Embed Size (px)
Citation preview

PROYECTJ IlllCIfiL P2 .A SGRVICIO SOCII%
,CL3$ Jh.onbre : Agustín Jorbnimo :.vila Sz.kilr
Teléfono p a r t i c u l e r : 5-95-10-38
MntrZcule: 81 33721 5
Cerrara : E i o1 o zía CAZ
4 r e a . de concentrac ibn : E i o l o g í a '?'xperimontr?l
Tr imestre : es-?
Lugar en donde se reclizcrh: Departameato de F i s i o l o g í a y B i o f f s i c c d e l CIE;VELT:,V-IPN
13 de j u n i o de 1985
13 de diciembre de 1 9 9
Fechr de i n i c i o :
J Fech. de terminncibn:
1 Lonibre d e l t u t o r :
/ 1Yttulo:
Dr. Fnrique S t e f a n i , P r o f e s o r T i t u l a r
ñespuestc i*!ecSnica de l Iddsculo E s q u e l é t i c o en h e s e n c i a de íian y o t r o s Floqucantts ae lii Per- m z b i l i i a d a l Cdcio

SUSTIFICRCION Y NIiTUFtkLEZA DEL FRGYECT0.-
. ," Este proyecto es oe investigación básica y forma parte de una ser ie de investighciones que se hm realizado desde l a década pasada a cerca del papel del Caz' eri e l aecanisuo de acoplaAento entre excitaciún y con- tracciún.
.., .,
" 1.1 I > ~ . > . & l J ~ ~ l ~ j < . -
/-
Es un iiecho establecido que durante l a hctividad d e l mÚsculo Usque- l 6 t i c o de la rana ocurre un i n f l u j o de iones Ca" del medio extracelular a l doplasma. Cuando se bloquean los corrientes sal ientes de K+ con TEA y se sustituyen los iones GI.- con SO:-o CH,SO;, es posible detectar un potencial de acción dependiente de Caw, con un curso temporal lento (4) . 1,:ediante l a técnica de vol ta je controlado, se ha deidostrado que e l ienó- meno iónico responsable de dicho potencial es una corriente entrante, lle- vada principalmente por k*, l a cual es insensit ive a TTX jr e6 bloqueada p o r GoN, Ni2+, Cd* y P600 (6). Esta corriente sigue una c iné t i ca que se a j u s t a a l a relacidn m3h de Hodgkin-lluxley y se inactiva completamente con despolarización sostenida (16).
sistema tu,bular con la membrana superf ic ia l (mediante tratamiento con g l i - cero l ) y l a corriente de Cay ha demostrado que l o s canales de Caz' se en- cuentran ix incipalmnts en el sistema t u b u l a r (8). E l decaimiento de l a corriente de Cal+ parece es tar gobernado fundamentalmente p o r un proceso de inactivación propio de los canales de Ca", aunque tanbien es posible que part ic ipe un fenómeno de deplecióo o agotamiento de Caz' del sistema tubu-
Recientemente ha sido posible reg is t rar una respuesta e l é c t r i c a de- pendiente de Ca" en f ibras musculares de -€fer0 (rata) . Esta respues- ta piirece estar mediada por cónales sirnilares a l o s de l a s f ibras de an- f i b i o s , ya que su c inét i ca y propiedades farmcol6gicas son semejantes a l a s de aquellas. Sin embargo, hay algunas diferencias importantes entre I t s respuestas de f ibras lentas y rápidas en ratas (14).
Otros iones también permean p o r e l canel de Ca y pueden comportarse COUIO bloqueantes ,de l a conductoncia a i Caz'. l e n t e s , cuya secuericia de permeabilidades es SCi2'>Sí.t>,*~~,>,,''(9).
p l i c d o en e l necanisuo ds exci tac i5n-~ontracción, y se han postulado varius alternetivas de cónio ocurriría ésto. La corriente entrante de Caz',
La correlación l i n e a l entre e l grado de discontinuidad e l é c t r i c a d e l
'
lar (10, 15, 17) .
Se t r a t a de cationes diva-
rjesde un principio se supuso que e l influjo ae Ca2'podria estar i m -
. p ü r ejemplo, podría intarac.twar de alguna ncmera con lzs corrientes capo-
x

c i t i v a s asim0tricas del sistema t u b u l a r , l a s cuales aparentemente parti- cipan en e l acoplaaiento entre e x c i t a i ó n y contraccibn. (2). También Se ha sugerido que l a entrada de Caz' durante despolarizaciones prolongadas e s s u f i c i e n t e para contribuir a l a tensión generada (13).
que l a sustitución de Caz+ por. otros i o n e s permeantes, como Baa* y Sr) puedo tener sobre l a respuesta mecánica de f ibras aisladas cuando éstas son estimuladas eléctricamente.
RhTEV3 DKiV 11:s. -
Por l o anteriormente mencionado, e s iaportante determinar e l efecto
ria sido establecido desde 1973 que existe una dependencia entre l a s contracturas de k' y e l Caz+ extracelular (1, 3). concentración externa de üa" a valores 6 lo+ Y produce una depresión war- cada de las contracturns de K'. a m p l i t u d de l a s contracciones simples, a s i como l a amplitud y duracidn de l a s contrecturas de K+ (1, 5). sencia de k*' en e l medio produce una aumento s igni f i ca t ivo d e l uriibral mecánico para pulsos brevesi es te efecto puede deberse a una reducción de l a e f i c ienc ia del mecanismo por e l cual l a despolarización tubular dispara la l iberación de Caz' del re t í cu lo sarcoplismico (7). go, e l ca* externo no es esencial para que se produzca tensión, ya que es posible regis trar contracciones.aln después de exposición prolongada a soluciones l i b r e s de Ca". Kecientenente se ha deteruinado e l efecto que t iene l a reducción o elimin&ción del Caw extracelular sobre la ciné- t i c a de 18s contracturas de K+ (11, 12). Mientras que l a s propiedades e l é c t r i c a s permanecen prbcticarwrnte inalteradas, l a tensión máxima dis- uinuye,. la duración es menor y l a velocidad de rela jución espontanea aumenta; l a curva de activación se desplhza hacia valores d s p o s i t i v o s y la curva de inactivación se desplaza hacia valores d s negativos. t o s resultados apoyan e l que e l c;a* extracelular contribuye a mantener la: tensión durante l a contractura.
OBJE TIVUS. -
La disminución de la
Se v i 6 también que e l h'' disminuye i a
Posteriormente se observó que l a au-
Sin embar-
Es-
En esto trabajo se tratará de establecer e l efecto del Ba*y otros bloqueantes de la permeabilidad al Ca" sobre la respuesta u e c h i c a de fibras aisladas de mfisculo esquelético de l a rana. sentar un cuadro d s congleto de resultsdos se real izará priijero una se- r i e de experilietitos sobre contracturas ae K+, en donde se probará también e i e fecto tiel &'.
Con el f i n de pre-
PHO%ALÜí Y UI'ODGLOSIA E iU1siJO.-
81 programa tie t r a b a j o inclu,e t r e s partes. La prij.cra consiste en L L ¿ntrei~apiiento en l a técnica ue disección de f ibras y r e g i s t r o ae con- L *I , acturas ,. y cofiLrúcciui~e3 siirirles bajo conLicioues n ú r i d e ~ . Esta fase es iwiispeosable pai'a poder l l eva r a c ~ b o Lou e x y e r i ; s r ; t i t o ~ sa t i s fac tor ia - mente. La ae6uii.a darte cocsicite en aatai i iecer si oi &*' tiene algún

5
senaltdos, yero puede hdber t r a s l a p e . Es probable que los primeros ex- perimentos s e r e a l i c e n desde j u n i o y también es f a c t i b l e que en noviem- b r e aún se n e c e s i t e h a c e r o r e p e t i r algdn experimento.
LITERAí'LJRA CITADA.-
G.J. Chiarandini B E. S t e f a n i . " E f f e c t s o f mnganese on the elec- t r i c a l and mechanical p r o p e r t i e s of f r o g s k e l e t a l muscle fi- bres." J. P h y s i o l . (Lond.] 2 3 2 ; 129-147, 1973.
L.F. Schneider dc W.K. Chandler. V o l t a g e dependent charge movenient i n s k e l e t a l muscle: t~ p o s s i b l e s t e p i n e x c i t u t i o n - c o n t r a c t i o n coupling." i iature 242: 244-246, 1973.
E. S t e f a n i rC U.J. Ciiiarandini. " S k e l e t a l muscle; dependence o f potassium c o n t r a c t u r e s on e x t r a c e l l u l a r calc i i in . " Pf lügers .- Arch. 3 4 3 ; 143-150, 1973.
GA. i>eaty & E. S t e f a n i . "Calcium depezdent electrical a c t i v i t y i n t w i t c h muscle f i b r e s o f the frog." Proc. R. Sac. Lotrd. 3 194: 141-1509 l 9 7 3 .
D.J. L l i a r a n d i n i & E. S t e f a n i . "Ca emd excitation-contraction cou- p l i n g i n f r o g s k e l e t a l muscle f i b r e s , " en E l e c t r o b i o l o w o f p e r v e , Synaose and Lluscl~. Ed. J.P. Reuben, D.P. Purpura, h.V.L. a e n o e t t & E.R. iiandel. Raven Press, ii.Y., 1976.
J.A. Sdnchez & E. S t e f a n i . " l n u a r d calcium current i n twitch muscle f i b r e s o f t h e frog." J. Physiol . (Lond.3 283: 197-209, 1978.
D.J. Chiarandini , J.A. Sdnchea D E. Stefani. " E f f e c t o f calcium bitiidrawal on ffiechanicel threshold i n s k e l e t a l uuscle f i b r e s o f t h e frog." J. Physiol . (Lond.) 303: 153-163, 1980
L. i i i c o l a S i r i , J.A. Sanchez ú. 2. S t e f a n i . " E f f e c t o f rr lycerol t r e a t - - _ iiierit on t h e calciuu current o f f r o g s k e l e t a l muscle." (Lond.) 3 0 5 ; 67-96, 1980.
3. F h y s i a l .
41. Aliliaers & P.T. Palada. "Slon caLcium and p a t a s s i m currents a c r o s s lleasurenents ' c i t h a vaseline-gap technique." Crog muscle membrane:
J. i'hysiol. (Locd.) 312: i55-1'76, 198i.
d . L l u e r s , K. r i n k S Pii. P a k d e . "Calciuin deplet ion in f r o g uuscle t u b u l e s ; ti;e u e c l i n e o f c u l c i u u current under maintained deuo- 1 .ar izt~t ion." J. Physiol . (Lond.) 312: 177-2507, 1981.
C"
Y


. ..
, .-
Agustín Jerbniuio Avi l t i Sakar

. . .
*""
....
I.
RESUIIEM
be estudiaron en f i b ras a is ladas de PuIscElo esque lé t ico de l a ra2a 10s efectoai producidos p o r 1ú. privación de Ca , su sust i tución por Ba y
y e l uso de l o s bloqueantes d i l t i a z em y Nit+, sobre la amplitud y curso temgoral de l a s sacudidas simples.
En ausencia de Caw, con EGTA 1 y 2 mlb, se observó una reducción de l a amplitud, duracidn til máximo y área bajo l a curva, as€ como de dP/dt a l i n i c i o de l a c~~ntracc ión . La recuperacidn fue gradual y se ajustó a una curva sxponencial de exponente negativo.
y generalmente también d e l tiempo a l d x i m o y área bajo l a curva. aumento de l a amplitud fue gradual y se ajustó a una función exponenciai de exsonente negativo. La recuperación no eiempre fue t o t a l . En algunos casos l a ea.cuaaaa parmtmeció potenciaati mucnos unusoe aesrués ae na&r repuesto l e solución normal. E l regreso a l a amplitud o r i g i n a l también se ajustd a una función exponencial.
gradual de l a tensi611 pasiva.
p!d, potenciación de l a contracoión.
en el acople excitación-contracción y ee cree que l o s e f ec tos observados se deben a modificaciones eo s i t i o s externos de l a membrana p l a s d t i c a , probablemente relacionudos con el movimiento de cargas d e l sistema de túbuloa transversos.
hl sus t i tu f r el Cd+ por Baa+, se observó un aumento de i a amd l i tud El
El d i l t i a z em produjo una potenciacidn de l a sacudida y un aumento
I:l Niz*, en concentración de 500 ,u& produjo disminución, y con 10
Se interpretaron los resultados en base a loa procesos implicados
INSHUDUCCI ON
Es un hecho estab lec ido que durante l a act iv idad de l músculo eaque- l 6 t i c o de l a rana ocurre un i n f l u j o de iones ca" d e l medio extrace lu lar a l miopiasma. y se sustituyen los iones C l - con SO;- o CH3SOi, es pos ib l e detectar un potencial de acción dependiente de Cae, con un curso temporal l en to (Beaty & Ste fani , 1976). Mediante la técnica de v o l t a j e controlado, se ha demos- trado que el fenómeno idn ico responsable de dicrio potenc ia l es una corrien- te entrante, l l evada principalmente por Ca*, l a cual es insens i t i va a TTX y es bloqueada por CO"Y Ni", Cd'+y D-600 (Sánchez y Ste fani , 1978). Esta corr iente sigue una c iné t i ca que 68 ajusta a l a re lac ión m'h de Hodgkin- Huxley y se inac t i va completamente con depolar ización sostenida (Snchez PC Stefani , 1983).
Bistemu tubular con l a membrana super f i c i a l jmediaote tratamiento con g l i - cerol) y l a cor r i ente de Caz* ha demostrado que l o s canales de Caw se en- cuentran principalmente en el sistema tubular (h i co l a S i r i e t al., 1980). E l decaimiento de l a corr iente parece es tar gobernado fundamentalmente por un proceso de inact ivac ión propio de l o s canales de Ca", aunc,ue tam- bien es pos ib le que pa r t i c i pe un fenómeno de depleción o agotamiento de
Cuando ee bloqueen l a s corr ientes sa l i en tes de K4 con TEA'
Le corre lac ión l i n e a l entre el grado de discontinuidad e l é c t r i c a de l

2
.' .
..
. ..
.
, .
, ~ * .
.-,
b e d e l sistema tubular (Almers, e t al., 1 9 8 1 ; Gota, e t a l . , 1983; Gota, e t al., 1984).
Recientemente ha s ido p o s j B e r e g i s t r a r una r e s p u e s t a e l é c t r i c a de- pendiente de @ e n f i b r a s musculares de mamífero (rata). parece e s t s r mediada por canales: s i m i l a r e s a los de las f ibras de anfi- b i o , ya que su c i n b t i c a y propiedades f.rirmacológicas 8on semejantes a los de aouellas. respuestas de f ibras l e n t a s y rd.!.:idas en ratas (Chiarandini & S t e f a n i ,
como bloqueantes de l a conductancia a l Ca". l e n t e s , cuya secuencia de perme&lidades es Br?>C?>Ca 7 b >&>+(fiilmers 6c Palade, 1981).
Desde un p r i n c i p i o se supuso que e l i n f l u j o de Caz+ podría estar i m - p l icado en e l mecanismo de acople exci tocibn-contracción. e s t e fenómeno e s mucho más com;ilicudo y en r e a l i d a d aún se conoce poco a c e r c a de é l . Se ha propuesto, por ejemplo, que e l aumento en la concen- t r a c i ó n de Caz' i n t r a c e l u l a r funciona como disparador de l a l i b e r a c i ó n de Caz+ d e l r e t i c u l o sarcoplásmico. que l a l i b e r a c i ó n de Caz' es un procerjo r e g e n e r a t i r o , en e l c u a l e l &mis- mo produce la l i b e r a c i ó n de más Caw del r e t í c u l o sarcoplásmico (Endo, e t al . , 1370) . S i n embargo, una s e r i e de evidencias apuntan en contra de que e l i n f l u j o de iones Ca" p a r t i c i p e de una pLELnera d i r e c t a en e l acople e x c i t u c i 5 n - c o n t r a c c i ó n . La c o r r i e n t e de Ce*+ no ee a c t i v a d a p o r l a depo- l a r i z a c i h rbyida de un p o t e n c i a l de a c c i 6 n (Sánchez & S t e f a n i , 1978) . E s t o , a su vez, apoya l a e x p e r i e n c i a de que muticulos y f ibras aisladas siguen c o n t r a y h d o s e p o r d s de 20 minutos después de reducir l a concen- t r a c i ó n d e #Caz+ e x t r a c e l u l a r a menos de IO-' bl (Armstrong, e t 8.1., 1972) . S in embargo, las c o n t r a c t u r a s de p o t a s i o sf s e ven vfect.tidos en amplitud y duracióri Id disminuir lo. concentración e x t r a c e l u l a r de @en l a misma magnitud ( S t e f a n i C. C h i a r a n d i n i , 1973) . La tensión máxima, la duración y l e consteinte de tiempo de l a r e l o j a c i ó n espontánea decrecen progresi - vamente (Cota & S t e f a n i , 1 9 8 1 j . con n i f e d i p i n a y d i l t i a z e m produce una reducción de l a duración y a m p l i - t u d de las c o n t r a c t u r a s de K+((;Umboa-A.ldeco, e t a l . , en preparación) y e l blocueo con hi" reduce l a f a s e sostenida de l a contractura (Huerta, e t el., 198!i). mantener l a tensión durante l a s c o n t r a c t u r a s de K+ mediante alguno de l o s s i g u i e n t e s mecanismos: a ) aumentando l a concentraci5n de Ca" d e l mioplasma [ l"otreiiu Fi Raymond, 1 9 8 0 b ) , b) cargando e l r e t i c u l o sarcoplas- mico, proporcionando a s í más CF? disponible pzrz su l i b e r a c i ó n (biicola S i r i , e t al . , 1380), c) provocando l a l i b e r a c i ó n de & d e l r e t í c u l o sar- coplásmico mediante e l mecanismo regenerat ivo menoiontido anteri . ,rments (Endo, e t a l . , 1 9 7 0 ) o d ) interactuündu externa- o internamente con las c o r r i e n t e s c a p a c i t i v a s a s i m é t r i c s s de los t ú b u l o s
r e g i s t r a n simultáneamente l o s cambios tranuituriozc d e l Ca" citopl6smico (mediante e1 uso u e l c o l o r a n t e I ~ r i t i p y r y l a z o 111) y l a s c o r r i e n t e s t r a n s - membranales, s e ha v i s t o que l e disminución de l a concentrsción de &'* e x t r a c e l u l a r reduce l a f a s e i n i c i a l ascendente d e l t r a n s i e n t e de C á y
E s t a respuesta
S in embargo, hay algunas d i f e r e n c i a s importantes entre las
1983) . Otros i o n e s también permean por d c a n a l de Cast y pueden comportarse
Se t ra taze c z t i o n e s diva-
S i n embago,
De hecho, se demostró en f i b r a s desnudas
i.demRs, e l blocueu de los cenales de Cs'*
1-5 p o s i b l e que l a c o r r i e n t e l e n t a de C z c o n t r i b u y a a
( S h l e v i n , 1979) . Por o t r o lado, estudios r e d i z a d o s en f i b r u c o r t & S a , en donde :;e
c

3
la cantidad de &* contribuido sor e i ret iculo siircoplismico, sin l a intervención de l a corriente de Ca"(Brum e t ai., en tjrensa). queo con nifedipina produce resultados simi~8XeS Y I Puesto corriente d e Ca': se piensa que e l s i t i o de acción debe ser extracelular, e s decir , que e l Caz+ no necesitt e n t r b r u l a célula para perticipar en e l acople excitación-contracción, sino que podría mós bien interactuar con los com,ponentes d e l sistema tubular sensibles a l vol ta je , l o s cuales son los responsables del movimiento asimétrico de chrgos ourante laa depolerizaciones de It: membrana (Schneider & Chandler, 1973). De hecho, se ha visto que l o s parámetros del movimiento de cargas i v y k ) cambian de acuerdo con l a concentración extracelular de Ca"? r o t o s cambio8 -on cnozi::t,mtss en direccidn y generalmente también en m a g n i t u d con los efectos reportados de la concentración de Ca" sobre e i umbral mecánico de lan f imas (Shlevin, 1979). iones o t r o e cationes divalentes) tienen un papel de upantallmiento sobre l a sukjerf icie externa d e 1~ membrana p l a s k t i c a , de modo que loe cambios en :La concentración de Caz' extracelular pueden modificar e l po- tencia l ae superficie y e l campo e léc t r i co dentro de l a propia membrana, lo cual, a 13u vez, se traduce en un cambio de la relación entre e l poten- c i a l de membrana y e l umbral mecánico (Chiarandini, e t a l . , 1980).
no decrece significativamente durante depolariraciones de hasta 2 segun- dos de duración a potenciales de meabrana.de -60 a -30 mV y tiende a desaparecer en fuacidn del tiempo de exposición a l a solución hipertd- nica de registro ( C o t a & Stefani , 1985; Cota 6. Stefani , en prensa). Cuando l a s f ibras se incuban por varias horas en TEA+ y Cs+, o cuando se t raba ja en f i b r a cortada con 'TEA+ y EGTP. intraceluler , no ee regia- t r a l a corrj.ente mencionada (Cota, et al . , 1983; Wmers d Palade, 1981). Es posible clue l a existencia de canales rhpidos de Ca" depender de compo- nentes intrmcelulares l f ib i les , f á c i l de perderse en esas condiciones; o bien, l a ausiencia de la corriente podría r e f l e j a r un efecto bloqueante por ucumuiación de TI-A+(Almers & Palade, 1981).
S i bien, p o r su cinét ica , podría esperarse que l a corriente rápida de Ca"se activara durante cada sacudida simple, hay evidencia en contra de oue ésto suceda. el diltiazem, p o r ejemplo, no tiene ningún efecto depresor sobre e l fenómeno de acople excit~ci6n-contracci6n (ver resul- tadosj. Ms bien produce una potenciación de le. sacudida simple ( b n z á - les- ierratos , e t al., 1982). aunque no depresor, a s í como e l efecto mencionado de l a nifedipina (en un s i t i o externo) sobre l a liberaci6n de Caz* del ret iculo sarcoplásmico, indica que existe una relacidn estrecha entre los canales de CaL+y s i - t i o s de l a membrana implicados en e l acople excitaci6n-contracción, de manera i;ue e:l Ca*+extraceluiar ( y otros iones divalentes o bloqueantes de Ca") module l a l iberación de Cadel ret iculo sarcoplésmico, no a trri- v6s de corrientes entrantes, sino de su interacción con los componentes de l movimiento intramembrand de cargas oensibles a l voltaje.
Se han propuesto otras hipótesis recientemente a cerca del mecanie- no del acople excitaci6n-c~~ntracci6n, como por ejemplo, 1t del inosi tol t r i fos fa to (1:Ej ), un producto de l a degradación del f o s f a t i d i l inosi tol . Se ha visto en d s c u l o l i s o que es ta sustancia se produce como respuesta
El blo- no hay
Esto puede explicarse en base a L,ue los
Recientemente se ha descrito una corriente rápida de Ca", l a cual
No obstante, e l que ef tenga un efecto,
I

