Embed Size (px)
Citation preview

Rekonstruktion der Phylogenie der
Opisthobranchia (Mollusca, Gastropoda) mit Hilfe
molekularer Methoden
Dissertation zur Erlangung des Grades
eines Doktors der Naturwissenschaften
der Fakultät für Biologie
der Ruhr-Universität Bochum
angefertigt am
Lehrstuhl für Spezielle Zoologie
vorgelegt von
Verena Vonnemann
aus
Essen
Bochum, 2005

Inhaltsverzeichnis 1 Einleitung 1 1.1 Die Systematik 1 1.2 Die phylogenetische Systematik 3 1.3 Die molekulare Phylogenetik 3 1.4 Die Gastropoda 4 1.5 Die Opisthobranchia 6 1.6 Bisherige Erkenntnisse über die Phylogenie der Opisthobranchia 12 1.7 Molekulare Marker 15 1.8 Ziele der vorliegenden Arbeit 18 2 Material und Methoden 23 2.1 Untersuchte Taxa und Herkunft des Tiermaterials 23 2.2 Untersuchte Gene 29 2.3 Labormethoden 32 2.3.1 Fixierung des Tiermaterials 32 2.3.2 DNA-Isolierung 32 2.3.3 Die Polymerase-Kettenreaktion 33 2.3.4 Aufreinigung von PCR-Produkten 36 2.3.5 Klonierung von PCR-Fragmenten 36 2.3.6 Aufreinigung der Plasmid-DNA 37 2.3.7 DNA-Sequenzierung 38 2.4 Methoden der Auswertung 42 2.4.1 Erstellen einer Konsensussequenz 42 2.4.2 Kontrolle der erzeugten Sequenzen 42 2.4.3 Alinierung der DNA-Sequenzen 43 2.4.4 Verfahren zur Erstellung von Dendrogrammen 44 2.4.4.1 Distanzmethoden 44 2.4.4.2 Maximum Parsimonie-Verfahren 47 2.4.4.3 maximum likelihood-Verfahren 48 2.4.4.4 Bayes’sche Verfahren 49 2.5 bootstrap-Verfahren 50 2.6 Prüfung der Plausibilität von Dendrogrammen 51 2.6.1 Vergleich mit Dendrogrammen auf der Basis alternativer Daten 51 2.6.2 Vergleich mit Fossilfunden 51 2.6.3 Vergleich mit biologischen Daten 52 2.7 Statistische Tests 52 2.7.1 Analyse der Basenzusammensetzung 52 2.7.2 Berechnung der Substitutionssättigung 52 2.7.3 Der relative rate test 53 2.8 Phylogenetisches Signal 53

3 Ergebnisse 55 3.1 Alinierungsergebnisse 55 3.2 Statistische Tests 57 3.2.1 Sequenzlängen, Basenzusammensetzung und X2-Test 57 3.2.2 Berechnung der Substitutionssättigung 64 3.2.3 relative rate test 69 3.3 Phylogenetisches Signal 71 3.4 Intragenomische Variabilität 73 3.5 Phylogenie der Opisthobranchia 74 3.5.1 18S rDNA 74 3.5.1.1 Maximum Parsimonie-Analyse 74 3.5.1.2 Distanzanalyse 75 3.5.1.3 maximum likelihood-Analyse 80 3.5.1.4 Bayes’sche Analyse 80 3.5.1.5 Zusammenfassung 18S rDNA 80 3.5.2 28S rDNA 81 3.5.2.1 Maximum Parsimonie-Analyse 81 3.5.2.2 Distanzanalyse 81 3.5.2.3 maximum likelihood-Analyse 82 3.5.2.4 Bayes’sche Analyse 82 3.5.2.5 Zusammenfassung 88 3.5.3 16S rDNA 88 3.5.3.1 Maximum Parsimonie-Analyse 88 3.5.3.2 Distanzanalyse 89 3.5.3.3 maximum likelihood-Analyse 89 3.5.3.4 Bayes’sche Analyse 90 3.5.3.5 Zusammenfassung 16S rDNA 90 3.6 Phylogenie der Anaspidea (inklusive Pteropoda) 96 3.6.1 18S rDNA 96 3.6.1.1 Maximum Parsimonie-Analyse 96 3.6.1.2 Distanzanalyse 96 3.6.1.3 maximum likelihood-Analyse 101 3.6.1.4 Bayes’sche Analyse 101 3.6.1.5 Zusammenfassung 101 3.6.2 ITS1-5.8S-ITS2 102 3.6.2.1 Maximum Parsimonie-Analyse 102 3.6.2.2 Distanzanalyse 102 3.6.2.3 maximum likelihood-Analyse 102 3.6.2.4 Bayes’sche Analyse 102 3.2.6.5 Zusammenfassung 104 3.7 Phylogenie von Phyllodesmium 104 3.8 Umbraculum 107

4 Diskussion 108 4.1 Auswahl der Taxa 108 4.2 Untersuchte Gene 109 4.3 Maximum Parsimonie-Analysen 114 4.4 Distanzanalysen 115 4.5 maximum likelihood-Analysen 116 4.6 Bayes’sche Analysen 117 4.7 Statistische Tests 118 4.8 Phylogenetisches Signal 119 4.9 Phylogenie der Opisthobranchia 119 4.9.1 Wahl der Außengruppe 119 4.9.2 Status der Opisthobranchia, Pulmonata und Euthyneura 120 4.9.3 Pulmonata 121 4.9.4 Pyramidellidae 125 4.9.5 Opisthobranchia 126 4.9.5.1 Nudibranchia 126 4.9.5.2 Pleurobranchoidea 127 4.9.5.3 Nudipleura 129 4.9.5.4 Tylodinoidea 130 4.9.5.5 Acteonoidea 132 4.9.5.6 Anaspidea und Pteropoda 134 4.9.5.7 Cephalaspidea s.str. 136 4.9.5.8 Cephalaspidea und Anaspidea 138 4.9.5.9 Sacoglossa 139 4.9.5.10 Acochlidiacea 141 4.10 Prüfung der Plausibilität der
rekonstruierten Verwandtschaftshypothesen 142 4.10.1 Vergleich mit Fossilfunden 142 4.10.2 Ernährungsweise 145 4.10.3 Chromosomenzahlen 152 4.11 Phylogenie der Anaspidea 153 4.12 Phylogenie von Phyllodesmium 156 4.13 Umbraculum 158 4.14 Zusammenfassung, Schlusswort und Ausblick 158
Literaturverzeichnis 162 Danksagung 184 Lebenslauf 185
Anhang 186
I Abkürzungen und Symbole 186 II Chemikalien, Lösungen, Medien, Geräte und Kits 187
Chemikalien 187 Rezepte für Lösungen und Medien 187

Kits 188 Geräte 189
III relative rate test 189 Opisthobranchia-Datensatz (18S rDNA) 189 Opisthobranchia-Datensatz (28S rDNA) 217 Opisthobranchia-Datensatz (16S rDNA) 229 Anaspidea-Datensatz (18S rDNA) 240 Anaspidea-Datensatz (ITS1-5.8S-ITS2) 242 Aeolidoidea-Datensatz (18S rDNA) 243
IV Distanzen 247 Opisthobranchia-Datensatz (18S rDNA) 247 Opisthobranchia-Datensatz (16S rDNA) 258 Opisthobranchia-Datensatz (28S rDNA) 262 Anaspidea-Datensatz (18S rDNA) 264 Anaspidea-Datensatz (ITS1-5.8S-ITS2) 264 Aeolidoidea-Datensatz (18S rDNA) 265
V Chromosomenzahlen 266

1
1 Einleitung 1.1 Die Systematik Überall in unserem Alltag begegnen uns die verschiedensten systematischen Klassifikationen, z.B. bei der Anordnung der Produkte in einem Supermarkt oder der Bücher in einer Bibliothek. Diese Klassifikationen sind nicht nur nützliche, sondern essentielle Werkzeuge, um sich in unserer komplexen Umwelt zurechtfinden zu können. Dabei können durchaus verschiedene, sich überschneidende Ordnungen nebeneinander existieren, die je nach Situationen herangezogen werden. Der Ausgangspunkt für das Errichten solcher Ordnungen ist die systematische Klassifikation der Sinneseindrücke, denen unterschiedliche Konzepte von Dingen gegenübergestellt werden. Zu den Methoden für die Erstellung von Begriffs-konzeptionen gehören das Beobachten und Vergleichen, sowie das Erkennen und Beschreiben. Als zentrales Prinzip zur Klassifikation dient dabei die wahrgenommene Ähnlichkeit. Das Ziel einer Klassifikation ist das Erstellen einer allgemeinen und natürlichen Ordnung der Dinge. Sie beinhaltet im Idealfall eine komplette Auflistung aller natürlichen Klassen von Dingen und darüber hinaus alle Wechselbeziehungen der Dinge untereinander. Die Systematik in der Biologie hat sich den Versuch einer Klassifikation auf der Grundlage der phylogenetischen Beziehungen aller Organismen zur Aufgabe gestellt. Dabei stellt die Systematik eine historische Wissenschaft dar, da man die Stammesgeschichte der Organismen ohne Zeitmaschine nicht beobachten, sondern nur rekonstruieren kann. Daher werden alle Verwandtschaftshypothesen immer Hypothesen bleiben. Die Geschichte der Systematik entstand wohl mit Beginn der Menschheitsgeschichte selbst, durch die Fähigkeit des Menschen, im Gedächtnis Gespeichertes willentlich und spontan abrufen zu können. Nur wenn man sich daran erinnern kann etwas Ähnliches bereits wahrgenommen zu haben, kann man auf die Idee kommen, es handele sich um Vertreter des gleichen Typs von Wahrnehmung und sich so einen Begriff davon machen. Vielleicht bildete aber auch erst die Entstehung der Sprache das notwendige Werkzeug für die Entwicklung der Systematik. Erste Überlieferungen biologischer Klassifikationen stammen aus der Antike, von Aristoteles. Er teilte die ihm bekannten Lebewesen auf der Grundlage von vergleichenden Untersuchungen in „Bluttiere“ (Fische, Amphibien, Säuger, Vögel) einerseits und „blutlose Tiere“ (Schalen-, Krusten-, Weichtiere) andererseits und nahm damit bereits die auch heute noch gültige Unterscheidung von Vertebraten und Invertebraten vorweg. Mit dem Zusammenbruch der griechisch-römischen Kultur allerdings sollte die Beobachtung und der empirische Vergleich nicht länger als Basis für die Klassifikation dienen. Während des Mittelalters wurde nicht zwischen Beobachtung, Dokument und Fabel unterschieden. Verfasser von Enzyklopädien verließen sich nicht auf ihre eigene Augen sondern stützten sich auf Erzählungen und Werke

2
anderer Autoren. Berichte von seltsamen Tieren und Fabelwesen wurden als beobachtete Fakten behandelt. Darüber hinaus galt ein grundlegend anderes Verständnis von Natur. Man war beispielsweise der Auffassung, dass Lebewesen Markierungen besitzen, die Aussagen über dessen Wesen und Eigenschaften machen, z.B. herzförmige Blätter – Einfluss auf das menschliche Herz, oder dass Verwandtschaft sich in räumlicher Nähe ausdrückt. Aus diesen „Geschichten der Natur“ wurde in der Mitte des 17. Jahrhunderts durch Jan Jonston und seine „Historia naturalis de quadripedibus libri“ die Naturgeschichte. Beobachtung, Dokument und Fabel waren von da an klar voneinander getrennt, es ging um die eindeutige Benennung des Sichtbaren. Dies war auch die Geburtsstunde der großen Zeit biologischer Klassifikationen, z.B. von Carl von Linné und Georges Louis de Buffon. Das Ziel war die Entwicklung einer Art „Primärsprache“, durch die Beobachtung und Beschreibung vom jeweiligen Beobachter unabhängig werden sollen. Damit war der Begriff der Struktur geboren, der zwischen Wahrnehmung und Sprache vermitteln sollte. Die Theorie des taxonomischen Merkmals entstand. Ein solches Merkmal beschreibt gleichzeitig das Einzelwesen und seine Stellung im System aller Lebewesen. Das Binomen wurde als Artbezeichnung eingeführt. Nicht alle Strukturen können naturgemäß als taxonomisches Merkmal dienen. Die Schwierigkeit besteht darin, die geeigneten unter einer unermesslichen Zahl von möglichen Merkmalen auszuwählen. Wegen der klaren und häufig sehr stabilen Formen der Pflanzen kam es zu einem „Boom“ der Botanik zu dieser Zeit. Mit Hilfe der Naturgeschichte sollte der göttliche Schöpfungsplan aufgedeckt werden, der Evolutionsgedanke war zu dieser Zeit noch nicht gedacht. Durch eine Wende in der Philosophie durch Kant Ende des 18. Jahrhunderts rückte die Zoologie in den Mittelpunkt der Klassifikationsforschung. Anfang des 19. Jahrhunderts kam der Begriff der Organisation auf, er bezeichnete den inneren Körperbau eines Lebewesens. Der Begriff der Funktion erhielt eine zentrale Bedeutung, da durch ihn eine Wertung der Merkmale in wichtige und weniger wichtige möglich wurde. Lamarck trennte schließlich die Nomenklatur von der Klassifikation, d.h. die Stellung im künstlichen Ordnungssystem von der natürlichen Stellung im „System des Lebens“. Er hat dadurch der Naturgeschichte ein Ende gesetzt und die Wissenschaft der Biologie erschaffen. Mit Cuvier kam der Gedanke von Grundformen und Organtypen auf, er stellte der Funktion die verschiedenen entsprechenden Organe gegenüber und versuchte über den Vergleich der Organe auf Ähnlichkeiten zu schließen. Im Laufe der Entwicklung der Systematik entstanden neben den verschiedenen Disziplinen der Biologie auch verschiedene Klassifikationssysteme, durch unterschiedliche Fragestellung und die betrachtete Organisationsebene bedingt. Der Morphologie und Paläontologie in Person von Charles Darwin und Alfred Russel Wallace blieb es jedoch vorbehalten, die moderne Evolutionstheorie zu formulieren. In Deutschland vertrat Ernst Haeckel Darwins Thesen und entwickelte erste konkrete Rekonstruktionen der Stammesgeschichte von Tieren. Er prägte den Begriff der

3
Phylogenie für die Abstammungsgeschichte. Damit war der Forschungszweig der Phylogenetik geboren, der seitdem die Taxonomie von der Systematik getrennt hat. 1.2 Die phylogenetische Systematik Von Willi Hennig ging in den 50er Jahren unseres Jahrhunderts eine neue Form der Verwandtschaftsforschung der Tiergruppen aus. Er verknüpfte bereits vorher vorhandene Techniken zu einer begründeten phylogenetischen Systematik. Unter einem phylogenetischen System verstand er eine natürliche Klassifikation, die die phylogenetischen Verwandtschaftsbeziehungen zwischen den einzelnen Organismen und Organismengruppen widerspiegelt. Zur Aufdeckung verwandtschaftlicher Beziehungen musste der Begriff des homologen Merkmals stärker differenziert werden. Es war zu unterscheiden zwischen Symplesiomorphien, gemeinsamen ursprünglichen Merkmalen, die sich nicht für die Begründung stammesgeschichtlicher Beziehungen eignen und Synapomorphien, gemeinsamen abgeleiteten Merkmalen. Nach diesem als Kladistik bezeichneten Prinzip sollte jedes aufgestellte Taxon eine monophyletische Gruppe repräsentieren, sich also von einem gemeinsamen Vorfahren ableiten lassen und alle Nachkommen dieses Vorfahren umfassen. Paraphyletische Einheiten sollten nach Möglichkeit vollständig aus dem wissenschaftlichen Sprachgebrauch verschwinden. In der vorliegenden Arbeit werden allgemein als Paraphyla angesehene Gruppierungen durch Anführungszeichen kenntlich gemacht. 1.3 Die molekulare Phylogenetik Die Molekulare Phylogenetik beschäftigt sich damit, die Entstehungsgeschichte und die Verwandtschaftsverhältnisse von Lebewesen aufgrund molekularer Merkmale (DNA, RNA, Proteine) zu rekonstruieren. Bis in die Anfänge des 20. Jahrhunderts hinein bildeten morphologische Merkmale allein die Grundlage für die Untersuchung verwandtschaftlicher Beziehungen, und auch heute ist der Informationsgehalt komplexer, morphologischer Merkmale zur Begründung monophyletischer Gruppen vielfach höher zu bewerten als der von Sequenzanalysen. Sequenzen weisen dagegen in anderen Bereichen Vorteile auf. Sie sind beispielsweise bei der Unterscheidung morphologisch identischer Arten von entscheidender Bedeutung und vermögen Homologien für sehr weit entfernte Taxa, z.B. Protisten und Wirbeltiere, zu liefern. Der Grundstein für die Entstehung der molekularen Systematik wurde bereits zu Beginn des 20. Jahrhunderts gelegt. Die Biologen Nuttal und Uhlenhuth (Nuttal 1904) mischten Sera und Antisera verschiedener Spezies mit dem Ziel, eine Art „Blutverwandtschaft“ aufzudecken. Die zugrundeliegende Theorie besagte, dass je näher Arten verwandt sind, die Reaktion zwischen Serum und Antiserum umso stärker ausfällt. Heute wissen wir, dass das von der genetischen Übereinstimmung der Arten abhängig ist. Durch diese Versuche etablierten Nuttal und Uhlenhuth das

4
zentrale Prinzip der molekularen Evolution: Der Grad der evolutiven Verwandtschaft spiegelt sich im Grad genetischer Ähnlichkeit wider. Eines der ersten Beispiele für molekulare Systematik ist die Arbeit von Sarich und Wilson aus dem Jahre 1967. Durch Messung der immunologischen Reaktion von Serumalbuminen verschiedener Primaten konnten die Autoren zeigen, dass Menschen, Schimpansen und Gorillas im Bezug auf das untersuchte Protein genetische Übereinstimmung aufweisen und sich vom Orang Utang deutlich abgrenzen lassen. Aus ihren Ergebnissen konnten sie die phylogenetische Entstehung von Mensch, Schimpanse und Gorilla auf einen Zeitpunkt vor ca. 5 Millionen Jahren datieren und lösten damit heftige Diskussionen aus, denn die bis zu diesem Tage gültige Theorie ging von ca. 15 Millionen Jahren getrennter Entwicklung aus. Erst 1982 (Andrew & Cronin) wurde die Theorie von Sarich und Wilson durch neue Fossilfunde bestätigt. Inzwischen sind die molekularen Methoden der Verwandtschaftsforschung aus der Biologie nicht mehr wegzudenken. In den Anfängen der molekularen Phylogenetik als Wunder- und Allheilmittel gepriesen, haben sie sich inzwischen zu einem essentiellen aber durchaus auch kritisch beurteilten Hilfsmittel gewandelt. 1.4 Die Gastropoda Die Gastropoden sind eine der mannigfaltigsten Tiergruppen überhaupt, das gilt gleichermaßen für ihr Aussehen und ihre Lebensweise. Die Gastropoden gehören zu den ältesten und aus evolutionärer Sicht erfolgreichsten Tiergruppen der Erde. Sie haben nahezu alle marinen, limnischen und terrestrischen Habitate besiedelt und eine Diversität auf allen taxonomischen Ebenen erreicht, die nur von der der Insekten übertroffen wird. Schätzungen der heute existierenden Artenzahl reichen von 40.000 (Boss 1971) bis mehr als 100.000 (Götting 1974), sogar bis 150.000 (Ponder & Lindberg 1997), mit über 13.000 benannten Gattungen rezenter und fossiler Arten (Bouchet & Rocroi 1992). Die Monophylie der Gastropoda ist nicht unbestritten (Stasek 1972), es gibt nur eine Synapomorphie die allgemein akzeptiert wird, die Torsion. Einige Autoren sehen z.B. die Bellerophonten (paläozoische Mollusken mit planspiraler Schale) als Gruppierung an, aus der sich die rezenten, paraphyletischen Gastropoden in mehreren Linien entwickelt haben (Edlinger 1988). Die Entwicklungsgeschichte der Gastropoden begann vor schätzungsweise 570 Millionen Jahren (Runnegar 1983), die ihrer systematischen Erforschung vor 200 bis 250 Jahren (Cuvier, Linné). Im 19. Jahrhundert existierten bereits zahlreiche unterschiedliche Theorien zur Entwicklungsgeschichte der Schnecken, in den meisten Fällen basierend auf der Betrachtung nur eines Organsystems (z.B. der Radulae (Troschel 1856-93) oder des Nervensystems (Bouvier 1887)). Alle wesentlichen Unsicherheiten wurden vorläufig durch Thiele (1929-31) beendet, er lieferte eine Synthese und Verfeinerung früherer Klassifikationen. Er etablierte und

5
modifizierte das Konzept der 3 Unterklassen „Prosobranchia“, Opisthobranchia und Pulmonata von Milne-Edwards (1848) und gliederte die „Prosobranchia“ in die ursprünglichen Archaeogastropoda, die „mittleren“ Mesogastropoda und die abgeleiteten Stenoglossa (=Neogastropoda, Wenz 1938-44). Opisthobranchia und Pulmonata wurden dabei den „Prosobranchia“ als Euthyneura (Spengel 1881) gegenübergestellt. Diese Klassifikation wurde auf breiter Basis akzeptiert und wird es von einigen Autoren bis heute. Auf der anderen Seite dauerte es nicht lange bis erste Modifikationen am Thieleschen System vorgenommen wurden. Die seit Thiele wahrscheinlich bedeutendsten, jedoch auch kontrovers diskutierten Beiträge zur Phylogenie der Gastropoden stammen von Salvini-Plawen und Haszprunar (Salvini-Plawen 1980, 1990; Salvini-Plawen & Haszprunar 1987; Haszprunar 1985b, 1988a,b). Haszprunar stellte 1985 das sog. Heterobranchia-Konzept vor, ein nicht grundsätzlich neues, sondern durch umfassende Analysen begründetes und modifiziertes System der Gastropoden. Nach Haszprunar bilden die paraphyletischen Opisthobranchia mit den Gymnomorpha und den Pulmonaten die Pentaganglionata (nov. = Euthyneura), hauptsächlich gestützt durch das Vorhandensein von 2 zusätzlichen (parietalen) Ganglien. Die Euthyneurie stellt dagegen eine konvergente Erscheinung innerhalb der neubenannten Gruppierung dar. Die Schwestergruppe der Pentaganglionata sind die triganglionaten Allogastropoda (Architectonicoidea, Pyramidelloidea, Nerineoidea). Beide Gruppen werden als Heterobranchia zusammengefasst und den restlichen „Prosobranchia“ gegenübergestellt. In den letzten 10 Jahren ist eine Fülle von Arbeiten publiziert worden, in denen Sequenzdaten zur Untersuchung der phylogenetischen Verwandtschafts-beziehungen innerhalb der Gastropoden verwendet wurden, hauptsächlich basierend auf 18S und 28S rDNA-Sequenzen (z.B. Tillier et al. 1992, 1994; Rosenberg et al. 1994, 1997; Winnepenninckx et al. 1994, 1996, 1998; Harasewych et al. 1997a, 1997b, 1998; Colgan et al. 2000; Harasewych & McArthur 2000). Im Wesentlichen haben diese Arbeiten übereinstimmend dieselben monophyletischen Gruppen rekonstruiert: Patellogastropoda („echte“ Napfschnecken), Cocculinoidea und Lepetelloidea (Tiefsee-Napfschnecken), Neritopsina (terrestrisch, limnisch und marin) Neomphalina (endemisch in Faunengemeinschaft heißer Tiefseequellen), Vetigastropoda (z.B. Schlitzbandschnecken mit “lebendem Fossil“ Pleurotomaria, Meerohr Haliotis, Kreiselschnecken, Lochschnecken), Caenogastropoda (z.B. limnische Apfelschnecken, riesige Fechterschnecken, Pantoffelschnecken, Kauris, Wellhornschnecken, giftige Kegelschnecken) und Heterobranchia (Allogastropoda, Opisthobranchia, Pulmonata). Uneinigkeit besteht allerdings in der Stellung dieser Taxa zueinander. McArthur & Harasewych (2003) nutzten die Daten aus 11 Publikationen über die Phylogenie der Gastropoda, von „präkladistischen“, morphologischen Arbeiten (Thiele 1929-31) bis hin zu molekularen Analysen (Colgan et al. 2000), um mit Hilfe der supertree-Methode (Baum 1992, Ragan 1992) eine Art Konsensustopologie zu erstellen. Die resultierende Verwandtschaftshypothese erscheint ziemlich

6
konventionell, mit basaler Stellung der Patellogastropoda und Monophylie aller Hauptlinien, mit Ausnahme der Vetigastropoda (Abb. 1). Die Monophylie der Euthyneura (Opisthobranchia + Pulmonata) gilt, basierend auf morphologischen (z.B. Salvini-Plawen & Steiner 1996, Dayrat & Tillier 2002) und molekularen Arbeiten (z.B. Wade & Mordan 2000) als gut gestützt und die Pulmonata werden ebenfalls von der überwiegenden Anzahl der Autoren als monophyletisch angesehen (z.B. Wade & Mordan 2000, Dayrat & Tillier 2002). Dagegen wird der Status der Opisthobranchia als Monophylum (Gosliner & Ghiselin 1984, Salvini-Plawen 1991, Wade & Mordan 2000, Medina et al. 2001, Ueshima 2001, Grande et al. 2002, Mikkelsen 2002) oder Paraphylum (Haszprunar 1985, Tillier et al. 1994, 1996, Salvini-Plawen & Steiner 1996, Ponder & Lindberg 1997, Winnepenninckx et al. 1998, Thollesson 1999a, Wollscheid & Wägele 1999, Dayrat et al. 2001, Vonnemann et al. 2005, Wägele et al. 2003) kontrovers diskutiert. 1.5 Die Opisthobranchia Die Opisthobranchia oder Hinterkiemer sind eine Vereinigung morphologisch sehr heterogener Schnecken, die weltweit alle marinen Habitate besiedelt haben. Einige Vertreter der Hinterkiemer, die Acochlidiacea, sind sogar ins Süßwasser vorgedrungen. Ihr Aussehen reicht von unscheinbaren, im Sand grabenden „Prosobranchia“- ähnlichen Vertretern, bis hin zu äußerst auffallend gefärbten, höchst abgeleiteten, schwimmenden Formen. Die kleinsten Individuen (Microhedylidae, Acochlidiacea) messen nur wenig mehr als 1mm (Arnaud, Poizat & Salvini-Plawen 1986), die größte Art (Aplysia vaccaria Winkler, 1955; Anaspidea) kann eine Länge von fast 1m erreichen (Behrens 1980). Die ältesten Fossilfunde von Opisthobranchiern wurden auf das Untere Karbon (ca. 310 Millionen Jahre) datiert und erinnern an rezente Vertreter der Scaphandridae
Patellogastropoda
Cocculinoidea
Lepetelloidea
Neritopsina
Caenogastropoda
Heterobranchia
Neomphalina
Pleurotomarioidea
Vetigastropoda
Abb 1: Phylogenie der Gastropoda (aus McArthur & Harasewych 2003, verändert). Konsensustopolgie
basierend auf Daten aus 11 Publikationen über die Phylogenie der Gastropoda, erstellt mit Hilfe der
supertree-Methode (Baum 1992, Ragan 1992).

7
und Retusidae (Cephalaspidea) (Gosliner 1981). Die Anzahl rezenter Arten der Opisthobranchia wird auf etwa 6000 geschätzt (Wägele et al. in Vorbereitung) und variiert erheblich zwischen den einzelnen Subtaxa (z.B. Tylodinoidea mit etwa 15 Arten, Nudibranchia mit annähernd 3000 Arten). Trotz aller Heterogenität innerhalb der Opisthobranchia, lassen sich einige morphologische Trends klar erkennen. Die Reduktion der Schale bis hin zu ihrem völligen Verlust hat unabhängig in mehreren Linien stattgefunden. Als ursprünglich gelten dabei „Prosobranchier“-ähnliche, spiralige Gehäuse, davon ausgehend ist eine Entwicklung über involute, „bulloide“ Schalen bis hin zu internen, vom Mantelgewebe überwachsenen Schalen vorstellbar. Ein letzter Schritt wäre anschließend die vollständige Reduktion des Gehäuses. Diese Transformation ist in mehreren Gruppen anhand von Übergangsformen nachvollziehbar. Verbunden mit dem Verlust der Schale als Schutzorgan (bzw. schon vorher entstanden, Präadaption), ist die Entwicklung von diversen Verteidigungsstrategien innerhalb der Opisthobranchia. Die sehr häufig bei Hinterkiemern zu beobachtende auffällige Färbung ist meist ein Signal für toxische oder schlecht schmeckende Substanzen. Diese können entweder mit der Nahrung, z.B. Schwämmen, aufgenommen werden (z.B. Tylodinoidea, Ebel et al. 1999) oder de novo von der Schnecke synthetisiert werden (z.B. Sacoglossa, Jensen 1997a). Ein weiterer Verteidigungsmechanismus ist das Abwerfen von Körperteilen (Autotomie), z.B. von Körperanhängen (Phyllodesmium, Nudibranchia) oder des Mantelrandes (Berthella, Pleurobranchoidea). Bei den Anaspidea besitzen die meisten Vertreter Drüsen, die ein farbiges Sekret produzieren, das bei Gefahr ausgestoßen werden kann. Die wohl erstaunlichste Abwehrstrategie ist bei den Aeolidoidea (Nudibranchia) verwirklicht, die sich von Cnidariern ernähren. Mit der Nahrung werden auch die Nesselzellen aufgenommen, die den Darmtrakt unbeschadet passieren und durch Ausläufer der Mitteldarmdrüse in die Spitzen von Körperanhängen gelangen. Dort werden sie gespeichert und bei Berührung zur Feindabwehr ausgestoßen. Ein weiterer Trend innerhalb der Hinterkiemer ist die Aufhebung der Torsion, die bei larvalen Opisthobranchiern ebenso wie bei allen „prosobranchiaten“ Formen zu beobachten ist. Durch diese Detorsion nimmt der Körper eine, zumindest äußerliche, Bilateralsymmetrie ein. Zusammen mit Schalenreduktion und Detorsion, aber weitestgehend unabhängig von diesen beiden Prozessen, findet eine Entwicklung in Richtung Entspiralisierung und Streckung des Körpers statt. Verbunden mit dieser Entspiralisierung ist zum Beispiel die Entwicklung von der streptoneuren Organisation mit überkreuzten Nervensträngen hin zum euthyneuren Zustand. Die Euthyneurie ist, genau wie der Verlust der Schale, mehrfach unabhängig entstanden.

8
Die Opisthobranchia sind fast ausnahmslos zwittrig, mit einem Trend zur Ausbildung von separaten Gängen für männliche und weibliche Gameten sowie für Eigen- und Fremdspermien. Für die Erscheinung dieser zahlreichen konvergenten Entwicklungen ist im englischsprachigen Raum der Begriff des rampant parallelism („zügelloser Parallelismus“, Gosliner & Ghiselin 1984) geprägt worden. Er ist wohl als Hauptgrund für die trotz zahlreicher Untersuchungen immer noch bestehenden Unsicherheiten hinsichtlich der Phylogenie der Opisthobranchia anzusehen. Denn eine Folge dieses Parallelismus ist die geringe Anzahl von synapomorphen Merkmalen zur Begründung von Verwandtschaftsbeziehungen zwischen den verschiedenen Großgruppen. Dagegen lassen sich die einzelnen Subtaxa der Opisthobranchia aufgrund morphologischer und ökologischer Merkmale meist gut voneinander abgrenzen. Cephalaspidea P. Fischer 1883 (Kopfschildschnecken) Die Cephalaspidea s.str. (Abb. 2A) stellen nach Ausgliederung der basalen Acteonoidea eine monophyletische Gruppierung (Mikkelsen 1996) mit ca. 900 Arten dar. Der deutsche Name Kopfschildschnecken bezieht sich auf den meist schildartig verbreiterten Kopf, mit dem viele Arten während der Nahrungssuche (z.B. Polychaeten, Foraminiferen, Muscheln, Schnecken) durch den Boden pflügen. Bei diesem Merkmal handelt es sich eher um eine ökophänotypische Anpassung an die charakteristische, grabende Lebensweise als um eine Synapomorphie der Gruppe. Als Apomorphien betrachtet man z.B. 3 gehärtete Platten im Kaumagen, die für die Nahrungszerkleinerung von Bedeutung sind (Mikkelsen 1996). Die Cephalaspidea s.str. besitzen teilweise äußere Gehäuse, teilweise vom Mantel überwachsene, seltener sind sie ganz reduziert. Ihre Kiemen liegen, dem plesiomorphen Zustand bei den Opisthobranchia entsprechend, in einer Mantelhöhle. Acteonoidea d'Orbigny, 1842 Die Acteonoidea (Abb. 2B), die man früher zu den Cephalaspidea gezählt hat, bilden eine basale, eventuell paraphyletische Gruppierung innerhalb der Opisthobranchia oder werden sogar ganz aus den Opisthobranchia ausgeschlossen (Mikkelsen 2002). Die Acteonoidea umfassen ca. 200 Arten mit überwiegend deutlich ausgeprägten Gehäusen. Lange Zeit galt die namensgebende Gattung Acteon als Paradebeispiel eines ursprünglichen Hinterkiemers, da sie sowohl „prosobranche“ als auch opisthobranche Merkmale in sich vereint. Gleichzeitig weist Acteon aber auch einige „hochentwickelte“ anatomische Merkmale auf, z.B. eine deutlich abgeleitete Radulabezahnung. Die Stellung der Acteonoidea scheint daher nicht so eindeutig wie man oft vermutet hat. Die Acteonoidea sind carnivor und ernähren sich z.B. von Polychaeten und anderen Invertebraten.

9
Abb 2: Vertreter der opisthobranchiaten Großgruppen. A: Chelidonura sp. (Cephalaspidea s.str.,
www.coralreefnetwork.com), B: Bullina lineata (Acteonoidea, www.daveharasti.com), C: Aplysia californica
(Anaspidea, www.uoregon.edu), D: Creseis acicula (Thecosomata, www.dafni.com), E: Clione limacina
(Gymnosomata, www.ii.uam.es), F: Thuridilla lineolata (Sacoglossa, www.edge-of-reef.com), G: Pontohedyle sp.
(Acochlidiacea, © Michael Schrödl), H: Pleurobranchus forskalii (Pleurobranchoidea, www.edge-of-reef.com), I:
Umbraculum umbraculum (Tylodinoidea, © Wolfgang Seifarth), J: Hypselodoris bullockii (Nudibranchia,
www.visiondive.com), K: Phyllodesmium jakobsenae (Nudibranchia, © Ingo Burghardt).
A
C
D
H
K
B
E F
G I
J

10
Anaspidea P. Fischer, 1883 (Seehasen) Die Anaspidea heißen wegen der ohrförmigen Tentakel vieler Arten Seehasen und ihre 80 Arten besitzen entweder eine kleine interne, eine deutliche externe oder eine völlig reduzierte Schale. Die Gattung Aplysia (Abb. 2C) ist wegen ihrer Riesenneurone als Modellorganismus für Neurobiologen bekannt. Die Anaspidea sind ausschließlich herbivor und leben überwiegend in küstennahen Seegraswiesen und Kelpwäldern. Sie werden aufgrund von zahlreichen Apomorphien (Gosliner 1991, Mikkelsen 1996, Medina & Walsh 2000) als Monophylum akzeptiert. Dazu gehören z.B. 2 besondere Merkmale des Verdauungsapparates: das Vorhandensein einer Filterkammer zwischen Kaumagen und Magen und die Existenz eines Magenblindsacks. Einige Vertreter der Anaspidea enthalten pharmazeutisch bedeutsame Inhaltsstoffe, z.B. gilt das aus der Art Dolabella auricularia isolierte Dolastatin (Pettit et al. 1987) als einer der vielversprechendsten Wirkstoffe zur Krebsbekämpfung (Carté 1996). Thecosomata Blainville, 1824 (Meeresschmetterlinge) Die Thecosomata (Abb. 2D) stellen die größte Gruppe planktonischer Gastropoden im Bezug auf Arten- und Individuenzahl dar (Lalli & Wells 1978). Der ursprüngliche Fuß der Schnecken hat sich zu einem Paar „Flügel“ (Parapodien) umgewandelt, die es ihnen ermöglichen, in der Wassersäule zu schweben. Ihr Gehäuse ist vorhanden oder reduziert und teilweise wurden sekundäre Gehäuse ausgebildet (Pseudoconcha). Ihre Kiemen sind ebenfalls reduziert. Die Thecosomata ernähren sich von Phytoplankton, das mit Hilfe von Wimpern herangestrudelt wird. Die Monophylie der Gruppe ist aufgrund der spezialisierten Lebensweise und der damit verbundenen morphologischen Anpassungen als wahrscheinlich anzusehen, ist aber bisher nicht durch phylogenetische Analysen gestützt. Gymnosomata Blainville, 1824 Die Gymnosomata (Abb. 2E) zeigen wie die Thecosomata eine planktonische Lebensweise. Ihre Schale ist jedoch bei allen Vertretern vollständig reduziert, ebenso wie die Kiemen. Die Gymnosomata ernähren sich vor allem von den Thecosomata und haben im Zuge dieser carnivoren Lebensweise zum Teil diverse Fangvorrichtungen, wie z.B. Saugarme entwickelt. Ebenso wie für Thecosomata wird ihre Monophylie angenommen, phylogenetische Untersuchungen fehlen aber bisher. Einige Arten der Gymnosomata und Thecosomata kommen in riesigen Schwärmen in arktischen und antarktischen Gewässern vor und stellen eine wichtige Nahrungsquelle für Bartenwale dar.

11
Sacoglossa Von Ihering, 1876 (Schlundsackschnecken) Die Sacoglossa oder Schlundsackschnecken umfassen ungefähr 350 Arten und tragen ihren Namen aufgrund eines Blindsackes, in den funktionslos gewordene Radulazähne aufgenommen werden und der eine der Apomorphien dieser Gruppe darstellt (Jensen 1996, Mikkelsen 1998). Einige Vertreter haben ihre Schale komplett reduziert (Abb. 2F), andere besitzen noch deutlich ausgeprägte äußere Gehäuse. Trotz ihres teilweise sehr unterschiedlichen Aussehens sind sie eine ökologisch sehr einheitliche Gruppe, nahezu alle Sacoglossa saugen Cytoplasma aus marinen Pflanzen. Dabei bleiben bei manchen Arten die mit der Nahrung aufgenommenen Chloroplasten funktionsfähig und gehen eine symbiontische Beziehung mit der Schnecke ein. Wie bei den Anaspidea liegen die Kiemen bei schalentragenden Formen in einer Mantelhöhle, bei gehäuselosen Sacoglossen sind sie reduziert und die Atmung erfolgt über die Epidermis. Zu den Sacoglossa zählen auch die ungewöhnlichen Gattungen Berthelinia, Julia und Midorigai, die eine muschelähnliche, zweiklappige Schale besitzen. Durch embryologische Untersuchungen konnte gezeigt werden, dass die linke Schalenhälfte dem Gastropodengehäuse homolog ist, während die rechte durch Kalkabscheidungen des Mantels entsteht. Acochlidiacea Odhner, 1937 Die im Sandlückensystem lebenden, wurmförmigen Acochlidiacea (Abb. 2G) mit ihren ca. 20 bekannten Arten zeigen eine einzigartige äußere Organisation. Ihr Eingeweidesack ist gut entwickelt und vom Rest des Körpers deutlich abgesetzt, dabei kann der Kopf-Fuß-Komplex zumindest teilweise in den Eingeweidesack zurückgezogen werden. Das ist die deutlichste Apomorphie dieser Gruppe (Sommerfeldt & Schrödl, im Druck). Ihr Gehäuse ist, wenn überhaupt vorhanden, rudimentär und die Kiemen sind vollständig reduziert. Die Acochlidiacea sind die einzigen Hinterkiemerschnecken die mit einigen Arten auch das Süßwasser besiedelt haben. Pleurobranchoidea de Férussac, 1822 (Seitenkiemer) Die etwa 80 Arten der Pleurobranchoidea (Abb. 2H) oder Seitenkiemer besitzen jeweils eine Kieme, die mit der der übrigen Opisthobranchia (außer der der Nudibranchia) homologisiert werden kann. Sie liegt auf der rechten Körperseite zwischen Mantelrand und Fuß. Die Monophylie der Pleurobranchoidea wird durch zahlreiche Apomorphien gestützt, z.B. durch die vom Mantel bedeckte, nahezu rechteckige Schale oder das Vorhandensein einer sog. Fußdrüse, deren Sekret der Partnerfindung dient.

12
Die Pleurobranchoidea sind carnivor und ernähren sich von Seeanemonen, Seescheiden, Schwämmen und anderen Opisthobranchiern; auch Kannibalismus ist beobachtet worden (Jensen 1997b). Tylodinoidea Gray, 1847 (Schirmschnecken) Die Tylodinoidea oder Schirmschnecken (Abb. 2I) umfassen nur 15 Arten und werden z.B. aufgrund ihrer externen, schirmförmigen Schale als Monophylum vereint. Ein weiteres synapomorphes Merkmal ist ein kutikularisierter Ring im Mund, der wahrscheinlich unterstützend bei der Nahrungsaufnahme wirkt. Genau wie die Pleurobranchoidea besitzen die Schirmschnecken eine seitlich gelegene Kieme. Die Tylodinoidea ernähren sich hauptsächlich von Schwämmen, aus denen sie Substanzen zur Feindabwehr gewinnen. Nudibranchia Blainville, 1814 (Nacktkiemer) Die Nudibranchia stellen etwa 60% der Hinterkiemerarten und tragen ihren Namen Nacktkiemer aufgrund der freien respiratorischen Anhänge. Dabei sind diese Anhänge den anderen Opisthobranchierkiemen wahrscheinlich nicht homolog. Bei allen Nudibranchia ist die Schale vollständig reduziert. Die Nudibranchia werden aufgrund von bemerkenswerten und hochentwickelten Verteidigungs- und Ernährungsstrategien zahlreicher Vertreter als sehr abgeleitet innerhalb der Opisthobranchia angesehen. Viele ihrer Arten sind ausgesprochene Nahrungsspezialisten, deren Beutespektrum manchmal nur auf eine einzige Art beschränkt ist. Oft sind sie strukturell und farblich perfekt an ihre sessile Beute, z.B. Korallen, Moostierchen oder Schwämme angepasst und dadurch für potentielle Fressfeinde kaum zu entdecken. Eine von vielen besonderen Ernährungsstrategien zeigen z.B. Vertreter der Gattung Tethys. Ihr bis zu 30 cm großer Körper besitzt einen riesigen Kopflappen, der am Rand mit Cirren versehen ist und wie ein Schöpftrichter über die Beute (Plankton, Jungfische, Krebse, Stachelhäuter) gestülpt wird. Die Monophylie der Nudibranchia wird durch mehrere Apomorphien gestützt: Neben der vollständigen Reduktion der Schale, die allerdings unabhängig auch in anderen Gruppen stattgefunden hat, z.B. durch den Besitz eines besonderen Epithels mit zahlreichen Vakuolen, des sog. Spezialvakuolenepithels (Wägele & Willan 2000). Dessen Funktion ist bis jetzt noch nicht geklärt. In Abbildung 2J ist ein Vertreter der Anthobranchia dargestellt, Abbildung 2K zeigt ein Individuum der Cladobranchia. 1.6 Bisherige Erkenntnisse über die Phylogenie der Opisthobranchia Bis heute ist die Phylogenie der Opisthobranchia in weiten Teilen ungeklärt, da auf kladistischen Prinzipien basierende, umfassende morphologische Arbeiten bisher fehlen und zudem durch die zahlreichen Konvergenzerscheinungen innerhalb der

13
Opisthobranchia erschwert werden. Die durchgeführten molekularen Analysen haben untereinander, und im Vergleich mit den morphologischen Daten, widersprüchliche Ergebnisse geliefert, sofern die verwandtschaftlichen Beziehungen überhaupt aufgelöst werden konnten. Während die Monophylie der einzelnen Subtaxa recht gut gestützt ist, erscheinen die postulierten Verwandtschaftsbeziehungen der Großguppen zueinander noch sehr hypothetisch. Die Monophylie des gebräuchlichen Taxons Notaspidea (Tylodinoidea + Pleurobranchoidea) ist von mehreren Autoren (Minichev & Starobogatov 1978, Schmekel 1985, Salvini-Plawen 1991, Salvini-Plawen & Steiner 1996, Wägele & Willan 2000) verworfen worden, da die vereinenden Merkmale, z.B. Kieme auf der rechten Körperseite, als plesiomorph erkannt wurden. Schmekel hat 1985 ein Schwestergruppenverhältnis von Nudibranchia und Pleurobranchoidea vorgeschlagen, das seitdem durch einige Analysen bestätigt werden konnte (z.B. Salvini-Plawen 1990, 1991, Wägele & Willan 2000). Von Wägele & Willan (2000) wurde der Name Nudipleura für den Zusammenschluß von Nudibranchia und Pleurobranchoidea eingeführt. Mikkelsen hat in ihrer umfassenden morphologischen Analyse (1996) der Cephalaspidea s.l. ein Schwestergruppenverhältnis zwischen Cephalaspidea s.str. und Anaspidea gefunden, das durch die molekulare Analyse der 16S rDNA von Thollesson (1999) und des 18S rDNA, 16S rDNA und COI Gens von Wägele et al. (2003) gestützt werden konnte. Das aus der von Mikkelsen (2002) durchgeführten, kombinierten Analyse morphologischer Merkmale (Willan 1987, Mikkelsen 1996, Jensen 1996, Wägele & Willan 2000) resultierende Kladogramm zeigte eine recht geringe Auflösung. Der am meisten abgeleitete Zweig bestand aus Tylodinoidea und Nudipleura, die zusammen eine monophyletische Gruppierung mit Anaspidea und Cephalaspidea s.str bildeten. Alle Taxa zusammen stellten die Schwestergruppe der Sacoglossa dar. Die übrigen Großgruppen der Opisthobranchia, z.B. Acochlidiacea, wurden nicht berücksichtigt. Die basale Stellung der Sacoglossa ist auch in der molekularen Analyse von Grande et al. (2004a,b) rekonstruiert worden, weiterhin ein gut gestütztes Monophylum aus Anaspidea, Cephalaspidea s.str. und Tylodinoidea. Sehr wenig ist bekannt über die Position der Acochlidiacea, Thecosomata und Gymnosomata, da kaum phylogenetische Daten verfügbar sind. Gosliner & Ghiselin (1984) präsentierten die Acochlidiacea als Schwestergruppe der Sacoglossa auf der Grundlage von morphologischen Merkmalen. Dayrat et al. (2001) hat basierend auf partiellen 28S rDNA-Sequenzen eine monophyletische Gruppierung aus Thecosomata, Gymnosomata und Anaspidea rekonstruiert, dagegen stehen die Gymnosomata in der von Thollesson (1999) aufgestellten Verwandtschaftshypothese außerhalb der Euthyneura. Die Acteonoidea stellen nach überwiegender Meinung eine ganz basale Gruppierung innerhalb der Opisthobranchia dar, Mikkelsen (2002) ordnet sie sogar in die basalen Heterobranchia ein. Umso erstaunlicher ist es, dass die Acteonoidea bei Grande et al. (2004a,b), basierend auf molekularen Merkmalen, als Schwestergruppe der als abgeleitet angesehenen Nudipleura gruppieren.

14
Abbildung 3 zeigt vier Stammbaumhypothesen der Opisthobranchia, basierend auf morphologischen und molekularen Arbeiten.
Pleurobranchoidea
Nudibranchia
Acteonoidea
Cephalaspidea s.str.
Tylodinoidea
Anaspidea
Pulmonata
Sacoglossa
Pyramidellidae
Pulmonata D
Nudibranchia
Cephalaspidea s.str.
Pteropoda
Anaspidea
Pulmonata
Sacoglossa
Acteonoide
Pulmonata
Tylodinoidea
Pleurobranchoidea C
Nudibranchia
Pleurobranchoidea
Tylodinoidea
Acochlidiacea
Rhodopidae
Gymnosomata
Cephalaspidea s.str.
Anaspidea
Sacoglossa
Cephalaspidea s.str.
Acteonoidea
Thecosomata
Pulmonata
Pyramidellidae A
Nudibranchia
Pleurobranchoidea
Tylodinoidea
Cephalaspidea s.str.
Anaspidea
Sacoglossa
Acteonoidea B
Abb. 3: Stammbaumhypothesen der Opisthobranchia, basierend auf morphologischen und molekularen
Arbeiten. A: Salvini-Plawen & Steiner 1996, morphologisch, B: Mikkelsen 2002, morphologisch, C: Dayrat
& Tillier 2001, molekular, D: Grande et al. 2004a, molekular

15
1.7 Molekulare Marker Für phylogenetische Fragestellungen verwendete Gene, sogenannte molekulare Marker, sollten bestimmte Eigenschaften aufweisen und Bedingungen erfüllen, von denen die wichtigsten nachfolgend kurz dargestellt werden. Ein idealer molekularer Marker sollte im haploiden Genom entweder als Einzelkopie vorliegen oder, beim Vorhandensein multipler Loci, identische Kopien aufweisen. Das ist mit wenigen Ausnahmen bei allen mitochondrialen und nukleären, ribosomalen Genen der Fall. Die Substitutionsrate des untersuchten Gens muss hoch genug sein, um genügend informative Positionen zur Verfügung stellen. Für nah verwandte Arten sollte z.B. ein Gen mit hoher Mutationsrate gewählt werden, damit in der kurzen Zeit seit der Divergenz der Taxa, ausreichend Mutationen akkumulieren konnten. Auf der anderen Seite darf die Substitutionsrate nicht zu hoch sein, da das phylogenetische Signal ansonsten durch multiple Mutationen „verrauscht“ werden könnte. Ein idealer Marker sollte zudem eine annähernd homogene Basenzusammensetzung aufweisen, weil es durch ungleiche Basenfrequenzen zu einer Zunahme an Homoplasien kommen kann. Zusätzlich sollte die Basenzusammensetzung bei allen Taxa eines untersuchten Datensatzes übereinstimmen, da sonst die Gefahr besteht, dass nicht verwandte Taxa nur aufgrund ähnlicher Basenhäufigkeiten zusammen gruppieren. Weiterhin sollte die Homogenität der Substitutionsraten gegeben sein, und zwar sowohl zwischen den Taxa eines Datensatzes, als auch über die ganze Länge des untersuchten Genbereichs innerhalb einer Sequenz hinweg. In der Realität erfüllt kein einziger molekularer Marker alle erwähnten Bedingungen, je nach Fragestellung muss der am besten geeignete ausgewählt werden. In der vorliegenden Arbeit wurden das 18S rRNA-, 28S rRNA-, 16S rRNA-Gen, die Regionen ITS1-ITS2, inklusive des 5.8S rRNA-Gens, sowie die Untereinheit 1 der mitochondrialen Cytochromoxidase untersucht und auf verschiedene Datensätze auf unterschiedlichem taxonomischen Niveau angewendet (siehe Kapitel 1.8). Die Ribosomen sind diejenigen Zellorganellen, die für die Biosynthese von Proteinen zuständig sind. Alle bakteriellen und eukaryontischen Ribosomen bestehen aus zwei Untereinheiten unterschiedlicher Größe, die bei den Eukaryonten als 60S- und 40S-Untereinheit bezeichnet werden; dabei steht das S für Svedberg-Einheit, ein Maß für die Sedimentationsgeschwindigkeit in der Ultrazentrifuge. Die größere 60S-Untereinheit setzt sich zusammen aus drei Arten ribosomaler RNA (28S rRNA, 5,8S rRNA, 5S rRNA) und etwa 40 Proteinen, die kleinere 40S-Untereinheit aus nur einer Sorte von RNA (18S rRNA) und etwa 30 Proteinen.

16
Die Gene für die ribosomale RNA sind im Genom in zahlreichen Kopien hintereinander angeordnet (z.B. 100-120 Kopien bei der Hefe, 120-240 bei Insekten, gilt für große rRNAs). Diese große Anzahl hat sich wahrscheinlich aus der Notwendigkeit heraus entwickelt, große Mengen an rRNA für die Bildung von Ribosomen zur Verfügung zu stellen. Anders als bei proteinkodierenden Genen, bei denen durch einen einzigen Transkriptionsvorgang und durch wiederholte Translation der dabei entstandenen mRNA mehrere Moleküle des Genprodukts entstehen können, liefert die Transkription eines rRNA-Gens immer nur ein Exemplar des Endprodukts. Zwischen den kodierenden Sequenzen befinden sich eine Reihe von sogenannten spacer Regionen. Das sind nicht-kodierende DNA-Sequenzen, die aber für die Bildung des rRNA Transkripts notwendige Signale enthalten. Dazu gehören ein external transcribed spacer (ETS) und zwei internal transcribed spacers (ITS1 und ITS2). Neben diesen gibt es andere spacer-Regionen die offensichtlich außer der eines Platzhalters keine weitere Funktion haben. Die Gene für die ribosomale RNA bilden zusammen mit den spacern eine Transkriptionseinheit, da nur ein einziges RNA-Transkript erzeugt wird. Die einzelnen Transkriptionseinheiten werden durch sog. non-transcribed spacer (NTS) voneinander getrennt (Abb. 4).
Mitochondrien sind die Energielieferanten der Zellen. Ihre DNA ist ringförmig, in der Regel um 16 kb groß und mit wenigen Ausnahmen für 37 Gene kodierend. Mitochondrien besitzen, genau wie die Chloroplasten der Pflanzen, ihr eigenes Translationssystem, bestehend aus 2 Molekülen ribosomaler RNA (16S rRNA: große Untereinheit, 12S rRNA: kleine Untereinheit) und zahlreichen Proteinen. Die ribosomalen Proteine werden allerdings, im Gegensatz zu denen für die rRNAs, von Genen des Zellkerns kodiert, wie überhaupt der überwiegende Teil der mitochondrialen Proteine. Eines der von der mitochondrialen DNA kodierten Proteine ist die mitochondriale Cytochromoxidase (COI). Die Cytochromoxidase ist ein Enzym in der Atmungskette und besteht aus einem Häm-Proteinkomplex, aufgebaut aus 6 Untereinheiten, der die innere Mitochondrienmembran durchsetzt. Ihre Hauptaufgabe ist die Aufnahme von Elektronen im letzten Schritt der Atmungskette.
28S 18S ETS ETS NTS NTS
ITS1 ITS2
5,8S
Abb. 4: Typische eukaryotische Transkriptionseinheit (nach verschiedenen Autoren).

17
Die Verwendung von ribosomalen Genen zur phylogenetischen Rekonstruktion reicht von Untersuchungen der basalen Stammlinien des Lebens bis hin zur Erforschung von Verwandtschaftsbeziehungen auf Populationsebene. Die Gründe für diese Vielseitigkeit liegen in der Existenz unterschiedlicher Evolutionsraten bei verschiedenen rDNA-Genen. So wird im Allgemeinen das Gen für die kleine ribosomale Untereinheit zur Aufklärung präkambrischer Divergenzereignisse herangezogen, da die18S rDNA sehr konserviert ist, also sehr langsam evolviert. Die rRNAs der großen Untereinheit dagegen, dienen aufgrund der schnelleren Evolution einiger Regionen der Untersuchung paläozoischer und mesozoischer Ereignisse. Die mitochondriale rRNA (16S, 12S) wird schließlich zur Analyse von känozoischen Aufspaltungen eingesetzt (Hillis & Dixon 1991) und die nicht-kodierenden spacer-Regionen auf Art- und Populationsebene. Diese Einteilung ist natürlich stark vereinfacht und dementsprechend nicht für alle Fragestellungen und jede Organismengruppe gültig. Die 18S rDNA ist wohl das für phylogenetische Fragestellungen meistverwendete Gen und wurde auf verschiedenen taxonomischen Ebenen eingesetzt, z.B. für die Aufklärung der basalen Verwandtschaftsverhältnisse innerhalb des Tierreichs (Abouheif et al. 1998), für die Phylogenie der Mollusca (Passamaneck et al. 2004), der Gastropoda (Winnepenninckx et al. 1996), der Nudibranchia (Wollscheid & Wägele 1999) und der Gattung Lymnaea (Bargues & Mas-Coma 1997). Das Anwendungsgebiet für das 28S rDNA-Gen entspricht im Wesentlichen dem der 18S rDNA (z.B. Dayrat et al. 2001, Passamanneck et al. 2004, Williams & Reid 2004). Das 16S rDNA-Gen ist gemäß seiner höheren Evolutionsgeschwindigkeit nicht für Untersuchungen auf der Ebene der Tierstämme geeignet, davon abgesehen reicht seine Verwendung von Studien innerhalb einer Gattung (Aplysia, Medina et al. 2001) bis hin zur phylogenetischen Analyse der Euthyneura (Thollesson 1999). ITS1- und ITS2-Sequenzen sind bisher nur auf niederem taxonomischem Niveau verwendet worden, z.B. auf Populationsebene (Raahauge & Kristensen 2000), innerhalb einer Gattung (Stothard et al. 1996, Korte & Armbruster 2003) bis hin zu phylogenetischen Untersuchungen innerhalb einer Familie (van Moorsel et al. 2000, Bargues et al. 2001). Mit Hilfe des COI-Gens sind z.B. die Verwandtschaftsbeziehungen innerhalb der gesamten Gastropoden (Remigio & Hebert 2003) untersucht worden, die Evolution der Euthyneura (Grande et al. 2004a), die Phylogenie der Nudibranchia (Wollscheid-Lengeling et al. 2001), bis hin zur Phylogenie einer Gattung (Fahey 2003).

18
1.8 Ziele der vorliegenden Arbeit Das vorrangige Ziel dieser Arbeit ist die Rekonstruktion der Phylogenie der Opisthobranchia. Dabei stehen die immer noch ungeklärten, verwandtschaftlichen Beziehungen der opisthobranchiaten Hauptlinien im Vordergrund. Das 18S rRNA-Gen wurde für die Rekonstruktion gewählt, da es sich aufgrund universeller Primer gut amplifizieren lässt, eine große aber noch handhabbare Sequenzlänge aufweist und gute Ergebnisse auf verschiedenen taxonomischen Ebenen mit diesem Gen erzielt wurden. Zur Ergänzung des 18S-Datensatzes (91 Sequenzen) wurde ein zweiter Datensatz, basierend auf partiellen 28S rDNA-Sequenzen (54 Sequenzen) erstellt. Dayrat et al. (2001) hat die Bereiche D1 und D2 der 28S rRNA bereits zur phylogenetischen Untersuchung der Euthyneura verwendet und es hat sich in dieser Analyse gezeigt, dass einige Gruppierungen gut gestützt werden, die Auflösung insgesamt aber nicht sehr hoch ist. Mit dem Ziel einer Zunahme an phylogenetischer Information wurde in der vorliegenden Arbeit zusätzlich der Bereich D3 sequenziert. Ein dritter Datensatz für die Opisthobranchia besteht aus der partiellen 16S rDNA (45 Sequenzen). Durch Arbeiten anderer Autoren (z.B. Wollscheid-Lengeling et al. 2001, Thollesson 1999, Medina & Walsh 2000) ist bereits große Anzahl von 16S rDNA-Sequenzen für die Opisthobranchia vorhanden, einige Gruppen wurden aber bisher nicht berücksichtigt, z.B. die Acochlidiacea. In dieser Arbeit wurde der bestehende Datensatz durch neue Sequenzen, z.B. der Acochlidiacea, ergänzt. Neben der Rekonstruktion der Phylogenie der Opisthobranchia, wurden 3 weitere Teilprojekte mit unterschiedlichen Fragestellungen durchgeführt: 1. Phylogenie der Anaspidea Die etwa 80 Arten der Anaspidea verteilen sich auf 10 Gattungen, dabei entfallen allein auf die Gattung Aplysia mehr als 50 Arten. Die bisherigen Klassifikationen teilen die Anaspidea in die beiden Familien Aplysiidae und Akeridae (Beeman 1968, Morton 1972, Thompson & Seaward 1989) ein, von denen die letztere monogenerisch ist (Akera spp.). Aufgrund von morphologischen Merkmalen, z.B. der Form der Parapodien und der Radulazähne werden vier Unterfamilien unterschieden (Eales 1944, Pruvot-Fol 1954, Beeman 1968, Marcus 1972), die Aplysiinae (Aplysia, Syphonota), Dolabellinae (Dolabella), Dolabriferinae (Dolabrifera, Petalifera, Phyllaplysia) und Notarchinae (Stylocheilus, Bursatella, Notarchus). Basierend auf der Länge von Nervensträngen, werden die beiden ersten Unterfamilien zu dem Taxon Longicommissurata zusammengefasst, die beiden letzteren zu den Brevicommissurata (Pruvot-Fol 1954, Marcus 1972). Die Phylogenie der Anaspidea ist bis heute aufgrund widersprüchlicher Verwandtschaftshypothesen noch nicht restlos geklärt. Medina und Walsh (2000)

19
präsentierten einen Stammbaum von 14 Anaspidea-Arten basierend auf den Sequenzen von 3 mitochondrialen Genen; Unterschiede zur traditionellen Klassifikation bestanden in der Paraphylie der Dolabriferinae und Notarchinae. In der umfassenden Analyse der Aplysiidae basierend auf morphologischen und histologischen Merkmalen von Klussmann-Kolb (2004), konnte dagegen die Monophylie der 4 Unterfamilien bestätigt werden, die Longicommissurata erwiesen sich dabei als paraphyletisch. Ziel der vorliegenden Arbeit ist es, die Phylogenie der Anaspidea zu klären. Zu diesem Zweck wurden das komplette 18S rDNA-Gen sowie der Bereich ITS1 - 5.8S - ITS2 von mehreren Vertretern der Anaspidea, Thecosomata und Gymnosomata sequenziert. Die Einbeziehung der beiden letztgenannten Gruppen erfolgte aufgrund der engen Verwandtschaft der 3 Taxa in der Topologie von Dayrat et al. (2001). Bezüglich der 18S rDNA bestehen Überschneidungen zu dem Datensatz für die Opisthobranchia. In dem Anaspidea-Datensatz sind zusätzlich noch 6 weitere Sequenzen enthalten, so dass er insgesamt 20 Sequenzen umfasst. Die Alinierung der ITS1 - 5.8S - ITS2-Bereiche beinhaltet 11 Sequenzen. Neben der Aufklärung der Phylogenie, soll die Eignung der beiden unterschiedlich schnell evolvierenden DNA-Bereiche für eine Untersuchung auf dieser taxonomischen Ebene geprüft werden. 2. Umbraculum Innerhalb der Tylodinoidea existieren 2 Familien mit insgesamt 3 Gattungen. Umbraculum Schuhmacher, 1817 ist die einzige Gattung der Umbraculidae und repräsentiert einige der sonderbarsten Vertreter der Opisthobranchia. Der Körper von Umbraculum (Abb. 2I) ist groß und erhaben, nahezu halbkugelförmig und wird von unzähligen Tuberkeln bedeckt. Er wird gekrönt durch eine zentrale, äußere Schale. Diese ist schirmförmig bis völlig flach und viel zu klein um den ganzen Körper zu bedecken, dazu gewöhnlich von einem dichten pflanzlichen und tierischen Bewuchs überzogen. Unter den Rhinophoren wird der Körper von einer Spalte durchzogen, in der die Mundöffnung liegt und sich der permanent hervorstehende Penis befindet. Die Umbraculum-Arten ernähren sich hauptsächlich von Schwämmen, dementsprechend ist ihre Radula breit und mit vielen kleinen raspelnden Zähnen besetzt (Willan 1984). Die Gattung kommt in einer Vielzahl unterschiedlicher Habitate vor, von Höhlen und Spalten in exponierten Riffen bis hin zu geschützten Riffebenen. Die Gattung Umbraculum ist, mit Ausnahme der arktischen und antarktischen Regionen, weltweit verbreitet und die Artabgrenzung erfolgte im Wesentlichen auf der Basis geographischer Verbreitung. Umbraculum mediterraneum (Lamarck 1812) ist hauptsächlich aus dem Mittelmeer und dem Ostatlantik bekannt (Madeira, Azoren, Kapverdische Inseln, St Helena), Umbraculum umbraculum (Roeding 1798) dagegen hauptsächlich aus dem indopazifischen Raum. Umbraculum sinicum

20
(Gmelin 1793) scheint kosmopolitisch zu sein, wohingegen Umbraculum plicatulum (v. Martens 1881) eine karibische und atlantische Verbreitung zeigt. Die Anzahl der Arten in dieser Gattung wird, ebenso wie ihre Artmerkmale, kontrovers diskutiert. Burn (1959) schreibt von nur einer Art mit weltweiter Verbreitung, die er U. sinicum nennt. Damit übereinstimmend, abgesehen vom gewählten Artnamen, ist die Aussage von Willan (1998): “The majority of opisthobranch systematists maintain that there is only a single species in the family, Umbraculum umbraculum, with a cosmopolitan distribution throughout tropical and warm temperate Indo-pacific seas…”. (Willan 1998). Thompson (1970) unterscheidet dagegen eine atlantisch und mediterran verbreitete Art (U. mediterranea) von einer indopazifischen Art (U. sinicum) und Marcus (1985) erwähnt eine dritte Art (U. plicatulum) aus der Karibik. Guangyu (1981) beschreibt eine neue Art (U. pulchrum sp. nov.) aus China und unterscheidet daneben 5 weitere (U. umbraculum, U. mediterraneum, U. pictum, U. ovalis, U. plicatulum). Während einer Expedition im Jahr 1998 mit dem deutschen Forschungsschiff „Meteor“ im Ostatlantik, wurden einige Exemplare der Gattung Umbraculum auf dem Unterwasserplateau “Große Meteorbank”, westlich der Kanaren, gesammelt. Dieses Material konnte bisher keiner der beiden Arten U. mediterraneum und U. umbraculum eindeutig zugeordnet werden und wurde im Rahmen der vorliegenden Arbeit zum ersten Mal gründlich morphologisch untersucht. Dabei wurden ebenfalls Individuen anderer Lokalitäten, bestimmt als U. umbraculum (Azoren, Australien) und U. mediterraneum (Griechenland), in die Analysen mit einbezogen. Zusätzlich wurden molekulare Untersuchungen der Individuen von der „Großen Meteorbank“, den Azoren und Australien mit Hilfe verschiedener Gene (18S rDNA, 16S rDNA, COI) durchgeführt. Die Fragestellung besteht darin, ob U. mediterraneum (Lamarck, 1812) und U. umbraculum (Lightfoot, 1786) zwei unterscheidbare Arten repräsentieren oder ob es sich um eine einzige, weltweit verbreitete Art handelt? Dabei sollen Möglichkeiten und Probleme der molekularen Artabgrenzung untersucht werden. 3. Phylogenie der Gattung Phyllodesmium Die Gattung Phyllodesmium Ehrenberg 1831 gehört zur Familie der Facelinidae (Nudibranchia, Aeolidoidea) und umfasst 16 beschriebene Arten, die bisher nur aus dem indopazifischen Raum bekannt sind (Rudman 1981, 1991, Baba 1949, 1991, Avila et al. 1998, Ortiz & Gosliner 2003, Burghardt & Wägele 2004). Für weitere 11 unbeschriebene Phyllodesmium-Arten sind Informationen im Internet verfügbar (www.seaslugforum.net).

21
Vertreter der Gattung Phyllodesmium (Abb. 2K) können eine Körperlänge von bis zu 14 cm erreichen (Rudman 1991) und weisen, wie die meisten Aeolidier, einen langgestreckten Körper mit zahlreichen Cerata auf. Diese sind leicht bis stark abgeflacht und besitzen, im Gegensatz zu vielen anderen Fadenschnecken, keine Cnidosäcke zum Speichern von Nesselzellen. Die Gattung Phyllodesmium hat dagegen eine alternative Verteidigungsstrategie entwickelt, die Autotomie von einzelnen Cerata. Diese produzieren ein klebriges Sekret und bewegen sich für einen relativ langen Zeitraum autonom umher. Dadurch verursachen sie offenbar genügend Ablenkung für potentielle Fressfeinde, um der Schnecke die Flucht zu ermöglichen. Die Artabgrenzung innerhalb der Gattung erfolgte hauptsächlich aufgrund unterschiedlicher Färbung, der Form der Rückenanhänge (Cerata), der Verzweigung der Mitteldarmdrüse und Unterschieden in der Radulabezahnung. Aufgrund ihrer Ernährung von Octokorallen (hauptsächlich Alcyonaria), sind die Phyllodesmium-Arten einzigartig unter den Aeolidoidea, die sich ansonsten ausschließlich von Hydroiden und Seeanemonen ernähren. Eine weitere Besonderheit der Gattung ist die Symbiose mit Zooxanthellen, einzelligen Algen der Gattung Symbiodinium (Dinophyceae), die allerdings mehrmals unabhängig innerhalb der Aeolidoidea entstanden ist. Die Zooxanthellen werden mit der Nahrung aufgenommen und in Zellen der Mitteldarmdrüse in den Cerata gespeichert, in denen sie photosynthetisch aktiv bleiben. Die Arten von Phyllodesmium zeigen eine große Bandbreite im Grad der Anpassung an eine symbiotische Beziehung mit den Zooxanthellen. Bei Formen ohne Zooxanthellen sind die Ausläufer der Mitteldarmdrüse in den Cerata unverzweigt, bei allen Arten mit Symbiose, sind sie zur Oberflächenvergrößerung in unterschiedlichem Maße verzweigt, bis hin zu feinen Verästelungen in Körperwand (z.T. sogar in den Rhinophoren und Oraltentakeln). In frühen Stadien der Symbiose, werden die Algen in normalen Zellen der Mitteldarmdrüse gespeichert, bei stärkerer Anpassung in modifizierten „Containerzellen“, die größenreduziert sind und keine sekretorische Funktion mehr aufweisen. In Verbindung mit steigender Effizienz der Symbiose werden die Cerata in zunehmendem Maße abgeflacht, um eine bessere Lichtexposition der Algen zu erzielen. Weiterhin ist sowohl eine Größenzunahme der Cerata als auch der allgemeinen Körpergröße zu beobachten (Rudman 1981, 1991). Die von Rudman (1991) vorgestellte Phylogenie von Phyllodesmium basiert auf dem oben beschriebenen Grad der Anpassung an eine Symbiose mit den Zooxanthellen, dabei werden Formen ohne Zooxanthellen als ursprünglich angesehen. Davon ausgehend haben sich die Mitglieder der Gattung vermutlich in zwei Linien entwickelt. In der einen scheinen sich die Schnecken permanent in ihrer Nahrungskoralle aufzuhalten und die Speicherung von Zooxanthellen ist wohl nur kurzzeitig möglich. Aus der Abhängigkeit von der Nahrung und der kurzen Speicherungzeit schließt Rudman (1991), dass es sich um ein frühes Stadium einer

22
Symbiose handelt. Die Arten der anderen evolutiven Linie weisen dagegen eine gut entwickelte, „echte“ Symbiose mit den Zooxanthellen auf. Sie zeigen keine obligate Ortsgebundenheit und obwohl sie in ihrer Futterkolonie sehr gut getarnt sind, werden sie häufig entfernt davon beobachtet. In der vorliegenden Arbeit soll die Phylogenie der Gattung mit Hilfe molekularer Methoden untersucht werden, um das von Rudman (1981, 1991) postulierte Evolutionsgeschehen zu stützen oder zu widerlegen. Da sich die 18S rDNA innerhalb der Nudibranchia als sehr variabel erwiesen hat (Wollscheid & Wägele 1999, Wägele et al. 2003), scheint sie geeignet, um auf diesem niederen taxonomischen Niveau gute Resultate zu erzielen. Der Datensatz umfasst neben 6 Sequenzen von unterschiedlichen Phyllodesmium-Arten noch 6 Sequenzen anderer Vertreter der Facelinidae und 13 Sequenzen aus anderen Familien der Aeolidoidea.

- 23 -
2 Material und Methoden 2.1 Untersuchte Taxa und Herkunft des Tiermaterials In der folgenden Tabelle ist die Herkunft aller untersuchten Taxa angegeben. Die jeweils sequenzierten Gene sind durch die Angabe der accession number bzw. durch den Eintrag „+“ in der betreffenden Spalte gekennzeichnet. Nicht bearbeitete Gene sind mit „o“ markiert. Das Tiermaterial ist entweder an der Zoologischen Staatssammlung München (ZSM) oder am Lehrstuhl für Spezielle Zoologie der Ruhr-Universität Bochum hinterlegt.
Taxon Familie Fundort komplette 18S rDNA
partielle 28S rDNA (D1-D3)
ITS 1+2, 5.8S rDNA
partielle 16S rDNA
partielle COI
NUDIBRANCHIA Bathydoris clavigera Thiele, 1912
Bathydorididae Antarktis AY165754
AY427444 o AF249222 o
Archidoris pseudoargus (Rapp, 1827)
Archidorididae Genbank o o o AF430347 o
Chromodoris krohni (Verany, 1846)
Chromodorididae Spanien, Atlantik AJ224774
AY427445 o AF249239 o
Hypselodoris villafranca (Risso, 1818)
Chromodorididae Spanien, Atlantik AJ224780
AY427446 o AF249237 o
Discodoris concinna (Alder & Hancock, 1864)
Discodorididae Genbank o o o AF249228 o
Dendrodoris fumata (Rüppell & Leuckart, 1830)
Dendrodorididae Australien, Großes Barriere Riff, Pazifik
AF249216
AY427447 o o o
Dendrodoris nigra (Stimpson, 1855)
Dendrodorididae Genbank o o o AF249242 o
Goniodoris nodosa (Montagu, 1808)
Goniodorididae Genbank o o o AF249226 o
Cuthona nana (Alder & Hancock, 1842)
Tergipedidae Helgoland, Nordsee AY165760
AY427448 o o o
Cuthona sibogae (Bergh, 1905)
Tergipedidae Australien, QLD, Lizard Isl., Pazifik
AY165761 o o o o
Cuthona caerulea (Montagu, 1804)
Tergipedidae Kattegat AF249199 o o o o
Embletonia sp.
Tergipedidae Frankreich, Mittelmeer + o o o o
Tergipes tergipes (Forskål, 1775)
Tergipedidae Kattegat AF249197 o o o o
Eubranchus exiguus (Alder & Hancock, 1848)
Eubranchidae Helgoland, Nordsee AJ224787 o o AF249246 o
Eubranchus sp.
Eubranchidae Spanien, Atlantik AJ224786 o o o o
Flabellina babai (Schmekel, 1972)
Flabellinidae Spanien, Mittelmeer AY165768
AY427449 o o o
Flabellina pedata (Montagu, 1815)
Flabellinidae Genbank o o o AF249247 o
Flabellina affinis (Gmelin, 1791)
Flabellinidae Spanien, Mittelmeer AY165767 o o o o
Flabellina verrucosa (M. Sars, 1829)
Flabellinidae USA, Nordatlantik AF249198 o o o o
Flabellina pedata (Montagu, 1815)
Flabellinidae Helgoland, Nordsee AJ224788 o o o o

- 24 -
Flabellina sp.
Flabellinidae Australien, QLD, Townsville, Paz.
AY165769 o o o o
Notaeolidia depressa Eliot, 1905
Notaeolidiidae Antarktis AY165770 o o o o
Phyllodesmium briareum (Bergh, 1896)
Facelinidae Australien, QLD, Lizard Isl., Pazifik
+ o o o o
Phyllodesmium colemani Rudman, 1991
Facelinidae Indonesien, Sulawesi, Lembeh Strait, Pazifik
+ o o o o
Phyllodesmium crypticum Rudman, 1981
Facelinidae Australien, NSW, Cockle Bay, Pazifik
+ o o o o
Phyllodesmium jakobsenae Burghardt & Wägele, 2004
Facelinidae Indonesien, Sulawesi, Bunaken Isl., Pazifik
+ o o o o
Phyllodesmium longicirrum (Bergh, 1905)
Facelinidae Australien, QLD, Lizard Isl., Pazifik
+ o o o o
Phyllodesmium cf. magnum
Facelinidae Indonesien, Sulawesi, Lembeh Strait, Pazifik
+ o o o o
Godiva banyulensis (Garcia & Garcia, 1985)
Facelinidae Spanien, Mittelmeer + o o o o
Baeolidia nodosa (Haefelfinger & Stamm, 1959)
Facelinidae Elba, Mittelmeer + o o o o
Facelina bostoniensis Thompson, 1843
Facelinidae Kieler Föhrde, Ostsee + o o o o
Facelina auriculata O. F. Müller, 1776
Facelinidae unbekannt + o o o o
Phidiana lynceus Bergh, 1896
Facelinidae Niederl. Antillen, Curaçao, Karibik
+ o o o o
Cratena peregrina (Gmelin, 1791)
Facelinidae + o o o o
Dendronotus dalli Bergh, 1879
Dendronotidae USA, Nordatlantik AY165757
AY427450 o AF249252 o
PLEUROBRANCHOIDEA Euselenops luniceps (Cuvier, 1817)
Pleurobranchae- idae
Australien, Großes Barriere Riff, Pazifik
AF249218
AY427451 o o o
Tomthompsonia antarctica (Thiele, 1912)
Pleurobranchidae Antarktis AY427492
AY427452 o + o
Bathyberthella antarctica Willan & Bertsch, 1987
Pleurobranchidae Antarktis, Weddelmeer AF249219
AY427453 o o o
Berthellina citrina (Rüppell & Leuckart, 1828)
Pleurobranchidae Italien, Elba, Mittelmeer
AY427493
AY427454 o o o
Pleurobranchus peroni Cuvier, 1804
Pleurobranchidae Australien, NSW, Wollongong, Pazifik
AY427494
AY427455 o + o
Berthella sideralis (Lovén,1846)
Pleurobranchidae Genbank o o o AJ225181 o
Berthella stellata (Risso, 1826)
Pleurobranchidae Italien, Elba, Mittelmeer
AY427495
AY427456 o o o
Berthella platei (Bergh, 1898)
Pleurobranchidae Scotia-Bogen, Atlantik + o o + o
TYLODINOIDEA Umbraculum mediterraneum (Lamarck, 1829)
Umbraculidae Atlantik, Große Meteor Bank
AY165753 AY427457 0 + +
Umbraculum cf. umbraculum
Umbraculidae Azoren, Atlantik + o o + +
- 25 -
Umbraculum umbraculum (Lightfoot, 1786)
Umbraculidae Australien, NSW, Shellharbour
+ o o + +
Tylodina perversa (Gmelin, 1791)
Tylodinidae Spanien, Mittelmeer AY427496
AY427458 o o AF249809
SACOGLOSSA Plakobranchus ocellatus van Hasselt, 1824
Plakobranchidae Australien, QLD, Magnetic Island, Pazifik
AY427497
AY427459 o o o
Bosellia mimetica Trinchese, 1891
Boselliidae Italien, Elba, Mittelmeer
AY427498
AY427460 o o o
Thuridilla bayeri Marcus, 1965
Elysiidae Australien, Großes Barriere Riff, Pazifik
AF249220
AY427461 o o o
Thuridilla ratna (Marcus, 1965)
Elysiidae Genbank o o o AF249256 o
Elysia viridis (Montagu, 1804)
Elysiidae Mittelmeer AY427499
AY427462 o AJ223398 o
Elysiella pusilla Bergh, 1872
Elysiidae Australien, QLD, Hydeaway Bay, Pazifik
+ o o o o
Cyerce nigricans (Pease, 1866)
Polybranchidae Australien, QLD, Eagle Island, Pazifik
AY427500
AY427463 o o o
Calliopaea bellula d’Orbigny, 1837
Limapontiidae Italien, Elba, Mittelmeer
AY427501
AY427464 o o o
Limapontia capitata (O.F. Müller, 1773)
Limapontiidae Nordsee AJ224920
AY427465 o o o
Placida cremoniana (Alder & Hancock, 1843)
Stiligeridae Indonesien, Sulawesi, Bitung, Pazifik
+ o o o o
Placida dendritica (Alder & Hancock, 1843)
Limapontiidae Genbank o o o AJ223399 o
Polybranchia orientalis (Kelaart, 1858)
Polybranchidae Antarktis, Bird Island + o o o o
ANASPIDEA Akera bullata Müller, 1776
Akeridae Dänemark, Kattegat AY427502
AY426746 + AF156127 o
Aplysia californica Cooper, 1863
Aplysiidae Genbank o
AY026366 o o o
Aplysia dactylomela Rang, 1828
Aplysiidae Australien, QLD, Hydeaway Bay, Pazifik
+ o o o o
Aplysia gigantea Sowerby, 1869
Aplysiidae Australien, Rottnest Island, Ind. Ozean
+ o + o o
Aplysia juliana Quoy & Gaimard, 1832
Aplysiidae Australien, NSW, Wollongong, Paz.
+ o + o o
Aplysia oculifera Adams & Reeve, 1850
Aplysiidae Australien, Rottnest Island, Ind. Ozean
+ o + AF192302 o
Aplysia parvula Guilding in Mörch, 1863
Aplysiidae Madeira, Atlantik + o o o o
Aplysia sydneyensis Sowerby, 1869
Aplysiidae Australien, NSW, Wollongong, Paz.
+ o o o o
Syphonota geographica (Adams & Reeve, 1850)
Aplysiidae Australien, Magnetic Island, Pazifik
+ o o o o
Bursatella leachii de Blainville, 1817
Aplysiidae Australien, QLD, Dingo Beach, Pazifik
+ o + AF156130 o
Dolabella auricularia (Lightfoot, 1786)
Aplysiidae Japan, Okinawa, Pazifik
AY427503
AY427467 + AF156132 o
Dolabrifera brazieri Sowerby 1870
Aplysiidae Australien, NSW, Hastings Point, Pazifik
+ o + o o
Dolabrifera dolabrifera (Cuvier, 1817)
Aplysiidae Japan, Okinawa, Pazifik
+ o + AF156133 o

- 26 -
Notarchus indicus Schweigger, 1820
Aplysiidae Genbank o o o AF156135 o
Petalifera petalifera (Rang, 1828)
Aplysiidae Australien, Magnetic Island, Pazifik
+ o + o o
Petalifera ramosa Baba, 1959
Aplysiidae Genbank o o o AF156152 o
Phyllaplysia taylori Dall, 1900
Aplysiidae Genbank o o o AF156139 o
Stylocheilus longicauda (Quoy & Gaimard, 1825)
Aplysiidae Australien, NSW, Wollongong, Pazifik
+ o o AF156140 o
CEPHALASPIDEA s.str. Haminoea hydatis (Linné, 1758)
Haminoeidae Italien, Elba, Mittelmeer
AY427504
AY427468 o o o
Haminoea cymbalum (Quoy and Gaimard, 1835)
Haminoeidae Genbank o o o AF249258 o
Phanerophthalmus smaragdinus (Ruppell & Leuckart, 1831)
Haminoeidae Australien, QLD, Lizard Island, Pazifik
AY427505
AY427469 o o o
Smaragdinella sp.
Haminoeidae Genbank AJ224789 o o AF249257 o
Bullacta exarata (Philippi, 1848)
Haminoeidae Genbank AF188675 o o o o
Atys semistriatus Pease, 1860
Haminoeidae Australien, QLD, Dingo Beach, Pazifik
AY427506
AY427470 + o o
Bulla gouldiana Pilsbry, 1893
Bullidae Genbank o o o AF156125 o
Odontoglaja guamensis Rudman, 1978
Aglajidae Australien, QLD, Orpheus Island, Pazifik
AY427507
AY427471 o o o
Navanax inermis (Cooper, 1863)
Aglajidae Mexico, La Paz, Golf von Kalifornien
AY427508
AY427472 o o o
Chelidonura inornata Baba, 1949
Aglajidae Australien, QLD, Lizard Island, Pazifik
AY165752
AY427473 + o o
Philinopsis pilsbryi (Eliot, 1900)
Aglajidae Australien, QLD, Magnetic Island, Pazifik
AY427509
AY427474 o o o
Philina aperta (Linné,1767)
Philinidae Genbank o o o AJ223402 o
Philinoglossa praelongata Salvini-Plawen, 1973
Philinoglossidae Kroatien, Rovinj, Mittelmeer
AY427510
AY427475 o o o
cf. Retusa sp.
Retusidae Italien, Elba, Mittelmeer
AY427511
AY427476 o o o
Bulla cf. striata Bruguière, 1792
Bullidae Italien, Elba, Mittelmeer
AY427512
AY427477 o o o
Sagaminopteron psychedelicum Carlson & Hoff, 1974
Gastropteridae Australien, QLD, Magnetic Island, Pazifik
AY427513
AY427478 o o o
Runcina adriatica Thompson, 1980
Runcinidae Italien, Elba, Mittelmeer
AY427514
AY427479 o o o
Cylichna gelida (Smith, 1907)
Cylichnidae o. Scaphandridae
Scotia-Bogen, Atlantik + o o o o
Scaphander lignarius (Linné, 1758)
Cylichnidae o. Scaphandridae
Daten von H. Wägele + o o o o
Scaphander punctostriatuss (Mighels & Adams, 1842)
Cylichnidae o. Scaphandridae
Genbank o o o AJ223403 o
Colpodaspis Diaphanidae Indonesien, Sulawesi, + o o o o
- 27 -
thomsoni Brown, 1979
Nain Island
Diaphana cf. inflata (Strebel, 1908)
Diaphanidae Antarktis + o o o o
Diaphana minuta Brown, 1827
Diaphanidae Genbank 0 0 0 AJ223404 o
Toledonia globosa ?
Diaphanidae Scotia-Bogen, Atlantik + o o o o
Newnesia antarctica Smith, 1902
Diaphanidae Antarktis + o o o o
ACTEONOIDEA Hydatina physis (Linné, 1758)
Hydatinidae Australien, NSW, Hastings Point, Pazifik
AY427515
AY427480 o + o
Bullina lineata (Gray, 1825)
Hydatinidae Australien, NSW, Shellharbour
+ o o o o
Acteon tornatilis (Linné, 1758)
Acteonidae Genbank o o o AF355186 o
Acteon sp.
Acteonidae Antarktis + o o o o
Pupa solidula (Linné, 1758)
Acteonidae Australien, QLD, Dingo Beach, Pazifik
AY427516
AY427481 o + o
ACOCHLIDIACEA Unela glandulifera (Kowalewsky, 1901)
Microhedylidae Kroatien, Rovinj, Mittelmeer
AY427517
AY427482 o + o
Parahedyle cryptophthalma (Westheide & Wawra, 1974)
Microhedylidae Italien, Elba, Mittelmeer
AY427518
AY427483 o + o
Pontohedyle milatchevitchi (Kowalewsky, 1901)
Microhedylidae Italien, Elba, Mittelmeer
AY427519
AY427484 o + o
Hedylopsis spiculifera (Kowalewsky, 1901)
Hedylopsidae Kroatien, Rovinj, Mittelmeer
AY427520
AY427485 o o o
THECOSOMATA Cavolinia sp.
Cavoliniidae Frankreich, Mittelmeer o o o + o
Clio pyramidata Linné, 1767
Cavoliniidae Spanien, Kanarische Inseln, Atlantik
+ o o o o
Creseis acicula (Rang, 1828)
Cavoliniidae Frankreich, Mittelmeer + o o o o
Diacra quadridentata (Lesueur in Blainville, 1821)
Cavoliniidae Spanien, Kanarische Inseln, Atlantik
+ o o o o
GYMNOSOMATA Clione limacina (Phipps, 1774)
Clionidae Genbank o o o AJ223406 o
Spongiobranchaea australis d'Orbigny, 1836
Pneumoderma- tidae
Scotia-Bogen, Atlantik + o o o o
PULMONATA Auriculinella bidentata (Montagu, 1808)
Ellobiidae Italien, Venedig, Mittelmeer
+ o o o o
Pythia scarabaeus (Linné, 1758)
Ellobiidae Indonesien, Sulawesi, Bunaken Isl.
+ o o + o
Cassidula nucleus (Gmelin, 1791)
Ellobiidae Indonesien, Sulawesi, Bunaken Isl.
+ o o o o
Ovatella firminii (Payraudeau, 1827)
Ellobiidae Italien, Venedig, Mittelmeer
+ o o o o
Melampus luteus Quoy & Gaimard, 1832
Ellobiidae Indonesien, Sulawesi, Bunaken Isl.
+ o o o o
Onchidella floridana (Dall, 1885)
Onchidiidae Bermudas, Flatts Village, Atlantik
AY427521
AY427486 o o o
Onchidium cf. Onchidiidae Australien, NSW, AY427522 AY427487 o o o

- 28 -
verruculatum (Cuvier, 1830)
Cockle Bay, Pazifik
Siphonaria alternata (Say, 1826)
Siphonariidae Bermudas, Flatts Village, Atlantik
AY427523
AY427488 o o o
Cepaea hortensis (OF Müller 1774)
Helicidae BRD, Grietherbusch, Rhein
AY427524
AY427489 o o o
Lymnaea stagnalis (Linnaeus, 1758)
Lymnaeidae BRD, Grietherbusch, Rhein
AY427525
AY427490 o o o
ALLOGASTROPODA Pyramidella sulcata tessellata (Adams A. 1854)
Pyramidellidae Indonesien, Sulawesi, Bunaken Isl.
+ o o o o
Odostomia sp. Pyramidellidae Frankreich, Banyuls sur Mer, Mittelmeer
AY427526
AY427491 o + o
„PROSOBRANCHIA“ Nodilittorina punctata (Gmelin, 1791)
Littorinidae Genbank Y11755 o o o o
Nodilittorina unifasciata
Littorinidae Genbank o AJ488662 o o o
Littoraria undulata (Gray, 1839)
Littorinidae Genbank Y11756 o o o o
Littorina striata (King, 1832)
Littorinidae Genbank Y11750 o o o o
Littorina saxatilis (Olivi, 1792)
Littorinidae Genbank Y11751 o o o o
Littorina littorea (Linné, 1758)
Littorinidae Genbank X91970 AJ488672 o o o
Littorina scutulata Gould, 1849
Littorinidae Genbank o o o AY163396 o
Littoraria undulata Gray, 1839
Littorinidae Genbank o AJ488676 o o o
Melaraphe neritoides (Linné,1758)
Littorinidae Genbank Y11754 AJ488649 o o o
Tectarius viviparus (Rosewater, 1982)
Littorinidae Genbank o o o U66352 o
Tectarius striatus
Littorinidae Genbank o AJ488685 o o o
Crepidula adunca Sowerby, 1825
Calyptraeidae Genbank X94277 o o o o
Balcis eburnea
Eulimidae Genbank AF120519 o o o o
Fasciolaria lignaria (Linné,1758)
Fasciolariidae Genbank X94275 o o o o
Pisania striata (Gmelin, 1791)
Buccinidae Genbank X94272 o o o o
Bursa rana (Linné, 1758)
Bursidae Genbank X94269 o o o o
Truncatella guerinii Villa & Villa 1841
Truncatellidae Genbank AF120518 o o o o
Viviparus georgianus (Lea, 1834)
Viviparidae Genbank AF120516 o o o o
Tabelle 1: Herkunft des untersuchten Tiermaterials und jeweils sequenzierter DNA-Bereich.

- 29 -
Abb. 5 Sekundärstruktur einer typischen eukaryotischen 18S rRNA (aus Ueda-Nishimura & Mikata 2000, verändert)
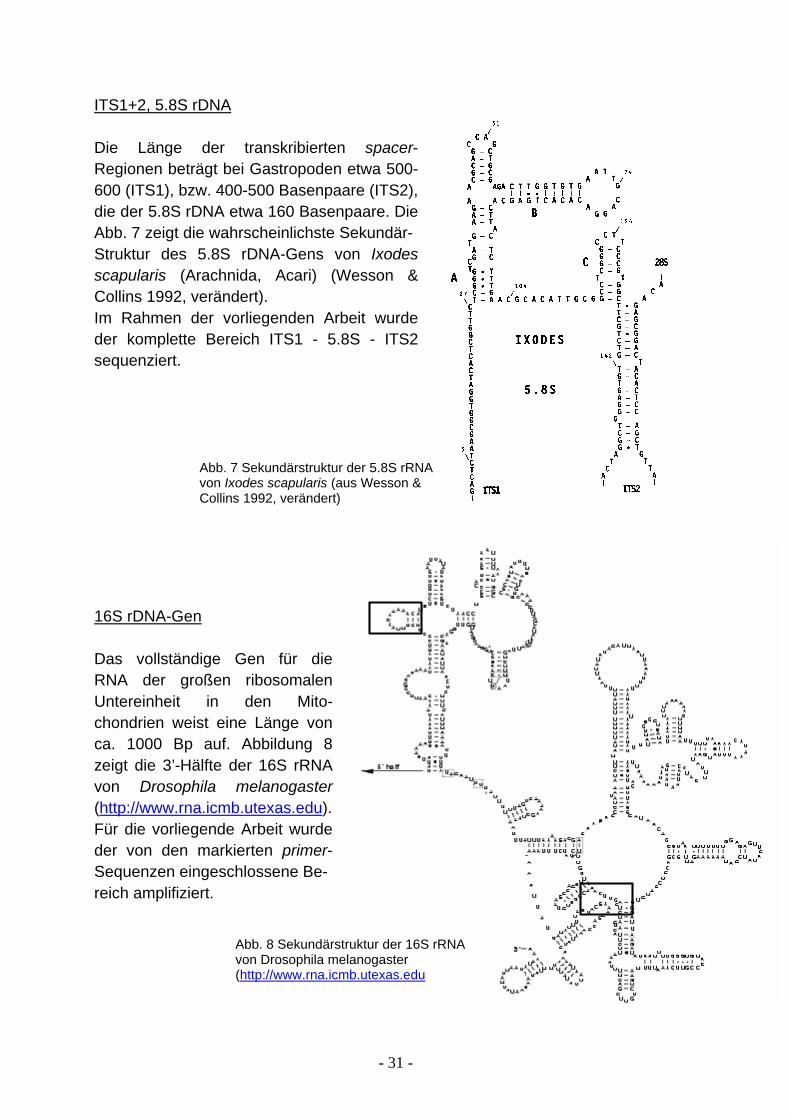
2.2 Untersuchte Gene Alle RNA-Moleküle bilden Sekundärstrukturen aus, dabei entstehen doppelsträngige, sog. stem- und einzelsträngige loop-Bereiche. Diese verschiedenen Genabschnitte weisen eine unterschiedliche Variabilität auf. In den einzelsträngigen Bereichen treten Modifikationen in der Nukleotidsequenz häufig in größerer Anzahl auf als in den doppelsträngigen, da eine Mutation im gepaarten Bereich die Bindung durch Wasserstoffbrücken unterbricht. Dieser Zusammenhang ist jedoch nicht universell gültig, es gibt durchaus stem-Regionen mit hoher Variabilität und loop-Bereiche die extrem konserviert sind. Ausschlaggebend für die Höhe der Substitutionsrate ist allein der Selektionsdruck, der wiederum von der funktionellen Bedeutung der Molekülregion abhängig ist. Für die Bezeichnung bestimmter Unterstrukturen gibt es allgemeingültige Regeln: Von 5’- in 3’-Richtung des RNA-Moleküls sind Unterstrukturen durchgehend nummeriert; daneben gibt es übergeordnete Bereiche, die sogenannten variable areas, die mit dem Buchstaben V gekennzeichnet werden. Die folgenden Gene wurden im Rahmen dieser Arbeit sequenziert: 18S rDNA-Gen Die Länge des kompletten 18S rDNA-Gens beträgt etwa 1900 Basenpaare. Abb. 5 zeigt die Sekundär- struktur einer typischen eu-karyotischen 18S rRNA, dabei sind relativ kon-servierte Regionen fett gedruckt und variable mit V1-V9 gekennzeichnet (aus Ueda-Nishimura & Mikata 2000, verändert). Für die vorliegende Arbeit wurde das 18S rDNA-Gen vollständig sequenziert. 28S rDNA-Gen Das komplette 28S rDNA-Gen umfasst etwa 4700 Basenpaare. In Abb. 6 ist die Sekundärstruktur der 28S rRNA von Saccharomyces cerevisiae (de Rijk et al. 1994, verändert) dargestellt. Die variablen Regionen sind besonders gekennzeichnet (V1-

- 30 -
V12). Für die vorliegende Arbeit wurden die Bereiche V1-V3 inklusive der dazwischen liegenden Regionen sequenziert.
Abb. 6 Sekundärstruktur der 28S rRNA von Saccharomyces cerevisiae (aus de Rijk et al. 1994, verändert)
- 31 -
ITS1+2, 5.8S rDNA Die Länge der transkribierten spacer-Regionen beträgt bei Gastropoden etwa 500-600 (ITS1), bzw. 400-500 Basenpaare (ITS2), die der 5.8S rDNA etwa 160 Basenpaare. Die Abb. 7 zeigt die wahrscheinlichste Sekundär- Struktur des 5.8S rDNA-Gens von Ixodes scapularis (Arachnida, Acari) (Wesson & Collins 1992, verändert). Im Rahmen der vorliegenden Arbeit wurde der komplette Bereich ITS1 - 5.8S - ITS2 sequenziert. 16S rDNA-Gen Das vollständige Gen für die RNA der großen ribosomalen Untereinheit in den Mito-chondrien weist eine Länge von ca. 1000 Bp auf. Abbildung 8 zeigt die 3’-Hälfte der 16S rRNA von Drosophila melanogaster (http://www.rna.icmb.utexas.edu). Für die vorliegende Arbeit wurde der von den markierten primer-Sequenzen eingeschlossene Be- reich amplifiziert.
Abb. 7 Sekundärstruktur der 5.8S rRNA von Ixodes scapularis (aus Wesson & Collins 1992, verändert)
Abb. 8 Sekundärstruktur der 16S rRNA von Drosophila melanogaster (http://www.rna.icmb.utexas.edu

- 32 -
Gen für die Untereinheit 1 der mitochondrialen Cytochromoxidase (COI) Das komplette Gen für die Untereinheit 1 der mitochondrialen Cytochromoxidase umfasst rund 1500 Basenpaare. Im Rahmen der vorliegenden Arbeit wurde der in der unten gezeigten Aminosäureabfolge grau unterlegte Bereich sequenziert. MRWLCSTNHKDIGTLYMVFGMWCGLVGTGLSLLIRFELGTAGALLGDDHFYNVIVTAHAFVMIFFMVMPLMIGGFGNWMVPLLIG
APDMSFPRMNNMSFWLLPPSLILLLSSSLMEGGAGTGWTVYPPLSGAMGHTGCSVDLAIFSLHLAGMSSLLGAVNFITTIFNMRA
PGMTMERVSLFVWSVLVTAFLLLLSLPVLAGAITMLLTDRNFNTSFFDPAGGGDPILYQHLFWFFGHPEVYILILPGFGMISHMLSN
YASKPAFGTLGMIYAMVSIGILGFIVWAHMFTVGMDVDTRAYFTAATMVIAIPTGIKVFSWLMTLYGTQGPLPASMWWVLGFIFTL
GGLTGIVLSNSSLDIVLHDTYYVVAHFHYVLSMGAVFAIFGGFVYWFPVMTGMMLHEGFAKAHFLVMFFAVNMTFFPQHFLGLAG
MPRRYSDYPDAYFKWNQISSYGSLFSVFAILMFIFMVWEALLSQRNALFSMAPSMSREWDGVLPPDFHSNTETSVSPF
2.3 Labormethoden 2.3.1 Fixierung des Tiermaterials Die Fixierung des bearbeiteten Tiermaterials erfolgte in 95%igem Ethanol. Nicht selbst gesammeltes Material lag zum Teil in 70%igem Ethanol vor, doch beide Konzentrationen erwiesen sich bei der weiteren Bearbeitung als gleichwertig. 2.3.2 DNA-Isolierung Die DNA der Tiere wurde mit dem Blood and Tissue-Kit der Firma Qiagen anhand des beiliegenden Protokolls isoliert. Das Ergebnis der DNA-Extraktion wurde anschließend mit Hilfe einer Agarose-Gelelektrophorese überprüft. Elektrophorese-Verfahren beruhen auf der Bewegung geladener Moleküle in einem elektrischen Feld. Nukleinsäuren besitzen eine Vielzahl geladener oder ionisierbarer Gruppen und eignen sich deswegen hervorragend für eine Auftrennung durch Elektrophorese. Das Standard-Verfahren ist die Elektrophorese in Agarose-Gelen. Die Geschwindigkeit mit der die DNA-Moleküle durch das Gel wandern, hängt in erster Linie von der Größe der DNA-Fragmente ab: lineare, doppelsträngige DNA-Moleküle bewegen sich durch die Agarose-Matrix mit einer Geschwindigkeit, die umgekehrt proportional zum Logarithmus ihrer Größe ist, d.h. kleine Moleküle wandern schneller als große. Weitere Faktoren für die Wanderungsgeschwindigkeit im Gel sind die Stromstärke, die Pufferbedingungen und vor allem die Agarose-Konzentration. Für eine Fragmentgröße von ca. 1800 Bp wie sie die 18S rDNA aufweist, hat sich z.B. ein 1%iges Agarose-Gel bewährt. Der Nachweis der DNA-Fragmente im Gel erfolgt durch Ethidiumbromid. Das flache, mehrgliedrige Ringsystem des Ethidiumbromid

- 33 -
interkaliert zwischen die Basenpaare der DNA und kann durch seine Fluoreszenz bei Bestrahlung mit UV-Licht sichtbar gemacht werden. 2.3.3 Die Polymerase-Kettenreaktion (PCR) Die Methode der Polymerase-Kettenreaktion (polymerase chain reaction, PCR, Saiki et al. 1988) wurde Mitte der 80er Jahre von Kary Mullis und Fred Faloona entwickelt und stellt einen Meilenstein in der Entstehung der molekularen Systematik dar. Ihre Bedeutung läßt sich durch ihre beiden wesentlichen Eigenschaften ausdrücken: 1. Man kann die DNA-Polymerase gezielt zur Synthese eines speziellen DNA-
Bereichs einsetzen. 2. Es findet eine Vervielfältigung des gewünschten DNA-Bereichs statt. Ablauf der PCR: Da die DNA-Polymerase zur Synthese von DNA auf eine einzelsträngige Matrize angewiesen ist, müssen die beiden Stränge der Ausgangs-DNA durch Hitzedenaturierung getrennt werden. Zusätzlich benötigt die Polymerase ein kurzes Stück doppelsträngiger DNA, um mit der Synthese zu beginnen. Man fügt dem Reaktionsansatz deshalb zwei Oligonukleotide (primer) zu, die so gewählt sind, daß sie antiparallel zueinander an die beiden Stränge der DNA hybridisieren und den zu vervielfältigenden Bereich zwischen sich einschließen. Zusätzlich werden dem Reaktionsansatz alle 4 Desoxynukleotide zugefügt. Nach Trennung der komplementären Stränge bei 94°C wird die Temperatur auf 30-65°C gesenkt, damit sich die primer an die nun einzelsträngigen DNA-Moleküle anlagern können. Dabei spielt die sogenannte Schmelztemperatur Tm eines Primers eine entscheidende Rolle. Sie kann annäherungsweise aus der Anzahl der GC- und AT-Paare berechnet werden und repräsentiert diejenige Temperatur, bei der die Hälfte der primer-Moleküle eine Bindung mit der DNA eingegangen sind. Bei den meisten PCR-Experimenten wird eine annealing-Temperatur gewählt, die ca. 5°C unter der berechneten Tm liegt. Bei dieser Temperatur wird ein Primer nur an seine vollständig komplementäre Sequenz binden, da bei Basenfehlpaarungen keine effiziente Doppelstrangbildung möglich ist. Bei steigender Temperatur können zunehmend weniger primer gebunden werden und die Ausbeute an PCR-Produkten wird geringer, während bei abnehmender Temperatur zwar die meisten Bindungsstellen besetzt sind, es aber auch vermehrt zu unspezifischen Hybridisierungen mit ähnlichen Sequenzbereichen kommen kann. Nach der annealing-Phase findet die DNA-Synthese bei 72°C statt. Das ist das Temperaturoptimum der hitzestabilen Taq-Polymerase aus dem in heißen Quellen lebenden Bakterium Thermus aquaticus. Zur Trennung der neusynthetisierten DNA-Stränge wird die Temperatur wieder auf 94°C erhöht.

- 34 -
Der ganze Zyklus – Erhitzen zur Trennung der Stränge, Binden der primer und Synthese mit Hilfe der DNA-Polymerase – wiederholt sich 25-50mal. Nach 30 Zyklen beispielsweise, beträgt die Anzahl der doppelsträngigen Zielmoleküle bereits 268 435 456 (bei 1 Ausgangsmolekül). Nach dem letzten Zyklus wird ein mehrminütiger Polymerisationsschritt angeschlossen, um es der Taq-Polymerase zu ermöglichen, alle synthetisierten DNA-Stränge zu vervollständigen. Routinemäßig wurde bei jeder PCR ein sogenannter hot-start durchgeführt. Da die Taq-Polymerase bereits bei Raumtemperatur aktiv ist, kann es zur Verlängerung von nicht spezifisch angelagerten primern kommen, oder von primern, die miteinander in Wechselwirkung treten. Die Bildung solch störender Nebenprodukte, die sogar das Fehlschlagen der PCR zur Folge haben kann, wird durch den hot start wirkungsvoll reduziert. Das Prinzip einer hot start-PCR besteht darin, dass die enzymatische Aktivität der Taq-Polymerase erst dann zur Verfügung gestellt wird, wenn der Reaktionsansatz eine erhöhte Temperatur erreicht hat. In der praktischen Anwendung wird die Polymerase erst nach der Aufheizung der Proben im thermocycler dazugegeben. Zur Amplifikation der DNA wurden die Taq-Polymerasen der Firmen Qiagen und Eppendorf verwendet. Die PCR-Reaktionen wurden in Anlehnung an die Herstellerangaben und unter Berücksichtigung eigener Erfahrungen angesetzt. PCR-Reaktionsansatz Qiagen-Taq PCR-Reaktionsansatz Eppendorf-Taq 5 µl 10x PCR-Puffer 5 µl 10x PCR-Puffer 10 µl Q-Solution (PCR-Additiv) 5 µl dNTP-Mix (2 mM je dNTP) 5 µl dNTP-Mix (2 mM je dNTP) 0,5 µl primer 1 (50 pmol/µl) 0,5 µl primer 1 (50pmol/µl) 0,5 µl primer 2 (50 pmol/µl) 0,5 µl primer 2 (50pmol/µl) 0,3 µl Taq-Polymerase 0,3 µl Taq-Polymerase 0,5-3,0 µl genomische DNA 0,5-3,0 µl genomische DNA x µl H2O (dest.) x µl H2O (dest.) ______________________________ ____________________________ 50 µl 50 µl Alternativ wurde die PCR aus Kostengründen mit halben Ansätzen durchgeführt. Verwendete PCR-primer 18S rRNA Gen: 18A1 5‘-CCTACTCTGGTTGATCCTGCCAGT-3‘ 1800 5‘-TAATGATCCTTCCGCAGGTT-3‘

- 35 -
28S rRNA-Gen (partiell): 28SC1 5’-ACC CGC TGA ATT TAA GCA T-3’ 28SD3 5’-GAC GAT CGA TTT GCA CGT CA-3’ 16S rRNA-Gen (partiell): 16a-3 5’-CGC CTG TTT ATC AAA AAC AT-3’ 16b-3 5’-CCG GTC TGA ACT CAG ATC ATG T-3’ ITS1+2 und 5.8S rRNA-Gen 1800F 5’-GTA AAA GTC GTA ACA AGG TTT CCG TA-3’ 28SC1R 5’-ATA TGC TTA AAT TCA GCG GGT-3’ COI-Gen (partiell) HCO 5’-TAA ACT TCA GGG TGA CCA AAA AAT CA-3’ LCO 5’-GGT CAA CAA ATC ATA AAG ATA TTG G-3’ PCR-Reaktionsprofil für 18S, 28S, ITS PCR-Reaktionsprofil für 16S, COI Initiale Denaturierung: Initiale Denaturierung: 95°C 4 min 95°C 4 min Denaturierung, primer-Hybridisierung Denaturierung, primer-Hybridisierung und Polymerisation: und Polymerisation: 94°C 30 sec 94°C 45 sec 42-54°C 50 sec 48-56°C 45 sec 72°C 2,30 min 72°C 1,30 min Abschließende Polymerisation: Abschließende Polymerisation: 72°C 10 min 72°C 7 min 4°C 8 4°C 8 Die Amplifikation aller DNA-Sequenzen erfolgte im thermischen Zykler „Progene“ der Firma Techne. Zur Kontrolle der entstandenen PCR-Produkte wurden 3µl je Probe mit derselben Menge Bromphenolblau vermischt und auf ein 1%iges Agarose-Gel aufgetragen. Durch Zugabe eines Längen-Markers konnte die Größe der PCR-Produkte abgeschätzt und dadurch der Erfolg der PCR bestimmt werden. Bei jeder PCR wurde zusätzlich ein PCR-Ansatz ohne DNA hergestellt, um etwaige Verunreinigungen der Komponenten aufzudecken.
38 x 35 x

- 36 -
2.3.4 Aufreinigung von PCR-Produkten Eine Aufreinigung der PCR-Produkte ist erforderlich, um unverbrauchte Reagenzien und Salze aus den Proben zu entfernen und dadurch eine direkte Sequenzierung sowie die Klonierung der Amplifikate zu ermöglichen. Zur Aufreinigung der PCR-Produkte wurde das QIAquick PCR Purification Kit der Firma QIAGEN verwendet. Die beiliegenden Säulen sind bei einer gegebenen Salzkonzentration durchlässig für DNA-Fragmente mit weniger als 100 Bp. Auf diese Weise können überschüssige primer und primer-Dimere aus dem PCR-Produkt entfernt werden, ebenso wie überschüssige Nukleotide, Polymerasemoleküle und Puffer-Salze. Die Aufreinigung wurde anhand des beiliegenden Protokolls durchgeführt. 2.3.5 Klonierung von PCR-Fragmenten Unter einem Zellklon versteht man Zellen, die durch Zellteilung aus einer Stammzelle hervorgegangen sind und deshalb untereinander genetisch identisch sind. Analog bezeichnet „klonierte“ DNA die durch Replikation von einer Ausgangs-DNA hergestellten identischen Kopien. Die übliche Zielsetzung bei der Anwendung von Klonierungstechniken besteht in der Vermehrung der zu untersuchenden DNA für nachfolgende Analysen oder in der Untersuchung unbekannter Gene mit Hilfe des Einsatzes universeller primer. Das Klonieren von DNA ist auch dann sinnvoll, wenn bei der Amplifikation eines Gens während der PCR mehrere Produkte entstehen, die nicht anderweitig, z.B. aufgrund unterschiedlicher Länge, isoliert werden können. Unabhängig von der Fragestellung erfolgt stets eine Replikation der zu untersuchenden DNA in Bakterienzellen, z.B. E. coli. Da freie DNA sich nicht autonom in Bakterienzellen vervielfältigt, bedient man sich biologischer Träger oder Vektoren. Ein geeigneter DNA-Vektor muß drei Bedingungen erfüllen: Er muß die Fremd-DNA aufnehmen können, er muß als eigenständiges Replikon in der Wirtszelle repliziert werden können und er sollte mit hoher Effizienz in die Wirtszelle eingebracht werden können. Als Vektoren eignen sich daher ringförmige DNA-Moleküle, wie sie in der Natur als Plasmide oder als Virus-DNA vorkommen. Zum Einbringen der Passagier-DNA z.B. in Plasmide wird mindestens eine, im Idealfall singuläre, Schnittstelle für Restriktionsendonukleasen benötigt. An dieser Stelle wird das Plasmid geöffnet und das zu klonierende PCR-Produkt eingefügt. Die verwendeten Plasmid-Vektoren pCR 2.1-TOPO (Invitrogen) und pGem -T (Promega) werden schon linearisiert geliefert und tragen einen Thymidin-Überhang am 3‘-Ende. Da die Taq-Polymerase dem 3‘-Ende von PCR-Produkten bei der PCR ein Adenosin anhängt, erweist sich die Ligation als sehr effizient. Durch eine kovalent am Vektor gebundene Topoisomerase werden PCR-Produkt und Plasmid schließlich verknüpft.

- 37 -
In der Praxis wird die Ligation durch Mischung von PCR-Produkt und Vektor und anschließender Inkubation bei Raumtemperatur herbeigeführt. Die Transformation des Ligationsproduktes in die Bakterienzelle erfolgt durch die „Hitzeschock-Methode“. Dabei wird der Ansatz mit dem Ligationsprodukt und den bereits im kompetenten Zustand gelieferten E.coli-Zellen einer Temperatur von 42°C ausgesetzt, wodurch es zur Plasmidaufnahme kommt. Die Selektion rekombinanter Plasmide erfolgt durch eine Farbreaktion. Der verwendete Vektor trägt einen Teil des lac-Operons von E.coli und ist dadurch in der Lage den aminoterminalen Teil der ß-Galactosidase zu synthetisieren. Als Wirtszelle werden Bakterien eingesetzt, die den fehlenden, carboxyterminalen Bereich der ß-Galactosidase bilden. Wenn solche Zellen mit dem Vektor infiziert werden, bildet sich aus dem vektorkodierten N-terminalen und dem wirtszellkodierten C-terminalen Teil eine funktionsfähige ß-Galactosidase und die infizierten Zellen bilden bei Zugabe des Farbstoffes X-Gal blaue Kolonien. Da die Insertion von Fremd-DNA innerhalb des lac-Operons stattfindet und dadurch das Leseraster innerhalb des Gens unterbochen wird, kann durch den Einbau von Fremd-DNA keine ß-Galactosidase mehr gebildet werden und die enstehenden Kolonien bleiben weiß. Die Anwesenheit eines Plasmids in der Wirtszelle und damit der erfolgreiche Abschluß einer Transformation wird mit Hilfe von Antibiotikaresistenz-Genen getestet. Die Ligation von PCR-Produkt und Vektor und die anschließende Transformation des Ligationsproduktes in die Bakterienzellen wurde anhand des beiliegenden Protokolls durchgeführt. Nach Übernachtinkubation der Bakterien auf IPTG, X-Gal und Ampicillin enthaltenden Agarplatten, wurden von jeder Agarplatte 1-2 weiße Einzelkolonien in LB-Medium (siehe Anhang II) überimpft und wiederum 14-16 Stunden zur Vermehrung der Zellen im Schüttelbrutschrank belassen. 2.3.6 Aufreinigung der Plasmid-DNA Durch eine Aufreinigung wird die Plasmid-DNA von den anderen Bestandteilen des Reaktionsansatzes, z.B. Zellfragmenten von E. coli, getrennt und somit einer nachfolgenden DNA-Sequenzierung zugänglich gemacht. Die Aufreinigung der Plasmid-DNA erfolgte mit Hilfe des FastPlasmidTM Mini-Kits (Eppendorf) anhand des mitgelieferten Protokolls. Zur Kontrolle der Plasmid-DNA wurden 3µl des Isolates mit 3µl Bromphenolblau vermischt und auf einem Agarosegel bei 130 Volt für 20 min elektrophoretisch aufgetrennt. Zum Längenvergleich wurde ein Plasmid ohne insert aufgetragen, das sich durch eine geringere Lauflänge im Agarosegel unterscheiden lässt.

- 38 -
2.3.7 DNA-Sequenzierung Es existieren gegenwärtig zwei prinzipiell verschiedene Methoden DNA-Sequenzen zu erhalten. Zum Einen das von Maxam und Gilbert (1977) entwickelte Verfahren, bei dem basenspezifische Chemikalien zu Kettenabbrüchen führen, zum Anderen die enzymatische Methode von Sanger et al. (1977), die DNA-Polymerasen und Desoxynucleotide verwendet. Das enzymatische Verfahren hat sich einer Automation als zugänglich erwiesen und deshalb unter den Molekularbiologen weite Verbreitung gefunden. Das Didesoxyverfahren oder Kettenabbruchverfahren von Sanger basiert auf der enzymatisch katalysierten Synthese einer Population von basenspezifisch terminierten DNA-Fragmenten, die nach ihrer Größe gelelektrophoretisch aufgetrennt werden können. Aus dem in einem Polyacrylamidgel entstehenden Bandenmuster kann anschließend die Sequenz rekonstruiert werden. Ausgehend von einem zu sequenzierenden DNA-Abschnitt wird durch Zugabe eines Sequenzierungs-primers (analog zur PCR ein kurzes DNA-Oligonukleotid), eines Nukleotidgemisches und einer DNA-Polymerase die Synthese eines komplementären DNA-Stranges initiiert. Um die Reaktionsprodukte im Gel nachweisen zu können, werden diese mit Fluoreszenzfarbstoffen markiert. Die Reaktion wird in 4 parallelen Reaktionen angesetzt, die sich nur durch die Verwendung eines unterschiedlichen Nukleotidgemisches unterscheiden. Die mit A, C, G und T bezeichneten Ansätze enthalten eine Mischung aus den natürlich vorkommenden 2‘-Desoxynukleotiden (dNTP) und jeweils nur einem Typ synthetischer 2‘,3‘-Didesoxynukleotide (ddNTP), der sogenannten Terminatoren. Während der Strangsynthese durch die schrittweise Kondensation von Nukleosidtriphosphaten kann es nun zu zwei unterschiedlichen Reaktionsereignissen kommen: Beim Einbau eines normalen dNTPs kommt es zur weiteren Verlängerung des DNA-Moleküls, bei einer Kondensationsreaktion zwischen dem DNA-Strang und einem 2‘,3‘-Didesoxynukleotid dagegen, ist nach dem Einbau eine Verlängerung des Stranges nicht mehr möglich, da keine freie 3‘-Hydroxygruppe vorhanden ist. Der Strang ist somit terminiert. Es entstehen also in jedem der 4 Reaktionsansätze unterschiedlich lange DNA-Produkte, die jeweils nur auf einen Basentyp enden. Durch einen Überschuß an dNTPs, z.B. bei einem molaren Verhältnis dNTP:ddNTP von 200:1, sind Terminationsereignisse relativ selten und es entstehen lange Reaktionsprodukte von bis zu 1000 Bp Länge. Die Anwendung von thermostabilen DNA-Polymerasen ermöglicht in der DNA-Sequenzierung – analog zur PCR – die gleichzeitige Amplifikation und DNA-Sequenzierung. Dieses Verfahren wird cycle-sequencing genannt. Im Gegensatz zur PCR befindet sich in der Reaktion nur ein primer, es wird also lediglich linear und nicht exponentiell amplifiziert. Das Prinzip der cycle-sequencing-Reaktion besteht darin, daß die 4 Ansätze mit einem Gemisch aus primer, Ausgangs-DNA (template), Taq-Polymerase, dNTPs und ddNTPs, in einer zyklisch durchlaufenen Sequenzierungsreaktion verschiedenen Temperaturen unterworfen werden.

