Embed Size (px)
Citation preview

RAPHAEL MOLINARO COELHO
O PAPEL DO CCR4 SOBRE AS CÉLULAS T REGULATÓRIAS NA SEPSE
GRAVE
UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
INSTITUTO DE MICROBIOLOGIA PROF PAULO DE GÓES
RIO DE JANEIRO
FEVEREIRO DE 2009

Livros Grátis
http://www.livrosgratis.com.br
Milhares de livros grátis para download.

II
RAPHAEL MOLINARO COELHO
O PAPEL DO CCR4 SOBRE AS CÉLULAS T REGULATÓRIAS NA SEPSE
GRAVE
Dissertação de mestrado apresentada ao Programa de Pós-
Graduação em Ciências (Microbiologia), Instituto de Microbiologia
Prof. Paulo de Góes da Universidade Federal do Rio de Janeiro,
como requisito para obtenção do título de Mestre em ciências
(microbiologia).
Orientadores:
Marcelo Torres Bozza
Claudia Farias Benjamim
UNIVERSIDADE FEDERAL DO RIO DE JANEIRO
INSTITUTO DE MICROBIOLOGIA PROF PAULO DE GÓES
RIO DE JANEIRO
FEVEREIRO DE 2009

III
RAPHAEL MOLINARO COELHO
O PAPEL DO CCR4 SOBRE AS CÉLULAS T REGULATÓRIAS NA SEPSE GRAVE.
Rio de Janeiro, 18 de Fevereiro de 2009.
Aprovado por:
_____________________________________
Prof. Dra. Marcela Lopes - IBCCF- UFRJ
_____________________________________
Dr. Hugo Castro Faria – FIOCRUZ
____________________________________
Prof. Dr. Alexandre Morrot- IMPPG-UFRJ
_____________________________________
Prof. Dra. Luciana Arruda - IMPPG-UFRJ (REVISORA)
Fevereiro de 2009

IV
O presente trabalho foi realizado no Laboratório de Inflamação e Câncer,
Departamento de Farmacologia, Instituto de Cências Biomédicas (ICB), Centro de
Ciências da Saúde (CCS), Universidade Federal do Rio de Janeiro, sob a orientação
da Prof(a) Claudia Farias Benjamim e do Prof. Marcelo Torres Bozza.

V
Ficha Catalográfica
Molinaro, Raphael Coelho
O papel do CCR4 sobre as células T regulatórias na sepse grave /
Raphael Molinaro Coelho - Rio de Janeiro, 2009.
XIV, f.83
Dissertação (Mestrado em Ciências Biológicas)
Universidade Federal do Rio de Janeiro / Instituto de Microbiologia
Prof. Paulo de Góes, 2009
Orientador: Marcelo Torres Bozza
Referências bibliográficas: f. 72-80.
1. Quimiocinas 2. CCR4 3. Sepse 4. Imunossupressão 5.
Células Tregulatórias 6. Inflamação I. Bozza, Marcelo II. UFRJ,
Instituto de Microbiologia Prof Paulo de Góes, Mestrado em Ciências
Biológicas III. O papel do CCR4 sobre as células T regulatórias na
sepse grave

VI
“ Passarinho que come pedra,
sabe o futuro que tem.”
Vanessa Martins

VII
AGRADECIMENTOS
A Prof.a Claudia Farias Benjamim que além de orientadora e professora, foi uma
grande amiga durante esses anos. A sua orientação com determinação e
brilhantismo trouxe lucros a todos que você incentivou. E de uma forma muito
especial, você pode demonstrar que com garra e muito trabalho nossas idéias
ganham gráficos e figuras. A sua eterna dedicação, paciência e atenção foram
apenas uma das muitas qualidades que me ajudou a me torna um profissional que
luta com alma e entusiasmo. A você, muitíssimo obrigado.
Ao Prof. Marcelo Torres Bozza por ter me ensinado da definição da palavra
caráter e sabedoria. Você foi para o meu profissional, um dos grandes incentivos por
dar tanta beleza e fé na ciência.
À minha “partner” de bancada, Cyntia Pecli, que desenvolveu grande parte deste
trabalho comigo e sempre foi companheira nos MEGA experimentos até as longas
tardes da noite com as Treginhas e os revoltadissímos neutrófilos. Gostaria de
agradecer a Deus pela sua amizade, e por me ensinar tantas coisas e deixar eu te
ensinar apenas algumas. Se eu fosse seu aluno, diria que seria o melhor aprendiz
do mundo, mas na verdade eu sou e sempre serei seu amigo. Sentirei muita falta
dos nossos risos. Obrigado Cyntia, por ter feito eu dizer: - Valeu!!!
Ao Carlos Alberto que sempre está disposto a ajudar e deixar compartilhar um
projeto comigo. A Cristiane Sécca que me ensinou a correr atrás do sucesso e por
me fazer rir de tantas loucuras que contamos no laboratório. Ao Leandro Lasdislau
que com sua insistência e fé tem demonstrado ao laboratório que tudo é possível. A
nova aquisição do lab, a aluna Janaina, por estar participando e apreendendo no
lab. E a Vanessa Martins pela sua majestosa alegria e descontração. Além de ter
dado o primeiro artigo para o Lab, me ajudou durante todos esses anos. A Ariane
mesmo estando longe que me viu crescer cientificamente.

VIII
À minha extensão de laboratório que foi com a galera do Marcelo Bozza: Beth,
Fabiano, Daniel Feijó, Tatiana, Guilherme, Raquel, Jaci, Marta, Claudia Paiva,
Camila, Cristine, Isadora, Iranaia e a Letícia. A essa galera eu devo de tudo, vocês
foram meus exemplos de pesquisadores.
A expansão da minha amizade não se limitou apenas ao laboratório do Marcelo
Bozza. A Ana Paula, Carla e a Tati pelos papos e risos intermináveis. A Taiane e a
Aline que me incentivaram e ajudaram em muitas decisões técnicas. Ao Paulo
Emílio pela demonstração de parceria e colaboração, além de tudo um amigo.
Ao Steve Kunkel por ter confiado na nossa capacidade, investido no nosso
laboratório e por ser um magnífico exemplo de pesquisador com toda essa
espantosa humildade. Você me fez ainda mais apaixonado pela pesquisa devido as
nossas conversas no congresso.
Aos professores e doutores da UFRJ, FIOCRUZ, UFF, UERJ, UMICH e do INCA,
que ajudaram e confiaram tanto em mim jamais serão esquecidos. Desejo que
continuemos com as colaborações cada vez mais fortes. E a todos os professores,
alunos e funcionários do Departamento da Farmacologia – UFRJ, inclusive o Prof.
Newton que foi espetacular por deixado às chaves do seu laboratório e sua
confiança conosco.
Aos órgãos de fomento CNPq, FAPERJ, pelo suporte financeiro. E a todas as
pessoas que me ajudaram a sanar as minhas intermináveis listas de pedidos, sem
vocês não teria conseguido.
À minha família, meus pais Antonio Jorge e Maria Helena, a Cássia e o
Alessandro, pelo carinho e amor. Por ter especialmente lutado comigo pelos meus
sonhos a vida inteira. Amo vocês demais.

IX
LISTA DE ABREVIATURAS
ASP Aspergillus fumigatus
BAL Lavado broncoalveolar
BSA Albumina de soro bovino
cAMP AMP cíclico
CARS Síndrome da resposta compensatória anti-inflamatória
CCR4 Receptor de quimiocina CC do tipo 4
CCS Centro de Ciências da Saúde
CD “Cluster” de diferenciação
CEUA Comissão de uso de animais
CFU Unidade Formadora de Colônia
CLP Ligadura e perfuração do ceco
CpG Seqüência de DNA de citosina seguida de guanina
CTLA-4 Antígeno 4 dos linfócitos T citotóxicos
DCs Células dendríticas
ELISA Ensaio imunoenzimático
EPM Erro padrão médio
FACS Citometria de fluxo
FITC Isotiocianato de fluoresceína
Foxp3 Fator de transcrição forkhead-box p3
FSH Detector dispersão frontal
H&E Hematoxilina e Eosina
HLA Antígenos leucocitários humanos
i.t. Intratraqueal
IL Interleucina

X
iNOS Oxído Nítrico Sintase Induzida
IPA Aspergilose pulmonar invasiva
IRAK Quinase associada ao receptor IL-1 - M
KC Quimiocinas derivada de Queratinocitos
kD Kilodaltons
L Sepse Grave
LNM Linfonodo mesentérico
LPS Lipopolissacarídeo
LTB4 Leucotrieno B4
MCP-1 Proteína quimiotática de macrófago
MDC Quimiocina derivada de macrófagos
MFI Intensidade média de fluorescência
MHC II Complexo de histocompatibilidade principal do tipo II
MIP Proteína inflamatória produzida por macrófagos
MPO Mieloperoxidase
NK Natural “killer”
NO Óxido nítrico
OPD o-fenilenediamina-dihidrocloreto
Pam3CysKKKK Lipopeptideo Pam 3 Cys-Ser-(Lys)4
PBS Solução salina de fosfato tamponada
PerCP Proteína clorofila peridinina
PMA Acetato de formol miristato
PMN polimorfonucleares
ROS Espécies reativas de oxigênio
RPM Rotações por minuto
SAL Solução fisiológica de NaCl 0,9%
SFB Soro fetal bovino

XI
SIRS Síndrome da resposta inflamatória sistêmica
SL Sepse Moderada
SN Sobrenadante
SSC Detector dispersor lateral
TARC Quimiocinas ativadas e reguladas pelo timo
TGF Fator crescimento transformante
Th Célula T auxiliar
TLR Receptores semelhantes a toll
TNF-α Fator de necrose tumoral alfa
Tregs T regulatórias
UTI Unidade de Tratamento Intensivo
WT Selvagens

XII
RESUMO
O PAPEL DO CCR4 SOBRE AS CÉLULAS T REGULATÓRIAS NA SEPSE.
Autor: Raphael Molinaro Coelho
Orientador: Marcelo Torres Bozza
Resumo da dissertação de mestrado submetida ao Programa de Pós-
Graduação em Ciências (Microbiologia), Instituto de Microbiologia Prof. Paulo de
Góes da Universidade Federal do Rio de Janeiro, como parte dos requisitos
necessários para a obtenção do título de Mestre em Ciências Biológicas.
A sepse é uma resposta inflamatória sistêmica contra uma infecção. Essa
resposta é muita intensa, visto que induz a produção de diversos mediadores
inflamatórios, a expressão de seus receptores, como também a ativação celular.
Esses receptores e seus mediadores também estão envolvidos em uma das
conseqüências da sepse, que é a evolução a um quadro de imunossupressão. Um
dos possíveis receptores envolvidos na evolução deste quadro é o CCR4, que tem
sido descrito em diversas patologias inflamatórias e está expresso em uma das
células capazes de modular negativamente o sistema imune, as células T
regulatórias (Tregs). O nosso objetivo foi investigar o papel do receptor CCR4 sobre
as células Tregs durante a sepse grave e na imunossupressão. Para isto, animais
selvagens (WT) e CCR4 deficientes (CCR4-/-) foram submetidos a uma sepse
polimicrobiana experimental através do modelo ligação e perfuração do ceco (CLP).
Os animais CCR4-/- sépticos apresentaram uma melhor sobrevida quando
submetidos à sepse grave. A resistência do animal foi acompanhada de uma maior
migração de neutrófilos para o peritôneo, redução das bactérias no peritônio e no
sangue, menor infiltrado neutrofílico pulmonar e uma menor produção de citocinas
inflamatória na fase aguda da sepse. O número de Tregs circulantes e esplênicas
parecem influenciar no controle da intensidade da resposta após 1 dia da cirurgia,
mas após 5 dias da cirurgia não observamos diferenças das células Tregs nos

XIII
órgãos linfóides. Os animais CCR4-/- também foram mais resistentes a uma infecção
secundária. E isto pode ser observado pela integridade dos pulmões destes animais.
As Tregs dos animais CCR4-/- não apresentaram a função supressora ativa sobre os
neutrófilos “in vivo” e “in vitro”. Além disso, o sobrenadante das Tregs dos CCR4-/-
não inibem a produção de ROS pelos neutrófilos. Portanto, os nossos dados
sugerem que o CCR4 possui um papel prejudicial na resposta inata contra a
infecção ocasionando o desenvolvimento de uma resposta imune exacerbada e
assim, evoluindo a um quadro de imunossupressão.
Palavras-chave: 1. Quimiocinas, 2. CCR4, 3. Sepse, 4. Inflamação, 5.
Células T regulatórias, 6. Imunossupressão.
Rio de Janeiro
Fevereiro de 2009

XIV
ABSTRACT
THE ROLE OF CCR4 ON T REGULATORY CELLS IN SEPSIS.
Author: Raphael Molinaro Coelho
Marcelo Torres Bozza
Abstract da dissertação de mestrado submetida ao Programa de Pós-Graduação
em Ciências (Microbiologia), Instituto de Microbiologia Prof. Paulo de Góes da
Universidade Federal do Rio de Janeiro, como parte dos requisitos necessários para
a obtenção do título de Mestre em Ciências Biológicas.
Sepsis is a systemic inflammatory response against infection. Overwhelming
inflammatory response induces the production of many mediators, expression of
theirs receptors and activation of different cell types. The mediators receptors have a
critical role to the development of immunossupression. CCR4 is one of these
chemokine receptors, which has been described to be involved in many inflammatory
diseases. This receptor is expressed on T regulatory (Tregs) cells and presents a
suppressive effect on immune response. Our aim was to investigate the role of CCR4
on Tregs cells during sepsis and immunosupression. Wild Type (WT) and CCR4
knockout (CCR4-/-) mice were subjected to cecal ligation and puncture (CLP) model.
Septic CCR4-/- mice showed higher survival rate to lethal sepsis as compared to
septic WT mice. The CCR4-/- resistance was due to a better neutrophils migration to
peritoneum, decreased bacterial count, less MPO levels and less inflammatory
cytokines on acute sepsis. The number of circulating and splenic Tregs cells were
increased in CCR4-/- mice on 1 day after surgery, but did not show differences after
5 days in lymphoid tissue. CCR4-/- mice were also resistant to secondary infection.
Tregs cells from CCR4-/- mice did not present suppressive function on neutrophils “in

XV
vivo” and “in vitro”. Furthermore, the supernatants of Tregs cells from septic CCR4-/-
mice did not inhibit ROS production by neutrophils. Our results suggest that CCR4
has a deleterious effect in innate immune response against the infection and
contributes to immunosuppression stage.
Keywords: 1. Chemokines, 2. CCR4, 3. Sepsis, 4. Inflammation, 5. T
regulatory cells, 6. Immunossupression.
Rio de Janeiro
Fevereiro de 2009

SUMÁRIO
1. INTRODUÇÃO ........................................................................................................1
1.1. MEDIADOR INFLAMATÓRIO: QUIMIOCINAS .............................................................................................. 1
1.2. SEPSE................................................................................................................................................... 7
1.2.1. MODELO EXPERIMENTAL DE SEPSE ...................................................................................................... 12
1.3. CÉLULAS T REGULATÓRIAS (TREGS) ............................................................................................... 16
2. OBJETIVOS..........................................................................................................21
2.1. OBJETIVOS ESPECÍFICOS....................................................................................................................... 21
3. METODOLOGIA ...................................................................................................22
3.1. ANIMAIS.............................................................................................................................................. 22
3.2. MODELO DE LIGAÇÃO DO CECO E PERFURAÇÃO (CLP)....................................................................... 22
3.3. AVALIAÇÃO DA RESPOSTA IMUNE INATA EM ANIMAIS WT E CCR4-/- DEPOIS DE 6 HORAS DE SUBMETIDOS A
UMA SEPSE GRAVE E MODERADA. ................................................................................................................ 23
3.4. CONTAGEM TOTAL DE LEUCÓCITOS. ..................................................................................................... 24
3.5. CONTAGEM DIFERENCIAL DE LEUCÓCITOS. ........................................................................................... 24
3.6. QUANTIFICAÇÃO DE BACTÉRIAS NO SANGUE E EXSUDATO PERITONEAL.................................................... 25
3.7. QUANTIFICAÇÃO DE CITOCINAS. .......................................................................................................... 25
3.8. ENSAIO DA ATIVIDADE DA ENZIMA MIELOPEROXIDASE (MPO)................................................................ 26
3.9. ESFREGAÇO SANGUÍNEO E DIFERENCIAL DE MEDULA ÓSSEA. .................................................................. 26
3.10. ANÁLISE TEMPORAL DAS CÉLULAS TREGS. ............................................................................................ 27
3.11. CITOMETRIA DE FLUXO. ...................................................................................................................... 28
3.12. CULTIVO DE ASP............................................................................................................................... 28
3.13. AVALIAÇÃO DA IMUNOSSUPRESSÃO PÓS-SEPSE. .................................................................................. 29
3.14. LAVADO BRONCOALVEOLAR (BAL). .................................................................................................. 30
3.15. HISTOLOGIA. ..................................................................................................................................... 30
3.16. PURIFICAÇÃO E MARCAÇÃO DE CÉLULAS CD4+FOXP3+ NO PULMÃO.................................................. 30
3.17. ISOLAMENTO DE CÉLULAS T REGULATÓRIAS. ......................................................................................... 31
3.18. INIBIÇÃO DA MIGRAÇÃO DE LEUCÓCITOS PARA UM FOCO INFECCIOSO “IN VIVO” VIA CÉLULAS T
REGULATÓRIAS............................................................................................................................................ 32
3.19. PURIFICAÇÃO DE NEUTRÓFILOS DE MEDULA ÓSSEA. ............................................................................. 33
3.20. INIBIÇÃO DA MIGRAÇÃO DE NEUTRÓFILOS “IN VITRO” VIA CÉLULAS T REGULATÓRIAS. ............................ 33
3.21. INIBIÇÃO DA PRODUÇÃO DE ROS POR NEUTRÓFILOS ESTIMULADOS COM PMA MEDIADA POR TREGS.... 34
3.22. ANÁLISE ESTATÍSTICA. ......................................................................................................................... 35

4. RESULTADOS......................................................................................................36
4.1. SOBREVIDA DE ANIMAIS WT E CCR4-/- AO CLP.................................................................................. 36
4.2. MIGRAÇÃO CELULAR APÓS ESTÍMULO DE SEPSE GRAVE E MODERADA..................................................... 37
4.3. AVALIAÇÃO DO NÚMERO DE BACTÉRIAS NA CAVIDADE PERITONEAL....................................................... 39
4.4. QUANTIFICAÇÃO DE CITOCINAS LOCAIS. .............................................................................................. 40
4.5. DETERMINAÇÃO DA ATIVIDADE DA MIELOPEROXIDASE NO PULMÃO........................................................ 42
4.6. QUANTIFICAÇÃO DE CITOCINAS SISTÊMICAS.......................................................................................... 43
4.7. AVALIAÇÃO DO LEUCOGRAMA E CELULARIDADE DA MEDULA ÓSSEA. .................................................... 44
4.8. PRESENÇA DAS CÉLULAS TREGS NO SANGUE PERIFÉRICO E ÓRGÃOS LINFÓIDES DURANTE A SEPSE GRAVE. . 45
4.9. SOBREVIDA DOS ANIMAIS WT E CCR4-/- SUBMETIDOS AO CLP OU SHAM OPERADOS DESAFIADOS COM
CONÍDIOS DE ASP. ..................................................................................................................................... 47
4.10. AVALIAÇÃO DA MIGRAÇÃO CELULAR PARA O PULMÃO FRENTE A UM ESTÍMULO INFECCIOSO. ............... 48
4.11. ANÁLISE HISTOLÓGICA DO PULMÃO DE ANIMAIS WT E CCR4-/- SUBMETIDOS À SEPSE E DESAFIADOS COM
ASP........................................................................................................................................................... 49
4.12. PRESENÇA DAS CÉLULAS TREGS NO SANGUE PERIFÉRICO E ÓRGÃOS LINFÓIDES 5 DIAS APÓS CLP. ......... 51
4.13. PRESENÇA DE CÉLULAS TREGS NO PULMÃO DE ANIMAIS PÓS-SÉPTICOS DESAFIADOS COM ASP OU SALINA.
.................................................................................................................................................................. 53
4.14. INIBIÇÃO DA MIGRAÇÃO DE NEUTRÓFILOS PARA O ESPAÇO BRONCOALVEOLAR VIA TREGS OBTIDAS DE
ANIMAIS WT E CCR4-/- SUBMETIDOS OU NÃO AO CLP. .............................................................................. 55
4.15. INIBIÇÃO DA MIGRAÇÃO DE NEUTRÓFILOS “IN VITRO” VIA TREG OBTIDAS DE ANIMAIS WT E CCR4-/-
SUBMETIDOS OU NÃO AO CLP..................................................................................................................... 57
4.16. INIBIÇÃO DA PRODUÇÃO DE ROS POR NEUTRÓFILOS ESTIMULADOS COM PMA PELO SN DE TREGS
OBTIDAS DE ANIMAIS WT E CCR4-/- SUBMETIDOS OU NÃO AO CLP. ............................................................ 59
5. DISCUSSÃO .........................................................................................................61
6. CONCLUSÕES.....................................................................................................71
7. BIBLIOGRAFIA ....................................................................................................72
8. ARTIGO I: .............................................................................................................79

ÍNDICE DE FIGURAS
FIGURA 01. Relação dos receptores de quimiocinas e seus ligantes........................ 2
FIGURA 02. Relação dos receptores de quimiocinas e doenças relacionadas.......... 4
FIGURA 03. Relação entre incidência e mortalidade na sepse.................................. 8
FIGURA 04. Processo de migração de neutrófilos do sangue para sítio infeccioso... 9
FIGURA 05. Curva temporal da sepse até a imunossupressão.. ............................. 11
FIGURA 06. Seqüência de eventos que permitem a imunossupressão observada
seguida da sepse...................................................................................................... 14
FIGURA 07. Alvos das células Tregs. ...................................................................... 15
FIGURA 08. Envolvimento das Tregs em diferentes estágios patológicos............... 17
FIGURA 09. Sobrevida ao CLP. ............................................................................... 22
FIGURA 10. Avaliação da resposta imune inata. ..................................................... 23
FIGURA 11. Análise temporal das células Tregs. .................................................... 26
FIGURA 12. Imunossupressão pós-sepse. .............................................................. 28
FIGURA 13. Isolamento e marcação de CD4+FOXP3+ pulmonar. ........................... 30
FIGURA 14. Inibição da migração celular “in vivo” pelas células tregs. ................... 31
FIGURA 15. Inibição da função de neutrófilos “in vitro” pelas células Tregs............ 33
FIGURA 16. Sobrevida de animais WT e CCR4-/- submetidos à sepse grave. ....... 35
FIGURA 17. Migração de neutrófilos para a cavidade peritoneal durante a sepse .. 36
FIGURA 18. Quantificação do número de CFU no lavado peritoneal e sangue de
animais submetidos a sepse moderada (SL) ou a sepse grave (L). ......................... 38
FIGURA 19. Quantificação de citocinas presentes no exsudato na sepse............... 39
FIGURA 20. Determinação do infiltrado neutrofílico pela atividade da MPO............ 40
FIGURA 21. Quantificação de citocinas no plasma de animais sépticos. ................ 41
FIGURA 22. Leucograma e contagem de neutrófilos na medula óssea................... 42

FIGURA 23. População de células Tregs no sangue periférico, baço e linfonodo
mesentérico 1 dia após a cirurgia. ............................................................................ 44
FIGURA 24. Curva de sobrevida dos animais WT e CCR4-/- desafiados com 7,5x107
conídios de ASP depois de serem submetidos ao CLP ou Sham ............................46
FIGURA 25. Migração de neutrófilos para o BAL após 14 e 48 horas do desafio com
asp ou salina depois de serem submetidos ao CLP ou Sham...................................47
FIGURA 26. Analise histológica do pulmão de animais WT e CCR4-/- desafiados
com ASP................................................................................................................... 48
FIGURA 27. Presença de células Tregs no sangue periférico, baço e linfonodo
mesentérico após 5 dias da cirurgia. ........................................................................ 50
FIGURA 28. Presença de Tregs no pulmão de animais WT e CCR4-/- submetidos ao
CLP e desafiados com ASP. .................................................................................... 52
FIGURA 29. Migração de neutrófilos para o lavado broncoalveolar de animais
desafiados com ASP + células Tregs ....................................................................... 54
FIGURA 30. Inibição da migração de neutrófilos “in vitro” via tregs obtidas de
animais WT e CCR4-/-.............................................................................................. 56
FIGURA 31. Inibição da produção de ROS pelo sobrenadante de cultura de células
Tregs. ....................................................................................................................... 57
FIGURA 32. Importância do CCR4 na sepse para a evolução da
imunossupressão.......................................................................................................68

1
1. INTRODUÇÃO
Atualmente, sabe-se que o processo inflamatório está envolvido em várias
doenças como obesidade, diabetes, neurodegeneração, sepse e câncer, e esse
envolvimento em muitas delas era desconhecido até recentemente. Algumas
doenças consideradas antes como degenerativas ou próprias do envelhecimento,
incluem a inflamação como parte de seu processo fisiopatológico.
O processo inflamatório é uma resposta do organismo a uma injúria. Entende-
se como injúria qualquer processo capaz de causar lesão celular ou tecidual. A
resposta a lesão é comum em vários tipos de tecidos e é mediada por diversos
mediadores inflamatórios produzidos pelas células danificadas e/ou por células
residentes ou recrutadas ao sitio inflamatório. Neste sentido, o nosso laboratório
vem investigando a participação de mediadores inflamatórios e de células da
resposta imune na evolução de doenças inflamatórias, incluindo a sepse.
1.1. Mediador inflamatório: quimiocinas
As quimiocinas são importantes moléculas envolvidas no controle da
migração de células, principalmente células do sistema imune, para determinados
sítios no organismo. As quimiocinas são secretadas por diversos tipos celulares,
geralmente quando estas células são estimuladas por algum estímulo inflamatório,
ou secretadas de forma constitutiva. As quimiocinas pertencem à superfamília das
citocinas e são pequenas moléculas protéicas de 8 a 15 kD que possuem diversas
atividades biológicas. A definição das quimiocinas se deve à sua seqüência de 4
resíduos de cisteína na porção amino-terminal. Elas são subdivididas em quatro
subfamílias distintas: CC, CXC, C e CX3C, sendo X um aminoácido qualquer.
Existem duas grandes subfamílias, uma delas é composta por 28 ligantes (CCL1 a
CCL28) e a outra contém 16 ligantes (CXCL1 a CXCL16). As outras 2 menores são
representadas por uma que contém apenas 2 ligantes (XCL1 e XCL2) e a última que
tem apenas o CX3CL1 (Figura 01) (COMERFORD & NIBBS, 2005).