4
et a estirnulo?i por ejenteu co l inérg i cos y ,;revoca l a l iberac i6n de Ca de l r e t í c u l o sarcoplásmico. Se ha estudiado muy poco a i IP, en músculo es- que lé t ico , pero se piensa cue puede funcionar como "segundo mensajero", enlazando químicamente e l fen6rnrmo de l a depolar izacidn de los túbulos T con l a l ib f i racidn de Ca" de l r e t í c u l o sarcoplásmico (Somlyo, 1985).
OBJETIVOS
E l oh je t i vo de los experimentos rea l i zados en es t e t rabajo es l a determinación d n l o s e f e c t os producidos por l a privacidn de Casi, su suo- t i tuc idn p:;r Bo sobre IC amp l i tud y curso temporal de l a s contracciones simples en d s - culo esque lé t ico de l a rana.
y l a adic ión de i l l y n o s bloqueantes de l canal de Caz+,
U T E R I & Y METODOS
Los experimentos fueron rea l i zados a temperatura ambiente en f i b ras ais ladas o fasc iculos pequenos de l t i b i a l i s anticus longus, semitendino- .~ sus o i l i o f i b u l a r i s de Rana PipiLnE.
Las ranas eran t ra idas de s i t i o s cercanos a l D.F., aproximadamente cada 1 5 d ías y se mantenían en un estanc.ue a temperatura ambiente y con agua c irculante. Se l a s aliment6 con saltamontes o escarabajos v ivos , cuando l o s hubo. Se s a c r i f i c a b a por decapitación y se l e s extraian to- dos&is milrsculo# requeridos p a r ~ . e l dia (no 5610 para l o s experimentos de es te trabajo, sino también para los d e d s experimentos l l evados a cabo en e l laborator io ) . Lh mayoria de l as veces se emplearon músculos fres- cos, disecados entre 1 y 3 horas antes de l experimento, pero algunas ve- ces se mantuvieron en re f r i ge rac idn , ya fuera e l músculo completo o fas- c i cu los pequeños, tior var ias horas (por ejemplo, durante l a noche).
Diseccidn y montaje de fibras.-
Una vez extra ído un mlsculo, se f i j a b a por l o s tendones mediante dos peeueños d f i l e r e s ("minucius") a l a base de p l ás t i co de una ca ja de P e t r i en l a cual hubiese su f i c i en t e cantidad de Ringer normal. go, con otro p&r de minucias, se extendía laterolmente e l músculo, de modo que quedar8 expuestr una mayor área de 61. DespuBs, con unas ti- j e r e s f inas se iban cortando l & ~ f i b ras 8u;er f ic ia lea y con una pinza se separaban para i r dejando una capa cada vez más delgada de f ibras . De esa capa delgada, ::,e sepiraban fascfculos pequeños, l o s cuales se dejaban reposar unos minutos para poder v i sue l i za r l a s f i b ras intactas mbs fdcilmente y d i s t ingu i r l e s de l a s danadas (La diseccidn se efectua- bu bajo una lupa de disección con campo c laro) . Después se iban qu i - tando l a s f i b r8s dañadas con cuidado de no tocar e l a s que estuviesen bien. Por último, 8s eliminaban una a una l a s f i b ras restantes hasta
Lue-

5
dejar una sola fibra intacta . una navaja e l t e j i d o conectivo {que mantiene unidas a l a s fibras. Cuando l a diseccilin se hacia para medir e l potencial de reposo, la primera por- t e de l a dlsección del núsculo ere igual , p e r o en vez de quedarse con una sola fibra,, era necesario quedarse con un fascículo de una sola capa de f ibras extendido de t a l mnera que 6stEis pudieran visualizarse b ien para penetrarlas con microelectrodos.
t a l de luc i ta . Uno de los extremos de la f i b r a se f i j aba mediante una minucia a l fondo de l a camarilla, mientras que e l otro extremo se engun- chaba a l a palenquite de un transductor mecano-el6ctrico (Cambridge Tech- nology, Inc . , modelo FX-O). Cerca d e l extremo f i j o de l a f ibra se encon- traba l a entrada p o r l a cual se efectuaba e l cambio de soluciones y en e l o t r o lado de l a camarilla se colocaba una manguerita de succión. LRS so- luciones se poniaii en jeringas dispuestas verticalmente sobre una conexión con válvulas, de nodo p e por gravedad se hacia pasar un f l u j o constante y no turbulento a través de l a cam;Arilla. La velocidad del recambio se regulaba mediante l a s v5.lvulos. Pora un cambio de soluciones se pasaba un f lu jo d e aproximadamente 10 ml/min y durante un lavado prolongado !se ;,usaba entre 1 y 2 ml/min. Tl f i u j o , entonces, se podia hacer l o sufi- cientemente rápido aara l o g a r en pocos segundos los cambios de crincen- tración deseados p a r u e l experimento.
T1 funcionamiento del transductor mecano-eléctrica empleado est& basado en un capacitor de espaciamiento varicihle, donde un c5pacitor ac- t ivo ( e l que mide l a fuerza, propiamente) es comparado eléctricamente con un dispositivo de referencia de igual tamano y construcción que es tá montado j u n t o a 61. proporcional a l valor de tanto el capacitor de referencia como e l de me- dición d? :fuerza. do no hay fuerza aplicada, l a s dos corrientes a s í generadas son iguales, y su sustracci6n electrónica da cero. S i se aplica una fuerza, e l valor de l a capacidad debido a l a fuerza cambia, l o cual hace c,ue l a resta electrónica ya no dé cer i . La diferen- c i a es proporcional a l cmbio de l a ca~~acidad, y p o r lo tanto a l a fuerza aplicada. La e!;uivalencia del transductor es 20 mV/mg y puede registriir cambios de tensión del orden de nicrogramos. un extremo de l a f ibra estaba fijo e l fondo de l a cxm3rills y e l o t r o su- j e t o a l trrrnsductor, l a tensidn se registró isométricamente en todos 1.0s casos.
C o n t r o l del. espaciamiento sarcom6rico.-
Para ésto era necesario ir cortando con
Una voz aislada l a f ibra , se transportaba a una camerille experimen-
Un sistema elsctrdnico genera una señal de corriente
Cuando l a s d o s ca;acidades son iguales, es decir , cuan-
l a cantidad de corriente generada
&Puesto que, como ya se d i j o ,
Es un hecho conocido que 1fl magni tud de l a tensión depende de l a lon- g i t u d sarcomérica, de t a l forma que a un espaciamiento ó p t i m o (2.0 a 2.25 pm) l a tensión es máxima (Peachey, e t al . , 1983). estiramiento de l a f ibra de aproximadamente 10% m6s que su l o n g i t u d anató- mica. Para controlar e l espaciamiento sarcomérico en reposo durante todos los experimentos, se empleó un aparato de rayos laser (Coherent, CR-90- 21HP helio-neón de 2 mH de potencia), cuyo haz luminoso, de l o n g i t u d de onda conocida ( 4 . 9 6 ~ 1 0 - ~ c m ) , se hacia pasar a través de l a fibra.
Esto corresponde a un
En una
I .
,. .
..
e..
I
,I.
,,~.
I-
.‘- r<_.
1. .
...
...
.-
.
.-.
, ...
... .
. -.
.. ..,
6
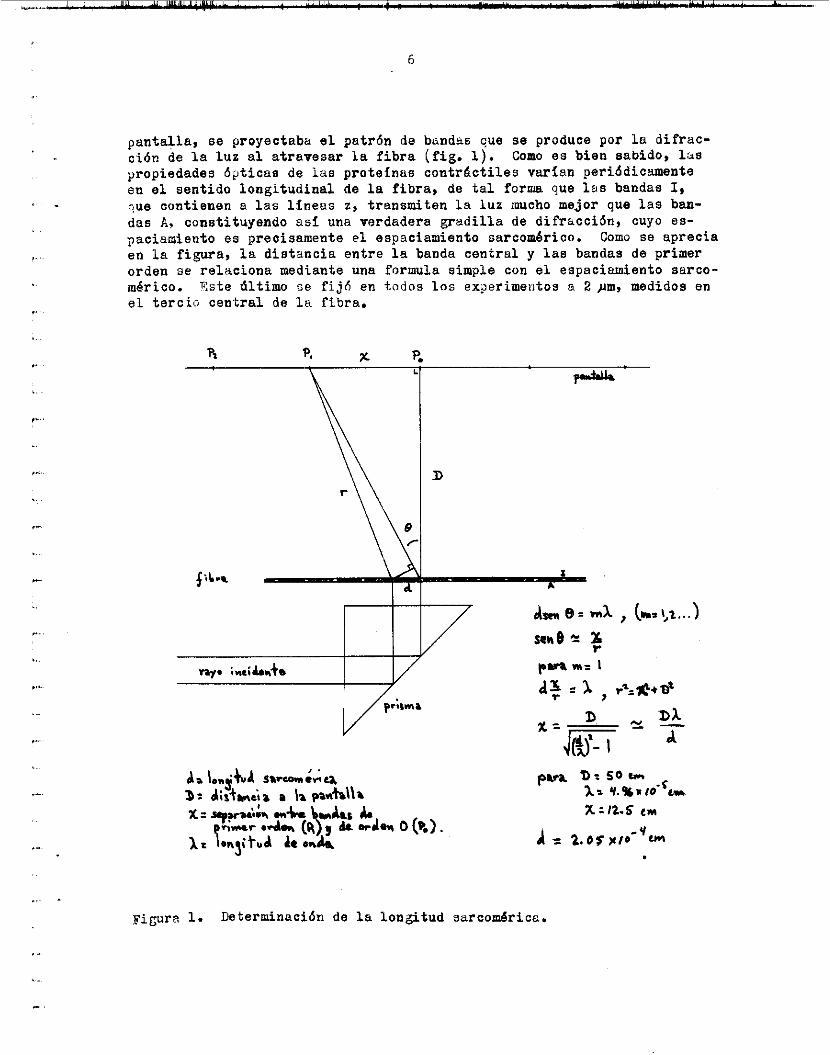
pantalla, se proyectaba el patrón de bmdus cue se produce por l a difrac- c i ó n de l a l u n a l atravesar l a f ibra (fig. 1 ) . propiedades ópticas de i a s proteínas contrdctiles varían periódicamente en el sentido longltudinal de la f ibra , de t a l forma que l a s bandas I , tue contienen a l a s l íneas z, transmiten l a l u z inucho mejor que l a s ban- das A, constituyendo as€ una verdadera gradil la de difracción, cuyo es- pacismiento es precisamente e l espaciamiento sarcom6rico. Como se aprecia en l a figura, l a distancia entre l a banda central y l a s bandas de primer orden oe relaciona mediante una formula simple con el espaciamiento sarco- mérito. % t e ú l t i m o se f i j 6 en t o d o s los experimeutos a 2 pm, medidos en e l t e rc ib central de l a fibra.
Como e s bien s a b i d o , 14s
Figura 1. Determinación de l a longitud sarcomérice.

7
Solucidn
Ringer Norm1 CaO,Mg3 * C~OY@~,EGTAZ Ba 3.2 K 5 M l t 10OuM Xi 500pM Ki lopid
CaO,Mg3,EGTAl:2.5
K+ Na' ~ 1 - caw MT HE PES EGTA Ya'' lDiit iezem x i Z +
h.5 117.5 123.6 1.8 --- 5.0 --- --- --- --- 2.5 117.5 126.0 --- 3.0 5.0 --- --- --- -..-
117.5 126.0 --- 3.0 5.0 1.0 --- --- . --- 2.5 117.5 126.0 --- 3.0 5.0 2.0 --- --- --- 2.5 117.5 125.4 -I- --- 5.0 --- 3.2 --- --- 2.5 117.5 124.5 1.8 --- 5.0 --- --- --- --- 2.5 117.5 123.6 1.8 --- 5.0 --- --- 9.100 --- 2.5 117.5 123.6 1.8 --- 5.0 --- --- --- .500
.o10 2.5 117.5 123.L 1.8 --- p .o --- --- ---

.
,...
,, -
. .
....
,. - ., .
..
.-
I
,, .
.
.. .
. .
-.
8
'El Ca-presente como contaminación en l o s reac t i vos se calcula en
*)La ad ic ián de EGTA 1mY a l a soluci5n s in Ca? disminuye l a concen- 3 x N 7 M (Chiarandini & Ste fani , 1976).
t rac ión de e s t e ión en e l medio ex t race lu lar a menos de 1x10-* hf (Amstrong, e t al., 1972; Chiarandini & Ste fani , 1976).
En la e lecc idn de 1:s concentraciones de l a s iones se tom6 en cuenta no sólo e l potencial de membrana s ino también potencial de superf ic ie . For esta razón, se sustituyó e l &?1.8d $or big 3mM o p o r BZ3.2-, ya que estas c,oncentracioces compenscn e l cambio en e l potenc ia l de superf i - c l e producido por l a eiirninacidt de l Ca"+ ( X i l l e , e t al., 1975; Chifirnndi- n i , e t al., 19EO).
su f i c i ente tiempo de ant ic ipacidn p m a asegurar e l e q u i l i b r i o térmico du- rante l o s experimentos.
3eg i s t ro y i iná l i s i s de l o s experimentos.-
h s so:luciones eren conservE.dcs er. re f r i ge rac ión y se sacaban con
¿a sa l ida de l transductor mecano-el6ctrico estaba directamente co- nectada a l o s dos canales de un aparato de r e g i o t r o Gould (modelo Brush 200) de t razo r e c t i l í n e o , a. un osc i l osco2 io de doble bcne de tiempo (Tek- tror1ics, Inc), 8 la entrade de unii grab:,.iora de v ideocassettes (sony beta- SL-2700)- Y a l a entrada de un convert idor analóg ico-d ig i ta l (Tee-, I ~ ~ . ) cuya SoEal de sa l ida era adquirida en uaa computadora (IBU-AT),
uso de un juego de programas especialmente diseriadoe para experimentos e l ec t ro f i s i o16g icos , pero cuyos parámetros fueron adaptados sa t i s f ac to - riamente para los experimentos de e s t e trabajo. Este juego de programas r ec ibe el nombre de pCLAyip (copyr ight Axon Instruments, 1984) y es tá di- señado para dos t i p o s de experimentos: tiempo y dependientes de un evento. genera secuencias de v o l t a j e s comando y r e g i s t r a simult8neamente i o 5 da- t o s de uno o dos canales. La duracidn de los in te rva los dentro y entre l a s secuencias se establece mediante los p a r b e t r o e de l programa. Para los experimentos dependientee de un evento, l a computadora espera en e l modo de adquis ic ión de datos hasta que ocurre e l evento aleator io . auceso dispara a l a computadora para que r e g i s t r e e l evento (incluyendo datos anteri.ores a l disparo). Lo8 prograsas que constituyen p c L W son cuatro' U:WEX (para adquisicibn de experimentos de v o l t a j e controlado dependientes de un intervalo ) CLiWAii (para a n á l i s i s de datos adquiridos por CLAMPEX), FXTCKEX (para experimentos dependientes de señales o even- tos espontáneos) y F%TCHAi< (para aná l i s i s de datos adquiridos por FETCIBX).
Fa l o s primeros ensayos se empleb FTTCILTX corn62 programa de adquisi- ción a parti.r de los r eg i s t r os grabados en videocassette. con este programa requiere de un elemento importante: e l discriminador de ventana (Window Mscriminator , modelo D-400, Y. Walsh Electronics). Este es un discriminador de amplitud de dos n ive les , Gon carac te r i e t i cas ad ic ionales de c r i t e r i o temporal. La señal de entrada es ampl i f icada de 0.5 a 10 veces mediante contro les d e l tablero. hlediante dos potenció-
La adquis ic ión de l as seiiales en l a computadora 5e h izo mediante e l
dependientes de un i n t e r va l o de Para l o s primeros, l a computadora
Este
La adquisicidn
metros de 10 vueltas se f i j a n cuales e l primero corresponde
dos n i v e l e s de DC (la ventana), de 105
a l n i v e l umbral i n f e r i o r de disparo. BX

v
. .
. ,
a , .
. .
._
. , .
, .,
... ".
. ..,
...
I ."
e -
".
I . .
. .. 1..
-.
9
segundo sigue a l primero y siempre es positivo rescecto a 61. c a l compuesta es l a suma de las t r e e señales mencionadas y un n i v e l 1 6 - R i c o i n d i c a l a presencia de l a senal de entrada entre l o s d o s niveles de DC. Otra salida de c r i t e r i o de duración indica, que l a señal estuvo dentro de l a ventsnn p o r e l tiempo especificado para l a adquisición.
este modo, cuando l a senal de entrada ( e n este caso, 10 que viene de le grabadora) permanece dentro d e l rango establecido p o r l a ventansi durante un tiempo determinado, e l discriminador manda un nivel lógico a l convertidor anal6gico-digital para que l a computadora adquiera la senal. tran en l a figurn 2.
con l a computadora mediante CLAWXX. Con este prograna se especif ica un i n t e r s d o de tiempo entre los p u n t o s de una muestra de 512 puntos. Los volta jes comando píirt: l o s ex;>erimeotos de registro de tensión son irreleventen y s610 se emplearon para disparar al estimulador ( a tra- vés del convertidor zne16~;ico-di,:ital). La magnitud, durución y r e t x - d o d e l e s t l r t u l o , como y a se d i j o , se controlaban mediante e l te6lero d e l estiIiu1::dor. Los parbetros de : :d;,uisici$n con 3, X se muestran en la fi.pri:. 3.
i't.rt: el. aná l i s i s u e Los experimentos adquiridos por FE'I'(;?SX, se emsleó e l ip.og;?mt. conplementari's Fi?TCíii4, con 01 cual se generoron t.rchivos de eventos promediados. Estos, a su vez, fueron unalizados con LOTUS, que es un programe exteoso de ordenamiento y procesamiento de datos en arreglos me.triciales. L o s datos de C!JJdYi?X fueron anali- zados directamente con cZAMP~%'. Fmpleando estos programas, se obtu- v ieron amplitudes, duraciones y Breas bajo l a s curvas de eventos a i s - lados o promediedos, e s i Corno s u gruficacidn, ésto ú l t i m o haciendo ueo de un graficador FiP-7470A. (He,alett-Packurd).
Far8 la medic ión d e l potenc,ial de reposo de l a s f ibras se emplea- ron dos microelectrodos (uno intracelulrr y o t r o extracelular) de aprox- imr.damente 40 Ir;Lz de res is tencia , conectados a un seguidor de 'vo l ta je , y de ahí a d o a entradns de un am,Aif icador diferencial de 3 canales (dise- liado y fabricado en e l Dpto. de Fisiología del GINVEYTAV-IPN). La d i f e - rencia de potencio.l entre e l electrodo intracelular y e l extracelular era seguida en un oscilosco>io y le ida en un voltimetro digi ta l .
Pari l o s exrerimenios en que :e midi6 simult6neamente l a tensidn y e l potencisl de acci6n extraceLular se empleó un par de electrodos de alambre fino de p la ts , o , i $ l a d o s en toda su longitud excepto en l a punti-. y con una separaci6n de aproximadamente 1 mm. conectados i i l seguidores de vol ta je y éstos, a su vez, cada uno a un ea- nal de un a m p l i f i c a d o r diferencial de dos canales (diseñado y construido en el. @,to. de Fisiologia del CII.íVESTAV-TPK). E l potencial de accidn se regietraba, entonces, como l a diferencia entre las señales de ambos cn- nules y entraba, después de pasar p o r un amplificador operaciona.1 con ga- ntincia 100, a uno de los canales de un osciloscopio de doble base de tiempo (En e l o t r o canal, por su,uesto, se registraba simultáneamente l a tensión 'con otra base de tiempo).
Una se-
Lffi parhetros de adquisicidn de 22 sacudida9 ::i.rnple:: se mues-
F.n los experimentos restantes, I& adquisicidn se h i s o directamente
L:os electrodos estaban

10
r- Parameter- F ' i l e (None) * ..
# Value Explanat ion m Stimir l .~is recovery t ime (5) i n Uuota mode 512 # o+ C;amp.les / Episode (U, 16, 24... !'76U)
L. 2 2 # o f Episodes/ T r i a l ( 1. .. 1024) 14 h0L1 C::lock I n t e r v a l (usee) í::. :36 1
(" ~
I." 1 H . 85 F-r-act:ion of episode t o take a f t e r t r i g g e r 1 9 0 üe l ay, t r i g g e r - t o - s t i mu1 cis (usee ::. 4)
c 20 0 Di.irat.icm, 5tinii.il.us (Timer 3 ) í ~ i s e c ,': 64000) 22 ....I Quota: L C J W ~ ~ l i m i t , contro: i /st . imulus episodes (-1 = Continuous inode) L'li 0 iAinta: IJpper 1 i m i t c o n t r o l / s t i m u l u s episodes s. - ,_ ... " 24 255 Number of r e t r i e s t o g e t an event ( (356 )
C..
Analog Out.pt.it , Channel 0: Hniding Level (mV) L i">' L >.I 0 .I,.'. 1. ü a i n from DCIC t o c e l l I m V 62 c e l l / mV 62 ü A C ) ._.I
c-
1- :?:5 0 Envi r orimen t a l I n f or mat. i on- .For r e f erence on l y ::,b ...I Cell i d e n t i f i e r
'lemperatcire í C) C r i t e r i o n Time (ms) (use -1 f o r zero) 'Threshold Current ( P A )
r i i j i t i n g ~ararns I..
Parameter to modify . ; E N ' l E K t o continue::.
Parameter F i l e (None) r L.... ~ # Vaii.ie Explanat ion
40 0 AC cc)~ipi i n g t ime constant (seconds) ,-41 su10 L.owpass I'i 1 ter CLitof f -frequency í t i z )
'-43 1 1:)isplay dat.a au tomat i ca l l y ai:ter t r i a l (0 = no) I:"olari.t:y? stimcilcis out ( 0 = a c t i v e 10, l = a c t i v e h i )
' f , , 44 1 45 1 F'o la r i t , y? lockout. out í '
c.1
*._ 46 1 P o l a r i t y , d i s c r i m i n a t o r i n (@=: fa l l ing edqe, l : - r i s i ng edge)
b.07 Una log I n #@I: Gain from c e l l to AUC ímV i? AiJC /pA 62 c e l l ) Debug: :Xi=dpy segments dur ing i o ; :::.l-don ' t c o l l e c t data -.,
d.<: 0 5 :<smment 49 :
f"'
r L.,
L ..
r - ! *..^
Edj.tring Params F'arameter t.n modify .::ENTER t o exit::. - Figura 2. Pardmetros de adquisición de FETCIiEX.
L.,
I

r'.
I._
# Value ?.. 1 1 " .3
? 2 1.
Parametei- F:'i I e t:.wj, t,ch E:.: p I ai-i a t 1. on
Time Elet.ween S t a r t u.f Episodes (seconds) # u+ fictnsi"rr-i.al.
# of: Episodee/Hun :1. .3:?>, 1s.t Clock Inter-val. íc.amples 1-296) .fs ( :;. 3v1, even 1-10. )
I)elay aft.er- Scope t r i g g e r (msec:) E C e t 'to 1 .for Acitromatic t r i a l r e p e t i t i o n 3 Ctart:.j,rig Episode # (normal ly 1 )
C:ommanci Mode: 2jC;tep 3 j . 2 Scaled Steps S)li'amp Cycle 6)F'rops. Pas.
i) j ,gi t .al C1~itp~its: Sample # +or- pu lse on Y 5 1 3 Kh. I First Episode a t w h i c h 951C. ch. 1 2 F'ir-e l....ast C:pisode a t which 9513 ch. 1 E* 2 F i r e Carnp1.e # +or pu lse on '7'51'5 Cli" 2
............... _ _
Analoq Output, C h a n n e l 0: i iolciina Level (nig) 2nd (I Level (rni.3) Increment 2 n d A Level. ( m g ) 2nd I3 Level í m g f Increment 2nd 13 Level ( n q ) 11i.tratrion 1st A Level (samples)
t.o change ( h i t .::FIETURN;. t u proceed) '?
r- F'arameter- F i l e t w i t c h ~ # V a l u e ExpIariatj.nri
*-..:>4: ..y ,.> v1 peak !, rng (Ramp-Clamp)
*- :jq @ _. .- ._ -
L. 51 0 (Outpcit . c:hannel 0, cont inued) Center, mg (Ramp-Clamp)
: .#y,; Ciain .+rum »AT: t o c e l l (mg @ c e l l / mV (a DAC)
- .......... .... "'I c ..::si, 0 i 6 v1 i .3 / 0
30 0 fTac:t.or- q t.o d i v i d e command vo l tage ::'7' la Niwri of average 40 ist CI Level (mg) C o n l y i n mode 63 41. VI increment i s t A L.evel (mg) I: on.iy i n mude 61 '$2 ....n0 1st L< Level (mg) I. on ly i n mode 6 3 43 v1 iricretnent 1st Ec Level (mgf Conly i n mode 6 1 44 1.IE3 Dcira.l:i.on E l l L.eve1 (samples) Conly i n mode 6 3 45 0 Increment du ra t i on A l Level r- 4ci w I ric r ement. d t.tr a t i on I3 I Level
l.,. 4 7 kl L n d Clock I n t e r v a l (samples 257--5lL), 0 =:> use #14
~ - . 4*'? ;;!n Analog I n #0: üa in from c e l l t o AD[: (rnV @ ADC /mg (3 c e l l ) Clna1c:)g i n # i : Gain from c e l l t u AUC í m V (0 (WC /mV @ c e l l ) ~ iie i
"Parame.ter t o chancle (hit *:RETURN:::. t o proceed) ?
c I _ ........ ... .... -. ... .- -,. -
CI c':
48 0 Mult ip l .exer Code: l a s t ADC: channel # ( 0 or 1 )
I
F .
6.. Fitwra 3 . Pzirámt~tros de ~ d q u i s i c i ó n de &,&p~x.
r- /<..
F-
I

