Embed Size (px)
Citation preview

REGULACION HORMONAL DE LA EXPRESIONDEL GEN QUE CODIFICA A LA ENZIMA
α2,6-SIALILTRANSFERASA QUE SIALIZA A FSHEN HIPOFISIS DE RATA HEMBRA
PRESENTA
ABRIL SAINT MARTIN CASTELLANOS
Para obtener el grado de
LICENCIADA EN BIOLOGIA EXPERIMENTAL
ASESOR: DR. PABLO DAMIAN MATZUMURA
DEPTO. BIOLOGIA DE LA REPRODUCCIONLaboratorio de Endocrinología Molecular
Presentado en mayo del 2002

1
REGULACIÓN HORMONAL DE LA EXPRESIÓN DEL GEN QUE CODIFICA A LA ENZIMA
α2,6-SIALILTRANSFERASA QUE SIALIZA A FSH EN HIPÓFISIS DE RATA HEMBRA.
I. INTRODUCCIÓN
A. LA IMPORTANCIA DEL ÁCIDO SIÁLICO
El término de ácido siálico se utiliza para nombrar a los miembros de la familia de amino azúcares (monosacáridos) de naturaleza ácida , derivados del ácido neuroamínico, que se encuentran presentes como los residuos terminales de las cadenas de carbohidratos de gliconjugados (glicoproteínas y glicolípidos) involucrados en muchos fenómenos biológicos y patológicos presentes en la mayoría de las células, secreciones, fluidos y tejidos de mamíferos. El ácido N-acetilneuroa mínico (NANA por sus siglas en inglés) es el miembro de la familia más abundante, por lo que comunmente se refiere a éste como el único integrante, por lo que en este texto el término de ácido siálico se empleará para nombrar a todos los miembros posibles de esta familia. La estructura particular de este monosacárido es usada para activar o inhibir interacciones intermoleculares e intercelulares por su virtual carga negativa a pH fisiológico, sin embargo, el ácido siálico puede actuar también como un componente crítico en el reconocimiento de ligandos de una variedad de proteínas animales, vegetales y de origen microbiano (ácido siálico unido a lectinas). El reconocimiento se puede afectar por variaciones estructurales específicas y modificaciones de la estructura del ácido siálico, sus enlaces a una cadena de carbohidratos fundamental, la estructura de estas cadenas y la naturaleza de los glicoconjugados a los cuales está unido (Kelm y Schauer, 1997; Schauer, 2000). La gran diversidad funcional del ácido siálico es expuesta por una gran cantidad de organismos como los virus, las bacterias y sus toxinas, la totalidad de las células eucariontes, principalmente en funciones de reconocimiento célula-célula a través de sialoglicoproteínas y sialoglicolípidos que le confieren alta especificidad en el reconocimiento y la unión a receptores celulares (Sillanaukee y cols., 1999) . Las glicoproteínas sializadas se encuentran en gran abundanc ia en las membranas plasmáticas y siempre se ubican de tal manera que los grupos de carbohidratos se encuentren únicamente en la superficie externa de la membrana celular, func ionando como sitios de reconocimiento de receptores membranales, en los cuales el ácido siálico es el último monosacárido adicionado a la cadena de oligosacáridos, por lo que funciona como sitio de contacto y reconocimiento (Varki, 1997; Martin y cols., 1998; Schauer, 2000). En lo que se refiere al sistema inmune, el ácido siálico juga un papel fundamental en la regulación del sistema puesto que los “Siglecs” (receptores, miembros de la superfamilia de las inmunoglobulinas que se unen al ácido siálico en forma altamente específica) forman parte de las moléculas de adhesión que permiten el reclutamiento de neutrófilos, eosinófisos y algunas subclases de linfocitos T a los sitios de inflamación e interacción entre células B y células B y con células T (Crocker y Varki, 2001, a y b). Además, estos sialoconjugados juegan un papel crucial en la interacción entre diferentes virus y bacterias con los receptores

2
sializados de las células huésped, así como la inducción de la proliferación de linfocitos T periféricos (Varki, 1997). Debido a que el cerebro y el sistema nervioso central contiene n un gran número de compuestos sializados, se ha propuesto que el ácido siálico tiene un papel fundamental en el desarrollo embrionario, en la infancia y el desarrollo de sus funciones. Además se ha encontrado que algunos compuestos sializados pueden inhibir la adhesión de toxinas, bacterias o virus a receptores en la superficie de células epiteliales y que tienen efectos que promueven el crecimiento de bifidobacterias y lactobacilos predominantemente presentes en la flora intestinal de infantes alimentados con leche materna , la cual tiene gran cantidad de compuestos sializados (oligosacáridos, glicolípidos y glicoproteínas) ausentes en la lecha de vaca, lo cual le confiere al ácido siálico un probable efecto protector contra infecciones entéricas (Mc Veagh y Miller, 1997; Nakano y cols., 2001). El ácido siálico juega un papel indispensable para la protección y adaptación de la vida, sin embargo, muchos aspectos de la regulación de su metabolismo a nivel enzimático y gé nico, así como sus funciones siguen siendo un misterio (Schauer, 2000).
B. PAPEL DE LOS CARBOHIDRATOS EN LA FUNCIÓN DE LA FSH
La hormona estimulante de los folículos (FSH) pertenece a la familia de hormonas glicoproteícas
constituida por la hormona luteinizante (LH), la hormona estimulante de los tirotropos (TSH), todas de origen hipofisiario, y la gonadotropina coriónica human (hCG) de origen placentario. Las gonadotropinas hipofisiarias FSH y LH son sintetizadas en los gonadotropos de la hipófisis anterior y sus vidas media plasmáticas son cortas, siendo de aproximadamente 110 minutos para la primera y de <20 minutos para la segunda en la rata (Gharib y cols., 1990). Estas hormonas interaccionan selectivamente con sus respectivos receptores presentes en células de las gónadas de machos y hembras, interviniendo principalmente en la esteroidogénesis, en la gametogénesis y la inducción de la ovulación (Lambalk y cols., 2001).
En el humano, la FSH estimula el crecimiento testicular y facilita el desarrollo de la espermatogénesis a través de la inducción de la síntesis de la proteína que une andrógenos por las células de Sertoli; mientras que es la responsable del crecimiento y el desarrollo de los folículos ováricos en la mujer, así como de la estimulación de la producción de estrógenos por los folículos en maduración (células de la granulosa). Por otro lado, la LH estimula la síntesis de testosterona por las células intersticiales de los testículos (células de Leydig) y en la mujer induce la maduración final de los folículos ováricos , induce la luteinización, la formación del cuerpo lúteo y la secreción de progesterona y, en conjunto con la FSH, estimula la síntesis de estrógenos (Lambalk y cols., 2001). La FSH estimula específicamente la activación del sistema de enzimas que catalizan la conversión de la testosterona en estrógenos, las aromatasas, y también estimula la producción de las enzimas relacionadas con la activación del plasminógeno, enzima clave en el rompimiento folicular durante la ovulación (Galway y cols., 1990b). Se pensaba que la LH era la única responsable de iniciar los procesos ovulatorios, sin embargo se ha observado que la FSH puede inducir por sí misma el desarrollo folicular y la ovulación (Galway y cols., 1990 a y b). La FSH tiene un peso molecular aproximado de 30,000 daltones y está formada por dos cadenas polipeptídicas glicosiladas que son codificadas por dos genes diferentes (Bousfield y cols., 1994). Estas cadenas están unidas por enlaces no covalentes que estabilizan la estructura terciaria de la molécula, además , cada subunidad contiene dos cadenas de oligosacáridos del tipo complejo o ramificado, cada una ligada covalentemente al grupo amino de la cadena lateral de la asparagina (Asn). La subunidad alfa (α-FSH) de la rata está constituida por 96 aminoácidos, mientras que la subunidad beta (β-FSH) por 111 aminoácidos . La subunidad α se transcribe a partir de un mismo gen en diferentes tipos

3
celulares y el péptido es idéntico para las cuatro hormonas glicoproteicas (α-FSH/LH/TSH/hCG), siendo específico para cada especie. Por otro lado, la subunidad β es específica para cada hormona (β-FSH, β-LH, β-TSH, β-CG) y también varía de especie a especie. Esta propiedad le confiere a cada hormona características inmunológicas y biológicas únicas sobre sus células blanco. En el caso de la FSH de rata , ambas subunidades presentan dos sitios de N-glicosilación, la subunidad α en los residuos 56 y 82 (Asn56 y Asn82) y la ß-FSH en Asn7 y en Asn24. Al igual que la humana, los oligosacáridos de la FSH de rata terminan en su mayoría en ácido siálico, mientras que la FSH ovina y la bovina tienen terminaciones mixtas de ácido siálico y de sulfatos (Green y Baenziger, 1988a y b; Bousfield y cols., 1994 y 2000).
A pesar de que la LH y la FSH de humano presentan una subunidad α común y de que sus subunidades ß tienen un cierto grado de homología, sus oligosacáridos difieren en sus carbohidratos terminales, mientras que la LH tiene terminaciones mayormente sulfatadas, la FSH presenta terminaciones principalemente sializadas (Baenziger y Green, 1988; Green y Baenziger, 1988a; Bousfield y cols., 1994 y 2000). La gran variedad de estructuras de carbohidratos presentes en la FSH y LH indican que la síntesis de oligosacáridos sulfatados y sializados está regulada y que podría tener importancia funcional (Smith y cols., 1990). La presencia de cantidades variables de residuos terminales de carbohidratos con carga eléctrica negativa en la FSH le confiere características aún más especiales ya que la misma hormona puede presentar diversas formas moleculares , denominadas isoformas o isohormonas de FSH (Chappel, 1995), característica que ha sido empleada para separarlas con base en su carga (Ulloa-Aguirre y cols., 1988 y 1995). Para la β-FSH, los carbohidratos son necesarios para la conformación de la hormona plegada que faciliten el apareamiento adecuado de los puentes disulfuro (Stockell Hartree y Renwick, 1992), para la determinación de su vida media plasmática y la correcta interacción con su receptor (Bishop y cols., 1994). Una de las funciones menos estudiadas del ácido siálico, que será explorada en esta ocasión, es su intervención en la regulación de la actividad de glicohormonas como es el caso de la hormona estimulante de los folículos (FSH), de origen hipofisiario, en donde los residuos de ácido siálico unidos a sus subunidades son críticos para el ensamblaje del dímero, integridad y secreción, asimismo en la transducción de señales, así como en la dimerización y secreción, de igual forma juegan un papel crucial en eliminación del dímero de la circulación (Ulloa-Aguirre y cols., 1999; Padmanabhan y cols., 1991).
C. PROCESO DE GLICOSILACIÓN DE LA FSH
Como muchas otras glicoproteínas de secreción, las subunidades α-FSH y ß-FSH son transcritas a partir de diferentes genes y traducidas en los ribosomas que se encuentran asociados a la membrana del retículo endoplásmico, lugar donde se ensamblan las subunidades y se inicia el proceso de maduración post-traduccional por glicosilación (Hirschberg y Snider, 1987; Gharib y cols., 1990; Verbert y Cacan, 1999).
La glicosilación de la FSH involucra la adición y maduración de sus cadenas de carbohidratos , la cual se realiza solamente por uniones covalentes del tipo N-ligado, en donde los carbohidratos se adicionan en el grupo amino de la asparagina, a diferencia del tipo O-ligado, donde los carbohidratos de adicionan en grupos hidroxilo de la serina o la treonina y, en algunos casos en el grupo hidroxilo de la hidroxilisina o hidroxiprolina. La fase inicial de la glicosilación de la FSH ocurre en el retículo endoplásmico, donde una estructura única de oligosacáridos (núcleo de oligosacáridos) es activada por un lípido transportador denominado dolicol fosfato para ser adicionada al grupo amino radical de las asparaginas con secuencia N-Asn-[X]-(Ser/Thr)-C, donde X es cualquier aminoácido excepto prolina, proceso que se conoce como glicosilación nuclear. Este núcleo de oligosacáridos consiste en dos unidades de N acetil glucosamina (GlcNAc), tres unidades de glucosa (Glu) y nueve unidades de manosa (Man), siendo la GlcNAc el azúcar que se une directamente a la asparagina (Verbert y Cacan, 1999; Becker y cols, 2000)

