melocotón (>90%) y de
manzana (>80%). Por el contrario, la inhibición obtenida con las LTPs de
artemisa y castaña fue sensiblemente menor, tanto inhibiéndose
82

Resultados
mutuamente (63-59%), como frente a los alérgenos de melocotón (46-41%)
o de manzana (61-40%).
Por último, las 4 LTPs purificadas en este trabajo se emplearon para
realizar ensayos in vivo, por prueba cutánea, en 47 pacientes (Tabla 4 de
Material y Métodos). Todos ellos eran alérgicos a melocotón; 24 habían
tenido procesos adversos con manzana, y sólo 10 con castaña. Por otra
parte, 39 de ellos sufría polinosis no ligada a Bet v 1.
En la Tabla 15 se resumen los resultados de las pruebas cutáneas. El
91,5% de los pacientes reaccionaron con la LTP de melocotón, y el 76,6%
con la de manzana. Sin embargo, el reconocimiento de las LTPs de castaña
y artemisa fue menor (23,4 y 36,1% respectivamente). Debe señalarse que
todos los pacientes que presentaban síntomas clínicos a castaña (Tabla 4 de
Material y Métodos) mostraron respuesta positiva a su LTP (Cs). El conjunto
de resultados obtenidos permitió establecer tres tipos principales de
asociaciones en la respuesta a las LTPs ensayadas. El grupo mayoritario de
los pacientes (46,8%) reconocieron sólo los alérgenos de melocotón y
manzana, un 21,3% reaccionó con las 4 LTPs, y un 10,6% con las proteínas
de ambos frutos de rosáceas y artemisa. Ningún paciente mostró prueba
positiva al alérgeno de castaña y no al de artemisa. Por último, un 10,6% de
los casos únicamente reconoció la LTP de melocotón.
Los datos presentados en este epígrafe indican que las LTPs son
alérgenos principales de melocotón y manzana, en poblaciones alérgicas a
estas frutas no sensibilizadas a Bet v 1. Estas proteínas se han denominado
Pru p 3 y Mal d 3, respectivamente, de acuerdo a las normas de
nomenclatura de alérgenos. Además, las LTPs de semilla de castaña y polen
de artemisa, denominadas provisionalmente Cas s 3 y Art v 3, son reactivas,
aunque en menor proporción, tanto in vitro como in vivo.
83
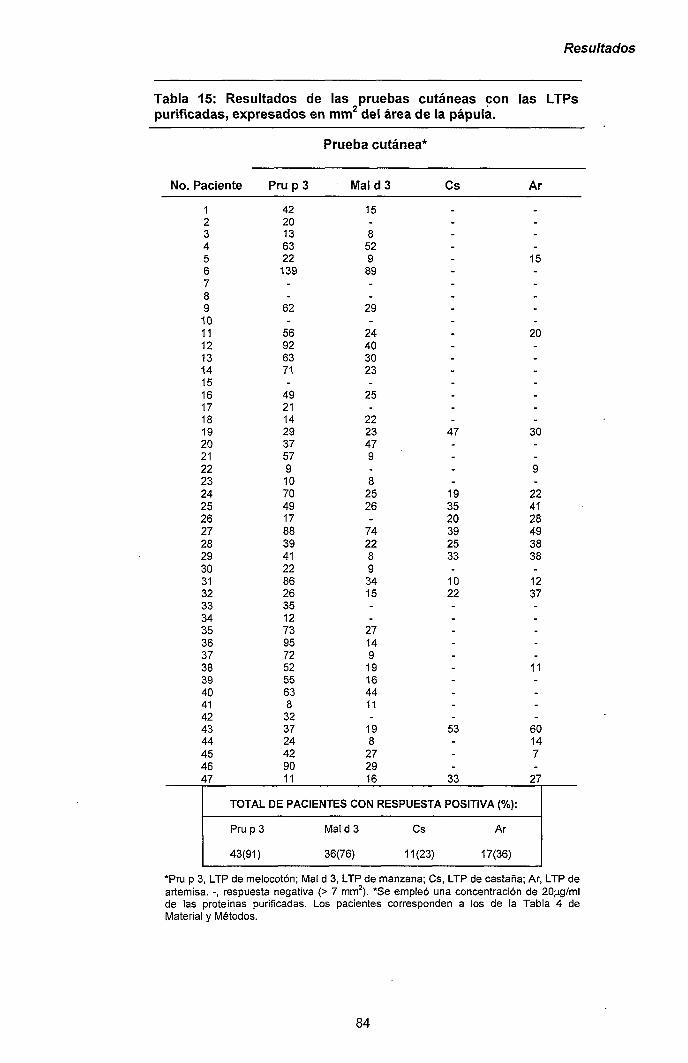
Resultados
Tabla 15: Resultados de las pruebas cutáneas con las LTPs purificadas, expresados en mm^ del área de la pápula.
Prueba cutánea*
No. Paciente
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47
Prup3
42 20 13 63 22 139 -. 62 -56 92 63 71 -49 21 14 29 37 57 9 10 70 49 17 88 39 41 22 86 26 35 12 73 95 72 52 55 63 8 32 37 24 42 90 11
Mald3
15 -8 52 9 89 --29 -24 40 30 23 -25 -22 23 47 9 -8 25 26 -74 22 8 9 34 15 --27 14 9 19 16 44 11 -19 8 27 29 16
Cs
_ -----------------47 ----19 35 20 39 25 33 -10 22 ----------53 ---33
Ar
_ ---15 -----20 -------30 --9 -22 41 28 49 38 38 -12 37 -----11 ----60 14 7 -27
TOTAL DE PACIENTES CON RESPUESTA POSITIVA {%):
Prup3
43(91)
MaldS
36(76)
Cs
11(23)
Ar
17(36)
*Pru p 3, LTP de melocotón; Mal d 3, LTP de manzana; Cs, LTP de castaña; Ar, LTP de artemisa. -, respuesta negativa (> 7 mm^). *Se empleó una concentración de 20ng/ml de las proteínas purificadas. Los pacientes corresponden a los de la Tabla 4 de Material y Métodos.
84

Resultados
ill.2.2.- Clonaje de los ADNc correspondientes a los alérgenos Pru
p 3 y Mal d 3.
A diferencia de lo que se conoce en otras especies (p.e. cereales,
almendra, etc.), no se ha descrito ningún clon de ADNc ni genómico de LTPs
de manzana o melocotón. Las propiedades alergénicas de estas proteínas
justifican el aislamiento y caracterización de sus clones de ADNc, con objeto
de producir, posteriormente, las correspondientes proteínas recombinantes.
Mald3
1 SI ATA ACÁ TGT GGC CAÁ OTO ACC AGC AGC CTT GCA CCA TGC ATT GGC TAC GTG I T C G O V T S S L A P C I G Y V
52 102 AGG AGT GGC GGA GCT GTC CCT CCA GCT TGC TGC AAT GGA ATC AGA ACC ATT R S G G A V P F A C C N G I R T I 103 153 AAC GGC TTG GCC AGG ACC ACC GCT GAC CGC CAG ACT GCT TGC AAC TGC CTG N G L A R T T A D R Q T A C N C L 154 204 AAG AAT CTT GCC GGC AGC ATC AGT GGT GTT AAC CCT AAC AAT GCA GCA GGG K N L A G S I S G V N P N N A A G
205 255 CTT CCT GGA AAG TGT GGA GTC AAC GTC CCC TAC AAG ATC AGC ACC TCC ACC L P G K C G V N V P Y K I S T S T
256 276 AAC TGC GCC ACC GTG AAG TAA N C A T V K *
Pru p3
1 51 ATA ACÁ TGT GGC CAÁ GTG TCC AGC AGC CTT GCA CCA TGC ATA CCC TAC GTG I T C G O V S S S L A P C I P Y V 52 102 AGA GGC GGT GGA GCT GTG CCT CCA GCT TGC TGC AAC GGC ATT AGG AAC GTC R G G G A V P P A C C N G I R N V 103 153 AAC AAC TTG GCA AGG ACC ACC CCT GAC CGC CAG GCC GCT TGC AAC TGC CTG N N L A R T T P D R Q A A C N C L 154 204 AAA CAG CTT TCC GCC AGC GTC CCC GGA GTC AAC CCT AAC AAT GCC GCA GCG K Q L S A S V P G V N P N N A A A
205 255 CTT CCC GGC AAG TGT GGA GTT AGT ATT CCT TAC AAG ATT AGC GCC TCC ACC L P G K C G V S I P Y K I S A S T
256 276 AAC TGC GCC ACC GTG AAG TGA N C A T V K *
Figura 35: Secuencia de los ADNc correspondientes a las LTPs de manzana (IVlal d 3) y melocotón (Pru p 3). Bajo la secuencia de nucleótidos se ha añadido la secuencia de aminoácidos y se ha subrayado la secuencia aminoterminal publicada por Sánchez-Monge ef al., (1999).
Para ello, se extrajo ARN total de piel de melocotón y manzana para
poder obtener ios ADNc correspondientes. A su vez, estos ADNc sirvieron
85

Resultados
para amplificar por PCR los ADNc completos correspondientes a las LTPs
de manzana y melocotón, utilizando para ello oligonucleótidos 5' específicos.
Dichos cebadores fueron diseñados teniendo en cuenta tanto las secuencias
N-terminales descritas por Sánchez-Monge et al. (1999), como la secuencia
completa de aminoácidos de Pru p 3 (Pastorello etal., 1999c), y la secuencia
de nucleótidos de la LTP de almendra (No. Accesión X96714), con la que
Pru p 3 y Mal d 3 comparten más de un 80% de las secuencias parciales
publicadas. De esta forma, se aislaron dos clones correspondientes a Pru p
3 y Mal d 3, cuyas secuencias se muestran en la figura 35.
P.pérsica M. domestica P.dulcis P.avium P. communis H. annuus Z.mays H. vulgare D. carota Par j 1
ITCGQVSSS LAPCIPYVRG GGAVPPA-CC NGIRNVNNLA RTTPDRQA ITCGQViiSS LAPClEYVRaGGAVPPA-CC NGIRgENgLA RTT^ORQÜ^
:RNVNNLA RTT
iN i
QVN rVQH KEKE^Ki
KTI 'KSL
P.pérsica M.domestica P.dulcis P. avium P.communis H. annuus Z.mays H. vulgare D. carota Par j l
Figura 36: Alineamiento de las secuencias de aminoácidos de LTPs de melocotón {Prunus pérsica), manzana (Malus domestica), almendra (P. dulcis; No. Accesión Q43017), cereza (P. avium; No. Accesión AAF26449), pera [Pyrus communis; No. Accesión AAF26451), girasol {IHelianthus annuus; No. Accesión Q39950), maíz {Zeas mays; No. Accesión P19656), cebada {Hordeum vulgare; No. Accesión P07597), zanahoria (Daucus carota; No. Accesión P27631) y del alérgeno de Parietaria judaica Par j 1 (No. Accesión P43217). Se han indicado los porcentajes de identidad de secuencia con Pru p 3.
La secuencia de aminoácidos correspondiente al ADNc obtenido de
melocotón presentó un único residuo diferente en la posición 76 (H/S),
comparada con la publicada por Pastorello et al. (1999c). Por otra parte, la
secuencia completa de la LTP de manzana y melocotón compartían un alto
porcentaje de identidad (81%).
86

Resultados
En la Figura 36 se presenta el alineamiento de las secuencias de Pru
p 3 y Mal d 3 con las de otras LTPs presentes en alimentos o pólenes (Par j
1). Mientras las proteínas de frutos de rosáceas muestran altos porcentajes
de identidad de secuencia (98-94%), estos valores disminuyen al comparar
Pru p 3 con sus homólogos de girasol (67%), cereales (63-52%) o zanahoria
(56%). La similitud con el alérgeno de Parietaria, Par j 1, es aún menor
(29%).
87