12
RESUL TAD05
privacitn de hw.-
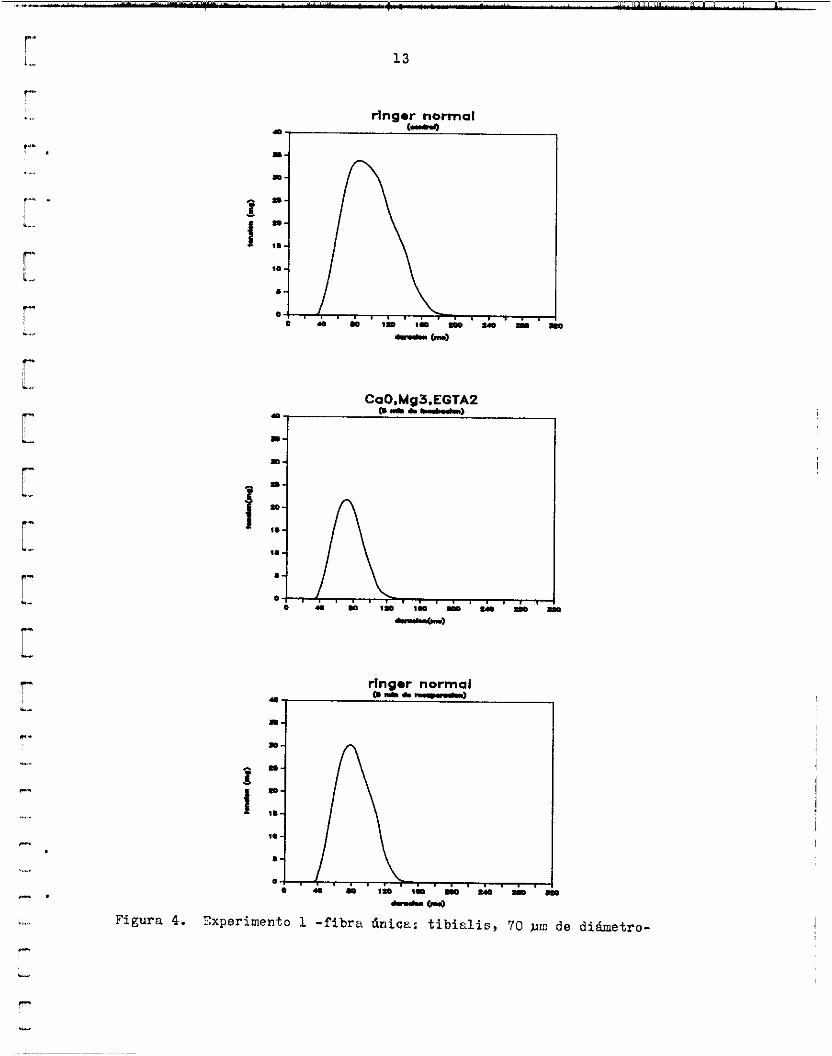
~l primer experimento fue grabado en c inta magnbtica y adquirido e11 l a computadora mediante FETCHEX. La solución prueba contenía EGTA W y la estimulación se hizo mediante dos electrodos colocados poralelamente 2 ::mhos lhdos de l a f ibra ei? toda su lcngitud. A l o s 5 minutos de estar l a f ibra en Ir solución prueba, l a amplitud d e l twitch se redujo a un 64.36% de su valor c o n t r o l en Ringer normal ( f ig . 4) . imo se redujo de 50.4 ms a 38.4 ms, 76.19% de su v a l o r c o n t r o l . La recu- peracidn de este f i b r a se midi6 snlo a l o s 5 ninutos de lavado en Ringer normel. Xn ese t iempo alcanzó e l 89.22% del control y eu dureci6n E;1 máx- i m o el 84.52i. valor c o n t r o l y en l a recuperacidn alcanzó e l 62.62&, mientras que e l g- rea de O a l máximo disminuyó de 9 1 ú mg.ms a 499 mg-ms, 49.046 d e l control, y en la recuperhcida alcanzó e l 74.7'0% ( f ig . 5). La morfología de la con- tracción no fue marcadamente diferente en l a solución sin @, S i n embar- go, hay algunas diferencias s ignif icat ivas en l a s pendientes a l i n i c i o de l a fase ascendente, medidas entre los 2.4 y 12.0 m s de tensidn positiva. La5 pendientes de l a s curvas para ese intervalo fueron 0.6285 mg/ms (Rin- ger normal), 0.6242 mg/ms (CaOYMg3,EQTA2) y 0.6515 &/ms (Ringer norml, recuperacidn).
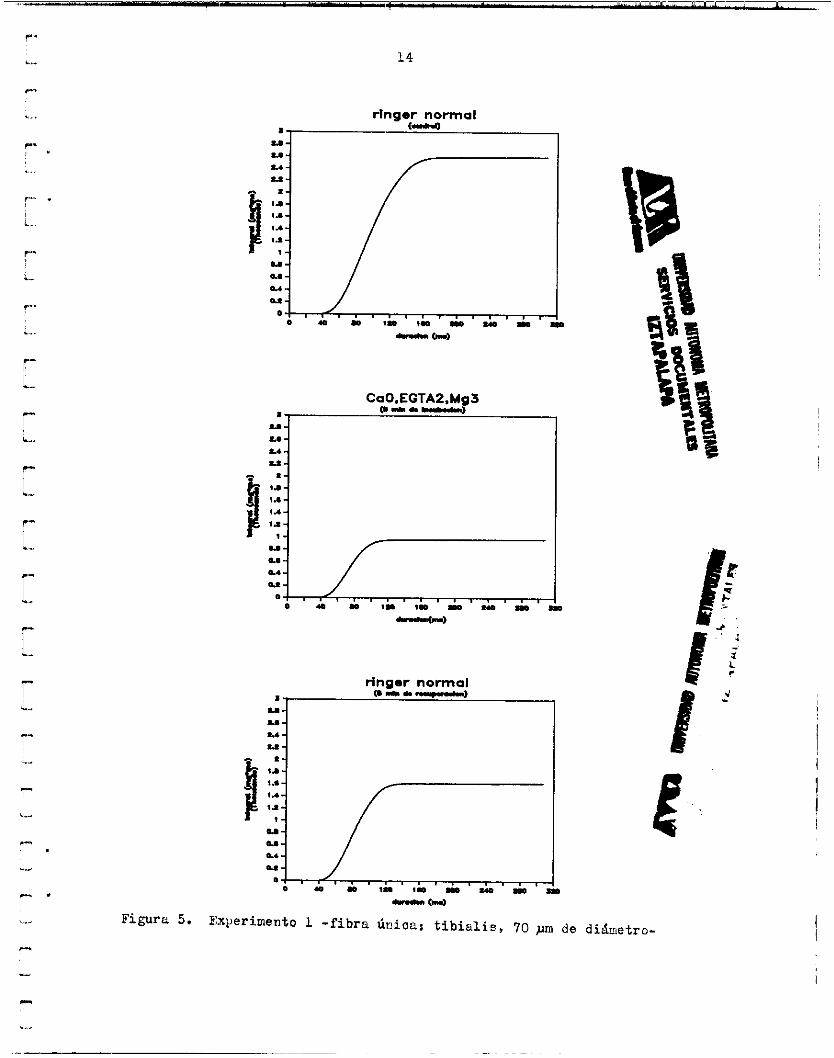
es t imlacidn se h i z o mediante un par de electrodos de alambre f i n o de plata situados cerca de uno de los extremos de la fibra ( e l extremo fi- j o ) . La disminución de l a contracción observada en el primer experimen- t o se r e p i t i ó consistentemente en l o s d e d s ( L O en total) . Sin embwgo, l a magnitud de dicha disminucidn y e l curso temporal de l a recuperación no fue similar en todos l o s casos. En algunas f ibras , l a tensidn se re- dujo sólarnente a un 8G.06$ d e l c o n t r o l y , en otras , l a contracción prác- ticamente desapareció (0.24% del control). En promedio, l a tensión dis- minuyd a un 34.20'24.70$ del c m t r o l y l a duracidn a l máximo a un 76.902 21.13%. En general, se observd que l e caída de tensión en l a fase de re- la jac ión de 1.8 sacudida fue I& suave en CaOYYg3,EGTA1 que en Ringer nor- mal. Debido a ésto , l a duracidn total de la contraccidn en algunos casos no disminuyó notablemente en l a solución prueba, mientras que e l tiempo a l máximo s í lo hizo. Las áreas tota les se redujeron a un 31.04-27.80$ del control y l a s áreas de O a l dximo a un 30.64&29.18p (Los valores se- ñalados son promedios de 9 f ibras y desviación standard).
En todoc; los casos considerados hubo recuperacidn de l a f ibra en Ringer normal. Descartando dos casos en que hubo potenciaci6n de l a sa- cudida u1 volver a bañar l a f ibra con Ringer norm1 (ver abajo) y uno en que l a f i b r a seguramente se dañd, l a magnitud alcanzada entre 10 y 49 m i - nutos de racuperac~ión fue del 95.03f6.12$ del valor control (promedio de 5). La mag- n i t u d de l a contracci6n después de 20 rninutos de recuperación en e88 so- iucidn fue del 97.927; del valor original an Ringer normal.
registra en e l aparato Gould a a l t a velocidad.
La dureción a l &x-
E l Brea t o t a l b a j o la curva se redujo a un 37.42;'. de su
En l o s siguientes experimentos, se u t i l i z ó sieffipre EGTsilu@d, y l a
En un experimento e l lavado se h i z o con CaO,Mg3 ( s in EGTA).
La figura 6 muestra e l curso de un experimento t ip ico , t a l como se En este caso, l a magnitud
P f
P f
13
ringer normal (rnr)
-w
a '1
ringor normal On**- ..
o
Figura 4. Zxperirnento 1 -f ibra dtiice: t i b i a l i s , 70 nim de diámetro-
14
ringor normal (rrrg
J 7 i u- U- la-
) /;: M,
CaO.EGTA2.Mg3
la
ringer normal
U la
(;i g a ; c
Figura 5. Experimento 1 - f i b ra único.: t i b i a l i s , 70 ~III de diámetro-
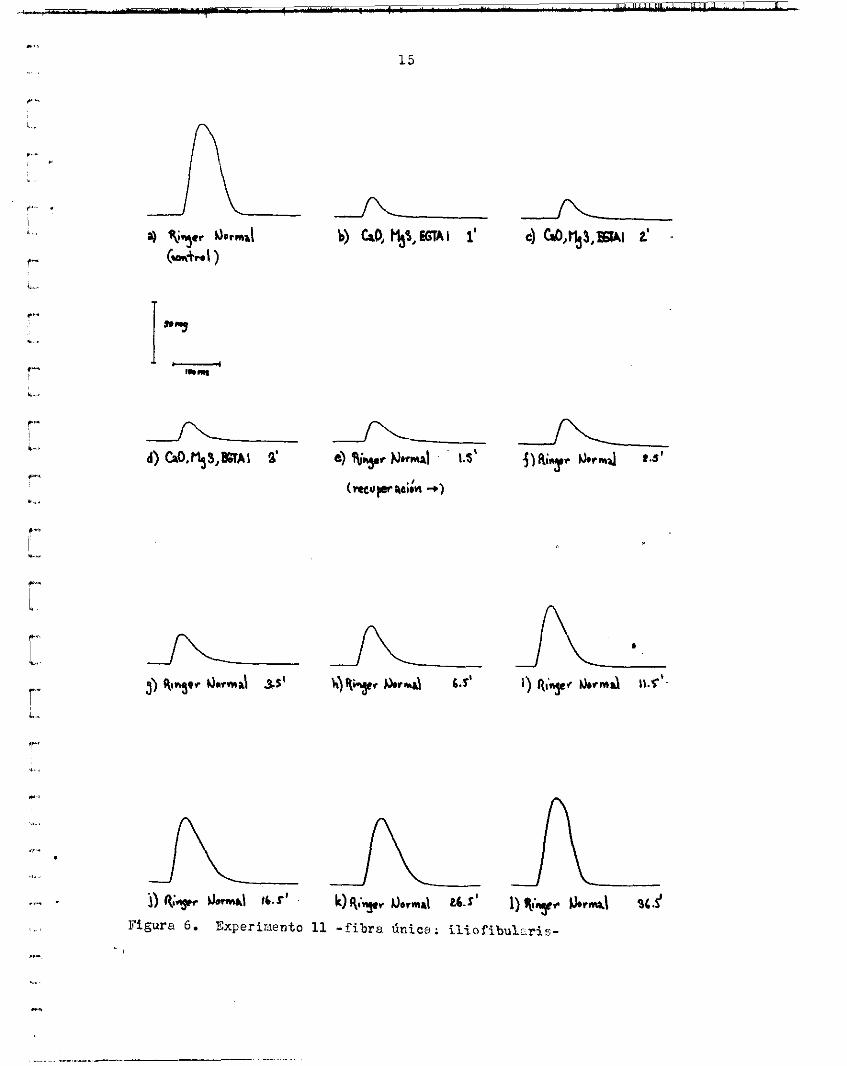
1 5
n
c