4
Posteriormente, en el aparato de Golgi se realiza la modificación f inal mediante la adición y remoción de los carbohidratos necesaria para ser clasificadas como proteína de secreción. Durante la maduración de sus oligosacáridos, la FSH se va trasladando de la cisterna cis hacia la cisterna media y finalmente a la cisterna trans por medio de un transporte vesicular, secuencia de eventos que se denomina glicosilación terminal. Dentro del proceso de glicosilación que tiene lugar en el complejo de Golgi, dos grupos de enzimas son las más importantes que intervienen en este proceso, las glicosiltransferasas y las glucosintetasas. Las transferasas son el instrumento en la adición de azucares a las glicoproteínas, y las sintetasas catalizan la formación de polisacáridos a partir de azucares de nucleótidos. Estos grupos de enzimas se encuentran distribuidas dentro de las cisternas del complejo de Golgi de manera ordenada, por lo que cada cisterna contiene sus muy distintivas enzimas según el progreso de la glicosilación (Hirschberg y Snider, 1987; Baenziger, 1994). El ensamblado de oligosacáridos terminales a la molécula de FSH en el complejo de Golgi produce parte de la señal distintiva que da la vía de secreción, ya sea secretada constitutivamente hacia el exterior de la célula (no regulada) o almacenada en gránulos de secreción en la red trans-Golgi hasta que un estímulo induzca su secreción (Hirschberg y Snider, 1987; Becker y cols., 2000).
D. PROCESO DE SIALIZACIÓN DE LA FSH
Uno de los procesos terminales de glicosilación más importantes para la FSH es la sialización, en la cual son adicionados monosacáridos del ácido siálico en las posiciones terminales de las cadenas bi, tri y tetra-antenarias de sus oligosacáridos. El ácido siálico posee una importancia especial puesto que es el único residuo de azúcar con una carga negativa neta a pH fisiológico (Lambert y cols., 1998; Ulloa-Aguirre y cols., 1999). Este proceso general de sialización ocurre en la cisterna trans del complejo de Golgi, siendo el último monosacárido adicionado. Las enzimas encargas de esta modificación terminal reciben el nombre de sialiltransferasas , las cuales son miembros de una familia de al menos 15 diferentes enzimas que se encuentran relacionadas entre sí por su función común de transferir ácido siálico a las posiciones terminales de los oligosacáridos, principalmente a los residuos de galactosa, de las glicoproteínas y los glicolípidos, usando como sustrato donador al ácido neuroamín-citosín-monofosfato (CMP-NeuAc) y generando un enlace de alta especificidad (Kitagawa y Paulson, 1994b; Tsuji y cols., 1996; Keppler y cols., 2001) Fig. 1.
FIGURA 1. Adición de ácido siálico terminal a la cadena de oligosacáridos de una glicoproteína en la región trans del aparato de Golgi. 1 Adición de diferentes monosacáridos hasta llegar a la terminación –Gal-NAcGlu. 2 La sialiltransferasa reconoce al nucleótido donador del ácido siálico y a la secuencia aceptora y realiza el enlace en la posición específica (Tomado de Keppler y cols., 2001).

5
Se ha definido la topología de las sialiltransferasas en la membrana de la cisterna trans del complejo de Golgi, describiéndose como proteínas transmembranales del tipo II, que consisten de una región en el extremo amino terminal de 9 aminoácidos hacia el citoplasma (dominio citoplasmático), un dominio de anclaje transmembranal de 17 aminoácidos, un "tallo" de 35 aminoácidos sensible a la proteólisis y un dominio catalítico de longitud variable que se proyecta hacia el lumen del Complejo de Golgi (Kleene y Berger, 1993). Las regiones del amino terminal y del tallo , que pueden ayudar a la enzima a tomar su lugar dentro de la membrana (Colley y cols, 1992), no son esenciales para su actividad catalítica ya que la pérdida de este anclaje convierte a la enzima en soluble pero conserva su actividad (Colley y cols, 1989; Dahdal y Colley, 1993). Todas las sialiltransferasas contienen una región altamente conservada de 48-49 aminoácidos en el centro del dominio catalítico denominada sitio de sialización ("sialilmotif"), la cual participa en la unión del CMP-NeuAc a la cadena de oligosacáridos receptora (Datta y Paulson, 1995). La secuencia de esta región ha sido empleada en la caracterización y el análisis del funcionamiento de otros miembros de esta familia de enzimas (Livingston y Paulson, 1993). Se ha descrito, en algunas sialiltransferasas, otra región de homología, por lo que se ha denominando al primero "L-sialilmotif" (del inglés, “long” por ser la más larga) y a la recién descubierta "S-sialilmotif" (del inglés short, por su menor longitud) que partic ipa en la interacción tanto del donador como del aceptor (Datta y Paulson, 1997; Datta y cols., 1998). Cada sialiltransferasa es altamente específica en el reconocimiento de la secuencia del oligosacárido aceptor (la posición de los últimos dos monosacáridos) y en la formación del enlace anomérico formado entre el ácido siálico y el azúcar al cual es adherido, pudiendo ser enlaces de tipo α o β. El enlace entre los últimos dos carbohidratos aceptores y el tipo de enlace anomérico es lo que da el nombre a cada uno de los miembros de esta familia de sialiltransferasas (Tsuji y cols, 1996). Hasta 1987 solamente se tenía información del ADNc (ADN complementario al ARNm) de la Galß1,4 GlcNAc α2,6-sialiltransferasa (ST6N), (donde: Gal=galactosa; GlcNAc=N-acetilglucosamina), la cual se ais ló y purificó por la técnica de la reacción en cadena de la polimerasa (PCR) empleando oligonucleótidos cebadores cuyas secuencias de bases se sintetizaron a partir de su secuencia de aminoácidos (Weinstein y cols., 1987). En 1992 se logró clonar y expresar el ADNc de una segunda sialiltransferasa, la Galß1,3 GalNAc α2,3-sialiltransferasa (ST3O) (Gillespie y cols., 1992), que es responsable de la síntesis de la secuencia: ácido siálico-α2,3 Galβ1,3 - GalNAc encontrada en oligosacáridos O-ligados, así como también de la secuencia terminal de ciertos ganglósidos. Se observó que esta enzima presenta dos formas derivadas de un procesamiento post-transcripcional alternativo, una de 47-49 KDa y otra de 45 KDa. Ambas isoenzimas encontradas en el hígado de la rata presentan una región de homología de 45 aminoácidos con 65% de identidad (Wen y cols., 1992b). Posteriormente se aisló, en hígado de rata, el ADNc de otra sialiltransferasa, la Galß1,3(4) GlcNAc α2,3-sialiltransferasa (ST3N) (Wen y cols., 1992a), misma que también fue aislada de placenta humana (ST3N, HP-1) (Kitagawa y Paulson, 1993), encontrándose un 91% de homología de nucleótidos y un 97% de aminoácidos entre ambas especies; sin embargo, se informó que la enzima de placenta humana prefiere cadenas aceptoras Galß1,4 GalNAc (Tipo 2), mientras que la ST3N de hígado de rata tiene mayor afinidad tanto por cadenas aceptoras Galß1,3 GlcNAc (Tipo 1) como para cadenas Galß1,3 GalNAc (Tipo 3) (Kitagawa y Paulson, 1994a; Kitagawa y cols., 1996). Estudios de l análisis de l ARNm por la técnica de hibridación por Northern blot reveló que el gen que codifica a la ST3N, HP -1 se expresa en bajas concentraciones en la placenta humana, pero mantiene expresión abundante en el músculo esquelético y otros tejidos fetales humanos (Kitagawa y Paulson, 1994b).

6
En el caso particular de la FSH de rata, la sialización la realizan específicamente dos enzimas, la Gal ß1,3(4) GlcNAc α 2,3-sialiltransferasa (ST3N) y la Gal β1,4 GlcNAc α2,6-sialiltransferasa (ST6N), por el tipo de unión que presentan los residuos de ácido siálico terminales con las galactosas de sus oligosacáridos, formándose las secuencias ÁCIDO SIÁLICO-α2,3 Gal ß1,3(4) GlcNAc y ÁCIDO SIÁLICO-α2,6 Gal ß1,4 GlcNAc encontradas en los complejos terminales de las cadenas de oligosacáridos N-ligados de ambas subunidades de la FSH (Baenziger y Green, 1988; Bousfield y cols., 2000).
E. REGULACIÓN DE LA ACTIVIDAD DE LA FSH
La síntesis y liberación cíclica de la FSH es controlada mediante sistemas de retroalimentación positiva (estimulación) y negativa (inhibición) por la acción de diferentes factores entre los que se encuentran los esteroides sexuales (estrógenos, progestágenos y andrógenos), el péptido hipotalámico denominado hormona liberadora de las gonadotropinas (GnRH) y los péptidos gonadales inhibinas, activinas y folistatina (Gharib y cols., 1990) (Figura 2). Estudios in vivo e in vitro han demostrado que la síntesis y secreción de la FSH y la LH se lleva a cabo en los mismos tipos celulares de la adenohipófisis, los gonadotropos, donde son almacenadas en diferentes gránulos secretores; su secreción se encuentra regulada de forma independiente pero estrechamente coordinada (Haisenleder y cols., 1994; Ulloa-Aguirre y cols., 1995).
La GnRH es un decapéptido sintetizado por neuronas especializadas localizadas en el núcleo arcuato de la sección mediobasal del hipotálamo y el área pre-óptica de la sección anterior del hipotálamo. Este decapéptido regula la síntesis de gonadotropinas adenohipofisiarias, actuando como un neurotransmisor o un neuromodulador el cual, al interaccionar con su receptor membranal (GnRHR), estimula la secreción de gonadotropinas FSH o LH a través de la activación de la transcripción de los genes que codifican para las diferentes subunidades de ambas gonadotropinas en los gonadotropos (Kaiser, 1998); la selectividad de la transactivación de los genes que codifican para cada una de las subunidades de una u otra hormona adenohipofisiaria, va a depender de la frecuencia pulsátil e intensidad del estímulo de GnRH, así como de la capacidad de interacción que éste tendrá con su receptor. La GnRH tiene un papel importante en la regulación de la heterogeneidad de la FSH. Estudios realizados en ratas hembras prepúberes, donde se piensa que la secreción de GnRH se incrementa, mostraron que el patrón de distribución de las isoformas de FSH cambia (Ulloa-Aguirre y cols., 1995). Empleando cultivos celulares de hipófisis de rata suplementados con GnRH se observó que contenían y secretaban mayores cantidades de las isoformas de FSH menos ácida, con respecto a las células control, mientras que el estradiol no modificó dicho patrón de distribución (Ulloa-Aguirre y cols.,1992). Estos resultados son contradictorios con otras especies, especialmente en ovinos, donde el fenómeno es inverso (Padmanabhan y cols.,1997).
Por otro lado, los esteroides gonadales regulan directamente la síntesis y la secreción de gonadotropinas a nivel hipofisiario, modificando la transcripción de los genes que codifican para sus diferentes subunidades, así como también tienen efecto sobre el gen que codifica para el receptor de GnRH. En el hipotálamo, sus efectos son ejercidos a través de la regulación de la función de las neuronas las cuales estimulan directa o indirectamente sobre las neuronas que sintetizan GnRH. (Haisenleder y cols., 1994)
Los estrógenos pueden inducir o inhibir la síntesis de gonadotropinas, dependiendo de su dinámica de secreción y las concentraciones alcanzadas durante el ciclo menstrual. Durante la fase folicular, el incremento progresivo en las concentraciones de estradiol, así como de inhibina A en suero, causan una inhibición de la síntesis y secreción de FSH. Esta retroalimentación negativa es revertida cuando las concentraciones de estradiol se incrementan drásticamente a 200-300 pg/ml en un período de 48 horas, lo que ocurre durante la fase folicular tardía y la fase pre-ovulatoria. En este período de tiempo, el estradiol evoca un efecto de retroalimentación positiva que se muestra como un incremento en la síntesis y secreción de ambas gonadotropinas, particularmente de LH, efecto que es amplificado por el aumento paralelo de las
7
concentraciones de progesterona secretada por células diferenciadas de la granulosa. Durante la fase lútea el estradiol, en conjunto con la progesterona y la inhibina A, suprimen notablemente la síntesis de FSH. Con la involución del cuerpo lúteo, la fuente principal de esteroides sexuales e inhibina en la fase folicular, aumenta la síntesis y secreción de FSH que va a estimular el reclutamiento de nuevos folículos para el siguiente ciclo (Ulloa-Aguirre y Timossi, 2000).
La regulación de la síntesis de FSH por factores gonadales no esteroideos está dada por péptidos gonadales como son las inhibinas, las activinas y la folistatina, donde las inhibinas y la folistatina suprimen la síntesis y secreción de FSH a nivel hipofisiario, mientras que las activinas inducen la síntesis y secreción de FSH (Blumenfeld y Ritter, 2001).
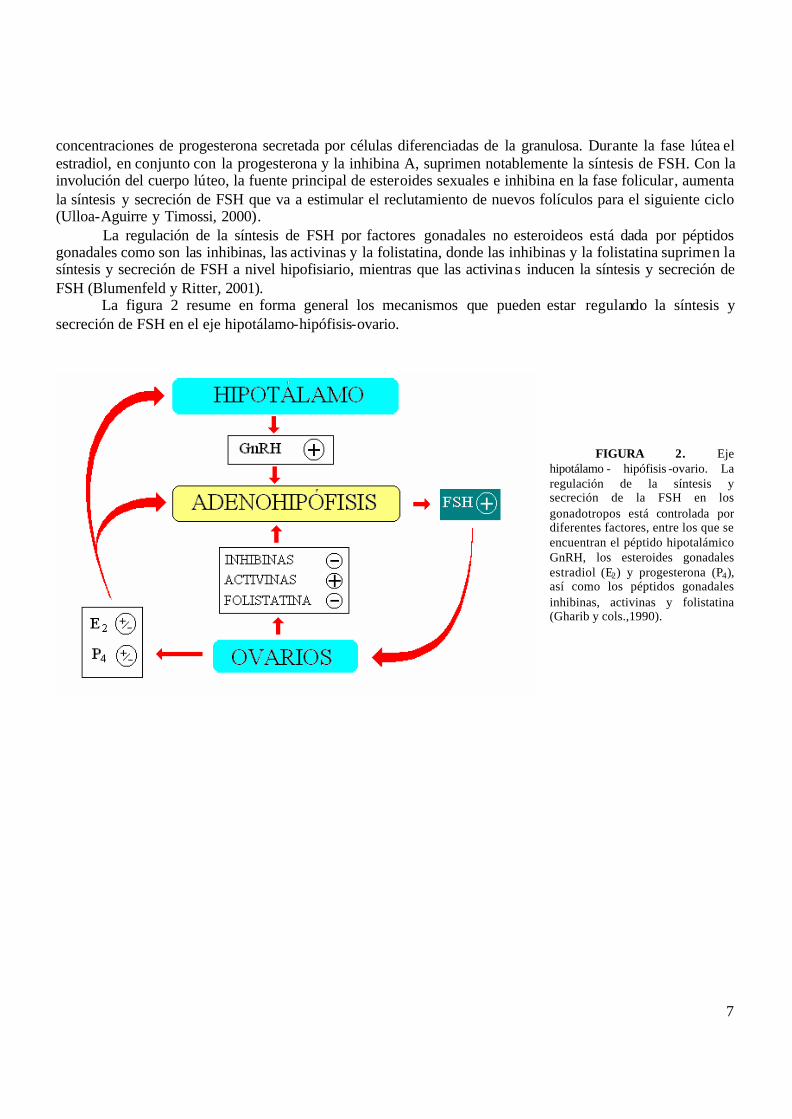
La figura 2 resume en forma general los mecanismos que pueden estar regulando la síntesis y secreción de FSH en el eje hipotálamo-hipófisis-ovario.
FIGURA 2. Eje
hipotálamo - hipófisis -ovario. La regulación de la síntesis y secreción de la FSH en los gonadotropos está controlada por diferentes factores, entre los que se encuentran el péptido hipotalámico GnRH, los esteroides gonadales estradiol (E2) y progesterona (P4), así como los péptidos gonadales inhibinas, activinas y folistatina (Gharib y cols.,1990).