2
Receptores Ligantes
CCR1 CCL3, CCL5, CCL7, CCL13, CCL14, CCL15, CCL16, CCL23
CCR2 CCL2, CCL7, CCL8, CCL13, CCL16
CCR3 CCL5, CCL7, CCL8, CCL11, CCL13, CCL15, CCL16, CCL24, CCL26, CCL28
CCR4 CCL17, CCL22
CCR5 CCL3, CCL4, CCL5, CCL8, CCL11, CCL14, CCL16
CCR6 CCL20
CCR7 CCL19, CCL21
CCR8 CCL1
CCR9 CCL25
CCR10 CCL27, CCL28
CCR11 CCL19,CCL21, CCL25
CXCR1 CXCL6, CXCL7, CXCL8
CXCR2 CXCL1, CXCL2, CXCL3, CXCL5, CXCL6, CXCL7, CXCL8
CXCR3-A CXCL9, CXCL10, CXCL11
CXCR3-B CXCL4, CXCL9, CXCL10, CXCL11
CXCR4 CXCL12
CXCR5 CXCL13
CXCR6 CXCL16
CXCR7 CXCL12
XCR1 XCL1, XCL2
CXXXCR1 CXXXCL1
Figura 01. Relação dos receptores de quimiocinas e seus ligantes.
Adaptado de COMERFORD, 2005.
As quimiocinas se ligam aos receptores de sete alças transmembranares
associado às proteínas Gs. Dentro de cada subfamília, particularmente entre as
quimiocinas que participam do processo inflamatório, observa-se uma grande
promiscuidade, em que um receptor pode ser alvo de ligação de várias quimiocinas
ou vice-versa. As quimiocinas atuam através destes receptores de alta afinidade
expostos na superfície das células. Atualmente já foram identificados onze
receptores diferentes para quimiocinas CC (chamadas de CCR1 a CCR11) e sete
para quimiocinas CXC (chamadas CXCR1 a CXCR7)(COMERFORD & NIBBS,
2005).

3
As quimiocinas também podem ser divididas em dois grupos: constitutivas ou
induzidas, dependendo da sua expressão. As quimiocinas constitutivas são
expressas primariamente em órgãos linfóides secundários onde terão papel
importante para direcionar os leucócitos aos órgãos alvos. As quimiocinas induzidas
ou inflamatórias têm um papel muito importante em direcionar as células para o foco
inflamatório (MOSER & LOETSCHER, 2001) e estas são expressas em altas
concentrações quando são induzidas através de estímulos inflamatórios como, por
exemplo, TNF-α, IL-1β e/ou LPS.
As quimiocinas e seus receptores junto com as moléculas de adesão
(integrinas e selectinas) atuam sobre diversos tipos celulares direcionando-os
seletivamente para seus tecidos alvos (LEY, 2003; ONO et al., 2003). Por exemplo,
o recrutamento de células T virgens, células dendríticas maduras e algumas células
T de memória para o linfonodo depende dos ligantes CCL9 e CCL21. A interação
destas células no órgão linfóide é importante para que ocorra uma resposta imune
dependente de células T (KUNKEL & BUTCHER, 2002).
A resposta imune requer uma migração coordenada dos leucócitos da
circulação sanguínea e linfática para os tecidos, e dentro destes para sítios
determinados seguindo um gradiente de concentração de quimioatraentes. Este
processo é regulado por uma extensa rede de quimioatraentes, como quimiocinas,
mediadores lipídicos, produtos microbianos e proteínas do sistema complemento.
As quimiocinas e seus receptores não são simples moléculas quimiotáticas.
Com o avanço das pesquisas foram descritos outros papéis além da função
quimiotática. As quimiocinas também estão envolvidas no desenvolvimento,
regulação e ativação celular. As quimiocinas atuam em inúmeros processos
incluindo a angiogênese/angiostase, diferenciação e ativação celular, injúria,
crescimento tumoral e metástase, localização e desenvolvimento de tecidos
linfóides, na diferenciação e balanço da resposta imune Th1/Th2 e sobre ativação e
migração de células T regulatórias (ROSSI & ZLOTNIK, 2000; D'AMBROSIO;
PANINA-BORDIGNON & SINIGAGLIA, 2003; ROSENKILDE & SCHWARTZ, 2004;
YI et al., 2006). Além disso, as quimiocinas participam da interação entre a resposta
imune inata e adquirida, sendo esta interação fundamental para uma resposta
automática, dinâmica, duradoura e regulada da homeostase do organismo. É

4
importante considerar a resposta imune inata e adquirida como parte de um
processo integrado (BEUTLER & POLTORAK, 2001; MAZZONI & SEGAL, 2004).
Vários trabalhos demonstram que quimiocinas CC são componentes
importantes para o recrutamento e ativação de células T, monócitos e eosinófilos,
normalmente associadas com a inflamação crônica (KUNKEL & BUTCHER, 2002;
OLSON & LEY, 2002). Com base em estudos clínicos, as quimiocinas CC e seus
receptores estão também associados a diversas patologias tornando-as moléculas
de grande importância médica para o desenvolvimento de novos alvos terapêuticos
de diversas doenças (Figura 02).
Alvo Terapêutico Indicações
CCR1 Esclerose múltipla, artrite reumatóide, câncer, rejeição ao transplante, doença renal
CCR2 Esclerose múltipla, artrite reumatóide, diabete, obesidade, aterosclerose, câncer, asma
CCR3 Asma, rinite alérgica
CCR4 Asma, doença alérgica
CCR5 HIV, transplante, câncer
CCR6 Asma
CCR7 Câncer
CCR9 Doença intestinal
CXCR1 e CXCR2 Sepse, Aterosclerose, artrite reumatoide, doença pulmonar crônica obstrutiva
CXCR3 Transplante, psoríase, esclerose múltipla, artrite reumatoide, câncer
CXCR4 HIV, câncer
CX3CR Aterosclerose
Figura 02. Relação dos receptores de quimiocinas e doenças relacionadas.
Adaptado de COMERFORD, 2005.
Receptores de quimiocinas da subfamília CC, como o CCR1, CCR2, CCR4,
CCR5, CCR6, CCR8 e seus ligantes estão envolvidos tanto na resposta imune

5
adquirida como na resposta imune inata. Um dos aspectos interessantes é que eles
podem modular a expressão de citocinas. Por exemplo, o CCL2 (ligante do receptor
CCR2) promove uma resposta inflamatória do tipo Th2 em modelo de granuloma
pulmonar induzido por Shistosoma mansoni via aumento da expressão de IL-4,
como também inibe a produção de IL-12 por macrófagos no granuloma (CHENSUE
et al., 1996). Além de modular citocinas, o CCR2 é capaz de modular o recrutamento
de macrófagos e células CD8 frente a uma infecção por Listeria monocytogenes
(KURIHARA et al., 1997). Também foi demonstrado que o CCR6 participa no
recrutamento de células T de memória para a pele (KLEINEWIETFELD et al., 2005).
Em modelo de peritonite aguda, a neutralização do CCL2 por anticorpos é capaz de
diminuir as quantidades de IL-12 e aumentar de IL-10 expressas no pulmão
(MATSUKAWA et al., 2000).
Dentre os receptores de quimiocinas o CCR4 é um receptor muito
interessante. O CCR4 possui alta afinidade por CCL17 (TARC) e CCL22 (MDC). O
RNAm do receptor é expresso no baço e em leucócitos, incluindo células
dendríticas, células T, basófilos, monócitos, macrófagos e plaquetas. Os primeiros
estudos sobre o receptor CCR4 na resposta imune demonstraram que este receptor
era importante no recrutamento de células relacionadas à resposta imune tipo Th2
(CHVATCHKO et al., 2000). Estudos mais recentes demonstram que o CCR4 possui
um papel essencial na imunidade inata e adquirida (ISHII et al., 2008). Neste
contexto, SCHUH et al. demonstraram que animais deficientes para o CCR4
apresentam uma resposta antifúngica caracterizada pelo aumento da função
neutrofílica e aumento do recrutamento de macrófagos no espaço broncoalveolar.
Estes dados demonstram que o CCR4 possui um papel regulatório no recrutamento
e/ou ativação das células inflamatórias (SCHUH et al., 2002). Em um modelo
experimental de endotoxemia, animais deficientes para CCR4 que foram desafiados
com LPS são extremamente resistentes à mortalidade, exibem uma menor produção
de citocinas inflamatórias no local e um maior recrutamento de macrófagos
peritoneais (CHVATCHKO et al., 2000). Adicionalmente, esses animais deficientes
também são mais resistentes à mortalidade induzida por outros ligantes de TLR
(Toll-Like receptors) além do LPS (TLR4), como o Pam3Cys (TLR2) e CpG (TLR9).
Essa resistência está associada a um recrutamento mais rápido de leucócitos,

6
aumento da expressão de TLR, desvio no perfil de citocinas/quimiocinas para o tipo
Th1 e uma melhor eliminação da infecção local (NESS et al., 2006). Entretanto, um
dado inicialmente contraditório é que a administração do ligante CCL22 possui um
papel protetor e a inibição do CCL22 é prejudicial à sepse induzida por CLP (ligação
do ceco e perfuração) (ABRAHAM, 2005). Mas como o CCR4 possui mais de um
ligante, provavelmente o CCL17 é o mediador responsável pelos efeitos deletérios
observados. Em um modelo de sepse polimicrobiana, a ausência do CCR4 possui
um papel essencial na resistência a sepse e pode atuar na resposta imune inata
através da diferenciação de macrófagos alternativos em animais sépticos via TLR9
(ISHII et al., 2008).
Em outros modelos inflamatórios auto-imunes, como em modelo de lesão
cutânea por linfoma, há uma grande expressão do CCL17 pelas células endoteliais
nas lesões. Junto com as lesões, observa-se um aumento da expressão de E-
selectina e ICAM-1, indicando um microambiente favorável para a migração das
células T para o local inflamatório (FIVENSON & NICKOLOFF, 1992; UCCINI et al.,
1993).
O aumento da expressão do CCR4 e de seus ligantes está associado a
diversas patologias como fibrose pulmonar, inflamação hepática, desenvolvimento
de granulomas e diabetes (BELPERIO et al., 2004; FUJIMURA et al., 2004;
JAKUBZICK et al., 2004). Todas essas doenças apresentam infiltração de células T
expressando o CCR4.
A partir destes dados, pressupomos que o CCR4 possui um papel importante
na imunidade inata e adquirida em processos inflamatórios com a participação de
células T. O nosso grupo vem estudando o papel de mediadores envolvidos nas
doenças inflamatórias, dentre elas a sepse. Investigar o papel do CCR4 na evolução
da sepse até a imunossupressão pós-sepse foi o nosso objetivo, pois a perspectiva
de podermos modular a ação das quimiocinas em uma resposta inflamatória intensa,
torna-se um dos maiores incentivos para estudos básicos e clínicos nesta área.

7
1.2. SEPSE
A manifestação clínica da sepse é conseqüência de uma resposta imune
intensa do hospedeiro contra um patógeno, que pode se manifestar por hipotensão,
coagulopatia e disfunção dos órgãos (BONE, 1994). Durante uma infecção, o
organismo responde inicialmente de forma não específica, liberando mediadores
inflamatórios na tentativa de eliminar o agente agressor. Caso a resposta inata local
não elimine o agente infeccioso, ocorre a proliferação e disseminação do agente
patogênico com liberação exacerbada de mediadores inflamatórios locais e
sistêmicos, atingindo órgãos distais podendo culminar na falência múltipla de órgãos
e então, na morte (COHEN, 2002).
A definição da sepse, de acordo com DELINGER et al. 2008, é uma resposta
inflamatória sistêmica à infecção. Assim, quando o paciente séptico apresenta um
quadro clínico de hipoperfusão ou hipotensão tecidual e disfunção orgânica (p.e.
acidose lática, oligúria ou confusão mental) determina-se que evoluiu a uma sepse
grave. Além disso, o quadro pode evoluir até choque séptico, o qual se caracteriza
por um quadro de hipotensão arterial não revertida pela expansão volêmica
adequada e administração de vasoconstrictores.
A sepse apresenta altos índices de mortalidade nos hospitais do mundo todo.
No Brasil, o índice de mortalidade de sepse grave em hospitais públicos é
significativamente maior do que em hospitais de rede privada. Além disso, observa-
se uma alta taxa de mortalidade em hospitais do Brasil (56%) quando comparados a
de outros países em desenvolvimento como a Argentina (45%) e em países
desenvolvidos (33%) Este trabalho demonstrou que a alta taxa de mortalidade
acomete tanto países desenvolvidos como em países em desenvolvimento (TELES
et al., 2008).
Nos Estados Unidos o tempo médio de internação dos pacientes sépticos em
UTIs é de 20 dias o que acarreta em um alto custo, ultrapassando os US$ 16 bilhões
de dólares /ano. Ocorre cerca de 215.000 mortes/ano em pacientes internados por
sepse o que corresponde a 9,3% de todas as mortes (O'BRIEN et al., 2007).
No Brasil, um estudo multicêntrico com 2419 pacientes, realizado em 50
hospitais de todas as regiões do Brasil – SEPSE BRASIL – revelou que a incidência

8
de sepse, sepse grave e choque séptico nos pacientes internados em unidades de
terapia intensiva foi de 16,9% e a taxa de mortalidade de 15%, 35,6% e 48%,
respectivamente (SILVA et al., 2004). Outro estudo demonstrou uma incidência de
17,4% de pacientes com sepse grave entre os pacientes sépticos de UTI, com uma
taxa de mortalidade de 46,9%. Assim como nos Estados Unidos, a taxa de
mortalidade no Brasil cresce de acordo com a gravidade da doença. A taxa de
incidência dos graus de severidade da sepse é inversamente proporcional à
mortalidade (Figura 03) (SILVA et al., 2004).
Mortalidade
ChoqueSéptico14,6%
52,2%
46,9%Sepse grave17,4%
33,9%Sepse30%
Incidência
Figura 03. Relação entre incidência e mortalidade na sepse.
Adaptado de SILVA et al., 2004.
Apesar do crescimento do conhecimento em pesquisa básica, avanço
tecnológico e significativo incremento no arsenal terapêutico, não houve um
correspondente decréscimo na mortalidade relacionada à sepse. Isso ocorre
provavelmente devido à natureza invasiva de diversos procedimentos médicos, do
uso indiscriminado de antibióticos e aumento da vida média da população (TELES et
al., 2008). O nosso estudo atual possui uma grande relevância para o
desenvolvimento de novas terapias a fim de melhorarmos a sobrevida de pacientes.
Vários autores têm demonstrado a importância das células inflamatórias no
controle e na resolução das infecções (FERREIRA, 1980; DONG et al., 1993;

9
BENJAMIM; FERREIRA & CUNHA, 2000). Nesse contexto, FAURSCHOU &
BORREGAARD demonstraram que a depleção de neutrófilos, induzida por
quimioterápicos, está fortemente associada às infecções bacterianas e fúngicas de
alta letalidade (FAURSCHOU & BORREGAARD, 2003).
Um dos principais processos da resposta imune inata é a migração de
neutrófilos para o local de injúria. Os neutrófilos são as primeiras células recrutadas
para um sítio infeccioso, em resposta, principalmente a citocinas como IL-1 β, TNF-
α, mediadores lipídicos e quimiocinas. A migração é um processo regulado por um
amplo espectro de moléculas como citocinas e mediadores lipídicos, liberados pelas
células residentes e pelas células recrutadas ao foco infeccioso como mostra na
figura 04.
Esses mediadores também estão envolvidos na ativação leucocitária,
estimulando a fagocitose e mecanismos efetores da ação microbicida. Animais
deficientes de neutrófilos (VERDRENGH & TARKOWSKI, 1997) apresentaram sinais
clínicos graves de sepse e alta mortalidade na fase inicial da infecção por
Staphylococcus aereus, sugerindo que a fagocitose por neutrófilos é uma etapa
crítica para o controle da infecção.
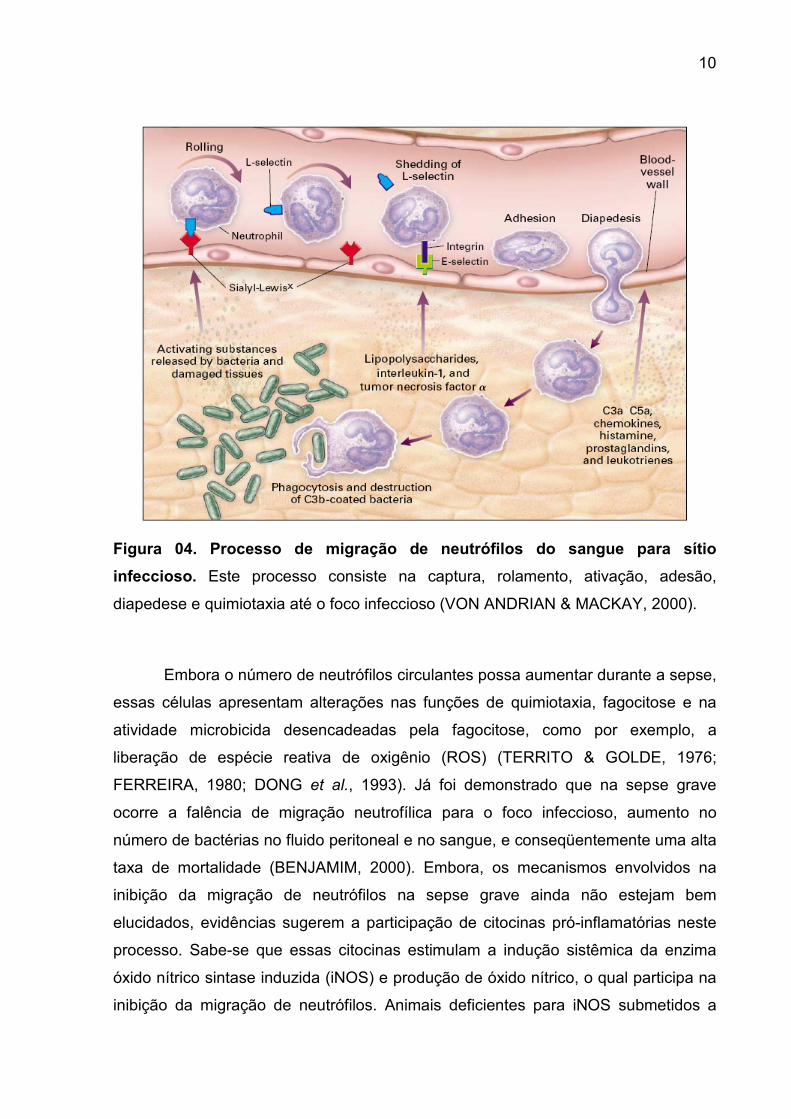
10
Figura 04. Processo de migração de neutrófilos do sangue para sítio
infeccioso. Este processo consiste na captura, rolamento, ativação, adesão,
diapedese e quimiotaxia até o foco infeccioso (VON ANDRIAN & MACKAY, 2000).
Embora o número de neutrófilos circulantes possa aumentar durante a sepse,
essas células apresentam alterações nas funções de quimiotaxia, fagocitose e na
atividade microbicida desencadeadas pela fagocitose, como por exemplo, a
liberação de espécie reativa de oxigênio (ROS) (TERRITO & GOLDE, 1976;
FERREIRA, 1980; DONG et al., 1993). Já foi demonstrado que na sepse grave
ocorre a falência de migração neutrofílica para o foco infeccioso, aumento no
número de bactérias no fluido peritoneal e no sangue, e conseqüentemente uma alta
taxa de mortalidade (BENJAMIM, 2000). Embora, os mecanismos envolvidos na
inibição da migração de neutrófilos na sepse grave ainda não estejam bem
elucidados, evidências sugerem a participação de citocinas pró-inflamatórias neste
processo. Sabe-se que essas citocinas estimulam a indução sistêmica da enzima
óxido nítrico sintase induzida (iNOS) e produção de óxido nítrico, o qual participa na
inibição da migração de neutrófilos. Animais deficientes para iNOS submetidos a

11
sepse grave não apresentaram inibição da migração de neutrófilos. Entretanto,
esses animais deficientes apresentaram alta mortalidade, o que sugere que o óxido
nítrico (NO) participa na falência de neutrófilos e na atividade microbicida dessas
células (BENJAMIM et al., 2002).
Os eventos fisiopatológicos na sepse podem ser subdivididos em uma fase
inicial e uma fase tardia. A fase inicial é caracterizada pela grande liberação de
mediadores pró-inflamatórios, incluindo TNF-α, IL-1β, IL-6 e quimiocinas, sendo
conhecida como Síndrome da Resposta Inflamatória Sistêmica [do inglês “Systemic
Inflammatory Response Syndrome”; SIRS]. A resposta pró-inflamatória é
compensada pela liberação de mediadores antiinflamatórios, como a IL-10 e TGF-β.
Esta fase tardia foi denominada de Síndrome da Resposta Anti-inflamatória
Compensatória [do inglês “Compensatory Anti-inflammatory Response Syndrome”;
CARS] (Figura 05) (DINARELLO & ABRAHAM, 2002).
Figura 05. Curva temporal da sepse até a imunossupressão.
Adaptado de BONE, 1994.
Este desequilíbrio de mediadores pró-inflamatórios com mediadores
regulatórios resulta em uma “tempestade” de citocinas. Essa liberação excessiva
dos mediadores antiinflamatórios e as alterações no processo reparativo são alguns
dos fatores responsáveis pelo desenvolvimento do quadro de imunossupressão, pois
ocorrem diversas alterações funcionais duradouras no sistema imune. Diversos
mecanismos celulares e moleculares já foram descritos para explicar a
IMUNOSSUPRESSÃO
Resposta Anti-inflamatória
Resposta Pró-inflamatória
Recuperação

12
imunossupressão pós-sepse. Entre eles estão o aumento de células T regulatórias,
a diminuição da produção de citocinas pró-inflamatórias, o aumento da expressão de
moléculas inibitórias intracelulares da sinalização da via de TLR (como por exemplo
a IRAK-M), a redução da expressão de HLA-DR, o grande número de células
apoptóticas, além de fatores epigenéticos (WANG & DENG, 2008). Essas alterações
podem prejudicar a resposta imune por desativação ou por falta de células capazes
de disparar uma resposta Th1 eficiente. Em conseqüência, o paciente séptico pode
apresentar incapacidade de erradicar a infecção primária causadora da sepse, e,
além disso, ficar mais propenso a adquirir infecções nosocomiais secundárias
(WANG & DENG, 2008).
A imunossupressão seguida de uma inflamação aguda grave não é exclusiva
da sepse, também é observada em outras doenças, incluindo a isquemia e
reperfusão por órgão transplantado (NAGANO & TILNEY, 1997), alterações na
recuperação de infecções por vírus sincicial respiratório (OPENSHAW; DEAN &
CULLEY, 2003), após queimaduras e traumas intensos (RODGERS et al., 2000;
KOBAYASHI et al., 2002) e pancreatites (NAGANO & TILNEY, 1997; RODGERS et
al., 2000; OPENSHAW; DEAN & CULLEY, 2003).
Dados na literatura demonstram que pacientes que se recuperam de um
quadro séptico grave apresentam alto índice de mortalidade no período de um ano.
Nestes trabalhos, 80% dos pacientes acometidos de sepse grave que obtiveram
alta, faleceram em um período de 8 anos por causas não sépticas, mas devido a
doenças como câncer, problemas cardiovasculares e pulmonares. Foi observado
que a vida-média do paciente séptico correlaciona-se com a severidade da sepse
(PERL et al., 1995; QUARTIN et al., 1997).
1.2.1. Modelo experimental de sepse
Embora existam muitas controvérsias no campo da sepse, é bastante
uniforme o pensamento de que essa síndrome é extremamente complexa devido ao
envolvimento de muitos sistemas, tecidos, tipos celulares e mediadores
inflamatórios. Muitos modelos animais têm sido usados para estudar a fisiopatologia
da sepse (OSUCHOWSKI et al., 2006), com o objetivo de recapitular as alterações

13
observadas na sepse humana. Utilizando um modelo murino experimental de sepse,
a ligação e perfuração do ceco (CLP) (BAKER et al., 1983), neste modelo o ceco
dos animais são expostos para fora da cavidade peritoneal e na porção distal do
ceco é realizado uma ligadura no ceco a fim de formar uma bolsa com conteúdo
fecal. A intensidade do estímulo irá depender do número de perfurações e do calibre
da agulha utilizada no ceco. Um alto número de furos e um grande calibre da agulha
irão desencadear uma grande quantidade de conteúdo fecal na cavidade peritoneal
induzindo a uma intensa resposta inflamatória, e consequentemente, uma alta
mortalidade (BENJAMIM, 2000). Este modelo experimental é o que mais mimetiza,
tanto qualitativamente como quantitativamente, observações clínicas de peritonite
polimicrobiana (FINK & HEARD, 1990). Semelhante ao que se observa na clínica, a
morte do animal no modelo de CLP é causada pelos efeitos diretos das bactérias e
por uma intensa resposta pró-inflamatória sistêmica contra os patógenos seguida de
um quadro anti-inflamatório (BONE, 1994).
A partir deste modelo, o nosso grupo vem estudando como as alterações
celulares e moleculares ocorridas na sepse podem alterar a interação da resposta
imune inata e adquirida e como conseqüência, desenvolver uma imunossupressão.
Nesse contexto, durante o processo infeccioso, as citocinas e outros mediadores
poderão induzir respostas imunes efetoras de proteção ou de lesão ao hospedeiro,
que dependerão principalmente do padrão de citocinas liberadas, bem como sua
concentração no sítio infeccioso e na circulação. Embora o papel de diversos
mediadores e células estejam bem descritos, na sepse, não existe atualmente um
tratamento eficaz, pois não está esclarecido como todos esses mediadores agem e
quais os papeis que cada um deles desempenha (RIEDEMANN; GUO & WARD).
Uma das conseqüências da imunossupressão pós-sepse é que o organismo
se torna mais suscetível a infecções secundárias. Recentemente foi demonstrado
que 28,3% dos pacientes sépticos na população chinesa desenvolvem aspergilose
pulmonar invasiva (IPA). O pulmão é o principal órgão afetado e o Aspergillus sp. é o
segundo principal causador da infecção (XIE et al., 2008). Além do Aspergillus,
outros patógenos são importantes causas de infecção. Assim como observado em
animais, pacientes sépticos são mais suscetíveis a infecções nosocomiais no