de l a contracci ln disminuyó a un 21.22.$ d e l contro l (promedio de 3 sa- cudidas) y se fue recuperando paulatinamente en Ringer normal durante 3u.5 minuto:i, h a s t a a lcanzar e l :LOO,;. El á r e a t o t a l ba jo l a curva (me-- dida de O a 120 ms) disminuyó a un 14.9bP y e l a rea de O al máximo u un 10.29% en la solución prueba (CaiI,tlg3,XG'T~l). E l Aros cubier ta despuks de 31i.5 minutos de recuperación es igua l p o r e e l i n t e r v a l o de O al d x - imo y l igeramente menor para e l i i i l e rva lo de l á r e a t o t a l (91.48% del con- t r a l ) . La durr ic ih de l a contracción (de O a l máximoj disminuyó n un 50s d e l contro l y aumentó nuevamente en Xioger normal, clcanzando e l ve lor coritrol en :!6.5 minutos.
cu i r ido directnnente en le comimtadvra con G¿;I,YYX. La tensión dismi- nuyb a 4¿. l5$ de l control (prcmedio de 5 sscudidas) entre 5 y 1 0 m i n u - t o ? en &OOMg3,PC'í'A1 y l a d u r o c i h a l tn5ximo .fue de 65.61,; de l c o n t r o l . il? tensión se recu:erÓ graduaimente durante 25 niinutos, alcanzando e l 92.31$ de su magnitud o r i g i n a l eri Ririger normal. Los valores r e l a t i v o s de l a s &reus t o t a l e s (de O i? 120 ms) para Ringer normal ( c o n t r o l ) , CaO, U ~ ~ , Z : I ; T K ~ y Ringer normal ( recuceracihn) fupron loo$, 33.337; y 94.735 respectivamente; para l a s 6 r e : : ~ ate O u1 dximo, e s t o s valores fueron 100$, 37.12$: y 96.@9$.
Como ;'n se mencionó, en un ensayo e l lava.do de recuperación se h i - zo en Ringer s i n Cal' y con Ei:$3mh'l, pero s i n EGTA (CaO,Xg3). En l a fi- guru 6 eparecen l o ~ t razos de l a tens ión corres; ;ondientes a Ringer nor- w-1, CaOyKg3,EGTrll y CaO,Vi3 en e l oec i loscoy~io con d o s barr idos d i f e - r e n t e s . k d e d s , s e muestra con mucha a m p i i f i c \ ~ c i b n e l i n i c i o de l a con- t r a c c i ó n y se ve ,;ue también hay un e f e c t o sobre l a r e l a j a c i ó n de i a t e n - c i a . La u n i . , l i t u d di: 1~ sacudidp. ue redufio a un 37.167; de l va lor en Rin- ger normal y el" tiempo a l mhxiico a un 76.477; (promedio de 2 sacudidas j . L:i f c s e de r e l i i j a c i ó n se hizo más suave y l a duración t o t a l de l a con- t r a c c i ó n no disminuyó n3tablemente. La amplitud de l a r e l a j a c i d n de l a t e n c i a en CaOYMg3,EGTAl disminuyó d 20 @g a partir de 50 pg en R i n - g e r normal y en CaOYb(g3 zu valor e s i g u a l a l control . L a duracián de 1.1 l a t e n c i a , medida de l punto de est imulaci6n a l i n i c i o de generaci6n de tensión p o s i t i v a , se redujo de 5.4 ms a 4.8 mo. En e e t e último pun- t o , l a s pendientes en 3inEer normal, CaO,klg3,XGTAl y CuO,Ug3 fueron r e a - pectivamente 0.2133, 0.1111 y 0.2156 mg/ms ( l o cue1 i n d i c a que l a ve lo - cidud con q u e se VC generando tensinn a l i n i c i o de 1c1 contracción es menor en CaO,Ng3,EG'I'I.l y l igeramente mayor en l a recuperación con CaO, Mg3, en comparación con e l va lor en R i t i y r normal).
í.n l e misma f i b r e se ;'robó d P s p u é S In secuencia Ringer normal, CaOli4g3,CGT,YL, Ringer normal y se obtuvieron nuewmente l o s r e s u l t a - do$ de disminución de l a terisión, seguida de una recuperación paulo- t ina ( f i g . 9).
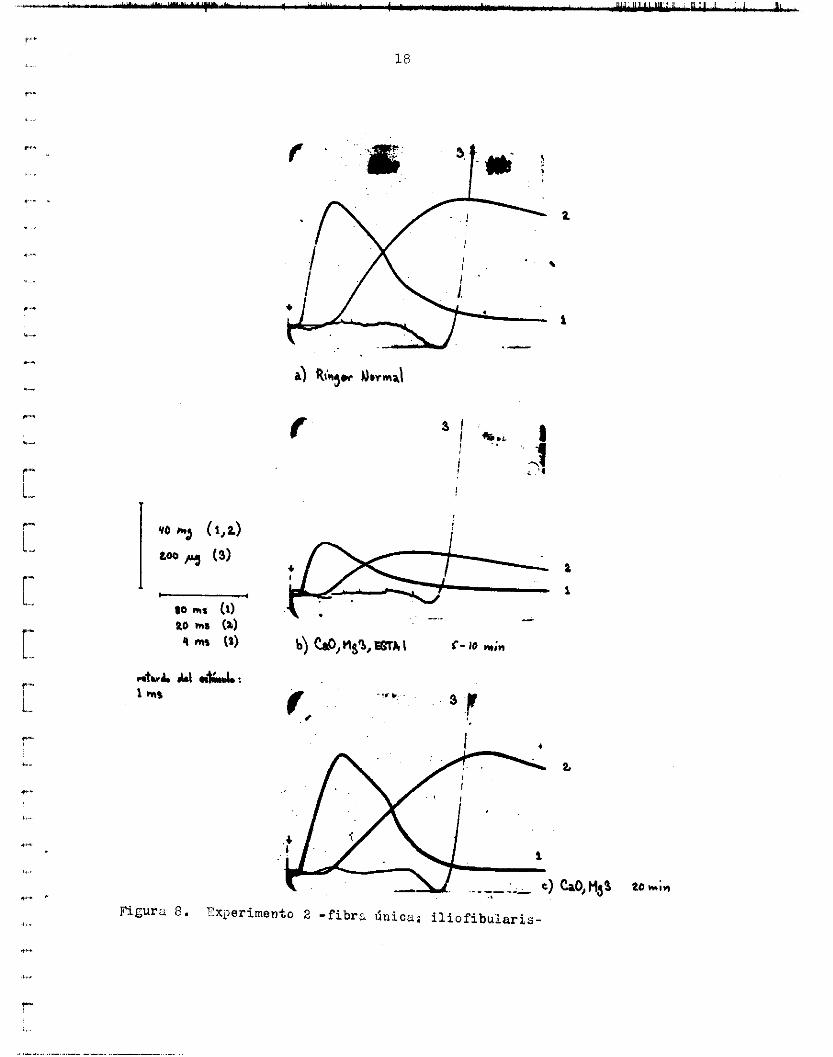
s i ó n y e l potenc ia l de acc ión e x t r a c e l u l a r . En uno de e l l o s ( f i g . lo), l e amplitud d e l potenc ia l en Ringer normi.1 y en CnOYE,~3,F:GTAZ e s igual (0.12 mV). La forma y dur>ición también eon i g u a l e s en ambas soluciones (durución t o t a l : 1.4 ma; del pico p o s i t i v o a l negat ivo; 0.3 a s ) . La ve- loc idad de conducci6nY s in embargo, aumenta en CaQ,Mg3,EGTM, y a que e l tiempo entre e l i n i c i o del Firtefacto de l est imula y e l i n i c i o d e l poten-
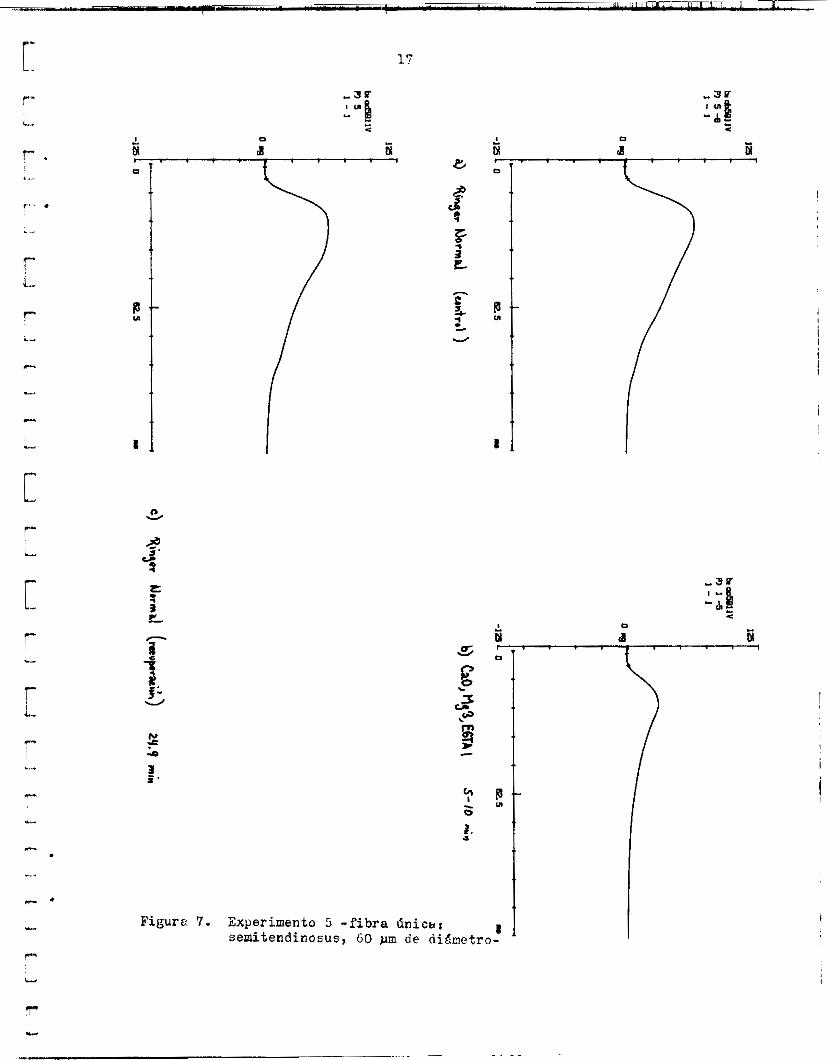
IC1 experinento de l a f igura 7 e s eirnil2.r a l a n t e r i o r , tjero fue ad-
En los :s iguientes experimentos, se r e g i s t r ó simultánenmente 1 ~ . ten-
ái r o
P Ll
8
Figura 7. Experimento 5 -fibra Cinicar semitendinosus, 60 pm de di.4netro-
CI
L
.- L..,.
.- L"
v-.
1.-
c
i,.
r.*
1 mr 3 p .r c
Figura 8 . ?x,>erimento 2 - f ibra Única: iliofibularis-
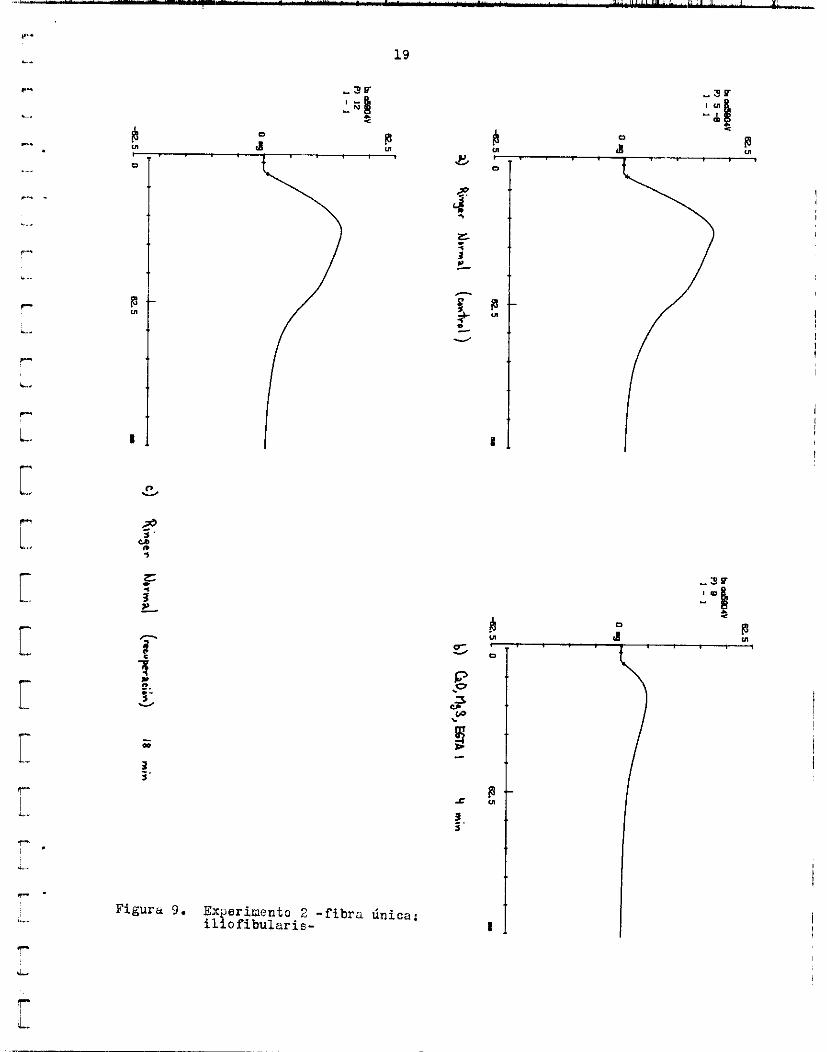
19
B ln I-
O
P YI
e
20
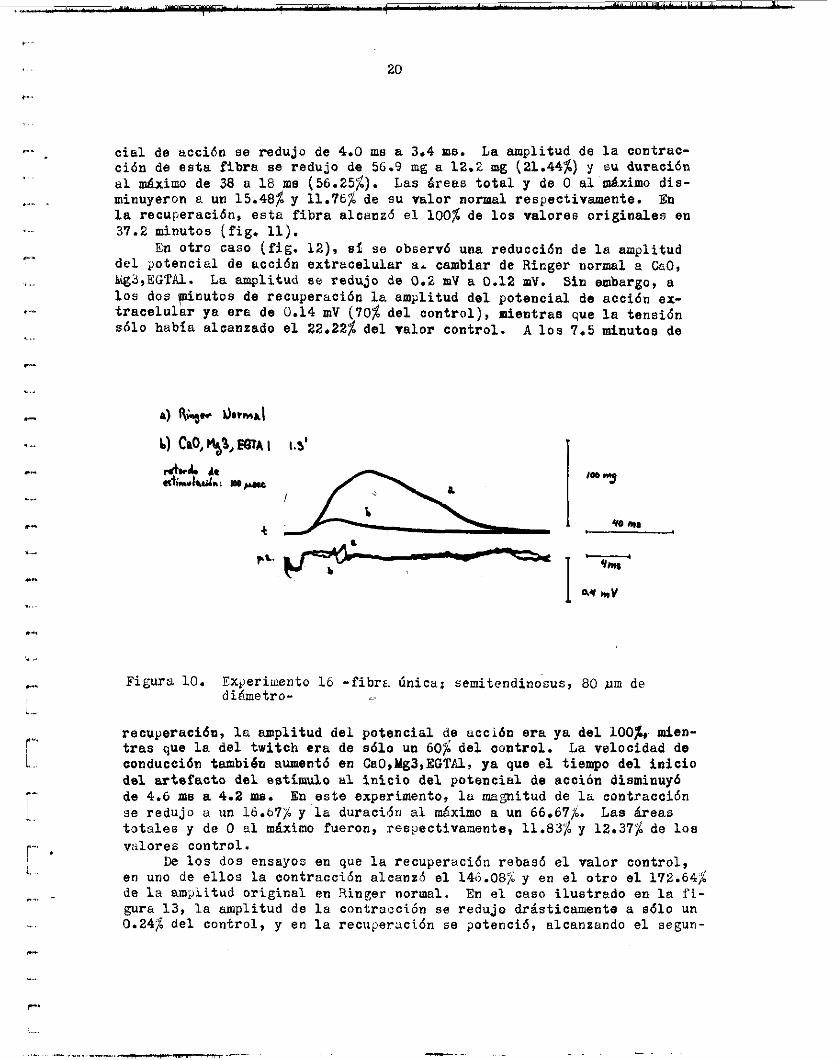
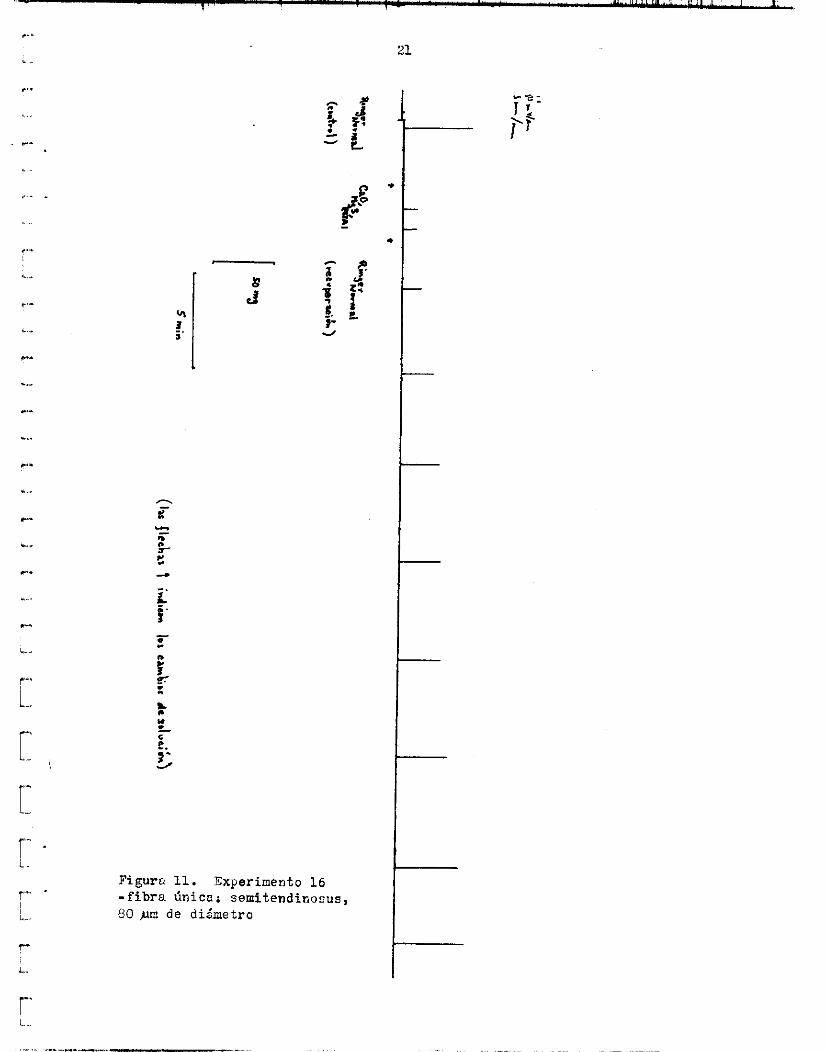
c i a 1 de a c c i 6 n s e redujo de 4.0 ms a 3.4 me. La a m p l i t u d de l a contrac- c i ó n de e s t a fibra se redujo de 56.9 mg a 12.2 ng (21.44%) y su duracidn a l máximo de 38 a 18 me (56.25%). Las á r e a s t o t a l y de O a l máximo d i s - minuyeron L: un 15.48$ y 11.767; de su v a l o r normal respectivamente. l a recuperacidn, e s t a f i b r a alcanzd e l 100% de l o s v a l o r e s o r i g i n a l e s en 37.2 minutos ( f i g . 11).
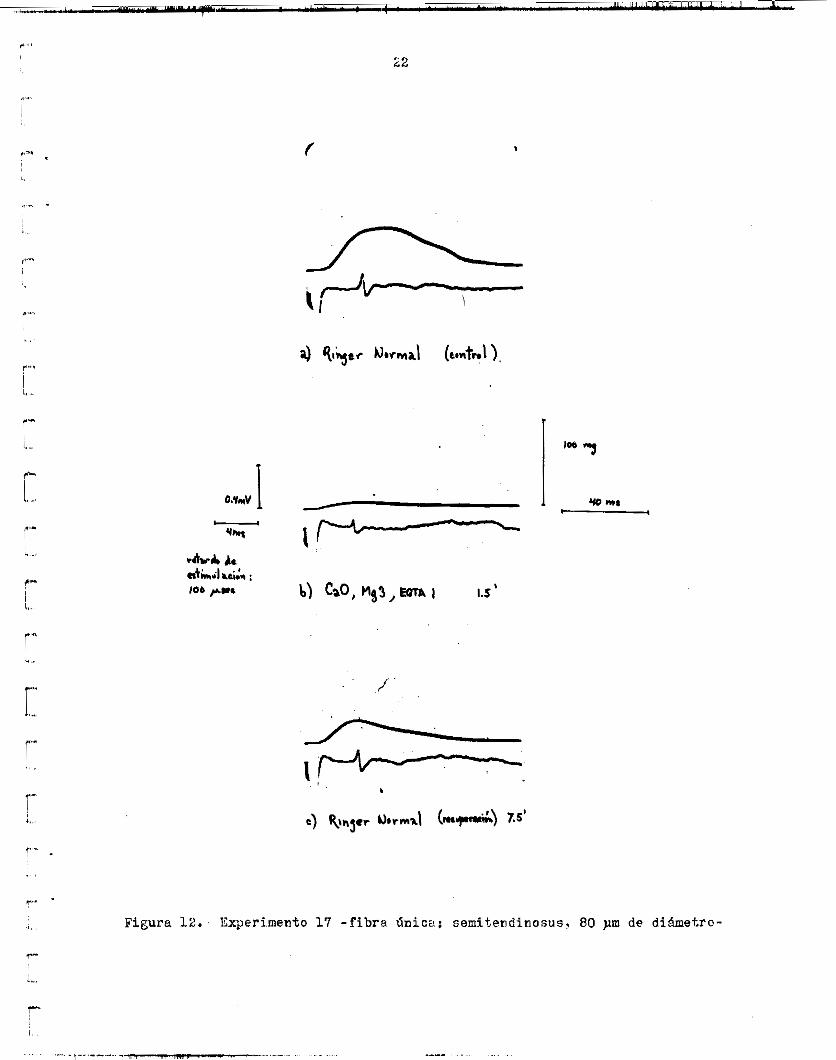
En o t r o caso ( f i g . 12), si se observó una reduccidn de l a a m p l i t u d d e l gotencia1 d e a c c i ó n e x t r a c e l u l a r a i cambiar de Ringer normal a &,O, Ligs,EGTU. l o s dos p i n u t o s de recuperación In a m p l i t u d d e l p o t e n c i a l de accidn ex- t r a c e l u l a r ya e r a de 0.14 mV (70% d e l c o n t r o l ) , mientras que la tensidn sólo había alcanzado e l 22.22$ d e l v a l o r c o n t r o l . A los 7.5 minutos de
En
La amplitud se redujo de O.% mV a 0.12 mV. S i n embargo, a
t
PL\..
1.3'
Figura 10. Experiiuento 16 - f i b r a Única: semitendinosus, 80 urn de d iáme tr o - c
r e c u y e r a c i b o , l a a m p l i t u d d e l p o t e n c i a l de acción e r a y a del 100% mien- tras que l a del t w i t c h e r a de sólo un 607: d e l c o n t r o l . conducción también aumentd en CaO,Mg3,EGTAl., ya que e l tiempo del i n i c i o del a r t e f a c t o del est ímulo a l i n i c i o d e l p o t e n c i a l de acc ión disminuyó de 4.6 us a 4.2 ma. s e redujo a un 16.671. y l a duración a l máximo a un 66.67jb. t o t a l e s y de O 01 m&ximo fueron, respectivamente, 11.83% y 12.37% de l o s v d o r e s c o n t r o l .
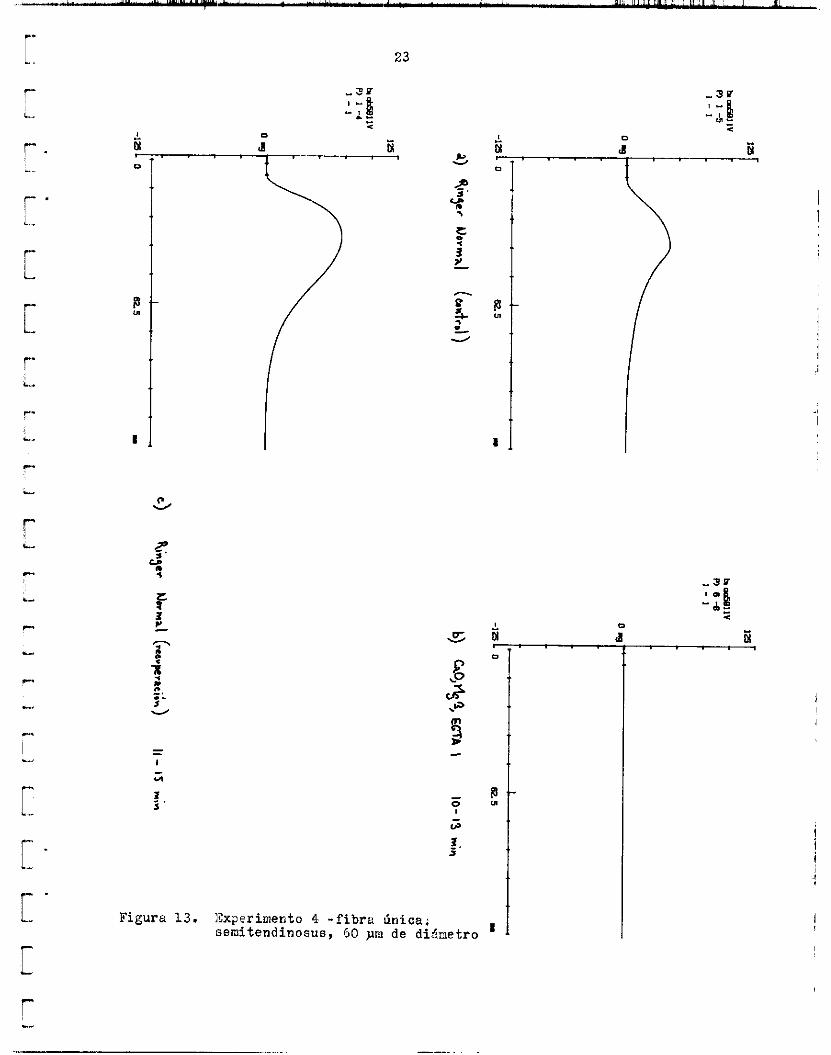
De los d o s ensayos en que l a recuperación rebasd e l v a l o r c o n t r o l , en uno de e l l o s l a contracción a l r r t n z ó e l 14~.08,~ y en e l o t r o e l 172.64;6 d e la a m ; i l i t u d o r i g i n a l en Ringer normal. En e l caso i l u s t r a d o en l a fi- gurb 1 3 , l a a m p l i t u d de l a contracción se redujo drásticamente a s d l o un 0.24$ d e l c o n t r o l , y en l a reciiSerdción se p o t e n c i á , alcanzando e l segun-
La velocidad de
En e s t e experimento, In magnitud de l a contracción Las á r e a s
-
f Y
3- 6 c. c/
Figura 11. Experimento 16 -fibra único2 semitendinosus, 80 ).un d e d i h e t r o
21
t-
7.2
f
i ,&- \
Figura 12. E x p e r i m e n t o 17 - f ibra Única: s e m i t e n d i n o s u s , 80 jan de diámetro-
23
D c 5 5 P
.e
- B - O UI
w - z. J
I 13xperimento 4 -fibra Única; semitendinosus, 60 JIN de diánetro
Figure 13.
I '
I
.I
I