8
II. ANTECEDENTES
El estudio de la regulación hormonal de las enzimas que le adicionan residuos de ácido siálico a la molécula de FSH está sustentada por reportes recientes que demuestran que la expresión de diferentes glicosiltrans ferasas que modifican a hormonas glicoproteicas de la misma familia de la FSH son reguladas hormonalmente. En el caso de las sulfotransferasas, que adicionan grupos SO4 a la LH, son reguladas por estrógenos (Dharmesh y Baenziger, 1993), la expresión de galactosiltransferasas, manosidasas y algunas sialiltransferasas, que modifican los carbohidratos de la TSH estan reguladas por hormonas tiroideas (Helton y Magner, 1994a y b). De igual manera, otras glicosiltransferasas que modifican a la prolactina se encuentran reguladas hormonalemente (Manzella y cols., 1997) y a la tiroglobulina (Vankatesh y Deshpande, 1999).
Se ha propuesto que la sialización confiere importantes propiedades biológicas a la FSH (Ulloa-Aguirre y cols., 2001), por lo cual nos hemos dado la tarea de estudiar a las dos sialiltransferasas reportadas que sializan a esta hormona, la Gal β1,3(4) GlcNAc α 2,3-sialiltransferasa (ST3N) y la Gal β1,4 GlcNAc- α2,6-sialiltransferasa (ST6N). En estudios previos sobre la regulación de la expresión del gen que codifica para la ST3N se demostró una correlación inversa entre la concentración relativa del ARNm de esta enzima y la concentración de estradiol en suero durante el ciclo estral de la rata, observandose que cuando la concentración de estradiol es baja se incrementa la expresión del ARNm de la enzima ST3N y, de manera contraria, conforme aumenta la concentración de estradiol, la expresión del ARNm de la enzima ST3N disminuye. Lo que sugirió que el estradiol puede influir en la síntesis, velocidad de transcripción o estabilidad del ARNm de la ST3N (Damián-Matsumura y cols., 1999).
Para apoyar esta hipótesis se han realizado experimentos en los cuales se demuestra que la administración de benzoato de estradiol (un poderoso agonista del 17β−estradiol) en animales con 28 días de castración, inhibió el incremento en la expresión del ARNm-ST3N en comparación con los animales testigo. Sin embargo, la correlación entre las concentraciones del ARNm-ST3N, la concentración de estradiol en suero y la distribución por carga de las isoformas de FSH fue parcial (Damián-Matsumura y cols., 1999), lo que sugiere la participación concomitante de otra sialiltransferasa, por lo que la ST6N pudiera estar también implicada en la regulación de la actividad de la FSH. III. OBJETIVOS
Objetivo General
Comparar la expresión del ARNm que codifica a las enzimas α2,3-sialiltransferasa y α2,6-sialiltransferasa que adicionan ácido siálico a la molécula de FSH.
Objetivos Particulares • Estandarizar la técnica de Northern Blot para determinar la expresión del ARNm de la ST6N. • Determinar la expresión del ARNm de las enzimas ST6N y ST3N durante las diferentes fases del ciclo estral en animales intactos.

9
• Determinar la expresión del ARNm de las enzimas ST6N y ST3N en animales ovactomizados a tiempo corto (7 días) y un tiempo largo (28 días). • Determinar la expresión del ARNm de las enzimas ST6N y ST3N en animales ovactomizados en tiempos largos e inyectados con benzoato de estradiol (BE).
IV. HIPÓTESIS
Se ha demostrado que la enzima ST3N que adiciona ácido siálico a la FSH, se encuentra regulada estrogenicamente. Si el mecanismo de regulación para la enzima ST6N (la otra enzima que sializa a FSH) es semejante, entonces la expresión del ARNm que codifica para esta enzima se encontrará también regulada estrogénicamente.
V. MÉTODO
A. MODELOS EXPERIMENTALES
Se utilizaron ratas hembras adultas con más de 60 días de edad (250-300g) de la cepa Wistar (Rattus norvergicus, bioterio del Instituto Nacional de Ciencias Médicas y de la Nutrición, Salvador Zubirán, México D.F.). Para todos los experimentos se utilizaron únicamente ratas que presentaron al menos 3 ciclos regulares continuos de 4 días cada uno.
EXPERIMENTO 1. Animales intactos. Para determinar si la expresión de los ARNm de la enzima ST6N se modificaba durante el ciclo estral de la rata, se usaron grupos de 6 ratas, que mostraron 4 ciclos regulares. Estas fueron sacrificadas en cada fase del ciclo estral (proestro, estro, metaestro y diestro) entre las 09:00 y 10:00h. Se obtuvieron las hipófisis y se almacenaron en tubos estériles individualmente a -70°C, así como los sueros de cada animal, para la determinación por RIA de estradiol, FSH y LH.
EXPERIMENTO 2. Ratas castradas (OVX).
Con la finalidad de determinar el efecto de la castración sobre la expresión del ARNm de la ST6N, grupos de 6 ratas se castraron bilateralmente por incisión abdominal bajo anestesia de “eter”, entre las 09:00 y las 10:00 h del día del proestro. Se mantuvieron en el bioterio hasta el día del sacrificio a los 7 y 28 días post-castración. Las hipófisis fueron lavadas con solución salina estéril, removidas y almacenadas a -70°C en tubos individuales, de igual forma los sueros de cada animal se almacenaron individualmente para la determinación de las hormonas estradiol, FSH y LH por RIA.

10
EXPERIMENTO 3. Ratas OVX con Benzoato de estradiol (BE)
Con la finalidad de determinar si altas dosis de estradiol tenían algún efecto sobre la expresión del ARNm-ST6N, animales castrados fueron inyectados por vía subcutanea con 10µg de benzoa to de estradiol (BE; Sigma Co.) en 100µl de aceite de maíz comercial (Mazola), en dosis única a las 10:00h de los días 7 y 28 post-castración y fueron sacrificados a las 48 h. Los grupos control recibieron solame nte el vehículo y se sacrificaron al mismo tiempo. Las hipófisis fueron procesadas en la forma antes descrita para el posterior análisis del ARNm de la ST6N.
B. EXTRACCIÓN DEL ARN El ARN total se extra jo de hipófisis individuales usando el reactivo de TRIZol™ (Gibco BRL, MD) según el método de aislamiento de RNA en un solo paso (Chomczynski y Sacchi, 1987) empleando 0.5 ml del reactivo por hipófisis. Las hipófisis fueron homogeneizadas en los mismos tubos en que fueron almacenadas usando un homogenizador de tejidos (Fisher tipo Politrón) a máxima velocidad, por 10 segundos sobre hielo. El homogenado fue resuspendido en 20 µl de formamida ultra pura (Sigma) y se cuantificó la concentración del ARN de cada muestra por espectrofotometría a 260 nm. Las muestras se guardaron a -70°C hasta el momento de la electroforesis.
C. ANÁLISIS DE LA EXPRESIÓN DEL ARN (Hibridación por "NORTHERN BLOT")
a. Electroforesis de ARN en condiciones desnaturalizantes Los ARN se separaron según su peso molecular mediante una electroforesis en geles de agarosa al 1.0% (Gibco BRL, MD; grado Biología Molecular) disuelto en la solución amortiguadora de MOPS 1X y formaldehido al 7.4% como agente desnaturalizante. Para preparar las muestras se alicuotaron 15 µg de ARN total de cada una, ajustando los volúmenes con ddH2O-DEPC a un volumen final de 10 µl en un tubo Eppendorff de 0.5 ml. A cada muestra, incluyendo el marcador de peso molecular, se le adicionaron 10 µl de la solución desnaturalizante FMF (50% formamida, 1X MOPS, 6.5% formaldehido) y se calentaron a 75°C en un baño de aceite (Boekel Industries Inc., USA) por 5 minutos, seguidos de 5 minutos en hielo y una breve centrifugación a 4°C. Después de desnaturalizar, a cada muestra se les adicionaron 2.0 µl de la solución de carga para ARN (30% glicerol, 0.1% azul de bromofenol y 0.1% xilen cianol) y 0.2 µl de una solución de bromuro de etidio 10 µg/ml. Una vez solidificado el gel se cubrió totalemente con 1X MOPS , para posteriormente depositar las muestras en los pozos del gel. El corrimiento electroforético se efectuó a 75 volts por 1.5 horas.
b. Transferencia por capilaridad (Blotting) Transcurrido el tiempo de la electroforesis (cuando el colorante de bajo peso molecular alcanzó las 2/3 partes de la longitud total del gel), se observaró el gel en el transiluminador, y se montó el dispositivo para la transferencia por capilariadad a una membrana de nylon Z-Probe GT® (Bio Rad, CA) del tamaño del gel. Este proceso de transferencia se realizó por un período de 18 a 24 h usando 10X SSC para inducir el ascenso capilar.