Discusión

Discusión
IV.1.- PROTEÍNAS DE DEFENSA Y ALERGIA.
La relación entre las características químicas y funcionales de una
proteína y su capacidad alergénica está aún por determinar. Aquellos
alérgenos que han sido caracterizados bioquímicamente presentan
propiedades muy dispares. Tampoco se han identificado funciones
biológicas, a excepción de la actividad proteasa del alérgeno de D.
pteronyssinus Der p 1 y de algunos alérgenos de hongos (Bufe, 1998;
Shakib et al., 1998; Shakib y Gough, 2000), que se hayan relacionado
directamente con los incrementos en los niveles de IgE específica. En este
contexto, y aún sin establecer pautas claras en la relación estructura-
función-capacidad alergénica, es significativa la interacción establecida en
los últimos años entre los alérgenos alimentarios de origen vegetal y las
proteínas de defensa en plantas (Breiteneder y Ebner, 2000; Hoffmann-
Sommergruber, 2000). De hecho, en este trabajo se han caracterizado dos
nuevas familias de proteínas alergénicas, quitinasas de clase I y LTPs, que
previamente ya habían sido incluidas en las familias PR-3 y PR-14 de
proteínas relacionadas con la patogénesis (van Loon y van Strier, 1999).
Las proteínas de defensa se acumulan en respuesta a ataques por
patógenos y/o insectos, y, en algunos casos, a estrés abióticos (Atkinson et
al., 1996). Existen pocos trabajos en los que se estudie la regulación de la
expresión de los alérgenos, aunque el cambio en su concentración relativa
puede suponer un serio problema de salud para aquellas personas que
estén sensibilizadas (Taylor, 1997). En este sentido, la inducción de
proteínas de defensa por distintas vías puede tener relevancia clínica. Así,
por ejemplo, se ha descrito la inducción en nabo {Brassica rapa) por etileno,
ácido salicílico o herida, de una proteína de la familia PR-4, homologa a la
proheveína de látex, que es reconocida por más del 80% de los sueros de
pacientes alérgicos a látex, y que provoca prueba cutánea en 4 de 6
pacientes analizados (Hanninen et al., 1999). Del mismo modo, en este
trabajo se ha demostrado que la inducción por etileno de la quitinasa de
clase I de judía verde aumenta el reconocimiento de los correspondientes
89

Discusión
extractos por una mezcla de sueros de pacientes con alergia a látex y frutas.
Por tanto, el uso de algunos tratamientos para mejorar la resistencia a
patógenos o tratamientos inductores de la maduración de frutos, podría estar
afectando al potencial alergénico del alimento vegetal. La caracterización de
alérgenos y el estudio de su expresión podría, en consecuencia, prevenir
contactos no deseados en pacientes alérgicos a alimentos de origen vegetal
(Hoffmann-Sommergruber etal., 1999).
IV.2.- ALÉRGENOS IMPLICADOS EN EL SÍNDROME LÁTEX-FRUTAS.
La alergia al látex se está incrementando en los últimos años,
llegando a ser un grave problema de salud no sólo por la incidencia, sino por
sus manifestaciones clínicas (Slater, 1995; Sussman y Beezhold, 1995;
Turjanmaa et al., 1996). Más del 50% de los pacientes que sufren
reacciones adversas al látex, están sensibilizados a frutas, razón por la cual
se propuso que a este tipo de asociación se la denominase síndrome látex-
frutas (Blanco eí al., 1994). Esta asociación está bien establecida a iiivel
clínico, aunque los componentes responsables de las reacciones cruzadas
no habían sido identificados. Distintos grupos habían observado, por
ensayos de inmunodetección, bandas de 30-35 kDa en varios frutos que
fijaban IgE de pacientes alérgicos a látex, y habían sugerido su potencial
importancia en la cosensibilización (Lavaud et al., 1995; Wadee et al., 1990;
Delbourg etal., 1996, Alenius etal., 1996b).
IV.2.1.- Las quitínasas de clase I son alérgenos mayoritarios en
castaña, aguacate y plátano.
Los frutos que más se han asociado con la alergia a látex son
castaña, aguacate, plátano y kiwi. En este trabajo, se utilizaron extractos de
castaña, aguacate y plátano para identificar los alérgenos mayoritarios
responsables de la asociación de estos frutos con la alergia a látex. En estos
extractos, se observó una única banda reactiva frente a sueros de pacientes
90

Discusión
alérgicos a látex y frutas de unos 32 kDa en castaña y aguacate (Flg. 5), y
dos bandas reactivas de 32 y 34 kDa en plátano (Fig. 12). Estas mismas
bandas fueron reconocidas por anticuerpos policlonales monoespecíficos
frente a qultinasas (Figs. 5 y 12), lo que, junto con su peso molecular,
sugería que se trataba de quitinasas de clase I. Esta hipótesis fue
confirmada con la purificación y caracterización de las cuatro proteínas
reactivas. La actividad quitinasa fue comprobada en todos los casos (Tablas
5 y 8), así como su reconocimiento por anticuerpos frente a quitinasas (Figs.
7 y 12). Sus secuencias N-terminales (Figs. 8 y 12) mostraron la presencia
de dominios heveína característicos de quitinasas de clase I, con altos
niveles de identidad de secuencia (> 80%) con la heveína de látex. La
capacidad alergénica de las proteínas purificadas fue confirmada in vitro por
su fijación de IgE específica en inmunodetección y por sus altos niveles de
inhibición de la fijación de IgE específica por los correspondientes extractos
crudos, tanto en ensayos de inmunoinhibición (Fig. 9 y 15), como utilizando
CAP comerciales (Fig. 11 y 16). Los ensayos de inmunoinhibición
permitieron, además, descartar el papel de posibles alérgenos no
relacionados con similares pesos moleculares aparentes.
La reactividad in vitro de los cuatro alérgenos aislados se constató por
pruebas cutáneas en un grupo de pacientes alérgicos a látex y frutas (Tablas
7 y 9). La proteína de castaña. Cas s 5, fue reconocida en un 72% de los
casos, el alérgeno de aguacate, Prs a 1, en un 67% de los individuos
analizados, y los dos componentes de plátano. Mus a 1.1 y Mus a 1.2 en un
53% y un 60% de los mismos.
La reactividad cruzada con látex de las qultinasas de clase I
estudiadas se comprobó utilizando extractos crudos de castaña y aguacate,
así como los alérgenos purificados Cas s 5 y Prs a 1 en ensayos de
inmunoinhibición (Fig. 10). La preincubación de la mezcla de sueros de
pacientes alérgicos con un extracto de látex, bloqueó por completo la unión
de IgE a las quitinasas alergénicas. Esto indica que hay epítopos comunes
mayoritarios compartidos por algún(os) componente(s) de látex, Cas s 5 y
Prs a 1. Si bien existen referencias de una quitinasa de clase I en látex
91

Discusión
(Breiteneder y Scheiner, 1998), de la que aún no se han publicado datos
bioquímicos ni inmunológicos, ésta parece ser un alérgeno minoritario (no ha
sido descrita su actividad alergénica en ningún trabajo sobre alergia a látex).
En consecuencia, parece que existen otros componentes de látex
responsables de la cosensibilización con frutas.
Todos los datos anteriormente comentados permiten asumir que las
quitinasas de clase I son los alérgenos principales de castaña, aguacate y
plátano implicadas en el síndrome látex-frutas. Cas s 5 ha sido identificada
como la quitinasa de clase I de castaña, Ch3, previamente descrita (Collada
et al., 1992; Aliona et al., 1995). La actividad antifúngica de este enzima ha
sido estudiada, y los residuos que conforman su centro activo se han
localizado por mutagénesis dirigida de su correspondiente clon de ADNc y la
expresión heteróloga de las enzimas mutantes (Collada et al., 1992; García-
Casado eí al., 1998). Sin embargo, no se ha realizado ningún otro trabajo
sobre sus propiedades alergénicas, al margen del expuesto en esta
memoria. Por el contrario, datos obtenidos por otros grupos confirman
plenamente los resultados presentados sobre los alérgenos de aguacate y
plátano. Sowka et al. (1998a) han aislado un clon de ADNc de Prs a 1,
expresado la proteína recombinante en Pichia pastoris y mostrado su
capacidad de ligar IgE específica, así como su posible reactividad cruzada
con proheveína de látex. Prs a 1 ha sido también identificado por Chen eí al.
(1998) y por Posh et al. (1999), que han demostrado por ensayos de CAP de
inhibición que la heveína de látex bloquea completamente la unión de IgE a
extractos de aguacate, mientras no se produce el resultado contrario. Su
conclusión tentativa es que la sensibilización a aguacate en pacientes
alérgicos a látex está causada exclusivamente por epítopos IgE presentes
en la heveína. Posch et al. (1999) observaron además que Prs a 1 no perdía
su capacidad de bloquear la unión de IgE a un extracto crudo de aguacate
después de haber sido tratada con un fluido gástrico simulado. La estabilidad
de las propiedades alergénicas en el tracto digestivo ha sido citada como
una de las características de los alérgenos alimentarios (Astwood eí al.,
1996). Finalmente, Mus a 1.1 y Mus a 1.2 han sido también localizados en
92

Discusión
plátano como alérgenos responsables del cruce con látex por Mikkola et al.
(1998), aunque en el caso de sus proteínas purificadas sólo se obtuvo
prueba cutánea positiva en 1 de 4 pacientes analizados.
En los extractos de castaña y aguacate, se detectaron quitinasas de
clase II junto a las de clase I. No así en plátano maduro, probablemente
debido a que durante la maduración natural de este fruto se reduce la
expresión de los genes que codifican para enzimas de clase II, y se
incrementa la de los homólogos de clase í (Cledennen y May, 1997). Las
quitinasas de clase II purificadas de castaña, Csll 1, y aguacate, Pall 1, aún
mostrando actividad enzimática y un fuerte reconocimiento por los
anticuerpos policlonales monoespecíficos frente a Csll 1, no parecen poseer
capacidad alergénica. No fueron reconocidas por los sueros de pacientes
alérgicos, ni ejercieron ninguna inhibición en los ensayos correspondientes
(Fig. 7 y 12), y su reactividad in vivo fue prácticamente nula (Tabla 7, sólo
Pall 1 fue reconocida por 2/18 pacientes). Únicamente en pruebas de
inhibición utilizando CAP comerciales se detectó alguna potencial unión a
IgE (Fig.11; valores máximos de inhibición de un 40%). El conjunto de los
datos referentes a las quitinasas de clase II indican que estas enzimas no
tienen un papel relevante (probablemente no juegan ninguno) en el síndrome
látex-frutas, y apuntan a la importancia esencial del dominio heveína en las
reacciones alérgicas. También podrían explicar el papel menor de la
hevamina, una quitinasa de clase III sin dominio heveína, como alérgeno de
látex (Alenius etal., 1996b).
IV.2.2.- Las quitinasas de cíase I como potenciales panaíergenos
vegetales.
Las quitinasas de clase I son una familia de proteínas ampliamente
distribuida en el reino vegetal, y dada su similitud de secuencia, era plausible
suponer que fueran responsables de reacciones cruzadas entre el látex y
otros alimentos, además de castaña, aguacate y plátano.
93