14
pulmão, principalmente pela Pseudomonas aeruginosa (RICHARDSON et al., 1982;
MUSTARD et al., 1991).
O Aspergillus fumigatus (ASP) é um fungo que está comumente presente no
ar e se reproduz através da propagação de conídios que são liberados no ambiente.
Esses conídios possuem uma superfície hidrofóbica o que lhes permite sobreviver
no ar por um tempo prolongado. Para a maioria dos indivíduos, a inalação de
conídios é inócua. No trato respiratório, o hospedeiro é protegido pela imunidade
inata através da geração de um processo inflamatório capaz de eliminar esses
conídios. O ASP é um fungo oportunista que acomete as vias aéreas e que tornou-
se uma importante causa de morte em casos de infecções pulmonares. Nas três
últimas décadas, a incidência da mortalidade por sepse causada por fungos cresceu
mais de 200% em pacientes imunocomprometidos (MARTIN et al., 2003), sendo os
principais agentes a Candida albicans e o Aspergillus fumigatus. Em indivíduos
imunossuprimidos eles podem se desenvolver em hifas e invadir o tecido, causando
pneumonia grave e progressiva que pode se disseminar a outros órgãos (MEHRAD;
STRIETER & STANDIFORD, 1999). O diagnóstico é difícil e quando realizado
tardiamente, o tratamento não é eficaz, consequentemente, a mortalidade pode
chegar até 50% (MEERSSEMAN et al., 2007).
A primeira linha de defesa no pulmão contra o ASP é composta por
macrófagos alveolares e neutrófilos. Animais depletados destas células inflamatórias
são mais suscetíveis a infecções por fungo, pois não apresentam uma eliminação do
patógeno de forma eficaz (MEHRAD; MOORE & STANDIFORD, 2000). Os conídios
que chegam às vias aéreas são fagocitados e mortos por essas células através de
uma via fagocítica oxidase-dependente. Já foi demonstrado que animais tratados
com anti-CCL2, ou animais depletados de neutrófilos com o bloqueio de TNF-α, ou
animais deficientes de TLR2 são mais suscetíveis a desenvolverem IPA por não
eliminarem a infecção (MORRISON et al., 2003; BALLOY et al., 2005; BENJAMIM et
al., 2005).
BENJAMIM et al., utilizando o modelo do CLP, demonstraram que após a
instilação intrapulmonar de células dendríticas obtidas da medula óssea de
camundongos falso-operados houve proteção dos camundongos pós-sépticos
desafiados a uma infecção secundária por A. fumigatus. Portanto, o
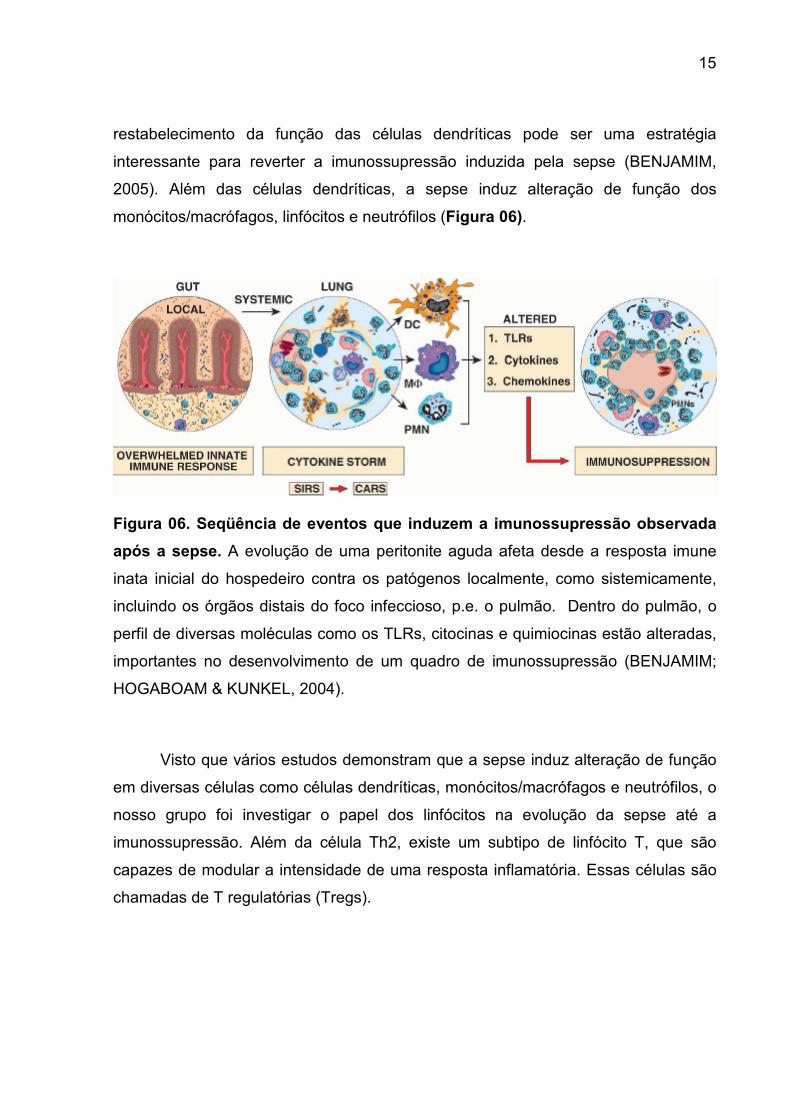
15
restabelecimento da função das células dendríticas pode ser uma estratégia
interessante para reverter a imunossupressão induzida pela sepse (BENJAMIM,
2005). Além das células dendríticas, a sepse induz alteração de função dos
monócitos/macrófagos, linfócitos e neutrófilos (Figura 06).
Figura 06. Seqüência de eventos que induzem a imunossupressão observada
após a sepse. A evolução de uma peritonite aguda afeta desde a resposta imune
inata inicial do hospedeiro contra os patógenos localmente, como sistemicamente,
incluindo os órgãos distais do foco infeccioso, p.e. o pulmão. Dentro do pulmão, o
perfil de diversas moléculas como os TLRs, citocinas e quimiocinas estão alteradas,
importantes no desenvolvimento de um quadro de imunossupressão (BENJAMIM;
HOGABOAM & KUNKEL, 2004).
Visto que vários estudos demonstram que a sepse induz alteração de função
em diversas células como células dendríticas, monócitos/macrófagos e neutrófilos, o
nosso grupo foi investigar o papel dos linfócitos na evolução da sepse até a
imunossupressão. Além da célula Th2, existe um subtipo de linfócito T, que são
capazes de modular a intensidade de uma resposta inflamatória. Essas células são
chamadas de T regulatórias (Tregs).

16
1.3. CÉLULAS T REGULATÓRIAS (Tregs)
Em uma resposta imune existe uma população de linfócitos T CD4+CD25+
que pode agir como células efetoras ou supressoras da resposta imune
apresentando um perfil de citocinas com características distintas das células Th1 ou
Th2. Estas células Tregs expressam na superfície o CD4 e o CD25, e
exclusivamente o fator de transcrição Foxp3 no citoplasma, sendo este considerado
atualmente o melhor marcador para células Tregs naturais. As células Tregs são
produzidas desde o terceiro dia de vida dos camundongos no timo, estão presentes
em 5-10% dos CD4 periféricos e podem inibir a ativação de diversas células
efetoras. As Tregs são capazes de inibir desde uma resposta imune inata, como por
exemplo a migração de neutrófilos até uma resposta imune adaptativa através da
inibição da proliferação de células T (Figura 07).
Tregs
Figura 07. Alvos das células Tregs.
(Fonte: Laboratório de Inflamação e Câncer – UFRJ; 2008)

17
O mecanismo de supressão das células Tregs ocorre por diversas vias,
incluindo a secreção local de citocinas como IL-10 e TGF-β, a competição pela IL-2
impedindo a proliferação de célula T efetora, ou por contato célula – célula, através
da ligação de moléculas de superfície das células regulatórias e efetoras, como o
CTLA-4 e CD80/CD86, respectivamente (VON BOEHMER, 2005). Recentemente
foram descritos novos mecanismos de supressão pelas células Tregs, por exemplo,
através da liberação de adenosina e o cAMP (DEAGLIO et al., 2007). As células
Tregs podem agir de forma sistêmica ou local. A sua presença em tecidos
especializados como fígado e olhos é capaz de controlar imunopatologias e proteger
estes órgãos (SUVAS et al., 2004).
Durante uma infecção aguda, as células Tregs naturais devem contribuir no
controle da resposta inflamatória. Conforme o processo inflamatório evolui e torna-se
crônico, com a participação das células Th1 e/ou Th2, outros processos regulatórios
participam modulando a resposta. No inicio da infecção, as células Tregs naturais
evitam um excessivo dano tecidual. Conforme a infecção evolui a um processo
crônico, independente da intensidade do processo inflamatório, são formadas novas
células Tregs Foxp3+ convertidas que junto com as células Tregs antígenos-
específicos acumulam-se no sítio de infecção (Figura 08)(BELKAID, 2007). As
células Tregs no local recrutam e formam mais células supressoras com um papel
regulatório importante no desenvolvimento do processo crônico. E esse acúmulo de
células também pode permanecer após a instalação da fase crônica da infecção.

18
Figura 08. Envolvimento das Tregs em diferentes estágios patológicos.
(BELKAID, 2007).
Trabalhos recentes têm demonstrado que essas células proliferam frente a
antígenos próprios como também a antígenos expressos por micróbios, como por
exemplo, Leishmania major, Helicobacter hepaticus, Listeria monocytogenesis e
Candida albicans (WALKER, 2004). Durante a infecção de camundongos com
Candida albicans, reduções de células Tregs naturais causam um melhor controle
da infecção, mas em contrapartida também causam aumento de uma inflamação
gastrointestinal (MONTAGNOLI et al., 2002).
Em 2004, um estudo demonstrou que após 28 dias do quadro infeccioso,
pacientes sépticos que não sobreviveram apresentavam maior porcentagem de
células Tregs no sangue do que os pacientes sépticos sobreviventes, além de
apresentarem menor expressão de HLA-DR. No ano seguinte, o mesmo grupo
demonstrou que esse aumento pode ser explicado devido às células Tregs
CD4+CD25+ serem mais resistentes a apoptose durante o choque séptico
comparado as CD4+CD25- (MONNERET et al., 2003; VENET et al., 2004). Em um
modelo murino de CLP foi observado um aumento de células CD4+CD25+ esplênicas
quando comparados aos animais Sham-operados. Observou-se também que as

19
células Tregs esplênicas de animais sépticos possuem maior capacidade supressora
e apresentam maior expressão de IL-10 intracelular após serem estimuladas,
indicando que, além das células Tregs naturais, as células Tregs induzidas também
participam na sepse. O autor ainda demonstrou que o tratamento de animais
sépticos com anti-CD25 e anti-IL-10 não melhorou a sobrevida dos animais sépticos
(SCUMPIA et al., 2006). A transferência adotiva de células Tregs CD4+CD25+
estimuladas in vitro com anti-CD3 e anti-CD28 para animais submetidos ao CLP
melhorou a sobrevida e diminuiu a quantidade de unidades formadoras de colônia
(CFU) presente no sítio de infecção (HEUER et al., 2005).
Algumas quimiocinas estimulam a migração de ambas às células CD4+CD25+
e CD4+CD25-. As CD25+ expressam especificamente CCR4 e CCR8 (IELLEM et al.,
2001) e migram em resposta aos seus ligantes CCL17 e CCL1, respectivamente
(COLANTONIO et al., 2002). Em um estudo clínico observou-se que os 104
indivíduos com carcinoma de ovário apresentavam células Tregs preferencialmente
se movendo e acumulando nos tumores e ascites, mas raramente entrando nos
linfonodos drenantes e que esse recrutamento era mediado pela produção de
CCL22 pelas células tumorais e macrófagos residentes naquele microambiente
(CURIEL et al., 2004). Estudos realizados em camundongos mostram que ligantes
do CCR5 atraem preferencialmente as células Tregs CD25+, que de fato expressam
este receptor na superfície (BYSTRY et al., 2001). A falta do CCR5 preveniu
fracamente a letalidade por doenças de enxerto vs. hospedeiro (WYSOCKI et al.,
2005). Em modelo de infecção com Paracoccidioides brasiliensis, o CCR5 foi
importante para a migração das Tregs para o sítio de infecção. No local, essas Tregs
diminuem a resposta imune ao fungo favorecendo a formação tardia do granuloma
(CAVASSANI et al., 2006). Assim como o CCR5, o CCR7 é importante para as
células Tregs migrarem para os linfonodos. Estas células suprimem uma resposta de
células T no local e assim, evitam o desenvolvimento de uma doença inflamatória
intestinal (SCHNEIDER et al., 2007). Já o receptor CXCR4 é importante para reter
células Tregs na medula óssea (ZOU et al., 2004).
As células Tregs também são importantes na tolerância. O recrutamento das
células Tregs Foxp3+ para o enxerto de transplante cardíaco tratado com CD154
(CD40L) é dependente de CCR4. A presença das células Tregs no órgão

20
transplantado induz a tolerância (LEE et al., 2005). Bloquear o CCR4 e o CCR6
bloqueia parcialmente a migração das células T virgem e Tregs, entretanto o
bloqueio do CCL2, CCL5 e CXCR3 inibe a migração das células CD4+CD25- sem
inibir a migração das células Tregs. Esse dado sugere que as Tregs possuem um
perfil de receptores diferente das outras células, embora apresentem também
receptores em comum com outras células que direcionam a migração para sítios
inflamatórios (CHEN & BROMBERG, 2006). O CCR4 está expresso em 75% das
células Tregs periféricas em humanos, estas células já parecem estar capazes para
suprimir a proliferação de células CD8. Quando as células T CCR4+ foram
depletadas observou-se uma maior polarização Th1 de linfócitos T CD4 virgens e
um aumento na função das células CD8 contra antígenos tumorais (BAATAR et al.,
2007). Esses dados suportam a idéia de que as quimiocinas participam na função e
na migração das células Tregs.
As Tregs são fortes candidatas a desempenharem um papel decisivo na
imunossupressão seguida da sepse grave, visto que essas células possuem um
papel modulador na resposta inflamatória e essa alteração pode afetar a infecção e
o dano tecidual. Os mecanismos envolvidos na imunossupressão ainda estão pouco
esclarecidos, entretanto, dados obtidos em nosso laboratório sugerem que o CCR4
tem um papel importante sobre a disposição e função das células Tregs no
organismo durante a sepse e a imunossupressão.

21
2. OBJETIVOS
O objetivo do nosso estudo é caracterizar o papel do CCR4 sobre as células
Tregs em diferentes fases da sepse, inclusive na imunossupressão pós-sepse. A
partir deste objetivo, nos propomos a investigar se os animais CCR4-/- são mais
resistentes à sepse e como as células Tregs poderiam participar desta resistência.
2.1. Objetivos Específicos
1 – Caracterizar a resposta inflamatória local e sistêmica dos animais WT e
CCR4-/- na sepse grave ou moderada através da migração celular, quantificação de
citocinas e mieloperoxidase no tecido pulmonar.
2 – Caracterizar o papel do CCR4 na localização das células T regulatórias no
pulmão e nos principais tecidos linfóides de diferentes fases da sepse grave.
3. Avaliar a sobrevida dos animais C57BL/6 (WT) e os CCR4-/- após o CLP e
após o desafio com ASP.
4. Analisar a migração celular e a integridade pulmonar após o desafio com
ASP em animais WT e CCR4-/- após a sepse.
5. Analisar a capacidade supressora das células T regulatórias obtidas de
animais WT e CCR4-/- sépticos sobre a migração de células inflamatórias “in vivo” e
“in vitro”, e na inibição da produção de ROS pelos neutrófilos.

22
3. METODOLOGIA
3.1. Animais.
Camundongos selvagens C57BL/6 (WT), pesando entre 20-24g, de ambos os
sexos, foram gentilmente doados pelo biotério da Fundação Oswaldo Cruz
(FIOCRUZ) e comprados do biotério central da Universidade Federal Fluminense
(UFF). Os animais foram mantidos sob condições controladas com ciclo de
claro/escuro e acesso livre a ração e água. Os camundongos deficientes
homozigotos para o receptor CCR4 (CCR4-/-) com fundo genético dos WT, também
foram mantidos nas mesmas condições no biotério da FIOCRUZ.
Os procedimentos com os animais de experimentação foram realizados de
acordo com as diretrizes da Comissão de Uso de Animais (CEUA) do centro de
ciências da saúde (CCS) da UFRJ, conforme o protocolo n° DFBCICB 028.
3.2. Modelo de ligação do ceco e perfuração (CLP).
Para avaliar a sepse grave e a imunossupressão, animais WT e CCR4-/-
foram previamente anestesiados intraperitonealmente com cetamina 112,5 mg/kg e
xilazina 10 mg/kg. Após estarem anestesiados, a cavidade abdominal foi aberta com
uma incisão de 1,5 cm, o ceco foi identificado, exposto para fora da cavidade e
ligado com linha de algodão (4-0), de forma a não obstruir a passagem entre íleo e
ceco. Em seguida, o ceco foi perfurado nove ou três vezes com uma agulha de 21G
para induzir a sepse grave (grupo CLP ou em alguns experimentos foi designado
como grupo L) ou uma sepse moderada (grupo SL), respectivamente. O ceco foi
recolocado na cavidade abdominal que foi fechada com auxílio de grampo cirúrgico
9 mm. Após o procedimento cirúrgico, os animais receberam a administração
subcutânea de solução salina (1,0 mL), para reposição volêmica. Os animais
controles ou falso operados (grupo Sham) foram abertos e o ceco exposto, mas não
foi ligado e perfurado (BAKER et al., 1983).

23
Os animais submetidos ao CLP ou Sham receberam tratamento com
antibióticos (Ertapenem, Merck) que foi preparado no dia do uso em solução salina
estéril na concentração de 75 mg/kg. O antibiótico foi administrado
intraperitonealmente nos animais CLP e Sham após 6, 24 e 48 horas da cirurgia.
Os animais foram acondicionados em caixas apropriadas para a observação
diária da evolução da sepse, durante o período de sete dias (Figura 09). No gráfico,
o dia da cirurgia foi considerado o dia zero.
Desenho Experimental:
Figura 09. Sobrevida ao CLP.
3.3. Avaliação da resposta imune inata em animais WT e CCR4-/- depois
de 6 horas de submetidos a uma sepse grave e moderada.
Animais WT e CCR4-/- foram submetidos a uma sepse moderada (SL) ou
grave (L). Como controles, os animais foram falso operado (Sham). Os animais
foram eutanaziados após 6 horas da cirurgia pela administração de uma injeção letal
de hidrato de cloral (800 mg/kg via i.p) (Figura 10).
Para avaliarmos a resposta imune local, a cavidade peritoneal foi lavada com
2 mL de PBS gelado e o volume foi recolhido. A partir dos exsudatos coletados
foram realizados diversos ensaios como: contagem total e diferencial de leucócitos,
a quantificação de citocinas e de CFU.
Para avaliarmos a resposta imune sistêmica, o sangue foi coletado por
punção cardíaca com auxílio de seringa contendo anticoagulante (EDTA 5% em

24
PBS, pH 7,4). A partir de uma amostra do sangue quantificamos citocinas (TNF-α e
IL-10) e CFU. Além do sangue, o pulmão foi perfundido pelo coração com 10 mL de
salina 0,9% e então, foi guardado no nitrogênio líquido até o processamento da
amostra.
Desenho experimental
Figura 10. Avaliação da resposta imune inata.
3.4. Contagem total de leucócitos.
A partir do exsudato recuperado o número total de leucócitos foi contado
microscopicamente utilizando hemocitômetro em solução de turk (acido acético 2%,
cristal violeta 0,01% em PBS).
3.5. Contagem diferencial de leucócitos.
Uma parte do exsudato foi centrifugada a 450 RPM durante 5 minutos em
citocentrífuga (Hettich Universal 16R). As células foram fixadas com metanol por 5
minutos e coradas por hematoxilina-eosina (Bio-Cor). As células foram examinadas
em microscópio óptico, sendo contadas 100 células por lâmina, as quais foram
diferenciadas em neutrófilos, eosinófilos e células mononucleares. A quantidade de
cada tipo celular presente na cavidade foi calculada pela porcentagem dessas
células contadas em relação ao valor total obtido. O valor obtido para 1 ml foi

25
multiplicado pelo volume de tampão utilizado para lavar a cavidade. Os resultados
foram expressos como n° de células por cavidade (nº de células/cavidade).
3.6. Quantificação de bactérias no sangue e exsudato peritoneal.
Após 6 horas da cirurgia, o sangue e o lavado peritoneal dos animais WT e
CCR4-/- submetidos à sepse grave ou moderada foram coletados como descrito
anteriormente. Os exsudatos do peritônio obtidos de animais submetidos à sepse
moderada (SL) foram diluídos 100 vezes em PBS e dos animais submetidos à sepse
grave (L) foram de 10000 vezes. A partir desta diluição, 10 µL foram semeados em
placas de petri contendo meio Ágar Mueller Hinton. Amostras de 10 µL de sangue do
grupo SL foram semeadas sem diluições, e no grupo L as amostras foram diluídas
10 e 100 vezes. Todo procedimento foi realizado sob condição estéril. Após as
semeaduras, as placas foram incubadas por 18h a 37°C e o número de unidades
formadoras de colônias (CFU) foram contadas e calculadas.
3.7. Quantificação de citocinas.
Uma parte do lavado peritoneal coletado 6 horas após a cirurgia foi utilizado
para a quantificação de TNF-α, IL-10, e MIP-2. O volume recolhido foi centrifugado e
o sobrenadante isento de células foi guardado em freezer à – 20°C até o dia da
dosagem. Para obtenção do plasma, o sangue foi coletado com auxílio de seringa
contendo anticoagulante por punção cardíaca (EDTA 5% em PBS, pH 7,4). As
amostras foram centrifugadas e o plasma foi transferido e guardado junto com o
exsudato à – 20°C até o dia da dosagem. A quantificação das citocinas foi realizada
através do método imunoenzimático (ELISA) descrito a seguir:
Placas de 96 poços foram cobertas com anticorpo específico (Pharmigen, San
Diego, CA,USA) diluídos em PBS e incubados por 18 horas a 4°C. As placas então
foram lavadas três vezes com PBS + 0,05% Tween20 (PBS-T20) (Sigma). As
ligações não específicas foram bloqueadas com PBS + 1% BSA por 2 horas a
temperatura ambiente. Os padrões das citocinas e as amostras foram adicionados
as placas e incubadas por 24 horas a 4°C. Então, as placas foram lavadas com
PBS-T20 e foram adicionados os anticorpos biotinilados específicos para cada

26
citocina. Após 1 hora, as placas foram lavadas com PBS-T20, o conjugado avidina-
peroxidase foi adicionado em cada poço e incubados por mais 30 minutos. As placas
foram lavadas com PBS-T20 e incubadas com o substrato OPD (o-fenilenediamina-
dihidrocloreto, Sigma) em tampão citrato pH 5,0. Após 20 minutos, a reação foi
interrompida com HCl 6 N e a densidade óptica quantificada em espectrofotômetro a
490nm (Spectra Max-250, Molecular Devices).
3.8. Ensaio da atividade da enzima mieloperoxidase (MPO).
Os pulmões retirados dos animais foram utilizados para dosagem de
mieloperoxidase. O pulmão foi colocado em tampão gelado (NaCl 0,1 M, Na2PO4
0,02 M, EDTA 12 mM pH 4,7) e homogeneizado em pollytron. O homogenato foi em
seguida centrifugado a 3000 RPM por 15 minutos a 4°C. Os eritrócitos foram
retirados com solução de lise ACK (NH4Cl 0,15 M, KHCO3 1 mM e EDTA 0,1 mM).
As células foram ressuspensas em uma solução de Na2PO4 contendo 0,5% de
brometo de hexadeciltrimetilamônio (HTAB). As células foram rompidas por
congelamento/aquecimento. O lisado foi centrifugado a 10.000g por 15 minutos.
Para reação da mieloperoxidase, 50 µL do sobrenadante do extrato celular foi
incubado com TMB (3,3,3,3–tetrametilbenzidina) e com H2O2 por 5 minutos. A
reação foi interrompida com H2SO4 2 N. A quantificação de neutrófilos foi feita a
partir da curva padrão de neutrófilos (obtidos de animais injetados com carragenina
e diluídos de forma seriada ½ na concentração inicial de 2x105 células/ poço). A
densidade óptica das amostras seriadas foi quantificada em espectrofotômetro a 450
nm (Spectra Max-250, Molecular Devices).
3.9. Esfregaço sanguíneo e diferencial de medula óssea.
Para avaliarmos a porcentagem de neutrófilos na médula óssea retiramos o
conteúdo da medula óssea de um fêmur através da lavagem com PBS gelado. As
amostras de medula dos animais normal WT e CCR4-/- foram analisadas pela
contagem total e diferencial, respectivamente. No esfregaço sanguíneo, uma gota de
sangue foi espalhada numa lâmina de vidro e corada com H&E. Em ambas as

27
amostras, contamos 100 células, diferenciando-os em neutrófilos ou mononucleares.
No gráfico expressamos apenas a contagem total e diferencial dos neutrófilos
presentes por mL de sangue ou por fêmur na medula óssea. Os dados foram
expressos em média ± EPM.
3.10. Análise temporal das células Tregs.
Animais WT e CCR4-/- foram eutanaziados 1 ou 5 dias após serem
submetidos ao CLP ou Sham operados (Figura 11).
O baço e o linfonodo mesentérico foram coletados separadamente e
macerados numa placa de 6 poços (Nunc, Dinamarca) contendo 4 mL de PBS. A
suspensão celular foi centrifugada a 400g por 5 minutos. Os eritrócitos foram
retirados com solução de lise ACK. O número de células foi determinado por
hemocitômetro e ajustado para 106 células/ mL. A suspensão celular foi fixada em
paraformaldeído 1% (Sigma) por 15 minutos à 4°C. As células foram lavadas 3
vezes com tampão de fluxo (1% SFB; 0,01% NaN3 em PBS) e guardadas à 4°C no
escuro.
O sangue foi coletado por punção cardíaca com auxílio de seringa contendo
anticoagulante (EDTA 5% em PBS, pH 7,4) e foi separado para marcação de
antígenos celulares por citometria de fluxo.
Desenho Experimental:

28
Figura 11. Análise temporal das células Tregs.
3.11. Citometria de Fluxo.
A caracterização fenotípica das células foi realizada através da marcação com
anticorpos monoclonais conjugado a flouróforos para moléculas presentes na
superfície celular e intracelular. Para marcação da superfície, 100 µL de amostra do
sangue ou 106 células do baço ou linfonodo mesentério foram incubadas com anti-
CD16/CD32 (B&D Pharmingen) para bloqueio do receptor Fc por 15 minutos a 4°C e
depois com anti-CD4-PerCP por 30 minutos a 4°C no escuro (B&D Pharmingen). A
permeabilização foi feita por 5 minutos com 0,1% de saponina em tampão de fluxo
para marcação intracelular com anti-Foxp3-FITC e incubado por 30 minutos a 4°C
no escuro. Lavamos três vezes para retirar o excesso de marcadores com tampão
de fluxo. As células foram analisadas no FACSCalibur (CellQuestTM software;
Becton and Dickinson, Mountain View, CA) e os dados foram analisados no
programa FCS Express 3.00. Foram adquiridos 30.000 eventos, e no “dot plot” do
FSH/SSC a população de linfócito foi selecionada e analisada. Os gráficos
mostrados em “dot plot” (CD4 e Foxp3) representam uma amostra de cada grupo
(n=3-5) e o gráfico de barras a direita expressa à média ± EPM dos valores
individuais.
3.12. Cultivo de ASP.
Utilizamos como ferramenta de nossos estudos a cepa 13073 de A. fumigatus
(American Type Culture Collection, Manassas, VA), pois esta cepa tem sido usada
para induzir IPA em animais imunocomprometidos (STEINHAUSER et al., 1999;
BENJAMIM et al., 2003). O organismo foi cultivado em ágar Sabouraud (Becton
Dickinson, Cockeysville, MD) por 7–10 dias a 37°C. No dia do inóculo, a superfície
da placa foi lavada com 50 mL de PBS-Tween80 0.1% (PBS-t) (Vetec). A suspensão
de conídios foi filtrada usando gaze estéril para remover agregados celulares e hifas.
Em seguida, centrifugamos a 1000g por 15 minutos. A concentração foi determinada
em hemocitômetro.