11
Una vez terminada la transferencia, se desmontó el sistema y se marcarón los carriles del gel sobre la membrana, el ARN se fijó a la membrana por entrecruzamiento con luz ultravioleta usando el programa del aparato "UV-crosslinker" (BioRad, CA) que correspondía al tipo de membrana usada. Cada una se guardó a 4°C en una bolsa de plástico sellada con calor para poder ser hibridada posteriormente con la sonda de ADNc correspondiente.
c. Síntesis de las sondas (ADNc) de ST6N y CYC
El ADNc de la enzima α2,6-sialiltransferasa (ADNc-ST6N) usado como sonda, se sintetizó con base en la estructura primaria del ADNc (figura 3) informado previmente (Weinstein y cols., 1987). Para tal efecto se mandaron sintetizar dos oligonucleótidos de 20 bases cada uno (Gibco BRL, MD), cuyas secuencias fueron seleccionadas mediante el programa computacional “Oligos” versión 4.0 de Wojciech Richlijk, donde se tomó en cuenta la temperatura óptima, la adecuada relación de bases GC/AT y se evitó que se formaran enlaces inter e intracatenarios. Las sencuenas de los oligonucléotidos son las siguientes:
5’ –GAG TTA CCG AAG GCT GAA CC- 3’ (oligonucléotido superior o sentido) posiciones 837-856 5’ –CGA ATG TTC CGG AAG CCA GA- 3’ (oligonucléotido inferior o antisentido) posiciones 1206-1186
1 cggtttttga tcatcctgag aaaaatgagc cttggcctcc agacctagtg aagtaacctc 61 tttctcatgg agaacagtgc tggctcctga ggatctggag ggcctgcagc cccagaggga 121 ttagccagaa gcaggcgtgg ttcctgctct gcacagtggc tctcctgtct ggaccattca 181 ttatgattca taccaacttg aagaaaaagt tcagcctctt catcctggtc tttctcctgt 241 tcgcagtcat ctgtgtttgg aagaaaggga gcgactatga ggcccttaca ctgcaagcca 301 aggaattcca gatgcccaag agccaggaga aagtggccat ggggtctgct tcccaggttg 361 tgttctcaaa cagcaagcaa gaccctaagg aagacattcc aatcctcagt taccacaggg 421 tcacagccaa ggtcaaacca cagccttcct tccaggtgtg ggacaaggac tccacatact 481 caaaacttaa ccccaggctg ctgaagatct ggagaaacta tctgaacatg aacaaatata 541 aagtatccta caagggaccg gggccaggag tcaagttcag cgtagaagca ctgcgttgcc 601 accttcgaga ccatgtgaac gtgtctatga tagaggccac agattttccc ttcaacacca 661 ctgagtggga gggttacctg cccaaggaga actttagaac caaggttggg ccttggcaaa 721 ggtgtgccgt cgtctcttct gcaggatctc tgaaaaactc ccagcttggt cgagagattg 781 ataatcatga tgcagttctg aggtttaatg gggcccctac cgacaacttc caacaggatg 841 tgggctcaaa aactaccatt cgcctaatga actctcagtt agtcaccaca gaaaagcgct 901 tcctcaagga cagtttgtac accgaaggaa tcctaattgt atgggaccca tccgtgtatc 961 atgcagatat cccaaagtgg tatcagaaac cagactacaa tttcttcgaa acctataaga 1021 gttaccgaag gctgaacccc agccagccat tttatatcct caagccccag atgccatggg 1081 aactgtggga catcattcag gaaatctctg cagatctgat tcagccaaat cccccatcct 1141 ccggcatgct gggtatcatc atcatgatga cgctgtgtga ccaggtagat atttacgagt 1201 tcctcccatc caagcgcaag acggacgtgt gctattatca ccaaaagttc tttgacagcg 1261 cttgcacgat gggtgcctac gacccgctcc tcttcgagaa gaatatggtg aagcatctca 1321 atgagggaac agatgaagac atttatttgt ttgggaaagc caccctttct ggcttccgga 1381 acattcgttg ttgagtacct agccaggcac ccttatcctt ctccatacgt cattttatgg 1441 ctactctcct ggttaccgct gcttgaagga gtgtttttat tcaacaggcc cagcctgctt 1501 cctgcgctct agggaatttt gttggcaaga gttctggggc ctccagcctg cctccctggg 1561 gccaccgagg atgggagtcc agattcttgc cacactcatt cctcctagac agcgtcctct 1621 cctccttctg catgggtagg gaaag
FIGURA 3. Secuencia completa de nucléotidos de la α2,6 sialiltransferasa (tomada de Weinstein y cols., 1987).

12
La síntesis y amplificación del ADNc-ST6N se realizó mediante las técnicas de retrotranscripción seguida de la reacción en cadena de la polimerasa (RT-PCR) empleando el estuche comercial de amplificación Super Script II (GIBCO BRL).
Los oligos que se usados para la síntesis de la sonda de ST3N fueron los previamente reportados por
Damian-Matsumura y cols., 1999: 5’-TCA AGT GGC TGA AGT ACA TC- 3’
(oligonucleótido superior o sentido) posiciones 704 al 723 5’-GTT GTG TGT CCA GGA CTC TT- 3’ (oligonucleótido inferior o antisentido) posiciones 1054 al 1035
La síntesis y amplificación del ADNc-ST3N fue usando el mismo estuche comercial que para la síntesis de ADNc-ST6N
Finalmente, para la síntesis del ADNc de ciclofilina (Cyc), usada como sonda de normalización, se
utilizó el mismo estuche comercial que para ST6N y para ST3N. Los oligos que se usaron fueron los siguientes:
5’-CTC TTT TCG CCG CTT GC-3’ (oligonucléotido superior o sentido) posiciones 19 al 35. 5’-GCT CCA TGG CTT CCA CA-3’ (oligonucléotido superior o sentido) posiciones 472 al 453. Estos oligos se obtuvieron de la siguiente secuencia reportada por Danielson y cols., 1988:
1 ctttgcagac gccgctgtct cttttcgccg cttgctgcag acatggtcaa ccccaccgtg 61 ttcttcgaca tcacggctga tggcgagccc ttgggtcgcg tctgcttcga gctgtttgca 121 gacaaagttc caaagacagc agaaaacttt cgtgctctga gcactgggga gaaaggattt 181 ggctataagg gttcctcctt tcacagaatt attccaggat tcatgtgcca gggtggtgac 241 ttcacacgcc ataatggcac tggtggcaag tccatctacg gagagaaatt tgaggatgag 301 aacttcatcc tgaagcatac aggtcctggc atcttgtcca tggcaaatgc tggaccaaac 361 acaaatggtt cccagttttt tatctgcact gccaagactg agtggctgga tggcaagcat 421 gtggtctttg ggaaggtgaa agaaggcatg agcattgtgg aagccatgga gcgttttggg 481 tccaggaatg gcaagaccag caagaagatc accatctccg actgtggaca actctaattt 541 ctttgacttg cgggcatttt acccatcaaa ccattccttc tgtagctcag gagagcaccc 601 ccaccccatc tgctcgcaat accctgtaat ctctgctctc actgaagttc tttgggttcc 661 atattttcct cattcccctt caagtctagc aggattgcaa agttaagttt atgattatga 721 ataaaaacta aatgagaagt gtt
• RETROTRANSCRIPCIÓN (RT)
Para la síntesis del ADNc a partir de ARN, se realizó el proceso de retrotranscripción (RT), para el
cual se elaboró una mezcla de 50µl en un tubo estéril para PCR de 0.2 ml con los siguientes componentes: 1 µl ARN total de cerebro de rata hembra adulta (50ng/µl), 1µl Oligo d(T)12-18 (0.5µg/µl), 10µl H2O DEPC, esta mezcla se incubó a 70°C en un baño de aceite durante 10 minutos y 1 minuto en hielo, se centrifugó

13
brevemente a 4°C. Posteriormente a esta mezcla se le agregó 2µl de 10X PCR buffer, 2µl de 25 mM MgCl2 , 1µl de 10 mM dNTP mix y 2µl de 0.1 M DTT. Posteriormente se le agregó 1µl de Super Script II (200 unidades), se agitó y se centrifugó brevemente antes de incubarse a 42°C durante 50 minutos en un incubador metabólico tipo Duffnof (Precision). Transcurrido el tiempo, se incubó la mezcla a 70°C en un baño de aceite (Boekel Industries Inc., USA) durante 15 minutos para terminar la reacción de RT. Finalmente se agregó 1µl de Rnasa H y se incubó a 37°C durante 20 minutos. Los RTs que se elaborarón fueron almacenados a -70°C hasta su posterior utilización.
• AMPLIFICACIÓN DEL ADNc (PCR: Polimerase Chain Reation) La sonda ST6N se amplificó por la técnica de Reacción en Cade na de la Polimerasa (PCR), para lo
cual en un tubo para PCR de 0.2 ml se mezclaron los siguientes componentes: 5µl 10X PCR buffer, 3µl 25 mM MgCl2, 1µl 10 mM dNTP mix, 2µl oligo sentido para ST6N, 2µl de oligo anti-sentido para ST6N, 2µl del producto de RT y se ajustó el volumen a 49.7µl con agua bidestilada estéril. Se centrifugó brevemente antes de la adición de 0.3µl de Taq DNA polimerasa (2 a 5 unidades/ µl), se agitó y se volvió a centrifugar brevemente la mezcla. Las muestras se calentaron a 95°C por 5 minutos y la amplificación se llevó acabo de la siguiente manera: 30 ciclos de 3 temperaturas (95°C por 30 segundos, 51°C por 30 segundo y 72°C por 45 segundos), se finaliza la reacción a 72°C durante 7 minutos y a 4°C durante 5 minutos, empleandose el termociclador Perkin Elmer modelo 9600. Con el objetivo de tener resultados comparativos entre la expresión de ST3N y ST6N, se sintetizó y amplificó el ADNc de ST3N por el método de RT/PCR empleando el mismo ARN de cerebro de rata hembra, unicamente se cambió la temperatura de alineación de los oligos durante el proceso de PCR, que en este caso fue de 60°C durante 30 segundos (Damián-Matsumura y cols., 1999). Así mismo, con la finalidad de demostrar que las muestras usadas en la hibridación por Northern Blot contienían la misma cantidad de ARN o en su caso hacer el ajuste correspondiente, se sintetizó el ADNc de ciclofilina de rata por el mismo método de RT/PCR, utilizando la misma temperatura de alineación de oligos que se uso para ST6N que fue de 51°C. Los productos de RT/PCR de ST6N, ST3N y Cyc tuvieron un peso molecular de 369pb, 345pb y 453pb respectivamente. Estos productos fueron separados por electroforesis en geles de agarosa 1.7% y purificados mediante columnas de exclusión molecular y por precipitación etanolica (Glenn y Glenn, 1994).
d. Marcaje de las sondas de ADNc (ST6N, ST3N y Ciclofilina) La marcación de las sondas de ADNc de la ST6N, ST3N y de la sonda de normalización Cyc se realizó empleando el protocolo de marcaje al azar empleando el estuche comercial "RadPrime DNA Labeling System®" (Gibco BRL, MD), el cual se detalla a continuación: En un tubo cónico tipo Eppendorf de 0.5 ml se desnaturalizarón 50-100 ng del ADNc en un volumen máximo de 20 µl de TE en el baño de aceite a 95°C por 5 minutos, e inmediatamente se dejó e hielo. Mientras tanto se mezclaron los siguientes componentes del estuche : 1 µl 0.5M dATP (en 3 mM Tris-HCl, pH 7.0, 0.2 mM EDTA). 1 µl 0.5M dGTP (en 3 mM Tris-HCl, pH 7.0, 0.2 mM EDTA). 1 µl 0.5M dTTP (en 3 mM Tris-HCl, pH 7.0, 0.2 mM EDTA). 20 µl de 2.5X Random Primers Solution [125 mM Tris -acetato, pH 6.8, 2.5 mM Acetato de magnesio, 25 mM 2-mercaptoetanol, 50 µg/ml cebadores oligodesoxirribonucleótidos (octámeros al azar)]. 5 µl de [32P]-dCTP.

14
ddH2O hasta un volumen de 49 µl (incluyendo el ADNc). A esta mezcla que incluye el ADNc desnaturalizado se le agregó 1 µl de la enzima DNA polimerasa (fragmento de Klenow; 4 U/µl) y se centrifugó brevemente. La mezcla de reacción se incubó a 37°C por 1 hora y se detuvo la reacción con 5 µl de solución de Stop Buffer (1mM EDTA a pH 7.5).
Las sondas que fueron marcadas se purificaron de los nucleótidos que no se incorporaron, por medio de una cromatografía de exclusión molecular, empleando columnas comerciales Micro Bio-Spin P-30 Tris® (BioRad®).
e. Prehibridación de la membrana de Nylon Con la finalidad de bloquear homogéneamente las posibles uniones no específicas de la sonda de ADNc sobre la membrana, se empleó el protocolo descrito por el fabricante de la membrana Z-Probe® GT, donde la prehibridación se realiza en una bolsa sellada conteniendo 150 µl de la solución de prehibridación/hibridación (1 mM EDTA pH 8.0, 0.25 M NaHPO4 pH 7.2, y 7 % SDS) por cm2 de membrana, durante 3 horas a 65°C con agitación constante (140 r.p.m.).
f. Hibridación de la membrana de Nylon con la sonda marcada
Una vez marcada la sonda de ADNc, se desechó la solución de prehibridación de la bolsa que contiena a la membrana y se adicionó la solución de prehibridación/ hibridación recién preparada (150 µl de solución de hibridación/cm2 de membrana). La sonda se desnaturalizó en el baño de aceite a 95°C por 5 minutos e inmediatamente después se dejó en hielo para evitar su renaturalización. La sonda se depositó en la bolsa de hibridación, se selló e incubó a 65°C en agitación constante durante 18 horas. Posteriormente se lavó la membrana a 65°C por 30 minutos con la solución de lavado (1 mM EDTA, 40 mM NaHPO4 , pH 7.2, 5 % SDS) usando 350 µl de la solución por cm2 de membrana. Después de los lavados, se secó suavemente el exceso de solución de la membrana con papel absorbente, se colocó en una bolsa nueva y se selló, ya que no se debe permitir que se seque totalmente si se volverá a hibridar, ya que el ADNc se unirá covalentemente al ARN de la membrana. Posteriormente se puso en un cassette para autorradiografía junto con una placa autorradiográfica el tiempo necesario (5 a 96 h en función de la intensidad de la señal) a -70°C y se reveló la placa.
g. Análisis de datos
Las autoradiografías obtenidas de la cuantificación de los ARNm para ST6N, ST3N y Cyc fueron sometidas a un análisis densitométrico en un sistema de análisis de imágenes “Eagle Eye II” (Stratagene, La Jolla, CA); los resultados se expresan en pixeles contenidos en el área analizada y se ajustó la intensidad del fondo a un mismo valor arbitrario. La densidad óptica relativa representa la relación entre el valor obtenido para cada muestra hibridada con ST6N y ST3N, con respecto al valor de la misma muestra normalizada con Cyc.
Para el análisis estadístico se utilizó el análisis de varianza de una vía y la prueba de “t” de Student no pareada para determinar diferencias entre grupos.