Discusión
El número de alimentos relacionados con alergia al látex se ha ido
incrementado en los últimos años (Blanco et al., 1994; Beezhold et al., 1996;
Brehier et al., 1997; Lavaud et al., 1997; Levy eí al., 2000). Por esa razón, se
seleccionaron 20 alimentos con distinto grado de implicación en el síndrome
látex-frutas, desde papaya o fruta de la pasión, claramente implicados por
estudios clínicos, hasta harina de trigo que no había sido asociada
previamente con el síndrome. Los anticuerpos policlonales monoespecíficos
frente a quitinasas sirvieron como herramienta para detectar quitinasas de
clase I y II en los extractos analizados (Fig. 17 C). La mayoría de las
preparaciones (16 de 21) presentaban al menos una banda reactiva con
dichos anticuerpos. Entre los componentes que fueron reconocidos, había
algunos de peso molecular comprendido entre 23 y 28 kDa (posiblemente
quitinasas de clase II, no alergénicas) y otros de 30 a 40 kDa
(probablemente quitinasas de clase I, alergénicas). Hubo otras bandas que
fueron reconocidas por los anticuerpos frente a quitinasas, con menor (16 a
20 kDa) y mayor (50 a 70 kDa) peso molecular aparente, de naturaleza
desconocida. Estos componentes (las bandas de 50-70 kDa) podrían
representar quitinasas con tamaños moleculares atípleos, agregados de las
enzimas de clase I y/o II, o proteínas relacionadas con las quitinasas.
Varias de las bandas reactivas con el suero antiquitinasas,
especialmente aquellas con peso molecular comprendido entre 30 y 45 kDa,
también fueron reconocidas por la mezcla de sueros de pacientes alérgicos
a látex y frutas (Fig. 17 D). Estas bandas fueron observadas en los extractos
crudos de chirimoya, fruta de la pasión, kiwi, papaya, mango, tomate y trigo.
Estos resultados sugerían que los componentes que fijaban IgE se podían
incluir en la familia de quitinasas de clase I. En cambio, las bandas con
pesos moleculares de 10-20 kDa, de 23-28 kDa y de 50-70 kDa, reconocidas
por anticuerpos frente a quitinasas, no reaccionaron con la mezcla de
sueros. Los datos anteriores confirman la asociación entre frutas tropicales
(p.e. papaya, fruta de la pasión, y kiwi) y tomate, con el síndrome látex-
frutas, previamente establecida por distintos grupos clínicos (Blanco eí al.,
1994; Brehier et al., 1997; Levy eí al., 2000). En este contexto, la ausencia
94

Discusión
de componentes reactivos con la mezcla de sueros de pacientes, tanto en
pina (una fruta tropical) como en patata (muy relacionada con tomate),
resultaba contradictoria. Sin embargo, la pina fue la única fruta tropical que
no presentó bandas reconocidas por el suero anti-quitinasas. En el caso de
patata, los componentes que reaccionaron con este suero (21 y 55-75 kDa)
no mostraban pesos moleculares aparentes compatibles con los de
quitinasas de clase I (30-40 kDa). Además, se ha sugerido que la
cosensibilización a látex y a patata está originada por la homología existente
entre el alérgeno Hev b 7 y la patatina, proteína presente en el tubérculo que
no está relacionada con quitinasas de clase I (Beezhold et al., 1994; Kostyal
etal., 1998; Sowka etal., 1998b).
Un dato sorprendente es la detección en la harina de trigo de una
putativa quitinasa de unos 33kDa, que era reconocida por los sueros de
pacientes alérgicos a látex y frutas. El resultado sugiere una posible
asociación de este cereal con la alergia a látex, lo que no había sido
detectado hasta el momento. Sin embargo, la inactivación de las quitinasas
de clase I por calor (ver epígrafe siguiente) invalida en gran parte la posible
relevancia clínica de esta observación.
En el caso de látex, la mezcla de sueros de pacientes reconoció
bandas de 6-10 kDa y de 20 kDa, además de los componentes de 35-45 kDa
que reaccionaban con los anticuerpos frente a quitinasas. Los componentes
de bajo peso molecular probablemente corresponden a proheveína (Hev b
6.01), su dominio C-terminal (Hev b 6.03), y la propia heveína (Hev b 6.02),
que no deben ser reconocidas por sueros antiquitinasas de clase II sin
domino heveína. La presencia de hevamina entre las bandas de 35-45 kDa,
aunque no puede descartarse, resulta dudosa. Esta quitinasa de clase III de
látex, está muy poco relacionada con las de clase II y I (Jekel et al., 1991), y
además presenta muy baja afinidad por la IgE de sueros de pacientes con el
síndrome látex-frutas (Yagami etal., 1998).
La implicación de las quitinasas de clase I en el síndrome látex-frutas,
fue confirmada por ensayos de inmunoinhibición. Prs a 1, el alérgeno
mayoritario de aguacate, inhibió la unión a IgE de todas las bandas reactivas
95

Discusión
en los distintos extractos (Flg. 18). Esto sugiere que diciías bandas (uno o
dos componentes dependiendo del alimento), que, a su vez, son ios únicos
potenciales alérgenos reconocidos por los sueros de pacientes con el
síndrome látex-frutas, representan quitinasas de clase I que poseen epítopos
comunes con Prs a 1. La desaparición de estas mismas bandas al
preincubar la mezcla de sueros con un extracto de látex, mostró la
reactividad cruzada entre los putativos alérgenos de los alimentos
analizados y componentes del látex.
Por otra parte, cuando se empleó suero de pacientes alérgicos a
látex, pero no a frutas, el patrón de reconocimiento fue diferente (Fig. 17 E).
Sólo se observaron bandas que fijaban IgE en el extracto de látex, con un
peso molecular aparente entre 30 y 60 kDa. Ni la quitinasa de clase I de
castaña, ni sus posibles homólogos en chirimoya, fruta de la pasión, papaya,
klwi, mango, tomate, o trigo, así como tampoco las bandas de bajo peso
molecular de látex, todas ellas reconocidas por el suero de los pacientes con
el síndrome látex-frutas, reaccionaron con la nueva mezcla de sueros. Estos
resultados sugieren que hay distintos patrones de sensibilización en ambos
grupos de pacientes, y que las quitinasas de clase I de alimentos juegan un
papel específico en las reacciones cruzadas con látex, pero no en alergia a
látex que no va acompañada por sensibilización a frutas. Los datos de
Lavaud et al. (1995) referentes a extractos de plátano y aguacate, indican
que las quitinasas de clase I de estos alimentos (Prs a 1 y Mus a 1)
muestran el mismo tipo de reconocimiento diferencial (positivo en el caso de
látex-frutas y negativo en el de látex-no frutas).
Se ha propuesto que los N-oligosacáridos complejos de glicoproteínas
tienen un papel destacado en la cosensibilización a látex y a pólenes de
árboles y herbáceas (Fuchs et al., 1998). De hecho, se ha demostrado que
los agentes causantes de algunas reacciones cruzadas son estos
oligosacáridos (Petersen etal., 1996; Aalbersey van Ree, 1997). Por ello, se
utilizó un suero específico frente a los mismos, para determinar si en el
síndrome látex-frutas tenían alguna relevancia (Fig. 17 B). Las drásticas
diferencias entre los componentes reactivos frente a este suero y los
96

Discusión
reconocidos por la IgE de pacientes alérgicos, junto al hecho de que varios
extractos con una fuerte respuesta a anticuerpos específicos frente a
azúcares no mostraron ningún componente que ligara IgE específica, indican
que los N-oligosacáridos complejos no están involucrados de forma
relevante en las reacciones cruzadas del síndrome látex-frutas.
IV.2.3.- Los panalergenos del síndrome látex-frutas son
termolábiles e inducibies por etiíeno.
Los resultados discutidos en el anterior epígrafe sugieren que
quitinasas de clase I potencialmente alergénicas están presentes en algunos
alimentos, como harina de trigo, no asociados con el síndrome látex-frutas.
De hecho, considerando la amplia distribución de estos enzimas en las
fuentes alimentarias de origen vegetal (p.e. cereales y leguminosas), así
como el alto grado de similitud estructural entre las mismas, podría
esperarse que los pacientes con dicho síndrome estuviesen sensibilizados a
la mayoría de los alimentos con quitinasas de clase I. Sin embargo, éste no
es el caso de una gran parte de las materias primas vegetales, como los
cereales y las leguminosas, que han sido raramente relacionadas con la
alergia a látex (Blanco et al., 1994; Brehier et al., 1997). La judía verde,
ampliamente consumida, se encuadra en este tipo de alimentos: los datos
clínicos disponibles no la asocian al síndrome látex-frutas, aún cuando está
demostrada la presencia de quitinasas de clase I en sus extractos (Broglie et
al., 1986), que, además, provocan una respuesta positiva en pacientes con
este síndrome cuando se ensayan por prueba cutánea (Tabla 11). Estas
características determinaron su elección como sistema modelo, para tratar
de dilucidar la aparente contradicción que se deriva de la presencia de
quitinasas reactivas y la no relación del alimento con la alergia a látex.
Por ensayos de inmunodetección, se observó la presencia de una
única banda de 32 kDa, también reconocida por los anticuerpos frente a
quitinasas, que fijaba IgE de sueros de pacientes con síndrome látex-frutas,
en extractos crudos de judía verde (Fig. 19). Esta proteína reactiva, llamada
97

Discusión
PvChl, fue identificada como una quitinasa de clase i (Fig. 22 y Tabla 10),
altamente relacionada con el alérgeno de aguacate, Prs a 1. La reactividad
cruzada entre ambas proteínas se confirmó por ensayos de inmunoinhibición
(Fig. 23). PvChI fue tan reactiva como Prs a 1, también, en ensayos in vivo
(Tabla 11), siendo reconocida por 7 de los 8 pacientes analizados. Por tanto,
puede concluirse que la judía verde contiene un alérgeno homólogo a los
que se han descrito en castaña, aguacate y plátano.
Por otro lado, cabe destacar el incremento en la expresión de PvChI
cuando las vainas de judía verde fueron tratadas con etileno (Fig. 19 y 20).
Este hecho puede ser relevante si se tiene en cuenta que tratamientos
similares son usados para acelerar la maduración después del
almacenamientos de frutos climatéricos como manzana, plátano, aguacate o
tomate. Este tipo de tratamientos puede, por tanto, cambiar sustancialmente
el contenido de alérgenos en los frutos maduros.
En contraste con los datos obtenidos en judía verde, no se detectó
ninguna banda que fijase IgE en la semilla madura de judía (judía blanca), lo
que sugiere que los niveles de PvChI podrían disminuir en maduración. Sin
embargo, se localizó un componente de 32 kDa que era reconocido por los
anticuerpos frente a quitinasas (Figs. 19 y 21), caracterizado como una
fitohemaglutinina (Fig. 22), previamente descrita (Hoffman y Donaldson,
1985). Esta proteína purificada, PvPhy, no presentó actividad alergénica, ni
in vitro ni in vivo.
La judía verde no se consume, normalmente, de forma cruda, sino
que previamente se la somete a un proceso de cocción. Por ello, se estudió
el efecto de un tratamiento térmico sobre las propiedades alergénicas de
PvChI, utilizando además Prs a 1 como control. Dicho tratamiento tuvo como
resultado la pérdida completa de la alergenicidad de las quitinasas de clase I
tanto de judía verde como de aguacate (Fig. 24 y Tabla 11). Esta pérdida de
actividad está acompañada por la ausencia de detección de las proteínas
tratadas en geles de SDS-PAGE, lo que probablemente pueda deberse a la
desnaturalización y agregación de las muestras calentadas. La inactivación
también se ha observado en extractos de judías verdes cocidas, así como en
98

Discusión
productos comerciales enlatados o congelados de las mismas, que suelen
sufrir un tratamiento térmico durante el procesamiento industrial.
Si el comportamiento de Prs a 1 y PvChl es común para las quitinasas
de clase I alergénicas, parece probable que el procesado y cocinado de los
alimentos que las contienen, inactive estos potenciales alérgenos. Este
hecho podría explicar el porqué los frutos frescos son los principalmente
asociados al síndrome látex-frutas, mientras que alimentos consumidos
después de un tratamiento térmico, aun conteniendo quitinasas alergénicas,
raramente se relacionan con la alergia al látex.
IV.2.4.- El dominio heveína, y no el catalítico, es responsable de
la alergenícidad de las quitinasas de clase I.
Los resultados discutidos en los epígrafes anteriores permiten concluir
que:
- Las quitinasas de clase I son los alérgenos principales de los frutos
más estrechamente asociados con la alergia al látex.
- Las quitinasas de clase II de estos frutos, que carecen de dominio
heveína N-terminal, pero que tienen dominios catalíticos homólogos a
las de clase I, no presentan capacidad alergénica.
- Hay componentes en látex que comparten epítopos IgE con las
quitinasas de clase I.
La consideración conjunta de estos hechos sugiere que el dominio
heveína juega un papel central en la alergenícidad de las quitinasas, y que la
heveína es el componente de látex responsable de las reacciones cruzadas.
Además de las quitinasas, existen otras proteínas vegetales con
dominios heveína que podrían considerarse como potenciales alérgenos.
Entre ellas se encuentra la aglutinina de germen de trigo, con cuatro de
estos dominios repetidos en tándem, alguno de los cuales (domino 3)
presenta hasta un 60% de identidad con la heveína de látex. Sin embargo,
esta lectina no fue reconocida por los sueros de pacientes alérgicos
utilizados en este trabajo, ni inhibió la fijación de IgE a Cas s 5 cuando fue
99