29
3.13. Avaliação da imunossupressão pós-sepse.
Animais WT e CCR4-/- submetidos ao CLP ou Sham foram desafiados com
conídios de ASP após 4 dias da cirurgia. Após o desafio, os animais foram avaliados
para diversos parâmetros, como sobrevida à infecção secundária, migração celular
para foco infeccioso e histologia. Nestes experimentos, animais WT e CCR4-/- foram
desafiados intratraquealmente (i.t.) com 7,5x107 conídios de ASP em 30µL de salina
por animal, e os animais controles receberam apenas salina. Os animais foram
acondicionados em caixas apropriadas para a observação diária da sobrevida após
a sepse. O dia do desafio com o fungo foi considerado o dia zero e os animais foram
observados por mais sete dias. Para avaliar a migração celular após 14 e 48 horas
do desafio foi realizado o lavado broncoalveolar nestes animais. Em seguida, o lado
direito do pulmão foi congelado para histologia. (Figura 12).
Desenho Experimental:
Figura 12. Imunossupressão pós-sepse.

30
3.14. Lavado broncoalveolar (BAL).
Após o desafio, os animais foram eutanaziados, a traquéia foi exposta e na
cavidade toráxica foi feito um orifício. Expomos a traquéia e inserimos uma agulha
de 21G na traquéia por onde injetamos 1 mL de PBS. O lavado broncoalveolar (BAL)
foi recuperado e analisado quanto ao número total e diferencial de leucócitos,
conforme descrito.
3.15. Histologia.
Após 48 horas do desafio, os animais foram eutanaziados e o ventrículo
direito foi perfundido com 10 mL de salina 0,9% gelada para remoção do sangue. O
pulmão foi inflado com 100 µL de meio de inclusão OCT (Optimal Cutting
Temperature). O pulmão direito foi recolhido e congelado no meio de inclusão direto
no nitrogênio líquido por 10 minutos. O material foi cortado na espessura de 5 µm e
logo fixado com acetona gelada por 10 minutos, depois foi corado com hematoxilina
e eosina.
3.16. Purificação e marcação de células CD4+Foxp3+ no pulmão.
Após 4 dias da cirurgia os animais WT e CCR4-/- foram desafiados com ASP
ou salina, e após 3 dias (Figura 13) os animais foram eutanaziados e o ventrículo
direito foi perfundido com salina para remoção do sangue. O pulmão foi coletado e
digerido com colagenase tipo IV (Sigma) por 1 hora a 37°C. A suspensão celular foi
tratada com ACK para retirar eritrócitos. Para purificarmos as células CD4
pulmonares, as células foram ressuspensas na concentração celular de 107/ 90µl de
Tampão MACS (BSA 1%, 2 mM de EDTA em PBS) e incubadas com anti-CD4-PE
por 15 minutos. Adicionamos anti-PE conjugada a microesferas, incubamos por 10
minutos e lavamos. A seleção positiva para obtenção de linfócitos T CD4+ foi
realizada em coluna MS (Myltenic biotec, EUA). A partir destas células CD4, fixamos
e marcamos para Foxp3.

31
Desenho experimental
Figura 13. Isolamento e marcação de células T CD4+Foxp3+ pulmonares.
As células foram analisadas por citometria de fluxo e adquiridos 10.000
eventos. Os gráficos mostrados em “dot plot” (CD4 e Foxp3) representam uma
amostra de cada grupo com o seu valor porcentual. O gráfico de barras expressa a
média ± EPM.
3.17. Isolamento de células T regulatórias.
Para purificarmos as células T regulatórias, utilizamos o kit de separação de
células T regulatórias (Myltenic biotec, EUA). O baço e o linfonodo mesentérico
foram coletados dos animais. As células foram ressuspensas na concentração
celular de 108/ 400 µl de tampão MACS (BSA 1%, 2 mM de EDTA em PBS) e
incubadas com coquetel de anticorpos biotinilados para seleção negativa de CD4+
mais anti-CD25-PE por 15 minutos. Lavamos e incubamos com estreptavidina
conjugada a microesferas por 15 minutos. Novamente lavamos e isolamos as CD4+
através de seleção negativa em coluna LD num suporte magnético em que somente
as células CD4+ não marcadas fluem pela coluna magnética. As células CD4+ foram

32
quantificadas e ajustadas numa concentração de 107 células/ 90 µL de tampão
MACS. Adicionamos anti-PE conjugado a microesferas, incubamos por 10 minutos e
lavamos. A seleção positiva foi realizada em coluna MS permitindo a obtenção de
células CD4+CD25+ com pureza de 92-96%.
3.18. Inibição da migração de leucócitos para um foco infeccioso “in
vivo” via células T regulatórias.
Animais WT e CCR4-/- foram submetidos ao CLP ou Sham-operados. Após 4
dias, o baço e o linfonodo dos animais de cada grupo foram unidos em uma única
amostra e as células T regulatórias CD4+CD25+ foram obtidas de acordo com o kit
de separação de células regulatórias. O ASP foi cultivado, isolado e ajustado na
concentração de 5x107 conídios/ 30 µL.
Animais WT recipientes foram divididos em 4 diferentes grupos. Em cada
grupo, os animais (n = 5-6) receberam intratraquealmente (i.t) salina ou ASP na
presença ou ausência de células Tregs obtidas de animais sépticos WT ou CCR4-/-.
Os diferentes grupos estão descritos como mostra na Figura 14.
Desenho Experimental:
Figura 14. Inibição da migração celular pelas células Tregs “in vivo”.

33
Após 24 horas do desafio, eutanaziamos os animais e o BAL de cada animal
foi coletado. O número total e diferencial de leucócitos foi quantificado e os
distinguimos em células mononucleares ou neutrófilos. O gráfico foi expresso em
média ± EPM dos neutrófilos que migraram para a cavidade.
3.19. Purificação de neutrófilos de medula óssea.
Os fêmures e as tíbias foram retirados de animais WT. Em seguida, o
conteúdo da medula óssea foi lavado com PBS gelado. A suspensão celular foi
centrifugada em gradientes de 65% e 72% de Percoll (AmerSham Pharmacia) a
1200g por 32 minutos com aceleração e desaceleração lenta, a temperatura
ambiente. A banda retirada entre os dois gradientes corresponde aos neutrófilos. A
pureza encontrada é de 94 a 96% determinada morfologicamente e por citometria.
3.20. Inibição da migração de neutrófilos “in vitro” via células T
regulatórias.
Células Tregs foram purificadas, conforme descrito anteriormente, de animais
WT e CCR4-/- submetidos ao CLP ou Sham após 4 dias da cirurgia, e incubadas em
RPMI-1640 suplementado (2 mM de L-glutamina e 10% de SFB) por 18 horas a
37°C/ 5% CO2. As células foram usadas no ensaio de migração celular e o
sobrenadante destas células foi recolhido e guardado a – 20°C para o ensaio de
inibição de produção de espécies reativas de oxigênio (ROS) produzidas pelos
neutrófilos (Figura 15). Os neutrófilos foram purificados no dia do ensaio conforme
descrito acima.
As células Tregs e neutrófilos foram pré-incubadas na proporção de 1:39 por
2 horas a 37ºC e 5% CO2. Após a pré-incubação, a migração celular foi realizada em
placas de quimiotaxia de 24 poços transwell (Costar, cat# 3421) com poro de 5 µm
por 2 horas a 37ºC e 5% CO2. Nos poços inferiores foram adicionados 600 µL de
RPMI com 1% de SFB contendo ou não o estímulo para a migração de neutrófilos, 1
nM de LTB4 (Cayman). A suspensão celular foi ressuspensa em um volume de
2x105 células/ 200µL de RPMI + 1% de SFB e adicionada no poço superior. Após a
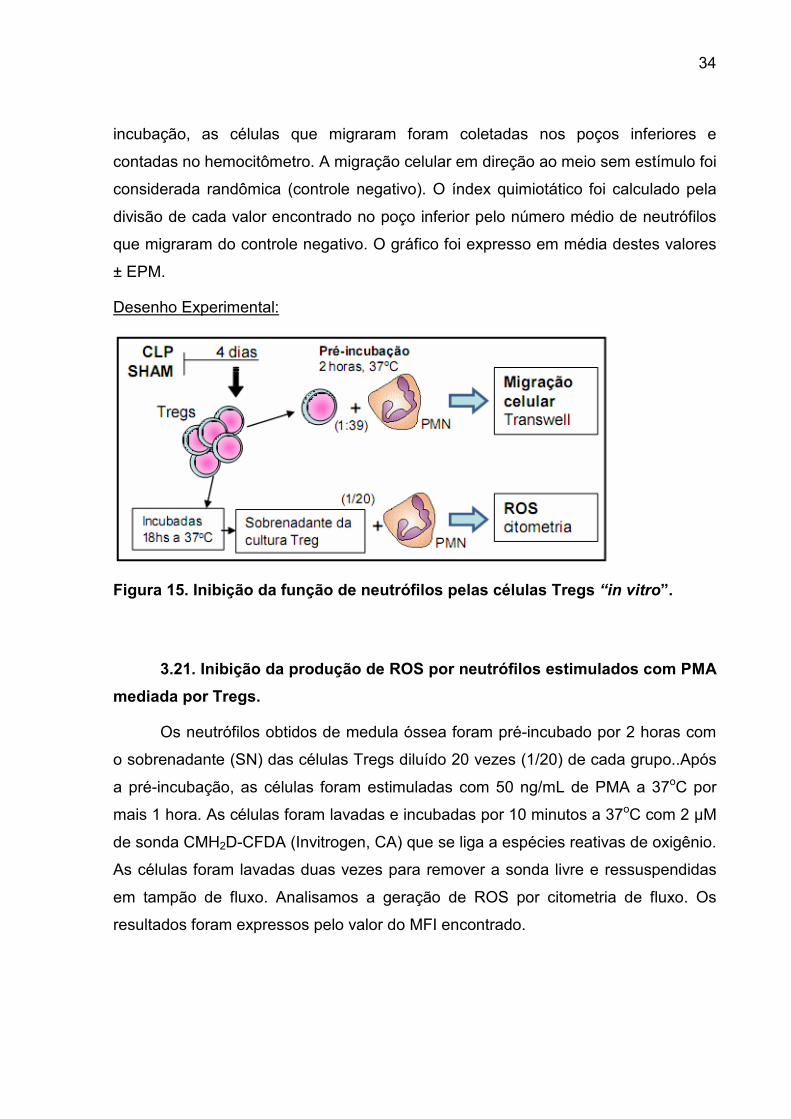
34
incubação, as células que migraram foram coletadas nos poços inferiores e
contadas no hemocitômetro. A migração celular em direção ao meio sem estímulo foi
considerada randômica (controle negativo). O índex quimiotático foi calculado pela
divisão de cada valor encontrado no poço inferior pelo número médio de neutrófilos
que migraram do controle negativo. O gráfico foi expresso em média destes valores
± EPM.
Desenho Experimental:
Figura 15. Inibição da função de neutrófilos pelas células Tregs “in vitro”.
3.21. Inibição da produção de ROS por neutrófilos estimulados com PMA
mediada por Tregs.
Os neutrófilos obtidos de medula óssea foram pré-incubado por 2 horas com
o sobrenadante (SN) das células Tregs diluído 20 vezes (1/20) de cada grupo..Após
a pré-incubação, as células foram estimuladas com 50 ng/mL de PMA a 37oC por
mais 1 hora. As células foram lavadas e incubadas por 10 minutos a 37oC com 2 µM
de sonda CMH2D-CFDA (Invitrogen, CA) que se liga a espécies reativas de oxigênio.
As células foram lavadas duas vezes para remover a sonda livre e ressuspendidas
em tampão de fluxo. Analisamos a geração de ROS por citometria de fluxo. Os
resultados foram expressos pelo valor do MFI encontrado.

35
3.22. Análise estatística.
Os resultados obtidos foram avaliados estatisticamente através dos testes
Mantel-Cox LogRank para curvas de sobrevivência. Os valores quantitativos foram
representados como média e erro padrão da média (EPM) e testados para testes
não-paramétricos Mann-Whitney. Foram consideradas diferenças significativas os
valores de p< 0,05.

36
4. RESULTADOS
4.1. Sobrevida de animais WT e CCR4-/- ao CLP.
Para investigar o papel do CCR4 na sepse grave, utilizamos o modelo já
estabelecido de sepse polimicrobiana em animais WT por BENJAMIM et al., 2003.
Todos os animais submetidos a este modelo de sepse grave experimental que não
foram tratados com antibióticos após a cirurgia morrem, enquanto os animais do
grupo Sham sobrevivem 100% (BENJAMIM et al., 2003). Com a finalidade de obter
um modelo mais próximo da clínica, os animais WT e CCR4-/- submetidos ao CLP
ou Sham foram tratados após 6, 24 e 48 horas da cirurgia com antibioticoterapia. A
sobrevida dos animais WT e CCR4-/- com sepse grave foi acompanhada por até 7
dias após a cirurgia. Após 6 horas da cirurgia, os animais WT do grupo CLP já
apresentavam um quadro semelhante ao choque endotóxico em que os olhos
produzem secreções, piloereção, letargia e taquipnéia. Esses sinais não foram
observados nos animais CCR4-/-. Os animais do grupo Sham apresentaram 100%
de sobrevivência. O grupo CLP do CCR4-/- apresentou 92% de sobrevivência e não
foi diferente estatisticamente dos grupos Sham de ambas as linhagens (WT e CCR4-
/-). E os animais do grupo CLP dos WT apresentaram 48% de sobrevivência quando
comparada aos outros grupos. O grupo CLP do WT apresentou mortalidade até o 3°
dia enquanto o grupo CLP do CCR4-/- a mortalidade foi observada apenas nas
primeiras 24 horas após a cirurgia (Figura 16). Estes resultados indicam que os
animais CCR4-/- são mais resistentes à sepse grave induzida por CLP comparado
ao WT.
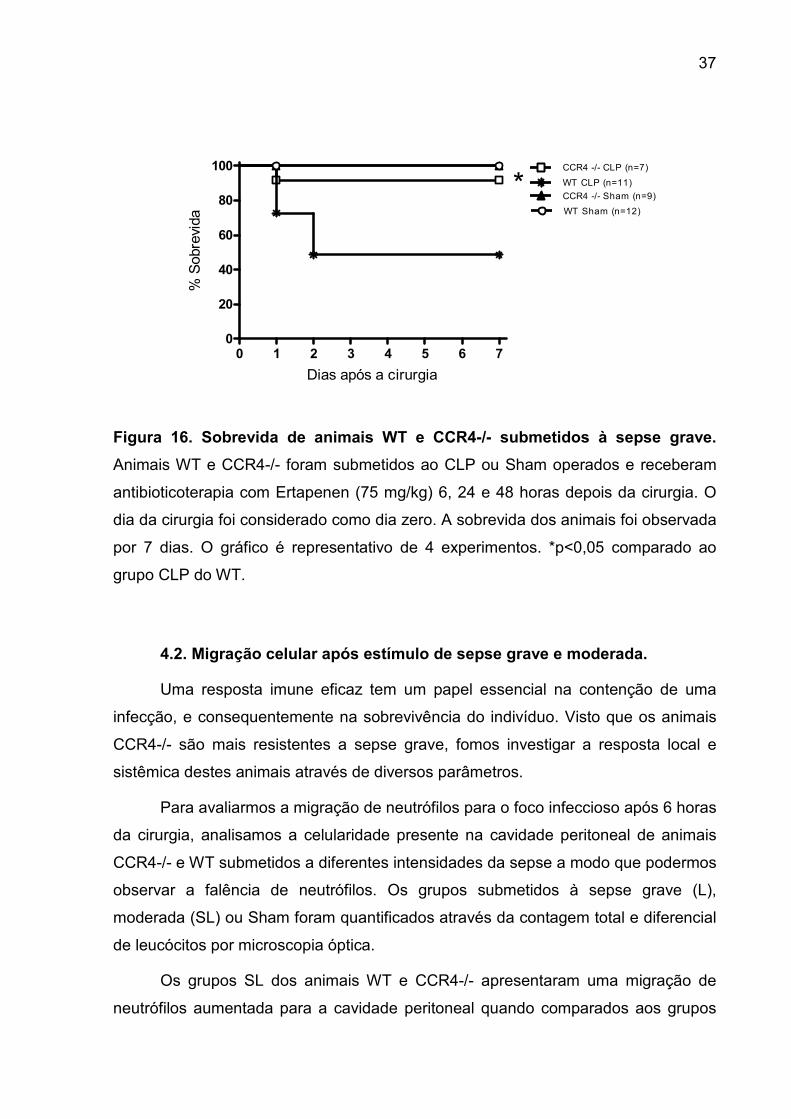
37
0 1 2 3 4 5 6 70
20
40
60
80
100
*
Dias após a cirurgia
% S
obre
vida
CCR4 -/- CLP (n=7)
WT CLP (n=11)
CCR4 -/- Sham (n=9)
WT Sham (n=12)
Figura 16. Sobrevida de animais WT e CCR4-/- submetidos à sepse grave.
Animais WT e CCR4-/- foram submetidos ao CLP ou Sham operados e receberam
antibioticoterapia com Ertapenen (75 mg/kg) 6, 24 e 48 horas depois da cirurgia. O
dia da cirurgia foi considerado como dia zero. A sobrevida dos animais foi observada
por 7 dias. O gráfico é representativo de 4 experimentos. *p<0,05 comparado ao
grupo CLP do WT.
4.2. Migração celular após estímulo de sepse grave e moderada.
Uma resposta imune eficaz tem um papel essencial na contenção de uma
infecção, e consequentemente na sobrevivência do indivíduo. Visto que os animais
CCR4-/- são mais resistentes a sepse grave, fomos investigar a resposta local e
sistêmica destes animais através de diversos parâmetros.
Para avaliarmos a migração de neutrófilos para o foco infeccioso após 6 horas
da cirurgia, analisamos a celularidade presente na cavidade peritoneal de animais
CCR4-/- e WT submetidos a diferentes intensidades da sepse a modo que podermos
observar a falência de neutrófilos. Os grupos submetidos à sepse grave (L),
moderada (SL) ou Sham foram quantificados através da contagem total e diferencial
de leucócitos por microscopia óptica.
Os grupos SL dos animais WT e CCR4-/- apresentaram uma migração de
neutrófilos aumentada para a cavidade peritoneal quando comparados aos grupos

38
Sham de ambas as linhagens. Corroborando com dados já publicados de nosso
grupo, o grupo L dos animais WT apresentaram falência na migração de neutrófilos
para a cavidade, quando comparado ao grupo SL dos animais WT. Entretanto, a
migração de neutrófilos para a cavidade no grupo L dos animais CCR4-/- foi
surpreendentemente elevada quando comparamos aos grupos L dos animais WT
(Figura 17). Em todos os grupos não observamos diferenças na quantidade de
células mononucleares entre os animais WT e CCR4-/- (dados não mostrados).
Estes resultados sugerem que os animais CCR4-/- submetidos ao modelo de sepse
grave não sofrem falência na migração de neutrófilos. O aumento de neutrófilos na
cavidade deve participar como um fator favorável na sobrevida destes animais
durante a sepse.
Sham SL L Sham SL L0
2
4
6
8
10
12
14
16
*#*
WT CCR4 -/-
**
( x1
06 n
eutr
ófilo
s /
cavi
dade
)
Figura 17. Migração de neutrófilos para a cavidade peritoneal durante a sepse.
Animais WT e CCR4-/- foram eutanaziados 6 horas depois de submetidos à sepse
grave (L= 9 furos, n=5), sepse moderada (SL= 3 furos, n= 5) e falsos operados
(Sham, n=3). A cavidade peritoneal foi lavada e do volume recuperado foram
realizadas a contagem dos neutrófilos presentes. Os resultados foram expressos
como média ± EPM. O gráfico é representativo de 2 experimentos. * p<0,05
comparados ao grupo Sham. ** p<0,05 comparado ao grupo SL do WT. # p<0,05
comparado ao grupo L do WT.

39
4.3. Avaliação do número de bactérias na cavidade peritoneal.
Com o intuito de caracterizar a resposta imune a infecção bacteriana nos animais
CCR4-/-, avaliamos a carga bacteriana na cavidade peritoneal e no sangue de
animais WT e CCR4-/- submetidos a sepse grave ou moderada. A quantificação foi
realizada através de amostras do exsudato e do sangue obtidos 6 horas após a
cirurgia. O grupo Sham foi excluído das analises por não apresentar nenhuma CFU
detectável no lavado peritoneal e nem no sangue (dados não mostrados). No lavado
peritoneal (Figura 18A) e no sangue (Figura 18B) do grupo L dos animais CCR4-/-
observamos redução do número de CFU quando comparado ao grupo L dos animais
WT. Quando induzimos um estímulo moderado (grupo SL) não observamos
diferenças entre as linhagens quanto ao número de bactérias presentes em ambos
locais. Estes resultados indicam que os animais CCR4-/- apresentam uma menor
carga bacteriana no peritônio e no sangue, sugerindo uma resposta imune mais
eficiente destes animais na sepse grave.

40
SL L SL L0
20
40
60
80
WT CCR4 -/-
*
**
x10
7 C
FU
/ ca
vid
ade
SL L SL L0
50
100
150
200
x105
CF
U/m
L d
e sa
ng
ue
**
WT CCR4 -/-
*
Figura 18. Quantificação do número de CFU no lavado peritoneal e sangue de
animais submetidos a sepse moderada (SL) ou a sepse grave (L).
(A) Os lavados peritoneais dos animais WT e CCR4-/- do grupo L (n=6) e do SL
(n=6) foram coletados após 6 horas da cirurgia e diluídos 100, 1000 e 10000 vezes.
(B) O sangue dos animais foi coletado por punção cardíaca e foi diluído 10 e 100
vezes. As diluições do lavado peritoneal e do sangue foram semeadas diretamente
em Ágar Mueller Hinton e incubadas por 24 horas a 37°C. As unidades formadoras
de colônias (CFU) foram contadas e os resultados foram expressos em valores
individuais e a mediana. * p<0,05 comparados ao grupo SL. ** p<0,05 comparado ao
grupo L do WT.
4.4. Quantificação de citocinas locais.
A partir do exsudato peritoneal recolhido após 6 horas da cirurgia de animais
WT e CCR4-/- submetidos ao Sham, SL ou L, algumas citocinas e quimiocinas que
são importantes no recrutamento e ativação de células inflamatórias, como o TNF-α,
IL-10 e o MIP-2, foram quantificadas. No exsudato de animais CCR4-/- do grupo L
A B
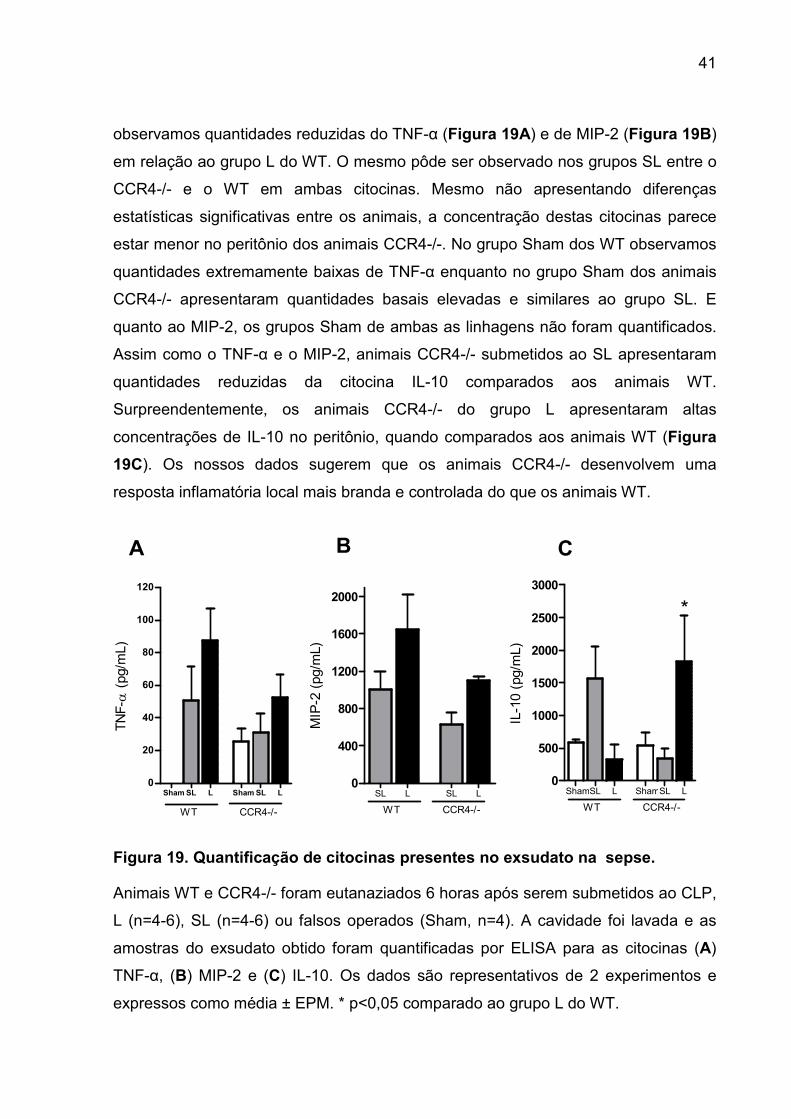
41
observamos quantidades reduzidas do TNF-α (Figura 19A) e de MIP-2 (Figura 19B)
em relação ao grupo L do WT. O mesmo pôde ser observado nos grupos SL entre o
CCR4-/- e o WT em ambas citocinas. Mesmo não apresentando diferenças
estatísticas significativas entre os animais, a concentração destas citocinas parece
estar menor no peritônio dos animais CCR4-/-. No grupo Sham dos WT observamos
quantidades extremamente baixas de TNF-α enquanto no grupo Sham dos animais
CCR4-/- apresentaram quantidades basais elevadas e similares ao grupo SL. E
quanto ao MIP-2, os grupos Sham de ambas as linhagens não foram quantificados.
Assim como o TNF-α e o MIP-2, animais CCR4-/- submetidos ao SL apresentaram
quantidades reduzidas da citocina IL-10 comparados aos animais WT.
Surpreendentemente, os animais CCR4-/- do grupo L apresentaram altas
concentrações de IL-10 no peritônio, quando comparados aos animais WT (Figura
19C). Os nossos dados sugerem que os animais CCR4-/- desenvolvem uma
resposta inflamatória local mais branda e controlada do que os animais WT.
Sham SL L Sham SL L0
20
40
60
80
100
120
WT CCR4-/-
TN
F- α
(pg
/mL)
ShamSL L ShamSL L0
500
1000
1500
2000
2500
3000
WT CCR4-/-
*
IL-1
0 (p
g/m
L)
SL L SL L0
400
800
1200
1600
2000
WT CCR4-/-
MIP
-2 (
pg/m
L)
A B C
Figura 19. Quantificação de citocinas presentes no exsudato na sepse.
Animais WT e CCR4-/- foram eutanaziados 6 horas após serem submetidos ao CLP,
L (n=4-6), SL (n=4-6) ou falsos operados (Sham, n=4). A cavidade foi lavada e as
amostras do exsudato obtido foram quantificadas por ELISA para as citocinas (A)
TNF-α, (B) MIP-2 e (C) IL-10. Os dados são representativos de 2 experimentos e
expressos como média ± EPM. * p<0,05 comparado ao grupo L do WT.