15
D. RADIOINMUNOESNSAYOS (RIA’s)
Los RIA’s se realizaron con los estuches proporcionados por el Programa Nacional de la Pituitaria de los Institutos Nacionales de Salud, EAU (National Pituitary Program, NIADDK, Bethesda MD), según lo descrito (Ulloa-Aguirre y cols, 1988). Para la FSH se empleó el extracto purificado de FSH de rata (rFSH-I-7), el cual fue radiomarcado con I125-Na por el método de la Cloramina T; se utilizó el rFSH-RP-2 para construir las curvas estándar, expresándose los resultados en términos de ng/ml de FSH-RP-2; y el anti suero policlonal anti-rFSH (conejo vs. rata; NIDDK-A-RAT FSH S-11) fue utilizado en la dilución de trabajo 1:30,000. Para evitar variaciones interensayo, se analizaron todas las muestras de cada cromatoenfoque en un mismo ensayo.
Para LH se trabajó de la misma manera, se radiomarcó la preparación LH-I-6 para ser usado como ligando marcado, la preparación de referencia empleada fue rLH-RP2 para la elaboración de la curva estándar y el antisuero policlonal anti-rLH (conejo vs. rata; NIADDK-A-RAT-LHS-8) a la dilución final de trabajo de 1:40,000. La determinación hormonal de FSH y LH de rata se realizó en alícuotas de 100ml de cada una de las fracciones obtenidas en el cromatoenfoque por duplicado, o en volúmenes menores (diluciones) si las concentraciones de las hormonas eran altas.
El RIA de 17β-estradiol fue realizado utilizando el estuche preparado en el Laboratorio de Hormonas Esteroides, Departamento de Biología de la Reproducción del Instituto Nacional de Ciencias Medicas y Nutrición Salvador Zubirán, México.