- 39 -
Bei hohen Temperaturen, z.B. 97°C, werden primer und template thermisch dissoziiert, bei der niedrigsten im Prozeß auftauchenden Temperatur, z.B. 55°C, lagert sich der primer an den komplementären DNA-Abschnitt an und bei einer mittleren Temperatur von z.B. 68°C finden Kettenverlängerung und Termination statt. Kriterien für den Entwurf von Sequenzier-primern (und PCR-primern): 1. Die primer müssen lang genug sein, um eine spezifische Bindung innerhalb der
DNA zu gewährleisten; in der Regel bestehen sie aus ca. 18-30 Basen. 2. Es darf nicht zur Ausbildung von Sekundärstrukturen innerhalb der primer
kommen. 3. Die primer dürfen nicht untereinander hybridisieren. 4. Am 3‘-Ende sollten Guanine und Cytosine liegen, da sie eine energetisch
stabilere Bindung als Adenine und Thymine aufweisen. 5. Der GC-Gehalt insgesamt sollte bei 40-60% liegen. Zur Überprüfung dieser Kriterien und Optimierung der zu entwerfenden primer wurde das Programm Oligo (Rychlik & Rhoads 1989) verwendet. Verwendete Oligonukleotide: Bei den mit Universal und Reverse bezeichneten Sequenzier-primern handelt es sich um universelle Oligonukleotide, die innerhalb des Kloniervektors, also außerhalb des eingefügten PCR-Produktes, hybridisieren. Die übrigen primer binden innerhalb des klonierten Produktes, wobei die Zahlen die ungefähre Lage innerhalb des Gens andeuten und die Buchstaben den kodierenden (F), bzw. den dazugehörigen Gegenstrang (R) kennzeichnen. Alle verwendeten Oligonukleotide wurden von der Firma MWG-Biotech synthetisiert und am 5‘-Ende mit einem Fluoreszenzfarbstoff markiert. Bezeichnung Sequenz (5‘ - 3‘) Quelle
Universal AGG GTT TTC CCA GTC ACG ACG TT Messing et al. 1981
Reverse TCA CAC AGG AAA CAG CTA TGA C Messing et al. 1981
18S rDNA 18A1 seq CTG GTT GAT CCT GCC AGT CAT ATG C Spears (pers. Mitteilung.)
400F seq ACG GGT AAC GGG GAA TCA GGG Wollscheid, Dreyer & Englisch
470F seq CAG CAG GCA CGC AAA TTA CCC Wollscheid, Dreyer & Englisch
700F seq GTC TGG TGC CAG CAG CCG CG Wollscheid, Dreyer & Englisch
1155F seq CTG AAA CTT AAA GGA ATT GAC GG Wollscheid, Dreyer & Englisch
1600F seq CGT CCC TGC CCT TTG TAC ACA CC Wollscheid, Dreyer & Englisch
1800R seq GAT CCT TCC GCA GGT TCA CCT ACG Spears (pers. Mitteilung)
1500R seq CAT CTA GGG CAT CAC AGA CC Wollscheid, Dreyer & Englisch
1155R seq CCG TCA ATT CCT TTA AGT TTC AG Wollscheid, Dreyer & Englisch
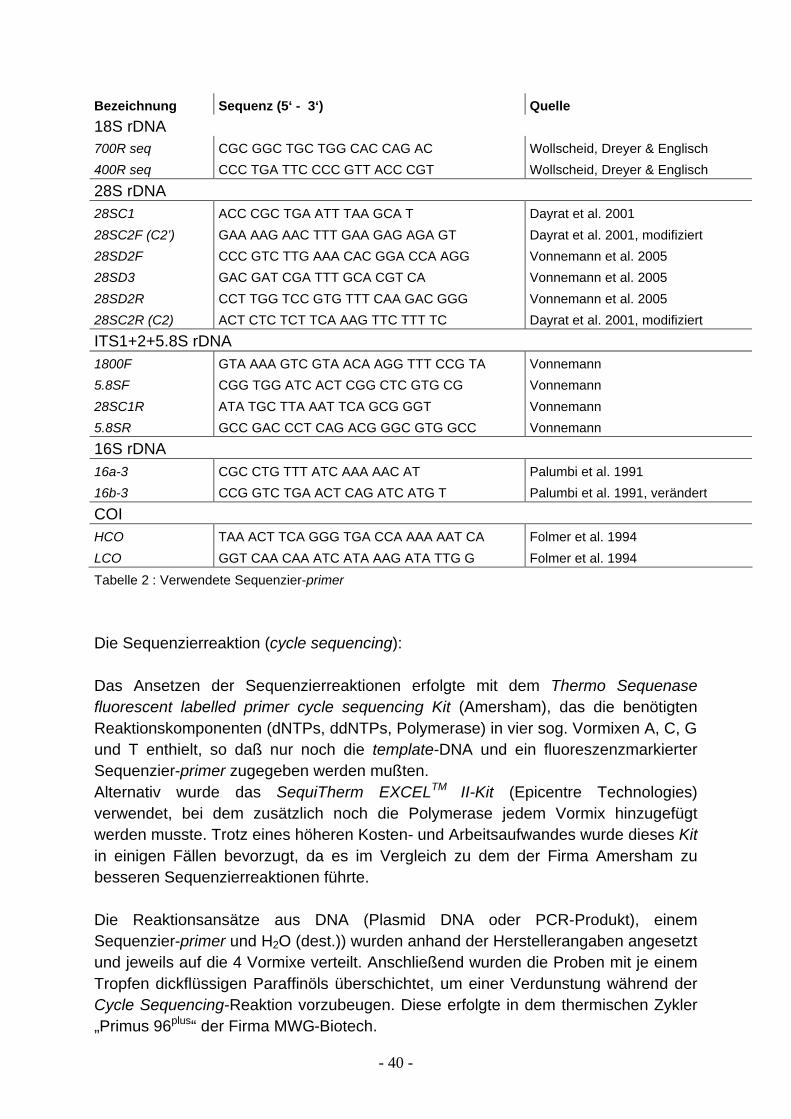
- 40 -
Bezeichnung Sequenz (5‘ - 3‘) Quelle
18S rDNA
700R seq CGC GGC TGC TGG CAC CAG AC Wollscheid, Dreyer & Englisch
400R seq CCC TGA TTC CCC GTT ACC CGT Wollscheid, Dreyer & Englisch
28S rDNA 28SC1 ACC CGC TGA ATT TAA GCA T Dayrat et al. 2001
28SC2F (C2’) GAA AAG AAC TTT GAA GAG AGA GT Dayrat et al. 2001, modifiziert
28SD2F CCC GTC TTG AAA CAC GGA CCA AGG Vonnemann et al. 2005
28SD3 GAC GAT CGA TTT GCA CGT CA Vonnemann et al. 2005
28SD2R CCT TGG TCC GTG TTT CAA GAC GGG Vonnemann et al. 2005
28SC2R (C2) ACT CTC TCT TCA AAG TTC TTT TC Dayrat et al. 2001, modifiziert
ITS1+2+5.8S rDNA 1800F GTA AAA GTC GTA ACA AGG TTT CCG TA Vonnemann
5.8SF CGG TGG ATC ACT CGG CTC GTG CG Vonnemann
28SC1R ATA TGC TTA AAT TCA GCG GGT Vonnemann
5.8SR GCC GAC CCT CAG ACG GGC GTG GCC Vonnemann
16S rDNA 16a-3 CGC CTG TTT ATC AAA AAC AT Palumbi et al. 1991
16b-3 CCG GTC TGA ACT CAG ATC ATG T Palumbi et al. 1991, verändert
COI HCO TAA ACT TCA GGG TGA CCA AAA AAT CA Folmer et al. 1994
LCO GGT CAA CAA ATC ATA AAG ATA TTG G Folmer et al. 1994
Tabelle 2 : Verwendete Sequenzier-primer
Die Sequenzierreaktion (cycle sequencing): Das Ansetzen der Sequenzierreaktionen erfolgte mit dem Thermo Sequenase fluorescent labelled primer cycle sequencing Kit (Amersham), das die benötigten Reaktionskomponenten (dNTPs, ddNTPs, Polymerase) in vier sog. Vormixen A, C, G und T enthielt, so daß nur noch die template-DNA und ein fluoreszenzmarkierter Sequenzier-primer zugegeben werden mußten. Alternativ wurde das SequiTherm EXCELTM II-Kit (Epicentre Technologies) verwendet, bei dem zusätzlich noch die Polymerase jedem Vormix hinzugefügt werden musste. Trotz eines höheren Kosten- und Arbeitsaufwandes wurde dieses Kit in einigen Fällen bevorzugt, da es im Vergleich zu dem der Firma Amersham zu besseren Sequenzierreaktionen führte. Die Reaktionsansätze aus DNA (Plasmid DNA oder PCR-Produkt), einem Sequenzier-primer und H2O (dest.)) wurden anhand der Herstellerangaben angesetzt und jeweils auf die 4 Vormixe verteilt. Anschließend wurden die Proben mit je einem Tropfen dickflüssigen Paraffinöls überschichtet, um einer Verdunstung während der Cycle Sequencing-Reaktion vorzubeugen. Diese erfolgte in dem thermischen Zykler „Primus 96plus“ der Firma MWG-Biotech.

- 41 -
Reaktionsprofil: Initiale Denaturierung: 95°C 2 min Denaturierung, primer-Hybridisierung und Polymerisation: 94°C 25 sec 46-56°C 25 sec 70°C 25 sec Abschließende Polymerisation: 4°C 8 Nach Beendigung der Reaktion wurden die Proben mit jeweils 3,0 µl Stop-Puffer überschichtet und für 30 sec bei 95°C denaturiert. Anschließend wurden die Reaktionen bis zum Auftragen bei –20°C gelagert. Die Polyacrylamid-Gelelektrophorese: Die Auftrennung des erzeugten Fragmentgemisches erfolgte durch Elektrophorese auf einem 6%igen, denaturierenden Polyacrylamidgel in den automatischen Sequenziergeräten „4000“ und „4200IR2“ der Firma LI-COR. Das verwendete Polyacrylamidgel wurde durch die Mischung dreier Komponenten erzeugt: 24 ml Sequagel XR 6% (National Diagnostics) 6 ml Sequagel Complete Buffer Reagent (National Diagnostics) 300 µl Ammoniumpersulfat (10%) Das flüssige Gel wurde in das nach Herstellerangaben vorbereitete, gründlich gereinigte Glasplattensystem des Sequenziergerätes gegossen und ein Vorkamm als Platzhalter eingesetzt. Das Gel polymerisierte anschließend innerhalb von 1 bis 2 Stunden aus. Die gewünschte Schichtdicke von 0,25 Millimetern wurde durch Kunststoffstreifen derselben Stärke, die sich zwischen den Glasplatten befanden, erzeugt. Nach dem Einbau des Gelsystems in den Sequenzierer, wurden die beiden Puffertanks mit insgesamt 1 Liter TBE-Laufpuffer (siehe Anhang 5.2.2) befüllt und der Vorkamm aus dem oberen Bereich des Gels entfernt. Die damit freigelegte Gelkante wurde von Gelresten und Harnstoffverunreinigungen befreit und anschließend wurde das Polyacrylamidgel auf 50°C vorgeheizt. Nach Erreichen dieser Temperatur wurde die Gelkante erneut gereinigt und der Probenkamm zur Aufnahme der Sequenzierreaktionen eingesetzt.
33 x

- 42 -
Die durch den Kamm geformten 64 Taschen wurden mit einer geringen Menge jeder Probe beschickt. Da jeder Sequenzieransatz aus vier Fragmentgemischen besteht, die in der Reihenfolge A, C, G und T aufgetragen werden, können durch eine Elektrophoresereaktion 16 verschiedene Ansätze aufgetrennt werden. Durch einen im Gerät integrierten Laser und Detektor konnten die im vertikalen Gel wandernden fluoreszenzmarkierten DNA-Moleküle erkannt werden. Die Laufbedingungen für eine Polyacrylamid-Gelelektrophorese stellen sich wie folgt dar: 31 Watt 50°C 1500 Volt 12 Stunden 2.4 Methoden der Auswertung 2.4.1 Erstellen einer Konsensussequenz Nach Aufnahme der Sequenzierdaten erzeugt die Rechnereinheit des Sequenziergerätes mit Hilfe des Programmes BaseImage IR einen Sequenziervorschlag für jede Probe unter Auswertung der vier parallel laufenden Teilreaktionen. Nicht eindeutig identifizierbare Basen werden als solche markiert und müssen später, durch Vergleich mit der Base an der entsprechenden Stelle im Gegenstrang, manuell korrigiert werden. Die Erstellung einer Konsensussequenz aus den einzelnen Teilfragmenten sowie die Korrektur der „Rohdaten“ erfolgte mit Hilfe des Programmes AlignIR (LI-COR Biosciences). 2.4.2 Kontrolle der erzeugten Sequenzen Eine Kontrolle der erhaltenen Sequenzen wird durchgeführt, um eine Kontamination der Proben mit Fremd-DNA auszuschließen und die Identität der Sequenzen zu bestätigen. Dazu werden die Sequenzdaten mit Hilfe des Programmes BLAST (http://www.genome.ad.jp/htbin/nph-blast) mit bereits in den internationalen Gendatenbanken veröffentlichten Daten verglichen. Die Identität der eigenen Sequenzen ist als gesichert anzusehen, wenn die größten erzielten Übereinstimmungen mit nah verwandten Taxa bestehen.

- 43 -
2.4.3 Alinierung der DNA-Sequenzen
Um aus den gewonnenen Sequenzdaten phylogenetische Schlüsse ziehen zu können, müssen die einzelnen DNA-Sequenzen eines Gens miteinander verglichen werden, wobei homologe Abschnitte aliniert, das heißt „übereinander angeordnet“ werden. Durch die Annahme von Substitutionen und das Einfügen von Lücken, stellvertretend für Insertionen/Deletionen, besteht die Möglichkeit, selbst stark differierende Nukleotidabfolgen zu alinieren. Wenn allerdings das Einfügen von Lücken unbegrenzt möglich wäre, könnten selbst nicht verwandte Sequenzen so aliniert werden, dass Nukleotide entweder perfekt passen oder durch Lücken getrennt werden. Es ist offensichtlich, dass eine solche Alinierung keinerlei Information enthält. Es ist deshalb notwenig, das Einfügen von Lücken zu beschränken. Das geschieht in der Praxis durch ein Bewertungssystem, in dem Übereinstimmungen einen positiven numerischen Wert erhalten und Lücken einen negativen, die sog. gap penalty. Das zentrale Prinzip der Sequenzalinierung ist es nun, die Alinierung mit dem höchstmöglichen Wert zu generieren. Die Alinierung der DNA-Sequenzen erfolgte mit Hilfe des Programmes CLUSTAL X 1.64b (Thompson 1997). Die implementierten Alinierungsparameter wurden dabei nicht verändert, da sie sich als am besten geeignet herausgestellt haben (Hall 2001). In einem ersten Schritt wird durch paarweise Alinierung aller Sequenzen eine Distanzmatrix erstellt, auf deren Grundlage eine ungewurzelte Baumtopologie erzeugt wird. Anschließend erfolgt eine Wurzelung des Baumes an derjenigen Stelle, die zu gleichen Astlängen auf jeder Seiten der Wurzel führt. Nach einer Gewichtung der einzelnen Sequenzen anhand ihrer Distanz zur Wurzel, werden die beiden terminalen Sequenzen mit der geringsten Distanz zueinander aliniert. Zu dem entstehenden Cluster wird dann die jeweils nächststehende Sequenz hinzugefügt. Der gesamte Alinierungsprozeß wird durch die Berücksichtigung der Sequenz an der Basis der Wurzel beendet. Alle verfügbaren Alinierungsprogramme besitzen den Nachteil, daß die erzeugte Alinierung von der Reihenfolge der eingelesenen Sequenzen abhängig ist. Zudem können die Programme nicht zwischen konservierten und hochvariablen Sequenzbereichen unterscheiden, so daß die Kosten für das Einfügen von Lücken in den konservierten Bereichen zu niedrig, in den variablen Bereichen dagegen zu hoch angesetzt werden. Aufgrund dieser Unzulänglichkeiten wurde eine manuelle Nachbearbeitung der computergenerierten Alinierung mit Hilfe des Programmes Genedoc (Nicholas & Nicholas 1998) notwendig.

- 44 -
Die manuelle Überarbeitung erfolgte nach verschiedenen, teils subjektiven Kriterien: o Beseitigung offensichtlicher Fehler, z.B. unnötige Lücken oder verschobene
Sequenzabschnitte o Optimierung homologer Merkmale bei morphologisch gut gestützten Taxa o Maximierung von Übereinstimmungen durch das Einfügen von Lücken 2.4.4 Verfahren zur Erstellung von Dendrogrammen Die molekularen Stammbaumrekonstruktionsverfahren lassen sich im Wesentlichen in vier Gruppen unterteilen: 1. Distanzmethoden 2. Maximum Likelihood-Verfahren 3. Maximum Parsimonie-Methoden 4. Bayes’sche Verfahren Die drei letztgenannten Verfahren können unter dem Begriff „Diskrete Methoden“ zusammengefaßt und den Distanzmethoden gegenübergestellt werden. Die genannte Einteilung basiert auf der Art und Weise wie die Sequenzdaten ausgewertet werden. Bei den Distanzmethoden werden die Daten zuerst in eine Distanzmatrix übersetzt, die die Basis für eine Stammbaumrekonstruktion bildet, wohingegen die diskreten Methoden die Informationen der Sequenzunterschiede direkt nutzen. 2.4.4.1 Distanzmethoden Distanzmethoden zu phylogenetischen Rekonstruktion basieren auf der Grundannahme, dass bei Kenntnis der aktuellen evolutionären Distanz zwischen allen Sequenzen eines Datensatzes, die evolutionäre Geschichte dieser Sequenzen unmittelbar rekonstruiert werden kann. Die einfachste Methode der Distanzabschätzung zweier Nukleotidsequenzen besteht in der Feststellung der Anzahl unterschiedlicher Nukleotidpositionen. Bei stärker differierenden Sequenzen ist diese Zahl allerdings ein schlechtes Maß für die aufgetretenen evolutionären Veränderungen, da Mehrfachsubstitutionen, also Substitutionen an exakt derselben Stelle in einem Gen, nicht erfaßt werden können. Mit steigender Divergenzzeit zweier Taxa wird demnach die tatsächlich aufgetretene Zahl von Mutationen in zunehmendem Maße unterschätzt, so dass z.B. Taxa, die sich vor 20 Millionen Jahren entwicklungsgeschichtlich getrennt haben, keine höhere totale Sequenzdivergenz aufweisen als Taxa, die seit 15 Millionen Jahren getrennt evolvieren. Die Berücksichtigung dieser multiplen Substitutionen ist von entscheidender Bedeutung, da Distanzverfahren nur dann die korrekte

- 45 -
Rekonstruktion einer Phylogenese erlauben, wenn die Distanzwerte zwischen Sequenzpaaren auch ein Maß für die Divergenzzeit sind. Zur „Korrektur“ der sichtbaren Distanzen wurde eine Reihe unterschiedlicher Modelle entwickelt, die größtenteils hierarchisch sind, sich also nur in der Anzahl implementierter Parameter unterscheiden. Im Folgenden werden einige Sequenzevolutionsmodelle und ihre Unterschiede beispielhaft vorgestellt: Jukes-Cantor-Modell (JC, Jukes & Cantor 1969) Dieses einfachste Modell der Sequenzevolution nimmt an, daß alle 4 Basen die gleiche Häufigkeit innerhalb des Datensatzes aufweisen und daß alle Substitutionen gleich wahrscheinlich sind. Kimuras 2-Parameter-Modell (K80, Kimura 1980) In diesem Modell werden, im Vergleich zum vorhergehenden, unterschiedliche Wahrscheinlichkeiten für Transitionen und Transversionen berücksichtigt. Felsenstein-Modell (F81, Felsenstein 1981) Das Modell von Felsenstein erlaubt das Auftreten unterschiedlicher Basenfrequenzen; Transitionen und Transversionen werden aber als gleichwahrscheinlich angesehen. Hasegawa-Kishino-Yano-Modell (HKY, Hasegawa et al. 1985) Das Modell von Hasegawa, Kishino und Yano kombiniert die beiden vorhergehenden, indem sowohl unterschiedlichen Basenfrequenzen als auch unterschiedlichen Substitutionsraten für Transitionen und Transversionen Rechnung getragen wird. General reversible-Modell (GTR, Rodríguez et al. 1990, Yang et al. 1994) Das sog. general reversible-Modell repräsentiert das komplexeste der vorgestellten Modelle. Nicht nur Transitionen und Transversionen haben unterschiedliche Raten, sondern alle 6 möglichen Substitutionen (A - C, A - G, A - T, C - G, C - T, G - T) weisen eine eigene Wahrscheinlichkeit auf. Daneben werden unterschiedliche Basenhäufigkeiten berücksichtigt. Neben Basenfrequenzen und Substitutionswahrscheinlichkeiten existieren weitere Parameter, die in die Korrektur der sichtbaren Distanzen einfließen können und sollten. Alle vorgestellten Modelle implizieren, dass die Substitutionsraten über die komplette Sequenz einer Sequenz hinweg konstant sind. Das ist in der Realität nachgewiesenermaßen nicht der Fall. Der meistverwendete Ansatz zur Berücksichtigung dieser Variabilität nutzt die sog. Gamma-Verteilung. Diese Verteilung hat den Formparameter a, der den Umfang der Variation der

- 46 -
Substitutionsraten unter den Nukleotiden einer Sequenz spezifiziert. Kleine a-Werte resultieren in einer extremen Variabilität der Raten, je größer a ist, desto geringer wird die Variationsbreite. Untersuchungen (Yang 1996) zeigen, dass die Werte für den Formparameter a bei nukleären und mitochondrialen Genen zwischen 0,16 und 1,37 liegen, das bedeutet, die meisten Nukleotidpositionen sind invariabel, während wenige Positionen sehr hohe Substitutionsraten aufweisen. Der Anteil dieser invariablen Positionen an der Gesamtanzahl der Nukleotide ist ein weiterer Parameter den es zu berücksichtigen gilt (Page & Holmes 1998). Die Problematik besteht nun in der Auswahl des für den zu untersuchenden Datensatz am besten geeigneten Sequenzevolutionsmodells. Ein Modell erscheint umso realistischer, je mehr unterschiedliche Parameter eingesetzt werden. Da aber der Wert jedes hinzugefügten Parameters aus dem vorliegenden Datensatz abgeschätzt werden muß, addieren sich bei steigender Parameterzahl auch Ungenauigkeiten in den Schätzungen. Dadurch verlieren die Sequenzdaten an Aussagekraft. Einfache Modelle können dagegen die reale Sequenzevolution nur unzureichend widerspiegeln. Das Ziel ist es daher ein Modell zu wählen, das so einfach wie möglich und so komplex wie nötig ist. Ein bekanntes Verfahren um das für einen bestimmten Datensatz am besten geeignete Sequenzevolutionsmodell zu ermitteln, ist der sog. likelihood ratio test (Goldman 1993). Likelihood ratio test Der likelihood ratio test (LRT) ist ein statistischer Test, mit dessen Hilfe überprüft werden kann, ob ein komplexeres Modell gegenüber einem einfacheren Modell eine signifikante Verbesserung der Wahrscheinlichkeit für eine Topologie bewirkt. Ist die Verbesserung nicht signifikant, ist es sinnvoll, das einfachere Sequenzevolutionsmodell beizubehalten. In der praktischen Anwendung des LRT erfolgt der Vergleich zwischen 2 Modellen anhand einer Prüfstatistik A, mit A = log L1 – log L0. Dabei ist L0 die maximale Wahrscheinlichkeit der Nullhypothese, also des einfacheren Modells und L1 die maximale Wahrscheinlichkeit der alternativen Hypothese, das heißt des komplexeren, parameter-reicheren Modells. Der LRT ist nur für den Vergleich von hierarchisch verknüpften Modellen gültig, das heißt, das komplexere Modell darf sich von dem einfacheren nur durch die Addition eines oder mehrerer zusätzlicher Parameter unterscheiden. Das Hinzufügen weiterer Parameter wird grundsätzlich in einer Erhöhung des Likelihood Wertes resultieren, doch ab einem bestimmten Punkt ist diese Erhöhung statistisch gesehen nicht mehr signifikant. Für die Durchführung des LRT wurde das Programm Modeltest Version 3.06 (Posada & Crandall 1998) verwendet. Es erlaubt den Vergleich von insgesamt 56 verschiedenen Modellvarianten. Darunter sind neben den 5 oben vorgestellten Modellen (JC, K80, F81, HKY, GTR) noch 9 weitere. Diese 14 Modelle sind jeweils in

- 47 -
4 Varianten implementiert: unter Berücksichtigung der Ratenvariabilität, unter Berücksichtigung der Anzahl invariabler Positionen, mit beiden zusätzlichen Parametern oder ohne beide. Nach der Distanztransformation der sichtbaren Distanzen mit Hilfe der Modelle zur Sequenzevolution in „reale“, evolutionäre Distanzen, kann eine Rekonstruktion der Phylogenie erfolgen. Um auf der Basis einer Distanzmatrix einen Stammbaum zu erstellen, sind verschiedene Methoden verfügbar, von denen das neighbor-joining genannte, heuristische Suchverfahren am geeignetsten erscheint. Das sog. UPGMA-(unweighted pairgroup method using arithmetic averages) Verfahren setzt z.B., im Gegensatz zum neighbor-joining-Verfahren, voraus, dass die Substitutionsrate in allen Linien bis hin zu den terminalen Taxa identisch ist. In der Realität ist das selten der Fall, wodurch es zu falschen Ergebnissen bei der Baumkonstruktion kommen kann. Mit Hilfe des neighbor-joining-Verfahrens kann unter dem minimum evolution-Aspekt der kürzestmögliche Baum gesucht werden, das heißt derjenige, dessen Summe aller Äste minimal ist (minimum evolution tree). Das zugrundeliegende Prinzip ist, genau wie bei einer Maximum Parsimonie-Analyse, das sog. Sparsamkeitsprinzip. Das neighbor-joining-Verfahren Auf der Grundlage der korrigierten Distanzmatrix werden die Distanzen aller Taxapaare berechnet. Für jedes Taxon kann daraufhin durch Addition eine Gesamtdistanz zu allen anderen Taxa des Datensatzes ermittelt werden. Die Unterschiede in der Substitutionsrate zwischen homologen Merkmalen werden durch Mittelung ausgeglichen. Dieser gemittelte Distanzwert für 2 Taxa i und j ergibt sich aus Mij = dij – ri + rj / N – 2, wobei dij die evolutionäre Distanz zwischen 2 Taxa repräsentiert, ri und rj die Gesamtdistanzen der Taxa i und j und N die Gesamtzahl aller Taxa. In einer neuen Matrix aus M-Werten sind diejenigen Arten Nachbarn, die den negativsten M-Wert aufweisen. Für sie wird ein Grundmuster u angenommen, das Mij in einer neuen Matrix ersetzt. Für u wird wiederum die Gesamtdistanz bestimmt. Alle Distanzen zu den Taxa i und j werden aus dem Datensatz entfernt und N um 1 verringert. Anschließend beginnt die ganze Prozedur mit der Erstellung einer Distanzmatrix, in der 2 Taxa i und j durch ihr Grundmuster u ersetzt wurden, von neuem. 2.4.4.2 Maximum Parsimonie-Verfahren Nach dem Prinzip der sparsamsten Erklärung (Parsimonie-Prinzip) wird in der Wissenschaft diejenige Hypothese bevorzugt, die mit den wenigsten Zusatzannahmen auskommt, die also die einfachste mögliche Erklärung eines Sachverhaltes liefert.

- 48 -
In Anwendung dessen beruht das Maximum Parsimonie-Verfahren auf der Auswahl des Stammbaumes, der die wenigsten evolutionären Veränderungen voraussetzt. Eine Merkmalsänderung ist dabei entweder das Auftreten eines neuen Mermals, das vorher nicht existierte oder die Zustandsänderung einer Homologie (Wägele 2000). Der einer Parsimonie-Analyse zugrundeliegende Datensatz besteht dabei aus den Nukleotidsequenzen und ihren taxonspezifischen Unterschieden. Als parsimonie-informativ gelten Merkmale, die mit zwei oder mehr Zuständen bei jeweils mindestens zwei terminalen Taxa vorkommen. Diese Merkmale erzeugen in einem Datensatz einen split, beziehungsweise in einem Stammbaum eine Verzweigung. Das Auffinden der sparsamsten, also kürzesten Topologie durch gezielte Berechnung ist allerdings nicht möglich. Vielmehr müssen alle möglichen Topologien erzeugt und ihre jeweilige Länge berechnet werden. In der Praxis ist eine solche, exakte Suche wegen des enormen Rechenaufwandes nicht durchführbar, so dass auf heuristische Suchverfahren zurückgegriffen werden muß. Eine heuristische, sowie eine exakte Suche beginnen mit der Wahl beliebiger 3 Taxa, aus denen ein erster ungewurzelter Baum konstruiert wird. Weitere Taxa werdem nacheinander an allen Kanten des Baumes eingefügt, wodurch bei 4 Taxa genau 3 verschiedene Topologien entstehen, bei 5 Taxa 15, bei 10 Taxa bereits rund 2 Millionen unterschiedliche Bäume. Bei einer heuristischen Suche wird nach der Addition eines neuen Taxons nur der kürzeste Baum beibehalten, der dann die Grundlage für die Addition des nächsten, zufällig gewählten Taxons bildet. Im Gegensatz zur exakten Suche gibt es bei einer heuristischen Suche keine Sicherheit für das Auffinden des sparsamsten Baumes, da möglicherweise nur ein lokales, anstelle des globalen Optimums (siehe z.B. Wägele 2000) erreicht wurde. Im Anschluß an eine heuristische Suche wird deshalb durch Verlagerung von Ästen (branch swapping) versucht, eine noch sparsamere Topologie zu finden. Für die Verlagerung von Ästen werden 3 unterschiedliche Strategien angewendet. Beim nearest neighbor interchange (NNI) werden 2 der jeweils 4 mit einer inneren Kante verknüpften Äste ausgetauscht, beim subtree pruning and regrafting (SPR) wird ein Ast abgetrennt und an einer beliebigen Stelle der Topologie wieder eingefügt und beim tree bisection-reconnection (TBR) teilt man eine Topologie an einer Stelle in 2 Dendrogramme, die dann auf andere Weise wieder miteinander verbunden werden. Die NNI-Methode erfordert den geringsten Zeitaufwand, die TBR-Verlagerung ist am zeitaufwendigsten. Bei der Exaktheit der Ergebnisse verhält es sich in der Regel entsprechend umgekehrt. 2.4.4.3 maximum likelihood-Verfahren Bei Anwendung des maximum likelihood-Verfahrens wird nach demjenigen Stammbaum gesucht, der die vorliegenden Daten auf der Grundlage der ausgewählten Baumtopologie am wahrscheinlichsten erscheinen läßt. Die Berechnung der likelihood (LD) erfolgt nach

- 49 -
LD = Pr (D/H), wobei Pr die Wahrscheinlichkeit des vorliegenden Datensatzes (D) auf der Basis des gegebenen Stammbaumes (H) darstellt. Den ersten Schritt bei Anwendung des maximum likelihood-Verfahrens stellt die Auswahl eines Dendrogrammes dar. Für die Dendrogrammkonstruktion können die bereits erwähnten Methoden, z.B. neighbor joining angewendet werden. Der zweite Schritt besteht in der Wahl eines Modells zur Sequenzevolution (siehe Kapitel 2.4.4.1). Ausgehend davon wird die Wahrscheinlichkeit berechnet, daß bei der gegebenen Topologie und den vorausgesetzten Substitutionsraten die terminalen Sequenzen aus der vorgegebenen Alinierung entstehen können. Dieser Vorgang wird für alle möglichen Stammbäume durchgeführt und dann diejenige Topologie ausgewählt, für die die Wahrscheinlichkeit am höchsten ist. In der praktischen Anwendung stößt dieses Verfahren durch seinen großen Zeitaufwand bei der Bearbeitung umfangreicher Datensätze schnell an die Grenzen der Praktikabilität. 2.4.4.4 Bayes’sche Verfahren Thomas Bayes war ein englischer Presbyterianerpriester, der im 18. Jahrhundert in der Nähe von London gelebt hat. Neben seiner Tätigkeit als Priester hat er sich mit Mathematik, besonders mit der Zufallslehre, beschäftigt. Erst nach seinem Tod wurde seine Wahrscheinlichkeitstheorie unter dem Titel: “An Essay towards solving a problem in the doctrine of chances“ (Bayes 1763) veröffentlicht. Seinen Ansatz kann man den sogenannten „klassischen Methoden“ der Wahrscheinlichkeitstheorie gegenüberstellen. Bei den klassischen Methoden wird die Wahrscheinlichkeit als relative Häufigkeit interpretiert, d.h. die Wahrscheinlichkeit eines Ereignisses ist ein fester Wert, gegen den die relative Häufigkeit des Ereignisses konvergiert, wenn man das Experiment unendlich oft durchführt. Das Bayes’sche Verfahren nutzt dagegen Informationen, die bereits vor Erhebung der Daten vorliegen, sog. a priori-Informationen (prior probability). Diese a priori-Informationen können aus vorherigen Versuchen stammen oder rein subjektive Einschätzungen sein. Im Gegensatz zu den klassischen Verfahren ist die Wahrscheinlichkeit hier kein fester Wert sondern eine zufällige Größe. Vor der Analyse der Daten wird mit Hilfe der a priori-Informationen ein sog. a priori-Verteilungsmodell für die unbekannten Parameter erstellt, z.B. die Wahrscheinlichkeit. Anschließend nutzt man die vorliegenden Daten um diese erste Einschätzung der Wahrscheinlichkeit „korrektur zu lesen“. Das Ergebnis besteht dann in einem sog. a posteriori-Verteilungsmodell (posterior probality distribution).

- 50 -
Die einfachste Formulierung des Bayes'schen Theorems ist die folgende, die in der Literatur im Allgemeinen als Bayes'sche Formel bezeichnet wird: P(B A) P(A) P(A B) = P(B) Dabei stellen A und B zwei voneinander unabhängige Ereignisse dar. In Anwendung der Bayes’schen Verfahren auf phylogenetische Fragestellungen bedeutet das: P(Daten | Baum) P(Baum) P(Baum | Daten) = P(Daten) Dabei repräsentiert P(Baum | Daten) die posterior probability distribution, P(Daten | Baum) die likelihood einer Topologie (siehe Kap. 2.4.4.3) und P(Baum) ihre a priori-Wahrscheinlichkeit. Die Wahrscheinlichkeit der Daten, P(Daten), berechnet sich aus der Summation der Produkte aller möglichen Topologien und ihrer jeweiligen a priori-Wahrscheinlichkeit. In der Praxis werden gewöhnlich alle auf der Grundlage eines Datensatzes konstruierbaren Stammbäume a priori als gleichwahrscheinlich betrachtet, damit gilt: P(Baum) = 1/ Anzahl aller möglichen Topologien (Huelsenbeck, Ronquist & Hall: MrBayes, Manual). Die a posteriori-Wahrscheinlichkeitsverteilung kann nicht analytisch berechnet werden, stattdessen wird eine Simulationstechnik verwendet um die Wahrscheinlichkeitsverteilung zu schätzen, die Markov chain Monte Carlo (MCMC, Metropolis et al. 1953, Hastings 1970) genannt wird. Die Bayes’schen Analysen zur phylogenetischen Rekonstruktion wurden mit Hilfe des Programmes MrBayes v3.0b4 (Huelsenbeck & Ronquist 2001a) durchgeführt. 2.5 bootstrap-Verfahren Die Ursache für eine unsichere oder falsche Darstellung der realen Verwandtschaftsverhältnisse liegt nicht immer, oder nur zum Teil, in den Unzulänglichkeiten des angewendeten Rekonstruktionsverfahrens, sondern auch in der Beschaffenheit des zugrundeliegenden Datensatzes selbst. Durch das Vorhandensein von Homoplasien in der Alinierung nimmt die Auswahl der Merkmale,

- 51 -
also der phylogenieinformativen Nukleotidpositionen, eine entscheidende Stellung bei der Generierung von Dendrogrammen ein. Die Konsequenz besteht in der Abhängigkeit der Baumtopologie von der Auswahl der Merkmale. Um das Ausmaß dieses sampling error abschätzen zu können, wird das bootstrap-Verfahren (Efron 1979) eingesetzt. Beim Bootstrap-Test, der von Felsenstein (1985) in die molekulare Phylogenetik eingeführt wurde, werden aus einer gegebenen Alinierung Zufallsdatensätze gleichen Umfangs generiert. Dabei können manche Nukleotidpositionen häufiger, andere dagegen gar nicht in diesen neuen Alinierungen enthalten sein. Aus jedem der 100 bis 1000 Datensätze wird unter Anwendung des ausgewählten Rekonstruktionsverfahrens ein Dendrogramm erstellt. Nach Abschluß aller Replikationen werden die Ergebnisse in einem Konsensusbaum dargestellt, wobei für jeden Knotenpunkt berechnet wird, in wie vielen der Dendrogramme dieser Knoten aufgetreten ist. Knotenpunkte mit bootstrap-Werten von mehr als 95 Prozent werden als das Ergebnis eines guten phylogenetischen Signals angesehen (Felsenstein 1985). Der bootstrap-Test kann jedoch weder die Zuverlässigkeit des ausgewählten Rekonstruktionsverfahren belegen, noch läßt sich eine Aussage über die Qualität des Datensatzes treffen. Es ist lediglich möglich zu beurteilen, inwiefern eine auf einem gegebenen Datensatz basierende Topologie vom gewählten Rekonstruktionsverfahren unterstützt wird. 2.6 Prüfung der Plausibilität von Dendrogrammen 2.6.1 Vergleich mit Dendrogrammen auf der Basis alternativer Daten Durch Übereinstimmungen in Dendrogrammen, die auf der Basis unterschiedlicher Daten erzeugt wurden, kann mit relativ hoher Wahrscheinlichkeit auf eine real existierende Verwandtschaft geschlossen werden. Allerdings ist zu beachten, dass auch nicht-zufällige Übereinstimmungen unter bestimmten Voraussetzungen falsch sein können. Beispielsweise können sich bei Verwendung derselben Rekonstruktionsmethode systematische Fehler einschleichen oder es kann bei rascher Radiation eines Taxons zu einer Überlagerung ursprünglich vorhandener synapomorpher Merkmale kommen (Wägele 2000). 2.6.2 Vergleich mit Fossilfunden Die Projektion von Fossildaten auf eine Stammbaumhypothese kann ebenfalls Aufschluß geben über die Plausibilität eines Dendrogrammes. Die im Stammbaum basal stehenden Taxa sollten demnach die erdgeschichtlich ältesten Gruppen repräsentieren, während die abgeleiteten Formen jüngeren Ursprungs sein sollten. Dieser Zusammenhang ist allerdings nur gegeben, wenn das erste zeitliche Auftreten einer Tiergruppe mit dem anhand von Fossilien datierten Alter übereinstimmt.

- 52 -
2.6.3 Vergleich mit biologischen Daten Neben Fossildaten können auch biologische Merkmale auf vorhandene Stammbaumtopologien projiziert werden, um die Plausibilität der rekonstruierten Verwandtschaftshypothese zu prüfen. Es eignen sich dafür unter anderem biogeographische oder ethologische Daten, z.B. unterschiedliche Ernährungs- oder Verteidigungsstrategien. Auch Einzelmerkmale, z.B. Chromosomenzahlen, können zur Bewertung von Dendrogrammen beitragen. Im Allgemeinen werden dabei phylogenetische Szenarien als eher unwahrscheinlich bewertet, bei denen es wiederholt zur Entstehung und anschließenden Reduktion von bestimmten Merkmalen kommt. Bei Existenz alternativer Stammbaumhypothesen kann so diejenige ausgewählt werden, die die Evolution der jeweils betrachteten Merkmale oder Daten am plausibelsten erscheinen lässt. 2.7 Statistische Tests 2.7.1 Analyse der Basenzusammensetzung Da der Informationsgehalt von Nukleotidsequenzen auf der Anordnung von nur vier verschiedenen Merkmalen, der vier Nukleotide beruht, entstehen Ähnlichkeiten zwischen Sequenzen sehr oft durch zufällige Übereinstimmungen. Dabei erhöhen Abweichungen vom 1:1:1:1-Verhältnis der Basen die Zahl zufälliger Übereinstimmungen, woraus eine unzutreffende Baumtopologie resultieren kann (Steel et al. 1993). Zur Überprüfung der Homogenität der Basenzusammensetzung wird der in PAUP (Swofford 1999) implementierte X2 – (Chiquadrat-) Test angewendet. 2.7.2 Berechnung der Substitutionssättigung Man spricht von der Sättigung einer Nukleotidsequenz, wenn sich die sichtbare genetische Distanz verschiedener Taxa mit steigender evolutionärer Distanz nicht mehr vergrößert. Das bedeutet, daß durch Mehrfachsubstitutionen die reale Divergenzzeit unterschätzt wird. Zur Korrektur der beobachteten Distanzen sind zahlreiche Modelle entwickelt worden, die bereits in Kapitel 2.2.4.1 vorgestellt worden sind. Da aufgrund von Mehrfachsubstitutionen Konvergenzerscheinungen auftreten können, die die verwandtschaftlichen Beziehungen verfälschen können, sollte ein für phylogenetische Fragestellungen herangezogener Datensatz keine allzu starke Substitutionssättigung aufweisen. Der Sättigungsgrad von Sequenzen kann abgeschätzt werden, indem man die korrigierten Distanzen gegen die beobachteten graphisch aufträgt.

- 53 -
2.7.3 Der relative rate test Wären Distanzen zwischen Organismen perfekt ultrametrisch, d.h. wären die Distanzen der Divergenzzeit direkt proportional, gäbe es genau 1 Topologie auf die sie passen würden. Die Stammbaumrekonstruktion würde dadurch zu einem sehr simplen Vorgang. In der Realität haben Distanzen leider äußerst selten, vielleicht auch niemals die Eigenschaft ultrametrisch zu sein. Um festzustellen, ob die Substitutionsraten bei verschiedenen Taxa voneinander abweichen, kann der relative rate test (Sarich & Wilson 1973) angewendet werden. Dabei werden die Unterschiede in der Anzahl der Substitutionen zwischen zwei nah verwandten Taxa im Vergleich mit einem dritten, phylogenetisch weiter entfernten Taxon festgestellt. S13 und S23 seien die evolutionären Distanzen zwischen Taxon 1 und Taxon 2 einerseits und Außengruppentaxon 3 andererseits. Ist die Differenz von S13 und S23
positiv, dann evolvierte Taxon 1 schneller, ist sie negativ, weist Taxon 2 eine höhere Substitutionsgeschwindigkeit auf. Die Differenz wird als signifikant angesehen, wenn sie größer als die doppelte Standardabweichung ist. Wenn signifikant unterschiedliche Evolutionsraten beobachtet werden, sollten die Ergebnisse der Stammbaumrekonstruktion in Frage gestellt werden. Der Test relativer Substitutionsraten wurde mit dem Programm K2WuLi (Jermiin 1997) durchgeführt. Es werden hier die nach Kimura (1980) für Mehrfachsubstitutionen korrigierten Distanzen verglichen. 2.8 Phylogenetisches Signal Das von Wägele und Rödding (1998) entwickelte Computerprogramm PHYSID (Phylogenetic Signal Detection) erlaubt es, potentielle Apomorphien auf Sequenzebene darzustellen. Die Einschätzung des Informationsgehaltes erfolgt a priori, basierend auf einer Sequenzalinierung, d.h. sie ist unabhängig von einer Baumtopologie oder einem Modell der Sequenzevolution. Grundlage dieser Untersuchung bildet die Tatsache, daß bei nahe verwandten Taxa weniger Substitutionen zu erwarten sind als bei entfernter verwandten Taxa. Mit Hilfe des Programmes kann nach Positionen mit positionsspezifischen, identischen Nukleotiden innerhalb eines hypothetischen Monophylums gesucht und mit denen der korrespondierenden Außengruppe, bestehend aus den verbleibenden Taxa der Alinierung, verglichen werden. Sowohl innerhalb der Innengruppe als auch in der Außengruppe kann ein sogenanntes Rauschen zugelassen werden. In der Außengruppe besteht dieses Rauschen aus zufälligen Übereinstimmungen mit der Innengruppe, in letzterer aus Abweichungen vom Grundmuster. Die potentiellen Apomorphien in Innen- und Außengruppe lassen sich in drei Arten von positionsspezifischen Apomorphien gliedern. Bei den sogenannten symmetrischen Positionen tritt in beiden Gruppen nur jeweils eine Sorte von Nukleotiden auf, z.B.

- 54 -
befindet sich an Position 345 in der Alinierung bei allen Innengruppentaxa ein A, bei allen Außengruppentaxa ein G. Die sogenannten asymmetrischen Positionen sind gekennzeichnet durch das Auftreten von unterschiedlichen Nukleotiden nur in der Außengruppe, wobei aber keine Übereinstimmungen mit der Innengruppe auftreten. Den überwiegenden Anteil der potentiellen Apomorphien bilden sogenannte verrauschte Positionen. In beiden Gruppen dürfen Übereinstimmungen mit der jeweils anderen Gruppe bestehen, allerdings nur bis zu einem festgelegten Prozentsatz. In der vorliegenden Arbeit wurde ein 25%-iges Rauschen in Innen- und Außengruppe zugelassen. Sind die splits mit den meisten potentiellen Apomorphien untereinander und mit den Verzweigungen in anschließend rekonstruierten Stammbäumen kompatibel, kann auf die Existenz eines deutlichen phylogenetischen Signals geschlossen werden (Wägele 1996, Wägele & Rödding 1998).

55
3. Ergebnisse 3.1 Alinierungsergebnisse 18S rDNA (Opisthobranchia-Datensatz) Die Alinierung der 90 kompletten 18S rDNA-Sequenzen umfasste nach manueller Korrektur und Entfernung der primer-Sequenzen 2451 Basenpaare. Drei Nukleotidabschnitte mussten wegen unsicherer Positionshomologie aus der Gesamtalinierung entfernt werden. Das betraf die Positionen 254-310 (Struktur E10_1, Bereich V2), 824-1073 (E23_3, V4) und 1821-2005 (43, V7) (Robakowski 2002). Obwohl im stem-loop-Bereich E10_1 keine ausgeprägte Längenvariabilität zwischen den einzelnen Sequenzen vorlag, war eine eindeutige Homologisierung der Nukleotide aufgrund von zu vielen Alternativen nicht möglich. In der Unterstruktur E23 variierten die Sequenzlängen sehr stark, von 59 Nukleotiden bei Chromodoris krohni bis hin zu 344 Nukleotiden bei Berthella platei. Alle Taxa der Pleurobranchoidea wiesen eine lange Insertion auf in den Bereichen E23_1 und E23_2 auf. Diese neue Unterstruktur ist als E23_3 bezeichnet worden (Wuyts et al. 2000) und beträgt 165-282 Basenpaare. In der Struktur 43 besitzen die Nudibranchia Sequenzlängen von über 100 Nukleotiden, mit einem Maximum bei Bathydoris clavigera (256 Bp). Alle anderen Taxa zeigen Längen zwischen 60 und 70 Nukleotiden. Von den verbleibenden 1959 Positionen der Alinierung erwiesen sich 1005 als konstant, 289 als variabel aber nicht Parsimonie-informativ und 665 als Parsimonie-informativ. 28S rDNA (Opisthobranchia-Datensatz) Die Alinierung der 55 partiellen 28S rDNA-Sequenzen umfasste 1547 Basenpaare. Nicht alinierbare Bereiche entfielen auf die Positionen 491-539, 667-729 und 1113-1398 und konnten im Sekundärstrukturmodell von Saccharomyces cerevisiae (De Rijk et al. 1998) identifiziert werden (491-539: Struktur C1, Bereich V2; 667-729: Struktur C1, Bereich V2; 1113-1398: Struktur D5, Bereich V3). Bei Bathydoris clavigera ist eine sehr lange Insertion im Bereich V3 (Struktur D5) zu beobachten, im Bereich V2 besitzen vor allem die Nudipleura Insertionen. Nach Ausschluß der genannten Bereiche verblieben 1076 Nukleotidpositionen (429 konstant, 140 variabel aber nicht Parsimonie-informativ, 507 Parsimonie-informativ) als Grundlage für die Stammbaumrekonstruktionen. 16S rDNA (Opisthobranchia-Datensatz) Die Alinierung der 45 partiellen 16S rDNA-Sequenzen führte zu einem Datensatz von insgesamt 559 Basenpaaren. Die Positionen 1-60 (Strukturen 66, 67, 68), 252-309 (75, 75’), 334-404 (81’, 84, 88, Abb. 20) wurden wegen unsicherer Homologie nicht

56
berücksichtigt. Im Bereich der Strukturen 66-68, 81’, 84 und 88 konnte eine eindeutige Homologisierung aufgrund zu großer Sequenzdivergenz nicht erfolgen. Im stem-loop-Bereich 75/75’ traten unterschiedlich lange Insertionen bei den Nudipleura, Clione limacina, den Sacoglossa, den Acochlidiacea, Pythia scarabaeus, Odostomia sp. und den Littorinidae auf. Die Identifizierung der Bereiche in denen die genannten Abschnitte lokalisiert sind, erfolgte anhand des Sekundärstrukturmodells von Spodoptera frugiperda (Buckley et al. 2000). Von den 370 verbleibenden Positionen des 16S rDNA-Datensatzes erwiesen sich 92 als konstant, 56 als variabel aber nicht Parsimonie-informativ und 222 als Parsimonie-informativ. 18S rDNA (Anaspidea-Datensatz) Zusätzlich zu den 7 Anaspidea-Sequenzen und den 4 Sequenzen der Thecosomata und Gymnosomata die bereits im Opisthobranchia 18S Datensatz enthalten sind, beinhaltet diese Alinierung 7 weitere Anaspideensequenzen und 2 Außengruppentaxa (Cephalaspidea s.str). Die Alinierung dieser 20 Taxa führte zu einem Datensatz aus 1813 Positionen. Die hohe Konserviertheit des 18S rDNA-Gens und die im Vergleich zu dem Opisthobranchia-Datensatz enge Verwandtschaft der Taxa ermöglichte eine sichere Positionshomologie, so dass keine Bereiche aus der Alinierung entfernt werden mussten. Von den 1813 Positionen des Datensatzes sind 1591 konstant, 118 variabel aber nicht Parsimonie-informativ und 104 Parsimonie-informativ. ITS1-5.8S rDNA-ITS2 (Anaspidea-Datensatz) Die sequenzierten DNA-Bereiche der 11 Arten (9 Arten der Anaspidea, 2 Arten der Cephalaspidea s.str.) zeigten eine extrem hohe Sequenzdivergenz in vielen Bereichen, die die Alinierung außerordentlich erschwerte. Eine weitere Problematik bestand im Auftreten zahlreicher Insertionen bzw. Deletionen einzelner Taxa. Die folgenden Nukleotidpositionen wurden aus der ursprünglich 1707 Basenpaare umfassenden Alinierung entfernt: 31-168, 304-360, 401-465, 493-546, 581-708, 792-966 (ITS1) und 1206-1309, 1594-1682 (ITS2). Von den 899 zur Analyse verbleibenden Positionen erwiesen sich 300 als konstant, 195 als variabel aber nicht Parsimonie-informativ und 404 als Parsimonie-informativ. 18S rDNA (Aeolidoidea-Datensatz) Neben den 6 Arten der Gattung Phyllodesmium beinhaltet der Datensatz 6 weitere Sequenzen aus der Familie der Facelinidae und 13 Arten anderer Aeolidoidea-Familien. Er weist eine Gesamtlänge von 2272 Nukleotiden auf, von denen 236 Positionen als nicht alinierbar unberücksichtigt blieben. Die Positionen 766-901 sind lokalisiert in der Struktur E23_2 (Bereich V4) (Robakowski 2002) und sind gekennzeichnet durch unterschiedlich lange und zum Teil nicht homologisierbare

57
Insertionen bei den Facelinidae und einigen Flabellina-Arten (Flabellinidae). Diese Beobachtung trifft ebenfalls auf die Positionen 1709-1808 (stem-loop-Struktur 43, Bereich V7) zu. Der den Stammbaumanalysen zugrunde liegende Datensatz beinhaltete nach Ausschluß dieser Positionen 2036 Nukleotide (1086 konstant, 203 variabel aber nicht Parsimonie-informativ, 747 Parsimonie-informativ). 18S rDNA, 16S rDNA, COI (Umbraculum spp.) Die 18S rDNA-Sequenzen wiesen eine Länge von jeweils 1789 Nukleotiden auf, die des 16S rRNA-Gens von jeweils 477 Basenpaaren. Da Tylodina perversa für die 16S rDNA-Alinierung nicht als Außengruppe zur Verfügung stand, wurde ein Vertreter der Anaspidea (Aplysia oculifera) gewählt, da molekulare Analysen (Vonnemann et al. 2005, Grande et al. 2004a,b) eine nahe Verwandtschaft der Tylodinoidea zu den Gruppen Anaspidea und Cephalaspidea s.str. angedeutet haben. Die Sequenzen der Cytochromoxidase I (Umbraculum spp. und Tylodina perversa als Außengruppe) zeigten Sequenzlängen von 649-650 Basenpaaren. 3.2 Statistische Tests 3.2.1 Sequenzlängen, Basenzusammensetzung und X2-Test Die in den folgenden Tabellen dargestellten Basenzusammensetzungen basieren auf den reduzierten Datensätzen, d.h. den Alinierungen ohne Berücksichtigung der nicht homologisierbaren Positionen. Zusätzlich sind die Sequenzlängen des kompletten Datensatzes angegeben, auf den sich die berechnete Längenvariabilität bezieht. Der chi-Quadrat-Test wurde auf der Grundlage des reduzierten Datensatzes durchgeführt, da dieser den Stammbaumrekonstruktionen zugrunde liegt. 18S rDNA (Opisthobranchia-Datensatz) Die 18S rDNA-Sequenzen weisen im kompletten Datensatz eine Längenvariabilität von 16,91 % auf. Aufgrund der Insertionen im Bereich E23_3 besitzen die Pleurobranchoidea die größten Sequenzlängen mit einer maximalen Nukleotidanzahl von 2105 bei Tomthompsonia antarctica. Die kürzeste Sequenz ist unter den Außengruppentaxa (Littorinidae) zu finden (Balcis eburnea, 1743 Bp). Die Basenzusammensetzung entspricht einem Verhältnis von 0,24 : 0,24 : 0,28 : 0,24 (A:C:G:T). Die deutlichste Abweichung von diesen Mittelwerten zeigen die Nudibranchia (0,21 : 0,28 : 0,31 : 0,20), unter diesen besonders die drei Arten der Cladobranchia (Dendronotus dalli, Cuthona nana, Flabellina babai). Die aus der Betrachtung der Basenzusammensetzung gewonnenen Eindrücke lassen sich statistisch durch den chi-Quadrat-Test (X2-Test, Tabelle X) belegen:
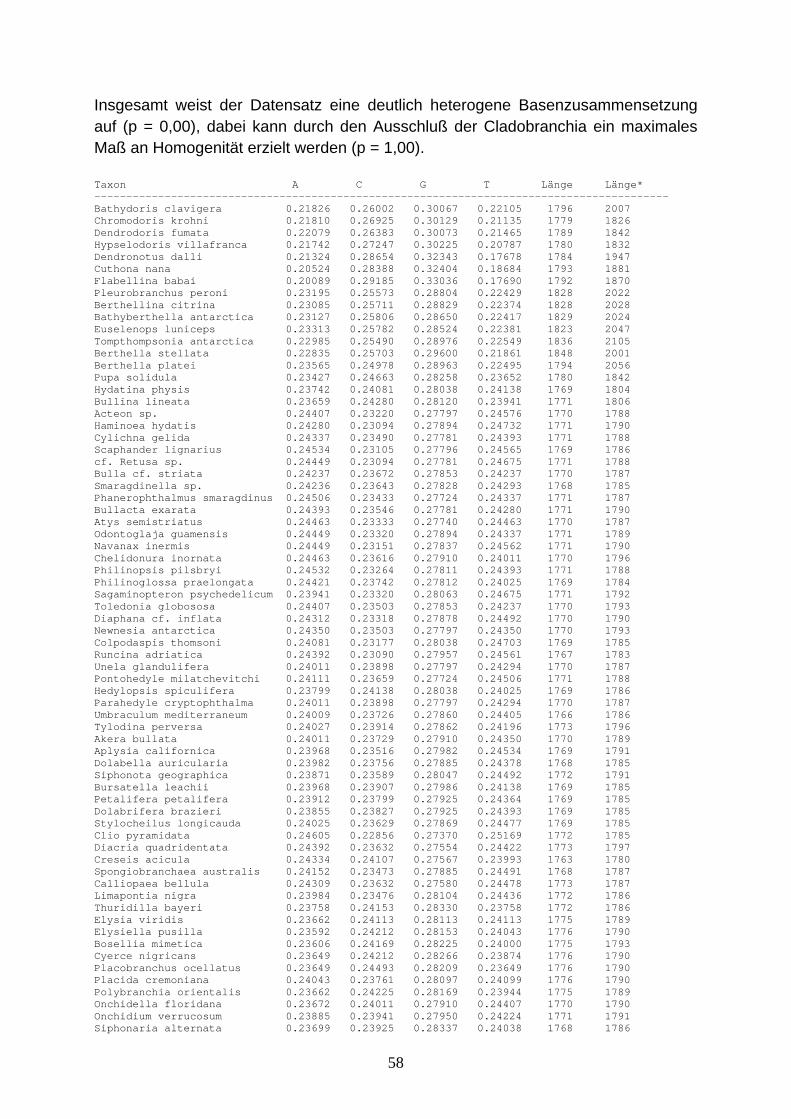
58
Insgesamt weist der Datensatz eine deutlich heterogene Basenzusammensetzung auf (p = 0,00), dabei kann durch den Ausschluß der Cladobranchia ein maximales Maß an Homogenität erzielt werden (p = 1,00). Taxon A C G T Länge Länge* ------------------------------------------------------------------------------------------ Bathydoris clavigera 0.21826 0.26002 0.30067 0.22105 1796 2007 Chromodoris krohni 0.21810 0.26925 0.30129 0.21135 1779 1826 Dendrodoris fumata 0.22079 0.26383 0.30073 0.21465 1789 1842 Hypselodoris villafranca 0.21742 0.27247 0.30225 0.20787 1780 1832 Dendronotus dalli 0.21324 0.28654 0.32343 0.17678 1784 1947 Cuthona nana 0.20524 0.28388 0.32404 0.18684 1793 1881 Flabellina babai 0.20089 0.29185 0.33036 0.17690 1792 1870 Pleurobranchus peroni 0.23195 0.25573 0.28804 0.22429 1828 2022 Berthellina citrina 0.23085 0.25711 0.28829 0.22374 1828 2028 Bathyberthella antarctica 0.23127 0.25806 0.28650 0.22417 1829 2024 Euselenops luniceps 0.23313 0.25782 0.28524 0.22381 1823 2047 Tompthompsonia antarctica 0.22985 0.25490 0.28976 0.22549 1836 2105 Berthella stellata 0.22835 0.25703 0.29600 0.21861 1848 2001 Berthella platei 0.23565 0.24978 0.28963 0.22495 1794 2056 Pupa solidula 0.23427 0.24663 0.28258 0.23652 1780 1842 Hydatina physis 0.23742 0.24081 0.28038 0.24138 1769 1804 Bullina lineata 0.23659 0.24280 0.28120 0.23941 1771 1806 Acteon sp. 0.24407 0.23220 0.27797 0.24576 1770 1788 Haminoea hydatis 0.24280 0.23094 0.27894 0.24732 1771 1790 Cylichna gelida 0.24337 0.23490 0.27781 0.24393 1771 1788 Scaphander lignarius 0.24534 0.23105 0.27796 0.24565 1769 1786 cf. Retusa sp. 0.24449 0.23094 0.27781 0.24675 1771 1788 Bulla cf. striata 0.24237 0.23672 0.27853 0.24237 1770 1787 Smaragdinella sp. 0.24236 0.23643 0.27828 0.24293 1768 1785 Phanerophthalmus smaragdinus 0.24506 0.23433 0.27724 0.24337 1771 1787 Bullacta exarata 0.24393 0.23546 0.27781 0.24280 1771 1790 Atys semistriatus 0.24463 0.23333 0.27740 0.24463 1770 1787 Odontoglaja guamensis 0.24449 0.23320 0.27894 0.24337 1771 1789 Navanax inermis 0.24449 0.23151 0.27837 0.24562 1771 1790 Chelidonura inornata 0.24463 0.23616 0.27910 0.24011 1770 1796 Philinopsis pilsbryi 0.24532 0.23264 0.27811 0.24393 1771 1788 Philinoglossa praelongata 0.24421 0.23742 0.27812 0.24025 1769 1784 Sagaminopteron psychedelicum 0.23941 0.23320 0.28063 0.24675 1771 1792 Toledonia globososa 0.24407 0.23503 0.27853 0.24237 1770 1793 Diaphana cf. inflata 0.24312 0.23318 0.27878 0.24492 1770 1790 Newnesia antarctica 0.24350 0.23503 0.27797 0.24350 1770 1793 Colpodaspis thomsoni 0.24081 0.23177 0.28038 0.24703 1769 1785 Runcina adriatica 0.24392 0.23090 0.27957 0.24561 1767 1783 Unela glandulifera 0.24011 0.23898 0.27797 0.24294 1770 1787 Pontohedyle milatchevitchi 0.24111 0.23659 0.27724 0.24506 1771 1788 Hedylopsis spiculifera 0.23799 0.24138 0.28038 0.24025 1769 1786 Parahedyle cryptophthalma 0.24011 0.23898 0.27797 0.24294 1770 1787 Umbraculum mediterraneum 0.24009 0.23726 0.27860 0.24405 1766 1786 Tylodina perversa 0.24027 0.23914 0.27862 0.24196 1773 1796 Akera bullata 0.24011 0.23729 0.27910 0.24350 1770 1789 Aplysia californica 0.23968 0.23516 0.27982 0.24534 1769 1791 Dolabella auricularia 0.23982 0.23756 0.27885 0.24378 1768 1785 Siphonota geographica 0.23871 0.23589 0.28047 0.24492 1772 1791 Bursatella leachii 0.23968 0.23907 0.27986 0.24138 1769 1785 Petalifera petalifera 0.23912 0.23799 0.27925 0.24364 1769 1785 Dolabrifera brazieri 0.23855 0.23827 0.27925 0.24393 1769 1785 Stylocheilus longicauda 0.24025 0.23629 0.27869 0.24477 1769 1785 Clio pyramidata 0.24605 0.22856 0.27370 0.25169 1772 1785 Diacria quadridentata 0.24392 0.23632 0.27554 0.24422 1773 1797 Creseis acicula 0.24334 0.24107 0.27567 0.23993 1763 1780 Spongiobranchaea australis 0.24152 0.23473 0.27885 0.24491 1768 1787 Calliopaea bellula 0.24309 0.23632 0.27580 0.24478 1773 1787 Limapontia nigra 0.23984 0.23476 0.28104 0.24436 1772 1786 Thuridilla bayeri 0.23758 0.24153 0.28330 0.23758 1772 1786 Elysia viridis 0.23662 0.24113 0.28113 0.24113 1775 1789 Elysiella pusilla 0.23592 0.24212 0.28153 0.24043 1776 1790 Bosellia mimetica 0.23606 0.24169 0.28225 0.24000 1775 1793 Cyerce nigricans 0.23649 0.24212 0.28266 0.23874 1776 1790 Placobranchus ocellatus 0.23649 0.24493 0.28209 0.23649 1776 1790 Placida cremoniana 0.24043 0.23761 0.28097 0.24099 1776 1790 Polybranchia orientalis 0.23662 0.24225 0.28169 0.23944 1775 1789 Onchidella floridana 0.23672 0.24011 0.27910 0.24407 1770 1790 Onchidium verrucosum 0.23885 0.23941 0.27950 0.24224 1771 1791 Siphonaria alternata 0.23699 0.23925 0.28337 0.24038 1768 1786

59
Cepaea hortensis 0.24011 0.23672 0.28079 0.24237 1770 1788 Lymnaea stagnalis 0.24099 0.23367 0.27928 0.24606 1776 1800 Cassidula nuclaeus 0.23869 0.23925 0.28167 0.24038 1768 1786 Melampus luteus 0.23842 0.23955 0.28079 0.24124 1770 1792 Auriculinella bidentata 0.23842 0.23811 0.28053 0.24294 1770 1788 Ovatella firminii 0.23925 0.23925 0.28054 0.24095 1768 1788 Pythia scarabaeus 0.23828 0.23828 0.28176 0.24167 1771 1789 Pyramidella sulcata 0.23885 0.24111 0.28007 0.23998 1771 1791 Odostomia sp. 0.23828 0.23885 0.28007 0.24280 1771 1791 Littoraria undulata 0.24839 0.22715 0.27483 0.24963 1769 1777 Nodilittorina punctata 0.24533 0.23094 0.27359 0.25014 1771 1779 Littorina striata 0.24447 0.22888 0.27647 0.25017 1767 1775 Littorina saxatilis 0.24728 0.23051 0.27373 0.24848 1763 1770 Littorina littorea 0.24530 0.23201 0.27451 0.24818 1767 1775 Melaraphe neritoides 0.24618 0.23145 0.27421 0.24817 1767 1775 Crepidula adunca 0.24411 0.23394 0.27294 0.24902 1761 1765 Fasciolaria lignaria 0.24496 0.22992 0.27183 0.25330 1761 1763 Pisania striata 0.24320 0.23265 0.27468 0.24947 1764 1766 Bursa rana 0.24315 0.23242 0.27440 0.25003 1766 1769 Truncatella guerinii 0.24229 0.23500 0.27482 0.24790 1783 1818 Viviparus georgianus 0.24213 0.22697 0.27921 0.25169 1780 1786 Balcis eburnea 0.24154 0.23321 0.27424 0.25101 1743 1749 --------------------------------------------------------------------------------------- Mittelwert 0.23833 0.24071 0.28200 0.23896 1776 1814
Tabelle 3: Basenzusammensetzung der 18S rDNA-Sequenzen des Opisthobranchia-Datensatzes
(reduzierter Datensatz). In der Spalte Länge* sind die einzelnen Sequenzlängen der kompletten
Alinierung angegeben.
Untersuchte Gruppierung X2 Freiheitsgrade p Alle Taxa 342,83 270 0,00 Nudibranchia 33,55 18 0,01 Alle Taxa, außer Nudibranchia 76,08 249 1,00 Alle Taxa, außer Cladobranchia 148,85 261 1,00 Tabelle 4: X2-Test der 18S rDNA-Sequenzen der Opisthobranchia (reduzierter Datensatz).
28S rDNA (Opisthobranchia-Datensatz) Die Sequenzen der partiellen 28S rDNA weisen eine Längenvariabilität von 25,74 % auf. Die kürzesten Sequenzen sind mit 1027 Bp unter den Außengruppentaxa zu finden (Nodilittorina unifasciata, Tectarius striatus), aber auch innerhalb der Nudibranchia (Cuthona nana). Aufgrund einer Insertion in Bereich V3 (Struktur D5) besitzt Bathydoris clavigera (Nudibranchia) die längste Sequenz (1383 Bp). Die Basenzusammensetzung entspricht im Mittel einem Verhältnis von 0,21 : 0,28 : 0,35 : 0,16 (A:C:G:T). Auffällige Abweichungen von diesen Werten zeigen, genau wie im 18S rDNA-Datensatz, die Taxa der Nudibranchia. Ihre Basenzusammensetzung entspricht 0,19 : 0,31 : 0,37 : 0,13. Der X2-Test (Tabelle X) zeigt die Heterogenität in der Basenzusammensetzung des 28S rDNA-Datensatzes (p = 0,26). Bei Ausschluß der Nudibranchia wird wiederum eine homogene Basenzusammensetzung erzielt (p = 1,00). Im Gegensatz zur 18S rDNA ist innerhalb der Nudibranchia keine Heterogenität der Basenfrequenzen festzustellen (p = 0,94).
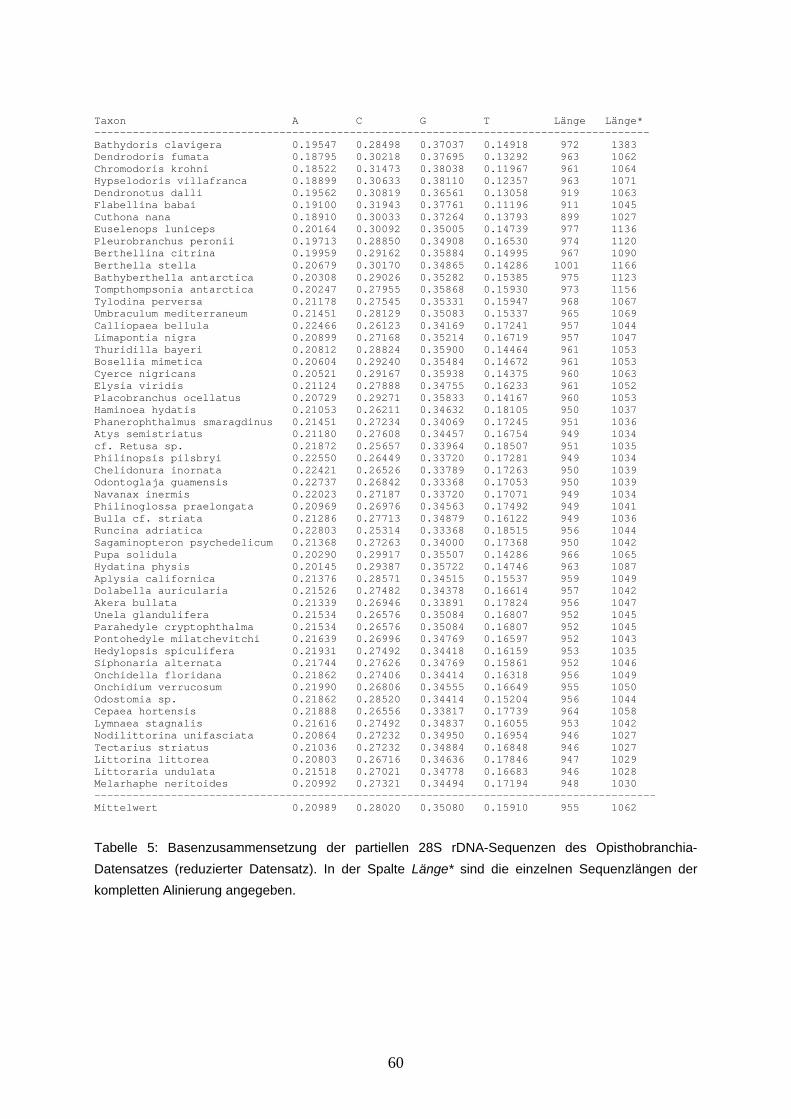
60
Taxon A C G T Länge Länge* --------------------------------------------------------------------------------------- Bathydoris clavigera 0.19547 0.28498 0.37037 0.14918 972 1383 Dendrodoris fumata 0.18795 0.30218 0.37695 0.13292 963 1062 Chromodoris krohni 0.18522 0.31473 0.38038 0.11967 961 1064 Hypselodoris villafranca 0.18899 0.30633 0.38110 0.12357 963 1071 Dendronotus dalli 0.19562 0.30819 0.36561 0.13058 919 1063 Flabellina babai 0.19100 0.31943 0.37761 0.11196 911 1045 Cuthona nana 0.18910 0.30033 0.37264 0.13793 899 1027 Euselenops luniceps 0.20164 0.30092 0.35005 0.14739 977 1136 Pleurobranchus peronii 0.19713 0.28850 0.34908 0.16530 974 1120 Berthellina citrina 0.19959 0.29162 0.35884 0.14995 967 1090 Berthella stella 0.20679 0.30170 0.34865 0.14286 1001 1166 Bathyberthella antarctica 0.20308 0.29026 0.35282 0.15385 975 1123 Tompthompsonia antarctica 0.20247 0.27955 0.35868 0.15930 973 1156 Tylodina perversa 0.21178 0.27545 0.35331 0.15947 968 1067 Umbraculum mediterraneum 0.21451 0.28129 0.35083 0.15337 965 1069 Calliopaea bellula 0.22466 0.26123 0.34169 0.17241 957 1044 Limapontia nigra 0.20899 0.27168 0.35214 0.16719 957 1047 Thuridilla bayeri 0.20812 0.28824 0.35900 0.14464 961 1053 Bosellia mimetica 0.20604 0.29240 0.35484 0.14672 961 1053 Cyerce nigricans 0.20521 0.29167 0.35938 0.14375 960 1063 Elysia viridis 0.21124 0.27888 0.34755 0.16233 961 1052 Placobranchus ocellatus 0.20729 0.29271 0.35833 0.14167 960 1053 Haminoea hydatis 0.21053 0.26211 0.34632 0.18105 950 1037 Phanerophthalmus smaragdinus 0.21451 0.27234 0.34069 0.17245 951 1036 Atys semistriatus 0.21180 0.27608 0.34457 0.16754 949 1034 cf. Retusa sp. 0.21872 0.25657 0.33964 0.18507 951 1035 Philinopsis pilsbryi 0.22550 0.26449 0.33720 0.17281 949 1034 Chelidonura inornata 0.22421 0.26526 0.33789 0.17263 950 1039 Odontoglaja guamensis 0.22737 0.26842 0.33368 0.17053 950 1039 Navanax inermis 0.22023 0.27187 0.33720 0.17071 949 1034 Philinoglossa praelongata 0.20969 0.26976 0.34563 0.17492 949 1041 Bulla cf. striata 0.21286 0.27713 0.34879 0.16122 949 1036 Runcina adriatica 0.22803 0.25314 0.33368 0.18515 956 1044 Sagaminopteron psychedelicum 0.21368 0.27263 0.34000 0.17368 950 1042 Pupa solidula 0.20290 0.29917 0.35507 0.14286 966 1065 Hydatina physis 0.20145 0.29387 0.35722 0.14746 963 1087 Aplysia californica 0.21376 0.28571 0.34515 0.15537 959 1049 Dolabella auricularia 0.21526 0.27482 0.34378 0.16614 957 1042 Akera bullata 0.21339 0.26946 0.33891 0.17824 956 1047 Unela glandulifera 0.21534 0.26576 0.35084 0.16807 952 1045 Parahedyle cryptophthalma 0.21534 0.26576 0.35084 0.16807 952 1045 Pontohedyle milatchevitchi 0.21639 0.26996 0.34769 0.16597 952 1043 Hedylopsis spiculifera 0.21931 0.27492 0.34418 0.16159 953 1035 Siphonaria alternata 0.21744 0.27626 0.34769 0.15861 952 1046 Onchidella floridana 0.21862 0.27406 0.34414 0.16318 956 1049 Onchidium verrucosum 0.21990 0.26806 0.34555 0.16649 955 1050 Odostomia sp. 0.21862 0.28520 0.34414 0.15204 956 1044 Cepaea hortensis 0.21888 0.26556 0.33817 0.17739 964 1058 Lymnaea stagnalis 0.21616 0.27492 0.34837 0.16055 953 1042 Nodilittorina unifasciata 0.20864 0.27232 0.34950 0.16954 946 1027 Tectarius striatus 0.21036 0.27232 0.34884 0.16848 946 1027 Littorina littorea 0.20803 0.26716 0.34636 0.17846 947 1029 Littoraria undulata 0.21518 0.27021 0.34778 0.16683 946 1028 Melarhaphe neritoides 0.20992 0.27321 0.34494 0.17194 948 1030 ---------------------------------------------------------------------------------------- Mittelwert 0.20989 0.28020 0.35080 0.15910 955 1062
Tabelle 5: Basenzusammensetzung der partiellen 28S rDNA-Sequenzen des Opisthobranchia-
Datensatzes (reduzierter Datensatz). In der Spalte Länge* sind die einzelnen Sequenzlängen der
kompletten Alinierung angegeben.
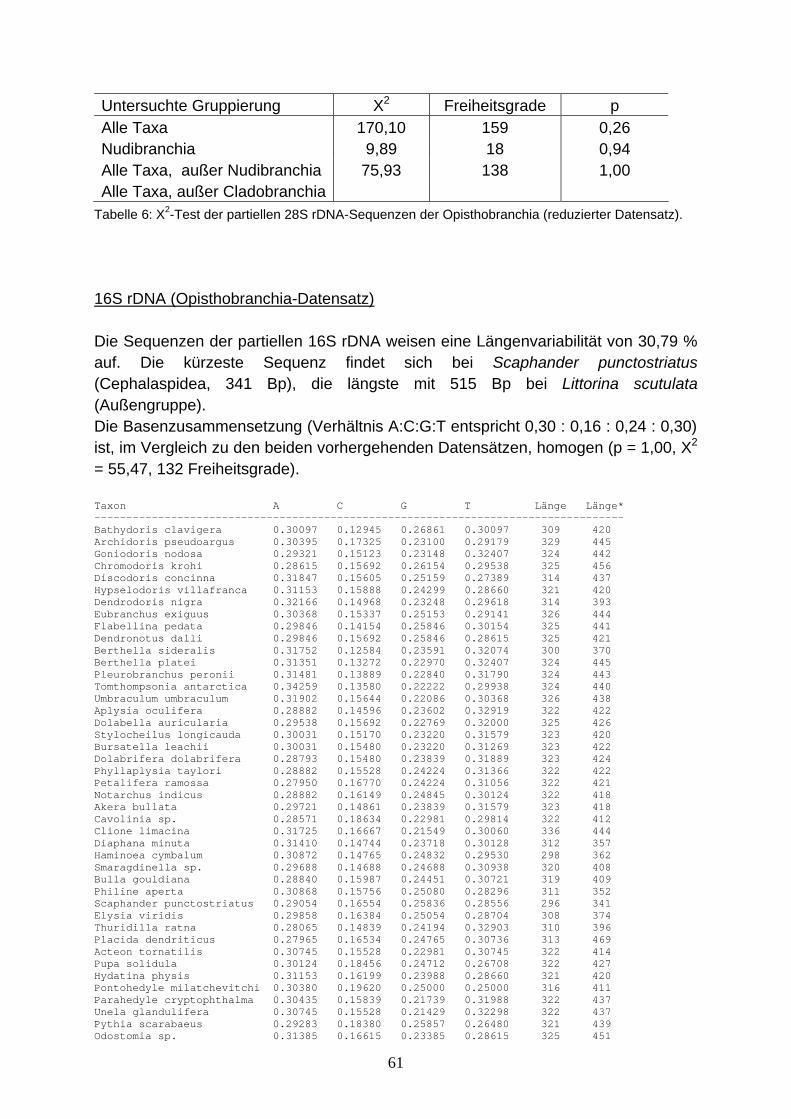
61
Untersuchte Gruppierung X2 Freiheitsgrade p Alle Taxa 170,10 159 0,26 Nudibranchia 9,89 18 0,94 Alle Taxa, außer Nudibranchia 75,93 138 1,00 Alle Taxa, außer Cladobranchia
Tabelle 6: X2-Test der partiellen 28S rDNA-Sequenzen der Opisthobranchia (reduzierter Datensatz).
16S rDNA (Opisthobranchia-Datensatz) Die Sequenzen der partiellen 16S rDNA weisen eine Längenvariabilität von 30,79 % auf. Die kürzeste Sequenz findet sich bei Scaphander punctostriatus (Cephalaspidea, 341 Bp), die längste mit 515 Bp bei Littorina scutulata (Außengruppe). Die Basenzusammensetzung (Verhältnis A:C:G:T entspricht 0,30 : 0,16 : 0,24 : 0,30) ist, im Vergleich zu den beiden vorhergehenden Datensätzen, homogen (p = 1,00, X2 = 55,47, 132 Freiheitsgrade). Taxon A C G T Länge Länge* ----------------------------------------------------------------------------------- Bathydoris clavigera 0.30097 0.12945 0.26861 0.30097 309 420 Archidoris pseudoargus 0.30395 0.17325 0.23100 0.29179 329 445 Goniodoris nodosa 0.29321 0.15123 0.23148 0.32407 324 442 Chromodoris krohi 0.28615 0.15692 0.26154 0.29538 325 456 Discodoris concinna 0.31847 0.15605 0.25159 0.27389 314 437 Hypselodoris villafranca 0.31153 0.15888 0.24299 0.28660 321 420 Dendrodoris nigra 0.32166 0.14968 0.23248 0.29618 314 393 Eubranchus exiguus 0.30368 0.15337 0.25153 0.29141 326 444 Flabellina pedata 0.29846 0.14154 0.25846 0.30154 325 441 Dendronotus dalli 0.29846 0.15692 0.25846 0.28615 325 421 Berthella sideralis 0.31752 0.12584 0.23591 0.32074 300 370 Berthella platei 0.31351 0.13272 0.22970 0.32407 324 445 Pleurobranchus peronii 0.31481 0.13889 0.22840 0.31790 324 443 Tomthompsonia antarctica 0.34259 0.13580 0.22222 0.29938 324 440 Umbraculum umbraculum 0.31902 0.15644 0.22086 0.30368 326 438 Aplysia oculifera 0.28882 0.14596 0.23602 0.32919 322 422 Dolabella auricularia 0.29538 0.15692 0.22769 0.32000 325 426 Stylocheilus longicauda 0.30031 0.15170 0.23220 0.31579 323 420 Bursatella leachii 0.30031 0.15480 0.23220 0.31269 323 422 Dolabrifera dolabrifera 0.28793 0.15480 0.23839 0.31889 323 424 Phyllaplysia taylori 0.28882 0.15528 0.24224 0.31366 322 422 Petalifera ramossa 0.27950 0.16770 0.24224 0.31056 322 421 Notarchus indicus 0.28882 0.16149 0.24845 0.30124 322 418 Akera bullata 0.29721 0.14861 0.23839 0.31579 323 418 Cavolinia sp. 0.28571 0.18634 0.22981 0.29814 322 412 Clione limacina 0.31725 0.16667 0.21549 0.30060 336 444 Diaphana minuta 0.31410 0.14744 0.23718 0.30128 312 357 Haminoea cymbalum 0.30872 0.14765 0.24832 0.29530 298 362 Smaragdinella sp. 0.29688 0.14688 0.24688 0.30938 320 408 Bulla gouldiana 0.28840 0.15987 0.24451 0.30721 319 409 Philine aperta 0.30868 0.15756 0.25080 0.28296 311 352 Scaphander punctostriatus 0.29054 0.16554 0.25836 0.28556 296 341 Elysia viridis 0.29858 0.16384 0.25054 0.28704 308 374 Thuridilla ratna 0.28065 0.14839 0.24194 0.32903 310 396 Placida dendriticus 0.27965 0.16534 0.24765 0.30736 313 469 Acteon tornatilis 0.30745 0.15528 0.22981 0.30745 322 414 Pupa solidula 0.30124 0.18456 0.24712 0.26708 322 427 Hydatina physis 0.31153 0.16199 0.23988 0.28660 321 420 Pontohedyle milatchevitchi 0.30380 0.19620 0.25000 0.25000 316 411 Parahedyle cryptophthalma 0.30435 0.15839 0.21739 0.31988 322 437 Unela glandulifera 0.30745 0.15528 0.21429 0.32298 322 437 Pythia scarabaeus 0.29283 0.18380 0.25857 0.26480 321 439 Odostomia sp. 0.31385 0.16615 0.23385 0.28615 325 451

62
Littorina scutulata 0.31034 0.15517 0.21839 0.31609 348 515 Tectarius viviparius 0.32808 0.18927 0.23028 0.25237 317 427 ------------------------------------------------------------------------------------ Mittelwert 0.30272 0.15729 0.23926 0.30073 320 419
Tabelle 7: Basenzusammensetzung der partiellen 16S rDNA-Sequenzen des Opisthobranchia-
Datensatzes (reduzierter Datensatz). In der Spalte Länge* sind die einzelnen Sequenzlängen der
kompletten Alinierung angegeben.
18S rDNA (Anaspidea-Datensatz) Aufgrund der geringen Sequenzdifferenzen innerhalb des Datensatzes gab es keine Bereiche mit unsicherer Positionshomologie, die Längenvariabilität ist entsprechend gering (0,67 %). Das Frequenzverhältnis A:C:G:T entspricht 0,24 : 0,24 : 0,28 : 0, 24, die Basen- zusammensetzung ist homogen (p = 1,00, X2 = 3,25, 57 Freiheitsgrade). Taxon A C G T Länge ----------------------------------------------------------------------------- Akera bullata 0.23868 0.23756 0.28060 0.24315 1789 Aplysia gigantea 0.23618 0.23618 0.28141 0.24623 1791 Aplysia oculifera 0.23618 0.23618 0.28141 0.24623 1791 Aplysia sydneyensis 0.23687 0.23575 0.28156 0.24581 1790 Aplysia parvula 0.23605 0.23717 0.28181 0.24498 1792 Aplysia juliana 0.23618 0.23730 0.28197 0.24456 1791 Aplysia dactylomela 0.23618 0.23674 0.28141 0.24567 1791 Siphonota geographica 0.23618 0.23645 0.28141 0.24596 1791 Bursatella leachii 0.23754 0.23973 0.28072 0.24202 1785 Dolabella auricularia 0.23754 0.23754 0.28011 0.24482 1785 Petalifera petalifera 0.23697 0.23810 0.28011 0.24482 1785 Dolabrifera brazieri 0.23641 0.23837 0.28011 0.24510 1785 Stylocheilus longicauda 0.23810 0.23641 0.27955 0.24594 1785 Dolabrifera dolabrifera 0.23613 0.23838 0.28011 0.24538 1785 Creseis acicula 0.24270 0.24213 0.27584 0.23933 1780 Clio pyramidata 0.24426 0.22913 0.27507 0.25154 1785 Diacria quadridentata 0.24122 0.23706 0.27743 0.24430 1797 Spongiobranchaea australis 0.24105 0.23602 0.27908 0.24385 1788 Atys semistriatus 0.24231 0.23335 0.27924 0.24510 1787 Chelidonura inornata 0.24165 0.23775 0.28062 0.23998 1796 ----------------------------------------------------------------------------- Mittelwert 0.23842 0.23686 0.27998 0.24474 1788 Tabelle 8: Basenzusammensetzung der 18S rDNA-Sequenzen des Anaspidea-Datensatzes.
ITS1 - 5.8S rDNA - ITS2 (Anaspidea-Datensatz) Die Längenvariabilität der Sequenzen dieses Datensatzes beträgt 30,52 %. Diese hohe Variabilität lässt sich im Wesentlichen auf die hohe Sequenzlänge von Bursatella leachii (1350 Bp) und die geringe von Chelidonura inornata (938 Bp) zurückführen, bei Nichtberücksichtigung dieser beiden Taxa beträgt sie nur noch 13,36 %. Bei der Betrachtung der Basenzusammensetzung entsteht der Eindruck einer Heterogenität in der Basenverteilung. Dieser Eindruck kann durch die Ergebnisse des X2-Tests bestätigt werden, der eine deutliche Variabilität der Basenverteilung anzeigt (p = 0,24, X2 = 35,05, 30 Freiheitsgrade).

63
Taxon A C G T Länge Länge* ----------------------------------------------------------------------------------- Bursatella leachii 0.17097 0.32226 0.31734 0.18942 813 1350 Petalifera petalifera 0.15276 0.33203 0.32380 0.19141 768 1082 Dolabrifera brazieri 0.14304 0.31139 0.32911 0.21646 790 1087 Dolabrifera dolabrifera 0.14518 0.31665 0.32040 0.21777 799 1113 Dolabella auricularia 0.15274 0.32715 0.32408 0.19603 755 1110 Aplysia juliana 0.16757 0.32703 0.32162 0.18378 740 1169 Aplysia gigantea 0.17158 0.32708 0.33110 0.17024 746 1198 Aplysia oculifera 0.18355 0.31424 0.32452 0.17768 681 1107 Akera bullata 0.16598 0.29355 0.32099 0.21948 729 1198 Atys semistriatus 0.18947 0.28872 0.31579 0.20602 665 1038 Chelidonura inornata 0.18059 0.27685 0.30352 0.23903 661 938 ----------------------------------------------------------------------------------- Mittelwert 0.16497 0.31324 0.32134 0.20044 741 1126
Tabelle 9: Basenzusammensetzung der ITS1-5.8S rDNA-ITS2 - Sequenzen des Anaspidea-
Datensatzes. In der Spalte Länge* sind die einzelnen Sequenzlängen der kompletten Alinierung
angegeben.
18S rDNA (Aeolidoidea-Datensatz) Die Sequenzlängen der 18S rDNA weisen innerhalb der Aeolidoidea Unterschiede von 8,65 % auf, dabei ist das 18S rRNA-Gen bei den Facelinidae (Baeolidia nodosa, Godiva banyulensis, Cratena peregrina, Phidiana lynceus, Facelina spp., Phyllodesmium spp.) mit durchschnittlich 1995 Basenpaaren deutlich länger als bei den übrigen Aeolidoidea (durchschnittlich 1899 Bp, Ausnahme: Flabellina affinis). Die Basenzusammensetzung innerhalb des Datensatzes ist homogen (p = 1,00, X2 = 39,81, 72 Freiheitsgrade). Taxon A C G T Länge Länge* ----------------------------------------------------------------------------------- Baeolidia nodosa 0.19280 0.27658 0.34264 0.18797 1862 2025 Godiva banyulensis 0.19685 0.27461 0.33931 0.18923 1839 2046 Cratena peregrina 0.19348 0.27935 0.33967 0.18750 1840 2002 Phidiana lynceus 0.19283 0.27920 0.34003 0.18794 1841 2011 Facelina bostoniensis 0.19694 0.27660 0.33715 0.18931 1833 1982 Facelina drummondi 0.19694 0.27660 0.33715 0.18931 1833 1981 Phyllodesmium briareum 0.19597 0.27599 0.33805 0.18998 1837 1988 Phyllodesmium crypticum 0.19478 0.27530 0.33841 0.19151 1838 1990 Phyllodesmium colemani 0.19597 0.27599 0.33805 0.18998 1837 1988 Phyllodesmium longicirrum 0.19563 0.27545 0.33840 0.19053 1837 1988 Phyllodesmium jakobsenae 0.19408 0.27657 0.33695 0.19240 1795 1946 Phyllodesmium cf. magnum 0.19321 0.27880 0.33948 0.18852 1835 1989 Embletonia sp. 0.20142 0.29221 0.32655 0.17982 1824 1898 Cuthona nana 0.20184 0.28649 0.32284 0.18882 1843 1880 Cuthona caerulea 0.20022 0.28486 0.32067 0.19425 1843 1888 Cuthona sibogae 0.20098 0.28952 0.32374 0.18577 1841 1878 Eubranchus exiguus 0.20305 0.29287 0.32118 0.18291 1837 1876 Eubranchus spec. 0.20206 0.29169 0.32482 0.18142 1841 1879 Tergipes tergipes 0.20596 0.28347 0.31924 0.19133 1845 1887 Flabellina babai 0.19772 0.29441 0.32863 0.17925 1841 1869 Flabellina pedata 0.20513 0.28369 0.32079 0.19040 1833 1870 Flabellina verrucosa 0.20351 0.29183 0.32200 0.18267 1823 1907 Flabellina sp. 0.21087 0.27488 0.31415 0.20011 1859 1938 Flabellina affinis 0.20299 0.27350 0.31250 0.21100 1872 2018 Notaeolidia depressa 0.19608 0.30338 0.32952 0.17102 1836 1901 ------------------------------------------------------------------------------------ Mittelwert 0.19886 0.28254 0.33006 0.18854 1839 1945
Tabelle 10: Basenzusammensetzung der 18S rDNA-Sequenzen des Aeolidoidea-Datensatzes. In der
Spalte Länge* sind die einzelnen Sequenzlängen der kompletten Alinierung angegeben.

64
3.2.2 Berechnung der Substitutionssättigung Für die graphische Darstellung der Substitutionssättigung wurden die korrigierten Sequenzdistanzen gegen die absoluten Distanzen aufgetragen. Die der Korrektur zugrunde liegenden Modelle der Sequenzevolution wurden für jeden Datensatz unabhängig mit Hilfe hierarchischer likelihood ratio tests (Modeltest Version 3.06, Posada & Crandall 1998) ermittelt. 18S rDNA (Opisthobranchia-Datensatz) Die Graphik in Abb. 9 zeigt, dass die absolute Anzahl der Transitionen deutlich größer ist als die der Transitionen, besonders im Bereich der höchsten Sequenzdivergenzen. Während die Anzahl der Transitionen mit steigender Distanz linear zunimmt, ist bei den Transversionen ein Sättigungseffekt zu beobachten. In der Graphik können unterschiedliche Regionen identifiziert werden, die mit den Zahlen 1 bis 5 gekennzeichnet sind. Die höchste Sequenzdivergenz (1) zeigen die drei Arten der Cladobranchia (Nudibranchia) im Vergleich zu Berthella stellata (Pleurobranchoidea). Bereich 2 repräsentiert die Sequenzdistanzen zwischen den Cladobranchia und den Außengruppentaxa (Littorinidae), Bereich 3 diejenigen zwischen den Cladobranchia und den übrigen Innengruppentaxa. Die beiden mit 4 gekennzeichneten Punktwolken repräsentieren korrigierte Distanzen (k.D.) von Berthella stellata zu den Littorinidae einerseits (k.D. = 0,42-0,45) und zu den verbleibenden Taxa andererseits (k.D. = 0,33-0,39). Der Distanzvergleich aller übrigen Taxa des Datensatzes ergab den mit 5 markierten Abschnitt des Punktdiagramms. 28S rDNA (Opisthobranchia-Datensatz) Im Gegensatz zur 18S rDNA wird bei der graphischen Darstellung der Substitutionssättigung der 28S rDNA-Sequenzen (Abb. 10) deutlich, dass beide Substitutionsklassen mit zunehmender korrigierter Distanz eine leichte Sättigung aufweisen. Dabei ist die Anzahl beobachteter Transitionen geringfügig höher als die der Transversionen. Eine deutliche Gliederung in mehrere Abschnitte, wie in Abb. 9, ist hier nicht zu beobachten. Es können nur zwei Regionen unterschieden werden, repräsentiert durch Sequenzdistanzen der Cladobranchia, sowie von Berthella stellata zu allen übrigen Taxa einerseits (1) und der übrigen Taxa untereinander andererseits (2). 16S rDNA (Opisthobranchia-Datensatz) Im Vergleich zur 18S und 28S rDNA ist anhand der graphischen Darstellung der korrigierten und absoluten Distanzen des 16S rDNA-Gens (Abb. 11), eine größere Anzahl von Transversionen als von Transitionen festzustellen. Die Transitionen weisen dabei eine stärkere Sättigung auf als die Transversionen.

65
Die größte Sequenzdistanz repräsentiert den Vergleich zwischen Odostomia sp. (Pyramidellidae) und Bulla gouldiana (Cephalaspidea s.str.), die höchste Anzahl von Transversionen resultiert aus der Gegenüberstellung von Cavolinia sp. (Thecosomata) und verschiedenen anderen Taxa. 18S rDNA (Anaspidea-Datensatz) Die graphische Darstellung der korrigierten und absoluten Sequenzdistanzen der Taxa des Anaspidea-Datensatzes (Abb. 12) zeigt keine Sättigungserscheinung der 18S rDNA, d.h. die Anzahl der Transitionen und Transversionen nimmt mit steigender Distanz linear zu. Die im Vergleich zum Opisthobranchia-Datensatz sehr geringen Sequenzdistanzen resultieren aus der viel näheren Verwandtschaft der untersuchten Taxa. Die größte Sequenzdivergenz (1) weisen die beiden Vertreter der Außengruppe (Cephalaspidea s.str.) im Vergleich zu Creseis acicula (Thecosomata) auf, Bereich 2 repräsentiert die Gegenüberstellung von Creseis acicula und den übrigen Arten des Datensatzes. Die mit 3 gekennzeichneten Punkte stellen die Sequenzdistanz zwischen den Außengruppentaxa und den übrigen Vertretern der Thecosomata dar, Bereich 4 ergibt sich aus dem Vergleich aller restlichen Sequenzen. ITS1 - 5.8S rDNA - ITS2 (Anaspidea-Datensatz) Wie bei der 16S rDNA ist im ITS1-5.8S rDNA-ITS2 - Datensatz eine größere Anzahl Transversionen als Transitionen festzustellen, dabei weisen die Transitionen eine leichte Sättigung auf (Abb. 13). Insgesamt zeigt der untersuchte DNA-Bereich wesentliche höhere Sequenzdivergenzen als das 18S rDNA-Gen derselben Taxa. Die beiden größten Sequenzdistanzen repräsentieren den Vergleich der zwei Außengruppentaxa mit Bursatella leachii (Anaspidea). 18S rDNA (Aeolidoidea-Datensatz) Aufgrund der sehr hohen Distanzwerte (siehe Anhang 5.4) zwischen den Facelinidae einerseits und den restlichen Aeolidoidea andererseits konnten für die graphische Darstellung der Substitutionssättigung nur die Distanzen innerhalb der beiden Gruppen berücksichtigt werden. Die Ergebnisse (Abb. 14) entsprechen denen des Anaspidea-Datensatzes (18S rDNA), d.h. es sind mehr Transitionen als Transversionen zu beobachten und beide Substitutionsklassen zeigen keine Sättigungserscheinung.

66
Substitutionssättigung 18S rDNA
0
50
100
150
200
250
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
korrigierte Distanzen
abso
lute
Dis
tan
zen
Transitionen
Transversionen
Substitutionssättigung 28S rDNA
0
20
40
60
80
100
120
140
160
0 0,2 0,4 0,6 0,8 1
korrigierte Distanzen
abso
lute
Dis
tan
zen
Transitionen
Transversionen
Abb. 9: Substitutionssättigung der 18S rDNA-Sequenzen (Opisthobranchia-Datensatz)
Abb. 10: Substitutionssättigung der 28S rDNA-Sequenzen (Opisthobranchia-Datensatz)
1
2
2 3
3
4 4
5
1
2

67
Substitutionssättigung 16S rDNA
0
10
20
30
40
50
60
70
0 0,2 0,4 0,6 0,8 1 1,2
korrigierte Distanzen
abso
lute
Dis
tan
zen
Transitionen
Transversionen
Substitutionssättigung 18S rDNA (Anaspidea)
0
10
20
30
40
50
60
70
80
0 0,02 0,04 0,06 0,08 0,1 0,12
korrigierte Distanzen
abso
lute
Dis
tan
zen
Transitionen
Transversionen
Abb. 11: Substitutionssättigung der 16S rDNA-Sequenzen (Opisthobranchia-Datensatz)
Abb. 12: Substitutionssättigung der 18S rDNA-Sequenzen (Anaspidea-Datensatz)
1
2
3
4

68
Substitutionssättigung 18S rDNA (Aeolidoidea)
0
20
40
60
80
100
120
0 0,05 0,1 0,15 0,2 0,25 0,3
korrigierte Distanzen
abso
lute
Dis
tan
zen
Transitionen
Transversionen
Substitutionssättigung ITS1-5.8S-ITS2 (Anaspidea)
0
20
40
60
80
100
120
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7
korrigierte Distanzen
abso
lute
Dis
tan
zen
Transitionen
Transversionen
Abb. 13: Substitutionssättigung der ITS1-5.8S-ITS2 – Sequenzen (Anaspidea-Datensatz)
Abb. 14: Substitutionssättigung der 18S rDNA-Sequenzen (Aeolidoidea-Datensatz)

69
3.2.3 relative rate-Test 18S rDNA (Opisthobranchia-Datensatz) Für die korrekte Einschätzung der relativen Substitutionsraten innerhalb eines Datensatzes ist es wichtig, dass das gewählte Referenztaxon eine möglichst geringe Distanz zur untersuchten Innengruppe aufweist, da besonders die evolutive Distanz zur Außengruppe durch Sättigungseffekte verfälscht wird. Je näher die Außengruppe mit der Innengruppe verwandt ist, desto höher ist also die Wahrscheinlichkeit, dass Unterschiede in den relativen Raten korrekt eingeschätzt, bzw. überhaupt erkannt werden können (Tajima 1993, Robinson et al. 1998) Unter Berücksichtigung dessen wurden keine Außengruppentaxa der phylogenetischen Analysen (Littorinidae) als Referenztaxon verwendet, sondern ein Vetreter der basalen Heterobranchia (Odostomia sp., Pyramidellidae). Die Sequenzen der Pulmonata wurden nicht berücksichtigt. Die höchsten Evolutionsraten innerhalb des 18S rDNA-Datensatzes sind bei den Cladobranchia (Nudibranchia) zu beobachten. Sie weisen Z-Werte von 2,32 bis 15,48 auf und evolvieren damit schneller als alle übrigen untersuchten Taxa. Die Anthobranchia (Nudibranchia) zeigen erhöhte Raten (1,96 - 10,10) gegenüber allen anderen Taxa, exklusive der Cladobranchia und einiger Vertreter der Pleurobranchoidea. Die Pleurobranchoidea insgesamt zeigen eine höhere Substitutionsgeschwindigkeit als die übrigen Gruppen, mit Ausnahme der Nudibranchia. Innerhalb der Pleurobranchoidea ist die relative Evolutionsrate von Berthella stellata auffällig, die fast die Werte der Cladobranchia erreicht. Abgesehen von den Nudipleura zeigen die Acteonoidea eine erhöhte Substitutionsgeschwindigkeit, das gilt allerdings nicht für Acteon sp. Unter den verbleibenden Taxa der Opisthobranchia besitzen die Thecosomata die höchsten Z-Werte. Am langsamsten evolvierten die Anaspidea und Acochlidiacea (außer Pontohedyle milatchevitchi). Auffällig ist, dass die Substitutionsraten gruppenspezifisch, d.h. innerhalb der einzelnen Subtaxa der Opisthobranchia relativ homogen sind. Ausnahmen bilden z.B. Colpodaspis thomsoni (Cephalaspidea s.str.) und Calliopaea bellula (Sacoglossa). Die folgende Zeile gibt in vereinfachter Darstellung die Rangfolge der Evolutionsgeschwindigkeiten innerhalb des Datensatzes wieder: Cladobranchia (Nudibranchia) > Anthobranchia (Nudibranchia) > Pleurobranchoidea > Acteonoidea > Thecosomata > restliche Taxa

70
28S rDNA (Opisthobranchia-Datensatz) Wie bei der 18S rDNA wurde Odostomia sp. als Außengruppetaxon verwendet und die „Prosobranchia“ nicht berücksichtigt. Innerhalb des 28S rDNA-Datensatzes sind, genau wie im 18S rDNA-Datensatz, die höchsten Evolutionsraten bei den Cladobranchia (Nudibranchia) zu verzeichnen (Z-Werte von 2,43 – 10,63), sie weisen signifikant erhöhte Raten gegenüber allen anderen Opisthobranchiern auf. An zweiter Stelle stehen die Anthobranchia (Nudibranchia), mit Evolutionsraten von Z = 1,97 – 6,72; nur gegenüber einigen Pleurobranchoida zeigen sie keine erhöhte Evolutionsgeschwindigkeit. Die Pleurobranchoida weisen ebenfalls erhöhte Raten auf (1,97 – 9,22), mit auffallend hohen Werten bei Berthella stellata. Die Z-Werte dieser Art sind deutlich größer als die der übrigen Pleurobranchoidea und übertreffen sogar die der Anthobranchia. Unter den verbleibenden Taxa der Opisthobranchia besitzen die Acteonoidea die höchsten Z-Werte. Wie beim 18S rDNA-Datensatz ist unten in vereinfachter Weise die Höhe der relativen Substitutionsgeschwindigkeit innerhalb der Opisthobranchia angegeben. Cladobranchia (Nudibranchia) > Anthobranchia (Nudibranchia) > Pleurobranchoidea > Acteonoidea > übrige Taxa 16S rDNA (Opisthobranchia-Datensatz) Wie in den beiden vorhergehenden Datensätzen wurde Odostomia sp. als Referenzart zur Untersuchung der relativen Substitutionsgeschwindigkeit innerhalb der Opisthobranchia verwendet. Die Verteilung der Evolutionsraten im 16S rDNA-Datensatz ist im Vergleich zur 18S und 28S rDNA relativ homogen. Nur Clione limacina (Gymnosomata) weist eine höhere Evolutionsgeschwindigkeit als die meisten übrigen Taxa auf. Gegenüber den Sacoglossa, Hydatina physis (Acteonoidea) und den Acochlidiacea (ohne Pontohedyle milatchevitchi) zeigen die meisten Taxa signifikant erhöhte Substitutionsraten. 18S rDNA (Anaspidea-Datensatz) Zur Berechnung der relativen Substitutionsraten wurde Atys semistriatus (Cephalaspidea s.str.) als Referenzart verwendet, da dieses Taxon zur Innengruppe (Anaspidea, Thecosomata, Gymnosomata) eine geringere genetische Distanz aufweist als das andere Außengruppentaxon Chelidonura inornata (Cephalaspidea s.str.). Signifikant erhöhte Evolutionsraten sind bei den thecosomaten Taxa Creseis acicula, Diacria quadridentata und Clio pyramidata zu beobachten. Die höchsten Z-Werte innerhalb des Datensatzes zeigt Creseis acicula (3,86 - 7,17), dabei weist diese Art sogar eine signifikant höhere Substitutionsgeschwindigkeit als die beiden anderen

71
Arten der Thecosomata auf. Spongiobranchaea australis (Gymnosomata) evolviert im Vergleich zu einigen Arten ebenfalls schneller. Unter den übrigen Taxa ist eine im Wesentlichen homogene Verteilung der Evolutionsgeschwindigkeiten zu beobachten. ITS1 - 5.8S rDNA - ITS2 (Anaspidea-Datensatz) Wie beim Anaspidea-Datensatz auf der Grundlage der 18S rDNA, wurde auch hier Atys semistriatus (Cephalaspidea s.str.) als Referenztaxon verwendet. Alle untersuchten Taxa zeigen gegenüber Akera bullata eine erhöhte Substitutionsgeschwindigkeit, der höchster Z-Wert tritt beim Vergleich von Bursatella leachii mit Akera bullata auf (3,95). Bursatella leachii weist gegenüber den meisten übrigen Arten ebenfalls eine höhere Evolutionsgeschwindigkeit auf. Die Art Petalifera petalifera evolviert ebenfalls schneller als einige andere Arten, abgesehen davon ist die Verteilung der Substitutionsraten homogen. 18S rDNA (Aeolidoidea-Datensatz) Innerhalb des Aeolidoidea-Datensatzes wurde die als ursprünglich angesehene Art Notaeolidia depressa als Referenzart verwendet, alle übrigen Taxa bildeten die zu untersuchende Innengruppe. Die Arten der Facelinidae (Baeolidia nodosa, Godiva banyulensis, Cratena peregrina, Phidiana lynceus, Facelina spp., Phyllodesmium spp.) zeigen gegenüber den restlichen Aeolidoidea eine signifikant erhöhte Substitutionsgeschwindigkeit (Z-Werte von 12,1 bis 16,6), innerhalb der Familie ist die Ratenverteilung homogen. Unter den übrigen Aeolidoidea weisen die Mitglieder der Flabellinidae (Flabellina spp.) und Tergipes tergipes erhöhte Raten gegenüber den restlichen Taxa auf. 3.3 Phylogenetisches Signal Mit Hilfe des Programmes PHYSID (Wägele & Rödding 1998) wurde die Anzahl der potentiellen Apomorphien für hypothetische Monophyla in jedem Datensatz festgestellt. Die Ergebnisse der Analyse sind in den Tabellen 11 und 12 dargestellt. Aufgrund von in der Vergangenheit aufgetretenen Unzulänglichkeiten wurden nicht alle Funktionen des Programmes genutzt. Da nicht sichergestellt werden konnte, dass tatsächlich alle existierenden splits innerhalb der Datensätze ermittelt werden können, wurde PHYSID nur angewendet, um gezielt nach der Stützung bestimmter Gruppen zu suchen. Auf die Berechnung von Spektren wurde verzichtet.

72
Opisthobranchia-Datensätze (18S rDNA, 28S rDNA, 16S rDNA) Tabelle 11 zeigt, dass die Datensätze der Opisthobranchia alle relativ verrauscht sind, da kaum symmetrische und asymmetrische stützende Positionen für die hypothetischen Monophyla vorhanden sind. Die Anzahl der potentiellen Apomorphien variiert erwartungsgemäß sowohl zwischen den Gruppierungen eines Gens, als auch bei Vergleich der verschiedenen Datensätze, dabei zeigt der 18S rDNA-Datensatz die höchste Variabilität. Die in den18S und 28S rDNA-Alinierungen am besten gestützten Gruppierungen sind die Nudibranchia, Pleurobranchoidea und Nudipleura, sowie die Opisthobranchia als Gesamtheit (nur 18S rDNA). Im 16S rDNA-Datensatz weisen die Opisthobranchia die höchste Anzahl potentieller Apomorphien auf. Die Anaspidea und Acochlidiacea zeigen mit jeweils 3 potentiellen Apomorphien die geringste Anzahl stützender Positionen (18S rDNA). Um Aussagen über die Stärke des phylogenetischen Signals der verschiedenen Sequenzalinierungen treffen zu können, muss die Anzahl der potentiellen Apomorphien der Taxa mit der Sequenzlänge des jeweiligen Datensatzes in Beziehung gebracht werden. Dabei zeigt sich, dass der untersuchte Abschnitt der 16S rDNA nach quantitativen Maßstäben das stärkste phylogenetische Signal aufweist. Im 16S rDNA-Datensatz entsprechen z.B. 50 stützende Positionen etwa 14 % der Gesamtalinierung, bei der 28S rDNA sind das nur etwa 5 % aller Positionen, bei der 18S rDNA sogar nur rund 3 %.
18S rDNA 28S rDNA 16S rDNA Hypothetisches Monophylum Anzahl potentieller Apomorphien
Gesamtzahl (verrauschte/symm./asymm. Positionen) Opisthobranchia 60 (60/0/0) 23 (23/0/0) 51 (51/0/0) Nudibranchia 70 (61/8/1) 77 (70/4/3) 25 (25/0/0) Pleurobranchoidea 75 (72/1/2) 69 (65/2/2) 32 (32/0/0) Nudipleura 49 (40/8/1) 69 (66/2/1) 23 (23/0/0) Tylodinoidea 9 (7/0/2) 26 (25/0/1) * Anaspidea 3 (3/0/0) 29 (27/0/2) 38 (38/0/0) Cephalaspidea s.str. 10 (10/0/0) 35 (35/0/0) 24 (24/0/0) Anaspidea + Cephalaspidea s.str. 12 (12/0/0) 38 (38/0/0) 27 (27/0/0) Acteonoidea 28 (28/0/0) 35 (28/2/5) 26 (26/0/0) Acochlidiacea 3 (3/0/0) 31 (31/0/0) 36 (34/1/1) Sacoglossa 17 (15/0/2) 39 (38/0/1) 36 (34/2/0) Thecosomata 15 (13/1/1) ** * Thecosomata + Gymnosomata 9 (9/0/0) ** 20 (19/0/1) Tabelle 11: Stützung hypothetischer Monophyla innerhalb des Opisthobranchia-Datensatzes durch die
Anzahl potentieller Apomorphien
* nur 1 Sequenz in diesem Taxon
** keine Sequenz in diesem Taxon

73
Anaspidea-Datensätze (18S rDNA, ITS1 - 5.8S rDNA - ITS2) Die beiden Datensätze der Anaspidea erscheinen ebenfalls relativ verrauscht, symmetrische und asymmetrische stützende Positionen zeigen sich, wie bei den Opisthobranchia-Datensätzen, nur bei den am stärksten gestützten Gruppierungen. Zu letzteren gehören die gesamte Innengruppe, bestehend aus Anaspidea und Pteropoda (Thecosomata und Gymnosomata), sowie die Anaspidea. Im ITS1-5.8S rDNA-ITS2 - Datensatz entsprechen 50 stützende Positionen etwa 6 % der Gesamtalinierung, im Vergleich zu 3 % bei der 18S rDNA.
18S rDNA ITS1 - 5.8S rDNA - ITS2 Hypothetisches Monophylum Anzahl potentieller Apomorphien
Gesamtzahl (verrauschte/symm./asymm. Positionen)
Aplysia spp. 11 (11/0/0) 42 (40/1/1) Aplysiinae 11 (11/0/0) 42 (40/1/1) Longicommissurata 13 (13/0/0) 28 (27/1/0) Dolabriferinae 8 (8/0/0) 48 (42/3/3) Notarchinae 5 (5/0/0) * Brevicommissurata 10 (9/0/1) 37 (37/0/0) Aplysiidae 17 (17/0/0) 51 (44/4/3) Anaspidea 26 (26/0/0) 99 (84/8/7) Anaspidea + Pteropoda 50 (37/13/0) * Tabelle 12: Stützung hypothetischer Monophyla innerhalb des Anaspidea-Datensatzes durch die
Anzahl potentieller Apomorphien
* nur 1 Sequenz in diesem Taxon
Aeolidoidea-Datensatz (18S rDNA) Die Facelinidae weisen 341 potentielle Apomorphien auf, davon sind 89 verrauscht, 222 symmetrisch und 30 asymmetrisch. Die Gattung Phyllodesmium wird durch 331 (329/1/1) potentielle Apomorphien gestützt. 3.4 Intragenomische Variabilität 16S rDNA Die Analyse der intragenomischen Variabilität beim 16S rDNA-Gen ergab keine signifikanten Unterschiede zwischen den Sequenzen (jeweils 479 Basenpaare), das heißt, alle sequenzierten Klone eines Individuums von Unela glandulifera (Acochlidiacea) stimmten zu 99,6 % überein. Die zu beobachtenden 0,4 % Unterschiede wurden durch 2 Positionen in der Alinierung verursacht. An diesen beiden Positionen unterschied sich jeweils 1 Klon durch ein anderes Nukleotid von

74
den übrigen. In Anbetracht der zum Teil geringen Qualität der Sequenzierreationen, kann diese Variabilität mit sehr hoher Wahrscheinlichkeit auf Sequenzierungsfehler zurückgeführt werden. ITS1-5.8S rDNA-ITS2 Zur Untersuchung der intragenomischen Variabilität innerhalb der internal transcribed spacer wurden 8 Klone von Dolabrifera brazieri (Anaspidea) sequenziert. Im Gegensatz zum 16S rRNA-Gen wiesen die ITS-Sequenzen geringe Längendifferenzen auf (1083-1087 Bp). Von den insgesamt 1090 Positionen der Alinierung konnte an 23 Nukleotidpositionen (13 Positionen in ITS1, 1 in 5.8S, 9 in ITS2) in mindestens einem Klon ein abweichendes Nukleotid festgestellt werden. Überwiegend handelt es sich bei diesen Unterschieden wohl um Sequenzierungsfehler, allerdings fallen 2 markante Stellen auf, an denen jeweils die gleichen 4 Klone durch eine Insertion/Deletion (Indel) von 2 bzw. 3 Nukleotiden zusammengefasst werden. Daneben existieren weitere 2 Stellen, an denen jeweils die gleichen 3 Klone ein Indel von einer Base aufweisen. Ohne Berücksichtigung der potentiellen Sequenzierungsfehler liegen mindestens 3 paraloge Gene vor. 3.5 Phylogenie der Opisthobranchia In allen Analysen zur phylogenetischen Rekonstruktion der Opisthobranchia wurden die im jeweiligen Datensatz enthaltenen Vertreter der „Prosobranchia“ als Außengruppentaxon verwendet. 3.5.1 18S rDNA 3.5.1.1 Maximum Parsimonie- (MP-) Analyse In der strikten Konsensustopologie der MP-Analyse (Abb. 15) erscheinen die Opisthobranchia als Paraphylum, da die Allogastropoda (Pyramidellidae) und Pulmonata innerhalb der Opisthobranchia gruppieren. Die Monophylie aller Großgruppen der Hinterkiemer ist gegeben, mit Ausnahme der Anaspidea, die eine monophyletische Gruppierung mit den Pteropoda (Thecosomata und Gymnosomata) bilden. Die Nudibranchia und Pleurobranchoidea (Nudipleura) zeigen eine basale Stellung innerhalb der Innengruppe. Abgesehen von den Nudipleura existiert noch ein weiteres Schwestergruppenverhältnis zwischen zwei opisthobranchiaten Großgruppen, dasjenige zwischen Cephalaspidea s.str. und Anaspidea (inklusive Pteropoda). Die Tylodinoidea bilden mit den beiden letztgenannten Taxa ein Monophylum, das wiederum die Schwestergruppe zu den Acochlidiacea darstellt. Die Cephalaspidea s.str., Anaspidea (inklusive Pteropoda), Tylodinoidea, Acochlidiacea,

75
Pulmonata und Pyramidellidae bilden zusammen mit den Sacoglossa und Lymnaea stagnalis (Pulmonata) ein nicht aufgelöstes Monophylum, gemeinsam repräsentieren sie die Schwestergruppe der Acteonoidea. Eine bootstrap-Analyse konnte aufgrund der Größe des Datensatzes nicht durchgeführt werden. 3.5.1.2 Distanzanalyse Die Parameter für das zur Korrektur der sichtbaren Distanzen verwendete Sequenzevolutionsmodell, wurden mit Hilfe hierarchischer likelihood ratio tests (Modeltest, Posada & Crandall 1998) ermittelt. Die aufgrund der geringen Distanzen schlecht zu erkennenden Großgruppenverzweigungen werden in einem Dendrogramm unter Punkt 3.4.1.5 im Vergleich zu den Ergebnissen der MP-, ML- (Maximum Likelihood-) und Bayes’schen Analyse dargestellt. Der Übersichtlichkeit halber erfolgt die Angabe der bootstrap-Werte ebenfalls in dieser Topologie. Sowohl die Opisthobranchia als auch die Pulmonata werden in der Verwandtschaftshypothese der Distanzanalyse (Abb. 16) wieder als paraphyletische Gruppierungen rekonstruiert. Im Vergleich zum MP-Stammbaum sind die Acochlidiacea und die Cephalaspidea s.str. nicht monophyletisch und die Thecosomata gruppieren nicht zusammen mit den Gymnosomata und Anaspidea. Übereinstimmend ist das Schwestergruppenverhältnis zwischen Nudibranchia und Pleurobranchoidea, einen weiteren Unterschied bildet dagegen das Schwestergruppenverhältnis der Nudipleura zu den Acteonoidea. Die Sacoglossa und Lymnaea stagnalis bilden die Schwestergruppe zu einer Vereinigung aus Tylodinoidea, Pyramidellidae und Auriculinella bidentata (Ellobiidae, Pulmonata), alle zusammen stellen die Schwestergruppe der restlichen Ellobiidae und Acochlidiacea (ohne Pontohedyle milatchevitchi) dar. Ganz basal innerhalb der Innengruppe stehen die Thecosomata, gefolgt von Pontohedyle milatchevitchi (Acochlidiacea), den Anaspidea (inklusive Gymnosomata), den Cephalaspidea s.str. (inklusive Cepaea hortensis, Pulmonata) und einem Monophylum aus Pulmonaten, den Acteonoidea und den Nudipleura. Besonders lange Äste, gleichbedeutend mit erhöhten Substitutionsraten, sind innerhalb der Nudipleura zu beobachten, weiterhin fallen vergleichsweise lange Äste bei Calliopaea bellula (Sacoglossa), Lymnaea stagnalis (Pulmonata), den Acteonoidea, wenigen Cephalaspidea s.str. (z.B. Runcina adriatica), Spongiobranchaea australis (Gymnosomata), Pontoheyle milatchevitchi (Acochlidiacea) und den Thecosomata auf.