42
4.5. Determinação da atividade da mieloperoxidase no pulmão.
Para caracterizar se a inflamação evoluiu para um quadro sistêmico,
decorrente da infecção induzida pelo CLP, avaliamos a presença de infiltrados
neutrofílicos no pulmão de animais WT e CCR4-/- após 6 horas da cirurgia. Para
quantificarmos este infiltrado tecidual, determinamos o número de células presentes
no tecido pulmonar através da quantificação da atividade de uma enzima específica
nos neutrófilos, a mieloperoxidase (MPO). Os pulmões dos animais CCR4-/- e WT
foram macerados e tratados a fim de obtermos a enzima mieloperoxidase livre para
a quantificação de sua atividade. Nossos dados demonstram que o grupo L de
animais CCR4-/- possui um menor infiltrado de células, comparado aos animais WT
do grupo L. Essa diferença não foi observada nos grupos SL entre os animais WT e
CCR4-/- (Figura 20). Assim, os animais CCR4-/- submetidos à sepse grave (grupo
L) possuem uma reação inflamatória sistêmica mais branda do que nos animais WT
provavelmente devido a uma melhor resposta inflamatória local.
0
2
4
6
8
10
12
14
WT CCR4 -/-SL L SL L
*
**
Neu
tróf
ilos
(x 1
03ce
lúla
s / m
g de
pul
mão
)
Figura 20. Determinação do infiltrado neutrofílico pela atividade da MPO.
Animais WT e CCR4-/- foram eutanaziados após 6 horas de terem sido submetidos
a sepse grave (L; n=5) ou moderada (SL; n=5). O pulmão foi obtido para determinar
a atividade da enzima mieloperoxidase (MPO). ** p<0,01 comparado ao grupo L dos
animais WT.
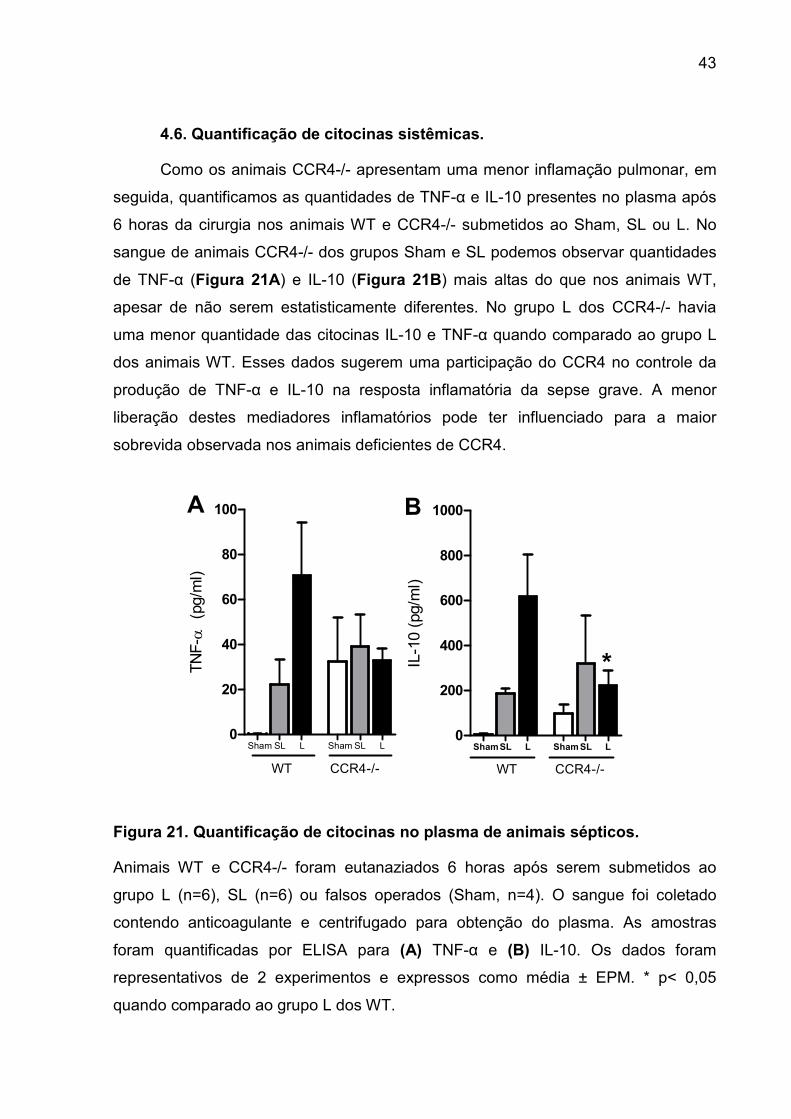
43
4.6. Quantificação de citocinas sistêmicas.
Como os animais CCR4-/- apresentam uma menor inflamação pulmonar, em
seguida, quantificamos as quantidades de TNF-α e IL-10 presentes no plasma após
6 horas da cirurgia nos animais WT e CCR4-/- submetidos ao Sham, SL ou L. No
sangue de animais CCR4-/- dos grupos Sham e SL podemos observar quantidades
de TNF-α (Figura 21A) e IL-10 (Figura 21B) mais altas do que nos animais WT,
apesar de não serem estatisticamente diferentes. No grupo L dos CCR4-/- havia
uma menor quantidade das citocinas IL-10 e TNF-α quando comparado ao grupo L
dos animais WT. Esses dados sugerem uma participação do CCR4 no controle da
produção de TNF-α e IL-10 na resposta inflamatória da sepse grave. A menor
liberação destes mediadores inflamatórios pode ter influenciado para a maior
sobrevida observada nos animais deficientes de CCR4.
Sham SL L Sham SL L0
20
40
60
80
100
WT CCR4-/-
TN
F- α
(p
g/m
l)
ShamSL L Sham SL L0
200
400
600
800
1000
WT CCR4-/-
*IL-1
0 (p
g/m
l)
A B
Figura 21. Quantificação de citocinas no plasma de animais sépticos.
Animais WT e CCR4-/- foram eutanaziados 6 horas após serem submetidos ao
grupo L (n=6), SL (n=6) ou falsos operados (Sham, n=4). O sangue foi coletado
contendo anticoagulante e centrifugado para obtenção do plasma. As amostras
foram quantificadas por ELISA para (A) TNF-α e (B) IL-10. Os dados foram
representativos de 2 experimentos e expressos como média ± EPM. * p< 0,05
quando comparado ao grupo L dos WT.

44
4.7. Avaliação do leucograma e celularidade da medula óssea.
Como controle dos nossos experimentos, avaliamos se os animais CCR4-/-
possuem diferenças na quantidade de neutrófilos presentes no sangue e na medula
óssea, podendo influenciar na migração celular e na contenção da infecção. Para tal,
foi feita a análise da celularidade do sangue e da medula óssea por microscopia
óptica. A porcentagem e o número de neutrófilos observados entre os animais
CCR4-/- e WT não apresentaram diferença no esfregaço de sangue (Figura 22A) e
nem na medula óssea (Figura 22B). Visto que não houve diferenças na quantidade
de neutrófilos entre os animais CCR4-/- e WT podemos sugerir que o aumento da
migração de células para o foco infeccioso após 6 horas da cirurgia nos CCR4-/- não
se deve a alterações na produção dos neutrófilos nestes animais.
WT CCR4 -/-0
10
20
30
40
% d
e N
eutr
ófi
los
WT CCR4 -/-0
1
2
3
4
5
6
7
8
Neu
tró
filo
s
(x10
6 c
élul
as /
fêm
ur )
A B
Figura 22. Leucograma e contagem de neutrófilos na medula óssea.
O sangue e a medula óssea de animais WT e CCR4-/- foram coletados. (A) As
células no esfregaço sanguíneo foram diferenciadas pela morfologia em
microscopia. (B) A medula óssea destes animais foram citocentrifugadas e coradas
para análise da contagem diferencial através da morfologia. Os dados foram
representativos de 2 experimentos e expresso como média ± EPM (n=3 / grupo).

45
4.8. Presença das células Tregs no sangue periférico e órgãos linfóides
durante a sepse grave.
Diversos modelos experimentais demonstram que a presença das células
Tregs (CD4+Foxp3+) no local pode regular a magnitude da resposta imune contra a
infecção. Dessa forma, fomos investigar o papel do CCR4 sobre a localização
destas células no sangue periférico e nos diferentes tecidos linfóides durante a
sepse. No sangue, observamos que as porcentagens de células Tregs do grupo
sepse grave CCR4-/- estavam elevadas em relação aos outros grupos (Figura 23A).
Este aumento foi de 45% nas porcentagens de células Tregs no sangue entre os
grupos CLP do CCR4-/- e os WT. Os animais WT submetidos à sepse grave (CLP)
não apresentaram diferenças entre os grupos CLP e Sham, diferente do observado
nos animais CCR4-/-.
No baço, o aumento na porcentagem de células Tregs entre os grupos CLP
dos animais CCR4-/- e WT foi de 70% (Figura 23B). O linfonodo não apresentou
diferenças nas porcentagens de células Tregs entre os grupos CLP do CCR4-/- e
WT (apenas uma discreta redução de 4%). Mas quando comparamos entre os
grupos CLP e Sham, os animais CCR4-/- apresentaram menor aumento do que
observado nos animais WT (Figura 23C). Os aumentos observados no baço e no
sangue, mas não no linfonodo, dos animais CCR4-/- do grupo CLP em relação aos
WT podem estar contribuindo para um controle de uma resposta inflamatória
sistêmica exarcebada e conseqüentemente, favorecendo a sobrevida.
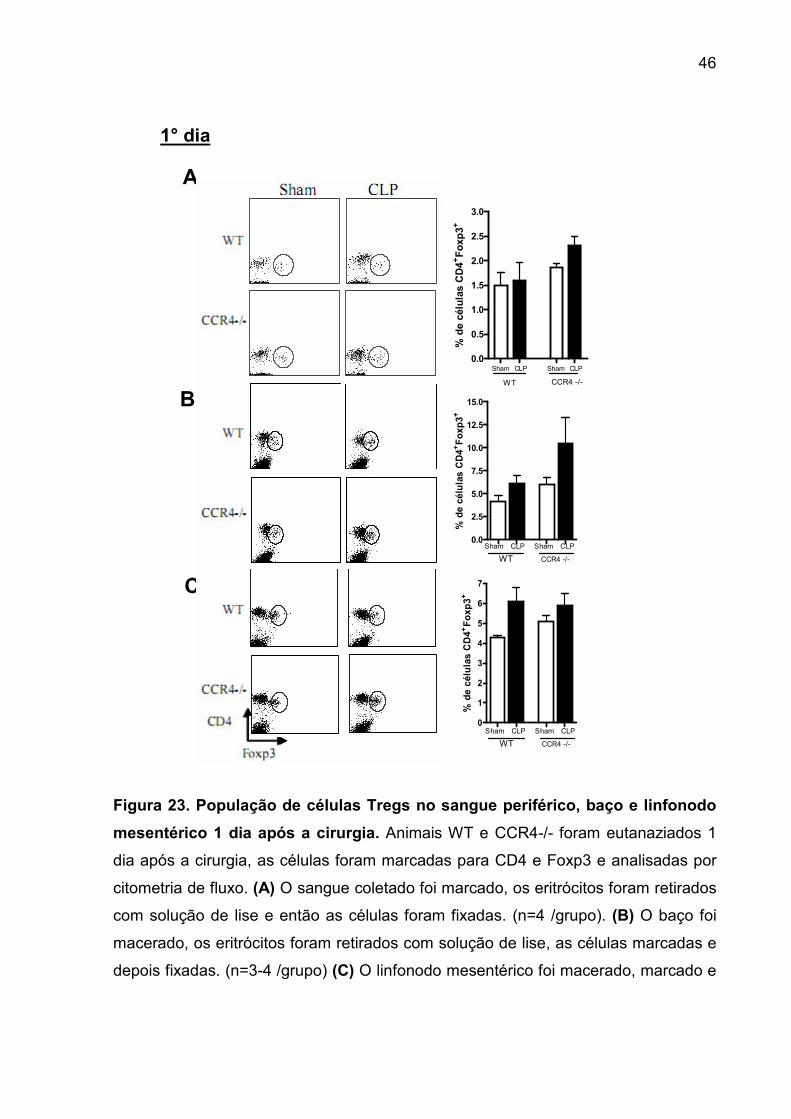
46
1° dia
A
B
C
Figura 23. População de células Tregs no sangue periférico, baço e linfonodo
mesentérico 1 dia após a cirurgia. Animais WT e CCR4-/- foram eutanaziados 1
dia após a cirurgia, as células foram marcadas para CD4 e Foxp3 e analisadas por
citometria de fluxo. (A) O sangue coletado foi marcado, os eritrócitos foram retirados
com solução de lise e então as células foram fixadas. (n=4 /grupo). (B) O baço foi
macerado, os eritrócitos foram retirados com solução de lise, as células marcadas e
depois fixadas. (n=3-4 /grupo) (C) O linfonodo mesentérico foi macerado, marcado e
0.0
2.5
5.0
7.5
10.0
12.5
15.0
WT
Sham CLP Sham CLP
CCR4 -/-
% d
e cé
lula
s C
D4+
Fo
xp3+
0
1
2
3
4
5
6
7
WT
Sham CLP Sham CLP
CCR4 -/-
% d
e cé
lula
s C
D4+
Fo
xp3+
0.0
0.5
1.0
1.5
2.0
2.5
3.0
WT
Sham CLP Sham CLP
CCR4 -/-
% d
e cé
lula
s C
D4+
Fo
xp3+
C

47
fixado. (n=3-4 /grupo). O conjunto de dot plot (esquerda) representa uma amostra de
cada grupo e o gráfico (direita) foi expresso como média ± EPM.
4.9. Sobrevida dos animais WT e CCR4-/- submetidos ao CLP ou Sham
operados desafiados com conídios de ASP.
Com o intuito de caracterizar a imunossupressão pós-sepse grave,
desafiamos os animais CCR4-/- e WT após 4 dias do CLP ou Sham-operados com
7,5x107 conídios de ASP. O desafio com um patógeno é uma ferramenta importante
para demonstrar a susceptibilidade à infecção secundária. Após o desafio
(considerado como dia zero), a sobrevida foi acompanhada por 7 dias. Os animais
do grupo Sham do WT e CCR4-/- apresentaram 100% de sobrevivência nos três
primeiros dias após o desafio, mas apenas 22% dos animais do grupo Sham do
CCR4-/- morreram no quarto dia após o desafio. Os animais do grupo CLP dos WT
apresentaram uma redução significativa na sobrevivência, a sua taxa de sobrevida
foi de 34%. O grupo CLP dos CCR4-/- apresentou 90% de sobrevida e não foi
diferente dos grupos Sham de ambas as linhagens (Figura 24). Portanto, os animais
CCR4-/- submetidos a sepse grave possuem maior resistência frente a infecções
secundárias por conídios de ASP comparados aos animais WT pós-sépticos.

48
0 1 2 3 4 5 6 70
20
40
60
80
100
*
CCR4-/- Sham (n=9)
WT Sham (n=7)
WT CLP (n=9)
CCR4-/- CLP (n=11)
Dias após a infecção secundária
% s
obre
vida
Figura 24. Curva de sobrevida dos animais WT e CCR4-/- desafiados com ASP
depois de submetidos ao CLP ou Sham-operados. Animais WT e CCR4-/- foram
submetidos ao CLP ou Sham-operados e tratados com antibióticos 6, 24 e 48 horas
após a cirurgia. Quatro dias após a cirurgia, os animais foram desafiados i.t. com
7,5x107 conídios de ASP. A partir deste dia (dia 0), a mortalidade foi acompanhada
por 7 dias. * p< 0,01 comparados aos grupos Sham dos WT e CCR4-/- e ao grupo
CLP dos CCR4-/-.
4.10. Avaliação da migração celular para o pulmão frente a um estímulo
infeccioso.
Para caracterizarmos a resposta imune dos animais CCR4-/- frente à infecção
secundária, analisamos a migração celular para o espaço broncoalveolar e a
integridade do tecido pulmonar. A análise foi realizada 14 e/ou 48 horas após o
desafio com o fungo. Primeiramente, fomos avaliar a migração celular nos animais
após 14 horas (Figura 25A) e 48 horas (Figura 25B) do desafio com o fungo (barra
preta) ou após administração de salina como controle (barra branca). Após 14 e 48
horas do desafio, a migração de neutrófilos induzida pelo fungo não foi diferente
entre os grupos CLP dos animais WT e dos CCR4-/-, embora o grupo CLP dos
CCR4-/- tenha apresentado uma ligeira diminuição na migração de neutrófilos
quando comparado ao grupo CLP dos WT. Por outro lado, o grupo CLP dos animais
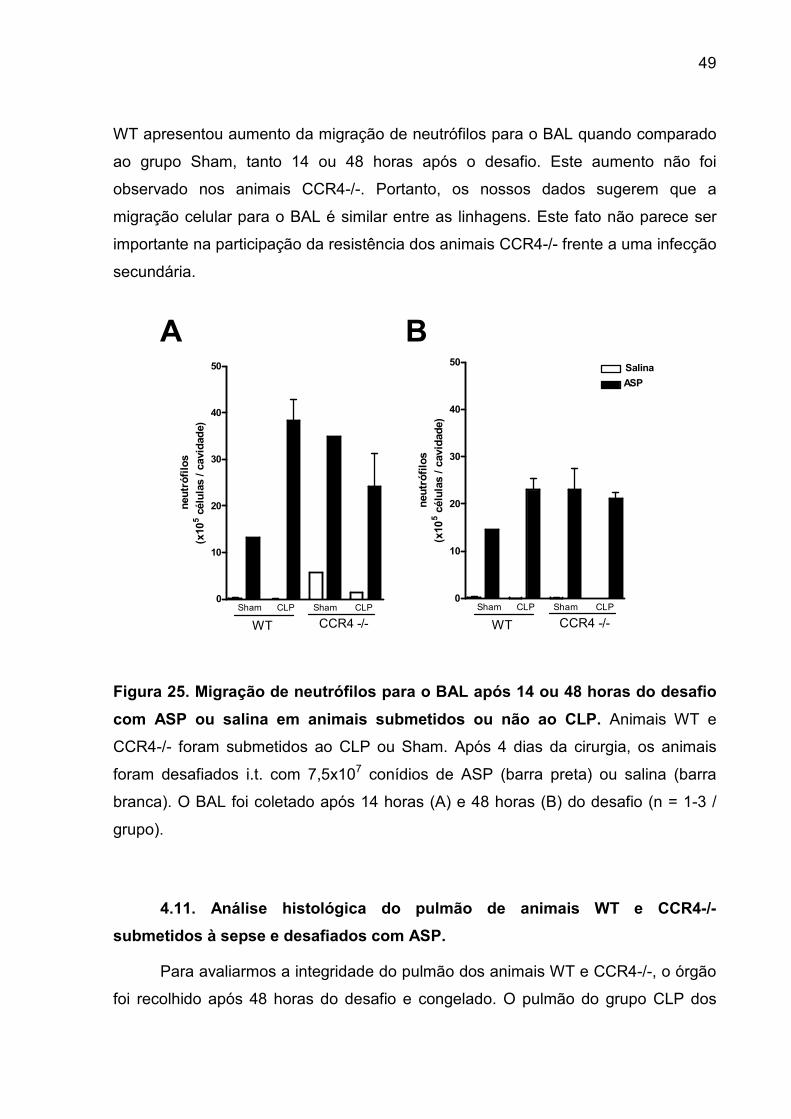
49
WT apresentou aumento da migração de neutrófilos para o BAL quando comparado
ao grupo Sham, tanto 14 ou 48 horas após o desafio. Este aumento não foi
observado nos animais CCR4-/-. Portanto, os nossos dados sugerem que a
migração celular para o BAL é similar entre as linhagens. Este fato não parece ser
importante na participação da resistência dos animais CCR4-/- frente a uma infecção
secundária.
0
10
20
30
40
50
Sham CLP
WT CCR4 -/-Sham CLP
neu
tró
filo
s
(x10
5 c
élu
las
/ ca
vid
ade)
0
10
20
30
40
50Salina
ASP
Sham CLP
WT CCR4 -/-Sham CLP
neu
tró
filo
s
(x10
5 c
élu
las
/ ca
vid
ade)
A B
Figura 25. Migração de neutrófilos para o BAL após 14 ou 48 horas do desafio
com ASP ou salina em animais submetidos ou não ao CLP. Animais WT e
CCR4-/- foram submetidos ao CLP ou Sham. Após 4 dias da cirurgia, os animais
foram desafiados i.t. com 7,5x107 conídios de ASP (barra preta) ou salina (barra
branca). O BAL foi coletado após 14 horas (A) e 48 horas (B) do desafio (n = 1-3 /
grupo).
4.11. Análise histológica do pulmão de animais WT e CCR4-/-
submetidos à sepse e desafiados com ASP.
Para avaliarmos a integridade do pulmão dos animais WT e CCR4-/-, o órgão
foi recolhido após 48 horas do desafio e congelado. O pulmão do grupo CLP dos
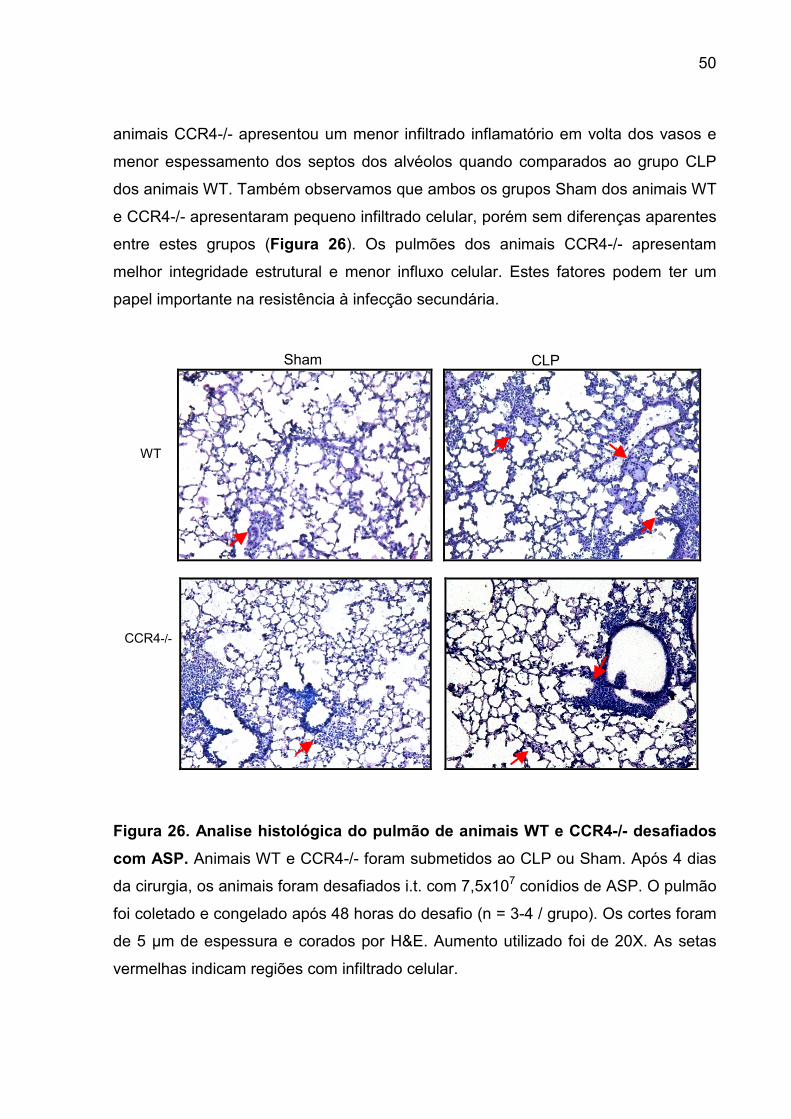
50
animais CCR4-/- apresentou um menor infiltrado inflamatório em volta dos vasos e
menor espessamento dos septos dos alvéolos quando comparados ao grupo CLP
dos animais WT. Também observamos que ambos os grupos Sham dos animais WT
e CCR4-/- apresentaram pequeno infiltrado celular, porém sem diferenças aparentes
entre estes grupos (Figura 26). Os pulmões dos animais CCR4-/- apresentam
melhor integridade estrutural e menor influxo celular. Estes fatores podem ter um
papel importante na resistência à infecção secundária.
Figura 26. Analise histológica do pulmão de animais WT e CCR4-/- desafiados
com ASP. Animais WT e CCR4-/- foram submetidos ao CLP ou Sham. Após 4 dias
da cirurgia, os animais foram desafiados i.t. com 7,5x107 conídios de ASP. O pulmão
foi coletado e congelado após 48 horas do desafio (n = 3-4 / grupo). Os cortes foram
de 5 µm de espessura e corados por H&E. Aumento utilizado foi de 20X. As setas
vermelhas indicam regiões com infiltrado celular.
WT
CCR4-/-
Sham CLP

