Embed Size (px)
Citation preview

Proc. Nati. Acad. Sci. USAVol. 82, pp. 3964-3968, June 1985Biochemistry
Purification, induction, and distribution of placental glutathionetransferase: A new marker enzyme for preneoplastic cells inthe rat chemical hepatocarcinogenesis
(hyperplastic nodule/detoxicating enzyme/in vitro translation)
KIMIHIKO SATOH, AKIO KITAHARA, YASUSHI SOMA, YUKIO INABA, ICHIRO HATAYAMA, AND KIYoMI SATO*
Second Department of Biochemistry, Hirosaki University School of Medicine, Hirosaki 036, Japan
Communicated by Sidney Weinhouse, February 7, 1985
ABSTRACT A polypeptide of Mr 26,000 and pI 6.7 thatwas markedly increased in rat livers bearing hyperplasticnodules (HNs) induced by chemical carcinogens was identifiedimmunochemically as the subunit of neutral glutathione (GSH)transferase (GSHTase; RX:glutathione R-transferase, EC2.5.1.18; also called GSH S-transferase) purified from placenta(GSHTase-P) and was demonstrated immunohistochemicallyto be localized in preneoplastic foci and HNs. In the presentstudy, GSHTase-P has been purified from the HN-bearingliver, and the distribution and inducibility have been examinedquantitatively using anti-GSHTase-P antibody. Elevation ofGSHTase-P in the HN-bearing livers was also confirmed by invitro translation of mRNAs isolated from the HN-bearinglivers. The purified GSHTase-P was homogeneous in size buthad two charge isomers on two-dimensional gel electrophore-sis. In normal tissues, including liver, placenta, and fetal liver,the protein content of GSHTase-P was generally low but wassignificantly high in kidney and pancreas. In contrast, theamount of GSHTase-P in HN-bearing livers (primaryhepatomas) and transplantable Morris hepatoma 5123D wereseveral 10-fold higher than that in normal liver but wereundetectably low in transplantable Yoshida ascites hepatomaAH 130. Different from ordinary drug-metabolizing enzymes,GSHTase-P was uninducible by administration of drugs andcarcinogens prior to appearance of the preneoplastic foci andHNs. In addition, species specificity of GSHTase-P was low asit was crossreactive among rat, hamster, and human.
Much attention has been focused on the morphological,histochemical, and biochemical properties of the preneoplas-tic cells such as enzyme-altered foci and hyperplastic nodules(HNs) induced at early stages of chemical hepatocarcinogen-esis (1-5). Among (pre)neoplastic markers so far found,epoxide hydrase (6), an isozyme of UDPglucuronosyltrans-ferase, and a molecular form ofglutathione (GSH) transferase(GSHTase; RX:glutathione R-transferase, EC 2.5.1.18; alsocalled GSH S-transferase) were remarkable since their activi-ties increased with the appearance of foci and HNs butdecreased with the dedifferentiation ofhepatomas (7). Never-theless, most of the enzymatic activities were significantlyhigh in normal hepatocytes and were easily inducible bydrugs and carcinogens. Thus, specific preneoplastic markersare required for the analysis of the multistep development ofcancer cells.
Several investigators (8-10, 34) have detected proteins thatare increased in the preneoplastic cells induced in livers ofrats administered diethylnitrosamine and N-2-fluorenylacet-amide according to Solt and Farber (4). Eriksson et al. (10)observed increased polypeptides of Mrs 27,000 and 21,000 inisolated HNs induced by six models for the chemical hepa-
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
3964
tocarcinogenesis. In our studies of isozymic alterationsduring chemical hepatocarcinogenesis, especially of drug-metabolizing enzymes (7, 11-15), we observed by two-dimensional gel electrophoretic analysis of the cytosolicproteins of the HN-bearing rat livers that a polypeptide ofMr26,000 and pI 6.7 [designated polypeptide (26/6.7)] increasedmost markedly together with three other proteins. Thispolypeptide was identified immunochemically as the subunitof a neutral GSHTase that was purified from rat placenta.This placental form (GSHTase-P) was demonstrated to belocalized in HNs and in foci positive for y-glutamyl-transferase (GluTase; y-glutamyl transpeptidase, EC 2.3.2.2)(14).
In the present study, GSHTase-P has been purified fromHN-bearing rat livers, some chemical properties have beencharacterized, and the content of this protein in normal,preneoplastic, and neoplastic tissues and its inducibility in ratlivers by drugs and carcinogens have been examined.