76
Acteon sp. Navanax inermis Odontoglaja guamensis Chelidonura inornata Philinopsis pilsbryi Philinoglossa praelongata Sagaminopteron psychedelicum Colpodaspis thomsoni Haminoea hydatis Atys semistriatus Bullacta exarata Phanerophthalmus smaragdinus Smaragdinella sp. Bulla cf. striata cf. Retusa sp. Cylichna gelida Scaphander lignarius Diaphana cf. inflata Toledonia globosa Newnesia antarctica Runcina adriatica Petalifera petalifera Stylocheilus longicauda Dolabrifera brazieri Bursatella leachii Clio pyramidata Creseis acicula Diacria quadridentata Spongiobranchaea australis Akera bullata Syphonota geographica Dolabella auricularia Aplysia californica Umbraculum mediterraneum Tylodina perversa Unela glandulifera Parahedyle cryptophthalma Pontohedyle milatchevitchi Hedylopsis spiculifera Onchidiella floridana Onchidium verrucosum Siphonaria alternata Cepaea hortensis Melampus luteus Cassidula nucleus Ovatella firminii Pythia scarabaeus Auriculinella bidentata Pyramidella sulcata Odostomia sp. Elysia viridis Elysiella pusilla Thuridilla bayeri Placobranchus ocellatus Bosellia mimetica Polybranchia orientalis Cyerce nigricans Calliopaea bellula Limapontia nigra Placida cremoniana Lymnaea stagnalis Hydatina physis Bullina lineata Pupa solidula Pleurobranchus peroni Bathyberthella antarctica Euselenops luniceps Tompthompsonia antarctica Berthellina citrina Berthella stellata Berthella platei Chromodoris krohni Hypselodoris villafranca Dendrodoris fumata Bathydoris clavigera Cuthona nana Flabellina babai Dendronotus dalli Fasciolaria lignaria Pisania striata Bursa rana Crepidula adunca Truncatella guerinii Viviparus georgianus Balcis eburnea Melaraphe neritoides Littoraria undulata Littorina striata Nodilittorina punctata Littorina saxatilis Littorina littorea
Cephalaspidea s.str
Anaspidea
Pteropoda
Anaspidea
Tylodinoidea
Acochlidiacea
Pulmonata
Pyramidellidae
Sacoglossa
Pulmonata
Acteonoidea
Pleurobranchoidea
Nudibranchia
Abb. 15: Strikter Maximum Parsimonie-Konsensusbaum aus 1.000.000 kürzesten Bäumen auf der Grundlage des 18S rDNA-Datensatzes. Suchparameter: stepwise addition = random, branch swapping = TBR, Alinierungslücken als 5. Nukleotid bewertet. Baumlänge: 3193, CI: 0,51, RI: 0,79, RC: 0,4.
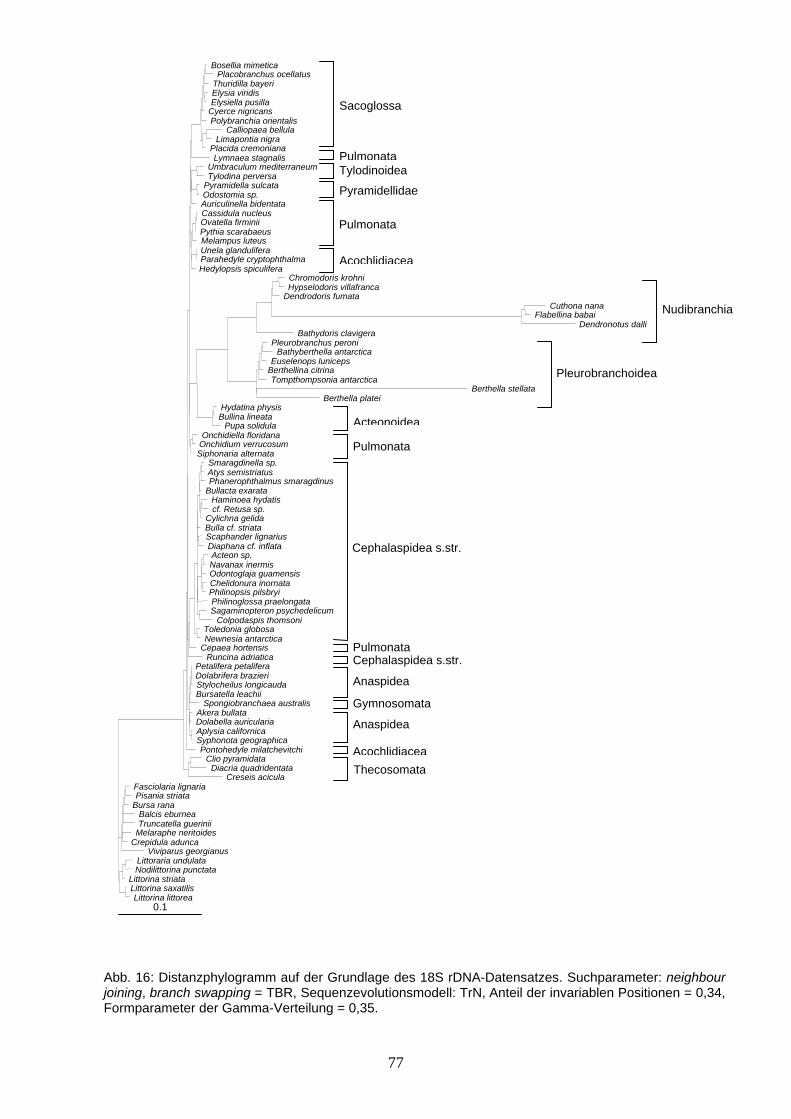
77
0.1
Bosellia mimetica Placobranchus ocellatus
Thuridilla bayeri Elysia viridis Elysiella pusilla
Cyerce nigricans Polybranchia orientalis
Calliopaea bellula Limapontia nigra
Placida cremoniana Lymnaea stagnalis
Umbraculum mediterraneum Tylodina perversa
Pyramidella sulcata Odostomia sp. Auriculinella bidentata Cassidula nucleus Ovatella firminii Pythia scarabaeus Melampus luteus Unela glandulifera Parahedyle cryptophthalma Hedylopsis spiculifera
Chromodoris krohni Hypselodoris villafranca
Dendrodoris fumata Cuthona nana
Flabellina babai Dendronotus dalli
Bathydoris clavigera Pleurobranchus peroni
Bathyberthella antarctica Euselenops luniceps
Berthellina citrina Tompthompsonia antarctica
Berthella stellata Berthella platei
Hydatina physis Bullina lineata
Pupa solidula Onchidiella floridana
Onchidium verrucosum Siphonaria alternata
Smaragdinella sp. Atys semistriatus Phanerophthalmus smaragdinus
Bullacta exarata Haminoea hydatis cf. Retusa sp.
Cylichna gelida Bulla cf. striata Scaphander lignarius Diaphana cf. inflata Acteon sp. Navanax inermis Odontoglaja guamensis Chelidonura inornata Philinopsis pilsbryi Philinoglossa praelongata Sagaminopteron psychedelicum
Colpodaspis thomsoni Toledonia globosa Newnesia antarctica
Cepaea hortensis Runcina adriatica
Petalifera petalifera Dolabrifera brazieri Stylocheilus longicauda Bursatella leachii
Spongiobranchaea australis Akera bullata Dolabella auricularia Aplysia californica Syphonota geographica Pontohedyle milatchevitchi
Clio pyramidata Diacria quadridentata
Creseis acicula Fasciolaria lignaria Pisania striata
Bursa rana Balcis eburnea Truncatella guerinii
Melaraphe neritoides Crepidula adunca
Viviparus georgianus Littoraria undulata Nodilittorina punctata
Littorina striata Littorina saxatilis Littorina littorea
Cephalaspidea s.str.
Anaspidea
Thecosomata
Anaspidea
Tylodinoidea
Acochlidiacea
Pulmonata
Pyramidellidae
Sacoglossa
Pulmonata
Acteonoidea
Pulmonata
Acochlidiacea
Cephalaspidea s.str.
Gymnosomata
Pulmonata
Pleurobranchoidea
Nudibranchia
Abb. 16: Distanzphylogramm auf der Grundlage des 18S rDNA-Datensatzes. Suchparameter: neighbour joining, branch swapping = TBR, Sequenzevolutionsmodell: TrN, Anteil der invariablen Positionen = 0,34, Formparameter der Gamma-Verteilung = 0,35.

78
0.1
Acteon sp. Navanax inermis Odontoglaja guamensis
Chelidonura inornata Philinopsis pilsbryi
Philinoglossa praelongata Sagaminopteron psychedelicum
Colpodaspis thomsoni Bulla cf. striata
Phanerophtalmus smaragdinus Atys semistriatus Smaragdinella sp. Haminoea hydatis
Bullacta exarata Cylichna gelida
cf. Retusa sp. Scaphander lignarius Diaphana cf. inflata
Toledonia globosa Newnesia antarctica Runcina adriatica
Clio pyramidata Creseis acicula
Diacria quadridentata Petalifera petalifera
Spongiobranchaea australis Stylocheilus longicauda Dolabrifera dolabrifera Bursatella leachii Aplysia californica Syphonota geographica Akera bullata Dolabella auricularia
Elysia viridis Elysiella pusilla Thuridilla ratna
Bosellia mimetica Placobranchus ocellatus
Polybranchia orientalis Cyerce nigricans
Calliopaea bellula Limapontia nigra
Placida cremoniana Lymnaea stagnalis
Pyramidella sulcata Odostomia sp. Umbraculum mediterraneum Tylodina perversa
Auriculinella bidentata Hedylopsis spiculifera
Pleurobranchus peroni Berthella stellata
Bathyberthella antarctica Euselenops luniceps Tompthompsonia antarctica
Berthellina citrina Berthella platei
Chromodoris krohni Hypselodoris villafranca
Dendrodoris fumata Bathydoris clavigera
Dendronotus dalli Cuthona nana
Flabellina babai Cassidula nucleus Ovatella firminii Pythia scarabaeus Unela glandulifera Parahedyle cryptophthalma
Hydatina physis Bullina lineata
Pupa solidula Onchidiella floridana
Onchidium verrucosum
Pontohedyle milatchevitchi Melampus luteus Cepaea hortensis
Siphonaria alternata Truncatella guerinii
Viviparus georgianus Fasciolaria lignaria Pisania striata
Bursa rana Crepidula adunca Balcis eburnea
Melaraphe neritoides Littorina saxatlis
Littorina striata Littoraria undulata
Nodilittorina punctata Littorina littorea
Tylodinoidea
Acochlidiacea
Pulmonata
Pyramidellidae
Sacoglossa
Pulmonata
Acteonoidea
Pulmonata
Pleurobranchoidea
Nudibranchia
Cephalaspidea s.str.
Anaspidea
Thecosomata
Anaspidea
Pulmonata Acochlidiacea
Gymnosomata
Pulmonata
Acochlidiacea
Abb. 17: Phylogramm der Bayes’schen Analyse auf der Grundlage des 18S rDNA-Datensatzes. Sequenzevoluitonsmodell: TrN, Anteil der invariablen Positionen = 0,34, Formparameter der Gamma-Verteilung = 0,35. Parameter der MCMC-Simulationen entsprachen den Grundeinstellungen von MrBayes (Huelsenbeck & Ronquist 2001a), burnin value = 1000.

79
Cephalaspidea s.str.
Anaspidea inkl. Pteropoda
Tylodinoidea
Acochlidiacea
Pulmonata
Pyramidellidae
Sacoglossa
Pulmonata
Acteonoidea
Pleurobranchoidea
Nudibranchia
Sacoglossa
Pulmonata
Tylodinoidea
Pyramidellidae
Pulmonata
Pulmonata
Acochlidiacea
Pleurobranchoidea
Nudibranchia
Acteonoidea
Pulmonata
Cephalaspidea s.str.
Pulmonata
Cephalaspidea s.str.
Anaspidea inkl. Gymnosomata
Acochlidiacea
Thecosomata
Abb. 18: Stammbaumhypothesen auf der Grundlage des 18S rDNA-Datensatzes. Arten wurden zu monophyletischen Gruppen zusammengefasst, unterschiedliche Astlängen wurden nicht berücksichtigt. A: Topologie der Maximum Parsimonie-Analyse, B: Topologie der Distanzanalyse, C: Topologie der Bayes’schen Analyse, nur posterior probabilities der Großgruppen angegeben.
Cephalaspidea s.str.
Anaspidea inkl. Pteropoda
Sacoglossa
Pulmonata
Pyramidellidae
Tylodinoidea
Pulmonata
Acochlidiacea
Pulmonata
Pulmonata
Acochlidiacea
Acteonoidea
Pleurobranchoidea
Nudibranchia
0,53
0,94
0,94
0,66 0,99
0,63 0,72
0,5
0,99
0,94
0,98
0,98
0,99
0,62
0,63
0,99
0,95
A
B
C

80
3.5.1.3 maximum likelihood- (ML-) Analyse Eine Stammbaumrekonstruktion mit Hilfe des Maximum Likelihood-Verfahrens konnte aufgrund der Größe des Datensatzes und des damit verbundenen Rechenaufwandes nicht durchgeführt werden. 3.5.1.4 Bayes’sche Analyse Für die mit Hilfe des Programmes MrBayes (Huelsenbeck & Ronquist 2001a) durchgeführte Analyse wurden ebenfalls die durch hierarchische likelihood ratio tests ermittelten Parameter der Sequenzevolution eingesetzt. In Abb. 17 ist der Konsensusbaum (50% majority rule) der Bayes’schen Analyse dargestellt, die posterior probabilities werden der Übersichtlichkeit halber erst in Abb. 18 unter Punkt 3.5.1.5 angegeben. Wie in den beiden vorangegangenen Bäumen, sind auch hier Pulmonata und Opisthobranchia paraphyletisch, die Subtaxa der Opisthobranchia dagegen mit Ausnahme der Acochlidiacea monophyletisch. Ein Schwestergruppenverhältnis zwischen Nudibranchia und Pleurobranchoidea ist zu beobachten, ebenso wie eines zwischen Cephalaspidea s.str. und Anaspidea (inklusive Pteropoda). Ein drittes Monophylum besteht aus den Sacoglossa und Lymnaea stagnalis, den Pyramidellidae, Tylodinoidea, Auriculinella bidentata (Pulmonata) und Hedylopsis spiculifera (Acochlidiacea). Darüber hinaus konnten die Verwandtschaftsverhältnisse zwischen den Taxa des Datensatzes nicht aufgelöst werden. 3.5.1.5 Zusammenfassung 18S rDNA In Abb. 18 sind die auf der Grundlage von ML-, Distanz- und Bayes’schem Verfahren rekonstruierten Verwandtschaftshypothesen dargestellt. Der Übersichtlichkeit halber und zum leichteren Vergleich der verschiedenen Topologien, wurden die einzelnen Arten zu Großgruppenmonophyla zusammengefasst und es wurde auf die Darstellung von Astlängen verzichtet. Übereinstimmungen zwischen den Bäumen bestehen zum einen im Status der Opisthobranchia und Pulmonata als Paraphyla, zum anderen in der Monophylie der einzelnen Großgruppen. Ausnahmen stellen die Anaspidea dar, die paraphyletisch sind, bzw. deren Verwandtschaft zu den Pteropoda nicht aufgelöst werden konnte. Weiterhin sind die Cephalaspidea s.str. im Distanzbaum paraphyletisch und die Acochlidiacea im Distanz- und Bayes’schen Phylogramm. Das Schwestergruppenverhältnis zwischen Nudibranchia und Pleurobranchoidea wird durch alle drei Verfahren rekonstruiert, das zwischen Sacoglossa und Lymnaea stagnalis (Pulmonata) im den durch Distanz- und Bayes’sche Analyse erstellten Topologien. Das Schwestertaxonverhältnis zwischen Cephalaspidea s.str. und

81
Anaspidea (inklusive Pteropoda) ist im Stammbaum der MP- und Bayes’schen Analyse zu beobachten, im Distanzphylogramm dagegen nicht. Die Stellung der Tylodinoidea erscheint variabel, entweder als Schwestergruppe zu Cephalapidea s.str. und Anaspidea (inklusive Pteropoda), zu den Pyramidellidae oder in einer monophyletischen Gruppierung mit den Sacoglossa, Pulmonaten und Pyramidellidae. Die Sacoglossa zeigen eine verwandtschaftliche Nähe zu den Pyramidellidae und Pulmonaten im MP-Baum und zusätzlich zu den Tylodinoidea in den beiden anderen Topologien. Die Stellung der Acochlidiacea ist ebenfalls äußerst variabel, ihre Vertreter bilden entweder die Schwestergruppe zu den Cephalaspidea s.str., Anaspidea (inklusive Pteropoda) und Tylodinoidea (Parsimonie), zu den Pulmonaten (ohne Pontohedyle) oder zu Pulmonaten einerseits (Unela, Parahedyle) und zu Sacoglossa, Tylodinoidea, Pulmonaten und Pyramidellidae andererseits (Hedylopsis). Die Acteonoidea repräsentieren entweder die Schwestergruppe aller Taxa (ohne Nudipleura) oder die der Nudipleura. 3.5.2 28S rDNA 3.5.2.1 Maximum Parsimonie-Analyse In dem in Abb. 19 gezeigten strikten Konsensusbaum bilden die Opisthobranchia, ebenso wie die Pulmonata, ein paraphyletisches Taxon. Die meisten Subtaxa der Opisthobranchia erscheinen als Monophyla (bootstrap-Werte: 67-100), mit Ausnahme der paraphyletischen Acochlidiacea. Auch die Nudibranchia sind in diesem Stammbaum paraphyletisch, da die Cladobranchia die Schwestergruppe zu den Pleurobranchoidea darstellen. In der Konsensustopologie der bootstrap-Analyse repräsentieren sie dagegen ein monophyletisches Taxon (bootstrap-Wert: 57). Ein weiteres, schon in den Stammbäumen auf der Grundlage des 18S-Datensatzes rekonstruiertes Monophylum, resultiert aus dem Schwestergruppenverhältnis von Cephalaspidea s.str und Anaspidea. Das Schwestertaxonverhältnis von Sacoglossa und Tylodinoidea konnte durch die bootstrap-Analyse nicht gestützt werden. Die Nudipleura nehmen eine basale Stellung innerhalb der Innengruppe ein, gefolgt von den Acteonoidea. Die übrigen Taxa gruppieren in zwei Monophyla, bestehend aus Cephalaspidea s.str. und Anaspidea einerseits und Pulmonata, Pyramidellidae, Acochlidiacea, Tylodinoidea und Sacoglossa andererseits. 3.5.2.2 Distanzanalyse Die Parameter des Sequenzevolutionsmodells wurden, wie beim 18S rDNA- Datensatz, mit Hilfe hierarchischer likelihood ratio tests, implementiert im Programm Modeltest (Posada & Crandall 1998) ermittelt.

82
Die Opisthobranchia repräsentieren in diesem Distanzphylogramm (Abb. 20) eine paraphyletische Gruppierung, die Pulmonaten bilden zusammen mit Odostomia sp. (Pyramidellidae) ein Monophylum. Die opisthobranchiaten Subtaxa sind ohne Ausnahme monophyletisch (bootstrap-Werte: 94-100) und sowohl das Schwestergruppenverhältnis zwischen Cephalaspidea s.str. und Anaspidea, als auch das zwischen Nudibranchia und Pleurobranchoidea ist vorhanden. Darüber hinaus bestehen keine wesentlichen Unterschiede zu dem durch das MP-Verfahren erzeugten Stammbaum. Die Verhältnisse der Astlängen entsprechen im Wesentlichen denen der 18S-Phylogramme. Die meisten Substitutionen sind innerhalb der Nudipleura zu beobachten, relativ lange Äste weisen zudem die Acteonoidea, Runcina adriatica (Cephalaspidea s.str.) und Cepaea hortensis (Pulmonata) auf. 3.5.2.3 maximum likelihood-Analyse Für die Stammbaumrekonstruktion mittels des Maximum Likelihood-Verfahrens, wurden ebenfalls die durch das Programm Modeltest (Posada & Crandall 1998) ermittelten Parameter eingesetzt. Die Opisthobranchia repräsentieren in dieser Verwandtschaftshypothese (Abb. 21), ebenso wie die Pulmonaten, ein parahyletisches Taxon, die einzelnen Gruppen sind mit Ausnahme der Acochlidiacea und der Cephalaspidea s.str. monophyletisch. Die Nudipleura (Nudibranchia und Pleurobranchoidea) stehen an der Basis der Innengruppe, gefolgt von den paraphyletischen Cephalaspidea s.str., den Anaspidea, Hedylopsis spiculifera (Acochlidiacea) und Lymnaea stagnalis (Pulmonata). Die restlichen Taxa gruppieren in zwei Monophyla, bestehend aus Sacoglossa und Acteonoidea einerseits und Acochlidiacea, Pulmonata, Pyramidellidae und Tylodinoidea andererseits. 3.5.2.4 Bayes’sche Analyse Für die Bayes’sche Analyse wurden wiederum die mit Modeltest (Posada & Crandall 1998) ermittelten Parameter verwendet. Die Opisthobranchia erscheinen in diesem Phylogramm (Abb. 22) als Paraphylum, der Status der Pulmonaten ist aufgrund mangelnder Auflösung unklar. Die Subtaxa der Opisthobranchia sind, mit Ausnahme der Cephalaspidea s.str. und Acochlidiacea, monophyletisch und werden durch hohe posterior probabilities gestützt (0,99). Das Schwestergruppenverhältnis von Nudibranchia und Pleurobranchoidea ist vorhanden, das von Cephalaspidea s.str. und Anaspidea dagegen nicht. Die Anaspidea bilden das Schwestertaxon zu einer monophyletischen Gruppierung aus Sacoglossa, Tylodinoidea, Pulmonata, Pyramidellidae,

83
Cyerce nigricans Placobranchus ocellatus Thuridilla bayeri Bosellia mimetica Elysia viridis Calliopaea bellula Limapontia nigra Tylodina perversa Umbraculum mediterraneum Odostomia sp. Cepaea hortensis Lymnaea stagnalis Unela glandulifera Parahedyle cryptophthalma Pontohedyle milatchevitchi Onchidiella floridana Onchidium verrucosum Siphonaria alternata Hedylopsis spiculifera Phanerophthalmus smaragdinus Atys semistriatus Haminoea hydatis cf. Retusa sp. Bulla cf. striata Sagaminopteron psychedelicum Philinoglossa praelongata Chelidonura inornata Odontoglaja guamensis Philinopsis pilsbryi Navanax inermis Runcina adriatica Aplysia californica Dolabella auricularia Akera bullata Pupa solidula Hydatina physis Pleurobranchus peroni Berthellina citrina Euselenops luniceps Bathyberthella antarctica Tompthompsonia antarctica Berthella stellata Flabellina babai Cuthona nana Dendronotus dalli Chromodoris krohni Hypselodoris villafranca Dendrodoris fumata Bathydoris clavigera Nodilittorina unifasciata Tectarius striatus Littoraria undulata Littorina littorea Melarhaphe neritoides
Tylodinoidea
Pyramidellidae
Sacoglossa
Pulmonata
Cephalaspidea s.str
Anaspidea
Acochlidiacea
Acochlidiacea
Pulmonata
Acteonoidea
Nudibranchia
Pleurobranchoidea
54 57
57 84
100
99
98
100 99
100
85 90
62
97
99
81
67
56
76
100
84 100
100
100 100
97 52
98
98 99
99 100
Abb. 19: Strikter Maximum Parsimonie-Konsensusbaum aus 3 Bäumen auf der Grundlage des 28S rDNA-Datensatzes. Suchparameter: stepwise addition = random, branch swapping = TBR, Alinierungslücken als 5. Nukleotid bewertet. Baumlänge: 3069, CI = 0,44, RI = 0,66, RC = 0,29. Zahlen repräsentieren bootstrap-Werte.

84
0.1
Onchidiella floridana Onchidium verrucosum
Siphonaria alternata Cepaea hortensis
Lymnaea stagnalis Odostomia sp. Unela glandulifera Parahedyle cryptophthalma Pontohedyle milatchevitchi
Hedylopsis spiculifera Tylodina perversa
Umbraculum mediterraneum Thuridilla bayeri Placobranchus ocellatus
Bosellia mimetica Cyerce nigricans
Elysia viridis Calliopaea bellula
Limapontia nigra Chelidonura inornata
Odontoglaja guamensis Navanax inermis Philinopsis pilsbryi Philinoglossa praelongata Sagaminopteron psychedelicum Phanerophthalmus smaragdinus
Atys semistriatus Haminoea hydatis cf. Retusa sp. Bulla cf. striata
Runcina adriatica Aplysia californica
Dolabella auricularia Akera bullata
Pupa solidula Hydatina physis
Chromodoris krohni Hypselodoris villafranca
Dendrodoris fumata Bathydoris clavigera
Dendronotus dalli Flabellina babai
Cuthona nana Berthella stellata
Bathyberthella antarctica Euselenops luniceps
Pleurobranchus peroni Berthellina citrina
Tompthompsonia antarctica Nodilittorina unifasciata Tectarius striatus Littoraria undulata Littorina littorea
Melarhaphe neritoides
Tylodinoidea
Pyramidellidae
Sacoglossa
Pulmonata
Cephalaspidea s.str.
Anaspidea
Acochlidiacea
Acteonoidea
Nudibranchia
Pleurobranchoidea
79
100 100
99
71 77
100
100
58
99 84
70
74 82 76
100 60
52
100
100
81
100
98 79
99
51 100
94
93 96
100
Abb. 20: Distanzphylogramm auf der Grundlage des 28S rDNA-Datensatzes. Suchparameter: neighbour joining, branch swapping = TBR, Sequenzevolutionsmodell: TrN, Anteil der invariablen Positionen = 0,28, Formparameter der Gamma-Verteilung = 0,54. Zahlen repräsentieren bootstrap-Werte.

85
0.1
Tylodina perversa Umbraculum mediterraneum
Siphonaria alternata Odostomia sp.
Onchidiella floridana Onchidium verrucosum
Cepaea hortensis Unela glandulifera Parahedyle cryptophthalma
Pontohedyle milatchevitchi Cyerce nigricans Placobranchus ocellatus Thuridilla bayeri
Bosellia mimetica Elysia viridis
Calliopaea bellula Limapontia nigra
Pupa solidula Hydatina physis
Lymnaea stagnalis Hedylopsis spiculifera
Aplysia californica Dolabella auricularia
Akera bullata Phanerophthalmus smaragdinus
Atys semistriatus Haminoea hydatis cf. Retusa sp. Chelidonura inornata Odontoglaja guamensis Philinopsis pilsbryi
Navanax inermis Philinoglossa praelongata
Sagaminopteron psychedelicum Bulla cf. striata Runcina adriatica
Chromodoris krohni Hypselodoris villafranca
Dendrodoris fumata Bathydoris clavigera
Flabellina babai Cuthona nana Dendronotus dalli
Pleurobranchus peroni Berthellina citrina
Euselenops luniceps Bathyberthella antarctica
Berthella stellata Tompthompsonia antarctica
Tectarius striatus Littoraria undulata
Nodilittorina unifasciata Littorina littorea Melarhaphe neritoides
Tylodinoidea
Pyramidellidae
Sacoglossa
Pulmonata
Cephalaspidea s.str.
Anaspidea
Acochlidiacea
Acteonoidea
Acochlidiacea
Pulmonata
Nudibranchia
Pleurobranchoidea
Abb. 21: Maximum likelihood-Phylogramm auf der Grundlage des 28S rDNA-Datensatzes. Suchparameter: neighbour joining, branch swapping = TBR, Sequenzevolutionsmodell: TrN, Anteil der invariablen Positionen = 0,28, Formparameter der Gamma-Verteilung = 0,54.

86
0.1
Cyerce nigricans Placobranchus ocellatus
Thuridilla bayeri Bosellia mimetica
Elysia viridis Calliopaea bellula
Limapontia nigra Tylodina perversa
Umbraculum mediterraneum Odostomia sp.
Onchidella floridana Onchidium verrucosum
Cepaea hortensis Unela glandulifera Parahedyle cryptophthalma
Pontohedyle milatchevitchi Pupa solidula Hydatina physis
Hedylopsis spiculifera Lymnaea stagnalis
Siphonaria alternata Aplysia californica Dolabella auricularia
Akera bullata Phanerophtalmus smaragdinus
Atys semistriatus Haminoea hydatis cf. Retusa sp.
Philinoglossa praelongata Sagaminopteron psychedelicum
Bulla cf. striata Chelidonura inornata Odontoglaja guamensis Philinopsis pilsbryi
Navanax inermis Runcina adriatica
Chromodoris krohni Hypselodoris villafranca
Dendrodoris fumata Bathydoris clavigera
Flabellina babai Cuthona nana Dendronotus dalli
Pleurobranchus peroni Berthellina citrina
Euselenops luniceps Bathyberthella antarctica
Berthella stellata Tompthompsonia antarctica
Tectarius striatus Littoraria undulata
Nodilittorina unifasciata Littorina littorea
Melarhaphe neritoides
0,50 0,98 0,99
0,99
0,99
0,99
0,99
0,57 0,99
0,90
0,98
1,00
1,00 0,99
0,94
0,91 0,99
0,98 1,00
0,93
0,88
0,52 0,88
0,98
0,99 1,00 0,68
1,00
0,99 0,86
0,98
0,99 0,97
0,99
0,99
0,93 0,93
0,94 0,98
0,80 0,93
0,88 1,00
0,96
0,99
Tylodinoidea
Pyramidellidae
Sacoglossa
Pulmonata
Cephalaspidea s.str.
Anaspidea
Acochlidiacea
Acteonoidea
Acochlidiacea
Pulmonata
Nudibranchia
Pleurobranchoidea
Abb. 22: Phylogramm der Bayes’schen Analyse auf der Grundlage des 18S rDNA-Datensatzes. Sequenzevolutionsmodell: TrN, Anteil der invariablen Positionen = 0,28, Formparameter der Gamma-Verteilung = 0,54. Zahlen repräsentieren posterior probabilities. Parameter der MCMC-Simulationen entsprachen den Grundeinstellungen von MrBayes (Huelsenbeck & Ronquist 2001a), burnin value = 1000.

87
Acochlidiacea und Acteonoidea. Darüber hinaus zeigt sich, im Gegensatz zu den durch die anderen Rekonstruktionsmethoden ermittelten Stammbäumen, und in Übereinstimmung mit dem Phylogramm der Bayes’schen Analyse auf der Grundlage der 18S rDNA, eine sehr geringe Auflösung.
Sacoglossa
Tylodinoidea
Pyramidellidae
Pulmonata
Pulmonata
Acochlidiacea
Cephalaspidea s.str.
Anaspidea
Acteonoidea
Pleurobranchoidea
Nudibranchia
Pulmonata
Pyramidellidae
Acochlidiacea
Tylodinoidea
Sacoglossa
Cephalaspidea s.str.
Anaspidea
Acteonoidea
Nudibranchia
Pleurobranchoidea
Tylodinoidea
Pulmonata
Pyramidellidae
Pulmonata
Acochlidiacea
Sacoglossa
Acteonoidea
Pulmonata
Acochlidiacea
Anaspidea
Cephalaspidea s.str.
Nudibranchia
Pleurobranchoidea
Sacoglossa
Tylodinoidea
Pyramidellidae
Pulmonata
Acochlidiacea
Acteonoidea
Anaspidea
Cephalaspidea s.str.
Nudibranchia
Pleurobranchoidea
Abb. 23: Stammbaumhypothesen auf der Grundlage des 28S rDNA-Datensatzes. Arten wurden zu monophyletischen Gruppen zusammengefasst, unterschiedliche Astlängen wurden nicht berücksichtigt. A: Topologie der Maximum Parsimonie-Analyse, B: Topologie der Distanzanalyse, C: Topologie der Bayes’schen Analyse, D: Topologie der Bayes’schen Analyse.
B
C
D
D

88
3.5.2.5 Zusammenfassung 28S rDNA In Abb. 23 sind die vier Verwandtschaftshypothesen auf der Basis des 28S rDNA-Datensatzes vergleichend dargestellt. Auf die Berücksichtigung von Astlängen wurde verzichtet und die einzelnen Arten wurden zu Taxa höherer Ordnung zusammengefasst. Die Opisthobranchia und Pulmonata erscheinen übereinstimmend als paraphyletische Gruppierungen, die Monophylie der einzelnen Großgruppen wird dagegen, mit Ausnahme der Cephalaspidea s.str. und Acochlidiacea, gut gestützt. Die Nudibranchia und Pleurobranchoidea zeigen ein Schwestergruppenverhältnis auf der Grundlage aller Rekonstruktionsverfahren, bei der MP-Analyse allerdings nur in der bootstrap-Topologie. Die Monophylie von Cephalaspidea s.str. und Anaspidea konnte nur durch die MP- und Distanzanalyse rekonstruiert werden. Die Stellung der Sacoglossa erscheint, wie auf der Grundlage der 18S rDNA, recht variabel. Sie repräsentieren entweder die Schwestergruppe der Tylodinoidea (bootstrap-Wert: 0) oder Acteonoidea, alternativ bilden sie ein Monophylum mit den Tylodinoidea, Acochlidiacea, Pulmonata und Pyramidellidae oder mit den letztgenannten Taxa plus Acteonoidea. Die Tylodinoidea zeigen ebenfalls deutliche Unterschiede bezüglich der postulierten Verwandtschaftsbeziehungen. Sie gruppieren entweder als Schwestertaxon zu den Sacoglossa, zu Siphonaria alternata (Pulmonata), zu Odostomia sp. (Pyramidellidae) oder sie bilden eine monophyletische Einheit mit den Acochlidiacea, Pulmonata und Pyramidellidae. Die Acochlidiacea gruppieren immer in Monophyla mit den Pulmonata, Pyramidellidae, Tylodinoidea, Sacoglossa und Acteonoidea, ihre Stellung innerhalb dieser Gruppierung erscheint allerdings variabel. Die Acteonoidea zeigen entweder ein Schwestergruppenverhältnis zu allen Taxa der Innengruppe (mit Ausnahme der Nudipleura) oder zu den Sacoglossa. Alternativ bilden sie ein nicht aufgelöstes Monophylum mit den Sacoglossa, Tylodinoidea, Acochlidiacea, Pulmonata und Pyramidellidae. 3.5.3 16S rDNA 3.5.3.1 Maximum Parsimonie-Analyse Die Opisthobranchia stellen in dieser strikten Konsensustopologie (Abb. 24), wie in den Stammbaumhypothesen der vorangegangenen Datensätze, eine paraphyletische Gruppierung dar. Die Nudibranchia und Acteonoidea sind paraphyletisch, da Dendrodoris nigra (Nudibranchia) als Schwestergruppe von Pupa solidula (Acteonoidea) und die Cladobranchia (Nudibranchia) als Schwestertaxon der Pleurobranchoidea erscheinen. Die übrigen Subtaxa der Opisthobranchia sind monophyletisch.

89
Die Monophylie der Nudipleura ist aus den oben genannten Gründen nicht gegeben, auch die Cephalaspidea s.str. und Anaspidea, sowie die Theco- und Gymnosomata, stehen nicht in einem Schwestergruppenverhältnis zueinander. Die Taxa der Acteonoidea und Nudipleura bilden eine monophyletische Gruppierung mit den Anaspidea und Tylodinoidea und stellen die Schwestergruppe der Cephalaspidea s.str. dar. Alle genannten Taxa zusammen repräsentieren die Schwestergruppe zu einer Vereinigung aus Acochlidiacea, Pulmonata, Thecosomata und Pyramidellidae. Clione limacina (Gymnosomata) nimmt eine ganz basale Stellung in der Innengruppe ein, in Richtung höherer Ableitung gefolgt von den Sacoglossa. 3.5.3.2 Distanzanalyse Die Parameter für das zugrundeliegende Sequenzevolutionsmodell wurden wiederum mit Hilfe hierarchischer likelihood ratio tests (Modeltest, Posada & Crandall 1998) ermittelt und ebenfalls in der ML- und Bayes’schen Analyse berücksichtigt. In Übereinstimmung mit dem Parsimoniebaum auf der Grundlage der 16S rDNA sind die Opisthobranchia im gezeigten Distanzphylogramm (Abb. 25) paraphyletisch, die einzelnen Großgruppen innerhalb der Opisthobranchia sind mit Ausnahme der Nudibranchia und Acteonoidea monophyletisch. Die Anaspidea stellen das Schwestertaxon der Nudipleura dar, die beiden zusammen bilden mit den Sacoglossa, Acteonoidea, Pyramidellidae, Acochlidiacea, Pulmonata, Thecosomata und Dendrodoris nigra (Nudibranchia) ein Monophylum. Alle gennanten Taxa repräsentieren die Schwestergruppe der Cephalaspidea s.str. Clione limacina (Gymnosomata) zeigt eine basale Stellung innerhalb der Innengruppe. Die Differenzen in den Astlängen der einzelnen Taxa sind hier nicht so ausgeprägt wie auf der Grundlage der beiden vorangegangenen Datensätze. Die längsten Äste in diesem Phylogramm weisen Dendrodoris nigra (Nudibranchia), Cavolinia sp. (Thecosomata) und Odostomia sp. auf. 3.5.3.3 maximum likelihood-Analyse Die Opisthobranchia werden im Phylogramm der ML-Analyse (Abb. 26) wieder als paraphyletische Gruppierung rekonstruiert, weitere Übereinstimmungen zu den beiden vorhergehenden Topologien bestehen im Status all ihrer Subtaxa. Die Nudipleura bilden mit Pupa solidula (Acteonoidea) eine monophyletische Gruppierung, die im Schwestergruppenverhältnis zu den Anaspidea steht. Gemeinsam mit den Tylodinoidea bilden diese Taxa die Schwestergruppe zu den Sacoglossa. Die Gruppierung aus allen erwähnten Gruppen stellt das Schwestertaxon der Thecosomata dar.

90
Ein zweites Monophylum besteht aus Hydatina physis (Acteonoidea), Sacoglossa, Acochlidiacea, Pulmonata und Pyramidellidae. Clione limacina (Gymnosomata) nimmt wieder eine ganz basale Stellung innerhalb der Innengruppe ein. 3.5.3.4 Bayes’sche Analyse Die Opisthobranchia bilden in der Verwandtschaftshypothese der Bayes’schen Analyse (Abb. 27), genau wie in allen bisher rekonstruierten Topologien, eine paraphyletische Gruppierung. Mit Ausnahme der Acteonoidea und der Nudibranchia sind alle Großgruppen innerhalb der Opisthobranchia monophyletisch, die Nudibranchia (ohne Dendrodoris nigra) bilden ein Monophylum mit den Pleurobranchoidea. Weiterhin besteht ein Schwestergruppenverhältnis sowohl zwischen den Tylodinoidea und Anaspidea, als auch zwischen den Acochlidiacea und Pythia scarabaeus (Pulmonata). Die beiden letztgenannten Taxa stellen die Schwestergruppe von Odostomia sp. (Pyramidellidae) dar und erscheinen zusammen mit Dendrodoris nigra als Schwestertaxon zu den Sacoglossa. Weitere Verwandtschaftsbeziehungen der Großgruppen zueinander konnten nicht aufgelöst werden. Die Stellung von Clione limacina (Gymnosomata) als Schwestergruppe zu allen übrigen Innengruppentaxa, stimmt mit der in den anderen Stammbäumen auf der Grundlage der 16S rDNA überein. 3.5.3.5 Zusammenfassung 16S rDNA In Abb. 28 sind die Ergebnisse der phylogenetischen Untersuchungen auf der Grundlage der 16S rDNA vergleichend dargestellt. Die einzelnen Arten wurden wiederum in Großgruppen zusammengefasst und die unterschiedlichen Astlängen nicht berücksichtigt. Die Opisthobranchia und Pulmonata stellen basierend auf dem 16S rRNA-Gen übereinstimmend paraphyletische Gruppierungen dar, die opisthobranchiaten Subtaxa erscheinen, mit Ausnahme der Nudibranchia und Acteonoidea, in allen Topologien als Monophyla. Übereinstimmungen bestehen daneben ausschließlich im Schwestergruppenverhältnis von Acochlidiacea und Pulmonata, sowie von Clione limacina (Gymnosomata) zu allen übrigen Innengruppentaxa. Davon abgesehen herrscht unter den phylogenetischen Beziehungen der Gruppen zueinander eine recht große Variabilität. Verwandtschaftliche Tendenzen zeigen die Anaspidea und Tylodinoidea, sowie die Nudipleura und Acteonoidea. Darüber hinaus lassen sich keine Gesetzmäßigkeiten erkennen.
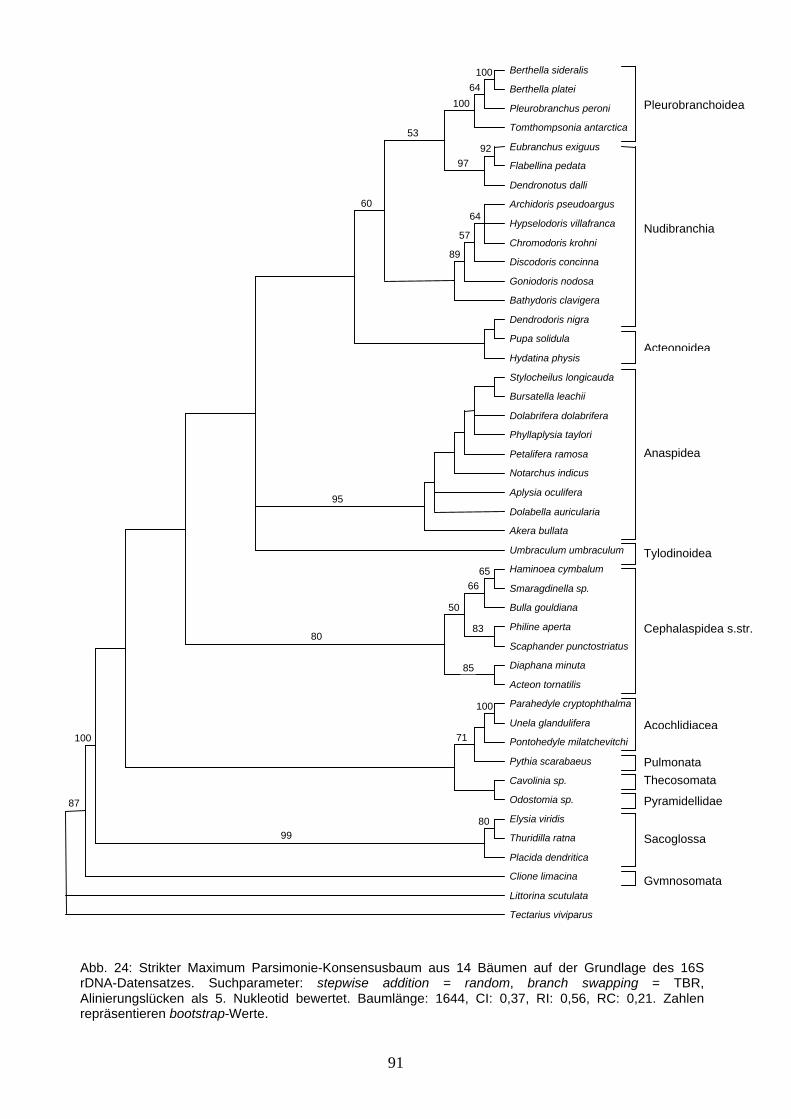
91
Berthella sideralis Berthella platei Pleurobranchus peroni Tomthompsonia antarctica Eubranchus exiguus Flabellina pedata Dendronotus dalli Archidoris pseudoargus Hypselodoris villafranca Chromodoris krohni Discodoris concinna Goniodoris nodosa Bathydoris clavigera Dendrodoris nigra Pupa solidula Hydatina physis Stylocheilus longicauda Bursatella leachii Dolabrifera dolabrifera Phyllaplysia taylori Petalifera ramosa Notarchus indicus Aplysia oculifera Dolabella auricularia Akera bullata Umbraculum umbraculum Haminoea cymbalum Smaragdinella sp. Bulla gouldiana Philine aperta Scaphander punctostriatus Diaphana minuta Acteon tornatilis Parahedyle cryptophthalma Unela glandulifera Pontohedyle milatchevitchi Pythia scarabaeus Cavolinia sp. Odostomia sp. Elysia viridis Thuridilla ratna Placida dendritica Clione limacina Littorina scutulata Tectarius viviparus
100 64
100
53
92 97
64
57
89
60
95
65 66
50
83 80
85
100
71
80 99
87
100
Tylodinoidea
Pyramidellidae
Sacoglossa
Thecosomata
Cephalaspidea s.str.
Anaspidea
Gymnosomata
Acteonoidea
Acochlidiacea
Pulmonata
Nudibranchia
Pleurobranchoidea
Abb. 24: Strikter Maximum Parsimonie-Konsensusbaum aus 14 Bäumen auf der Grundlage des 16S rDNA-Datensatzes. Suchparameter: stepwise addition = random, branch swapping = TBR, Alinierungslücken als 5. Nukleotid bewertet. Baumlänge: 1644, CI: 0,37, RI: 0,56, RC: 0,21. Zahlen repräsentieren bootstrap-Werte.

92
0.1
Eubranchus exiguus Flabellina pedata
Dendronotus dalli Bathydoris clavigera
Berthella sideralis Berthella platei
Pleurobranchus peroni Tomthompsonia antarctica
Archidoris pseudoargus Hypselodoris villafranca
Chromodoris krohni Discodoris concinna
Goniodoris nodosa Phyllaplysia taylori Petalifera ramosa
Dolabrifera dolabrifera Bursatella leachii Aplysia oculifera Dolabella auricularia
Notarchus indicus Stylocheilus longicauda Akera bullata
Elysia viridis Thuridilla ratna
Placida dendritica Hydatina physis
Odostomia sp. Parahedyle cryptophthalma Unela glandulifera
Pontohedyle milatchevitchi Pythia scarabaeus
Dendrodoris nigra Cavolinia sp.
Pupa solidula Haminoea cymbalum Smaragdinella sp. Bulla gouldiana
Philine aperta Scaphander punctostriatus
Diaphana minuta Acteon tornatilis
Umbraculum umbraculum Clione limacina
Littorina scutulata Tectarius viviparus
64 73
55
93
84 73
98
98
Tylodinoidea
Pyramidellidae
Sacoglossa
Thecosomata
Cephalaspidea s.str.
Anaspidea
Gymnosomata
Acteonoidea
Acochlidiacea
Pulmonata
Nudibranchia
Pleurobranchoidea
Nudibranchia
Nudibranchia
Acteonoidea
99
61
50 59
51
56
70
95
59 83
98
100
97 85
89
60
97 75
Abb. 25: Distanzphylogramm auf der Grundlage des 16S rDNA-Datensatzes. Suchparameter: neighbour joining, branch swapping = TBR, Sequenzevolutionsmodell: TVM, Anteil der invariablen Positionen = 0,25, Formparameter der Gamma-Verteilung = 0,64. Zahlen repräsentieren bootstrap-Werte.

93
0.1
Berthella sideralis Berthella platei
Tomthompsonia antarctica Pleurobranchus peroni
Eubranchus exiguus Flabellina pedata
Dendronotus dalli Archidoris pseudoargus
Hypselodoris villafranca Chromodoris krohni
Discodoris concinna Goniodoris nodosa
Dendrodoris nigra Bathydoris clavigera
Pupa solidula Dolabrifera dolabrifera Phyllaplysia taylori
Bursatella leachii Stylocheilus longicauda
Petalifera ramosa Aplysia oculifera Dolabella auricularia
Notarchus indicus Akera bullata
Umbraculum umbraculum Haminoea cymbalum
Smaragdinella sp. Bulla gouldiana Philine aperta Scaphander punctostriatus
Diaphana minuta Acteon tornatilis
Cavolinia sp. Parahedyle cryptophthalma Unela glandulifera
Pontohedyle milatchevitchi Pythia scarabaeus
Odostomia sp. Elysia viridis
Thuridilla ratna Placida dendritica
Hydatina physis Clione limacina
Littorina scutulata Tectarius viviparus
Tylodinoidea
Anaspidea
Acteonoidea
Pleurobranchoidea
Cephalaspidea s.str.
Nudibranchia
Pyramidellidae
Sacoglossa
Pulmonata
Gymnosomata
Thecosomata
Acteonoidea
Acochlidiacea
Abb. 26: Maximum likelihood-Phylogramm auf der Grundlage des 16S rDNA-Datensatzes. Suchparameter: neighbour joining, branch swapping = TBR, Sequenzevolutionsmodell: TVM, Anteil der invariablen Positionen = 0,25, Formparameter der Gamma-Verteilung = 0,64.

94
0,1
Berthella sideralis Berthella platei
Tomthompsonia antarctica Pleurobranchus peroni
Eubranchus exiguus Flabellina pedata
Dendronotus dalli Archidoris pseudoargus
Hypselodoris villafranca Chromodoris krohni
Discodoris concinna Goniodoris nodosa
Bathydoris clavigera Aplysia oculifera Dolabella auricularia
Notarchus indicus Akera bullata
Petalifera ramosa Dolabrifera dolabrifera Phyllaplysia taylori Bursatella leachii
Stylocheilus longicauda Umbraculum umbraculum
Parahedyle cryptophthalma Unela glandulifera
Pontohedyle milatchevitchi Pythia scarabaeus
Odostomia sp. Dendrodoris nigra
Elysia viridis Thuridilla ratna
Placida dendritica Haminoea cymbalum
Smaragdinella sp. Bulla gouldiana Philine aperta
Scaphander lignarius Diaphana minuta Acteon tornatilis
Cavolinia sp. Hydatina physis
Pupa solidula Clione limacina
Littorina scutulata Tectarius viviparus
1,00 0,90
1,00
0,93
1,00 1,00
0,55 0,62
1,00
0,60
0,99 1,00
0,84
0,51
0,74
0,67
0,52
0,73
1,00
0,50
1,00 0,91
0,70
0,56
0,53
0,52
0,87 0,99
0,99 0,99
0,76
0,99 0,94
1,00
0,99
0,93
0,53
Tylodinoidea
Anaspidea
Acteonoidea
Pleurobranchoidea
Nudibranchia
Cephalaspidea s.str.
3
Sacoglossa
2
Gymnosomata
Thecosomata
4
1
1 Acochlidiacea 2 Pulmonata 3 Pyramidellidae 4 Nudibranchia
Abb. 27: Phylogramm der Bayes’schen Analyse auf der Grundlage des 16S rDNA-Datensatzes. Sequenzevolutionsmodell: TVM, Anteil der invariablen Positionen = 0,25, Formparameter der Gamma-Verteilung = 0,64. Zahlen repräsentieren posterior probabilities. Parameter der MCMC-Simulationen entsprachen den Grundeinstellungen von MrBayes (Huelsenbeck & Ronquist 2001a), burnin value = 1000.

95
Abb. 28: Stammbaumhypothesen auf der Grundlage des 16S rDNA-Datensatzes. Arten wurden zu monophyletischen Gruppen zusammengefasst, unterschiedliche Astlängen wurden nicht berücksichtigt. A: Topologie der Maximum Parsimonie-Analyse, B: Topologie der Distanzanalyse, C: Topologie der maximum likelihood-Analyse, D: Topologie der Bayes’schen Analyse.
Anaspidea
Tylodinoidea
Cephalaspidea s.str.
Pleurobranchoidea
Nudibranchia
Acochlidiacea
Pulmonata
Nudibranchia
Thecosomata
Pyramidellidae
Sacoglossa
Acteonoidea
Gymnosomata
Pleurobranchoidea
Nudibranchia
Nudibranchia
Acteonoidea
Anaspidea
Tylodinoidea
Cephalaspidea s.str.
Thecosomata
Acochlidiacea
Pulmonata
Pyramidellidae
Sacoglossa
Acteonoidea
Gymnosomata
Pleurobranchoidea
Nudibranchia
Anaspidea
Tylodinoidea
Acochlidiacea
Pulmonata
Pyramidellidae
Nudibranchia
Sacoglossa
Cephalaspidea s.sr.
Thecosomata
Acteonoidea
Gymnosomata
Pleurobranchoidea
Nudibranchia
Nudibranchia
Acteonoidea
Anaspidea
Tylodinoidea
Cephalaspidea s.str.
Acochlidiacea
Pulmonata
Thecosomata
Pyramidellidae
Sacoglossa
Gymnosomata A B
C D

96
3.6 Phylogenie der Anaspidea (inklusive Pteropoda) 3.6.1 18S rDNA 3.6.1.1 Maximum Parsimonie-Analyse Die Ergebnisse der Maximum Parsimonie-Analyse, basierend auf dem 18S rRNA-Gen, sind in Abb. 29 dargestellt. Die Anaspidea erscheinen hier als monophyletische Gruppierung, da die Pteropoda ein basales Paraphylum bilden. Spongiobranchaea australis (Gymnosomata) gruppiert dabei als Schwestergruppe zu den Anaspidea und Thecosomata. Innerhalb der Anaspidea stellt Bursatella leachii die Schwestergruppe der übrigen Anaspideen dar, die Stellung von Petalifera petalifera, Dolabrifera spp. und Stylocheilus longicauda wird dabei nicht aufgelöst. Die übrigen Taxa repräsentieren ein monophyletisches Taxon mit Dolabella auricularia an der Basis und Akera bullata als Schwestergruppe der Aplysiinae (Aplysia spp., Syphonota geographica). 3.6.1.2 Distanzanalyse Die Parameter für das zur Korrektur der sichtbaren Distanzen verwendete Sequenzevolutionsmodell, wurden mit Hilfe hierarchischer likelihood ratio tests (Modeltest, Posada & Crandall 1998) ermittelt und auch in der nachfolgenden ML- und Bayes’schen Analyse berücksichtigt. Im Distanzphylogramm auf der Grundlage des 18S rRNA-Gens (Abb. 30) erscheinen die Anaspidea nicht als monophyletisches Taxon, da Spongiobranchaea australis (Gymnosomata) die Schwestergruppe von Akera bullata darstellt. Die Thecosomata sind monophyletisch und repräsentieren die Schwestergruppe der Anaspidea (inklusive Gymnosomata). Die übrigen Innengruppentaxa bilden zwei Monophyla. Das eine besteht aus den monophyletischen Aplysiinae (Aplysia spp., Syphonota geographica), die zusammen mit Spongiobranchaea australis und Akera bullata die Schwestergruppe von Dolabella auricularia repräsentieren. Im zweiten monophyletischen Taxon steht Petalifera petalifera basal und bildet mit Stylocheilus longicauda die Schwestergruppe von Dolabrifera spp. Beide genannten Monophyla zusammen, stellen die Schwestergruppe von Bursatella leachii dar. Die Thecosomata zeigen die höchsten Substitutionsraten innerhalb des Datensatzes, zudem sind vergleichsweise lange Äste bei Spongiobranchaea australis und Akera bullata zu beobachten. Die restlichen Taxa weisen überwiegend sehr geringe Distanzen auf, besonders innerhalb der Aplysiinae.
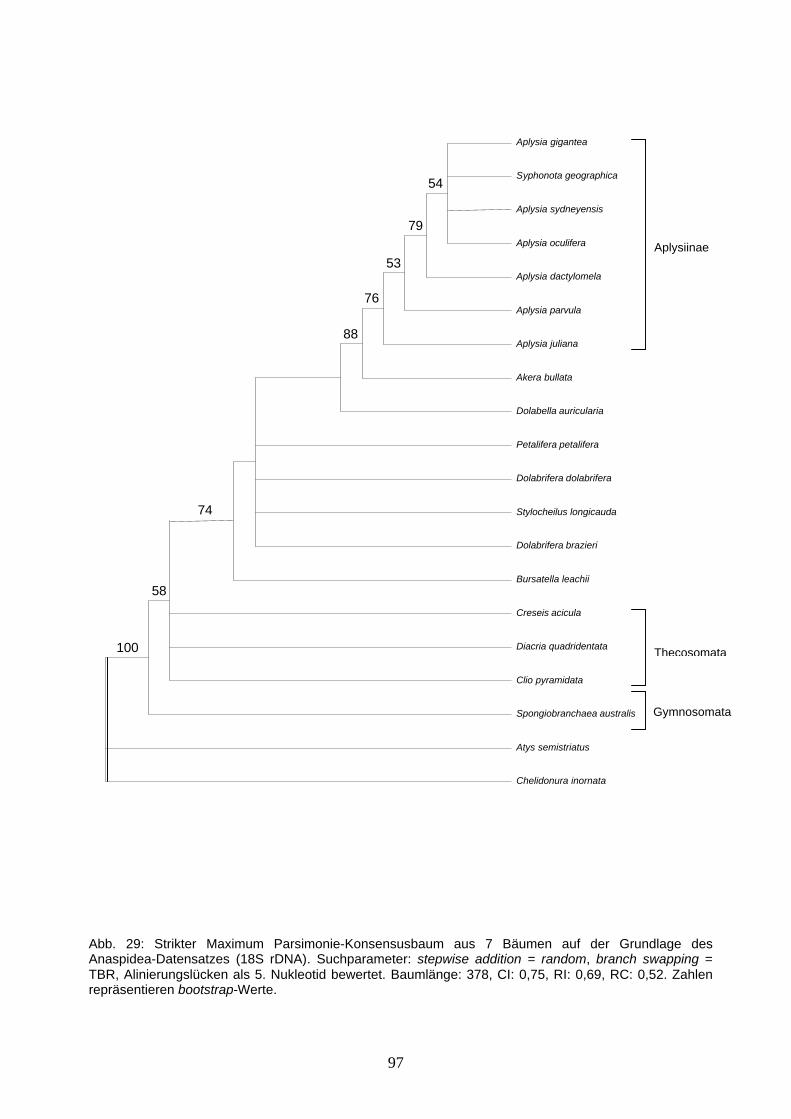
97
Aplysia gigantea
Syphonota geographica
Aplysia sydneyensis
Aplysia oculifera
Aplysia dactylomela
Aplysia parvula
Aplysia juliana
Akera bullata
Dolabella auricularia
Petalifera petalifera
Dolabrifera dolabrifera
Stylocheilus longicauda
Dolabrifera brazieri
Bursatella leachii
Creseis acicula
Diacria quadridentata
Clio pyramidata
Spongiobranchaea australis
Atys semistriatus
Chelidonura inornata
Thecosomata
Gymnosomata
Aplysiinae
74
88
76
53
79
54
58
100
Abb. 29: Strikter Maximum Parsimonie-Konsensusbaum aus 7 Bäumen auf der Grundlage des Anaspidea-Datensatzes (18S rDNA). Suchparameter: stepwise addition = random, branch swapping = TBR, Alinierungslücken als 5. Nukleotid bewertet. Baumlänge: 378, CI: 0,75, RI: 0,69, RC: 0,52. Zahlen repräsentieren bootstrap-Werte.

98
0.01
Aplysia sydneyensis
Aplysia juliana
Aplysia dactylomela
Aplysia gigantea
Aplysia oculifera
Syphonota geographica
Aplysia parvula
Akera bullata
Spongiobranchaea australis
Dolabella auricularia
Dolabrifera brazieri
Dolabrifera dolabrifera
Stylocheilus longicauda
Petalifera petalifera
Bursatella leachii
Creseis acicula
Clio pyramidata
Diacria quadridentata
Atys semistriatus
Chelidonura inornata
Thecosomata
Gymnosomata
Aplysiinae
90
80
100
63
72
56
91
Abb. 30: Distanzphylogramm auf der Grundlage des Anaspidea-Datensatzes (18S rDNA). Suchparameter: neighbour joining, branch swapping = TBR, Sequenzevolutionsmodell: TrNef, Anteil der invariablen Positionen = 0,75, Formparameter der Gamma-Verteilung = 0,64. Zahlen repräsentieren bootstrap-Werte.

99
0.01
Aplysia sydneyensis
Aplysia juliana
Aplysia parvula
Aplysia dactylomela
Aplysia gigantea
Syphonota geographica
Aplysia oculifera
Akera bullata
Spongiobranchaea australis
Dolabella auricularia
Dolabrifera brazieri
Dolabrifera dolabrifera
Petalifera petalifera
Stylocheilus longicauda
Bursatella leachii
Creseis acicula
Clio pyramidata
Diacria quadridentata
Atys semistriatus
Chelidonura inornata
Gymnosomata
Aplysiinae
Thecosomata
Abb. 31: Maximum likelihood-Phylogramm auf der Grundlage des Anaspidea-Datensatzes (18S rDNA). Suchparameter: neighbour joining, branch swapping = TBR, Sequenzevolutionsmodell: TrNef, Anteil der invariablen Positionen = 0,75, Formparameter der Gamma-Verteilung = 0,64.

100
0.01
Aplysia gigantea
Syphonota geographica
Aplysia dactylomela
Aplysia juliana
Aplysia parvula
Aplysia sydneyensis
Aplysia oculifera
Akera bullata
Spongiobranchaea australis
Dolabella auricularia
Petalifera petalifera
Dolabrifera dolabrifera
Stylocheilus longicauda
Dolabrifera brazieri
Bursatella leachii
Creseis acicula
Clio pyramidata
Diacria quadridentata
Atys semistriatus
Chelidonura inornata
0,96
0,96
0,92
0,93
0,78
0,97
0,89
1,00
1,00
Gymnosomata
Thecosomata
Aplysiinae
Abb. 32: Phylogramm der Bayes’schen Analyse auf der Grundlage des Anaspidea-Datensatzes (18S rDNA). Sequenzevolutionsmodell: TrNef, Anteil der invariablen Positionen = 0,75, Formparameter der Gamma-Verteilung = 0,64. Zahlen repräsentieren posterior probabilities. Parameter der MCMC-Simulationen entsprachen den Grundeinstellungen von MrBayes (Huelsenbeck & Ronquist 2001a), burnin value = 1000.

101
3.6.1.3 Maximum Likelihood-Analyse Die Ergebnisse der ML-Analyse (Abb. 31) stimmen größtenteils mit denen der Distanzanalyse überein. Der einzige Unterschied besteht in der Paraphylie der Gruppierung aus Dolabrifera spp., Stylocheilus longicauda und Petalifera petalifera, die in der Topologie der Distanzanalyse als Monophylum erscheint. 3.6.1.4 Bayes’sche Analyse Die auf der Basis einer Bayes’schen Analyse erstellte Verwandtschaftshypothese (Abb. 32), zeigt keine wesentlichen Unterschiede zu den beiden vorhergehenden Phylogrammen, abgesehen von der weitaus geringeren Auflösung. Diese betrifft zum einen die Aplysiinae (Aplysia spp., Syphonota geographica) und zum anderen die Taxa Dolabrifera spp., Stylocheilus longicauda und Petalifera petalifera. 3.6.1.5 Zusammenfassung Die durch Distanz-, Maximum Likelihood- und Bayes’schem Verfahren erzeugten Topologien unterscheiden sich nur im Status von Dolabrifera spp., Petalifera petalifera und Stylocheilus longicauda als mono- oder paraphyletische Gruppierung und im Grad der Auflösung. In Abb. 33 ist ein Konsensusbaum aus den Topologien der drei genannten Verfahren dem durch eine MP-Analyse erzeugten Baum gegenübergestellt. Auf die Darstellung von Astlängen wurde dabei verzichtet und einzelne Arten wurden zu Taxa höherer Ordnung zusammengefasst.
Aplysiinae
Akera bullata
Dolabella auricularia
Petalifera petalifera
Dolabrifera spp.
Stylocheilus longicauda
Bursatella leachii
Thecosomata
Gymnosomata
Aplysiinae
Akera bullata
Gymnosomata
Dolabella auricularia
Petalifera petalifera
Dolabrifera spp.
Stylocheilus longicauda
Bursatella leachii
Thecosomata
Abb. 33: Stammbaumhypothesen auf der Grundlage des Anaspidea-Datensatzes (18S rDNA). Arten wurden zu monophyletischen Gruppen zusammengefasst, unterschiedliche Astlängen wurden nicht berücksichtigt. A: Topologie der Maximum Parsimonie-Analyse, B: Konsensustopologie der Distanz-, maximum likelihood- und Bayes’schen Analyse.
A B

102
3.6.2 ITS1-5.8S-ITS2 3.6.2.1 Maximum Parsimonie-Analyse In dem auf der Grundlage des ITS1-5.8S-ITS2 - Datensatzes rekonstruierten Stammbaum (Abb. 34A), erscheint Akera bullata als Schwestertaxon zu allen übrigen Vertretern der Anaspidea. Bursatella leachii und Petalifera petalifera, sowie Dolabrifera brazieri und Dolabrifera dolabrifera stehen jeweils in einem Schwestergruppenverhältnis zueinander und bilden mit Dolabella auricularia ein Monophylum. Diese Gruppierung repräsentiert zusammen mit Aplysia spp. ein monophyletisches Taxon, dabei stellen Aplysia gigantea und Aplysia oculifera Schwestertaxa dar. 3.6.2.2 Distanzanalyse Das Sequenzevolutionsmodell für die Distanz-, ML- und Bayes’sche Analyse, wurde auf der Grundlage hierarchischer likelihood ratio tests (Modeltest, Posada & Crandall 1998) ausgewählt. Der durch die Distanzanalyse rekonstruierte Stammbaum (Abb. 34B) stimmt weitestgehend mit dem durch das MP-Verfahren erzeugten Baum überein. Beide Topologien unterscheiden sich nur durch die Monophylie von Aplysia spp., deren verwandtschaftliche Beziehungen im MP-Baum nicht aufgelöst werden konnten. Die Verteilung der Astlängen erweist sich als relativ homogen, allein Bursatella leachii fällt durch eine vergleichsweise hohe Substitutionsrate auf. 3.6.2.3 Maximum Likelihood-Analyse Das in Abb. 34C gezeigte Phylogramm der ML-Analyse weist keine wesentlichen Unterschiede zu den beiden vorhergehenden Topologien auf, die einzige Diskrepanz besteht in der Paraphylie der Gattung Aplysia. 3.6.2.4 Bayes’sche Analyse Die nach dem Bayesschen Verfahren erzeugte Stammbaumhypothese (Abb. 34D) der Anaspidea entspricht genau derjenigen, die durch Anwendung des ML-Verfahrens rekonstruiert wurde.

103
0.1
Bursatella leachii
Petalifera petalifera
Dolabrifera brazieri
Dolabrifera dolabrifera
Dolabella auricularia
Aplysia gigantea
Aplysia oculifera
Aplysia juliana
Akera bullata
Atys semistriatus
Chelidonura inornata
75
97
100
100
100
97
91
100
0.1
Bursatella leachii
Petalifera petalifera
Dolabrifera brazieri
Dolabrifera dolabrifera
Dolabella auricularia
Aplysia juliana
Aplysia gigantea
Aplysia oculifera
Akera bullata
Atys semistriatus
Chelidonura inornata
0.1
Dolabrifera brazieri
Dolabrifera dolabrifera
Bursatella leachii
Petalifera petalifera
Dolabella auricularia
Aplysia juliana
Aplysia gigantea
Aplysia oculifera
Akera bullata
Atys semistriatus
Chelidonura inornata
1,0
1,00
0,97 1,0
0,8
1,0
0,9
1,0
Abb. 34: Stammbaumhypothesen auf der Grundlage des Anaspidea-Datensatzes (ITS1-5.8S-ITS2). A: Strikter Maximum Parsimonie-Konsensusbaum aus 2 Bäumen. Suchparameter: stepwise addition = random, branch swapping = TBR, Alinierungslücken als 5. Nukleotid bewertet. Baumlänge: 1360, CI: 0,77, RI: 0,67, RC: 0,52. B: Distanzphylogramm, Suchparameter: neighbour joining, branch swapping = TBR, Sequenzevolutionsmodell: TrN, Formparameter der Gamma-Verteilung = 0,48. C: maximum likelihood-Phylogramm, Parameter wie bei C. D: Phylogramm der Bayes’schen Analyse. Parameter der MCMC-Simulationen entsprachen den Grundeinstellungen von MrBayes (Huelsenbeck & Ronquist 2001a), burnin value = 1000. Zahlen repräsentieren bootstrap-Werte, bzw. posterior probabilities.
Bursatella leachii
Petalifera petalifera
Dolabrifera brazieri
Dolabrifera dolabrifera
Dolabella auricularia
Aplysia gigantea
Aplysia oculifera
Aplysia juliana
Akera bullata
Atys semistriatus
Chelidonura inornata
95
100 100
100
93
100
100
A B
C D

104
3.6.2.5 Zusammenfassung Zwischen den durch unterschiedliche phylogenetische Rekonstruktionsverfahren erstellten Verwandtschaftshypothesen sind nur geringe Unterschiede zu beobachten, die einzigen Widersprüche ergeben sich durch Mono-, bzw. Paraphylie der Gattung Aplysia. Die Topologie der MP-Analyse (Abb. 34A) kann als strikter Konsensusbaum aller erhaltenen Stammbäume angesehen werden. 3.7 Phylogenie von Phyllodesmium Alle durchgeführten phylogenetischen Rekonstruktionen mit Notaeolidia depressa als Außengruppe ergaben sowohl die Monophylie der Facelinidae (Baeolidia nodosa, Godiva banyulensis, Cratena peregrina, Phidiana lynceus, Facelina spp., Phyllodesmium spp.), als auch die der Gattung Phyllodesmium. Abb. 35 zeigt die durch eine MP-Analyse rekonstruierte Phylogenie der Taxa in einem Dendrogramm, zur Veranschaulichung der höchst unterschiedlichen Astlängen ist zusätzlich das Phylogramm der ML-Analyse dargestellt (Abb. 36). Innerhalb der Gattung Phyllodesmium zeigt P. cf. magnum eine basale Stellung. Die übrigen Taxa bilden zwei Monophyla, bestehend aus P. crypticum und P. jakobsenae einerseits und P. colemani, P. longicirrum und P. briareum andererseits.

105
Phyllodesmium colemani
Phyllodesmium longicirrum
Phyllodesmium briareum
Phyllodesmium crypticum
Phyllodesmium jakobsenae
Phyllodesmium cf. magnum
Godiva banyulensis
Baeolidia nodosa
Facelina bostoniensis
Facelina auriculata
Phidiana lynceus
Cratena peregrina
Flabellina sp.
Flabellina affinis
Flabellina verrucosa
Cuthona nana
Cuthona sibogae
Cuthona caerulea
Tergipes tergipes
Eubranchus exiguus
Eubranchus sp.
Flabellina pedata
Flabellina babai
Embletonia sp.
Notaeolidia depressa
Facelinidae
54
59
53
60 85
59
100
100
69
74
67
99
65
54
Abb. 35: Maximum Parsimonie-Baum (1 kürzester Baum) auf der Grundlage des Aeolidoidea-Datensatzes (18S rDNA). Suchparameter: stepwise addition = random, branch swapping = TBR, Alinierungslücken als 5. Nukleotid bewertet. Baumlänge: 1863, CI: 0,74, RI: 0,93, RC: 0,68. Zahlen repräsentieren bootstrap-Werte.

106
1
Phyllodesmium colemani
Phyllodesmium longicirrum
Phyllodesmium briareum
Phyllodesmium crypticum
Phyllodesmium jakobsenae
Phyllodesmium cf. magnum
Facelina bostoniensis
Facelina auriculata
Phidiana lynceus
Cratena peregrina
Godiva banyulensis
Baeolidia nodosa
Flabellina sp.
Flabellina affinis
Flabellina verrucosa
Cuthona nana
Cuthona caerulea
Cuthona sibogae
Flabellina pedata
Eubranchus exiguus
Eubranchus sp.
Tergipes tergipes
Embletonia sp.
Flabellina babai
Notaeolidia depressa
Facelinidae
Abb. 36: Distanzphylogramm auf der Grundlage des Aeolidoidea-Datensatzes (18S rDNA). Suchparameter: neighbour joining, branch swapping = TBR. Sequenzevolutionsmodell: TrNef, Anteil der invariablen Positionen = 0,75, Formparameter der Gamma-Verteilung = 0,64.

107
3.8 Umbraculum Die 18S rDNA-Sequenzen der drei Individuen von Umbraculum zeigen keinerlei Nukleotidunterschiede. Die Sequenzen des 16S rRNA-Gens differieren in zwei Positionen, dabei weisen jeweils die Individuen von der Großen Meteorbank und aus Australien ein übereinstimmendes Nukleotid auf. Ein Vergleich zwischen Innen- und Außengruppe ergab, dass es sich bei den beiden erwähnten Unterschieden in den drei Sequenzen von Umbraculum um Symplesiomorphien der Taxa von der Großen Meteobank und aus Australien handelt, entsprechend also um Autapomorphien für das Individuum von den Azoren. Die Sequenzen der Cytochromoxidase I zeigen Unterschiede in insgesamt 12 Positionen der Alinierung, die übereinstimmend die dritte Codonposition betreffen. Die Untersuchung der Aminosäuresequenz ergab eine völlige Identität. Abbildung 36 zeigt die apomorphen und plesiomorphen Nukleotidpositionen der drei Umbraculum-Individuen, projiziert auf den zugehörigen Stammbaum der Taxa. Aus der Abbildung wird ersichtlich, dass die Individuen aus Australien und von der Großen Meteor Bank durch 5 synapomorphe Merkmale, die Nukleotidpositionen 202, 247, 271, 304 und 343, vereint werden. Die Ergebnisse der anatomischen und histologischen Untersuchungen an Umbraculum sind ausführlich in der Arbeit von Wägele et al. (im Druck) dargelegt und werden hier nicht mehr zusätzlich aufgeführt.
Australien
Große Meteorbank
Azoren
Tylodina perversa
Abb. 36: Maximum Parsimonie-Baum auf der Grundlage der COI-Sequenzen dreier Individuen von Umbraculum (Australien, Große Meteor Bank, Azoren) und Tylodina perversa als Außengruppentaxon. Zahlen kennzeichnen die Nukleotidposition in der Alinierung (650 Bp), Buchstaben die Base an der jeweiligen Stelle. Positionen der Stammlinie (links) repräsentieren den plesiomorphen, rechts den autapomorphen oder synapomorphen Zustand.
166 457 C A
424 457 578 C G T
202 247 271 304 343 G C G A C
259 271 337 424 457 494 G A T T G C
166 202 247 259 271 304 337 343 424 457 494 578 T A T A T G C T T T T C

108
4 Diskussion 4.1 Auswahl der Taxa Die Auswahl der Taxa für eine phylogenetische Untersuchung kann einen nicht zu vernachlässigenden Einfluß auf das Ergebnis der Verwandtschaftsanalyse ausüben. Im Idealfall würde man alle Arten eines zu untersuchenden Taxons berücksichtigen, was bei der Artenzahl der Opisthobranchia, zumindest im Rahmen einer Doktorarbeit, nicht realisierbar ist. Es muß also eine geeignete Auswahl getroffen werden. Unglücklicherweise ist es nicht möglich zu testen, ob die Artenstichprobe ausreicht, um die Phylogenie zu rekonstruieren. Je mehr Taxa berücksichtigt werden, umso geringer wird zumindest die Gefahr, dass paraphyletische Gruppen durch Symplesiomorphien gestützt als Monophyla erscheinen oder durch Analogien (long branch attraction) zusammen gruppieren. Eine unzureichende Probennahme der untersuchten Taxa wird oft als bedeutende Fehlerquelle für phylogenetische Analysen genannt und dementsprechend wird die Erlangung eines möglichst großen Datensatzes befürwortet (z.B. Lecointre et al. 1993, Yoder & Irwin 1999, Johnson 2001). Computersimulationen auf der Basis realer Sequenzdaten von Säugern haben dagegen gezeigt, dass eine unvollständige Taxonauswahl einen weitaus geringeren Einfluß auf die korrekte Rekonstruktion einer Phylogenie ausübt als z.B. die Anzahl der Nukleotidpositionen oder die Verteilung der Substitutionsraten (Rosenberg & Kumar 2001). Bei der Untersuchung höherer taxonomischer Einheiten stehen Individuen stellvertretend für Arten, Gattungen (wie bei der Analyse der Opisthobranchia) oder sogar Taxa höherer Ordnung. Das ausgewählte Individuum sollte demnach „repräsentativ“ sein, also nicht zu sehr vom Grundmuster des zu vertretenden Taxons abweichen. Anders ausgedrückt heißt das, es sollte möglichst viele plesiomorphe und synapomorphe Merkmale besitzen und möglichst wenig Autapomorphien. Diesem Ziel können die meisten phylogenetischen Analysen nicht gerecht werden, da die Auswahl der Taxa in erster Linie von der Verfügbarkeit und nicht von der Eignung des Tiermaterials abhängig ist. Trotzdem ist es in der vorliegenden Arbeit gelungen, insgesamt eine recht ausgewogene und „repräsentative“ Taxonzusammenstellung zu erzielen, wobei einige Einschränkungen nicht unerwähnt bleiben sollen: Im Datensatz zur Rekonstruktion der Phylogenie der Opisthobranchia sind einige Großgruppen, z.B. die Gymnosomata zahlenmäßig unterrepräsentiert und in den einzelnen Gruppen sind einige wichtige Linien gar nicht vertreten, z.B. die beschalten Sacoglossa. Im 18S-Datensatz der Anaspidea sind die Gattungen Notarchus und Phyllaplysia nicht enthalten, im ITS1-5.8S-ITS2 - Datensatz fehlen weitaus mehr. Bei der Untersuchung der Evolution von Phyllodesmium konnten wichtige Arten nicht berücksichtigt werden, z.B. die als ursprünglich angesehenen Vertreter ohne symbiontische Beziehungen. Bei der Analyse der Umbraculum-Arten schließlich,

109
wäre die Einbeziehung weiterer Individuen von anderen Lokalitäten, z.B. dem Mittelmeer wünschenswert gewesen. 4.2 Untersuchte Gene 18S rDNA Das Gen für die RNA der kleinen ribosomalen Untereinheit ist aufgrund seiner niedrigen Evolutionsgeschwindigkeit sehr konserviert. Diese Tatsache drückt sich besonders deutlich im zahlenmäßigen Verhältnis von konstanten zu variablen Positionen in den Alinierungen der 18S rDNA-Sequenzen aus. Im Opisthobranchia-Datensatz, der neben den Opisthobranchia auch Vertreter der Pulmonata, Allogastropoda und Caenogastropoda enthält, erwiesen sich 1005 Positionen als konstant, das entspricht mehr als 50% aller Nukleotidpositionen. Bei Betrachtung des Anaspidea-Datensatzes erhöht sich der Anteil der konstanten Positionen aufgrund der geringeren Sequenzdivergenzen innerhalb der Alinierung auf knapp 90%. Die Untersuchung der drei Individuen von Umbraculum ergab erwartungsgemäß eine völlige Übereinstimmung der Sequenzen. Eine Gruppierung innerhalb der Opisthobranchia ist bezogen auf die Konserviertheit der 18S rDNA auffällig: Die Nudipleura (Nudibranchia + Pleurobranchoidea) zeigen eine im Vergleich zu den anderen Taxa erhöhte Variabilität. Werden die Nudipleura aus dem Datensatz entfernt, steigt der Anteil der konstanten Nukleotidpositionen auf über 75%. Diese größere Variabilität innerhalb der Nudipleura wird auch durch signifikant erhöhte Substitutionsraten (relative rate test) belegt. Besonders augenfällig wird die hohe Evolutionsgeschwindigkeit der Nudipleura durch ihre extrem langen Äste in den Phylogrammen der Distanz-, maximum likelihood- und Bayes’schen Analysen. Innerhalb der Nudipleura nehmen die drei Sequenzen der Cladobranchia (Nudibranchia) eine Sonderstellung ein, da sie neben den höchsten Substitutionsraten des gesamten Datensatzes, als einzige Gruppierung eine abweichende Basenzusammensetzung aufweisen. Eine Konvergenz in den Basenfrequenzen nicht verwandter Abstammungslinien ist ein Faktor, der die korrekte Rekonstruktion einer Phylogenie beeinträchtigen kann. Da die abweichende Basenzusammensetzung im Fall der 18S rDNA eine mit hoher Wahrscheinlichkeit monophyletische Gruppierung betrifft (Tholleson 1999, Wägele & Willan 2000, Wägele et al. 2003), sind keine negativen Auswirkungen auf die Stammbaumrekonstruktion zu erwarten. Zudem haben Untersuchungen ergeben, daß ein sehr hohes Maß an Konvergenz nötig ist, bevor z.B. das Parsimonieverfahren eine falsche Topologie bevorzugt (Conant & Lewis 2001). Bezüglich der Variabilität der Evolutionsraten verhält es sich ähnlich wie bei der Basenzusammensetzung. Insgesamt zeigt der 18S rDNA-Datensatz der Opisthobranchia sehr große Differenzen in den Substitutionsgeschwindigkeiten der einzelnen Sequenzen. Die Substitutionsraten sind allerdings gruppenspezifisch, d.h.

110
innerhalb der einzelnen (als monophyletisch angesehenen) Subtaxa der Opisthobranchia sind sie relativ homogen, so dass die Gefahr von long branch-Effekten zwischen nicht näher verwandten Taxa als relativ gering eingestuft wird. Die Analyse der Substitutionssättigung hat ergeben, dass die Transversionen einen recht hohen Grad an Sättigung aufweisen, die Transitionen dagegen keine Sättigungserscheinung zeigen. Dieses Ergebnis ist sehr ungewöhnlich, da Transitionen leichter entstehen als Transversionen und somit durch multiple Substitutionen schneller überlagert werden. Zudem weist das 18S rDNA-Gen eine sehr geringe Evolutionsgeschwindigkeit auf, so dass keinerlei Sättigung zu erwarten gewesen wäre. Die Ursache für die genannten Beobachtungen liegt möglicherweise in den signifikant erhöhten Substitutionsraten der Cladobranchia und von Berthella stellata (Pleurobranchoidea). Bei Nichtberücksichtigung dieser Taxa ergibt sich das erwartete Bild einer linearen Zunahme von Transitionen und Transversionen bei steigender evolutionärer Distanz. Da eine Substitutionssättigung nur die genannten Taxa innerhalb des Datensatzes betrifft ist das Auftreten von Konvergenzerscheinungen aufgrund von Mehrfachsubstitutionen, die die verwandtschaftlichen Beziehungen verfälschen können, als unwahrscheinlich zu bewerten. 28S rDNA Im Datensatz der partiellen 28S rDNA-Sequenzen sind knapp 40% der Nukleotidpositionen konstant, das sind rund 10% weniger als bei der 18S rDNA. Insgesamt weist die 28S rDNA eine höhere Variabilität als die 18S rDNA auf, dabei fallen wiederum die Nudipleura und unter diesen die Cladobranchia auf. Bei Nichtberücksichtigung der Nudipleura steigt der Anteil der konstanten Positionen auf mehr als 60%. Bei der Untersuchung der Evolutionsgeschwindigkeit der einzelnen Taxa, zeigt sich ein ähnliches Bild wie auf der Grundlage der 18S rDNA-Sequenzen, d.h. die Unterschiede in den Substitutionsraten sind ebenfalls gruppenspezifisch, mit den höchsten verzeichneten Werten bei den Cladobranchia (Nudibranchia). Auch die unterschiedlichen Basenfrequenzen werden nicht als Risiko für die korrekte phylogenetische Rekonstruktion angesehen, da nur die Nudibranchia eine (in sich homogene) von den übrigen Taxa abweichende Basenzusammensetzung aufweisen. Die Analyse der Substitutionssättigung hat ergeben, dass beide Substitutionsklassen eine leichte Sättigung aufweisen. Genau wie bei der Analyse der 18S rDNA-Sequenzen ist bei Nichtberücksichtigung der Cladobranchia und von Berthella stellata eine lineare Zunahme von Transitionen und Transversionen bei steigender evolutionärer Distanz zu beobachten.

111
ITS1-5.8S rDNA-ITS2 Das Verhältnis von konstanten zu variablen Positionen erscheint in dieser DNA-Region günstiger als bei der 18S und 28S rDNA, da sich nur knapp über 30% aller Nukleotidpositionen als konstant erweisen. Von diesen konstanten Positionen liegt der weitaus größte Anteil in der konservierten 5.8S rDNA, dagegen zeichnen sich die internal transcribed spacer-Regionen durch eine extrem hohe Sequenzdivergenz aus. Die Analyse der Basenzusammensetzung ergab eine deutliche Inhomogenität der Basenfrequenzen innerhalb des Datensatzes, dabei konnte keine Taxonspezifität wie bei den vorhergehenden Datensätzen festgestellt werden. Die Ergebnisse der Stammbaumrekonstruktionen müssen also dahingehend geprüft werden, ob (falsche) Schwestergruppenverhältnisse auf Übereinstimmungen in der Basenverteilung zurückgeführt werden können. Die beiden Arten Bursatella lechii und Petalifera petalifera weisen gegenüber den meisten übrigen Arten eine höhere Evolutionsgeschwindigkeit auf, ein Schwestergruppenverhältnis der beiden Sequenzen könnte demnach durch eine long branch attraction verursacht worden sein. Bei der Untersuchung der Substitutionssättigung zeigen die Transitionen eine leichte Sättigung, die Transversionen akkumulieren linear mit der Zunahme der evolutionären Distanz. Unerwartet ist die gegenüber den Transitionen höhere Anzahl an Transversionen bei nahezu allen paarweisen Vergleichen. Diese Beobachtung wurde auch von Lydeard et al. (2000) bei der Untersuchung der 16S rDNA verschiedener Molluskentaxa gemacht. In vorherigen Analysen (Lydeard et al. 1997, 1998) der 16S rDNA bei Pleuroceriden („Prosobranchia“) konnte bis zu einer Sequenzdivergenz von etwa 20% eine größere Anzahl Transitionen festgestellt werden, bei zunehmenden Distanzen kehrten sich die Verhältnisse um. Die Autoren erklärten die beobachteten Ergebnisse mit einer Sättigung der Transitionen aufgrund großer Sequenzdivergenz. Tatsächlich ist in der graphischen Darstellung der korrigierten und absoluten Distanzen der ITS1-5.8S rDNA-ITS2 – Sequenzen ab einer Divergenz von etwa 20% eine deutliche Zunahme der Transversionen gegenüber den Transitionen festzustellen, wohingegen beide Substitutionsklassen bei Werten unter 20% etwa gleiche Häufigkeiten aufweisen. Möglicherweise sind also die beim 16S rRNA-Gens bezüglich der Substitutionssättigung gemachten Beobachtungen auch auf andere Gene übertragbar. Da spacer-Regionen keinem oder nur einem sehr geringen Evolutionsdruck unterliegen, zeichnen sie sich durch sehr hohe Substitutionsraten aus. Obwohl alle Gene für die RNA der Ribosomen und die spacer-Regionen in einem Individuum gemeinsam evolvieren (concerted evolution), kann es trotzdem zur Existenz von unterschiedlichen Kopien in einem Genom kommen und zwar immer dann, wenn die concerted evolution langsamer abläuft als die Speziation (Harris & Crandall 2000).

112
ITS1- und ITS2-Sequenzen bilden routinemäßig die Grundlage für phylogenetische Rekonstruktionen, die Untersuchung genetischer Variabilität und die Unterscheidung nah verwandter Arten. Obwohl das Vorhandensein von intragenomischer Variabilität in ITS1 und ITS2 bekannt ist, wird es in phylogenetischen Studien weitestgehend ignoriert. In Abhängigkeit von der taxonomischen Untersuchungsebene kann das zu fehlerhaften Ergebnissen führen. Harris & Crandall (2000) konnten z.B. durch Untersuchungen an Crustaceen zeigen, dass die intragenomische Variation der Individuen groß genug ist, um Analysen auf Populationsebene zu verhindern, die Verwandtschaft zwischen Arten konnte dagegen abgeschätzt werden werden. Die in der vorliegenden Arbeit festgestellte intragenomische Variabilität der 8 Klone von Dolabrifera brazieri (Anaspidea) liegt bei etwa 1,4%, dagegen kann eine Sequenzdivergenz von knapp 10% zwischen Dolabrifera brazieri und Dolabrifera dolabrifera beobachtet werden. Da die hier durchgeführten phylogenetischen Untersuchungen auf Gattungsebene stattfinden, ist ein negativer Einfluß der intragenomischen Variabilität auf die Rekonstruktion der Verwandtschafts- beziehungen innerhalb der Anaspidea auszuschließen. Das Alter, ab dem eine Auflösung von Verwandtschaftsbeziehungen mit Hilfe von ITS-Sequenzen nicht mehr möglich ist, wird auf etwa 40-60 Millionen Jahre geschätzt (Oliverio 2002), wobei aufgrund unterschiedlicher Substitutionsraten bei verschiedenen Taxa keine allgemeingültigen Aussagen gemacht werden können. Die Anaspidea sind, basierend auf Fossildaten (Tracey et al. 1994), vor ca. 165 Millionen Jahren entstanden, die Aplysiidae sind wahrscheinlich bedeutend jüngeren Ursprungs (ca. 25 Mio Jahre, Geiger & Young 1996, Valdés & Lozouet 2000). Da mit Ausnahme der Gattung Akera alle untersuchten Taxa zu den Aplysiidae gehören, erscheint die Untersuchung dieses DNA-Bereichs erfolgversprechend. 16S rDNA Die 16S rDNA-Alinierung zeigt mit etwa 25 % unter allen untersuchten Datensätzen den geringsten Anteil konstanter Positionen. Die Variabilität innerhalb der Alinierung ist, im Gegensatz zu den 18S und 28S rDNA-Datensätzen, bei allen untersuchten Taxa in etwa gleich. Die Nudipleura zeigen hier keine Auffälligkeiten. Sowohl die Basenzusammensetzung, als auch die Verteilung der Substitutionsraten ist relativ homogen, allein Clione limacina (Gymnosomata) weist eine erhöhte Substitutionsgeschwindigkeit auf.

113
Die Untersuchung der Substitutionssättigung ergab eine deutliche Sättigung der Transitionen, dabei ist in Übereinstimmung mit den ITS1-5.8S rDNA-ITS2 – Sequenzen eine höhere Anzahl Transversionen als Transitionen zu beobachten. Mögliche Gründe für diese Beobachtung wurden bereits dargelegt. Die festgestellte Sättigung der Transitionen kann möglicherweise zu Problemen bei der korrekten phylogenetischen Rekonstruktion führen, allerdings legen Untersuchungen nahe, dass das Problem der Sättigung in der Vergangenheit überbewertet worden ist und dass im Gegenteil das Fehlen von phylogenetischer Information viel stärker zu gewichten ist (Yang 1998). Nukleäre Pseudogene sind nicht-funktionelle Kopien von funktionellen Genen im Genom und unterscheiden sich mehr oder weniger deutlich von letzteren. In der Regel können diese Pseudogene aufgrund von Mutationen nicht in funktionelle Proteine oder rRNAs translatiert werden (Lewin 2000). Durch den geänderten Selektionsdruck in Pseudogenen akkumulieren Substitutionen viel schneller als in funktionellen Genen. Pseudogene aus lang zurückliegenden Duplikationsereignissen stellen meist kein Problem für phylogenetische Untersuchungen dar, denn sie werden durch ihre große Divergenz zu den funktionellen Genen als solche identifizierbar, wenn eine Amplifikation überhaupt noch möglich ist. Das Vorkommen von „jungen“ Pseudogenen kann dagegen unentdeckt bleiben und so zur Postulierung falscher Verwandtschaftshypothesen führen. Obwohl die Existenz von Pseudogenen bei der 16S rDNA seit fast 20 Jahren bekannt ist, wurden die für phylogenetische Analysen daraus resultierenden Probleme weitestgehend ignoriert. Pseudogene kommen dabei viel häufiger vor als allgemein erwartet (Harrison et al., 2001; Zhang et al. 2002; Zhang et al. 2003a, 2003b), doch bisher gab es noch keine Anzeichen für mitochondriale Pseudogene bei den Mollusca. Jüngste Untersuchungen (Hebert et al. 2003a, 2003b) der Verwandtschafts- beziehungen verschiedener Taxa auf der Grundlage des COI-Gens ergaben einige sehr fragwürdige Ergebnisse, z.B. die Polyphylie der Mollusca, die auf der Basis morphologischer Analysen und anderer Gene als gesichert angesehen wird. Möglicherweise beruhen diese unerwarteten Resultate auf der unbemerkten Einbeziehung von Pseudogenen der COI. Die untersuchten 16S rDNA-Klone eines Individuums von Unela glandulifera (Acochlidiacea) stimmten zu 99,6 % überein, die restlichen 0,4 % lassen sich wahrscheinlich auf Sequenzierungsfehler zurückführen. Um das Vorhandensein von Pseudogenen allerdings mit Sicherheit ausschließen zu können, hätte es weiterer Untersuchungen mit einer größeren Anzahl von Klonen und Individuen bedurft.

114
4.3 Maximum Parsimonie- (MP-) Analysen Im Vergleich mit den maximum likelihood- und Distanzmethoden zur phylogenetischen Rekonstruktion, weist das Maximum Parsimonie-Verfahren scheinbar den Vorteil auf, dass keine expliziten Modelle zum Prozeß der Sequenzevolution benötigt werden. Allerdings entsprechen die im Verfahren implizierten Annahmen zur Sequenzevolution, d.h. die Konstanz der Substitutionsraten der 4 Nukleotide, die Homogenität der Raten innerhalb des gesamten Genbereichs und gleiche Astlängen innerhalb des Baumes, nur in Ausnahmefällen dem realen Evolutionsgeschehen (Yang 1996). Die zentrale Frage bezüglich des Maximum Parsimonie-Verfahrens besteht darin, ob der sparsamste Baum, im Vergleich zu einem Stammbaum der mehr Schritte erfordert, der realen Phylogenie am besten entspricht. Es wird nämlich dadurch impliziert, dass evolutionäre Veränderungen an sich seltene Ereignisse darstellen (Page & Holmes 1998). Felsenstein (1973, 1978, 1988) hat darauf hingewiesen, dass die Parsimonie-Methode eine akzeptable Annäherung an das maximum likelihood-Verfahren darstellt, wenn die Menge an evolutiven Veränderungen gering ist und die Evolutionsraten sich als relativ konstant erweisen. Neuere Untersuchungen haben dagegen gezeigt, dass sogar die Existenz einer molekularen Uhr und geringe evolutive Veränderungen die statistische Konsistenz der Parsimonie-Methode nicht garantieren können. Durch die Einbeziehung weiterer Sequenzen konnte sogar eine Verschlechterung der Situation beobachtet werden (Hendy & Penny 1989, Zharkikh & Li 1993, Takezaki & Nei 1994). Vergleichende Untersuchungen der unterschiedlichen phylogenetischen Rekonstruktionsmethoden durch Computer-simulationen deuten darauf hin, dass die Fähigkeit, die korrekte Baumtopologie darzustellen, bei Anwendung des Maximum Parsimonie-Verfahrens oft geringer ist als bei Distanz- und maximum likelihood-Verfahren (Hasegawa & Fujiwara 1993, Huelsenbeck & Hillis 1993, Kuhner & Felsenstein 1994). Ein wesentlicher Nachteil des Maximum Parsimonie-Verfahrens liegt in dessen Unvermögen, unterschiedliche Astlängen, gleichbedeutend mit unterschiedlichen Substitutionsraten, bei der phylogenetischen Rekonstruktion zu berücksichtigen. Ein daraus resultierendes Problem ist unter dem Bergriff long branch attraction, im deutschen Sprachraum auch als Analogieproblem bekannt. Es kommt dabei aufgrund von zufälligen Übereinstimmungen durch hohe Substitutionsraten in zwei oder mehr Stammlinien zur Postulierung falscher Schwestergruppenverhältnisse zwischen den long branch-Taxa. Unter Berücksichtigung dieses Aspekts ist den Taxa mit signifikant erhöhten Substitutionsraten, z.B. Clione limacina im 16S rDNA-Datensatz, eine besondere Aufmerksamkeit zu schenken. Zu berücksichtigen ist allerdings, dass lange Stammlinien bei Schwestertaxa auch auf realen Verwandtschaftsbeziehungen beruhen können. Siddall & Whiting (1999) konnten zeigen, dass unerwartete monophyletische Gruppierungen in phylogenetischen Analysen oft vorschnell als das Ergebnis einer

115
long branch attraction betrachtet werden, ohne diese Ergebnisse einer näheren Untersuchung, z.B. einem Vergleich mit morphologischen Daten, zu unterziehen. Neben falscher Stützung von Gruppierungen durch long branch-Effekte, können auch Plesiomorphien Ursache für die Postulierung unglaubhafter Hypothesen sein. Dieser Fall kann eintreten, wenn eine falsche Gruppierung durch scheinbare Synapomorphien gestützt wird, die in Wirklichkeit Symplesiomorphien sind und aufgrund von multiplen Substitutionen nicht als solche erkannt werden konnten. Der Umgang mit Alinierungslücken, die durch Insertionen oder Deletionen in einer oder mehreren Sequenzen entstehen, stellt ebenfalls ein nicht zu vernachlässigendes Problem bei der phylogenetischen Rekonstruktion dar. Das verwendete Computerprogramm PAUP (Swofford 1999) ermöglicht die unterschiedliche Bewertung und Gewichtung von Alinierungslücken, entweder als „fehlende Information“ oder als „fünftes Nukleotid“, wobei sich die ermittelten Topologien deutlich unterscheiden können. Bei der Wahl der Bewertung gilt, den Informationsverlust durch die Nichtberücksichtigung der Lücken gegen eine unsichere Positionshomologie in den variableren Bereichen abzuwägen. Da die Sequenzbereiche mit sehr hoher Variabilität bei den phylogenetischen Rekonstruktionen nicht berücksichtigt wurden, erscheint die Wertung als zusätzliche Information gerechtfertigt. Einen weiteren kritischen Punkt bei Anwendung des Maximum Parsimonie-Verfahrens stellt die Gewichtung der Nukleotidpositionen nach ihrer Homologiewahrscheinlichkeit dar. Werden keine Gewichtungen vorgenommen, kann die Topologie eines Dendrogrammes durch wenige bedeutungslose (zufällige) Merkmale signifikant verändert werden, allerdings ist die Beurteilung von Merkmalen als „wertvoll“ oder „weniger wertvoll“ oft subjektiv und daher kritisch zu beurteilen. Da durch Ausschluß der hochvariablen Bereiche der Einfluß von zufälligen Merkmalen weitgehend minimiert wurde, ist auf eine Gewichtung der Merkmale verzichtet worden. 4.4 Distanzanalysen Distanzanalysen haben gegenüber Maximum Parsimonie- und besonders maximum likelihood-Verfahren den Vorteil, sehr schnelle Berechnungen zu ermöglichen. Sie eignen sich besonders für die Untersuchung konservierter Genbereiche und nicht zu langer Divergenzzeiten der Taxa, da die zugrundeliegenden Modelle der Sequenzevolution den realen Prozessen immer weniger gerecht werden können, je mehr Zeit verstreicht und je entfernter Taxa verwandt sind. Ein grundsätzlicher Nachteil von Distanzanalysen liegt in dem Informationsverlust, den die Transformation der Sequenzdaten in eine Distanzmatrix mit sich bringt.

116
Dieser Informationsverlust wird darin ersichtlich, dass einer Distanzmatrix zahlreiche unterschiedliche Sequenzdatensätze zugrunde liegen können. Eine weitere Einschränkung besteht darin, dass triviale Merkmale und Plesiomorphien, die nur bei einem Außengruppentaxon vorkommen, bei der Aufstellung einer Distanzmatrix mitberücksichtigt werden, wohingegen es die Anwendung des Maximum Parsimonie-Verfahrens erlaubt, die trivialen Merkmale auszuschließen. Während letztere Merkmale und zufällige Übereinstimmungen von Nukleotiden bei der Größe des 18S rDNA-Datensatzes als „Hintergrundrauschen“ eher vernachlässigt werden können, führen Plesiomorphien häufig in die sog. „Symplesiomorphiefalle“, d.h. die falsche Stützung von Gruppen durch Plesiomorphien kann zur Postulierung unglaubhafter Verwandtschaftshypothesen führen. Ein bekanntes und nicht zu vernachlässigendes Problem bei der Durchführung von Distanzanalysen besteht in der Anfälligkeit der bestehenden Algorithmen für die Reihenfolge der eingelesenen Sequenzen, d.h. die Dendrogrammtopologie ist unter Umständen abhängig von der Reihenfolge der Sequenzen im untersuchten Datensatz. In der vorliegenden Arbeit konnte allerdings anhand von Tests festgestellt werden, dass die Topologie von einer veränderten Taxareihenfolge nicht beeinflusst wird. 4.5 maximum likelihood- (ML-) Analysen Die maximum likelihood-Methode der phylogenetischen Rekonstruktion wird von vielen Autoren als den anderen Verfahren überlegen betrachtet (Huelsenbeck & Hillis 1993, Kuhner & Felsenstein 1994, Huelsenbeck 1995a+b). Leider stößt seine Anwendbarkeit aufgrund des hohen Rechenaufwandes bei großen Datensätzen schnell an Grenzen. Die Notwendigkeit zur Berücksichtigung von Modellen zur Sequenzevolution ist unter dem Aspekt der korrekten phylogenetischen Rekonstruktion kontrovers zu beurteilen. Die notwendige Auswahl der Parameter zur Sequenzevolution macht die Annahmen bewusst, die den Stammbaumberechnungen zugrunde liegen und in anderen Verfahren nur implizit vorhanden sind. Andererseits birgt die Wahl eines Modells, genau wie bei den Distanzanalysen, Risiken und Schwierigkeiten die nicht exakt eingeschätzt werden können und unter Umständen zu einer fehlerhaften Interpretation der Ergebnisse führen. Sehr einfache Modelle tragen dem realen Evolutionsgeschehen oft nur ungenügend Rechnung, bei komplexen Modellen kann es dagegen leicht zu einer Akkumulation von Ungenauigkeiten bei der Abschätzung der Parameter kommen. Die allgemeine Auffassung, dass die Wahl eines nicht zutreffenden Modells oft zur Postulierung nicht korrekter Verwandtschaftsverhältnisse führt, ist dagegen aber nicht zutreffend. Yang et al. (1994, 1995) und Yang (1996) konnten anhand von Computersimulationen zeigen, dass die Likelihood-Werte der Baumtopologien bei Anwendung eines „falschen“ Modells zwar insgesamt sinken, die Wahrscheinlichkeit die reale Topologie

117
darzustellen aber sogar steigt. Zu übereinstimmenden Ergebnissen kommen Schoeninger & von Haeseler (1993), sowie Tajima & Takezaki (1994) bei der Untersuchung von Distanzmethoden. 4.6 Bayes’sche Analysen Genau wie bei den maximum likelihood- und Distanzverfahren muß hier ein Modell zur Sequenzevolution ausgewählt werden, mit allen damit verbundenen Vor- und Nachteilen. Obwohl MCMC (Markov chain Monte Carlo) die Analyse von sehr komplexen Modellen möglich gemacht hat, ist es kein Patentrezept, da es Markov-Ketten unter bestimmten Umständen nicht möglich ist, sich der posterior probability distribution zu nähern (Huelsenbeck et al. 2001). Um dieses Problem zu minimieren, werden mehrere Ketten parallel durchgeführt. Zudem ist es schwierig zu bestimmen, wie lange Markov-Ketten laufen müssen, um eine vernünftige Annäherung an die Aposteriori-Wahrscheinlichkeitsverteilung zu repräsentieren. Ein bedeutender Vorteil einer Bayes’schen Analyse liegt in den sehr geringen Rechenzeiten, die darauf beruhen, dass nicht der optimale Baum unter unendlich vielen möglichen gesucht wird, sondern die Bäume entsprechend ihrer posterior probability gesammelt werden. Diese Stichprobe kann dann dazu verwendet werden, um eine Konsensustopologie zu erstellen. Trotz dieser Effizienz entspricht das Verfahren qualitativ einer maximum likelihood-Analyse mit anschließendem bootstrap-Test (Huelsenbeck et al. 2001). Zudem sind posterior probabilities wesentlich leichter zu interpretieren als bootstrap-Werte, da die Aposteriori-Wahrscheinlichkeit eines Astes die Wahrscheinlichkeit angibt, mit der das zugehörige Taxon der realen Phylogenie (auf der Grundlage des Datensatzes und des gewählten Modells der Sequenzevolution) entspricht. bootstrap-Werte erlauben dagegen nur Aussagen über die Übereinstimmung von Topologie und zugrunde liegenden Daten. Ein Vorteil gegenüber dem Maximum Parsimonie-Verfahren besteht darin, dass nicht nur ein evolutionäres Szenario betrachtet wird, nämlich das mit den geringsten evolutiven Veränderungen, sondern viele mögliche, die ihrer Wahrscheinlichkeit nach gewichtet werden (Huelsenbeck et al. 2001). Die in den hier dargestelltenTopologien zu beobachtende geringe Auflösung stellt scheinbar einen gravierenden Nachteil der Bayes’schen Methode dar. Vielleicht ist aber gerade das ihre Stärke, da sie uns vor Augen führt, daß zu wenig phylogenetisches Signal in den Datensätzen vorhanden ist, um die zu untersuchenden Verwandtschaftsverhältnisse aufzulösen.

118
Kontrovers zu beurteilen ist, dass mit dem Bayes’schen Ansatz phylogenetische a priori-Informationen berücksichtigt werden können. Die Einbeziehung zusätzlicher Daten an sich ist sicherlich positiv zu bewerten, allerdings ist völlig unklar in welcher Form phylogenetische Ergebnisse vorheriger Analysen mit einfließen können. Außerdem wird die Bestimmung der a priori-Informationen teilweise als zu subjektiv und damit unwissenschaftlich angesehen (Huelsenbeck et al. 2002). Das Resultat dieser Problematik ist, daß bei phylogenetischen Analysen bisher alle Bäume a priori als gleichwahrscheinlich betrachtet wurden. 4.7 Statistische Tests Relative rate test Die korrekte Einschätzung der relativen Substitutionsraten ist abhängig von der Wahl des Referenztaxons und es sollte nicht übersehen werden, dass unterschiedliche Außengruppen zu unterschiedlichen Ergebnissen führen können. Weiterhin gilt es zu berücksichtigen, dass das im Programm K2WuLi implementierte Sequenzevolutionsmodell (K2P) die reale Sequenzevolution unter Umständen nur unzureichend widerspiegelt und Distanzwerte dadurch über- oder unterschätzt werden. Verschiedene Autoren betrachten es als unwahrscheinlich, daß mit Hilfe von relative rate tests moderate Level von Ratenvariabilität aufgedeckt werden können (Muse & Weir 1992, Bromham et al. 2000). Das könnte der Fall beim 16S rDNA-Gen sein, denn die zu beobachtende Variabilität der Astlängen korrespondiert nicht mit signifikanten Unterschieden bei der Analyse der Substitutionsraten. bootstrap-Analyse Im Gegensatz zu den posterior probabilities einer Bayes’schen Analyse sind bootstrap-Werte einer intuitiven Interpretation nicht so einfach zugänglich. Mit Hilfe einer bootstrap-Analyse können weder Aussagen über die Qualität der verwendeten Merkmale getroffen werden, noch kann das Fehlen von Information oder die Häufung von Analogien erkannt werden. Es ist lediglich möglich zu beurteilen, inwieweit eine Topologie mit der zugrunde liegenden Datenstruktur übereinstimmt. Hohe bootstrap-Werte können z.B. auch dann auftreten, wenn eine Gruppierung nur durch sehr wenige Apomorphien gestützt wird, es aber für alternative Gruppierungen keine Merkmale gibt, so dass nur ein einziges Merkmal ausreicht, um die Gruppe wiederzufinden (Wägele 2000).

119
4.8 Phylogenetisches Signal Beim Vergleich der Datensätze für die Untersuchung der Opisthobranchia zeigt sich, dass der untersuchte Abschnitt der 16S rDNA quantitativ das stärkste phylogenetische Signal aufweist, bei der 18S rDNA werden die einzelnen Gruppierungen, bezogen auf die Sequenzlänge, durch die geringste Anzahl potentieller Apomorphien gestützt. Bezüglich der Anaspidea erweisen sich die ITS1-5.8S rDNA-ITS2 - Sequenzen gegenüber denen der 18SrDNA als wesentlich informativer. Eine mögliche Ursache für das Fehlen phylogenetischer Information kann eine zu hohe Evolutionsgeschwindigkeit des untersuchten Gens sein, die dazu führt, dass die phylogenetischen Signale völlig verrauschen. Andererseits könnte bei einer zu langsamen Evolutionsrate phylogenetisches Signal möglicherweise gar nicht erst entstehen, so dass Gruppierungen allein auf zufälligen Übereinstimmungen der Taxa beruhen. Bei einer hohen rate heterogeneity among sites (positionsabhängige Variabilität der Substitutionsrate) könnte die Kombination aus beidem für das Fehlen eines deutlichen phylogenetischen Signals verantwortlich sein. Tatsächlich wird die Unfähigkeit der ribosomalen Gene viele Verwandtschaftsverhältnisse aufzulösen, auf eine ausgedehnte positionsabhängige Variabilität der Substitutionsraten zurückgeführt, die die Anzahl phylogenetisch informativer Positionen signifikant reduziert (Olsen & Woese 1993). Die positionsabhängige Variabilität der Substitutionsraten kann mit Hilfe der sog. Gamma-Verteilung beschrieben werden. Diese Verteilung hat den Formparameter a, der das Ausmaß der Ratenvariabilität spezifiziert. Sehr kleine a-Werte resultieren in einer Verteilung mit einer sehr hohen Variabilität, d.h. die meisten Nukleotidpositionen sind konstant, und einige wenige weisen eine sehr hohe Substitutionsrate auf. Die untersuchten Datensätze zeigen a-Werte von 0,35 (18S rDNA, Opisthobranchia–Datensatz) bis 0,64 (16SrDNA, Opisthobranchia-Datensatz; ITS1-2, Anaspidea-Datensatz) und damit eine sehr positionsspezifische Ratenvariabilität. Dabei ist eine Korrelation zwischen der Ratenheterogenität und der relativen Anzahl der potentiellen Apomorphien festzustellen, d.h. der Datensatz mit der größten Variabilität der Substitutionsraten weist bei allen Taxa die geringste relative Anzahl stützender Positionen auf. 4.9 Phylogenie der Opisthobranchia 4.9.1 Wahl der Außengruppe Um die Phylogenie einer Gruppe von Taxa richtig darstellen zu können, ist es von entscheidender Bedeutung, dass die gewählte Außengruppe aus phylogenetischer Sicht nicht Teil der Innengruppe ist. Zum anderen sollte die Außengruppe evolutiv nicht zu weit von der Innengruppe entfernt sein, damit die untersuchten Merkmale, in

120
der molekularen Phylogenetik sind das die 4 Nukleotide, noch eindeutig homologisierbar sind. Weiterhin sollte ein Außengruppentaxon nicht zu abgeleitet sein, damit plesiomorphe Merkmale nicht durch autapomorphe „verdeckt“ werden. Nach dem Heterobranchia-Konzept (Haszprunar 1985a) erscheint ein Vertreter der Allogastropoda (Architectonicidae, Pyramidellidae, Mathildidae) als Außengruppe für eine Innengruppe aus Opisthobranchiern und Pulmonaten am besten geeignet. Dem wurde in früheren Analysen (Vonnemann et al. 2005) durch die Verwendung von Odostomia sp. (Pyramidellidae) Rechnung getragen. In der vorliegenden Arbeit wurde dieses Taxon allerdings nicht mehr als Außengruppe verwendet, da eine Stellung der Pyramidellidae innerhalb der Euthyneura, oder sogar innerhalb der Opisthobranchia, nicht mit Sicherheit ausgeschlossen werden kann (Grande et al. 2004a,b). Zum anderen weisen die Pyramidellidae als hochspezialisierte Ektoparasiten auf morphologischer Ebene zahlreiche autapomorphe Merkmale auf, die sich möglicherweise auch auf molekularer Ebene manifestiert haben. Ein Taxon, das mit Sicherheit nicht innerhalb der Euthyneura gruppiert, stellen die unspezialisierten Littorinidae dar. Obwohl die Littorinidae im Vergleich zu den Euthyneura getrenntgeschlechtlich sind ist Gosliner (1981) der Auffassung, daß das Reproduktionssystem bei Vertretern der Littorinacea weitestgehend identisch mit dem von ursprünglichen Pulmonata und Opisthobranchia ist und zahlreiche weitere Übereinstimmungen bei Littorinidae und Euthyneura zu beobachten sind. Insgesamt sind die Littorinacea dasjenige Taxon der „Mesogastropoda“, das die meisten ancestralen Merkmale beibehalten hat. In Verbindung mit paläontologischen Befunden legen die Ergebnisse nahe, dass die Littorinacea ein mögliches Modell eines euthyneuren Vorfahren darstellen (Gosliner 1981). Unabhängig von der Wahl der Außengruppe ist bei einem Außengruppenvergleich immer zu beachten, daß rezente Taxa niemals die reale Ahnengruppe anderer rezenter Taxa darstellen können und das Innen- und Außengruppe unabhängig voneinander verschiedene Autapomorphien entwickeln, die die Unterscheidung plesiomorpher und apomorpher Merkmale erschweren kann. 4.9.2 Status der Opisthobranchia, Pulmonata und Euthyneura In den erstellten Verwandtschaftshypothesen, basierend auf dem 16S rRNA-, 18S rRNA- und 28S rRNA-Gen, erscheinen weder die Opisthobranchier, noch die Pulmonaten und die Euthyneura als monophyletische Gruppierungen, da die Pulmonaten innerhalb der Opisthobranchia gruppieren und die Pyramidellidae innerhalb der Euthyneura. Dieses evolutionäre Szenario erscheint unglaubwürdig, da auf der Basis morphologischer Merkmale zumindest die Monophylie der Pulmonata (z.B. Salvini-Plawen & Steiner 1996, Dayrat & Tillier 2002) und der Euthyneura (z.B. Haszprunar 1985a, 1988a, Salvini-Plawen & Steiner 1996, Dayrat & Tillier 2002) als gut gestützt angesehen wird. Haszprunar nennt z.B. eine ventrale Ganglienkette aus 5 Ganglien als stärkste Synapomorphie für die Euthyneura und schlägt die

121
Bezeichnung Pentaganglionata vor, da er die Euthyneurie als konvergent entstanden betrachtet. Auch molekulare Analysen sprechen für die Monophylie der Euthyneura (z.B. Wade & Mordan 2000, Dayrat et al. 2001). Die Opisthobranchia erscheinen in den meisten Untersuchungen auf der Basis morphologischer Merkmale als monophyletische Gruppierung, in molekularen Analysen sind sie dagegen überwiegend paraphyletisch. Tatsächlich widersprechen die meisten molekularen Analysen sowohl der Monophylie der Opisthobranchia als auch der der Pulmonata (z.B. Tillier & Ponder 1992, Rosenberg et al. 1994, Tillier et al. 1994, 1996, Winnepenninckx et al. 1998, Yoon & Kim 2000, Dayrat et al. 2001, Grande et al. 2004a,b). Auf der Grundlage der mitochondrialen Genanordnung konnte keine der drei Gruppierungen (Euthyneura, Pulmonata, Opisthobranchia) als monophyletisch erkannt werden (Grande et al. 2004a). Die Ergebnisse der vorliegenden Arbeit unterstützen übereinstimmend die Paraphylie der drei Gastropodengruppen. Um die Phylogenie der gesamten Heterobranchia aufzuklären müssten allerdings viel mehr Taxa in die Analysen einbezogen werden, sowohl aus den Pulmonaten, als auch aus den Allogastropoda, z.B. den Architectonicidae. Im Anschluß werden die Phylogenie der einzelnen Großgruppen der Euthyneura und Allogastropoda, sowie ihre möglichen Verwandtschaftsbeziehungen zu anderen Taxa zusammengefasst und diskutiert. 4.9.3 Pulmonata Zu den berücksichtigten Taxa der Pulmonata gehören die Ellobiidae (18S rDNA: 5 Sequenzen, 16S rDNA:1), die Systellomatophora (18S und 28S rDNA: jeweils 2 Sequenzen), die „Basommatophora“ (18S und 28S rDNA: jeweils 2 Sequenzen) und die Stylommatophora (18S und 28S rDNA: jeweils 1 Sequenz). Ellobiidae Die Ellobiidae (Cassidula nucleus, Ovatella firminii, Pythia scarabaeus, Melampus luteus, Auriculinella bidentata) erscheinen in den Bäumen auf der Grundlage der 18S rDNA als paraphyletische Gruppierung, ein übereinstimmendes Ergebnis haben Dayrat et al. (2001) basierend auf partiellen 28S rDNA-Sequenzen erzielt. In allen erzeugten Topologien werden Pythia scarabaeus, Cassidula nucleus und Ovatella firminii als monophyletische Gruppierung rekonstruiert, dabei repräsentieren die beiden letztgenannten Arten Schwestertaxa. Die Schwestergruppe der drei Sequenzen stellen entweder Melampus luteus (Distanzanalyse), Auriculinella bidentata (MP-Analyse) oder die Acochlidiacea (Bayes’sche Analyse) dar.