51
4.12. Presença das células Tregs no sangue periférico e órgãos linfóides
5 dias após CLP.
A maior resistência dos animais CCR4-/- durante a fase inicial da sepse grave
parece estar relacionada com as alterações nas porcentagens das células Tregs
CD4+Foxp3+ no sangue, no baço e no linfonodo. Para avaliarmos se, assim como na
fase aguda da sepse, o CCR4 pode influenciar a população das células Tregs pelos
órgãos linfóides, fomos analisar a presença destas células Tregs CD4+Foxp3+ no
sangue, baço e no linfonodo mesentérico 5 dias após a cirurgia. Um perfil distinto na
disposição das células Tregs nos órgãos linfóides pode estar participando na
supressão da resposta imune e influenciando na resistência dos camundongos
CCR4-/- ao ASP.
No sangue periférico 5 dias após a cirurgia, observamos que os animais
CCR4-/- do grupo CLP apresentaram uma redução de 17% de células Tregs em
relação ao grupo CLP dos WT. Nos grupos Sham não observamos diferenças
(menor que 1%) entre o CCR4-/- e o WT (Figura 27A). No baço observamos um
aumento de 45% de células CD4+Foxp3+ do grupo CLP dos animais CCR4-/- em
relação aos WT. Entretanto, as porcentagens de células Tregs do grupo Sham tanto
dos CCR4-/- como dos WT apresentaram-se elevadas em relação ao grupo do CLP
(Figura 27B). No linfonodo mesentérico, também observamos um aumento de 30%
na porcentagem de células CD4+Foxp3+ do grupo CLP dos animais CCR4-/- em
relação aos WT (Figura 27C). Porém, nenhuma das diferenças foi estatisticamente
significativa.
Visto que a disposição das células Tregs encontradas não apresentaram
alterações nos animais CCR4-/-, o nosso grupo foi avaliar a presença direta das
células Tregs no tecido pulmonar.
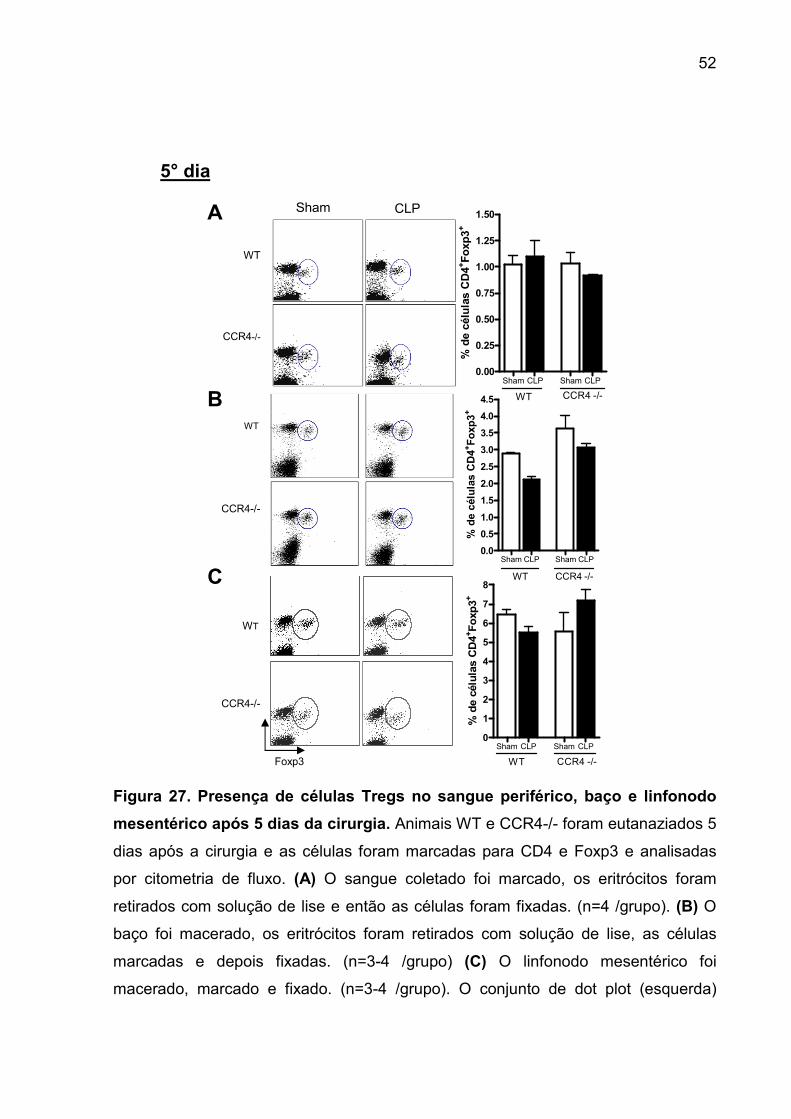
52
5° dia
A
B
C
Figura 27. Presença de células Tregs no sangue periférico, baço e linfonodo
mesentérico após 5 dias da cirurgia. Animais WT e CCR4-/- foram eutanaziados 5
dias após a cirurgia e as células foram marcadas para CD4 e Foxp3 e analisadas
por citometria de fluxo. (A) O sangue coletado foi marcado, os eritrócitos foram
retirados com solução de lise e então as células foram fixadas. (n=4 /grupo). (B) O
baço foi macerado, os eritrócitos foram retirados com solução de lise, as células
marcadas e depois fixadas. (n=3-4 /grupo) (C) O linfonodo mesentérico foi
macerado, marcado e fixado. (n=3-4 /grupo). O conjunto de dot plot (esquerda)
WT
CCR4-/-
Sham CLP
Sham CLP Sham CLP0.00
0.25
0.50
0.75
1.00
1.25
1.50
WT CCR4 -/-
% d
e cé
lula
s C
D4+
Fo
xp3+
CCR4-/-
WT
Sham CLP Sham CLP0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
4.5
WT CCR4 -/-
% d
e cé
lula
s C
D4+
Fo
xp3+
WT
CCR4-/-
Sham CLP Sham CLP0
1
2
3
4
5
6
7
8
WT CCR4 -/-
% d
e cé
lula
s C
D4+
Fo
xp3+
Foxp3

53
representa uma amostra de cada grupo e o gráfico (direita) foi expresso como média
± EPM.
4.13. Presença de células Tregs no pulmão de animais pós-sépticos
desafiados com ASP ou salina.
O próximo passo foi avaliar a presença de células Tregs no pulmão dos
animais WT e CCR4-/-, e se o perfil a ser observado se correlaciona com a
resistência a imunossupressão. Após 72 horas do desafio com ASP ou salina
(Controle), o pulmão de animais WT e CCR4-/- submetidos ao Sham ou CLP foram
retirados e digeridos com colagenase para obtenção de uma suspensão celular.
Cada grupo foi incubado com anticorpos conjugados a esferas magnéticas contra
CD4 a fim de purificarmos as células CD4+ para analisar as células CD4+Foxp3+ por
citometria de fluxo.
Nos grupos de animais que receberam salina como controle (Figura 28A),
observamos um aumento da porcentagem de células Treg nos grupos CLP
comparado ao grupo Sham, em ambas as linhagens, embora nos animais CCR4-/- a
porcentagem de células Tregs foram menores quando comparados aos animais WT.
Mas quando desafiamos os animais com ASP (Figura 28B) observamos que a
porcentagem de células Tregs do grupo CLP dos animais CCR4-/- aumentou em
relação aos animais WT. E ainda observamos que a proporção de células Tregs
diminuiu no grupo CLP comparado ao Sham dos animais WT. Os nossos dados
sugerem que o aumento de células Tregs no local de infecção após 3 dias do
desafio com ASP poderiam estar favorecendo a uma resposta contra o fungo e
participando na sobrevida. A partir destes dados, fomos então investigar a função
das células Tregs.
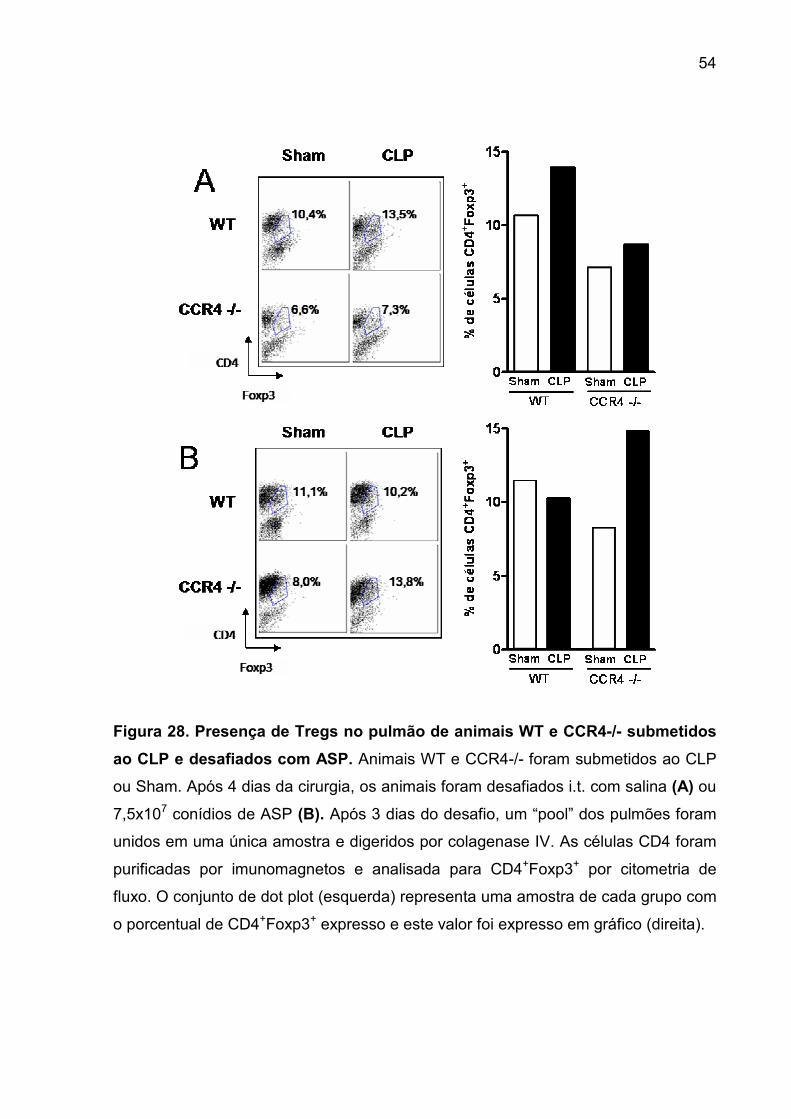
54
Figura 28. Presença de Tregs no pulmão de animais WT e CCR4-/- submetidos
ao CLP e desafiados com ASP. Animais WT e CCR4-/- foram submetidos ao CLP
ou Sham. Após 4 dias da cirurgia, os animais foram desafiados i.t. com salina (A) ou
7,5x107 conídios de ASP (B). Após 3 dias do desafio, um “pool” dos pulmões foram
unidos em uma única amostra e digeridos por colagenase IV. As células CD4 foram
purificadas por imunomagnetos e analisada para CD4+Foxp3+ por citometria de
fluxo. O conjunto de dot plot (esquerda) representa uma amostra de cada grupo com
o porcentual de CD4+Foxp3+ expresso e este valor foi expresso em gráfico (direita).

55
4.14. Inibição da migração de neutrófilos para o espaço broncoalveolar
via Tregs obtidas de animais WT e CCR4-/- submetidos ou não ao CLP.
Para avaliarmos a função das células Tregs “in vivo” sobre a migração de
leucócitos a um foco infeccioso, desafiamos os animais com 5x107 conídios de ASP
i.t. e co-injetamos células Tregs obtidas dos animais WT e CCR4-/- submetidos ao
CLP ou Sham-operados.
Os animais que receberam somente o desafio com o ASP tiveram um grande
aumento do número de neutrófilos para o espaço broncoalveolar. Os animais que só
receberam salina (grupo SAL) não apresentaram migração, como esperado. Quando
os animais foram desafiados com o ASP e receberam células Tregs obtidas do
grupo CLP dos animais CCR4-/- não observamos uma inibição da migração de
neutrófilos para o local, nem de mononucleares (dados não mostrados). Entretanto,
os animais que receberam células Tregs do grupo CLP dos WT apresentaram uma
clara inibição da migração de neutrófilos (Figura 29). Animais que receberam células
Tregs provenientes do grupo Sham do WT e CCR4-/- não foram capazes de inibir a
migração de células (dados não mostrados). Esses dados sugerem que as Tregs do
grupo CLP dos animais CCR4-/- não apresentam atividade supressora no que diz
respeito a inibição da migração de neutrófilos para o espaço broncoalveolar.
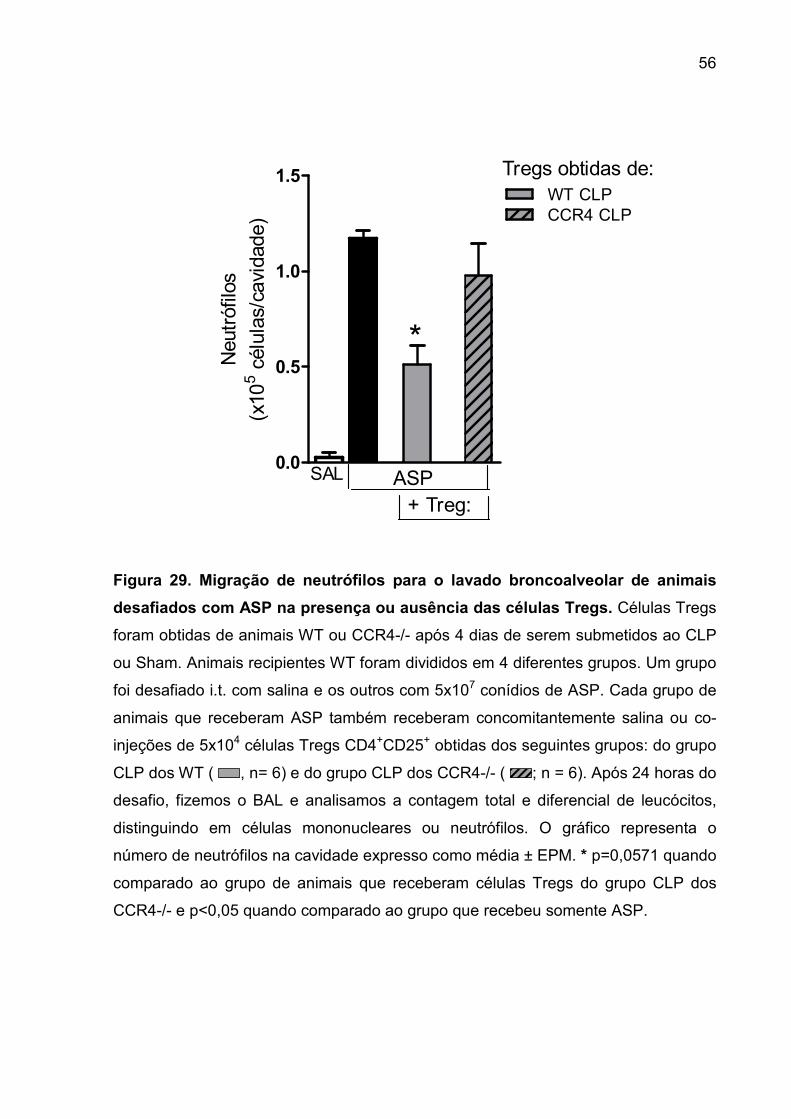
56
0.0
0.5
1.0
1.5
ASPSAL
Tregs obtidas de:
CCR4 CLP
+ Treg:
*
WT CLP
Neu
tróf
ilos
(x10
5 cél
ulas
/cav
idad
e)
Figura 29. Migração de neutrófilos para o lavado broncoalveolar de animais
desafiados com ASP na presença ou ausência das células Tregs. Células Tregs
foram obtidas de animais WT ou CCR4-/- após 4 dias de serem submetidos ao CLP
ou Sham. Animais recipientes WT foram divididos em 4 diferentes grupos. Um grupo
foi desafiado i.t. com salina e os outros com 5x107 conídios de ASP. Cada grupo de
animais que receberam ASP também receberam concomitantemente salina ou co-
injeções de 5x104 células Tregs CD4+CD25+ obtidas dos seguintes grupos: do grupo
CLP dos WT ( , n= 6) e do grupo CLP dos CCR4-/- ( ; n = 6). Após 24 horas do
desafio, fizemos o BAL e analisamos a contagem total e diferencial de leucócitos,
distinguindo em células mononucleares ou neutrófilos. O gráfico representa o
número de neutrófilos na cavidade expresso como média ± EPM. * p=0,0571 quando
comparado ao grupo de animais que receberam células Tregs do grupo CLP dos
CCR4-/- e p<0,05 quando comparado ao grupo que recebeu somente ASP.

57
4.15. Inibição da migração de neutrófilos “in vitro” via Treg obtidas de
animais WT e CCR4-/- submetidos ou não ao CLP.
Para investigarmos qual o papel do CCR4 na inibição direta das células Tregs
de animais pós-sépticos sobre os neutrófilos realizamos um ensaio de migração “in
vitro”. Neutrófilos isolados da medula óssea de um animal selvagem e células Tregs
obtidas de grupos CLP e Sham de animais WT e CCR4-/- foram pré-incubados e
depois colocados para migrar numa câmara de quimiotaxia. As células Tregs obtidas
de animais CCR4-/- do grupo CLP apresentaram uma fraca capacidade de inibir a
migração de neutrófilos quando comparado com células Tregs provenientes dos
animais WT do grupo CLP. Os neutrófilos incubados com células Tregs provenientes
dos animais WT do grupo Sham também não inibiram a migração do neutrófilo frente
a um gradiente quimiotático potente como o LTB4 (Figura 30).

58
0
4
8
12
16
20
24
RPMI
Tregs obtida de:
WT Sham
CCR4 CLP
+ TregSuperior
LTB4
WT CLP
Inferior
RPMI RPMIPoço
Inde
x qu
imio
tátic
o
Figura 30. Inibição da migração de neutrófilos “in vitro” via Tregs obtidas de
animais WT e CCR4-/-. Neutrófilos obtidos da medula óssea de animais WT foram
pré-incubados por 2 horas a 37°C com células Tregs CD4+CD25+ obtidas de animais
do grupo Sham ( ) ou CLP ( ) dos WT ou células do grupo CLP ( ) dos animais
CCR4-/-, na razão de 40 vezes. Após a pré-incubação, as células foram incubadas
em placas de 24 poços por 2 horas a 37°C frente ao LTB4. Como controle negativo
( ), os neutrófilos foram pré-incubados com apenas RPMI e incubados para migrar
sem um agente quimiotático. No controle positivo ( ), os neutrófilos foram pré-
incubados com RPMI e incubados para migrar frente ao LTB4. A migração de
neutrófilos foi avaliada pela contagem de células que migraram para o poço inferior
(n=2). O index quimiotático foi calculado através de cada valor obtido dividido pela
média dos valores do controle negativo. Os resultados foram expressos no gráfico
como média ± EPM.

59
4.16. Inibição da produção de ROS por neutrófilos estimulados com PMA
pelo SN de Tregs obtidas de animais WT e CCR4-/- submetidos ou não ao CLP.
Para investigarmos o mecanismo de ação das células Tregs sobre as funções
dos neutrófilos, estes obtidos da medula óssea de animais WT foram pré-incubados
por 2 horas com o SN das células Tregs obtidas do grupo Sham ou CLP de animais
WT ou CCR4-/- por 18 horas. Em seguida, os neutrófilos foram estimulados com
PMA para induzir a produção de ROS, o qual marcamos através de uma sonda
fluorescente. A quantificação de ROS gerada foi realizada por citometria de fluxo.
O SN das células Tregs obtidas do grupo Sham de ambas as linhagens e do
grupo CLP dos animais CCR4-/- não apresentaram uma forte inibição na produção
de ROS dos neutrófilos estimulados quando comparados ao SN das células Tregs
obtidas de animais WT do grupo CLP. O SN deste grupo apresentou uma inibição de
quase 100% em relação aos neutrófilos sem estímulo (Figura 31). Assim, o
sobrenadante obtido das células Tregs dos animais CCR4-/- pós-sépticos não
parece apresentar um fator solúvel capaz de modular a produção de ROS em
neutrófilos estimulados com PMA, como foi observado para os neutrófilos incubados
no sobrenadante das células Tregs dos animais pós-sépticos WT.
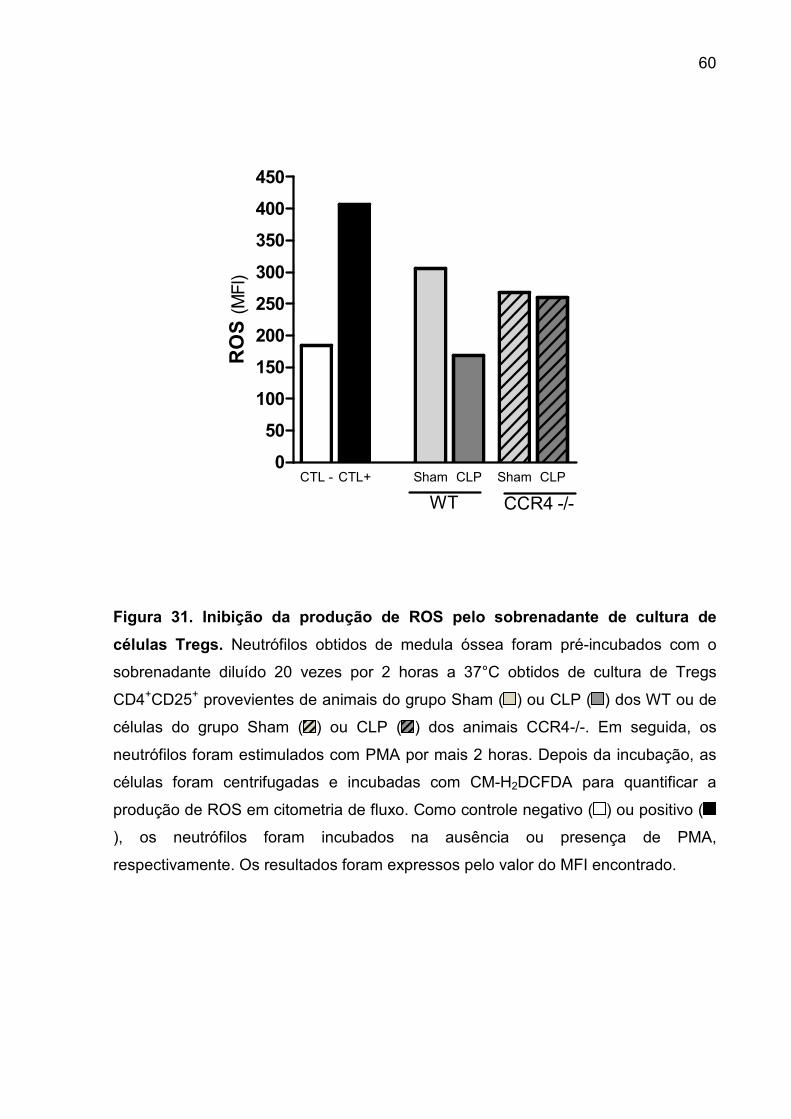
60
CTL - CTL+ Sham CLP Sham CLP0
50
100
150
200
250
300
350
400
450
WT CCR4 -/-
RO
S(M
FI)
Figura 31. Inibição da produção de ROS pelo sobrenadante de cultura de
células Tregs. Neutrófilos obtidos de medula óssea foram pré-incubados com o
sobrenadante diluído 20 vezes por 2 horas a 37°C obtidos de cultura de Tregs
CD4+CD25+ provevientes de animais do grupo Sham ( ) ou CLP ( ) dos WT ou de
células do grupo Sham ( ) ou CLP ( ) dos animais CCR4-/-. Em seguida, os
neutrófilos foram estimulados com PMA por mais 2 horas. Depois da incubação, as
células foram centrifugadas e incubadas com CM-H2DCFDA para quantificar a
produção de ROS em citometria de fluxo. Como controle negativo ( ) ou positivo (
), os neutrófilos foram incubados na ausência ou presença de PMA,
respectivamente. Os resultados foram expressos pelo valor do MFI encontrado.

61
5. Discussão
Embora existam muitas controvérsias no campo da sepse, é bastante
uniforme o pensamento de que essa síndrome é extremamente complexa devido ao
envolvimento de muitos sistemas, tecidos, tipos celulares e mediadores
inflamatórios. Muitos modelos animais têm sido usados para estudar a fisiopatologia
da sepse, com o objetivo de mimetizar as alterações observadas na sepse humana
(OSUCHOWSKI et al., 2006).
O modelo experimental murino de sepse utilizado pelo nosso grupo, CLP, que
consiste em 9 perfurações no ceco (BENJAMIM et al., 2003), nos permitiu investigar
o papel do CCR4 na sepse grave. Os nossos dados corroboram com dados da
literatura demonstrando que animais CCR4-/- são mais resistentes a baixas e altas
doses de LPS (CHVATCHKO et al., 2000), e ao Pam3Cys (NESS et al., 2006).
Mesmo sem o tratamento com antibióticos, os animais CCR4-/- possuem maior taxa
de sobrevivência por um tempo maior que os WT, os quais apresentam uma
mortalidade de 80 a 100% dentro das primeiras 48 horas após a cirurgia (dados não
mostrados). Momentos antes da primeira dose de antibiótico, 6 horas após a
cirurgia, foi observado que os animais CCR4-/- não apresentavam sinais aparentes
de endotoxemia como os WT, sintomas como letargia, olhos com secreções e
taquipnéia. A ausência do CCR4 preveniu o aparecimento destes sinais e a
evolução para um quadro séptico característico.
O processo inflamatório gerado pelo CLP induz a uma série de eventos
determinantes na sobrevida dos animais sépticos. Um evento importante é a
migração de neutrófilos para o foco infeccioso. BENJAMIM et al., 2000
demonstraram que animais submetidos a uma sepse grave apresentam falência na
migração de neutrófilos para cavidade peritoneal favorecendo a disseminação da
infecção, sendo determinante para o desfecho desfavorável com alta mortalidade.
Assim, a migração de neutrófilos para cavidade peritoneal é inversamente
proporcional ao número de perfurações no ceco. Também foi demonstrado que

62
neutrófilos obtidos de pacientes sépticos apresentam falhas na migração em câmara
de boyden e esse fato está diretamente relacionado com a gravidade e mortalidade
dos pacientes (TAVARES-MURTA et al., 2002). Nossos dados demonstraram que
nos animais CCR4-/- o número de neutrófilos obtido na cavidade peritoneal
aumentou significativamente, mostrando que não houve falência de migração como
nos animais WT.
Um importante parâmetro de gravidade na sepse é o número de bactérias
presentes na cavidade peritoneal de camundongos submetidos ao CLP. Se o foco
infeccioso não for contido pela defesa do organismo durante as primeiras horas após
a cirurgia, as bactérias migram para o sangue e atinge outros tecidos, como o
pulmão. O modelo de sepse moderada (SL) é um estímulo bem menos agressivo,
pois o volume de material fecal que entra na cavidade peritoneal é menor que o
modelo de sepse grave (L). A menor quantidade de bactérias na cavidade peritoneal
dos animais CCR4-/- provavelmente se deve ao grande número de neutrófilos
recrutados. Estas células tem um papel central na eliminação dos patógenos. Neste
contexto, a infecção contida na cavidade peritoneal impede a disseminação de
bactérias para o sangue. Em animais WT que demonstraram falência de neutrófilos
para a cavidade peritoneal, observamos grande quantidade de bactérias na
cavidade e na circulação, devido ao ineficaz controle da infecção. Os animais Sham,
tanto de animal WT como do CCR4-/-, apresentaram uma migração basal para a
cavidade devido ao processo cirúrgico recente e não foram detectadas bactérias
presentes na cavidade peritoneal ou no sangue. Um estudo em animais TLR9-/-
submetidos ao CLP demonstrou que estes animais possuem um grande número de
granulócitos e DC no peritônio e que estas células são importantes no controle da
infecção. A transferência de DC dos animais TLR9-/- é capaz de proteger o animal
WT do CLP e aumentar o número de neutrófilos na cavidade (PLITAS et al., 2008).
Os nossos dados mostram que o grande número de neutrófilos recrutados para a
cavidade peritoneal de animais CCR4-/- foi importante para conter a infecção e
impedir que as bactérias se disseminassem para o sangue, aumentando a sobrevida
dos animais operados. Assim como o CCR4, os CCR1, 2 e 5 são receptores que
também participam na migração de células num modelo de peritonite aguda
(SPEYER et al., 2004).