MATERIALS AND METHODSAnimals. Male Sprague-Dawley rats, weighing 150-160 g,
were obtained from Charles River-Oriental Yeast (Tokyo).Chemicals. Chemicals were obtained as follows;
diethylnitrosamine, 3'-methyl-4-dimethylaminoazobenzene(3'-Me-DAB), and 3-methylcholanthrene, Tokyo Chemicals;N-nitroso-bis(2-hydroxypropyl)amine, Nakarai Chemicals(Tokyo); butylated hydroxyanisol, Wako Chemicals (To-kyo); phenobarbital, Sankyo, Tokyo; PCB, KanegafuchiChemical (Osaka).
Induction of Preneoplastic Enzyme-Altered Foci, HNs, andHepatomas. Preneoplastic foci, HNs, and hepatomas wereinduced in rat livers by administration of diethylnitrosamine(200 mg/kg of body weight) and N-2-fluorenylacetamide(0.02% in diet) plus partial hepatectomy according to theprotocol of Solt and Farber (4). Foci and HNs were induced4 to 6 wk after diethylnitrosamine injection, and large anddense HNs were detectable with the naked eye after 6 to 8wk. Hepatomas were induced by diethylnitrosamine fol-lowed by N-2-fluorenylacetamide without hepatectomy or by3'-Me-DAB (0.06% in diet) alone as described (11, 15). Inlivers of the Syrian golden hamster, HNs were induced bys.c. injection of N-nitroso-bis(2-hydroxypropyl)amine in sa-line (500 mg/kg of body weight) once a week for 15 wk, andanimals were killed 1 wk later. Morris 5123D hepatomas,intramuscularly transplanted in male Buffalo rats, weresupplied by M. Watanabe (Tohoku University in Sendai) andwere used 1 month after inoculation. Yoshida ascites hepa-
Abbreviations: GSH, glutathione; GSHTase, glutathione transferase(EC 2.5.1.18); GSHTase-P, placental form of GSHTase; GluTase,y-glutamyltransferase (EC 2.3.2.2); HN, hyperplastic nodule; 3'-Me-DAB, 3'-methyl-4-dimethylaminoazobenzene; PCB, polychlorin-ated biphenyls; CIN2B, 1-chloro-2,4-dinitrobenzene; C12NB, 1,2-dichloro-4-nitrobenzene.*To whom reprint requests should be addressed.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 8,
202
1
Proc. Natl. Acad. Sci. USA 82 (1985) 3965
toma AH 130 was transplanted and used as reported (16).Fibrosarcomas were induced in Sprague-Dawley rats bysingle s.c. injection of 3-methylcholanthrene (50 mg/kg) inparaffin and were used 6 months after the injection.
Short-Term Administration of Drugs and Carcinogens. Ratswere administered drugs or carcinogens as follows: pheno-barbital (0.02%), butylated hydroxyanisol (0.75%), N-2-fluorenylacetamide (0.02%), and 3'-Me-DAB (0.06%) in basaldiet for 2 weeks; 3-methylcholanthrene, in corn oil by i.p.injection (25 mg/kg) every 2 days for 6 days (rats were killed1 wk later); and PCB in corn oil by i.p. injection once (500mg/kg; rats were killed after 1 wk). Rats were fastedovernight before decapitation.Enzyme Assays. GSHTase was assayed with 1-chloro-2,4-
dinitrobenzene (ClN2B) or 1,2-dichloro-4-nitrobenzene(Cl2NB) as substrate as described by Habig et al. (17).
Tissue Homogenate Preparation. Rat tissues were homog-enized with a Teflon or glass homogenizer in 5 vol of 150mMTris, pH 7.4/193 mM KCl/5 mM EDTA/6.25 mM dithio-threitol. The homogenates were centrifuged at 105,000 x gfor 45 min, and the supernatants were assayed.
Antibody Preparation. Antibody to GSHTase-P purifiedfrom HN-bearing rat livers was prepared in a rabbit asreported (18).