24
do = l o r a r r i b a mencionado. p l i c ó durante e l lavedo de recuperación.
paulat ina . nutos de lavado en Ringer nor1:ml s e rijuotarrm a una función exi ,onsncir l
$1 á r e a bajo l a curva p r a c t i c a n e n t e se du-
Como ye s e mencionó, l a recuperación en l a izayoría de l o s casos fue Lao amplitudes de 15s contracciones e n t r e O y de 20 a 40 mi-
d e l t i p o I -I-=TG#(I-+-~$)) , donde 'i&, eo l a tension or ig ina :L en Ringer n o r n n 1 , Z e s una c o n s t t n t e de tiempo, t. s i t ú a un i n i c i o de L a curvs y 'f ea l a tensión de 1 2 1 f i b r a en el tiempo t.
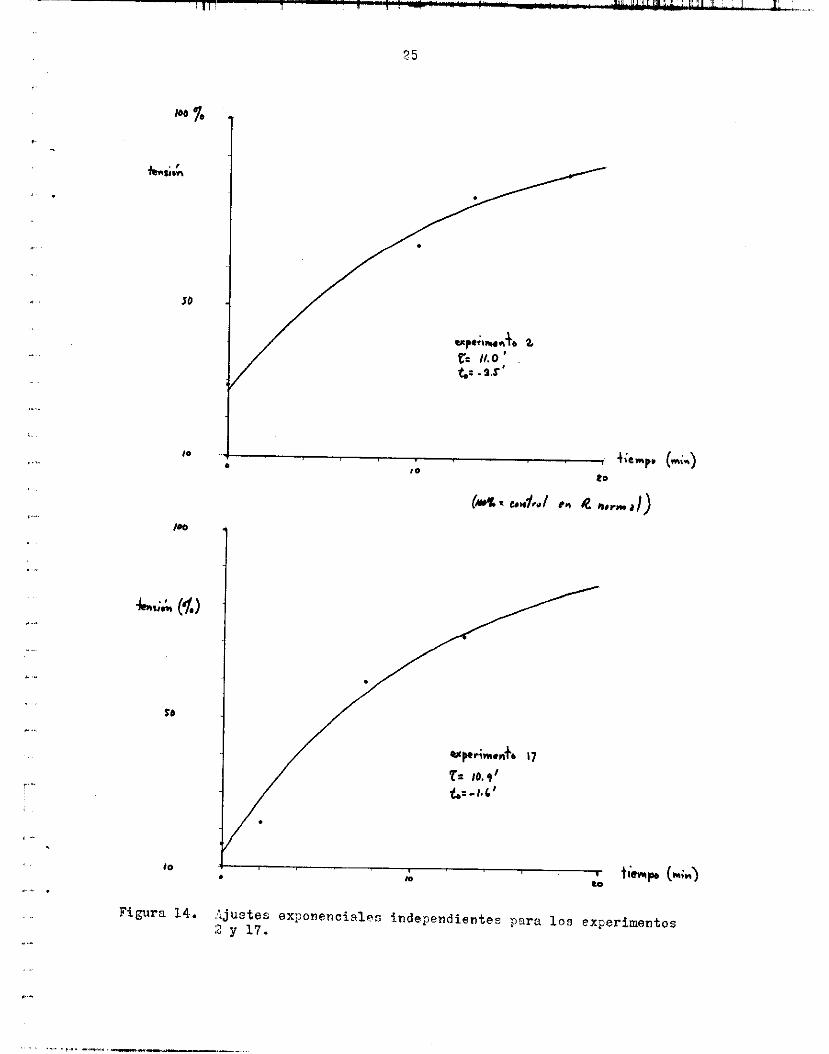
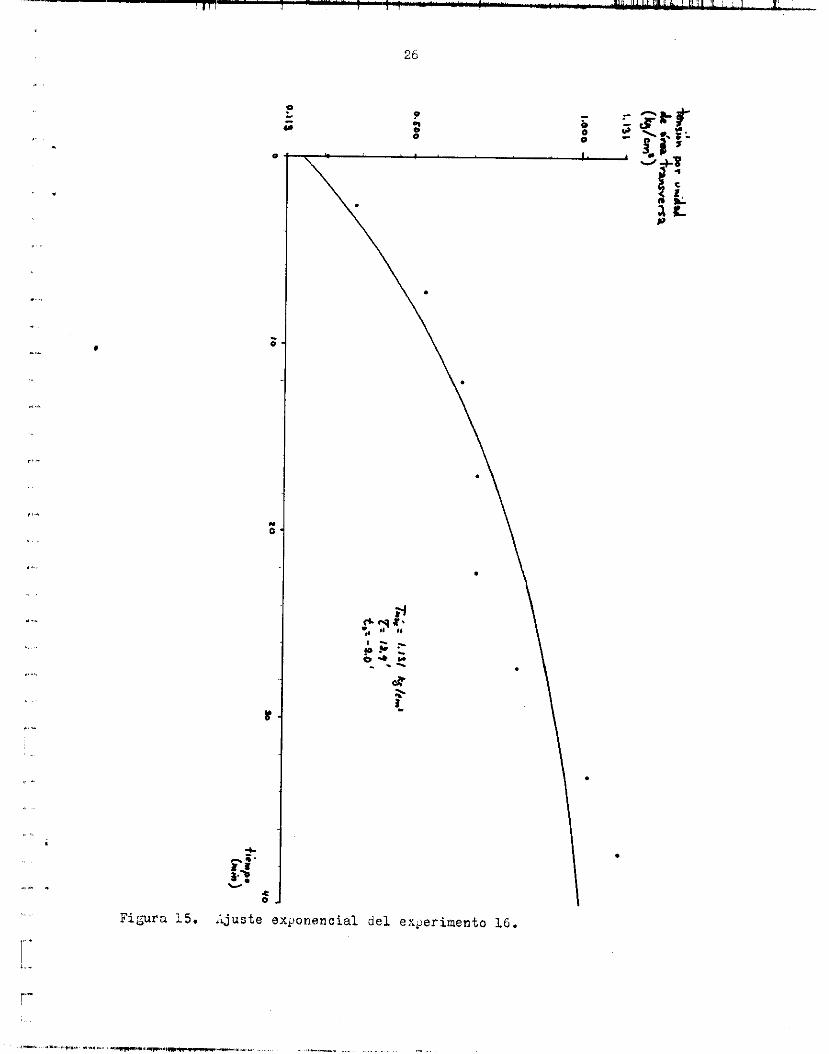
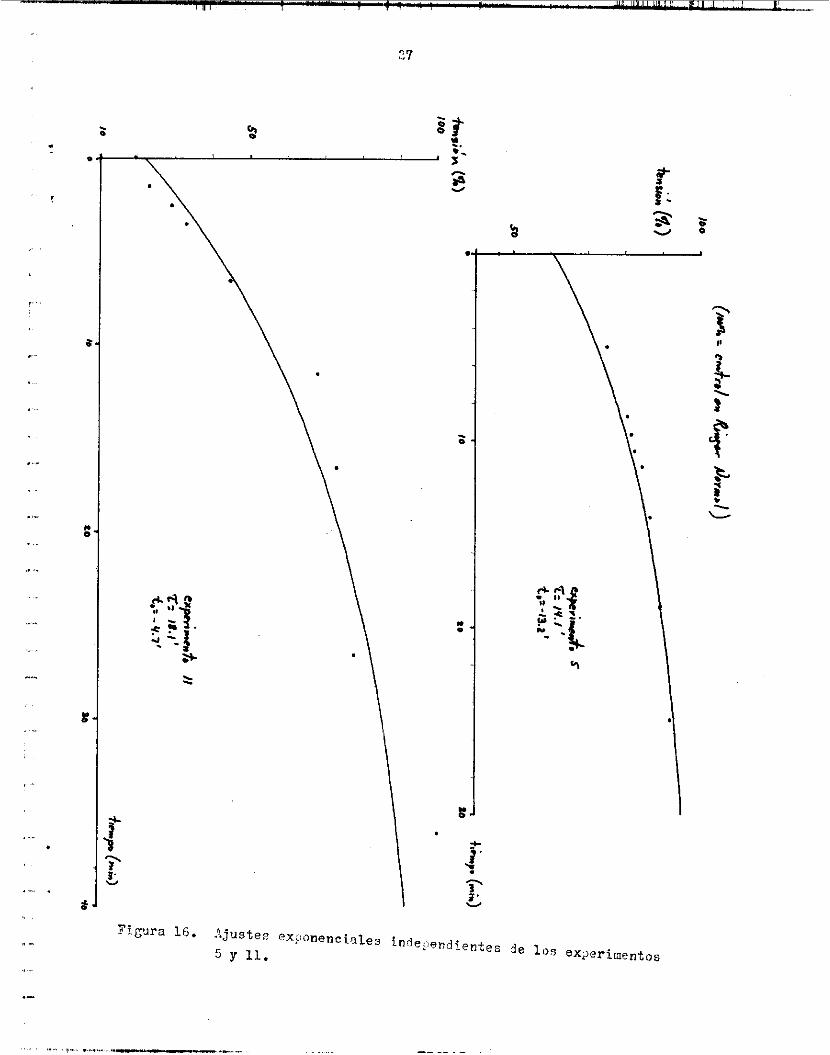
Lr.s f igura6 14, 1 5 y 1 6 muestran, independientemente, l o s puntos ex- perimentales de 5 experimentos :[ 16 curva exponencial correspondiente. En uno de e l l o c se expresa l a tensidn en tenoión/érea t r a n s v e r s a . t a n t e de tlempo promedio fue 14.6e3.8 m i n .
s ión en do:; experimentos con Z semejante y e l a j u s t e exponencial corres- pondiente.
pende de la del p o t e n c i a l de acc ión ( e l cual ,ia s e v i ó iue medido e x t r a - celularmente no s e modifica s i g n i f i c a t i v a m e n t e o s e recupera m6s r i p i d o que l a c o n t r a c c i ó n ) , y l b de Bste , a. su vez, de l a magnitud de l a depo- l a r i z a c i o n n e c e s a r i a pera :!lcaneur e l umbral e l é c t r i c o , semidid e l po- t e n c i a l de reposo en f i b s a s expuestas a CoO,Fdg3,EGTPl. Los r e s u l t a d o s s e resumen en l a s i g u i e n t e t a b l a .
La cons-
Los flgurae 1 7 y 1 8 muestran, cade une, le recupereción de l a t e n -
Puesto que la a m p l i t u d de :Ir; contracción en condi.ciones normales de-
Solucidn . No. de f i b r a s P o t e n c i a l de feu080 Des. standart
R. normalf 11 -95.0 mV 3.1 CaO,Yg3,EGTAl 1 5 -90.7 mV 3.2 R. normal" 20 -96.0 mV 3 . 1 -
* c o n t r o l '*des&s de la. solucidn prueba
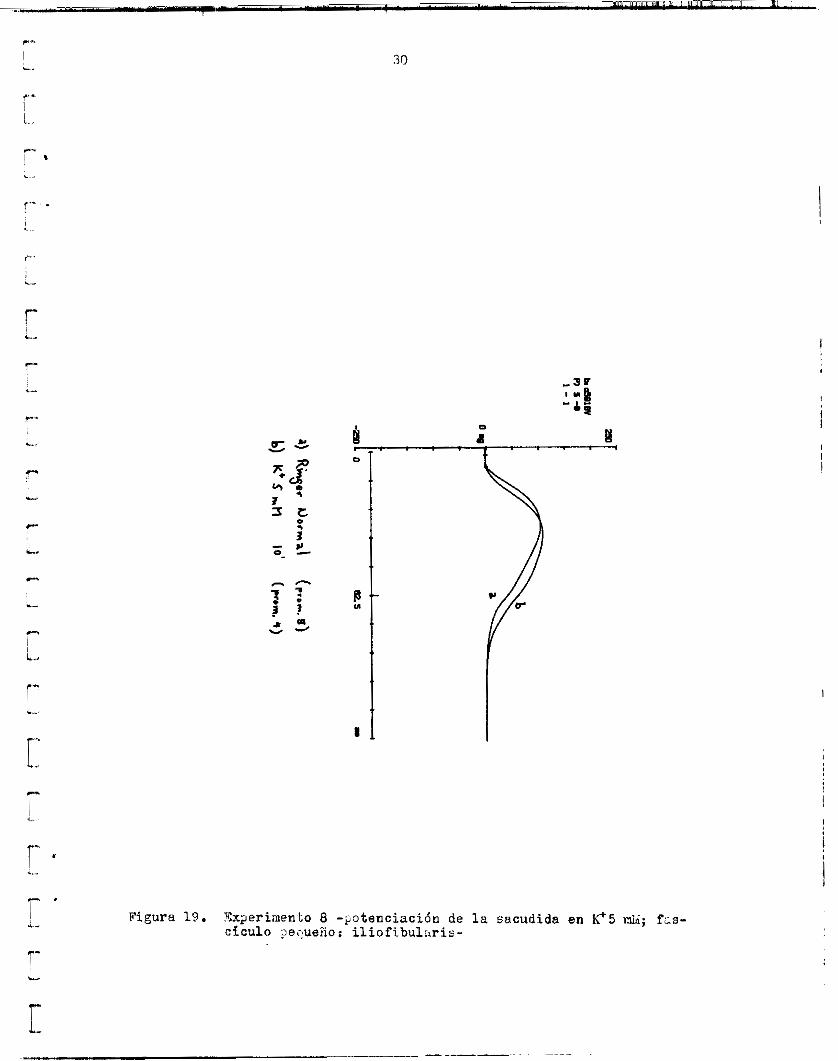
Se r e ~ l i z d o t r o experimento en que s e r e g i s t r d l a tensión en Ringer normal y despues en Ringer con 5 mM de K+. E s t a solucibn, de acuerdo con l a ecuacidii de Goldmann ( K a t z , 1 9 7 9 ) , produce una depolár irac i6n de 12nV, de modo que e l p o t e n c i a l de l a f i b r a s e encuentra más abttjo (en v a l o r ab- e o l u t o ) aue 1 0 5 -90.7 mV r e g i s t r a d o s en Ca09Mg3,EGUl en e l experimento a n t e r i o r . c idn l e v e , l e contraccidn se potenció en 5 mifi K+(fig. 19) . La a m p l i t u d aumentd 2.5 mg (2.381.) y e l tiempo a l máximo 3.5 ma. E l área a i máximo aumentó 118.b mg.ms (7.07%) y e l á r e a t o t ú i 485.8 mg.ms (10.87$).
Contrariamente a l o (que se podria esperar de l a d e p o l a r i z a -
E f e c t o d e l Bp.-
En Ocho ensayos r e a l i z a d o s se observo consiatentemente un aumento En promedio, l a am- de la t e n s i ó n a l sust i tuir e l C P por B8' 3.2 RIM.
p l i t u d de la contracción aumentJ a un 111.49+10.41~ d e l c o n t r o l duran- t e e l primer minuto de i n c u b a c i h en %*. Durante e s e primer minuto, e l
25
I O
Figura 14.
I tienp (mi.) . .O
)o
:justes exponencialas independientes para los exi>erimentos :z y 17.
26
. .. *
? 2 o
\.
. F i a r a 15. Lxjuste exponencial del ex,erinento 16.
-
;u.--
?,?
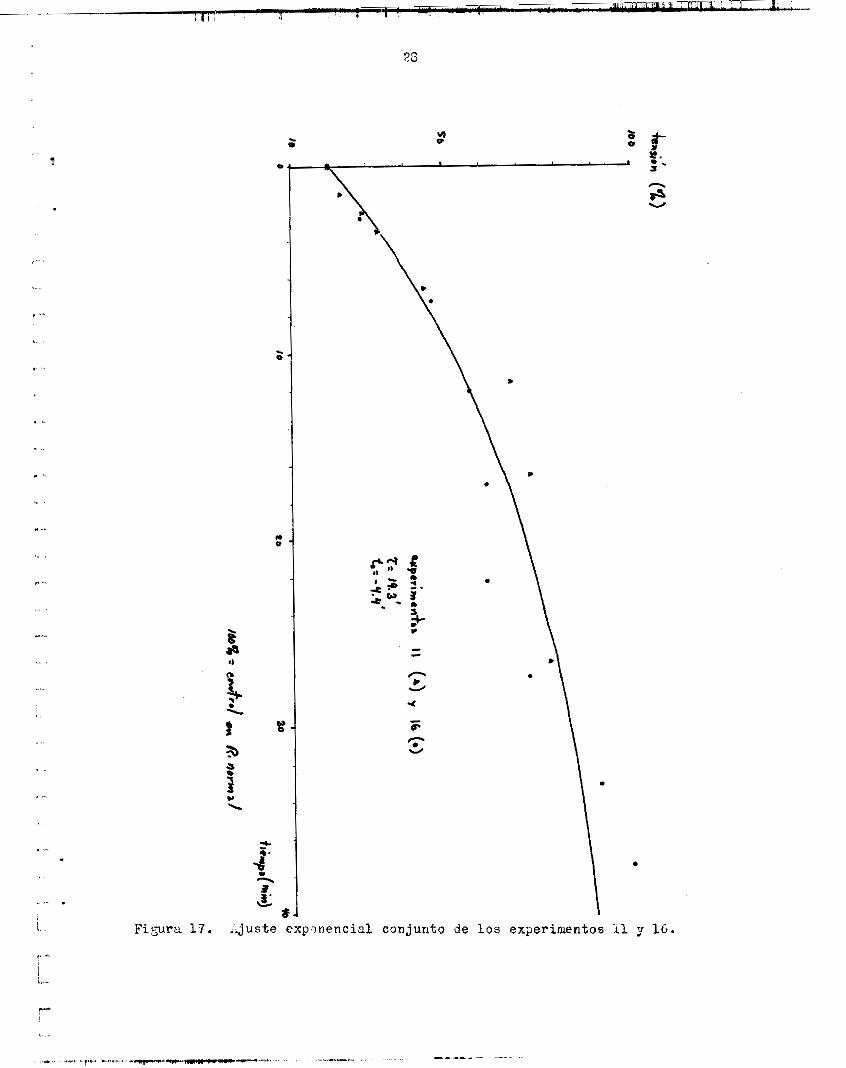
. Fi:ura 17. ..juste e x p l n e n c i a l c o n j u n t o de los experimentos 11 y 1b.
I .~
I..
...
*_
*“I
-...
”“.
100
so
o
,. .. . , . ... .
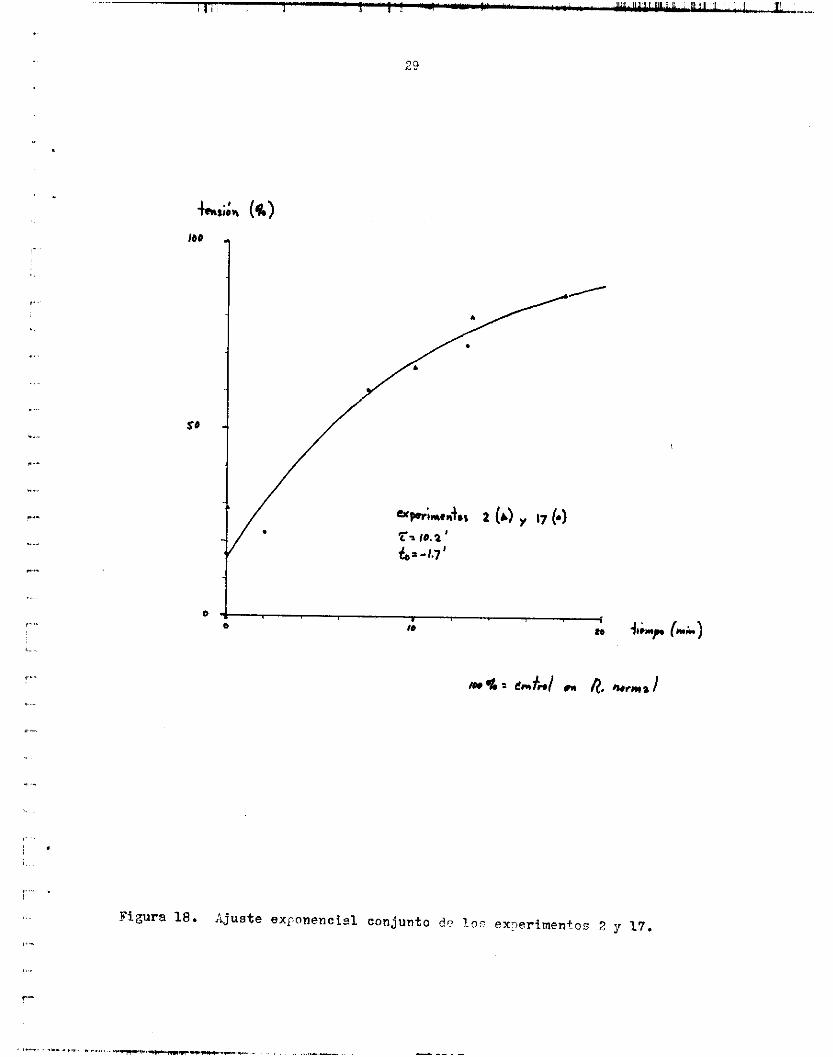
Figura 18. Ajuste exponenc is l conjunto d e los ex?srimentos 2 y 17. 4,.
,,..
w
30
f .
f ' L_
*-
F i g u r a 19 . Tx;ieriinenlo 8 - r o t e n c i a c i ó n de la s a c u d i d a en k*5 di; f,s- c i c u l o ,>equehor i l i o f i b u l s r i s -
I
!

31
comportamiento de las f ibra6 no fue muy homogéneo. mentd e l t iempo a l máxiloo, mientras que en dos disminuyó y en otras dos se mantuvo constant?. Tanto e l &rea hital como de O a l dximo aumenth en 5 f i b r a s y disminuyó en l a s o t r a s t r e s . Xn promedio, el Area to ta l au- nentó a 110.95~21.61;;b y el &ea de O a l máximo a 118.11t24.O2$ del con- t r o l (prom. de 8).
t iempo a la solucidn prueba, l a .tensión s iguió aumentando, a p r i x i d n d o m a un dximo. l a soluci6n prueba, a l comenzar e l lavado en Ringer normal,tendiÓ a se- guir aumentando l a tensión durante var ios minutos en algunas de e l l a s , y en o t r a s empezó a descender inmedtiitamente, pa:iroxidndose n un valor fi- n a l , e l ciral. a vcces f ue e l vaioi- c o n t r i t y o t r a s veces otri valor.
En l a figura 20 se muestra tm experimento reg is t rado con e l Gould, en e1 cual Lie observa un aumento gradual de l a t e n s i h durante 17 minu- t o s . Ls t e n s i ó n a lcanza un ii8.:!5$ d e l contro l . E l Area t o t a l aumenta. h a s t a i23.42$ y e l tire, a l mdximo a un 124.56,; de l V a l o r or ig in : . i . pués de alcmzar un m6ximo, 1% t í m s i ó n y e l á rea coriienzun a d i s m i n u i r . S i n embargo, a 1 0 s 23 m i n u t u s se observa un rlevamiento brusco de la ten- s i ó n , seg i ido de una disminución consideruble. La tenui6n se mantuvo más o metios cons'tante durainte l o s s i g u i e n t e s 10 minutos. 21 tiempo t a t a 1 que e s t a f i b r a estuvo ex;juestx a l a s . i luci6n c3n Si' fue 28 uiniitos.
tan notablemente dursnte 30 ;uir.ut,vs de expos ic i Jn a iia 3.2. Las magni- tudes de 1;i tendión, h . 1 t o t ü l y á r e a al d x i m o d c m z ü n un dico I l a s 20 minutos en l a w l u c i ó n p u e b z , y sus valores fueron, respectivamente, 171.58& 169.10!: y 177.31): de 10.1, originales. .A1 lhv-r l a f i b r a nueva- mrnte c m Rinser n o r m ? . , comenzó c d i : i m i n u i r l a tensión y e l área. Xn esa f igura puede a p e c i n r s e que li velocidad de generación de tensión en l a fase ascendente d e l twitch e s mayor en Ba 3.2 que en Fiinger normal.
3i6n y e l 2otenci t i l de acc ión e x t r u c e l u l a r . En e l primero ( f i g . 22), l a f i b r a estuva3 e x p e s t a a la solución con B$+ por 3 minutos, en i o s cuaiec se prodiijer'jn 3 sacudidas simples. La umplitud de l a contracciSn aumen- t ó a 112.00)& eL área t o t a l a 108.80;. y e l dren 01 máximo a 111.54% de su5 valores c o n t r o l , y se recuperaron en 5610 5 minuto.; a1 lavarse l a fi- bra nuevamente con Ringer normal. 21 tiempo 3 1 m5xiuo y ls morfologia iie Is contrucciN5n no s e modificeron notablemente. Tl potenc ia l de acc iSn au- mentó 0.02 mV en umpli . tud y su dur-cibn disminuyb 0.2 ms. La vei,,cidad de conducción ailment6 l igeramente, ya que e l tiempo entre e l i n i c i o de l s r t e - f r c t i ) de est.imui:ición y l r i ,:rimera def lex ión iositiv:i d e i poteticid1 de ciSii dismiriiiyó 0.2 ms.
que e l ex2erlmento de l a f i g u r a 113) se observó también un aumento de l a tensión y de l a s Areas t o t a l y a l máximo (fig. 2 3 j . dos a l o o 4.2 minutos fueron resi~i?ctivamente 121.74$, 127.RSj0 y 182.48,. de l o s contro les . Es i n t e r e s a n t e , con rec;,-ecto a l a morfologia de estar: sacudidas, que hay, además de l máximo, o t r o p i n t o en e l cual 18 pendiente cambia más o menos bruscamente (aprox. a l ' J S 17 ms). E s t e punto, en F i i n - ger n o r m l , se encuentra antes d e l d x i m o , y, a l aumentar l a amplitud de
En tres de e l las au-
En 1 0 3 experimentos en que se dejaron l a s f i b r a s expuestas p o r d s
Sa l a s fibras que 3610 se dejaron e n t r e 1 y 3 minutos en
Des-
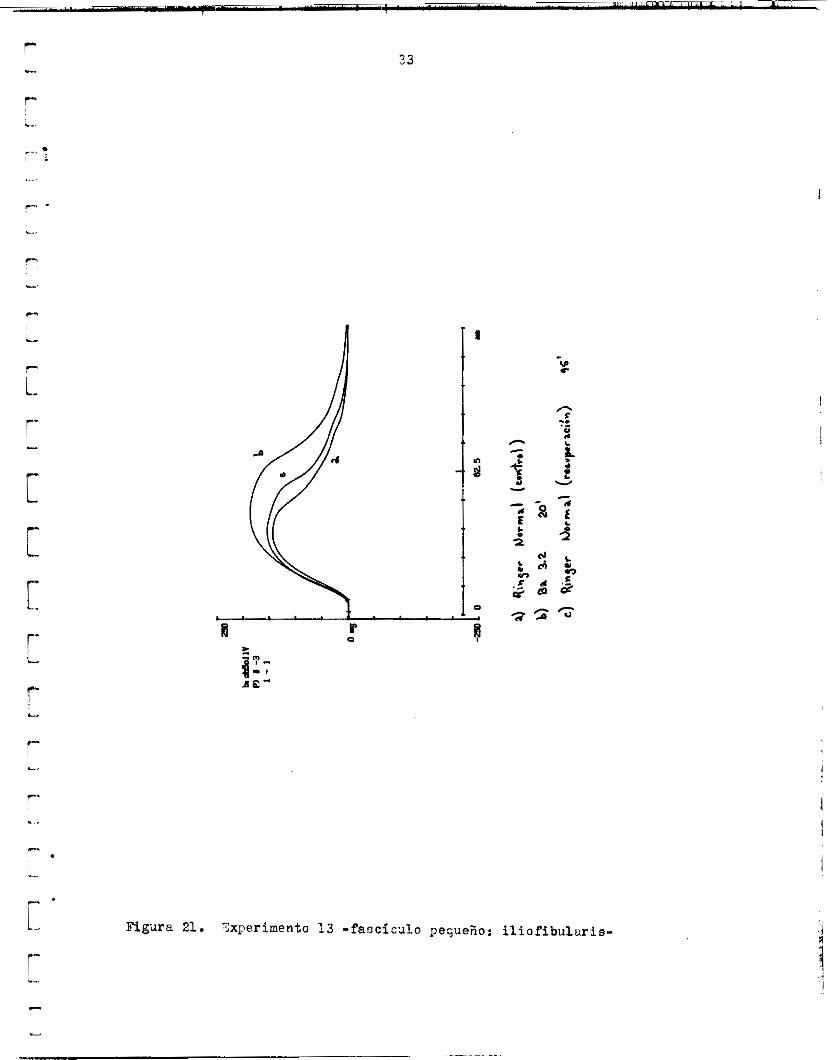
En o t r o experimentir s i m i l a r ( f i z . 21), l a ampiitud y el Srea aumen-
En 10s s i g y i e n t e s dos experimentos, se midíó oimultánenwente l a ten-
Zn e l segundo experimento iq.ue, > o r c i e r t o , se h i z o en la mism f i b r a
Los valores a lcania -
..I
" . ,
.*
32
..,. .
c-
i..
CI
._ .
.,".
._.
..-
._
c
.. . r-
.... c
.._+
P I
m. r
-.
1 : ’ Figdra 21. Txperimento 1 3 - f a o c i r u l o pequeño: i l i o f i b u l u r i s -
I
34
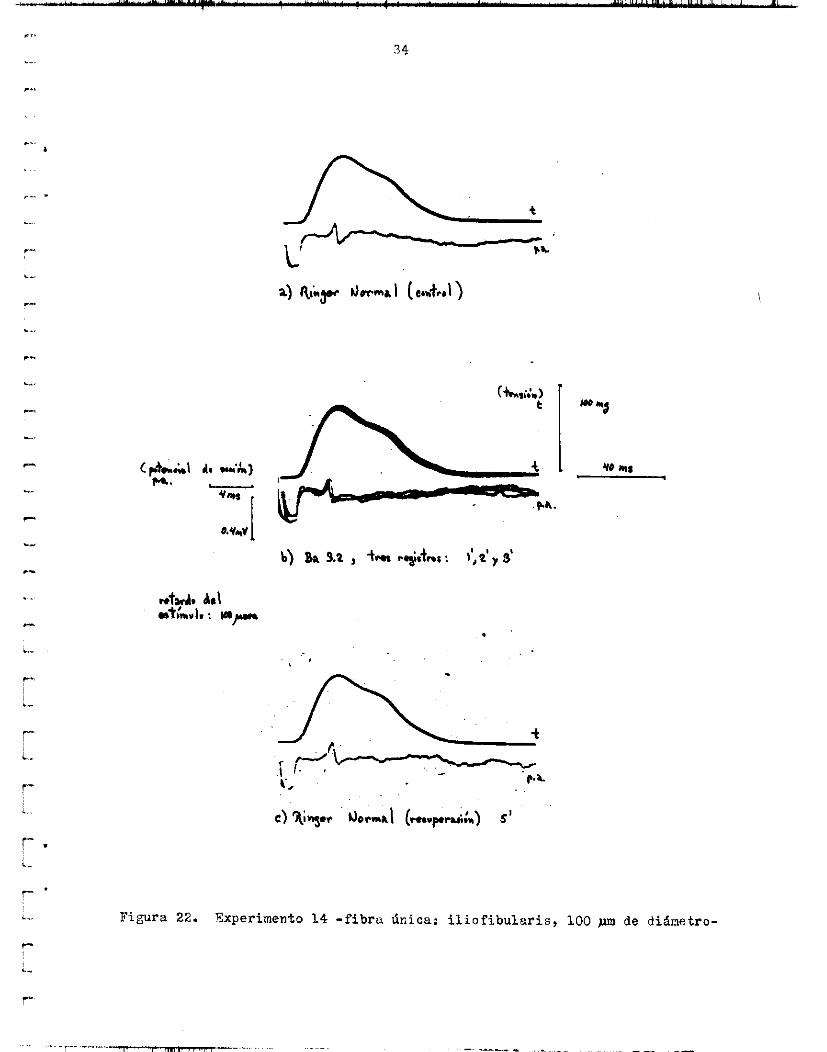
Fimra 22. Experimento 14 - f ibra h i c n t i l i o f i b u l a r i s , 100 de diánetro-
e .* -. .
I
35
r"- . il
f" l.,.
r" ' ..,
I '
F.
G..
4.2'
r- I
r- L
15.3' 20.3'
r' L.
r-
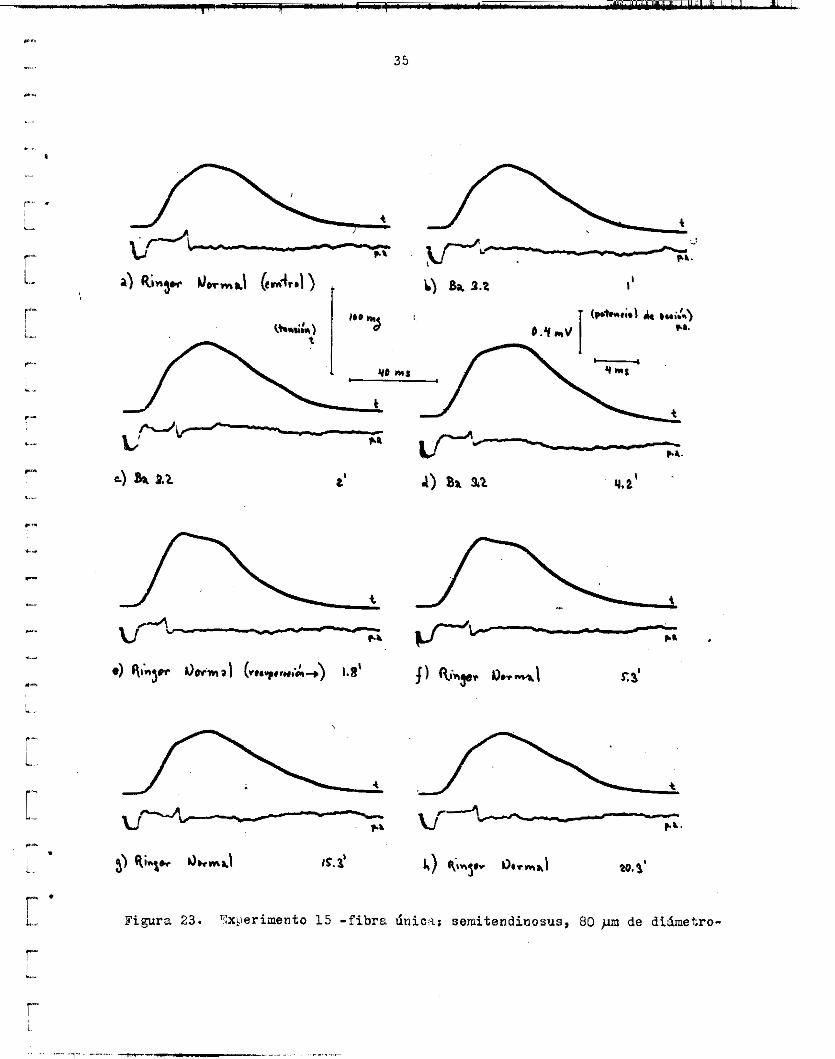
Figura 23. '-k$erirnento 1 5 - f ibra h i c - i : semitendinosus, 80 prn de didmetro-
, 4..
r-