122
Die taxonomische Gliederung der Ellobiidae basiert im Wesentlichen auf dem Vergleich des Geschlechts- und Nervensystems, dabei werden dabei 5 Unterfamilien unterschieden, entsprechend den 5 Organisationstypen des Reproduktionssystems (Morton 1955). Von diesen fünf Gruppierungen sind drei im 18S rDNA-Datensatz vertreten. Die hier monophyletischen Pythiinae (Pythia scarabaeus, Ovatella firminii, Cassidula nucleus) werden von de Frias Martins (1996) auf der Basis morphologischer Untersuchungen als Kerngruppe betrachtet, von der aus sich die Ellobiinae (Auriculinella bidentata), Melampinae (Melampus luteus) und die übrigen beiden Unterfamilien unabhängig entwickelt haben. Die Ellobiinae müssen sich stammesgeschichtlich sehr früh von der Ursprungsgruppe getrennt haben, da sie das ursprünglichste Nervensystem beibehalten haben. Auriculinella bidentata gruppiert in zwei Topologien nicht mit anderen Ellobiidae, was möglicherweise als Hinweis für eine große Sequenzdistanz zu den übrigen Ellobiiden gedeutet werden kann. Die Melampinae weisen auf der Grundlage morphologischer Daten die meisten abgeleiteten Merkmale innerhalb der Ellobiidae auf (de Frias Martins 1996), eine entsprechende Stellung innerhalb der Topologien konnte aber nicht beobachtet werden. Die Ellobiidae wurden traditionell als lebende Vertreter der ursprünglichsten Pulmonaten angesehen und gleichzeitig mit ursprünglichen Opisthobranchiern in Verbindung gebracht (de Frias Martins 1996). Nach Meinung einiger Autoren stellen sie wahrscheinlich eine paraphyletische Gruppe an der Basis der Eupulmonata (Ellobiidae, Onchidiidae, Soleolifera, Stylommatophora) dar (z.B. Dayrat & Tillier 2003). Bei Grande et al. (2004a) nehmen sie dagegen, basierend auf mitochondrialen Genen, eine ganz basale Position innerhalb der Euthyneura (inkl. Pyramidella, Allogastropoda) ein. Die Stellung der Ellobiidae in den rekonstruierten Stammbäumen ist sehr variabel. Im Baum der MP-Analyse (18S rDNA) bilden sie ein Monophylum mit den Onchidiidae, Siphonariidae, Stylommatophora, Acochlidiacea, Tylodinoidea, Cephalaspidea, Anaspidea und Pteropoda, im 18S rDNA-Distanzbaum mit den Acochlidiacea, Pyramidellidae, Tylodinoidea, „Basommatophora“ und Sacoglossa. Die Bayes’sche Analyse ergab wegen geringer Auflösung nur ein Schwestergruppenverhältnis von Pythiinae und Acochlidiacea, sowie die Monophylie von Auriculinella und Tylodinoidea, Pyramidellidae, Lymnaea stagnalis und Sacoglossa. Auf der Grundlage der 16S rDNA-Alinierung, die nur eine einzige Ellobiiden-Sequenz enthält (Pythia scarabaeus), erscheinen die Ellobiidae übereinstimmend als Schwestergruppe der Acochlidiacea. Die einzige Übereinstimmung zwischen den Topologien auf der Basis beider Gene besteht in einer näheren Verwandtschaft der Ellobiidae mit den Acochlidiacea. Mit Hilfe des Programms PHYSID (Wägele & Rödding 1998) konnten für das hypothetische Monophylum aus Acochlidiacea und Pythia scarabaeus 25 potentielle Apomorphien auf der Grundlage des 16S rDNA-Datensatzes festgestellt werden. Allerdings konnten ebenso viele oder sogar mehr stützende Positionen für alternative Gruppierungen gefunden werden, z.B. 27 potentielle Apomorphien für P. scarabaeus und Sacoglossa oder 28 Positionen für P. scarabaeus und Anaspidea. Diese

123
Beobachtungen lassen sich auf das Fehlen eines deutlichen phylogenetischen Signals zurückführen, ein enges Verwandtschaftsverhältnis zwischen Ellobiidae und Acochlidiacea ist daher fragwürdig. Die verwandtschaftlichen Beziehungen der Ellobiidae zu anderen euthyneuren Taxa bleiben damit offen, ebenso wie der Status der Gruppe als Mono- oder Paraphylum. Allein die Monophylie der Pythiinae kann als relativ gesichert angesehen werden. Systellomatophora Die in den 18S und 28S rDNA-Datensätzen enthaltenen Vertreter der Systellomatophora oder Gymnomorpha, Onchidium verrucosum und Onchidiella floridana (Onchidiidae), bilden in allen konstruierten Bäumen eine monophyletische Einheit. Die Monophylie der Systellomatophora gilt, auf der Basis molekularer (Dutra-Clarke et al. 2001) und morphologischer Analysen (Haszprunar 1985a) als gut gestützt, dagegen werden ihre verwandtschaftlichen Beziehungen zu anderen Taxa recht kontrovers diskutiert. In der Vergangenheit wurde sowohl die Stellung der Onchidiidae innerhalb der Opisthobranchier (Fretter 1943, Boettger 1954) als auch innerhalb der Pulmonaten (Ghiselin 1966) diskutiert. Nach Boettger (1954) bilden die Onchidiidae eine monophyletische Gruppierung mit den Tylodinoidea, Pleurobranchoidea und Nudibranchia. Im Gegensatz zu Boettgers Meinung betrachten die meisten Systematiker die Systellomatophora heute als Pulmonaten (z.B. Delhaye & Bouillon 1972, Solem 1979), bei Tillier et al. (1996, molekulare Daten) und Dayrat et al. (2001, morphologische Daten) stellen sie z.B. die Schwestergruppe der Stylommatophora dar. Alternativ zur Stellung innerhalb der Opisthobranchia oder Pulmonata werden die Systellomatophora als eigenständige evolutive Linie angesehen (Salvini-Plawen 1970, Haszprunar 1985a). In den rekonstruierten Verwandtschaftshypothesen auf der Grundlage der 18S rDNA, bilden die Onchidiidae ein Monophylum mit Siphonaria alternata („Basommatophora“), wobei dieses Schwestergruppenverhältnis im Baum der Bayes’schen Analyse wegen zu geringer Auflösung nicht gefunden werden konnte. In den Topologien basierend auf dem 28S rRNA-Gen gruppieren die Onchidiidae entweder mit Cepaea hortensis (Stylommatophora, Bayes’sche und ML-Analyse) oder ebenfalls mit Siphonaria alternata (Distanz-, MP-Analyse), dabei konnte durch bootstrap-Analysen nur die Monophylie der Onchidiidae gestützt werden. Auf der Grundlage mitochondrialer Gene erscheint Onchidiella celtica bei Grande et al. (2004b) als Schwestergruppe zu einer Vereinigung aus Opisthobranchia, Siphonaria pectinata und Pyramidella dolabrata (Allogastropoda), Dayrat et al. (2001) rekonstruierten auf der Grundlage partieller 28S rDNA-Sequenzen ein Monophylum aus Ellobiiden und Onchidium striatum.

124
In der vorliegenden Arbeit stehen die beiden Vertreter der Systellomatophora zwar immer in einem Schwestergruppenverhältnis mit pulmonaten Arten, das gesamte Monophylum gruppiert aber innerhalb der Opisthobranchia. Die verwandtschaftlichen Beziehungen der Onchidiidae bleiben somit weiterhin unklar. Siphonaria alternata und Lymnaea stagnalis Die beiden Arten werden in die als paraphyletisch erkannten “Basommatophora” eingeordnet, dabei repräsentiert die marine Gattung Siphonaria den wahrscheinlich ursprünglichsten Vertreter dieser Gruppierung (Haszprunar 1985a), wohingegen Lymnaea (limnisch) als eher abgeleitet angesehen wird. Die taxonomische Position der Siphonarioidea innerhalb der „Basommatophora“ hat in der Vergangenheit reichlich Anlaß zur Diskussion gegeben (siehe dazu z.B. Köhler 1893, Haszprunar & Huber 1990, Nordsieck 1992). Jüngste Untersuchungen der „Basommatophora“ (Tillier et al. 1996, Yoon & Kim 2000, Dutra-Clarke et al. 2001), basierend auf 18S und 28S rRNA-Genen, zeigen eine deutliche Zweiteilung der Gruppe in marine Formen einerseits und Süßwasserformen andererseits. Dabei zeigen die marinen Taxa wie die Siphonariidae Tendenzen zur Verwandschaft mit den Opisthobranchia und Systellomatophora, die limnischen dagegen mit den Stylommatophora (Dutra-Clarke et al. 2001). Im Gegensatz dazu stellt Siphonaria in der Analyse von Wade & Mordan (2000) die Schwestergruppe zu den übrigen untersuchten Pulmonaten dar. Die in der vorliegenden Arbeit durchgeführten phylogenetischen Analysen unterstützen klar die Verwandtschaft der Siphonariidae zu den Systellomatophora und Opisthobranchia. In Anbetracht derselben zugrundeliegenden Genbereiche, erscheint diese Übereinstimmung aber nicht verwunderlich und es bedarf der Analyse alternativer Daten, um die postulierten Verwandtschaftsbeziehungen zu bewerten. Lymnaea stagnalis erscheint in den Topologien auf der Basis des 18S rDNA-Datensatzes als Schwestergruppe der Sacoglossa (keine Auflösung im Baum der Bayes’schen Analyse), basierend auf den partiellen 28S rDNA-Sequenzen ist eine nahe Verwandtschaft zu den Sacoglossa, Tylodinoidea, Acochlidiacea und den übrigen Pulmonaten zu beobachten. Die Stellung von Lymnaea als Schwestergruppe zu Cepaea hortensis (Distanzanalyse) stützt die Ergebnisse der Untersuchungen von Tillier et al. (1996), Yoon & Kim (2000) und Dutra-Clarke et al. (2001). Zu berücksichtigen ist allerdings wiederum, dass dieselben Gene verwendet wurden. Die Verwandtschaft von Lymnaea zu den Sacoglossa erscheint fragwürdig, doch phylogenetische Analysen auf der Grundlage der 16S rDNA (Thollesson 1999) ergaben ebenfalls die Monophylie von Sacoglossa + Pulmonata.

125
Cepaea hortensis Die Stellung der Landlungenschnecke Cepaea hortensis ist in den erzeugten Verwandtschaftshypothesen recht variabel. So zeigt sie Beziehungen zu den Siphonariidae und Onchidiidae, den Cephalaspidea, Lymnaea und Odostomia. Für eine Verwandtschaft mit den Onchidiidae spricht das Ergebnis einer Analyse von Dayrat & Tillier (2002) auf der Basis morphologischer Daten, die die Monophylie von Stylommatophora + Systellomatophora ergab. Eine engere Beziehung zu Lymnaea unterstützen die molekularen Analysen von Tillier et al. (1996), Yoon & Kim (2000) und Dutra-Clarke et al. (2001). 4.9.4 Pyramidellidae Die Pyramidellidae sind hochspezialisierte Ektoparasiten sessiler Polychaeten, Mollusken und Echinodermaten, die in Anpassung an ihre Lebensweise die Radula reduziert haben (Fretter & Graham 1949, Fretter et al. 1986). Sie werden entweder als Schwestergruppe der Euthyneura angesehen bzw. in die „niederen“ Heterobranchia eingeordnet (z.B. Gosliner 1981, Haszprunar 1985a, Salvini-Plawen & Steiner 1996) oder als Angehörige der Euthyneura aufgefasst (z.B. Fretter & Graham 1949, Thompson 1976, Dayrat & Tillier 2002). Die postulierte Zugehörigkeit der Pyramidelliden zu den Euthyneura ist fragwürdig, da die als vereinend betrachteten Merkmale nur eine oberflächliche Ähnlichlichkeit aufweisen oder symplesiomorphen Charakters sind (Haszprunar 1985a). Auf der Basis molekularer Analysen scheint die Einbeziehung der Pyramidellidae in die Euthyneura dagegen gerechtfertigt (Grande et al. 2004a, basierend auf 16S und partial COI). Sekundärstruktur-Untersuchungen der 16S rRNA scheinen diese Ergebnisse durch die übereinstimmende Existenz eines Deletionsereignisses in beiden Gruppen zu belegen (Schander 1997). In den rekonstruierten Stammbäumen auf der Grundlage von 16S, 18S und 28S rDNA-Sequenzen gruppieren die Pyramidellidae ausnahmslos innerhalb der Euthyneura. Sie zeigen dabei in Abhängigkeit vom zugrundeliegenden Datensatz und der verwendeten Rekonstruktionsmethode eine engere Verwandtschaft mit Vertretern der Tylodinoidea, Sacoglossa, Acochlidiacea und Pulmonata. Aufgrund der geringen Anzahl berücksichtigter Pulmonaten- und Allogastropodentaxa, können keine gesicherten Aussagen über die phylogenetische Stellung der Pyramidellidae gemacht werden.

126
4.9.5 Opisthobranchia 4.9.5.1 Nudibranchia Auf der Grundlage des 18S rDNA- und des 28S rDNA-Datensatzes, erscheinen die Nudibranchia in allen durch die verschiedenen Rekonstruktionsverfahren erzeugten Bäumen, mit Ausnahme des MP-Baumes der 28S rDNA, übereinstimmend als monophyletische Gruppierung. Basierend auf dem 16S rRNA Gen werden sie dagegen als Polyphylum rekonstruiert. Die Polyphylie in den Topologien des 16S rRNA-Gens lässt sich entweder auf die Stellung von Dendrodoris nigra (MP-, Distanz-, Bayes’sche Analyse), oder von Bathydoris clavigera zurückführen (ML-Analyse). Diese beiden Arten gruppieren mit unterschiedlichen anderen Taxa des Datensatzes, z.B. bildet Dendrodoris nigra im MP-Baum ein Monophylum mit Pupa solidula und Hydatina physis (Acteonoidea), im Distanzbaum mit Cavolinia sp. (Thecosomata) und Odostomia sp. (Pyramidellidae). Obwohl mit Hilfe des relative rate test nur bei Clione limacina (Gymnosomata) eine signifikant erhöhte Substitutionsgeschwindigkeit festgestellt werden konnte (vergleiche dazu Kap. 4.7), fallen eher andere Arten in den Phylogrammen durch besonders lange Äste auf. Zu diesen gehören vor allem Dendrodoris nigra, Cavolinia sp. und Odostomia sp. Die abweichende Stellung von Dendrodoris kann demnach, zumindest in einigen Fällen, mit einiger Wahrscheinlichkeit auf eine long branch attraction zwischen Taxa mit erhöhten Substitutionsraten zurückgeführt werden. Doch auch bei Nichtberücksichtigung der „aberranten“ Stellung von Dendrodoris nigra und Bathydoris clavigera bleiben die Nudibranchia paraphyletisch, da die Pleurobranchoidea in allen erzeugten Topologien innerhalb der Nudibranchia gruppieren. Übereinstimmende Resultate erbrachten Analysen von Thollesson (1999a) und Grande et al. (2004a,b), ebenfalls auf der Grundlage der 16S rDNA, bzw. 16S rDNA und COI. In den von Grande et al. präsentierten Stammbäumen stellen die Pleurobranchoidea allerdings die Schwestergruppe der Anthobranchia dar, und nicht, wie in den eigenen Bäumen und bei Thollesson, die der Cladobranchia. Auf Grundlage der 18S und 28S rDNA-Sequenzen erscheinen die Nudibranchia monophyletisch, das entspricht sowohl den Ergebnissen anderer Analysen derselben Gene (Wollscheid-Lengeling et al. 2001, Wägele et al. 2003), als auch denen morphologischer Untersuchungen (Wägele & Willan 2000). Als synapomorphes Merkmal für die Nudibranchia nennen die letztgenannten Autoren z.B. die Existenz eines sog. Spezialvakuolenepithels. Die Analyse des phylogenetischen Signals erweist sich für die Interpretation der Beobachtungen als aufschlussreich. Während Im 18S rDNA-Datensatz 70 (davon 87% verrauscht) potentielle Apomorphien für die Nudibranchia und nur 66 (98% verrauscht) für das hypothetische Monophylum aus Cladobranchia und Pleurobranchoidea gefunden wurden, existieren im 16S Datensatz mehr stützende Positionen für letztere Gruppierung (36 > 25). Die abweichende Gruppierung von

127
Dendrodoris nigra mit Cavolinia sp. und Odostomia sp. weist z.B. 39 potentielle Apomorphien auf, im Vergleich zu nur 25 Positionen bei einer Stellung innerhalb der Nudibranchia. Die Monophylie der Nudibranchia ist wegen der widersprüchlichen Ergebnisse, sowohl zwischen molekularen und morphologischen Daten, als auch beim Vergleich unterschiedlicher Gene als wahrscheinlich, aber nicht restlos gesichert anzusehen. Die Stellung der Taxa innerhalb der Nudibranchia in den 18S und 28S rDNA-Bäumen entspricht den postulierten Verwandtschaftsverhältnissen auf der Basis morphologischer Merkmale zum Teil sehr gut. Drei Topologien (18S rDNA: MP-Analyse, 28S rDNA: ML- und Bayes’sche Analyse) zeigen eine völlige Übereinstimmung mit den morphologischen Daten von Wägele & Willan (2000): Die Anthobranchia (Chromodoris krohni, Hypselodoris villafranca, Dendrodoris fumata, Bathydoris clavigera und Cladobranchia (Dendronotus dalli, Cuthona nana, Flabellina babai) stellen Schwestergruppen dar, dabei gruppieren sowohl Chromodoris und Hypselodoris (Chromodorididae) zusammen, als auch Cuthona und Flabellina (Aeolidoidea). Innerhalb der Anthobranchia bildet Bathydoris das Schwestertaxon zu den übrigen Vertretern der Anthobranchia. Die Monophylie der Anthobranchia und Cladobranchia konnte auf der Grundlage verschiedener Gene (Wollscheid-Lengeling et al. 2001, Wägele et al. 2003), sowie auf der Basis morphologischer Merkmale (Wägele & Willan 2000, Valdés 2002) gezeigt werden. Die basale Stellung der Gattung Bathydoris innerhalb der Anthobranchia ist ebenfalls als gesichert anzusehen (z.B. Wägele & Willan 2000, Valdés 2002, Wägele et al. 2003). 4.9.5.2 Pleurobranchoidea Die Pleurobranchoidea erscheinen auf der Basis aller drei Datensätze und in allen rekonstruierten Topologien als monophyletische Gruppierung und werden durch hohe bootstrap-Werte (97-100) und posterior probabilities (0,94 -1,00) gestützt. Die Monophylie der Gruppe ist auf der Basis morphologischer Merkmale nie in Frage gestellt worden, als Apomorphien werden z.B. die rechteckige Schale und die Existenz einer Fußdrüse genannt (Willan 1987). In der molekularen Analyse von Grande et al. (2004) erscheinen die drei untersuchten Sequenzen ebenfalls als Monophylum. Die vorliegende Arbeit hat für die Pleurobranchoidea eine als gewichtig beurteilte Synapomorphie auf Sequenzebene aufgedeckt, und zwar eine lange Insertion (E23_3) im Bereich V4 der 18S rDNA. Die a priori-Analyse des phylogenetischen Signals (PHYSID, Wägele & Rödding 1998) ergab für die Pleurobranchoidea eine im Vergleich zu den meisten anderen Großgruppen der Opisthobranchia sehr hohe Anzahl stützender Positionen. Die Stellung der Taxa innerhalb der Pleurobranchoidea ist bei Vergleich der verschiedenen Datensätze und Rekonstruktionsverfahren sehr variabel. Aufgrund

128
morphologischer Daten werden gegenwärtig die beiden Familien Pleurobranchidae und Pleurobranchaeidae unterschieden (Willan 1987, Cervera et al. 2000), dabei gehören alle untersuchten Arten, bis auf Euselenops luniceps, zu den Pleurobranchidae. Bei Übereinstimmug mit der durch morphologische Daten begründeten Phylogenie, sollte Euselenops luniceps demnach die Schwestergruppe zu den übrigen Vertretern der Pleurobranchoidea darstellen. Das ist allerdings in keinem der ermittelten Dendrogramme der Fall. Die Stellung von Euselenops zeigt sich sehr variabel, sowohl innerhalb eines Datensatzes, als auch bei Vergleich der Alinierungen. Innerhalb der Pleurobranchidae unterscheidet Willan (1987) weiterhin das Taxon Berthellinae, mit den hier betrachteten Gattungen Bathyberthella, Berthella, Berthellina und Tomthompsonia (Cervera et al. 2000) und das Taxon Pleurobranchinae mit der einzigen Gattung Pleurobranchus. In den Analysen auf der Basis morphologischer Merkmale (Willan 1987, Cervera et al. 2000) gruppiert die Gattung Pleurobranchus als Schwestergruppe zu den übrigen Mitgliedern der Pleurobranchidae, deren Verwandtschaftsverhältnisse nicht aufgelöst werden konnten. Das genannte Schwestergruppenverhältnis konnte durch keine Topologie bestätigt werden. Tatsächlich erscheint Pleurobranchus in meisten Bäumen als sehr abgeleitet, dagegen repräsentiert die Gattung nach Willan (1987) die ursprünglichste innerhalb der Pleurobranchinae. Die Gattung Berthella zeichnet sich in den Bäumen auf der Grundlage der 18S und 28S rDNA durch sehr lange Äste aus, das stimmt mit der sehr hohen Substitutionsrate dieser Sequenzen überein. Dabei zeigt Berthella stellata gegenüber Berthella platei eine deutlich höhere Evolutionsgeschwindigkeit. In der Alinierung der 18S rDNA-Sequenzen wird diese durch eine extrem hohe Anzahl autapomorpher Merkmale, selbst in den hochkonservierten Bereichen des Gens, besonders augenfällig. Trotzdem erscheint die im 18S und 16S rDNA-Datensatz mit jeweils 2 Arten vertretene Gattung als Monophylum (Ausnahme: Bayes’sche Analyse, 18S rDNA). Willan (1987) hat basierend auf seinen morphologischen Untersuchungen vermutet, dass Berthella die Stammgruppe der Berthellinae bildet, von der ausgehend die übrigen Gattungen entstanden sind. Das erscheint bei Kenntnis der molekularen Daten wegen der großen Anzahl autapomorpher Merkmale unwahrscheinlich. Möglicherweise hat aber auch eine extrem hohe Substitutionsrate dazu geführt, dass vorhandene plesiomorphe Merkmale durch multiple Substitutionen überlagert worden sind. Insgesamt ist die Stellung der Taxa innerhalb der Pleurobranchoidea zu variabel, um postulierte Verwandschaftsverhältnisse bestätigen oder widerlegen zu können. Die hohe Variabilität lässt sich vermutlich auf das Fehlen eines deutlichen phylogenetischen Signals in allen drei Datensätzen zurückführen.

129
4.9.5.3 Nudipleura Das Schwestergruppenverhältnis von Nudibranchia und Pleurobranchoidea tritt, mit einer Ausnahme, in allen Verwandtschaftshypothesen auf der Grundlage des 18S und des 28S rRNA-Gens auf (bootstrap-Wert: 99, posterior probability: 0,99). Im Maximum Parsimonie-Baum auf der Grundlage des 28S rDNA-Datensatzes gruppieren die Cladobranchia als Schwestergruppe der Pleurobranchoidea, allerdings wird in 60% der bootstrap-Topologien die Monophylie der Nudibranchia rekonstruiert. Die Monophylie der Nudipleura wird ebenfalls durch die Analyse der 16S rDNA bestätigt, wenn man die gesonderte Stellung von Dendrodoris nigra auf eine long branch attraction zurückführt. Auf der Grundlage morphologischer Merkmale galt die Monophylie der Nudipleura bisher als gut gestützt (Schmekel 1985, Salvini-Plawen 1990, 1991, Wägele & Willan 2000, Mikkelsen 2002). Als synapomorph für diese Gruppierung interpretieren die Autoren die Existenz einer Blutdrüse, den Besitz eines androdiaulischen Reproduktionssystems und das Fehlen des Osphradiums. Die Blutdrüse kommt allerdings nicht bei allen Vertretern der Nudibranchia vor (Minichev 1970) und die genannte Ausprägung des Geschlechtssystems ist auch innerhalb der Acteonoidea und Sacoglossa zu finden (Ghiselin 1966, Mikkelsen 1996). Der Verlust des Osphradiums repräsentiert demnach die einzige bekannte Synapomorphie der Nudipleura. Da die vollständige Reduktion von Organen innerhalb der Opisthobranchia mehrfach unabhängig stattgefunden hat, ist es durchaus denkbar, dass auch es sich auch bei den Nudipleura um eine parallele Entwicklung handelt, die keinen Hinweis auf eine enge Verwandtschaft liefert. Auf der Grundlage molekularer Analysen ist die Monophylie der Nudipleura als recht gesichert anzusehen, bei Wägele et al. (2003) auf der Basis von 16S und 18S rDNA-Sequenzen, bei Grande et al. (2004a,b) auf der Grundlage verschiedener mitochondrialer Gene und bei Thollesson (1999a) basierend auf dem 16S rDNA-Gen. Dagegen bilden die beiden Vertreter der Nudipleura bei Dayrat et al. (2001) kein Monophylum. In dieser Arbeit wurde nur der Abschnitt D1 - D2 der 28S rDNA berücksichtigt, der im Vergleich zu dem hier untersuchten Bereich D1 - D3, anscheinend nicht genug phylogenetische Information enthält, um das erwähnte Schwestergruppenverhältnis aufzulösen. Möglicherweise ist auch die geringe Anzahl der nudipleuren Sequenzen für das von den anderen molekularen Analysen abweichende Ergebnis verantwortlich. Einen weiteren starken Beleg für die Monophylie der Nudipleura erbrachten Sekundärstrukturanalysen. Grande et al. (2004a) konnten die gemeinsame Existenz einer helical-loop structure in der Domäne III der 16S rRNA nachweisen. Die verwandtschaftlichen Beziehungen der Nudipleura zu anderen Opisthobranchia-Gruppen sind noch weitestgehend ungeklärt.

130
Diskutiert werden vor allem phylogenetische Verbindungen zu den Tylodinoidea (siehe Kap. 4.9.5.4) und zu den Acteonoidea (siehe Kap. 4.9.5.5). Die basale Stellung der Nudipleura in einigen Topologien (18S rDNA: MP-Analyse, 28S rDNA: MP-, ML- und Distanzanalyse) verdient besondere Erwähnung, da die Nudipleura gewöhnlich als höchst abgeleitet angesehen werden (z.B. Schmekel 1985, Mikkelsen 2002). Die Erklärung für diese Beobachtung beruht möglicherweise auf der Existenz unterschiedlicher Substitutionsraten bei den untersuchten Taxa. Grechko (2002) beschreibt einen sog. Akkumulationseffekt bei ribosomalen Genen, durch den die Stellung der Innengruppentaxa in erster Linie von ihrer Evolutionsrate abhängig ist. Das bedeutet, dass das am schnellsten evolvierende Taxon der Innengruppe, die Nudipleura, als erstes von der Stammlinie abzweigt, in der Topologie also ganz basal steht. Dieser Effekt lässt sich auf eine long branch attraction zwischen einem schnell evolvierenden Außengruppentaxon und einem Innengruppentaxon mit erhöhter Substitutionsrate zurückführen. Da die relativen Substitutionsraten der verwendeten Außengruppe (Littorinidae) nicht untersucht wurden, liefern die Astlängen in den Phylogrammen den einzigen Hinweis auf eine möglicherweise erhöhte Evolutionsrate. Allein in den Stammbäumen der Bayes’schen Analysen erscheint die Stammlinie der Littorinidae sehr lang, in den ML- und Distanzphylogrammen können dagegen keine langen Äste festgestellt werden. Die Möglichkeit einer „Fehlplazierung“ der Nudipleura aufgrund erhöhter Evolutionsraten muß aber zumindest in Betracht gezogen werden. 4.9.5.4 Tylodinoidea Die beiden Vertreter der Tylodinoidea, Tylodina perversa und Umbraculum mediterraneum, gruppieren in allen Stammbäumen auf der Grundlage von 18S und 28S rDNA zusammen (bootstrap-Wete: 98-99, posterior probabilities: 0,95-0,99), im 16S rDNA-Datensatz ist nur eine Sequenz dieses Taxons enthalten. In alternativen molekularen Analysen sind die Tylodinoidea mit maximal einer einzigen Sequenz vertreten, nur Grande et al. (2004a,b) konnten ihre Monophylie durch Berücksichtigung der beiden Gattungen Tylodina und Umbraculum bestätigen. Auf der Grundlage morphologischer Daten gilt die Monophylie dieser Gruppierung als gut gestützt, als Synapomorphie wird dabei die Existenz eines verdickten Ringes im Mund (Willan 1987) genannt. Die Stellung der Tylodinoidea innerhalb der Opisthobranchia erscheint sehr variabel. So bilden sie z.B. ein Monophylum mit den Anaspidea, Pteropoda und Cephalaspidea s.str. (18S rDNA: MP-Analyse; 16S rDNA: Distanzanalyse), mit den Nudipleura, Acteonoidea und Anaspidea (16S rDNA, MP-Analyse), oder sie stellen die Schwestergruppe der Pyramidellidae (18S rDNA: Distanzanalyse; 28S rDNA: Bayes’sche Analyse) oder der Sacoglossa (28S rDNA: MP-Analyse) dar. In keiner der erstellten Verwandtschaftshypothesen repräsentieren sie die Schwestergruppe der Pleurobranchoidea.

131
Die Tylodinoidea und Pleurobranchoidea wurden in der Vergangenheit traditionell zu den „Notaspidea“ zusammengefasst, basierend auf Merkmalen, die inzwischen als symplesiomorph erkannt worden sind (z.B. Schmekel 1985). Doch trotz der Nicht-Monophylie der „Notaspidea“, spricht vieles für einen gemeinsamen evolutiven Ursprung von Tylodinoidea und Pleurobranchoidea, bzw. Nudipleura (Guiart 1901, Odhner 1939, Ghiselin 1966). Die Tylodinoidea werden dabei als basal angesehen, tatsächlich erscheint das Reproduktionssystem von Tylodina ursprünglicher als das jedes anderen Vertreters der Opisthobranchia (Gosliner 1981). In der auf morphologischen Daten basierenden phylogenetischen Analyse der Opisthobranchia von Mikkelsen (2002), erscheinen die Tylodinoidea + Nudipleura monophyletisch, als Synapomorphien nennt Mikkelsen z.B. die Existenz eines Mundsegels und einer Blutdrüse. Einen weiteren (scheinbaren) Hinweis auf einen gemeinsamen Ursprung liefern Chromosomenstudien. Die Chromosomenanzahl gilt als sehr konservativ innerhalb der Euthyneura (z.B. Patterson 1969) und kann deshalb zur Begründung verwandtschaftlicher Beziehungen herangezogen werden. Die plesiomorphe Anzahl innerhalb der Opisthobranchia liegt bei 16-18 im haploiden Satz (Haszprunar 1985a) und ist bei den Cephalaspidea, Sacoglossa, Anaspidea und Gymnosomata verwirklicht. Dagegen weisen die „Notaspidea“ eine Anzahl von 12-13 Chromosomen auf, die Nudibranchia eine von 13 (Patterson 1969, Rasotto & Cardellini 1983). Bei genauer Betrachtung der Daten ist die geringe Chromosomenzahl als mögliche Synapomorphie für die Tylodinoidea und Nudipleura allerdings hinfällig, da die genannten Autoren als Vertreter der „Notaspidea“ nur Taxa der Pleurobranchoidea berücksichtigt haben. Chromosomenzahlen der Tylodinoidea sind bisher nicht bekannt. Schmekel (1985) hält die Tylodinoidea und Pleurobranchoidea für morphologisch so divergent, daß sie einen gemeinsamen Ursprung als unwahrscheinlich erachtet und stattdessen eine nähere Verwandtschaft der Tylodinoidea zu den Anaspidea befürwortet. Basierend auf der Analyse verschiedener mitochondrialer Gene haben Grande et al. (2002, 2004a+b) übereinstimmend ein Monophylum aus Tylodinoidea, Anaspidea und Cephalaspidea s.str. rekonstruiert, dabei gruppierten die Cephalaspidea s.str. entweder mit den Tylodinoidea oder den Anaspidea zusammen. Als morphologische Synapomorphien für die Gruppierung aus Tylodinoidea, Anaspidea und Cephalaspidea s.str geben die Autoren z.B. die Existenz einer Eiweißdrüse, Platten im Kaumagen und das Fehlen einer Blutdrüse an (Grande et al. 2004b). In der vorliegenden Arbeit konnte ein gemeinsamer Ursprung der drei Gruppierungen nur im MP-Baum auf der Grundlage der 18S rDNA sowie im Distanzbaum der 16S rDNA festgestellt werden. In der durch eine Bayes’sche Analyse rekonstruierten Topologie des letztgenannten Gens stellen die Tylodinoidea die Schwestergruppe der Anaspidea dar, die posterior probability liegt für dieses Monophylum allerdings nur bei 0,5.

132
Die Analyse des phylogenetischen Signals mit PHYSID (Wägele & Rödding 1998) ergab im 18S und 28S rDNA-Datensatz eine höhere Anzahl potentieller Apomorphien für die „Notaspidea“ (70/62 pot. Apomorphien) als für verwandtschaftliche Beziehungen zu den Cephalaspidea und/oder Anaspidea (3-12/27-39 pot. Apomorphien). Diese Beobachtung lässt sich möglicherweise auf die signifikant erhöhten Substitutionsraten der Gene innerhalb der Pleurobranchoidea zurückführen, die zu zufälligen Übereinstimmungen mit den beiden Sequenzen der Tylodinoidea geführt haben. Andererseits kann die Existenz eines stärkeren Signals für die „Notaspidea“ auch Ausdruck einer real existierenden Verwandtschaft sein. Aufgrund der Variabilität in den rekonstruierten Topologien und den Diskrepanzen zu Bäumen auf der Basis morphologischer Daten, müssen die verwandtschaftlichen Beziehungen der Tylodinoidea als ungeklärt eingestuft werden. 4.9.5.5 Acteonoidea Die Acteonoidea oder Architectibranchia werden durch vier Sequenzen im 18S rDNA-Datensatz (Hydatina physis, Bullina lineata, Pupa solidula, Acteon sp.), durch zwei im 28S rDNA-Datensatz (Hydatina physis, Pupa solidula) und durch drei in der 16S rDNA-Alinierung (Hydatina physis, Pupa solidula, Acteon tornatilis) repräsentiert. In den Bäumen auf der Grundlage der 28S rDNA erscheinen sie übereinstimmend monophyletisch, wohingegen sie in den Topologien der 18S und 16S rDNA-Datensätze paraphyletische Gruppierungen darstellen. Die Paraphylie lässt sich in beiden Genen auf die Gruppierung von Acteon innerhalb der Cephalaspidea s.str. zurückführen, davon abgesehen gruppieren in den Verwandtschaftshypothesen basierend auf der 16S rDNA auch die übrigen Vertreter der Acteonoidea nicht als Monophylum. Die Stellung von Acteon sp. und Acteon tornatilis innerhalb der Cephalaspidea s.str. ist bemerkenswert, da die Acteonidae, Hydatinidae und Ringiculidae aufgrund zahlreicher plesiomorpher Merkmale aus den Cephalaspidea s.l. ausgeschlossen worden sind (Mikkelsen 1996, 2002). Gerade Acteon wird von vielen Autoren als Prototyp eines „Ur-Opisthobranchiers“ angesehen (z.B. Rudman 1972, Thompson 1976, Salvini-Plawen 1990). In der phylogenetischen Analyse morphologischer Daten der Opisthobranchia (Mikkelsen 1996), erscheint Acteon in einem Schwestergruppenverhältnis zu Gegania (Mathildidae, basale Heterobranchia), als Synapomorphie nennt Mikkelsen die besondere Form des Receptaculum seminis. Haszprunar (1985b,c) erwähnt dasselbe Merkmal für zwei andere Arten der basalen Heterobranchia. Die basale Stellung von Acteon wird durch die Ergebnisse molekularer Analysen (Tillier et al.1992, Thollesson 1999, Dayrat et al. 2001, Wägele et al. 2003) gestützt.

133
Die unerwartete Stellung von Acteon innerhalb der Cephalaspidea s.str. lässt sich nicht durch einen long branch-Effekt erklären, da weder Acteon, noch die Cephalaspidea s.str. erhöhte Substitutionsraten aufweisen. Bei Betrachtung der 18S und 16S rDNA-Alinierungen wird deutlich, dass sich die Sequenzen von Acteon sp. und Acteon tornatilis sehr gut in das Sequenzmuster der Cephalaspidea s.str. einfügen, eine auf Unzulänglichkeiten der verwendeten Rekonstruktionsverfahren beruhende „Fehlplazierung“ kann demnach ausgeschlossen werden. Dagegen darf die Möglichkeit nicht unberücksichtigt bleiben, dass es sich um eine Verwechslung von Sequenzen, DNA oder Tiermaterial handelt. Die Paraphylie von Hydatina physis und Pupa solidula in den Stammbäumen des 16S rRNA-Gens lässt sich vermutlich durch das Fehlen eines deutlichen phylogenetischen Signals erklären, das durch die nicht aufgelöste Verwandtschaft im Baum der Bayes’schen Analyse augenfällig wird. Die Stellung der Acteonoidea (exklusive Acteon sp.) in den Verwandtschafts-hypothesen auf der Grundlage der 18S und 28S rDNA, erweist sich als recht variabel. Sie bilden entweder die Schwestergruppen zu allen Taxa der Innengruppe (außer Nudipleura), zu den Nudipleura oder zu den Sacoglossa. In keiner Topologie ist eine basale Stellung der Acteonoidea zu beobachten. Tatsächlich wird der Status der Acteonoidea als archetypische Opisthobranchier von einigen Autoren in Frage gestellt. Bis auf die sehr ursprüngliche Ausbildung von Schale und Mantelkomplex weisen alle Organsysteme zahlreiche Modifikationen auf. Besonders das Reproduktionssystem ist als höchst abgeleitet anzusehen (Ghiselin 1966, Gosliner 1981). Die Form der Radula bei Acteon spricht ebenfalls gegen die basale Stellung der Gruppe, da sie in Anpassung an eine Ernährung von Polychaeten eine sehr große Anzahl winziger, gleichförmiger Zähne aufweist. Eine Ableitung der übrigen innerhalb der Opisthobranchia vorkommenden Radulatypen von dieser ist schwer vorstellbar (Mikkelsen 2002). Das auf der Grundlage der 18S rDNA (Distanzanalyse) rekonstruierte Schwestergruppenverhältnis von Acteonoidea und Nudipleura scheint sich auf den ersten Blick auf eine long branch attraction zurückführen zu lassen, da sowohl die Nudipleura, als auch die Acteonoidea signifikant erhöhte Substitutionsraten aufweisen. Es gibt allerdings einige Hinweise, die auf eine reale Verwandtschaft der Gruppen hinweisen. Bereits Eliot (1910) hat eine Reihe von Ähnlichkeiten erwähnt, die eine Verwandtschaft von Acoela (Tylodinoida + Nudipleura) und Acteonoidea nahe legen. Ghiselin (1966) vereint die beiden Gruppen auf der Grundlage der Übereinstimmungen im Reproduktionssystem. Auch molekulare Daten stützen das postulierte Monophylum zum Teil. Sowohl die Analysen von Grande et al. (2004a+b) auf der Basis mitochondrialer Gene, als auch die von Vonnemann et al. (2005) auf der Basis von 18S und 28S rDNA-Sequenzen, ergaben übereinstimmend ein Schwestergruppenverhältnis von Nudipleura und

134
Acteonoidea. Im Gegensatz zu den vorliegenden Rekonstruktionen wurden bei Vonnemann et al. (2005) die Pyramidellidae als Außengruppe verwendet. Im Widerspruch zu morphologische Daten zeigten die Tylodinoidea allerdings keine phylogenetischen Beziehungen zu den beiden Taxa. Unter Berücksichtigung der genannten Fakten kann man vermuten, dass sich die Acteonoidea stammesgeschichtlich sehr früh von den anderen Opisthobranchiern getrennt haben, dafür spricht das „Mosaik“ aus ursprünglichen und abgeleiteten Merkmalen. Möglicherweise gilt für die Nudipleura dasselbe, die Übereinstimmungen im Reproduktionssystem würden demnach auf eine Parallelentwicklung zurückzuführen sein. Aufgrund der erhöhten Evolutionsraten in beiden Gruppen muß die Möglichkeit einer long branch attraction in Betracht gezogen werden, wobei die Monophylie von Acteonoidea + Nudipleura auch Ausdruck realer Verwandtschaft sein kann. 4.9.5.6 Anaspidea und Pteropoda Die Anaspidea erscheinen in allen Topologien auf der Basis der 28S und 16S rDNA als monophyletische Gruppierungen (bootstrap-Werte: 95-100, posterior probabilities: 0,99-1). In den Stammbaumhypothesen basierend auf dem 18S rRNA-Gen, bilden sie entweder ein Monophylum mit allen Pteropoda (Thecosomata und Gymnosomata, MP-, Bayes’sche Analyse) oder nur mit den Gymnosomata (Distanzanalyse) und stellen demnach übereinstimmend eine paraphyletische Gruppierung dar. Die Monophylie der Anaspidea gilt anhand morphologischer Merkmale als gut gestützt (z.B. Schmekel 1985, Mikkelsen 1996, 2002), als synapomorph gelten das Vorhandensein einer Filterkammer und eines Magenblindsacks (Mikkelsen 1996). Auch molekulare Untersuchungen (Thollesson 1999b, Medina & Walsh 2000, Wägele et al. 2003, Grande et al. 2004a,b) konnten die Monophylie der Anaspidea bestätigen. Die Stellung der Taxa innerhalb der Anaspidea wird eingehend unter Punkt 4.10 diskutiert. Die Pteropoda sind möglicherweise monophyletischen Ursprungs (Ghiselin 1966), werden aber überwiegend als diphyletisch angesehen (van der Spoel 1967). Chromosomenstudien zeigen z.B., dass die Thecosomata aufgrund einer auffälligen Diversität in der Chromosomenzahl eine isolierte Gruppe innerhalb der Opisthobranchia darstellen, wohingegen die untersuchten Chromosomenmerkmale bei den Gymnosomata alle plesiomorphen Charakters sind. Aufgrund dieser und anderer Merkmale wird eine lange, unabhängige Entwicklung ausgehend von einem frühen Cephalaspidea-artigen Vorfahren angenommen (Thiriot-Quiévreux 1987). Ghiselin (1966) sieht dagegen “... more sound evidence for relating the Thecosomata

135
to the Gymnosomata than to relating either group to any other opisthobranchs” (1966: 368). In den hier erstellten Verwandtschaftshypothesen sind Vertreter der Thecosomata und Gymnosomata nur in denen auf der Grundlage des 16S- und des 18S rDNA-Datensatzes enthalten. Sie gruppieren dabei nur in einer Topologie (18S rDNA: MP-Analyse) als Monophylum. Die Thecosomata erscheinen dagegen in allen 3 Stammbäumen auf der Grundlage der 18S rDNA monophyletisch, dabei werden die Verwandtschaftsbeziehungen ihrer Taxa nur im Distanzbaum aufgelöst. In diesem stellen Clio pyramidata und Diacra quadridenata Schwestergruppen dar. Für die phylogenetischen Beziehungen innerhalb der Thecosomata wurden unterschiedliche Hypothesen diskutiert, die Einteilung in die beiden Gruppen Euthecosomata und Pseudothecosomata wird aber von den meisten Autoren akzeptiert (Thiriot-Quievreux 1987). Die in der vorliegenden Arbeit untersuchten Arten gehören alle zur erstgenanten Gruppierung, deren postulierte Verwandtschaftshypothesen ausschließlich auf Arbeiten von van der Spoel (z.B. 1967) beruhen. In Übereinstimmung mit seinen Ergebnissen, stellen hier Clio und Diacra, im Vergleich zu Creseis, stärker abgeleitete Gattungen dar. Eine enge Verwandtschaft der Pteropoda mit den Anaspidea wird in allen drei Topologien deutlich, wobei die Thecosomata im Distanzbaum nicht in der monophyletischen Gruppierung aus Anaspidea und Gymnosomata enthalten sind. Eine Verwandtschaft zwischen Gymnosomata und Anaspidea wurde bereits von Pelseneer (1888) vermutet, er leitet die Gymnosomata von hochspezialisierten Anaspidea, die Thecosomata dagegen von nicht näher spezifizierten Cephalaspidea ab. Hoffmann (1932-40) schlägt dagegen Verbindungen beider pteropoden Gruppen zu einem Vorfahren der Anaspidea vor. Howells (1936) beschreibt bei Cymbulia peroni (Thecosomata) ein Drüsengewebe in einer Position identisch zu der der Buccaldrüse bei den Aplysiidae (Anaspidea). Falls sich die Homologie beider Strukturen bestätigen sollte, wäre eine enge phylogenetische Beziehung zu den Aplysiidae sehr wahrscheinlich. Minichev (1963) leitet die Gymnosomata von basalen Anaspidea (Akeratidae) und diese wiederum von den Acteonidae ab, wohingegen er die Thecosomata als von den Acteonoidea ausgehende unabhängige Entwicklungslinie ansieht. Unter Berücksichtigung aller vorhandenen Daten betrachtet Ghiselin (1966) die Pteropoda als monophyletische Gruppierung, die mit den Anaspidea zwar einen gemeinsamen Ursprung teilen, aber nicht innerhalb dieser entstanden sind. Im Vergleich dazu suggeriert das Ergebnis der Bayes’schen Analyse durch die abgeleitete Stellung der Pteropoda sehr wohl eine Entwicklung aus den Anaspidea. In der molekularen Analyse (28S rDNA) von Dayrat et al. (2001) erscheinen die Pteropoda und Anaspidea als gut gestütztes Monophylum. Als morphologische

136
Synapomorphie nennen Dayrat et al. (2003) die Entstehung von zahlreichen Kalkplatten im Kaumagen, im Vergleich zu drei Platten im plesiomorphen Zustand. Innerhalb der Pteropoda ist das Fehlen dieser Strukturen dann als sekundäre Reduktion anzusehen. In den Topologien auf der Basis des 16S rDNA-Datensatzes, lassen Clione limacina (Gymnosomata) und Cavolinia sp. (Thecosomata) weder phylogenetische Beziehungen zueinander, noch zu den Anaspidea erkennen. Clione limacina (AJ 223406, Tholleson 1999a) gruppiert dabei sogar außerhalb der Heterobranchia. Diese Stellung ist als sehr fragwürdig einzuschätzen, da die Plazierung der Gymnosomata innerhalb der Opisthobranchia als gesichert angesehen wird (Ghiselin 1966, Thiriot-Quievreux 1987, Huber 1993). Bei einem Abgleich mit Sequenzen der Genbank werden die größten Übereinstimmungen der Sequenz von Clione limacina ausschließlich mit Vertretern der Caenogastropoda erzielt. In der Alinierung wird die enge Verwandtschaft von Clione limacina zu den Außengruppentaxa durch mehrere synapomorphe Merkmale, z.B. eine Insertion im Bereich 89/89’, deutlich. Thollesson diskutiert die Möglichkeit, dass „...it may be a possible mistake or artefact...“ (1999a: 80), schließt aber eine Fehlbestimmung oder Kontamination mit Fremd-DNA aus. Eine potentielle Erklärung für den beobachteten Sachverhalt liefert die Existenz von Pseudogenen. In Anbetracht der großen Ähnlichkeit mit Caenogastropoden-sequenzen müsste das sequenzierte 16S rRNA-Pseudogen von Clione limacina aus einem lang zurückliegenden Duplikationsereignissen stammen, eine Abstammung der Gymnosomata aus Caenogastropoden vorausgesetzt. Cavolinia sp. zeigt eine variable Position in den Topologien und erscheint entweder als Schwestertaxon zu Dendrodoris nigra (Nudibranchia), zu Odostomia sp. (Pyramidellidae) oder zu einer Vereinigung aus Cephalaspidea s.str., Tylodinoidea, Anaspidea, Nudipleura und Pupa solidula (Acteonoidea). Da Cavolinia sp. ebenso wie Dendrodoris nigra und Odostomia sp. sehr lange Äste aufweisen, ist eine „fehlerhafte“ Gruppierung aufgrund einer long branch attraction sehr wahrscheinlich. 4.9.5.7 Cephalaspidea s.str. Die Cephalaspidea s.str. erscheinen, abgesehen von Runcina adriatica, in allen erzeugten Topologien als monophyletische Gruppierung. Bei einer Stellung innerhalb der Cephalaspidea s.str. stellt Runcina adriatica immer die Schwestergruppe zu den restlichen Vertretern des Taxons dar. Bei einer Gruppierung außerhalb (18S rDNA: Distanzanalyse; 28S rDNA: ML-, Bayes’sche Analyse), bildet die Art entweder die Schwestergruppe zu Cephalaspidea s.str. und Cepaea hortensis (Pulmonata) oder zu allen Innengruppentaxa, mit Ausnahme der Nudipleura. Der 16S rDNA-Datensatz beinhaltet keine Vertreter der Runcinidae. Die Stellung der Runcinidae wird kontrovers diskutiert (vergl. z.B. Odhner 1968, Kress 1977 und Schmekel 1985). In der Analyse morphologischer Daten der

137
Euthyneura gruppieren die Runcinoidea nicht innerhalb des Monophylums aus Anaspidea und Cephalaspidea s.str. (Dayrat et al. 2001). Bei Wägele & Klussmann-Kolb (in Vorbereitung) bilden die Runcinidae und Philinoglossidae, ebenfalls auf der Basis morphologischer Merkmale, eine monophyletische Gruppierung mit den Acochlidiacea. Ghiselin (1966) vereinigt dagegen die Retusidae, Philinoglossidae, Bullidae, Atyidae und Runcinidae aufgrund von Übereinstimmungen im Kopulationsapparat in einer Abstammungsgemeinschaft, gibt aber gleichzeitig an, dass es sich möglicherweise um eine polyphyletische Gruppierung handelt. Abgesehen von der Stellung der Runcinidae gilt die Monophylie der Cephalaspidea s.str. als gut gestützt. Mikkelsen (1996) nennt, ohne Berücksichtigung der Runcinidae, als Synapomorphien u.a. das Vorhandensein von drei Kauplatten gebogene Cilienstreifen in der Mantelhöhle. Die Analyse molekularer Daten führte zu übereinstimmenden Ergebnissen (Thollesson 1999a, Dayrat et al. 2001, Wägele et al. 2003, Grande et al. 2004a,b). Bei Grande et al. (2004b) gruppierte die kombinierte Sequenz einiger mitochondrialer Gene von Runcina coronata ebenfalls als Schwestergruppe zu den übrigen Sequenzen der Cephalaspidea s.str. Die unerwartete Stellung von Acteon innerhalb der Cephalaspidea wurde unter Punkt 4.9.5.5 diskutiert und wird als das Produkt einer Fehlbestimmung oder Verwechslung angesehen. Die Familienzugehörigkeit der untersuchten Arten spiegelt sich zum Teil sehr gut in den rekonstruierten Stammbäumen wider. Die Aglajidae (Navanax inermis, Odontoglaja guamensis, Chelidonura inornata, Philinopsis pilsbryi) werden in allen Topologien als monophyletische Gruppierung rekonstruiert (Bootstrap: 99, posterior probabilities: 0,99-1), in den Topologien basierend auf der 18S rDNA inklusive Acteon sp. Im 16S rDNA-Datensatz sind keine Vertreter der Aglajidae enthalten. Eine weitere Familie mit mehreren untersuchten Arten sind die Haminoeidae (Haminoea hydatis, Phanerophthalmus smaragdinus, Atys semistriatus, Smaragdinella sp. und Bullacta exarata). Die beiden letztgenannten Taxa sind im 28S rDNA-Datensatz nicht enthalten und in der 16S rDNA-Alinierung werden die Haminoeidae durch die beiden Sequenzen von Haminoea cymbalum und Smaragdinella sp. repräsentiert. In allen 16S und 28S rDNA-Bäumen ist diese Familie monophyletisch. Basierend auf der 18S rDNA konnte eine Monophylie nur durch die Bayes’sche Analyse rekonstruiert werden, im Distanzphylogramm erscheinen sie paraphyletisch und im MP-Dendrogramm konnten die Verwandtschaftsverhältnisse der Haminoeidae überhaupt nicht aufgelöst werden. Die Diaphanidae sind mit vier Arten (Diaphana cf. inflata, Toledonia globosa, Newnesia antarctica, Colpodaspis thompsoni) im 18S rDNA- und mit nur einer (Diaphana minuta) im 16S rDNA-Datensatz vertreten. Die Diaphanidae stellen in allen Topologien eine basale, paraphyletische Gruppierung dar, dabei bildet Colpodaspis thompsoni übereinstimmend die Schwestergruppe von Sagaminopteron psychedelicum (Gastropteridae). Die basale Stellung der Familie stimmt gut mit den

138
Ergebnissen morphologischer Untersuchungen überein, denn das Nervensystem der Diaphanidae gilt als sehr ursprünglich (Ghiselin 1966). Die taxonomische Definition der Familie gründet sich hauptsächlich auf Negativmerkmale, z.B. das Fehlen von Kiefern und Kauplatten. Die Unkenntnis synapomorpher Merkmale spricht für eine Paraphylie der Gruppe (Jensen 1996), die durch morphologische Untersuchungen von Wägele & Klussmann-Kolb (in Vorbereitung) bestätigt werden konnte. Die Gattungen Toledonia und Newnesia weisen dagegen einige übereinstimmende Merkmale auf, eine Tatsache, die durch ihr Schwestergruppenverhältnis in der Topologie der Distanzanalyse gestützt wird. Die beiden im 18S rDNA-Datensatz vertretenen Arten der Cylichnidae (Scaphander lignarius, Cylichna gelida) stellen in allen Fällen eine paraphyletische Gruppierung dar. Das entspricht dem Ergebnis auf der Basis morphologischer Daten (Mikkelsen 1996, Wägele & Klussmann-Kolb, in Vorbereitung). Weitere Familien werden nur durch jeweils eine Sequenz repräsentiert: cf. Retusa sp. (Retusidae), Bulla cf. striata (Bullidae), Philinoglossa praelongata (Philinoglossidae), Philine aperta (Philinidae), Runcina adriatica (Runcinidae), Sagaminopteron psychedelicum (Gastropteridae). Eine in allen Topologien wiederkehrende Verwandtschaftsbeziehung ist z.B. das Schwestergruppenverhältnis von Retusidae und Haminoeidae (28S rDNA), bzw. die Monophylie von Retusidae, Haminoeidae und Cylichna gelida (18S rDNA). Eine eindeutige Stützung durch morphologische Daten gibt es für diese Gruppierung nicht, allerdings wurde eine Beziehung der Retusidae zu den Bullidae und Haminoeidae von einigen Autoren vermutet (Ghiselin 1966, Mikkelsen 1993, 1994). Weitere Übereinstimmungen bestehen im Schwestergruppenverhältns von Philinoglossidae und Aglajidae auf der Grundlage der 18S rDNA, anhand der 28S rDNA-Datensatz konnte diese Verwandtschaft allerdings nicht aufgezeigt werden. Im Vergleich zu 18S und 28S rDNA ist in allen Topologien auf der Basis der 16S rDNA ein Monophylum aus Bullidae und Haminoeidae zu beobachten. Diese Verwandtschaft wurde ebenfalls von Mikkelsen (1996) auf der Grundlage morphologischer Daten postuliert aber nicht durch Untersuchungen von Wägele & Klussmann-Kolb (in Vorbereitung) bestätigt Alle Topologien auf der Grundlage der 16S rDNA-Alinierung zeigen ein Schwestergruppenverhältnis von Philine aperta und Scaphander punctostriatus, sowohl gestützt durch die Untersuchungen von Mikkelsen (1996), als auch durch die Analyse chemosensorischer Organe (z.B. Osphradium) von Edlinger (1980). Bei Wägele & Klussmann-Kolb (in Vorbereitung) erscheinen die beiden Gattungen allerdings als Paraphylum. 4.9.5.8 Cephalaspidea und Anaspidea Ein Schwestergruppenverhältnis von Cephalaspidea s.str. und Anaspidea konnte auf der Basis morphologischer Daten bereits von Mikkelsen (1996) gezeigt werden,

139
allerdings hat sie wichtige opisthobranchiate Linien dabei unberücksichtigt gelassen, z.B. Nudipleura, Tylodinoidea, Pteropoda und mehrere cephalaspide Familien. Als Synapomorphie für das erwähnte Monophylum nennt sie z.B die Entstehung eines Kaumagens. In der Topologie von Wägele & Klussmann-Kolb (in Vorbereitung) wird die Monophylie von Anaspidea und Cephalaspidea s.str. (exklusive Philinoglossidae und Runcinidae) bestätigt, allerdings stellen die Anaspidea die Schwestergruppe der Haminoeidae dar, die Cephalaspidea s.str. sind demnach paraphyletisch. Inzwischen sind zahlreiche molekulare Analysen der Euthyneura durchgeführt worden, die die Monophylie der beiden Gruppen zum größten Teil stützen: Thollesson (1999) auf der Basis der 16S rDNA, Wägele et al. (2003) basierend auf 16S und 18S rDNA-Sequenzen und Grande et al. (2002, 2004a) auf der Grundlage verschiedener mitochondriale Gene. Die Untersuchung der partiellen 28S rDNA von Dayrat et al. (2001) ergab ebenfalls eine enge Verwandtschaft von Cephalaspidea s.str. und Anaspidea (inkl. Pteropoda), allerdings gruppierte Archidoris tuberculata innerhalb dieser Gruppierung, als Schwestertaxon zu Runcina sp. Möglicherweise handelt es sich dabei um eine long branch attraction, falls die Sequenz von Runcina sp. ebenso wie die der hier untersuchten von Runcina adriatica im Vergleich zu den übrigen Cephalaspidea s.str. eine erhöhte Substitutionsrate aufweist. 4.9.5.9 Sacoglossa Die Sacoglossa werden in allen erstellten Verwandtschaftshypothesen als Monophylum rekonstruiert (Bootstrap: 99-100, posterior probabilities: 0,98-0,99). Gestützt werden diese Ergebnisse durch molekulare Untersuchungen von Wägele et al. (2003) auf der Basis von 16S rDNA-, 18S rDNA- und COI-Sequenzen, von Dayrat et al. (2001) auf der Grundlage des 28S rRNA-Gens und von Thollesson (1999a) basierend auf der 16S rDNA. In den genannten Arbeiten wurden maximal drei Sequenzen der Sacoglossa berücksichtigt. Die Monophylie der Sacoglossa gilt auf der Grundlage morphologischer Untersuchungen heute als unstrittig, zahlreiche Synapomorphien existieren im Verdauungs-, Reproduktions- und Nervensystem (Mikkelsen 1996, Wägele & Klussmann-Kolb, in Vorbereitung). Die Phylogenie der Sacoglossa wurde anhand umfassender morphologischer Daten von Jensen (1996) rekonstruiert. In ihrer Verwandtschaftshypothese erscheinen die schalentragenden Sacoglossen als Paraphylum, die schalenlosen als mono-phyletische Gruppierung. In der vorliegenden Arbeit konnten nur Vertreter der Letztgenannten berücksichtigt werden. Innerhalb der schalenlosen Sacoglossa, konnte Jensen zwei monophyletische Gruppierungen nachweisen, die Plakobranchoidea mit den hier untersuchten Gattungen Bosellia, Plakobranchus, Thuridilla, Elysia und Elysiella und die Limapontioidea mit Placida, Cyerce, Polybranchia, Placida, Calliopaea und Limapontia.

140
In den Bäumen auf der Grundlage des 18S rDNA-Datensatzes, repräsentieren die Plakobranchoidea übereinstimmend ein monophyletisches Taxon und die Limapontioidea ein paraphyletisches, in den Topologien auf der Basis der 28S rDNA bilden die Plakobranchoidea ein Monophylum mit Cyerce (Limapontioidea) und stellen dadurch eine paraphyletische Gruppierung dar. Im 16S rDNA-Datensatz sind nur drei Sacoglossen-Sequenzen enthalten, dabei bilden Elysia und Thuridilla (Plakobranchoidea) die Schwestergruppe zu Placida (Limapontioidea). Innerhalb der Plakobranchoidea sind drei Familien in den untersuchten Datensätzen vertreten, die Elysiidae (Elysia, Elysiella, Thuridilla), Boselliidae (Bosellia) und Plakobranchidae (Plakobranchus). Die Elysiidae sind nur im Stammbaum der Bayes’schen Analyse der 18S rDNA monophyletisch, ein Schwestergruppen-verhältnis von Elysia und Elysiella tritt aber in allen Fällen auf. In den Bäumen des 18S rRNA-Gens stellen Bosellia und Plakobranchus übereinstimmend Schwestertaxa dar, in den Topologien auf der Basis der 28S rDNA ist diese Verwandtschaft dagegen nie zu beobachten. Die untersuchten Taxa der Limapontioidea lassen sich taxonomisch in die Limapontiidae (Calliopaea, Limapontia, Placida) und Polybranchidae (Polybranchia, Cyerce) einteilen. In allen erstellten Stammbaumhypothesen (18S rDNA, 28S rDNA) erscheinen Calliopaea und Limapontia als Schwestergruppentaxa, Placida (nur 18S rDNA) bildet mit diesen ein Monophylum in den durch MP- und Distanzanalysen erzeugten Stammbäumen. Diese Stellung der drei Taxa entspricht exakt den morphologischen Ergebnissen (Jensen 1996, Mikkelsen 1998). Die Polybranchidae stellen in allen Topologien eine paraphyletische Gruppierung dar, dabei bilden sie mit den Plakobranchoidea ein Monophylum. In der Verwandtschaftshypothese von Jensen (1996) werden alle Familien als monophyletische Einheiten bestätigt, nur die Polybranchidae erscheinen dort, wie auch in den vorliegenden Analysen, als Paraphylum. Jensen (1996) bezeichnet die Limapontioidea als „problematischste“ Gruppierung innerhalb der Sacoglossa, da die morpholgischen Unterschiede zwischen den Gattungen und Arten viel größer sind als innerhalb der Plakobranchoidea. Auf Grundlage der vorliegenden Analysen sollte eine Paraphylie der Limapontioidea in Betracht gezogen werden. Die Stellung der Sacoglossa innerhalb der Opisthobranchia, bzw. Heterobranchia erweist sich beim Vergleich der erstellten Topologien als äußerst variabel. Zu beobachten ist u.a. ein Schwestergruppenverhältnis zu den Acteonoidea (28S rDNA, ML-Analyse), zu den Tylodinoidea (28S rDNA, MP-Analyse) oder zu Lymnaea stagnalis (Pulmonata, 18S rDNA, Distanz-, Bayes’sche Analyse). In der Stammbaumhypothese von Grande et al. (2004a), basierend auf 4 mitochondrialen Genen, stellen die Sacoglossa die Schwestergruppe zu allen übrigen Opisthobranchia (inkl. Siphonaria) dar, allerdings wurde nur eine einzige Sequenz der Sacoglossa berücksichtigt. Die Maximum Parsimonie-Analyse der 16S

141
rDNA (Thollesson 1999) ergab ebenfalls eine basale Position innerhalb der Euthyneura. In der Distanzanalyse desselben Datensatzes erschienen die Sacoglossa dagegen als Schwestergruppe der Pulmonata. In der anhand morphologischer Daten erstellten Verwandtschaftshypothese der Opisthobranchia (Mikkelsen 2002), bilden die Sacoglossa die Schwestergruppe der Nudipleura, Tylodinoidea, Cephalaspidea s.str. und Anaspidea, wohingegen sie im Stammbaum von Wägele & Klussmann-Kolb (in Vorbereitung) eine eher abgeleitete Stellung einnehmen. Sie bilden dort eine monophyletische Gruppierung mit den Runcinidae, Philinoglossidae, Acochlidicea und den Rhodopidae. Eine basale Stellung der Sacoglossa, die von den meisten phylogenetischen Analysen postuliert wird, kann durch die vorliegenden Ergebnisse nicht gestützt werden. Die Variabilität in den zu beobachtenden Verwandtschaftsbeziehungen kann am besten durch das Fehlen eines deutlichen phylogenetischen Signals erklärt werden, das scheint sowohl für molekulare als auch für morphologische Daten zu gelten. Laut Jensen ist es „…not possible to link the Sacoglossa with any other extant opisthobranch clade. Any apparent synapomorphy can be explained as parallel evolution or not quite homologous structures.” (1996: 117). 4.9.5.10 Acochlidiacea Die Acochlidiacea erscheinen in allen Topologien auf der Grundlage des 16S rDNA-Datensatzes, sowie im MP-Baum der 18S rDNA und im Distanzbaum der 28S rDNA als monophyletische Gruppierung. Die Paraphylie in den übrigen Stammbaum-hypothesen lässt sich im wesentlichen auf eine abweichende Stellung von Hedylopsis spiculifera zurückführen. In allen Topologien stellen Unela glandulifera und Parahedyle cryptophthalma Schwestertaxa dar, die mit Ausnahme des Distanzbaumes der 18S rDNA, ein Monophylum mit Pontohedyle milatchevitchi bilden. Die Monophylie der Acochlidiacea gilt auf der Grundlage morphologischer Daten als gut gestützt, in molekularen Analysen sind sie bisher nicht berücksichtigt worden. Die Verwandtschaftsbeziehungen der Acochlidiacea zu anderen Gruppierungen der Opisthobranchia sind aufgrund der vielen autapomorpen Merkmale, bedingt durch geringe Körpergröße und hohen Spezialisierungsgrad, noch völlig ungeklärt. Von Bergh (1895) ursprünglich in die Nudibranchia eingeordnet, wurden sie von anderen Autoren mit den Sacoglossa in Verbindung gebracht (Gosliner & Ghiselin 1984) oder in die Cephalaspidea gestellt (Boettger 1954, Salvini-Plawen 1973). Am weitesten verbreitet ist heute ihre Betrachtung als unabhängige opisthobranchiate Linie (Taylor & Sohl 1962, Hyman 1967), mit möglichen phylogenetischen Beziehungen zu den Philinoglossoidea und Platyhedyloidea (Rankin 1979). Tatsächlich stellen die Acochlidiacea in der auf morphologischen Merkmalen basierenden Verwandtschaftshypothese der Opisthobranchia (Wägele & Klussmann-

142
Kolb, in Vorbereitung) die Schwestergruppe der Runcinidae und Philinoglossidae dar. Allerdings ist es vorstellbar, dass diese Gruppierung allein auf (konvergenten) Übereinstimmungen in Verbindung mit der Lebensweise und der geringen Körpergröße beruht. Die phylogenetischen Beziehungen der Acochlidiacea zu anderen Taxa der Euthyneura stellen sich in den hier erstellten Topologien als recht variabel dar, doch eine nähere Verbindung zu verschiedenen Gruppen der Pulmonaten ist auffallend konstant. Abschließend betrachtet bleiben die verwandtschaftlichen Beziehungen der Acochlidiacea noch völlig offen. 4.10 Prüfung der Plausibilität der rekonstruierten Verwandtschaftshypothesen In den vorangegangenen Kapiteln wurden die einzelnen Großgruppen der Euthyneura bezüglich ihres Status als Mono- oder Paraphylum und ihrer möglichen verwandtschaftlichen Beziehungen zueinander diskutiert. Die auf der Grundlage der unterschiedlichen Datensätze und Rekonstruktionsverfahren postulierten phylo-genetischen Beziehungen der einzelnen Taxa konnten durch den Vergleich mit alternativen morphologischen oder molekularen Literaturdaten bewertet werden. Dabei wurden die erhaltenen Verwandtschaftshypothesen nie in ihrer Gesamtheit betrachtet, sondern nur aus dem Blickwinkel des jeweils behandelten Taxons. In den folgenden Abschnitten werden die erhaltenen Topologien durch den Vergleich mit Fossilfunden und die Methode des character mapping im Hinblick auf ihre Plausibilität geprüft. Bei der Projektion der biologischen Daten auf die Stammbäume wird das Parsimonie-Prinzip zugrundegelegt, d.h. das hypothetische evolutionäre Szenario, das zu den bei den terminalen Taxa beobachteten Merkmalen (z.B. der Chromosomenanzahl) geführt hat, soll möglichst wenige Änderungen aufweisen. 4.10.1 Vergleich mit Fossilfunden Aufgrund der mehrfach stattgefundenen und zum Teil kompletten Schalenreduktion innerhalb der Opisthobranchia ist der Vergleich mit Fossildaten sehr problematisch. Gruppen mit ausgeprägten Gehäusen, z.B. die Bullidae (Cephalaspidea s.str.) oder Acteonidae (Acteonoidea) besitzen einen sehr umfangreichen Fossilnachweis, Formen mit dünnschaligen Gehäusen, z.B. die Philinidae (Cephalaspidea s.str.) oder Akeridae (Anaspidea) sind dagegen sehr selten erhalten. Vollständig schalenlose Gruppen wie die Nudibranchia oder Plakobranchoidea (Sacoglossa) sind aus-schließlich aus der rezenten Fauna bekannt, machen aber einen Anteil von über 70% aller bekannten Opisthobranchia-Arten aus.

143
Ein weiteres Problem bei der Analyse von Fossildaten stellt die Zuordnung von fossilen Schalen zu rezenten Taxa dar, da die überwiegende Anzahl der taxonomisch wichtigen Merkmale auf den Weichkörper beschränkt ist. Die Altersangaben für die behandelten Taxa sind der Arbeit von Wägele et al. (in Vorbereitung) entnommen. Die Projektion der Fossildaten auf die Stammbaumhypothese der MP-Analyse des 18S rDNA-Datensatzes ist in Abb. 37 dargestellt. Im „Idealfall“ sollten die im Stammbaum basal stehenden Taxa die erdgeschichtlich ältesten Gruppen repräsentieren, während die abgeleiteten Formen jüngeren Ursprungs sein sollten. Dieser Zusammenhang ist bei der vorliegenden Topologie nicht gegeben. Tatsächlich zeigt das erdgeschichtlich jüngste Taxon, die Nudipleura, eine basale Stellung und eines der ältesten (Bullidae) eine höchst abgeleitete Position. Für diese Beobachtungen gibt es zwei alternative Erklärungen. Zum einen ist es möglich, dass die Phylogenie der untersuchten Taxa aufgrund von Unzu-länglichkeiten der zugrundeliegenden Daten oder der angewendeten Rekonstruktionsverfahren nicht korrekt dargestellt wurde. Zum anderen können sich die Diskrepanzen in den Altersangaben darauf zurückführen lassen, daß das erste zeitliche Auftreten einer Tiergruppe nicht mit dem anhand von Fossilien datierten Alter übereinstimmt. Es wird vermutet, dass im vorliegenden Fall beide Annahmen zutreffen. Bei Taxa mit einem umfangreichen Fossilnachweis ist die Wahrscheinlichkeit groß, dass die ältesten existierenden Formen auch gefunden worden sind, wohingegen bei spärlichen Fossilfunden nur zufällig das exakte Alter einer Gruppe „getroffen“ werden kann. Bei den Fossildaten der Taxa des Datensatzes zeigt sich ein deutlicher Zusammenhang zwischen der Schalenausbildung und dem anhand von Fossilien datierten Alter, indem die erdgeschichtlich ältesten Gruppen die robustesten Gehäuse aufweisen (z.B. Acteonoidea, Ellobiidae, Bullidae). Andererseits ist zusätzlich auch die Möglichkeit einer fehlerhaften phylogenetischen Rekonstruktion der Taxa in Betracht zu ziehen. Die basale Stellung der Nudipleura wurde bereits in Kapitel 4.9.5.3 diskutiert und entspricht möglicherweise nicht ihrer realen, stammesgeschichtlichen Position. Auch die Paraphylie der Pulmonata ist kritisch zu beurteilen. Früheste Pulmonatenfunde (Physidae, Carychidae) wurden auf ein Alter von etwa 340 Millionen Jahre datiert, wohingegen die erdgeschichtlich ältesten Taxa der Opisthobranchia (Hydatinidae) ein Alter von etwa 190 Millionen Jahren aufweisen (Tracey et al. 1993). Die in dieser Topologie postulierte Entstehung der Pulmonata aus opisthobranchiaten Formen lässt sich nicht mit diesen Fakten vereinbaren und könnte als das Ergebnis einer nicht korrekten phylogenetischen Rekonstruktion angesehen werden. Zudem wird die Monophylie der Pulmonata durch die Analyse morphologischer Daten gestützt. Abschließend betrachtet erweist sich die Analyse von Fossildaten der Opisthobranchia als nicht sehr aufschlussreich bei der Beurteilung der erhaltenen Topologien. Es wird deshalb darauf verzichtet, alle Stammbaumhypothesen einzeln zu diskutieren.

144
Acteon sp. Navanax inermis Odontoglaja guamensis Chelidonura inornata Philinopsis pilsbryi Philinoglossa praelongata Sagaminopteron psychedelicum Colpodaspis thomsoni Haminoea hydatis Atys semistriatus Bullacta exarata Phanerophthalmus smaragdinus Smaragdinella sp. Bulla cf. striata cf. Retusa sp. Cylichna gelida Scaphander lignarius Diaphana cf. inflata Toledonia globosa Newnesia antarctica Runcina adriatica Petalifera petalifera Stylocheilus longicauda Dolabrifera brazieri Bursatella leachii Clio pyramidata Creseis acicula Diacria quadridentata Spongiobranchaea australis Akera bullata Syphonota geographica Dolabella auricularia Aplysia californica Umbraculum mediterraneum Tylodina perversa Unela glandulifera Parahedyle cryptophthalma Pontohedyle milatchevitchi Hedylopsis spiculifera Onchidella floridana Onchidium verrucosum Siphonaria alternata Cepaea hortensis Melampus luteus Cassidula nucleus Ovatella firminii Pythia scarabaeus Auriculinella bidentata Pyramidella sulcata Odostomia sp. Elysia viridis Elysiella pusilla Thuridilla bayeri Placobranchus ocellatus Bosellia mimetica Polybranchia orientalis Cyerce nigricans Calliopaea bellula Limapontia nigra Placida cremoniana Lymnaea stagnalis Hydatina physis Bullina lineata Pupa solidula Pleurobranchus peroni Bathyberthella antarctica Euselenops luniceps Tompthompsonia antarctica Berthellina citrina Berthella stellata Berthella platei Chromodoris krohni Hypselodoris villafranca Dendrodoris fumata Bathydoris clavigera Cuthona nana Flabellina babai Dendronotus dalli Fasciolaria lignaria Pisania striata Bursa rana Crepidula adunca Truncatella guerinii Viviparus georgianus Balcis eburnea Melaraphe neritoides Littoraria undulata Littorina striata Nodilittorina punctata Littorina saxatilis Littorina littorea
50 Mio
140 Mio
160-190 Mio
30 Mio
70 Mio
150 Mio
150 Mio
150 Mio
50 Mio
54 Mio 54 Mio 54 Mio
190 Mio
54 Mio
60 Mio
70 Mio 70 Mio 70 Mio 70 Mio 70 Mio
180 Mio 160 Mio 150 Mio 150 Mio 160 Mio 160 Mio 160 Mio
Abb. 37: Projektion von Fossildaten auf die Stammbaumhypothese der Maximum Parsimonie-Analyse (18S rDNA, Opisthobranchia-Datensatz). Strikte Konsensustopologie aus 1.000.000 kürzesten Bäumen, Suchparameter: stepwise addition = random, branch swapping = TBR, Alinierungslücken als 5. Nukleotid bewertet. Baumlänge: 3193, CI: 0,51, RI: 0,79, RC: 0,4.
Cephalaspidea s.str.
Anaspidea
Pteropoda
Anaspidea
Tylodinoidea
Acochlidiacea
Pulmonata
Pyramidellidae
Sacoglossa
Pulmonata
Pleurobranchoidea
Nudibranchia

145
4.10.2 Ernährungsweise Das Erschließen unterschiedlichster Nahrungsquellen und die Entwicklung spezialisierter Ernährungsstrategien werden als eine treibende Kraft in der Evolution der Opisthobranchia angesehen. Dabei ist das Merkmal „Ernährungsweise“ sehr komplex, bestehend aus verschiedenen ökologischen und anatomischen Faktoren, die eine sinnvolle Kodierung für die Berücksichtigung in phylogenetischen Analysen nahezu unmöglich machen. Es reicht zum Beispiel nicht aus, nur zwischen carnivorer und herbivorer Lebensweise zu unterscheiden. Die Anaspidea und Sacoglossa ernähren sich beide ausschließlich von Algen, aber während die Anaspidea ganze Algenteile aufnehmen, saugen die Sacoglossa nur das Cytoplasma aus dem pflanzlichen Gewebe. Verbunden mit der Art und Weise des Nahrungserwerbs ist eine deutlich unterschiedliche Ausprägung der Radula in beiden Guppen. Bei der carnivoren Ernährung muß beispielsweise berücksichtigt werden, ob es sich um aktive Räuber handelt, die vagile Beute jagen (z.B. Aglajidae, Cephalaspidea s.str.) oder ob sessile Invertebraten nur „abgeweidet“ werden (z.B. Nudibranchia). Alternative Ernährungsstrategien repräsentieren Planktonstrudler (Thecosomata), Detritusfresser (Acochlidiacea), Parasiten (Pyramidellidae) und omnivore Formen (Bullidae, Cephalaspidea s.str.). Dabei ist es schwierig zu beurteilen, wie wahrscheinlich der Übergang von der einen zur anderen Lebensweise aus evolutionärer Sicht ist. Erfordert z.B. die Entstehung von herbivoren Weidegängern aus carnivoren Weidegängern ein größeres Maß an morphologischen Verän-derungen als die Entwicklung zur aktiv jagenden Ernährungsweise aus letzteren? Im Folgenden werden die rekonstruierten Verwandtschaftshypothesen mit Hilfe des Merkmals „Ernährungsweise“ auf ihre Plausibilität hin geprüft, Ernährungsdaten sind der Arbeit von Wägele (2004) entnommen. Es wird dabei zwei Aspekten eine besondere Aufmerksamkeit geschenkt, zum einen der Entwicklung eines Kaumagens innerhalb der Opisthobranchia und zum anderen der Ernähungsweise des hypothetischen „Ur-Opisthobranchiers“, bzw. „Ur-Heterobranchiers“. Bis zum heutigen Zeitpunkt herrscht Uneinigkeit über den plesiomorphen Zustand des Merkmals innerhalb der Opisthobranchia. Haszprunar (1985) sieht die Carnivorie als ursprünglich an, Gosliner (1994) vermutet einen omnivoren Weidegänger als Stammlinienvertreter. Mikkelsen (1996) postuliert dagegen in Übereinstimmung mit Purchon (1977) die Herbivorie als plesiomorphe Ausprägung. In den meisten Gruppen der Opisthobranchia ist innerhalb des Ösophagus eine muskuläre Struktur vorhanden, die üblicherweise eine Reihe von chitinigen Platten enthält und als Kaumagen bezeichnet wird. Die Anzahl, Form und Position dieser Platten sind charakteristisch für einzelne Linien innerhalb der Opisthobranchia und enthalten demnach phylogenetische Information (Gosliner 1994).

146
18S rDNA
Acteon sp. Navanax inermis Odontoglaja guamensis Chelidonura inornata Philinopsis pilsbryi Philinoglossa praelongata Sagaminopteron psychedelicum Colpodaspis thomsoni Haminoea hydatis Atys semistriatus Bullacta exarata Phanerophthalmus smaragdinus Smaragdinella sp. Bulla cf. striata cf. Retusa sp. Cylichna gelida Scaphander lignarius Diaphana cf. inflata Toledonia globosa Newnesia antarctica Runcina adriatica Petalifera petalifera Stylocheilus longicauda Dolabrifera brazieri Bursatella leachii Clio pyramidata Creseis acicula Diacria quadridentata Spongiobranchaea australis Akera bullata Siphonota geographica Dolabella auricularia Aplysia californica Umbraculum mediterraneum Tylodina perversa Unela glandulifera Parahedyle cryptophthalma Pontohedyle milatchevitchi Hedylopsis spiculifera Onchidella floridana Onchidium verrucosum Siphonaria alternata Cepaea hortensis Melampus luteus Cassidula nucleus Ovatella firminii Pythia scarabaeus Auriculinella bidentata Pyramidella sulcata Odostomia sp. Elysia viridis Elysiella pusilla Thuridilla bayeri Placobranchus ocellatus Bosellia mimetica Polybranchia orientalis Cyerce nigricans Calliopaea bellula Limapontia nigra Placida cremoniana Lymnaea stagnalis Hydatina physis Bullina lineata Pupa solidula Pleurobranchus peroni Bathyberthella antarctica Euselenops luniceps Tompthompsonia antarctica Berthellina citrina Berthella stellata Berthella platei Chromodoris krohni Hypselodoris villafranca Dendrodoris fumata Bathydoris clavigera Cuthona nana Flabellina babai Dendronotus dalli Fasciolaria lignaria Pisania striata Bursa rana Crepidula adunca Truncatella guerinii Viviparus georgianus Balcis eburnea Melaraphe neritoides Littoraria undulata Littorina striata Nodilittorina punctata Littorina saxatilis Littorina littorea
?
?
Parasiten
Planktonstrudler
herbivor
herbivor/Detritus
carnivor, vagile Beute
carnivor, sessile Beute
unbekannt, wenn nicht
näher bezeichnet
Abb. 38: Projektion von Ernährungsdaten auf die Stammbaumhypothese der Maximum Parsimonie-Analyse (18S rDNA, Opisthobranchia-Datensatz). Strikte Konsensustopologie aus 1.000.000 kürzesten Bäumen, Suchparameter: stepwise addition = random, branch swapping = TBR, Alinierungslücken als 5. Nukleotid bewertet. Baumlänge: 3193, CI: 0,51, RI: 0,79, RC: 0,4.
Cephalaspidea s.str.
Anaspidea
Pteropoda
Anaspidea
Tylodinoidea
Acochlidiacea
Pulmonata
Pyramidellidae
Sacoglossa
Pulmonata
Pleurobranchoidea
Nudibranchia

147
Auf der Grundlage des MP-Baumes (Abb. 38) erscheint die carnivore Lebensweise als plesiomorph, repräsentiert durch die Nudipleura als Weidegänger sessiler Invertebraten und die Acteonoidea als aktive Jäger von Polychaeten. Alternativ könnte sich aus einem omnivoren Vorfahren eine Nahrungsspezialisierung bei den Nudipleura und Acteonoidea entwickelt haben. Eine weniger sparsame, aber durchaus mögliche Erklärung liefert die Annahme eines herbivoren Vorgängers, von dem aus sich unabhängig in zwei Linien die Carnivorie ausgebildet hat. Abgesehen von den Nudipleura und Acteonoidea ist für die übrigen Taxa die Herbivorie mit großer Wahrscheinlichkeit als ursprünglich anzusehen, da sie in allen Sacoglossa, Pulmonata und Anaspidea verwirklicht ist. In verschiedenen evolutionären Linien innerhalb der Heterobranchia ist es zur sekundären Ausbildung anderer Ernährungsstrategien gekommen, z.B. zum Parasitismus bei den Pyramidellidae oder zur Ernährung von Detritus bei den interstitiellen Acochlidiacea. Die Tylodinoidea weiden sessile Organismen ab und aus den herbivoren Anaspidea sind die räuberischen Gymnosomata und planktonstrudelnden Thecosomata entstanden. Der in dieser Verwandtschaftshypothese ursprünglichste Vertreter der Cephalaspidea s.str., Runcina adriatica, weist eine Körpergröße von nur 2-3 mm auf und ernährt sich von Algen und Detritus. Die Nahrungsgrundlage der ebenfalls recht kleinen, paraphyletischen Diaphanidae bilden wahrscheinlich Foraminiferen. Der Stammlinienvertreter der Cephalaspidea s.str. war auf der Grundlage dieser Stammbaumhypothese vermutlich klein und omnivor oder zeigte eine herbivore Lebensweise wie der gemeinsame Vorfahre von Anaspidea und Cephalaspidea s.str. Ausgehend von den carnivoren Diaphanidae haben sich dann innerhalb der Bulloidea (Bullidae, Atyidae, Haminoeidae) wieder pflanzenfressende Formen entwickelt. Diese sekundäre Enstehung der Herbivorie ist nicht sehr befriedigend aber trotzdem nicht unmöglich. Die Entwicklung eines Kaumagens mit chitinigen Platten tritt erstmals in der Stammlinie von Anaspidea und Cephalaspidea s.str. auf, dagegen ist ein schwach ausgebildeter Kaumagen ohne chitinige Platten bereits bei den Acteonoidea und Sacoglossa vorhanden. Die Nudipleura besitzen primär keinen muskulösen Abschnitt im Ösophagus, nur einige Nudibranchia haben in Anpassung an eine spezialisierte Lebensweise eine Muskelpumpe zur Aufnahme von Schwammgewebe entwickelt. Der plesiomorphe Zustand innerhalb der Opisthobranchia ist demnach das Fehlen eines Kaumagens. Die Anaspidea besitzen einen Muskelmagen mit vielen kleinen und mindestens 10 größeren Platten. Bei den Thecosomata muß sich diese Anzahl im Laufe der Evolution auf 4 reduziert haben, während die Gymnosomata alle Hartstrukturen im Ösophagus verloren haben. Unabhängig von den Thecosomata hat in der Stammlinie der Cephalaspidea s.str. eine Reduktion der Kauplatten auf eine Anzahl von 4 bei Runcina adriatica stattgefunden, die Diaphanidae besitzen dagegen keinerlei Kauplatten. Da allerdings mit Ausnahme der Aglajidae alle übrigen Cephalaspidea s.str. drei große Chitinplatten aufweisen, müssen diese sekundär

148
entstanden sein. Dieses evolutionäre Szenario erscheint nicht sehr plausibel und könnte sich auf eine Fehlstellung der Diaphanidae zurückführen lassen. Im Distanzphylogramm auf der Grundlage der 18S rDNA (Abb. 16 und 18B) zeigt sich eine basale Stellung der Thecosomata und von Pontohedyle milatchevitchi (Acochlidiacea). Aufgrund der hochspezialisierten Lebensweise beider Taxa ist ihre Ursprünglichkeit in diesem Stammbaum mit sehr hoher Wahrscheinlichkeit auf Unzulänglichkeiten der Rekonstruktionsmethode und/oder auf das Fehlen eines deutlichen phylogenetischen Signals zurückzuführen. Bleibt die basale Stellung dieser Taxa unberücksichtigt, ergibt sich die Herbivorie als plesiomorpher Zustand innerhalb der Heterobranchia. Die carnivoren Acteonoidea und Nudipleura zeigen einen monophyletischen Ursprung, dabei scheint die Entwicklung von carnivoren Weidegängern aus herbivoren wahrscheinlicher als der Übergang von einer herbivoren zu einer aktiv räuberischen Lebensweise. Bei Berücksichtigung der Thecosomata als basales Taxon würden 4 Chitinplatten in einem muskulösen Kaumagen den plesiomorphen Zustand innerhalb der untersuchten Heterobranchia repräsentieren. Bei den Anaspidea hätte sich diese Anzahl dann auf 10 erhöht und wäre bei Runcina adriatica erhalten geblieben. In der Stammlinie der übrigen Taxa, sowie bei Pontohedyle milatchevitchi muß es zu einer vollständigen Reduktion des Muskelmagens und der Kauplatten gekommen sein, wobei die Existenz eines muskulären Ösophagusabschnitts bei den Acteonoidea als sekundäre Neuentstehung interpretiert werden müsste. Solange die Homologie der muskulären Strukturen bei den Acteonoidea, Anaspidea, Thecosomata und Cephalaspidea s.str. nicht zweifelsfrei belegt ist, spricht nichts gegen die Gültigkeit des bisher beschriebenen evolutiven Szenarios. Die Stellung von Cepaea hortensis (Pulmonata) innerhalb der Cephalaspidea s.str. entbehrt jeder morphologischen Grundlage und wird nicht weiter berücksichtigt. Ausgehend von 4 Chitinplatten bei Runcina adriatica muß es zum völligen Verlust dieser Strukturen gekommen sein und zu einer Neuentstehung von 3 Platten bei der Gruppierung aus Haminoeidae, Cylichnidae, Bullidae, Retusidae und Diaphana cf. inflata. Die postulierte Neuentstehung von Kauplatten ist als sehr unglaubwürdig zu bewerten. Eine von mehreren sparsamsten Erklärungungen bezüglich der zu beobachtenden Ernährungsweisen innerhalb der Cephalaspidea s.str. beinhaltet einen carnivoren Stammlinienvertreter, aus dem sich die herbivoren Runcinidae einerseits und die restlichen Formen andererseits entwickelt haben könnten. Allerdings müsste dann eine dreimal unabhängig entstandene sekundäre Herbivorie angenommen werden, die nicht sehr plausibel erscheint. Alternative Szenarien erscheinen nicht weniger unwahrscheinlich, so dass man von einer fehlerhaften Rekonstruktion der phylogenetischen Beziehungen innerhalb dieser Gruppierung ausgehen kann. Die mit Hilfe des Bayes’schen Verfahrens rekonstruierte Verwandtschaftshypothese (Abb. 17 und 18C) weist eine sehr geringe Auflösung auf und bleibt deshalb an dieser Stelle unberücksichtigt.

149
Beim Vergleich der Topologien auf der Grundlage der 18S rDNA erscheint der MP-Stammbaum etwas plausibler. Das auf der Basis alternativer morphologischer und molekularer Daten rekonstruierte Schwestergruppenverhältnis von Anaspidea + Cephalaspidea s.str. kann durch den MP-Baum gestützt werden, ebenso wie die Monophylie der hochspezialisierten Acochlidiacea. Die postulierte Ursprünglichkeit der Thecosomata und von Pontohedyle milatchevitchi (Acochlidiacea) im Distanzphylogramm entspricht sicherlich nicht der realen Phylogenese, allerdings ist die basale Stellung der Nudipleura im MP-Baum möglicherweise auch auf Unzulänglichkeiten des Rekonstruktionsverfahrens zurückzuführen (siehe Kapitel 4.9.5.3). Beide Stammbaumhypothesen erfordern gleichermaßen die sekundäre Neu-entstehung von Kauplatten innerhalb der Cephalaspidea s.str., davon abgesehen ist das evolutionäre Szenario der Merkmalsentwicklungen in beiden Topologien vorstellbar. Der plesiomorphe Zustand der Ernährungsweise bleibt weiterhin unklar. 28S rDNA Beim Vergleich der Stammbaumhypothesen auf der Grundlage der 28S rDNA erweist sich die mit Hilfe einer Distanzanalyse rekonstruierte Topologie (Abb. 20 und 23B) aus zweierlei Gründen als die wahrscheinlichste. Zum einen werden alle auf der Basis alternativer Daten als monophyletisch angesehenen Gruppen auch als solche rekonstruiert. Im Vergleich dazu erscheinen z.B. die Acochlidiacea im ML- und MP-Baum und die Gruppierung aus Anaspidea und Cephalaspidea s.str. im ML- und Bayes’schen Baum als Paraphyla. Der zweite Grund für die Bevorzugung der Distanztopologie besteht darin, dass sie im Vergleich zu den alternativen Stammbäumen die wenigsten Merkmalsänderungen bezüglich der Ernährungsweise erfordert. In Übereinstimmung mit dem auf der Grundlage der 18S rDNA rekonstruierten MP-Baum erscheinen die Nudipleura und Acteonoidea als basalste Taxa innerhalb der Heterobranchia. Damit kommen für den plesiomorphen Zustand der Ernährungsweise wiederum sowohl die Omnivorie als auch die Herbi- und Carnivorie in Frage. Der hypothetische Vorfahr der Anaspidea und Cephalaspidea s.str. war sicherlich ein Pflanzenfresser, von dem ausgehend sich innerhalb der Cephalaspidea s.str. einmal aktiv jagende Formen entwickelt haben (Aglajidae, Gastropteridae) und einmal carnivore Weidegänger (Retusidae). Ein muskulöser Kaumagen mit chitinigen Platten tritt anhand der Verwandtschafts-hypothese zum erstenmal in der Stammlinie der Anaspidea und Cephalaspidea s.str auf, dabei existieren im plesiomorphen Zustand entweder viele Platten wie bei den Anaspidea oder 4 wie bei Runcina adriatica. In der weiteren Entwicklung der Cephalaspidea hat sich eine Anzahl von 3 Platten konsolidiert, die dann bei den Aglajidae und Gastropteridae sekundär reduziert worden sind.

150
Das Vorhandensein eines Muskelmagens ohne Kauplatten bei den Acteonoidea repräsentiert entweder den plesiomorphen Zustand für alle Taxa außer den Nudipleura oder ist eine Konvergenzerscheinung zu den Anaspidea und Cephalaspidea s.str. Die Plesiomorphie des Merkmals setzt dann allerdings eine Reduktion in der Stammlinie von Sacoglossa, Tylodinoidea, Acochlidiacea, Pulmonata und Odostomia sp. (Pyramidellidae) voraus. Das beschriebene evolutionäre Szenario weist keine signifikanten Ungereimtheiten auf, allerding wurden einige wichtige Taxa, z.B. die Pteropoda und Diaphanidae nicht berücksichtigt. 16S rDNA Bei den Stammbaumhypothesen basierend auf der 16S rDNA ist die Benennung einer bevorzugten Topologie nicht ohne weiteres möglich. Die aufgrund alternativer Daten als unwahrscheinlich bewertete Paraphylie der Nudipleura tritt in allen vier Stammbäumen auf, ebenso wie die basale Stellung von Clione limacina (Gymnosomata), die ebenfalls nicht mit den Ergebnissen morphologischer Analysen zu vereinbaren ist. Im MP-Baum (Abb. 24 und 28A) stellt die herbivore Ernährungsweise, bei Nichtberücksichtigung von Clione limacina, zweifellos den ursprünglichen Zustand innerhalb der Heterobranchia dar. Ausgehend davon haben sich möglicherweise dreimal unabhängig carnivore Formen entwickelt und zwar in der Stammlinie der Gruppierung aus Nudipleura und Acteonoidea, bei den Tylodinoidea und mindestens einmal innerhalb der Cephalaspidea s.str. Die Stellung von Acteon tornatilis innerhalb der Cephalaspidea s.str. wurde bereits in Kapitel 4.9.5.5 diskutiert und wird als das Ergebnis einer Verwechslung oder Fehlbestimmung angesehen. Durch diese Fehlplazierung wird die Interpretation der Merkmalsentwicklung in dieser Gruppe erschwert. Für das Monophylum aus Nudipleura, Acteonoidea, Cephalaspidea s.str., Anaspidea und Tylodinoidea ist das Vorhandensein eines Muskelmagens mit Kauplatten als plesiomorph anzusehen. Dabei kommen als ursprüngliche Anzahl entweder die 10 der Anaspidea oder die 3 der Cephalaspidea s.str. in Frage. In jedem Fall muß es bei den Nudipleura, Acteonoidea und Tylodinoidea zu einer vollständigen Reduktion aller Hartstrukturen des Kaumagens gekommen und bei Cavolinia sp. (Thecosomata) zu einer Parallelentstehung von 4 Chitinplatten. Eine alternative Erklärung liefert die Annahme von 4 Platten im plesiomorphen Zustand für alle Taxa, eine anschließende Reduktion auf 3 bei den Cephalaspidea s.str. und eine Erhöhung auf 10 bei den Anaspidea. Diese Hypothese erfordert den unabhängigen Verlust dieser Strukturen bei den Acochlidiacea, Pulmonata und Pyramidellidae sowie bei den Nudipleura, Acteonoidea und Tylodinoidea. Beide potentiellen Evolutionsgeschehen erscheinen nicht sehr plausibel und deuten möglicherweise eine nicht korrekt rekonstruierte Stellung der Thecosomata an.

151
In der durch eine Distanzanalyse rekonstruierten Topologie (Abb. 25 und 28B) liefert die Annahme eines carnivoren Vorfahren die sparsamste Erklärung für die zu beobachtende Merkmalsverteilung. Ausgehend davon hätte sich dann unabhängig bei den Anaspidea, einigen Cephalaspidea s.str., den Sacoglossa sowie bei den Pulmonata und Acochlidiacea die Herbivorie entwickelt. Die Annahme von zahlreichen Konvergenzen erscheint an sich nicht sehr plausibel, doch gerade die Opishobranchia zeichnen sich durch eine Vielzahl an parallelen Entwicklungen aus, z.B. durch den mehrfach unabhängig entstandenen Verlust der Schale. Die Existenz von Kauplatten bei den Anaspidea, Cephalaspidea s.str. und Thecosomata ist anhand der postulierten Verwandtschaftshypothese nicht befriedigend zu erklären. Entweder muß eine unabhängige Entstehung bei den Thecosomata einerseits und den Anaspidea und Cephalaspidea s.str. andererseits angenommen werden oder man muß das Fehlen dieser Strukturen bei den übrigen Gruppen auf sekundäre Reduktionen zurückführen. Das erste Szenario liefert die sparsamere Erklärung und wird z.B. von Lalli & Gilmer (1989) und Gosliner (1994) favorisiert. In der durch eine maximum likelihood-Analyse erhaltenen Topologie (Abb. 26 und 28C) wird ebenfalls die Existenz eines carnivoren Vorfahren für die untersuchten Taxa unterstützt. Im Vergleich zum Stammbaum der Distanzanalyse erfordert der ML-Baum dabei nur eine dreimal unabhängige Entstehung der Herbivorie. Ein deulich ausgeprägter Kaumagen hat sich in dieser Verwandtschaftshypothese in der Stammlinie der Gruppierung aus Thecosomata, Anaspidea, Cephalaspidea s.str., Tylodinoidea, Nudipleura und Pupa solidula (Acteonoidea) entwickelt und wurde unabhängig sowohl bei den Nudipleura, Tylodinoidea und innerhalb der Cephalaspidea s.str. wieder reduziert. Der plesiomorphe Zustand der Kauplattenanzahl beträgt entweder 3 (Cephalaspidea s.str.), 4 (Thecosomata) oder 10 (Anaspidea). Die mit Hilfe des Bayes’schen Verfahrens auf der Grundlage der 16S rDNA rekonstruierte Verwandtschaftshypothese wird aufgrund ihrer geringen Auflösung nicht diskutiert. Beim Vergleich der Topologien auf der Grundlage der 16S rDNA erscheint der ML-Stammbaum bezüglich der Merkmalsentwicklung des Kaumagens plausibler als die beiden anderen Verwandtschaftshypothesen, da er weniger evolutive Veränderungen erfordert. Die Frage nach der ursprünglichen Ernährungsweise kann aufgrund widersprüchlicher Ergebnisse nicht eindeutig beantwortet werden.

152
4.10.3 Chromosomenzahlen Chromosomenzahlen haben sich innerhalb der Opisthobranchia als extrem konservativ erwiesen (Schmekel 1985) und ermöglichen daher phylogenetische Rückschlüsse auf hoher taxonomischer Ebene. In der Tabelle X (Anhang X) sind die Chromosomenzahlen aller untersuchten Taxa der Heterobranchia angegeben und in Abb. 39 auf eine Stammbaumhypothese (18S rDNA: MP-Analyse) projiziert. 18S rDNA Die mit Abstand am häufigsten zu beobachtende chromosomale Anzahl innerhalb der Heterobranchia liegt bei 16-18 und könnte den plesiomorphen Zustand repräsentieren. Allerdings weisen gerade die als basal rekonstruierten Nudipleura Werte von 12-13 auf und auch die Acteonoidea weichen mit 15 Chromosomen im haploiden Satz vom „Durchschnitt“ ab. Die Annahme eines Stammlinienvertreters mit 17 Chromosomen und einer anschließenden unabhängigen Reduktion bei den Nudipleura und Acteonoidea liefert dabei einen ebenso sparsame Erklärung wie die Entstehung von 17 aus 15 Chromosomen. Gegen die Plesiomorphie von 12 oder 13 Chromosomen spricht möglicherweise die Existenz von 15-17 bei der Außengruppe, den Littorinidae. Für alle Taxa der Innengruppe mit Ausnahme der Nudipleura und Acteonoidea ist sicherlich eine Anzahl von 17 Chromosomen als ursprünglich zu betrachten. Ausgehend davon haben sich in einigen Gruppen apomorphe Abweichungen ergeben, z.B. eine Erhöhung der Chromosomenzahl innerhalb der Stylommatophora (22-30) oder eine zahlenmäßige Variabilität innerhalb der Thecosomata (10-18). Der monophyletische Ursprung der Nudipleura kann durch die festgestellte Chromosomenverteilung deutlich gestützt werden. Besonders aufschlussreich wäre die Kenntnis der Chromosomenzahl bei den Tylodinoidea, da bei einer Anzahl von 17 die diskutierte Verwandtschaft zu den Nudipleura mit hoher Wahrscheinlichkeit ausgeschlossen werden könnte. Insgesamt ist das erhaltene evolutionäre Gesamtbild als durchaus vorstellbar anzusehen. Das Distanzphylogramm wurde auf der Grundlage des Vergleichs mit alternativen Daten als weniger plausibel bewertet und wird hier nicht weiter diskutiert. 28S rDNA Aus oben erwähnten Gründen wurde auf der Basis der 28S rDNA die Distanztopologie favorisiert, die übrigen Stammbaumhypothesen bleiben bei der Plausibilitätsüberprüfung mit Hilfe der Chromosomendaten unberücksichtigt.

153
In Übereinstimmung der MP-Topologie auf der Grundlage der 18S rDNA kann für den plesiomorphen Zustand innerhalb der Opisthobranchia eine Chromosomenzahl von 15 oder 17 als wahrscheinlich betrachtet werden. Bei der postulierten Merkmalsentwicklung ergeben sich aufgrund der Konserviertheit der Chromosomenzahlen keine Ungereimtheiten. Abweichungen vom Grundmuster, z.B. bei Cepaea hortensis (Pulmonata), können als Apomorphien angesehen werden. 16S rDNA In allen Topologien auf der Grundlage der 16S rDNA erscheint unter Berücksichtigung des Parsimonie-Prinzips eine Chromosomenzahl von 17 als der wahrscheinlichste plesiomorphe Zustand. Die basale Stellung von Clione limacina (Gymnosomata) wurde dabei nicht berücksichtigt (siehe Kapitel 4.9.5.6). Die Paraphylie der Nudipleura und Acteonoidea erfordert eine höhere Anzahl von Merkmalsänderungen als die auf der Grundlage der beiden anderen Datensätze rekonstruierte Monophylie der Taxa. Im ML-Baum (Abb. 26 und 28C) muß ausgehend von 17 Chromosomen für die Stammlinie der Gruppierung aus Nudipleura und Pupa solidula (Acteonoidea) eine Reduktion auf 12-13 Chromosomen und eine sekundäre Erhöhung auf 15 bei Pupa solidula angenommen werden. Die Reduktion auf 15 Chromosomen bei Hydatina physis (Acteonoidea) ist als konvergent entstanden zu betrachten. Auf der Grundlage der MP- und Distanztopologie sind genauso viele Merkmalsänderungen nötig um die terminalen Zustände zu erklären. Das Merkmal „Chromosomenanzahl“ ist daher nicht geeignet, um die unterschiedlichen Topologien anhand ihrer Plausibilität zu bewerten. Die postulierten evolutionären Szenarien erscheinen übereinstimmend unwahrscheinlich. 4.11 Phylogenie der Anaspidea Die Untersuchung der verwandtschaftlichen Beziehungen innerhalb der Anaspidea stützt sich in erster Linie auf die Anaspidea-Datensätze (18S rDNA, ITS1-5.8S-ITS2), mitberücksichtigt werden die Ergebnisse auf der Grundlage des 16S rDNA-Datensatzes der Opisthobranchia. Die Stellung der Gattung Akera innerhalb der Anaspidea wurde aufgrund ihrer zahlreichen plesiomorphen Merkmale lange in Frage gestellt (Thiele 1931, Marcus 1972, Thompson 1976, Schmekel 1985), ist heute allerdings als gesichert anzusehen. Die monogenerischen Akeridae bilden dabei die Schwestergruppe zu allen übrigen Anaspidea, den Aplysiidae (Mikkelsen 1996, Medina & Walsh 2000, Dayrat & Tillier 2002).

154
Acteon sp. Navanax inermis Odontoglaja guamensis Chelidonura inornata Philinopsis pilsbryi Philinoglossa praelongata Sagaminopteron psychedelicum Colpodaspis thomsoni Haminoea hydatis Atys semistriatus Bullacta exarata Phanerophthalmus smaragdinus Smaragdinella sp. Bulla cf. striata cf. Retusa sp. Cylichna gelida Scaphander lignarius Diaphana cf. inflata Toledonia globosa Newnesia antarctica Runcina adriatica Petalifera petalifera Stylocheilus longicauda Dolabrifera brazieri Bursatella leachii Clio pyramidata Creseis acicula Diacria quadridentata Spongiobranchaea australis Akera bullata Syphonota geographica Dolabella auricularia Aplysia californica Umbraculum mediterraneum Tylodina perversa Unela glandulifera Parahedyle cryptophthalma Pontohedyle milatchevitchi Hedylopsis spiculifera Onchidella floridana Onchidium verrucosum Siphonaria alternata Cepaea hortensis Melampus luteus Cassidula nucleus Ovatella firminii Pythia scarabaeus Auriculinella bidentata Pyramidella sulcata Odostomia sp. Elysia viridis Elysiella pusilla Thuridilla bayeri Placobranchus ocellatus Bosellia mimetica Polybranchia orientalis Cyerce nigricans Calliopaea bellula Limapontia nigra Placida cremoniana Lymnaea stagnalis Hydatina physis Bullina lineata Pupa solidula Pleurobranchus peroni Bathyberthella antarctica Euselenops luniceps Tompthompsonia antarctica Berthellina citrina Berthella stellata Berthella platei Chromodoris krohni Hypselodoris villafranca Dendrodoris fumata Bathydoris clavigera Cuthona nana Flabellina babai Dendronotus dalli Fasciolaria lignaria Pisania striata Bursa rana Crepidula adunca Truncatella guerinii Viviparus georgianus Balcis eburnea Melaraphe neritoides Littoraria undulata Littorina striata Nodilittorina punctata Littorina saxatilis Littorina littorea
12
13
17
16-17
17-18
15
10-18
16
17-18
16
17-19
18
22
17
15-17
Cephalaspidea s.str.
Anaspidea
Pteropoda
Anaspidea
Tylodinoidea
Acochlidiacea
Pulmonata
Pyramidellidae
Sacoglossa
Pulmonata
Pleurobranchoidea
Nudibranchia
Abb. 39: Projektion von Chromosomenzahlen auf die Stammbaumhypothese der Maximum Parsimonie-Analyse (18S rDNA, Opisthobranchia-Datensatz). Strikte Konsensustopologie aus 1.000.000 kürzesten Bäumen, Suchparameter: stepwise addition = random, branch swapping = TBR, Alinierungslücken als 5. Nukleotid bewertet. Baumlänge: 3193, CI: 0,51, RI: 0,79, RC: 0,4.

155
Dieses Schwestergruppenverhältnis ist, mit Ausnahme der Bayes’schen Analyse, in allen Topologien des 16S rRNA-Gens zu beobachten, weiterhin stellen Aplysia und Dolabella die Schwestergruppe von Akera in den Topologien auf der Grundlage der 28S rDNA dar. Basierend auf dem ITS1-5.8S-ITS2 – Datensatz konnte dieses Verwandtschaftsverhältnis ebenfalls bestätigt werden. Dagegen gruppiert Akera sowohl im Distanzphylogramm der 18S rDNA des Opisthobranchia-Datensatzes, als auch in allen Stammbäumen des Anaspidea-Datensatzes (18S rDNA) innerhalb der Aplysiidae (inklusive Spongiobranchaea, Gymnosomata). Auf der Grundlage aller bisherigen Erkenntnisse wird das Schwestergruppen-verhältnis von Akeridae und Aplysiidae als gesichert angesehen, Diskrepanzen in den vorliegenden Topologien werden auf fehlende Informationen im Datensatz und/oder Unzulänglichkeiten der Rekonstruktionsverfahren, z.B. long branch attraction, zurückgeführt. Die verwandtschaftlichen Beziehungen der Anaspidea zu den Thecosomata und Gymnosomata können nur auf der Grundlage der 18S rDNA-Datensätze untersucht werden. Allerdings kann über die Mono- oder Paraphylie der Pteropoda und Anaspidea zum jetzigen Zeitpunkt noch keine Aussage getroffen werden, da sich ihre phylogenetischen Beziehungen als sehr variabel darstellen. Nur die Monophylie der Thecosomata kann als relativ gesichert angesehen werden. Innerhalb der Aplysiidae werden taxonomisch die Aplysiinae (Aplysia, Syphonota), Dolabellinae (Dolabella), Notarchinae (Stylocheilus, Bursatella) und Dolabriferinae (Dolabrifera, Petalifera) unterschieden (Eales 1944, Pruvot-Fol 1954, Beeman 1968, Marcus 1972), die von einigen Autoren zu den Longicommissurata (Aplysiinae, Dolabellinae) und Brevicommissurata (Dolabriferinae, Notarchinae) zusammen-gefasst werden (Pruvot-Fol 1954, Marcus 1972). In der umfassenden morphologischen Untersuchung der Aplysiidae von Klussmann-Kolb (2004), erscheinen die Aplysiinae, Notarchinae und Dolabriferinae als monophyletische Einheiten, die Dolabellinae sind nur mit einer Art vertreten. Die als plesiomorph erkannten Merkmale der Longicommissurata führen zu deren Paraphylie in der Stammbaumhypothese, die Brevicommissurata bilden eine monophyletische Einheit. In den Stammbaumhypothesen auf der Grundlage der 18S rDNA sind die Aplysiidae übereinstimmend monophyletisch und auf der Basis mitochondrialer Gene (Medina & Walsh 2000) stellen die beiden untersuchten Arten von Aplysia ebenfalls Schwestertaxa dar. Basierend auf dem ITS1-5.8S-ITS2 – Datensatz, erscheinen sie zwar zum Teil als Paraphyla, doch in Anbetracht der Vielzahl stützender Topologien basierend auf morphologischen und molekularen Daten, erscheint ihre Monophylie sehr wahrscheinlich. Die Notarchinae repräsentieren in allen erzeugten Topologien eine paraphyletische Gruppierung, mit Ausnahme des MP-Baumes auf der Grundlage der 16S rDNA. In der Stammbaumhypothese von Medina & Walsh (2000) erscheinen sie je nach Gewichtung von Transversionen zu Transitionen mono- oder paraphyletisch. Da die

156
morphologische Unterstützung für das Schwestergruppenverhältnis von Bursatella und Stylocheilus ebenfalls recht schwach ist (Klussmann-Kolb 2004), muß eine Paraphylie der Notarchinae in Betracht gezogen werden. Allerdings weisen Bursatella und Petalifera im ITS1-5.8S-ITS2 – Datensatz erhöhte Substitutionsraten auf, ihr Schwestergruppenverhältnis in den erstelltenTopologien könnte sich also auch auf eine long branch attraction zurückführen lassen. Die Dolabriferinae bilden in sämtlichen Stammbäumen auf der Grundlage des ITS1-5.8S-ITS2 – Datensatzes eine monophyletische Gruppierung mit Bursatella, stellen also ein Paraphylum dar. In den Topologien der 18S rDNA sind ihre Verwandtschaftsverhältnisse entweder nicht aufgelöst oder sie erscheinen ebenfalls als Paraphylum. Auf der Grundlage morphologischer Daten werden die Dolabriferinae durch eine große Anzahl synapomorpher Merkmale charakterisiert, in den Stammbäumen auf der Basis mitochondrialer Gene (Medina & Walsh 2000) konnte ihre Monophylie dagegen ebenfalls nicht rekonstruiert werden. Die von Klussmann-Kolb (2004) postulierte Monophylie der Brevicommissurata konnte ebenfalls nicht durch molekulare Daten gestützt werden, dagegen ist ein Monophylum aus Brevicommissurata und Dolabellinae in allen Topologien des ITS1-5.8S-ITS2 – Datensatzes, sowie im Stammbaum von Medina & Walsh (2000) zu beobachten. Beim Vergleich der auf unterschiedlichen Genen und Rekonstruktionsverfahren beruhenden Topologien, ist eine große Variabilität in den Verwandtschafts-beziehungen der einzelnen Taxa zu beobachten. Möglicherweise beinhalten alle untersuchten Genabschnitte nicht genug phylogenetische Information, um die Beziehungen innerhalb der Anaspideen aufzulösen. Zur endgültigen Klärung, bzw. Stützung der von Klussmann-Kolb (2004) postulierten Verwandtschaftsbeziehungen, erscheint die Analyse weiterer anatomischer und histologischer Daten sinnvoller als die Untersuchung alternativer Sequenzabschnitte. Beim Vergleich der beiden verwendeten Alinierungen, zeigt sich der ITS1-5.8S-ITS2 – Datensatz als den auf morphologischen Daten basierenden Ergebnissen besser entsprechend. Zudem weist er eine größere Unabhängigkeit vom verwendeten Rekonstruktionsverfahren auf, das spricht für ein deutlicheres phylogenetisches Signal. Durch die Analyse mit PHYSID wird offensichtlich, dass der ITS1-5.8S-ITS2 – Datensatz ein stärkeres phylogenetisches Signal aufweist, als die 18S rDNA-Alinierung. Letztere scheint für das untersuchte taxonomische Niveau zu konserviert zu sein. 4.12 Phylogenie von Phyllodesmium Die durchgeführten phylogenetischen Rekonstruktionen zeigen übereinstimmend sowohl die Monophylie der Facelinidae, als auch die der Gattung Phyllodesmium. Die Facelinidae stellen innerhalb der Aeolidoidea eine sehr abgeleitete Gruppierung dar (Willan 1987), eine Tatsache, die durch den extrem langen Ast im Phylogramm

157
(Z-Werte von 12,1 bis 16,6) und die große Anzahl potentieller Apomorphien (341, davon 89 verrauscht, 222 symmetrisch, 30 asymmetrisch) deutlich wird. Die Monophylie von Phyllodesmium wird durch 331 (329/1/1) potentielle Apomorphien gestützt. Die zu beobachtenden Verwandtschaftsverhältnisse innerhalb der Gattung Phyllodesmium zeigen keine Abhängigkeit vom gewählten Rekonstruktionsverfahren, in allen Topologien stellt P. cf. magnum die Schwestergruppe der übrigen fünf Arten dar, die zwei Monophyla, bestehend aus P. crypticum und P. jakobsenae einerseits und P. colemani, P. longicirrum und P. briareum andererseits bilden. P. crypticum und die neu beschriebene Art P. jakobsenae (Burghardt & Wägele 2004) ernähren sich beide, wie die hier nicht untersuchten Arten P. pecten und P. hyalinum, ausschließlich von der Weichkorallen-Gattung Xenia (Rudman 1991). Laut Rudman (1981, 1991) befinden sich diese Arten in einem Frühstadium einer Symbiose mit Zooxanthellen, da die Verzweigung der Mitteldarmdrüse nicht sehr ausgeprägt ist und die aufgenommenen Algen nur temporär und in nicht-modifizierten Zellen gespeichert werden. Für die Art P. jakobsenae konnte dagegen ein hoher Verzweigungsgrad der Mitteldarmdrüse festgestellt werden und auf der Grundlage von Hungerversuchen konnte eine hohe Effizienz der Symbiose gezeigt werden. Die Arten P. colemani, P. longicirrum und P. briareum zeigen nach Rudman (1991) übereinstimmend einen höheren Anpassungsgrad an eine symbiontische Beziehung. Bei ihnen zeigen sich feine Verästelungen der Mitteldarmdrüse, die alle unbeschattenen Bereiche der Körperwand durchziehen. Die Zooxanthellen befinden sich bei ihnen in modifizierten „Containerzellen“ oder frei in kleinen Kanälen der Mitteldarmdrüse. Die beiden rekonstruierten Monophyla repräsentieren deutlich die beiden, von Vertetern ohne Symbiose ausgehenden, unterschiedlichen Evolutionslinien innerhalb der Gattung (Rudman 1991). Die kategorische Einteilung in Taxa mit weniger und mit höher effizienter Symbiose erweist sich durch Untersuchungen an P. jakobsenae jedoch als nicht zutreffend. Die Stellung von P. cf. magnum als Schwestertaxon der übrigen Arten stimmt nicht mit der von Rudman vorgeschlagenen Phylogenie der Gattung überein, denn die Art gehört aufgrund morphologischer Übereinstimmungen eindeutig in den Verwandtschaftskreis von P. colemani, P. longicirrum und P. briareum. Allerdings repräsentiert P. magnum nach Rudman (1991) innerhalb der letztgenannten evolutionären Linie einen eher ursprünglichen Vertreter, seine Stellung im hier rekonstruierten Stammbaum bildet demnach keinen sehr starken Widerspruch zu der von Rudman angenommenen Stellung. Unabhängig von den Ergebnissen der molekularen Untersuchungen, erscheint die Zuordnung des Individuums „Phyllodesmium cf. magnum“ zu der Art P. magnum aufgrund morphologischer Unterschiede fragwürdig (Burghardt, persönliche Mitteilung), möglicherweise handelt es sich dabei um eine neue, noch nicht beschriebene Art.