Single Radial Immunodiffusion Method. Single radialimmunodiffusion was performed with the antibody toGSHTase-P as described by Mancini et al. (19).Two-Dimensional Gel Electrophoresis. Two-dimensional
gel electrophoresis (isoelectric focusing/sodium dodecyl sul-fate electrophoresis) was performed as described byO'Farrell (21) and modified by Hirsch et al. (22).Amino Acid Analysis. Purified GSHTase-P (300 ,ug) and an
amino acid standard mixture were hydrolyzed in 6 M HCl at110°C for 20 hr, and the amino acid composition was analyzedwith an amino acid analyzer, model JLC-6AH, JEOL.In Vitro Translation ofmRNAs. mRNAs were isolated from
normal and HN-bearing rat livers, and in vitro translation ina rabbit reticulocyte lysate translation system was performedas reported (23).
RESULTS
Purification of GSHTase-P from HN-Bearing Rat Livers.HN-bearing rat livers (19.7 g) were homogenized in 5 vol of0.25 M sucrose and centrifuged at 10,000 x g for 10 min andfurther centrifuged at 105,000 x g for 45 min. The supernatantwas dialyzed against 10 mM KH2PO4 (pH 6.7) overnight andthen applied to a column of CM-Sephadex (3.5 x 8 cm) thathad been equilibrated with the phosphate buffer. Flow-through fractions were applied to a column of S-hexyl-GSH-Sepharose 6B (1.2 x 8 cm) (24) and washed with =200 ml of10 mM Tris, pH 7.8/0.2 M NaCl, and the adsorbedGSHTases were eluted with the Tris buffer containing 5 mMS-hexyl-GSH. Active fractions were dialyzed against 10 mMTris (pH 8.1) overnight and applied to a column of DE-52cellulose (1.5 x 20 cm), followed by elution with a NaCl lineargradient from 0 (200 ml) to 100 mM (200 ml). Three majorpeaks (I, II, and III) were observed in the elution profile, in
50Fraction
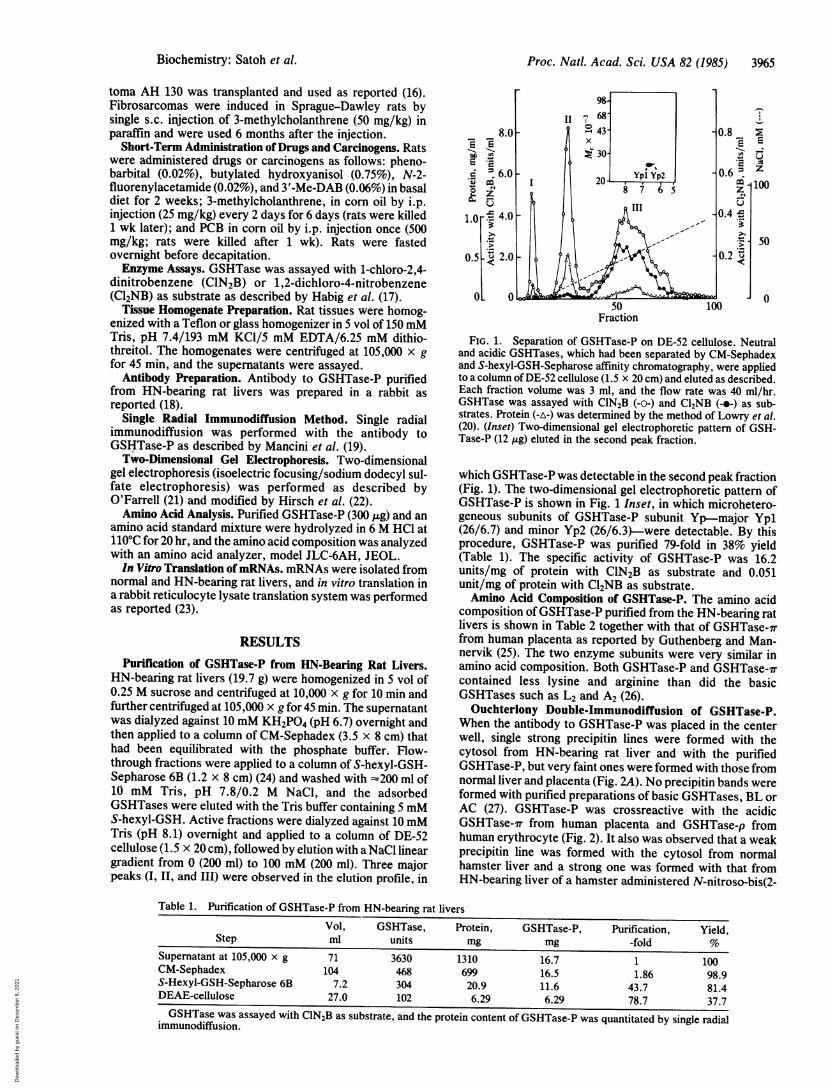
FIG. 1. Separation of GSHTase-P on DE-52 cellulose. Neutraland acidic GSHTases, which had been separated by CM-Sephadexand S-hexyl-GSH-Sepharose affinity chromatography, were appliedto a column ofDE-52 cellulose (1.5 x 20 cm) and eluted as described.Each fraction volume was 3 ml, and the flow rate was 40 ml/hr.GSHTase was assayed with ClN2B (-o-) and Cl2NB (-o-) as sub-strates. Protein (-A-) was determined by the method of Lowry et al.(20). (Inset) Two-dimensional gel electrophoretic pattern of GSH-Tase-P (12 ,g) eluted in the second peak fraction.