63
Um outro evento importante na resposta inflamatória durante a sepse grave é
a liberação exacerbada de mediadores inflamatórios, dentre elas citocinas e
quimiocinas, tanto no local como sistemicamente. Alguns trabalhos correlacionam a
presença de altas concentrações de algumas citocinas e quimiocinas com um papel
prejudicial no controle da infecção. WALLEY et al, 1997 demonstraram que
elevadas concentrações da quimiocina MIP-2 no peritônio estava correlacionada
com o agravamento da severidade da sepse (WALLEY et al., 1997). Em nosso
trabalho os animais CCR4-/- apresentaram menores concentrações de MIP-2 na
cavidade peritoneal, embora essa diferença não seja significativa em relação ao
grupo WT. Nossos dados corroboram com os dados da literatura de que a baixa
concentração de MIP-2 no peritônio nos animais CCR4-/- foi favorável para que
estes animais não desenvolvam um quadro grave de sepse. Outra hipótese é que
esta ligeira diminuição na produção de MIP-2 dos animais CCR4-/- após 6 horas da
cirurgia poderia ser devido aos animais CCR4-/- eliminarem mais rapidamente o foco
infeccioso e por isto apresentarem uma menor liberação deste mediador.
Assim como o MIP-2, o TNF-α produzido pelos animais CCR4-/- nos grupos
submetidos à sepse moderada e grave estava em menor quantidade no peritônio
quando comparado aos WT. Estes dados sugerem que estas citocinas e quimiocinas
pró-inflamatórias estão sendo produzidas e controladas no local e quando está em
menor quantidade desempenham um papel favorável no processo inflamatório. A IL-
10, uma citocina anti-inflamatória, também se apresentou reduzida nos animais
CCR4-/- quando submetidos à sepse moderada, porém elevada quando submetidos
a uma sepse grave. Este dado também nos sugere que a IL-10 no peritônio possui
um papel protetor na sepse grave.
A infecção local pode disseminar de tal modo que em órgãos como o pulmão,
rins e fígado é possível encontrar bactérias. A presença de bactérias nestes órgãos
induz a migração de neutrófilos para estes tecidos. Embora os neutrófilos não
migrem para o foco infeccioso na sepse grave, eles aderem a microcirculação de
órgãos distantes do foco infeccioso, como fígado, rins e pulmões. Esse fator
contribui para lesão tecidual, falência múltipla de órgãos e morte do hospedeiro
(COLLIN et al., 2004). Para determinar a presença de infiltrado de neutrófilos nos

64
pulmões dosamos a atividade da enzima mieloperoxidase, uma enzima específica
presente nos neutrófilos, no extrato pulmonar de animais WT e CCR4-/- submetidos
à sepse moderada ou grave. A presença de um menor infiltrado neutrofílico
pulmonar nos animais CCR4-/- demonstra mais uma vez que esses animais são
mais resistentes a inflamação sistêmica. Os animais CCR4-/- não desenvolveram
uma disseminação bacteriana de tamanha intensidade capaz de afetar este órgão.
É importante mencionar que grandes quantidades de citocinas no local da
infecção possuem um importante papel na ativação e recrutamento de leucócitos
sanguíneos para o sítio inflamatório com conseqüente controle do foco infeccioso.
Todavia, as mesmas citocinas são deletérias quando presentes em altas
concentrações na circulação, pois provocam a falência da migração de neutrófilos
para o sítio infeccioso e induzem alterações cardiovasculares que resultam no
choque com posterior hipoperfusão (BENJAMIM; FERREIRA & CUNHA, 2000;
BENJAMIM et al., 2002). Uma menor concentração de citocinas plasmáticas nos
animais CCR4-/- submetidos a sepse grave indica uma resposta mais favorável, o
que se correlaciona com uma maior sobrevivência.
Corroborando com os nossos dados, OSUCHOWSKI et al., 2006
demonstraram elevadas quantidades de citocinas plasmáticas pró-inflamatórias (IL-
6, TNF, IL-1β, KC, MIP-2, MCP-1 e eotaxina) e anti-inflamatórias (receptores
solúveis de TNF, IL-10, receptores antagonista da IL-1) em animais que morreram
de um quadro séptico agudo (nos 4 primeiros dias). Também foi demonstrado que
as quantidades de IL-10 estão diretamente correlacionadas com a severidade da
inflamação e o desenvolvimento de falência de órgãos no choque séptico (Friedman,
1997 #398). Assim, o TNF-α e a IL-10 são citocinas que podem servir de
marcadores da intensidade da resposta inflamatória sistêmica. De forma similar,
maiores quantidades do fator inibidor da migração de macrófagos (MIF) se
correlacionam com letalidade na sepse murina e humana (BOZZA et al., 1999;
BOZZA et al., 2004). Portanto, a falta do CCR4, e a conseqüente redução de
mediadores inflamatórios na circulação, mostrou-se favorável para o organismo
controlar e impedir a exarcebação de uma resposta inflamatória sistêmica na sepse
grave.

65
Algumas desordens ou mecanismos compensatórios podem surgir em
modelos de animais deficientes. A maior migração de neutrófilos observada nos
animais CCR4-/- poderia ser em parte decorrente de alterações na sua produção na
medula óssea. Quando fomos avaliar a celularidade destes animais observamos que
não apresentaram diferenças nas porcentagens de neutrófilos presentes no sangue
ou na medula. Isto confirma a nossa hipótese de que os animais CCR4-/-
apresentaram uma resposta inflamatória mais eficiente e provavelmente mais rápida,
e não devido a alterações na quantidade de células em condições basais.
Além da resposta no foco infeccioso, o encontro do patógeno e seus
antígenos com as células do sistema imunitário acontecem nos órgãos linfóides
periféricos como o sangue e outros tecidos periféricos. Diversas células, como os
linfócitos, estão recirculando no organismo e passam continuamente pelos órgãos
linfóides. O baço e o linfonodo mesentérico são os tecidos linfóides que drenam a
região acometida pela infecção no modelo do CLP. Dentre os linfócitos envolvidos
na resposta ao CLP, as células Tregs tem sido descritas como células importantes
no controle da resposta imune frente a diferentes estímulos infecciosos (WALKER &
ROTONDO, 2004). As células Tregs presentes podem ser ativadas por produtos de
microrganismos ou moléculas pró-inflamatórias que estão sendo liberadas pelo
hospedeiro (RUDENSKY & CAMPBELL, 2006). A disposição das células Tregs no
sangue, baço e no linfonodo mesentérico poderiam então estar influenciando o perfil
de migração e ativação de outras células e a evolução da sepse grave. Além disso,
o nosso interesse nas Tregs durante a sepse em animais CCR4-/- é devido as Tregs
expressarem o receptor CCR4, e esse receptor ser importante para a migração
desse tipo celular para o foco da inflamação (LEE et al, 2005). Ainda, não existe
nenhum dado na literatura sobre o papel desse receptor na ativação da Treg e na
participação dessa célula na resposta inata e na imunossupressão pós-sepse grave.
As porcentagens das células Tregs no sangue como no baço parecem estar
aumentadas no grupo CLP do CCR4-/- em relação ao WT durante a sepse. Esse
fato pode estar relacionado a uma maior resistência do animal CCR4-/- frente a
sepse grave, favorecendo uma menor inflamação sistêmica. Este fenômeno poderá

66
ser melhor explorado com um maior número de animais e uma análise mais
criteriosa em diferentes tempos.
O papel das Tregs em modelos de infecção é controverso. Alguns trabalhos
indicam que as células CD4+CD25+ não são capazes de aumentar a sobrevida dos
animais em diferentes modelos de infecção (STEINHAUSER et al., 1999;
SAKAGUCHI, 2003; SCUMPIA et al., 2006; SUFFIA et al., 2006; ZAKRZEWSKI et
al., 2006; ANDERSON et al., 2007). Por outro lado, a transferência adotiva de
células Tregs CD4+CD25+ estimuladas in vitro com anti-CD3 e anti-CD28 em animais
submetidos ao CLP melhorou a sobrevida e diminuiu a quantidade de CFU presente
no sítio de infecção (HEUER et al., 2005). Uma das conseqüências da sepse é o
quadro de imunossupressão observado em pacientes e animais pós-sépticos. Esses
pacientes que se recuperam apresentam alto índice de mortalidade no período de
um ano. Dos pacientes acometidos de sepse grave que obtiveram alta, 80%
faleceram em um período de 8 anos por causas não sépticas, mas devido a doenças
inflamatórias como câncer, problemas cardiovasculares e pulmonares (PERL et al.,
1995; QUARTIN et al., 1997). Os nossos resultados demonstram que a ausência do
CCR4 tem um papel favorável na sobrevida frente a uma infecção secundária, pois
os animais CCR4 -/- não desenvolveram uma resposta inflamatória inicial excessiva
e também não demonstraram um quadro de imunossupressão pós-séptica.
As primeiras células a chegarem ao pulmão em uma infecção fúngica são os
neutrófilos. Os animais CCR4-/- parecem responder de forma semelhante aos
animais WT pós-sépticos, embora o número de animais utilizados neste ensaio
tenha sido baixo para afirmarmos estes resultados. Ainda assim, os nossos dados
sugerem que a migração celular para o BAL não parece explicar a resistência destes
animais ao ASP. Dados anteriores demonstraram que as células dos animais
submetidos ao CLP produzem mediadores pró-inflamatórios (citocinas e
quimiocinas), porém não conseguem combater a infecção secundária, sugerindo
uma falha na atividade microbicida das células inflamatórias (BENJAMIM et al.,
2003).
Um dos tecidos acometidos pela sepse é o pulmão, especialmente devido ao
desenvolvimento do edema alveolar que representa apenas uma das manifestações

67
da resposta sistêmica (MALLOY et al., 1997). Mesmo após 48 horas da cirurgia,
constatou-se que animais sépticos sofrem um aumento da celularidade no tecido
pulmonar juntamente com espessamento do septo alveolar (ROCCO et al., 2001).
Em animais desafiados com ASP, diferente do observado para a migração celular
para o BAL, os pulmões dos CCR4-/- apresentaram menor influxo celular devido a
um processo de eliminação mais rápido do ASP enquanto o grupo CLP dos animais
WT não conseguem eliminar a infecção desencadeando uma resposta celular
intensa. Esses dados sugerem que os camundongos WT sépticos são suscetíveis à
IPA e que a incapacidade do sistema imune de resolver a infecção não está
associada á capacidade fagocítica e sim à incapacidade dos leucócitos de matarem
os conídios do fungo (BENJAMIM et al., 2003). Na sepse, a inflamação persistente
composta por neutrófilos nos alvéolos correlaciona com alta mortalidade (MATUTE-
BELLO et al., 1997).
Em conjunto, os nossos dados até o momento sugerem que a resistência dos
animais CCR4-/- à infecção pelo ASP não parece ser decorrente de um maior
recrutamento de neutrófilos para o tecido pulmonar ou para o BAL, já que estes
apresentaram influxo celular menor ou similar. A resistência do CCR4-/- a infecção
secundária tão pouco parece sofrer influência do aumento de células Tregs nos
órgãos linfóides após 5 dias da cirurgia ou na circulação.
Um estudo recente demonstrou que a presença das células CD4+Foxp3+ na
cavidade pleural é importante na susceptibilidade de animais BALB/C a L.
sigmodontis, pois estas células afetam a capacidade do animal em reduzir a carga
parasitária (TAYLOR et al., 2007). Esse mesmo trabalho demonstrou que a
presença das células Tregs no local neste tipo de infecção era importante. Então
com base neste trabalho avaliamos se as células Tregs poderiam migrar diretamente
para o pulmão de animais WT e causar uma supressão da resposta local,
favorecendo ao patógeno e prejudicando a sobrevida do animal. Diferente dos
órgãos linfóides, no pulmão dos animais CCR4-/- observamos uma menor
porcentagem de células Tregs quando comparado aos animais WT submetidos ao
CLP sem o desafio com o fungo. Isto poderia explicar a resposta imune mais
eficiente dos CCR4-/- no combate ao fungo nos pulmões. Entretanto, quando os

68
animais CCR4-/- foram desafiados com ASP, observamos um aumento das células
Tregs no pulmão comparado ao animal WT pós-séptico. Com base neste dado,
sugerimos que o aumento de células Tregs no pulmão dos animais CCR4-/- poderia
estar modulando uma resposta intensa do sistema imune frente a infecção com
ASP, o que evitaria danos teciduais e a falência funcional. Fato que poderia explicar
os dados da histologia, onde foi mostrado redução do influxo celular para o pulmão
dos animais desafiados com ASP.
Além de avaliarmos quantitativamente as células Tregs no sangue, pulmão e
tecidos linfóides, determinamos a capacidade supressora das células Tregs sobre
outras células inflamatórias. As células Tregs podem modular a resposta imune
contra infecções pela inibição da migração de células mononucleares e neutrófilos
para o sítio inflamatório. Estudos in vitro já haviam demonstrado que o LPS induz
nas células Tregs CD4+CD25+ aumento da capacidade supressora e a sua
sobrevivência/proliferação (CARAMALHO et al., 2003). Em nosso modelo
experimental, os animais desafiados com fungo que receberam as células Tregs dos
animais CCR4-/- submetidos ao CLP não apresentaram inibição da migração dos
neutrófilos para o foco infeccioso, fato observado para as Tregs dos animais
selvagens. Além disso, para excluirmos a possibilidade de um efeito indireto das
células Tregs, estas foram incubadas diretamente com os neutrófilos. Nos animais
WT observamos que a migração dos neutrófilos realmente foi inibida pelas células
Tregs, enquanto que as Tregs dos animais CCR4-/- não inibiram a migração. Os
nossos dados sugerem que o receptor CCR4 na sepse grave é importante para a
capacidade supressora das células Tregs quanto ao efeito na migração de
neutrófilos para o foco infeccioso. Logo, a redução de infiltrado leucocitário
observado no pulmão dos animais CCR4-/- após desafio com ASP não se deve a
uma atividade supressora das Tregs, com sugerimos anteriormente.
Em 2006, foi demonstrado que as células Tregs podem afetar a função e a
sobrevivência dos neutrófilos. As células Tregs ativadas por LPS podem promover a
apoptose, inibir a produção de ROS e a liberação de citocinas dos neutrófilos. Essa
inibição foi parcialmente dependente de IL-10 e TGF-β (LEWKOWICZ et al., 2006).
Durante a resposta inflamatória exacerbada existe uma grande quantidade de LPS

69
capaz de ativar as células Tregs. Isto acontece principalmente nos animais WT,
enquanto que nos animais CCR4-/- que conseguem conter a infecção e impedir a
evolução da sepse não observamos um caráter supressor nas Tregs pós-sépticas.
Estudos anteriores demonstraram que assim como os neutrófilos no pulmão,
os macrófagos alveolares desempenham um papel central contra conídios de ASP e
que a fagocitose dos conídios pelos macrófagos não está alterada na
imunossupressão induzida por corticosteróides (in vivo e in vitro). Entretanto, as
ROS são essenciais para eliminar os conídios após terem sido fagocitados pelos
macrófagos (IBRAHIM-GRANET et al., 2003). E de acordo com dados anteriores do
nosso grupo foi demonstrado que camundongos sépticos são suscetíveis ao ASP e
que a incapacidade do sistema imune de resolver a infecção não está associada á
capacidade fagocítica, e sim à incapacidade dos leucócitos de eliminarem os
conídios do fungo (BENJAMIM et al., 2003).
Podemos inferir com base em nossos dados que os neutrófilos incubados
com o sobrenadante das células Tregs dos animais CCR4-/- não tiveram sua
produção de ROS inibida. Mais um dado que sugere que essas células Tregs não
adquiriram capacidade supressora. Este resultado também sugere que existe algum
fator solúvel liberado pelas Tregs dos WT pós-sépticos, e não pelas Tregs dos
CCR4-/-, que inibe a produção de ROS nos neutrófilos.
Com base em todos os nossos resultados apresentados, a nossa hipótese,
como mostrado na Figura 32, propõe que a falta do CCR4 melhora a resposta inata
tornando-a mais eficiente para conter a infecção no local, impedindo sua
disseminação a outros órgãos. A ausência dessa intensa resposta inflamatória
aguda durante a sepse nos CCR4-/- parece ser importante para que as células
Tregs não adquiram um caráter supressor numa resposta pós-sepse. A falta dessa
capacidade supressora dessas células deve contribuir para que os animais CCR4-/-
não desenvolvam uma imunossupressão tardia e respondam quando desafiados a
um segundo insulto. As células Tregs dos CCR4-/- não inibem a migração de
neutrófilos para o foco infeccioso ou afetam a geração de ROS, o que torna esses
camundongos CCR4-/- resistentes e capazes de eliminar o fungo através da
ativação de seus neutrófilos.

70
Fig
ura
32.
Im
po
rtâ
nci
a d
o C
CR
4 n
a s
ep
se
pa
ra e
vo
luçã
o d
a i
mu
no
ss
up
ress
ão
.

71
6. CONCLUSÕES
1. Os animais CCR4-/- foram mais resistentes a sepse grave e não
apresentaram um perfil de imunossupressão 4 dias após o CLP quando desafiados
com A. fumigatus.
2. A resposta inata dos CCR4-/- demonstrou melhor capacidade de conter
o processo inflamatório no modelo de CLP grave, através de uma melhor contenção
da disseminação bacteriana.
3. Os animais CCR4-/- não apresentaram o desenvolvimento de uma
resposta inflamatória sistêmica exacerbada como os animais WT.
4. O aumento da porcentagem de células Tregs nos animais CCR4-/-
parece influenciar a sobrevida na fase inicial da sepse, entretanto o número de
Tregs na fase pós-sepse não parece explicar a resistência dos CCR4-/- à infecção
secundária.
5. As células Tregs dos animais CCR4-/- submetidos à sepse grave não
apresentam uma capacidade de supressão in vivo e in vitro sobre a migração
neutrófilos ou a geração de ROS por estas células.
Este trabalho demonstra o envolvimento do CCR4 desde a fase inicial da
sepse até a evolução da imunossupressão pós-sepse. O melhor entendimento do
papel deste receptor durante a sepse grave contribui para o desenvolvimento de
possíveis intervenções terapêuticas que possam levar uma melhora no quadro
clínico, um aumento na sobrevida dos pacientes na sepse e ainda, impedir o
desenvolvimento de um quadro de imunossupressão a longo prazo. Além disso, o
estudo sugere a participação essencial do CCR4 sobre a migração das células
Tregs e sobre possíveis mecanismos de inibição sobre outras células inflamatórias.

72
7. BIBLIOGRAFIA
ABRAHAM, E.; Alterations in cell signaling in sepsis. Clin Infect Dis, 41 Suppl 7, S459-464, 2005.
ANDERSON, C.F.; OUKKA, M.; KUCHROO, V.J. & SACKS, D.; CD4(+)CD25(-)Foxp3(-) Th1 cells are the source of IL-10-mediated immune suppression in chronic cutaneous leishmaniasis. J Exp Med, 204 (2), 285-297, 2007.
BAATAR, D.; OLKHANUD, P.; SUMITOMO, K.; TAUB, D.; GRESS, R. & BIRAGYN, A.; Human peripheral blood T regulatory cells (Tregs), functionally primed CCR4+ Tregs and unprimed CCR4- Tregs, regulate effector T cells using FasL. J Immunol, 178 (8), 4891-4900, 2007.
BAKER, C.C.; CHAUDRY, I.H.; GAINES, H.O. & BAUE, A.E.; Evaluation of factors affecting mortality rate after sepsis in a murine cecal ligation and puncture model. Surgery, 94 (2), 331-335., 1983.
BALLOY, V.; SI-TAHAR, M.; TAKEUCHI, O.; PHILIPPE, B.; NAHORI, M.A.; TANGUY, M.; HUERRE, M.; AKIRA, S.; LATGE, J.P. & CHIGNARD, M.; Involvement of toll-like receptor 2 in experimental invasive pulmonary aspergillosis. Infect Immun, 73 (9), 5420-5425, 2005.
BELKAID, Y.; Regulatory T cells and infection: a dangerous necessity. Nat Rev Immunol, 7 (11), 875-888, 2007.
BELPERIO, J.A.; DY, M.; MURRAY, L.; BURDICK, M.D.; XUE, Y.Y.; STRIETER, R.M. & KEANE, M.P.; The role of the Th2 CC chemokine ligand CCL17 in pulmonary fibrosis. J Immunol, 173 (7), 4692-4698, 2004.
BENJAMIM, C., FERREIRA, SH , CUNHA, FQ; Role of nitric oxide in the failure of neutrophil migration in sepsis. Journal Infection Disease, 182, 214-223, 2000.
BENJAMIM, C.F.; HOGABOAM, C.M. & KUNKEL, S.L.; The chronic consequences of severe sepsis. J Leukoc Biol, 75 (3), 408-412, 2004.
BENJAMIM, C.F.; HOGABOAM, C.M.; LUKACS, N.W. & KUNKEL, S.L.; Septic mice are susceptible to pulmonary aspergillosis. Am J Pathol, 163 (6), 2605-2617, 2003.
BENJAMIM, C.F.; LUNDY, S.K.; LUKACS, N.W.; HOGABOAM, C.M. & KUNKEL, S.L.; Reversal of long-term sepsis-induced immunosuppression by dendritic cells. Blood, 105 (9), 3588-3595, 2005.

73
BENJAMIM, C.F.; SILVA, J.S.; FORTES, Z.B.; OLIVEIRA, M.A.; FERREIRA, S.H. & CUNHA, F.Q.; Inhibition of leukocyte rolling by nitric oxide during sepsis leads to reduced migration of active microbicidal neutrophils. Infect Immun, 70 (7), 3602-3610., 2002.
BENJAMIM, C.F.L.S.K.L.N.W.H.C.M.K.S.L.; Reversal of Long-term Sepsis-induced Immunosuppression by Dendritic Cell. Blood Journal, 105 (9), 3588-3595, 2005.
BEUTLER, B. & POLTORAK, A.; Sepsis and evolution of the innate immune response. Crit Care Med, 29 (7 Suppl), S2-6; discussion S6-7, 2001.
BONE, R.C.; Sepsis and its complications: the clinical problem. Crit Care Med, 22 (7), S8-11, 1994.
BOZZA, M.; SATOSKAR, A.R.; LIN, G.; LU, B.; HUMBLES, A.A.; GERARD, C. & DAVID, J.R.; Targeted disruption of migration inhibitory factor gene reveals its critical role in sepsis. J Exp Med, 189 (2), 341-346, 1999. BOZZA, F.A.; GOMES, R.N.; JAPIASSU, A.M.; SOARES, M.; CASTRO-FARIA-NETO, H.C.; BOZZA, P.T. & BOZZA, M.T.; Macrophage migration inhibitory factor levels correlate with fatal outcome in sepsis. Shock, 22 (4), 309-313, 2004.
BYSTRY, R.S.; ALUVIHARE, V.; WELCH, K.A.; KALLIKOURDIS, M. & BETZ, A.G.; B cells and professional APCs recruit regulatory T cells via CCL4. Nat Immunol, 2 (12), 1126-1132, 2001.
CARAMALHO, I.; LOPES-CARVALHO, T.; OSTLER, D.; ZELENAY, S.; HAURY, M. & DEMENGEOT, J.; Regulatory T cells selectively express toll-like receptors and are activated by lipopolysaccharide. J Exp Med, 197 (4), 403-411, 2003.
CAVASSANI, K.A.; CAMPANELLI, A.P.; MOREIRA, A.P.; VANCIM, J.O.; VITALI, L.H.; MAMEDE, R.C.; MARTINEZ, R. & SILVA, J.S.; Systemic and local characterization of regulatory T cells in a chronic fungal infection in humans. J Immunol, 177 (9), 5811-5818, 2006.
CHEN, D. & BROMBERG, J.S.; T regulatory cells and migration. Am J Transplant, 6 (7), 1518-1523, 2006.
CHENSUE, S.W.; WARMINGTON, K.S.; RUTH, J.H.; SANGHI, P.S.; LINCOLN, P. & KUNKEL, S.L.; Role of monocyte chemoattractant protein-1 (MCP-1) in Th1 (mycobacterial) and Th2 (schistosomal) antigen-induced granuloma formation: relationship to local inflammation, Th cell expression, and IL-12 production. J Immunol, 157 (10), 4602-4608, 1996.
CHVATCHKO, Y.; HOOGEWERF, A.J.; MEYER, A.; ALOUANI, S.; JUILLARD, P.; BUSER, R.; CONQUET, F.; PROUDFOOT, A.E.; WELLS, T.N. & POWER, C.A.; A key role for CC chemokine receptor 4 in lipopolysaccharide-induced endotoxic shock. J Exp Med, 191 (10), 1755-1764, 2000.
COHEN; The immunopathogenesis of sepsis. Nature, 420, 885-891, 2002.

74
COLANTONIO, L.; IELLEM, A.; SINIGAGLIA, F. & D'AMBROSIO, D.; Skin-homing CLA+ T cells and regulatory CD25+ T cells represent major subsets of human peripheral blood memory T cells migrating in response to CCL1/I-309. Eur J Immunol, 32 (12), 3506-3514, 2002.
COLLIN, M.; ROSSI, A.; CUZZOCREA, S.; PATEL, N.S.; DI PAOLA, R.; HADLEY, J.; COLLINO, M.; SAUTEBIN, L. & THIEMERMANN, C.; Reduction of the multiple organ injury and dysfunction caused by endotoxemia in 5-lipoxygenase knockout mice and by the 5-lipoxygenase inhibitor zileuton. J Leukoc Biol, 76 (5), 961-970, 2004.
COMERFORD, I. & NIBBS, R.J.; Post-translational control of chemokines: a role for decoy receptors? Immunol Lett, 96 (2), 163-174, 2005.
CURIEL, T.J.; COUKOS, G.; ZOU, L.; ALVAREZ, X.; CHENG, P.; MOTTRAM, P.; EVDEMON-HOGAN, M.; CONEJO-GARCIA, J.R.; ZHANG, L.; BUROW, M.; ZHU, Y.; WEI, S.; KRYCZEK, I.; DANIEL, B.; GORDON, A.; MYERS, L.; LACKNER, A.; DISIS, M.L.; KNUTSON, K.L.; CHEN, L. & ZOU, W.; Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat Med, 10 (9), 942-949, 2004.
D'AMBROSIO, D.; PANINA-BORDIGNON, P. & SINIGAGLIA, F.; Chemokine receptors in inflammation: an overview. J Immunol Methods, 273 (1-2), 3-13, 2003.
DEAGLIO, S.; DWYER, K.M.; GAO, W.; FRIEDMAN, D.; USHEVA, A.; ERAT, A.; CHEN, J.F.; ENJYOJI, K.; LINDEN, J.; OUKKA, M.; KUCHROO, V.K.; STROM, T.B. & ROBSON, S.C.; Adenosine generation catalyzed by CD39 and CD73 expressed on regulatory T cells mediates immune suppression. J Exp Med, 204 (6), 1257-1265, 2007.
DINARELLO, C.A. & ABRAHAM, E.; Does blocking cytokines in sepsis work? Am J Respir Crit Care Med, 166 (9), 1156-1157, 2002.
DONG, Y.L.; ABDULLAH, K.; YAN, T.Z.; RUTAN, T.; BROEMELING, L.; ROBSON, M.; HERNDON, D.N. & WAYMACK, J.P.; Effect of thermal injury and sepsis on neutrophil function. J Trauma, 34 (3), 417-421, 1993.
FAURSCHOU, M. & BORREGAARD, N.; Neutrophil granules and secretory vesicles in inflammation. Microbes Infect, 5 (14), 1317-1327, 2003.
FERREIRA, S.H.; Are macrophages body's alarm cells? Agents and Actions, 10, 229-230, 1980.
FINK, M.P. & HEARD, S.O.; Laboratory models of sepsis and septic shock. J Surg Res, 49 (2), 186-196, 1990.
FIVENSON, D.P. & NICKOLOFF, B.J.; Immunodiagnosis in cutaneous T cell lymphoma: how does gene expression of the variable region of the T cell receptor fit into the diagnostic and pathophysiological picture of T cell neoplasia. J Cutan Pathol, 19 (1), 1-5, 1992.