16
VI. RESULTADOS Y DISCUSIÓN
A. SÍNTESIS DEL ADNc-ST6N
La sonda de ADNc para determinar la expresión del ARNm-ST6N fue sintetizada mediante las
técnicas de retrotranscripción y de reacción en cadena de la polimerasa (RT/PCR) empleando los oligonucleótidos reportados en la metodología, así como los ADNc de ST3N y Cyc. Estos productos fueron separados por electroforesis en geles de agarosa al 1.7% (fig.4) para demostrar que el peso molecular está de acuerdo a lo reportado, que es para el caso de ST6N de aproximadamente 369 pb (carril 2, fig.4), para ST3N de aproximadamente 345 pb (carril 3, fig.4) y para Cyc 453 pb (carril 4, fig.4). La fotografía del gel muestra los estándares de peso molecular de 100 pb (100 bp DNA ladder) señalando la banda de 600 pb (carril 1).
Una vez que se separaron los productos de RT/PCR en el gel de agarosa al 1.7%, se cortaron y se purificaron por columnas de exclusión molecular y por precipitación etanólica, el producto de RT/PCR se volvió a separar por electroforesis en un gel de agarosa 1.7% para determinar su pureza y cuantificar su concentración por comparación con el estándar de peso molecular, para poder ser utilizado en la técnica de hibridación Northern blot.
Figura. 4 Separación electroforética del ADNc-ST6N, ADNc-ST3N, ADNc-CYC. Fotografía de un gel de agarosa al 1.7% teñido con bromuro de etidio. El carril 1 corresponde al marcador de peso molecular de 100 pares de bases (3 µl), el carril 2 al ADNc -ST6N (5 µl), el carril 3 al ADNc-ST3N (5 µl) y el carril 4 al ADNc-CYC (5 µl).
17
B. EXPRESIÓN DEL ARNm-ST6N
Las autorradiografías obtenidas de todos los experimentos fueron analizadas densitométicamaete en un analizador de imágenes Eagle EyeR y los resultados se expresaron en forma del cociente del área de la densidad óptica relativa calculada para la expresión del ARNm-ST6N entre el área calculada para la expresión de ARNm-ST3N y el área calculada para la sonda de normalización de la ciclofilina, ARNm-Cyc, cuya expresión es constitutiva. En todas las figuras se muestra una autorradiografía representativa del análisis del ARNm hibridado con la sonda ST6N y ST3N.
La talla molecular del ARNm-ST6N es de aproximadamente 4.7 Kb (kilobases), mientras que el peso molecular de ARNm-ST3N es de alrededor de 2.5 Kb y el peso molecular del ARNm-Cyc es de 1.1 Kb.
EXPERIMENTO 1. Animales intactos.
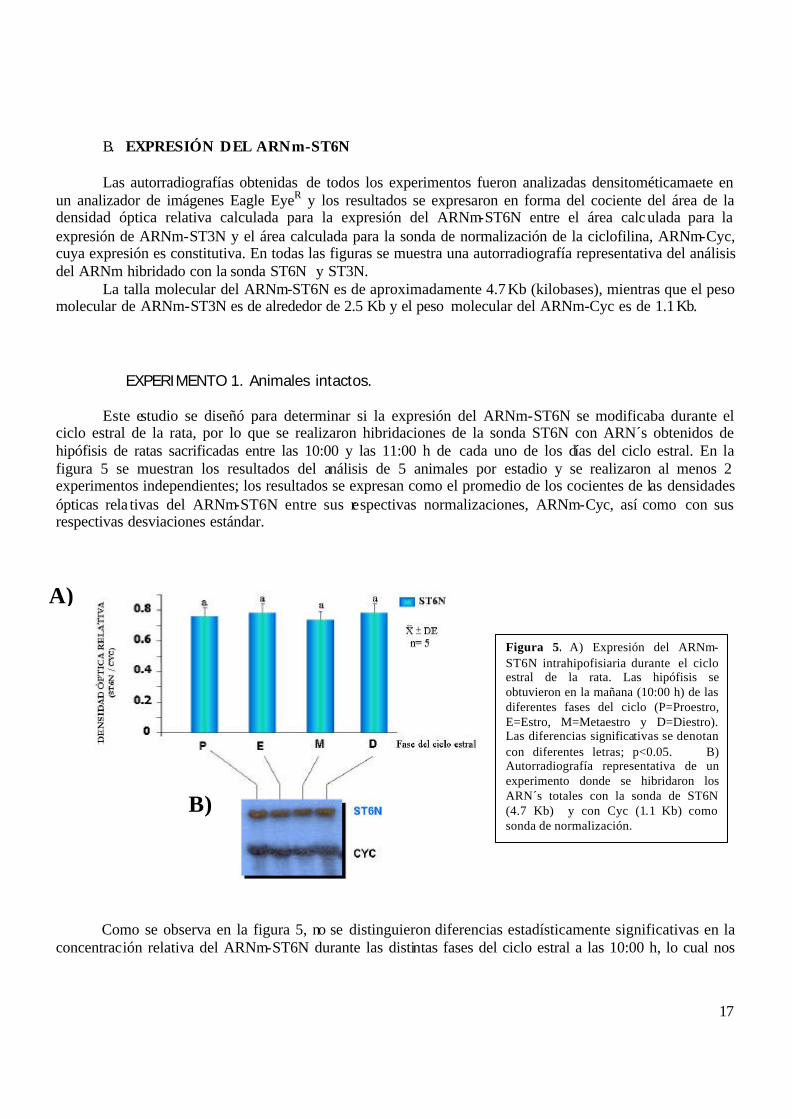
Este estudio se diseñó para determinar si la expresión del ARNm-ST6N se modificaba durante el ciclo estral de la rata, por lo que se realizaron hibridaciones de la sonda ST6N con ARN´s obtenidos de hipófisis de ratas sacrificadas entre las 10:00 y las 11:00 h de cada uno de los días del ciclo estral. En la figura 5 se muestran los resultados del análisis de 5 animales por estadio y se realizaron al menos 2 experimentos independientes; los resultados se expresan como el promedio de los cocientes de las densidades ópticas rela tivas del ARNm-ST6N entre sus respectivas normalizaciones, ARNm-Cyc, así como con sus respectivas desviaciones estándar.
Como se observa en la figura 5, no se distinguieron diferencias estadísticamente significativas en la concentración relativa del ARNm-ST6N durante las distintas fases del ciclo estral a las 10:00 h, lo cual nos
Figura 5. A) Expresión del ARNm-ST6N intrahipofisiaria durante el ciclo estral de la rata. Las hipófisis se obtuvieron en la mañana (10:00 h) de las diferentes fases del ciclo (P=Proestro, E=Estro, M=Metaestro y D=Diestro). Las diferencias significativas se denotan con diferentes letras; p<0.05. B) Autorradiografía representativa de un experimento donde se hibridaron los ARN´s totales con la sonda de ST6N (4.7 Kb) y con Cyc (1.1 Kb) como sonda de normalización.
A)
B)
18
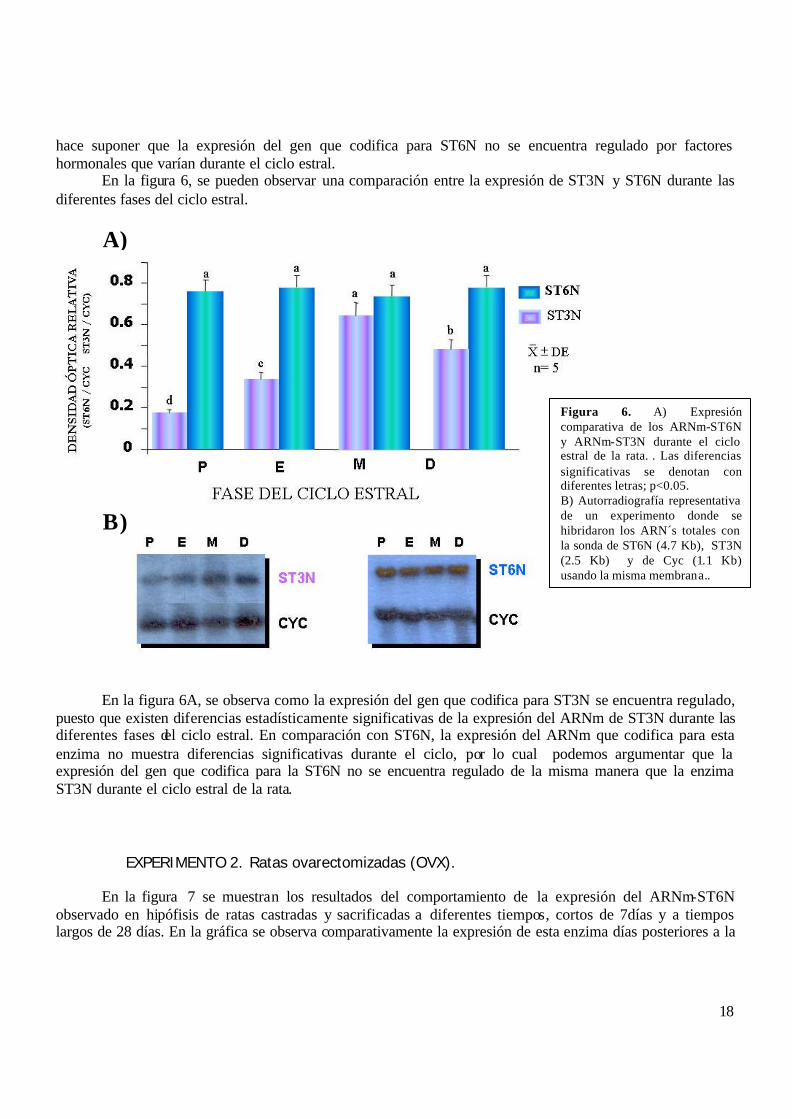
hace suponer que la expresión del gen que codifica para ST6N no se encuentra regulado por factores hormonales que varían durante el ciclo estral. En la figura 6, se pueden observar una comparación entre la expresión de ST3N y ST6N durante las diferentes fases del ciclo estral.
En la figura 6A, se observa como la expresión del gen que codifica para ST3N se encuentra regulado,
puesto que existen diferencias estadísticamente significativas de la expresión del ARNm de ST3N durante las diferentes fases del ciclo estral. En comparación con ST6N, la expresión del ARNm que codifica para esta enzima no muestra diferencias significativas durante el ciclo, por lo cual podemos argumentar que la expresión del gen que codifica para la ST6N no se encuentra regulado de la misma manera que la enzima ST3N durante el ciclo estral de la rata.
EXPERIMENTO 2. Ratas ovarectomizadas (OVX). En la figura 7 se muestran los resultados del comportamiento de la expresión del ARNm-ST6N observado en hipófisis de ratas castradas y sacrificadas a diferentes tiempos, cortos de 7días y a tiempos largos de 28 días. En la gráfica se observa comparativamente la expresión de esta enzima días posteriores a la
Figura 6. A) Expresión comparativa de los ARNm-ST6N y ARNm-ST3N durante el ciclo estral de la rata. . Las diferencias significativas se denotan con diferentes letras; p<0.05. B) Autorradiografía representativa de un experimento donde se hibridaron los ARN´s totales con la sonda de ST6N (4.7 Kb), ST3N (2.5 Kb) y de Cyc (1.1 Kb) usando la misma membrana..
A)
B)
19
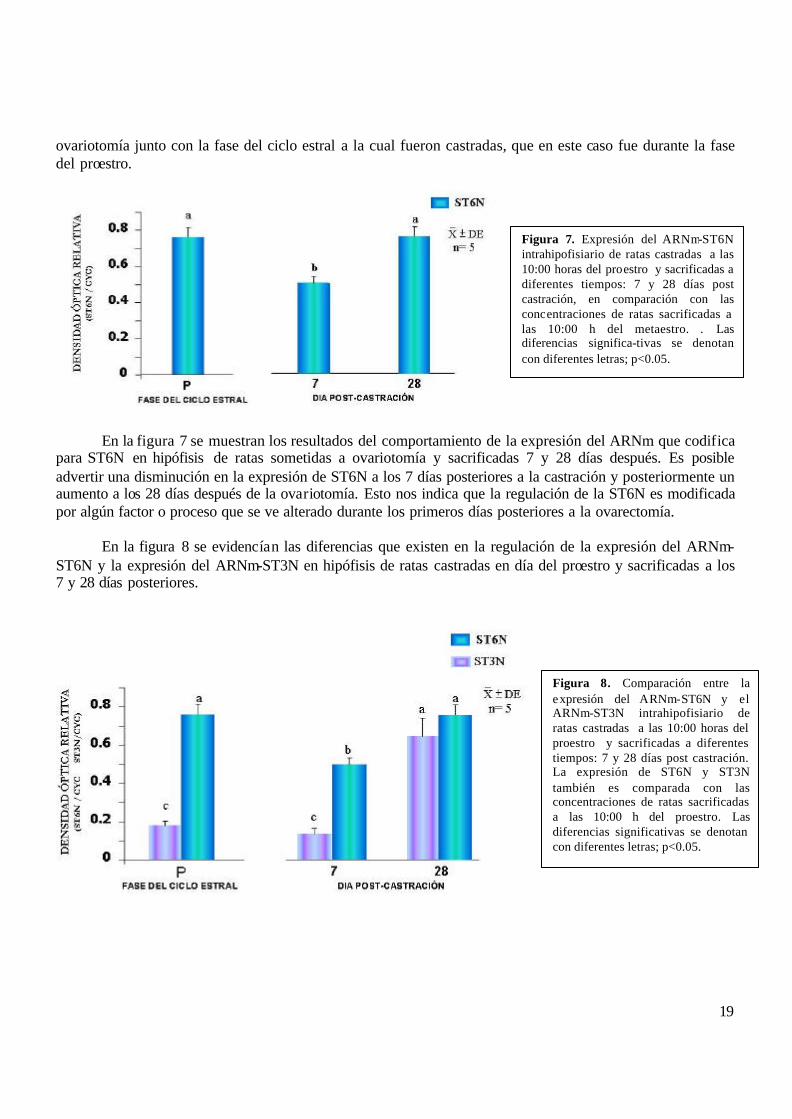
ovariotomía junto con la fase del ciclo estral a la cual fueron castradas, que en este caso fue durante la fase del proestro.
En la figura 7 se muestran los resultados del comportamiento de la expresión del ARNm que codifica para ST6N en hipófisis de ratas sometidas a ovariotomía y sacrificadas 7 y 28 días después. Es posible advertir una disminución en la expresión de ST6N a los 7 días posteriores a la castración y posteriormente un aumento a los 28 días después de la ovariotomía. Esto nos indica que la regulación de la ST6N es modificada por algún factor o proceso que se ve alterado durante los primeros días posteriores a la ovarectomía. En la figura 8 se evidencían las diferencias que existen en la regulación de la expresión del ARNm-ST6N y la expresión del ARNm-ST3N en hipófisis de ratas castradas en día del proestro y sacrificadas a los 7 y 28 días posteriores.
Figura 7. Expresión del ARNm-ST6N intrahipofisiario de ratas castradas a las 10:00 horas del proestro y sacrificadas a diferentes tiempos: 7 y 28 días post castración, en comparación con las concentraciones de ratas sacrificadas a las 10:00 h del metaestro. . Las diferencias significa-tivas se denotan con diferentes letras; p<0.05.
Figura 8. Comparación entre la expresión del ARNm-ST6N y el ARNm-ST3N intrahipofisiario de ratas castradas a las 10:00 horas del proestro y sacrificadas a diferentes tiempos: 7 y 28 días post castración. La expresión de ST6N y ST3N también es comparada con las concentraciones de ratas sacrificadas a las 10:00 h del proestro. Las diferencias significativas se denotan con diferentes letras; p<0.05.
20
Con la finalidad de hacer una comparación directa, se incluyeron en el mismo experimento ARN de hipófis is de ratas sacrificadas en la fase del ciclo en la cual se ha reportado la mayor expresión de la enzima, metaestro a las 10:00 horas (Damian-Matsumura y cols., 1999). En la figura 8 observa que a los 7 días post-castración, que la expresión del ARNm-ST3N se mantiene en la misma concentración que la encontrada en las hipófisis de ratas intactas sacrificadas en la mañana del proestro, puesto que a pesar de la remoción de los ovarios aun se detectan concentraciones de estradiol en la circulación (Damian-Matsumura y cols. , 1999). En este mismo momento en que se observa un decremento de aproximadamente del 30% con respecto a las concentraciones de la expresión del ARNm-ST6N. A los 28 días post-castración como se observa en la figura 8, se incrementa la concentración de la expresión tanto ST3N como ST6N hasta llegar a concentraciones similares a las reportadas para la fase del metaestro de la rata.
Estos resultados en donde observamos un decremento en la expresión del ARNm-ST6N a los 7 días post-castración, nos permiten sugerir que el gen que codifica para ST6N se encuentra regulado posiblemente por algún factor hormonal.
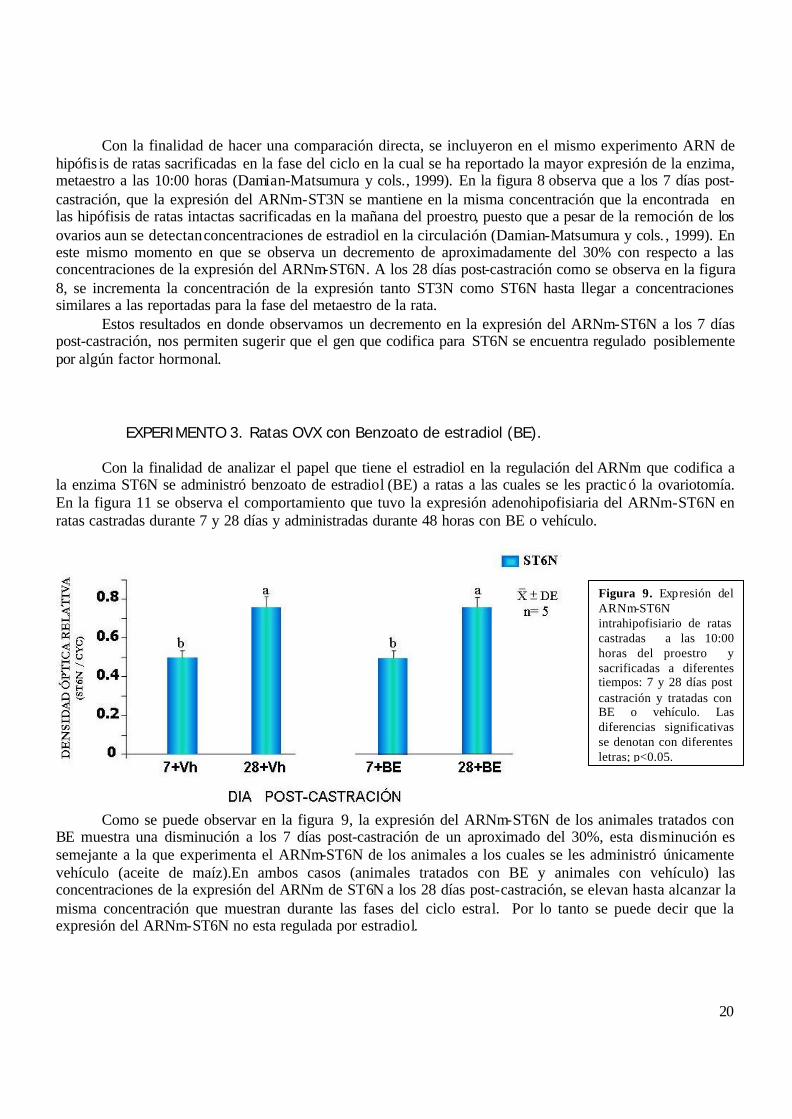
EXPERIMENTO 3. Ratas OVX con Benzoato de estradiol (BE). Con la finalidad de analizar el papel que tiene el estradiol en la regulación del ARNm que codifica a la enzima ST6N se administró benzoato de estradiol (BE) a ratas a las cuales se les practicó la ovariotomía. En la figura 11 se observa el comportamiento que tuvo la expresión adenohipofisiaria del ARNm-ST6N en ratas castradas durante 7 y 28 días y administradas durante 48 horas con BE o vehículo.
Como se puede observar en la figura 9, la expresión del ARNm-ST6N de los animales tratados con BE muestra una disminución a los 7 días post-castración de un aproximado del 30%, esta disminución es semejante a la que experimenta el ARNm-ST6N de los animales a los cuales se les administró únicamente vehículo (aceite de maíz).En ambos casos (animales tratados con BE y animales con vehículo) las concentraciones de la expresión del ARNm de ST6N a los 28 días post-castración, se elevan hasta alcanzar la misma concentración que muestran durante las fases del ciclo estral. Por lo tanto se puede decir que la expresión del ARNm-ST6N no esta regulada por estradiol.
Figura 9. Expresión del ARNm-ST6N intrahipofisiario de ratas castradas a las 10:00 horas del proestro y sacrificadas a diferentes tiempos: 7 y 28 días post castración y tratadas con BE o vehículo. Las diferencias significativas se denotan con diferentes letras; p<0.05.
21
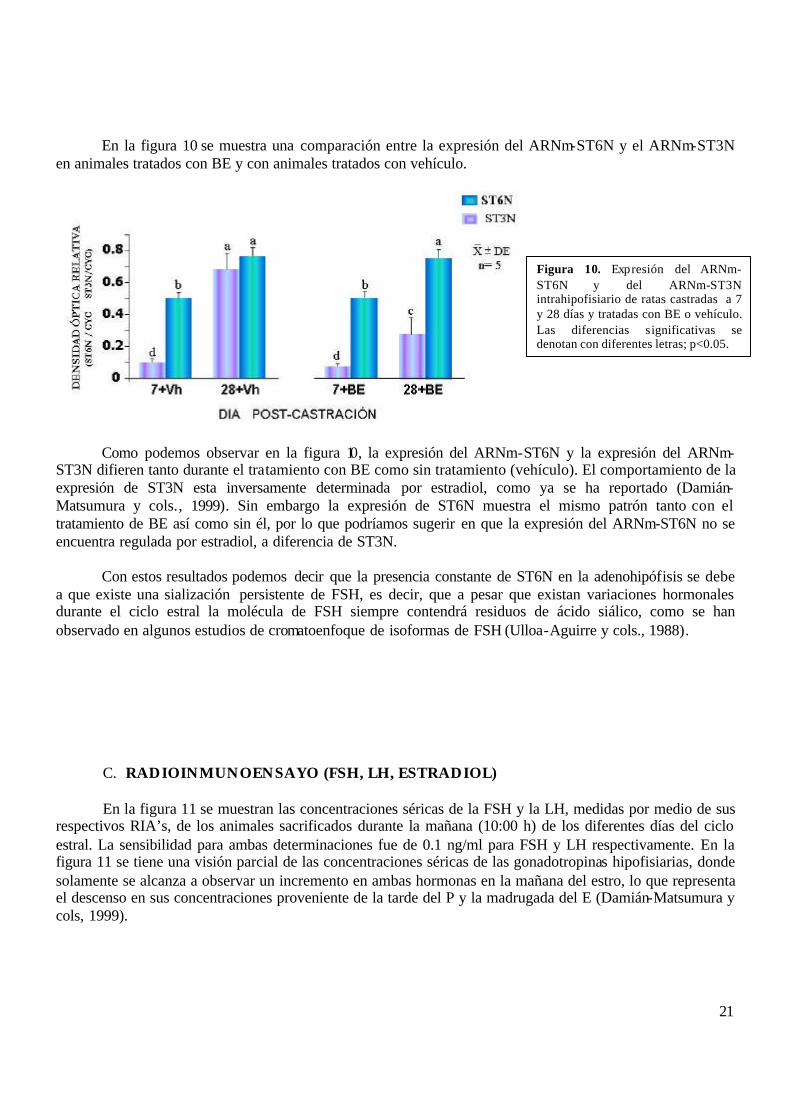
En la figura 10 se muestra una comparación entre la expresión del ARNm-ST6N y el ARNm-ST3N en animales tratados con BE y con animales tratados con vehículo.
Como podemos observar en la figura 10, la expresión del ARNm-ST6N y la expresión del ARNm-ST3N difieren tanto durante el tratamiento con BE como sin tratamiento (vehículo). El comportamiento de la expresión de ST3N esta inversamente determinada por estradiol, como ya se ha reportado (Damián-Matsumura y cols., 1999). Sin embargo la expresión de ST6N muestra el mismo patrón tanto con el tratamiento de BE así como sin él, por lo que podríamos sugerir en que la expresión del ARNm-ST6N no se encuentra regulada por estradiol, a diferencia de ST3N. Con estos resultados podemos decir que la presencia constante de ST6N en la adenohipófisis se debe a que existe una sialización persistente de FSH, es decir, que a pesar que existan variaciones hormonales durante el ciclo estral la molécula de FSH siempre contendrá residuos de ácido siálico, como se han observado en algunos estudios de cromatoenfoque de isoformas de FSH (Ulloa-Aguirre y cols., 1988).
C. RADIOINMUNOENSAYO (FSH, LH, ESTRADIOL)
En la figura 11 se muestran las concentraciones séricas de la FSH y la LH, medidas por medio de sus respectivos RIA’s, de los animales sacrificados durante la mañana (10:00 h) de los diferentes días del ciclo estral. La sensibilidad para ambas determinaciones fue de 0.1 ng/ml para FSH y LH respectivamente. En la figura 11 se tiene una visión parcial de las concentraciones séricas de las gonadotropinas hipofisiarias, donde solamente se alcanza a observar un incremento en ambas hormonas en la mañana del estro, lo que representa el descenso en sus concentraciones proveniente de la tarde del P y la madrugada del E (Damián-Matsumura y cols, 1999).
Figura 10. Expresión del ARNm-ST6N y del ARNm-ST3N intrahipofisiario de ratas castradas a 7 y 28 días y tratadas con BE o vehículo. Las diferencias significativas se denotan con diferentes letras; p<0.05.