which GSHTase-P was detectable in the second peak fraction(Fig. 1). The two-dimensional gel electrophoretic pattern ofGSHTase-P is shown in Fig. 1 Inset, in which microhetero-geneous subunits of GSHTase-P subunit Yp-major Ypl(26/6.7) and minor Yp2 (26/6.3)-were detectable. By thisprocedure, GSHTase-P was purified 79-fold in 38% yield(Table 1). The specific activity of GSHTase-P was 16.2units/mg of protein with ClN2B as substrate and 0.051unit/mg of protein with Cl2NB as substrate.Amino Acid Composition of GSHTase-P. The amino acid
composition ofGSHTase-P purified from the HN-bearing ratlivers is shown in Table 2 together with that of GSHTase-trfrom human placenta as reported by Guthenberg and Man-nervik (25). The two enzyme subunits were very similar inamino acid composition. Both GSHTase-P and GSHTase-ircontained less lysine and arginine than did the basicGSHTases such as L2 and A2 (26).Ouchterlony Double-Immunodiffusion of GSHTase-P.
When the antibody to GSHTase-P was placed in the centerwell, single strong precipitin lines were formed with thecytosol from HN-bearing rat liver and with the purifiedGSHTase-P, but very faint ones were formed with those fromnormal liver and placenta (Fig. 2A). No precipitin bands wereformed with purified preparations of basic GSHTases, BL orAC (27). GSHTase-P was crossreactive with the acidicGSHTase-ir from human placenta and GSHTase-p fromhuman erythrocyte (Fig. 2). It also was observed that a weakprecipitin line was formed with the cytosol from normalhamster liver and a strong one was formed with that fromHN-bearing liver of a hamster administered N-nitroso-bis(2-
Table 1. Purification of GSHTase-P from HN-bearing rat liversVol, GSHTase, Protein, GSHTase-P, Purification, Yield,
Step ml units mg mg -fold SSupernatant at 105,000 x g 71 3630 1310 16.7 1 100CM-Sephadex 104 468 699 16.5 1.86 98.9S-Hexyl-GSH-Sepharose 6B 7.2 304 20.9 11.6 43.7 81.4DEAE-cellulose 27.0 102 6.29 6.29 78.7 37.7GSHTase was assayed with CIN2B as substrate, and the protein content of GSHTase-P was quantitated by single radialimmunodiff-usion.
Biochemistry: Satoh et al.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 8,
202
1

Proc. Natl. Acad. Sci. USA 82 (1985)
Table 2. Amino acid composition of the subunits of GSHTase-Pfrom HN-bearing rat liver and GSHTase-# from human placenta
Amino acid GSHTase-P GSHTase-irAsx 25.8 21.1Thr 8.2 9.2Ser 11.7 10.1Glx 18.9 24.6Pro 12.2 11.8Gly 19.4 18.7Ala 14.8 16.0Cys 4.0 4.0Val 12.3 14.1Met 1.8 2.4Ile 7.3 6.8Leu 28.8 31.2Tyr 11.4 12.0Phe 7.0 6.9His 4.2 2.0Lys 13.7 12.1Trp ND 1.7Arg 9.5 8.0
Values are amino acid residues per subunit. Amino acid composi-tion of GSHTase-ir was cited from ref. 25. Cysteine was determinedas cysteic acid after performic acid oxidation ofGSHTase-P. ND, notdetected.
hydroxypropyl)amine. These results indicate that the placen-tal GSHTase is crossreactive among rat, hamster, and hu-man.