158
4.13 Umbraculum Aufgrund der hohen Konserviertheit der 18S rDNA, ist die völlige Übereinstimmung der drei Sequenzen von Umbraculum (von der Großen Meteor Bank, den Azoren und aus Australien) nicht unerwartet. Da aber die Existenz von hochvariablen Insertionen bei Vertretern der Opisthobranchia, z.B. bei den Pleurobranchoidea, nachgewiesen ist, wurde dieses Gen mitberücksichtigt. Bei der 16S rDNA beträgt die Sequenzdivergenz zwischen den drei Umbraculum-Individuen nur 0,4 %. Bei Austrodoris kerguelenensis (Nudibranchia) konnte im Vergleich dazu z.B. eine intraspezifische Variabilität von 1,5 % festgestellt werden. Die höchste Sequenzdivergenz der drei Umbraculum-Individuen ist mit 2,3 % innerhalb der Sequenzen der Cytochromoxidase zu beobachten. Dieser Wert liegt im Rahmen der innerartlichen Variabilität bei anderen Taxa. Bucklin et al. (2003) fanden z.B. bei der Untersuchung der COI von Copepoden (Crustacea) eine intraspezifische Variabilität von 1-4 %, die Divergenz zwischen verschiedenen Arten lag bereits bei 9-25 %. Einen weiteren Beleg für das Vorliegen einer einzigen Art bildet die Tatsache, dass eine größere genetische Ähnlichkeit zwischen den Individuen von der Großen Meteor Bank und Australien, als zwischen denen des Nordatlantiks (Große Meteor Bank und Azoren) vorliegt. Es sind keine Strömungen oder geographisch bedingten Barrieren bekannt, die einen Genaustausch zwischen den beiden Lokalitäten im Atlantik verhindern könnten, demnach lassen sich die Unterschiede nur als zufällige Übereinstimmungen aufgrund von intraspezifischer Variabilität interpretieren. Die morphologischen Untersuchungen (Wägele et al. im Druck) der verschiedenen Individuen von Umbraculum ergaben geringe anatomische Unterschiede, z.B. bezüglich der Größe und Ausdehnung der Manteldrüse, der Eingänge der Mitteldarmdrüse und der Position der Gonade. Da ein gewisses Maß an Variabilität auch unter den verschiedenen Individuen eines Fundortes (Große Meteor Bank) zu beobachten ist, werden die beobachteten Unterschiede auf die Existenz intraspezifischer Variabilität zurückgeführt. Unter Berücksichtigung aller anatomischen, histologischen und molekularen Ergebnisse, werden die hier untersuchten Individuen von Umbraculum als Mitglieder einer Art, Umbraculum umbraculum (Lightfoot, 1786) betrachtet. 4.14 Zusammenfassung, Schlußwort und Ausblick Der Schwerpunkt der vorliegenden Arbeit lag in der Rekonstruktion der Phylogenie der Opisthobranchia mit Hilfe von 18S, 28S und 16S rDNA-Sequenzen. Der 18S rDNA-Datensatz umfasst die kompletten Sequenzen von 78 Vertretern der Opisthobranchia, Pulmonata und Pyramidellidae, sowie von 13 „Prosobranchiern“ (Außengruppe). Die Anwendung statistischer Tests ergab eine abweichende

159
Basenzusammensetzung bei den Cladobranchia (Nudibranchia), ebenso wie die höchsten Substitutionsraten dieses Taxons. Weiterhin konnte eine signifikant erhöhte Evolutionsgeschwindigkeit bei den Anthobranchia (Nudibranchia), Pleurobranchoidea und in geringerem Maße bei den Acteonoidea festgestellt werden. Die Analyse der Substitutionssättigung ergab für die Transversionen eine Sättigung im Bereich hoher Sequenzdivergenz, im Wesentlichen verursacht durch die Sequenzen der Cladobranchia und von Berthella stellata (Pleurobranchoidea). Aufgrund der hohen Konserviertheit der 18S rDNA (> 50% konstante Nukleotidpositionen) und der ausgeprägten positionsspezifischen Ratenvariabilität ergab die Analyse des phylogenetischen Signals erwartungsgemäß nur eine sehr geringe Anzahl potentieller Apomorphien für die jeweiligen hypothetischen Monophyla. Die mit Hilfe von MP-, Distanz-, ML- und Bayes’schen Verfahren rekonstruierten Stammbäume zeigen mit Ausnahme der Nudipleura (Nudibranchia + Pleurobranchoidea) eine deutliche Variabilität in den postulierten Verwandtschafts-beziehungen der opisthobranchiaten Gruppierungen. Die einzelnen Taxa können dagegen im Wesentlichen als Monophyla gestützt werden. Die Opisthobranchia, Pulmonata und Euthyneua repräsentieren übereinstimmend Paraphyla. Durch den Vergleich mit alternativen morphologischen und molekularen Daten erscheint der MP-Baum plausibler als die Stammbaumhypothese der Distanzanalyse, die Bayes’sche Topologie zeigt eine sehr geringe Auflösung. Der zweite Datensatz umfasst 54 partielle 28S rDNA-Sequenzen, davon gehören 43 den Opisthobranchia an, 5 den Pulmonata, eine den Pyramidellidae und 5 den „Prosobranchia“. Knapp 40% der Nukleotidpositionen erwiesen sich als konstant, dabei ergab die Analyse des phylogenetischen Signals eine etwas höhere Anzahl stützender Positionen als beim 18S rDNA-Datensatz. Abgesehen von den Nudibranchia zeigte sich eine homogene Basenzusammensetzung, bei der Untersuchung der relativen Substitutionsraten fielen wiederum die Cladobranchia, Anthobranchia, Pleurobranchoidea und Acteonoidea durch signifikant erhöhte Werte auf. Die graphische Darstellung der Substitutionssättigung ergab mit zunehmender korrigierter Distanz eine leichte Sättigung von Transitionen und Transversionen. Beim Vergleich der durch die unterschiedlichen phylogenetischen Verfahren erhaltenen Topologien erwies sich das Distanzphylogramm aufgrund von Überein-stimmungen mit den Ergebnissen morphologischer Untersuchungen und den Ergebnissen des character mapping als am plausibelsten. Der 16S rDNA-Datensatz beinhaltet 43 Innengruppen- (Opisthobranchia, Pulmonata, Pyramidellidae) und 2 Außengruppensequenzen. Die Basenzusammensetzung erwies sich als homogen, ebenso wie die Verteilung der relativen Evolutionsraten. Nur Clione limacina (Gymnosomata) zeigte eine höhere Evolutionsgeschwindigkeit als die meisten übrigen Taxa. Bei der Analyse der Substitutionssättigung konnte eine größere Anzahl von Transversionen als von Transitionen festgestellt werden, die Transitionen wiesen dabei eine stärkere Sättigung auf. Der untersuchte Abschnitt der 16S rDNA enthielt im Vergleich zu den beiden anderen Datensätzen nach

160
quantitativen Maßstäben das stärkste phylogenetische Signal und den geringsten Anteil an konstanten Positionen (25%). Beim Vergleich der erstellten Topologien erscheint der ML-Stammbaum auf der Grundlage des character mapping plausibler als die anderen Verwandtschafts-hypothesen, da er weniger evolutive Veränderungen erfordert. Abgesehen davon ist die Benennung einer bevorzugten Topologie nicht ohne weiteres möglich, da in allen Bäumen gleichermaßen Inkongruenzen zu den Ergebnissen alternativer molekularer und morphologischer Analysen auftreten. Abschließend betrachtet müssen sowohl die Stellung der Opisthobranchia innerhalb der Heterobranchia, als auch die phylogenetischen Beziehungen der meisten ihrer Subtaxa als ungeklärt angesehen werden. Allerdings konnte die Monophylie aller opisthobranchiaten Hauptlinien durch die meisten Analysen bestätigt werden. Darüberhinaus erscheint ein Schwestergruppenverhältnis von Nudibranchia und Pleurobranchoidea einerseits und Cephalaspidea s.str. und Anaspidea andererseits als sehr wahrscheinlich. Der Vergleich der Ergebnisse im Hinblick auf die unterschiedlichen Rekonstruktions-verfahren lässt keinen Schluß auf die Eignung der verschiedenen Methoden zu. Basierend auf dem 18S rDNA-Datensatz erscheint der MP-Baum am plausibelsten, bei der 28S rDNA-Alinierung ist es das Distanzphylogramm und auf der Grundlage der 16S rDNA schließlich der ML-Baum. Die geringe Auflösung aller Bayes’schen Bäume repräsentiert die Widersprüchlichkeit der Ergebnisse und das Fehlen von phylogenetischer Information. Es ist ebenfalls nicht möglich, eines der Gene als besser geeignet zu bewerten, wahrscheinlich haben die untersuchten DNA-Bereiche alle nicht das Potential, um Verwandtschaftsbeziehungen auf dieser Hierarchieebene aufzulösen. Möglicher-weise lässt sich das Fehlen eines phylogenetischen Signals auch nicht durch längere Sequenzen und mehr Taxa kompensieren, so dass eine Analyse anderer Gene oder anderer Datenquellen, z.B. der mitochondriale Genanordnung, notwendig wird. Einige vielversprechende Gene die in zukünftigen Arbeiten untersucht werden könnten sind z.B. tRNAs oder Entwicklungsgene, wie die Hox Gene (z.B. Degnan et al. 1995). Weitere Quellen für phylogenetische Untersuchungen könnten Schalenproteine (z.B. Bowen & Tang 1996), Hämocyanine (z.B. Lieb et al. 2000), Histone (z.B. Okusu et al. 2003, Huff et al. 2004) oder das Radulamyoglobin (Brito et al. 2001) darstellen. Eines der drei zusätzlichen Projekte die im Rahmen dieser Arbeit durchgeführt wurden stellt die Rekonstruktion der Phylogenie der Anaspidea dar. Zu diesem Zweck wurden das komplette 18S rDNA-Gen sowie der Bereich ITS1-5.8S-ITS2 von mehreren Vertretern der Anaspidea, Thecosomata und Gymnosomata sequenziert. Neben der Aufklärung der Phylogenie sollte die Eignung der beiden unterschiedlich schnell evolvierenden DNA-Bereiche für eine Untersuchung auf dieser taxo-nomischen Ebene geprüft werden.

161
Bei den phylogenetischen Analysen war in Abhängigkeit vom Rekonstruktions-verfahren und dem zugrunde liegenden Datensatz eine große Variabilität in den Verwandtschaftsbeziehungen der einzelnen Taxa der Anaspidea zu beobachten. Auch die stammesgeschichtlichen Beziehungen zwischen den Pteropoda und Anaspidea konnten aufgrund widersprüchlicher Ergebnisse nicht geklärt werden. Beim Vergleich der beiden verwendeten Alinierungen zeigte sich der ITS1-5.8S-ITS2 – Datensatz als den auf morphologischen Daten basierenden Ergebnissen besser entsprechend und wies zudem eine größere Unabhängigkeit vom verwendeten Rekonstruktionsverfahren auf. Die 18S rDNA scheint für das untersuchte taxonomische Niveau, bzw. das untersucht Taxon zu konserviert zu sein. Die Fragestellung des zweiten Projekts bestand darin, ob Umbraculum mediterraneum (Lamarck, 1812) und Umbraculum umbraculum (Lightfoot, 1786) zwei unterscheidbare Arten repräsentieren oder ob es sich um eine einzige, weltweit verbreitete Art handelt. Dazu wurden Individuen beider „Arten“ von verschiedenen Lokalitäten mit Hilfe morphologischer und molekularer Methoden untersucht. Die morphologischen Untersuchungen der verschiedenen Individuen von Umbraculum ergaben nur geringe anatomische Unterschiede und da ein gewisses Maß an Variabilität auch unter den verschiedenen Individuen eines Fundortes zu beobachten war, wurden die beobachteten Unterschiede auf die Existenz intraspezifischer Variabilität zurückgeführt. Die molekularen Untersuchungen der 18S rDNA, 16S rDNA und COI von drei Individuen unterschiedlicher Fundorte zeigten eine Variabilität von 0%, 0,4% und 3,2% und lagen damit im Rahmen der innerartlichen Variabilität bei anderen Taxa. Das dritte Teilprojekt bestand in der phylogenetischen Untersuchung der Nudibranchia-Gattung Phyllodesmium, die sich durch eine Symbiose mit Zooxanthellen, einzelligen Algen der Gattung Symbiodinium (Dinophyceae) auszeichnet. Das von Rudman (1981, 1991) anhand morphologischer Merkmale postulierte Evolutionsgeschehen sollte dabei mit Hilfe der 18S rDNA gestützt oder widerlegt werden. Aufgrund der hohen Sequenzvariabilität innerhalb der Nudibranchia erwies sich die 18S rDNA trotz der nahen Verwandtschaft der untersuchten Taxa als geeignet. Die beiden von Rudman (1991) erkannten evolutiven Linien konnten auf der Grundlage der molekularen Untersuchungen bestätigt werden, die kategorische Einteilung in Taxa mit weniger und mit höher effizienter Symbiose erwiesen sich allerdings durch Untersuchungen an der neu beschriebenen Art P. jakobsenae als nicht zutreffend.

162
Abouheif E., Zardoya R. & Meyer A. (1998): Limitations of metazoan 18S rRNA sequence data: implications for reconstructing a phylogeny of the animal kingdom and inferring the reality of the Cambrian explosion. Journal of Molecular Evolution 47: 394-405. Andrews P. & Cronin J. E. (1982): The relationships of Sivapithecus and Ramapithecus and the evolution of the orang-utan. Nature 297: 541-545. Avila C., Ballesteros M., Slattery M. & Paul V. J. (1998): Phyllodesmium guamensis (Nudibranchia, Aeolidoidea), a new species from Guam (Micronesia). Journal of Molluscan Studies 64: 147-160. Arnaud P. M., Poizat C. & Salvini-Plawen L. V. (1986): Marine-interstitial Gastropoda (including one freshwater interstitial species). In Stygofauna Mundi (L. Botosaneanu, Hrsg.), Seiten 153-176. Brill, Leiden. Baba K. (1991): Taxonomical study on some speciesof the genus Phyllodesmium from Cape Muroto-misaki, Shikoku and Okinawa Province, Southern Japan (Nudibranchia: Facelinidae). Venus 50: 109-123. Bargues M.D. & Mas-Coma S. (1997): Phylogenetic analysis of Lymnaeid snails based on 18S rDNA sequences. Molecular Biology and Evolution 14 (5): 569-577. Bargues M. D., Vigo M., Horak P., Dvorak J., Patzner R. A., Pointter J. P., Jachewicz M., Meier-Brook C. & Mas-Coma S. (2001): European Lymnaeidae (Mollusca: Gastropoda), intermediate hosts of trematodiases, based on nuclear ribosomal DNA ITS-2 sequences. Infec. Gen. Evol. 1: 85-107. Baum B. R. (1992): Combining trees as a way of combining datasets fpr phylogeneic inference, and the desirability of combining gene trees. Taxon 41: 3-10. Bayes T. (1763): An essay towards solving a problem in the doctrine of chances. Philosophical Transactions of the Royal Society of London 53: 370-418. Beeman R. D. (1968): The order Anaspidea. The Veliger, Supplement 3: 87-102. Behrens D. W. (1980): Pacific Coast Nudibranchs. Sea Challengers Natural History Books, Danville. Bergh R. (1895): Die Hedyliden, eine Familie der kladohepatischen Nudibranchien. Verhandlungen der Zoologisch-botanischen Gesellschaft in Wien 45: 4-12. Boettger C. (1954): Die Systematik der euthyneuren Schnecken. Verhandlungen der Deutschen Zoologischen Gesellschaft in Tübingen 18: 253-280.

163
Boss K. J. (1971): Critical estimate of the number of recent Mollusca. Occas. Pap. Mollusks 3: 81-135. Bouchet P. & Rocroi J.-P. (1992): Supraspecific names of molluscs: a quantitative review. Malacologia 34: 75-86. Bouvier E. L. (1887): Système nerveux, morphologie génèrale et classification des Gastéropodes Prosobranches. Annales des Sciences Naturelle. Zoologie 7(3): 1-510. Bowen C. E. & Tang H. (1996): Conchiolin-protein in aragonite shells of mollusks. Comparative Biochemistry and Physiology Part A 115: 269-275. Brito C., Lourenco P., Medeiros R., Rebello J. F., de Wolf H., Jordaens K. & Backeljau T. (2001): Radular myoglobin as a molecular marker in littorinid systematics (Caenogastropoda). Journal of Shellfish Research 20: 411-414. Bromham L., Penny D., Rambaut A. & Hendy M. D. (2000): The power of relative rate tests depends on the data. Journal of Molecular Evolution 50: 296-301. Buckley T. R., Simon C., Flook P. K. & Misof B. (2000): Secondary structure and conserved motifs of the frequently sequenced domains IV and V of the insect mitochondrial large subunit rRNA gene. Insect Molecular Biology 9(6): 565-580. Bucklin A., Frost B. W., Bradford-Grieve J., Allen L. D. & Copley N. J. (2003): Molecular systematic assessment of thirty-four calanoid copepod species of the Calanidae and Clausocalanidae using DNA sequences of mtCOI and nuclear 18S rRNA. Marine Biology 142(2): 333-343. Burch J. B. (1960): Chromosome studies of aquatic pulmonate snails. Nucleus 3: 177-208. Burch J. B. (1965): Chromosome numbers and systematics in euthyneuran snails. Proc. First Europ. Malacol. Congr. 1962: 215-241. Burch J. B. & Natarajan R. (1967): Chromosomes of some opisthobranchiate mollusks from Eniwetok Atoll, western Pacific. Pacific Science 21(2): 252-259. Burghardt I. & Wägele H. (2004): A new solar powered species of the genus Phyllodesmium Ehrenberg, 1831 (Mollusca: Nudibranchia: Aeolidoidea) from Indonesia with analysis of its photosynthetic activity and notes on biology. Zootaxa 596: 1-18.

164
Burn R. (1959): Comments on the Australian Umbraculum Mollusca. Journal of the Malacological Society of Australia 3: 28-30. Carté B. K. (1996): Biomedical potential of marine natural products. Bioscience 46(4): 271-285. Cervera J. L., Gosliner T. M., Garcia Gomez J. C. & Ortea J. A. (2000): A new species of Berthella Blainville, 1824 (Opisthobranchia: Notaspidea) from the Canary Islands (Eastern Atlantic Ocean), with a re-examination of the phylogenetic relationships of the Notaspidea. Journal of Molluscan Studies 66: 301-311. Choudhury R. C., Pandit R. K. & Sahu T. (1992): Chromosomes of a freshwater gastropod, Lymnaea luteola Lamarck (Lymnaeidae: Basommatophora). Cytologia 57: 143-147. Colgan D. J., Ponder W. F. & Eggler P. E. (2000): Gastropod evolutionary rates and phylogenetic relationships assessed using partial 28S rDNA and histone H3 sequences. Zoologica Scripta 29: 29-63. Conant G. C. & Lewis P. O. (2001): Effects of nucleotide composition bias on the success of the parsimony criterion in phylogenetic inference. Molecular Biology and Evolution 18(6): 1024-1033. Dayrat B., Tillier A., Lecointre G. & Tillier S. (2001): New clades of euthyneuran gastropods (Mollusca) from 28S rRNA sequences. Molecular Phylogenetics and Evolution 19:225-235. Dayrat B. & Tillier S. (2002): Evolutionary relationships of euthyneuran gastropods (Mollusca): a cladistic re-evaluation of morphological characters. Zool. J. Linn. Soc. 135: 403-470. Dayrat B. & Tillier S. (2003): Goals and limits of phylogenetics. The euthyneuran gastropods. In Lydeard C., Lindberg D. (Hrsg.). Molecular systematics and phylogeography of Mollusks. Smithsonia Books. Washington, London. 161-184. Degnan B. M., Degnan S. M., Giusti A. & Morse D. E. (1995): A hox/hom homeobox gene in sponges. Gene 155: 175-177. Delhaye W & Bouillon J (1972): L’évolution et l’adaptation de l’organe excréteur chez les gastéropodes pulmonés. III. Histophysiologie comparée du rein chez les soléolifères et conclusions générales pour tous les stylommatophores. Bulletin biologique 106: 295-314.

165
De Rijk P., Van de Peer Y., Chapelle S., De Wachter R. (1994) : Database on the structure of large ribosomal subunit RNA. Nucleic Acids Research 22: 3495-3501. De Rijk P., Caers A., Van de Peer Y. & De Wachter R. (1998): Secondary structure model for X. laevis LSU rRNA, (http://rrna.uia.ac.be/ ). Database on the structure of large ribosomal subunit RNA. Nucleic Acids Research 26(1):183-186. Dutra-Clarke A. V. C., Williams C., Dickstein R., Kaufer N. & Spotila J. R. (2001): Inferences on the phylogenetic relationships of Succineidae (Mollusca, Pulmonata) based on 18S rRNA gene. Malacologia 43: 223-236. Eales N. B. (1944): Aplysiids from the Indian Ocean. Proceedings of the Malacological Society London 26: 1-22. Ebel, R., Marin, A. and Proksch, P. (1999) Organspecific distribution of dietary alkaloids in the marine opisthobranch Tylodina perversa. Biochemical Systematics and Ecology 27: 769-777. Edlinger K. (1980): Zur Phylogenie der chemischen Sinnesorgane einiger Cephalaspidea (Mollusca, Opisthobranchia). Zeitschrift für Zoologische Systematik und Evolutionsforschung 18: 241-256. Edlinger K. (1988): Torsion in gastropods: a phylogenetic model. In Prosobranch Phylogeny. Ponder W. F. (Hrsg.). Malacological Review. Supplement 4: 241-250. Efron B. (1979): Bootstrap methods: Another look at the jacknife. Ann. Statist. 7: 1-26. Eliot C. (1910): A monograph of the British nudibranchiate Mollusca, Part 8 (Supplement). Ray Society, London. Fahey S. J. (2003): Phylogeny of Halgerda (Mollusca:Gastropoda) based on combined analysis of mitochondrial COI and morphology. Invertebr. Syst. 17: 617-624. Felsenstein J. (1981): Evolutionary trees from DNA sequences: a maximum likelihood approach. Journal of Molecular Evolution 17: 368-376. Felsenstein J. (1985): Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39: 783-791. Folmer O., Black M., Hoeh W., Lutz R. & Vrijenhoek R. (1994) DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Molecular Marine Biology and Biotechnology 3: 294-299.

166
Fretter V. (1943): Studies in the functional morphology and embryology of Onchidella celtica (Forbes and Hanley) and their bearing on its relationships. Journal of Marine Biology Association, U. K. 25: 685-720. Fretter V. & Graham A. (1949): The structure and mode of life of the Pyramidellidae, parasitic opisthobranchs. Journal of Marine Biology Association, U. K. 28: 493-532. Fretter V., Graham A. & Andrews E. (1986): The Prosobranch Molluscs of Britain and Denmark. Part. 9. Pyramidellacea. Journal of Molluscan Studies, Supplement 16: 557-649. Frias Martins A. M. d. (1996): Anatomy and sytematics of the western Atlantic Ellobiidae (Gastropoda: Pulmonata). Malacologia 37(2): 163-332. Geiger D. L. & Jung P. (1996): A shell of Floribella aldrichi (Dall, 1890) , a large sea hare (Mollusca: Opisthobranchia: Aplysiidae) from the neocene of the northern Dominican Republic. Journal of Conchology London 35: 437-444. Götting K.-J. (1974): Malakozoologie: Grundriss der Weichtierkunde. Stuttgart: Gustav Fischer. 320 Seiten. Ghiselin M. T. (1965): Reproductive function and the phylogeny of opisthobranch gastropods. Malacologia 3(3): 327-378. Goldman N. 1993. Simple diagnostic statistical test of models for DNA substitution. Journal of Molecular Evolution, 37: 650-661 Gosliner T. M. (1981): Origins and relationships of primitive members of the Opisthobranchia (Mollusca: Gastropoda). Biological Journal of the Linnean Society, 16: 197-225. Gosliner T.M. & Ghiselin M.T. (1984): Parallel evolution in opisthobranch gastropods and its implications for phylogenetic methodology. Systematic Zoology 33(3): 255-274. Gosliner T. M. (1994): Gastropoda: Opisthobranchia. In Microscopic anatomy of Invertebrates, Volume 5, Mollusca I (F. W. Harrison & A. J. Kohn, Hrsg.), Seiten 253-355. Wiley Liss, New York. Grande C., Templado J., Cervera J. L. & Zardoya R. (2002): The complete mitochondrial genome of the nudibranch Roboastra europaea (Mollusca: Gastropoda) supports the monophyly of opisthobranchs. Molecular Biology and Evolution 19:1672-1685.

167
Grande C., Templado J., Cervera J. L. & Zardoya R. (2004a): Molecular phylogeny of Euthyneura (Mollusca: Gastropoda). Molecular Biology and Evolution 21: 303-313. Grande C., Templado J., Cervera J. L. & Zardoya R. (2004b): Phylogenetic relationships among Opisthobranchia (Mollusca: Gastropoda) based on mitochondrial cox 1, trnV, and rrnL genes. Molecular Phylogenetics and Evolution 33: 378-388. Grechko V. V. (2002): Molecular DNA markers in phylogeny and systematics. Russian Journal of Genetics 38(8): 851-686. Übersetzt aus Genetika 38(8): 1013-1033. Guangyu L. (1981): A new species of the genus Umbraculum (Opisthobranchia) from China. Oceanologia et Limnologia Sinica 12(3): 287-289. Guiart J. (1901): Contribution a l’étude des gasteropodes opisthobranchs et en particulier de cephalaspides. Mémoires de la Société de Zoologie de France 14 : 5-219. Hall B. G. (2001): Phylogenetic trees made easy. Sinauer, Sunderland, Mass. Harasewych M. G., Adamkewicz S. L., Blake J. A., Saudek D., Spriggs T. & Bult C. J. (1997a): Phylogeny and relationships of pleurotomarid gastropods (Mollusca: Gastropoda): an assessment based on partial 18S rDNA and cytochrome oxidase I sequences. Molecular Marine Biology and Biotechnology 6:1-20. Harasewych M. G., Adamkewicz S. L., Blake J. A., Saudek D., Spriggs T. & Bult C. J. (1997b): Neogastropod phylogeny: A molecular perspective. Journal of Molluscan Studies 63: 327-351. Harasewych M. G., Adamkewicz S. L., Plassmeyer M. & Gillevet P. M. (1998): Phylogenetic relationships for the lower Caenogastropoda (Mollusca, Gastropoda, Architaeniogossa, Campaniloidea, Cerithioidea) as determined by partial 18S rDNA sequences. Zoologica Scripta 27: 361-372. Harasewych M. G. & McArthur A. G. (2000): A molecular phylogeny of the Patellogastropoda (Mollusca: Gastropoda). Marine Biology 137: 183-194. Harris D. J. & Crandall K. A. (2000): Intragenomic variation within ITS1 and ITS2 of freshwater crayfishes (Decapoda: Cambaridae): implications for phylogenetic and microsattelite studies. Molecular Biology and Evolution 17(2): 284-291.

168
Harrison P., Echols N. & Gerstein M. (2001): Digging for dead genes: An analysis of the characteristics of the pseudogene population in the C. elegans genome. Nucleic Acids Research 29: 818-830. Hasegawa M., Kishino H. & Yano T.-A. (1985): Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. Journal of Molecular Evolution 22: 160-174. Hasegawa M. & Fujiwara M. (1993): Relative efficiencies of the maximum likelihood, maximum parsimony, and neighbour joining methods for estimating protein phylogeny. Molecular Phylogenetics and Evolution 2: 1-5. Hastings W. K. (1970): Monte Carlo sampling methods using markov chains and their applications. Biometrika 57: 97-109. Haszprunar, G. (1985a): The Heterobranchia – a new concept of the phylogeny of the higher Gastropoda. Zeitschrift für Zoologische Systematik und Evolutionsforschung 23: 15-37. Haszprunar G. (1985b): On the anatomy and systematic position of the Mathildidae (Mollusca, Allogastropoda). Zoologica Scripta 14: 201-213. Haszprunar G. (1985c): Zur Anatomie und systematischen Stellung der Architectonicidae (Mollusca, Aloastropoda). Zoologica Scripta 14: 25-43. Haszprunar, G. (1988a): On the origin and evolution of major gastropod groups, with special reference to the Streptoneura. Journal of Molluscan Studies 54: 367-441. Haszprunar G. (1988b): A preliminary phylogenetic analysis of the streptoneurous gastropods. In Prosobranch Phylogeny, Ponder W. F. (Hrsg.). Malacological Review. Supplement 4: 7-16. Haszprunar G. & Huber G. (1990): On the central nervous system of Smeagolidae and Rhodopidae, two families questionably allied with the Gymnomorpha (Gastropoda: Euthyneura). Journal of Zoology 220: 185-199. Hebert P.D.N., Cywinska A., Ball S.L., & deWaard,J.R. (2003a): Biological identifications through barcodes. Proceedings of the Royal Society of London B 270: 313-321. Hebert P.D.N., Ratnasingham S. & deWaard J.R. (2003b): Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proceedings of the Royal Society of London, Supplement.

169
Hendy M. D. & Penny D. (1989): A framework for the quantitative study of evolutionary trees. Systematic Zoology 38: 297-309. Hillis D. M. & Dixon M. T. (1991): Ribosomal DNA: Molecular evolution and phylogenetic inference. The Quaterly Review of Biology 66(4): 411-446. Hoffmann H. (1932-40): Opisthobranchia. In Dr. H. G. Bronns, Klassen und Ordnungen des Tierreichs. 3. Band: Mollusca. II. Abteilung: Gastropoda. 3. Buch: Opisthobranchia. Leipzig, Akademische Verlagsgesellschaft. Howells H. H. (1936): The anatomy and histology of the gut of Cymbulia peronii (Blainville). Proceedings of the Malacological Society London 22: 62-73. Huber G. (1993): On the cerebral nervous system of marine Heterobranchia (Gastropoda). Journal of Molluscan Studies 59: 381-420. Huelsenbeck J. P. & Hillis D. M. (1993): Success of phylogenetic methods in the four-taxon case. Systematic Biology 42: 247-264. Huelsenbeck J. P. (1995a): Performance of phylogenetic methods in simulation. Systematic Biology 44: 17-48. Huelsenbeck J. P. (1995b): The robustness of two phylogenetic methods: Four-taxon simulations reveal a slight superiority of maximum likelihood over neighbor joining. Molecular Biology and Evolution 12(5): 843-849 Huelsenbeck J. P. & Ronquist F. (2001a): MrBayes. Bayesian inference of phylogeny. Bioinformatics 17: 754-755. Huelsenbeck J. P., Ronquist F., Nielsen R. & Bollback J. P. (2001b): Bayesian inference of phylogeny and its impact on evolutionary biology. Science’s Compass Review 294: 2310-2314. Huelsenbeck J. P., Larget B., Miller R. E. & Ronquist F. (2002): Potential applications and pitfalls of Bayesian inference of phylogeny. Systematic Biology 51(5): 673-688. Huff S. W., Campbell D., Gustafson D. L., Lydeard C., Altaba C. R. & Giribet G. (2004): Investigations into the phylogenetic relationships of the threatened freshwater pearl-mussels (Bivalvia, Unionoidea, Margaritiferidae) based on molecular data: implications for their taxonomy and biogeography. Journal of Molluscan Studies 70: 379-388. Hyman L. H. (1967): The Invertebrates: Vol. 6, Mollusca I. New York, McGraw-Hill, 792 Seiten.

170
Inaba A. & Hirota R. (1958): A chromosome survey in ten species of nudibranchs (Gastropoda, Mollusca). Jap. J. Zool. 12: 157-162. Inaba A. (1959) : Cytological studies in molluscs, III. A chromosome survey in the opisthobranchiate Gastropoda. Annotationes Zoologicae Japonenses 32(2): 81-88. Inaba A. (1961) : The chromosomes in some opisthobranchs. Dobutsugaku Zasshi 70: 24 Inaba A. & Saiki Y. (1967): A chromosome survey in sixteen species of nudibranchs. Venus, Jap. J. Malacol. 26(1) : 31-39. Jensen K. R. (1996): Phylogenetic systematics and classification of the Sacoglossa (Mollusca, Gastropoda, Opisthobranchia). Philosophical Transactions of the Royal Society of London B 351: 91-122. Jensen K. R. (1997a): Evolution of the Sacoglossa (Mollusca, Opisthobranchia) and the ecological associations with their food plants. Evolutionary Ecology 11: 301-335. Jensen K. R. (1997b): Cannibalism in an opisthobranch mollusc, Pleurobranchaea brockii Bergh, 1897 from Hong Kong waters. Phuket Marine Biological Center Special Publication 17(1): 41-46. Johnson K. P. (2001): Taxon sampling and the phylogenetic position of Passeriformes: Evidence from 916 avian cytochrome c sequences. Systematic Biology 50: 128-136. Jukes T. H. & Cantor C. R. (1969): Evolution of protein molecules. In: Munro H. N. (Hrsg.): Mammalian protein metabolism. Academic Press, New York: 21-132. Kimura M. (1980): A simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. Journal of Molecular Evolution 16: 111-120. Klussmann-Kolb A. (2004): Phylogeny of the Aplysiidae (Gastropoda, Opisthobranchia) with new aspects of the evolution of seahares. Zoologica Scripta 33: 439-462. Köhler A. (1893): Beiträge zur Anatomie der Gattung Siphonaria. Zoologische Jahrbücher, Abteilung Anatomie 7: 1-92.

171
Korte A. & Armbruster G.F.J. (2003): Apomorphic and plesiomorphic ITS-1 rDNA patterns in morphologically similar snails (Stylommatophora: Vallonia), with estimates on divergence time. Journal of Zoology, London 260 (3): 275-283. Kress A. (1977): Runcina ferruginea (Cephalaspidea: Opisthobranchia: Gastropoda), a new runcinid from Great Britain. J. Mar. Biol. Assoc. U. K. 57: 201-211. Kuhner M. K. & Felsenstein J. (1994): A simulation of phylogeny algorithms under equal and unequal evolutionary rates. Molecular Biology and Evolution 11: 459-468. Lalli C. M. & Gilmer R. W. (1989): Pelagic snails: the biology of holoplanktonic gastropod mollusks. Stanford University Press, Stanford, California. Lalli C. M. & Wells F. E. (1978): Reproduction in the genus Limacina (Opisthobranchia: Thecosomata). Journal of Zoology, London 186: 95-108. Lecointre G., Philippe H., Vân Lê H. L. & Le Guyader H. (1993) : Species sampling has a major impact on phylogenetic inference. Molecular Phylogenetics and Evolution 2(3): 205-224. Lewin B. (2000): Genes VII. New York: Oxford University Press. Lieb B., Altenhein B. & Markl J. (2000): The sequence of a gastropod hemocyanin (HtH1) from Haliotis tuberculata. Journal of Biological Chemistry 275: 5675-5681. Mancino G. & Sordi M. (1964): Il corredo cromosomico i alcuni Opistobranchi Sacoglossi del Mar Tirreno. Ibid., Ser. B 71: 30-39. Marcus E. (1972): On the Anaspidea (Gastropoda: Opisthobranchia) of the warm waters of the western Atlantic. Bulletin of Marine Science 22: 841-874. Marcus, E. (1985): Catalogue of the Western Atlantic warm water Opisthobranchia. 10.: The western Atlantic warm water Notaspidea (Gastropoda, Opisthobranchia), Part 3: Umbraculacea. Bolm. Zool. Univ. S. Paulo 9: 1-15. Maxam A. M. & Gilbert W. (1977): A new method of sequencing DNA. Proceedings of the National Academy of Sciences USA 74: 560-564. McArthur A. G. & Harasewych M. G. (2003): Molecular systematics of the major lineages of the Gastropoda. In: Lydeard C., Lindberg D. (eds). Molecular systematics and phylogeography of Mollusks. Smithsonian Books. Washington, London, 140-160.

172
Medina M. & Walsh P. J. (2000): Systematics of the order Anaspidea based on mitochondrial DNA sequence (12S, 16S and COI). Molecular Phylogenetics and Evolution 15: 41-58. Medina M., Collins T. M. & Walsh P. J. (2001): mtDNA ribosomal gene phylogeny of sea hares in the genus Aplysia (Gastropoda, Opisthobranchia, Anaspidea): Implications for comparative neurobiology. Systematic Biology 50: 676-688. Messing J., Crea R. & Seeburg P. H. (1981): A system for shotgun DNA sequencing. Nucleic Acids Research 9: 309-321. Metropolis N., Rosenbluth A. W., Rosenbluth M. N., Teller A. H. & Teller E. (1953): Equations of state calculations by fast computing machines. J. Chem. Phy. 21: 1087-1091. Mikkelsen P. M. (1993): Monophyly versus the Cephalaspidea (Gastropoda, Opisthobranchia) with an analysis of traditional cephalaspid characters. Bolletino Malacologico 29: 115-138. Mikkelsen P. M. (1994): The evolutionary relationships of Cephalaspidea s.l. (Gastropoda: Opisthobranchia): a phylogenetic analysis. 236 Seiten, Doktorarbeit, Florida Institute of Technology, Melbourne, Florida. Mikkelsen P. M. (1996): The evolutionary relationships of Cephalaspidea s.l. (Gastropoda: Opisthobranchia): a phylogenetic analysis. Malacologia 37(2): 375-442. Mikkelsen P. M. (1998): Cylindrobulla und Ascobulla in the western Atlantic (Gastropoda, Opisthobranchia, Sacoglossa): Systematic review, description of a new species, and phylogenetic reanalysis. Zoologica Scripta 27(1): 49-71. Mikkelsen P. M. (2002): Shelled opisthobranchs. Adv. Mar. Biol. 42: 67-136. Milne-Edwards H. (1848): Note sur la classification naturelle chez Mollusques Gasteropodes. Annales des Sciences Naturelles, Series 3 9: 102-112. Minichev Y. S. (1963) : Anatomy of Anopsia gaudichaudii (Souleyet) and systematic position of Gymnosomata (Opisthobranchia). Zoologischeskii Zhurnal 42: 1317-1328. Minichev Y. S. (1970): On the origin and system of nudibranchiate molluscs (Gastropoda, Opisthobranchia). Monit. Zool. Ital. 4: 169-182. Minichev Y. S. & Starobogatov Y. I. (1978): The subclasses of Gastropoda and their phylogenetic relations. Zoologischeskii Zhurnal 58: 293-305.

173
Moorsel C. H. v., Dijkstra E. G. & Gittenberger E. (2000): Molecular evidence for repetitive parallel evolution of shell structure in Clausiliidae (Gastropoda, Pulmonata) Molecular Phylogenetics and Evolution 17(2): 200-208. Morton J. E. (1955): The evolution of the Ellobiidae with adiscussion on the origin of the Pulmonata. Proceedings of the Zoological Society of London 125: 127-168. Morton J. E. (1972): The form and functioning of the pallial organs in the opisthobranch Akera bullata with a discussion on the nature of the gill in Notaspidea and other tectibranchs. The Veliger 14: 337-349. Muse S. V. & Weir B. S. (1992): Testing for equality of evolutionary rates. Genetics 132: 269-276. Natarajan R. (1958): Chromosomes of some estuarine basommatophorous snails (Mollusca: Pulmonata) from Porto Novo. J. Zool. Soc. India 10(2): 103-107. Natarajan R. (1959): Chromosomes of the slug Onchidium verruculatum Cuv. (Mollusca: Gastropoda: Pulmonata). J. Zool. Soc. India 11: 30-33. Natarajan R. & Burch J. B. (1966): Chromosomes of some Archaeopulmonata (Mollusca: Basommatophora). Cytologia 31(2): 109-116. Natarajan R. (1970): Cytological studies of Indian mollusks: chromosomes of some opisthobranchs from Porto Novo, South India. Malacological Review 3: 19-23. Nicholas K. B. & Nicholas H. B. Jr. (1997): GeneDoc: Analysis and Visualization of Genetic Variation, http://www.cris.com/~Ketchup/genedoc.shtml Nishikawa S. (1962): A comparative study of chromosomes in marine gastropods, with some remarks on cytotaxonomy and phylogeny. J. Shimonoseki Coll. Fish. 11(3): 149-186. Nordsieck H. (1992): Phylogeny and system of the Pulmonata (Gastropoda). Archiv für Molluskenkunde 117: 93-116. Nuttal G. H. F. (1904): Blood immunity and blood relationship. Cambridge University Press, Cambridge. Odhner N. H. (1939): Opisthobranchiate Mollusca from the western and northern coasts of Norway. Kongelige NorskeVidenskabernes selskabs skrifter 1939: 1-93. Odhner N. H. (1968): Systematique in sous-classe des opisthobranches. In Franc A. (ed.), Traitéde Zoologie. Masson, Paris, Seiten 834-893.

174
Okusu A., Schwabe E., Eernisse D. J. & Giribet G. (2003): Towards a phylogeny of chitons (Mollusca, Polyplacophora) based on combined analysis of five molecular loci. Org. Divers. Evol. 3: 281-302. Oliverio M., Cervelli M. & Mariottini P. (2002): ITS2 rRNA evolution and its congruence with the phylogeny of muricid neogastropods (Caenogastropoda, Muricoidea). Molecular Phylogenetics and Evolution 25: 63-69. Olsen G. J. & Woese C. R. (1993): Ribosomal RNA: a key to phylogeny. FASEB J. 7: 113-123. Ortiz D. M. & Gosliner T. M. (2003): A new species of Phyllodesmium Ehrenberg 1831 (Mollusca, Nudibranchia) from the tropical Indo-Pacific. Proceedings of the California Academy of Sciences 54: 161-168. Page R. D. M. & Holmes E. C. (1998): Molecular Evolution. A phylogenetic approach. Blackwell Science: Oxford, London, Edinburgh, Malden, Victoria, Paris: 346 S. Palumbi S., Martin R.A., Romano S., McMillan W.O., Stice L. & Grabowski G. (1991): The simple fool's Guide to PCR, Version 2. University of Hawaii Zoology Department, Honululu. Passamaneck Y.J., Schander C. & Halanych K.M. (2004): Investigation of molluscan phylogeny using large-subunit and small-subunit nuclear rRNA sequences. Molecular Phylogenetics and Evolution 32 (1): 25-38. Patterson C. M. (1969): Chromosomes of molluscs. Seiten 635-686, in: Proceedings of Symposium on Mollusca, Part II. Marine Biological Associaion of India. Pelseneer P. (1888): Report on the Pteropoda collected by H. M. S. Challenger during the years 1873-76. Part III, Anatomy. Voyage of H. M. S. Challenger, Zool. 23: 97 Seiten. Perrot M. (1938): Étude de cytologie compareé chez les gastéropodes pulmonés. Rev. Suisse Zool. 45(20): 487-566. Pettit G. R., Kamano Y., Herald C. L., Tuinman A. A., Boettner F. E., Kizu H., Schmidt J. M., Baczynskyj L., Tomer K. B. & Bontems R. (1987): The isolation and structure of a remarkable marine animal antineoplastic constituent: Dolastatin 10. J. Am. Chem. Soc. 109: 6883-6885.

175
Ponder W. F.& Lindberg D. R. (1997): Towards a phylogeny of gastropod molluscs: an analysis using morphological characters. Zoological Journal of the Linnean Society 119: 83-265. Posada D. & Crandall K. A. (1998): Modeltest: testing the model of DNA substitution. Bioinformatics 14(9): 817-818. Pruvot-Fol A. (1954): Mollusques Opisthobranches. In Faune de France. Paris: Paul Lechevalier. Purchon R. D. (1977): The Biology of the Mollusca, 2nd edn. Pergamon Press, New York. Raahauge P. & Kristensen T. K. (2000): A comparison of Bulinus africanus group species (Planorbidae; Gastropoda) by use of the internal transcribed spacer 1 region combined by morphological and anatomical characters. Acta Tropica 75: 85-94. Ragan M. A. (1992): Phylogenetic inference based on matrix representation of trees. Molecular Phylogenetics and Evolution 1: 53-58. Rainer M. (1967): Chromosomenuntersuchungen an Gastropoden (Stylommatophora). Malacologia 5(3): 341-373. Rankin J. J. (1979): A freshwater shell-less mollusc from the Caribbean: Structure, biotics, and contribution to a new understanding of the Acochlidioidea. Life Sciences Contributions Royal Ontario Museum 116: 1-123. Rasotto M. & Cardellini P. (1983): The chromosomes of Dendrodoris limbata and Dendrodoris grandiflora (Gastropoda, Mollusca). Caryologia 36: 175-181. Remigio E. A. & Hebert P. D. (2003): Testing the utility of partial COI sequences for phylogenetic estimates of gastropod relationships. Molecular Phylogenetics and Evolution 29(3): 641-647. Robakowski S. (2002): Die Sekundärstruktur des 18S rRNA Gens bei Opisthobranchiern im Vergleich. Schriftliche Hausarbeit im Rahmen der ersten Staatsprüfung für die Lehrämter für die Sekundarstufe II/I. Lehrstuhl für Spezielle Zoologie, Ruhr-Universität Bochum. Robinson M., Gouy M., Gautier C. & Mouchiroud D. (1998): Sensitivity of the relative-rate test to taxonomic sampling. Molecular Biology and Evolution 15: 1091-1098. Rodríguez F. J., Oliver A., Marìn A. & Medina J. R. (1990): The general stochastic model of nucleotide substitution. J. Theor. Biol. 142: 485-501.

176
Rosenberg G., Kuncio G. S., Davis G. M. & Harasewych M. G. (1994): Preliminary ribosomal RNA phylogeny of gastropod an unioidean bivalve mollusks. The Nautilus, Supplement 2: 111-121. Rosenberg G., Tillier S., Tillier A., Kuncio G., Hanlon R. T., Masselot M. & Williams C. J. (1997): Ribosomal RNA phylogeny of selected major clades in the Mollusca. Journal of Molluscan Studies 63: 301-309. Rosenberg M. S. & Kumar S. (2001): Incomplete taxon sampling is not a problem for phylogenetic inference. PNAS 98(19): 10751-10756. Rudman W. B. (1972): Studies on the primitive opisthobranch genera Bullina Férussac and Micromelo Pilsbry. Zoological Journal of the Linnean Society 51: 105-119. Rudman W. B. (1981): The anatomy and biology of alcyonarian-feeding aeolid opisthobranch molluscs and their development of a symbiosis with zooxanthellae. Zoological Journal of the Linnean Society, London 72: 219-262. Rudman W. B. (1991): Further studies on the taxonomy and biology of the octocoral-feeding genus Phyllodesmium Ehrenberg 1831 (Nudibranchia, Aeolidacea). Journal of Molluscan Studies 57: 167-203. Runnegar B. (1983): Molluscan phylogeny revisited. Memoirs of the Association of Australasian Palaeontologists 1: 121-144. Rychlik W. & Rhoads R. E. (1989): A computer program for choosing optimal oligonucleotides for filter hybridization, sequencing and in vitro amplification of DNA. Nucleic Acids Research 21: 8543-8551. Saiki, R. K., Gelfand, D. H., Stoffel, S., Scharf, S. J., Higuchi, R., Horn, G. T., Mullis, K. B. & Erlich, H. A. 1988. Primer-directed enzymatic amplification of DNA with a thermostable DNA polymerase. Science 239: 487-491. Salvini-Plawen L. v. (1973): Zur Kenntnis der Philinoglossacea und der Acochlidiacea mit Platyhedylidae fam. nov. (Gastropoda, Cephalaspidea). Zeitschrift für Zoologische Systematik und Evolutionsforschung 11: 110-133. Salvini-Plawen L. v. (1980): A reconsideration of systematics in the Mollusca (Phylogeny and higher classification). Malacologia 19: 249-278. Salvini-Plawen L. v. (1990): Origin, phylogeny and classification of the phylum Mollusca. Iberus 9: 1-33.

177
Salvini-Plawen L. v. (1991): The status of the Rhodopidae (Gastropoda: Euthyneura). Malacologia 32: 301-311. Salvini-Plawen L. v. & Haszprunar G. (1987): The Vetigastropoda and the systematics of streptoneurous Gastropoda (Mollusca). Journal of Zoology, London 211: 747-770. Salvini-Plawen L. v. & Steiner G. (1996): Synapomorphies and plesiomorphies in higher classification of Mollusca. In Origin and evolutionary radiation of the Mollusca (J. Taylor, Hrsg.), Seiten 29-51. Oxford University Press, New York. Sanger F., Miklen S. & Coulson A. R. (1977): DNA sequencing with chain-termination inhibitors. Proceedings of the National Academy of Sciences U.S.A. 74: 5463-5467. Sarich V. M. & Wilson A. C. (1973): Immunological time scale for hominoid evolution. Science 158: 1200-1203. Schander C. (1997): Taxonomy and phylogeny of the Pyramidellidae (Mollusca, Gastropoda, Heterobranchia). Doktorarbeit, Universität Göteborg. Schmekel L. (1985): Aspects of Evolution within the opisthobranchs. In: Trueman R. & Clarke, M.R. (eds.). The Mollusca. Vol. 10. Evolution. Academic Press, New York. Seiten 221-267. Schoeninger M. & von Haeseler A. (1993): A simple method to improve the reliability of tree reconstructions. Molecular Biology and Evolution 10: 471-483. Siddal M. E. & Whiting M. F. (1999): Long-branch abstractions. Cladistics 15: 9-24. Solem A. (1979): A theory of land snail biogeographic patterns through time. In Pathways in Malacology (S. van der Spoel, A. C. van Bruggen & J. Lever, Hrsg.), Junk, Den Haag: 225-249. Sommerfeldt N. & Schrödl M. (im Druck): Microanatomy of Hedylopsis wallantinei,a new interstitial acochlidian gastropod from the Red Sea, and ist significance from phylogeny. Journal of Molluscan Studies. Spengel J. W. (1881): Die Geruchsorgane und das Nervensystem der Mollusken. Zeitschrift für wissenschaftliche Zoologie 35: 333-383.

178
Spoel S. van der (1967): Euthecosomata, a group with remarkable development stages (Gastropoda, Pteropoda): 1-375 (Doktorarbeit, Universität Amsterdam; J. Noorduijn & Zoon, Gorinchem). Stasek C. R. (1972): The molluscan framework. In Chemical Zoology, Vol. VII, Mollusca, M. Florkin, B. T. Scheer (Hrsg.). Seiten 1-44. New York: Academic Press. Steel M. A., Lockhart P. J. & Penny D. (1993): Confidence in evolutionary trees from biological sequence data. Nature 364: 440-442. Stothard J. R., Hughes S. & Rollinson D. (1996): Variation within the internal transcribed spacer (ITS) of ribosomal DNA genes of intermediate snail hosts within the genus Bulinus (Gastropoda: Planorbidae). Acta Tropica 61: 19-29. Swofford D. L. (1999): PAUP*. Phylogenetic Analysis Using Parsimony (*and other methods. Sinauer Associates, Sunderland, Massachusetts. Tajima F. (1993): Simple methods for testing the molecular clock hypothesis. Genetics 135: 599–607. Tajima F. & Takezaki N. (1994): Estimation of evolutionary distance for reconstructing molecular phylogenetic trees. Molecular Biology and Evolution 11: 277-286. Takezaki N. & Nei M. (1994): Inconsistency of the maximum parsimony method when the rate of nucleotide substitution is constant. Journal of Molecular Evolution 39: 210-218. Taylor D. W. & Sohl N. F. (1962): An outline of gastropod classification. Malacologia 1: 7-32. Thiele J. (1929-1931): Handbuch der systematischen Weichtierkunde. Band 1. Jena. Thiriot-Quiévreux C. (1987): Chromosome studies in pelagic opisthobranch molluscs. Canadian Journal of Zoology 66: 1460-1473. Thollesson M. (1999b): Phylogenetic analysis of dorid nudibranchs (Gastropoda: Doridacea) using the mitochondrial 16s rRNA gene. Journal of Molluscan Studies 65: 335-353. Thollesson M. (1999a): Phylogenetic analysis of Euthyneura (Gastropoda) by means of the 16S rRNA gene: use of a "fast" gene for "higher-level" phylogenies. Proceedings of the Royal Society of London B 266:75-83.

179
Thompson J. D., Gibson T. J., Plewniak F., Jeanmougin F. & Higgins D. G. (1997): The Clustal X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research, 25(24): 4876-4882. Thompson T. E. (1970): Eastern Australian Pleurobranchomorpha (Gastropoda, Opisthobranchia). Journal of Zoology, London, 160: 173-198. Thompson T. E. (1976): Biology of opisthobranch molluscs. The Ray Society, London. Thompson T. E. & Seaward D. R. (1989): Ecology and taxonomic status of the aplysiomorph Akera bullata in the British Isles. Journal of Molluscan Studies 55: 489-496. Tillier S. & Ponder W. F. (1992): New species of Smeagol from Australia and New Zealand, with a discussion of the affinities of the genus (Gastropoda: Pulmonata). Journal of Molluscan Studies 58: 135-155. Tillier S., Masselot M., Herve P. & Tillier A. (1992): Phylogénie moléculaire des Gastropoda (Mollusca) fondée sur le séquençage partiel de l’ARN ribosomique 28S. Comptes Rendus Academie de Sciences (Paris) 134 : 79-85. Tillier S., Masselot M., Guerdox J. & Tillier A. (1994): monophyly of major gastropod taxa tested from partial 28S rRNA sequences, with emphasis on Euthyneura and hot-vent limpets Peltospiroidea. The Nautilus, Supplement 2: 122-140. Tillier S., Masselot M. and Tillier A. (1996): Phylogenetic relationships of the pulmonate gastropods from rRNA sequences, and tempo and age of the stylommatophoran radiation. Pp 267-284. In Origin and evolutionary radiation of the Mollusca. J. Taylor (Hrsg.). Oxford: Oxford Univ. Press. Tracey S, Todd J. A. & Erwin D. H. (1993): Mollusca: Gastropoda. In The fossil record 2, M. J. Benton (Hrsg.). Seiten 131-167, London: Chapman & Hall. Troschel F. H. (1856-1893): Das Gebiss der Schnecken, zur Begründung einer natürlichen Classification. Berlin. 2 Bände. Ueda-Nishimura K. & Mikata K. (2000): Two distinct 18S rRNA secondary structures in Dipodascus (Hemiascomycetes). Microbiol. 146: 1045-1051. Ueshima R. (2001): Phylogeny of opisthobranch gastropods inferred from mitochondrial gene arrangement: A case study for comparing phylogenetic performance between gene sequence and geneorder data. Seite 361. In World

180
Congress of Malacology (L. v. Salvini-Plawen, J. Woltzow, H. Sattmann & G. Steiner, Hrsg.). Vienna, Austria, Unitas Malacologica. Valdés A. & Lozouet P. (2000) : Opisthobranch molluscs from the tertiary of the Aquitaine Basin (South-Western France), with descriptions of seven new species and a new genus. Palaeontology 43: 457-479. Valdés A. (2002): Phylogenetic systematics of “Bathydoris” s.l. Bergh, 1884 (Mollusca, Nudibranchia), with the description of a new species from New Caledonian deep waters. Canadian Journal of Zoology 80: 1084-1099. Vitturi R., Rasotto M., Parrinello N. & Catalano E. (1982): Spermatocyte chromosomes in some species of the family Aplysiidae (Gastropoda, Opisthobranchia). Caryologia 35(3): 327-333. Vonnemann V., Schrödl M., Klussmann-Kolb A. & Wägele H. (2005): Reconstruction of the phylogeny of the Opisthobranchia (Mollusca, Gastropoda) by means of 18S and 28S rRNA gene sequences. Journal of Molluscan Studies Wade C. M. & Mordan P. B. (2000): Evolution within the gastropod molluscs; using the ribosomal RNA gene-cluster as an indicator of phylogenetic relationships. Journal of Molluscan Studies 66: 565-570. Wägele J.-W. (1996): First principles of phylogenetic systematics, a basis for numerical methods used for morphological and molecular characters. Vie Milieu 46: 125-138. Wägele J.-W. & Rödding F. (1998): A priori estimation of phylogenetic information conserved in aligned sequences. Molecular Phylogenetics and Evolution 9: 358-365. Wägele J.-W. (2000): Grundlagen der phylogenetischen Systematik. Verlag Dr. Friedrich Pfeil, München. Wägele H. & Willan R. C. (2000): On the phylogeny of the Nudibranchia. Zoological Journal of the Linnean Society 130: 83-181. Wägele H., Vonnemann V. & Wägele, J.-W. (2003): Towards a phylogeny of the Opisthobranchia. In Molecular systematics and phylogeography of Mollusks. : Lydeard C., Lindberg D. (Hrsg.). Smithsonian Books. Washington, London, Seiten 185-228. Wägele H. (2004): Potential key characters in Opisthobranchia (Gastropoda, Mollusca) enhancing adaptive radiation. Organisms, Diversity and Evolution 4: 175-188.

181
Wägele H., Vonnemann V. & Rudman W. B. (im Druck): Umbraculum umbraculum (Lightfoot, 1786) (Gastropoda, Opisthobranchia, Tylodinoidea) and the synonymy of U. mediterraneum (Lamarck, 1812). In Opisthobranchia (G. Brodie & S. Fahey, Hrsg.), Records of Western Australian Museum, Supplement. Wägele H., Klussmann-Kolb A., Vonnemann V. & Medina M. (in Vorbereitung): Towards an understanding of opisthobranch evolution. In Phylogeny of Molluscs (W. F. Ponder & D. R. Lindberg, Hrsg.). Wägele H. & Klussmann-Kolb (im Druck): Opisthobranchia (Mollusca, Gastropoda) – more than just slimy slugs. Shell reduction and its implications on defense and foraging. Frontiers in Zoology. Wenz W. (1938-1944): Gastropoda, Teil 1: Allgemeiner Teil und Prosobranchia. In Handbuch der Paläozoologie (O. H. Schindewolf, Hrsg.). Gebrüder Borntraeger, Berlin. Wesson D. M. and Frank H. Collins (1992): Sequence and secondary structure of 5.8S rRNA in the tick, Ixodes scapularis. Nucleic Acids Research 20(5): 1139 Willan R. C. (1984): A review of diets in the Notaspidea (Mollusca: Opisthobranchia). Journal of the Malacological Society of Australia 6(3-4): 125-142. Willan R. C. (1987): Phylogenetic systematics of the Notaspidea (Opisthobranchia) with reappraisal of families and genera. American Malacological Bulletin 5: 215-241. Willan R. C. (1998): Order Notaspidea. Pp 977-979. In Beesley P. L., Ross G. J. B. & Wells A. (Hrsg.) Mollusca: The southern Synthesis. Fauna of Australia Vol 5. CSIRO Publishing: Melbourne, Part B, Seiten 565-1234. Williams S. T. & Reid D. G. (2004): Speciation and diversity on tropical rocky shores: a global phylogeny of snails of the genus Echinolittorina. Evolution 58 (10): 2227-2251. Winnepenninckx B., Backeljau T. & De Wachter R. (1994): Small ribosomal subunit RNA and the phylogeny of the Mollusca. The Nautilus, Supplement 2: 98-110. Winnepenninckx B., Backeljau T. & De Wachter R. (1996): Investigation of molluscan phylogeny on the basis of 18S rRNA sequences. Molecular Biology and Evolution 13: 1306-1317.

182
Winnepenninckx B., Steiner G., Backeljau T. & De Wachter R. (1998): Details of gastropod phylogeny inferred from 18S rRNA sequences. Molecular Phylogenetics and Evolution 9: 55-63. Wollscheid E. & Wägele H. (1999): Initial results on the molecular phylogeny of the Nudibranchia (Gastropoda, Opisthobranchia) based on 18S rDNA data. Molecular Phylogenetics and Evolution 13: 215-226. Wollscheid-Lengeling E., Boore J., Brown W. & Wägele H. (2001): The phylogeny of Nudibranchia (Opisthobranchia, Gastropoda, Mollusca) reconstructed by three molecular markers. Organisms, Diversity and Evolution 1: 241-256. Wuyts J., de Rijk P., van de Peer Y., Pison G., Rousseeuw P. & de Wachter R. (2000): Comparative analysis of more than 3000 sequences reveals the existence of two pseudoknots in area V4 of eukaryotic small subunit ribosomal RNA. Nucleic Acids Research 28: 4698-4708. Yang Z., Goldman N. & Friday A. (1994): Comparison of models for nukleotide substitution used in maximum likelihood phylogenetic estimation. Molecular Biology and Evolution 11: 316-324. Yang Z., Goldman N. & Friday A. (1995): Maximum likelihood trees from DNA sequences: a peculiar statistical estimation problem. Systematic Biology 44: 384-399. Yang Z. (1996): Phylogenetic analyses using parsimony and likelihood methods. Journal of Molecular Evolution 42: 294-307. Yang Z. (1998): On the best evolutionary rate for phylogenetic analyses. Systematic Biology 47(1): 125-133. Yang Z., Goldman N. & Friday A. (1994): Comparison of models for nucleotide substitution used in maximum likelihood phylogenetic estimation. Molecular Biology and Evolution 11: 316-324. Yoder A. D. & Irwin J. A. (1999): Phylogeny of the Lemuridae: effects of taxon and character sampling on resolution of species relationships within Eulemur. Cladistics 15: 351-361. Yoon S. H. & Kim W. (2000): Phylogeny of some gastropod mollusks derived from 18S rRNA sequences with emphasis on the Euthyneura. The Nautilus 114: 84-92. Zarnick B. (1911): Über den Chromosomencyclus bei Pteropoden. Ver. Dtsch. Zool. Ges. 20: 205-215.

183
Zhang Z., Harrison P., & Gerstein M. (2002): Identification and analysis of over 2000 ribosomal protein pseudogenes in the human genome. Genome Research 12: 1466-1482. Zhang Z. & Gerstein M. (2003a): Identification and characterization of over 100 mitochondrial ribosomal protein pseudogenes in the human genome. Genomics 81: 468-480. Zhang Z. & Gerstein M. (2003b): Patterns of nucleotide substitution, insertion and deletion in the human genome inferred from pseudogenes. Nucleic Acids Research 31: 5338-5348. Zharkikh A. & Li W. H. (1993): Inconsistency of the maximum-parsimony method: the case of five taxa with a molecular clock. Systematic Biology 42: 113-125.

184
Danksagung Für das Gelingen dieser Arbeit möchte ich verschiedenen Personen und aus unterschiedlichen Gründen danken:
- H. Wägele für all ihre Unterstützung und Begeisterung für meine Arbeit. - J.-W. Wägele für die Aufnahme an seinem Lehrstuhl. - Th. Stützel für die Begutachtung dieser Arbeit. - Meinen unzähligen lieben Kollegen des Lehrstuhls für Spezielle Zoologie für
unermüdliche Hilfe, Unterhaltung, Ablenkung, konstruktive Kritik und Zerstreuung.
- M. & R. Schrödl-Carvalho, E. Schwabe, D. Eheberg und I. Burghardt für wunderbare Erinnerungen an Reisen in ferne Länder.
- S. Janett und I. Manstedt für Hilfe in bürokratischen Angelegenheiten und am Computer.
- B. Hackethal und M. Rudschewski für den reibungslosen Ablauf der Laborarbeit.
- C. Brefeld für die Bearbeitung der Fernleihbestellungen. - J. Bohn (München), B. Bolland (Okinawa), J. Brodie (Townsville), P. Durán
(Santiago de Chile), D. Eheberg (München), A. Fahrner (München), G. Haszprunar (München), A. Klussmann-Kolb (Frankfurt), N. Michiels (Münster), M. Raupach (Bochum), E. Schwabe (München), M. Schrödl und G. Steiner (Wien) für die Bereitstellung von Tiermaterial.
- Der Deutschen Forschungsgemeinschaft für die finanzielle Unterstützung.

185
Lebenslauf Verena Vonnemann Geboren am 16. Februar 1975 in Essen. Schulische Ausbildung: 1981-1984 Grundschule Essen-Fischlaken 1985-1994 Gymnasium Essen-Werden, Abitur 1994 Wissenschaftliche Ausbildung: 1994-2000 Studium der Biologie an der RUB
2000-2001 Diplomarbeit am Lehrstuhl für Spezielle Zoologie 2001-2004 Wissenschaftliche Mitarbeiterin am Lehrstuhl für Spezielle Zoologie
Publikationen: Vonnemann V., Schrödl M., Klussmann-Kolb A. & Wägele H. (im Druck): Reconstruction of the phylogeny of the Opisthobranchia (Mollusca, Gastropoda) by means of 18S and 28S rRNA gene sequences. Journal of Molluscan Studies Wägele H., Vonnemann V. & Rudman W. B. (im Druck): Umbraculum umbraculum (Lightfoot, 1786) (Gastropoda, Opisthobranchia, Tylodinoidea) and the synonymy of U. mediterraneum (Lamarck, 1812). In Opisthobranchia (G. Brodie & S. Fahey, Hrsg.), Records of Western Australian Museum, Supplement. Wägele H., klussmann-Kolb A., Vonnemann V. & Medina M. (in Vorbereitung): Towards an understanding of opisthobranch evolution. In Phylogeny of Molluscs (W. F. Ponder & D. R. Lindberg, Hrsg.).

186
Anhang I Abkürzungen und Symbole 8 unendlich > größer als/ schneller als & und % Prozent °C Grad Celsius A Adenin Abb. Abbildung Bp Basenpaar(e) BPH Bromphenolblau (3´,3´´,5´,5´´-Tetrabromophenolsulfonphthalein) bzw. beziehungsweise C Cytosin cm Zentimeter COI Untereinheit 1 der mitochondrialen Cytochromoxidase dest. destilliert ddNTP 2´, 3´-Didesoxynukleotidtriphosphat dNTP 2´-Desoxynukleotidtriphosphat dGTP Desoxyguanosin-5´-triphosphat DNA Desoxyribonukleinsäure („desoxyribonucleid acid“) dTTP Desoxythymidin-5´-triphosphat E. coli Escherichia coli EDTA Ethylendiamintetraessigsäure EthBr Ethidiumbromid ETS external transcribed spacer et al. und weitere (et alii) F „forward” g Gramm G Guanin inkl. inklusive ITS internal transcribed spacer IPTG Isopropyl-b-thiogalactopyranosid kb Kilobasen l Liter LB Luria-Bertani M Mol min Minute(n) ml Milliliter mm Millimeter mM Millimolar mmol Millimol ML maximum likelihood

187
MP Maximum Parsimonie N Nukleotid ng Nanogramm NSW New South Wales pH pars Hydrogenium PCR Polymerase-Kettenreaktion (polymerase chain reaction) pmol Picomol QLD Queensland R „reverse“ RNA Ribonukleinsäure (ribonucleic acid) sec Sekunde(n) S Svedberg-Einheit T Thymin Tab. Tabelle Taq Thermophilus aquaticus TAE Tris/Natrium-Acetat-Puffer mit EDTA TBE Tris/Borat-Puffer mit EDTA Tm Schmelztemperatur Ts Transitionen Tv Transversionen u. a. unter anderem vgl. vergleiche X-Gal 5-Brom-4-chlor-3-indolyl-b-D-galacto-pyranosid z.B. zum Beispiel II Chemikalien, Lösungen, Medien, Geräte und Kits Chemikalien Die verwendeten Chemikalien wurden von den Fimen Serva (Heidelberg), Biozym (Hamburg), Promega (Mannheim), Merck (Darmstadt), Roth (Karlsruhe), J.T. Baker (Griesheim), National Diagnostics (Hess. Oldendorf), AppliChem (Darmstadt), Biomol (Hamburg) und DIFCO (Hamburg) bezogen. Rezepte für Lösungen und Medien Ampicillin-Stammlösung (10%) 1,0 g in 10 ml H2O (reinst) Agarose-Gel (1%) 2,0 g Agarose in 200 ml TAE-Puffer (1x) APS-Lösung (10%) 0,5 g Ammoniumpersulfat in 5 ml H2O (reinst)

188
Bromphenolblau-Ladepuffer 100 mM EDTA
43% (v/v) Glycerol 0,05% (w/v) BPB
dNTP-Mix 2 mM dATP 2 mM dCTP 2 mM dGTP 2 mM dTTP
EthBr-Stammlösung (1%) 100 mg in 10 ml H2O (reinst) LB-Medium 10 g/l Tryptone Pepton
5 g/l Hefe-Extrakt 5 g/l NaCl in 1 l H2O (reinst) gelöst und auto-klaviert
LB-Agar-Ampicilin-Platten 15 g/l Agar in LB-Medium mit 100 µg/ml Ampicillin, 100 µg/ml IPTG und 100 µg/ml X-Gal steril gegossen
Polyacrylamid-Gel 24 ml SequagelTM XR 6%
6 ml SequagelTM Complete Buffer Reagent 250 µl Ammoniumpersulfat (10%)
TAE-Puffer (50x, pH 8,3) 2 M Tris-Base 0,5 M Na-Acetat 50 mM EDTA in H2O (reinst)
TBE-Puffer (10x, pH 8,3) 1340 mM Tris-Base 45 mM Borsäure 25 mM EDTA in H2O (reinst)
Kits FastPlasmidTM Mini Kit Eppendorf AG, Hamburg NucleoSpin®-Extract Kit Macherey-Nagel, Düren NucleoSpin®-Tissue Kit Macherey-Nagel, Düren Original TA Cloning® Kit Version N Invitrogen BV De Schelp, NL QIAquick PCR-Purification Kit Qiagen GmbH, Hilden QIAamp Tissue Kit Qiagen GmbH, Hilden

189
Thermo-Sequenase fluorescent Labelled primer cycle sequencing kit Amersham Pharmacia Biotech, Freiburg Geräte Autoklaviergerät SANO Claw, Wolf Brutschrank GFL 3032, Gesellschaft für Labortechnik mbH DNA-Sequenziergerät DNA-Sequenzierer 4000 und 4200, LI-COR Inc. Dokumentationssysteme Gel Print 1000i, BioPhotonics Corporation
Sony Black and White Monitor SSM-930 CE, Sony Feinwaage FA-1500-2, Faust Pipetten ABIMED, Langenfeld 8-Kanal Pipette Gel Loading Syringe, Hamilton Bonaduz AG Spannungsregler Power Pac 200, Bio-Rad Laboratories GmbH Thermische Zykler Primus96plus Thermocycler, MWG-Biotech AG
Progene Thermocycler, Techne Ltd. PTC-200 Peltier Thermal Cycler, MJ Reserach
UV-Leuchttisch TFX-20.M UV-Leuchttisch, Vilber Lourmat Vortexer Vortex Genie 2, Scientific Industries Wasserbad Wasserbad Typ 1002,
Gesellschaft für Labortechnik mbH Zentrifuge Biofuge pico, Heraeus Instruments GmbH III Relative rate test Opisthobranchia-Datensatz (18S rDNA) Ergebnisse der Analyse der relativen Substitutionsraten auf der Grundlage des 18S rDNA-Datensatzes mit Odostomia sp. als Außengruppe (1). Eine signifikant unterschiedliche Substitutionsgeschwindigkeit ist gegeben, wenn der „Z-score“ (k12-k13/SD) höher als 1,96 ist. Positive „Z-score“-Werte bedeuten, dass Taxon (2) eine größere Evolutionsgeschwindigkeit aufweist; negative Werte stehen für eine erhöhte Substitutionsrate des Taxons (3). (2) (3) K12 - K13 SD Z-score Bathydoris vs Chromod -0.003432 0.005991 -0.572841 Bathydoris vs Dendrod 0.001803 0.006025 0.299225 Bathydoris vs Hypselo -0.000779 0.005937 -0.131226 Bathydoris vs Dendron -0.132502 0.012544 -10.563299 Bathydoris vs Cuthona -0.120902 0.012320 -9.813439 Bathydoris vs Flabell -0.117843 0.012100 -9.739433 Bathydoris vs Pleurob 0.015766 0.008037 1.961572 Bathydoris vs Berthel 0.017691 0.008039 2.200727 Bathydoris vs Bathybe 0.012551 0.008255 1.520467 Bathydoris vs Euselen 0.015107 0.008154 1.852728 Bathydoris vs Tomptho 0.018343 0.007993 2.294924 Bathydoris vs Berthel -0.087113 0.011397 -7.643216

190
Bathydoris vs Berthel -0.015883 0.009318 -1.704681 Bathydoris vs Pupa_so 0.049449 0.008108 6.098836 Bathydoris vs Hydatin 0.053673 0.008198 6.546891 Bathydoris vs Bullina 0.056730 0.008111 6.994278 Bathydoris vs Acteon_ 0.064378 0.008046 8.001700 Bathydoris vs Haminoe 0.063778 0.008187 7.790368 Bathydoris vs Cylichn 0.070317 0.007911 8.888279 Bathydoris vs Scaphan 0.068483 0.008014 8.545329 Bathydoris vs cf._Ret 0.065552 0.008020 8.173604 Bathydoris vs Bulla_c 0.070882 0.007843 9.038073 Bathydoris vs Smaragd 0.068113 0.008057 8.453393 Bathydoris vs Phanero 0.066776 0.008037 8.308368 Bathydoris vs Bullact 0.069735 0.007986 8.732634 Bathydoris vs Atys_se 0.067396 0.008066 8.355445 Bathydoris vs Odontog 0.066769 0.007944 8.404521 Bathydoris vs Navanax 0.066151 0.007972 8.298037 Bathydoris vs Chelido 0.066791 0.007951 8.400507 Bathydoris vs Philino 0.066184 0.007979 8.294710 Bathydoris vs Philino 0.063187 0.007921 7.977502 Bathydoris vs Sagamin 0.064370 0.007750 8.306122 Bathydoris vs Toledon 0.073318 0.007965 9.204577 Bathydoris vs Diaphan 0.067677 0.008084 8.371897 Bathydoris vs Newnesi 0.070354 0.008071 8.716743 Bathydoris vs Colpoda 0.055946 0.007986 7.005278 Bathydoris vs Runcina 0.061458 0.008106 7.581490 Bathydoris vs Unela_g 0.075120 0.007896 9.513319 Bathydoris vs Pontohe 0.066833 0.008007 8.346427 Bathydoris vs Hedylop 0.077415 0.007814 9.907285 Bathydoris vs Parahed 0.075074 0.007891 9.513404 Bathydoris vs Umbracu 0.069792 0.008059 8.660228 Bathydoris vs Tylodin 0.069115 0.008001 8.638153 Bathydoris vs Akera_b 0.072005 0.007883 9.134321 Bathydoris vs Aplysia 0.073817 0.007762 9.509908 Bathydoris vs Dolabel 0.073863 0.007767 9.509824 Bathydoris vs Siphono 0.073289 0.007839 9.349546 Bathydoris vs Bursate 0.075108 0.007720 9.728760 Bathydoris vs Petalif 0.075595 0.007736 9.772071 Bathydoris vs Dolabri 0.075006 0.007762 9.663610 Bathydoris vs Styloch 0.073832 0.007764 9.509518 Bathydoris vs Clio_py 0.055967 0.008353 6.700431 Bathydoris vs Diacria 0.051720 0.008421 6.141855 Bathydoris vs Creseis 0.032617 0.008717 3.741626 Bathydoris vs Spongio 0.065546 0.007942 8.252830 Bathydoris vs Calliop 0.055414 0.008462 6.548946 Bathydoris vs Limapon 0.061476 0.008099 7.590552 Bathydoris vs Thuridi 0.068719 0.008061 8.525245 Bathydoris vs Elysia_ 0.067941 0.008023 8.468063 Bathydoris vs Elysiel 0.067886 0.008094 8.387067 Bathydoris vs Boselli 0.067904 0.008114 8.368289 Bathydoris vs Cyerce_ 0.072074 0.008039 8.965030 Bathydoris vs Placobr 0.062002 0.008265 7.501345 Bathydoris vs Placida 0.069762 0.008097 8.616003 Bathydoris vs Polybra 0.067930 0.007925 8.571842 Chromodori vs Dendrod 0.005851 0.003679 1.590369 Chromodori vs Hypselo 0.001346 0.002911 0.462418 Chromodori vs Dendron -0.126511 0.011995 -10.547023 Chromodori vs Cuthona -0.117957 0.011901 -9.911870 Chromodori vs Flabell -0.113371 0.011583 -9.787781 Chromodori vs Pleurob 0.017364 0.007762 2.237107 Chromodori vs Berthel 0.018631 0.007643 2.437598 Chromodori vs Bathybe 0.014130 0.007823 1.806244 Chromodori vs Euselen 0.017370 0.007754 2.240179 Chromodori vs Tomptho 0.019289 0.007648 2.521917 Chromodori vs Berthel -0.082685 0.011111 -7.441987 Chromodori vs Berthel -0.015793 0.009031 -1.748699 Chromodori vs Pupa_so 0.050503 0.007932 6.367166 Chromodori vs Hydatin 0.052912 0.008058 6.566160 Chromodori vs Bullina 0.056600 0.007894 7.170308 Chromodori vs Acteon_ 0.065564 0.008088 8.106060 Chromodori vs Haminoe 0.063760 0.008182 7.792781 Chromodori vs Cylichn 0.070345 0.008000 8.793404 Chromodori vs Scaphan 0.068503 0.007953 8.613527 Chromodori vs cf._Ret 0.064952 0.008184 7.936787 Chromodori vs Bulla_c 0.070345 0.007960 8.837479 Chromodori vs Smaragd 0.068127 0.008172 8.336251 Chromodori vs Phanero 0.066780 0.008149 8.195334 Chromodori vs Bullact 0.069759 0.008000 8.720284 Chromodori vs Atys_se 0.067404 0.008131 8.289630

191
Chromodori vs Odontog 0.067374 0.008035 8.384937 Chromodori vs Navanax 0.066752 0.008088 8.253027 Chromodori vs Chelido 0.067998 0.008085 8.410232 Chromodori vs Philino 0.066786 0.008090 8.255438 Chromodori vs Philino 0.063167 0.007988 7.907857 Chromodori vs Sagamin 0.063760 0.008063 7.907469 Chromodori vs Toledon 0.073368 0.007929 9.252669 Chromodori vs Diaphan 0.067687 0.008147 8.308261 Chromodori vs Newnesi 0.070339 0.007980 8.814849 Chromodori vs Colpoda 0.057177 0.008135 7.028335 Chromodori vs Runcina 0.062055 0.007974 7.781702 Chromodori vs Unela_g 0.075134 0.007726 9.725325 Chromodori vs Pontohe 0.066165 0.007888 8.388504 Chromodori vs Hedylop 0.077444 0.007667 10.101039 Chromodori vs Parahed 0.075087 0.007721 9.725430 Chromodori vs Umbracu 0.071089 0.007980 8.908603 Chromodori vs Tylodin 0.070405 0.007946 8.860718 Chromodori vs Akera_b 0.072641 0.007740 9.385581 Chromodori vs Aplysia 0.073870 0.007768 9.509792 Chromodori vs Dolabel 0.073868 0.007644 9.663742 Chromodori vs Siphono 0.073292 0.007692 9.528048 Chromodori vs Bursate 0.074445 0.007766 9.586223 Chromodori vs Petalif 0.075608 0.007712 9.803673 Chromodori vs Dolabri 0.075015 0.007738 9.693811 Chromodori vs Styloch 0.073833 0.007741 9.538230 Chromodori vs Clio_py 0.056513 0.008198 6.893351 Chromodori vs Diacria 0.052901 0.008466 6.248958 Chromodori vs Creseis 0.035509 0.008652 4.104276 Chromodori vs Spongio 0.065573 0.007902 8.298493 Chromodori vs Calliop 0.054118 0.008409 6.435929 Chromodori vs Limapon 0.062098 0.008046 7.717574 Chromodori vs Thuridi 0.069187 0.007836 8.829117 Chromodori vs Elysia_ 0.067980 0.007739 8.783753 Chromodori vs Elysiel 0.068524 0.007811 8.773085 Chromodori vs Boselli 0.068540 0.007809 8.776659 Chromodori vs Cyerce_ 0.072708 0.007753 9.377726 Chromodori vs Placobr 0.062569 0.007883 7.936999 Chromodori vs Placida 0.069786 0.007790 8.958163 Chromodori vs Polybra 0.068540 0.007732 8.864332 Dendrodori vs Hypselo -0.003251 0.003720 -0.874054 Dendrodori vs Dendron -0.130710 0.012144 -10.763613 Dendrodori vs Cuthona -0.121516 0.011893 -10.217427 Dendrodori vs Flabell -0.117100 0.011581 -10.110988 Dendrodori vs Pleurob 0.013367 0.007483 1.786272 Dendrodori vs Berthel 0.014631 0.007392 1.979302 Dendrodori vs Bathybe 0.010145 0.007568 1.340551 Dendrodori vs Euselen 0.013368 0.007498 1.782722 Dendrodori vs Tomptho 0.015284 0.007392 2.067518 Dendrodori vs Berthel -0.086976 0.011090 -7.842863 Dendrodori vs Berthel -0.017732 0.008819 -2.010617 Dendrodori vs Pupa_so 0.045711 0.007707 5.930728 Dendrodori vs Hydatin 0.048079 0.007808 6.158001 Dendrodori vs Bullina 0.052388 0.007798 6.717916 Dendrodori vs Acteon_ 0.060746 0.007912 7.677537 Dendrodori vs Haminoe 0.060143 0.008061 7.460976 Dendrodori vs Cylichn 0.066689 0.007803 8.546277 Dendrodori vs Scaphan 0.064828 0.007805 8.306333 Dendrodori vs cf._Ret 0.061328 0.007988 7.677242 Dendrodori vs Bulla_c 0.066690 0.007814 8.534764 Dendrodori vs Smaragd 0.064474 0.007976 8.083248 Dendrodori vs Phanero 0.063145 0.007956 7.936636 Dendrodori vs Bullact 0.066107 0.007803 8.471574 Dendrodori vs Atys_se 0.063763 0.007935 8.035312 Dendrodori vs Odontog 0.063140 0.007913 7.979230 Dendrodori vs Navanax 0.062519 0.007865 7.948481 Dendrodori vs Chelido 0.063755 0.007843 8.129235 Dendrodori vs Philino 0.062549 0.007797 8.022640 Dendrodori vs Philino 0.059555 0.007791 7.644113 Dendrodori vs Sagamin 0.059542 0.007821 7.613463 Dendrodori vs Toledon 0.069692 0.007758 8.983402 Dendrodori vs Diaphan 0.064028 0.007975 8.028773 Dendrodori vs Newnesi 0.066725 0.007762 8.595803 Dendrodori vs Colpoda 0.052966 0.007887 6.715551 Dendrodori vs Runcina 0.059083 0.007682 7.691568 Dendrodori vs Unela_g 0.072085 0.007456 9.668118 Dendrodori vs Pontohe 0.063837 0.007598 8.402278 Dendrodori vs Hedylop 0.074384 0.007397 10.056330 Dendrodori vs Parahed 0.072040 0.007451 9.668219

192
Dendrodori vs Umbracu 0.067422 0.007709 8.745848 Dendrodori vs Tylodin 0.066747 0.007700 8.667822 Dendrodori vs Akera_b 0.068976 0.007469 9.235132 Dendrodori vs Aplysia 0.070196 0.007497 9.362707 Dendrodori vs Dolabel 0.070830 0.007399 9.572391 Dendrodori vs Siphono 0.069620 0.007470 9.319735 Dendrodori vs Bursate 0.070766 0.007469 9.474260 Dendrodori vs Petalif 0.071924 0.007415 9.700181 Dendrodori vs Dolabri 0.071335 0.007441 9.586146 Dendrodori vs Styloch 0.070160 0.007444 9.425413 Dendrodori vs Clio_py 0.052335 0.007934 6.596162 Dendrodori vs Diacria 0.049348 0.008248 5.983438 Dendrodori vs Creseis 0.032077 0.008419 3.809929 Dendrodori vs Spongio 0.061948 0.007630 8.119352 Dendrodori vs Calliop 0.050531 0.008210 6.155043 Dendrodori vs Limapon 0.058456 0.007770 7.522945 Dendrodori vs Thuridi 0.064398 0.007626 8.444429 Dendrodori vs Elysia_ 0.064337 0.007592 8.473894 Dendrodori vs Elysiel 0.064852 0.007509 8.636306 Dendrodori vs Boselli 0.064896 0.007539 8.607686 Dendrodori vs Cyerce_ 0.069039 0.007535 9.162685 Dendrodori vs Placobr 0.058997 0.007671 7.691251 Dendrodori vs Placida 0.065467 0.007611 8.601405 Dendrodori vs Polybra 0.064896 0.007434 8.729405 Hypselodor vs Dendron -0.127824 0.012064 -10.595347 Hypselodor vs Cuthona -0.119994 0.011938 -10.051302 Hypselodor vs Flabell -0.116993 0.011714 -9.987553 Hypselodor vs Pleurob 0.014741 0.007686 1.917883 Hypselodor vs Berthel 0.016009 0.007623 2.099932 Hypselodor vs Bathybe 0.012134 0.007765 1.562566 Hypselodor vs Euselen 0.015377 0.007702 1.996596 Hypselodor vs Tomptho 0.016665 0.007596 2.193973 Hypselodor vs Berthel -0.085370 0.011210 -7.615800 Hypselodor vs Berthel -0.017115 0.009070 -1.886916 Hypselodor vs Pupa_so 0.048451 0.007803 6.209126 Hypselodor vs Hydatin 0.050851 0.007955 6.392686 Hypselodor vs Bullina 0.054536 0.007789 7.001843 Hypselodor vs Acteon_ 0.062894 0.008105 7.759738 Hypselodor vs Haminoe 0.062290 0.008129 7.662301 Hypselodor vs Cylichn 0.068857 0.007897 8.719423 Hypselodor vs Scaphan 0.067008 0.007900 8.482346 Hypselodor vs cf._Ret 0.063479 0.008082 7.854810 Hypselodor vs Bulla_c 0.068857 0.007857 8.764111 Hypselodor vs Smaragd 0.066641 0.008070 8.257956 Hypselodor vs Phanero 0.065301 0.008046 8.115406 Hypselodor vs Bullact 0.068272 0.007947 8.590905 Hypselodor vs Atys_se 0.065881 0.008024 8.210827 Hypselodor vs Odontog 0.065295 0.008006 8.155802 Hypselodor vs Navanax 0.064674 0.008059 8.025077 Hypselodor vs Chelido 0.065916 0.008009 8.230437 Hypselodor vs Philino 0.064706 0.008064 8.024495 Hypselodor vs Philino 0.061699 0.007885 7.825009 Hypselodor vs Sagamin 0.061687 0.007937 7.772443 Hypselodor vs Toledon 0.071871 0.007826 9.183349 Hypselodor vs Diaphan 0.065527 0.008037 8.153400 Hypselodor vs Newnesi 0.068894 0.007932 8.685681 Hypselodor vs Colpoda 0.055089 0.008081 6.817326 Hypselodor vs Runcina 0.060587 0.007998 7.575420 Hypselodor vs Unela_g 0.073633 0.007673 9.596358 Hypselodor vs Pontohe 0.065359 0.007868 8.307130 Hypselodor vs Hedylop 0.075937 0.007562 10.041322 Hypselodor vs Parahed 0.073587 0.007668 9.596460 Hypselodor vs Umbracu 0.069594 0.007950 8.754372 Hypselodor vs Tylodin 0.068914 0.007891 8.733764 Hypselodor vs Akera_b 0.071148 0.007764 9.164051 Hypselodor vs Aplysia 0.072373 0.007664 9.443384 Hypselodor vs Dolabel 0.072371 0.007643 9.468864 Hypselodor vs Siphono 0.071795 0.007588 9.461915 Hypselodor vs Bursate 0.072946 0.007662 9.520582 Hypselodor vs Petalif 0.074106 0.007608 9.740565 Hypselodor vs Dolabri 0.073516 0.007634 9.629488 Hypselodor vs Styloch 0.072337 0.007688 9.408822 Hypselodor vs Clio_py 0.053818 0.008124 6.624474 Hypselodor vs Diacria 0.051459 0.008319 6.186086 Hypselodor vs Creseis 0.033530 0.008566 3.914398 Hypselodor vs Spongio 0.064098 0.007850 8.165703 Hypselodor vs Calliop 0.052670 0.008400 6.269995 Hypselodor vs Limapon 0.060589 0.007987 7.586242

193
Hypselodor vs Thuridi 0.067870 0.007801 8.700018 Hypselodor vs Elysia_ 0.066497 0.007733 8.599217 Hypselodor vs Elysiel 0.067041 0.007759 8.640874 Hypselodor vs Boselli 0.067056 0.007706 8.702000 Hypselodor vs Cyerce_ 0.071212 0.007650 9.309286 Hypselodor vs Placobr 0.061139 0.007785 7.853400 Hypselodor vs Placida 0.067702 0.007838 8.638013 Hypselodor vs Polybra 0.067056 0.007602 8.820769 Dendronotu vs Cuthona 0.011793 0.007496 1.573230 Dendronotu vs Flabell 0.017235 0.007022 2.454342 Dendronotu vs Pleurob 0.147791 0.012760 11.582625 Dendronotu vs Berthel 0.148348 0.012645 11.731817 Dendronotu vs Bathybe 0.144179 0.012889 11.186329 Dendronotu vs Euselen 0.148047 0.012742 11.619158 Dendronotu vs Tomptho 0.149653 0.012802 11.689375 Dendronotu vs Berthel 0.046177 0.013769 3.353838 Dendronotu vs Berthel 0.110906 0.013055 8.495242 Dendronotu vs Pupa_so 0.176229 0.012907 13.654280 Dendronotu vs Hydatin 0.177956 0.013004 13.684984 Dendronotu vs Bullina 0.181826 0.012888 14.107914 Dendronotu vs Acteon_ 0.192186 0.013255 14.499127 Dendronotu vs Haminoe 0.194040 0.013234 14.661836 Dendronotu vs Cylichn 0.200105 0.013118 15.254325 Dendronotu vs Scaphan 0.199455 0.013201 15.109426 Dendronotu vs cf._Ret 0.193420 0.013272 14.573430 Dendronotu vs Bulla_c 0.200093 0.013061 15.319885 Dendronotu vs Smaragd 0.198006 0.013101 15.113933 Dendronotu vs Phanero 0.196487 0.013187 14.899979 Dendronotu vs Bullact 0.198903 0.013134 15.144430 Dendronotu vs Atys_se 0.197218 0.013168 14.976583 Dendronotu vs Odontog 0.195866 0.013254 14.777852 Dendronotu vs Navanax 0.194025 0.013259 14.633902 Dendronotu vs Chelido 0.193932 0.013189 14.704055 Dendronotu vs Philino 0.195379 0.013168 14.836971 Dendronotu vs Philino 0.192785 0.013195 14.610164 Dendronotu vs Sagamin 0.193427 0.013161 14.696568 Dendronotu vs Toledon 0.202524 0.013136 15.417891 Dendronotu vs Diaphan 0.197571 0.013274 14.883832 Dendronotu vs Newnesi 0.198189 0.013189 15.026342 Dendronotu vs Colpoda 0.187439 0.013214 14.184457 Dendronotu vs Runcina 0.190589 0.013291 14.339922 Dendronotu vs Unela_g 0.202029 0.013225 15.276353 Dendronotu vs Pontohe 0.193974 0.013209 14.685254 Dendronotu vs Hedylop 0.204869 0.013153 15.575366 Dendronotu vs Parahed 0.201887 0.013215 15.277089 Dendronotu vs Umbracu 0.198082 0.013386 14.797621 Dendronotu vs Tylodin 0.198349 0.013317 14.894159 Dendronotu vs Akera_b 0.201119 0.013101 15.351546 Dendronotu vs Aplysia 0.202299 0.013162 15.370495 Dendronotu vs Dolabel 0.201678 0.013146 15.341327 Dendronotu vs Siphono 0.201183 0.013116 15.338188 Dendronotu vs Bursate 0.203172 0.013124 15.480519 Dendronotu vs Petalif 0.202879 0.013105 15.480999 Dendronotu vs Dolabri 0.202276 0.013125 15.411885 Dendronotu vs Styloch 0.201074 0.013143 15.299329 Dendronotu vs Clio_py 0.185162 0.013268 13.955140 Dendronotu vs Diacria 0.184920 0.013376 13.824847 Dendronotu vs Creseis 0.166650 0.013180 12.644493 Dendronotu vs Spongio 0.194823 0.013216 14.740948 Dendronotu vs Calliop 0.183432 0.013436 13.652608 Dendronotu vs Limapon 0.190820 0.013257 14.394238 Dendronotu vs Thuridi 0.196834 0.013144 14.975358 Dendronotu vs Elysia_ 0.197246 0.013191 14.952805 Dendronotu vs Elysiel 0.197688 0.013111 15.077878 Dendronotu vs Boselli 0.197715 0.013107 15.084250 Dendronotu vs Cyerce_ 0.201369 0.013067 15.410103 Dendronotu vs Placobr 0.192276 0.013159 14.611392 Dendronotu vs Placida 0.199099 0.013115 15.181164 Dendronotu vs Polybra 0.198327 0.013031 15.219459 Cuthona_na vs Flabell 0.003902 0.005163 0.755799 Cuthona_na vs Pleurob 0.136542 0.012345 11.060400 Cuthona_na vs Berthel 0.137095 0.012292 11.153366 Cuthona_na vs Bathybe 0.132895 0.012504 10.628298 Cuthona_na vs Euselen 0.136751 0.012374 11.051087 Cuthona_na vs Tomptho 0.138398 0.012387 11.173294 Cuthona_na vs Berthel 0.032939 0.013680 2.407834 Cuthona_na vs Berthel 0.102403 0.012801 7.999503 Cuthona_na vs Pupa_so 0.169605 0.012518 13.548899

194
Cuthona_na vs Hydatin 0.171956 0.012638 13.606015 Cuthona_na vs Bullina 0.175787 0.012541 14.016526 Cuthona_na vs Acteon_ 0.186512 0.013018 14.327525 Cuthona_na vs Haminoe 0.185931 0.012941 14.367476 Cuthona_na vs Cylichn 0.193158 0.012897 14.976764 Cuthona_na vs Scaphan 0.192367 0.012990 14.809284 Cuthona_na vs cf._Ret 0.187125 0.013012 14.380968 Cuthona_na vs Bulla_c 0.193750 0.012902 15.017373 Cuthona_na vs Smaragd 0.191065 0.012940 14.765827 Cuthona_na vs Phanero 0.188972 0.012955 14.586992 Cuthona_na vs Bullact 0.191373 0.012959 14.767338 Cuthona_na vs Atys_se 0.190292 0.012949 14.695242 Cuthona_na vs Odontog 0.189086 0.012971 14.577177 Cuthona_na vs Navanax 0.188333 0.012944 14.549781 Cuthona_na vs Chelido 0.188243 0.012956 14.529214 Cuthona_na vs Philino 0.189064 0.012990 14.554628 Cuthona_na vs Philino 0.186506 0.012991 14.356421 Cuthona_na vs Sagamin 0.186524 0.012976 14.374845 Cuthona_na vs Toledon 0.194351 0.012905 15.059997 Cuthona_na vs Diaphan 0.191204 0.012981 14.729318 Cuthona_na vs Newnesi 0.191867 0.012839 14.944209 Cuthona_na vs Colpoda 0.179734 0.012947 13.882502 Cuthona_na vs Runcina 0.184243 0.013017 14.153629 Cuthona_na vs Unela_g 0.195672 0.012897 15.171396 Cuthona_na vs Pontohe 0.185651 0.012868 14.427033 Cuthona_na vs Hedylop 0.198496 0.012846 15.452467 Cuthona_na vs Parahed 0.195536 0.012888 15.172113 Cuthona_na vs Umbracu 0.192272 0.013024 14.763330 Cuthona_na vs Tylodin 0.190666 0.013015 14.649855 Cuthona_na vs Akera_b 0.192305 0.012912 14.893096 Cuthona_na vs Aplysia 0.194236 0.012900 15.057634 Cuthona_na vs Dolabel 0.193620 0.012961 14.939009 Cuthona_na vs Siphono 0.193800 0.012853 15.077633 Cuthona_na vs Bursate 0.195843 0.012889 15.194668 Cuthona_na vs Petalif 0.196157 0.012907 15.198125 Cuthona_na vs Dolabri 0.194962 0.012928 15.080130 Cuthona_na vs Styloch 0.194362 0.012967 14.988624 Cuthona_na vs Clio_py 0.175173 0.013027 13.446475 Cuthona_na vs Diacria 0.173487 0.013210 13.133110 Cuthona_na vs Creseis 0.153909 0.012973 11.863495 Cuthona_na vs Spongio 0.187231 0.012985 14.419035 Cuthona_na vs Calliop 0.176782 0.013051 13.545214 Cuthona_na vs Limapon 0.183868 0.012950 14.198432 Cuthona_na vs Thuridi 0.188551 0.012791 14.741315 Cuthona_na vs Elysia_ 0.188972 0.012893 14.656932 Cuthona_na vs Elysiel 0.190038 0.012906 14.724620 Cuthona_na vs Boselli 0.188845 0.012866 14.677889 Cuthona_na vs Cyerce_ 0.193072 0.012788 15.097289 Cuthona_na vs Placobr 0.182605 0.012867 14.191173 Cuthona_na vs Placida 0.191409 0.012814 14.937170 Cuthona_na vs Polybra 0.189449 0.012811 14.787528 Flabellina vs Pleurob 0.133292 0.012141 10.978690 Flabellina vs Berthel 0.134499 0.012000 11.208679 Flabellina vs Bathybe 0.129643 0.012289 10.549186 Flabellina vs Euselen 0.133495 0.012165 10.974138 Flabellina vs Tomptho 0.135149 0.012183 11.092984 Flabellina vs Berthel 0.031429 0.013520 2.324702 Flabellina vs Berthel 0.102342 0.012621 8.108701 Flabellina vs Pupa_so 0.165622 0.012560 13.186889 Flabellina vs Hydatin 0.168800 0.012613 13.383051 Flabellina vs Bullina 0.172629 0.012516 13.793063 Flabellina vs Acteon_ 0.182609 0.012950 14.100926 Flabellina vs Haminoe 0.182027 0.012893 14.118471 Flabellina vs Cylichn 0.189255 0.012752 14.841648 Flabellina vs Scaphan 0.189260 0.012878 14.696448 Flabellina vs cf._Ret 0.183222 0.012906 14.196784 Flabellina vs Bulla_c 0.189848 0.012737 14.905418 Flabellina vs Smaragd 0.187154 0.012774 14.650685 Flabellina vs Phanero 0.185069 0.012810 14.447667 Flabellina vs Bullact 0.187469 0.012756 14.696597 Flabellina vs Atys_se 0.186385 0.012765 14.601423 Flabellina vs Odontog 0.185180 0.012884 14.372852 Flabellina vs Navanax 0.184430 0.012915 14.280404 Flabellina vs Chelido 0.184341 0.012850 14.345862 Flabellina vs Philino 0.185158 0.012845 14.415105 Flabellina vs Philino 0.182605 0.012866 14.193252 Flabellina vs Sagamin 0.183230 0.012834 14.276837 Flabellina vs Toledon 0.190448 0.012779 14.903495

195
Flabellina vs Diaphan 0.187276 0.012893 14.525335 Flabellina vs Newnesi 0.187965 0.012751 14.740880 Flabellina vs Colpoda 0.175834 0.012723 13.820121 Flabellina vs Runcina 0.180338 0.012876 14.005699 Flabellina vs Unela_g 0.191766 0.012809 14.970779 Flabellina vs Pontohe 0.181754 0.012762 14.242091 Flabellina vs Hedylop 0.194594 0.012719 15.299544 Flabellina vs Parahed 0.191633 0.012800 14.971465 Flabellina vs Umbracu 0.188362 0.012901 14.600177 Flabellina vs Tylodin 0.186765 0.012928 14.446926 Flabellina vs Akera_b 0.189232 0.012710 14.888689 Flabellina vs Aplysia 0.190336 0.012735 14.946197 Flabellina vs Dolabel 0.189721 0.012739 14.893399 Flabellina vs Siphono 0.189896 0.012688 14.966399 Flabellina vs Bursate 0.191108 0.012692 15.057716 Flabellina vs Petalif 0.192253 0.012742 15.088510 Flabellina vs Dolabri 0.191059 0.012744 14.991710 Flabellina vs Styloch 0.190458 0.012764 14.921343 Flabellina vs Clio_py 0.172103 0.012800 13.445104 Flabellina vs Diacria 0.170413 0.013043 13.065303 Flabellina vs Creseis 0.150840 0.012788 11.795697 Flabellina vs Spongio 0.184153 0.012832 14.350714 Flabellina vs Calliop 0.175063 0.012968 13.499556 Flabellina vs Limapon 0.182140 0.012803 14.226105 Flabellina vs Thuridi 0.184643 0.012625 14.625640 Flabellina vs Elysia_ 0.185894 0.012763 14.564680 Flabellina vs Elysiel 0.186962 0.012699 14.722631 Flabellina vs Boselli 0.185769 0.012736 14.585799 Flabellina vs Cyerce_ 0.189995 0.012659 15.009280 Flabellina vs Placobr 0.180136 0.012661 14.227224 Flabellina vs Placida 0.188330 0.012665 14.870401 Flabellina vs Polybra 0.186972 0.012703 14.719153 Pleurobran vs Berthel 0.001267 0.002957 0.428349 Pleurobran vs Bathybe -0.003876 0.003168 -1.223501 Pleurobran vs Euselen -0.000653 0.003101 -0.210664 Pleurobran vs Tomptho 0.001917 0.003195 0.599987 Pleurobran vs Berthel -0.100228 0.009818 -10.208964 Pleurobran vs Berthel -0.032040 0.006572 -4.875560 Pleurobran vs Pupa_so 0.032217 0.007354 4.380852 Pleurobran vs Hydatin 0.035847 0.007383 4.855300 Pleurobran vs Bullina 0.040115 0.007372 5.441361 Pleurobran vs Acteon_ 0.049085 0.007309 6.715547 Pleurobran vs Haminoe 0.047267 0.007187 6.576854 Pleurobran vs Cylichn 0.054383 0.007140 7.616449 Pleurobran vs Scaphan 0.052414 0.007181 7.299273 Pleurobran vs cf._Ret 0.049636 0.007331 6.771041 Pleurobran vs Bulla_c 0.054973 0.007167 7.670583 Pleurobran vs Smaragd 0.052142 0.007193 7.248985 Pleurobran vs Phanero 0.050261 0.007148 7.031172 Pleurobran vs Bullact 0.053216 0.007094 7.501751 Pleurobran vs Atys_se 0.051458 0.007206 7.141135 Pleurobran vs Odontog 0.050247 0.007097 7.080005 Pleurobran vs Navanax 0.050225 0.007281 6.898393 Pleurobran vs Chelido 0.051450 0.007309 7.039566 Pleurobran vs Philino 0.050248 0.007234 6.945721 Pleurobran vs Philino 0.047845 0.007040 6.795934 Pleurobran vs Sagamin 0.047849 0.007150 6.692212 Pleurobran vs Toledon 0.056159 0.007141 7.864380 Pleurobran vs Diaphan 0.051073 0.007185 7.108667 Pleurobran vs Newnesi 0.053823 0.007230 7.444009 Pleurobran vs Colpoda 0.040716 0.007426 5.482649 Pleurobran vs Runcina 0.046209 0.007150 6.463010 Pleurobran vs Unela_g 0.059121 0.006903 8.564983 Pleurobran vs Pontohe 0.051488 0.006863 7.501977 Pleurobran vs Hedylop 0.062042 0.006766 9.169558 Pleurobran vs Parahed 0.059085 0.006898 8.565016 Pleurobran vs Umbracu 0.055076 0.007045 7.817191 Pleurobran vs Tylodin 0.053215 0.006982 7.621885 Pleurobran vs Akera_b 0.056134 0.007008 8.010115 Pleurobran vs Aplysia 0.056719 0.006926 8.189756 Pleurobran vs Dolabel 0.056715 0.006981 8.124475 Pleurobran vs Siphono 0.056181 0.006930 8.107522 Pleurobran vs Bursate 0.056736 0.006875 8.251997 Pleurobran vs Petalif 0.058458 0.006921 8.446414 Pleurobran vs Dolabri 0.057286 0.006950 8.242587 Pleurobran vs Styloch 0.056697 0.007005 8.094161 Pleurobran vs Clio_py 0.038924 0.007417 5.247630 Pleurobran vs Diacria 0.035866 0.007563 4.742563

196
Pleurobran vs Creseis 0.016846 0.007737 2.177453 Pleurobran vs Spongio 0.050259 0.007069 7.109339 Pleurobran vs Calliop 0.038924 0.007505 5.186450 Pleurobran vs Limapon 0.046158 0.007001 6.592769 Pleurobran vs Thuridi 0.052131 0.006832 7.629970 Pleurobran vs Elysia_ 0.051401 0.006899 7.450708 Pleurobran vs Elysiel 0.052545 0.006868 7.650854 Pleurobran vs Boselli 0.051966 0.006730 7.721359 Pleurobran vs Cyerce_ 0.056095 0.006809 8.238775 Pleurobran vs Placobr 0.045460 0.006708 6.777378 Pleurobran vs Placida 0.053150 0.006920 7.680332 Pleurobran vs Polybra 0.051966 0.006756 7.691357 Berthellin vs Bathybe -0.005129 0.002986 -1.717731 Berthellin vs Euselen -0.001281 0.002766 -0.463177 Berthellin vs Tomptho 0.000650 0.002815 0.230873 Berthellin vs Berthel -0.102732 0.009706 -10.584699 Berthellin vs Berthel -0.033972 0.006336 -5.361684 Berthellin vs Pupa_so 0.030931 0.007188 4.303255 Berthellin vs Hydatin 0.034558 0.007244 4.770495 Berthellin vs Bullina 0.038822 0.007207 5.387142 Berthellin vs Acteon_ 0.047786 0.007327 6.521480 Berthellin vs Haminoe 0.045970 0.007204 6.381375 Berthellin vs Cylichn 0.053082 0.007132 7.442542 Berthellin vs Scaphan 0.051104 0.007173 7.124460 Berthellin vs cf._Ret 0.047740 0.007325 6.517783 Berthellin vs Bulla_c 0.053672 0.007106 7.553510 Berthellin vs Smaragd 0.050839 0.007210 7.051514 Berthellin vs Phanero 0.048963 0.007138 6.858972 Berthellin vs Bullact 0.051915 0.007057 7.356546 Berthellin vs Atys_se 0.050158 0.007222 6.944766 Berthellin vs Odontog 0.048949 0.007112 6.882203 Berthellin vs Navanax 0.048927 0.007299 6.702994 Berthellin vs Chelido 0.050150 0.007275 6.893784 Berthellin vs Philino 0.048948 0.007201 6.797782 Berthellin vs Philino 0.046548 0.007031 6.620087 Berthellin vs Sagamin 0.046552 0.007143 6.516914 Berthellin vs Toledon 0.054856 0.006974 7.865918 Berthellin vs Diaphan 0.049767 0.007175 6.936054 Berthellin vs Newnesi 0.052522 0.007036 7.464245 Berthellin vs Colpoda 0.039410 0.007338 5.370512 Berthellin vs Runcina 0.044269 0.007027 6.299412 Berthellin vs Unela_g 0.057817 0.006785 8.521651 Berthellin vs Pontohe 0.050187 0.006800 7.380014 Berthellin vs Hedylop 0.060109 0.006724 8.939445 Berthellin vs Parahed 0.057782 0.006781 8.521685 Berthellin vs Umbracu 0.053785 0.007009 7.673451 Berthellin vs Tylodin 0.051902 0.006997 7.418208 Berthellin vs Akera_b 0.054833 0.006837 8.020543 Berthellin vs Aplysia 0.055417 0.006835 8.107430 Berthellin vs Dolabel 0.055413 0.006809 8.138407 Berthellin vs Siphono 0.054879 0.006839 8.024409 Berthellin vs Bursate 0.055422 0.006754 8.205592 Berthellin vs Petalif 0.057156 0.006803 8.401107 Berthellin vs Dolabri 0.055985 0.006833 8.193699 Berthellin vs Styloch 0.055396 0.006833 8.106630 Berthellin vs Clio_py 0.037630 0.007358 5.114016 Berthellin vs Diacria 0.034577 0.007504 4.607817 Berthellin vs Creseis 0.015568 0.007652 2.034554 Berthellin vs Spongio 0.048961 0.006953 7.041189 Berthellin vs Calliop 0.037005 0.007386 5.009917 Berthellin vs Limapon 0.044873 0.006938 6.467871 Berthellin vs Thuridi 0.050830 0.006739 7.542613 Berthellin vs Elysia_ 0.050103 0.006862 7.301846 Berthellin vs Elysiel 0.051247 0.006775 7.563618 Berthellin vs Boselli 0.050668 0.006692 7.570958 Berthellin vs Cyerce_ 0.054795 0.006743 8.126017 Berthellin vs Placobr 0.044167 0.006642 6.649750 Berthellin vs Placida 0.051865 0.006803 7.623413 Berthellin vs Polybra 0.050668 0.006691 7.572745 Bathyberth vs Euselen 0.003222 0.003174 1.014967 Bathyberth vs Tomptho 0.005780 0.003345 1.728223 Bathyberth vs Berthel -0.097817 0.009889 -9.891477 Bathyberth vs Berthel -0.030086 0.006598 -4.559646 Bathyberth vs Pupa_so 0.036121 0.007421 4.867553 Bathyberth vs Hydatin 0.039127 0.007475 5.234271 Bathyberth vs Bullina 0.043399 0.007463 5.815407 Bathyberth vs Acteon_ 0.053015 0.007530 7.040366 Bathyberth vs Haminoe 0.051193 0.007385 6.932226

197
Bathyberth vs Cylichn 0.058316 0.007338 7.946595 Bathyberth vs Scaphan 0.056379 0.007382 7.637543 Bathyberth vs cf._Ret 0.052967 0.007527 7.037098 Bathyberth vs Bulla_c 0.059537 0.007343 8.107987 Bathyberth vs Smaragd 0.056083 0.007416 7.562848 Bathyberth vs Phanero 0.054190 0.007371 7.351780 Bathyberth vs Bullact 0.057147 0.007318 7.809381 Bathyberth vs Atys_se 0.055356 0.007423 7.457649 Bathyberth vs Odontog 0.054176 0.007345 7.375868 Bathyberth vs Navanax 0.054155 0.007502 7.218442 Bathyberth vs Chelido 0.055383 0.007504 7.380166 Bathyberth vs Philino 0.054180 0.007457 7.265496 Bathyberth vs Philino 0.051769 0.007159 7.230823 Bathyberth vs Sagamin 0.051777 0.007398 6.998691 Bathyberth vs Toledon 0.059501 0.007262 8.193448 Bathyberth vs Diaphan 0.055024 0.007358 7.478391 Bathyberth vs Newnesi 0.057164 0.007325 7.803914 Bathyberth vs Colpoda 0.044620 0.007591 5.878013 Bathyberth vs Runcina 0.048894 0.007394 6.612254 Bathyberth vs Unela_g 0.062471 0.007107 8.790381 Bathyberth vs Pontohe 0.055423 0.007041 7.871894 Bathyberth vs Hedylop 0.064766 0.006968 9.294121 Bathyberth vs Parahed 0.062433 0.007102 8.790423 Bathyberth vs Umbracu 0.058400 0.007294 8.006085 Bathyberth vs Tylodin 0.056536 0.007181 7.872803 Bathyberth vs Akera_b 0.059474 0.007208 8.251171 Bathyberth vs Aplysia 0.060060 0.007075 8.488791 Bathyberth vs Dolabel 0.060056 0.007130 8.422996 Bathyberth vs Siphono 0.059523 0.007079 8.408228 Bathyberth vs Bursate 0.060693 0.007052 8.605922 Bathyberth vs Petalif 0.062431 0.007048 8.858227 Bathyberth vs Dolabri 0.061257 0.007077 8.656330 Bathyberth vs Styloch 0.060667 0.007131 8.507745 Bathyberth vs Clio_py 0.043481 0.007538 5.768272 Bathyberth vs Diacria 0.040411 0.007630 5.296149 Bathyberth vs Creseis 0.021378 0.007875 2.714693 Bathyberth vs Spongio 0.054223 0.007195 7.536402 Bathyberth vs Calliop 0.041604 0.007644 5.443046 Bathyberth vs Limapon 0.049439 0.007171 6.894612 Bathyberth vs Thuridi 0.056104 0.006960 8.060587 Bathyberth vs Elysia_ 0.055362 0.007052 7.849937 Bathyberth vs Elysiel 0.056505 0.007022 8.046881 Bathyberth vs Boselli 0.055925 0.006886 8.121674 Bathyberth vs Cyerce_ 0.059431 0.006959 8.539509 Bathyberth vs Placobr 0.048777 0.006859 7.111317 Bathyberth vs Placida 0.056481 0.007097 7.958891 Bathyberth vs Polybra 0.055294 0.006907 8.005005 Euselenops vs Tomptho 0.002559 0.002820 0.907553 Euselenops vs Berthel -0.101036 0.009853 -10.253823 Euselenops vs Berthel -0.032052 0.006355 -5.043886 Euselenops vs Pupa_so 0.032309 0.007245 4.459481 Euselenops vs Hydatin 0.034678 0.007373 4.703244 Euselenops vs Bullina 0.038955 0.007335 5.310621 Euselenops vs Acteon_ 0.048545 0.007349 6.605407 Euselenops vs Haminoe 0.046725 0.007225 6.466879 Euselenops vs Cylichn 0.053852 0.007154 7.527663 Euselenops vs Scaphan 0.051876 0.007195 7.210304 Euselenops vs cf._Ret 0.049098 0.007319 6.708623 Euselenops vs Bulla_c 0.054442 0.007180 7.582252 Euselenops vs Smaragd 0.051606 0.007231 7.136409 Euselenops vs Phanero 0.049723 0.007187 6.918753 Euselenops vs Bullact 0.052682 0.007132 7.386700 Euselenops vs Atys_se 0.050922 0.007244 7.029419 Euselenops vs Odontog 0.050338 0.007189 7.001786 Euselenops vs Navanax 0.049689 0.007321 6.787176 Euselenops vs Chelido 0.050915 0.007323 6.953063 Euselenops vs Philino 0.049711 0.007301 6.808660 Euselenops vs Philino 0.047303 0.007025 6.733717 Euselenops vs Sagamin 0.047309 0.007244 6.530570 Euselenops vs Toledon 0.055037 0.007075 7.778769 Euselenops vs Diaphan 0.050534 0.007223 6.995968 Euselenops vs Newnesi 0.052696 0.007139 7.381034 Euselenops vs Colpoda 0.040150 0.007412 5.416826 Euselenops vs Runcina 0.045049 0.007162 6.289926 Euselenops vs Unela_g 0.058007 0.006916 8.386828 Euselenops vs Pontohe 0.050952 0.006932 7.350270 Euselenops vs Hedylop 0.060934 0.006805 8.953938 Euselenops vs Parahed 0.057971 0.006912 8.386861