22
Las concentraciones séricas de las gonadotropinas hipofisiarias de ratas sacrificadas a diferentes intervalos de tiempo después de ser castradas, en la mañana del día del P, se muestran en la figura 12. Para hacer el análisis estadístico, los resultados se comparan con respecto a las concentraciones encontradas en los animales sacrificados después de 7 días de castración.
En el caso de las concentraciones séricas de FSH en animales castrados, estas se incrementan rápidamente a los 7 días de castración y a los 28 días hay un aumento del 35% aproximadamente. Los
Figura 11. Concentraciones séricas de las hormonas FSH y LH en los cuatro días del ciclo estral de ratas sacrificadas a las 10:00 h. Las diferencias significativas entre los grupos se denotan con distintas letras: p<0.05.
Figura 12. Concentraciones séricas de las hormonas de rata FSH y LH de animales castrados. La diferencias significativas con respecto a los animales sacrificados después de 7 días de castración se denotan con *p<0.05.
23
incrementos de la LH sérica se comportan de una manera similar a la FSH, solo que a los 28 días post-castración el incremento con respecto a los 7 días postcastración es del 60% aproximadamente.
Congruente con lo informado (Ward y cols, 1991), el incremento de moderado a alto en las
concentraciones séricas de LH y FSH es debido a la pérdida de la retroalimentación negativa de los esteroides gonadales estradiol y progesterona (Freeman, 1994).
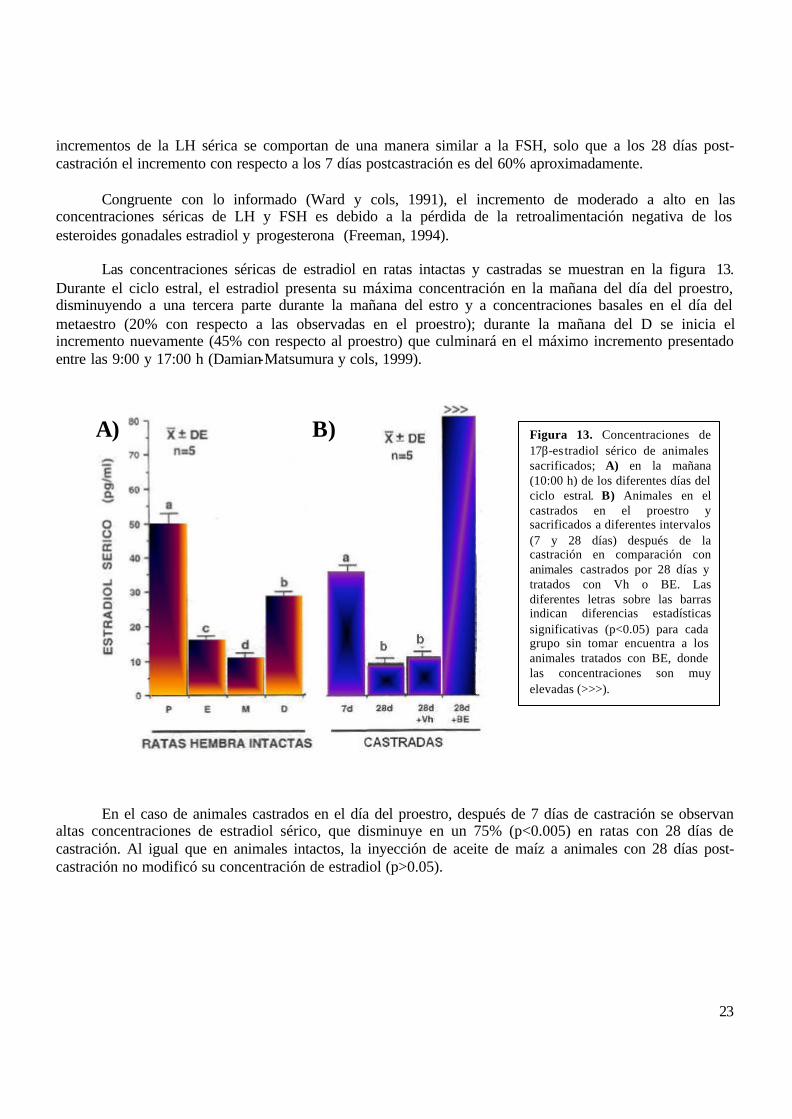
Las concentraciones séricas de estradiol en ratas intactas y castradas se muestran en la figura 13. Durante el ciclo estral, el estradiol presenta su máxima concentración en la mañana del día del proestro, disminuyendo a una tercera parte durante la mañana del estro y a concentraciones basales en el día del metaestro (20% con respecto a las observadas en el proestro); durante la mañana del D se inicia el incremento nuevamente (45% con respecto al proestro) que culminará en el máximo incremento presentado entre las 9:00 y 17:00 h (Damian-Matsumura y cols, 1999).
En el caso de animales castrados en el día del proestro, después de 7 días de castración se observan altas concentraciones de estradiol sérico, que disminuye en un 75% (p<0.005) en ratas con 28 días de castración. Al igual que en animales intactos, la inyección de aceite de maíz a animales con 28 días post-castración no modificó su concentración de estradiol (p>0.05).
Figura 13. Concentraciones de 17β-estradiol sérico de animales sacrificados; A) en la mañana (10:00 h) de los diferentes días del ciclo estral. B) Animales en el castrados en el proestro y sacrificados a diferentes intervalos (7 y 28 días) después de la castración en comparación con animales castrados por 28 días y tratados con Vh o BE. Las diferentes letras sobre las barras indican diferencias estadísticas significativas (p<0.05) para cada grupo sin tomar encuentra a los animales tratados con BE, donde las concentraciones son muy elevadas (>>>).
A) B)

24
VII. CONCLUSIONES
Se ha demostrado previamente que los cambios en la cantidad de residuos de ácido siálico pueden
tener efectos biológicos tan variados como la modificación en la distribución por carga eléctrica de la FSH, generando diferentes isohormonas modulando las propiedades biológicas de la FSH circulante, por lo que es indudable que éstos juegan un papel fisiológico importante sobre el ciclo reproductivo, ya que contribuyen a ajustar la actividad estimulante de los folículos a necesidades temporales (Ulloa-Aguirre y cols., 1995; Padmanhabhan y cols., 1999)
Experimentos previos han demostrado que la expresión diferencial del ARNm de la enzima ST3N se encuentra regulada por estrógenos (Damián-Matsumura y cols., 1999), sin embargo ST3N no es la única enzima que sializa a FSH, ST6N también es una enzima encargada de esta tarea. No obstante, la regulación de estas enzimas es diferente. Con este trabajo se demostró que la expresión del ARNm de la enzima ST6N no presenta cambios significativos durante el ciclo estral a las 10:00, lo que sugiere que ST6N no se encuentra regulada por factores hormonales que oscilan durante el ciclo estral. Así mismo, se manifestó el comportamiento que tuvo ST6N a los 7 días post-castración donde la expresión del ARNm de ST6N disminuyó con respecto a la observada durante el ciclo estral, sin embargo a los 28 días post-castración la expresión del ARNm de ST6N se incrementa hasta los niveles presentes durante el ciclo estral, lo que sugiere que algún factor que removido durante el proceso de castración interviene en la regulación directa o indirecta de la expresión del ARNm que codifica para la enzima ST6N. De la misma manera, el comportamiento de la expr esión de ARNm de ST6N a 7 y 28 días post-castración no se ve modificado por la administración de Benzoato de Estradiol (BE), por lo cual podemos decir que la enzima ST6N no se encuentra regulada estrogenicamente. Como siguiente meta se analizará el papel que desempeña GnRH como posible factor que interviene en la expresión del ARNm-ST6N en hipófisis de ratas sometidas a diversas condiciones fisiológicas y experimentales.

25
VIII. BIBLIOGRAFÍA 1. Baenziger JU. (1994). Protein-specific glycosyltransferases: how and why they do it! FASEB J. 8 (13):
1019-1025. 2. Baenziger JU, Green ED. (1988). Pituitary glycoprotein hormone oligosaccharides: structure, synthesis
and function of the asparagine-linked oligosaccharides on lutropin, follitropin and thyrotropin. Biochim Biophys Acta. 947 (2): 287-306.
3. Becker WM, Reece JB, Poenie MF. (2000). The World of the Cell. Fourth edition. The Benjamin/Cummings Publishing Company, CA Ed. pp 583 594.
4. Bishop LA, Robertson DM, Cahir N y Schofield PR. (1994). Specific roles for the asparagine-linked carbohydrate residues of recombinant human follicle stimulating hormone in receptor binding and signal transduction. Mol Endocrinol. 8 (6): 722-731.
5. Blumenfeld Z, Ritter M. (2001). Inhibin, activin, and follistatin in human fetal pituitary and gonadal physiology. Ann N Y Acad Sci. 943:34-48.
6. Bousfield GR, Perry WM, Ward DN. (1994). Gonadotropins. Chemistry and Biosynthesis. En: The Physiology of Reproduction. (Knobil E & Neil J D, Eds.). Raven Press, Ltd. New York, USA. 2nd Ed. Vol. 1. pp. 1749-1792.
7. Bousfield GR, Baker VL, Gotschall RR, Butnev VY. (2000). Carbohydrate analysis of glycoprotein hormones. Methods. 21(1):15-39.
8. Chappel SC. (1995). Heterogeneity of follicle stimulating hormone: control and physiological function. Hum Reprod Update. 1 (5): 479-487.
9. Chomczynski P y Sacchi N. (1987). Single-step method of RNA isolation by acid guanidinium thiocyanate- phenol-chloroform extraction. Anal Biochem. 162 (1): 156-159.
10. Colley KJ, Lee EU y Paulson JC. (1992). The signal anchor and stem regions of the ß-galactoside α2,6- sialyltransferase may each act to localize the enzyme to the Golgi apparatus. J Biol Chem. 267 (11): 7784-7793.
11. Colley KJ, Lee EU, Adler B, Browne JK y Paulson JC. (1989). Conversion of a Golgi apparatus sialyltransferase to a secretory protein by replacement of the NH2-terminal signal anchor with a signal peptide. J Biol Chem. 264 (30): 17619-17622.
12. Crocker PR, Varki A. (2001a). Siglecs in the immune system. Immunology. 103(2):137-45 13. Crocker PR, Varki A. (2001b). Siglecs, sialic acids and innate immunity. Trends Immunol. 22(6):337-
42 14. Dahdal RY y Colley KJ. (1993). Specific sequences in the signal anchor of the ß-galactoside α2,6-
sialyltransferase are not essential for Golgi localization. Membrane flanking sequences may specify Golgi retention. J Biol Chem. 268 (35): 26310-26319.
15. Damián-Matsumura P, Zaga V, Maldonado A, Sánchez-Hernández C, Timossi C, Ulloa-Aguirre A. (1999). Oestrogens regulate pituitary α2,3-sialyltransferase messenger ribonucleic acid levels in the female rat. Journal of Molecular Endocrinology. 23, 153-165.
16. Danielson PE, Forss-Petter S, Brow MA, Calavetta L, Douglass J, Milner RJ, Sutcliffe JG. (1988). p1B15: a cDNA clone of the ral mRNA encoding cyclophilin. DNA. 7 (4): 261-267.
17. Datta AK, Paulson JC. (1995). The sialyltransferase "sialylmotif" participates in binding the donor substrate CMP-NeuAc. J Biol Chem. 270 (4): 1497-1500.
18. Datta AK, Paulson JC. (1997). Sialylmotifs of sialyltransferases. Indian J Biochem Biophys. 34(1-2):157-65
19. Datta AK, Sinha A y Paulson JC. (1998). Mutation of the sialyltransferase S-sialylmotif alters the kinetics of the donor and acceptor substrates. J Biol Chem. 273 (16): 9608-9614.