Protein Content of GSHTase-P in Normal, Preneoplastic,and Neoplastic Tissues. The protein contents ofGSHTase-P innormal tissues, HN-bearing livers, and hepatomas weredetermined by single radial immunodiffusion. The proteincontent of GSHTase-P was generally very low in adult andfetal livers, placenta, heart, and other tissues (30-60 Ag/g oftissue) but significantly higher in kidney and pancreas (Table3). Significant but weak induction of GSHTase-P (-2-fold)was observed by 70% partial hepatectomy. The proteincontents in HN-bearing livers and hepatomas induced bydiethylnitrosamine and N-2-fluorenylacetamide plus partialhepatectomy were 20- to 50-fold and 10- to 30-fold higher thanthat in the normal liver, which corresponded to 0.97 ± 0.43%
A B
FIG. 2. Ouchterlony double-immunodiffusion test of GSHTase-P. Center wells contained antibody to GSHTase-P (35 ILI) (well Ab).(A) Wells 1, 3, and 6 are 105,000 x g supernatants (35 pl) fromHN-bearing liver, placenta, and normal liver, respectively, and wells2, 4, and 5 are purified preparations of GSHTase-P (15 ug),GSHTase-BL (14.5 Ag), and GSHTase-AC (13 jug), respectively. Allantigens (1-6) were from rat tissues. (B) Wells 1, 2, and 6 are purifiedpreparations ofrat GSHTase-P (15 .g), human placental GSHTase-vr(15 j.g), and human erythrocyte GSHTase-p (10 jtg); wells 3, 4, andS are 105,000 x g supernatants (35 1l) from HN-bearing rat liver,HN-bearing hamster liver, and normal hamster liver, respectively.GSHTase-IT and -p were purified by the method of Guthenberg andMannervik (25).
and 0.49 + 0.20% (mean + SD) of the soluble cytosolicproteins, respectively. The protein contents in hepatomasinduced by 3'-Me-DAB and Morris hepatoma 5123D were ashigh as those in the above HN-bearing livers induced bydiethylnitrosamine and N-2-fluorenylacetarnide. However,GSHTase-P was undetectably low in AH 130, indicating thatit seems to decrease with increased dedifferentiation ofhepatomas. GSHTase-P was also undetectable in primary3-methylcholanthrene-induced fibrosarcoma.
Induction of GSHTase-P by Short-Term Administration ofDrugs or Carcinogens. Degrees of induction of GSHTase-Pwere low in livers of rats administered phenobarbital, PCB,N-2-fluorenylacetamide, 3'-Me-DAB, and 3-methylchol-anthrene; however, administration of butylated hydroxyan-isole induced about twice the amount of GSHTase-P as incontrol liver.
Table 3. GSHTase-P levels in normal, preneoplastic, and neoplastic tissues
GSHTase-P, GSHTase-P,Tissue n jkg/g of tissue Tissue n ,ug/g of tissue
Normal tissue Primary hepatomaNormal liver 6 34.3 ± 9.3 Et2Nan/AcNHFln-induced 21 591 ± 237Placenta 6 60.5 ± 8.2 3'-Me-DAB-induced 6 1127 ± 164Fetal liver 6 44.5 ± 4.8 Primary sarcomaRegenerating liver Fibrosarcoma, 3-MC-induced 3 <5
24 hr 3 73.7 ± 6.5 Transplantable hepatoma48 hr 3 62.2 ± 11.6 Morris 5123D 3 1120 ± 6772 hr 3 55.8 ± 11.2 Yoshida AH 130 3 <5
Brain 5 34.9 ± 8.7 Livers of rats administered drugs orKidney 6 287 ± 76.9 carcinogenLung 6 98.3 ± 12.2 Phenobarbital 5 38.8 ± 7.5Testis 6 49.9 ± 7.3 BHA 6 66.8 ± 6.0Prostate 4 51.0 ± 14.6 PCB 4 37.7 ± 7.3Spleen 5 104 ± 21.9 AcNHFln 5 47.5 ± 2.1Pancreas 5 163 ± 42 3'-Me-DAB 5 40.2 ± 5.0Heart 5 48.0 ± 6.5 3-MC 3 27.5 ± 2.0Muscle 5 <5
Preneoplastic tissueHN-bearing livers,Et2Nan/AcNHFln-induced 20 1160 ± 520
Values are means + SD; n = number of tissues examined.diethylnitrosamine; AcNHFln, N-2-fluorenylacetamide.