198
Euselenops vs Umbracu 0.054560 0.007113 7.669977 Euselenops vs Tylodin 0.052074 0.006993 7.446077 Euselenops vs Akera_b 0.055014 0.006941 7.926331 Euselenops vs Aplysia 0.055600 0.006912 8.043817 Euselenops vs Dolabel 0.055596 0.006968 7.979331 Euselenops vs Siphono 0.055060 0.006916 7.961393 Euselenops vs Bursate 0.055603 0.006831 8.139558 Euselenops vs Petalif 0.057343 0.006880 8.334690 Euselenops vs Dolabri 0.056169 0.006909 8.129391 Euselenops vs Styloch 0.055578 0.006965 7.980172 Euselenops vs Clio_py 0.038369 0.007458 5.144421 Euselenops vs Diacria 0.035307 0.007552 4.675412 Euselenops vs Creseis 0.016259 0.007751 2.097745 Euselenops vs Spongio 0.049125 0.007003 7.014902 Euselenops vs Calliop 0.037762 0.007415 5.092278 Euselenops vs Limapon 0.045627 0.007015 6.504606 Euselenops vs Thuridi 0.051629 0.006791 7.602385 Euselenops vs Elysia_ 0.050898 0.006886 7.391618 Euselenops vs Elysiel 0.052045 0.006855 7.592478 Euselenops vs Boselli 0.051465 0.006743 7.632698 Euselenops vs Cyerce_ 0.055604 0.006768 8.216134 Euselenops vs Placobr 0.044944 0.006749 6.659758 Euselenops vs Placida 0.052652 0.006908 7.622318 Euselenops vs Polybra 0.051465 0.006715 7.664631 Tompthomps vs Berthel -0.101406 0.009764 -10.385797 Tompthomps vs Berthel -0.033319 0.006461 -5.156569 Tompthomps vs Pupa_so 0.031526 0.007236 4.356561 Tompthomps vs Hydatin 0.034513 0.007288 4.735646 Tompthomps vs Bullina 0.038775 0.007327 5.291921 Tompthomps vs Acteon_ 0.047134 0.007267 6.485657 Tompthomps vs Haminoe 0.045318 0.007245 6.255063 Tompthomps vs Cylichn 0.052433 0.007123 7.361556 Tompthomps vs Scaphan 0.050447 0.007163 7.042863 Tompthomps vs cf._Ret 0.047089 0.007315 6.437286 Tompthomps vs Bulla_c 0.053650 0.007126 7.528374 Tompthomps vs Smaragd 0.050187 0.007251 6.921600 Tompthomps vs Phanero 0.048313 0.007206 6.704125 Tompthomps vs Bullact 0.051267 0.007152 7.168236 Tompthomps vs Atys_se 0.049507 0.007263 6.815994 Tompthomps vs Odontog 0.048298 0.007078 6.824025 Tompthomps vs Navanax 0.048275 0.007263 6.646346 Tompthomps vs Chelido 0.049499 0.007291 6.788992 Tompthomps vs Philino 0.048296 0.007217 6.691774 Tompthomps vs Philino 0.045897 0.006995 6.561886 Tompthomps vs Sagamin 0.045900 0.007160 6.410436 Tompthomps vs Toledon 0.054208 0.007070 7.667856 Tompthomps vs Diaphan 0.049113 0.007140 6.878123 Tompthomps vs Newnesi 0.051872 0.007133 7.271831 Tompthomps vs Colpoda 0.038767 0.007355 5.270490 Tompthomps vs Runcina 0.044217 0.007151 6.183501 Tompthomps vs Unela_g 0.057169 0.006911 8.271687 Tompthomps vs Pontohe 0.049536 0.006873 7.206813 Tompthomps vs Hedylop 0.059462 0.006797 8.748731 Tompthomps vs Parahed 0.057134 0.006907 8.271715 Tompthomps vs Umbracu 0.053100 0.007078 7.502428 Tompthomps vs Tylodin 0.051846 0.006988 7.419667 Tompthomps vs Akera_b 0.054190 0.006882 7.874567 Tompthomps vs Aplysia 0.054775 0.006853 7.992491 Tompthomps vs Dolabel 0.054771 0.006908 7.928028 Tompthomps vs Siphono 0.054236 0.006857 7.909521 Tompthomps vs Bursate 0.055408 0.006777 8.176297 Tompthomps vs Petalif 0.057142 0.006826 8.371525 Tompthomps vs Dolabri 0.055970 0.006855 8.165120 Tompthomps vs Styloch 0.055382 0.006910 8.014761 Tompthomps vs Clio_py 0.037613 0.007379 5.097238 Tompthomps vs Diacria 0.034559 0.007473 4.624674 Tompthomps vs Creseis 0.015548 0.007672 2.026518 Tompthomps vs Spongio 0.048944 0.006975 7.016668 Tompthomps vs Calliop 0.036987 0.007383 5.009603 Tompthomps vs Limapon 0.044228 0.006958 6.356448 Tompthomps vs Thuridi 0.051403 0.006733 7.634787 Tompthomps vs Elysia_ 0.050675 0.006802 7.450308 Tompthomps vs Elysiel 0.051818 0.006797 7.624082 Tompthomps vs Boselli 0.051240 0.006658 7.695921 Tompthomps vs Cyerce_ 0.054738 0.006733 8.129680 Tompthomps vs Placobr 0.044116 0.006688 6.596115 Tompthomps vs Placida 0.051797 0.006845 7.566662 Tompthomps vs Polybra 0.050022 0.006709 7.455661

199
Berthella_ vs Berthel 0.071313 0.010531 6.771708 Berthella_ vs Pupa_so 0.130776 0.010936 11.958458 Berthella_ vs Hydatin 0.133449 0.011064 12.061171 Berthella_ vs Bullina 0.137798 0.011055 12.464834 Berthella_ vs Acteon_ 0.145482 0.011194 12.995957 Berthella_ vs Haminoe 0.144786 0.011191 12.937960 Berthella_ vs Cylichn 0.150205 0.011083 13.553101 Berthella_ vs Scaphan 0.146922 0.011095 13.242674 Berthella_ vs cf._Ret 0.144778 0.011246 12.873769 Berthella_ vs Bulla_c 0.152096 0.011109 13.690713 Berthella_ vs Smaragd 0.148792 0.011199 13.286392 Berthella_ vs Phanero 0.146611 0.011128 13.174641 Berthella_ vs Bullact 0.149609 0.011046 13.544527 Berthella_ vs Atys_se 0.147898 0.011155 13.258782 Berthella_ vs Odontog 0.148410 0.011145 13.316440 Berthella_ vs Navanax 0.146573 0.011190 13.098372 Berthella_ vs Chelido 0.146679 0.011196 13.100912 Berthella_ vs Philino 0.146063 0.011158 13.090537 Berthella_ vs Philino 0.144098 0.011048 13.043478 Berthella_ vs Sagamin 0.145393 0.011149 13.040516 Berthella_ vs Toledon 0.151401 0.011065 13.682919 Berthella_ vs Diaphan 0.146465 0.011109 13.184893 Berthella_ vs Newnesi 0.148487 0.011058 13.428484 Berthella_ vs Colpoda 0.135582 0.011309 11.988840 Berthella_ vs Runcina 0.143036 0.010959 13.052177 Berthella_ vs Unela_g 0.155651 0.010885 14.299097 Berthella_ vs Pontohe 0.147285 0.010900 13.512424 Berthella_ vs Hedylop 0.158452 0.010838 14.620393 Berthella_ vs Parahed 0.155548 0.010878 14.299422 Berthella_ vs Umbracu 0.152862 0.011095 13.778019 Berthella_ vs Tylodin 0.150165 0.010913 13.759916 Berthella_ vs Akera_b 0.152020 0.010944 13.890120 Berthella_ vs Aplysia 0.152561 0.010901 13.994966 Berthella_ vs Dolabel 0.152541 0.010941 13.941595 Berthella_ vs Siphono 0.151353 0.010901 13.884431 Berthella_ vs Bursate 0.153289 0.010873 14.098116 Berthella_ vs Petalif 0.154991 0.010943 14.163207 Berthella_ vs Dolabri 0.153802 0.010945 14.052408 Berthella_ vs Styloch 0.153199 0.010944 13.998252 Berthella_ vs Clio_py 0.135228 0.011246 12.024509 Berthella_ vs Diacria 0.133504 0.011341 11.771428 Berthella_ vs Creseis 0.117256 0.011312 10.365980 Berthella_ vs Spongio 0.146822 0.011043 13.295786 Berthella_ vs Calliop 0.136547 0.011212 12.178620 Berthella_ vs Limapon 0.144744 0.010980 13.182249 Berthella_ vs Thuridi 0.152058 0.010964 13.869397 Berthella_ vs Elysia_ 0.150441 0.010974 13.708489 Berthella_ vs Elysiel 0.150936 0.010906 13.839903 Berthella_ vs Boselli 0.151546 0.010905 13.896311 Berthella_ vs Cyerce_ 0.153836 0.010917 14.090807 Berthella_ vs Placobr 0.145000 0.010872 13.337359 Berthella_ vs Placida 0.151428 0.010859 13.944345 Berthella_ vs Polybra 0.150822 0.010940 13.786430 Berthella_ vs Pupa_so 0.066375 0.008575 7.740181 Berthella_ vs Hydatin 0.069375 0.008640 8.029397 Berthella_ vs Bullina 0.073758 0.008729 8.450023 Berthella_ vs Acteon_ 0.081663 0.008814 9.265397 Berthella_ vs Haminoe 0.081690 0.008647 9.447274 Berthella_ vs Cylichn 0.087748 0.008635 10.162188 Berthella_ vs Scaphan 0.087299 0.008574 10.181621 Berthella_ vs cf._Ret 0.082892 0.008784 9.436793 Berthella_ vs Bulla_c 0.089016 0.008711 10.218520 Berthella_ vs Smaragd 0.085545 0.008688 9.846481 Berthella_ vs Phanero 0.083527 0.008665 9.639893 Berthella_ vs Bullact 0.086551 0.008567 10.102356 Berthella_ vs Atys_se 0.084777 0.008670 9.777909 Berthella_ vs Odontog 0.082267 0.008689 9.467508 Berthella_ vs Navanax 0.082272 0.008839 9.307562 Berthella_ vs Chelido 0.082892 0.008860 9.355681 Berthella_ vs Philino 0.081712 0.008750 9.338477 Berthella_ vs Philino 0.079813 0.008617 9.262067 Berthella_ vs Sagamin 0.079835 0.008771 9.102380 Berthella_ vs Toledon 0.089016 0.008521 10.446352 Berthella_ vs Diaphan 0.083927 0.008714 9.631797 Berthella_ vs Newnesi 0.086590 0.008502 10.185049 Berthella_ vs Colpoda 0.074343 0.008822 8.427441 Berthella_ vs Runcina 0.078781 0.008496 9.272297 Berthella_ vs Unela_g 0.090220 0.008474 10.646351

200
Berthella_ vs Pontohe 0.083563 0.008555 9.767889 Berthella_ vs Hedylop 0.092563 0.008418 10.995938 Berthella_ vs Parahed 0.090162 0.008469 10.646457 Berthella_ vs Umbracu 0.086701 0.008481 10.223259 Berthella_ vs Tylodin 0.084705 0.008394 10.090788 Berthella_ vs Akera_b 0.087722 0.008393 10.451863 Berthella_ vs Aplysia 0.088314 0.008345 10.582350 Berthella_ vs Dolabel 0.088314 0.008418 10.491206 Berthella_ vs Siphono 0.088316 0.008321 10.613431 Berthella_ vs Bursate 0.088377 0.008351 10.582889 Berthella_ vs Petalif 0.090152 0.008394 10.740312 Berthella_ vs Dolabri 0.088949 0.008420 10.563714 Berthella_ vs Styloch 0.088345 0.008421 10.491286 Berthella_ vs Clio_py 0.075687 0.008648 8.752069 Berthella_ vs Diacria 0.070670 0.008906 7.934802 Berthella_ vs Creseis 0.050506 0.009018 5.600274 Berthella_ vs Spongio 0.082373 0.008457 9.740196 Berthella_ vs Calliop 0.068845 0.009064 7.595664 Berthella_ vs Limapon 0.077010 0.008666 8.886131 Berthella_ vs Thuridi 0.084408 0.008541 9.882510 Berthella_ vs Elysia_ 0.083518 0.008614 9.695891 Berthella_ vs Elysiel 0.085333 0.008467 10.078302 Berthella_ vs Boselli 0.084130 0.008468 9.934604 Berthella_ vs Cyerce_ 0.087706 0.008458 10.369236 Berthella_ vs Placobr 0.077394 0.008513 9.091103 Berthella_ vs Placida 0.084675 0.008533 9.922668 Berthella_ vs Polybra 0.082852 0.008513 9.732277 Pupa_solid vs Hydatin 0.003600 0.002874 1.252449 Pupa_solid vs Bullina 0.007240 0.002532 2.859323 Pupa_solid vs Acteon_ 0.012567 0.005662 2.219515 Pupa_solid vs Haminoe 0.011361 0.005535 2.052643 Pupa_solid vs Cylichn 0.019073 0.005384 3.542501 Pupa_solid vs Scaphan 0.016221 0.005437 2.983347 Pupa_solid vs cf._Ret 0.013132 0.005594 2.347510 Pupa_solid vs Bulla_c 0.020268 0.005419 3.739924 Pupa_solid vs Smaragd 0.016745 0.005631 2.973728 Pupa_solid vs Phanero 0.014954 0.005616 2.662750 Pupa_solid vs Bullact 0.017906 0.005485 3.264694 Pupa_solid vs Atys_se 0.016127 0.005589 2.885733 Pupa_solid vs Odontog 0.015546 0.005685 2.734608 Pupa_solid vs Navanax 0.014320 0.005627 2.544885 Pupa_solid vs Chelido 0.016119 0.005525 2.917371 Pupa_solid vs Philino 0.014917 0.005564 2.681241 Pupa_solid vs Philino 0.013152 0.005461 2.408521 Pupa_solid vs Sagamin 0.011952 0.005693 2.099575 Pupa_solid vs Toledon 0.020859 0.005484 3.803823 Pupa_solid vs Diaphan 0.014989 0.005488 2.731225 Pupa_solid vs Newnesi 0.018491 0.005522 3.348283 Pupa_solid vs Colpoda 0.004802 0.005798 0.828199 Pupa_solid vs Runcina 0.011411 0.005538 2.060374 Pupa_solid vs Unela_g 0.025548 0.005073 5.035572 Pupa_solid vs Pontohe 0.015551 0.005167 3.009582 Pupa_solid vs Hedylop 0.027275 0.004927 5.535508 Pupa_solid vs Parahed 0.025532 0.005070 5.035572 Pupa_solid vs Umbracu 0.020893 0.005282 3.955756 Pupa_solid vs Tylodin 0.019677 0.005311 3.704769 Pupa_solid vs Akera_b 0.022042 0.005146 4.283029 Pupa_solid vs Aplysia 0.023214 0.005146 4.511425 Pupa_solid vs Dolabel 0.023215 0.005147 4.510819 Pupa_solid vs Siphono 0.022650 0.005150 4.398356 Pupa_solid vs Bursate 0.024426 0.004941 4.943774 Pupa_solid vs Petalif 0.024973 0.005041 4.953619 Pupa_solid vs Dolabri 0.023804 0.005044 4.718884 Pupa_solid vs Styloch 0.023215 0.005149 4.508241 Pupa_solid vs Clio_py 0.004197 0.005916 0.709448 Pupa_solid vs Diacria 0.003643 0.005757 0.632795 Pupa_solid vs Creseis -0.014756 0.006387 -2.310346 Pupa_solid vs Spongio 0.017356 0.005328 3.257564 Pupa_solid vs Calliop 0.004824 0.005985 0.805921 Pupa_solid vs Limapon 0.013218 0.005502 2.402292 Pupa_solid vs Thuridi 0.020355 0.005294 3.844618 Pupa_solid vs Elysia_ 0.019703 0.005285 3.728355 Pupa_solid vs Elysiel 0.020274 0.005250 3.861819 Pupa_solid vs Boselli 0.020287 0.005180 3.916462 Pupa_solid vs Cyerce_ 0.023799 0.005031 4.730195 Pupa_solid vs Placobr 0.014390 0.005681 2.533262 Pupa_solid vs Placida 0.020283 0.005312 3.818613 Pupa_solid vs Polybra 0.019095 0.004966 3.844961

201
Hydatina_p vs Bullina 0.003645 0.001693 2.153688 Hydatina_p vs Acteon_ 0.010187 0.005402 1.885724 Hydatina_p vs Haminoe 0.008982 0.005199 1.727478 Hydatina_p vs Cylichn 0.016106 0.005078 3.171968 Hydatina_p vs Scaphan 0.013225 0.005055 2.616449 Hydatina_p vs cf._Ret 0.010755 0.005262 2.043824 Hydatina_p vs Bulla_c 0.016694 0.005113 3.265261 Hydatina_p vs Smaragd 0.013767 0.005337 2.579619 Hydatina_p vs Phanero 0.011980 0.005323 2.250837 Hydatina_p vs Bullact 0.014938 0.005185 2.881001 Hydatina_p vs Atys_se 0.013154 0.005293 2.485061 Hydatina_p vs Odontog 0.013168 0.005493 2.397215 Hydatina_p vs Navanax 0.011944 0.005433 2.198453 Hydatina_p vs Chelido 0.013146 0.005430 2.420982 Hydatina_p vs Philino 0.011942 0.005402 2.210499 Hydatina_p vs Philino 0.010176 0.005365 1.896960 Hydatina_p vs Sagamin 0.008973 0.005436 1.650724 Hydatina_p vs Toledon 0.018486 0.005078 3.640324 Hydatina_p vs Diaphan 0.013201 0.005110 2.583661 Hydatina_p vs Newnesi 0.016116 0.005153 3.127285 Hydatina_p vs Colpoda 0.002417 0.005548 0.435674 Hydatina_p vs Runcina 0.009032 0.005238 1.724110 Hydatina_p vs Unela_g 0.022592 0.004935 4.578109 Hydatina_p vs Pontohe 0.013763 0.005060 2.719921 Hydatina_p vs Hedylop 0.024324 0.004784 5.084131 Hydatina_p vs Parahed 0.022578 0.004932 4.578108 Hydatina_p vs Umbracu 0.017925 0.004934 3.632683 Hydatina_p vs Tylodin 0.017320 0.004934 3.510308 Hydatina_p vs Akera_b 0.019659 0.004934 3.984188 Hydatina_p vs Aplysia 0.020243 0.004788 4.227911 Hydatina_p vs Dolabel 0.020245 0.004863 4.163128 Hydatina_p vs Siphono 0.019676 0.004792 4.105754 Hydatina_p vs Bursate 0.021450 0.004795 4.473727 Hydatina_p vs Petalif 0.022588 0.004750 4.754997 Hydatina_p vs Dolabri 0.021419 0.004754 4.505875 Hydatina_p vs Styloch 0.020830 0.004864 4.282066 Hydatina_p vs Clio_py 0.001188 0.005532 0.214701 Hydatina_p vs Diacria 0.000033 0.005783 0.005712 Hydatina_p vs Creseis -0.019021 0.006511 -2.921423 Hydatina_p vs Spongio 0.014966 0.005047 2.965389 Hydatina_p vs Calliop 0.001229 0.005894 0.208579 Hydatina_p vs Limapon 0.009034 0.005268 1.714849 Hydatina_p vs Thuridi 0.016761 0.005225 3.208100 Hydatina_p vs Elysia_ 0.016118 0.005146 3.131971 Hydatina_p vs Elysiel 0.016685 0.005109 3.265865 Hydatina_p vs Boselli 0.016704 0.005039 3.315022 Hydatina_p vs Cyerce_ 0.020829 0.004889 4.259887 Hydatina_p vs Placobr 0.010216 0.005488 1.861424 Hydatina_p vs Placida 0.016720 0.005002 3.342915 Hydatina_p vs Polybra 0.016122 0.005107 3.156663 Bullina_li vs Acteon_ 0.007127 0.005280 1.349759 Bullina_li vs Haminoe 0.005925 0.005075 1.167517 Bullina_li vs Cylichn 0.013041 0.004950 2.634331 Bullina_li vs Scaphan 0.010143 0.004998 2.029393 Bullina_li vs cf._Ret 0.007696 0.005139 1.497478 Bullina_li vs Bulla_c 0.013637 0.004949 2.755207 Bullina_li vs Smaragd 0.010697 0.005213 2.051888 Bullina_li vs Phanero 0.008920 0.005131 1.738611 Bullina_li vs Bullact 0.011874 0.005059 2.347301 Bullina_li vs Atys_se 0.010090 0.005170 1.951787 Bullina_li vs Odontog 0.010104 0.005304 1.904837 Bullina_li vs Navanax 0.008883 0.005311 1.672636 Bullina_li vs Chelido 0.010082 0.005172 1.949598 Bullina_li vs Philino 0.008879 0.005280 1.681570 Bullina_li vs Philino 0.007120 0.005242 1.358291 Bullina_li vs Sagamin 0.005916 0.005349 1.106072 Bullina_li vs Toledon 0.015416 0.004985 3.092524 Bullina_li vs Diaphan 0.010124 0.005088 1.989962 Bullina_li vs Newnesi 0.013049 0.005097 2.560232 Bullina_li vs Colpoda -0.000628 0.005429 -0.115681 Bullina_li vs Runcina 0.005375 0.005220 1.029678 Bullina_li vs Unela_g 0.018930 0.004801 3.942633 Bullina_li vs Pontohe 0.010697 0.004967 2.153709 Bullina_li vs Hedylop 0.020663 0.004648 4.445712 Bullina_li vs Parahed 0.018918 0.004798 3.942631 Bullina_li vs Umbracu 0.014264 0.004913 2.903179 Bullina_li vs Tylodin 0.013661 0.004912 2.780911 Bullina_li vs Akera_b 0.016009 0.004804 3.332628

202
Bullina_li vs Aplysia 0.016594 0.004730 3.508319 Bullina_li vs Dolabel 0.016595 0.004730 3.508206 Bullina_li vs Siphono 0.016025 0.004733 3.385542 Bullina_li vs Bursate 0.017805 0.004661 3.819969 Bullina_li vs Petalif 0.018947 0.004616 4.104397 Bullina_li vs Dolabri 0.017777 0.004619 3.848683 Bullina_li vs Styloch 0.017188 0.004733 3.631632 Bullina_li vs Clio_py -0.002462 0.005582 -0.440998 Bullina_li vs Diacria -0.003618 0.005640 -0.641485 Bullina_li vs Creseis -0.022673 0.006412 -3.536141 Bullina_li vs Spongio 0.010721 0.004993 2.147348 Bullina_li vs Calliop -0.001811 0.005841 -0.309982 Bullina_li vs Limapon 0.005381 0.005248 1.025298 Bullina_li vs Thuridi 0.013108 0.005028 2.606983 Bullina_li vs Elysia_ 0.012475 0.005019 2.485284 Bullina_li vs Elysiel 0.013051 0.004983 2.619313 Bullina_li vs Boselli 0.013062 0.004910 2.660598 Bullina_li vs Cyerce_ 0.017176 0.004644 3.698358 Bullina_li vs Placobr 0.006568 0.005334 1.231368 Bullina_li vs Placida 0.013671 0.004875 2.804274 Bullina_li vs Polybra 0.012472 0.004944 2.522532 Acteon_sp. vs Haminoe -0.001195 0.003434 -0.348093 Acteon_sp. vs Cylichn 0.006494 0.002939 2.209439 Acteon_sp. vs Scaphan 0.004152 0.002976 1.395078 Acteon_sp. vs cf._Ret 0.000569 0.003280 0.173608 Acteon_sp. vs Bulla_c 0.007091 0.003053 2.322342 Acteon_sp. vs Smaragd 0.004143 0.003178 1.303695 Acteon_sp. vs Phanero 0.002389 0.003428 0.696988 Acteon_sp. vs Bullact 0.005332 0.003111 1.713886 Acteon_sp. vs Atys_se 0.003550 0.003226 1.100435 Acteon_sp. vs Odontog 0.003557 0.002697 1.319066 Acteon_sp. vs Navanax 0.002340 0.002038 1.148047 Acteon_sp. vs Chelido 0.004127 0.002695 1.531483 Acteon_sp. vs Philino 0.002927 0.002274 1.287480 Acteon_sp. vs Philino 0.001774 0.003678 0.482288 Acteon_sp. vs Sagamin -0.000018 0.003534 -0.005187 Acteon_sp. vs Toledon 0.008269 0.003533 2.340523 Acteon_sp. vs Diaphan 0.002360 0.003240 0.728556 Acteon_sp. vs Newnesi 0.005314 0.003779 1.406048 Acteon_sp. vs Colpoda -0.005944 0.004056 -1.465476 Acteon_sp. vs Runcina 0.000039 0.004577 0.008464 Acteon_sp. vs Unela_g 0.012362 0.004114 3.004712 Acteon_sp. vs Pontohe 0.002965 0.004339 0.683421 Acteon_sp. vs Hedylop 0.014679 0.004147 3.539836 Acteon_sp. vs Parahed 0.012354 0.004112 3.004710 Acteon_sp. vs Umbracu 0.008295 0.004556 1.820654 Acteon_sp. vs Tylodin 0.007702 0.004515 1.705895 Acteon_sp. vs Akera_b 0.010631 0.004113 2.584715 Acteon_sp. vs Aplysia 0.011212 0.004028 2.783761 Acteon_sp. vs Dolabel 0.011216 0.004115 2.725858 Acteon_sp. vs Siphono 0.010642 0.004116 2.585789 Acteon_sp. vs Bursate 0.011811 0.003942 2.996401 Acteon_sp. vs Petalif 0.012369 0.003849 3.213887 Acteon_sp. vs Dolabri 0.011207 0.003895 2.877062 Acteon_sp. vs Styloch 0.010616 0.003942 2.693346 Acteon_sp. vs Clio_py -0.008352 0.004756 -1.755950 Acteon_sp. vs Diacria -0.011323 0.004832 -2.343322 Acteon_sp. vs Creseis -0.027280 0.006140 -4.442718 Acteon_sp. vs Spongio 0.004181 0.004410 0.948034 Acteon_sp. vs Calliop -0.007113 0.005438 -1.307939 Acteon_sp. vs Limapon -0.001147 0.005120 -0.224018 Acteon_sp. vs Thuridi 0.005375 0.004963 1.083010 Acteon_sp. vs Elysia_ 0.005947 0.004890 1.216114 Acteon_sp. vs Elysiel 0.006528 0.004886 1.336175 Acteon_sp. vs Boselli 0.006537 0.004952 1.320080 Acteon_sp. vs Cyerce_ 0.010639 0.004765 2.232646 Acteon_sp. vs Placobr 0.000647 0.005307 0.121977 Acteon_sp. vs Placida 0.007123 0.004774 1.492031 Acteon_sp. vs Polybra 0.006537 0.004884 1.338297 Haminoea_h vs Cylichn 0.007685 0.002484 3.094205 Haminoea_h vs Scaphan 0.005354 0.002715 1.971747 Haminoea_h vs cf._Ret 0.001764 0.002816 0.626473 Haminoea_h vs Bulla_c 0.008281 0.002934 2.822287 Haminoea_h vs Smaragd 0.005338 0.002642 2.020025 Haminoea_h vs Phanero 0.003583 0.002818 1.271324 Haminoea_h vs Bullact 0.006524 0.002427 2.688295 Haminoea_h vs Atys_se 0.004743 0.002353 2.015782 Haminoea_h vs Odontog 0.004165 0.003383 1.230901

203
Haminoea_h vs Navanax 0.002947 0.003172 0.929213 Haminoea_h vs Chelido 0.004735 0.003584 1.320884 Haminoea_h vs Philino 0.003535 0.003490 1.013047 Haminoea_h vs Philino 0.002381 0.003862 0.616555 Haminoea_h vs Sagamin 0.000588 0.003725 0.157965 Haminoea_h vs Toledon 0.009460 0.003275 2.888057 Haminoea_h vs Diaphan 0.003560 0.003181 1.119134 Haminoea_h vs Newnesi 0.006506 0.003587 1.813795 Haminoea_h vs Colpoda -0.005337 0.004015 -1.329407 Haminoea_h vs Runcina 0.000647 0.004578 0.141222 Haminoea_h vs Unela_g 0.013550 0.004279 3.166974 Haminoea_h vs Pontohe 0.003561 0.004496 0.792084 Haminoea_h vs Hedylop 0.015865 0.004429 3.582188 Haminoea_h vs Parahed 0.013542 0.004276 3.166970 Haminoea_h vs Umbracu 0.008897 0.004704 1.891405 Haminoea_h vs Tylodin 0.008883 0.004701 1.889728 Haminoea_h vs Akera_b 0.011811 0.004197 2.814049 Haminoea_h vs Aplysia 0.012393 0.004071 3.043973 Haminoea_h vs Dolabel 0.012396 0.004281 2.895417 Haminoea_h vs Siphono 0.011832 0.004200 2.817041 Haminoea_h vs Bursate 0.013600 0.004202 3.236282 Haminoea_h vs Petalif 0.014156 0.004072 3.476621 Haminoea_h vs Dolabri 0.013584 0.004076 3.332654 Haminoea_h vs Styloch 0.012404 0.004241 2.924789 Haminoea_h vs Clio_py -0.007159 0.005007 -1.429627 Haminoea_h vs Diacria -0.010129 0.005044 -2.008025 Haminoea_h vs Creseis -0.025482 0.006173 -4.128058 Haminoea_h vs Spongio 0.005366 0.004565 1.175488 Haminoea_h vs Calliop -0.005913 0.005694 -1.038452 Haminoea_h vs Limapon 0.000051 0.005190 0.009761 Haminoea_h vs Thuridi 0.007759 0.005143 1.508561 Haminoea_h vs Elysia_ 0.007140 0.005100 1.399875 Haminoea_h vs Elysiel 0.007720 0.004995 1.545539 Haminoea_h vs Boselli 0.007729 0.004956 1.559399 Haminoea_h vs Cyerce_ 0.011828 0.004878 2.424805 Haminoea_h vs Placobr 0.001843 0.005311 0.347055 Haminoea_h vs Placida 0.008314 0.004955 1.677849 Haminoea_h vs Polybra 0.007729 0.005063 1.526451 Cylichna_g vs Scaphan -0.002397 0.001964 -1.220606 Cylichna_g vs cf._Ret -0.005921 0.002344 -2.525679 Cylichna_g vs Bulla_c 0.000597 0.002181 0.273814 Cylichna_g vs Smaragd -0.002365 0.002196 -1.076916 Cylichna_g vs Phanero -0.004102 0.002542 -1.613624 Cylichna_g vs Bullact -0.001161 0.001838 -0.631594 Cylichna_g vs Atys_se -0.002947 0.001845 -1.597126 Cylichna_g vs Odontog -0.003525 0.002874 -1.226707 Cylichna_g vs Navanax -0.004738 0.002761 -1.716009 Cylichna_g vs Chelido -0.002955 0.002877 -1.027289 Cylichna_g vs Philino -0.004154 0.002759 -1.505476 Cylichna_g vs Philino -0.005313 0.003316 -1.602108 Cylichna_g vs Sagamin -0.007097 0.003108 -2.283626 Cylichna_g vs Toledon 0.001770 0.002549 0.694390 Cylichna_g vs Diaphan -0.004162 0.002632 -1.581533 Cylichna_g vs Newnesi -0.001183 0.002993 -0.395365 Cylichna_g vs Colpoda -0.013036 0.003440 -3.789548 Cylichna_g vs Runcina -0.007060 0.004158 -1.697903 Cylichna_g vs Unela_g 0.005861 0.003788 1.547368 Cylichna_g vs Pontohe -0.002944 0.004202 -0.700525 Cylichna_g vs Hedylop 0.008181 0.003868 2.115268 Cylichna_g vs Parahed 0.005858 0.003786 1.547366 Cylichna_g vs Umbracu 0.001791 0.004217 0.424778 Cylichna_g vs Tylodin 0.001197 0.004256 0.281165 Cylichna_g vs Akera_b 0.004120 0.003552 1.160000 Cylichna_g vs Aplysia 0.004701 0.003454 1.361185 Cylichna_g vs Dolabel 0.004706 0.003649 1.289588 Cylichna_g vs Siphono 0.004134 0.003554 1.163068 Cylichna_g vs Bursate 0.005893 0.003555 1.657688 Cylichna_g vs Petalif 0.006458 0.003456 1.868708 Cylichna_g vs Dolabri 0.005882 0.003507 1.677102 Cylichna_g vs Styloch 0.004706 0.003652 1.288794 Cylichna_g vs Clio_py -0.014857 0.004512 -3.293071 Cylichna_g vs Diacria -0.017824 0.004742 -3.758648 Cylichna_g vs Creseis -0.032609 0.005884 -5.542328 Cylichna_g vs Spongio -0.002329 0.004020 -0.579279 Cylichna_g vs Calliop -0.013607 0.005287 -2.573676 Cylichna_g vs Limapon -0.007650 0.004744 -1.612570 Cylichna_g vs Thuridi 0.000047 0.004764 0.009888 Cylichna_g vs Elysia_ -0.000554 0.004647 -0.119178

204
Cylichna_g vs Elysiel 0.000032 0.004532 0.007047 Cylichna_g vs Boselli 0.000040 0.004603 0.008687 Cylichna_g vs Cyerce_ 0.004138 0.004481 0.923504 Cylichna_g vs Placobr -0.005851 0.004980 -1.174857 Cylichna_g vs Placida 0.000627 0.004490 0.139682 Cylichna_g vs Polybra 0.000035 0.004607 0.007563 Scaphander vs cf._Ret -0.003574 0.002721 -1.313633 Scaphander vs Bulla_c 0.002999 0.002583 1.161052 Scaphander vs Smaragd -0.000583 0.002525 -0.230810 Scaphander vs Phanero -0.002340 0.002840 -0.824100 Scaphander vs Bullact 0.001225 0.002443 0.501483 Scaphander vs Atys_se -0.001173 0.002372 -0.494357 Scaphander vs Odontog -0.001157 0.003089 -0.374723 Scaphander vs Navanax -0.002381 0.002860 -0.832530 Scaphander vs Chelido -0.000582 0.003092 -0.188158 Scaphander vs Philino -0.001791 0.002981 -0.600757 Scaphander vs Philino -0.002959 0.003461 -0.854877 Scaphander vs Sagamin -0.004760 0.003518 -1.353297 Scaphander vs Toledon 0.004183 0.002516 1.662611 Scaphander vs Diaphan -0.001787 0.002452 -0.728620 Scaphander vs Newnesi 0.001205 0.002851 0.422660 Scaphander vs Colpoda -0.010141 0.003864 -2.624491 Scaphander vs Runcina -0.004122 0.004290 -0.960779 Scaphander vs Unela_g 0.007719 0.003882 1.988440 Scaphander vs Pontohe -0.002365 0.004207 -0.562260 Scaphander vs Hedylop 0.010060 0.003961 2.539459 Scaphander vs Parahed 0.007714 0.003880 1.988437 Scaphander vs Umbracu 0.003612 0.004391 0.822518 Scaphander vs Tylodin 0.003611 0.004387 0.823134 Scaphander vs Akera_b 0.007148 0.003834 1.864325 Scaphander vs Aplysia 0.007148 0.003693 1.935236 Scaphander vs Dolabel 0.006562 0.003836 1.710305 Scaphander vs Siphono 0.005977 0.003648 1.638405 Scaphander vs Bursate 0.007155 0.003647 1.962066 Scaphander vs Petalif 0.007725 0.003546 2.178402 Scaphander vs Dolabri 0.007143 0.003597 1.986037 Scaphander vs Styloch 0.005956 0.003648 1.632667 Scaphander vs Clio_py -0.013187 0.004482 -2.941854 Scaphander vs Diacria -0.014959 0.004714 -3.173637 Scaphander vs Creseis -0.032318 0.005944 -5.436927 Scaphander vs Spongio -0.000539 0.004025 -0.133841 Scaphander vs Calliop -0.010719 0.005342 -2.006713 Scaphander vs Limapon -0.004119 0.004868 -0.846145 Scaphander vs Thuridi 0.002450 0.004818 0.508603 Scaphander vs Elysia_ 0.002434 0.004736 0.514018 Scaphander vs Elysiel 0.003023 0.004622 0.654148 Scaphander vs Boselli 0.003030 0.004693 0.645668 Scaphander vs Cyerce_ 0.006569 0.004533 1.449166 Scaphander vs Placobr -0.002906 0.005069 -0.573220 Scaphander vs Placida 0.003026 0.004618 0.655288 Scaphander vs Polybra 0.003027 0.004620 0.655198 cf._Retusa vs Bulla_c 0.006518 0.002689 2.423563 cf._Retusa vs Smaragd 0.003570 0.002948 1.210844 cf._Retusa vs Phanero 0.001819 0.003213 0.566087 cf._Retusa vs Bullact 0.004760 0.002813 1.692328 cf._Retusa vs Atys_se 0.002978 0.002691 1.106692 cf._Retusa vs Odontog 0.002400 0.003383 0.709379 cf._Retusa vs Navanax 0.001183 0.003120 0.379209 cf._Retusa vs Chelido 0.002970 0.003225 0.920832 cf._Retusa vs Philino 0.001770 0.003230 0.548024 cf._Retusa vs Philino 0.000018 0.003671 0.004891 cf._Retusa vs Sagamin -0.001175 0.003487 -0.337053 cf._Retusa vs Toledon 0.007695 0.003274 2.350145 cf._Retusa vs Diaphan 0.001787 0.002886 0.619243 cf._Retusa vs Newnesi 0.004741 0.003537 1.340633 cf._Retusa vs Colpoda -0.007702 0.003965 -1.942667 cf._Retusa vs Runcina -0.001122 0.004540 -0.247027 cf._Retusa vs Unela_g 0.011785 0.004319 2.728917 cf._Retusa vs Pontohe 0.002393 0.004574 0.523171 cf._Retusa vs Hedylop 0.014102 0.004309 3.272897 cf._Retusa vs Parahed 0.011779 0.004316 2.728913 cf._Retusa vs Umbracu 0.007720 0.004628 1.668004 cf._Retusa vs Tylodin 0.007118 0.004587 1.551882 cf._Retusa vs Akera_b 0.010045 0.004111 2.443755 cf._Retusa vs Aplysia 0.010627 0.003982 2.668686 cf._Retusa vs Dolabel 0.010631 0.004196 2.533285 cf._Retusa vs Siphono 0.010065 0.004114 2.446782 cf._Retusa vs Bursate 0.011832 0.004116 2.874807

205
cf._Retusa vs Petalif 0.012390 0.003941 3.143674 cf._Retusa vs Dolabri 0.011816 0.003987 2.963952 cf._Retusa vs Styloch 0.010638 0.004115 2.584837 cf._Retusa vs Clio_py -0.008926 0.004863 -1.835511 cf._Retusa vs Diacria -0.011895 0.005043 -2.358771 cf._Retusa vs Creseis -0.027254 0.006257 -4.355917 cf._Retusa vs Spongio 0.003600 0.004448 0.809319 cf._Retusa vs Calliop -0.008276 0.005499 -1.505054 cf._Retusa vs Limapon -0.002315 0.005149 -0.449616 cf._Retusa vs Thuridi 0.005390 0.005168 1.042965 cf._Retusa vs Elysia_ 0.004777 0.005023 0.950956 cf._Retusa vs Elysiel 0.005359 0.004952 1.082155 cf._Retusa vs Boselli 0.005367 0.005017 1.069758 cf._Retusa vs Cyerce_ 0.009466 0.004904 1.930191 cf._Retusa vs Placobr -0.000520 0.005368 -0.096902 cf._Retusa vs Placida 0.006549 0.004951 1.322860 cf._Retusa vs Polybra 0.005366 0.005021 1.068641 Bulla_cf._ vs Smaragd -0.002964 0.002555 -1.159830 Bulla_cf._ vs Phanero -0.004699 0.002607 -1.802291 Bulla_cf._ vs Bullact -0.001758 0.002406 -0.730600 Bulla_cf._ vs Atys_se -0.003544 0.002549 -1.390178 Bulla_cf._ vs Odontog -0.004122 0.003047 -1.352840 Bulla_cf._ vs Navanax -0.005335 0.002882 -1.851223 Bulla_cf._ vs Chelido -0.003552 0.002812 -1.263220 Bulla_cf._ vs Philino -0.004752 0.002881 -1.649642 Bulla_cf._ vs Philino -0.006494 0.003319 -1.956708 Bulla_cf._ vs Sagamin -0.007693 0.003161 -2.433768 Bulla_cf._ vs Toledon 0.001172 0.002982 0.393091 Bulla_cf._ vs Diaphan -0.005351 0.002888 -1.853020 Bulla_cf._ vs Newnesi -0.001781 0.003320 -0.536408 Bulla_cf._ vs Colpoda -0.014216 0.003590 -3.959676 Bulla_cf._ vs Runcina -0.007657 0.004283 -1.787975 Bulla_cf._ vs Unela_g 0.005263 0.003962 1.328479 Bulla_cf._ vs Pontohe -0.003542 0.004240 -0.835318 Bulla_cf._ vs Hedylop 0.007583 0.003908 1.940364 Bulla_cf._ vs Parahed 0.005260 0.003959 1.328478 Bulla_cf._ vs Umbracu 0.000611 0.004214 0.144934 Bulla_cf._ vs Tylodin 0.000607 0.004333 0.140024 Bulla_cf._ vs Akera_b 0.003529 0.003737 0.944388 Bulla_cf._ vs Aplysia 0.004111 0.003644 1.127969 Bulla_cf._ vs Dolabel 0.004115 0.003739 1.100717 Bulla_cf._ vs Siphono 0.003536 0.003693 0.957385 Bulla_cf._ vs Bursate 0.005294 0.003647 1.451811 Bulla_cf._ vs Petalif 0.005860 0.003643 1.608416 Bulla_cf._ vs Dolabri 0.005283 0.003692 1.430878 Bulla_cf._ vs Styloch 0.004108 0.003741 1.098152 Bulla_cf._ vs Clio_py -0.015449 0.004663 -3.312841 Bulla_cf._ vs Diacria -0.018415 0.004850 -3.796785 Bulla_cf._ vs Creseis -0.033796 0.006034 -5.600923 Bulla_cf._ vs Spongio -0.002920 0.004228 -0.690670 Bulla_cf._ vs Calliop -0.014788 0.005317 -2.781283 Bulla_cf._ vs Limapon -0.008832 0.004851 -1.820733 Bulla_cf._ vs Thuridi -0.001137 0.004723 -0.240757 Bulla_cf._ vs Elysia_ -0.001735 0.004717 -0.367845 Bulla_cf._ vs Elysiel -0.001149 0.004604 -0.249481 Bulla_cf._ vs Boselli -0.001141 0.004674 -0.244031 Bulla_cf._ vs Cyerce_ 0.002958 0.004477 0.660809 Bulla_cf._ vs Placobr -0.007030 0.004942 -1.422634 Bulla_cf._ vs Placida 0.000622 0.004565 0.136172 Bulla_cf._ vs Polybra -0.001145 0.004678 -0.244858 Smaragdine vs Phanero -0.001747 0.002423 -0.720968 Smaragdine vs Bullact 0.001202 0.002041 0.588819 Smaragdine vs Atys_se -0.000587 0.001869 -0.313943 Smaragdine vs Odontog -0.001167 0.003064 -0.380781 Smaragdine vs Navanax -0.002384 0.003070 -0.776457 Smaragdine vs Chelido -0.000595 0.003062 -0.194365 Smaragdine vs Philino -0.001797 0.003069 -0.585680 Smaragdine vs Philino -0.002957 0.003430 -0.862112 Smaragdine vs Sagamin -0.004748 0.003227 -1.471359 Smaragdine vs Toledon 0.004141 0.003167 1.307460 Smaragdine vs Diaphan -0.001795 0.003187 -0.563252 Smaragdine vs Newnesi 0.001181 0.003490 0.338311 Smaragdine vs Colpoda -0.010697 0.003880 -2.757310 Smaragdine vs Runcina -0.005300 0.004537 -1.168194 Smaragdine vs Unela_g 0.008242 0.004025 2.047358 Smaragdine vs Pontohe -0.001175 0.004380 -0.268364 Smaragdine vs Hedylop 0.010565 0.003972 2.659943 Smaragdine vs Parahed 0.008237 0.004023 2.047355

206
Smaragdine vs Umbracu 0.004164 0.004435 0.938857 Smaragdine vs Tylodin 0.003565 0.004549 0.783703 Smaragdine vs Akera_b 0.006497 0.003981 1.632046 Smaragdine vs Aplysia 0.007080 0.003893 1.818587 Smaragdine vs Dolabel 0.007084 0.003894 1.819156 Smaragdine vs Siphono 0.006513 0.003895 1.672130 Smaragdine vs Bursate 0.008279 0.003806 2.175346 Smaragdine vs Petalif 0.008843 0.003713 2.381933 Smaragdine vs Dolabri 0.008266 0.003761 2.198107 Smaragdine vs Styloch 0.007087 0.003897 1.818539 Smaragdine vs Clio_py -0.012523 0.004720 -2.653385 Smaragdine vs Diacria -0.015498 0.004978 -3.113171 Smaragdine vs Creseis -0.030907 0.005968 -5.179118 Smaragdine vs Spongio 0.000034 0.004329 0.007873 Smaragdine vs Calliop -0.010675 0.005371 -1.987492 Smaragdine vs Limapon -0.005298 0.005015 -1.056477 Smaragdine vs Thuridi 0.002421 0.004927 0.491379 Smaragdine vs Elysia_ 0.001813 0.004885 0.371159 Smaragdine vs Elysiel 0.002399 0.004775 0.502302 Smaragdine vs Boselli 0.002407 0.004807 0.500734 Smaragdine vs Cyerce_ 0.006515 0.004651 1.400760 Smaragdine vs Placobr -0.003497 0.005172 -0.676089 Smaragdine vs Placida 0.002994 0.004659 0.642661 Smaragdine vs Polybra 0.002403 0.004774 0.503440 Phaneropht vs Bullact 0.002941 0.002346 1.254051 Phaneropht vs Atys_se 0.001158 0.002113 0.548072 Phaneropht vs Odontog 0.000580 0.003164 0.183235 Phaneropht vs Navanax -0.000636 0.003275 -0.194065 Phaneropht vs Chelido 0.001150 0.003111 0.369576 Phaneropht vs Philino -0.000050 0.003117 -0.015891 Phaneropht vs Philino -0.001206 0.003619 -0.333175 Phaneropht vs Sagamin -0.002994 0.003574 -0.837755 Phaneropht vs Toledon 0.005875 0.003264 1.800028 Phaneropht vs Diaphan -0.000040 0.003230 -0.012486 Phaneropht vs Newnesi 0.002921 0.003375 0.865543 Phaneropht vs Colpoda -0.008926 0.004047 -2.205879 Phaneropht vs Runcina -0.002359 0.004451 -0.529910 Phaneropht vs Unela_g 0.010552 0.004141 2.548051 Phaneropht vs Pontohe 0.001756 0.004332 0.405232 Phaneropht vs Hedylop 0.012868 0.004215 3.053180 Phaneropht vs Parahed 0.010546 0.004139 2.548050 Phaneropht vs Umbracu 0.005900 0.004502 1.310579 Phaneropht vs Tylodin 0.005308 0.004575 1.160026 Phaneropht vs Akera_b 0.008234 0.004015 2.050690 Phaneropht vs Aplysia 0.008815 0.003928 2.244007 Phaneropht vs Dolabel 0.009406 0.003973 2.367337 Phaneropht vs Siphono 0.008243 0.003928 2.098537 Phaneropht vs Bursate 0.010007 0.003797 2.635365 Phaneropht vs Petalif 0.010567 0.003837 2.753959 Phaneropht vs Dolabri 0.009993 0.003841 2.601630 Phaneropht vs Styloch 0.008815 0.003930 2.243016 Phaneropht vs Clio_py -0.010153 0.004854 -2.091575 Phaneropht vs Diacria -0.013709 0.005105 -2.685064 Phaneropht vs Creseis -0.028485 0.006075 -4.689275 Phaneropht vs Spongio 0.001199 0.004276 0.280410 Phaneropht vs Calliop -0.009500 0.005521 -1.720530 Phaneropht vs Limapon -0.003538 0.005107 -0.692839 Phaneropht vs Thuridi 0.004163 0.004985 0.835038 Phaneropht vs Elysia_ 0.003552 0.005014 0.708538 Phaneropht vs Elysiel 0.004135 0.004907 0.842632 Phaneropht vs Boselli 0.004144 0.004902 0.845284 Phaneropht vs Cyerce_ 0.008243 0.004713 1.748829 Phaneropht vs Placobr -0.001744 0.005226 -0.333653 Phaneropht vs Placida 0.005325 0.004760 1.118659 Phaneropht vs Polybra 0.004142 0.004906 0.844304 Bullacta_e vs Atys_se -0.001785 0.001856 -0.961826 Bullacta_e vs Odontog -0.002363 0.002815 -0.839679 Bullacta_e vs Navanax -0.003577 0.002943 -1.215402 Bullacta_e vs Chelido -0.001793 0.003052 -0.587679 Bullacta_e vs Philino -0.002993 0.002821 -1.061026 Bullacta_e vs Philino -0.004151 0.003521 -1.178891 Bullacta_e vs Sagamin -0.005936 0.003321 -1.787452 Bullacta_e vs Toledon 0.002932 0.002618 1.119639 Bullacta_e vs Diaphan -0.002996 0.002890 -1.036413 Bullacta_e vs Newnesi -0.000022 0.002996 -0.007272 Bullacta_e vs Colpoda -0.011873 0.003586 -3.310661 Bullacta_e vs Runcina -0.005896 0.004074 -1.447146 Bullacta_e vs Unela_g 0.007022 0.003877 1.811069

207
Bullacta_e vs Pontohe -0.001780 0.004120 -0.432181 Bullacta_e vs Hedylop 0.009341 0.003956 2.361440 Bullacta_e vs Parahed 0.007018 0.003875 1.811068 Bullacta_e vs Umbracu 0.002954 0.004217 0.700480 Bullacta_e vs Tylodin 0.002365 0.004336 0.545359 Bullacta_e vs Akera_b 0.005289 0.003455 1.530755 Bullacta_e vs Aplysia 0.005871 0.003456 1.698703 Bullacta_e vs Dolabel 0.005875 0.003555 1.652323 Bullacta_e vs Siphono 0.005297 0.003507 1.510265 Bullacta_e vs Bursate 0.007057 0.003508 2.011585 Bullacta_e vs Petalif 0.007621 0.003600 2.116875 Bullacta_e vs Dolabri 0.007045 0.003556 1.981273 Bullacta_e vs Styloch 0.005869 0.003604 1.628484 Bullacta_e vs Clio_py -0.013688 0.004510 -3.034921 Bullacta_e vs Diacria -0.016654 0.004743 -3.511230 Bullacta_e vs Creseis -0.031443 0.005825 -5.397998 Bullacta_e vs Spongio -0.001159 0.004190 -0.276735 Bullacta_e vs Calliop -0.012445 0.005255 -2.368165 Bullacta_e vs Limapon -0.006486 0.004709 -1.377436 Bullacta_e vs Thuridi 0.001212 0.004765 0.254254 Bullacta_e vs Elysia_ 0.000608 0.004649 0.130799 Bullacta_e vs Elysiel 0.001193 0.004457 0.267574 Bullacta_e vs Boselli 0.001201 0.004568 0.262970 Bullacta_e vs Cyerce_ 0.005300 0.004483 1.182255 Bullacta_e vs Placobr -0.004689 0.004947 -0.947830 Bullacta_e vs Placida 0.001789 0.004414 0.405369 Bullacta_e vs Polybra 0.001197 0.004609 0.259749 Atys_semis vs Odontog -0.000579 0.003169 -0.182592 Atys_semis vs Navanax -0.001794 0.003063 -0.585715 Atys_semis vs Chelido -0.000008 0.003168 -0.002628 Atys_semis vs Philino -0.001208 0.003062 -0.394646 Atys_semis vs Philino -0.002366 0.003474 -0.680985 Atys_semis vs Sagamin -0.004154 0.003327 -1.248428 Atys_semis vs Toledon 0.004719 0.003049 1.548031 Atys_semis vs Diaphan -0.001204 0.002949 -0.408265 Atys_semis vs Newnesi 0.001764 0.003275 0.538657 Atys_semis vs Colpoda -0.010092 0.003731 -2.704961 Atys_semis vs Runcina -0.004109 0.004411 -0.931436 Atys_semis vs Unela_g 0.008813 0.003931 2.241755 Atys_semis vs Pontohe 0.000008 0.004414 0.001815 Atys_semis vs Hedylop 0.011132 0.004008 2.777140 Atys_semis vs Parahed 0.008808 0.003929 2.241752 Atys_semis vs Umbracu 0.004743 0.004348 1.090816 Atys_semis vs Tylodin 0.004145 0.004541 0.912661 Atys_semis vs Akera_b 0.007072 0.003886 1.819968 Atys_semis vs Aplysia 0.007653 0.003796 2.016265 Atys_semis vs Dolabel 0.007657 0.003887 1.969912 Atys_semis vs Siphono 0.007088 0.003798 1.866333 Atys_semis vs Bursate 0.008256 0.003751 2.201184 Atys_semis vs Petalif 0.009414 0.003706 2.540236 Atys_semis vs Dolabri 0.008838 0.003754 2.354459 Atys_semis vs Styloch 0.007661 0.003890 1.969329 Atys_semis vs Clio_py -0.011914 0.004634 -2.571195 Atys_semis vs Diacria -0.014884 0.005005 -2.973950 Atys_semis vs Creseis -0.029665 0.006019 -4.928307 Atys_semis vs Spongio 0.000620 0.004239 0.146324 Atys_semis vs Calliop -0.010665 0.005395 -1.976635 Atys_semis vs Limapon -0.004699 0.005003 -0.939236 Atys_semis vs Thuridi 0.003004 0.004954 0.606360 Atys_semis vs Elysia_ 0.002396 0.004912 0.487720 Atys_semis vs Elysiel 0.002980 0.004803 0.620344 Atys_semis vs Boselli 0.002988 0.004835 0.618073 Atys_semis vs Cyerce_ 0.007089 0.004680 1.514638 Atys_semis vs Placobr -0.002904 0.005129 -0.566286 Atys_semis vs Placida 0.003575 0.004689 0.762435 Atys_semis vs Polybra 0.002985 0.004802 0.621637 Odontoglaj vs Navanax -0.001216 0.002042 -0.595280 Odontoglaj vs Chelido 0.000570 0.002497 0.228436 Odontoglaj vs Philino -0.000034 0.002502 -0.013468 Odontoglaj vs Philino -0.001787 0.003482 -0.513150 Odontoglaj vs Sagamin -0.003576 0.003432 -1.041818 Odontoglaj vs Toledon 0.005298 0.003215 1.647937 Odontoglaj vs Diaphan -0.000623 0.003396 -0.183376 Odontoglaj vs Newnesi 0.002343 0.003581 0.654246 Odontoglaj vs Colpoda -0.010107 0.004141 -2.440972 Odontoglaj vs Runcina -0.004125 0.004374 -0.942971 Odontoglaj vs Unela_g 0.009391 0.004103 2.288591 Odontoglaj vs Pontohe -0.000007 0.004251 -0.001764

208
Odontoglaj vs Hedylop 0.011710 0.004051 2.890248 Odontoglaj vs Parahed 0.009386 0.004101 2.288589 Odontoglaj vs Umbracu 0.005322 0.004429 1.201616 Odontoglaj vs Tylodin 0.004137 0.004464 0.926849 Odontoglaj vs Akera_b 0.007658 0.004060 1.886143 Odontoglaj vs Aplysia 0.008240 0.003974 2.073309 Odontoglaj vs Dolabel 0.008244 0.004062 2.029696 Odontoglaj vs Siphono 0.007668 0.004063 1.887355 Odontoglaj vs Bursate 0.008836 0.003797 2.327240 Odontoglaj vs Petalif 0.009398 0.003794 2.477428 Odontoglaj vs Dolabri 0.008823 0.003841 2.297224 Odontoglaj vs Styloch 0.007645 0.003888 1.966530 Odontoglaj vs Clio_py -0.011327 0.004823 -2.348642 Odontoglaj vs Diacria -0.014297 0.004895 -2.920469 Odontoglaj vs Creseis -0.029085 0.005983 -4.861118 Odontoglaj vs Spongio 0.001207 0.004321 0.279367 Odontoglaj vs Calliop -0.010086 0.005494 -1.835827 Odontoglaj vs Limapon -0.004122 0.005041 -0.817571 Odontoglaj vs Thuridi 0.002988 0.004699 0.635948 Odontoglaj vs Elysia_ 0.002974 0.004769 0.623665 Odontoglaj vs Elysiel 0.003558 0.004730 0.752166 Odontoglaj vs Boselli 0.004151 0.004688 0.885542 Odontoglaj vs Cyerce_ 0.007668 0.004680 1.638265 Odontoglaj vs Placobr -0.001735 0.004989 -0.347870 Odontoglaj vs Placida 0.004748 0.004729 1.003999 Odontoglaj vs Polybra 0.003564 0.004874 0.731200 Navanax_in vs Chelido 0.001786 0.002123 0.841150 Navanax_in vs Philino 0.000586 0.001956 0.299771 Navanax_in vs Philino -0.000569 0.003487 -0.163276 Navanax_in vs Sagamin -0.002359 0.003336 -0.706983 Navanax_in vs Toledon 0.006511 0.003385 1.923626 Navanax_in vs Diaphan 0.000598 0.003134 0.190960 Navanax_in vs Newnesi 0.003557 0.003640 0.977181 Navanax_in vs Colpoda -0.008886 0.003927 -2.263104 Navanax_in vs Runcina -0.002905 0.004420 -0.657296 Navanax_in vs Unela_g 0.010602 0.003986 2.659853 Navanax_in vs Pontohe 0.001208 0.004174 0.289499 Navanax_in vs Hedylop 0.012919 0.004062 3.180294 Navanax_in vs Parahed 0.010595 0.003983 2.659850 Navanax_in vs Umbracu 0.006535 0.004319 1.513122 Navanax_in vs Tylodin 0.005348 0.004314 1.239795 Navanax_in vs Akera_b 0.008871 0.004028 2.202462 Navanax_in vs Aplysia 0.009452 0.003941 2.398441 Navanax_in vs Dolabel 0.009457 0.004030 2.346734 Navanax_in vs Siphono 0.008880 0.004030 2.203449 Navanax_in vs Bursate 0.010048 0.003761 2.671444 Navanax_in vs Petalif 0.010608 0.003758 2.822846 Navanax_in vs Dolabri 0.010034 0.003805 2.636756 Navanax_in vs Styloch 0.008856 0.003853 2.298453 Navanax_in vs Clio_py -0.010101 0.004793 -2.107595 Navanax_in vs Diacria -0.013070 0.004865 -2.686523 Navanax_in vs Creseis -0.027843 0.006078 -4.580661 Navanax_in vs Spongio 0.002425 0.004290 0.565233 Navanax_in vs Calliop -0.008863 0.005435 -1.630654 Navanax_in vs Limapon -0.002903 0.005048 -0.575039 Navanax_in vs Thuridi 0.004204 0.004781 0.879258 Navanax_in vs Elysia_ 0.004190 0.004778 0.876853 Navanax_in vs Elysiel 0.004772 0.004739 1.007047 Navanax_in vs Boselli 0.004781 0.004807 0.994528 Navanax_in vs Cyerce_ 0.008879 0.004763 1.864170 Navanax_in vs Placobr -0.001107 0.005170 -0.214195 Navanax_in vs Placida 0.005365 0.004773 1.124225 Navanax_in vs Polybra 0.004778 0.004883 0.978617 Chelidonur vs Philino -0.001200 0.002050 -0.585289 Chelidonur vs Philino -0.002952 0.003432 -0.860135 Chelidonur vs Sagamin -0.004146 0.003540 -1.171213 Chelidonur vs Toledon 0.004728 0.003528 1.340222 Chelidonur vs Diaphan -0.001195 0.003293 -0.363063 Chelidonur vs Newnesi 0.001773 0.003866 0.458502 Chelidonur vs Colpoda -0.011272 0.003966 -2.842292 Chelidonur vs Runcina -0.004704 0.004457 -1.055511 Chelidonur vs Unela_g 0.009407 0.004023 2.338238 Chelidonur vs Pontohe 0.000008 0.004172 0.001920 Chelidonur vs Hedylop 0.011726 0.003970 2.953610 Chelidonur vs Parahed 0.009402 0.004021 2.338236 Chelidonur vs Umbracu 0.004751 0.004352 1.091650 Chelidonur vs Tylodin 0.003567 0.004427 0.805754 Chelidonur vs Akera_b 0.007088 0.003893 1.820803

209
Chelidonur vs Aplysia 0.007670 0.003979 1.927446 Chelidonur vs Dolabel 0.008261 0.004024 2.052921 Chelidonur vs Siphono 0.007096 0.004108 1.727561 Chelidonur vs Bursate 0.008265 0.003709 2.228188 Chelidonur vs Petalif 0.008827 0.003798 2.324009 Chelidonur vs Dolabri 0.008252 0.003845 2.145869 Chelidonur vs Styloch 0.007074 0.003893 1.817390 Chelidonur vs Clio_py -0.011304 0.004790 -2.359900 Chelidonur vs Diacria -0.014867 0.004897 -3.035618 Chelidonur vs Creseis -0.030254 0.006048 -5.002059 Chelidonur vs Spongio 0.000637 0.004369 0.145852 Chelidonur vs Calliop -0.011251 0.005430 -2.071868 Chelidonur vs Limapon -0.005289 0.005043 -1.048715 Chelidonur vs Thuridi 0.001821 0.004661 0.390729 Chelidonur vs Elysia_ 0.001810 0.004767 0.379622 Chelidonur vs Elysiel 0.002394 0.004693 0.510195 Chelidonur vs Boselli 0.002402 0.004725 0.508484 Chelidonur vs Cyerce_ 0.006503 0.004605 1.412130 Chelidonur vs Placobr -0.003490 0.005096 -0.684993 Chelidonur vs Placida 0.004177 0.004839 0.863191 Chelidonur vs Polybra 0.002399 0.004802 0.499590 Philinopsi vs Philino -0.001157 0.003387 -0.341488 Philinopsi vs Sagamin -0.002946 0.003340 -0.882207 Philinopsi vs Toledon 0.005928 0.003486 1.700613 Philinopsi vs Diaphan 0.000010 0.003189 0.003018 Philinopsi vs Newnesi 0.002973 0.003688 0.806046 Philinopsi vs Colpoda -0.009479 0.003835 -2.471878 Philinopsi vs Runcina -0.002907 0.004344 -0.669145 Philinopsi vs Unela_g 0.010608 0.004074 2.603544 Philinopsi vs Pontohe 0.001807 0.004182 0.431997 Philinopsi vs Hedylop 0.012926 0.004149 3.115547 Philinopsi vs Parahed 0.010602 0.004072 2.603541 Philinopsi vs Umbracu 0.005952 0.004439 1.340735 Philinopsi vs Tylodin 0.004765 0.004434 1.074708 Philinopsi vs Akera_b 0.008289 0.003898 2.126278 Philinopsi vs Aplysia 0.008871 0.003899 2.275088 Philinopsi vs Dolabel 0.009462 0.003945 2.398670 Philinopsi vs Siphono 0.008298 0.004030 2.058890 Philinopsi vs Bursate 0.009466 0.003717 2.546671 Philinopsi vs Petalif 0.010027 0.003806 2.634416 Philinopsi vs Dolabri 0.009452 0.003853 2.453102 Philinopsi vs Styloch 0.008274 0.003810 2.171652 Philinopsi vs Clio_py -0.010107 0.004681 -2.158984 Philinopsi vs Diacria -0.013665 0.004866 -2.808325 Philinopsi vs Creseis -0.028450 0.006021 -4.724932 Philinopsi vs Spongio 0.001839 0.004333 0.424380 Philinopsi vs Calliop -0.009456 0.005303 -1.783037 Philinopsi vs Limapon -0.003492 0.004906 -0.711827 Philinopsi vs Thuridi 0.003619 0.004925 0.734698 Philinopsi vs Elysia_ 0.003605 0.004922 0.732533 Philinopsi vs Elysiel 0.004189 0.004740 0.883796 Philinopsi vs Boselli 0.004197 0.004844 0.866423 Philinopsi vs Cyerce_ 0.008298 0.004690 1.769150 Philinopsi vs Placobr -0.001695 0.005206 -0.325651 Philinopsi vs Placida 0.004782 0.004698 1.017768 Philinopsi vs Polybra 0.004194 0.004811 0.871758 Philinoglo vs Sagamin -0.001792 0.003863 -0.463910 Philinoglo vs Toledon 0.007088 0.003853 1.839661 Philinoglo vs Diaphan 0.001770 0.003782 0.467837 Philinoglo vs Newnesi 0.004131 0.003994 1.034369 Philinoglo vs Colpoda -0.007727 0.004007 -1.928547 Philinoglo vs Runcina -0.002337 0.004908 -0.476153 Philinoglo vs Unela_g 0.011763 0.004149 2.834957 Philinoglo vs Pontohe 0.003559 0.004335 0.821088 Philinoglo vs Hedylop 0.014663 0.004019 3.648776 Philinoglo vs Parahed 0.011756 0.004147 2.834954 Philinoglo vs Umbracu 0.007697 0.004473 1.720717 Philinoglo vs Tylodin 0.007096 0.004509 1.573854 Philinoglo vs Akera_b 0.009445 0.004391 2.151105 Philinoglo vs Aplysia 0.010027 0.004270 2.348017 Philinoglo vs Dolabel 0.010031 0.004272 2.348251 Philinoglo vs Siphono 0.009455 0.004234 2.233266 Philinoglo vs Bursate 0.010624 0.004106 2.587695 Philinoglo vs Petalif 0.011184 0.004144 2.699031 Philinoglo vs Dolabri 0.010610 0.004188 2.533633 Philinoglo vs Styloch 0.009431 0.004272 2.207698 Philinoglo vs Clio_py -0.008948 0.004938 -1.812056 Philinoglo vs Diacria -0.012516 0.005078 -2.464736

210
Philinoglo vs Creseis -0.026682 0.006076 -4.391743 Philinoglo vs Spongio 0.002995 0.004635 0.646148 Philinoglo vs Calliop -0.008294 0.005365 -1.546011 Philinoglo vs Limapon -0.002332 0.005044 -0.462379 Philinoglo vs Thuridi 0.005962 0.004854 1.228341 Philinoglo vs Elysia_ 0.005347 0.004883 1.094840 Philinoglo vs Elysiel 0.005928 0.004774 1.241680 Philinoglo vs Boselli 0.005937 0.004771 1.244499 Philinoglo vs Cyerce_ 0.010036 0.004651 2.157861 Philinoglo vs Placobr 0.000051 0.005169 0.009924 Philinoglo vs Placida 0.005939 0.004805 1.235990 Philinoglo vs Polybra 0.005348 0.004806 1.112656 Sagaminopt vs Toledon 0.008871 0.003858 2.299155 Sagaminopt vs Diaphan 0.002968 0.003693 0.803838 Sagaminopt vs Newnesi 0.005917 0.004129 1.433200 Sagaminopt vs Colpoda -0.006525 0.003785 -1.723956 Sagaminopt vs Runcina 0.000647 0.004544 0.142273 Sagaminopt vs Unela_g 0.013550 0.004439 3.052719 Sagaminopt vs Pontohe 0.004759 0.004656 1.022168 Sagaminopt vs Hedylop 0.015865 0.004392 3.612422 Sagaminopt vs Parahed 0.013542 0.004436 3.052715 Sagaminopt vs Umbracu 0.008897 0.004668 1.905760 Sagaminopt vs Tylodin 0.008294 0.004627 1.792443 Sagaminopt vs Akera_b 0.011222 0.004158 2.699182 Sagaminopt vs Aplysia 0.011804 0.004114 2.869300 Sagaminopt vs Dolabel 0.012396 0.004160 2.980183 Sagaminopt vs Siphono 0.011243 0.004280 2.626655 Sagaminopt vs Bursate 0.013010 0.004120 3.157450 Sagaminopt vs Petalif 0.013567 0.004074 3.329904 Sagaminopt vs Dolabri 0.012994 0.004119 3.155049 Sagaminopt vs Styloch 0.011815 0.004243 2.784259 Sagaminopt vs Clio_py -0.007757 0.004902 -1.582481 Sagaminopt vs Diacria -0.010718 0.005252 -2.040573 Sagaminopt vs Creseis -0.025472 0.006142 -4.147214 Sagaminopt vs Spongio 0.004777 0.004644 1.028624 Sagaminopt vs Calliop -0.006502 0.005660 -1.148750 Sagaminopt vs Limapon -0.000539 0.005186 -0.103972 Sagaminopt vs Thuridi 0.007168 0.005240 1.368063 Sagaminopt vs Elysia_ 0.006551 0.005061 1.294285 Sagaminopt vs Elysiel 0.007131 0.005061 1.409197 Sagaminopt vs Boselli 0.007140 0.005056 1.412342 Sagaminopt vs Cyerce_ 0.011239 0.005048 2.226319 Sagaminopt vs Placobr 0.001254 0.005437 0.230630 Sagaminopt vs Placida 0.007725 0.005022 1.538326 Sagaminopt vs Polybra 0.007140 0.005128 1.392230 Toledonia_ vs Diaphan -0.005943 0.002771 -2.144267 Toledonia_ vs Newnesi -0.002955 0.002118 -1.395354 Toledonia_ vs Colpoda -0.014817 0.004051 -3.657791 Toledonia_ vs Runcina -0.008839 0.003989 -2.215973 Toledonia_ vs Unela_g 0.004094 0.003786 1.081367 Toledonia_ vs Pontohe -0.005309 0.004202 -1.263583 Toledonia_ vs Hedylop 0.006416 0.003684 1.741639 Toledonia_ vs Parahed 0.004091 0.003783 1.081367 Toledonia_ vs Umbracu 0.000610 0.004257 0.143405 Toledonia_ vs Tylodin -0.000566 0.004086 -0.138623 Toledonia_ vs Akera_b 0.002358 0.003502 0.673349 Toledonia_ vs Aplysia 0.002939 0.003194 0.920399 Toledonia_ vs Dolabel 0.002944 0.003403 0.865028 Toledonia_ vs Siphono 0.002363 0.003249 0.727357 Toledonia_ vs Bursate 0.004121 0.003249 1.268378 Toledonia_ vs Petalif 0.004689 0.003191 1.469672 Toledonia_ vs Dolabri 0.004111 0.003194 1.286887 Toledonia_ vs Styloch 0.002936 0.003301 0.889571 Toledonia_ vs Clio_py -0.016633 0.004471 -3.720387 Toledonia_ vs Diacria -0.019600 0.004629 -4.233843 Toledonia_ vs Creseis -0.034409 0.005949 -5.784065 Toledonia_ vs Spongio -0.004096 0.003840 -1.066562 Toledonia_ vs Calliop -0.015387 0.005221 -2.947110 Toledonia_ vs Limapon -0.008837 0.004784 -1.847252 Toledonia_ vs Thuridi -0.001729 0.004689 -0.368734 Toledonia_ vs Elysia_ -0.002326 0.004609 -0.504659 Toledonia_ vs Elysiel -0.001739 0.004494 -0.386945 Toledonia_ vs Boselli -0.001731 0.004566 -0.379131 Toledonia_ vs Cyerce_ 0.002370 0.004402 0.538316 Toledonia_ vs Placobr -0.007626 0.004838 -1.576300 Toledonia_ vs Placida -0.001136 0.004413 -0.257482 Toledonia_ vs Polybra -0.001737 0.004569 -0.380198 Diaphana_c vs Newnesi 0.002975 0.003020 0.985328

211
Diaphana_c vs Colpoda -0.010116 0.003945 -2.564230 Diaphana_c vs Runcina -0.004108 0.004273 -0.961440 Diaphana_c vs Unela_g 0.010053 0.004171 2.410328 Diaphana_c vs Pontohe 0.000615 0.004192 0.146743 Diaphana_c vs Hedylop 0.012381 0.004161 2.975753 Diaphana_c vs Parahed 0.010047 0.004168 2.410325 Diaphana_c vs Umbracu 0.005967 0.004529 1.317445 Diaphana_c vs Tylodin 0.004785 0.004487 1.066419 Diaphana_c vs Akera_b 0.008314 0.004085 2.035308 Diaphana_c vs Aplysia 0.008898 0.003910 2.275849 Diaphana_c vs Dolabel 0.008902 0.004087 2.178375 Diaphana_c vs Siphono 0.008323 0.003956 2.103644 Diaphana_c vs Bursate 0.010686 0.003916 2.728983 Diaphana_c vs Petalif 0.011248 0.003910 2.876934 Diaphana_c vs Dolabri 0.010671 0.003914 2.726525 Diaphana_c vs Styloch 0.009488 0.004002 2.370574 Diaphana_c vs Clio_py -0.010159 0.004848 -2.095722 Diaphana_c vs Diacria -0.013142 0.004996 -2.630423 Diaphana_c vs Creseis -0.028591 0.006138 -4.658112 Diaphana_c vs Spongio 0.002427 0.004386 0.553204 Diaphana_c vs Calliop -0.009514 0.005257 -1.809742 Diaphana_c vs Limapon -0.003525 0.004890 -0.720803 Diaphana_c vs Thuridi 0.004215 0.004947 0.852015 Diaphana_c vs Elysia_ 0.003601 0.004869 0.739659 Diaphana_c vs Elysiel 0.004187 0.004795 0.873137 Diaphana_c vs Boselli 0.004195 0.004863 0.862735 Diaphana_c vs Cyerce_ 0.007724 0.004706 1.641383 Diaphana_c vs Placobr -0.002310 0.005258 -0.439306 Diaphana_c vs Placida 0.004793 0.004717 1.016054 Diaphana_c vs Polybra 0.003604 0.004828 0.746403 Newnesia_a vs Colpoda -0.011858 0.004142 -2.862573 Newnesia_a vs Runcina -0.005877 0.003906 -1.504552 Newnesia_a vs Unela_g 0.007049 0.003705 1.902491 Newnesia_a vs Pontohe -0.002353 0.004211 -0.558632 Newnesia_a vs Hedylop 0.009369 0.003787 2.473986 Newnesia_a vs Parahed 0.007045 0.003703 1.902489 Newnesia_a vs Umbracu 0.003571 0.004267 0.836825 Newnesia_a vs Tylodin 0.002389 0.004262 0.560412 Newnesia_a vs Akera_b 0.005315 0.003609 1.472737 Newnesia_a vs Aplysia 0.005896 0.003412 1.728037 Newnesia_a vs Dolabel 0.005901 0.003414 1.728584 Newnesia_a vs Siphono 0.005322 0.003414 1.558766 Newnesia_a vs Bursate 0.007084 0.003364 2.105701 Newnesia_a vs Petalif 0.007648 0.003306 2.313131 Newnesia_a vs Dolabri 0.007071 0.003310 2.136179 Newnesia_a vs Styloch 0.005895 0.003310 1.781037 Newnesia_a vs Clio_py -0.013672 0.004476 -3.054697 Newnesia_a vs Diacria -0.016641 0.004787 -3.476455 Newnesia_a vs Creseis -0.031438 0.005989 -5.249725 Newnesia_a vs Spongio -0.001137 0.003848 -0.295528 Newnesia_a vs Calliop -0.012430 0.004986 -2.493121 Newnesia_a vs Limapon -0.005875 0.004793 -1.225758 Newnesia_a vs Thuridi 0.001234 0.004771 0.258645 Newnesia_a vs Elysia_ 0.000631 0.004657 0.135431 Newnesia_a vs Elysiel 0.001216 0.004543 0.267673 Newnesia_a vs Boselli 0.001224 0.004689 0.261084 Newnesia_a vs Cyerce_ 0.005325 0.004491 1.185569 Newnesia_a vs Placobr -0.004667 0.004987 -0.935729 Newnesia_a vs Placida 0.001811 0.004343 0.416981 Newnesia_a vs Polybra 0.001220 0.004617 0.264152 Colpodaspi vs Runcina 0.005399 0.004927 1.095803 Colpodaspi vs Unela_g 0.018913 0.004686 4.035909 Colpodaspi vs Pontohe 0.011306 0.004891 2.311690 Colpodaspi vs Hedylop 0.021811 0.004787 4.555923 Colpodaspi vs Parahed 0.018902 0.004683 4.035908 Colpodaspi vs Umbracu 0.014842 0.004940 3.004633 Colpodaspi vs Tylodin 0.014827 0.005075 2.921725 Colpodaspi vs Akera_b 0.016589 0.004534 3.658571 Colpodaspi vs Aplysia 0.017172 0.004535 3.786582 Colpodaspi vs Dolabel 0.017174 0.004460 3.850913 Colpodaspi vs Siphono 0.016605 0.004539 3.658266 Colpodaspi vs Bursate 0.018362 0.004458 4.119060 Colpodaspi vs Petalif 0.018932 0.004534 4.175952 Colpodaspi vs Dolabri 0.018361 0.004538 4.046334 Colpodaspi vs Styloch 0.017177 0.004576 3.753982 Colpodaspi vs Clio_py -0.001815 0.005233 -0.346784 Colpodaspi vs Diacria -0.004186 0.005338 -0.784239 Colpodaspi vs Creseis -0.018897 0.006327 -2.986851

212
Colpodaspi vs Spongio 0.010134 0.005024 2.017101 Colpodaspi vs Calliop -0.000571 0.005708 -0.100083 Colpodaspi vs Limapon 0.005399 0.005369 1.005518 Colpodaspi vs Thuridi 0.013710 0.005392 2.542398 Colpodaspi vs Elysia_ 0.012487 0.005319 2.347456 Colpodaspi vs Elysiel 0.013064 0.005149 2.537092 Colpodaspi vs Boselli 0.013074 0.005280 2.476012 Colpodaspi vs Cyerce_ 0.017179 0.005172 3.321442 Colpodaspi vs Placobr 0.007186 0.005554 1.293870 Colpodaspi vs Placida 0.013074 0.005179 2.524618 Colpodaspi vs Polybra 0.012484 0.005315 2.348861 Runcina_ad vs Unela_g 0.014109 0.003940 3.581046 Runcina_ad vs Pontohe 0.005900 0.004176 1.412592 Runcina_ad vs Hedylop 0.016427 0.003889 4.224438 Runcina_ad vs Parahed 0.014100 0.003938 3.581045 Runcina_ad vs Umbracu 0.009467 0.004363 2.169669 Runcina_ad vs Tylodin 0.008266 0.004315 1.915525 Runcina_ad vs Akera_b 0.010616 0.003372 3.148227 Runcina_ad vs Aplysia 0.011199 0.003569 3.137855 Runcina_ad vs Dolabel 0.011787 0.003617 3.258543 Runcina_ad vs Siphono 0.010621 0.003618 2.935529 Runcina_ad vs Bursate 0.012985 0.003526 3.682874 Runcina_ad vs Petalif 0.012958 0.003758 3.448036 Runcina_ad vs Dolabri 0.011785 0.003760 3.134643 Runcina_ad vs Styloch 0.011201 0.003761 2.978079 Runcina_ad vs Clio_py -0.006627 0.004758 -1.392865 Runcina_ad vs Diacria -0.010778 0.004941 -2.181407 Runcina_ad vs Creseis -0.026173 0.005965 -4.387615 Runcina_ad vs Spongio 0.004152 0.004123 1.006945 Runcina_ad vs Calliop -0.005968 0.005239 -1.139011 Runcina_ad vs Limapon 0.000603 0.004691 0.128583 Runcina_ad vs Thuridi 0.008312 0.004756 1.747527 Runcina_ad vs Elysia_ 0.007099 0.004677 1.517765 Runcina_ad vs Elysiel 0.007679 0.004485 1.712173 Runcina_ad vs Boselli 0.007690 0.004519 1.701835 Runcina_ad vs Cyerce_ 0.011799 0.004512 2.615020 Runcina_ad vs Placobr 0.001795 0.004867 0.368816 Runcina_ad vs Placida 0.007691 0.004553 1.689292 Runcina_ad vs Polybra 0.007101 0.004747 1.495796 Unela_glan vs Pontohe -0.009406 0.003105 -3.029643 Unela_glan vs Hedylop 0.002325 0.002392 0.972103 Unela_glan vs Parahed 0.000000 0.000000 Not deter. Unela_glan vs Umbracu -0.004073 0.003248 -1.254296 Unela_glan vs Tylodin -0.005243 0.003450 -1.519661 Unela_glan vs Akera_b -0.002899 0.003138 -0.924047 Unela_glan vs Aplysia -0.002317 0.003138 -0.738477 Unela_glan vs Dolabel -0.001731 0.002969 -0.583142 Unela_glan vs Siphono -0.002315 0.003192 -0.725426 Unela_glan vs Bursate -0.002316 0.002968 -0.780128 Unela_glan vs Petalif -0.001155 0.002968 -0.389270 Unela_glan vs Dolabri -0.001737 0.003026 -0.574120 Unela_glan vs Styloch -0.002909 0.003030 -0.960267 Unela_glan vs Clio_py -0.020732 0.004395 -4.716855 Unela_glan vs Diacria -0.023694 0.004777 -4.959902 Unela_glan vs Creseis -0.039129 0.005667 -6.904581 Unela_glan vs Spongio -0.009359 0.003608 -2.594334 Unela_glan vs Calliop -0.020060 0.004829 -4.154005 Unela_glan vs Limapon -0.013519 0.004194 -3.223306 Unela_glan vs Thuridi -0.005833 0.004091 -1.425678 Unela_glan vs Elysia_ -0.006421 0.003959 -1.621843 Unela_glan vs Elysiel -0.005251 0.003870 -1.356704 Unela_glan vs Boselli -0.005823 0.003865 -1.506437 Unela_glan vs Cyerce_ -0.001723 0.003675 -0.468811 Unela_glan vs Placobr -0.011720 0.004301 -2.724905 Unela_glan vs Placida -0.005813 0.003949 -1.471876 Unela_glan vs Polybra -0.006412 0.003867 -1.658165 Pontohedyl vs Hedylop 0.011727 0.003262 3.595392 Pontohedyl vs Parahed 0.009401 0.003103 3.029640 Pontohedyl vs Umbracu 0.004159 0.003939 1.055889 Pontohedyl vs Tylodin 0.002973 0.004067 0.730923 Pontohedyl vs Akera_b 0.005914 0.003622 1.632816 Pontohedyl vs Aplysia 0.006497 0.003670 1.770192 Pontohedyl vs Dolabel 0.007084 0.003524 2.010124 Pontohedyl vs Siphono 0.006505 0.003670 1.772588 Pontohedyl vs Bursate 0.007092 0.003425 2.070492 Pontohedyl vs Petalif 0.008244 0.003519 2.342411 Pontohedyl vs Dolabri 0.007072 0.003521 2.008685 Pontohedyl vs Styloch 0.006489 0.003572 1.816437

213
Pontohedyl vs Clio_py -0.010148 0.004328 -2.344873 Pontohedyl vs Diacria -0.013693 0.004526 -3.025499 Pontohedyl vs Creseis -0.032067 0.005578 -5.748887 Pontohedyl vs Spongio -0.000555 0.004122 -0.134513 Pontohedyl vs Calliop -0.011871 0.004959 -2.393795 Pontohedyl vs Limapon -0.005899 0.004377 -1.347676 Pontohedyl vs Thuridi 0.002406 0.004364 0.551332 Pontohedyl vs Elysia_ 0.001798 0.004319 0.416229 Pontohedyl vs Elysiel 0.002383 0.004196 0.567870 Pontohedyl vs Boselli 0.002392 0.004190 0.570760 Pontohedyl vs Cyerce_ 0.006500 0.003971 1.636756 Pontohedyl vs Placobr -0.004099 0.004748 -0.863391 Pontohedyl vs Placida 0.002988 0.004190 0.713040 Pontohedyl vs Polybra 0.001801 0.004107 0.438555 Hedylopsis vs Parahed -0.002324 0.002391 -0.972102 Hedylopsis vs Umbracu -0.006401 0.003293 -1.943864 Hedylopsis vs Tylodin -0.007564 0.003187 -2.373017 Hedylopsis vs Akera_b -0.005222 0.002963 -1.762396 Hedylopsis vs Aplysia -0.004640 0.002785 -1.665939 Hedylopsis vs Dolabel -0.004054 0.002722 -1.489380 Hedylopsis vs Siphono -0.004639 0.002845 -1.630528 Hedylopsis vs Bursate -0.004055 0.002660 -1.524540 Hedylopsis vs Petalif -0.003478 0.002593 -1.341449 Hedylopsis vs Dolabri -0.004644 0.002721 -1.706598 Hedylopsis vs Styloch -0.005231 0.002788 -1.876297 Hedylopsis vs Clio_py -0.023046 0.004183 -5.509186 Hedylopsis vs Diacria -0.026004 0.004658 -5.582598 Hedylopsis vs Creseis -0.042028 0.005563 -7.554991 Hedylopsis vs Spongio -0.011677 0.003553 -3.287014 Hedylopsis vs Calliop -0.022957 0.004565 -5.028821 Hedylopsis vs Limapon -0.016424 0.003979 -4.127646 Hedylopsis vs Thuridi -0.008744 0.003689 -2.370366 Hedylopsis vs Elysia_ -0.009325 0.003592 -2.595981 Hedylopsis vs Elysiel -0.008156 0.003495 -2.333681 Hedylopsis vs Boselli -0.008726 0.003490 -2.500268 Hedylopsis vs Cyerce_ -0.004629 0.003280 -1.411214 Hedylopsis vs Placobr -0.014621 0.004007 -3.648890 Hedylopsis vs Placida -0.008131 0.003724 -2.183380 Hedylopsis vs Polybra -0.009315 0.003492 -2.667664 Parahedyle vs Umbracu -0.004071 0.003246 -1.254298 Parahedyle vs Tylodin -0.005240 0.003448 -1.519662 Parahedyle vs Akera_b -0.002898 0.003136 -0.924050 Parahedyle vs Aplysia -0.002316 0.003136 -0.738480 Parahedyle vs Dolabel -0.001730 0.002967 -0.583145 Parahedyle vs Siphono -0.002314 0.003190 -0.725429 Parahedyle vs Bursate -0.002314 0.002966 -0.780131 Parahedyle vs Petalif -0.001155 0.002966 -0.389271 Parahedyle vs Dolabri -0.001736 0.003024 -0.574122 Parahedyle vs Styloch -0.002908 0.003028 -0.960268 Parahedyle vs Clio_py -0.020720 0.004393 -4.716858 Parahedyle vs Diacria -0.023680 0.004774 -4.959904 Parahedyle vs Creseis -0.039106 0.005664 -6.904599 Parahedyle vs Spongio -0.009354 0.003605 -2.594335 Parahedyle vs Calliop -0.020048 0.004826 -4.154006 Parahedyle vs Limapon -0.013511 0.004192 -3.223305 Parahedyle vs Thuridi -0.005829 0.004089 -1.425680 Parahedyle vs Elysia_ -0.006417 0.003957 -1.621843 Parahedyle vs Elysiel -0.005248 0.003868 -1.356704 Parahedyle vs Boselli -0.005819 0.003863 -1.506437 Parahedyle vs Cyerce_ -0.001722 0.003673 -0.468814 Parahedyle vs Placobr -0.011713 0.004299 -2.724903 Parahedyle vs Placida -0.005810 0.003947 -1.471878 Parahedyle vs Polybra -0.006408 0.003865 -1.658165 Umbraculum vs Tylodin -0.001178 0.003092 -0.380977 Umbraculum vs Akera_b 0.001167 0.003499 0.333361 Umbraculum vs Aplysia 0.001749 0.003498 0.500076 Umbraculum vs Dolabel 0.002337 0.003243 0.720692 Umbraculum vs Siphono 0.001172 0.003595 0.325988 Umbraculum vs Bursate 0.002342 0.003350 0.699276 Umbraculum vs Petalif 0.003501 0.003397 1.030500 Umbraculum vs Dolabri 0.002332 0.003398 0.686521 Umbraculum vs Styloch 0.001745 0.003350 0.520823 Umbraculum vs Clio_py -0.016082 0.004704 -3.418787 Umbraculum vs Diacria -0.019636 0.005168 -3.799583 Umbraculum vs Creseis -0.036228 0.006035 -6.003413 Umbraculum vs Spongio -0.005299 0.004101 -1.292023 Umbraculum vs Calliop -0.017197 0.005150 -3.338854 Umbraculum vs Limapon -0.010632 0.004515 -2.354705

214
Umbraculum vs Thuridi -0.002933 0.004221 -0.694811 Umbraculum vs Elysia_ -0.003529 0.004134 -0.853704 Umbraculum vs Elysiel -0.002945 0.004002 -0.735737 Umbraculum vs Boselli -0.002932 0.004210 -0.696421 Umbraculum vs Cyerce_ 0.001177 0.004118 0.285672 Umbraculum vs Placobr -0.008837 0.004579 -1.929727 Umbraculum vs Placida -0.002336 0.004209 -0.554936 Umbraculum vs Polybra -0.003522 0.004373 -0.805465 Tylodina_p vs Akera_b 0.002341 0.003496 0.669529 Tylodina_p vs Aplysia 0.002922 0.003294 0.887181 Tylodina_p vs Dolabel 0.003510 0.003397 1.033190 Tylodina_p vs Siphono 0.002346 0.003399 0.690335 Tylodina_p vs Bursate 0.004689 0.003192 1.469038 Tylodina_p vs Petalif 0.004676 0.003191 1.465315 Tylodina_p vs Dolabri 0.003509 0.003297 1.064353 Tylodina_p vs Styloch 0.002918 0.003352 0.870649 Tylodina_p vs Clio_py -0.015482 0.004592 -3.371333 Tylodina_p vs Diacria -0.018427 0.004917 -3.747330 Tylodina_p vs Creseis -0.034381 0.005908 -5.818992 Tylodina_p vs Spongio -0.004113 0.004140 -0.993550 Tylodina_p vs Calliop -0.015387 0.005177 -2.972187 Tylodina_p vs Limapon -0.008845 0.004551 -1.943649 Tylodina_p vs Thuridi -0.001160 0.004298 -0.269991 Tylodina_p vs Elysia_ -0.002348 0.004255 -0.551777 Tylodina_p vs Elysiel -0.001178 0.004252 -0.276993 Tylodina_p vs Boselli -0.001752 0.004208 -0.416379 Tylodina_p vs Cyerce_ 0.002349 0.004116 0.570700 Tylodina_p vs Placobr -0.007646 0.004651 -1.644090 Tylodina_p vs Placida -0.001157 0.004327 -0.267434 Tylodina_p vs Polybra -0.002341 0.004369 -0.535872 Akera_bull vs Aplysia 0.000581 0.002013 0.288847 Akera_bull vs Dolabel 0.001168 0.001927 0.606023 Akera_bull vs Siphono 0.000004 0.002251 0.001919 Akera_bull vs Bursate 0.001172 0.002015 0.581356 Akera_bull vs Petalif 0.002333 0.002173 1.073354 Akera_bull vs Dolabri 0.001167 0.002176 0.536406 Akera_bull vs Styloch 0.000576 0.002253 0.255828 Akera_bull vs Clio_py -0.017218 0.003928 -4.383054 Akera_bull vs Diacria -0.021957 0.004351 -5.045999 Akera_bull vs Creseis -0.036756 0.005501 -6.682141 Akera_bull vs Spongio -0.005865 0.003092 -1.896412 Akera_bull vs Calliop -0.017747 0.004796 -3.700401 Akera_bull vs Limapon -0.011200 0.004158 -2.693810 Akera_bull vs Thuridi -0.003506 0.004051 -0.865462 Akera_bull vs Elysia_ -0.004101 0.004003 -1.024624 Akera_bull vs Elysiel -0.002932 0.003871 -0.757450 Akera_bull vs Boselli -0.003504 0.003910 -0.896177 Akera_bull vs Cyerce_ 0.000598 0.003813 0.156801 Akera_bull vs Placobr -0.009400 0.004383 -2.144394 Akera_bull vs Placida -0.003495 0.003906 -0.894800 Akera_bull vs Polybra -0.004095 0.003999 -1.023991 Aplysia_ca vs Dolabel 0.000586 0.001746 0.335438 Aplysia_ca vs Siphono -0.000577 0.001294 -0.446220 Aplysia_ca vs Bursate 0.000590 0.001836 0.321209 Aplysia_ca vs Petalif 0.001751 0.001835 0.954602 Aplysia_ca vs Dolabri 0.000585 0.001837 0.318689 Aplysia_ca vs Styloch -0.000005 0.002097 -0.002456 Aplysia_ca vs Clio_py -0.018398 0.003791 -4.853128 Aplysia_ca vs Diacria -0.022541 0.004146 -5.437433 Aplysia_ca vs Creseis -0.037326 0.005500 -6.786837 Aplysia_ca vs Spongio -0.006445 0.002976 -2.165435 Aplysia_ca vs Calliop -0.018324 0.004795 -3.821290 Aplysia_ca vs Limapon -0.011780 0.004027 -2.925019 Aplysia_ca vs Thuridi -0.004090 0.003964 -1.031674 Aplysia_ca vs Elysia_ -0.004682 0.003872 -1.209086 Aplysia_ca vs Elysiel -0.003513 0.003782 -0.928798 Aplysia_ca vs Boselli -0.004085 0.003777 -1.081615 Aplysia_ca vs Cyerce_ 0.000015 0.003677 0.004049 Aplysia_ca vs Placobr -0.009980 0.004096 -2.436467 Aplysia_ca vs Placida -0.004077 0.003863 -1.055465 Aplysia_ca vs Polybra -0.004676 0.004000 -1.169003 Dolabella_ vs Siphono -0.000582 0.001745 -0.333355 Dolabella_ vs Bursate 0.000004 0.001741 0.002283 Dolabella_ vs Petalif 0.001166 0.001739 0.670407 Dolabella_ vs Dolabri 0.000000 0.001534 0.000000 Dolabella_ vs Styloch -0.000591 0.001640 -0.360503 Dolabella_ vs Clio_py -0.018985 0.003837 -4.947834 Dolabella_ vs Diacria -0.022543 0.004189 -5.380942

215
Dolabella_ vs Creseis -0.037937 0.005402 -7.022687 Dolabella_ vs Spongio -0.007035 0.002798 -2.514276 Dolabella_ vs Calliop -0.018326 0.004722 -3.880862 Dolabella_ vs Limapon -0.011782 0.004114 -2.864114 Dolabella_ vs Thuridi -0.004093 0.003964 -1.032461 Dolabella_ vs Elysia_ -0.004686 0.003961 -1.182999 Dolabella_ vs Elysiel -0.003517 0.003783 -0.929744 Dolabella_ vs Boselli -0.004089 0.003868 -1.057267 Dolabella_ vs Cyerce_ 0.000011 0.003677 0.002967 Dolabella_ vs Placobr -0.009983 0.004263 -2.341817 Dolabella_ vs Placida -0.004080 0.003773 -1.081256 Dolabella_ vs Polybra -0.004679 0.003913 -1.195754 Siphonota_ vs Bursate 0.001168 0.001925 0.607049 Siphonota_ vs Petalif 0.002326 0.002008 1.158327 Siphonota_ vs Dolabri 0.001160 0.002010 0.576747 Siphonota_ vs Styloch 0.000573 0.002175 0.263335 Siphonota_ vs Clio_py -0.017819 0.003652 -4.878667 Siphonota_ vs Diacria -0.021977 0.004150 -5.295114 Siphonota_ vs Creseis -0.036177 0.005404 -6.694583 Siphonota_ vs Spongio -0.005871 0.003033 -1.935648 Siphonota_ vs Calliop -0.017759 0.004797 -3.701872 Siphonota_ vs Limapon -0.011210 0.004115 -2.724016 Siphonota_ vs Thuridi -0.003514 0.003966 -0.885992 Siphonota_ vs Elysia_ -0.004108 0.003963 -1.036621 Siphonota_ vs Elysiel -0.002937 0.003874 -0.758202 Siphonota_ vs Boselli -0.003511 0.003826 -0.917694 Siphonota_ vs Cyerce_ 0.000592 0.003679 0.160989 Siphonota_ vs Placobr -0.009410 0.004182 -2.249804 Siphonota_ vs Placida -0.003503 0.003865 -0.906336 Siphonota_ vs Polybra -0.004101 0.004002 -1.024895 Bursatella vs Petalif 0.001158 0.001296 0.893695 Bursatella vs Dolabri -0.000007 0.001299 -0.005725 Bursatella vs Styloch -0.000594 0.001540 -0.385914 Bursatella vs Clio_py -0.018399 0.003746 -4.911755 Bursatella vs Diacria -0.022544 0.003973 -5.673898 Bursatella vs Creseis -0.037343 0.005330 -7.005798 Bursatella vs Spongio -0.006450 0.002917 -2.210955 Bursatella vs Calliop -0.019536 0.004798 -4.071447 Bursatella vs Limapon -0.012981 0.004199 -3.091388 Bursatella vs Thuridi -0.004688 0.003920 -1.195830 Bursatella vs Elysia_ -0.005278 0.004003 -1.318431 Bursatella vs Elysiel -0.004107 0.003915 -1.048916 Bursatella vs Boselli -0.004681 0.003911 -1.196888 Bursatella vs Cyerce_ -0.000576 0.003677 -0.156776 Bursatella vs Placobr -0.009393 0.004259 -2.205376 Bursatella vs Placida -0.005259 0.003995 -1.316333 Bursatella vs Polybra -0.005270 0.003912 -1.347077 Petalifera vs Dolabri -0.001165 0.000820 -1.421343 Petalifera vs Styloch -0.001752 0.001164 -1.505023 Petalifera vs Clio_py -0.019559 0.003603 -5.428014 Petalifera vs Diacria -0.023701 0.003972 -5.967679 Petalifera vs Creseis -0.038497 0.005364 -7.177502 Petalifera vs Spongio -0.007610 0.002671 -2.848839 Petalifera vs Calliop -0.020086 0.004901 -4.098351 Petalifera vs Limapon -0.013540 0.004317 -3.136737 Petalifera vs Thuridi -0.005257 0.004045 -1.299490 Petalifera vs Elysia_ -0.005844 0.004041 -1.446426 Petalifera vs Elysiel -0.004675 0.003954 -1.182375 Petalifera vs Boselli -0.005248 0.003949 -1.328809 Petalifera vs Cyerce_ -0.001148 0.003718 -0.308844 Petalifera vs Placobr -0.011141 0.004298 -2.592293 Petalifera vs Placida -0.005826 0.004033 -1.444491 Petalifera vs Polybra -0.005837 0.003951 -1.477327 Dolabrifer vs Styloch -0.000586 0.001161 -0.504761 Dolabrifer vs Clio_py -0.018384 0.003698 -4.971708 Dolabrifer vs Diacria -0.022535 0.003930 -5.734380 Dolabrifer vs Creseis -0.037326 0.005365 -6.956838 Dolabrifer vs Spongio -0.007033 0.002606 -2.698442 Dolabrifer vs Calliop -0.018913 0.004902 -3.858187 Dolabrifer vs Limapon -0.012368 0.004318 -2.864239 Dolabrifer vs Thuridi -0.004087 0.004047 -1.009859 Dolabrifer vs Elysia_ -0.004677 0.004043 -1.156772 Dolabrifer vs Elysiel -0.003507 0.003956 -0.886499 Dolabrifer vs Boselli -0.004081 0.003951 -1.032834 Dolabrifer vs Cyerce_ 0.000016 0.003721 0.004432 Dolabrifer vs Placobr -0.009972 0.004300 -2.319262 Dolabrifer vs Placida -0.004661 0.004035 -1.155019 Dolabrifer vs Polybra -0.004670 0.003953 -1.181310

216
Stylocheil vs Clio_py -0.017799 0.003653 -4.872808 Stylocheil vs Diacria -0.021942 0.004104 -5.347050 Stylocheil vs Creseis -0.036738 0.005431 -6.764142 Stylocheil vs Spongio -0.005852 0.002737 -2.138331 Stylocheil vs Calliop -0.018332 0.004940 -3.710762 Stylocheil vs Limapon -0.011784 0.004361 -2.702361 Stylocheil vs Thuridi -0.003498 0.004176 -0.837810 Stylocheil vs Elysia_ -0.004090 0.004171 -0.980655 Stylocheil vs Elysiel -0.002921 0.004002 -0.729906 Stylocheil vs Boselli -0.003495 0.004082 -0.856116 Stylocheil vs Cyerce_ 0.000605 0.003858 0.156732 Stylocheil vs Placobr -0.009387 0.004421 -2.123194 Stylocheil vs Placida -0.004075 0.004080 -0.998754 Stylocheil vs Polybra -0.004084 0.004084 -0.999983 Clio_pyram vs Diacria -0.003572 0.004174 -0.855840 Clio_pyram vs Creseis -0.020728 0.005468 -3.790884 Clio_pyram vs Spongio 0.011360 0.004319 2.630473 Clio_pyram vs Calliop 0.000036 0.005759 0.006281 Clio_pyram vs Limapon 0.006021 0.005291 1.137900 Clio_pyram vs Thuridi 0.014353 0.005279 2.718847 Clio_pyram vs Elysia_ 0.013715 0.005236 2.619176 Clio_pyram vs Elysiel 0.014281 0.005165 2.764932 Clio_pyram vs Boselli 0.014302 0.005198 2.751456 Clio_pyram vs Cyerce_ 0.018412 0.005017 3.669911 Clio_pyram vs Placobr 0.008409 0.005443 1.544848 Clio_pyram vs Placida 0.014294 0.005123 2.790216 Clio_pyram vs Polybra 0.013711 0.005163 2.655290 Diacria_qu vs Creseis -0.016532 0.005561 -2.972756 Diacria_qu vs Spongio 0.016112 0.004555 3.537189 Diacria_qu vs Calliop 0.003599 0.005882 0.611911 Diacria_qu vs Limapon 0.009588 0.005357 1.789608 Diacria_qu vs Thuridi 0.017325 0.005343 3.242824 Diacria_qu vs Elysia_ 0.016679 0.005301 3.146336 Diacria_qu vs Elysiel 0.017252 0.005267 3.275697 Diacria_qu vs Boselli 0.017265 0.005228 3.302664 Diacria_qu vs Cyerce_ 0.021375 0.005052 4.230902 Diacria_qu vs Placobr 0.011384 0.005601 2.032670 Diacria_qu vs Placida 0.017853 0.005263 3.391928 Diacria_qu vs Polybra 0.017270 0.005165 3.343607 Creseis_ac vs Spongio 0.031494 0.005675 5.549526 Creseis_ac vs Calliop 0.019596 0.006534 2.998984 Creseis_ac vs Limapon 0.026206 0.006115 4.285468 Creseis_ac vs Thuridi 0.033368 0.006139 5.435836 Creseis_ac vs Elysia_ 0.032685 0.006186 5.283505 Creseis_ac vs Elysiel 0.033839 0.006032 5.609562 Creseis_ac vs Boselli 0.033261 0.006062 5.486849 Creseis_ac vs Cyerce_ 0.037377 0.005935 6.298126 Creseis_ac vs Placobr 0.027369 0.006311 4.336508 Creseis_ac vs Placida 0.033840 0.006028 5.613722 Creseis_ac vs Polybra 0.033261 0.005999 5.544584 Spongiobra vs Calliop -0.011300 0.005228 -2.161620 Spongiobra vs Limapon -0.004737 0.004606 -1.028528 Spongiobra vs Thuridi 0.002968 0.004511 0.657850 Spongiobra vs Elysia_ 0.002357 0.004504 0.523251 Spongiobra vs Elysiel 0.002940 0.004426 0.664288 Spongiobra vs Boselli 0.002951 0.004497 0.656143 Spongiobra vs Cyerce_ 0.007056 0.004293 1.643601 Spongiobra vs Placobr -0.002943 0.004665 -0.630851 Spongiobra vs Placida 0.002955 0.004417 0.669071 Spongiobra vs Polybra 0.002361 0.004499 0.524784 Calliopaea vs Limapon 0.007175 0.003703 1.937460 Calliopaea vs Thuridi 0.014879 0.004228 3.519276 Calliopaea vs Elysia_ 0.014241 0.004096 3.476739 Calliopaea vs Elysiel 0.015410 0.004181 3.686004 Calliopaea vs Boselli 0.014827 0.004174 3.552068 Calliopaea vs Cyerce_ 0.018927 0.003824 4.949779 Calliopaea vs Placobr 0.008947 0.004623 1.935225 Calliopaea vs Placida 0.014241 0.003822 3.726517 Calliopaea vs Polybra 0.014238 0.003963 3.592585 Limapontia vs Thuridi 0.007714 0.003537 2.180702 Limapontia vs Elysia_ 0.007092 0.003430 2.067809 Limapontia vs Elysiel 0.007665 0.003273 2.342076 Limapontia vs Boselli 0.007683 0.003271 2.348797 Limapontia vs Cyerce_ 0.011790 0.002937 4.014732 Limapontia vs Placobr 0.001789 0.003775 0.473937 Limapontia vs Placida 0.007099 0.002992 2.372338 Limapontia vs Polybra 0.007093 0.003327 2.131643 Thuridilla vs Elysia_ -0.000605 0.002416 -0.250313

217
Thuridilla vs Elysiel 0.000567 0.002416 0.234814 Thuridilla vs Boselli -0.000008 0.002336 -0.003530 Thuridilla vs Cyerce_ 0.004104 0.002182 1.880705 Thuridilla vs Placobr -0.005910 0.002994 -1.974349 Thuridilla vs Placida 0.000590 0.002982 0.197845 Thuridilla vs Polybra -0.000600 0.002805 -0.213760 Elysia_vir vs Elysiel 0.001759 0.001851 0.950196 Elysia_vir vs Boselli 0.000595 0.002101 0.283108 Elysia_vir vs Cyerce_ 0.004697 0.002259 2.079429 Elysia_vir vs Placobr -0.005297 0.003101 -1.708116 Elysia_vir vs Placida 0.000602 0.002738 0.219962 Elysia_vir vs Polybra 0.000005 0.002743 0.001963 Elysiella_ vs Boselli -0.000574 0.002021 -0.283963 Elysiella_ vs Cyerce_ 0.003529 0.002184 1.615841 Elysiella_ vs Placobr -0.006466 0.002989 -2.163477 Elysiella_ vs Placida 0.000018 0.002860 0.006438 Elysiella_ vs Polybra -0.001163 0.002681 -0.433713 Bosellia_m vs Cyerce_ 0.004100 0.001841 2.227734 Bosellia_m vs Placobr -0.005888 0.002550 -2.309498 Bosellia_m vs Placida 0.000008 0.002795 0.002809 Bosellia_m vs Polybra -0.000589 0.002543 -0.231763 Cyerce_nig vs Placobr -0.009992 0.002805 -3.562400 Cyerce_nig vs Placida -0.003506 0.002176 -1.611384 Cyerce_nig vs Polybra -0.004689 0.002102 -2.231381 Placobranc vs Placida 0.006489 0.003465 1.872809 Placobranc vs Polybra 0.005299 0.003417 1.550671 Placida_cr vs Polybra -0.000596 0.002974 -0.200531
Opisthobranchia-Datensatz (28S rDNA) Ergebnisse der Analyse der relativen Substitutionsraten auf der Grundlage des 28S rDNA-Datensatzes mit Odostomia sp. als Außengruppe (1). Eine signifikant unterschiedliche Substitutionsgeschwindigkeit ist gegeben, wenn der „Z-score“ (k12-k13/SD) höher als 1,96 ist. Positive „Z-score“-Werte bedeuten, dass Taxon (2) eine größere Evolutionsgeschwindigkeit aufweist; negative Werte stehen für eine erhöhte Substitutionsrate des Taxons (3). (2) (3) K12 - K13 SD Z-score Bathydoris vs Dendrod 0.028929 0.013659 2.118037 Bathydoris vs Chromod 0.021155 0.013104 1.614404 Bathydoris vs Hypselo 0.010826 0.013799 0.784538 Bathydoris vs Dendron -0.128378 0.023126 -5.551319 Bathydoris vs Flabell -0.120664 0.022029 -5.477459 Bathydoris vs Cuthona -0.104832 0.021715 -4.827652 Bathydoris vs Euselen 0.046410 0.017005 2.729133 Bathydoris vs Pleurob 0.018625 0.018065 1.031048 Bathydoris vs Berthel 0.046390 0.017429 2.661558 Bathydoris vs Berthel -0.061487 0.021597 -2.847048 Bathydoris vs Bathybe 0.032482 0.017597 1.845866 Bathydoris vs Tomptho 0.033613 0.017251 1.948406 Bathydoris vs Tylodin 0.107034 0.017372 6.161306 Bathydoris vs Umbracu 0.107977 0.017742 6.086059 Bathydoris vs Calliop 0.061397 0.018841 3.258616 Bathydoris vs Limapon 0.096570 0.017686 5.460335 Bathydoris vs Thuridi 0.102822 0.017040 6.034287 Bathydoris vs Boselli 0.104037 0.017217 6.042731 Bathydoris vs Cyerce_ 0.104316 0.017156 6.080536 Bathydoris vs Elysia_ 0.085172 0.018085 4.709505 Bathydoris vs Placobr 0.101556 0.017369 5.847014 Bathydoris vs Haminoe 0.099142 0.017491 5.668021 Bathydoris vs Phanero 0.109360 0.017541 6.234630 Bathydoris vs Atys_se 0.113844 0.017167 6.631593 Bathydoris vs cf._Ret 0.102601 0.017582 5.835505 Bathydoris vs Philino 0.096207 0.017447 5.514270 Bathydoris vs Chelido 0.102529 0.017581 5.831881 Bathydoris vs Odontog 0.098717 0.017608 5.606359 Bathydoris vs Navanax 0.101042 0.017582 5.746853 Bathydoris vs Philino 0.081187 0.017244 4.708172

218
Bathydoris vs Bulla_c 0.093025 0.017958 5.180055 Bathydoris vs Runcina 0.062856 0.018532 3.391772 Bathydoris vs Sagamin 0.087156 0.017830 4.888236 Bathydoris vs Pupa_so 0.082909 0.016810 4.932107 Bathydoris vs Hydatin 0.077573 0.016864 4.599961 Bathydoris vs Aplysia 0.107423 0.017558 6.118326 Bathydoris vs Dolabel 0.112473 0.017011 6.611822 Bathydoris vs Akera_b 0.099568 0.017520 5.683226 Bathydoris vs Unela_g 0.105271 0.017453 6.031709 Bathydoris vs Parahed 0.105271 0.017453 6.031709 Bathydoris vs Pontohe 0.096491 0.017805 5.419263 Bathydoris vs Hedylop 0.113752 0.016915 6.725005 Dendrodori vs Chromod -0.006523 0.008944 -0.729332 Dendrodori vs Hypselo -0.016727 0.009585 -1.745142 Dendrodori vs Dendron -0.159375 0.021442 -7.432735 Dendrodori vs Flabell -0.144025 0.021080 -6.832313 Dendrodori vs Cuthona -0.137403 0.020811 -6.602447 Dendrodori vs Euselen 0.008888 0.016232 0.547593 Dendrodori vs Pleurob -0.013318 0.016821 -0.791765 Dendrodori vs Berthel 0.011609 0.016261 0.713936 Dendrodori vs Berthel -0.099401 0.020764 -4.787147 Dendrodori vs Bathybe -0.003500 0.016507 -0.212035 Dendrodori vs Tomptho -0.003545 0.016112 -0.220026 Dendrodori vs Tylodin 0.076902 0.015911 4.833108 Dendrodori vs Umbracu 0.079455 0.015947 4.982364 Dendrodori vs Calliop 0.036862 0.017425 2.115483 Dendrodori vs Limapon 0.069330 0.016196 4.280623 Dendrodori vs Thuridi 0.076847 0.015901 4.832915 Dendrodori vs Boselli 0.075486 0.015899 4.747772 Dendrodori vs Cyerce_ 0.075734 0.016026 4.725680 Dendrodori vs Elysia_ 0.056551 0.016532 3.420619 Dendrodori vs Placobr 0.074319 0.016038 4.633873 Dendrodori vs Haminoe 0.069321 0.016149 4.292620 Dendrodori vs Phanero 0.079514 0.015911 4.997289 Dendrodori vs Atys_se 0.085467 0.015711 5.439921 Dendrodori vs cf._Ret 0.074034 0.016520 4.481385 Dendrodori vs Philino 0.067819 0.016028 4.231377 Dendrodori vs Chelido 0.074178 0.016470 4.503773 Dendrodori vs Odontog 0.070361 0.016448 4.277907 Dendrodori vs Navanax 0.074067 0.015948 4.644171 Dendrodori vs Philino 0.054104 0.015750 3.435170 Dendrodori vs Bulla_c 0.064457 0.015998 4.029143 Dendrodori vs Runcina 0.034376 0.017434 1.971764 Dendrodori vs Sagamin 0.057550 0.016037 3.588526 Dendrodori vs Pupa_so 0.052975 0.015510 3.415651 Dendrodori vs Hydatin 0.047555 0.015780 3.013592 Dendrodori vs Aplysia 0.079166 0.015738 5.030113 Dendrodori vs Dolabel 0.086552 0.015284 5.662917 Dendrodori vs Akera_b 0.071198 0.016297 4.368871 Dendrodori vs Unela_g 0.076676 0.016443 4.663114 Dendrodori vs Parahed 0.076676 0.016443 4.663114 Dendrodori vs Pontohe 0.067885 0.016545 4.103018 Dendrodori vs Hedylop 0.085347 0.015998 5.335005 Chromodori vs Hypselo -0.008888 0.006557 -1.355478 Chromodori vs Dendron -0.154606 0.021624 -7.149605 Chromodori vs Flabell -0.141980 0.020881 -6.799555 Chromodori vs Cuthona -0.135589 0.020880 -6.493594 Chromodori vs Euselen 0.009953 0.016148 0.616402 Chromodori vs Pleurob -0.011058 0.017105 -0.646445 Chromodori vs Berthel 0.014172 0.015937 0.889244 Chromodori vs Berthel -0.095017 0.020548 -4.624082 Chromodori vs Bathybe 0.000240 0.016535 0.014521 Chromodori vs Tomptho -0.003953 0.016134 -0.245026 Chromodori vs Tylodin 0.078589 0.016171 4.859768 Chromodori vs Umbracu 0.081174 0.016196 5.011968 Chromodori vs Calliop 0.042305 0.017848 2.370275 Chromodori vs Limapon 0.074810 0.016441 4.550179 Chromodori vs Thuridi 0.081087 0.015989 5.071509 Chromodori vs Boselli 0.079713 0.015931 5.003784 Chromodori vs Cyerce_ 0.078703 0.016011 4.915550 Chromodori vs Elysia_ 0.061975 0.016557 3.743091 Chromodori vs Placobr 0.078545 0.016015 4.904481 Chromodori vs Haminoe 0.074894 0.016511 4.536047 Chromodori vs Phanero 0.082502 0.016257 5.074908 Chromodori vs Atys_se 0.087151 0.015988 5.450861 Chromodori vs cf._Ret 0.075639 0.016526 4.576958 Chromodori vs Philino 0.073190 0.016298 4.490772 Chromodori vs Chelido 0.081019 0.016657 4.864041

219
Chromodori vs Odontog 0.077183 0.016741 4.610362 Chromodori vs Navanax 0.082266 0.016198 5.078653 Chromodori vs Philino 0.059529 0.015797 3.768452 Chromodori vs Bulla_c 0.068526 0.016002 4.282369 Chromodori vs Runcina 0.038285 0.017695 2.163652 Chromodori vs Sagamin 0.063000 0.016333 3.857259 Chromodori vs Pupa_so 0.059846 0.015430 3.878636 Chromodori vs Hydatin 0.054380 0.015642 3.476587 Chromodori vs Aplysia 0.083465 0.015833 5.271675 Chromodori vs Dolabel 0.092336 0.015631 5.907121 Chromodori vs Akera_b 0.074058 0.016259 4.554823 Chromodori vs Unela_g 0.079666 0.016518 4.822971 Chromodori vs Parahed 0.079666 0.016518 4.822971 Chromodori vs Pontohe 0.072095 0.016834 4.282805 Chromodori vs Hedylop 0.089792 0.016170 5.552956 Hypselodor vs Dendron -0.142088 0.021326 -6.662650 Hypselodor vs Flabell -0.126544 0.020891 -6.057445 Hypselodor vs Cuthona -0.123003 0.021031 -5.848750 Hypselodor vs Euselen 0.022934 0.016386 1.399632 Hypselodor vs Pleurob -0.000714 0.017158 -0.041608 Hypselodor vs Berthel 0.025782 0.016256 1.585954 Hypselodor vs Berthel -0.084143 0.020696 -4.065723 Hypselodor vs Bathybe 0.011912 0.016792 0.709402 Hypselodor vs Tomptho 0.009066 0.016691 0.543136 Hypselodor vs Tylodin 0.091260 0.016697 5.465602 Hypselodor vs Umbracu 0.093830 0.016612 5.648404 Hypselodor vs Calliop 0.052353 0.018179 2.879831 Hypselodor vs Limapon 0.086184 0.016965 5.080006 Hypselodor vs Thuridi 0.092442 0.016512 5.598573 Hypselodor vs Boselli 0.091055 0.016400 5.552304 Hypselodor vs Cyerce_ 0.090095 0.016426 5.484769 Hypselodor vs Elysia_ 0.073372 0.017171 4.272949 Hypselodor vs Placobr 0.089907 0.016484 5.454329 Hypselodor vs Haminoe 0.087568 0.016868 5.191396 Hypselodor vs Phanero 0.095118 0.016888 5.632318 Hypselodor vs Atys_se 0.099748 0.016350 6.100811 Hypselodor vs cf._Ret 0.088292 0.017077 5.170117 Hypselodor vs Philino 0.085879 0.016925 5.074224 Hypselodor vs Chelido 0.093685 0.017054 5.493422 Hypselodor vs Odontog 0.089868 0.017187 5.228703 Hypselodor vs Navanax 0.094943 0.016706 5.683286 Hypselodor vs Philino 0.072304 0.016528 4.374519 Hypselodor vs Bulla_c 0.081255 0.016450 4.939419 Hypselodor vs Runcina 0.051132 0.018018 2.837779 Hypselodor vs Sagamin 0.075716 0.016840 4.496184 Hypselodor vs Pupa_so 0.068484 0.015599 4.390396 Hypselodor vs Hydatin 0.063006 0.015678 4.018851 Hypselodor vs Aplysia 0.094709 0.016238 5.832501 Hypselodor vs Dolabel 0.103574 0.015884 6.520661 Hypselodor vs Akera_b 0.084016 0.016695 5.032353 Hypselodor vs Unela_g 0.092272 0.017141 5.383282 Hypselodor vs Parahed 0.092272 0.017141 5.383282 Hypselodor vs Pontohe 0.083458 0.017184 4.856625 Hypselodor vs Hedylop 0.100937 0.016620 6.073116 Dendronotu vs Flabell 0.021341 0.017404 1.226254 Dendronotu vs Cuthona 0.022862 0.016501 1.385546 Dendronotu vs Euselen 0.170652 0.022348 7.636245 Dendronotu vs Pleurob 0.144940 0.022706 6.383229 Dendronotu vs Berthel 0.171749 0.022425 7.658669 Dendronotu vs Berthel 0.069169 0.024196 2.858738 Dendronotu vs Bathybe 0.160422 0.022624 7.090687 Dendronotu vs Tomptho 0.161343 0.022817 7.071172 Dendronotu vs Tylodin 0.230949 0.024191 9.546853 Dendronotu vs Umbracu 0.233696 0.023737 9.845344 Dendronotu vs Calliop 0.196421 0.023764 8.265452 Dendronotu vs Limapon 0.229853 0.023434 9.808351 Dendronotu vs Thuridi 0.233914 0.023554 9.931138 Dendronotu vs Boselli 0.234864 0.023357 10.055445 Dendronotu vs Cyerce_ 0.235267 0.023396 10.055662 Dendronotu vs Elysia_ 0.217796 0.023584 9.234742 Dendronotu vs Placobr 0.235223 0.023446 10.032482 Dendronotu vs Haminoe 0.234964 0.023964 9.804954 Dendronotu vs Phanero 0.239078 0.023714 10.081555 Dendronotu vs Atys_se 0.248872 0.023832 10.442786 Dendronotu vs cf._Ret 0.229449 0.024124 9.511210 Dendronotu vs Philino 0.230641 0.023973 9.621062 Dendronotu vs Chelido 0.235995 0.023937 9.859105 Dendronotu vs Odontog 0.230842 0.024108 9.575256