Discusión
preincubada con dichos sueros (Fig. 26). Estos datos indican que no todas
las proteínas con dominios homólogos a la heveína son alergénicas.
Nuestros datos en este contexto son contradictorios con los de otros autores
que han analizado sueros de pacientes alérgicos al látex en poblaciones
noreuropeas o noramericanas (Alenius etal., 1996c; Beezhold et al., 1997).
En estos casos, se detectó fijación de IgE específica por WGA, así como una
inhibición parcial del reconocimiento de la heveína de látex al preincubar con
la aglutinina.
Si bien todos los resultados indicaban que el dominio heveína era el
único responsable de la capacidad alergénica de los alérgenos implicados
en el síndrome látex-frutas, no existía ningún dato que lo corroborara de
forma directa. Era posible que también hubiera epítopos en la parte
diferencial entre el domino catalítico de las quitinasas de clase I y las de
clase II. La forma directa de conocer si el domino heveína y/o el catalítico de
las quitinasas de clase I tenían capacidad alergénica, era poder expresarlos
como proteínas independientes, y comparar su reactividad frente a sueros
de pacientes con alergia a látex y frutas, tomando como referencia a la
proteína completa. Por ello, se abordó la expresión de Cas s 5 y de sus dos
dominios de forma independiente, en el vector de expresión extracelular
pPIC 9 (Fig. 3), en la levadura Pichia pastoris (el esquema de las
construcciones está indicado en la Fig. 4).
La expresión se llevó a cabo en esta levadura, debido a que
presentaba la ventaja de ser un organismo eucariota, y por tanto, con una
maquinaria de plegamiento tridimensional más similar a la de plantas.
También se eligió este organismo porque las quitinasas de clase I no eran
tóxicas para la levadura y se podía, potencialmente, obtener una cantidad
apreciable de producto recombinante.
El alérgeno completo recombinante, rCas s 5, así como su dominio
catalítico, rCAT, se expresaron a niveles de 30 mg/l de cultivo en medio
mínimo. Por el contrario, las dificultades de detección y purificación del
dominio heveína, tanto en medio mínimo como completo, después de
100

Discusión
analizar más de 15 transformantes distintos, determinaron prescindir de este
producto recombinante en pruebas posteriores.
Tanto rCas s 5 como rCAT eran los componentes mayoritarios en los
sobrenadantes de los cultivos correspondientes (Fig. 28), a partir de los
cuales se purificaron. Las proteínas recombinantes purificadas presentaron
las secuencias N-terminales esperadas, incluido el primer glutámico (E) que
se añadió a la construcción del domino catalítico para facilitar el corte del
péptido señal. Sin embargo, se observó un cambio de movilidad
electroforética en rCas s 5, con respecto a Cas s5, en SDS-PAGE (Fig. 29).
Ese cambio pudo atribuirse a la presencia de azúcares en las proteínas
recombinantes, demostrado por el método de detección de azúcares por
digoxigenina (Fig. 29). Se ha descrito que Pichia pastoris puede glicosilar
inespecíficamente en todos las secuencias diana para la N-glicosilación. En
la secuencia de aminoácidos de Cas s 5, se localizó una de estas dianas en
el dominio catalítico (indicada en la Fig. 4). Si bien en las proteínas nativas
no se detectó glicosilación, ambas proteínas recombinantes sí estaban
glicosiladas. Este tipo de modificación ha sido previamente descrito en varios
alérgenos expresados en P. pastoris, incluido Prs a 1 (Sow/ka etal., 1998a).
El procesamiento, y, probablemente, plegamiento correcto de rCas s 5
y rCAT, está sustentado por su reconocimiento por anticuerpos frente a
quitinasas, y por su actividad enzimática (Tabla 12), similar a la de las
quitinasas de clase I (Cas s 5) y II (Csll 1) aisladas de semillas de castaña.
La deleción del dominio heveína conlleva una pérdida completa de la
capacidad alergénica de rCas s 5, al menos in vitro. Mientras la quitinasa
recombinante completa es tan activa como el alérgeno natural Cas s 5, tanto
en ensayos de inmunodetección como de inmunoinhibición (Fig. 29 y 30),
rCAT no es reconocida por sueros de pacientes alérgicos y su preincubación
con la mezcla de sueros no tiene efecto alguno en la fijación de IgE por rCas
s 5 o Cas s 5. Este comportamiento es similar al de la quitinasa natural de
clase II de castaña, Csll 1. Basándose en esto datos, puede concluirse que
todos los epítopos IgE principales de la quitinasa de clase I de castaña se
encuentran localizados en el dominio heveína. Si se considera la alta
101

Discusión
similitud de secuencia de este dominio con el de otras quitinasas alergénicas
(75-80% de identidad con Prs a 1, Mus a 1.1 ó PvChl), junto a la falta de
reactividad de Pali 1 (quitinasa de clase II de aguacate), la anterior
conclusión parece generalizable a los alérgenos de frutos implicados en el
síndrome látex-frutas.
La reactividad cruzada entre el alérgeno de castaña (natural y
recombinante) con la heveína de látex, se confirmó en ensayos de
inmunoinhibición. La heveína purificada inhibió en gran parte la fijación de
IgE por Cas s 5 o rCas s 5 (Fig. 30). Resultados similares han sido obtenidos
por Chen et al. (1998) para Prs a 1 analizando extractos crudos de aguacate,
y por Mikkola et al. (1998) para el alérgeno natural de plátano. Estos datos
suponen la presencia de epítopos comunes en la heveína de látex y en los
dominios N-terminales de las quitinasas alergénicas. De hecho, no sólo' hay
una alta identidad de secuencia (>70%) entre los dominios heveína de estos
alérgenos y el péptido de látex, sino que además, de los dos epítopos IgE
mapeados en la heveína (H1, residuos 14-25, y H2, residuos 31-38;
Beezhold et al., 1997), la secuencia de H1 se encuentra altamente
conservada en el dominio de las quitinasas alergénicas. Sin embargo, la
débil inhibición de la fijación de IgE por la heveína ejercida por las quitinasas
de plátano y aguacate (Chen et al., 1998; Mikkola et al., 1998), sugiere que
algunos epítopos de unión a IgE de la heveína no están presentes en los
alérgenos de frutas (p.e. el denominado H2, cuya secuencia ha variado
drásticamente en Cas s 5, Prs a 1 o Mus a 1.1).
La producción de rCas s 5 puede tener, en un futuro, aplicaciones
clínicas en el diagnóstico del síndrome látex-frutas, en la misma línea
propuesta para otros alérgenos recombinantes (Valenta eí al., 1999a). Por
otro lado, el pequeño tamaño de su dominio heveína hace factible abordar
un mapeo de epítopos usando un número razonable de péptidos sintéticos, y
posteriormente la producción de mutantes hipoalergénicos (si se solventan
los problemas de su expresión como péptido recombinante).
102

Discusión
IV.3.- LTPs: UNA FAMILIA DE PANALERGENOS CUYOS MIEMBROS
PRESENTAN DIFERENTE ACTIVIDAD ALERGÉNICA.
Como se ha comentado en la Introducción, la alergia alimentaria a
frutos de la familia de las rosáceas está asociada, frecuentemente, a polen
de abedul en el área del Centro y Norte de Europa, Estados Unidos y
Canadá. Los síntomas que provoca suelen estar limitados al síndrome de
alergia oral, posiblemente debido a que los agentes responsables de la
cosensibilización, homólogos a Bet v 1, son inactivados en la digestión
gastrointestinal (van Ree eí al., 2000b). Un claro contraste presentan varios
grupos de pacientes alérgicos, con o sin polinosis asociada, del área
mediterránea, en los que la alergia a frutos de rosáceas no se relaciona con
Bet V 1, y en los que, además del síndrome de alergia oral, pueden
manifestarse síntomas sistémicos severos (Ortolani etal., 1988; Pastorello eí
al., 1994, 1998; Fernández-Rivas etal., 1997; Rodríguez etal., 2000). Las
LTPs han sido identificadas como potenciales alérgenos de melocotón (Pru p
3), manzana (Mal d 3) y albaricoque (Pru ar 3), específicos de estas
poblaciones mediterráneas (Sánchez-Monge eí al., 1999; Pastorello eí al.,
1999a y b, 2000).
Par j 1, el alérgeno principal de Paríetaria judaica (Duro et al., 1997),
es otra LTP, alejada en secuencia de las de frutos de rosáceas (Fig. 36), y
LTPs alergénicas parece que también se han purificado de extractos de
maíz, zanahoria, y brócoli (Pastorello eí al., 1998; Asero eí al., 2000),
aunque sus características bioquímicas, inmunológicas o clínicas aún no se
han publicado. Bandas de bajo peso molecular que ligan IgE específica,
probablemente LTPs, se han descrito en frutos de otras rosáceas (ciruela,
cereza, mora; Pastorello eí al., 1994; Navarro eí al., 1997), y en productos
procesados de especies filogeneticamente distantes (cerveza; Curioni eí al.,
1999). Además, alérgenos de pequeño tamaño molecular (w7kDa),
débilmente relacionados con las LTPs, pero que conservan la posición de la
mayoría de las cisteínas y de algunos residuos adyacentes, se han
103

Discusión
identificado en polen de Brassica rapa (Toriyama et al., 1998) y en cascara
de soja (González et al., 1995).
Este panorama, junto a la distribución ubicua de las LTPs en especies
vegetales (Kader, 1996), sugiere que estas proteínas pueden constituir un
nuevo tipo de panalergenos vegetales. Como en otras familias de proteínas
de defensa, su acumulación en tejidos específicos, y su inducción por estrés
biótico o abiótico (García-Olmedo et al., 1995; Kader, 1996), hace esperar
que se produzcan cambios sustanciales en los niveles de alérgenos
dependiendo de las condiciones de recolección y almacenamiento, así como
de la parte del alimento que se analice (p.e. piel o pulpa en frutos;
Fernández-Rivas y Cuevas, 1999). Otras propiedades de las LTPs, como
son su estabilidad térmica (Brenna et al., 2000), o el mantener su capacidad
de ligar IgE después de ser tratadas con pepsina (Asero et al., 2000),
apoyan su potencial papel como alérgenos.
Con un escenario como el anteriormente expuesto, sorprende que no
haya estudios que comparen la actividad alergénica de LTPs purificadas de
diferentes especies vegetales no relacionadas filogeneticamente, y aún más,
que no se haya publicado ningún estudio de ensayos in vivo (pruebas
cutáneas) utilizando estas proteínas. La purificación de LTPs de manzana,
melocotón, polen de artemisa y semilla de castaña se ha llevado a cabo en
este trabajo tratando de cubrir estos objetivos. El aislamiento de las LTPs de
las dos últimas fuentes se basó en la sensibilización a polen de artemisa de
un número significativo de pacientes alérgicos a frutos de rosáceas
(Hernández et al., 1995; Cuesta-Herranz et al., 1999), y en la asociación
clínica de la castaña con algunos casos de alergias a frutas, como manzana
o melocotón (Hernández et al., 1995), así como en la distancia filogenética
entre A. vulgaris, C. sativa y especies de la familia Rosaceae.
Los polipéptidos purificados de artemisa (Art v 3) y castaña (Cas s 3)
(Fig. 31 y 32) tenían un tamaño molecular similar al de Pru p 3 y Mal d 3,
pero la determinación de su secuencia aminoterminal mostró que, aún
perteneciendo a la familia de LTPs, su similitud con los homólogos de
104