3-MC, 3-methylcholanthrene; BHA, butylated hydroxyanisole; Et2Nan,
3966 Biochemistry: Satoh et al.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 8,
202
1

Proc. Natl. Acad. Sci. USA 82 (1985) 3967
A98-
68-
¶ 43-
x
30-
20-
B
Vc I%A
YiiIyal ypi yp2
I
8 7 6pH
I I
5 8 7
pH
FIG. 3. Two-dimensional gel electrophoretic patterns of in vitrotranslated subunits of GSHTase from mRNAs of normal and HN-bearing livers. In vitro translation was performed in the rabbitreticulocyte lysate translation system. Incubation was carried out at370C for 60 min. [35S]Methionine-labeled translation products frommRNAs of normal and HN-bearing livers (2.04 x 106 cpm and 2.61x 106 cpm, respectively) were applied to separate columns ofS-hexyl-GSH-Sepharose 6B (0.6 x 5 cm) and were eluted with 10mMTris, pH 7.8/0.2 M NaCl/5 mM S-hexyl-GSH. (A and B)Fluorographic patterns of the column-bound fractions of the transla-tion products of mRNAs isolated from normal and the HN-bearinglivers, respectively. Ya, Yb, Yb', Yc, and Yp, including theirmicroheterogeneous subunits were identified immunochemically asdescribed (15).
In Vitro Translation of mRNAs Isolated from Normal andHN-Bearing Livers. Two-dimensional gel electrophoretic pat-terns of the glutathione-affinity column-bound GSHTasestranslated from mRNAs of normal and HN-bearing livers areshown in Fig. 3A and B, respectively. The amounts of all fivesubunits (Ya, Yb, Yb', Yc, and Yp) synthesized frommRNAs of preneoplastic liver were much higher than thoseof normal liver. Among subunits that are present in normalliver, the increased amount of Yb was prominent, as noted invivo (15). However, the Yp subunit was increased mostmarkedly as it was practically undetectable in the normalliver pattern. Microheterogeneous subunits (charge isomers)were detectable not only for the Yp subunit as Ypl and Yp2but also for Yb and Yb' subunits as Yb1 and Yb2, and Yb'land Yb'2, respectively. The molecular weight and pI valuesof these heterogeneous and microheterogeneous subunitswere identical with those observed in the liver cytosol.Microheterogeneity of the GSHTase subunits will be re-ported elsewhere.
DISCUSSIONWe have demonstrated immunohistochemically (14) thatGSHTase-P is localized in HNs; in this study, GSHTase-Pwas purified from whole rat livers bearing HNs induced byadministration of diethylnitrosamine and N-2-fluorenylacet-amide. Basic GSHTases were removed selectively by ad-sorption to a column of CM-Sephadex, and unbound neutraland acidic GSHTases were subjected to GSH-Sepharose 6Baffinity chromatography. GSHTase-P was finally separatedfrom the acidic and residual basic forms on DE-52 cellulosechromatography. The purified enzyme was homogeneous insize, but two charge isomers were detectable on two-dimensional gel electrophoresis. They were named Ypl(26/6.7) and Yp2 (26/6.3) but were indistinguishable im-munochemically.GSHTase-P was different from basic GSHTases in im-
munochemical properties (15) and amino acid composition(26). GSHTase-P was crossreactive among rat, hamster, andhuman, showing that the antibody to GSHTase-P is useful for
immunohistochemical staining of hamster hepatic and pan-creatic preneoplastic foci and HNs (35).GSHTase-P could most conveniently be quantitated by
single radial immunodiffusion as described by Mancini et al.(19). The protein content ofGSHTase-P was generally low innormal tissues, including adult and fetal livers, regeneratinglivers, and others, except that it was significantly high inkidney and pancreas. GSHTase-P was also low in placenta,although it was isolated originally from that tissue where it isthe predominant molecular form of GSHTase. In contrast,GSHTase-P level was high in preneoplastic and neoplastictissues. GSHTase-P contents in the HN-bearing livers andhepatomasinducedbydiethylnitrosamine/N-2-fluorenylacet-amide and in hepatomas induced by 3'-Me-DAB were 30- to50-fold higher than that in the normal liver. It is noteworthythat GSHTase-P was also high in the transplantable moder-ately differentiated Morris hepatoma 5123D but wasundetectably low in the poorly differentiated Yoshida asciteshepatoma AH 130.