, .
r..
., ,
,. . ,
.... \.
(. -
,.,..
..,_
.- . .
« .
,... ,.. .
..”.
. .
,..,
Y -
* .
.-
. .
t-
,.
36
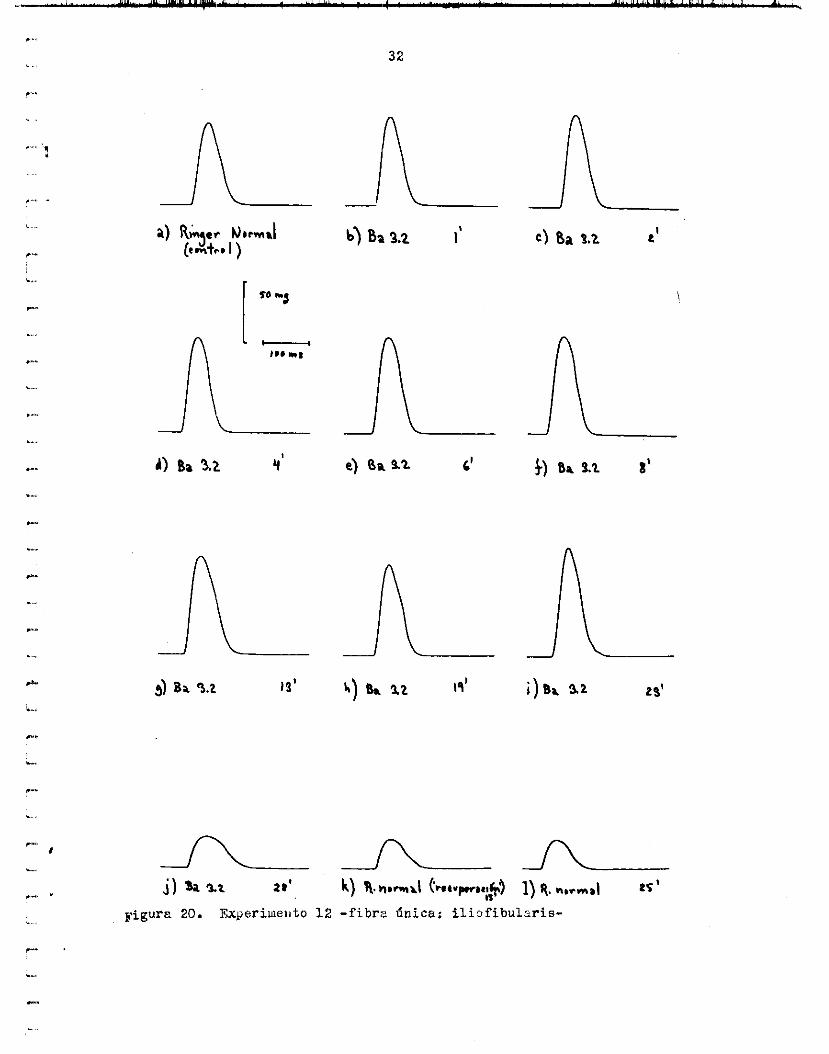
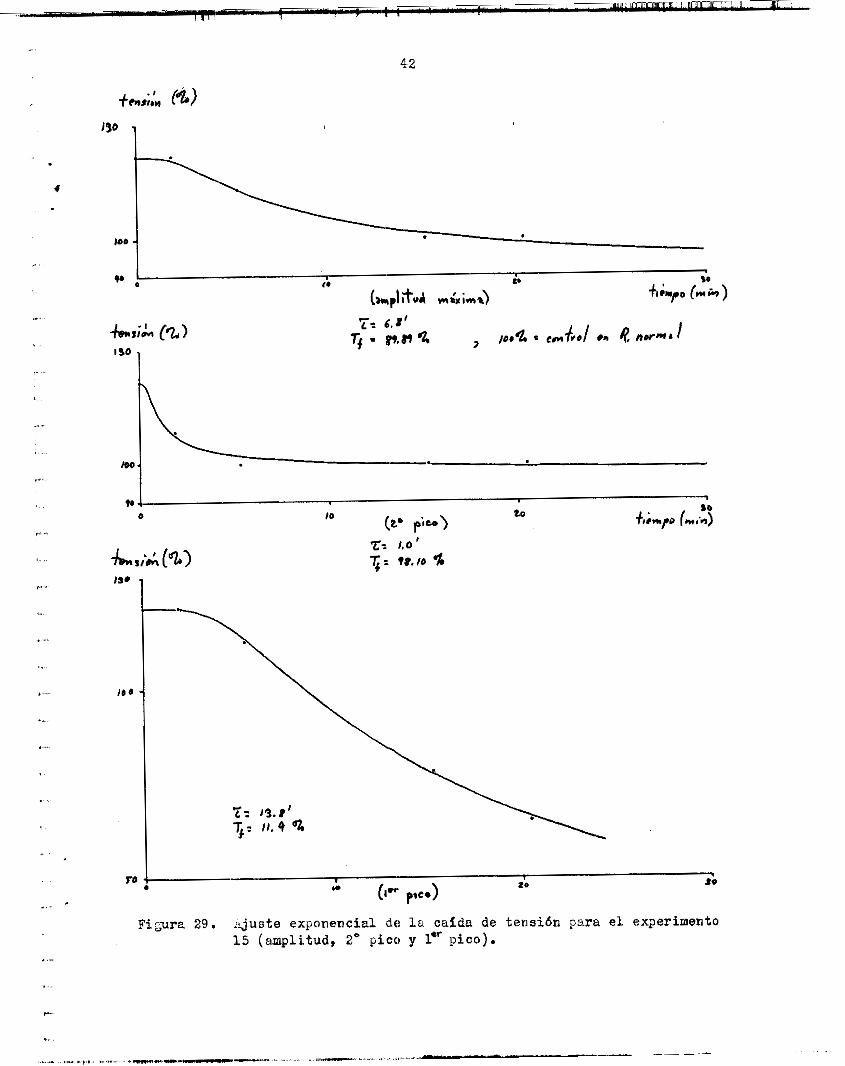
l a sacudida en Ba 3.2, IC tensitin crece d s rápidamente en ese punto, d e illanera %ue rebasa a l mdximo 0r igina. t y se convierte en e l nuevo uiáximo durante l o s primeros minutos de recureración en Ringer iiormai. c r e c e y l a sacudida vuelve a su form or ig in21 a los 25 mioutos de recu- peración- ICn e l a j u s t e exponencial d e l crecimiento y recu.,eracibn de l a
tens ibn (ver a b a j o ) s e considera. e s t a s i , tuuci6n p r t i c i i ’ i a r . La f i g u r a 2 4 riiuestru e l curso de e s t e exgerimento en e l Gould.
mejante. En e l s i g u i e n t e caso ( f i g . 2 5 ) , p o r ejemplo, s i bien aumentó la ampl.itud de l a sacudida en Ba. 3.2 (103.70$), e l Area ba jo l a curva disminuyó notablemente (Area t o t a l ! 88.00$, á r e a a l m&xz 92.98); de l con- t r o l ) . En la recuperecidn, s iguió creciendo un i ; ~ ~ ~ l a tens ión , y e l k - r e a tsmbien iiuinentó para des9ués volver a decrecer .
En o t r o s d o s experimentas (en l o s que 12s f i b r a s estuvieron expues- t a s sólo un minuto a Ba 3 . 2 ) , l a tensión y e l Brea s igz ieron aumentando durante var ios minutos en e l lav..do ciin Ringer normal. ñn l o s t r e s ex- perimentos inencicmhdos l a tensi6ri mixima ulcmzada fue 132.57*18.02$ d e l contro l . 169.75i24.44f4 respectivamente.
Tr.nto ,el aumento de l a amplitud de l a contracción en 3a 3.2 como l a recu:)ersci6in en Xinger normal fueron a justados a funciones exponenciales. Para l a f a s e de aumento de l a tensihr: , Ins puntos experimentales de CUB- t r o experimeritos se a jus taron independientemente a una f u n c i ó n de l t i p o ;
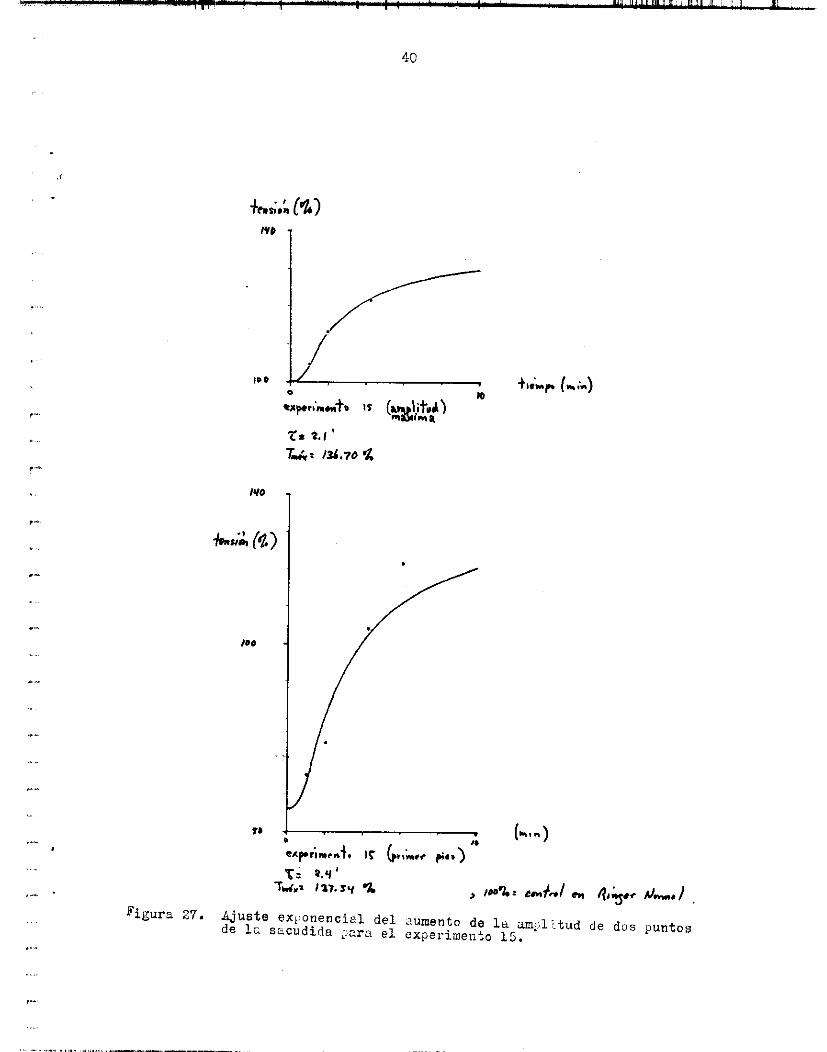
doriae TO e s l a tensibri en Rin,ner rj.jrrn:.l, in& es LEI tensidn ioSxim a l o cual l o s vúl.orez expxinientnles :se ticercan exponencialmente, Z e s una constante de t i e m p , t o e s el tiempo i n i c i a l y T e s la tensióri en el t ieEpo t. LL!S curvas se ituestralq en l e f i g u r a 29 y en la 27. De los experimentos i l u s t r a d o s , ya se seiia16 que en uno de e l l o s (exp. 1 5 ) dos puntos d i f e r e n t e s de l a controcci3n crecen y decrecen a. d i s t i n t o r i tmo, l o cual s e t i a n i f i e s t a en U% Z d i f e r e n t e . d i o pera esos cuatro experimento!: e s 1.9tü.9 m i n .
piintos experimentales de 3 experi.mentos a una función de l t i p o :
donde T+ e s l a t e n s i l n final c Ir. cual se ncercun l o s valores experinien- t a l e s , T,i es l a a m p l i t u d de l a sacudida innedie tmente antes de comen- zar e l lcvildo en Ring@r normal, Z e s una constánte de tiempo, ta e s e l tiempo i n i c i e 1 y T es l a tensiór: en el tiempo t. 29 s e muestran l o s puntos experimentales y l e s funciories exponenciales a jus tadas . 9.9f9.3 m i n .
E f e c t o s de o t r o s agentes.-
E1titizem.-
t o l e n t o de l a terisibri p x i v a , segaido de una r e l a j a c i ó n gradual hac ia l a bssal o r i g i n a l . En uiiibos casos , EB u t i l i z ó Dil i iazem 100 pM. % e l p r i - mer ensayo, IC tensión pasiva tiument6 7 mg en 6.1 m i n u t o s y decreció 3 ng
Luego de-
Como ya se mencionó, no todas l o s f i b r a s respondieron de manera se-
Lirs Breas t n t u l y a l maxim0 uunientaron a un 144.75t24.70jb y
T= T.+ (T&-T.) ezp cz /(t-td) ,
Li. constante de tiempo prome-
Pzra l a fane de recuperacibn, se c.justeron independientemente l o s
T= T++(Tmt.->)(l- ew(-’C/[t-t,))) ,
En las f igurns 28 y
La constante de tiempo p-omedio p:3.r:i eso6 experimentos e s
Yn aos ,ansa;.os rea l izados sobre l a inisma f i b r a se observd un uumen-