26
20. Dharmesh SM y Baenziger JU. (1993). Estrogen modulates expression of the glycosyltransferases that synthesize sulfated oligosaccharides on lutropin. Proc Natl Acad Sci U S A. 90 (23): 11127-11131.
21. Freeman ME. (1994). The neuroendocrine control of the ovarian cycle of the rat. En: The Physiology of Reproduction. (Knobil E & Neil JD, Eds). Raven Press, Ltd. New York, USA. 2nd Ed. Vol.2 pp. 613-658.
22. Galway AB, Hsueh AJ, Keene JL, Yamoto M, Fauser BC y Boime I. (1990a). In vitro and in vivo bioactivity of recombinant human follicle- stimulating hormone and partially deglycosylated variants secreted by transfected eukaryotic cell lines. Endocrinology. 127 (1): 93-100.
23. Galway AB, Lapolt PS, Tsafriri A, Dargan CM, Boime I y Hsueh AJ. (1990b). Recombinant follicle-stimulating hormone induces ovulation and tissue plasminogen activator expression in hypophysectomized rats. Endocrinology. 127 (6): 3023-3028.
24. Gharib SD, Wierman ME, Shupnik MA y Chin WW. (1990). Molecular biology of the pituitary gonadotropins. Endocr Rev. 11 (1): 177-199.
25. Gillespie W, Kelm S, Paulson JC. (1992). Cloning and expression of the Gal ß-1,3 GalNAc α2,3- sialyltransferase. J Biol Chem. 267 (29): 21004-21010.
26. Glenn TC y Glenn SJ. (1994). Rapid elution of DNA from agarose gels using polyester plug spin inserts (PEPSIs). Trends Genet. 10 (10): 344.
27. Green ED y Baenziger JU. (1988a). Asparagine-linked oligosaccharides on lutropin, follitropin, and thyrotropin. I. Structural elucidation of the sulfated and sialylated oligosaccharides on bovine, ovine, and human pituitary glycoprotein hormones. J Biol Chem. 263 (1): 25-35.
28. Green ED y Baenziger JU. (1988b). Asparagine-linked oligosaccharides on lutropin, follitropin, and thyrotropin. II. Distributions of sulfated and sialylated oligosaccharides on bovine, ovine, and human pituitary glycoprotein hormones. J Biol Chem. 263 (1): 36-44.
29. Haisenleder DJ, Dalkin AC, Marshall JC. (1994). Regulation of gonadotropin gene expression. En: The Physiology of Reproduction. (Knobil E & Neil J D, Eds.). Raven Press, Ltd. New York, USA. 2nd Ed. Capítulo 31, Vol. 2. pp. 1793-1813.
30. Helton TE y Magner JA. (1994a). Sialyltransferase messenger ribonucleic acid increases in thyrotrophs of hypothyroid mice: an In situ hybridization study. Endocrinology. 134 (6): 2347-2353.
31. Helton TE y Magner JA. (1994b). ß-1,4-galactosyltransferase and α-mannosidase-II messenger ribonucleic acid levels increase with different kinetics in thyrotrophs of hypothyroid mice. Endocrinology. 135 (5): 1980-1985.
32. Hirschberg CB y Snider MD. (1987). Topography of glycosylation in the rough endoplasmic reticulum and Golgi apparatus. Annu Rev Biochem. 56 63-87.
33. Kaiser UB. (1998). Molecular mechanisms of the regulation of gonadotropin gene expression by gonadotropin-releasing hormone. Mol Cells. 31;8(6):647-56.
34. Kelm S, Schauer R. (1997). Sialic acids in molecular and cellular interactions. Int Rev Cytol. 175:137-240
35. Keppler OT, Horstkorte R, Pawlita M, Schmidt C, Reutter W. (2001). Biochemical engineering of the N-acyl side chain of sialic acid: biological implications. Glycobiology. 11(2):11R-18R.
36. Kitagawa H y Paulson JC. (1993). Cloning and expression of human Gal ß1,3 (4) GlcNAc α2,3- sia lyltransferase. Biochem Biophys Res Commun. 194 (1): 375-382.
37. Kitagawa H y Paulson JC. (1994a). Cloning of a novel α2,3-sialyltransferase that sialylates glycoprotein and glycolipid carbohydrate groups. J Biol Chem. 269 (2): 1394-1401.
38. Kitagawa H y Paulson JC. (1994b). Differential expression of five sialyltransferase genes in human tissues. J Biol Chem. 269 (27): 17872-17878.
39. Kitagawa H, Mattei MG, Paulson JC. (1996). Genomic organization and chromosomal mapping of the Gal ß1,3 Gal NAc/Gal ß1,4 Glc NAc α2,3-sialyltransferase. J Biol Chem. 271 (2): 931-938.

27
40. Kleene R y Berger EG. (1993). The molecular and cell biology of glycosyltransferases. Biochim Biophys Acta. 1154 (3-4): 283-325.
41. Lambalk CB, Delemarre -van HA, Findlay JK, Kenemans P. (2001). Basic concept s of the female reproductive system. En: Endocrinology and Metabolism (Pinchera A. Ed.) McGraw-Hill International (UK) Ltd. Londres. Cap.43; pp. 435-464.
42. Lambert A, Talbot JA, Anobile CJ, Robertson WR. (1998). Gonadotrophin heterogeneity and biopotency: implications for assisted reproduction. Mol Hum Reprod. 4(7):619-29
43. Livingston BD y Paulson JC. (1993). Polymerase chain reaction cloning of a developmentally regulated member of the sialyltransferase gene family. J Biol Chem. 268 (16): 11504-11507.
44. Manzella SM, Dharmesh SM, Cohick CB, Soares MJ, Baenziger JU. (1997). Developmental regulation of a pregnancy-specific oligosaccharide structure, NeuAc α-2,6 Gal NAc α-1,4 Glc NAc, on select members of the rat placental prolactin family. J Biol Chem. 272 (8): 4775-4782.
45. Martin A, Rambal C, Berger V, Perier S, Louisot P. (1998). Availability of specific sugars for glycoconjugate biosynthesis: a need for further investigations in man. Biochimie 80(1):75-86.
46. McVeagh P, Miller JB. (1997). Human milk oligosaccharides: only the breast. J Paediatr Child Health. 33(4):281-6
47. Nakano T, Sugawara M, Kawakami H. (2001). Sialic acid in human milk: composition and functions. Acta Paediatr Taiwan. 42(1):11-7.
48. Padmanabhan V, Sairam MR, Hassing JM, Brown MB, Ridings JW, Beitins IZ. (1991). Follicle-stimulating hormone signal transduction: role of carbohydrate in aromatase induction in immature rat Sertoli cells. Mol Cell Endocrinol. 79 (1-3): 119-128.
49. Padmanabhan V, McFadden K, Mauger DT, Karsch FJ, Midgley AR, Jr. (1997). Neuroendocrine control of follicle-stimulating hormone (FSH) secretion. I. Direct evidence for separate episodic and basal components of FSH secretion. Endocrinology. 138 (1): 424-432.
50. Padmanabhan V, Lee JS, Beitins IZ. (1999). Follicle -stimulating isohormones: regulation and biological significance. J Reprod Fertil Suppl 54:87-99
51. Schauer R. (2000). Achievements and challenges of sialic acid research. Glycoconj J. 17(7-9):485-99. 52. Sillanaukee P, Ponnio M, Jaaskelainen IP. (1999). Occurrence of sialic acids in healthy humans and
different disorders. Eur J Clin Invest. 29(5):413-25 53. Smith PL, Kaetzel D, Nilson J, Baenziger JU. (1990). The sialylated oligosaccharides of recombinant
bovine lutropin modulate hormone bioactivity. J Biol Chem. 265 (2): 874-881. 54. Stockell Hartree A y Renwick AG. (1992). Molecular structures of glycoprotein hormones and
functions of their carbohydrate components. Biochem J. 287 (3): 665-679. 55. Tsuji S, Datta AK y Paulson JC. (1996). Systematic nomenclature for sialyltransferases. Glycobiology.
6 (7): v-vii. 56. Ulloa-Aguirre A, Timossi C. (2000). Biochemical and funcional aspects of gonadotrophin-releasing
hormona and gonadotrophins. Reproductive BioMedicine. 1(2):41-55. 57. Ulloa-Aguirre A, Espinoza R, Damian-Matsumura P, Larrea F, Flores A, Morales L, Dominguez
R. (1988). Studies on the microheterogeneity of anterior pituitary follicle-stimulating hormone in the female rat. Isoelectric focusing pattern throughout the estrous cycle. Biol Reprod. 38 (1): 70-78.
58. Ulloa-Aguirre A, Schwall R, Cravioto A, , Damian-Matsumura P. (1992). Effects of gonadotrophin -releasing hormone, recombinant human activin-A and sex steroid hormones upon the follicle-stimulating isohormones secreted by rat anterior pituitary cells in culture. J Endocrinol. 134 (1): 97-106.
59. Ulloa-Aguirre A, Midgley AR, Jr., Beitins IZ, Padmanabhan V. (1995). Follicle-stimulating isohormones: characterization and physiological relevance. Endocr Rev. 16 (6): 765-787.
60. Ulloa-Aguirre A, Timossi C, Damian-Matsumura P, Dias JA. (1999). Role of glycosylation in function of follicle-stimulating hormone. Endocrine. 11(3):205-15.

28
61. Ulloa-Aguirre A, Timossi C, Mendez JP. (2001). Is there any physiological role for gonadotrophin oligosaccharide heterogeneity in humans? I. Gondatrophins are synthesized and released in multiple molecular forms. A matter of fact. Hum Reprod. 16(4):599-604
62. Varki A. (1997). Sialic acids as ligands in recognition phenomena. FASEB J. 11(4):248-55. 63. Venkatesh SG, Deshpande V. (1999). A comparative review of the structure and biosynthesis of
thyroglobulin. Comp Biochem Physiol C Pharmacol Toxicol Endocrinol. 122(1):13-20 64. Verbert A, Cacan R. (1999). Trafficking of oligomannosides released during N-glycosylation: a clearing
mechanism of the rough endoplasmic reticulum. Biochim Biophys Acta. 6;1473(1):137-46. 65. Ward DN, Bousfield GR y Moore KH. (1991). Gonadotropins. En: Reproduction in Domestic Animals.
Academic Press, Inc. London Ed. Capítulo 2. pp.25-79. 66. Weinstein J, Lee EU, McEntee K, Lai PH, Paulson JC. (1987). Primary structure of ß-galactoside
α2,6-sialyltransferase. Conversion of membrane-bound enzyme to soluble forms by cleavage of the NH2-terminal signal anchor. J Biol Chem. 262 (36): 17735-17743.
67. Wen DX, Livingston BD, Medzihrads zky KF, Kelm S, Burlingame AL, Paulson JC. (1992a). Primary structure of Gal ß1,3 (4 )Glc NAc α2,3-sialyltransferase determined by mass spectrometry sequence analysis and molecular cloning. Evidence for a protein motif in the sialyltransferase gene family. J Biol Chem. 267 (29): 21011-21019.
68. Wen DX, Svensson EC, Paulson JC. (1992b). Tissue-specific alternative splicing of the β-galactoside α2,6- sialyltransferase gene. J Biol Chem. 267 (4): 2512-2518.





