220
Dendronotu vs Navanax 0.234679 0.023959 9.795050 Dendronotu vs Philino 0.216513 0.023533 9.200385 Dendronotu vs Bulla_c 0.230694 0.023010 10.026031 Dendronotu vs Runcina 0.206788 0.024842 8.324199 Dendronotu vs Sagamin 0.222976 0.023925 9.319766 Dendronotu vs Pupa_so 0.218363 0.023525 9.282029 Dendronotu vs Hydatin 0.215487 0.023662 9.106803 Dendronotu vs Aplysia 0.238792 0.023729 10.063216 Dendronotu vs Dolabel 0.247426 0.023282 10.627321 Dendronotu vs Akera_b 0.226435 0.023652 9.573529 Dendronotu vs Unela_g 0.237328 0.023942 9.912587 Dendronotu vs Parahed 0.237328 0.023942 9.912587 Dendronotu vs Pontohe 0.230823 0.024185 9.544116 Dendronotu vs Hedylop 0.243703 0.023591 10.330235 Flabellina vs Cuthona 0.015717 0.014932 1.052519 Flabellina vs Euselen 0.163558 0.021247 7.698060 Flabellina vs Pleurob 0.135681 0.021598 6.282037 Flabellina vs Berthel 0.166783 0.021056 7.921041 Flabellina vs Berthel 0.066317 0.023853 2.780243 Flabellina vs Bathybe 0.153860 0.021564 7.135168 Flabellina vs Tomptho 0.156213 0.022121 7.061715 Flabellina vs Tylodin 0.218894 0.022733 9.628770 Flabellina vs Umbracu 0.217965 0.023041 9.459866 Flabellina vs Calliop 0.178872 0.023610 7.576075 Flabellina vs Limapon 0.213967 0.022732 9.412449 Flabellina vs Thuridi 0.215242 0.022396 9.610520 Flabellina vs Boselli 0.214914 0.022345 9.618035 Flabellina vs Cyerce_ 0.215267 0.022322 9.643747 Flabellina vs Elysia_ 0.198862 0.022372 8.888871 Flabellina vs Placobr 0.213915 0.022040 9.705875 Flabellina vs Haminoe 0.210975 0.022555 9.353874 Flabellina vs Phanero 0.215153 0.022494 9.564755 Flabellina vs Atys_se 0.221230 0.022387 9.882258 Flabellina vs cf._Ret 0.209277 0.023010 9.095229 Flabellina vs Philino 0.209023 0.022829 9.156001 Flabellina vs Chelido 0.216973 0.022663 9.574007 Flabellina vs Odontog 0.211774 0.022966 9.221008 Flabellina vs Navanax 0.214313 0.022834 9.385535 Flabellina vs Philino 0.196016 0.022743 8.618698 Flabellina vs Bulla_c 0.206597 0.022391 9.226575 Flabellina vs Runcina 0.179538 0.023352 7.688187 Flabellina vs Sagamin 0.198303 0.023219 8.540383 Flabellina vs Pupa_so 0.197995 0.022822 8.675747 Flabellina vs Hydatin 0.194797 0.022664 8.594940 Flabellina vs Aplysia 0.211796 0.022262 9.513735 Flabellina vs Dolabel 0.222410 0.022379 9.938265 Flabellina vs Akera_b 0.205095 0.022608 9.071913 Flabellina vs Unela_g 0.220019 0.023098 9.525309 Flabellina vs Parahed 0.220019 0.023098 9.525309 Flabellina vs Pontohe 0.212198 0.022986 9.231675 Flabellina vs Hedylop 0.227607 0.022623 10.060676 Cuthona_na vs Euselen 0.149732 0.021029 7.120220 Cuthona_na vs Pleurob 0.122257 0.021460 5.696837 Cuthona_na vs Berthel 0.149946 0.021154 7.088233 Cuthona_na vs Berthel 0.057343 0.023642 2.425468 Cuthona_na vs Bathybe 0.141012 0.021282 6.625983 Cuthona_na vs Tomptho 0.150873 0.021756 6.934769 Cuthona_na vs Tylodin 0.209051 0.022748 9.189828 Cuthona_na vs Umbracu 0.203654 0.022519 9.043835 Cuthona_na vs Calliop 0.165189 0.023106 7.149117 Cuthona_na vs Limapon 0.199405 0.022014 9.058086 Cuthona_na vs Thuridi 0.199405 0.022164 8.996973 Cuthona_na vs Boselli 0.200441 0.022379 8.956540 Cuthona_na vs Cyerce_ 0.202056 0.022289 9.065177 Cuthona_na vs Elysia_ 0.182917 0.022476 8.138297 Cuthona_na vs Placobr 0.196706 0.022231 8.848455 Cuthona_na vs Haminoe 0.201685 0.022268 9.057063 Cuthona_na vs Phanero 0.207135 0.021824 9.491342 Cuthona_na vs Atys_se 0.212086 0.022014 9.634134 Cuthona_na vs cf._Ret 0.200116 0.022235 9.000014 Cuthona_na vs Philino 0.197643 0.022519 8.776723 Cuthona_na vs Chelido 0.202938 0.022558 8.996342 Cuthona_na vs Odontog 0.195049 0.022732 8.580392 Cuthona_na vs Navanax 0.204223 0.022471 9.088257 Cuthona_na vs Philino 0.188582 0.022111 8.529037 Cuthona_na vs Bulla_c 0.197908 0.022154 8.933385 Cuthona_na vs Runcina 0.178648 0.022983 7.772904 Cuthona_na vs Sagamin 0.194749 0.022530 8.643898

221
Cuthona_na vs Pupa_so 0.188993 0.022658 8.341172 Cuthona_na vs Hydatin 0.185798 0.022670 8.195752 Cuthona_na vs Aplysia 0.200433 0.022117 9.062560 Cuthona_na vs Dolabel 0.213688 0.022187 9.631205 Cuthona_na vs Akera_b 0.188817 0.022042 8.566241 Cuthona_na vs Unela_g 0.208244 0.022521 9.246729 Cuthona_na vs Parahed 0.208244 0.022521 9.246729 Cuthona_na vs Pontohe 0.198994 0.022227 8.952872 Cuthona_na vs Hedylop 0.212202 0.022350 9.494307 Euselenops vs Pleurob -0.023510 0.009151 -2.569061 Euselenops vs Berthel 0.001379 0.007775 0.177411 Euselenops vs Berthel -0.102331 0.017344 -5.900012 Euselenops vs Bathybe -0.010985 0.007875 -1.394921 Euselenops vs Tomptho -0.008464 0.011658 -0.726043 Euselenops vs Tylodin 0.073029 0.014886 4.905847 Euselenops vs Umbracu 0.072881 0.014827 4.915411 Euselenops vs Calliop 0.026794 0.016843 1.590832 Euselenops vs Limapon 0.061806 0.015517 3.983114 Euselenops vs Thuridi 0.067948 0.015379 4.418239 Euselenops vs Boselli 0.070418 0.015332 4.592930 Euselenops vs Cyerce_ 0.066807 0.015538 4.299634 Euselenops vs Elysia_ 0.049030 0.015741 3.114739 Euselenops vs Placobr 0.065419 0.015507 4.218639 Euselenops vs Haminoe 0.064356 0.015296 4.207442 Euselenops vs Phanero 0.069416 0.015080 4.603269 Euselenops vs Atys_se 0.075256 0.014837 5.072065 Euselenops vs cf._Ret 0.065085 0.015629 4.164497 Euselenops vs Philino 0.062659 0.015805 3.964486 Euselenops vs Chelido 0.069118 0.015963 4.329794 Euselenops vs Odontog 0.064093 0.015994 4.007336 Euselenops vs Navanax 0.069152 0.015722 4.398494 Euselenops vs Philino 0.047728 0.015577 3.063955 Euselenops vs Bulla_c 0.061765 0.015381 4.015652 Euselenops vs Runcina 0.032092 0.016155 1.986490 Euselenops vs Sagamin 0.053807 0.015951 3.373198 Euselenops vs Pupa_so 0.046666 0.015886 2.937547 Euselenops vs Hydatin 0.045276 0.015602 2.901956 Euselenops vs Aplysia 0.070403 0.015380 4.577526 Euselenops vs Dolabel 0.075397 0.015145 4.978344 Euselenops vs Akera_b 0.058618 0.015591 3.759802 Euselenops vs Unela_g 0.070340 0.015491 4.540722 Euselenops vs Parahed 0.070340 0.015491 4.540722 Euselenops vs Pontohe 0.065389 0.015665 4.174288 Euselenops vs Hedylop 0.080503 0.014815 5.433944 Pleurobran vs Berthel 0.024943 0.008896 2.803840 Pleurobran vs Berthel -0.081737 0.018275 -4.472519 Pleurobran vs Bathybe 0.015261 0.009081 1.680493 Pleurobran vs Tomptho 0.015115 0.012437 1.215316 Pleurobran vs Tylodin 0.099438 0.015766 6.307234 Pleurobran vs Umbracu 0.100551 0.015816 6.357333 Pleurobran vs Calliop 0.051647 0.017328 2.980480 Pleurobran vs Limapon 0.086805 0.016223 5.350626 Pleurobran vs Thuridi 0.092890 0.016250 5.716305 Pleurobran vs Boselli 0.094105 0.016216 5.803183 Pleurobran vs Cyerce_ 0.093086 0.016246 5.729911 Pleurobran vs Elysia_ 0.073937 0.016272 4.543838 Pleurobran vs Placobr 0.091626 0.016202 5.655137 Pleurobran vs Haminoe 0.091569 0.016006 5.720852 Pleurobran vs Phanero 0.100428 0.016066 6.250924 Pleurobran vs Atys_se 0.107609 0.015967 6.739577 Pleurobran vs cf._Ret 0.093678 0.016656 5.624152 Pleurobran vs Philino 0.091280 0.016836 5.421655 Pleurobran vs Chelido 0.096449 0.016703 5.774408 Pleurobran vs Odontog 0.091392 0.016790 5.443385 Pleurobran vs Navanax 0.096472 0.016699 5.776993 Pleurobran vs Philino 0.076384 0.016370 4.666216 Pleurobran vs Bulla_c 0.090465 0.016411 5.512502 Pleurobran vs Runcina 0.059438 0.017061 3.483805 Pleurobran vs Sagamin 0.083749 0.016496 5.076979 Pleurobran vs Pupa_so 0.071483 0.016304 4.384335 Pleurobran vs Hydatin 0.068757 0.016090 4.273314 Pleurobran vs Aplysia 0.097912 0.016540 5.919767 Pleurobran vs Dolabel 0.102819 0.016269 6.320060 Pleurobran vs Akera_b 0.087276 0.016657 5.239509 Pleurobran vs Unela_g 0.097745 0.016526 5.914650 Pleurobran vs Parahed 0.097745 0.016526 5.914650 Pleurobran vs Pontohe 0.090269 0.016481 5.477037 Pleurobran vs Hedylop 0.107931 0.015680 6.883266

222
Berthellin vs Berthel -0.103831 0.017602 -5.898662 Berthellin vs Bathybe -0.013730 0.007877 -1.743093 Berthellin vs Tomptho -0.011112 0.011529 -0.963759 Berthellin vs Tylodin 0.069227 0.014854 4.660511 Berthellin vs Umbracu 0.069371 0.014942 4.642804 Berthellin vs Calliop 0.026838 0.016531 1.623527 Berthellin vs Limapon 0.061862 0.015281 4.048390 Berthellin vs Thuridi 0.065494 0.015253 4.293981 Berthellin vs Boselli 0.066711 0.015327 4.352494 Berthellin vs Cyerce_ 0.065612 0.015244 4.304018 Berthellin vs Elysia_ 0.046494 0.015575 2.985196 Berthellin vs Placobr 0.064225 0.015205 4.223952 Berthellin vs Haminoe 0.063164 0.015503 4.074413 Berthellin vs Phanero 0.070726 0.015369 4.601830 Berthellin vs Atys_se 0.075423 0.015063 5.007291 Berthellin vs cf._Ret 0.065103 0.015664 4.156285 Berthellin vs Philino 0.062676 0.016048 3.905455 Berthellin vs Chelido 0.069142 0.016096 4.295617 Berthellin vs Odontog 0.064113 0.016181 3.962149 Berthellin vs Navanax 0.069176 0.015811 4.375221 Berthellin vs Philino 0.047862 0.015600 3.068082 Berthellin vs Bulla_c 0.059332 0.015358 3.863171 Berthellin vs Runcina 0.032233 0.016333 1.973452 Berthellin vs Sagamin 0.052647 0.015930 3.304857 Berthellin vs Pupa_so 0.045554 0.015800 2.883115 Berthellin vs Hydatin 0.044158 0.015571 2.835824 Berthellin vs Aplysia 0.068009 0.015530 4.379242 Berthellin vs Dolabel 0.074230 0.015232 4.873348 Berthellin vs Akera_b 0.057439 0.015501 3.705506 Berthellin vs Unela_g 0.066731 0.015533 4.296196 Berthellin vs Parahed 0.066731 0.015533 4.296196 Berthellin vs Pontohe 0.061754 0.015375 4.016486 Berthellin vs Hedylop 0.079259 0.014973 5.293383 Berthella_ vs Bathybe 0.093900 0.017588 5.338945 Berthella_ vs Tomptho 0.098507 0.018160 5.424316 Berthella_ vs Tylodin 0.173820 0.019782 8.786814 Berthella_ vs Umbracu 0.180456 0.020275 8.900342 Berthella_ vs Calliop 0.134817 0.021423 6.293216 Berthella_ vs Limapon 0.168756 0.020338 8.297431 Berthella_ vs Thuridi 0.175120 0.020851 8.398842 Berthella_ vs Boselli 0.177688 0.020512 8.662696 Berthella_ vs Cyerce_ 0.176884 0.020570 8.599033 Berthella_ vs Elysia_ 0.158632 0.021085 7.523590 Berthella_ vs Placobr 0.175120 0.020794 8.421584 Berthella_ vs Haminoe 0.172806 0.020774 8.318316 Berthella_ vs Phanero 0.175247 0.020762 8.440707 Berthella_ vs Atys_se 0.179799 0.020665 8.700724 Berthella_ vs cf._Ret 0.172144 0.020840 8.260210 Berthella_ vs Philino 0.171096 0.020896 8.187942 Berthella_ vs Chelido 0.181595 0.020986 8.653325 Berthella_ vs Odontog 0.176556 0.021337 8.274591 Berthella_ vs Navanax 0.180467 0.020889 8.639239 Berthella_ vs Philino 0.158951 0.020728 7.668477 Berthella_ vs Bulla_c 0.169470 0.020701 8.186497 Berthella_ vs Runcina 0.141791 0.021193 6.690371 Berthella_ vs Sagamin 0.165149 0.021000 7.864403 Berthella_ vs Pupa_so 0.158053 0.020349 7.767178 Berthella_ vs Hydatin 0.155491 0.020358 7.638012 Berthella_ vs Aplysia 0.182857 0.020819 8.783267 Berthella_ vs Dolabel 0.189037 0.020280 9.321466 Berthella_ vs Akera_b 0.169386 0.020744 8.165550 Berthella_ vs Unela_g 0.176297 0.020656 8.534703 Berthella_ vs Parahed 0.176297 0.020656 8.534703 Berthella_ vs Pontohe 0.173800 0.020703 8.395028 Berthella_ vs Hedylop 0.189038 0.020498 9.222188 Bathyberth vs Tomptho 0.005210 0.011240 0.463563 Bathyberth vs Tylodin 0.087881 0.015807 5.559583 Bathyberth vs Umbracu 0.086518 0.015632 5.534670 Bathyberth vs Calliop 0.037632 0.017066 2.205153 Bathyberth vs Limapon 0.074037 0.015962 4.638324 Bathyberth vs Thuridi 0.080120 0.016069 4.986063 Bathyberth vs Boselli 0.082587 0.015917 5.188579 Bathyberth vs Cyerce_ 0.080280 0.015947 5.034277 Bathyberth vs Elysia_ 0.061228 0.015939 3.841289 Bathyberth vs Placobr 0.078859 0.016014 4.924484 Bathyberth vs Haminoe 0.079049 0.015874 4.979809 Bathyberth vs Phanero 0.085257 0.015718 5.424114 Bathyberth vs Atys_se 0.089784 0.015760 5.696827

223
Bathyberth vs cf._Ret 0.079774 0.016240 4.912263 Bathyberth vs Philino 0.078521 0.016360 4.799577 Bathyberth vs Chelido 0.085024 0.016560 5.134189 Bathyberth vs Odontog 0.080036 0.016641 4.809692 Bathyberth vs Navanax 0.085082 0.016229 5.242725 Bathyberth vs Philino 0.061308 0.016253 3.772071 Bathyberth vs Bulla_c 0.075291 0.015948 4.721027 Bathyberth vs Runcina 0.044241 0.016825 2.629420 Bathyberth vs Sagamin 0.067280 0.016409 4.100084 Bathyberth vs Pupa_so 0.061429 0.016189 3.794505 Bathyberth vs Hydatin 0.058756 0.015804 3.717785 Bathyberth vs Aplysia 0.083809 0.016289 5.145289 Bathyberth vs Dolabel 0.089992 0.015899 5.660260 Bathyberth vs Akera_b 0.074620 0.016068 4.643995 Bathyberth vs Unela_g 0.083760 0.016125 5.194251 Bathyberth vs Parahed 0.083760 0.016125 5.194251 Bathyberth vs Pontohe 0.080087 0.016124 4.966820 Bathyberth vs Hedylop 0.093848 0.015534 6.041245 Tompthomps vs Tylodin 0.082292 0.014885 5.528625 Tompthomps vs Umbracu 0.082591 0.015220 5.426379 Tompthomps vs Calliop 0.035081 0.017038 2.058986 Tompthomps vs Limapon 0.071351 0.015834 4.506101 Tompthomps vs Thuridi 0.077382 0.016102 4.805752 Tompthomps vs Boselli 0.079846 0.015950 5.005889 Tompthomps vs Cyerce_ 0.076270 0.016010 4.763999 Tompthomps vs Elysia_ 0.058516 0.016014 3.654073 Tompthomps vs Placobr 0.077334 0.016102 4.802755 Tompthomps vs Haminoe 0.078918 0.015789 4.998382 Tompthomps vs Phanero 0.082731 0.015568 5.314156 Tompthomps vs Atys_se 0.086048 0.015363 5.601048 Tompthomps vs cf._Ret 0.078401 0.015982 4.905632 Tompthomps vs Philino 0.078562 0.015592 5.038586 Tompthomps vs Chelido 0.086170 0.015805 5.452165 Tompthomps vs Odontog 0.082401 0.015876 5.190366 Tompthomps vs Navanax 0.086175 0.015605 5.522228 Tompthomps vs Philino 0.063755 0.015585 4.090772 Tompthomps vs Bulla_c 0.073955 0.015819 4.675002 Tompthomps vs Runcina 0.041551 0.016034 2.591491 Tompthomps vs Sagamin 0.064540 0.015975 4.040133 Tompthomps vs Pupa_so 0.057572 0.015678 3.672163 Tompthomps vs Hydatin 0.053499 0.015657 3.417013 Tompthomps vs Aplysia 0.081165 0.015884 5.109756 Tompthomps vs Dolabel 0.084562 0.015106 5.598018 Tompthomps vs Akera_b 0.066631 0.015407 4.324628 Tompthomps vs Unela_g 0.079950 0.015600 5.124919 Tompthomps vs Parahed 0.079950 0.015600 5.124919 Tompthomps vs Pontohe 0.076270 0.015662 4.869702 Tompthomps vs Hedylop 0.087351 0.015132 5.772434 Tylodina_p vs Umbracu 0.001346 0.009543 0.141062 Tylodina_p vs Calliop -0.041941 0.014016 -2.992321 Tylodina_p vs Limapon -0.007216 0.012694 -0.568511 Tylodina_p vs Thuridi 0.000103 0.012235 0.008416 Tylodina_p vs Boselli 0.001299 0.012045 0.107879 Tylodina_p vs Cyerce_ -0.002457 0.012159 -0.202051 Tylodina_p vs Elysia_ -0.019776 0.013276 -1.489658 Tylodina_p vs Placobr -0.001197 0.012169 -0.098406 Tylodina_p vs Haminoe -0.008290 0.012460 -0.665348 Tylodina_p vs Phanero -0.003146 0.012080 -0.260451 Tylodina_p vs Atys_se 0.003923 0.011844 0.331203 Tylodina_p vs cf._Ret -0.006289 0.012220 -0.514683 Tylodina_p vs Philino -0.009974 0.012655 -0.788174 Tylodina_p vs Chelido -0.002401 0.012402 -0.193631 Tylodina_p vs Odontog -0.006185 0.012665 -0.488333 Tylodina_p vs Navanax -0.002566 0.012640 -0.203009 Tylodina_p vs Philino -0.021056 0.012972 -1.623210 Tylodina_p vs Bulla_c -0.014813 0.012474 -1.187475 Tylodina_p vs Runcina -0.040400 0.013902 -2.906028 Tylodina_p vs Sagamin -0.018845 0.012875 -1.463751 Tylodina_p vs Pupa_so -0.025939 0.012698 -2.042733 Tylodina_p vs Hydatin -0.029905 0.012938 -2.311425 Tylodina_p vs Aplysia 0.003883 0.011658 0.333123 Tylodina_p vs Dolabel 0.008737 0.011958 0.730613 Tylodina_p vs Akera_b -0.007748 0.012078 -0.641498 Tylodina_p vs Unela_g 0.001350 0.011217 0.120363 Tylodina_p vs Parahed 0.001350 0.011217 0.120363 Tylodina_p vs Pontohe -0.007277 0.011752 -0.619212 Tylodina_p vs Hedylop 0.006330 0.010896 0.580878 Umbraculum vs Calliop -0.040657 0.013418 -3.030004

224
Umbraculum vs Limapon -0.004895 0.012265 -0.399138 Umbraculum vs Thuridi 0.002390 0.011592 0.206182 Umbraculum vs Boselli 0.003582 0.011473 0.312217 Umbraculum vs Cyerce_ -0.000145 0.011385 -0.012768 Umbraculum vs Elysia_ -0.016167 0.012354 -1.308649 Umbraculum vs Placobr -0.000094 0.011678 -0.008080 Umbraculum vs Haminoe -0.005969 0.011905 -0.501392 Umbraculum vs Phanero -0.003268 0.011608 -0.281507 Umbraculum vs Atys_se 0.003764 0.011134 0.338087 Umbraculum vs cf._Ret -0.005172 0.011518 -0.449022 Umbraculum vs Philino -0.008854 0.012160 -0.728141 Umbraculum vs Chelido -0.001302 0.011708 -0.111220 Umbraculum vs Odontog -0.005076 0.011990 -0.423313 Umbraculum vs Navanax -0.000213 0.011770 -0.018069 Umbraculum vs Philino -0.022431 0.012520 -1.791595 Umbraculum vs Bulla_c -0.012349 0.011799 -1.046605 Umbraculum vs Runcina -0.039205 0.012988 -3.018615 Umbraculum vs Sagamin -0.016432 0.011734 -1.400434 Umbraculum vs Pupa_so -0.024635 0.012444 -1.979727 Umbraculum vs Hydatin -0.029877 0.012251 -2.438710 Umbraculum vs Aplysia 0.002576 0.010651 0.241846 Umbraculum vs Dolabel 0.007417 0.010841 0.684129 Umbraculum vs Akera_b -0.006654 0.011342 -0.586653 Umbraculum vs Unela_g 0.002393 0.010926 0.219019 Umbraculum vs Parahed 0.002393 0.010926 0.219019 Umbraculum vs Pontohe -0.006258 0.011695 -0.535076 Umbraculum vs Hedylop 0.009829 0.010615 0.925927 Calliopaea vs Limapon 0.035891 0.009102 3.943361 Calliopaea vs Thuridi 0.043236 0.010513 4.112710 Calliopaea vs Boselli 0.044430 0.010151 4.377026 Calliopaea vs Cyerce_ 0.040803 0.010618 3.842936 Calliopaea vs Elysia_ 0.024565 0.011206 2.192102 Calliopaea vs Placobr 0.040748 0.010655 3.824268 Calliopaea vs Haminoe 0.037204 0.014025 2.652741 Calliopaea vs Phanero 0.043508 0.013826 3.146721 Calliopaea vs Atys_se 0.050553 0.013385 3.776867 Calliopaea vs cf._Ret 0.037930 0.013957 2.717620 Calliopaea vs Philino 0.035508 0.014479 2.452358 Calliopaea vs Chelido 0.043210 0.014463 2.987677 Calliopaea vs Odontog 0.039441 0.014909 2.645536 Calliopaea vs Navanax 0.041756 0.014340 2.911933 Calliopaea vs Philino 0.022113 0.014418 1.533722 Calliopaea vs Bulla_c 0.032142 0.014460 2.222839 Calliopaea vs Runcina 0.004088 0.014930 0.273802 Calliopaea vs Sagamin 0.025662 0.013864 1.850961 Calliopaea vs Pupa_so 0.017406 0.014285 1.218451 Calliopaea vs Hydatin 0.012157 0.014034 0.866252 Calliopaea vs Aplysia 0.045763 0.013830 3.309096 Calliopaea vs Dolabel 0.049306 0.013601 3.625210 Calliopaea vs Akera_b 0.034147 0.014071 2.426729 Calliopaea vs Unela_g 0.043148 0.012958 3.329930 Calliopaea vs Parahed 0.043148 0.012958 3.329930 Calliopaea vs Pontohe 0.035702 0.013403 2.663797 Calliopaea vs Hedylop 0.053105 0.012512 4.244183 Limapontia vs Thuridi 0.007350 0.008407 0.874268 Limapontia vs Boselli 0.008544 0.007759 1.101101 Limapontia vs Cyerce_ 0.004819 0.008526 0.565283 Limapontia vs Elysia_ -0.010044 0.009402 -1.068261 Limapontia vs Placobr 0.004859 0.008492 0.572187 Limapontia vs Haminoe 0.000093 0.012337 0.007560 Limapontia vs Phanero 0.007649 0.012258 0.623961 Limapontia vs Atys_se 0.014655 0.011842 1.237533 Limapontia vs cf._Ret 0.003279 0.012227 0.268150 Limapontia vs Philino -0.000442 0.012607 -0.035069 Limapontia vs Chelido 0.005966 0.012812 0.465639 Limapontia vs Odontog 0.002197 0.013295 0.165235 Limapontia vs Navanax 0.004598 0.012570 0.365766 Limapontia vs Philino -0.013902 0.012587 -1.104504 Limapontia vs Bulla_c -0.003901 0.012525 -0.311419 Limapontia vs Runcina -0.031969 0.013644 -2.342990 Limapontia vs Sagamin -0.010361 0.012526 -0.827147 Limapontia vs Pupa_so -0.017313 0.012648 -1.368774 Limapontia vs Hydatin -0.021310 0.012669 -1.681991 Limapontia vs Aplysia 0.011088 0.012032 0.921524 Limapontia vs Dolabel 0.017139 0.011912 1.438779 Limapontia vs Akera_b -0.000531 0.012387 -0.042840 Limapontia vs Unela_g 0.008539 0.011242 0.759553 Limapontia vs Parahed 0.008539 0.011242 0.759553

225
Limapontia vs Pontohe -0.000102 0.011930 -0.008512 Limapontia vs Hedylop 0.015877 0.010651 1.490693 Thuridilla vs Boselli 0.001193 0.005197 0.229510 Thuridilla vs Cyerce_ -0.002545 0.006480 -0.392741 Thuridilla vs Elysia_ -0.018621 0.007952 -2.341675 Thuridilla vs Placobr -0.002488 0.005767 -0.431348 Thuridilla vs Haminoe -0.004812 0.011876 -0.405147 Thuridilla vs Phanero 0.001535 0.011903 0.128930 Thuridilla vs Atys_se 0.008525 0.011010 0.774305 Thuridilla vs cf._Ret -0.002829 0.012139 -0.233072 Thuridilla vs Philino -0.006549 0.012718 -0.514956 Thuridilla vs Chelido 0.001048 0.012404 0.084469 Thuridilla vs Odontog -0.002716 0.012837 -0.211597 Thuridilla vs Navanax -0.000309 0.012032 -0.025670 Thuridilla vs Philino -0.018796 0.012490 -1.504833 Thuridilla vs Bulla_c -0.010019 0.012161 -0.823821 Thuridilla vs Runcina -0.039280 0.014031 -2.799455 Thuridilla vs Sagamin -0.015258 0.012147 -1.256151 Thuridilla vs Pupa_so -0.024643 0.012293 -2.004580 Thuridilla vs Hydatin -0.028636 0.012430 -2.303896 Thuridilla vs Aplysia 0.002475 0.011223 0.220545 Thuridilla vs Dolabel 0.008506 0.011524 0.738111 Thuridilla vs Akera_b -0.007872 0.012089 -0.651197 Thuridilla vs Unela_g 0.002433 0.011060 0.220017 Thuridilla vs Parahed 0.002433 0.011060 0.220017 Thuridilla vs Pontohe -0.006196 0.011425 -0.542319 Thuridilla vs Hedylop 0.009765 0.010533 0.927071 Bosellia_m vs Cyerce_ -0.003741 0.005623 -0.665247 Bosellia_m vs Elysia_ -0.019877 0.007480 -2.657232 Bosellia_m vs Placobr -0.003680 0.005592 -0.658142 Bosellia_m vs Haminoe -0.006009 0.011496 -0.522713 Bosellia_m vs Phanero 0.000338 0.011648 0.029055 Bosellia_m vs Atys_se 0.007327 0.010873 0.673919 Bosellia_m vs cf._Ret -0.004025 0.011755 -0.342458 Bosellia_m vs Philino -0.007747 0.012411 -0.624192 Bosellia_m vs Chelido -0.000151 0.012280 -0.012321 Bosellia_m vs Odontog -0.003915 0.012777 -0.306455 Bosellia_m vs Navanax -0.000263 0.011786 -0.022283 Bosellia_m vs Philino -0.019995 0.012314 -1.623791 Bosellia_m vs Bulla_c -0.010006 0.012019 -0.832575 Bosellia_m vs Runcina -0.039230 0.013968 -2.808558 Bosellia_m vs Sagamin -0.016457 0.012032 -1.367781 Bosellia_m vs Pupa_so -0.024613 0.012098 -2.034375 Bosellia_m vs Hydatin -0.028601 0.012235 -2.337617 Bosellia_m vs Aplysia 0.002472 0.011342 0.217959 Bosellia_m vs Dolabel 0.008496 0.011585 0.733333 Bosellia_m vs Akera_b -0.007862 0.011972 -0.656703 Bosellia_m vs Unela_g 0.002430 0.010933 0.222292 Bosellia_m vs Parahed 0.002430 0.010933 0.222292 Bosellia_m vs Pontohe -0.006189 0.011290 -0.548139 Bosellia_m vs Hedylop 0.008514 0.010380 0.820245 Cyerce_nig vs Elysia_ -0.016127 0.008535 -1.889465 Cyerce_nig vs Placobr 0.000051 0.005981 0.008583 Cyerce_nig vs Haminoe -0.003524 0.011916 -0.295718 Cyerce_nig vs Phanero 0.002837 0.011754 0.241351 Cyerce_nig vs Atys_se 0.009845 0.010908 0.902601 Cyerce_nig vs cf._Ret -0.001539 0.011792 -0.130544 Cyerce_nig vs Philino -0.005270 0.012773 -0.412608 Cyerce_nig vs Chelido 0.002351 0.012643 0.185966 Cyerce_nig vs Odontog -0.001422 0.013012 -0.109322 Cyerce_nig vs Navanax 0.002239 0.012336 0.181518 Cyerce_nig vs Philino -0.017542 0.012656 -1.386043 Cyerce_nig vs Bulla_c -0.008733 0.011992 -0.728276 Cyerce_nig vs Runcina -0.038083 0.013852 -2.749323 Cyerce_nig vs Sagamin -0.013996 0.011918 -1.174311 Cyerce_nig vs Pupa_so -0.022156 0.012350 -1.794033 Cyerce_nig vs Hydatin -0.026161 0.012357 -2.117110 Cyerce_nig vs Aplysia 0.005026 0.011851 0.424071 Cyerce_nig vs Dolabel 0.011074 0.011898 0.930730 Cyerce_nig vs Akera_b -0.005348 0.012455 -0.429403 Cyerce_nig vs Unela_g 0.004988 0.011445 0.435785 Cyerce_nig vs Parahed 0.004988 0.011445 0.435785 Cyerce_nig vs Pontohe -0.003663 0.011869 -0.308657 Cyerce_nig vs Hedylop 0.012338 0.010930 1.128801 Elysia_vir vs Placobr 0.016137 0.007962 2.026844 Elysia_vir vs Haminoe 0.011349 0.012063 0.940868 Elysia_vir vs Phanero 0.018912 0.012263 1.542260 Elysia_vir vs Atys_se 0.025931 0.011653 2.225288

226
Elysia_vir vs cf._Ret 0.013314 0.012376 1.075725 Elysia_vir vs Philino 0.010800 0.013079 0.825758 Elysia_vir vs Chelido 0.017236 0.012908 1.335259 Elysia_vir vs Odontog 0.013467 0.013271 1.014798 Elysia_vir vs Navanax 0.018378 0.012464 1.474511 Elysia_vir vs Philino -0.002632 0.012434 -0.211675 Elysia_vir vs Bulla_c 0.008651 0.012391 0.698151 Elysia_vir vs Runcina -0.019365 0.014488 -1.336572 Elysia_vir vs Sagamin 0.002197 0.012301 0.178567 Elysia_vir vs Pupa_so -0.004695 0.013141 -0.357277 Elysia_vir vs Hydatin -0.008672 0.013147 -0.659583 Elysia_vir vs Aplysia 0.022371 0.012632 1.770953 Elysia_vir vs Dolabel 0.027140 0.012786 2.122648 Elysia_vir vs Akera_b 0.012051 0.012858 0.937257 Elysia_vir vs Unela_g 0.019814 0.012030 1.647008 Elysia_vir vs Parahed 0.019814 0.012030 1.647008 Elysia_vir vs Pontohe 0.012439 0.012323 1.009430 Elysia_vir vs Hedylop 0.027142 0.011361 2.389002 Placobranc vs Haminoe -0.002315 0.012016 -0.192650 Placobranc vs Phanero 0.004028 0.011918 0.338009 Placobranc vs Atys_se 0.011022 0.011088 0.994070 Placobranc vs cf._Ret -0.000336 0.012338 -0.027192 Placobranc vs Philino -0.004056 0.012730 -0.318595 Placobranc vs Chelido 0.003548 0.012293 0.288598 Placobranc vs Odontog -0.000216 0.012728 -0.016997 Placobranc vs Navanax 0.002185 0.012169 0.179548 Placobranc vs Philino -0.016296 0.012561 -1.297314 Placobranc vs Bulla_c -0.007519 0.012170 -0.617809 Placobranc vs Runcina -0.036789 0.014108 -2.607751 Placobranc vs Sagamin -0.012758 0.012280 -1.038897 Placobranc vs Pupa_so -0.022152 0.012425 -1.782926 Placobranc vs Hydatin -0.026146 0.012686 -2.061019 Placobranc vs Aplysia 0.004963 0.011618 0.427158 Placobranc vs Dolabel 0.010997 0.011858 0.927357 Placobranc vs Akera_b -0.004136 0.012169 -0.339871 Placobranc vs Unela_g 0.004924 0.011405 0.431723 Placobranc vs Parahed 0.004924 0.011405 0.431723 Placobranc vs Pontohe -0.003706 0.011566 -0.320390 Placobranc vs Hedylop 0.012256 0.010892 1.125238 Haminoea_h vs Phanero 0.006349 0.007986 0.794951 Haminoea_h vs Atys_se 0.013353 0.007277 1.835044 Haminoea_h vs cf._Ret 0.001979 0.008698 0.227474 Haminoea_h vs Philino -0.000493 0.009923 -0.049642 Haminoea_h vs Chelido 0.005865 0.009217 0.636377 Haminoea_h vs Odontog 0.002101 0.009886 0.212545 Haminoea_h vs Navanax 0.004552 0.009143 0.497835 Haminoea_h vs Philino -0.013978 0.010014 -1.395852 Haminoea_h vs Bulla_c -0.005158 0.010049 -0.513341 Haminoea_h vs Runcina -0.032025 0.012941 -2.474809 Haminoea_h vs Sagamin -0.009283 0.009791 -0.948140 Haminoea_h vs Pupa_so -0.017352 0.012335 -1.406732 Haminoea_h vs Hydatin -0.022593 0.012329 -1.832576 Haminoea_h vs Aplysia 0.009747 0.011353 0.858567 Haminoea_h vs Dolabel 0.016947 0.011102 1.526508 Haminoea_h vs Akera_b 0.000584 0.010944 0.053337 Haminoea_h vs Unela_g 0.008446 0.010979 0.769275 Haminoea_h vs Parahed 0.008446 0.010979 0.769275 Haminoea_h vs Pontohe -0.001401 0.010780 -0.129930 Haminoea_h vs Hedylop 0.014573 0.010629 1.371084 Phaneropht vs Atys_se 0.006988 0.006665 1.048484 Phaneropht vs cf._Ret -0.004364 0.009066 -0.481342 Phaneropht vs Philino -0.008046 0.010144 -0.793100 Phaneropht vs Chelido -0.000491 0.010074 -0.048776 Phaneropht vs Odontog -0.004256 0.010255 -0.414973 Phaneropht vs Navanax -0.000604 0.009509 -0.063531 Phaneropht vs Philino -0.020335 0.010074 -2.018578 Phaneropht vs Bulla_c -0.011519 0.010508 -1.096195 Phaneropht vs Runcina -0.038333 0.012805 -2.993695 Phaneropht vs Sagamin -0.015634 0.009390 -1.665001 Phaneropht vs Pupa_so -0.024978 0.012596 -1.983057 Phaneropht vs Hydatin -0.028971 0.012365 -2.342997 Phaneropht vs Aplysia 0.002146 0.010723 0.200124 Phaneropht vs Dolabel 0.010627 0.011225 0.946745 Phaneropht vs Akera_b -0.004546 0.011013 -0.412830 Phaneropht vs Unela_g 0.000902 0.011303 0.079825 Phaneropht vs Parahed 0.000902 0.011303 0.079825 Phaneropht vs Pontohe -0.007739 0.011370 -0.680677 Phaneropht vs Hedylop 0.007062 0.010877 0.649266

227
Atys_semis vs cf._Ret -0.011358 0.009014 -1.260022 Atys_semis vs Philino -0.012590 0.009410 -1.337896 Atys_semis vs Chelido -0.006247 0.009487 -0.658480 Atys_semis vs Odontog -0.010006 0.009518 -1.051364 Atys_semis vs Navanax -0.007553 0.008383 -0.900944 Atys_semis vs Philino -0.026076 0.009861 -2.644331 Atys_semis vs Bulla_c -0.017348 0.009555 -1.815561 Atys_semis vs Runcina -0.044127 0.012351 -3.572879 Atys_semis vs Sagamin -0.021387 0.008546 -2.502616 Atys_semis vs Pupa_so -0.030712 0.012146 -2.528645 Atys_semis vs Hydatin -0.035935 0.011967 -3.002730 Atys_semis vs Aplysia -0.004847 0.010176 -0.476352 Atys_semis vs Dolabel 0.002378 0.010549 0.225433 Atys_semis vs Akera_b -0.012763 0.010236 -1.246826 Atys_semis vs Unela_g -0.006085 0.010907 -0.557901 Atys_semis vs Parahed -0.006085 0.010907 -0.557901 Atys_semis vs Pontohe -0.014737 0.010860 -1.357082 Atys_semis vs Hedylop 0.000040 0.010412 0.003824 cf._Retusa vs Philino -0.002467 0.010228 -0.241201 cf._Retusa vs Chelido 0.003885 0.009759 0.398081 cf._Retusa vs Odontog 0.000121 0.010476 0.011529 cf._Retusa vs Navanax 0.003778 0.009816 0.384901 cf._Retusa vs Philino -0.015958 0.009553 -1.670481 cf._Retusa vs Bulla_c -0.005925 0.009806 -0.604254 cf._Retusa vs Runcina -0.033963 0.013235 -2.566126 cf._Retusa vs Sagamin -0.011277 0.010027 -1.124678 cf._Retusa vs Pupa_so -0.020646 0.013103 -1.575636 cf._Retusa vs Hydatin -0.025907 0.013046 -1.985899 cf._Retusa vs Aplysia 0.007762 0.011613 0.668353 cf._Retusa vs Dolabel 0.015006 0.011368 1.320074 cf._Retusa vs Akera_b -0.001398 0.010627 -0.131597 cf._Retusa vs Unela_g 0.005266 0.011032 0.477367 cf._Retusa vs Parahed 0.005266 0.011032 0.477367 cf._Retusa vs Pontohe -0.003375 0.011755 -0.287146 cf._Retusa vs Hedylop 0.010122 0.010615 0.953501 Philinopsi vs Chelido 0.006345 0.007492 0.846786 Philinopsi vs Odontog 0.002585 0.008059 0.320793 Philinopsi vs Navanax 0.004974 0.006712 0.741109 Philinopsi vs Philino -0.014694 0.010604 -1.385749 Philinopsi vs Bulla_c -0.004663 0.010676 -0.436758 Philinopsi vs Runcina -0.031499 0.012637 -2.492622 Philinopsi vs Sagamin -0.010013 0.009937 -1.007625 Philinopsi vs Pupa_so -0.016914 0.012966 -1.304453 Philinopsi vs Hydatin -0.019645 0.012752 -1.540470 Philinopsi vs Aplysia 0.009026 0.011440 0.788981 Philinopsi vs Dolabel 0.016274 0.010998 1.479667 Philinopsi vs Akera_b 0.001073 0.011297 0.094953 Philinopsi vs Unela_g 0.008988 0.011776 0.763231 Philinopsi vs Parahed 0.008988 0.011776 0.763231 Philinopsi vs Pontohe 0.000344 0.011970 0.028700 Philinopsi vs Hedylop 0.013821 0.011266 1.226706 Chelidonur vs Odontog -0.003759 0.005617 -0.669251 Chelidonur vs Navanax -0.001360 0.006514 -0.208759 Chelidonur vs Philino -0.021045 0.009930 -2.119204 Chelidonur vs Bulla_c -0.012234 0.010507 -1.164353 Chelidonur vs Runcina -0.039095 0.013200 -2.961818 Chelidonur vs Sagamin -0.016357 0.009639 -1.697011 Chelidonur vs Pupa_so -0.023225 0.013129 -1.769033 Chelidonur vs Hydatin -0.027217 0.012974 -2.097821 Chelidonur vs Aplysia 0.002638 0.011225 0.234989 Chelidonur vs Dolabel 0.009861 0.010754 0.916997 Chelidonur vs Akera_b -0.005295 0.011143 -0.475216 Chelidonur vs Unela_g 0.002591 0.011542 0.224440 Chelidonur vs Parahed 0.002591 0.011542 0.224440 Chelidonur vs Pontohe -0.007268 0.011566 -0.628374 Chelidonur vs Hedylop 0.008704 0.011098 0.784302 Odontoglaj vs Navanax 0.002395 0.007015 0.341406 Odontoglaj vs Philino -0.017285 0.010110 -1.709683 Odontoglaj vs Bulla_c -0.008466 0.010771 -0.785926 Odontoglaj vs Runcina -0.035335 0.013273 -2.662127 Odontoglaj vs Sagamin -0.012598 0.009943 -1.266991 Odontoglaj vs Pupa_so -0.019456 0.013283 -1.464753 Odontoglaj vs Hydatin -0.023448 0.013068 -1.794383 Odontoglaj vs Aplysia 0.006402 0.011260 0.568543 Odontoglaj vs Dolabel 0.013625 0.011136 1.223527 Odontoglaj vs Akera_b -0.000317 0.011381 -0.027825 Odontoglaj vs Unela_g 0.006355 0.012136 0.523634 Odontoglaj vs Parahed 0.006355 0.012136 0.523634

228
Odontoglaj vs Pontohe -0.003504 0.011968 -0.292745 Odontoglaj vs Hedylop 0.012463 0.011642 1.070553 Navanax_in vs Philino -0.020960 0.010048 -2.085955 Navanax_in vs Bulla_c -0.010864 0.009766 -1.112432 Navanax_in vs Runcina -0.038984 0.012784 -3.049429 Navanax_in vs Sagamin -0.017468 0.009096 -1.920415 Navanax_in vs Pupa_so -0.024351 0.012727 -1.913281 Navanax_in vs Hydatin -0.027092 0.012572 -2.154896 Navanax_in vs Aplysia 0.001543 0.010815 0.142640 Navanax_in vs Dolabel 0.010009 0.010628 0.941711 Navanax_in vs Akera_b -0.005116 0.010869 -0.470686 Navanax_in vs Unela_g 0.002747 0.011622 0.236366 Navanax_in vs Parahed 0.002747 0.011622 0.236366 Navanax_in vs Pontohe -0.007105 0.011566 -0.614312 Navanax_in vs Hedylop 0.008786 0.011034 0.796315 Philinoglo vs Bulla_c 0.008857 0.010566 0.838229 Philinoglo vs Runcina -0.018050 0.013516 -1.335455 Philinoglo vs Sagamin 0.004700 0.009094 0.516810 Philinoglo vs Pupa_so -0.003425 0.012773 -0.268115 Philinoglo vs Hydatin -0.008697 0.012639 -0.688154 Philinoglo vs Aplysia 0.023703 0.010955 2.163545 Philinoglo vs Dolabel 0.030950 0.010794 2.867246 Philinoglo vs Akera_b 0.015828 0.011114 1.424122 Philinoglo vs Unela_g 0.022434 0.011492 1.952163 Philinoglo vs Parahed 0.022434 0.011492 1.952163 Philinoglo vs Pontohe 0.012575 0.011653 1.079191 Philinoglo vs Hedylop 0.029761 0.011098 2.681548 Bulla_cf._ vs Runcina -0.026882 0.013365 -2.011283 Bulla_cf._ vs Sagamin -0.005370 0.009547 -0.562543 Bulla_cf._ vs Pupa_so -0.013486 0.012501 -1.078809 Bulla_cf._ vs Hydatin -0.018758 0.012576 -1.491603 Bulla_cf._ vs Aplysia 0.013685 0.011587 1.181143 Bulla_cf._ vs Dolabel 0.020962 0.011544 1.815794 Bulla_cf._ vs Akera_b 0.005747 0.011780 0.487881 Bulla_cf._ vs Unela_g 0.012448 0.011539 1.078772 Bulla_cf._ vs Parahed 0.012448 0.011539 1.078772 Bulla_cf._ vs Pontohe 0.003793 0.011981 0.316547 Bulla_cf._ vs Hedylop 0.018556 0.011219 1.653929 Runcina_ad vs Sagamin 0.022838 0.012825 1.780736 Runcina_ad vs Pupa_so 0.014612 0.014553 1.003998 Runcina_ad vs Hydatin 0.010558 0.014365 0.734957 Runcina_ad vs Aplysia 0.041699 0.013046 3.196233 Runcina_ad vs Dolabel 0.047748 0.012896 3.702489 Runcina_ad vs Akera_b 0.032430 0.013110 2.473685 Runcina_ad vs Unela_g 0.039192 0.013560 2.890318 Runcina_ad vs Parahed 0.039192 0.013560 2.890318 Runcina_ad vs Pontohe 0.030549 0.013781 2.216814 Runcina_ad vs Hedylop 0.047719 0.013274 3.594956 Sagaminopt vs Pupa_so -0.005680 0.012546 -0.452740 Sagaminopt vs Hydatin -0.010956 0.012430 -0.881416 Sagaminopt vs Aplysia 0.017746 0.011078 1.601819 Sagaminopt vs Dolabel 0.023758 0.010983 2.163103 Sagaminopt vs Akera_b 0.009851 0.011326 0.869778 Sagaminopt vs Unela_g 0.017701 0.011397 1.553164 Sagaminopt vs Parahed 0.017701 0.011397 1.553164 Sagaminopt vs Pontohe 0.006564 0.011458 0.572903 Sagaminopt vs Hedylop 0.026213 0.011185 2.343674 Pupa_solid vs Hydatin -0.005205 0.005708 -0.911816 Pupa_solid vs Aplysia 0.027048 0.011977 2.258364 Pupa_solid vs Dolabel 0.033147 0.012053 2.750229 Pupa_solid vs Akera_b 0.017934 0.012210 1.468826 Pupa_solid vs Unela_g 0.027031 0.011840 2.283060 Pupa_solid vs Parahed 0.027031 0.011840 2.283060 Pupa_solid vs Pontohe 0.019596 0.012079 1.622225 Pupa_solid vs Hedylop 0.034371 0.011637 2.953643 Hydatina_p vs Aplysia 0.032276 0.011856 2.722306 Hydatina_p vs Dolabel 0.035924 0.011735 3.061417 Hydatina_p vs Akera_b 0.021954 0.012273 1.788812 Hydatina_p vs Unela_g 0.032263 0.011907 2.709564 Hydatina_p vs Parahed 0.032263 0.011907 2.709564 Hydatina_p vs Pontohe 0.024844 0.012025 2.066033 Hydatina_p vs Hedylop 0.039588 0.011879 3.332664 Aplysia_ca vs Dolabel 0.004920 0.007617 0.645971 Aplysia_ca vs Akera_b -0.010300 0.008580 -1.200463 Aplysia_ca vs Unela_g -0.001282 0.010619 -0.120720 Aplysia_ca vs Parahed -0.001282 0.010619 -0.120720 Aplysia_ca vs Pontohe -0.008664 0.010601 -0.817329 Aplysia_ca vs Hedylop 0.007241 0.010421 0.694840

229
Dolabella_ vs Akera_b -0.016463 0.008019 -2.053087 Dolabella_ vs Unela_g -0.007311 0.010822 -0.675574 Dolabella_ vs Parahed -0.007311 0.010822 -0.675574 Dolabella_ vs Pontohe -0.013449 0.010993 -1.223381 Dolabella_ vs Hedylop 0.000039 0.010191 0.003807 Akera_bull vs Unela_g 0.009111 0.010407 0.875444 Akera_bull vs Parahed 0.009111 0.010407 0.875444 Akera_bull vs Pontohe -0.000779 0.010615 -0.073375 Akera_bull vs Hedylop 0.017578 0.010183 1.726247 Unela_glan vs Parahed 0.000000 0.000000 Not deter. Unela_glan vs Pontohe -0.008619 0.007609 -1.132798 Unela_glan vs Hedylop 0.007332 0.008539 0.858717 Parahedyle vs Pontohe -0.008619 0.007609 -1.132798 Parahedyle vs Hedylop 0.007332 0.008539 0.858717 Pontohedyl vs Hedylop 0.015908 0.009555 1.664940
Opisthobranchia-Datensatz (16S rDNA) Ergebnisse der Analyse der relativen Substitutionsraten auf der Grundlage des 16S rDNA-Datensatzes mit Odostomia sp. als Außengruppe (1). Eine signifikant unterschiedliche Substitutionsgeschwindigkeit ist gegeben, wenn der „Z-score“ (k12-k13/SD) höher als 1,96 ist. Positive „Z-score“-Werte bedeuten, dass Taxon (2) eine größere Evolutionsgeschwindigkeit aufweist; negative Werte stehen für eine erhöhte Substitutionsrate des Taxons (3). (2) (3) K12 - K13 SD Z-score Bathydoris vs Archido 0.010849 0.044953 0.241343 Bathydoris vs Goniodo 0.033677 0.039296 0.857019 Bathydoris vs Chromod 0.032907 0.042391 0.776267 Bathydoris vs Discodo 0.008775 0.040541 0.216453 Bathydoris vs Hypselo 0.008593 0.046664 0.184138 Bathydoris vs Dendrod 0.004674 0.055026 0.084938 Bathydoris vs Eubranc 0.020003 0.042029 0.475929 Bathydoris vs Flabell 0.045486 0.040465 1.124072 Bathydoris vs Dendron 0.030369 0.041881 0.725135 Bathydoris vs Berthel 0.025524 0.048175 0.529807 Bathydoris vs Berthel 0.029526 0.045433 0.649883 Bathydoris vs Pleurob 0.008593 0.046596 0.184405 Bathydoris vs Tomthom 0.040631 0.045206 0.898814 Bathydoris vs Umbracu 0.081944 0.044270 1.850998 Bathydoris vs Aplysia 0.088174 0.043832 2.011625 Bathydoris vs Dolabel 0.066650 0.045109 1.477533 Bathydoris vs Styloch 0.065793 0.042492 1.548363 Bathydoris vs Bursate 0.054815 0.045106 1.215255 Bathydoris vs Dolabri 0.071675 0.044838 1.598541 Bathydoris vs Phyllap 0.059704 0.044803 1.332587 Bathydoris vs Petalif 0.060728 0.044986 1.349917 Bathydoris vs Notarch 0.067154 0.044401 1.512447 Bathydoris vs Akera_b 0.073943 0.044756 1.652139 Bathydoris vs Cavolin 0.007807 0.054115 0.144274 Bathydoris vs Clione_ -0.059270 0.054413 -1.089272 Bathydoris vs Diaphan 0.060686 0.044344 1.368526 Bathydoris vs Haminoe 0.034379 0.044465 0.773161 Bathydoris vs Smaragd 0.010558 0.045309 0.233014 Bathydoris vs Bulla_g -0.002908 0.048155 -0.060387 Bathydoris vs Philine 0.060221 0.044436 1.355215 Bathydoris vs Scaphan 0.006304 0.050071 0.125905 Bathydoris vs Elysia_ 0.095864 0.040390 2.373445 Bathydoris vs Thuridi 0.116240 0.040789 2.849775 Bathydoris vs Placida 0.109906 0.043761 2.511524 Bathydoris vs Acteon_ 0.051140 0.043404 1.178231 Bathydoris vs Pupa_so 0.019116 0.046156 0.414165 Bathydoris vs Hydatin 0.113423 0.041303 2.746126 Bathydoris vs Pontohe 0.045597 0.051025 0.893629 Bathydoris vs Parahed 0.125594 0.047198 2.661015 Bathydoris vs Unela_g 0.125594 0.047198 2.661015 Archidoris vs Goniodo 0.025502 0.031436 0.811230 Archidoris vs Chromod 0.018613 0.033072 0.562810 Archidoris vs Discodo -0.004575 0.034159 -0.133920

230
Archidoris vs Hypselo -0.019822 0.038512 -0.514703 Archidoris vs Dendrod 0.010658 0.052514 0.202957 Archidoris vs Eubranc 0.012277 0.048853 0.251306 Archidoris vs Flabell 0.036822 0.046478 0.792245 Archidoris vs Dendron 0.022136 0.046517 0.475863 Archidoris vs Berthel 0.032242 0.050728 0.635593 Archidoris vs Berthel 0.023012 0.047402 0.485464 Archidoris vs Pleurob 0.003141 0.046049 0.068204 Archidoris vs Tomthom 0.033682 0.043920 0.766895 Archidoris vs Umbracu 0.066883 0.044088 1.517032 Archidoris vs Aplysia 0.073401 0.043558 1.685145 Archidoris vs Dolabel 0.053305 0.045832 1.163051 Archidoris vs Styloch 0.052440 0.046262 1.133539 Archidoris vs Bursate 0.041941 0.045565 0.920464 Archidoris vs Dolabri 0.058090 0.045591 1.274147 Archidoris vs Phyllap 0.046473 0.045400 1.023635 Archidoris vs Petalif 0.047491 0.045646 1.040432 Archidoris vs Notarch 0.053194 0.044430 1.197244 Archidoris vs Akera_b 0.061091 0.044078 1.385977 Archidoris vs Cavolin -0.002582 0.054992 -0.046943 Archidoris vs Clione_ -0.061168 0.054057 -1.131541 Archidoris vs Diaphan 0.048819 0.045115 1.082106 Archidoris vs Haminoe 0.035609 0.049341 0.721693 Archidoris vs Smaragd -0.007874 0.049240 -0.159914 Archidoris vs Bulla_g -0.014494 0.048442 -0.299195 Archidoris vs Philine 0.060680 0.044533 1.362605 Archidoris vs Scaphan 0.001667 0.047776 0.034896 Archidoris vs Elysia_ 0.099924 0.044335 2.253840 Archidoris vs Thuridi 0.104687 0.045535 2.299040 Archidoris vs Placida 0.084691 0.043787 1.934182 Archidoris vs Acteon_ 0.032805 0.044292 0.740666 Archidoris vs Pupa_so 0.001378 0.050935 0.027051 Archidoris vs Hydatin 0.102561 0.042657 2.404338 Archidoris vs Pontohe 0.031201 0.051367 0.607412 Archidoris vs Parahed 0.116111 0.044663 2.599707 Archidoris vs Unela_g 0.116111 0.044663 2.599707 Goniodoris vs Chromod -0.006877 0.032518 -0.211471 Goniodoris vs Discodo -0.025027 0.030506 -0.820399 Goniodoris vs Hypselo -0.040804 0.040840 -0.999105 Goniodoris vs Dendrod -0.028323 0.049002 -0.577994 Goniodoris vs Eubranc -0.008598 0.041768 -0.205852 Goniodoris vs Flabell 0.015797 0.039308 0.401881 Goniodoris vs Dendron -0.010379 0.040432 -0.256708 Goniodoris vs Berthel -0.004293 0.045861 -0.093598 Goniodoris vs Berthel -0.009175 0.043407 -0.211380 Goniodoris vs Pleurob -0.029263 0.042543 -0.687832 Goniodoris vs Tomthom 0.001401 0.043312 0.032342 Goniodoris vs Umbracu 0.040394 0.039499 1.022657 Goniodoris vs Aplysia 0.047310 0.039507 1.197501 Goniodoris vs Dolabel 0.026628 0.040977 0.649812 Goniodoris vs Styloch 0.025675 0.041413 0.619986 Goniodoris vs Bursate 0.015133 0.041602 0.363750 Goniodoris vs Dolabri 0.031390 0.041665 0.753391 Goniodoris vs Phyllap 0.019450 0.041321 0.470694 Goniodoris vs Petalif 0.020558 0.041445 0.496036 Goniodoris vs Notarch 0.021023 0.040545 0.518499 Goniodoris vs Akera_b 0.028380 0.038933 0.728955 Goniodoris vs Cavolin -0.018038 0.051543 -0.349969 Goniodoris vs Clione_ -0.083175 0.052994 -1.569519 Goniodoris vs Diaphan 0.027274 0.040506 0.673323 Goniodoris vs Haminoe 0.013159 0.044272 0.297220 Goniodoris vs Smaragd -0.028924 0.043445 -0.665764 Goniodoris vs Bulla_g -0.028998 0.043874 -0.660946 Goniodoris vs Philine 0.033583 0.039899 0.841694 Goniodoris vs Scaphan -0.008340 0.044623 -0.186899 Goniodoris vs Elysia_ 0.079757 0.040646 1.962222 Goniodoris vs Thuridi 0.083612 0.042049 1.988436 Goniodoris vs Placida 0.075709 0.042990 1.761071 Goniodoris vs Acteon_ 0.012175 0.038952 0.312559 Goniodoris vs Pupa_so -0.019664 0.048153 -0.408362 Goniodoris vs Hydatin 0.087416 0.039845 2.193925 Goniodoris vs Pontohe 0.003742 0.047349 0.079023 Goniodoris vs Parahed 0.094619 0.042233 2.240386 Goniodoris vs Unela_g 0.094619 0.042233 2.240386 Chromodori vs Discodo -0.012131 0.035620 -0.340559 Chromodori vs Hypselo -0.027504 0.037666 -0.730210 Chromodori vs Dendrod -0.010332 0.050632 -0.204063 Chromodori vs Eubranc -0.007124 0.046187 -0.154238

231
Chromodori vs Flabell 0.017421 0.043009 0.405051 Chromodori vs Dendron -0.003689 0.041630 -0.088618 Chromodori vs Berthel 0.003321 0.048258 0.068823 Chromodori vs Berthel 0.003833 0.045482 0.084278 Chromodori vs Pleurob -0.016134 0.044942 -0.358994 Chromodori vs Tomthom 0.014333 0.043910 0.326409 Chromodori vs Umbracu 0.047977 0.043593 1.100569 Chromodori vs Aplysia 0.054904 0.043341 1.266811 Chromodori vs Dolabel 0.034353 0.044235 0.776618 Chromodori vs Styloch 0.027641 0.044581 0.620026 Chromodori vs Bursate 0.017110 0.044414 0.385242 Chromodori vs Dolabri 0.033389 0.042041 0.794186 Chromodori vs Phyllap 0.021364 0.042813 0.498997 Chromodori vs Petalif 0.022551 0.044660 0.504959 Chromodori vs Notarch 0.034176 0.043341 0.788531 Chromodori vs Akera_b 0.035944 0.041103 0.874466 Chromodori vs Cavolin -0.015849 0.052390 -0.302516 Chromodori vs Clione_ -0.074638 0.052273 -1.427864 Chromodori vs Diaphan 0.035202 0.043387 0.811364 Chromodori vs Haminoe 0.027494 0.045702 0.601591 Chromodori vs Smaragd -0.020882 0.044634 -0.467843 Chromodori vs Bulla_g -0.020838 0.046044 -0.452577 Chromodori vs Philine 0.047014 0.043827 1.072726 Chromodori vs Scaphan -0.000020 0.048421 -0.000423 Chromodori vs Elysia_ 0.097961 0.043595 2.247069 Chromodori vs Thuridi 0.096733 0.044483 2.174616 Chromodori vs Placida 0.088666 0.045819 1.935138 Chromodori vs Acteon_ 0.025055 0.042477 0.589841 Chromodori vs Pupa_so -0.022531 0.049655 -0.453761 Chromodori vs Hydatin 0.077589 0.042303 1.834135 Chromodori vs Pontohe 0.012346 0.049004 0.251943 Chromodori vs Parahed 0.091876 0.042004 2.187321 Chromodori vs Unela_g 0.091876 0.042004 2.187321 Discodoris vs Hypselo -0.017118 0.041366 -0.413826 Discodoris vs Dendrod -0.004209 0.052412 -0.080313 Discodoris vs Eubranc 0.016912 0.046947 0.360244 Discodoris vs Flabell 0.042234 0.044294 0.953497 Discodoris vs Dendron 0.014022 0.045806 0.306126 Discodoris vs Berthel 0.010807 0.049673 0.217566 Discodoris vs Berthel 0.008832 0.046586 0.189578 Discodoris vs Pleurob -0.011407 0.045254 -0.252075 Discodoris vs Tomthom 0.020839 0.043743 0.476394 Discodoris vs Umbracu 0.066741 0.042261 1.579256 Discodoris vs Aplysia 0.074190 0.043282 1.714094 Discodoris vs Dolabel 0.053522 0.043452 1.231760 Discodoris vs Styloch 0.052510 0.044092 1.190911 Discodoris vs Bursate 0.041474 0.043478 0.953899 Discodoris vs Dolabri 0.058498 0.043602 1.341636 Discodoris vs Phyllap 0.046143 0.043604 1.058224 Discodoris vs Petalif 0.047327 0.044773 1.057050 Discodoris vs Notarch 0.053424 0.044689 1.195453 Discodoris vs Akera_b 0.055138 0.042127 1.308829 Discodoris vs Cavolin 0.001468 0.055404 0.026497 Discodoris vs Clione_ -0.066681 0.054142 -1.231589 Discodoris vs Diaphan 0.053829 0.041542 1.295759 Discodoris vs Haminoe 0.038747 0.044622 0.868336 Discodoris vs Smaragd 0.000893 0.047476 0.018820 Discodoris vs Bulla_g 0.000940 0.046061 0.020404 Discodoris vs Philine 0.060304 0.042521 1.418217 Discodoris vs Scaphan 0.004964 0.045595 0.108866 Discodoris vs Elysia_ 0.100124 0.042427 2.359937 Discodoris vs Thuridi 0.099863 0.045033 2.217523 Discodoris vs Placida 0.090552 0.044601 2.030301 Discodoris vs Acteon_ 0.042704 0.041298 1.034045 Discodoris vs Pupa_so -0.006712 0.048890 -0.137277 Discodoris vs Hydatin 0.098871 0.041458 2.384826 Discodoris vs Pontohe 0.032290 0.051087 0.632060 Discodoris vs Parahed 0.112698 0.046456 2.425919 Discodoris vs Unela_g 0.112698 0.046456 2.425919 Hypselodor vs Dendrod 0.025594 0.052385 0.488571 Hypselodor vs Eubranc 0.026714 0.050562 0.528340 Hypselodor vs Flabell 0.051410 0.049623 1.036022 Hypselodor vs Dendron 0.023890 0.048389 0.493707 Hypselodor vs Berthel 0.015484 0.050848 0.304516 Hypselodor vs Berthel 0.031457 0.048913 0.643120 Hypselodor vs Pleurob 0.011169 0.047656 0.234362 Hypselodor vs Tomthom 0.042257 0.045727 0.924117 Hypselodor vs Umbracu 0.076026 0.046879 1.621744

232
Hypselodor vs Aplysia 0.082716 0.046318 1.785826 Hypselodor vs Dolabel 0.062343 0.046864 1.330291 Hypselodor vs Styloch 0.055613 0.047958 1.159624 Hypselodor vs Bursate 0.044899 0.047616 0.942942 Hypselodor vs Dolabri 0.061427 0.046930 1.308904 Hypselodor vs Phyllap 0.049449 0.047995 1.030301 Hypselodor vs Petalif 0.050594 0.049484 1.022427 Hypselodor vs Notarch 0.062302 0.047626 1.308141 Hypselodor vs Akera_b 0.075829 0.045430 1.669138 Hypselodor vs Cavolin 0.011745 0.054416 0.215839 Hypselodor vs Clione_ -0.071371 0.054783 -1.302792 Hypselodor vs Diaphan 0.045972 0.046280 0.993351 Hypselodor vs Haminoe 0.032907 0.051102 0.643945 Hypselodor vs Smaragd 0.012904 0.049879 0.258704 Hypselodor vs Bulla_g 0.006536 0.051326 0.127344 Hypselodor vs Philine 0.051332 0.043942 1.168165 Hypselodor vs Scaphan -0.001254 0.049742 -0.025216 Hypselodor vs Elysia_ 0.102895 0.045515 2.260662 Hypselodor vs Thuridi 0.107016 0.044563 2.401438 Hypselodor vs Placida 0.093590 0.046797 1.999888 Hypselodor vs Acteon_ 0.058137 0.046591 1.247816 Hypselodor vs Pupa_so 0.004673 0.050750 0.092079 Hypselodor vs Hydatin 0.112839 0.045713 2.468438 Hypselodor vs Pontohe 0.040742 0.054342 0.749732 Hypselodor vs Parahed 0.109996 0.047937 2.294605 Hypselodor vs Unela_g 0.109996 0.047937 2.294605 Dendrodori vs Eubranc -0.005261 0.054442 -0.096637 Dendrodori vs Flabell 0.026801 0.053110 0.504636 Dendrodori vs Dendron -0.000606 0.054636 -0.011093 Dendrodori vs Berthel -0.005348 0.052833 -0.101222 Dendrodori vs Berthel 0.000736 0.050648 0.014523 Dendrodori vs Pleurob -0.026150 0.051412 -0.508635 Dendrodori vs Tomthom 0.011903 0.050882 0.233935 Dendrodori vs Umbracu 0.047802 0.047552 1.005259 Dendrodori vs Aplysia 0.060266 0.048325 1.247087 Dendrodori vs Dolabel 0.039017 0.050062 0.779363 Dendrodori vs Styloch 0.038237 0.048140 0.794272 Dendrodori vs Bursate 0.021158 0.049254 0.429569 Dendrodori vs Dolabri 0.038237 0.048861 0.782562 Dendrodori vs Phyllap 0.025899 0.050311 0.514776 Dendrodori vs Petalif 0.026920 0.051113 0.526682 Dendrodori vs Notarch 0.038897 0.050047 0.777207 Dendrodori vs Akera_b 0.047587 0.049919 0.953292 Dendrodori vs Cavolin 0.005060 0.053647 0.094319 Dendrodori vs Clione_ -0.054153 0.057904 -0.935217 Dendrodori vs Diaphan 0.040790 0.051382 0.793854 Dendrodori vs Haminoe 0.033756 0.057474 0.587333 Dendrodori vs Smaragd -0.011487 0.054770 -0.209726 Dendrodori vs Bulla_g -0.019870 0.055850 -0.355779 Dendrodori vs Philine 0.040186 0.051857 0.774935 Dendrodori vs Scaphan 0.017770 0.055713 0.318964 Dendrodori vs Elysia_ 0.105856 0.049484 2.139180 Dendrodori vs Thuridi 0.117848 0.048114 2.449353 Dendrodori vs Placida 0.095908 0.052931 1.811956 Dendrodori vs Acteon_ 0.033518 0.050301 0.666355 Dendrodori vs Pupa_so -0.015845 0.054128 -0.292737 Dendrodori vs Hydatin 0.100120 0.046414 2.157120 Dendrodori vs Pontohe 0.002433 0.053829 0.045199 Dendrodori vs Parahed 0.081124 0.048514 1.672168 Dendrodori vs Unela_g 0.081124 0.048872 1.659933 Eubranchus vs Flabell 0.024696 0.026771 0.922521 Eubranchus vs Dendron 0.004405 0.034766 0.126714 Eubranchus vs Berthel 0.013219 0.043128 0.306512 Eubranchus vs Berthel 0.022493 0.041152 0.546591 Eubranchus vs Pleurob 0.002676 0.042404 0.063113 Eubranchus vs Tomthom 0.032958 0.043623 0.755507 Eubranchus vs Umbracu 0.071550 0.046188 1.549106 Eubranchus vs Aplysia 0.067533 0.046625 1.448435 Eubranchus vs Dolabel 0.047351 0.048675 0.972784 Eubranchus vs Styloch 0.040771 0.044699 0.912127 Eubranchus vs Bursate 0.030206 0.046692 0.646923 Eubranchus vs Dolabri 0.046486 0.046366 1.002590 Eubranchus vs Phyllap 0.034664 0.047420 0.731000 Eubranchus vs Petalif 0.035750 0.047475 0.753031 Eubranchus vs Notarch 0.058022 0.047459 1.222582 Eubranchus vs Akera_b 0.065772 0.046383 1.418005 Eubranchus vs Cavolin -0.003873 0.054499 -0.071065 Eubranchus vs Clione_ -0.054597 0.056521 -0.965947

233
Eubranchus vs Diaphan 0.048112 0.045701 1.052745 Eubranchus vs Haminoe 0.027308 0.042681 0.639808 Eubranchus vs Smaragd -0.001823 0.044435 -0.041033 Eubranchus vs Bulla_g -0.014602 0.043627 -0.334698 Eubranchus vs Philine 0.047889 0.043136 1.110176 Eubranchus vs Scaphan 0.006349 0.049799 0.127493 Eubranchus vs Elysia_ 0.108019 0.046419 2.327051 Eubranchus vs Thuridi 0.098150 0.044108 2.225235 Eubranchus vs Placida 0.091287 0.045807 1.992883 Eubranchus vs Acteon_ 0.032736 0.044244 0.739882 Eubranchus vs Pupa_so 0.004546 0.047474 0.095749 Eubranchus vs Hydatin 0.106552 0.044649 2.386451 Eubranchus vs Pontohe 0.017744 0.050981 0.348044 Eubranchus vs Parahed 0.102811 0.048432 2.122776 Eubranchus vs Unela_g 0.102811 0.048432 2.122776 Flabellina vs Dendron -0.020291 0.030327 -0.669075 Flabellina vs Berthel -0.014185 0.042870 -0.330883 Flabellina vs Berthel -0.002354 0.041339 -0.056940 Flabellina vs Pleurob -0.027571 0.039558 -0.696993 Flabellina vs Tomthom 0.002885 0.042750 0.067479 Flabellina vs Umbracu 0.046818 0.042760 1.094908 Flabellina vs Aplysia 0.047828 0.044424 1.076611 Flabellina vs Dolabel 0.022345 0.045128 0.495155 Flabellina vs Styloch 0.015766 0.042686 0.369351 Flabellina vs Bursate 0.005201 0.044509 0.116848 Flabellina vs Dolabri 0.021480 0.043448 0.494395 Flabellina vs Phyllap 0.009501 0.043772 0.217065 Flabellina vs Petalif 0.010587 0.045267 0.233885 Flabellina vs Notarch 0.032701 0.044989 0.726856 Flabellina vs Akera_b 0.034511 0.042262 0.816609 Flabellina vs Cavolin -0.034349 0.053058 -0.647388 Flabellina vs Clione_ -0.080409 0.054481 -1.475918 Flabellina vs Diaphan 0.027904 0.042859 0.651060 Flabellina vs Haminoe 0.006452 0.043434 0.148539 Flabellina vs Smaragd -0.021797 0.042699 -0.510482 Flabellina vs Bulla_g -0.034455 0.043247 -0.796707 Flabellina vs Philine 0.027946 0.043273 0.645809 Flabellina vs Scaphan -0.014155 0.048892 -0.289510 Flabellina vs Elysia_ 0.087053 0.044109 1.973578 Flabellina vs Thuridi 0.078118 0.041065 1.902291 Flabellina vs Placida 0.070539 0.044717 1.577472 Flabellina vs Acteon_ 0.013001 0.041825 0.310839 Flabellina vs Pupa_so -0.019538 0.044516 -0.438885 Flabellina vs Hydatin 0.076830 0.041537 1.849677 Flabellina vs Pontohe 0.005310 0.051688 0.102740 Flabellina vs Parahed 0.090032 0.046799 1.923799 Flabellina vs Unela_g 0.090032 0.046799 1.923799 Dendronotu vs Berthel 0.000325 0.042756 0.007592 Dendronotu vs Berthel 0.012740 0.041616 0.306123 Dendronotu vs Pleurob -0.007107 0.040793 -0.174215 Dendronotu vs Tomthom 0.023146 0.043251 0.535139 Dendronotu vs Umbracu 0.061718 0.044638 1.382629 Dendronotu vs Aplysia 0.062718 0.044309 1.415465 Dendronotu vs Dolabel 0.042618 0.044863 0.949964 Dendronotu vs Styloch 0.036080 0.043785 0.824035 Dendronotu vs Bursate 0.025571 0.045404 0.563188 Dendronotu vs Dolabri 0.041762 0.044565 0.937107 Dendronotu vs Phyllap 0.029989 0.045009 0.666298 Dendronotu vs Petalif 0.024694 0.045711 0.540227 Dendronotu vs Notarch 0.048005 0.045853 1.046949 Dendronotu vs Akera_b 0.049500 0.042947 1.152581 Dendronotu vs Cavolin -0.013795 0.054147 -0.254767 Dendronotu vs Clione_ -0.059888 0.052793 -1.134399 Dendronotu vs Diaphan 0.036930 0.045399 0.813452 Dendronotu vs Haminoe 0.029384 0.047785 0.614915 Dendronotu vs Smaragd -0.011779 0.048936 -0.240710 Dendronotu vs Bulla_g -0.024478 0.047513 -0.515171 Dendronotu vs Philine 0.036823 0.044518 0.827143 Dendronotu vs Scaphan 0.001377 0.049783 0.027650 Dendronotu vs Elysia_ 0.095073 0.044029 2.159319 Dendronotu vs Thuridi 0.098534 0.043798 2.249760 Dendronotu vs Placida 0.091773 0.044739 2.051298 Dendronotu vs Acteon_ 0.022847 0.045323 0.504091 Dendronotu vs Pupa_so 0.000943 0.042720 0.022065 Dendronotu vs Hydatin 0.097175 0.045019 2.158530 Dendronotu vs Pontohe 0.013355 0.050236 0.265854 Dendronotu vs Parahed 0.092291 0.045825 2.013997 Dendronotu vs Unela_g 0.092291 0.045825 2.013997

234
Berthella_ vs Berthel 0.005843 0.005142 1.136202 Berthella_ vs Pleurob -0.015920 0.027430 -0.580367 Berthella_ vs Tomthom 0.010932 0.030521 0.358197 Berthella_ vs Umbracu 0.054186 0.044289 1.223485 Berthella_ vs Aplysia 0.060830 0.043603 1.395070 Berthella_ vs Dolabel 0.038481 0.042884 0.897315 Berthella_ vs Styloch 0.043077 0.042327 1.017723 Berthella_ vs Bursate 0.031725 0.044251 0.716942 Berthella_ vs Dolabri 0.049250 0.041985 1.173043 Berthella_ vs Phyllap 0.036408 0.043790 0.831421 Berthella_ vs Petalif 0.037660 0.044086 0.854241 Berthella_ vs Notarch 0.044714 0.043153 1.036165 Berthella_ vs Akera_b 0.058491 0.041888 1.396355 Berthella_ vs Cavolin -0.022933 0.052021 -0.440853 Berthella_ vs Clione_ -0.085593 0.056672 -1.510323 Berthella_ vs Diaphan 0.020201 0.045734 0.441712 Berthella_ vs Haminoe 0.009672 0.048048 0.201297 Berthella_ vs Smaragd -0.028588 0.047620 -0.600328 Berthella_ vs Bulla_g -0.030923 0.045624 -0.677780 Berthella_ vs Philine 0.026826 0.045201 0.593489 Berthella_ vs Scaphan -0.005112 0.047124 -0.108481 Berthella_ vs Elysia_ 0.093046 0.046276 2.010689 Berthella_ vs Thuridi 0.087537 0.044939 1.947901 Berthella_ vs Placida 0.089593 0.048080 1.863425 Berthella_ vs Acteon_ 0.021280 0.045807 0.464570 Berthella_ vs Pupa_so 0.017567 0.046540 0.377460 Berthella_ vs Hydatin 0.105198 0.043787 2.402479 Berthella_ vs Pontohe 0.027131 0.050340 0.538957 Berthella_ vs Parahed 0.093879 0.043528 2.156765 Berthella_ vs Unela_g 0.093879 0.043528 2.156765 Berthella_ vs Pleurob -0.025048 0.024539 -1.020738 Berthella_ vs Tomthom 0.010491 0.028118 0.373097 Berthella_ vs Umbracu 0.053891 0.041760 1.290494 Berthella_ vs Aplysia 0.055227 0.041371 1.334924 Berthella_ vs Dolabel 0.030158 0.040615 0.742539 Berthella_ vs Styloch 0.023534 0.040801 0.576796 Berthella_ vs Bursate 0.013014 0.042495 0.306244 Berthella_ vs Dolabri 0.029248 0.040114 0.729124 Berthella_ vs Phyllap 0.017274 0.041728 0.413963 Berthella_ vs Petalif 0.018405 0.041975 0.438476 Berthella_ vs Notarch 0.035309 0.040587 0.869945 Berthella_ vs Akera_b 0.043185 0.039680 1.088329 Berthella_ vs Cavolin -0.031192 0.049679 -0.627873 Berthella_ vs Clione_ -0.077658 0.053456 -1.452763 Berthella_ vs Diaphan 0.024916 0.043456 0.573364 Berthella_ vs Haminoe 0.016305 0.044921 0.362975 Berthella_ vs Smaragd -0.024850 0.045151 -0.550377 Berthella_ vs Bulla_g -0.026784 0.043859 -0.610680 Berthella_ vs Philine 0.030454 0.042386 0.718492 Berthella_ vs Scaphan -0.004721 0.045963 -0.102708 Berthella_ vs Elysia_ 0.076200 0.043686 1.744275 Berthella_ vs Thuridi 0.081587 0.043098 1.893066 Berthella_ vs Placida 0.072680 0.045993 1.580239 Berthella_ vs Acteon_ 0.020540 0.043047 0.477155 Berthella_ vs Pupa_so -0.017303 0.046138 -0.375028 Berthella_ vs Hydatin 0.095497 0.041873 2.280650 Berthella_ vs Pontohe 0.008430 0.047659 0.176891 Berthella_ vs Parahed 0.080855 0.042210 1.915527 Berthella_ vs Unela_g 0.080855 0.042210 1.915527 Pleurobran vs Tomthom 0.030130 0.027700 1.087707 Pleurobran vs Umbracu 0.079265 0.041940 1.889982 Pleurobran vs Aplysia 0.080762 0.041508 1.945670 Pleurobran vs Dolabel 0.055353 0.042003 1.317837 Pleurobran vs Styloch 0.048770 0.042169 1.156534 Pleurobran vs Bursate 0.038306 0.041365 0.926028 Pleurobran vs Dolabri 0.054452 0.041484 1.312607 Pleurobran vs Phyllap 0.042709 0.042662 1.001101 Pleurobran vs Petalif 0.037499 0.042680 0.878604 Pleurobran vs Notarch 0.060799 0.042225 1.439885 Pleurobran vs Akera_b 0.068485 0.041622 1.645401 Pleurobran vs Cavolin -0.005655 0.051381 -0.110065 Pleurobran vs Clione_ -0.057324 0.052909 -1.083439 Pleurobran vs Diaphan 0.050923 0.043631 1.167131 Pleurobran vs Haminoe 0.043915 0.048271 0.909751 Pleurobran vs Smaragd 0.000000 0.047077 0.000000 Pleurobran vs Bulla_g -0.000911 0.045934 -0.019825 Pleurobran vs Philine 0.056283 0.043343 1.298527 Pleurobran vs Scaphan 0.022219 0.047653 0.466261

235
Pleurobran vs Elysia_ 0.108965 0.045022 2.420284 Pleurobran vs Thuridi 0.107727 0.044072 2.444353 Pleurobran vs Placida 0.106062 0.047064 2.253598 Pleurobran vs Acteon_ 0.046096 0.043343 1.063513 Pleurobran vs Pupa_so 0.009175 0.048400 0.189574 Pleurobran vs Hydatin 0.115558 0.043117 2.680106 Pleurobran vs Pontohe 0.027811 0.048507 0.573336 Pleurobran vs Parahed 0.112992 0.044586 2.534265 Pleurobran vs Unela_g 0.112992 0.044586 2.534265 Tomthompso vs Umbracu 0.037467 0.040014 0.936370 Tomthompso vs Aplysia 0.044609 0.040361 1.105239 Tomthompso vs Dolabel 0.024635 0.039778 0.619310 Tomthompso vs Styloch 0.018075 0.041629 0.434182 Tomthompso vs Bursate 0.007588 0.040721 0.186338 Tomthompso vs Dolabri 0.023756 0.038901 0.610681 Tomthompso vs Phyllap 0.011835 0.040895 0.289396 Tomthompso vs Petalif 0.012932 0.041501 0.311600 Tomthompso vs Notarch 0.023734 0.040307 0.588835 Tomthompso vs Akera_b 0.037663 0.041309 0.911723 Tomthompso vs Cavolin -0.036150 0.052122 -0.693570 Tomthompso vs Clione_ -0.076673 0.052938 -1.448338 Tomthompso vs Diaphan 0.020108 0.042337 0.474948 Tomthompso vs Haminoe 0.011354 0.046881 0.242185 Tomthompso vs Smaragd -0.036196 0.046945 -0.771028 Tomthompso vs Bulla_g -0.037518 0.046693 -0.803490 Tomthompso vs Philine 0.025646 0.043092 0.595134 Tomthompso vs Scaphan -0.009917 0.047399 -0.209233 Tomthompso vs Elysia_ 0.070343 0.040617 1.731845 Tomthompso vs Thuridi 0.081790 0.042705 1.915237 Tomthompso vs Placida 0.072955 0.044155 1.652267 Tomthompso vs Acteon_ 0.009853 0.041764 0.235921 Tomthompso vs Pupa_so -0.022493 0.048019 -0.468411 Tomthompso vs Hydatin 0.089883 0.042078 2.136108 Tomthompso vs Pontohe -0.002300 0.048632 -0.047298 Tomthompso vs Parahed 0.081091 0.041828 1.938662 Tomthompso vs Unela_g 0.081091 0.041828 1.938662 Umbraculum vs Aplysia 0.001507 0.030016 0.050222 Umbraculum vs Dolabel -0.018846 0.028974 -0.650451 Umbraculum vs Styloch -0.030653 0.030484 -1.005553 Umbraculum vs Bursate -0.041173 0.032499 -1.266891 Umbraculum vs Dolabri -0.024938 0.031040 -0.803434 Umbraculum vs Phyllap -0.037211 0.031876 -1.167376 Umbraculum vs Petalif -0.041088 0.031382 -1.309288 Umbraculum vs Notarch -0.019383 0.030624 -0.632928 Umbraculum vs Akera_b -0.017047 0.031673 -0.538214 Umbraculum vs Cavolin -0.068911 0.049070 -1.404353 Umbraculum vs Clione_ -0.121714 0.045343 -2.684285 Umbraculum vs Diaphan -0.019803 0.031716 -0.624369 Umbraculum vs Haminoe -0.037284 0.039448 -0.945145 Umbraculum vs Smaragd -0.080642 0.039466 -2.043321 Umbraculum vs Bulla_g -0.075967 0.039690 -1.913992 Umbraculum vs Philine -0.008505 0.036544 -0.232729 Umbraculum vs Scaphan -0.051934 0.041726 -1.244645 Umbraculum vs Elysia_ 0.037226 0.037177 1.001322 Umbraculum vs Thuridi 0.047299 0.038368 1.232768 Umbraculum vs Placida 0.031907 0.038789 0.822561 Umbraculum vs Acteon_ -0.028405 0.031999 -0.887691 Umbraculum vs Pupa_so -0.065772 0.045746 -1.437760 Umbraculum vs Hydatin 0.056783 0.033744 1.682754 Umbraculum vs Pontohe -0.047381 0.046511 -1.018714 Umbraculum vs Parahed 0.041571 0.038779 1.072013 Umbraculum vs Unela_g 0.041571 0.038779 1.072013 Aplysia_oc vs Dolabel -0.025405 0.020697 -1.227470 Aplysia_oc vs Styloch -0.031988 0.023973 -1.334350 Aplysia_oc vs Bursate -0.042452 0.024145 -1.758244 Aplysia_oc vs Dolabri -0.026306 0.022942 -1.146620 Aplysia_oc vs Phyllap -0.038520 0.022392 -1.720258 Aplysia_oc vs Petalif -0.042398 0.022338 -1.898026 Aplysia_oc vs Notarch -0.025929 0.020181 -1.284806 Aplysia_oc vs Akera_b -0.023456 0.025320 -0.926369 Aplysia_oc vs Cavolin -0.070449 0.047743 -1.475591 Aplysia_oc vs Clione_ -0.133646 0.046400 -2.880271 Aplysia_oc vs Diaphan -0.031571 0.035464 -0.890230 Aplysia_oc vs Haminoe -0.054933 0.041811 -1.313847 Aplysia_oc vs Smaragd -0.081684 0.041527 -1.967008 Aplysia_oc vs Bulla_g -0.082584 0.041781 -1.976624 Aplysia_oc vs Philine -0.019808 0.037638 -0.526297 Aplysia_oc vs Scaphan -0.069674 0.042218 -1.650350

236
Aplysia_oc vs Elysia_ 0.019625 0.037510 0.523183 Aplysia_oc vs Thuridi 0.035380 0.039405 0.897856 Aplysia_oc vs Placida 0.019980 0.039245 0.509101 Aplysia_oc vs Acteon_ -0.035090 0.035926 -0.976746 Aplysia_oc vs Pupa_so -0.055365 0.043955 -1.259567 Aplysia_oc vs Hydatin 0.055526 0.036203 1.533750 Aplysia_oc vs Pontohe -0.038553 0.046228 -0.833977 Aplysia_oc vs Parahed 0.050172 0.037499 1.337948 Aplysia_oc vs Unela_g 0.050172 0.037499 1.337948 Dolabella vs Styloche -0.006541 0.025463 -0.256896 Dolabella vs Bursatel -0.016951 0.024518 -0.691369 Dolabella vs Dolabrif -0.000892 0.022211 -0.040146 Dolabella vs Phyllapl -0.012903 0.024325 -0.530445 Dolabella vs Petalife -0.016951 0.020858 -0.812661 Dolabella vs Notarchu -0.005587 0.020502 -0.272499 Dolabella vs Akera_bu 0.001902 0.025346 0.075030 Dolabella vs Cavolini -0.044399 0.048765 -0.910463 Dolabella vs Clione_l -0.113001 0.045945 -2.459492 Dolabella vs Diaphana -0.005281 0.035255 -0.149791 Dolabella vs Haminoea -0.027527 0.041247 -0.667367 Dolabella vs Smaragdi -0.056071 0.040030 -1.400713 Dolabella vs Bulla_go -0.056972 0.040282 -1.414334 Dolabella vs Philine_ 0.006533 0.037292 0.175185 Dolabella vs Scaphand -0.041923 0.042170 -0.994137 Dolabella vs Elysia_v 0.040833 0.039617 1.030692 Dolabella vs Thuridil 0.062240 0.040277 1.545278 Dolabella vs Placida_ 0.046886 0.040967 1.144473 Dolabella vs Acteon_t -0.009663 0.034971 -0.276314 Dolabella vs Pupa_sol -0.034273 0.044037 -0.778283 Dolabella vs Hydatina 0.076599 0.038237 2.003250 Dolabella vs Pontohed -0.017488 0.048257 -0.362389 Dolabella vs Parahedy 0.076382 0.038015 2.009239 Dolabella vs Unela_gl 0.076382 0.038015 2.009239 Stylocheil vs Bursate -0.010409 0.022015 -0.472831 Stylocheil vs Dolabri 0.005650 0.022070 0.255995 Stylocheil vs Phyllap -0.006320 0.021716 -0.291041 Stylocheil vs Petalif -0.005213 0.021852 -0.238550 Stylocheil vs Notarch 0.006162 0.025553 0.241155 Stylocheil vs Akera_b 0.008465 0.029510 0.286855 Stylocheil vs Cavolin -0.037732 0.047626 -0.792242 Stylocheil vs Clione_ -0.106337 0.047157 -2.254948 Stylocheil vs Diaphan 0.001518 0.035033 0.043340 Stylocheil vs Haminoe -0.020331 0.042290 -0.480761 Stylocheil vs Smaragd -0.049381 0.040741 -1.212084 Stylocheil vs Bulla_g -0.050282 0.042011 -1.196865 Stylocheil vs Philine 0.013422 0.038159 0.351735 Stylocheil vs Scaphan -0.034903 0.043405 -0.804127 Stylocheil vs Elysia_ 0.047744 0.038180 1.250495 Stylocheil vs Thuridi 0.069220 0.038845 1.781940 Stylocheil vs Placida 0.053842 0.039998 1.346127 Stylocheil vs Acteon_ -0.002994 0.035152 -0.085170 Stylocheil vs Pupa_so -0.022076 0.042734 -0.516584 Stylocheil vs Hydatin 0.077525 0.036600 2.118184 Stylocheil vs Pontohe -0.016016 0.045918 -0.348803 Stylocheil vs Parahed 0.072580 0.037858 1.917129 Stylocheil vs Unela_g 0.072580 0.037858 1.917129 Bursatella vs Dolabri 0.016059 0.021189 0.757916 Bursatella vs Phyllap 0.004144 0.020097 0.206210 Bursatella vs Petalif 0.005252 0.021943 0.239321 Bursatella vs Notarch 0.021884 0.026704 0.819511 Bursatella vs Akera_b 0.018853 0.029914 0.630216 Bursatella vs Cavolin -0.027156 0.049037 -0.553776 Bursatella vs Clione_ -0.095658 0.047126 -2.029831 Bursatella vs Diaphan 0.012163 0.035875 0.339031 Bursatella vs Haminoe -0.009232 0.043943 -0.210098 Bursatella vs Smaragd -0.038828 0.042674 -0.909871 Bursatella vs Bulla_g -0.039728 0.041735 -0.951923 Bursatella vs Philine 0.024079 0.038130 0.631494 Bursatella vs Scaphan -0.023628 0.042637 -0.554173 Bursatella vs Elysia_ 0.063756 0.039436 1.616703 Bursatella vs Thuridi 0.080099 0.040661 1.969917 Bursatella vs Placida 0.064745 0.042873 1.510153 Bursatella vs Acteon_ 0.007481 0.036744 0.203610 Bursatella vs Pupa_so -0.011338 0.045955 -0.246731 Bursatella vs Hydatin 0.088227 0.036924 2.389438 Bursatella vs Pontohe -0.005541 0.045943 -0.120607 Bursatella vs Parahed 0.088607 0.039284 2.255548 Bursatella vs Unela_g 0.088607 0.039284 2.255548

237
Dolabrifer vs Phyllap -0.012002 0.012699 -0.945142 Dolabrifer vs Petalif -0.010895 0.016326 -0.667311 Dolabrifer vs Notarch 0.005650 0.020938 0.269839 Dolabrifer vs Akera_b 0.002815 0.026410 0.106601 Dolabrifer vs Cavolin -0.043479 0.047991 -0.905979 Dolabrifer vs Clione_ -0.112118 0.047066 -2.382157 Dolabrifer vs Diaphan -0.004296 0.036342 -0.118202 Dolabrifer vs Haminoe -0.026429 0.041916 -0.630521 Dolabrifer vs Smaragd -0.055129 0.040828 -1.350261 Dolabrifer vs Bulla_g -0.056029 0.041078 -1.363975 Dolabrifer vs Philine 0.007574 0.038725 0.195586 Dolabrifer vs Scaphan -0.041001 0.042687 -0.960494 Dolabrifer vs Elysia_ 0.047159 0.039368 1.197910 Dolabrifer vs Thuridi 0.063268 0.041277 1.532755 Dolabrifer vs Placida 0.047890 0.041108 1.164992 Dolabrifer vs Acteon_ -0.008708 0.036779 -0.236774 Dolabrifer vs Pupa_so -0.027958 0.044304 -0.631048 Dolabrifer vs Hydatin 0.071744 0.035873 1.999953 Dolabrifer vs Pontohe -0.021731 0.047314 -0.459293 Dolabrifer vs Parahed 0.072034 0.037130 1.940028 Dolabrifer vs Unela_g 0.072034 0.037130 1.940028 Phyllaplys vs Petalif 0.001119 0.015572 0.071862 Phyllaplys vs Notarch 0.017828 0.021866 0.815339 Phyllaplys vs Akera_b 0.014831 0.026130 0.567600 Phyllaplys vs Cavolin -0.031499 0.048694 -0.646869 Phyllaplys vs Clione_ -0.100519 0.047812 -2.102389 Phyllaplys vs Diaphan 0.008078 0.037465 0.215624 Phyllaplys vs Haminoe -0.013560 0.043231 -0.313668 Phyllaplys vs Smaragd -0.043237 0.041964 -1.030319 Phyllaplys vs Bulla_g -0.044147 0.042617 -1.035913 Phyllaplys vs Philine 0.020117 0.039745 0.506153 Phyllaplys vs Scaphan -0.028289 0.042435 -0.666627 Phyllaplys vs Elysia_ 0.059978 0.039387 1.522790 Phyllaplys vs Thuridi 0.076310 0.042831 1.781641 Phyllaplys vs Placida 0.060840 0.043405 1.401697 Phyllaplys vs Acteon_ 0.003423 0.037882 0.090359 Phyllaplys vs Pupa_so -0.015590 0.044642 -0.349213 Phyllaplys vs Hydatin 0.084377 0.036580 2.306630 Phyllaplys vs Pontohe -0.009710 0.047514 -0.204360 Phyllaplys vs Parahed 0.084867 0.038114 2.226689 Phyllaplys vs Unela_g 0.084867 0.038114 2.226689 Petalifera vs Notarch 0.011429 0.019098 0.598442 Petalifera vs Akera_b 0.013702 0.026591 0.515295 Petalifera vs Cavolin -0.032642 0.049766 -0.655902 Petalifera vs Clione_ -0.095541 0.046731 -2.044498 Petalifera vs Diaphan 0.012174 0.037901 0.321213 Petalifera vs Haminoe -0.009280 0.042938 -0.216120 Petalifera vs Smaragd -0.038861 0.041720 -0.931475 Petalifera vs Bulla_g -0.039772 0.041572 -0.956711 Petalifera vs Philine 0.018846 0.039675 0.475007 Petalifera vs Scaphan -0.023587 0.043217 -0.545783 Petalifera vs Elysia_ 0.053327 0.040528 1.315799 Petalifera vs Thuridi 0.075043 0.042296 1.774238 Petalifera vs Placida 0.064645 0.041760 1.548025 Petalifera vs Acteon_ 0.007528 0.038264 0.196739 Petalifera vs Pupa_so -0.016852 0.045833 -0.367676 Petalifera vs Hydatin 0.083291 0.037051 2.247997 Petalifera vs Pontohe -0.010885 0.047401 -0.229637 Petalifera vs Parahed 0.083664 0.038695 2.162123 Petalifera vs Unela_g 0.083664 0.038695 2.162123 Notarchus_ vs Akera_b -0.002828 0.023822 -0.118732 Notarchus_ vs Cavolin -0.049708 0.049483 -1.004540 Notarchus_ vs Clione_ -0.102268 0.047018 -2.175075 Notarchus_ vs Diaphan 0.000000 0.036325 0.000000 Notarchus_ vs Haminoe -0.011057 0.041685 -0.265255 Notarchus_ vs Smaragd -0.050455 0.040737 -1.238573 Notarchus_ vs Bulla_g -0.051377 0.039595 -1.297549 Notarchus_ vs Philine 0.017145 0.038025 0.450869 Notarchus_ vs Scaphan -0.030214 0.041598 -0.726332 Notarchus_ vs Elysia_ 0.051921 0.040536 1.280856 Notarchus_ vs Thuridi 0.072930 0.040889 1.783586 Notarchus_ vs Placida 0.057619 0.041240 1.397180 Notarchus_ vs Acteon_ -0.009189 0.036586 -0.251155 Notarchus_ vs Pupa_so -0.039263 0.044795 -0.876495 Notarchus_ vs Hydatin 0.067180 0.038055 1.765352 Notarchus_ vs Pontohe -0.028216 0.047715 -0.591346 Notarchus_ vs Parahed 0.066596 0.038435 1.732670 Notarchus_ vs Unela_g 0.066596 0.038435 1.732670

238
Akera_bull vs Cavolin -0.051767 0.047388 -1.092396 Akera_bull vs Clione_ -0.120726 0.046450 -2.599066 Akera_bull vs Diaphan -0.012833 0.036144 -0.355050 Akera_bull vs Haminoe -0.035263 0.041224 -0.855406 Akera_bull vs Smaragd -0.063692 0.041269 -1.543309 Akera_bull vs Bulla_g -0.058822 0.041219 -1.427059 Akera_bull vs Philine -0.006138 0.038016 -0.161470 Akera_bull vs Scaphan -0.050152 0.041231 -1.216378 Akera_bull vs Elysia_ 0.038784 0.040063 0.968063 Akera_bull vs Thuridi 0.049518 0.040963 1.208839 Akera_bull vs Placida 0.039042 0.042157 0.926110 Akera_bull vs Acteon_ -0.011358 0.036655 -0.309858 Akera_bull vs Pupa_so -0.042450 0.042333 -1.002748 Akera_bull vs Hydatin 0.063453 0.037191 1.706156 Akera_bull vs Pontohe -0.025306 0.045501 -0.556167 Akera_bull vs Parahed 0.063941 0.036798 1.737600 Akera_bull vs Unela_g 0.063941 0.036798 1.737600 Cavolinia_ vs Clione_ -0.063051 0.054098 -1.165508 Cavolinia_ vs Diaphan 0.044775 0.047836 0.936015 Cavolinia_ vs Haminoe 0.019739 0.052937 0.372879 Cavolinia_ vs Smaragd -0.005545 0.053164 -0.104294 Cavolinia_ vs Bulla_g -0.000792 0.052040 -0.015227 Cavolinia_ vs Philine 0.056524 0.048883 1.156297 Cavolinia_ vs Scaphan 0.011389 0.052678 0.216205 Cavolinia_ vs Elysia_ 0.107629 0.048837 2.203841 Cavolinia_ vs Thuridi 0.102504 0.047602 2.153352 Cavolinia_ vs Placida 0.097952 0.050085 1.955712 Cavolinia_ vs Acteon_ 0.055800 0.048520 1.150033 Cavolinia_ vs Pupa_so 0.038725 0.052619 0.735937 Cavolinia_ vs Hydatin 0.110659 0.046838 2.362582 Cavolinia_ vs Pontohe 0.004804 0.052062 0.092283 Cavolinia_ vs Parahed 0.094661 0.047363 1.998630 Cavolinia_ vs Unela_g 0.094661 0.047988 1.972607 Clione_lim vs Diaphan 0.101849 0.048187 2.113635 Clione_lim vs Haminoe 0.085604 0.050481 1.695747 Clione_lim vs Smaragd 0.045405 0.048619 0.933907 Clione_lim vs Bulla_g 0.037908 0.046606 0.813360 Clione_lim vs Philine 0.113818 0.049508 2.299002 Clione_lim vs Scaphan 0.071937 0.049559 1.451557 Clione_lim vs Elysia_ 0.160992 0.050832 3.167113 Clione_lim vs Thuridi 0.172557 0.052078 3.313397 Clione_lim vs Placida 0.169765 0.050008 3.394751 Clione_lim vs Acteon_ 0.103984 0.048656 2.137109 Clione_lim vs Pupa_so 0.066522 0.050245 1.323948 Clione_lim vs Hydatin 0.171479 0.049208 3.484748 Clione_lim vs Pontohe 0.098579 0.055367 1.780460 Clione_lim vs Parahed 0.172988 0.049918 3.465438 Clione_lim vs Unela_g 0.172988 0.049918 3.465438 Diaphana_m vs Haminoe -0.016948 0.036227 -0.467822 Diaphana_m vs Smaragd -0.051764 0.036200 -1.429937 Diaphana_m vs Bulla_g -0.046870 0.034926 -1.341991 Diaphana_m vs Philine 0.022511 0.031360 0.717818 Diaphana_m vs Scaphan -0.031066 0.036322 -0.855286 Diaphana_m vs Elysia_ 0.062658 0.035477 1.766144 Diaphana_m vs Thuridi 0.061376 0.036743 1.670437 Diaphana_m vs Placida 0.046446 0.037802 1.228641 Diaphana_m vs Acteon_ 0.000854 0.009546 0.089479 Diaphana_m vs Pupa_so -0.023103 0.046775 -0.493931 Diaphana_m vs Hydatin 0.066782 0.038673 1.726822 Diaphana_m vs Pontohe 0.000016 0.048011 0.000340 Diaphana_m vs Parahed 0.067058 0.039414 1.701382 Diaphana_m vs Unela_g 0.067058 0.039414 1.701382 Haminoea_c vs Smaragd -0.018124 0.024325 -0.745070 Haminoea_c vs Bulla_g -0.031528 0.035048 -0.899546 Haminoea_c vs Philine 0.040683 0.036866 1.103533 Haminoea_c vs Scaphan -0.014792 0.040137 -0.368531 Haminoea_c vs Elysia_ 0.070442 0.040896 1.722452 Haminoea_c vs Thuridi 0.080918 0.039644 2.041133 Haminoea_c vs Placida 0.058861 0.041067 1.433302 Haminoea_c vs Acteon_ 0.018005 0.036924 0.487638 Haminoea_c vs Pupa_so -0.008415 0.049606 -0.169633 Haminoea_c vs Hydatin 0.092807 0.043118 2.152393 Haminoea_c vs Pontohe 0.010367 0.049178 0.210800 Haminoea_c vs Parahed 0.087874 0.043986 1.997765 Haminoea_c vs Unela_g 0.087874 0.043986 1.997765 Smaragdine vs Bulla_g -0.006494 0.034467 -0.188399 Smaragdine vs Philine 0.074970 0.038225 1.961284 Smaragdine vs Scaphan 0.022597 0.040645 0.555951

239
Smaragdine vs Elysia_ 0.103973 0.041654 2.496115 Smaragdine vs Thuridi 0.107727 0.039787 2.707593 Smaragdine vs Placida 0.093590 0.043059 2.173512 Smaragdine vs Acteon_ 0.052346 0.036451 1.436065 Smaragdine vs Pupa_so 0.016624 0.047356 0.351037 Smaragdine vs Hydatin 0.128853 0.041990 3.068675 Smaragdine vs Pontohe 0.038865 0.049333 0.787798 Smaragdine vs Parahed 0.117852 0.043186 2.728955 Smaragdine vs Unela_g 0.117852 0.043186 2.728955 Bulla_goul vs Philine 0.069818 0.037967 1.838929 Bulla_goul vs Scaphan 0.016885 0.039155 0.431245 Bulla_goul vs Elysia_ 0.104926 0.044914 2.336123 Bulla_goul vs Thuridi 0.108590 0.042582 2.550122 Bulla_goul vs Placida 0.094496 0.044652 2.116287 Bulla_goul vs Acteon_ 0.047618 0.036128 1.318015 Bulla_goul vs Pupa_so 0.004608 0.047304 0.097421 Bulla_goul vs Hydatin 0.118168 0.044027 2.684001 Bulla_goul vs Pontohe 0.033591 0.050831 0.660850 Bulla_goul vs Parahed 0.120004 0.044966 2.668763 Bulla_goul vs Unela_g 0.120004 0.044966 2.668763 Philine_ap vs Scaphan -0.048256 0.031869 -1.514219 Philine_ap vs Elysia_ 0.046444 0.039809 1.166675 Philine_ap vs Thuridi 0.050435 0.039986 1.261329 Philine_ap vs Placida 0.035306 0.041143 0.858118 Philine_ap vs Acteon_ -0.021754 0.031864 -0.682721 Philine_ap vs Pupa_so -0.034343 0.046547 -0.737816 Philine_ap vs Hydatin 0.044908 0.041257 1.088508 Philine_ap vs Pontohe -0.023301 0.049092 -0.474631 Philine_ap vs Parahed 0.045498 0.040342 1.127821 Philine_ap vs Unela_g 0.045498 0.040342 1.127821 Scaphander vs Elysia_ 0.089905 0.043991 2.043711 Scaphander vs Thuridi 0.110616 0.044399 2.491394 Scaphander vs Placida 0.084940 0.044197 1.921847 Scaphander vs Acteon_ 0.037818 0.036667 1.031380 Scaphander vs Pupa_so 0.007665 0.049892 0.153625 Scaphander vs Hydatin 0.095416 0.044292 2.154260 Scaphander vs Pontohe 0.013266 0.051298 0.258610 Scaphander vs Parahed 0.097126 0.043342 2.240926 Scaphander vs Unela_g 0.097126 0.043342 2.240926 Elysia_vir vs Thuridi 0.004947 0.024095 0.205314 Elysia_vir vs Placida 0.004977 0.026442 0.188228 Elysia_vir vs Acteon_ -0.061092 0.034792 -1.755891 Elysia_vir vs Pupa_so -0.079729 0.042474 -1.877114 Elysia_vir vs Hydatin 0.005007 0.034425 0.145449 Elysia_vir vs Pontohe -0.065374 0.041279 -1.583704 Elysia_vir vs Parahed -0.003957 0.037321 -0.106029 Elysia_vir vs Unela_g -0.003957 0.037321 -0.106029 Thuridilla vs Placida -0.010427 0.027208 -0.383229 Thuridilla vs Acteon_ -0.060372 0.036305 -1.662937 Thuridilla vs Pupa_so -0.085873 0.044327 -1.937269 Thuridilla vs Hydatin 0.000000 0.037440 0.000000 Thuridilla vs Pontohe -0.071845 0.042455 -1.692256 Thuridilla vs Parahed -0.009573 0.037291 -0.256720 Thuridilla vs Unela_g -0.009573 0.037291 -0.256720 Placida_de vs Acteon_ -0.050713 0.037737 -1.343836 Placida_de vs Pupa_so -0.076521 0.045466 -1.683024 Placida_de vs Hydatin 0.010629 0.035995 0.295301 Placida_de vs Pontohe -0.071547 0.044948 -1.591778 Placida_de vs Parahed 0.006027 0.038115 0.158134 Placida_de vs Unela_g 0.006027 0.038115 0.158134 Acteon_tor vs Pupa_so -0.040986 0.047084 -0.870477 Acteon_tor vs Hydatin 0.069374 0.038457 1.803965 Acteon_tor vs Pontohe -0.011830 0.048760 -0.242608 Acteon_tor vs Parahed 0.049909 0.041113 1.213956 Acteon_tor vs Unela_g 0.049909 0.041113 1.213956 Pupa_solid vs Hydatin 0.117761 0.044186 2.665091 Pupa_solid vs Pontohe 0.006250 0.049763 0.125597 Pupa_solid vs Parahed 0.075210 0.046336 1.623152 Pupa_solid vs Unela_g 0.075210 0.046336 1.623152 Hydatina_p vs Pontohe -0.096804 0.040310 -2.401472 Hydatina_p vs Parahed -0.030584 0.038679 -0.790707 Hydatina_p vs Unela_g -0.030584 0.038679 -0.790707 Pontohedyl vs Parahed 0.067824 0.038680 1.753468 Pontohedyl vs Unela_g 0.067824 0.039067 1.736105 Parahedyle vs Unela_g 0.000000 0.006979 0.000000

240
Anaspidea-Datensatz (18S rDNA) Ergebnisse der Analyse der relativen Substitutionsraten auf der Grundlage des 18S rDNA-Datensatzes mit Atys semistriatus als Außengruppe (1). Eine signifikant unterschiedliche Substitutionsgeschwindigkeit ist gegeben, wenn der „Z-score“ (k12-k13/SD) höher als 1,96 ist. Positive „Z-score“-Werte bedeuten, dass Taxon (2) eine größere Evolutionsgeschwindigkeit aufweist; negative Werte stehen für eine erhöhte Substitutionsrate des Taxons (3). (2) (3) K12 - K13 SD Z-score Akera_bull vs Aplysia 0.003516 0.002539 1.385016 Akera_bull vs Aplysia 0.003516 0.002539 1.385016 Akera_bull vs Aplysia 0.003525 0.002605 1.353126 Akera_bull vs Aplysia 0.004109 0.002601 1.579535 Akera_bull vs Aplysia 0.001175 0.002726 0.431277 Akera_bull vs Aplysia 0.002926 0.002470 1.184669 Akera_bull vs Siphono 0.002933 0.002473 1.186211 Akera_bull vs Bursate 0.004100 0.002668 1.536496 Akera_bull vs Dolabel 0.002317 0.002399 0.965915 Akera_bull vs Petalif 0.005844 0.002601 2.247291 Akera_bull vs Dolabri 0.004675 0.002603 1.795880 Akera_bull vs Styloch 0.003498 0.002730 1.281235 Akera_bull vs Dolabri 0.004679 0.002605 1.795865 Akera_bull vs Creseis -0.033343 0.005700 -5.849149 Akera_bull vs Clio_py -0.005928 0.004245 -1.396544 Akera_bull vs Diacria -0.011260 0.004412 -2.552321 Akera_bull vs Spongio -0.002357 0.003412 -0.690817 Aplysia_gi vs Aplysia 0.000000 0.000000 Not deter. Aplysia_gi vs Aplysia 0.000007 0.000576 0.011619 Aplysia_gi vs Aplysia 0.001176 0.001170 1.005077 Aplysia_gi vs Aplysia -0.001754 0.001835 -0.956043 Aplysia_gi vs Aplysia -0.000589 0.000586 -1.005076 Aplysia_gi vs Siphono 0.000000 0.000000 Not deter. Aplysia_gi vs Bursate 0.000588 0.002010 0.292299 Aplysia_gi vs Dolabel -0.001786 0.001935 -0.923126 Aplysia_gi vs Petalif 0.002329 0.001919 1.213716 Aplysia_gi vs Dolabri 0.001162 0.001923 0.604222 Aplysia_gi vs Styloch -0.000015 0.002096 -0.007254 Aplysia_gi vs Dolabri 0.001163 0.001924 0.604152 Aplysia_gi vs Creseis -0.038065 0.005734 -6.638488 Aplysia_gi vs Clio_py -0.010035 0.003942 -2.545575 Aplysia_gi vs Diacria -0.014163 0.004199 -3.372754 Aplysia_gi vs Spongio -0.005868 0.003092 -1.897666 Aplysia_oc vs Aplysia 0.000007 0.000576 0.011619 Aplysia_oc vs Aplysia 0.001176 0.001170 1.005077 Aplysia_oc vs Aplysia -0.001754 0.001835 -0.956043 Aplysia_oc vs Aplysia -0.000589 0.000586 -1.005076 Aplysia_oc vs Siphono 0.000000 0.000000 Not deter. Aplysia_oc vs Bursate 0.000588 0.002010 0.292299 Aplysia_oc vs Dolabel -0.001786 0.001935 -0.923126 Aplysia_oc vs Petalif 0.002329 0.001919 1.213716 Aplysia_oc vs Dolabri 0.001162 0.001923 0.604222 Aplysia_oc vs Styloch -0.000015 0.002096 -0.007254 Aplysia_oc vs Dolabri 0.001163 0.001924 0.604152 Aplysia_oc vs Creseis -0.038065 0.005734 -6.638488 Aplysia_oc vs Clio_py -0.010035 0.003942 -2.545575 Aplysia_oc vs Diacria -0.014163 0.004199 -3.372754 Aplysia_oc vs Spongio -0.005868 0.003092 -1.897666 Aplysia_sy vs Aplysia 0.001170 0.001304 0.897168 Aplysia_sy vs Aplysia -0.001762 0.001743 -1.010884 Aplysia_sy vs Aplysia -0.000596 0.000822 -0.725107 Aplysia_sy vs Siphono -0.000007 0.000576 -0.011626 Aplysia_sy vs Bursate 0.000582 0.002092 0.278067 Aplysia_sy vs Dolabel -0.001794 0.002020 -0.888109 Aplysia_sy vs Petalif 0.002324 0.002005 1.159215 Aplysia_sy vs Dolabri 0.001156 0.002008 0.575617 Aplysia_sy vs Styloch -0.000022 0.002175 -0.010152 Aplysia_sy vs Dolabri 0.001156 0.002010 0.575400 Aplysia_sy vs Creseis -0.038094 0.005768 -6.604827

241
Aplysia_sy vs Clio_py -0.010048 0.003987 -2.520226 Aplysia_sy vs Diacria -0.014179 0.004242 -3.342682 Aplysia_sy vs Spongio -0.005878 0.003147 -1.867798 Aplysia_pa vs Aplysia -0.002930 0.001925 -1.522013 Aplysia_pa vs Aplysia -0.001765 0.001014 -1.739774 Aplysia_pa vs Siphono -0.001176 0.001171 -1.005080 Aplysia_pa vs Bursate -0.000006 0.002169 -0.002683 Aplysia_pa vs Dolabel -0.002379 0.002181 -1.090789 Aplysia_pa vs Petalif 0.001734 0.002085 0.831334 Aplysia_pa vs Dolabri 0.000568 0.002089 0.271690 Aplysia_pa vs Styloch -0.000610 0.002249 -0.271001 Aplysia_pa vs Dolabri 0.000567 0.002091 0.271331 Aplysia_pa vs Creseis -0.038638 0.005760 -6.707851 Aplysia_pa vs Clio_py -0.010625 0.003981 -2.668759 Aplysia_pa vs Diacria -0.015350 0.004238 -3.621873 Aplysia_pa vs Spongio -0.006459 0.003143 -2.055318 Aplysia_ju vs Aplysia 0.001165 0.001739 0.670034 Aplysia_ju vs Siphono 0.001755 0.001836 0.956047 Aplysia_ju vs Bursate 0.002922 0.002533 1.153516 Aplysia_ju vs Dolabel 0.000550 0.002601 0.211547 Aplysia_ju vs Petalif 0.004663 0.002462 1.894388 Aplysia_ju vs Dolabri 0.003496 0.002465 1.418545 Aplysia_ju vs Styloch 0.002320 0.002603 0.891437 Aplysia_ju vs Dolabri 0.003499 0.002467 1.418464 Aplysia_ju vs Creseis -0.035699 0.005943 -6.006867 Aplysia_ju vs Clio_py -0.007688 0.004278 -1.797206 Aplysia_ju vs Diacria -0.012419 0.004518 -2.748478 Aplysia_ju vs Spongio -0.003529 0.003455 -1.021270 Aplysia_da vs Siphono 0.000589 0.000586 1.005079 Aplysia_da vs Bursate 0.001176 0.002094 0.561376 Aplysia_da vs Dolabel -0.001197 0.002023 -0.591925 Aplysia_da vs Petalif 0.002918 0.002007 1.453722 Aplysia_da vs Dolabri 0.001751 0.002011 0.870441 Aplysia_da vs Styloch 0.000574 0.002177 0.263507 Aplysia_da vs Dolabri 0.001752 0.002013 0.870477 Aplysia_da vs Creseis -0.037474 0.005767 -6.498373 Aplysia_da vs Clio_py -0.009445 0.003988 -2.368573 Aplysia_da vs Diacria -0.013574 0.004242 -3.199919 Aplysia_da vs Spongio -0.005278 0.003149 -1.676407 Siphonota_ vs Bursate 0.000588 0.002011 0.292301 Siphonota_ vs Dolabel -0.001787 0.001936 -0.923137 Siphonota_ vs Petalif 0.002331 0.001920 1.213721 Siphonota_ vs Dolabri 0.001163 0.001924 0.604222 Siphonota_ vs Styloch -0.000015 0.002097 -0.007259 Siphonota_ vs Dolabri 0.001163 0.001926 0.604153 Siphonota_ vs Creseis -0.037460 0.005701 -6.571139 Siphonota_ vs Clio_py -0.010041 0.003945 -2.545582 Siphonota_ vs Diacria -0.014172 0.004202 -3.372760 Siphonota_ vs Spongio -0.005285 0.003038 -1.739561 Bursatella vs Dolabel -0.002369 0.002097 -1.129692 Bursatella vs Petalif 0.001740 0.001534 1.134122 Bursatella vs Dolabri 0.000574 0.001538 0.373451 Bursatella vs Styloch -0.000601 0.001748 -0.343753 Bursatella vs Dolabri 0.000574 0.001539 0.373103 Bursatella vs Creseis -0.038601 0.005694 -6.779069 Bursatella vs Clio_py -0.010024 0.003896 -2.572737 Bursatella vs Diacria -0.014748 0.004073 -3.621061 Bursatella vs Spongio -0.006449 0.003298 -1.955580 Dolabella_ vs Petalif 0.004113 0.001838 2.238197 Dolabella_ vs Dolabri 0.002945 0.001644 1.791304 Dolabella_ vs Styloch 0.001770 0.001834 0.964812 Dolabella_ vs Dolabri 0.002948 0.001645 1.792061 Dolabella_ vs Creseis -0.036252 0.005639 -6.428293 Dolabella_ vs Clio_py -0.008240 0.004076 -2.021430 Dolabella_ vs Diacria -0.012970 0.004328 -2.996827 Dolabella_ vs Spongio -0.004077 0.003250 -1.254416 Petalifera vs Dolabri -0.001166 0.000819 -1.423958 Petalifera vs Styloch -0.002343 0.001166 -2.009965 Petalifera vs Dolabri -0.001167 0.000820 -1.424153 Petalifera vs Creseis -0.040377 0.005634 -7.167241 Petalifera vs Clio_py -0.012363 0.003852 -3.209410 Petalifera vs Diacria -0.017086 0.004072 -4.196352 Petalifera vs Spongio -0.008193 0.003138 -2.610867 Dolabrifer vs Styloch -0.001176 0.001160 -1.013522 Dolabrifer vs Dolabri 0.000000 0.000000 Not deter. Dolabrifer vs Creseis -0.039192 0.005634 -6.956179 Dolabrifer vs Clio_py -0.011191 0.003941 -2.839714 Dolabrifer vs Diacria -0.015922 0.004032 -3.948835