37
\
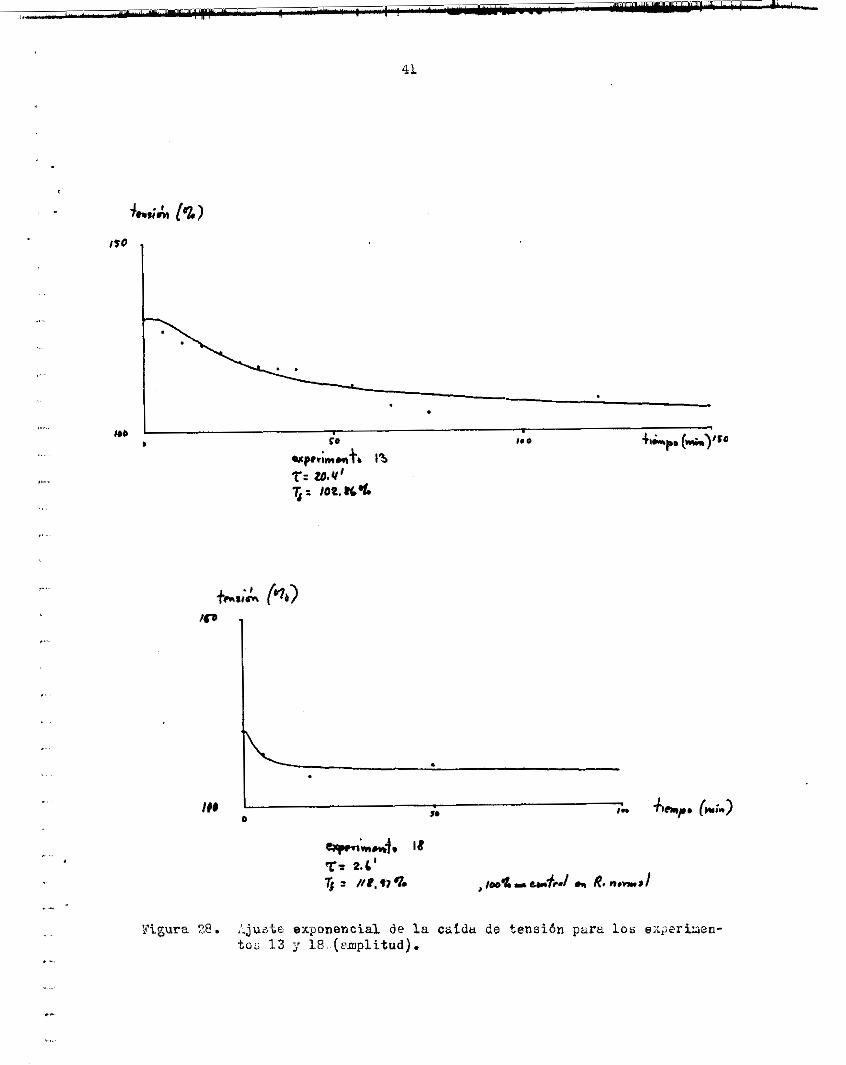
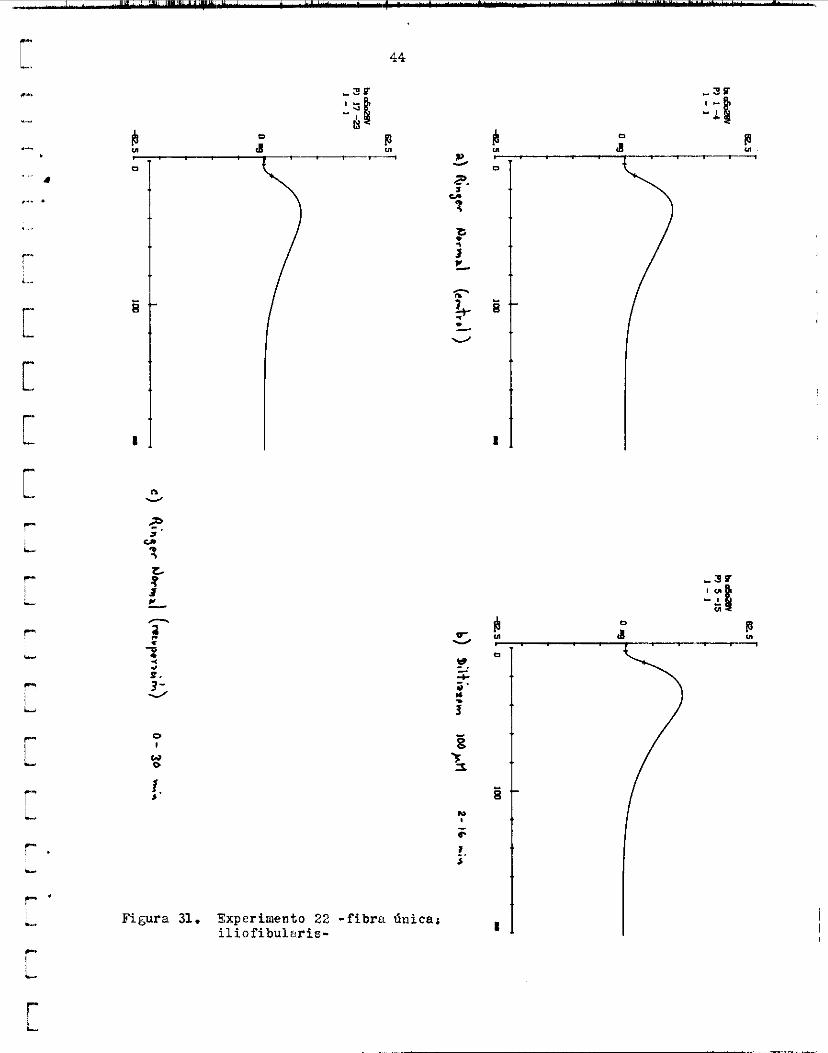
en 17.4 m i n u t o s E P a r t i r de ese míximo ( f i g . 30) . dl l a v a r nuevamente con R i n g e r normal, l a tensidn paeiva alcanzó l a l inea b a s a l o r i g i n a l en aprox. 7 rninutos. Litro e f e c t o observado fue que In sacudidn s e potenció er? Elt- iazem 100 LIE. La magnitud de l a contracción f u e , en p r o m e d i o 120.60% l e d e l or ig in i l l en Riayer n o r m d ( f i g , 31).
Después d e l lsvu~o eri R i rger norml, l a amplitud del twi tch descen- d i 6 u un 76.517; del c o n t r o l , pero s e mantuvo en ese va lor p o r bas tante tiempo, de modo que s e gudo r e a l i x u un segundo ensayo en l a misma f i b r a . En e s e segundo ensayo, s e volviC í' observar e l aumento de l a tens ión pa- oiva, el cual fue de 6 ng en 6 miriutos, y decrec id lentamente haJ ta a lcnn- zer l a l i n e a b a s a l o r i g i n a l en 25 minutos. La magnitud de l a potenciación en e s t e ensayo fue d e l 124.74%. La amplitud de la. contracción después d e l lavado en Ringer norual decrec ió kiesta 6 mg, lo c u d indicgt que l a f i b r a probablemente se dañd des2uén de e s t e ensayo; sin embargo, siguiócontra- yéndose con esa amplitud durante varios minutos más.
t r a c c i ó n l a r e l z j i i c i ó n l l e v a b a a l a tensidn R un valor menor que e l i n i - c i a l ( f i g . 31) . va subía nuevamente, pero no p r i m nloanzar e l va lor i n i c i a l d e l twitch, sino el valiir que temporalmente l e correspondla en l a curva de tens ión pa- s i v a .
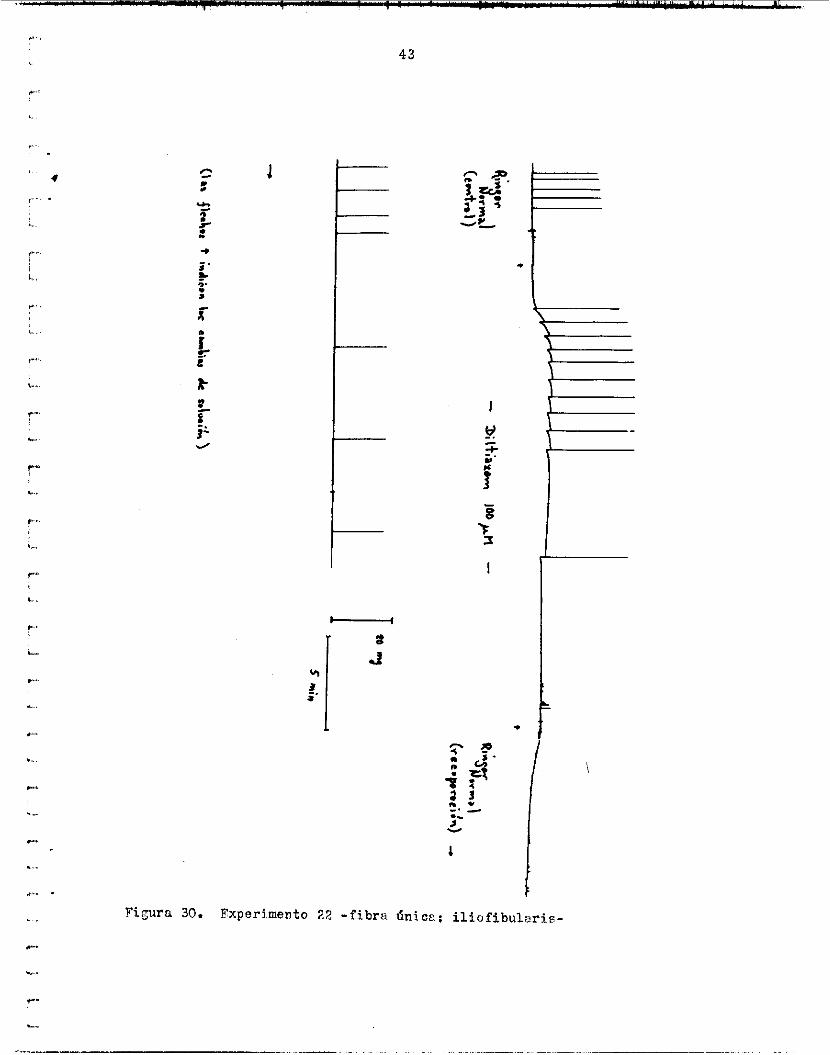
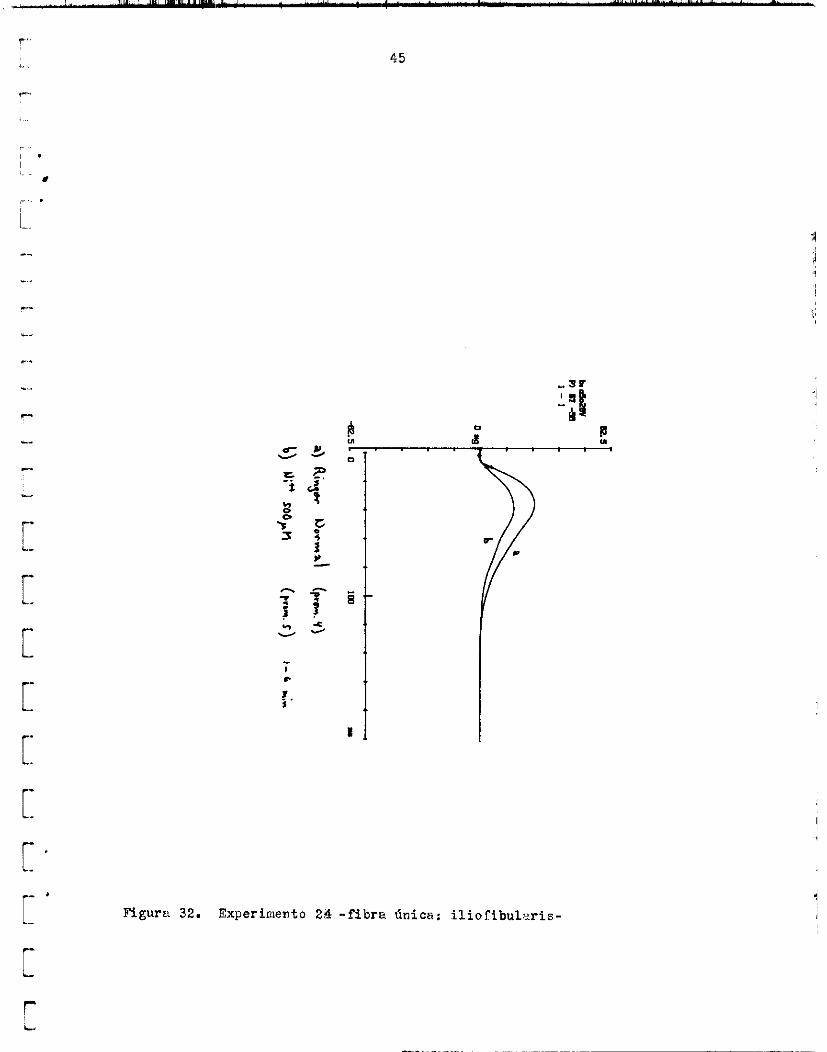
Niquel. - f e r e n t e s . Xn e l primero, s e observó una disminución de l a tens ión a l ?o- ner 500 PM de !Jiz' en e l Xinger. de su valor norma l . S i n embargo, l a f i b r a se dan6 y no se obtuvo recupe-
Es i n t e r e s a n t e que, en I n sol .uciSn con Mlt iazem 100 uM, en cada con-
Luego, como s e a p r e c i a en l a f i s r a 30 , l a tensi , in p a s i -
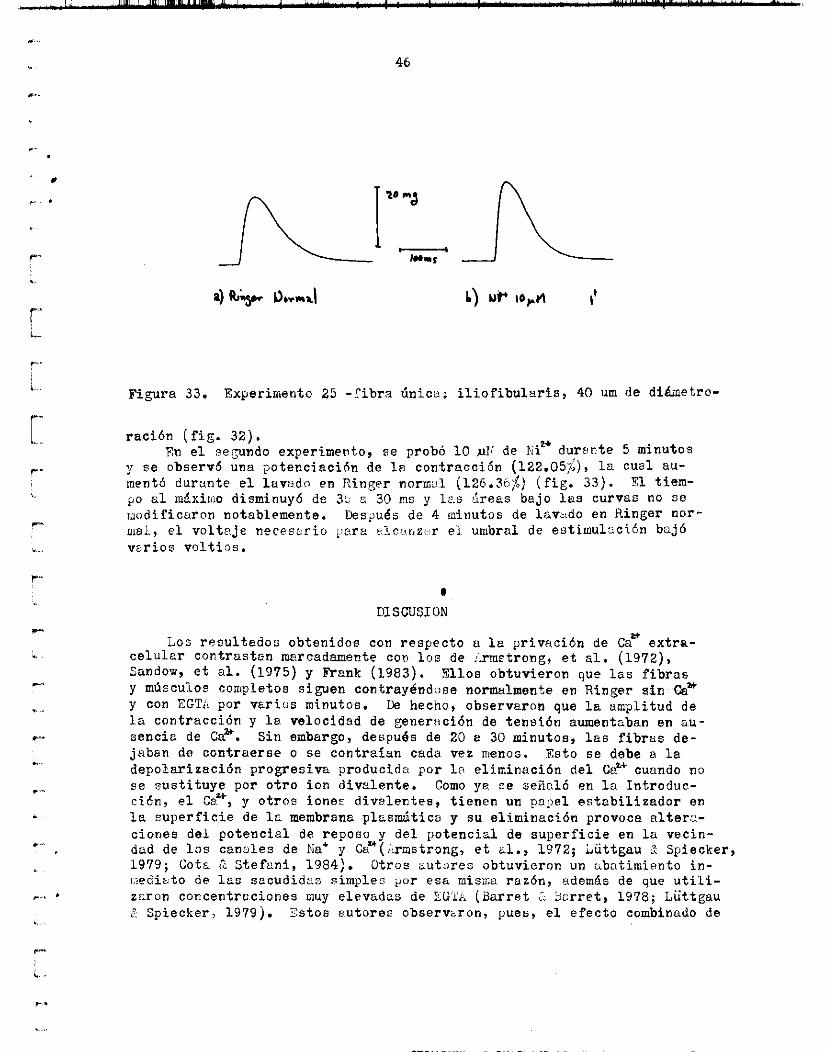
En d o s ex:ierimentos reali .zados con Ni:' se obtuvieron resul tados di-
La a m p l i t u d de la sacudida fue del 61.577;
'x I-
n
E
i
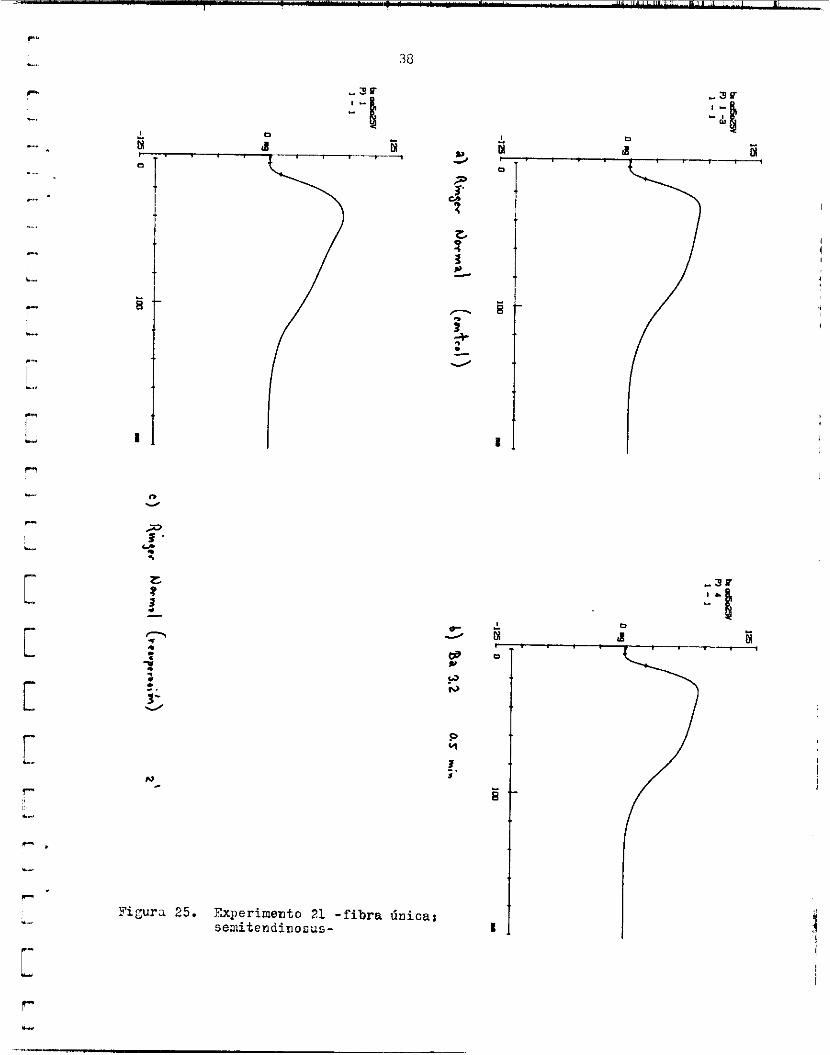
FiCura 25. Experimento 21 -fibra Única: oemitendinosus-
'x r o
E
I
'x I-
n
E
1
.. .
....
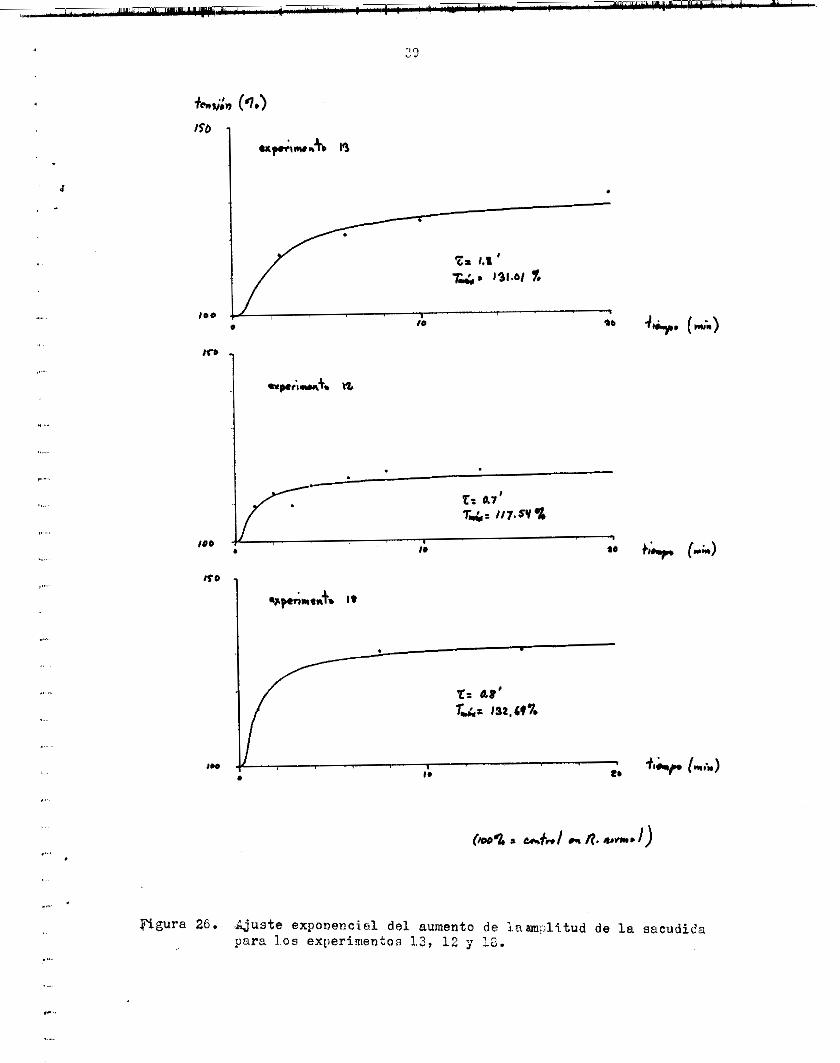
Figura 26. Ajuste exponenc id de1 aumento de lRam,ilitUd de l a sacudida para l o s experimenton 13, 12 7 16.
~...
.-
41
<. .
Yigura 28. :.jutit.& exponencial de la cüida de tensi6n pura l o s eqeririsn- tos 13 y 18 ( a m p l i t u d ) .
"O 1
Y 6- pea) t. S O
Figure 29. .guste exponencial d e 1; caida de tensidn ?ara e l ex;ierimento 15 (amplitud, Z 0 pico y ler pico).
... , 43
í' L ?. .. s
f E
Figurn 30.
.
- a I[ J
.Y
.-
Fxperirnento ZZ -fibrii única: iliofibularis-
44
o
E
I -
P o In . t-l P
In d 4
B
'-
0- u
F. r Y '
i
W O
1. *
Figura 31. Experimento 22 - f ibra dnicai i l i o f i b u l e r i s -
B UI r o
8
I
B In ?
D
8
1
P o
d UI
P d In
D
45
I ,
I..
c.
-_. F
c
c
c
Figure. 32. Experimento 2 4 -f ibre única: i l i o f i b u l e r i s - C I ' c r L-r
'I
C "
C . *
P"
r.
A
Figura 33. Exyerimanto 25 - f i b r a Únicci: i l i o f i b u l a r i s , 40 um de diáinetro-
r a c i ó n ( f i g . 32) .
y se observó una potenc iac ión de l a contracc ión (122.05$), l a cual au- En e l ?eJundo experimento, se probó 10 ,uii de P;iw d u r s r t e 5 m i n u t o 6
mentó durante e l lavado en Ringer norffial (126.30$) ( f i g . 3 3 ) . po al m&xiaio disminuyó de 3b a ?,O ms y 12,s a r e e s ba jo las curves no s e m o d i f i c a r o n notablemente. !.ks$u,és de 4 minutos de lavado en Ringer n o r - iiisl, e l volt?..je necesr.rio iari- s lcoi iz ; . r el. umbral de e s t i m u l z c i h ba jó vur ios v o l t i o s .
-;L tiem-
i)
111 SCüSI UN z+ Los r e s u l t a d o s obtenidos con r e s p e c t o a le privación de Ca e x t r a -
c e l u l a r contras ten marcadamente coa l o s de :.rm?trong, e t a l . (1972), Landow, e t a l . (1975) y Frank (1.983). y múscuioz completos siguen c o n t r a y é n d ~ s e normalmente en Ringer s i n Caw y con EGTíi p o r v ~ r i u s m i n u t o s . De hecho, observaron que 12 amplitud de l a contracc ión y l a velocidad de genernción de t e n s i ó n aumentaban en au- s e n c i a de C 8 . Sin embargo, después de 20 B 30 m i n u t o s , 12s f i b r u s de- j aban de c o n t r a e r s e o s e contra ian cada vez nienos. E s t o se debe a l a depolar ización progresiva producida. p o r 18 eliminación d e l Cew cuando no s e s u s t i t u y e por o t r o i o n d iva lente . Como ya. re aerial6 en l a I n t r o d u c - c i ó n , el Cay, y o t r o s ionec d i v o l e n t e a , t i e n e n un papel e s t a b i l i z a d o r en l a s u p e r f i c i e de IC membrana plasm&tico y su eliminación provoca a l t e r i - c iones d e l p o t e n c i a l de reposo y d e l p o t e n c i a l de s u p e r f i c i e en la vecin- dad de l o s cana les de Na' y Cs*(I;mustrong, e t d., 3972; Lüttgau i: Spiecker , 1 9 7 9 ; C o t 6 2 S t e f c n i , 1984). Otros autores obtuvieron un Lbtitimiento i n - i ,ieditto de l e s sacudidi:s simple:: ;lor e s a misaa razón, además de que u t i l i - z m o n concentrociones muy elevadas de kGYh ( B a r r e t C S c r r e t , 1978 ; Lüttgau r S p i e c k e r , 1979) . % t o s vmtoree observcron, pues, el e f e c t o combinado de
E l l o s obtuvieron que l a s f i b r a s

1
..
.-, . .
...
*,-
1.
c
I.
.-
...
...
...
.,.
I , .
...
,..
L ,
,.-
c
-
47
l a p r i v e c i ó n de caz+ y
a s { cióri siinpie.
membrane se a l t e r e o e n lo menos p o s i b l e , s e z:istituyú e i C 8 1 . 8 p o r Q3mhii ( H i l i e , e t al . , 19755 Cota C S t e f a n i , 13791 Lbiarandini, e t ai . , 1980) y s e emplearon concentraci.ones de EGTA que no danartin a las fi- b r a s , pero que s í redujesen l a c:oncentrrción de Ca* a v a l o r e s menores de loF8 Y (j;ri:istrong, e t a l . , 1 9 7 % ; Chiarandini & S t e f u n i , 1 9 7 6 ) . kde-
=As, f i b r a s pare. asegurar j u e l a s so luc iones difundieran completvnente y en i: o co ti empom . conuistentemente una reducción de l a amplitud de l a contracción simple a l cambiar l a f i b r a d 'Binger c o n una concentración de Caw menor que 10-8Mi. Lns determinaciones d e l p o t e n c i a l de reposo y e l r e g i s t r o d e l p o t e n c i a l de acc ión e x t r a c e l u l a r apoyan l a suposición tie ,,ue 1:s propiedades e l é c - t r i . c u s no se modificaron s igni f i ca t iv turente . No o b s t a n t e , es importante t e n e r presente que el p o t e n c i a l de acc ión de accidn medido i n t r a c e l u l u r - aiente y e l p o t e n c i a l de a c c i ó n e x t r a c e l u l a r s e re lac ionan cano una fun- c i ó n y su d.mivada, por l o que l o s cambios en e l p-iniero no s e traducen necesari8.mente de l a misma megnitud en e l segundo. Debido a eso serir convetiiente r e g i s t r a r in t race lu lnrmente el p o t e n c i a l de acc ión en l a s so luc iones prueba y v e r i f i c a r (.;lie efectivameote no cambie ( é s t o , !lor su- puer to , s e aplica también a los experimentos con Ba'*).
tud , pero se recuperd mucho más rdpidamente que l a tens ión . E s t o , l e j o s de i m p l i c a r que l a reduccidn del p o t e n c i a l de acciór. e s l a cause de l a caídü de Ir; tensicin, i n d i c a que, cuando hay e f e c t o sobre e l p o t e n c i a l de acc idn , B c t e es independiente d e l e f e c t o a n i v e l de acople e x c i t a c i d n - c o n t r ~ c c i ó n , e l c u a l , como :.e a p r e c i a en l o s s j u s t e s exponenciales, e s muy prolongado.
a l sumento de l a velocidad de conducción d e l p o t e n c i a l de acción, ya que, como se mencionó snter iormente , l a estimulación se hizo en un extremo de l a f i b r a y e l r e g i s t r o de t e n s i ó n y potencial d e acc ión en e l o t r o e x t r e - mo. Lc disminuci6n de l a amplitud de l a l a t e n c i a e s t á de acuerdo con l o s r e s u l t a d o s de Sandow, et a l . , !1975) , quienes obseivaron una rediiccidn d e l 45:; en Ringer s in Ca*y con EGTA 1 a.
L a disminucidn de l a tens idn máxima de l a s contraccLones está acom- paaadn de una reducción notzb le d e l área b a j o l a curva, indicando que l a l i b e r a c i ó n de Caw d e l r e t i c u l o sarcoplásmico t m b i é n disminuye. Adeds , contrariamente a l o observado por Sandoi7, e t al . ( 1 9 7 5 ) , e l incremento de l a t e n s i ó n ,por unidad de tiempo a l p r i n c i p i o de l a contracc ión e s menor en ausencia de Cas+ ( c o n TGTA 1 mu) que en Xinger normal. R s t o s e cumple, no sólo p a : i l o s primeros milisegundos, s ino para toda In f a s e ascendente de la c o n t r a c c i h , s i bipn en a lgunos casos en más marcado que en otros.
En CaO,,Mg3 ( s in XGTII), l a contrucci5n no s e nodifica en amplitud n i en duración. La l a t e n c i a y lu m;sgnitud de l a r e l a j a c i ó o de 1atenci : i son i g u a l e s que en Ringer normal. ? S t o implica que ,o bien la pequelia canti-
dei;olai-i.znciCn groducids p o r su e i iminac ixn , el e f e c t o de z l t t j s concentraciones de EGT!, sobre l a contrae-
63 e s t e t r a b a j o , p;;ra ~ s e g u r a r que 1-s p r o , i e d i d e s de c impo de 1P
siempre s e u t i l i z a r o n f i b r a s a i s l a d a s o f a s c í c u l o s de muy pocas
Como l o a t e s t i g u a e l a n d l i r r i ~ de 10 ensayos r e a l i z a d o s , se observó
Aflorcl b i e n , en un caso e l p o t e n c i a l de uccibn si disminuyó en ampli-
F o r o t r o l ado , l a disminucidn d e l tiempo de lettencie puede deberse