Consistent with the immunohistochemical observation thatGSHTase-P was localized in the GluTase-positive foci andHNs (14), degrees of elevation of GSHTase-P and GluTase inthe preneoplastic livers were similar (30- to 50-fold), butGluTase activity was relatively high in the hepatomas (28),whereas GSHTase-P level was rather low. Thus, it seems thatGSHTase-P is more characteristic of preneoplastic thanneoplastic cells. As noted above, it appears that GSHTase-Pis not an organ- or tissue-specific enzyme when comparedwith GluTase, whose activity is extremely high in kidney.The high GluTase activities of preneoplastic and neoplastictissues are much lower than that of normal kidney (28).GSHTase-P was practically uninducible in rat livers by
short-term administration of drugs and carcinogens such asphenobarbital, PCB, N-2-fluorenylacetamide, 3'-Me-DAB,and 3-methylcholanthrene; however, butylated hydroxy-anisole induced GSHTase-P slightly in periportal areas asdetected immunohistochemically.
Since GSHTase-P does not increase prior to the appear-ance of preneoplastic foci and HNs and since it is high in thetransplantable Morris hepatoma 5123D as well, GSHTase-Pincrease is not due simply to administration of drugs orcarcinogens.
Recently, it was confirmed that the Mr 21,000 protein foundby Farber and co-workers (9, 10) was identical with GSHTase-P when examined with the antibody to GSHTase-P preparedby us (E. Farber, personal communication). Although themolecular weight of the subunit of GSHTase-P was estimatedto be 26,000 by us with carbonic anhydrase (Mr 30,000) as thecritical standard, it could be estimated to be 21,000 as wellwith a-chymotrypsinogen (Mr 25,000) as the standard. Re-cently, Reddy et al. estimated the molecular weight ofGSHTase subunits Ya, Yb, and Yc to be 25,600, 27,000, and28,000, respectively, from cDNA analysis (29). Based onthese values, the molecular weight of the subunit ofGSHTase-P can be calculated to be 25,100.As observed by Farber et al., it is interesting to note that
the Mr 21,000 protein-i.e., GSHTase-P-is increased in thepreneoplastic cells independent of their mode of induction (9,10). It should also be noted that GSHTase-P is abundant inthe preneoplastic cells because the actual protein content inthe soluble cytosolic proteins must be several-fold higherthan the apparent one (l1%) in the HN-bearing livers bytaking into account its localization at the foci and HNs. Whenthe protein content of GluTase was estimated from theenzymatic activities, with its specific activity assumed to be1030 units/mg of protein (28), GSHTase-P was 200- to 400-fold higher than GluTase in protein content in the preneo-plastic livers.
Neutral and acidic GSHTases have been observed by
Biochemistry: Satoh et al.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 8,
202
1

Proc. Natl. Acad. Sci. USA 82 (1985)
several investigators (30-32). It is possible that GSHTase-Pis identical with the tissue-specific GSHTase of Mr 24,000,which was found by Tu and co-workers (29, 30).The function of GSHTase-P increase in HNs and hepato-
mas is unknown. However, it has been pointed out thatpreneoplastic cell populations are resistant to the cytotoxicaction ofcarcinogens (1, 33). Since the GSHTases are a groupof multifunctional enzymes involved in the detoxification ofvarious drugs and carcinogens, GSHTase-P, which is abun-dant in the preneoplastic cells, may be expected to have acrucial role in relation to the resistance mechanism or moredirectly to the growth of these cells.
The authors are grateful to Dr. Hajime Matsue for his assistancein the amino acid analysis and Dr. Minro Watanabe for supply ofMorris 5123D hepatomas. This work was supported in part byGrants-in-Aid for Cancer Research (59010009, 59010064, and59015006) from the Ministry of Education, Science, and Culture ofJapan and by a grant from the Karoji Memorial Fund for MedicalResearch in Hirosaki University.
1. Farber, E. & Cameron, R. (1980) Adv. Cancer Res. 31,125-226.
2. Peraino, C., Fry, R. J. M. & Staffelt, E. (1971) Cancer Res.31, 1506-1512.
3. Scherer, E. & Emmelot, P. (1975) Eur. J. Cancer 11, 145-153.4. Solt, D. & Farber, E. (1976) Nature (London) 263, 701-703.5. Pitot, H. C., Barsness, L. & Goldsworthy, T. (1978) Nature
(London) 261, 456-457.6. Levin, W., Lu, A. Y. H., Thomas, P. E., Kizer, D. E. &
Griffin, M. J. (1978) Proc. Nati. Acad. Sci. USA 75,3240-3243.