75
FUJIMURA, T.; AIBA, S.; YOSHINO, Y.; KUROKI, S.; KIMURA, Y.; KIKUCHI, K.; KUNII, T.; MATSUNAGA, J.; MATSUSHIMA, K. & TAGAMI, H.; CCR4 Expression by atypical T cells in systemic pilotropic lymphoma: its behavior under treatment with interferon gamma, topical PUVA and systemic retinoid. Dermatology, 208 (3), 221-226, 2004.
HEUER, J.G.; ZHANG, T.; ZHAO, J.; DING, C.; CRAMER, M.; JUSTEN, K.L.; VONDERFECHT, S.L. & NA, S.; Adoptive transfer of in vitro-stimulated CD4+CD25+ regulatory T cells increases bacterial clearance and improves survival in polymicrobial sepsis. J Immunol, 174 (11), 7141-7146, 2005.
IBRAHIM-GRANET, O.; PHILIPPE, B.; BOLETI, H.; BOISVIEUX-ULRICH, E.; GRENET, D.; STERN, M. & LATGE, J.P.; Phagocytosis and intracellular fate of Aspergillus fumigatus conidia in alveolar macrophages. Infect Immun, 71 (2), 891-903, 2003.
IELLEM, A.; MARIANI, M.; LANG, R.; RECALDE, H.; PANINA-BORDIGNON, P.; SINIGAGLIA, F. & D'AMBROSIO, D.; Unique chemotactic response profile and specific expression of chemokine receptors CCR4 and CCR8 by CD4(+)CD25(+) regulatory T cells. J Exp Med, 194 (6), 847-853, 2001.
ISHII, M.; HOGABOAM, C.M.; JOSHI, A.; ITO, T.; FONG, D.J. & KUNKEL, S.L.; CC chemokine receptor 4 modulates Toll-like receptor 9-mediated innate immunity and signaling. Eur J Immunol, 38 (8), 2290-2302, 2008.
JAKUBZICK, C.; WEN, H.; MATSUKAWA, A.; KELLER, M.; KUNKEL, S.L. & HOGABOAM, C.M.; Role of CCR4 ligands, CCL17 and CCL22, during Schistosoma mansoni egg-induced pulmonary granuloma formation in mice. Am J Pathol, 165 (4), 1211-1221, 2004.
KLEINEWIETFELD, M.; PUENTES, F.; BORSELLINO, G.; BATTISTINI, L.; ROTZSCHKE, O. & FALK, K.; CCR6 expression defines regulatory effector/memory-like cells within the CD25(+)CD4+ T-cell subset. Blood, 105 (7), 2877-2886, 2005.
KOBAYASHI, M.; TAKAHASHI, H.; SANFORD, A.P.; HERNDON, D.N.; POLLARD, R.B. & SUZUKI, F.; An increase in the susceptibility of burned patients to infectious complications due to impaired production of macrophage inflammatory protein 1 alpha. J Immunol, 169 (8), 4460-4466, 2002.
KUNKEL, E.J. & BUTCHER, E.C.; Chemokines and the tissue-specific migration of lymphocytes. Immunity, 16 (1), 1-4, 2002.
KURIHARA, T.; WARR, G.; LOY, J. & BRAVO, R.; Defects in macrophage recruitment and host defense in mice lacking the CCR2 chemokine receptor. J Exp Med, 186 (10), 1757-1762, 1997.
LEE, I.; WANG, L.; WELLS, A.D.; DORF, M.E.; OZKAYNAK, E. & HANCOCK, W.W.; Recruitment of Foxp3+ T regulatory cells mediating allograft tolerance depends on the CCR4 chemokine receptor. J Exp Med, 201 (7), 1037-1044, 2005.

76
LEWKOWICZ, P.; LEWKOWICZ, N.; SASIAK, A. & TCHORZEWSKI, H.; Lipopolysaccharide-activated CD4+CD25+ T regulatory cells inhibit neutrophil function and promote their apoptosis and death. J Immunol, 177 (10), 7155-7163, 2006.
LEY, K.; Arrest chemokines. Microcirculation, 10 (3-4), 289-295, 2003.
MACKAY, C.R.; Chemokines: immunology's high impact factors. Nat Immunol, 2 (2), 95-101, 2001.
MALLOY, J.; MCCAIG, L.; VELDHUIZEN, R.; YAO, L.J.; JOSEPH, M.; WHITSETT, J. & LEWIS, J.; Alterations of the endogenous surfactant system in septic adult rats. Am J Respir Crit Care Med, 156 (2 Pt 1), 617-623, 1997.
MARTIN, G.S.; MANNINO, D.M.; EATON, S. & MOSS, M.; The epidemiology of sepsis in the United States from 1979 through 2000. N Engl J Med, 348 (16), 1546-1554, 2003.
MATSUKAWA, A.; HOGABOAM, C.M.; LUKACS, N.W. & KUNKEL, S.L.; Chemokines and innate immunity. Rev Immunogenet, 2 (3), 339-358, 2000.
MATUTE-BELLO, G.; LILES, W.C.; RADELLA, F., 2ND; STEINBERG, K.P.; RUZINSKI, J.T.; JONAS, M.; CHI, E.Y.; HUDSON, L.D. & MARTIN, T.R.; Neutrophil apoptosis in the acute respiratory distress syndrome. Am J Respir Crit Care Med, 156 (6), 1969-1977, 1997.
MAZZONI, A. & SEGAL, D.M.; Controlling the Toll road to dendritic cell polarization. J Leukoc Biol, 75 (5), 721-730, 2004.
MEERSSEMAN, W.; LAGROU, K.; MAERTENS, J. & VAN WIJNGAERDEN, E.; Invasive aspergillosis in the intensive care unit. Clin Infect Dis, 45 (2), 205-216, 2007.
MEHRAD, B.; MOORE, T.A. & STANDIFORD, T.J.; Macrophage inflammatory protein-1 alpha is a critical mediator of host defense against invasive pulmonary aspergillosis in neutropenic hosts. J Immunol, 165 (2), 962-968, 2000.
MEHRAD, B.; STRIETER, R.M. & STANDIFORD, T.J.; Role of TNF-alpha in pulmonary host defense in murine invasive aspergillosis. J Immunol, 162 (3), 1633-1640, 1999.
MONNERET, G.; DEBARD, A.L.; VENET, F.; BOHE, J.; HEQUET, O.; BIENVENU, J. & LEPAPE, A.; Marked elevation of human circulating CD4+CD25+ regulatory T cells in sepsis-induced immunoparalysis. Crit Care Med, 31 (7), 2068-2071, 2003.
MONTAGNOLI, C.; BACCI, A.; BOZZA, S.; GAZIANO, R.; MOSCI, P.; SHARPE, A.H. & ROMANI, L.; B7/CD28-dependent CD4+CD25+ regulatory T cells are essential components of the memory-protective immunity to Candida albicans. J Immunol, 169 (11), 6298-6308, 2002.

77
MORRISON, B.E.; PARK, S.J.; MOONEY, J.M. & MEHRAD, B.; Chemokine-mediated recruitment of NK cells is a critical host defense mechanism in invasive aspergillosis. J Clin Invest, 112 (12), 1862-1870, 2003.
MOSER, B. & LOETSCHER, P.; Lymphocyte traffic control by chemokines. Nat Immunol, 2 (2), 123-128, 2001.
MUSTARD, R.A.; BOHNEN, J.M.; ROSATI, C. & SCHOUTEN, B.D.; Pneumonia complicating abdominal sepsis. An independent risk factor for mortality. Arch Surg, 126 (2), 170-175, 1991.
NAGANO, H. & TILNEY, N.L.; Chronic allograft failure: the clinical problem. Am J Med Sci, 313 (5), 305-309, 1997.
NESS, T.L.; EWING, J.L.; HOGABOAM, C.M. & KUNKEL, S.L.; CCR4 is a key modulator of innate immune responses. J Immunol, 177 (11), 7531-7539, 2006.
O'BRIEN, J.M., JR.; ALI, N.A.; ABEREGG, S.K. & ABRAHAM, E.; Sepsis. Am J Med, 120 (12), 1012-1022, 2007.
OLSON, T.S. & LEY, K.; Chemokines and chemokine receptors in leukocyte trafficking. Am J Physiol Regul Integr Comp Physiol, 283 (1), R7-28, 2002.
ONO, S.J.; NAKAMURA, T.; MIYAZAKI, D.; OHBAYASHI, M.; DAWSON, M. & TODA, M.; Chemokines: roles in leukocyte development, trafficking, and effector function. J Allergy Clin Immunol, 111 (6), 1185-1199; quiz 1200, 2003.
OPENSHAW, P.J.; DEAN, G.S. & CULLEY, F.J.; Links between respiratory syncytial virus bronchiolitis and childhood asthma: clinical and research approaches. Pediatr Infect Dis J, 22 (2 Suppl), S58-64; discussion S64-55, 2003.
OSUCHOWSKI, M.F.; WELCH, K.; SIDDIQUI, J. & REMICK, D.G.; Circulating cytokine/inhibitor profiles reshape the understanding of the SIRS/CARS continuum in sepsis and predict mortality. J Immunol, 177 (3), 1967-1974, 2006.
PERL, T.; L DVORAK ; HWANG, T. & WENZEL, R.; Long-term survival and function after suspected Gram-negative sepsis. JAMA, 274 (4), 338-345, 1995.
PLITAS, G.; BURT, B.M.; NGUYEN, H.M.; BAMBOAT, Z.M. & DEMATTEO, R.P.; Toll-like receptor 9 inhibition reduces mortality in polymicrobial sepsis. J Exp Med, 205 (6), 1277-1283, 2008.
QUARTIN, A.A.; SCHEIN, R.M.; KETT, D.H. & PEDUZZI, P.N.; Magnitude and duration of the effect of sepsis on survival. Department of Veterans Affairs Systemic Sepsis Cooperative Studies Group. Jama, 277 (13), 1058-1063, 1997.

78
RICHARDSON, J.D.; DECAMP, M.M.; GARRISON, R.N. & FRY, D.E.; Pulmonary infection complicating intra-abdominal sepsis: clinical and experimental observations. Ann Surg, 195 (6), 732-738, 1982.
RIEDEMANN, N.C.; GUO, R.F. & WARD, P.A.; Novel strategies for the treatment of sepsis. Nat Med, 9 (5), 517-524, 2003.
ROCCO, P.R.; NEGRI, E.M.; KURTZ, P.M.; VASCONCELLOS, F.P.; SILVA, G.H.; CAPELOZZI, V.L.; ROMERO, P.V. & ZIN, W.A.; Lung tissue mechanics and extracellular matrix remodeling in acute lung injury. Am J Respir Crit Care Med, 164 (6), 1067-1071, 2001.
RODGERS, G.L.; MORTENSEN, J.; FISHER, M.C.; LO, A.; CRESSWELL, A. & LONG, S.S.; Predictors of infectious complications after burn injuries in children. Pediatr Infect Dis J, 19 (10), 990-995, 2000.
ROSENKILDE, M.M. & SCHWARTZ, T.W.; The chemokine system -- a major regulator of angiogenesis in health and disease. Apmis, 112 (7-8), 481-495, 2004.
ROSSI, D. & ZLOTNIK, A.; The biology of chemokines and their receptors. Annu Rev Immunol, 18, 217-242, 2000.
RUDENSKY, A.Y. & CAMPBELL, D.J.; In vivo sites and cellular mechanisms of T reg cell-mediated suppression. J Exp Med, 203 (3), 489-492, 2006.
SAKAGUCHI, S.; Control of immune responses by naturally arising CD4+ regulatory T cells that express toll-like receptors. J Exp Med, 197 (4), 397-401, 2003.
SCHNEIDER, M.A.; MEINGASSNER, J.G.; LIPP, M.; MOORE, H.D. & ROT, A.; CCR7 is required for the in vivo function of CD4+ CD25+ regulatory T cells. J Exp Med, 204 (4), 735-745, 2007.
SCHUH, J.M.; POWER, C.A.; PROUDFOOT, A.E.; KUNKEL, S.L.; LUKACS, N.W. & HOGABOAM, C.M.; Airway hyperresponsiveness, but not airway remodeling, is attenuated during chronic pulmonary allergic responses to Aspergillus in CCR4-/- mice. Faseb J, 16 (10), 1313-1315, 2002.
SCUMPIA, P.O.; DELANO, M.J.; KELLY, K.M.; O'MALLEY, K.A.; EFRON, P.A.; MCAULIFFE, P.F.; BRUSKO, T.; UNGARO, R.; BARKER, T.; WYNN, J.L.; ATKINSON, M.A.; REEVES, W.H.; SALZLER, M.J. & MOLDAWER, L.L.; Increased natural CD4+CD25+ regulatory T cells and their suppressor activity do not contribute to mortality in murine polymicrobial sepsis. J Immunol, 177 (11), 7943-7949, 2006.
SILVA, E.; PEDRO MDE, A.; SOGAYAR, A.C.; MOHOVIC, T.; SILVA, C.L.; JANISZEWSKI, M.; CAL, R.G.; DE SOUSA, E.F.; ABE, T.P.; DE ANDRADE, J.; DE MATOS, J.D.; REZENDE, E.; ASSUNCAO, M.; AVEZUM, A.; ROCHA, P.C.; DE MATOS, G.F.; BENTO, A.M.; CORREA, A.D.; VIEIRA, P.C. & KNOBEL, E.; Brazilian Sepsis Epidemiological Study (BASES study). Crit Care, 8 (4), R251-260, 2004.

79
SPEYER, C.L.; GAO, H.; RANCILIO, N.J.; NEFF, T.A.; HUFFNAGLE, G.B.; SARMA, J.V. & WARD, P.A.; Novel chemokine responsiveness and mobilization of neutrophils during sepsis. Am J Pathol, 165 (6), 2187-2196, 2004.
STEINHAUSER, M.L.; HOGABOAM, C.M.; KUNKEL, S.L.; LUKACS, N.W.; STRIETER, R.M. & STANDIFORD, T.J.; IL-10 is a major mediator of sepsis-induced impairment in lung antibacterial host defense. J Immunol, 162 (1), 392-399., 1999.
SUFFIA, I.J.; RECKLING, S.K.; PICCIRILLO, C.A.; GOLDSZMID, R.S. & BELKAID, Y.; Infected site-restricted Foxp3+ natural regulatory T cells are specific for microbial antigens. J Exp Med, 203 (3), 777-788, 2006.
SUVAS, S.; AZKUR, A.K.; KIM, B.S.; KUMARAGURU, U. & ROUSE, B.T.; CD4+CD25+ regulatory T cells control the severity of viral immunoinflammatory lesions. J Immunol, 172 (7), 4123-4132, 2004.
TAVARES-MURTA, B.M.; ZAPAROLI, M.; FERREIRA, R.B.; SILVA-VERGARA, M.L.; OLIVEIRA, C.H.; MURTA, E.F.; FERREIRA, S.H. & CUNHA, F.Q.; Failure of neutrophil chemotactic function in septic patients. Crit Care Med, 30 (5), 1056-1061, 2002.
TAYLOR, M.D.; HARRIS, A.; BABAYAN, S.A.; BAIN, O.; CULSHAW, A.; ALLEN, J.E. & MAIZELS, R.M.; CTLA-4 and CD4+ CD25+ regulatory T cells inhibit protective immunity to filarial parasites in vivo. J Immunol, 179 (7), 4626-4634, 2007.
TELES, J.M.; SILVA, E.; WESTPHAL, G.; FILHO, R.C. & MACHADO, F.R.; Surviving sepsis campaign in Brazil. Shock, 30 Suppl 1, 47-52, 2008.
TERRITO, M.C. & GOLDE, D.W.; Granulocyte function in experimental human endotoxemia. Blood, 47 (4), 539-544, 1976.
UCCINI, S.; RUCO, L.P.; MONARDO, F.; LA PAROLA, I.L.; CERIMELE, D. & BARONI, C.D.; Molecular mechanisms involved in intraepithelial lymphocyte migration: a comparative study in skin and tonsil. J Pathol, 169 (4), 413-419, 1993.
VENET, F.; PACHOT, A.; DEBARD, A.L.; BOHE, J.; BIENVENU, J.; LEPAPE, A. & MONNERET, G.; Increased percentage of CD4+CD25+ regulatory T cells during septic shock is due to the decrease of CD4+CD25- lymphocytes. Crit Care Med, 32 (11), 2329-2331, 2004.
VERDRENGH, M. & TARKOWSKI, A.; Role of neutrophils in experimental septicemia and septic arthritis induced by Staphylococcus aureus. Infect Immun, 65 (7), 2517-2521, 1997.
VON ANDRIAN, U.H. & MACKAY, C.R.; T-cell function and migration. Two sides of the same coin. N Engl J Med, 343 (14), 1020-1034, 2000.
VON BOEHMER, H.; Mechanisms of suppression by suppressor T cells. Nat Immunol, 6 (4), 338-344, 2005.

80
WALKER, L.S.; CD4+ CD25+ Treg: divide and rule? Immunology, 111 (2), 129-137, 2004.
WALKER, W. & ROTONDO, D.; Prostaglandin E2 is a potent regulator of interleukin-12- and interleukin-18-induced natural killer cell interferon-gamma synthesis. Immunology, 111 (3), 298-305, 2004.
WALLEY, K.R.; LUKACS, N.W.; STANDIFORD, T.J.; STRIETER, R.M. & KUNKEL, S.L.; Elevated levels of macrophage inflammatory protein 2 in severe murine peritonitis increase neutrophil recruitment and mortality. Infect Immun, 65 (9), 3847-3851., 1997.
WANG, T.S. & DENG, J.C.; Molecular and cellular aspects of sepsis-induced immunosuppression. J Mol Med, 2008.
WYSOCKI, C.A.; JIANG, Q.; PANOSKALTSIS-MORTARI, A.; TAYLOR, P.A.; MCKINNON, K.P.; SU, L.; BLAZAR, B.R. & SERODY, J.S.; Critical role for CCR5 in the function of donor CD4+CD25+ regulatory T cells during acute graft-versus-host disease. Blood, 106 (9), 3300-3307, 2005.
XIE, G.H.; FANG, X.M.; FANG, Q.; WU, X.M.; JIN, Y.H.; WANG, J.L.; GUO, Q.L.; GU, M.N.; XU, Q.P.; WANG, D.X.; YAO, S.L.; YUAN, S.Y.; DU, Z.H.; SUN, Y.B.; WANG, H.H.; WU, S.J. & CHENG, B.L.; Impact of invasive fungal infection on outcomes of severe sepsis: a multicenter matched cohort study in critically ill surgical patients. Crit Care, 12 (1), R5, 2008.
YI, H.; ZHEN, Y.; JIANG, L.; ZHENG, J. & ZHAO, Y.; The phenotypic characterization of naturally occurring regulatory CD4+CD25+ T cells. Cell Mol Immunol, 3 (3), 189-195, 2006.
ZAKRZEWSKI, J.L.; KOCHMAN, A.A.; LU, S.X.; TERWEY, T.H.; KIM, T.D.; HUBBARD, V.M.; MURIGLAN, S.J.; SUH, D.; SMITH, O.M.; GRUBIN, J.; PATEL, N.; CHOW, A.; CABRERA-PEREZ, J.; RADHAKRISHNAN, R.; DIAB, A.; PERALES, M.A.; RIZZUTO, G.; MENET, E.; PAMER, E.G.; HELLER, G.; ZUNIGA-PFLUCKER, J.C.; ALPDOGAN, O. & VAN DEN BRINK, M.R.; Adoptive transfer of T-cell precursors enhances T-cell reconstitution after allogeneic hematopoietic stem cell transplantation. Nat Med, 12 (9), 1039-1047, 2006.
ZOU, L.; BARNETT, B.; SAFAH, H.; LARUSSA, V.F.; EVDEMON-HOGAN, M.; MOTTRAM, P.; WEI, S.; DAVID, O.; CURIEL, T.J. & ZOU, W.; Bone marrow is a reservoir for CD4+CD25+ regulatory T cells that traffic through CXCL12/CXCR4 signals. Cancer Res, 64 (22), 8451-8455, 2004.

81
8. ARTIGO I:

82
ATLa, an Aspirin-Triggered Lipoxin A4 Synthetic Analog, Prevents the
Inflammatory and Fibrotic Effects of Bleomycin-Induced Pulmonary Fibrosis1
Vanessa Martins*, Samuel S. Valença†, Francisco A. Farias-Filho‡, Raphael
Molinaro Coelho*, Rafael Loureiro Simões§, Tatiana Paula Teixeira Ferreira‡, Patrícia
M. R. e Silva‡, Cory M. Hogaboam||, Steven L. Kunkel||, Iolanda M. Fierro§, Claudio
Canetti2¶, and Claudia F. Benjamim2,3*
*Departamento de Farmacologia Básica e Clínica-ICB and ¶Instituto de Biofísica Carlos
Chagas Filho, Universidade Federal do Rio de Janeiro, and †Departamento de Histologia e
Embriologia and §Departamento de Farmacologia e Psicobiologia, IBRAG, Universidade do Estado do
Rio de Janeiro, and ‡Laboratório de Inflamação, Instituto Oswaldo Cruz, FIOCRUZ, Rio de Janeiro,
Brazil. ||Department of Pathology, University of Michigan, Ann Arbor, MI, USA.
Running title: Reversal of Lung Fibrosis by Lipoxin Analog ATLa
Keywords: Lipoxin; fibrosis; inflammation
Footnotes:
1 This work was supported by Conselho Nacional de Pesquisa (CNPq),
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and Fundação
Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ).
2 C.C. and C.F.B. contributed equally to this work.

83
Dear Prof. Benjamim:
Your manuscript titled "ATLa, an Aspirin-Triggered Lipoxin A4 Synthetic
Analog, Prevents the Inflammatory and Fibrotic Effects of Bleomycin-Induced
Pulmonary Fibrosis" is acceptable for publication in The Journal of Immunology
contingent upon revision.
Please see the comments below, which contain a summary of the concerns
that were raised by our referees. Please provide a point-by-point reply that indicates
how you have revised your manuscript to address each of these comments.
Should you decide not to revise your manuscript in response to a specific
comment, please justify your decision in detail. It is important that you address each
of these issues, either through revision or rebuttal, in order to increase the likelihood
the manuscript can be published by The Journal of Immunology. A thorough and
reasoned response to the referees' comments will help to complete the review
process as quickly as possible. Please note a revised manuscript not returned within
nine months of the date of the decision letter will be considered as NEW and will be
subject to a complete review.
Upon resubmission, you will need to upload a Word file of the manuscript,
marked to indicate changes. Please use yellow highlighting or red-colored font to
mark changes. Do not show deletions, because if the manuscript is accepted, this
version will be immediately sent for publication. You will also need to upload
publication-quality figures, preferably in TIFF or EPS format. Please check your
manuscript carefully, especially for the common errors in the checklists below. Please
be advised that changes at page-proof stage, other than correcting errors introduced
at copy-editing, may result in the manuscript needing to be returned to the editors
and thus delay publication.
Thank you for submitting your manuscript to The Journal of Immunology.
Sincerely,
Jeremy M. Boss, Ph.D.

84
Editor-in-Chief, The Journal of Immunology
Comments from Reviewer #2:
The manuscript by Martins et al is much improved. I have only the following
minor points to raise:
1. I suggest that the ALX Figure in the response to my comments be included
in the manuscript or as supplemental data. The authors should incorporate these
findings into the Results and Discussion sections.
2. The authors' state findings on CD8 and NK 1.1 positive cells in new results
but these data are not included in the revised manuscript. Either the numbers should
be provided in the Results section or a new Figure provided. Ok not to show the
dendritic cell data that was unchanged with ATLa.
3. The authors have changed ATL-1 to ATLa in the text and Figure legends,
but the Figures still show only ATL-1. This should also be changed to ATLa to match
the text.
4. I think the paragraph crafted in response to query 5 is appropriate for
inclusion in the Discussion (as written in the revised text).
5. There are still numerous problems with grammatical errors throughout the
manuscript.
Comments from the Deputy Editor:
Important work. The comments can be dealt with by text modifications. Please
have the paper edited by a native English speaker.
Livros Grátis( http://www.livrosgratis.com.br )
Milhares de Livros para Download: Baixar livros de AdministraçãoBaixar livros de AgronomiaBaixar livros de ArquiteturaBaixar livros de ArtesBaixar livros de AstronomiaBaixar livros de Biologia GeralBaixar livros de Ciência da ComputaçãoBaixar livros de Ciência da InformaçãoBaixar livros de Ciência PolíticaBaixar livros de Ciências da SaúdeBaixar livros de ComunicaçãoBaixar livros do Conselho Nacional de Educação - CNEBaixar livros de Defesa civilBaixar livros de DireitoBaixar livros de Direitos humanosBaixar livros de EconomiaBaixar livros de Economia DomésticaBaixar livros de EducaçãoBaixar livros de Educação - TrânsitoBaixar livros de Educação FísicaBaixar livros de Engenharia AeroespacialBaixar livros de FarmáciaBaixar livros de FilosofiaBaixar livros de FísicaBaixar livros de GeociênciasBaixar livros de GeografiaBaixar livros de HistóriaBaixar livros de Línguas
Baixar livros de LiteraturaBaixar livros de Literatura de CordelBaixar livros de Literatura InfantilBaixar livros de MatemáticaBaixar livros de MedicinaBaixar livros de Medicina VeterináriaBaixar livros de Meio AmbienteBaixar livros de MeteorologiaBaixar Monografias e TCCBaixar livros MultidisciplinarBaixar livros de MúsicaBaixar livros de PsicologiaBaixar livros de QuímicaBaixar livros de Saúde ColetivaBaixar livros de Serviço SocialBaixar livros de SociologiaBaixar livros de TeologiaBaixar livros de TrabalhoBaixar livros de Turismo