,..
r
I-
...
.- I..
...~ I"
..
...
I",
,..
<- .
I
I
r
,.. 9
.
.-
C"
-..I
48
* dc+d de Ca c o n t t m i n a n t e (3xlÓ' M:, C h i a r e n d i n i & S t e f a n i , 1376) e s s u f i - c i e n t e pare cumplir con sus func innes r e l a t i v a s a l a c o p l e e x c i t a c i ó n - c o n t r a c c i ó n , o que e l TCTA e s e l r e s p 3 n s ü b l e d i r e c t o d e l o s cambios ob- s e r v a d o s . ?:Sto i í l t imo e s una p o s i b i l i d a d que n o cia guede d e s c a r t a r con los r e s u l t e d o s ob ten idos . Xl EGTR p o d r í a i n t n r a c t i i a r con s i t i o s e s p e c i - f i c o s de l?. membrana p l a s m á t i c a . modif icsndo e l moviwiento de ctirgss, €lor e jemplo. Y s t & generalment,e i:cep;odo, sil , embargo, que cuando s e usa "Tii $11 e f e c t o se l imita a l a Gueliiciiin de i o n e s d e l t i p o Can, Bu*, e t c . , s i b i e n concent.racione3 s l e v a d % s pueden da2ictr l a s f i b r u s . Tomando en cuen- ta Q s t o : y cons ide rando l o s c m b i o s en e l movimiento de ciirgas a l modif i - c a r l a c o n c e ~ n t r a c i ó n de ca* e x t r a c e l u l - r ( S h l e v i n , 1379), se pueden i n t e r - p r e t c r l o s r e s u l t a d o s o b t e n i d o s ou;~oniendo que e l Ca" i n t e r a c t i i a COI, l o s cL>íii:..onentes d e l sistema. t i ibi i ler s e n s i b l e s e l v o l t a j e ; 13 I t i s enc ia de ¡;o,* produce una d i sminuc ión en e l v o l . t a j e de t r a n s i c i b n 0 y en l a coristtinta k, haciendo que una d e p o l c r i z a c i 5 n de magnitud normdl en el. sistema tii'bu- l a r s e t r a n s m i t 3 como una ser ia l a t e n u a d a a l r e t í c u l o r,rco&dsmici ( S h l e v i n , 1979; lirun, e t *l., en p r e n s a ) .
R inge r s i n C;", son rargs y no hay ex.Jli.caci6n r a z o n a b l e para e l l o s . cuan to 8.1 t r a t i e n t o de l a s fibi 'os c3n K' 5 mi&, l o s resu1:adgs concuerdan con l o o b t e n i d o p o r C h i a r a n d i n i ic S t e f a n i (1374). i i o t enc ian 1r.s s a c u d i d a s sinT1es.
Goii r e s p e c t o al e f e c t o d e l i3a*, se puede d e c i r poco, ya que l a varia- b i l i d a d de l a s r e s p u e s t a s f u e mayor que con CaO. S in embzrgo, e s c l a r o c;ue, en 8 ensayos reali.z!*dos, se r e p r o d u j o c o n s i s t e n t e m e n t e e l mismo ef'ec- t o i n i c i a l , es d e c i r , e l aumento d e l a t e n s i ó n máxima d e l e sacud ida . 51 c,ue en elgiinos cnsos l e r ecu ; s r ac ión en IIiFiger nnrnel baya s i d o nds rC:.,ida :.lie en o t r o s , p z r e c e e s t i r r e l a c i o n e d o co? e1 tiem,o de ex2osiciór . ti lz s o l u c i ó n prueba. LL.S f i b r o r E&.; t iempo t c rds ro r : mbs e n re- c u p e r h r s e o se daiiúron, x i c,ue e s t u v i e r o t i ~ O C O t iempo se r e - cuperaron t;:.oibién en coco tiempo,. Pero é s t o Último no s u c e d i ó sieffipre; en algunos casus , li. t e n s i ó n ::iguif iiumentmdo i n c l u s o desdiiés de bailar a 1 u f i b r a en R inge r norincil.
Es p o c o p r o b a b l e que e l e f e c t o e s t é r e l a c i o n r d o con una e n t r a d s de Ba a l miopl.asmn. S i b i e n , e? u17 hecho conocido ;ue e s t e i6n a t r a v i e s a e l c a m 1 de Ca"+, la c s r r i e n t e de 3aH(x1 ig?zcl GIie Ir de ,&+j no se n c t i - v:i d u r a n t e ::acudidas s imples . S!i p e n e t r a c i ó n deberi,: r e r o b i e n p o r e l c:.:ni.l r s y i d o de Caz+, o p o r : : l g n s . o t r a via aún no conocida (lo c u a l e s improbzble) . v e l d e l r e t i c u l o s z r c o p l i s i d c o , i nduc iendo le. l i b e r a c i h n de Ca': t a l como se hu v i s t o a l p r o d u c i r c o n t r a c c i o n e s mediante p o t e n c i a l e s de B a e n f i - b r a s n i s l e d a c (i 'otreau k Raymond, 1980) . !in es te caso, e l r e t i c u l o sar- cop l¿ sn ico tie c a r g c r i c después de a l g ú n tieniiio con Sa?, e l cixsl, a l ser l i b e r a d o a l c i t o p l a s m a p o r lo? c:-nalPn de Cui, p r o p i o s de e s e sistema, se- ri.r i n e f i c r i z p l r n Eererr!r t e n s i ó n (%do, e t a l . , 1970). Aunque en al;:~- nos expe r imen tos s e observó una d i ~ m i n u c i ó n b r u s c a de l a t e n s i ó n despues de v a r i o s minutos (20-30 m i n ) , é:?to mbs b i e n se deb ió a un dano de l a f : i - b ru que a su s a t u r a c i ó n con Baa+. E1 e f e c t o debe tener un s i t i o de accidn e x t r a c e l u l a r . En es te caso , como con los expe r imen tos de p r i r e c i ó n de Caw, es i m p o r t n n t e medir i n t r a c e l u l v r i n e n t e e l p o t e n c i a l de ~ c c i ó n en l a s o i t ~ c i ó n
L o s ca3os en que 1u siicudidi; s e p o t e n c i ó después de li: e x p o s i c i ó n u En
W p o l a r l z a c i o n e s leves
ti
Tn caso de ;>enet.ra:r, r e e s p r a r i z que su e f e c t o fuese o n i -
*

49
i!rueba, y" t p e , aunr;ue por l a s concentraciones escogidas e l potenc ia l de rep"so y e l p o t e n c i s l de s u p e r f i c i e no debieron a l t e r a r s e s i g n i f i c a - t ivamente, un p q u e n o aumento en Is r im; i l i tud d e l potenc ia l d e acc ión P O -
dria producir un aumento correspondiente oe l a tens ión , s i n que en e l potenc ia l de acc ión e x t r a c e l u l a r se detec tase e l cambio. -io obstante , e s t o no e x g i i c a r i u todos l o s hec'hos, pues e l e f e c t o d e l Baw, a l igua l cue e l de la privoción de CaB, 3 8 p r o l o n p mucho más tiempo que e l ne- cesar . io par:? r e p o l a r i z a r l a meubrbrnna con Ringer normal. so, cono se vé en l o s e . justes exponenciales de recuperación, l a f i b r a tardó haeta 2 horas en regresar a un valor de 108$ d e l normal .
con io^ resul tados obtenidos ::or h n z c i e z - S e r r c t o s , e t si. (19szj con el r.i3mo bloquiimte sobre l a a m i , l i t u d de l a s contracciones simples. embcrgo, ES de $&i"t,iculdr i o t e r e t e l aumento griidual de 1- tensi6n pi- s i n . i z r e e r i a como s i e l dilt,iazem p o v o c a r a una l i b e r a c i ó n gradur,l y sostenida ~8' del r e t i c u l o sarcoplésmico, semeja.nte a una contractura , sólo que muy prolongada. De hecko, como ya se seEul6 en l o s resul tudos, 18. tenf i i fn :pasiva comienza P. descender por s i s ó l u después de algunos minutos y su descenso e s acelerado ['or e l cambio u Rineer normal. Otro hecho i n t e r e s a n t e e s r.ue en czda contracción l a tensión de:jpués de l a f a s e de r e l a j c c i ó n ba jaba R un n i v e l menor (,<e e l i n i c i a l , para despues recuperor e1 valor cor-erpondiente de l a curve de tens ión pasiva. i p a - rentemente, -.urante li, f2se ascendente de l a sacudida, la l i b e r a c i h de C 8 e s t 6 f & c i l i t a d n p o r e l di l t iazeni (produciendo l a potenciec i6n obser- va.dej, y en la r e l o j a c i ó n , i a recz.ptación de Cz" por el r e t í c u l o ssrco- p l á m i c : , t m b i é n e s t i f a c i l i t a d u : de modo que hay un excedente de 6 r o - cóptado, e l c u d inmeidiotauiente comienza :-. s e r extruído nuevamente.
Con respec to a l e f e c t o de l Mi*, ?e puede d e c i r niiiy poco. 31 empleo de Ni 500 AIM probablemente fue d e n e z i a d o p::>r:i 1 7 f i b r a y se 2856, mien- t.rns qiie 10 MU p r o d u j o una poterNaieciót rentemente n o r e v e r s i b l e . I'uesto (,ue, en egte dit imo ccso, el. $.rea b: jo l o curva de tensión n o se moii i f icó, e l e fec to fue mbr bier! sobre 12 c i n é t i c u de crecimiento de 1:. tens ión que sobre l a cantida.d d e Cn" l iberado p o r e l r e t í c u l o sarcoplás- mico diirante l a contrnccicn.
Tn ai;pnos ca-
E l e fec to e e l d i l t iazem observado en e s t e t r a b ; i j o e s t é de ..ciierdo
S i n
&:
CClilCLUSIOW
E l u c o , l e exc i tac ión-contrccc ión e s un p o c e s o extremadamente com- : . l e j o , resul tado de l a in terhcc ión de vzr ios elementus, cuyas funciones son aún en gran p a r t e desconocidas. En e s t e t r a b a j o , 38 t r a t 6 de ver s i cambios que, por una r e r i e de evidencias experimentales, se sabe o supo- ne modificsn a l p n o de l o s elementos r e l ~ c i o n o d o s con e l mecmismo de acople , mostrabrn c l $ n e f e c t o sobre -1 eslc*bón f i n u l de e s t e Froceso, es d e c i r , sobre l a contrucción. Pert i determinar e l s i t i o exacto donde ocurren los modifichciones y su mecanisno de acc ión e s necesar io d isecar e l proceso, y anul izur cadti una de l a s per tes p o r separado. E l r e s u l t a - do f i s i o i 6 g i c 0 , s i n embwgo, purde r e s u l t m mucho mds complicado de in- t e r p r e t r r , debido ;; l a mult ipl ic idad de 12s i n t e r s c c i o n e s de l o s elemen-

50
tos p u r t i c i p s n t e s y P. la v e r i c b i l , i d n d , t a n t o de l a s condicione- exper i - mentc.les, como del pro,:io sistema b i o l ó g i c o en estudio.
I)e los r e s u l t a d o s obtenidos, s e puede afirmkr Gue e l Caz' e s t 6 i m - p l i cado en e l meccnismo de z c o p l e e x c i t c c i ó n - c o n t r a c c i ó n y que su ausen- c i a modifica, e s t e proceno, produciendo contracc iones de menor megnitud iue 12. nornúl. Alterne.tivamente, e l e f e c t i observado juede deberse u la inter¿tcción d e l XGT:. cor: zlg& s i t i o re lac ionado con e l acople. E1 Bo? tnnibiéri modifica e s t e proceso, produciendci u n a potenc iec ión de l a sacudida shple. Finalmente, e l e f e c t o d e l d i l t i a r e r r contrndico l a suposición de que e l influjo de Cia" pueda s e r e l in termediar io en e l s c o p l e , pero presenta más evidencia de qve un s i t i o ext.erno de 1; membrana, o a c c e s i b l e desde e l e x t e r i o r , sea el i m p l i c s d o en l a transmisión de i r s e ñ a l de los tú- bulos t ransversos a l r e t í c u l o srrcoplásmico, probhbleuiente e l mismo (que s 9 modific:ue »or l a rlusencie de Cae o su s u s t i t u c i ó n ijor be".
Su s i t i o de acc ión debe ser también e x t r a c e l u l a r .
LIERAWiA CITADA
Almers, 'T. rC Palade, P.T. (1981) . Slow calcirlrn and ijotasium c u r r e n t s ?cross f r o g muscle membrane ; measurements with a vaseline-gap technique. J. Phys io l . ( L o n u 312, 159-176.
Almers, 'Y., Fink, F. i- Fnlade, P.T. (1981). Calcium deple t ion i n f r o g m a c l e tubules : the d e c l i n e o f calcium current under rncintained d e p o l c r i z a t i o n . J. ?hysiol . (Lond.] 3 1 2 , 177-207.
Armstrong, C . L , B e z a n i l l a F A . & Horowicz, t'. (1972) . ' b i t c h e s in t h e presenci? o f e thylene g l y c o l bis@ - m i n o e t h y l e ther) -N,N'- tetraacet ic oc id . Uiochim. Biophys. Act;% 267, 605-608.
B a r r e t t , J.h. & B a r r e t t , E.F. ( 1 9 7 8 ) . E x c i t a t i o n - c o n t r a c t i o n coupling i n s k e l e t a l muscles o f celsi.um buffers. S c i e n c e , 1l.Y. 200, 1270-1272.
blockodi? by hi&h e x t r a c e l l u l a r concentra t ions
Deaty, G.1.. k Y t e f i n i , E. ( 1 9 7 6 ) . Calcium d e u m d e n t e l e c t r i c a l z c t i v i t v i n t v i t c h mii?clp f ibre3 o f ' t h e f rog . l/*l-lX.
Proc. R. Soc. Lond. B. 1 9 4 ,
Brun, G . , S t e f o n i , 7:. & Ilics, r;. ,;cn prenaa). z"ffect,s o f e x t r a c e l l u l a r cslci.um on t h e calcium movements of e x c i t i t i o n - c o n t r a c t i o n coupling i n s k e l e t a l muscle f i b e r s .
Chizrcndinl., D.J. C: U t e f r n i , X . (1.971). I b i t c h potenti-et inn by ;.otassium c o n t r a c t u r e s in s i n g l e muscle f i b r e s o f t h e frog. J. Physiol . (Lond.) 24Q, 1-14.
Chierandini , D.J. e¿ S t e f a n i , E. (1976) . Ga and e x c i t c t i o n - c o n t r a c t i o n coupling, i n f r o g s k e l e t a l muscle f i b e r s , en E l e c t r o b i o l o a v o f Ejerve.

51
Synapse, and Kuscle, ed i ted by J.P. Reuben, C.P. Purpure, E.V.L. Bennett, and F.R. Kandel. Rzven Press, Nes York.
Chirrandini, U.J. ct Stefnni , E. (1983). Calcium ect ion po tent ia l s ii! r a t f t s t - and slos:j-.twitch rr,uncle f ibren. J. Phyeio l . (Lond.) 335, 29-41.
Chiarandini, D.J., Sdnchez, J.A. 8; S t e f m i , E. (1980). X f f e c t o f calcium wit.hdrxra1 on mechanical threshold i n ske l e ta l muscle f i b r es o f the frog. J. Fhysiol. (Lond.) 303, 153-163.
Cota, G. & Stefuni, E. (19.79). E fec to de cationes d ivs lentes eobre e l potencial de suuer f i c i e externo en f i b r z r uiusculrree esiuel6ticc.s de rana.,. Prric. Xi11 Congreso Nucional de Ciencias F is io lbg i ccs , 1978, hjii8xico; Soc. Méx. Ciencias F is io lbg i cas .
Cote, G. e: Ste fsn i , E. (1981). E f f e c t s o f external calcium reduction on the k ine t i c s o f potessium contractures i n f r o g twi tch muscle f ibres . J. Fhysio l . (Lond.) 317, 303.-316.
Cote, G. : Ste fani , E. (1984). U.aturation o f calcium channel6 and sur- - face charge e f f e c t s i n ske l e ta l muscle f i b r es of the frog. (Lond.) 351, 135-151.
J. Fhysiol.
Cota, G. 4 S t e f a n i , E. (198Sj. Fast ú n d slow Ca channels in t d t c h nuscle f i b r es o f the frog. biophys. J. 47, b 5 a i
Cota, G. S te fmi , E. (en prensa). A f a s t ac t i va ted inward calcium cur- rent i n tw i t ch muscle f i b r e s o f the f r o g (Rana montezume). d. a L'hvs '
C o b , G., h i co la S i r i , L. :- Ctefuni, E. (1983). Calcium-channel gating i n f r o g ske l e ta l muscle membrane: e f f e c t o f temperature. J. Physiol. (Lond.) 338, 395-412.
%te., G., N ico la Sir;., L. Y: S t e f m i , E. (1964). Calcium channel inac t i - vat ion i n frog (Rum piy iens ,nd lbn i : cioctezuma) ske l e ta l muscle f i - bres. J. i'hysiol. (Lond.) 354, 99-108.
Endo, ;A, 'Iknaka, U. Ogawa, Y. (1970). Calcium induced re l ease o f cal- cium from the sarcoplasmic reticulum of skinned ske l e ta l muscle fi- bres. listure (Lond.) 228, 34:-36.
Frank, G.I.3. (1984). Blockade c f C:a*+ channels inh ib i t s I? contractures but, not, t::.itches i n ske l e ta l muscle. Can. J. Physiol. Phcrmacol. 52, 374-378.
GomboL-,,ldeco, R.., Huerta, K. 6. Stefani , E. E f f e c t o f Cae channel blockers end Ni* on K+ contractures i n frog t i t i tch f ib res . 3n prep~rnc i6n.

52
Gonzálee-Serratos, Ii., V a l l e - m i l e r a , R., Lathrop, D.A. & Garcia, María del Carmen. (1982). Slow inward calcium currents have no obvious r o l e i n muscle e x c i t a t i o n - c o n t r a c t i m cougling. Nature (Lond.) 298, 29 2-294.
H i l l e , B., .ioodhiill, A.K. P Shapiro, B.I. (1975). iqegative s u r f a c e charge near soiliiiin channels o f nerve; d i v a l e n t i o n s , monovalent i o n s , and pIi. i'hil. Trans. R. Soc. Lond. 13, 270, 301-318.
Huerta, L., Gamboa-iddeco, R. 5: Sitefani , E. (1985) . S f f e c t o f ca" channel blocker:; and I,ii* on K' coctr::ctiiro!: i n f rog twi tch f i b e r s . Ileurosci.. Abstr. V. 11, P r r t I . 409.
&
Ketz, B. (1?79) .
Lüttgau, H. Ch. 2; S p i e c k e r , :+. (1979) .
Islerv, i:uskel und S y n q s e . Gex-8 l'hiem9 VPrlag, jtiittprt,.
ilie e f f e c t s :if c:jlciiim deprivdtion mechenicrl a n d e lec t rophys io l .og ica1 parariieters i n s k e l e t a l nitecle E i - b r e s o f the frog. J. Fhysiol. . (bond.) 23u, 411-429.
l ' i c o l a S i r i , L., Cbnchez, J.R. & S t e f a n i , E. (1980). E f f e c t o f g l y c e r o l treutment on t h e calcium current o f k ~ g c k e l e t s l muscle. .(Land.) 305. 87-96.
J. Phyniol.
?eachey, L . , AdriLin, i.. 2 Geiger , S. S k e l e t a l b!u!uscle, en Handbook of i'hysiology. aethesda, El:nryl.,i.nd, 1983.
Potresu , E., & Raymond, G . (1980). Slow inward barium current and contrac- t i o n on f r o g s i n g l e muscle fibres. J. Physiol . (Lond.) 303, 91-103.
Fotroau, D., d Raymond, G. (1980b). Calcium-dependent e l e c t r i c a l a c t i v i t y and COntruCtiOn o f voltage-clamped frog s ing le muscle f i b r e s . J. P h j s i o l . (Lond.) 307, 9-12.
-
SBnchez, J.L. & S t e f a n i , E. (1978). f i b r e s o f t h s frog. J. Pnyuiol. (Lor1d.j 283, 197-20:).
(1983) .
Inward calcium current i n t w i t c h muscle
Sánchez, J.A. L : ;tefani, E. K i n e t i c p r o p e r t i e s o f calcium channels o f t w i t c h iauscle f i b r e s o f t h e frog. J. Fhyciol.. (Land.) 337, 1-17.
CJando~.~, I?., :Pagal, id. 8; Sphicas, 7. (1975) . Exci tRt ion-contract ion couplingr e f f e c t e o f "zero"-Ca medim. Biochim. Uio$hys. I.c% 4C4, 157-163.
Schneider, 1i.F. Y : Chendlm, N.K. :1197:). Voltace dependent chmge movenent i n skeIet?,l muscle; E p o s i b l e s t e p i n e x c i t a t i o n - c o n t r a c t i o n coupling. Future (Lona.) 242, 244-246.
S d e v i n , H.2. (1979). Sf fec t : . o f e x t e r n n l celcium concentra t ion a n d pH on ch6rge movemente i n frog r k e l e t a l muscle. J. Phys io l . (Lond.) 288, 12?-158.

Somlyo, !..P. (1955) . R e nesf;i.n,:er across t h e gap. hoture (Land.) 316, 238-209.
Stefmi, Y. ?: C;iiürandit:i, D.J. (1913). Skeletal muscle; Dependence of iiotass<um contrectures on extracellular calcium. Pf lÜgers .*rch. 343, 143-150.

c
, . ”.....,.