Discusión
rosáceas (53-43% de identidad en 35 residuos; Fig. 33), así como entre
ambos (50%), era muy inferior a la de Pru p 3 con Mal d 3 (81%).
A pesar de la baja similitud de secuencia, tanto la LTP de polen de
artemisa como la de castaña, fueron reconocidas por una mezcla de sueros
de pacientes alérgicos a frutas de rosáceas, en la que previamente se había
confirmado su reactividad frente a las LTPs de manzana y melocotón (Fig.
34). Sorprendentemente, el polipéptido aislado de castaña mostró una
reactividad con estos sueros aún mayor que las de los alérgenos de
melocotón y manzana. La base estructural de este resultado, contradictorio
con los que se comentarán a continuación, y confirmado con dos
preparaciones distintas de Cas s 3 en tres ensayos diferentes, es, hasta el
momento, desconocida.
En contraste con los datos de inmunodetección, los ensayos de
inmunoinhibición indicaron un comportamiento diferencial de las cuatro LTPs
purificadas. La inhibición de la unión a IgE por la banda correspondiente a
Pru p 3 en el extracto de melocotón, fue completa al utilizar dicha proteína
como inhibidor, mientras que Art v 3 y Cas s 3 no mostraron ninguna
reducción significativa. Un efecto similar al de Pru p 3 había sido
previamente demostrado para Mal d 3 (Sánchez-Monge et al., 1999). La
diferente capacidad de unir IgE in vitro por las LTPs aisladas se confirmó
mediante dos líneas de evidencia complementarias. Los valores de IgE
específica para las cuatro proteínas fueron mayores para Pru p 3 y Ma! d 3
que para Art v 3 y Cas s 3 (un 25% del nivel de Pru p 3 en los dos últimos
casos; Tabla 13). Por otro lado, ensayos de inhibición por ELISA (Tabla 14)
revelaron que cuando los alérgenos de manzana o melocotón se utilizaban
como inhibidores, se anulaba entre un 83 y 98% la unión de IgE por las
cuatro LTPs purificadas, mientras que Art v 3 y Cas s 3 sólo afectaban
parcialmente (63 a 40%) a esta unión.
Los resultados anteriores prueban que hay una reactividad cruzada
parcial entre las LTPs de rosáceas y las purificadas de polen de artemisa y
semilla de castaña. Parece que las dos últimas LTPs comparten algunos
105

Discusión
epítopos IgE con las de manzana y melocotón, pero que carecen de otros
importantes que están presentes en los alérgenos de frutos de rosáceas.
La purificación de cuatro LTPs con distintos niveles de identidad de
secuencia entre ellas, permitió también evaluar las diferencias en la
respuesta in vivo (pruebas cutáneas) de diferentes miembros de esta familia
de proteínas (Tabla 15). Las pruebas se realizaron en un grupo de 47
pacientes seleccionados por una historia clínica que sugería una alergia a
melocotón. De ellos, 24 habían sufrido reacciones adversas con manzana y
10 con castaña. La mayoría (39) tenían diagnosticada polinosis no asociada
con Bet v 1.
Las LTPs de melocotón y manzana fueron reconocidas por la mayoría
de los pacientes (91,5% y 76,6% respectivamente). Sólo 3 pacientes no
reaccionaron positivamente con la LTP de melocotón, 2 de los cuales
tampoco lo hicieron cuando se les realizó la prueba cutánea con el extracto
crudo de este fruto. Las respuestas positivas a Mal d 3 se elevan a un 92%
cuando se consideran únicamente los 24 pacientes con historia clínica
positiva a manzana. Estos resultados confirman que las LTPs son alérgenos
principales (reconocimiento >90%) en pacientes españoles alérgicos a
manzana y melocotón. Además, la alta sensibilidad de las pruebas cutáneas
utilizando las proteínas purificadas, similar a la obtenida con fruta fresca
{príck-prick test), es un dato prometedor para la utilización futura de las
mismas en ensayos diagnósticos in vivo de este tipo de alergia.
La respuesta positiva a la LTP de artemisa se observó en 17
pacientes (36%). Sin embargo, cuando se seleccionan sólo aquellos que
mostraron reactividad cutánea con el extracto de este polen (27/47 casos), la
cifra anterior alcanza el 55%. Por último. Cas s 3 fue reconocida por 11
pacientes (23%), 10 de los cuales eran los únicos del grupo estudiado que
presentaron una historia clínica positiva a castaña. De nuevo, si sólo se
consideran aquellos casos con respuesta positiva al extracto crudo de
castaña, las reacciones positivas a Cas s 3 representan un porcentaje muy
superior (61%).
106

Discusión
Los resultados del estudio in vivo confirman la reactividad cruzada
parcial entre las cuatro LTPs purificadas. En este contexto, los diferentes
patrones de reacción observados en el grupo de pacientes analizados
pueden sugerir aspectos adicionales del co-reconocimiento de estas
proteínas. La pauta más frecuente (46%), incluye únicamente a las dos LTPs
de rosáceas, lo que es consistente con la alta similitud estructural entre
ambos alérgenos, y con la asociación clínica entre las alergias a manzana y
melocotón. El segundo patrón (21%) supone un reconocimiento de las cuatro
proteínas, y un tercero agrupa a pacientes que reconocieron a Pru p 3, Mal d
3 y Art V 3, pero no a Cas s 3 (10%). Un dato sorprendente es que sólo se
obtuvieron pruebas positivas a Cas s 3 en aquellos pacientes que
reaccionaron con Art v 3, y que hay una correlación significativa entre las
pruebas cutáneas a estas dos LTPs (r=0,928; p< 0,001). Puede especularse
que se requiere un reconocimiento de Pru p 3 para que reaccione Art v 3, y
que el reconocimiento de este alérgeno es un factor necesario para extender
la reacción a Cas s 3.
Un estudio reciente de Asero et al. (2000) aporta datos adicionales a
los presentados en esta memoria sobre la reactividad cruzada entre LTPs de
diferentes especies. La evidencia aportada, aún siendo preliminar, sugiere
que estos polipéptidos pueden estar implicados en' la cosensibilización a
frutas de rosáceas y a un amplio número de alimentos, incluidos zanahoria,
apio, tomate, melón, kiwi, nuez, cacahuete y cereales. El establecer una
base experimental sólida que permita definir a las LTPs como una relevante
familia de panalergenos vegetales, requiere un conocimiento más profundo
de las relaciones estructurales y de los epítopos que comparten los
miembros de esta familia con mayor interés en el campo de la alergia. El
aislamiento y secuenciación de clones ADNc correspondientes a Pru p 3 y
í\/lal d 3 (Fig. 35) es un primer paso para cubrir estos objetivos. Las
estructuras primarias deducidas han corroborado la estrecha relación entre
ambos alérgenos, así como con sus homólogos, potencialmente reactivos,
de otra rosáceas (cereza, pera, almendra), y han permitido estimar la
divergencia con LTPs presentes en otros alimentos y pólenes (67 a 29%, de
107

Discusión
identidad de secuencia; Fig. 36). La obtención de las proteínas
recombinantes y el modelado de su estructura terciaria están abordándose
actualmente.
108

2iesu7nen y
concñisíones

Resumen y conclusiones
1.- Las quitinasas de clase I son alérgenos principales de aguacate, castaña
y plátano, tres de los alimentos más asociados clínicamente con el síndrome
látex-frutas. Los alérgenos purificados tienen un alto grado de similitud de
secuencia, y unos dominios N-terminales estrechamente relacionados con la
heveína de látex. Por el contrario, las quitinasas de clase II de estos
alimentos, con dominios catalíticos homólogos a los de clase I pero sin el
dominio heveína N-terminal, no muestran actividad alergénica ni en ensayos
in vitro ni en pruebas cutáneas in vivo.
2.- Otra serie de alimentos, relacionados en distinta medida con la alergia a
látex, como chirimoya, fruta de la pasión, kiwi, papaya, mango, tomate y
trigo, contienen también putativas quitinasas de clase I que son reconocidas
por sueros de pacientes alérgicos con el síndrome látex-frutas. Estas
enzimas parecen ser los panalergenos responsables de las reacciones
cruzadas descritas en dicho síndrome. Los N-oligosacáridos complejos,
implicados en algunos tipos de cosensibilización, no son relevantes en el
contexto de la asociación látex-frutas. La falta de reconocimiento de las
quitinasas de clase I por sueros de pacientes alérgicos a látex pero no a
frutas, indica que no juegan un papel específico en casos de alergia a látex
no relacionada con la ingestión de frutos.
3.- La judía verde, un alimento no asociado a la sensibilización a látex,
posee una quitinasa de clase I con una capacidad alergénica similar a la del
alérgeno principal de aguacate. La expresión de dicha quitinasa es inducible
por etileno. El tratamiento térmico provoca una pérdida completa de la
reactividad alergénica de las quitinasas de judía verde y aguacate. Este
hecho podría explicar por qué los alimentos consumidos en fresco son los
principalmente asociados al síndrome látex-frutas, mientras aquellos que
sufren un tratamiento térmico antes de ser ingeridos, aun conteniendo
quitinasas reactivas, no suelen relacionarse con la alergia a látex.
110

Resumen y conclusiones
4.- Se ha expresado la quitinasa alergénica de castaña y su dominio
catalítico en Pichia pastoris. La enzima recombinante completa tiene una
capacidad de ligar IgE específica similar a la del alérgeno natural, mientras
que el dominio catalítico recombinante no reacciona con sueros de pacientes
alérgicos. El dominio heveína de las quitinasas de clase I juega, portante, un
papel esencial en su reactividad como alérgenos. Sin embargo, otros
dominios heveína, como los de la aglutinina de germen de trigo, no son
reconocidos por los mismos sueros de pacientes alérgicos.
5.- Las LTPs de polen de artemisa y de semilla de castaña, dos fuentes
alergénicas relacionadas a diferente nivel con la alergia a frutas de rosáceas
en poblaciones mediterráneas, tienen alrededor de un 50% de identidad de
secuencia con los alérgenos homólogos de manzana y melocotón, con los
que además presentan reactividad cruzada. Sin embargo, se han observado
diferencias significativas en la capacidad de unir IgE específica entre estos
cuatro miembros de la familia de las LTPs, siendo las proteínas de rosáceas
considerablemente más activas.
6.- Las LTPs de melocotón y manzana provocan pruebas cutáneas positivas
en más de un 90% de pacientes con una historia clínica previa que sugiere
sensibilización a la ingestión de estos frutos. En el mismo grupo de
pacientes, aunque la reactividad de las LTPs de artemisa y castaña es
mucho menor, más de la mitad de los que muestran prueba cutánea positiva
al extracto de polen, y todos lo que tienen síntomas clínicos a castaña,
reaccionan positivamente con las LTPs correspondientes. Los patrones de
reactividad in vivo confirman, en términos generales, las reacciones
cruzadas observadas in vitro, así como el potencial papel de las LTPs como
panalergenos vegetales.
7.- El aislamiento y secuenciación de dones de ADNc de las LTPs de
melocotón y manzana muestra la estrecha relación estructural entre ambos
alérgenos, así como con las proteínas homologas de otras especies de
111

Resumen y conclusiones
rosáceas. La identidad de secuencia con otros alérgenos vegetales de la
misma familia, como el del polen de Parietaría judaica, es sensiblemente
menor, alcanzando cifras inferiores al 30%.
112