7. Sato, K., Kitahara, A., Yin, Z., Ebina, T., Satoh, K.,Hatayama, I., Nishimura, K., Yamazaki, T., Tsuda, H., Ito,N. & Dempo, K. (1983) Ann. N. Y. Acad. Sci. 417, 213-223.
8. Sugioka, Y., Fujii-Kuriyama, Y., Kitagawa, T. & Muramatsu,M. (1982) Proc. Jpn. Cancer Assoc., 274 (abstr.).
9. Roomi, M. W., Sharma, R. N., Ho, R. K., Eriksson, L. C.,Farber, E. & Murray, R. K. (1983) Proc. Am. Assoc. CancerRes. 24, 90 (abstr.).
10. Eriksson, L. C., Sharma, R. N., Roomi, M. W., Ho, R. K.,Farber, E. & Murray, R. K. (1983) Biochem. Biophys. Res.Commun. 117, 740-745.
11. Sato, K., Hatayama, I., Hoshino, K., Imai, F., Tsuchida,
S., Sato, T., Nishimura, K., Tatematsu, M. & Ito, N. (1981)Cancer Res. 41, 4147-4153.
12. Yin, Z., Sato, K., Tsuda, H. & Ito, N. (1982) Gann 73,239-248.
13. Kitahara, A., Satoh, K. & Sato, K. (1983) Biochem. Biophys.Res. Commun. 112, 20-28.
14. Sato, K., Kitahara, A., Satoh, K., Ishikawa, T., Tatematsu,M. & Ito, N. (1984) Gann 75, 199-202.
15. Kitahara, A., Satoh, K., Nishimura, K., Ishikawa, T., Ruike,K., Sato, K., Tsuda, H. & Ito, N. (1984) Cancer Res. 44,2698-2703.
16. Sato, K., Takaya, S., Imai, F., Hatayama, I. & Ito, N. (1978)Cancer Res. 38, 3086-3093.
17. Habig, W. H., Pabst, M. J. & Jakoby, W. B. (1974) J. Biol.Chem. 249, 7130-7139.
18. Satoh, K. & Sato, K. (1979) Biochem. Biophys. Acta 578,520-525.
19. Mancini, G., Carbonara, A. 0. & Heremans, J. F. (1965)Immunochemistry 2, 235-254.
20. Lowry, 0. H., Rosebrough, N. J., Farr, A. L. & Randall,R. J. (1951) J. Biol. Chem. 193, 265-275.
21. O'Farrell, P. H. (1975) J. Biol. Chem. 250, 4007-4021.22. Hirsch, R. W., Nall, K. N., Busch, F. N., Morris, H. P. &
Busch, H. (1978) Cancer Res. 38, 1514-1522.23. Satoh, K. & Busch, H. (1983) Cancer Res. 43, 2143-2149.24. Guthenberg, C. & Mannervik, B. (1979) Biochem. Biophys.
Res. Commun. 86, 1304-1310.25. Guthenberg, C. & Mannervik, B. (1981) Biochim. Biophys.
Acta 661, 255-260.26. Beale, D., Meyer, D. J., Taylor, J. B. & Ketterer, B. (1983)
Eur. J. Biochem. 137, 125-129.27. Mannervik, B. & Jensson, H. (1982) J. Biol. Chem. 257,
9909-9912.28. Tsuchida, S., Hoshino, K., Sato, T., Ito, N. & Sato, K. (1979)
Cancer Res. 39, 4200-4205.29. Reddy, C. C., Li, N. & Tu, C.-P. D. (1984) Biochem. Biophys.
Res. Commun. 121, 1014-1020.30. Tu, C.-P. D., Weiss, M. J., Li, N. & Reddy, C. C. (1983) J.
Biol. Chem. 258, 4659-4662.31. Hayes, J. D. (1983) Biochem. J. 213, 625-633.32. Maruyama, H. & Listowsky, I. (1984) J. Biol. Chem. 259,
12449-12455.33. Farber, E. (1984) Cancer Res. 44, 5463-5474.34. Sugioka, Y., Fujii-Kuriyama, Y., Kitagawa, T. & Muramatsu,
M. (1985) Cancer Res. 45, 365-378.35. Moore, M. A., Satoh, K., Kitahara, A., Sato, K. & Ito, N.
(1985) Gann 76, 1-4.
3968 Biochemistry: Satoh et al.
Dow
nloa
ded
by g
uest
on
Dec
embe
r 8,
202
1