SuTfiTfiary

Summary
Class I chitinases are relevant allergens of avocado, chestnut and banana,
three plant foods closely associated with the latex-fruit syndrome. The isolated
allergens show a high degree of sequence similarity, and N-terminal domains
homologues to látex heveln. By contrast, their class II counterparts without N-
termlnal heveln-like domaln, exert allergenic activity nelther in vitro ñor in vivo.
Other plant foods dlfferentially implicated In the syndrome, such as cherimoya,
passion fruit, kiwi, papaya, mango, tomato and flour wheat, also contaln putative
class i chitinases which are recognized by sera from patiens with látex and fruit
allergy. Thus, these enzymes seem to be the panallergens responsible for the
latex-fruit syndrome. By contrast, cross-reactive carbohydrate determinants are
not important structures in the context of these cross-sensitizatlon. The lack of
recognition of class I chitinases by sera from patiens allergic to látex but not to
fruit, indicates that they do not play a specific role ¡n allergy to látex without fruit
sensitization. Green beans, a food not associated with látex allergy, contain a
class I chitinase with an allergenic capacity similar to that of the major avocado
allergen. This bean protein is strongly induced by ethylene treatment. The
allergenic activity of both green bean and avocado chitinases seems to be
compietely lost by heating. This fact could explain why plant foods containing
these putative allergens that are consumed after cooking, are not usually
associated with the látex fruit syndrome. The chestnut allergenic chitinase, as well
as its catalytic domain, have been expressed in Pichia pastoris. The full
recombinant enzyme shows and IgE-binding capacity similar to that of the natural
allergen, whereas the recombinant catalytic domain does not react with sera from
allergic patients. Therefore, the hevein-like domain of class I chitinases plays an
essential role in their allergenic reactivity. However, other hevein-like domains,
such as those present in wheat germ agglutinin, are not recognized by the sera
from patients with allergy to látex and fruits.
Lipid Transfer Proteins, LTPs, from mugwort pollen and chestnut seed, two
allergenic agents differentially related with allergy to Rosaceae fruits, show around
50% of sequence identity with their homologues from peach and apple. Although
the four proteins crossreact, very different IgE-binding capacities are found among
them, the fruit allergens being much more active. LTPs from peach and apple
114

Summary
provoke up to 90% of positive responses when skin-prick tested in patients with
positive clinical history to the corresponding fruit. Those from mugwort pollen and
chestnut seed show significantly lower levéis of reactlvity In the same group of
patients. However, all individuáis with positive clinical history to chestnut react with
its seed LTP, and about 50% of those with positive skin-prick-test to mugwort
pollen recognize the mugwort protein. Finally, the isolation and sequencing of
cDNA clones for peach and apple LTPs have allowed to confirm their alose
relationship, and with other homologous polypeptides from Rosaceae species. The
sequence identity level to other plant allergens belonging to the same protein
family, such as that from Parietaria pollen, is substantially lower reaching valúes
below 30%.
115

St^íío^rafta

Bibliografía
Aalberse RC and van Ree R (1997): Crossreactive carbohydrate determinants. Clin. Rev. Allergy Immunol., 15: 375-387.
Alenius H, Kalkkinen N, Lukka M, Reunala T, Turjanmaa K, Makínen-Kiljunen S, Yip E and Palosuo T (1995): Prohevein from rubber tree {Hevea brasiliensis) is a major látex allergen. Clin. Exp. Allergy, 24: 659-665.
Alenius H, Kalkkinen N, Reunala T, Turjanmaa K and Palosuo T (1996a): The main IgE-binding epitope of a major látex allergen, prohevein, is present in its N-terminal 43-amino acidfragment, hevein. J. Immunol., 156: 1618-1625.
Alenius H, Makinen-Kiljunen S, Ahlroth M, Turjanmaa K, Reunala T and Palosuo T (1996b): Crossreactivity between allergens in natural rubber látex and banana studied by immunoblot inhibition. Clin. Exp. Allergy, 26: 341-348.
Alenius H, Kalkkinen N, Turjanmaa K, Lauerma A, Reunala T, and Palosuo T (1996c): Cross-reactivity between wheat germ agglutinin (WGA) and prohevein, a major látex allergen [abstract]. J. Allergy Clin. Immunol., 97: 321.
Aliona I, Collada C, Casado R, Paz-Ares J and Aragoncillo C (1996): Bacterial expression of an active class Ib chitinase from Castanea sativa cotyiedons. Plant Mol. Biol., 32: 1171-1176.
Asero R (1999): Detection and clinical characterization of patients with oral allergy syndrome caused by stable allergens in rosaceae fruits and nuts. Ann. Allergy Asthma Immunol., 83: 377-383.
Asero R, Mistrello G, Roncarolo D, Vries SC, Gautier MF, Ciruana C, Verbeek E, Mohammadi R, Knul-Brettiova V, Akkerdaas JH, Bulder I, Aalberse RC and van Ree R (2000): Lipid Transfer Protein: A pan-allergen in plant-derived foods that is highiy resistant to pepsin digestión. Int. Arch. Allregy Immunol., 122: 20-32.
Asensio JL, Cañada FJ, Bruix M, Rodriguez-Romero A and Jiménez-Barbero J (1995): The interaction of hevein with A/-acetyIglucosamine containing oligosaccharides. Solution structure of hevein complexed to chitobiose. Eur J. Biochem., 230: 621-633.
Asensio JL, Cañada FJ, Bruix M, González C, Khiar N, Rodriguez-Romero A and Jiménez-Barbero J (1997): NMR investigations of protein-carbohydrate interactions: refined three-dimensional structure of the complex between hevein and methyl p-chitobioside. Glycobiology, 8: 569-577.
Astwood JD, Leach JN and Fuchs L (1996): Stability of food allergens in digestión in vitro. Nature Biotechnol., 14: 1269-1273.
Atkinson RG, Perry J, Matsui T, Ross SG and Macrae EA (1996): A stress-, pathogenesis-, and allergen-related cDNA in apple fruit is also hpening-related. J. Crop Horticult. Se/., 24: 103-107.
Batanero EF, Crespo JF, Monsalve Rl, Martin-Esteban M, Villalba M and Rodríguez R (1999): IgE binding and histamine-release capabilities of main carbohydrate component isolated from the major allergen of olive tree pollenOlee 1. J. Allergy Clin. Immunol., 103:147-153.
Beezhold DH, Sussman GL, Kostyal DA and Chang NS (1994): Identification of a 46 kDa látex protein allergen in health care worker. Clin. Exp. Immunol., 98: 408-413.
Beezhold DH, Sussman GL, Liss GM, and Chang NS (1996): Látex allergy can induce clinical reactions to specific foods. Clin. Exp. Allergy, 26: 416-422.
117

Bibliografía
Beezhold DH, Kostyal DA and Sussman GL (1997): IgE epitopes analysis of the hevein preprotein; a major látex allergen. Clin. Exp. Immunol., 108:114-121.
Blanco C, Carrillo T, Castillo R, Quiralte J and Cuevas M (1994): Látex aliergy: cíinical features and cross-reactivity with fruits. Ann. Aliergy, 73: 309-314.
Bousquet J, Lockey R, Mailing H, and the WHO Panel members (1998): Allergen immunotherapy: therapeutic vaccines for allergic disease: A WHO position paper. J. Aliergy Clin. Immunol., 102: 558-562.
Bradford MM (1976): A rapid and sensitive method for the quantification of microgram quantities of protein utiiizing the principie of protein- dye binding. Anal. Biochem., 72: 248-254.
Brehier R, Theissen U, Mohr C and Luger T (1997): "Latex-Fruit Syndrome": Frequency of cross-reacting IgE antibodíes. Aliergy, 52:404-410.
Breiteneder H, Pettenburger K, Bito A, Kraft D, Rumpold H, Scheiner O and Breitenbach M (1989): The gene coding for the major birch pollen allergen Bet v 1, is highiy homologous to a pea disease resistance response gene. EMBO J., 8: 1935-1938.
Breiteneder H, Hoffmann-Sommergruber K, O'Riordain G, Susani M, Ahorn H, Ebner C, Kraft D and Scheiner O (1995): IVlolecular characterization of Api g 1, the major allergen of celery {Apium graveolens), and ¡ts immunological and structural relationship to a group of 17-kD tree pollen allergens. Eur J. Biochem., 233: 484-488.
Breiteneder H and Scheiner O (1998): Molecular and immunological characteristics of látex allergens. Int. Arch. Aliergy Immunol., 116: 83-92.
Breiteneder H and Ebner C (2000): Molecular and biochemical classification of plant-derived food allergens. J. Aliergy Clin. Immunol., 106: 27-36.
Brenna O, Pompei C, Ortolani C, Pravettoni V, Farioli L and Pastorelo EA (2000): Technological processes to decrease the allergenicity of peach juice and néctar. J. Agrie. Food Chem., 48:493-497.
Broekaert W, Parijs J, Leyns F, Jóos H and Peumans W (1986): Comparison of some molecular, enzymatic and antifungal properties of chitinases from tomate, apple, tobáceo and wheat. Physiol. Mol. Plañí Pathol., 33: 319-331.
Broekaert W, Cammue BPA, de Bolle M, Thevissen K, Samblans G and Osborn R (1997): Antimicrobial peptidesfrom plants. Crit. Rev. Plant Sel., 16: 297-323.
Broglie K, Gaynor J and Broglie R (1986): Ethylene-regulated gene expression: molecular cloning of the genes encoding an endochitinase from Phaseolus vulgarís. Proc. Nati. Acad. Sci. USA; 83: 6820-6824.
Broglie K, Chet I, Holliday M, Cressman R, Biddie P, Knowlton S, Mauvais C and Broglie R (1991): Transgenic plants with enhanced resistance to the fungal pathogen Rhizoctonia solani. Science, 254: 4308-4314.
Brujinzeel-Koomen C, Ortolani C, Aas D, Binslev-Jensen C, Bjorksten B, Moneret-Vautrin D and Wuthrich B (1995): Adverse reactions to food. Aliergy, 50: 623-635.
Bufe A (1998): The biogical function of allergens: relevant for the induction of allergic diseases?. int Arch. Aliergy Immunol., 117: 215-219.
118

Bibliografía
Clendennen SK and May GD (1997): Differential gene expression in ripening banana fruit. Plant Physiol., 115: 463-469.
Colombo P, Kennedy D, Ramsdale T, Costa MA, Duro G, Izzo V, Salvador! S, Guerrini R, Cocchíara R, Mirisola MG, Wood S and Geraci D (1998): Identification of an immunodonninant IgE epitope of Parietaria judaica nnajor allergen. J. Immunol., 160: 2780-2785.
Collada C, Casado R, Fraile A and Aragoncillo C (1992): Basic endcchitinases are major proteins in Castanea sativa cotyiedons. Plant Pliysioi., 100: 778-783.
Collinge DB, Kragh KM, Míkkelsen JD, Nielsen KK, Rasmussen U and Vad K (1993): Plant chitinases. Plant J., 3: 31-40.
Cookson W (1999): The ailiance of genes and environment in asthma and allergy. Nature, 402(suppl):B5-B11.
Corry DB and Kheradmand F (1999): Induction and regulation of the IgE response. Nature, 402(suppl.): B18-B22.
Crespo JF, Pascual C, Burks AW, Helm RM, and Esteban MM (1995): Frequency of food allergy in a pediatric population from Spain. Paedritr. Allergy Immunol., 6: 39-43.
Cuesta-Herranz J, Lázaro M, Martínez A, Fígueredo E, Palacios R, de las Heras M and Martínez J (1999): Rollen allergy in peach-allergic patients: sensitization and cross-reactivity to taxonomically unrelated pollens. J. Allergy Clin. Immunol., 104: 688-694.
Curíoní A, Santuccí B, Crístando A, Canistraci C, Pietravalle M, Simomato B and Gianmattasio M (1999): Urticaria from beer: an inmediate hypersensitivity reaction due to a 10-l<Da protein derived from barley. Clin. Exp. Allergy, 29: 407-413.
Chen Z, Posch A, Raulf-Heimsoth M, Ruhs HP and Baur X (1997a): Purification of rubber elongation factor from Hevea brasiliensis (Hev b 1) that acts as a major allergen in látex allergic patients with spina bifida. Allergy, 52: 79.
Chen Z, Posch A, Lohaus C, Raulf-Heímsoth M, Meyer HE and Baur X (1997b): Isolation and Identification of hevein as major IgE-binding polypeptide in Hevea látex. J. Allergy Clin. Immunol., 99: 402-409.
Chen A, Posch A, Cremer R, Raulf-Heímsoth M and Baur X (1998): Identification of hevein (Hev b 6.02) in Hevea látex as major cross-reacting allergen with avocado fruit in patients with látex allergy. J. Allergy Clin. Immunol., 102: 476-481.
Choi JK, Yoon SH, Hong HY, Choí DK and Yoo GS (1996): A modified Coomassie Blue staining of proteins in polyacrilamide geis with Bismark Brown R. Anal. Biochem., 236: 82-84.
Delbourg MF, Guilloux L, Moneret-Vuatrín DA and Ville G (1996): Hypersensitivity to banana in latex-allergic patients. Identification of two major banana allergens of 33 and 37 kD. Ann. Allergy Asthma Immunol., 76: 321-326.
Devíller P and Paulí G (1997): Crossreactions ¡nvolving plant allergens. Clin. Rev. Allergy Immunol., 15:405-413.
Duro G, Colombo P, Costa MA, Izo V, Porcasí R, Di Fiore R, Locorotondo G, Cocchíara R and Geraci D (1997): Isolation and characterization of two cDNA clones coding for isoforms of the Parietaria judaica major allergen Par J 1.0101. Int. Arch. Allergy Immunol., 112:348-355.
119

Bibliografía
Ebner C, Birkner T, Valenta E, Rumpold H, Breitenbach M, Scheniner O and Kraft D (1991): Common epitopes of birch pollen and apples. Studies by western and northern blot. J. Allergy Clin. ImmunoL, 88: 588-595.
Ebner C, Jensen-Jarolím E, Leitner A and Breiteneder H (1998): Characterization of allergens in plant-derived: Apiaceae spices, pepper (Piperaceae), and paprika (bell peppers, Solanaceae). Allergy, 53: 52-54.
Fernandez-Rivas M, van Ree R and Cuevas M (1997): Allergy to Rosaceae fruits without related pollinosis. J. Allergy Clin. Immunol., 100: 728-733.
Fernandez-Rivas M and Cuevas M (1999): Peéis of rosaceae fruits have a higlier allergenicity than pulps. Clin. Exp. Allergy, 29: 1239-1247.
Fritsch R, Ebner C and Kraft D (1997): Allergenic crossreactivities. Pollen and vegetable foods. Clin. Rev. Allergy Immunol, 15: 397-404.
Fuchs E (1979): Clinical diagnosis of allergy. Prax. Klin. Pneumol., 33: 268-276.
Fuchs T, Spitzauer S, Vente C, Hevier J, Kapíotis S, Rumpold H, Kraft K and Valenta R (1998): Natural látex, grass pollen, and weed pollen share IgE epitopes. J. Allergy Clin. Immunol, 100: 356-364.
García-Casado G, Sanchez-Monge R, Chrispeeis MJ, Armentia A, Salcedo G and Gómez L (1996a): Role of complex asparagine-linked glycans in the allergenicity of plant glycoproteins. Glycobiology, 6: 471-477.
García-Casado G, Armentia A, Sanchez-Monge R, Malpíca JM and Salcedo G (1996b): Rye flour allergens associated with baker's asthma. Correlation between in vivo and in vitro activities and comparison with their wheat and barley homologues. Clin. Exp. Allergy, 26: 428-435.
García-Casado G, Collada C, Aliona I, Casado R, Ráelos L, Aragoncíllo C and Gómez L (1998): Site-directed mutagenesis of active site residues in a class I endochitinase from chestnut seeds. Glycobiology, 8: 1021-1028.
García-Olmedo F, Molina A, Segura A and Moreno M (1995): The defensive role of non-specific lipid-transfer proteins in plants. Trends Microbio!., 3: 72-74.
Gídrol S, Chrestun H, Tan HL and Kush A (1994): Hevein a lectin like protein from Hevea brasiliensis (rubber tree) is involved in the coagulation of látex. J. Biol. Chem., 269: 9278.
González R, Valera J, Carreira J and Polo F (1995): Soybean hydrophobic protein and soybean hull alergy. Lancet, 346: 48-49.
Gultekin H and Heermann KH (1988): The use of polyvinylidenofluoride membranes as a general blotting matrix. Anal. Biochem., 172: 320-329.
Grísson R, Grezes-Besset B, Schneider M, Lucante N, Olsen L, Leguay J and Toppan A (1996): Field tolerance to fungal pathogens of Brassica napus constitutively expressing chimeric chitinase gene. Nature Biotechnol, 14: 643-646.
Hanninen AR, Mikkola JH, Kalkkinen N, Turjanmaa K, Ylitalo L, Reunala T and Palosuo T (1999): Increased allergen production in turnip {Brassica rapa) by treatments activating defense mechanisms. J. Allergy Clin. Immunol, 104:194-201.
120

Bibliografía
Hernández J, Garcia-Selles FJ, Pagan JA and Negro JM (1995): Hipersensibilidad inmediata a frutas y verduras y polinosis. Mergo!. Immunopathol., 13; 197-211.
Hirs GHW (1967): Performic acid oxidation. Meth. Enzymoí.AV. 197-199.
Hoffman LM and Donaldson DD (1985): Characterization of two Phaseolus vulgaris phytohemagglutinin genes closeiy iinked on the chromosome. EMBO J., 4: 883-889.
Hoffmann-Sommergruber K, O'Riordain G, Ahrn H, Ebner C, Laimer da Cámara Machado M, Puhringer H, Scheiner O and Breiteneder H (1999): Molecular characterization of Dau c 1, the Bet v 1 homoiogous protein from carrot and its cross-reactivity with Bet v 1 and Api g 1. Clin. Exp. Allergy, 29: 840-847.
Hoffmann-Sommergruber K (2000): Plant allergens and patogenesis-related proteins. Int. Arch. Allergy Immunol., 122:155-166.
Hseih LS, Moos M and Lin Y (1995): Characterization of appíe 18 and 31 kd allergens by microsequencing and evoíution of their content during storage and ripening. J. Allergy Clin. Immunol., 96: 960-970.
Inschlag C, Hoffmann-Sommergruber K, O'Riordain G, Ahorn H, Ebner C, Scheiner O and Breiteneder H (1998): Biochemical characterization of Pru a 2, a 23 kD thaumatin-like protein representing a potential major allergen in cherry {Prunus avium). Int. Arch. Allergy Immunol., 116:22-28.
Ipsen H and Hansen OC (1991): The NH2-terminal amino acid sequence of the immunochemically partial identical major allergens of Alder {Alnus glutinosa) Aln g I, birch {Betula verrucosa) Bet v I, hornbeam (Carpinus betulus) Car b I and oak {Quercus alba) Que a I pollens. Mol. Immunol.; 28: 1279-1288.
Ishizaka K, Ishizaka T and Hornbrook MIVI (1966): Physicochemical properties of human reagric antibody. J. Immunol., 97: 840-845.
Jach G, Gormhardt B, Mundy J, Logemann J, Pinsdorf E, Leah R, Schell J and IVIaas C (1995): Enhanced quantitative resistance against funga! disease by combinatorial expression of different bariey antifungal proteins in transgenic tabacco. Plant J., 8: 97-109.
James JM, Sixbey JP, Heím RM, Bannon GA and Burks AW (1997): Wheat a-amy!ase inhibitor: a second route of allergic sensitization. J. Allergy Clin. Immunol., 99: 239-244.
Jeckel PA, Hartmann JBM and Beintema JJ (1991): The primary structure of hevamine, and enzyme with lysozime/chitinase activity from Hevea brasiliensis látex. Eur. J. Biochem., 200: 123-130.
Johansson SGO and Bennich H (1967): Inmunological studies of an atypical (myeloma) immunogiobulin. Immunology, 13: 381-394.
Kader JO (1996): Lipid-transfer proteins: a puzziing family of plant proteins. Trends Plant Sci., 3: 66-70.
Kay AB (1997): Allergy and allergic diseases. Oxford: BlackweII Science.
Kazemi-Shirazi L, Pauli G, Purohit A, Spitzener S, Frosch! R, Hoffman-Sommergruber K, Breiteneder H, Scheiner O, Kraft D and Valenta R (2000): Quantitative IgE inhibition experiments with purified recombinant allergens indícate pollen-derived as the sensitizing agents responsible from many forms of plant food. J. Allergy Clin. Immunol., 105: 116-125.
121

Bibliografía
Kostyal DA, Hickey VL, Noti JD, Sussman GL and Beezhold DH (1998): Cloning and characterization of a látex allergen (Hev b 7): homology to patatin, a plant PLA(2). Clin. Exp. Immunol., 112: 355-362.
Kusaba-Nakayama M, Ki M, Iwamoto M, Shibata R, Sato M and Imaizumi K (2000): CMS one of the wheat a-amylase inhibitor subunits, and binding of IgE in sera from japanese with atopic dermatitis relatad to wheat. Food Chem. Toxicid., 38: 179-185.
Laemmli UK (1970): Cieavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
Lagrimini LM, Burkhart W, Moyer M and Rothstein S (1987): Molecular cloning of complementary DNA encodig the lignin forming peroxidases from tabacco: molecular analysis and tissue-specific expression. Proc. Nati. Acad. Sai. USA, 84: 7542-7546.
Lavaud F, Prevost A, Cossart C, Guerin L, Bernard J and Kochman S (1995): Allergy to látex, avocado pear, and banana: evidence for a 30 kDa antigen in immunobiotting. J. Allergy Clin. Immunol., 95: 557-564.
Lavaud F, Sabouraud D, Deschampas F and Pedu D (1997): Crossreactions involving natural rubber látex. Clin. Rev. Aliergy Immunol., 15: 429-446.
Lauriere M, Lauriere C, Chrispeeis MJ, Johnson KD and Sturm A (1989): Characterization of a xylose-specific antiserum that reacts with the complex asparagine-linked glycans of extraceliular and vacuolar glycoproteins. Plant Physiol., 90: 1182-1188.
Levy DA, Mounedji N, Noirot C and Leynadier F (2000): Allergic sensitization and clinical reactions to látex, food and poílen in aduit patiens. Clin. Exp. Allergy, 30: 270-275.
Mauch F, Mauch-Mani B and Boller T (1988): Antifunga! hydrolases in pea tissue. II. Inhibition of fungal growth by combination of chitinases and p-1,3-glucanases. Plant Physiol., 88: 936-942.
MeJns F, Fritíng B, Linthorst H, Mikkelsen JD, Neuthaus J and Ryals J (1994): Plant chitinases genes. Plant Mol. Biol. Rep., 12: 522-528.
Melchers LS and Striver MH (2000): Novel genes for disease-resistance breeding. Curr. Op//7.P/aA7fB/o/., 3:147-152.
Mikkola JH, Aleníus H, Kalkkinen N, Turjanmaa K, Palosuo T and Reunala T (1998): Hevein-lil<e protein domains as a possible cause for allergen cross-reactivity between látex and banana. J. Allergy Clin. Immunol., 102: 1005-1012.
Molano J, Duran A and Cabid E (1970): A rapid and sensitive assay for chitinase using tritiated chitin. Anal. Biochem., 83: 648: 656.
Molina A and García-Olmedo F (1993a): Developmental and pathogen-induced expression of three barley genes encoding lipid transfer protein. Plant J., 4: 983-991
Molina A, Segura A and Garcia-Olmedo F (1993b): Lipid transfer proteins (nsLTP) from barley and maize leaves are potent inhibitors of bacterial and fungal plant pathogens. FEBS Lefí., 316: 119-122.
Navarro AM, Orta JO, Sánchez MC, Delgado J, Barber D and Lombardero M (1997): Primary sensitization to Morus alba. Allergy, 52: 1144-1145.
Neuhaus J-M, Friting B, Linthorst HJM, Meins F, Mikkelsen JD and Ryals J (1996): A revised nomenclature for chitinase genes. Plant Mol. Biol. Rep., 14: 102-104.
122

Bibliografía
Niggemann B and Breiteneder H (2000): Látex allergy in children. Int. Arch. Allergy Immunol., 121: 98-107.
Olmsted JB (1986): Analysis of cytoskeletal structure using blot-purified monospecific antibodies. Methods Enzymol, 134: 467-472.
Ortoiani C, Ispano M, Pastorello E, Bigi A and Ansaloni R (1988): The oral allergy syndrome. Ann. Allergy, 61: 47-52.
Pastorello EA, Ortoiani C, Farioli L, Pravettoni V, Ispano M, Borga A, Bengtsson A, Incorvaia C, Berti C and Zanussi C (1994): Alíergenic cross-reactivity among peach, apricot, plum, and cherry in patients with oral allergy syndrome: an in vivo and in vitro study. J. Allergy Clin. Immunol., 94: 699-707.
Pastorello EA, Incorvaia C, Pravettoni V and Ortoiani C (1997): Crossreactions in food allergy. Clin. Rev. Allergy Immunol., 15: 415-427.
Pastorello EA, Incorvaia C, Pravettoni V, Farioli L, Conti A, Vigano G, Rivolta F, Ispano M, Rotondo F and Ortoiani C (1998): New allergens in fruits and vegetables. Allergy, 53 (suppl): 48-51.
Pastorello EA, Farioli L, Pravettoni V, Ortoiani C, Ispano M, Monza M, Barogiio C, Scibola E, Ansaloni R, Incorvaia C and Conti A (1999a): The major allergen of peach {Prunus pérsica) is a lipid transfer protein. J. Allergy Clin. Immunol., 103: 520-526.
Pastorello EA, Pravettoni V, Farioli L, Ispano M, Fortunato D, Monza M, Giuffrida MG, Rivolta F, Scibola E, Ansaloni R, Incorvaia C, Conti A and Ortoiani C (1999b): Clinical role of a lipid transfer protein that acts as a new apple-specific allergen. J. Allergy Clin. /mmt/no/., 104: 1099-1106.
Pastorello EA, Ortoiani C, Barogiio C, Pravettoni V, Ispano M, Giuffrida G, Fortunato D, Farioli L, Monza M, Napolitano L, Saxxo M, Scibolda E and Conti A (1999c): Complete amino acid sequence determination of the major allergen of peach {Prunus pérsica) Pru p 1. Biol. Chem., 380: 1315-1320.
Pastorello EA, D'Ambrosio FP, Pravettoni V, Farioli L, Giuffrida G, Monza M, Ansaloni R, Fortunato D, Scibola E, Rivolta F, Incorvaia C, Bengtsson A, Conti A and Ortoiani C (2000): Evidence for a lipid transfer protein as the major allergen of apricot. J. Allergy Clin. Immunol., 105: 371-377.
Petersen A, Vieths S, Aulepp H, Schlaak M and Becker W-M (1996); Ubiquitous structures responsible for IgE cross-reactivity between tomato fruit and grass pollen allergens. J. Allergy Clin. Immunol., 98: 805-815.
Posch A, Chen Z, Raulf-Heimsoth M and Baur X (1998): Látex allergens. Clin. Exp. Allergy, 28: 134-140.
Posch A, Wheeler CH, Chen Z, Flagge A, Dunn MJ, Papenfuss F, Raulf-Heimsoth M and Baur X (1999): Class i endochitinase containing a hevein domain is the causative allergen in latex-associated avocado allergy. Clin. Exp. Allergy, 29: 667-672.
Potter S, Uknes S, Lawton K, Winter AM, Chandier D, DiMaio J, Novitzky R and Ryals J (1993): Regulation of a hevein-like gene in Arabidopsis. Mol. Plant Microbe Interact, 6: 680-685.
123

Bibliografía
Rodríguez J, Crespo JF, Lopez-Rubio A, Cruz-Bertolo J, Fernando-Vivas P, Vives R and Daroca P (2000): Clinical cross-reactivity among foods of the Roseaceae family. J. AllergyCHn. Immunol., 106: 183-189.
Romagnani S (1998): The Th1/Th2 paradigm and allergic disorders. Allergy, 53 (suppl. 46): 12-15.
Salcedo G, Diaz-Perales A and Sancliez-IVIonge R (1999): Fruit allergy: plant defensa proteins as novel potential panallergens. Clin. Exp. Allergy, 29: 1158-1160.
Sanchez-Monge R, Lombardero M, Garcia-Selles FJ, Barber D and Salcedo G (1999): Lipid-transfer proteins are relevant ailergens in fruit allergy. J. Allergy Clin. Imn^unol., 103: 514-519.
Shakib F, Schuiz O and Svi ell H (1998): A mite subversiva: cleavage of CD23 y CD25 by Darp 1 enhances allergenicity. Immunol. Today, 117: 215-219.
Shal<ib F and Gough L (2000): The proteolytic activity of Der p 1 selectively enhances IgE synthesis: a link between allergenicity and cysteine protease activity. Clin. Exp. Allergy, 30: 751-752.
Scheurer S, Metzner K, IHanstein D and Vieths S (1997): Molecular cloning, expression and characterization of Pru a 1, the major cherry allergen. Mol. Immunol., 34: 619-629.
Schiumbaum A, Mauch F, Vogeli U and Bolier T (1986): Plant chitinases are potent inhibitors of fungal growth. Natura, 324: 365-367.
Sliewry PR and Lucas A (1997): Plant proteins that confer resistance to pest and pathogens. Adv. Bol Res., 26: 135-192.
Slater JE (1997): Látex allergy. IN: Kay AB, editor. Allergy and allergic disease. Oxford. BlackweII Science; p. 981-983.
Sloan AE and Powers ME (1986): A perspective on popular perceptions of adverse reactions to food. J. Allergy Clin. Immunol., 78: 127-133.
Smedley (2000): Occupational látex allergy: the magnitude of the probiem and its prevention. Clin. Exp. Allergy, 30: 458-460.
Smith PK, Krohn Rl, Hermanson GT, IVIallia AK, Gartner FH, Provenzano IVID, Fujimoto EK, Goel<e NM, Olson BJ and Klenk DC (1985): Measurement of protein using bicinchoninic acid. Anal. Biochem., 150: 76-85.
Soedjanaatmadja UMS, Subroto T, and Beijte MA (1995): Processed producís of the hevein precursor in the látex of the rubber tree {Hevea brasiliensis). FEBS Lett, 363: 211-213.
Sowka S, Li-Shan H, Krebitz lU!, Akasawa A, IVIartin BIVI, Starrett D, Petervauer CK, Scheiner O and Breiteneder H (1998a): Indentification and cloning of Prs a 1, a 32 kDa endochitinase and major allergen of avocado in the yeast Pichia pastoris. J. Biol. Chem., 273: 28091-28097.
Sowka S, Wagner S, Krebitz Wl, Arija-Mad-Arif S, Yusof F, Kinaciyan T, Brehier R, Sclieiner O and Breiteneder H (1998b): cDNA cloning of the 43-kDa látex allergen Hev b 7 with sequence similarity to patatins and its expression in the yeast Pichia pastoris. Eur. J. B/oc/ie/77., 255: 213-219.
124

Bibliografía
Stanley JS and Bannon GA (1999): Biochemistry of food allergens. Clin. Rev. Allergy Immunol., 17:279-291.
Sussman Gl and Beezhoid DH (1995): Allergy to látex rubber. Ann. Int. Med., 122: 43-46.
Taylor SL (1997): Food from genetically modified organisms and potential for food allergy. Env. Toxicol. Pharmacol., 4: 121-126.
Toriyama K, Hanaoka K, Okada T and Watanabe M (1998): Molecular cloning of a cDNA encoding a pollen extracellular protein as a potential source of a pollen allergen in Brassica rapa. FEBS Lett., 424: 234-238.
Towbin H, Staehelin T and Gordon J (1974): Electrophoretic transfer of proteins from poiyacrilamide geis to nitro-cellulose sheets: procedure and some applications. Proc. Nati. Acad. Sel. USA, 76: 4350-4354.
Turjanmaa K, Alenius H, Makinen-Kiljunen S, Reunala T and Palosuo T (1996): Natural rubber látex allergy. Allergy, 51: 593-602.
Valenta R, Breiteneder H, Pettenburger K, Breitenbach M, Rumpold H, Kraft D and Scheiner O (1991): Homoiogy of the major birch pollen allergen, Bet v 1, with the major pollen allergens of alder, hazeinut and hornbean at the nucleic acid level as determinad by cross-hybridation. J. Allergy. Clin, immunol., 87: 677-682.
Valenta R, Duchene M, Ebner C, Valent P, Sillaber C, Deviller, Ferrerira F, Fejkl M, Eddelmann H, Kraft D and Scheiner O (1992): Profilins constitute a novel family of functional plant pan-allergens. J. Exp. Med., 2: 377-381.
Valenta R, Lídholm J, Niederberger V, Hayek B, Kraft D and Gronlund H (1999a): The recombinant allergen-based concept of component-resolved diagnostics and immunotherapy. Clin. Exp. Allergy, 29: 896-904.
Valenta R, Vrtala S, Focke-Tejkl M, Bugajska-Schretter A, Ball T, Twardosz A, Spitzauer S, Gronlund H and Kraft D (1999b): Genetically engineered and synthetic allergen derivates: candidates for vaccination against type i allergy. Biol. Chem., 380: 815-824.
Van Damme JM, Charels D, Roy S, Tierens K, Barre A, Martins JC, Rouge P, van Leuven F, Does M and Peumans WJ (1999): A gene encoding a hevein-like protein from eiderberry fruits ¡s homologous to PR4 and class V quitinases genes. Plant PhysioL, 119: 1547-1556.
Van Loon LC and Van Strien EA (1999): The families of pathogenesis-related proteins, their activities, and comparative analysis of PR-1 type proteins. Physiol. Mol. Plant. PathoL, 55: 85-97.
Van Neerven RJJ (1999): The role of allergen-specific T cells in the allergic immune response: relevance to allergy vaccination. Allergy, 54: 552-561.
Van Parijs J, Broekaert WF, Goldstein IJ and Peumans WJ (1991): Hevein: an antifungai protein from rubber tree {Hevea brasiliensis) látex. Planta, 183: 258-264.
Van Ree R, Fernandez-Rivas M, Cuevas M, van Wijngaarden M and Aalberse RC (1995): Pollen-related allergy to peach and apple: an importan role for profilin. J. Allergy Clin. Immunol., 95: 726-734.
Van Ree R, Cabanes-Machetan M, Akerdaas J, Milazzo J-P, Louteher-Bourhis C, Rayón C, Vlllalba M, Koppelman L, Aalberse Re, Rodríguez R, Faye L and Lerouge P (2000a): P(1,2)-
125

Bibliografía
xylose and a(1,3)-fucose residues have a strong contribution in IgE binding to plant glycoallergens. J. Biol. Chem., 275:11451-11458.
Van Ree R, Akkerdaas J, Van Leeweb A, Fernandez-Rivas M, Asero R, Knul-Bretthova V, Knuist A and Aalberse RC (2000b): New perspectivas for the diagnosis of food allergy. Allergy Clin. Immunol. Int., 12: 7-12.
Vieths S, Janek K, Aulepp H and Petersen A (1995): isoiation and cliaracterization of the 18-kDa major apple aliergen and comparison with the major birch poílen ailergen (Bet v i ) . Allergy, 50: 421-430.
Vieths S and Schoning B (1996): Characterization of Mal d 1, the 18 kd major apple aliergen, at the molecular level. In: Wuethrich B, Ortolani C eds. Highiights in Food Allergy, Basel: Karger 63-72.
Vieths S (1997): Allergenic cross-reactivity, food allergy and políen. Env. Tox. Pharmacol., 4: 61-70.
Wadee AA, Boting L-A and Rabson AR (1990): Fruit allergy: Demostration of IgE antibodies to a 30 kd protein present in several fruits. J. Allergy Clin. Immunol., 85: 801-807.
Wirth SJ and Wolf GA (1990): Dye-labelled substrates for the assay and detection of chitinase and lysozime activity. J. Microbiol. Methods, 12: 197-205.
Yagami T, Soto M, Nakamura A, Komiyama T, Kitagawa K, Akasawa A and íkezawa A (1998): Plant defense-related enzymes as látex antigens. J. Allergy Clin. Immunol., 101: 379-385.
126