242
Dolabrifer vs Spongio -0.007618 0.003082 -2.471365 Stylocheil vs Dolabri 0.001177 0.001161 1.013952 Stylocheil vs Creseis -0.038027 0.005699 -6.672613 Stylocheil vs Clio_py -0.010014 0.003857 -2.596145 Stylocheil vs Diacria -0.014741 0.004161 -3.542359 Stylocheil vs Spongio -0.005848 0.003143 -1.860907 Dolabrifer vs Creseis -0.039230 0.005639 -6.956552 Dolabrifer vs Clio_py -0.011200 0.003944 -2.839665 Dolabrifer vs Diacria -0.015927 0.004076 -3.907124 Dolabrifer vs Spongio -0.007030 0.003139 -2.239632 Creseis_ac vs Clio_py 0.028041 0.005484 5.113001 Creseis_ac vs Diacria 0.022664 0.005867 3.863142 Creseis_ac vs Spongio 0.031586 0.006043 5.227013 Clio_pyram vs Diacria -0.004161 0.004135 -1.006372 Clio_pyram vs Spongio 0.004744 0.004527 1.048019 Diacria_qu vs Spongio 0.008876 0.004518 1.964653
Anaspidea-Datensatz (ITS1-5.8S-ITS2) Ergebnisse der Analyse der relativen Substitutionsraten auf der Grundlage des ITS1-5.8S-ITS2 - Datensatzes mit Atys semistriatus als Außengruppe (1). Eine signifikant unterschiedliche Substitutionsgeschwindigkeit ist gegeben, wenn der „Z-score“ (k12-k13/SD) höher als 1,96 ist. Positive „Z-score“-Werte bedeuten, dass Taxon (2) eine größere Evolutionsgeschwindigkeit aufweist; negative Werte stehen für eine erhöhte Substitutionsrate des Taxons (3). (2) (3) K12 - K13 SD Z-score Bursatella vs Petalif 0.011941 0.020990 0.568906 Bursatella vs Dolabri 0.042801 0.021114 2.027133 Bursatella vs Dolabri 0.040038 0.022206 1.803011 Bursatella vs Dolabel 0.025053 0.021957 1.140994 Bursatella vs Aplysia 0.049025 0.023056 2.126361 Bursatella vs Aplysia 0.068202 0.022832 2.987158 Bursatella vs Aplysia 0.066973 0.026674 2.510736 Bursatella vs Akera_b 0.106825 0.027058 3.947947 Petalifera vs Dolabri 0.030294 0.014410 2.102249 Petalifera vs Dolabri 0.027884 0.015201 1.834413 Petalifera vs Dolabel 0.010624 0.018284 0.581030 Petalifera vs Aplysia 0.039168 0.021102 1.856110 Petalifera vs Aplysia 0.053838 0.019847 2.712597 Petalifera vs Aplysia 0.051345 0.023920 2.146531 Petalifera vs Akera_b 0.090806 0.023596 3.848328 Dolabrifer vs Dolabri -0.002427 0.007137 -0.340101 Dolabrifer vs Dolabel -0.012149 0.018080 -0.671929 Dolabrifer vs Aplysia 0.004883 0.019490 0.250563 Dolabrifer vs Aplysia 0.011365 0.018219 0.623801 Dolabrifer vs Aplysia -0.000580 0.022071 -0.026293 Dolabrifer vs Akera_b 0.051296 0.021867 2.345800 Dolabrifer vs Dolabel -0.012085 0.018307 -0.660117 Dolabrifer vs Aplysia 0.014821 0.020068 0.738524 Dolabrifer vs Aplysia 0.025432 0.018641 1.364270 Dolabrifer vs Aplysia 0.017907 0.022768 0.786507 Dolabrifer vs Akera_b 0.062632 0.022339 2.803644 Dolabella_ vs Aplysia 0.007519 0.017464 0.430538 Dolabella_ vs Aplysia 0.023054 0.016127 1.429480 Dolabella_ vs Aplysia 0.015560 0.020995 0.741094 Dolabella_ vs Akera_b 0.063396 0.021933 2.890487 Aplysia_ju vs Aplysia 0.018359 0.013893 1.321438 Aplysia_ju vs Aplysia 0.015063 0.017714 0.850350 Aplysia_ju vs Akera_b 0.056164 0.020723 2.710186 Aplysia_gi vs Aplysia -0.007699 0.012906 -0.596536 Aplysia_gi vs Akera_b 0.040930 0.019736 2.073893 Aplysia_oc vs Akera_b 0.050949 0.021615 2.357117

243
Aeolidoidea-Datensatz (18S rDNA) Ergebnisse der Analyse der relativen Substitutionsraten auf der Grundlage des 18S rDNA-Datensatzes mit Notaeolidia depressa als Außengruppe (1). Eine signifikant unterschiedliche Substitutionsgeschwindigkeit ist gegeben, wenn der „Z-score“ (k12-k13/SD) höher als 1,96 ist. Positive „Z-score“-Werte bedeuten, dass Taxon (2) eine größere Evolutionsgeschwindigkeit aufweist; negative Werte stehen für eine erhöhte Substitutionsrate des Taxons (3). (2) (3) K12 - K13 SD Z-score Baeolidia_ vs Godiva_ 0.000597 0.005702 0.104785 Baeolidia_ vs Cratena 0.001709 0.005770 0.296217 Baeolidia_ vs Phidian 0.004342 0.005388 0.805884 Baeolidia_ vs Facelin 0.001765 0.005460 0.323283 Baeolidia_ vs Facelin 0.001765 0.005460 0.323283 Baeolidia_ vs Phyllod 0.002478 0.005154 0.480701 Baeolidia_ vs Phyllod 0.004232 0.005276 0.801994 Baeolidia_ vs Phyllod 0.003356 0.005217 0.643344 Baeolidia_ vs Phyllod 0.002480 0.005333 0.464948 Baeolidia_ vs Phyllod 0.003581 0.005629 0.636142 Baeolidia_ vs Phyllod 0.002548 0.005498 0.463568 Baeolidia_ vs Embleto 0.240680 0.015132 15.905354 Baeolidia_ vs Cuthona 0.235601 0.015122 15.580079 Baeolidia_ vs Cuthona 0.234661 0.015186 15.451990 Baeolidia_ vs Cuthona 0.234773 0.015149 15.497755 Baeolidia_ vs Eubranc 0.239484 0.015186 15.769589 Baeolidia_ vs Eubranc 0.240480 0.015219 15.801169 Baeolidia_ vs Tergipe 0.225370 0.015494 14.545447 Baeolidia_ vs Flabell 0.250476 0.015125 16.560637 Baeolidia_ vs Flabell 0.237759 0.015253 15.587781 Baeolidia_ vs Flabell 0.226747 0.015011 15.105546 Baeolidia_ vs Flabell 0.192339 0.015402 12.487566 Baeolidia_ vs Flabell 0.209799 0.015479 13.553391 Godiva_ban vs Cratena 0.001940 0.004566 0.425006 Godiva_ban vs Phidian 0.004630 0.004741 0.976506 Godiva_ban vs Facelin 0.002054 0.004961 0.414047 Godiva_ban vs Facelin 0.002054 0.004961 0.414047 Godiva_ban vs Phyllod 0.002766 0.004649 0.595074 Godiva_ban vs Phyllod 0.004520 0.004787 0.944270 Godiva_ban vs Phyllod 0.003647 0.004720 0.772604 Godiva_ban vs Phyllod 0.002769 0.004847 0.571295 Godiva_ban vs Phyllod 0.003886 0.005141 0.755909 Godiva_ban vs Phyllod 0.002836 0.004949 0.573169 Godiva_ban vs Embleto 0.237979 0.014945 15.923279 Godiva_ban vs Cuthona 0.232025 0.015037 15.429927 Godiva_ban vs Cuthona 0.230994 0.015063 15.335096 Godiva_ban vs Cuthona 0.231206 0.015117 15.294116 Godiva_ban vs Eubranc 0.235890 0.015049 15.675002 Godiva_ban vs Eubranc 0.236469 0.015108 15.651529 Godiva_ban vs Tergipe 0.221667 0.015278 14.508632 Godiva_ban vs Flabell 0.245783 0.014914 16.480518 Godiva_ban vs Flabell 0.235659 0.015227 15.476858 Godiva_ban vs Flabell 0.222410 0.014860 14.966881 Godiva_ban vs Flabell 0.188759 0.015144 12.464273 Godiva_ban vs Flabell 0.204664 0.015321 13.358326 Cratena_pe vs Phidian 0.002686 0.003819 0.703339 Cratena_pe vs Facelin -0.000775 0.003635 -0.213119 Cratena_pe vs Facelin -0.000775 0.003635 -0.213119 Cratena_pe vs Phyllod 0.000824 0.004479 0.184067 Cratena_pe vs Phyllod 0.002577 0.004469 0.576740 Cratena_pe vs Phyllod 0.001702 0.004551 0.374091 Cratena_pe vs Phyllod 0.000825 0.004534 0.181985 Cratena_pe vs Phyllod 0.001874 0.004832 0.387917 Cratena_pe vs Phyllod 0.000885 0.004339 0.203959 Cratena_pe vs Embleto 0.235062 0.014814 15.867326 Cratena_pe vs Cuthona 0.230609 0.014859 15.519596 Cratena_pe vs Cuthona 0.229574 0.014902 15.405354 Cratena_pe vs Cuthona 0.229802 0.014906 15.416280 Cratena_pe vs Eubranc 0.234458 0.014980 15.650890

244
Cratena_pe vs Eubranc 0.235045 0.014968 15.703307 Cratena_pe vs Tergipe 0.220262 0.015081 14.604809 Cratena_pe vs Flabell 0.244289 0.014830 16.472357 Cratena_pe vs Flabell 0.233262 0.015014 15.536077 Cratena_pe vs Flabell 0.221108 0.014734 15.006308 Cratena_pe vs Flabell 0.187510 0.015044 12.464256 Cratena_pe vs Flabell 0.204137 0.015242 13.392672 Phidiana_l vs Facelin -0.002583 0.003702 -0.697816 Phidiana_l vs Facelin -0.002583 0.003702 -0.697816 Phidiana_l vs Phyllod -0.001858 0.003973 -0.467716 Phidiana_l vs Phyllod -0.000103 0.003963 -0.025871 Phidiana_l vs Phyllod -0.000980 0.003883 -0.252435 Phidiana_l vs Phyllod -0.001860 0.003949 -0.470951 Phidiana_l vs Phyllod -0.000905 0.004326 -0.209171 Phidiana_l vs Phyllod -0.001815 0.004310 -0.421137 Phidiana_l vs Embleto 0.233777 0.014792 15.803956 Phidiana_l vs Cuthona 0.226621 0.014810 15.301785 Phidiana_l vs Cuthona 0.225567 0.014816 15.224937 Phidiana_l vs Cuthona 0.225819 0.014821 15.236738 Phidiana_l vs Eubranc 0.230457 0.014820 15.550492 Phidiana_l vs Eubranc 0.231043 0.014847 15.561185 Phidiana_l vs Tergipe 0.216244 0.015036 14.381569 Phidiana_l vs Flabell 0.240919 0.014687 16.403697 Phidiana_l vs Flabell 0.228604 0.014965 15.275849 Phidiana_l vs Flabell 0.218646 0.014791 14.782217 Phidiana_l vs Flabell 0.183202 0.014982 12.228117 Phidiana_l vs Flabell 0.200671 0.015167 13.230663 Facelina_b vs Facelin 0.000000 0.000000 Not deter. Facelina_b vs Phyllod 0.000721 0.004145 0.173859 Facelina_b vs Phyllod 0.002479 0.004136 0.599541 Facelina_b vs Phyllod 0.001601 0.004223 0.379089 Facelina_b vs Phyllod 0.000721 0.004310 0.167321 Facelina_b vs Phyllod 0.001762 0.004413 0.399260 Facelina_b vs Phyllod 0.000778 0.004695 0.165744 Facelina_b vs Embleto 0.236323 0.014843 15.921608 Facelina_b vs Cuthona 0.230009 0.014913 15.423721 Facelina_b vs Cuthona 0.228967 0.014919 15.347576 Facelina_b vs Cuthona 0.229200 0.014941 15.340036 Facelina_b vs Eubranc 0.233238 0.014961 15.589689 Facelina_b vs Eubranc 0.234448 0.014969 15.662494 Facelina_b vs Tergipe 0.219618 0.015102 14.542125 Facelina_b vs Flabell 0.243729 0.014827 16.438480 Facelina_b vs Flabell 0.232083 0.015093 15.377355 Facelina_b vs Flabell 0.221819 0.014834 14.953199 Facelina_b vs Flabell 0.186498 0.015044 12.396435 Facelina_b vs Flabell 0.203160 0.015243 13.328550 Facelina_d vs Phyllod 0.000721 0.004145 0.173859 Facelina_d vs Phyllod 0.002479 0.004136 0.599541 Facelina_d vs Phyllod 0.001601 0.004223 0.379089 Facelina_d vs Phyllod 0.000721 0.004310 0.167321 Facelina_d vs Phyllod 0.001762 0.004413 0.399260 Facelina_d vs Phyllod 0.000778 0.004695 0.165744 Facelina_d vs Embleto 0.236323 0.014843 15.921608 Facelina_d vs Cuthona 0.230009 0.014913 15.423721 Facelina_d vs Cuthona 0.228967 0.014919 15.347576 Facelina_d vs Cuthona 0.229200 0.014941 15.340036 Facelina_d vs Eubranc 0.233238 0.014961 15.589689 Facelina_d vs Eubranc 0.234448 0.014969 15.662494 Facelina_d vs Tergipe 0.219618 0.015102 14.542125 Facelina_d vs Flabell 0.243729 0.014827 16.438480 Facelina_d vs Flabell 0.232083 0.015093 15.377355 Facelina_d vs Flabell 0.221819 0.014834 14.953199 Facelina_d vs Flabell 0.186498 0.015044 12.396435 Facelina_d vs Flabell 0.203160 0.015243 13.328550 Phyllodesm vs Phyllod 0.001751 0.001643 1.065813 Phyllodesm vs Phyllod 0.000877 0.000823 1.065768 Phyllodesm vs Phyllod 0.000000 0.001346 0.000000 Phyllodesm vs Phyllod 0.001021 0.001838 0.555146 Phyllodesm vs Phyllod 0.000054 0.002727 0.019801 Phyllodesm vs Embleto 0.235490 0.014755 15.959959 Phyllodesm vs Cuthona 0.229195 0.014861 15.422233 Phyllodesm vs Cuthona 0.228154 0.014886 15.326928 Phyllodesm vs Cuthona 0.228386 0.014908 15.319489 Phyllodesm vs Eubranc 0.233038 0.014872 15.669553 Phyllodesm vs Eubranc 0.233620 0.014933 15.644786 Phyllodesm vs Tergipe 0.218835 0.015102 14.490045 Phyllodesm vs Flabell 0.243482 0.014775 16.479175 Phyllodesm vs Flabell 0.232091 0.015034 15.438146

245
Phyllodesm vs Flabell 0.220361 0.014751 14.938999 Phyllodesm vs Flabell 0.184942 0.014916 12.399224 Phyllodesm vs Flabell 0.201515 0.015205 13.253609 Phyllodesm vs Phyllod -0.000875 0.001421 -0.615617 Phyllodesm vs Phyllod -0.001753 0.001778 -0.985860 Phyllodesm vs Phyllod -0.000794 0.001387 -0.572601 Phyllodesm vs Phyllod -0.001710 0.002960 -0.577741 Phyllodesm vs Embleto 0.233006 0.014698 15.853008 Phyllodesm vs Cuthona 0.226726 0.014769 15.351675 Phyllodesm vs Cuthona 0.225673 0.014793 15.255777 Phyllodesm vs Cuthona 0.225921 0.014816 15.248269 Phyllodesm vs Eubranc 0.230563 0.014779 15.600810 Phyllodesm vs Eubranc 0.231147 0.014840 15.575499 Phyllodesm vs Tergipe 0.216349 0.015010 14.413511 Phyllodesm vs Flabell 0.241023 0.014683 16.415621 Phyllodesm vs Flabell 0.229595 0.014940 15.368005 Phyllodesm vs Flabell 0.218754 0.014692 14.889623 Phyllodesm vs Flabell 0.182424 0.014862 12.274248 Phyllodesm vs Flabell 0.199015 0.015113 13.168866 Phyllodesm vs Phyllod -0.000878 0.001064 -0.825148 Phyllodesm vs Phyllod 0.000112 0.001628 0.069012 Phyllodesm vs Phyllod -0.000829 0.002596 -0.319526 Phyllodesm vs Embleto 0.234604 0.014757 15.897854 Phyllodesm vs Cuthona 0.228316 0.014827 15.399020 Phyllodesm vs Cuthona 0.227270 0.014851 15.303516 Phyllodesm vs Cuthona 0.227508 0.014874 15.296050 Phyllodesm vs Eubranc 0.232155 0.014837 15.646957 Phyllodesm vs Eubranc 0.232738 0.014898 15.622002 Phyllodesm vs Tergipe 0.217951 0.015068 14.464805 Phyllodesm vs Flabell 0.242603 0.014740 16.458341 Phyllodesm vs Flabell 0.231201 0.014998 15.415078 Phyllodesm vs Flabell 0.220361 0.014751 14.938999 Phyllodesm vs Flabell 0.184055 0.014920 12.335794 Phyllodesm vs Flabell 0.200631 0.015170 13.225592 Phyllodesm vs Phyllod 0.001022 0.001967 0.519462 Phyllodesm vs Phyllod 0.000054 0.002813 0.019227 Phyllodesm vs Embleto 0.235663 0.014770 15.955898 Phyllodesm vs Cuthona 0.229363 0.014839 15.456778 Phyllodesm vs Cuthona 0.228321 0.014863 15.361433 Phyllodesm vs Cuthona 0.228553 0.014886 15.353680 Phyllodesm vs Eubranc 0.233208 0.014831 15.724345 Phyllodesm vs Eubranc 0.233790 0.014910 15.679719 Phyllodesm vs Tergipe 0.218996 0.015080 14.522271 Phyllodesm vs Flabell 0.243659 0.014753 16.515984 Phyllodesm vs Flabell 0.231428 0.015018 15.410476 Phyllodesm vs Flabell 0.221411 0.014798 14.962674 Phyllodesm vs Flabell 0.185082 0.014911 12.412043 Phyllodesm vs Flabell 0.200837 0.015189 13.222458 Phyllodesm vs Phyllod -0.000965 0.003132 -0.308207 Phyllodesm vs Embleto 0.237563 0.015108 15.723772 Phyllodesm vs Cuthona 0.231067 0.015180 15.222045 Phyllodesm vs Cuthona 0.230002 0.015206 15.125715 Phyllodesm vs Cuthona 0.230233 0.015229 15.118504 Phyllodesm vs Eubranc 0.235033 0.015191 15.471608 Phyllodesm vs Eubranc 0.235635 0.015254 15.446926 Phyllodesm vs Tergipe 0.220401 0.015432 14.281974 Phyllodesm vs Flabell 0.245783 0.015090 16.288041 Phyllodesm vs Flabell 0.234073 0.015361 15.238211 Phyllodesm vs Flabell 0.222890 0.015103 14.757974 Phyllodesm vs Flabell 0.185441 0.015282 12.134602 Phyllodesm vs Flabell 0.202540 0.015540 13.033721 Phyllodesm vs Embleto 0.235382 0.014757 15.950879 Phyllodesm vs Cuthona 0.229053 0.014902 15.370367 Phyllodesm vs Cuthona 0.228004 0.014946 15.255490 Phyllodesm vs Cuthona 0.228242 0.014950 15.267391 Phyllodesm vs Eubranc 0.232919 0.015002 15.525489 Phyllodesm vs Eubranc 0.232675 0.014962 15.550829 Phyllodesm vs Tergipe 0.218628 0.015161 14.420119 Phyllodesm vs Flabell 0.243431 0.014833 16.410922 Phyllodesm vs Flabell 0.231126 0.015026 15.381677 Phyllodesm vs Flabell 0.220217 0.014795 14.884661 Phyllodesm vs Flabell 0.184566 0.015122 12.205262 Phyllodesm vs Flabell 0.202975 0.015316 13.252744 Embletonia vs Cuthona -0.004145 0.005498 -0.753978 Embletonia vs Cuthona -0.006551 0.005427 -1.207122 Embletonia vs Cuthona -0.002405 0.005435 -0.442379 Embletonia vs Eubranc -0.001179 0.005657 -0.208402 Embletonia vs Eubranc -0.003460 0.005511 -0.627787

246
Embletonia vs Tergipe -0.016697 0.006211 -2.688442 Embletonia vs Flabell 0.009905 0.004649 2.130740 Embletonia vs Flabell -0.001778 0.005546 -0.320544 Embletonia vs Flabell -0.017326 0.005456 -3.175373 Embletonia vs Flabell -0.055456 0.007900 -7.019679 Embletonia vs Flabell -0.040157 0.007026 -5.715467 Cuthona_na vs Cuthona -0.001187 0.002671 -0.444444 Cuthona_na vs Cuthona 0.000556 0.002728 0.203783 Cuthona_na vs Eubranc 0.002367 0.004086 0.579282 Cuthona_na vs Eubranc 0.001278 0.004303 0.297041 Cuthona_na vs Tergipe -0.014870 0.004609 -3.226033 Cuthona_na vs Flabell 0.012771 0.004406 2.898265 Cuthona_na vs Flabell 0.002397 0.004053 0.591419 Cuthona_na vs Flabell -0.011331 0.005426 -2.088367 Cuthona_na vs Flabell -0.054651 0.007720 -7.078811 Cuthona_na vs Flabell -0.035841 0.006821 -5.254726 Cuthona_ca vs Cuthona 0.001746 0.002866 0.609165 Cuthona_ca vs Eubranc 0.003565 0.004143 0.860551 Cuthona_ca vs Eubranc 0.003069 0.004244 0.723284 Cuthona_ca vs Tergipe -0.011894 0.004662 -2.551435 Cuthona_ca vs Flabell 0.014590 0.004315 3.381610 Cuthona_ca vs Flabell 0.004198 0.004280 0.980638 Cuthona_ca vs Flabell -0.008998 0.005456 -1.649072 Cuthona_ca vs Flabell -0.052927 0.007904 -6.696154 Cuthona_ca vs Flabell -0.034146 0.006898 -4.949932 Cuthona_si vs Eubranc 0.001810 0.004049 0.446964 Cuthona_si vs Eubranc 0.000721 0.004108 0.175625 Cuthona_si vs Tergipe -0.015447 0.004766 -3.241360 Cuthona_si vs Flabell 0.012215 0.004335 2.817556 Cuthona_si vs Flabell 0.001243 0.004275 0.290677 Cuthona_si vs Flabell -0.010729 0.005470 -1.961434 Cuthona_si vs Flabell -0.055223 0.007720 -7.152908 Cuthona_si vs Flabell -0.037030 0.006936 -5.338703 Eubranchus vs Eubranc 0.000090 0.004154 0.021600 Eubranchus vs Tergipe -0.015484 0.005151 -3.006117 Eubranchus vs Flabell 0.010463 0.004567 2.290833 Eubranchus vs Flabell 0.000605 0.004585 0.131929 Eubranchus vs Flabell -0.013204 0.005635 -2.343148 Eubranchus vs Flabell -0.054112 0.007651 -7.072374 Eubranchus vs Flabell -0.039530 0.007048 -5.608510 Eubranchus vs Tergipe -0.016178 0.004901 -3.301290 Eubranchus vs Flabell 0.011524 0.004502 2.559666 Eubranchus vs Flabell 0.000522 0.004602 0.113439 Eubranchus vs Flabell -0.012668 0.005643 -2.244673 Eubranchus vs Flabell -0.054824 0.007737 -7.086015 Eubranchus vs Flabell -0.038411 0.006979 -5.503480 Tergipes_t vs Flabell 0.027693 0.005325 5.200767 Tergipes_t vs Flabell 0.017403 0.005222 3.332679 Tergipes_t vs Flabell 0.002381 0.006320 0.376753 Tergipes_t vs Flabell -0.036174 0.008050 -4.493627 Tergipes_t vs Flabell -0.022181 0.007096 -3.126000 Flabellina vs Flabell -0.009919 0.004478 -2.215180 Flabellina vs Flabell -0.023641 0.005112 -4.624248 Flabellina vs Flabell -0.065636 0.007496 -8.756590 Flabellina vs Flabell -0.049247 0.006328 -7.782396 Flabellina vs Flabell -0.010899 0.005513 -1.976869 Flabellina vs Flabell -0.054573 0.007833 -6.966844 Flabellina vs Flabell -0.038072 0.006803 -5.596533 Flabellina vs Flabell -0.041577 0.007170 -5.798705 Flabellina vs Flabell -0.027027 0.006407 -4.218318 Flabellina vs Flabell 0.013877 0.007783 1.782959

247
1 2 3 4 5 6 7 8 9 10 11
1 Bathydoris clavigera 0,0625 0,0664 0,0631 0,4336 0,4163 0,3913 0,1610 0,1605 0,1680 0,1666 2 Chromodoris krohni 0,0506 0,0208 0,0132 0,3641 0,3525 0,3251 0,1412 0,1376 0,1420 0,1410 3 Dendrodoris fumata 0,0532 0,0191 0,0228 0,3805 0,3578 0,3267 0,1311 0,1286 0,1335 0,1325 4 Hypselodoris villafranca 0,0511 0,0124 0,0208 0,3663 0,3565 0,3330 0,1365 0,1354 0,1386 0,1377 5 Dendronotus dalli 0,1757 0,1596 0,1647 0,1606 0,0889 0,0793 0,4491 0,4370 0,4702 0,4483 6 Cuthona nana 0,1746 0,1601 0,1611 0,1608 0,0640 0,0320 0,4000 0,3992 0,4171 0,4036 7 Flabellina babai 0,1693 0,1528 0,1534 0,1548 0,0585 0,0280 0,3687 0,3597 0,3898 0,3745 8 Pleurobranchus peroni 0,0979 0,0896 0,0852 0,0878 0,1804 0,1713 0,1652 0,0179 0,0173 0,0178 9 Berthellina citrina 0,0979 0,0879 0,0840 0,0872 0,1776 0,1707 0,1624 0,0164 0,0166 0,0160
10 Bathyberthella antarctica 0,1016 0,0905 0,0865 0,0891 0,1838 0,1752 0,1691 0,0159 0,0153 0,0179 11 Euselenops luniceps 0,1004 0,0898 0,0859 0,0885 0,1803 0,1729 0,1668 0,0165 0,0148 0,0165 12 Tompthompsonia antarctica 0,0968 0,0879 0,0840 0,0867 0,1839 0,1758 0,1708 0,0208 0,0186 0,0214 0,0159 13 Berthella stellata 0,1694 0,1592 0,1588 0,1619 0,2081 0,2076 0,2035 0,1297 0,1292 0,1322 0,1317 14 Berthella platei 0,1218 0,1130 0,1095 0,1140 0,1858 0,1811 0,1760 0,0666 0,0627 0,0662 0,0622 15 Pupa solidula 0,0960 0,0911 0,0871 0,0891 0,1805 0,1731 0,1757 0,0840 0,0805 0,0847 0,0813 16 Hydatina physis 0,0979 0,0937 0,0891 0,0923 0,1832 0,1770 0,1773 0,0838 0,0809 0,0852 0,0835 17 Bullina lineata 0,0960 0,0901 0,0890 0,0887 0,1793 0,1737 0,1738 0,0837 0,0802 0,0850 0,0827 18 Acteon sp. 0,0955 0,0947 0,0916 0,0961 0,1923 0,1891 0,1890 0,0814 0,0819 0,0855 0,0822 19 Haminoea hydatis 0,0989 0,0969 0,0950 0,0967 0,1923 0,1869 0,1874 0,0785 0,0790 0,0820 0,0793 20 Cylichna gelida 0,0925 0,0927 0,0892 0,0913 0,1883 0,1855 0,1831 0,0778 0,0777 0,0814 0,0780 21 Scaphander lignarius 0,0947 0,0914 0,0896 0,0918 0,1884 0,1872 0,1848 0,0782 0,0781 0,0818 0,0784 22 cf. Retusa sp. 0,0954 0,0974 0,0938 0,0949 0,1936 0,1897 0,1884 0,0829 0,0824 0,0860 0,0821 23 Bulla cf. striata 0,0908 0,0916 0,0892 0,0902 0,1867 0,1857 0,1827 0,0784 0,0772 0,0814 0,0786 24 Smaragdinella sp. 0,0954 0,0962 0,0927 0,0948 0,1878 0,1866 0,1836 0,0785 0,0790 0,0826 0,0793 25 Phanerophthalmus smaragdinus 0,0954 0,0962 0,0927 0,0948 0,1908 0,1873 0,1849 0,0778 0,0778 0,0820 0,0786 26 Bullacta exarata 0,0942 0,0927 0,0892 0,0924 0,1888 0,1872 0,1830 0,0767 0,0760 0,0808 0,0774 27 Atys semistriatus 0,0960 0,0957 0,0921 0,0942 0,1898 0,1869 0,1833 0,0790 0,0795 0,0832 0,0798 28 Odontoglaja guamensis 0,0931 0,0935 0,0916 0,0937 0,1923 0,1872 0,1870 0,0768 0,0773 0,0815 0,0787 29 Navanax inermis 0,0937 0,0946 0,0904 0,0949 0,1923 0,1868 0,1879 0,0807 0,0812 0,0849 0,0815 30 Chelidonura inornata 0,0931 0,0946 0,0898 0,0937 0,1904 0,1872 0,1859 0,0818 0,0812 0,0854 0,0820 31 Philinopsis pilsbryi 0,0936 0,0945 0,0887 0,0948 0,1895 0,1880 0,1855 0,0795 0,0789 0,0837 0,0809 32 Philinoglossa praelongata 0,0932 0,0929 0,0894 0,0904 0,1913 0,1894 0,1875 0,0763 0,0762 0,0781 0,0759 33 Sagaminopteron psychedelicum 0,0886 0,0938 0,0892 0,0919 0,1896 0,1882 0,1859 0,0779 0,0778 0,0826 0,0798 34 Toledonia globosa 0,0937 0,0911 0,0881 0,0897 0,1889 0,1857 0,1838 0,0778 0,0743 0,0796 0,0763 35 Diaphana cf. inflata 0,0955 0,0952 0,0922 0,0939 0,1910 0,1858 0,1851 0,0780 0,0779 0,0810 0,0788 36 Newnesia antarctica 0,0960 0,0922 0,0882 0,0920 0,1908 0,1840 0,1833 0,0796 0,0755 0,0809 0,0775 37 Colpodaspis thomsoni 0,0939 0,0952 0,0906 0,0950 0,1913 0,1878 0,1831 0,0836 0,0819 0,0867 0,0833 38 Runcina adriatica 0,0964 0,0927 0,0868 0,0935 0,1934 0,1895 0,1872 0,0783 0,0760 0,0830 0,0786 39 Unela glandulifera 0,0915 0,0865 0,0813 0,0863 0,1905 0,1844 0,1837 0,0727 0,0704 0,0763 0,0729 40 Pontohedyle milatchevitchi 0,0937 0,0900 0,0842 0,0904 0,1918 0,1856 0,1844 0,0722 0,0710 0,0752 0,0736 41 Hedylopsis spiculifera 0,0898 0,0854 0,0802 0,0841 0,1890 0,1834 0,1815 0,0704 0,0698 0,0740 0,0712 42 Parahedyle cryptophthalma 0,0914 0,0865 0,0813 0,0862 0,1904 0,1843 0,1836 0,0727 0,0704 0,0762 0,0729 43 Umbraculum mediterraneum 0,0957 0,0926 0,0873 0,0922 0,1956 0,1891 0,1872 0,0758 0,0752 0,0805 0,0771 44 Tylodina perversa 0,0952 0,0921 0,0874 0,0918 0,1943 0,1896 0,1888 0,0743 0,0748 0,0779 0,0745 45 Akera bullata 0,0910 0,0867 0,0814 0,0881 0,1873 0,1852 0,1804 0,0751 0,0717 0,0787 0,0736 46 Aplysia californica 0,0888 0,0878 0,0825 0,0864 0,1897 0,1854 0,1823 0,0734 0,0716 0,0758 0,0730 47 Dolabella auricularia 0,0887 0,0849 0,0802 0,0858 0,1891 0,1872 0,1824 0,0746 0,0711 0,0770 0,0742 48 Siphonota geographica 0,0905 0,0860 0,0819 0,0847 0,1878 0,1836 0,1805 0,0735 0,0717 0,0759 0,0731 49 Bursatella leachii 0,0880 0,0878 0,0820 0,0864 0,1885 0,1846 0,1804 0,0729 0,0700 0,0753 0,0714 50 Petalifera petalifera 0,0882 0,0867 0,0809 0,0854 0,1879 0,1851 0,1820 0,0735 0,0712 0,0754 0,0726 51 Dolabrifera brazieri 0,0882 0,0867 0,0809 0,0854 0,1879 0,1852 0,1815 0,0735 0,0712 0,0754 0,0726 52 Stylocheilus longicauda 0,0882 0,0868 0,0810 0,0866 0,1886 0,1864 0,1821 0,0747 0,0712 0,0766 0,0738 53 Clio pyramidata 0,1028 0,0979 0,0926 0,0972 0,1956 0,1916 0,1861 0,0852 0,0840 0,0871 0,0861 54 Diacria quadridentata 0,1040 0,1038 0,0996 0,1012 0,1965 0,1958 0,1921 0,0881 0,0869 0,0888 0,0878 55 Creseis acicula 0,1092 0,1067 0,1019 0,1054 0,1896 0,1881 0,1838 0,0903 0,0886 0,0928 0,0906 56 Spongiobranchaea australis 0,0934 0,0913 0,0860 0,0911 0,1911 0,1872 0,1840 0,0769 0,0745 0,0787 0,0754 57 Calliopaea bellula 0,1058 0,1039 0,1003 0,1047 0,1989 0,1900 0,1893 0,0875 0,0858 0,0900 0,0855 58 Limapontia nigra 0,0979 0,0961 0,0908 0,0946 0,1935 0,1877 0,1852 0,0774 0,0768 0,0803 0,0776 59 Thuridilla bayeri 0,0966 0,0909 0,0870 0,0909 0,1893 0,1822 0,1795 0,0727 0,0710 0,0746 0,0718
IV Distanzen Opisthobranchia-Datensatz (18S rDNA) Dargestellt sind die beobachteten (unten) und korrigierten (oben) Distanzen der 18S rDNA-Sequenzen. Die Parameter des für die Korrektur der beobachteten Distanzen verwendeten Sequenzevolutionsmodells wurden mit Hilfe hierarchischer likelihood ratio tests (Modeltest, Posada & Crandall 1998) ermittelt

248
1 2 3 4 5 6 7 8 9 10 11 60 Elysia viridis 0,0958 0,0886 0,0863 0,0895 0,1901 0,1848 0,1827 0,0743 0,0737 0,0767 0,0739 61 Elysiella pusilla 0,0981 0,0909 0,0851 0,0895 0,1884 0,1859 0,1815 0,0737 0,0719 0,0761 0,0733 62 Bosellia mimetica 0,0986 0,0908 0,0856 0,0883 0,1885 0,1848 0,1827 0,0713 0,0707 0,0738 0,0715 63 Cyerce nigricans 0,0962 0,0890 0,0850 0,0876 0,1864 0,1817 0,1796 0,0725 0,0713 0,0749 0,0715 64 Placobranchus ocellatus 0,1014 0,0919 0,0879 0,0906 0,1895 0,1847 0,1802 0,0702 0,0691 0,0726 0,0709 65 Placida cremoniana 0,0975 0,0897 0,0868 0,0918 0,1883 0,1830 0,1805 0,0748 0,0725 0,0778 0,0744 66 Polybranchia orientalis 0,0940 0,0891 0,0834 0,0860 0,1858 0,1830 0,1816 0,0720 0,0708 0,0744 0,0710 67 Onchidella floridana 0,0885 0,0859 0,0784 0,0851 0,1879 0,1820 0,1789 0,0727 0,0715 0,0773 0,0746 68 Onchidium verrucosum 0,0901 0,0858 0,0794 0,0844 0,1868 0,1812 0,1780 0,0732 0,0715 0,0773 0,0740 69 Siphonaria alternata 0,0891 0,0847 0,0801 0,0834 0,1861 0,1815 0,1801 0,0703 0,0680 0,0728 0,0694 70 Cepaea hortensis 0,0932 0,0883 0,0836 0,0869 0,1891 0,1858 0,1821 0,0744 0,0721 0,0763 0,0735 71 Lymnaea stagnalis 0,0949 0,0929 0,0906 0,0911 0,1918 0,1860 0,1863 0,0822 0,0810 0,0852 0,0807 72 Cassidula nucleus 0,0893 0,0855 0,0786 0,0835 0,1891 0,1845 0,1831 0,0727 0,0716 0,0763 0,0741 73 Melampus luteus 0,0892 0,0849 0,0796 0,0853 0,1870 0,1832 0,1801 0,0710 0,0681 0,0746 0,0729 74 Auriculinella bidentata 0,0910 0,0891 0,0844 0,0882 0,1882 0,1855 0,1801 0,0745 0,0716 0,0769 0,0753 75 Ovatella firminii 0,0888 0,0850 0,0792 0,0830 0,1886 0,1840 0,1826 0,0716 0,0704 0,0752 0,0730 76 Pythia scarabaeus 0,0886 0,0865 0,0801 0,0852 0,1883 0,1836 0,1810 0,0721 0,0709 0,0762 0,0740 77 Pyramidella sulcata 0,0879 0,0893 0,0852 0,0873 0,1921 0,1848 0,1829 0,0732 0,0731 0,0773 0,0739 78 Odostomia sp. 0,0885 0,0887 0,0852 0,0867 0,1909 0,1854 0,1835 0,0743 0,0731 0,0773 0,0739 79 Littoraria undulata 0,1238 0,1222 0,1231 0,1230 0,2092 0,2053 0,1996 0,1156 0,1133 0,1193 0,1166 80 Nodilittorina punctata 0,1223 0,1204 0,1214 0,1207 0,2067 0,2016 0,1960 0,1157 0,1129 0,1206 0,1162 81 Littorina striata 0,1213 0,1184 0,1199 0,1198 0,2019 0,1989 0,1951 0,1097 0,1063 0,1140 0,1101 82 Littorina saxatilis 0,1207 0,1219 0,1217 0,1228 0,2085 0,2028 0,2006 0,1135 0,1107 0,1184 0,1150 83 Littorina littorea 0,1220 0,1214 0,1218 0,1223 0,2069 0,2035 0,1989 0,1126 0,1109 0,1174 0,1147 84 Melaraphe neritoides 0,1251 0,1239 0,1254 0,1265 0,2082 0,2060 0,2004 0,1178 0,1143 0,1221 0,1199 85 Crepidula adunca 0,1196 0,1190 0,1182 0,1204 0,2025 0,1995 0,1960 0,1101 0,1095 0,1144 0,1111 86 Fasciolaria lignaria 0,1228 0,1218 0,1215 0,1238 0,2070 0,2027 0,2005 0,1143 0,1109 0,1175 0,1152 87 Pisania striata 0,1229 0,1195 0,1191 0,1214 0,2043 0,2030 0,1994 0,1167 0,1127 0,1192 0,1171 88 Bursa rana 0,1199 0,1181 0,1191 0,1196 0,2068 0,2004 0,1981 0,1115 0,1087 0,1146 0,1119 89 Truncatella guerinii 0,1222 0,1207 0,1210 0,1210 0,2080 0,2000 0,2001 0,1193 0,1147 0,1212 0,1179 90 Viviparus georgianus 0,1299 0,1249 0,1252 0,1257 0,2065 0,1994 0,1981 0,1263 0,1234 0,1277 0,1238 91 Balcis eburnea 0,1245 0,1244 0,1254 0,1241 0,2072 0,2033 0,1991 0,1157 0,1145 0,1212 0,1185
12 13 14 15 16 17 18 19 20 21 22
1 Bathydoris clavigera 0,1554 0,4199 0,2225 0,1481 0,1528 0,1491 0,1449 0,1532 0,1394 0,1424 0,1452 2 Chromodoris krohni 0,1367 0,3787 0,2006 0,1344 0,1399 0,1326 0,1382 0,1449 0,1366 0,1335 0,1455 3 Dendrodoris fumata 0,1286 0,3787 0,1908 0,1271 0,1307 0,1305 0,1346 0,1415 0,1302 0,1303 0,1388 4 Hypselodoris villafranca 0,1339 0,3884 0,2012 0,1300 0,1367 0,1295 0,1420 0,1434 0,1332 0,1334 0,1399 5 Dendronotus dalli 0,4655 0,6511 0,4652 0,4408 0,4497 0,4312 0,5020 0,4953 0,4783 0,4751 0,5055 6 Cuthona nana 0,4196 0,6313 0,4445 0,4014 0,4147 0,4008 0,4708 0,4586 0,4537 0,4574 0,4742 7 Flabellina babai 0,3923 0,6024 0,4192 0,4117 0,4160 0,4020 0,4658 0,4546 0,4392 0,4426 0,4649 8 Pleurobranchus peroni 0,0234 0,2614 0,0946 0,1272 0,1253 0,1244 0,1207 0,1121 0,1137 0,1134 0,1238 9 Berthellina citrina 0,0207 0,2613 0,0868 0,1194 0,1185 0,1167 0,1202 0,1124 0,1125 0,1122 0,1213
10 Bathyberthella antarctica 0,0239 0,2712 0,0935 0,1274 0,1265 0,1261 0,1284 0,1186 0,1203 0,1201 0,1295 11 Euselenops luniceps 0,0175 0,2707 0,0864 0,1208 0,1234 0,1215 0,1208 0,1129 0,1130 0,1128 0,1209 12 Tompthompsonia antarctica 0,2790 0,0953 0,1215 0,1211 0,1227 0,1183 0,1157 0,1132 0,1141 0,1221 13 Berthella stellata 0,1329 0,3296 0,3574 0,3569 0,3570 0,3700 0,3671 0,3585 0,3583 0,3815 14 Berthella platei 0,0666 0,1445 0,1818 0,1785 0,1839 0,1878 0,1764 0,1789 0,1733 0,1908 15 Pupa solidula 0,0816 0,1538 0,1069 0,0137 0,0106 0,0643 0,0605 0,0576 0,0592 0,0636 16 Hydatina physis 0,0821 0,1550 0,1064 0,0130 0,0046 0,0577 0,0526 0,0505 0,0502 0,0554 17 Bullina lineata 0,0831 0,1548 0,1085 0,0102 0,0045 0,0550 0,0499 0,0478 0,0492 0,0527 18 Acteon sp. 0,0807 0,1589 0,1097 0,0513 0,0468 0,0450 0,0209 0,0150 0,0149 0,0195 19 Haminoea hydatis 0,0801 0,1588 0,1057 0,0490 0,0433 0,0416 0,0192 0,0107 0,0125 0,0144 20 Cylichna gelida 0,0777 0,1558 0,1056 0,0467 0,0416 0,0398 0,0141 0,0102 0,0064 0,0100 21 Scaphander lignarius 0,0786 0,1564 0,1037 0,0480 0,0417 0,0411 0,0142 0,0119 0,0062 0,0131 22 cf. Retusa sp. 0,0824 0,1611 0,1102 0,0507 0,0450 0,0433 0,0181 0,0136 0,0096 0,0125 23 Bulla cf. striata 0,0778 0,1565 0,1073 0,0472 0,0422 0,0398 0,0153 0,0141 0,0079 0,0108 0,0124 24 Smaragdinella sp. 0,0801 0,1583 0,1063 0,0507 0,0456 0,0438 0,0164 0,0113 0,0079 0,0108 0,0147 25 Phanerophthalmus smaragdinus 0,0795 0,1569 0,1061 0,0506 0,0456 0,0427 0,0192 0,0130 0,0107 0,0136 0,0175 26 Bullacta exarata 0,0783 0,1546 0,1038 0,0484 0,0433 0,0415 0,0158 0,0096 0,0056 0,0097 0,0136 27 Atys semistriatus 0,0807 0,1576 0,1062 0,0501 0,0450 0,0433 0,0170 0,0090 0,0056 0,0096 0,0124 28 Odontoglaja guamensis 0,0772 0,1576 0,1068 0,0518 0,0484 0,0455 0,0119 0,0186 0,0136 0,0153 0,0192 29 Navanax inermis 0,0806 0,1588 0,1103 0,0507 0,0473 0,0455 0,0068 0,0164 0,0124 0,0130 0,0164 30 Chelidonura inornata 0,0818 0,1594 0,1107 0,0495 0,0478 0,0438 0,0124 0,0215 0,0141 0,0159 0,0181 31 Philinopsis pilsbryi 0,0795 0,1576 0,1079 0,0495 0,0467 0,0450 0,0085 0,0198 0,0124 0,0142 0,0175 32 Philinoglossa praelongata 0,0756 0,1554 0,1057 0,0484 0,0468 0,0450 0,0226 0,0249 0,0187 0,0205 0,0220 33 Sagaminopteron psychedelicum 0,0784 0,1577 0,1085 0,0518 0,0473 0,0461 0,0203 0,0226 0,0158 0,0198 0,0203 34 Toledonia globosa 0,0766 0,1552 0,1027 0,0484 0,0416 0,0404 0,0204 0,0175 0,0107 0,0102 0,0181 35 Diaphana cf. inflata 0,0773 0,1567 0,1064 0,0479 0,0416 0,0416 0,0170 0,0164 0,0113 0,0097 0,0141 36 Newnesia antarctica 0,0778 0,1548 0,1022 0,0490 0,0427 0,0421 0,0232 0,0209 0,0147 0,0136 0,0209

249
12 13 14 15 16 17 18 19 20 21 22 37 Colpodaspis thomsoni 0,0824 0,1626 0,1106 0,0536 0,0491 0,0474 0,0266 0,0260 0,0192 0,0238 0,0260 38 Runcina adriatica 0,0794 0,1532 0,1030 0,0498 0,0441 0,0441 0,0346 0,0346 0,0289 0,0302 0,0340 39 Unela glandulifera 0,0737 0,1503 0,1019 0,0421 0,0393 0,0376 0,0277 0,0300 0,0237 0,0250 0,0311 40 Pontohedyle milatchevitchi 0,0727 0,1501 0,1044 0,0428 0,0411 0,0399 0,0307 0,0329 0,0289 0,0291 0,0346 41 Hedylopsis spiculifera 0,0720 0,1496 0,1008 0,0404 0,0376 0,0359 0,0289 0,0328 0,0254 0,0267 0,0317 42 Parahedyle cryptophthalma 0,0737 0,1502 0,1019 0,0421 0,0393 0,0376 0,0277 0,0300 0,0237 0,0250 0,0311 43 Umbraculum mediterraneum 0,0774 0,1563 0,1022 0,0456 0,0394 0,0394 0,0340 0,0363 0,0295 0,0319 0,0352 44 Tylodina perversa 0,0749 0,1516 0,0996 0,0461 0,0399 0,0398 0,0334 0,0362 0,0300 0,0313 0,0351 45 Akera bullata 0,0733 0,1520 0,1003 0,0433 0,0394 0,0376 0,0278 0,0289 0,0210 0,0239 0,0284 46 Aplysia californica 0,0727 0,1508 0,0997 0,0433 0,0371 0,0365 0,0266 0,0272 0,0198 0,0222 0,0266 47 Dolabella auricularia 0,0734 0,1521 0,1009 0,0428 0,0382 0,0365 0,0278 0,0300 0,0221 0,0245 0,0295 48 Siphonota geographica 0,0728 0,1515 0,0993 0,0434 0,0371 0,0366 0,0278 0,0289 0,0210 0,0222 0,0283 49 Bursatella leachii 0,0706 0,1510 0,0994 0,0400 0,0377 0,0360 0,0261 0,0295 0,0216 0,0228 0,0290 50 Petalifera petalifera 0,0723 0,1528 0,1011 0,0417 0,0366 0,0349 0,0244 0,0272 0,0199 0,0211 0,0261 51 Dolabrifera brazieri 0,0723 0,1522 0,1011 0,0417 0,0365 0,0348 0,0250 0,0267 0,0199 0,0211 0,0261 52 Stylocheilus longicauda 0,0735 0,1522 0,1011 0,0428 0,0377 0,0360 0,0250 0,0289 0,0216 0,0216 0,0278 53 Clio pyramidata 0,0846 0,1610 0,1093 0,0577 0,0498 0,0510 0,0370 0,0409 0,0336 0,0331 0,0392 54 Diacria quadridentata 0,0863 0,1634 0,1136 0,0531 0,0536 0,0513 0,0380 0,0414 0,0368 0,0359 0,0419 55 Creseis acicula 0,0897 0,1614 0,1141 0,0643 0,0660 0,0643 0,0593 0,0598 0,0547 0,0555 0,0621 56 Spongiobranchaea australis 0,0756 0,1549 0,1026 0,0467 0,0417 0,0412 0,0323 0,0346 0,0272 0,0273 0,0334 57 Calliopaea bellula 0,0857 0,1607 0,1187 0,0603 0,0571 0,0565 0,0488 0,0533 0,0465 0,0479 0,0505 58 Limapontia nigra 0,0767 0,1547 0,1096 0,0519 0,0469 0,0469 0,0443 0,0454 0,0386 0,0405 0,0443 59 Thuridilla bayeri 0,0710 0,1534 0,1055 0,0478 0,0457 0,0428 0,0415 0,0442 0,0386 0,0393 0,0453 60 Elysia viridis 0,0726 0,1540 0,1071 0,0477 0,0445 0,0427 0,0402 0,0436 0,0368 0,0376 0,0430 61 Elysiella pusilla 0,0725 0,1523 0,1042 0,0472 0,0439 0,0421 0,0402 0,0419 0,0351 0,0359 0,0407 62 Bosellia mimetica 0,0702 0,1529 0,1041 0,0466 0,0433 0,0415 0,0419 0,0418 0,0368 0,0375 0,0424 63 Cyerce nigricans 0,0713 0,1528 0,1035 0,0437 0,0404 0,0370 0,0385 0,0401 0,0345 0,0352 0,0412 64 Placobranchus ocellatus 0,0702 0,1510 0,1047 0,0545 0,0500 0,0477 0,0470 0,0469 0,0418 0,0427 0,0486 65 Placida cremoniana 0,0736 0,1512 0,1058 0,0482 0,0421 0,0404 0,0385 0,0413 0,0345 0,0364 0,0418 66 Polybranchia orientalis 0,0714 0,1541 0,1053 0,0432 0,0444 0,0421 0,0407 0,0435 0,0367 0,0375 0,0424 67 Onchidella floridana 0,0737 0,1535 0,1013 0,0410 0,0382 0,0376 0,0312 0,0317 0,0249 0,0267 0,0312 68 Onchidium verrucosum 0,0737 0,1528 0,1013 0,0421 0,0387 0,0364 0,0283 0,0289 0,0215 0,0233 0,0278 69 Siphonaria alternata 0,0686 0,1512 0,0984 0,0376 0,0359 0,0325 0,0250 0,0261 0,0204 0,0211 0,0272 70 Cepaea hortensis 0,0732 0,1540 0,1015 0,0427 0,0371 0,0359 0,0272 0,0272 0,0192 0,0205 0,0255 71 Lymnaea stagnalis 0,0811 0,1567 0,1082 0,0535 0,0469 0,0457 0,0409 0,0409 0,0341 0,0359 0,0403 72 Cassidula nucleus 0,0720 0,1537 0,1020 0,0411 0,0360 0,0354 0,0296 0,0307 0,0244 0,0257 0,0312 73 Melampus luteus 0,0720 0,1530 0,1002 0,0399 0,0371 0,0348 0,0284 0,0318 0,0244 0,0256 0,0306 74 Auriculinella bidentata 0,0744 0,1536 0,1020 0,0417 0,0366 0,0348 0,0329 0,0306 0,0250 0,0274 0,0295 75 Ovatella firminii 0,0709 0,1526 0,1020 0,0411 0,0372 0,0354 0,0290 0,0301 0,0239 0,0251 0,0307 76 Pythia scarabaeus 0,0719 0,1535 0,1018 0,0399 0,0360 0,0342 0,0290 0,0301 0,0227 0,0240 0,0301 77 Pyramidella sulcata 0,0732 0,1557 0,1002 0,0410 0,0376 0,0342 0,0346 0,0345 0,0283 0,0307 0,0317 78 Odostomia sp. 0,0732 0,1563 0,1009 0,0439 0,0399 0,0370 0,0312 0,0323 0,0249 0,0273 0,0306 79 Littoraria undulata 0,1144 0,1759 0,1348 0,0918 0,0891 0,0873 0,0862 0,0856 0,0821 0,0829 0,0856 80 Nodilittorina punctata 0,1157 0,1739 0,1338 0,0889 0,0873 0,0855 0,0843 0,0837 0,0809 0,0817 0,0849 81 Littorina striata 0,1091 0,1702 0,1277 0,0872 0,0862 0,0844 0,0798 0,0803 0,0775 0,0771 0,0798 82 Littorina saxatilis 0,1135 0,1761 0,1349 0,0877 0,0849 0,0832 0,0785 0,0802 0,0773 0,0764 0,0808 83 Littorina littorea 0,1132 0,1755 0,1353 0,0892 0,0863 0,0846 0,0794 0,0816 0,0788 0,0773 0,0828 84 Melaraphe neritoides 0,1182 0,1760 0,1376 0,0856 0,0840 0,0822 0,0839 0,0804 0,0799 0,0807 0,0827 85 Crepidula adunca 0,1112 0,1705 0,1323 0,0859 0,0854 0,0830 0,0814 0,0808 0,0779 0,0776 0,0819 86 Fasciolaria lignaria 0,1137 0,1751 0,1330 0,0878 0,0861 0,0843 0,0838 0,0837 0,0809 0,0794 0,0843 87 Pisania striata 0,1160 0,1721 0,1361 0,0885 0,0863 0,0845 0,0853 0,0852 0,0823 0,0808 0,0863 88 Bursa rana 0,1109 0,1715 0,1320 0,0862 0,0840 0,0822 0,0846 0,0845 0,0816 0,0813 0,0868 89 Truncatella guerinii 0,1180 0,1757 0,1350 0,0858 0,0872 0,0854 0,0890 0,0878 0,0832 0,0852 0,0889 90 Viviparus georgianus 0,1254 0,1730 0,1398 0,0899 0,0917 0,0899 0,0901 0,0906 0,0872 0,0869 0,0912 91 Balcis eburnea 0,1172 0,1761 0,1383 0,0936 0,0897 0,0891 0,0858 0,0845 0,0816 0,0807 0,0857
23 24 25 26 27 28 29 30 31 32 33 1 Bathydoris clavigera 0,1338 0,1460 0,1455 0,1430 0,1469 0,1400 0,1410 0,1403 0,1409 0,1414 0,1300 2 Chromodoris krohni 0,1322 0,1440 0,1443 0,1365 0,1427 0,1360 0,1379 0,1399 0,1390 0,1363 0,1373 3 Dendrodoris fumata 0,1279 0,1373 0,1368 0,1300 0,1361 0,1334 0,1312 0,1305 0,1278 0,1299 0,1282 4 Hypselodoris villafranca 0,1289 0,1405 0,1408 0,1355 0,1391 0,1366 0,1385 0,1376 0,1391 0,1309 0,1327 5 Dendronotus dalli 0,4721 0,4752 0,4957 0,4839 0,4833 0,4983 0,4956 0,4928 0,4854 0,4977 0,4813 6 Cuthona nana 0,4519 0,4564 0,4630 0,4585 0,4576 0,4583 0,4562 0,4598 0,4637 0,4752 0,4641 7 Flabellina babai 0,4354 0,4397 0,4483 0,4363 0,4388 0,4545 0,4557 0,4506 0,4489 0,4636 0,4470 8 Pleurobranchus peroni 0,1146 0,1137 0,1126 0,1099 0,1146 0,1104 0,1187 0,1210 0,1166 0,1110 0,1137 9 Berthellina citrina 0,1113 0,1140 0,1119 0,1080 0,1149 0,1113 0,1183 0,1186 0,1139 0,1101 0,1123
10 Bathyberthella antarctica 0,1197 0,1219 0,1208 0,1179 0,1228 0,1201 0,1263 0,1277 0,1242 0,1145 0,1227 11 Euselenops luniceps 0,1138 0,1146 0,1135 0,1107 0,1155 0,1141 0,1189 0,1201 0,1179 0,1097 0,1158 12 Tompthompsonia antarctica 0,1127 0,1174 0,1163 0,1135 0,1183 0,1116 0,1181 0,1204 0,1160 0,1099 0,1140 13 Berthella stellata 0,3608 0,3665 0,3608 0,3508 0,3637 0,3647 0,3670 0,3713 0,3635 0,3552 0,3618
250
23 24 25 26 27 28 29 30 31 32 33 14 Berthella platei 0,1827 0,1792 0,1793 0,1726 0,1794 0,1809 0,1883 0,1905 0,1829 0,1798 0,1835 15 Pupa solidula 0,0582 0,0635 0,0638 0,0599 0,0627 0,0652 0,0630 0,0620 0,0618 0,0604 0,0651 16 Hydatina physis 0,0510 0,0562 0,0565 0,0527 0,0553 0,0602 0,0581 0,0593 0,0576 0,0578 0,0585 17 Bullina lineata 0,0478 0,0535 0,0522 0,0500 0,0526 0,0558 0,0553 0,0535 0,0549 0,0550 0,0564 18 Acteon sp. 0,0162 0,0175 0,0209 0,0169 0,0181 0,0125 0,0070 0,0131 0,0088 0,0250 0,0222 19 Haminoea hydatis 0,0150 0,0118 0,0137 0,0100 0,0094 0,0202 0,0175 0,0234 0,0214 0,0279 0,0250 20 Cylichna gelida 0,0082 0,0082 0,0113 0,0058 0,0058 0,0143 0,0130 0,0149 0,0130 0,0204 0,0169 21 Scaphander lignarius 0,0112 0,0112 0,0143 0,0100 0,0100 0,0162 0,0135 0,0167 0,0148 0,0223 0,0214 22 cf. Retusa sp. 0,0131 0,0156 0,0189 0,0144 0,0131 0,0208 0,0174 0,0194 0,0187 0,0245 0,0221 23 Bulla cf. striata 0,0112 0,0119 0,0100 0,0112 0,0162 0,0142 0,0143 0,0143 0,0203 0,0176 24 Smaragdinella sp. 0,0107 0,0100 0,0069 0,0057 0,0162 0,0161 0,0169 0,0162 0,0218 0,0183 25 Phanerophthalmus smaragdinus 0,0113 0,0096 0,0093 0,0076 0,0175 0,0187 0,0174 0,0168 0,0245 0,0230 26 Bullacta exarata 0,0096 0,0068 0,0090 0,0058 0,0137 0,0148 0,0168 0,0137 0,0231 0,0196 27 Atys semistriatus 0,0107 0,0057 0,0073 0,0056 0,0174 0,0161 0,0181 0,0162 0,0224 0,0195 28 Odontoglaja guamensis 0,0153 0,0153 0,0164 0,0130 0,0164 0,0069 0,0112 0,0105 0,0222 0,0209 29 Navanax inermis 0,0136 0,0153 0,0175 0,0141 0,0152 0,0068 0,0081 0,0063 0,0221 0,0194 30 Chelidonura inornata 0,0136 0,0158 0,0164 0,0158 0,0169 0,0107 0,0079 0,0075 0,0222 0,0226 31 Philinopsis pilsbryi 0,0136 0,0153 0,0158 0,0130 0,0153 0,0102 0,0062 0,0073 0,0209 0,0194 32 Philinoglossa praelongata 0,0187 0,0198 0,0220 0,0209 0,0204 0,0204 0,0204 0,0204 0,0192 0,0278 33 Sagaminopteron psychedelicum 0,0164 0,0170 0,0209 0,0181 0,0181 0,0192 0,0181 0,0209 0,0181 0,0249 34 Toledonia globosa 0,0147 0,0164 0,0175 0,0113 0,0153 0,0170 0,0187 0,0209 0,0198 0,0249 0,0243 35 Diaphana cf. inflata 0,0136 0,0164 0,0170 0,0136 0,0141 0,0187 0,0158 0,0181 0,0164 0,0238 0,0221 36 Newnesia antarctica 0,0181 0,0198 0,0186 0,0147 0,0175 0,0209 0,0215 0,0249 0,0220 0,0266 0,0277 37 Colpodaspis thomsoni 0,0209 0,0243 0,0266 0,0209 0,0226 0,0277 0,0249 0,0260 0,0238 0,0266 0,0232 38 Runcina adriatica 0,0307 0,0341 0,0329 0,0278 0,0323 0,0318 0,0323 0,0335 0,0312 0,0398 0,0340 39 Unela glandulifera 0,0260 0,0266 0,0283 0,0249 0,0254 0,0277 0,0260 0,0271 0,0271 0,0289 0,0322 40 Pontohedyle milatchevitchi 0,0295 0,0312 0,0306 0,0278 0,0318 0,0295 0,0284 0,0289 0,0284 0,0313 0,0352 41 Hedylopsis spiculifera 0,0260 0,0266 0,0300 0,0266 0,0271 0,0277 0,0277 0,0266 0,0288 0,0278 0,0322 42 Parahedyle cryptophthalma 0,0260 0,0266 0,0283 0,0249 0,0254 0,0277 0,0260 0,0271 0,0271 0,0289 0,0322 43 Umbraculum mediterraneum 0,0295 0,0324 0,0335 0,0295 0,0312 0,0323 0,0306 0,0318 0,0323 0,0329 0,0357 44 Tylodina perversa 0,0311 0,0340 0,0345 0,0311 0,0339 0,0328 0,0305 0,0328 0,0322 0,0340 0,0351 45 Akera bullata 0,0233 0,0261 0,0266 0,0198 0,0250 0,0272 0,0267 0,0250 0,0250 0,0323 0,0284 46 Aplysia californica 0,0221 0,0249 0,0255 0,0198 0,0238 0,0260 0,0255 0,0266 0,0249 0,0306 0,0277 47 Dolabella auricularia 0,0232 0,0250 0,0261 0,0210 0,0249 0,0272 0,0266 0,0272 0,0255 0,0306 0,0283 48 Siphonota geographica 0,0227 0,0250 0,0255 0,0204 0,0238 0,0272 0,0267 0,0284 0,0267 0,0301 0,0300 49 Bursatella leachii 0,0227 0,0244 0,0244 0,0210 0,0244 0,0244 0,0238 0,0238 0,0233 0,0290 0,0284 50 Petalifera petalifera 0,0221 0,0227 0,0244 0,0216 0,0227 0,0238 0,0233 0,0244 0,0238 0,0290 0,0272 51 Dolabrifera brazieri 0,0221 0,0227 0,0238 0,0204 0,0227 0,0238 0,0233 0,0244 0,0238 0,0290 0,0272 52 Stylocheilus longicauda 0,0227 0,0244 0,0250 0,0210 0,0244 0,0244 0,0238 0,0250 0,0233 0,0301 0,0289 53 Clio pyramidata 0,0358 0,0365 0,0387 0,0336 0,0353 0,0381 0,0375 0,0375 0,0358 0,0404 0,0392 54 Diacria quadridentata 0,0385 0,0403 0,0425 0,0368 0,0408 0,0391 0,0385 0,0386 0,0385 0,0426 0,0448 55 Creseis acicula 0,0575 0,0559 0,0581 0,0536 0,0570 0,0564 0,0581 0,0576 0,0570 0,0587 0,0593 56 Spongiobranchaea australis 0,0300 0,0312 0,0306 0,0295 0,0301 0,0312 0,0306 0,0312 0,0312 0,0363 0,0357 57 Calliopaea bellula 0,0471 0,0477 0,0505 0,0460 0,0482 0,0500 0,0488 0,0483 0,0465 0,0482 0,0528 58 Limapontia nigra 0,0403 0,0427 0,0443 0,0380 0,0426 0,0432 0,0432 0,0426 0,0409 0,0426 0,0455 59 Thuridilla bayeri 0,0380 0,0409 0,0420 0,0386 0,0414 0,0374 0,0386 0,0363 0,0408 0,0403 0,0460 60 Elysia viridis 0,0379 0,0402 0,0425 0,0368 0,0408 0,0385 0,0385 0,0379 0,0408 0,0408 0,0430 61 Elysiella pusilla 0,0362 0,0385 0,0408 0,0340 0,0391 0,0379 0,0379 0,0379 0,0379 0,0380 0,0430 62 Bosellia mimetica 0,0379 0,0396 0,0413 0,0362 0,0402 0,0379 0,0396 0,0379 0,0402 0,0385 0,0436 63 Cyerce nigricans 0,0345 0,0368 0,0379 0,0345 0,0373 0,0373 0,0384 0,0356 0,0373 0,0373 0,0430 64 Placobranchus ocellatus 0,0413 0,0447 0,0458 0,0413 0,0441 0,0418 0,0446 0,0430 0,0452 0,0453 0,0492 65 Placida cremoniana 0,0356 0,0368 0,0384 0,0334 0,0373 0,0379 0,0385 0,0390 0,0373 0,0396 0,0424 66 Polybranchia orientalis 0,0379 0,0390 0,0413 0,0367 0,0396 0,0407 0,0407 0,0390 0,0396 0,0390 0,0447 67 Onchidella floridana 0,0261 0,0283 0,0278 0,0266 0,0278 0,0295 0,0283 0,0300 0,0278 0,0335 0,0289 68 Onchidium verrucosum 0,0227 0,0249 0,0260 0,0215 0,0244 0,0266 0,0266 0,0266 0,0266 0,0289 0,0277 69 Siphonaria alternata 0,0210 0,0227 0,0221 0,0210 0,0232 0,0232 0,0232 0,0221 0,0232 0,0272 0,0272 70 Cepaea hortensis 0,0226 0,0232 0,0232 0,0209 0,0215 0,0249 0,0260 0,0260 0,0238 0,0272 0,0289 71 Lymnaea stagnalis 0,0358 0,0387 0,0403 0,0352 0,0381 0,0392 0,0392 0,0409 0,0392 0,0393 0,0409 72 Cassidula nucleus 0,0261 0,0273 0,0272 0,0261 0,0267 0,0250 0,0273 0,0278 0,0273 0,0284 0,0318 73 Melampus luteus 0,0238 0,0267 0,0272 0,0238 0,0272 0,0272 0,0255 0,0255 0,0244 0,0295 0,0289 74 Auriculinella bidentata 0,0239 0,0267 0,0284 0,0244 0,0272 0,0289 0,0306 0,0306 0,0289 0,0290 0,0289 75 Ovatella firminii 0,0256 0,0267 0,0267 0,0256 0,0261 0,0244 0,0267 0,0273 0,0267 0,0279 0,0312 76 Pythia scarabaeus 0,0244 0,0256 0,0261 0,0244 0,0250 0,0244 0,0267 0,0261 0,0255 0,0273 0,0295 77 Pyramidella sulcata 0,0255 0,0306 0,0311 0,0294 0,0311 0,0311 0,0323 0,0294 0,0317 0,0312 0,0351 78 Odostomia sp. 0,0238 0,0272 0,0289 0,0261 0,0278 0,0283 0,0294 0,0278 0,0289 0,0301 0,0317 79 Littoraria undulata 0,0793 0,0834 0,0838 0,0804 0,0822 0,0832 0,0861 0,0850 0,0862 0,0834 0,0816 80 Nodilittorina punctata 0,0780 0,0822 0,0814 0,0792 0,0809 0,0820 0,0855 0,0844 0,0843 0,0827 0,0792 81 Littorina striata 0,0746 0,0782 0,0792 0,0758 0,0787 0,0792 0,0821 0,0810 0,0809 0,0799 0,0769 82 Littorina saxatilis 0,0739 0,0780 0,0756 0,0750 0,0774 0,0790 0,0802 0,0785 0,0785 0,0803 0,0768 83 Littorina littorea 0,0759 0,0789 0,0782 0,0776 0,0788 0,0810 0,0817 0,0805 0,0805 0,0806 0,0776 84 Melaraphe neritoides 0,0764 0,0806 0,0798 0,0798 0,0799 0,0832 0,0839 0,0828 0,0827 0,0829 0,0793 85 Crepidula adunca 0,0744 0,0780 0,0785 0,0773 0,0779 0,0813 0,0819 0,0808 0,0808 0,0809 0,0767 86 Fasciolaria lignaria 0,0785 0,0810 0,0802 0,0803 0,0809 0,0837 0,0843 0,0838 0,0838 0,0839 0,0797 87 Pisania striata 0,0788 0,0818 0,0829 0,0817 0,0823 0,0857 0,0869 0,0858 0,0858 0,0847 0,0795 88 Bursa rana 0,0793 0,0829 0,0828 0,0810 0,0828 0,0833 0,0863 0,0857 0,0868 0,0846 0,0805 89 Truncatella guerinii 0,0814 0,0844 0,0855 0,0815 0,0844 0,0854 0,0884 0,0872 0,0872 0,0862 0,0837

251
90 Viviparus georgianus 0,0843 0,0873 0,0883 0,0877 0,0867 0,0901 0,0901 0,0890 0,0901 0,0909 0,0872 91 Balcis eburnea 0,0787 0,0800 0,0816 0,0804 0,0811 0,0851 0,0863 0,0852 0,0852 0,0835 0,0810
34 35 36 37 38 39 40 41 42 43 44
1 Bathydoris clavigera 0,1414 0,1456 0,1454 0,1399 0,1469 0,1358 0,1399 0,1328 0,1357 0,1480 0,1479 2 Chromodoris krohni 0,1331 0,1420 0,1350 0,1381 0,1366 0,1239 0,1303 0,1223 0,1238 0,1382 0,1380 3 Dendrodoris fumata 0,1278 0,1364 0,1277 0,1299 0,1248 0,1139 0,1195 0,1124 0,1138 0,1275 0,1284 4 Hypselodoris villafranca 0,1298 0,1383 0,1341 0,1372 0,1378 0,1230 0,1303 0,1191 0,1229 0,1378 0,1374 5 Dendronotus dalli 0,4828 0,4947 0,4901 0,4911 0,4959 0,4867 0,4907 0,4809 0,4861 0,5218 0,5180 6 Cuthona nana 0,4509 0,4541 0,4446 0,4574 0,4681 0,4429 0,4442 0,4383 0,4424 0,4749 0,4789 7 Flabellina babai 0,4386 0,4473 0,4366 0,4327 0,4488 0,4336 0,4329 0,4263 0,4331 0,4595 0,4702 8 Pleurobranchus peroni 0,1130 0,1138 0,1158 0,1250 0,1125 0,1032 0,1016 0,0989 0,1031 0,1094 0,1074 9 Berthellina citrina 0,1052 0,1131 0,1071 0,1206 0,1075 0,0979 0,0983 0,0974 0,0978 0,1076 0,1078
10 Bathyberthella antarctica 0,1165 0,1193 0,1182 0,1312 0,1222 0,1094 0,1068 0,1052 0,1094 0,1185 0,1141 11 Euselenops luniceps 0,1093 0,1146 0,1110 0,1235 0,1119 0,1023 0,1027 0,0996 0,1023 0,1106 0,1069 12 Tompthompsonia antarctica 0,1105 0,1122 0,1121 0,1223 0,1143 0,1044 0,1019 0,1018 0,1044 0,1117 0,1082 13 Berthella stellata 0,3538 0,3615 0,3498 0,3869 0,3468 0,3359 0,3321 0,3325 0,3356 0,3646 0,3466 14 Berthella platei 0,1710 0,1798 0,1689 0,1911 0,1694 0,1678 0,1732 0,1656 0,1677 0,1701 0,1635 15 Pupa solidula 0,0599 0,0595 0,0607 0,0683 0,0624 0,0509 0,0516 0,0486 0,0508 0,0572 0,0574 16 Hydatina physis 0,0503 0,0507 0,0519 0,0616 0,0541 0,0467 0,0490 0,0444 0,0467 0,0480 0,0482 17 Bullina lineata 0,0485 0,0506 0,0509 0,0587 0,0538 0,0444 0,0472 0,0422 0,0443 0,0480 0,0483 18 Acteon sp. 0,0220 0,0182 0,0253 0,0302 0,0406 0,0308 0,0348 0,0325 0,0308 0,0392 0,0385 19 Haminoea hydatis 0,0188 0,0177 0,0227 0,0293 0,0407 0,0338 0,0376 0,0376 0,0338 0,0425 0,0426 20 Cylichna gelida 0,0112 0,0119 0,0155 0,0211 0,0332 0,0259 0,0323 0,0281 0,0259 0,0334 0,0341 21 Scaphander lignarius 0,0106 0,0100 0,0143 0,0266 0,0346 0,0273 0,0324 0,0295 0,0273 0,0363 0,0355 22 cf. Retusa sp. 0,0195 0,0151 0,0227 0,0294 0,0397 0,0349 0,0396 0,0360 0,0349 0,0409 0,0409 23 Bulla cf. striata 0,0156 0,0143 0,0193 0,0230 0,0353 0,0287 0,0332 0,0290 0,0287 0,0335 0,0357 24 Smaragdinella sp. 0,0175 0,0176 0,0213 0,0275 0,0399 0,0294 0,0353 0,0297 0,0294 0,0372 0,0393 25 Phanerophthalmus smaragdinus 0,0188 0,0182 0,0201 0,0303 0,0382 0,0316 0,0345 0,0340 0,0316 0,0388 0,0402 26 Bullacta exarata 0,0118 0,0143 0,0154 0,0232 0,0317 0,0273 0,0309 0,0296 0,0273 0,0335 0,0357 27 Atys semistriatus 0,0162 0,0150 0,0187 0,0254 0,0376 0,0280 0,0359 0,0303 0,0280 0,0357 0,0392 28 Odontoglaja guamensis 0,0181 0,0201 0,0226 0,0315 0,0367 0,0307 0,0331 0,0310 0,0307 0,0370 0,0377 29 Navanax inermis 0,0199 0,0168 0,0230 0,0278 0,0372 0,0285 0,0316 0,0308 0,0285 0,0346 0,0347 30 Chelidonura inornata 0,0226 0,0194 0,0271 0,0294 0,0386 0,0300 0,0324 0,0296 0,0300 0,0363 0,0377 31 Philinopsis pilsbryi 0,0212 0,0175 0,0238 0,0266 0,0356 0,0299 0,0315 0,0322 0,0298 0,0368 0,0370 32 Philinoglossa praelongata 0,0277 0,0265 0,0298 0,0303 0,0476 0,0325 0,0356 0,0311 0,0325 0,0380 0,0393 33 Sagaminopteron psychedelicum 0,0269 0,0243 0,0309 0,0258 0,0396 0,0365 0,0403 0,0367 0,0365 0,0416 0,0408 34 Toledonia globosa 0,0131 0,0075 0,0300 0,0302 0,0259 0,0322 0,0254 0,0259 0,0343 0,0314 35 Diaphana cf. inflata 0,0124 0,0155 0,0278 0,0346 0,0315 0,0318 0,0325 0,0315 0,0389 0,0380 36 Newnesia antarctica 0,0073 0,0147 0,0313 0,0287 0,0246 0,0321 0,0267 0,0245 0,0342 0,0342 37 Colpodaspis thomsoni 0,0266 0,0249 0,0277 0,0486 0,0421 0,0454 0,0456 0,0421 0,0485 0,0510 38 Runcina adriatica 0,0267 0,0301 0,0255 0,0403 0,0286 0,0327 0,0287 0,0286 0,0363 0,0356 39 Unela glandulifera 0,0238 0,0283 0,0226 0,0362 0,0256 0,0175 0,0106 0,0000 0,0189 0,0214 40 Pontohedyle milatchevitchi 0,0290 0,0284 0,0290 0,0386 0,0290 0,0164 0,0202 0,0175 0,0292 0,0306 41 Hedylopsis spiculifera 0,0232 0,0289 0,0244 0,0385 0,0256 0,0102 0,0187 0,0106 0,0203 0,0189 42 Parahedyle cryptophthalma 0,0238 0,0283 0,0226 0,0362 0,0256 0,0000 0,0164 0,0102 0,0189 0,0214 43 Umbraculum mediterraneum 0,0301 0,0335 0,0301 0,0403 0,0313 0,0176 0,0262 0,0187 0,0176 0,0169 44 Tylodina perversa 0,0277 0,0328 0,0300 0,0419 0,0307 0,0198 0,0272 0,0175 0,0198 0,0159 45 Akera bullata 0,0204 0,0273 0,0216 0,0341 0,0188 0,0164 0,0222 0,0147 0,0164 0,0204 0,0204 46 Aplysia californica 0,0170 0,0249 0,0193 0,0340 0,0210 0,0164 0,0227 0,0136 0,0164 0,0210 0,0187 47 Dolabella auricularia 0,0193 0,0272 0,0193 0,0324 0,0216 0,0147 0,0205 0,0130 0,0147 0,0176 0,0193 48 Siphonota geographica 0,0176 0,0255 0,0193 0,0341 0,0216 0,0170 0,0227 0,0142 0,0170 0,0216 0,0193 49 Bursatella leachii 0,0182 0,0256 0,0193 0,0335 0,0211 0,0159 0,0199 0,0131 0,0159 0,0199 0,0176 50 Petalifera petalifera 0,0170 0,0250 0,0182 0,0341 0,0234 0,0148 0,0210 0,0119 0,0148 0,0193 0,0170 51 Dolabrifera brazieri 0,0165 0,0244 0,0176 0,0335 0,0228 0,0148 0,0205 0,0125 0,0148 0,0199 0,0176 52 Stylocheilus longicauda 0,0176 0,0256 0,0176 0,0341 0,0228 0,0148 0,0210 0,0131 0,0148 0,0193 0,0182 53 Clio pyramidata 0,0330 0,0382 0,0330 0,0444 0,0370 0,0318 0,0307 0,0285 0,0318 0,0370 0,0353 54 Diacria quadridentata 0,0352 0,0403 0,0374 0,0460 0,0404 0,0375 0,0335 0,0358 0,0374 0,0443 0,0402 55 Creseis acicula 0,0559 0,0588 0,0565 0,0633 0,0561 0,0508 0,0496 0,0491 0,0507 0,0583 0,0559 56 Spongiobranchaea australis 0,0250 0,0318 0,0250 0,0420 0,0284 0,0221 0,0290 0,0215 0,0221 0,0290 0,0295 57 Calliopaea bellula 0,0454 0,0455 0,0415 0,0540 0,0450 0,0386 0,0410 0,0347 0,0386 0,0438 0,0443 58 Limapontia nigra 0,0392 0,0404 0,0392 0,0483 0,0370 0,0307 0,0325 0,0279 0,0307 0,0348 0,0353 59 Thuridilla bayeri 0,0374 0,0409 0,0386 0,0482 0,0382 0,0290 0,0319 0,0239 0,0289 0,0307 0,0313 60 Elysia viridis 0,0363 0,0397 0,0368 0,0470 0,0365 0,0267 0,0313 0,0221 0,0267 0,0290 0,0306 61 Elysiella pusilla 0,0345 0,0385 0,0351 0,0442 0,0330 0,0255 0,0296 0,0216 0,0255 0,0267 0,0306 62 Bosellia mimetica 0,0362 0,0402 0,0379 0,0470 0,0341 0,0261 0,0301 0,0215 0,0261 0,0301 0,0306 63 Cyerce nigricans 0,0334 0,0373 0,0345 0,0447 0,0341 0,0232 0,0267 0,0187 0,0232 0,0289 0,0289 64 Placobranchus ocellatus 0,0396 0,0458 0,0418 0,0509 0,0393 0,0312 0,0375 0,0272 0,0312 0,0352 0,0362 65 Placida cremoniana 0,0334 0,0374 0,0323 0,0447 0,0352 0,0272 0,0295 0,0244 0,0272 0,0306 0,0317 66 Polybranchia orientalis 0,0362 0,0396 0,0368 0,0476 0,0375 0,0261 0,0290 0,0215 0,0261 0,0323 0,0328 67 Onchidella floridana 0,0244 0,0267 0,0244 0,0380 0,0256 0,0170 0,0244 0,0170 0,0170 0,0227 0,0238 68 Onchidium verrucosum 0,0193 0,0249 0,0204 0,0351 0,0222 0,0153 0,0204 0,0130 0,0153 0,0221 0,0238 69 Siphonaria alternata 0,0170 0,0233 0,0187 0,0335 0,0234 0,0130 0,0188 0,0096 0,0130 0,0227 0,0221 70 Cepaea hortensis 0,0210 0,0221 0,0210 0,0357 0,0273 0,0175 0,0244 0,0153 0,0176 0,0244 0,0244 71 Lymnaea stagnalis 0,0324 0,0387 0,0346 0,0460 0,0342 0,0290 0,0353 0,0262 0,0290 0,0331 0,0313 72 Cassidula nucleus 0,0244 0,0290 0,0244 0,0404 0,0273 0,0114 0,0194 0,0125 0,0114 0,0222 0,0233 73 Melampus luteus 0,0238 0,0267 0,0238 0,0364 0,0285 0,0142 0,0205 0,0131 0,0142 0,0210 0,0233
252
34 35 36 37 38 39 40 41 42 43 44 74 Auriculinella bidentata 0,0227 0,0284 0,0216 0,0347 0,0256 0,0159 0,0228 0,0136 0,0159 0,0199 0,0210 75 Ovatella firminii 0,0239 0,0284 0,0239 0,0398 0,0279 0,0102 0,0194 0,0114 0,0102 0,0222 0,0233 76 Pythia scarabaeus 0,0227 0,0273 0,0227 0,0387 0,0268 0,0097 0,0210 0,0108 0,0097 0,0194 0,0216 77 Pyramidella sulcata 0,0266 0,0300 0,0283 0,0385 0,0313 0,0181 0,0273 0,0147 0,0181 0,0204 0,0204 78 Odostomia sp. 0,0232 0,0289 0,0261 0,0374 0,0307 0,0187 0,0273 0,0159 0,0187 0,0221 0,0233 79 Littoraria undulata 0,0804 0,0816 0,0816 0,0852 0,0795 0,0776 0,0789 0,0759 0,0776 0,0819 0,0815 80 Nodilittorina punctata 0,0792 0,0798 0,0804 0,0867 0,0788 0,0775 0,0765 0,0759 0,0775 0,0818 0,0809 81 Littorina striata 0,0758 0,0758 0,0776 0,0845 0,0755 0,0736 0,0736 0,0731 0,0736 0,0778 0,0769 82 Littorina saxatilis 0,0751 0,0733 0,0739 0,0854 0,0759 0,0740 0,0722 0,0741 0,0740 0,0783 0,0762 83 Littorina littorea 0,0777 0,0748 0,0794 0,0863 0,0791 0,0772 0,0743 0,0761 0,0772 0,0820 0,0805 84 Melaraphe neritoides 0,0804 0,0788 0,0816 0,0892 0,0807 0,0794 0,0778 0,0771 0,0793 0,0813 0,0821 85 Crepidula adunca 0,0774 0,0757 0,0797 0,0872 0,0811 0,0780 0,0746 0,0757 0,0780 0,0799 0,0819 86 Fasciolaria lignaria 0,0786 0,0786 0,0798 0,0884 0,0811 0,0781 0,0764 0,0753 0,0781 0,0823 0,0814 87 Pisania striata 0,0794 0,0795 0,0818 0,0876 0,0825 0,0778 0,0796 0,0772 0,0778 0,0826 0,0811 88 Bursa rana 0,0776 0,0806 0,0800 0,0892 0,0796 0,0766 0,0772 0,0749 0,0765 0,0819 0,0787 89 Truncatella guerinii 0,0798 0,0833 0,0793 0,0903 0,0822 0,0781 0,0797 0,0764 0,0781 0,0835 0,0826 90 Viviparus georgianus 0,0844 0,0844 0,0867 0,0954 0,0878 0,0827 0,0844 0,0799 0,0827 0,0892 0,0883 91 Balcis eburnea 0,0811 0,0782 0,0834 0,0887 0,0824 0,0811 0,0795 0,0788 0,0811 0,0860 0,0856
45 46 47 48 49 50 51 52 53 54 55
1 Bathydoris clavigera 0,1347 0,1305 0,1308 0,1347 0,1303 0,1300 0,1300 0,1299 0,1605 0,1643 0,1769 2 Chromodoris krohni 0,1242 0,1270 0,1210 0,1234 0,1274 0,1251 0,1251 0,1248 0,1475 0,1613 0,1693 3 Dendrodoris fumata 0,1140 0,1166 0,1120 0,1157 0,1159 0,1138 0,1138 0,1134 0,1367 0,1530 0,1585 4 Hypselodoris villafranca 0,1263 0,1238 0,1220 0,1202 0,1241 0,1219 0,1219 0,1238 0,1448 0,1544 0,1648 5 Dendronotus dalli 0,4731 0,4828 0,4793 0,4729 0,4804 0,4767 0,4767 0,4798 0,5082 0,5210 0,4865 6 Cuthona nana 0,4439 0,4456 0,4522 0,4371 0,4460 0,4477 0,4462 0,4517 0,4735 0,5012 0,4675 7 Flabellina babai 0,4176 0,4293 0,4283 0,4211 0,4225 0,4313 0,4278 0,4298 0,4451 0,4809 0,4431 8 Pleurobranchus peroni 0,1074 0,1046 0,1065 0,1049 0,1037 0,1055 0,1051 0,1073 0,1284 0,1337 0,1395 9 Berthellina citrina 0,1000 0,1003 0,0991 0,1006 0,0977 0,1002 0,0999 0,0999 0,1245 0,1300 0,1353
10 Bathyberthella antarctica 0,1143 0,1089 0,1107 0,1091 0,1083 0,1084 0,1081 0,1103 0,1318 0,1350 0,1454 11 Euselenops luniceps 0,1037 0,1028 0,1046 0,1031 0,1001 0,1027 0,1024 0,1045 0,1285 0,1317 0,1396 12 Tompthompsonia antarctica 0,1035 0,1026 0,1036 0,1029 0,0988 0,1022 0,1019 0,1040 0,1259 0,1291 0,1378 13 Berthella stellata 0,3423 0,3380 0,3426 0,3421 0,3393 0,3473 0,3443 0,3449 0,3764 0,3917 0,3854 14 Berthella platei 0,1647 0,1640 0,1661 0,1622 0,1626 0,1677 0,1671 0,1671 0,1856 0,1981 0,1971 15 Pupa solidula 0,0525 0,0523 0,0515 0,0521 0,0478 0,0500 0,0498 0,0515 0,0745 0,0673 0,0860 16 Hydatina physis 0,0468 0,0436 0,0451 0,0435 0,0443 0,0429 0,0427 0,0443 0,0619 0,0680 0,0884 17 Bullina lineata 0,0445 0,0428 0,0428 0,0427 0,0422 0,0407 0,0406 0,0422 0,0638 0,0645 0,0856 18 Acteon sp. 0,0311 0,0296 0,0310 0,0310 0,0291 0,0270 0,0278 0,0277 0,0436 0,0450 0,0781 19 Haminoea hydatis 0,0325 0,0303 0,0337 0,0325 0,0333 0,0305 0,0297 0,0326 0,0492 0,0498 0,0782 20 Cylichna gelida 0,0229 0,0215 0,0240 0,0228 0,0236 0,0214 0,0215 0,0234 0,0389 0,0434 0,0704 21 Scaphander lignarius 0,0261 0,0241 0,0267 0,0241 0,0249 0,0229 0,0229 0,0235 0,0382 0,0420 0,0716 22 cf. Retusa sp. 0,0318 0,0296 0,0330 0,0318 0,0326 0,0289 0,0290 0,0311 0,0466 0,0506 0,0824 23 Bulla cf. striata 0,0256 0,0242 0,0255 0,0249 0,0249 0,0242 0,0243 0,0248 0,0419 0,0458 0,0750 24 Smaragdinella sp. 0,0290 0,0275 0,0275 0,0276 0,0271 0,0249 0,0249 0,0269 0,0428 0,0481 0,0719 25 Phanerophthalmus smaragdinus 0,0296 0,0282 0,0289 0,0283 0,0270 0,0270 0,0263 0,0277 0,0461 0,0514 0,0755 26 Bullacta exarata 0,0215 0,0215 0,0227 0,0221 0,0229 0,0235 0,0222 0,0228 0,0390 0,0434 0,0682 27 Atys semistriatus 0,0276 0,0261 0,0274 0,0262 0,0270 0,0248 0,0249 0,0269 0,0413 0,0487 0,0735 28 Odontoglaja guamensis 0,0303 0,0288 0,0301 0,0302 0,0270 0,0262 0,0263 0,0269 0,0450 0,0467 0,0734 29 Navanax inermis 0,0294 0,0280 0,0293 0,0293 0,0261 0,0254 0,0255 0,0260 0,0440 0,0456 0,0758 30 Chelidonura inornata 0,0274 0,0294 0,0301 0,0315 0,0262 0,0268 0,0268 0,0274 0,0439 0,0460 0,0749 31 Philinopsis pilsbryi 0,0274 0,0273 0,0280 0,0295 0,0255 0,0261 0,0261 0,0253 0,0418 0,0459 0,0740 32 Philinoglossa praelongata 0,0371 0,0348 0,0348 0,0341 0,0330 0,0328 0,0329 0,0342 0,0483 0,0519 0,0773 33 Sagaminopteron psychedelicum 0,0317 0,0310 0,0315 0,0340 0,0318 0,0303 0,0304 0,0324 0,0464 0,0546 0,0776 34 Toledonia globosa 0,0221 0,0182 0,0207 0,0189 0,0196 0,0183 0,0177 0,0190 0,0383 0,0411 0,0717 35 Diaphana cf. inflata 0,0304 0,0276 0,0302 0,0283 0,0283 0,0277 0,0270 0,0283 0,0452 0,0482 0,0768 36 Newnesia antarctica 0,0234 0,0208 0,0207 0,0208 0,0209 0,0196 0,0190 0,0189 0,0383 0,0441 0,0722 37 Colpodaspis thomsoni 0,0394 0,0393 0,0369 0,0392 0,0389 0,0396 0,0388 0,0395 0,0543 0,0567 0,0847 38 Runcina adriatica 0,0205 0,0232 0,0239 0,0239 0,0233 0,0261 0,0253 0,0253 0,0441 0,0490 0,0727 39 Unela glandulifera 0,0174 0,0174 0,0155 0,0181 0,0169 0,0155 0,0156 0,0155 0,0362 0,0440 0,0636 40 Pontohedyle milatchevitchi 0,0240 0,0246 0,0220 0,0247 0,0215 0,0228 0,0221 0,0228 0,0349 0,0387 0,0621 41 Hedylopsis spiculifera 0,0156 0,0143 0,0137 0,0150 0,0138 0,0125 0,0131 0,0137 0,0321 0,0419 0,0612 42 Parahedyle cryptophthalma 0,0174 0,0174 0,0155 0,0181 0,0169 0,0155 0,0156 0,0155 0,0362 0,0440 0,0636 43 Umbraculum mediterraneum 0,0223 0,0231 0,0191 0,0238 0,0218 0,0211 0,0218 0,0210 0,0438 0,0545 0,0766 44 Tylodina perversa 0,0223 0,0203 0,0210 0,0210 0,0190 0,0183 0,0190 0,0196 0,0409 0,0486 0,0721 45 Akera bullata 0,0076 0,0064 0,0088 0,0070 0,0082 0,0076 0,0082 0,0284 0,0359 0,0596 46 Aplysia californica 0,0074 0,0058 0,0029 0,0058 0,0058 0,0052 0,0070 0,0265 0,0322 0,0599 47 Dolabella auricularia 0,0062 0,0057 0,0052 0,0052 0,0052 0,0046 0,0052 0,0279 0,0336 0,0582 48 Siphonota geographica 0,0085 0,0028 0,0051 0,0064 0,0070 0,0070 0,0082 0,0244 0,0320 0,0564 49 Bursatella leachii 0,0068 0,0057 0,0051 0,0063 0,0029 0,0029 0,0040 0,0258 0,0294 0,0555 50 Petalifera petalifera 0,0080 0,0057 0,0051 0,0068 0,0028 0,0011 0,0023 0,0237 0,0294 0,0560 51 Dolabrifera brazieri 0,0074 0,0051 0,0045 0,0068 0,0028 0,0011 0,0023 0,0244 0,0287 0,0551 52 Stylocheilus longicauda 0,0080 0,0068 0,0051 0,0079 0,0040 0,0023 0,0023 0,0237 0,0308 0,0569 53 Clio pyramidata 0,0256 0,0239 0,0250 0,0222 0,0234 0,0216 0,0222 0,0216 0,0331 0,0590 54 Diacria quadridentata 0,0312 0,0283 0,0295 0,0284 0,0261 0,0261 0,0255 0,0272 0,0289 0,0633 55 Creseis acicula 0,0484 0,0484 0,0473 0,0462 0,0451 0,0456 0,0450 0,0462 0,0480 0,0501 56 Spongiobranchaea australis 0,0164 0,0153 0,0142 0,0159 0,0148 0,0125 0,0125 0,0136 0,0307 0,0335 0,0513
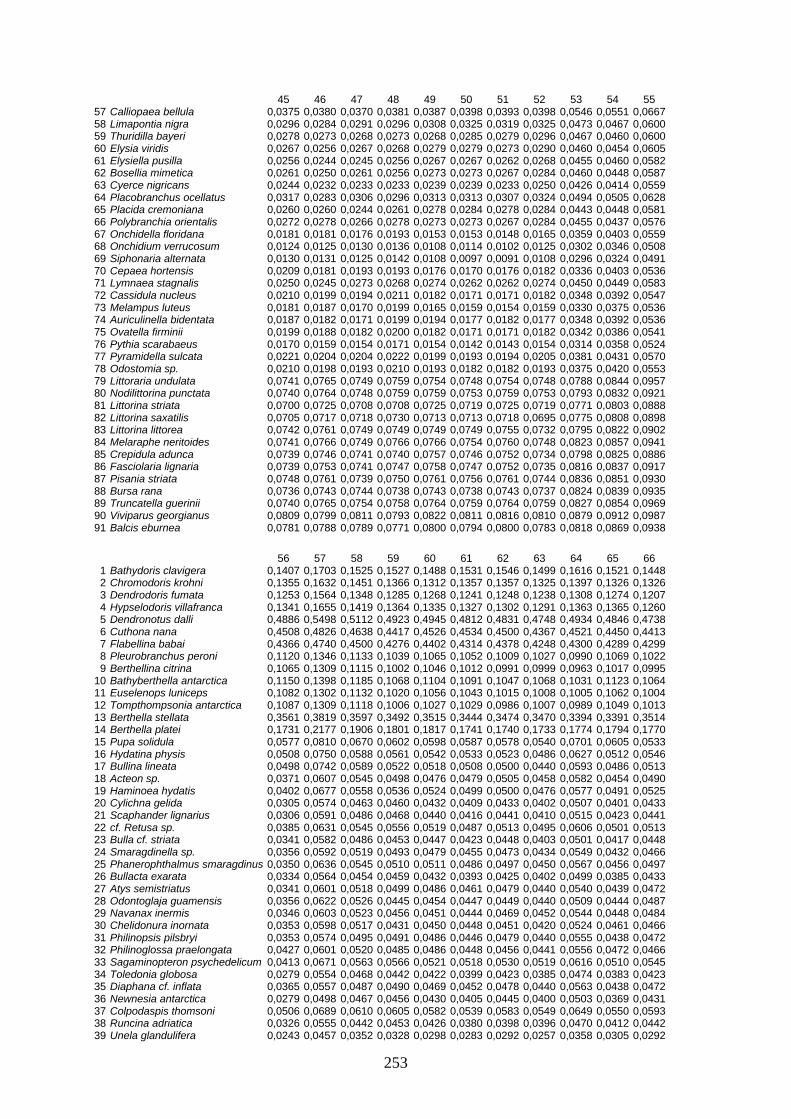
253
45 46 47 48 49 50 51 52 53 54 55 57 Calliopaea bellula 0,0375 0,0380 0,0370 0,0381 0,0387 0,0398 0,0393 0,0398 0,0546 0,0551 0,0667 58 Limapontia nigra 0,0296 0,0284 0,0291 0,0296 0,0308 0,0325 0,0319 0,0325 0,0473 0,0467 0,0600 59 Thuridilla bayeri 0,0278 0,0273 0,0268 0,0273 0,0268 0,0285 0,0279 0,0296 0,0467 0,0460 0,0600 60 Elysia viridis 0,0267 0,0256 0,0267 0,0268 0,0279 0,0279 0,0273 0,0290 0,0460 0,0454 0,0605 61 Elysiella pusilla 0,0256 0,0244 0,0245 0,0256 0,0267 0,0267 0,0262 0,0268 0,0455 0,0460 0,0582 62 Bosellia mimetica 0,0261 0,0250 0,0261 0,0256 0,0273 0,0273 0,0267 0,0284 0,0460 0,0448 0,0587 63 Cyerce nigricans 0,0244 0,0232 0,0233 0,0233 0,0239 0,0239 0,0233 0,0250 0,0426 0,0414 0,0559 64 Placobranchus ocellatus 0,0317 0,0283 0,0306 0,0296 0,0313 0,0313 0,0307 0,0324 0,0494 0,0505 0,0628 65 Placida cremoniana 0,0260 0,0260 0,0244 0,0261 0,0278 0,0284 0,0278 0,0284 0,0443 0,0448 0,0581 66 Polybranchia orientalis 0,0272 0,0278 0,0266 0,0278 0,0273 0,0273 0,0267 0,0284 0,0455 0,0437 0,0576 67 Onchidella floridana 0,0181 0,0181 0,0176 0,0193 0,0153 0,0153 0,0148 0,0165 0,0359 0,0403 0,0559 68 Onchidium verrucosum 0,0124 0,0125 0,0130 0,0136 0,0108 0,0114 0,0102 0,0125 0,0302 0,0346 0,0508 69 Siphonaria alternata 0,0130 0,0131 0,0125 0,0142 0,0108 0,0097 0,0091 0,0108 0,0296 0,0324 0,0491 70 Cepaea hortensis 0,0209 0,0181 0,0193 0,0193 0,0176 0,0170 0,0176 0,0182 0,0336 0,0403 0,0536 71 Lymnaea stagnalis 0,0250 0,0245 0,0273 0,0268 0,0274 0,0262 0,0262 0,0274 0,0450 0,0449 0,0583 72 Cassidula nucleus 0,0210 0,0199 0,0194 0,0211 0,0182 0,0171 0,0171 0,0182 0,0348 0,0392 0,0547 73 Melampus luteus 0,0181 0,0187 0,0170 0,0199 0,0165 0,0159 0,0154 0,0159 0,0330 0,0375 0,0536 74 Auriculinella bidentata 0,0187 0,0182 0,0171 0,0199 0,0194 0,0177 0,0182 0,0177 0,0348 0,0392 0,0536 75 Ovatella firminii 0,0199 0,0188 0,0182 0,0200 0,0182 0,0171 0,0171 0,0182 0,0342 0,0386 0,0541 76 Pythia scarabaeus 0,0170 0,0159 0,0154 0,0171 0,0154 0,0142 0,0143 0,0154 0,0314 0,0358 0,0524 77 Pyramidella sulcata 0,0221 0,0204 0,0204 0,0222 0,0199 0,0193 0,0194 0,0205 0,0381 0,0431 0,0570 78 Odostomia sp. 0,0210 0,0198 0,0193 0,0210 0,0193 0,0182 0,0182 0,0193 0,0375 0,0420 0,0553 79 Littoraria undulata 0,0741 0,0765 0,0749 0,0759 0,0754 0,0748 0,0754 0,0748 0,0788 0,0844 0,0957 80 Nodilittorina punctata 0,0740 0,0764 0,0748 0,0759 0,0759 0,0753 0,0759 0,0753 0,0793 0,0832 0,0921 81 Littorina striata 0,0700 0,0725 0,0708 0,0708 0,0725 0,0719 0,0725 0,0719 0,0771 0,0803 0,0888 82 Littorina saxatilis 0,0705 0,0717 0,0718 0,0730 0,0713 0,0713 0,0718 0,0695 0,0775 0,0808 0,0898 83 Littorina littorea 0,0742 0,0761 0,0749 0,0749 0,0749 0,0749 0,0755 0,0732 0,0795 0,0822 0,0902 84 Melaraphe neritoides 0,0741 0,0766 0,0749 0,0766 0,0766 0,0754 0,0760 0,0748 0,0823 0,0857 0,0941 85 Crepidula adunca 0,0739 0,0746 0,0741 0,0740 0,0757 0,0746 0,0752 0,0734 0,0798 0,0825 0,0886 86 Fasciolaria lignaria 0,0739 0,0753 0,0741 0,0747 0,0758 0,0747 0,0752 0,0735 0,0816 0,0837 0,0917 87 Pisania striata 0,0748 0,0761 0,0739 0,0750 0,0761 0,0756 0,0761 0,0744 0,0836 0,0851 0,0930 88 Bursa rana 0,0736 0,0743 0,0744 0,0738 0,0743 0,0738 0,0743 0,0737 0,0824 0,0839 0,0935 89 Truncatella guerinii 0,0740 0,0765 0,0754 0,0758 0,0764 0,0759 0,0764 0,0759 0,0827 0,0854 0,0969 90 Viviparus georgianus 0,0809 0,0799 0,0811 0,0793 0,0822 0,0811 0,0816 0,0810 0,0879 0,0912 0,0987 91 Balcis eburnea 0,0781 0,0788 0,0789 0,0771 0,0800 0,0794 0,0800 0,0783 0,0818 0,0869 0,0938
56 57 58 59 60 61 62 63 64 65 66
1 Bathydoris clavigera 0,1407 0,1703 0,1525 0,1527 0,1488 0,1531 0,1546 0,1499 0,1616 0,1521 0,1448 2 Chromodoris krohni 0,1355 0,1632 0,1451 0,1366 0,1312 0,1357 0,1357 0,1325 0,1397 0,1326 0,1326 3 Dendrodoris fumata 0,1253 0,1564 0,1348 0,1285 0,1268 0,1241 0,1248 0,1238 0,1308 0,1274 0,1207 4 Hypselodoris villafranca 0,1341 0,1655 0,1419 0,1364 0,1335 0,1327 0,1302 0,1291 0,1363 0,1365 0,1260 5 Dendronotus dalli 0,4886 0,5498 0,5112 0,4923 0,4945 0,4812 0,4831 0,4748 0,4934 0,4846 0,4738 6 Cuthona nana 0,4508 0,4826 0,4638 0,4417 0,4526 0,4534 0,4500 0,4367 0,4521 0,4450 0,4413 7 Flabellina babai 0,4366 0,4740 0,4500 0,4276 0,4402 0,4314 0,4378 0,4248 0,4300 0,4289 0,4299 8 Pleurobranchus peroni 0,1120 0,1346 0,1133 0,1039 0,1065 0,1052 0,1009 0,1027 0,0990 0,1069 0,1022 9 Berthellina citrina 0,1065 0,1309 0,1115 0,1002 0,1046 0,1012 0,0991 0,0999 0,0963 0,1017 0,0995
10 Bathyberthella antarctica 0,1150 0,1398 0,1185 0,1068 0,1104 0,1091 0,1047 0,1068 0,1031 0,1123 0,1064 11 Euselenops luniceps 0,1082 0,1302 0,1132 0,1020 0,1056 0,1043 0,1015 0,1008 0,1005 0,1062 0,1004 12 Tompthompsonia antarctica 0,1087 0,1309 0,1118 0,1006 0,1027 0,1029 0,0986 0,1007 0,0989 0,1049 0,1013 13 Berthella stellata 0,3561 0,3819 0,3597 0,3492 0,3515 0,3444 0,3474 0,3470 0,3394 0,3391 0,3514 14 Berthella platei 0,1731 0,2177 0,1906 0,1801 0,1817 0,1741 0,1740 0,1733 0,1774 0,1794 0,1770 15 Pupa solidula 0,0577 0,0810 0,0670 0,0602 0,0598 0,0587 0,0578 0,0540 0,0701 0,0605 0,0533 16 Hydatina physis 0,0508 0,0750 0,0588 0,0561 0,0542 0,0533 0,0523 0,0486 0,0627 0,0512 0,0546 17 Bullina lineata 0,0498 0,0742 0,0589 0,0522 0,0518 0,0508 0,0500 0,0440 0,0593 0,0486 0,0513 18 Acteon sp. 0,0371 0,0607 0,0545 0,0498 0,0476 0,0479 0,0505 0,0458 0,0582 0,0454 0,0490 19 Haminoea hydatis 0,0402 0,0677 0,0558 0,0536 0,0524 0,0499 0,0500 0,0476 0,0577 0,0491 0,0525 20 Cylichna gelida 0,0305 0,0574 0,0463 0,0460 0,0432 0,0409 0,0433 0,0402 0,0507 0,0401 0,0433 21 Scaphander lignarius 0,0306 0,0591 0,0486 0,0468 0,0440 0,0416 0,0441 0,0410 0,0515 0,0423 0,0441 22 cf. Retusa sp. 0,0385 0,0631 0,0545 0,0556 0,0519 0,0487 0,0513 0,0495 0,0606 0,0501 0,0513 23 Bulla cf. striata 0,0341 0,0582 0,0486 0,0453 0,0447 0,0423 0,0448 0,0403 0,0501 0,0417 0,0448 24 Smaragdinella sp. 0,0356 0,0592 0,0519 0,0493 0,0479 0,0455 0,0473 0,0434 0,0549 0,0432 0,0466 25 Phanerophthalmus smaragdinus 0,0350 0,0636 0,0545 0,0510 0,0511 0,0486 0,0497 0,0450 0,0567 0,0456 0,0497 26 Bullacta exarata 0,0334 0,0564 0,0454 0,0459 0,0432 0,0393 0,0425 0,0402 0,0499 0,0385 0,0433 27 Atys semistriatus 0,0341 0,0601 0,0518 0,0499 0,0486 0,0461 0,0479 0,0440 0,0540 0,0439 0,0472 28 Odontoglaja guamensis 0,0356 0,0622 0,0526 0,0445 0,0454 0,0447 0,0449 0,0440 0,0509 0,0444 0,0487 29 Navanax inermis 0,0346 0,0603 0,0523 0,0456 0,0451 0,0444 0,0469 0,0452 0,0544 0,0448 0,0484 30 Chelidonura inornata 0,0353 0,0598 0,0517 0,0431 0,0450 0,0448 0,0451 0,0420 0,0524 0,0461 0,0466 31 Philinopsis pilsbryi 0,0353 0,0574 0,0495 0,0491 0,0486 0,0446 0,0479 0,0440 0,0555 0,0438 0,0472 32 Philinoglossa praelongata 0,0427 0,0601 0,0520 0,0485 0,0486 0,0448 0,0456 0,0441 0,0556 0,0472 0,0466 33 Sagaminopteron psychedelicum 0,0413 0,0671 0,0563 0,0566 0,0521 0,0518 0,0530 0,0519 0,0616 0,0510 0,0545 34 Toledonia globosa 0,0279 0,0554 0,0468 0,0442 0,0422 0,0399 0,0423 0,0385 0,0474 0,0383 0,0423 35 Diaphana cf. inflata 0,0365 0,0557 0,0487 0,0490 0,0469 0,0452 0,0478 0,0440 0,0563 0,0438 0,0472 36 Newnesia antarctica 0,0279 0,0498 0,0467 0,0456 0,0430 0,0405 0,0445 0,0400 0,0503 0,0369 0,0431 37 Colpodaspis thomsoni 0,0506 0,0689 0,0610 0,0605 0,0582 0,0539 0,0583 0,0549 0,0649 0,0550 0,0593 38 Runcina adriatica 0,0326 0,0555 0,0442 0,0453 0,0426 0,0380 0,0398 0,0396 0,0470 0,0412 0,0442 39 Unela glandulifera 0,0243 0,0457 0,0352 0,0328 0,0298 0,0283 0,0292 0,0257 0,0358 0,0305 0,0292
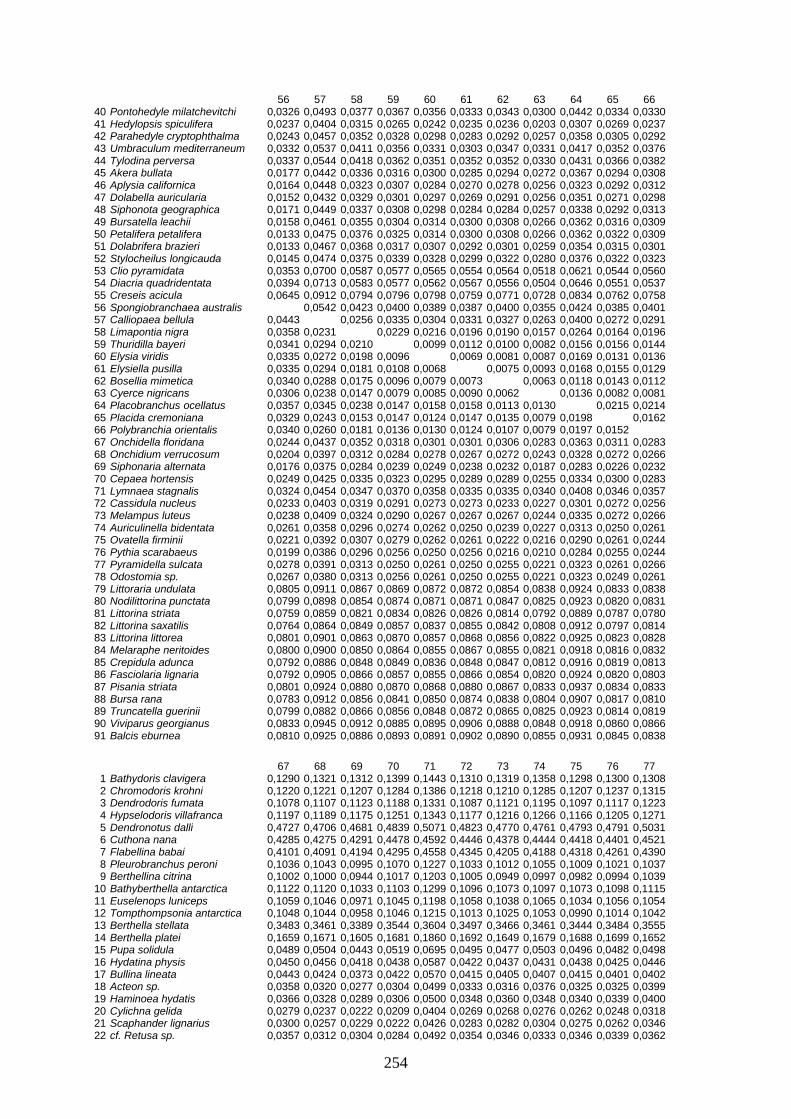
254
56 57 58 59 60 61 62 63 64 65 66 40 Pontohedyle milatchevitchi 0,0326 0,0493 0,0377 0,0367 0,0356 0,0333 0,0343 0,0300 0,0442 0,0334 0,0330 41 Hedylopsis spiculifera 0,0237 0,0404 0,0315 0,0265 0,0242 0,0235 0,0236 0,0203 0,0307 0,0269 0,0237 42 Parahedyle cryptophthalma 0,0243 0,0457 0,0352 0,0328 0,0298 0,0283 0,0292 0,0257 0,0358 0,0305 0,0292 43 Umbraculum mediterraneum 0,0332 0,0537 0,0411 0,0356 0,0331 0,0303 0,0347 0,0331 0,0417 0,0352 0,0376 44 Tylodina perversa 0,0337 0,0544 0,0418 0,0362 0,0351 0,0352 0,0352 0,0330 0,0431 0,0366 0,0382 45 Akera bullata 0,0177 0,0442 0,0336 0,0316 0,0300 0,0285 0,0294 0,0272 0,0367 0,0294 0,0308 46 Aplysia californica 0,0164 0,0448 0,0323 0,0307 0,0284 0,0270 0,0278 0,0256 0,0323 0,0292 0,0312 47 Dolabella auricularia 0,0152 0,0432 0,0329 0,0301 0,0297 0,0269 0,0291 0,0256 0,0351 0,0271 0,0298 48 Siphonota geographica 0,0171 0,0449 0,0337 0,0308 0,0298 0,0284 0,0284 0,0257 0,0338 0,0292 0,0313 49 Bursatella leachii 0,0158 0,0461 0,0355 0,0304 0,0314 0,0300 0,0308 0,0266 0,0362 0,0316 0,0309 50 Petalifera petalifera 0,0133 0,0475 0,0376 0,0325 0,0314 0,0300 0,0308 0,0266 0,0362 0,0322 0,0309 51 Dolabrifera brazieri 0,0133 0,0467 0,0368 0,0317 0,0307 0,0292 0,0301 0,0259 0,0354 0,0315 0,0301 52 Stylocheilus longicauda 0,0145 0,0474 0,0375 0,0339 0,0328 0,0299 0,0322 0,0280 0,0376 0,0322 0,0323 53 Clio pyramidata 0,0353 0,0700 0,0587 0,0577 0,0565 0,0554 0,0564 0,0518 0,0621 0,0544 0,0560 54 Diacria quadridentata 0,0394 0,0713 0,0583 0,0577 0,0562 0,0567 0,0556 0,0504 0,0646 0,0551 0,0537 55 Creseis acicula 0,0645 0,0912 0,0794 0,0796 0,0798 0,0759 0,0771 0,0728 0,0834 0,0762 0,0758 56 Spongiobranchaea australis 0,0542 0,0423 0,0400 0,0389 0,0387 0,0400 0,0355 0,0424 0,0385 0,0401 57 Calliopaea bellula 0,0443 0,0256 0,0335 0,0304 0,0331 0,0327 0,0263 0,0400 0,0272 0,0291 58 Limapontia nigra 0,0358 0,0231 0,0229 0,0216 0,0196 0,0190 0,0157 0,0264 0,0164 0,0196 59 Thuridilla bayeri 0,0341 0,0294 0,0210 0,0099 0,0112 0,0100 0,0082 0,0156 0,0156 0,0144 60 Elysia viridis 0,0335 0,0272 0,0198 0,0096 0,0069 0,0081 0,0087 0,0169 0,0131 0,0136 61 Elysiella pusilla 0,0335 0,0294 0,0181 0,0108 0,0068 0,0075 0,0093 0,0168 0,0155 0,0129 62 Bosellia mimetica 0,0340 0,0288 0,0175 0,0096 0,0079 0,0073 0,0063 0,0118 0,0143 0,0112 63 Cyerce nigricans 0,0306 0,0238 0,0147 0,0079 0,0085 0,0090 0,0062 0,0136 0,0082 0,0081 64 Placobranchus ocellatus 0,0357 0,0345 0,0238 0,0147 0,0158 0,0158 0,0113 0,0130 0,0215 0,0214 65 Placida cremoniana 0,0329 0,0243 0,0153 0,0147 0,0124 0,0147 0,0135 0,0079 0,0198 0,0162 66 Polybranchia orientalis 0,0340 0,0260 0,0181 0,0136 0,0130 0,0124 0,0107 0,0079 0,0197 0,0152 67 Onchidella floridana 0,0244 0,0437 0,0352 0,0318 0,0301 0,0301 0,0306 0,0283 0,0363 0,0311 0,0283 68 Onchidium verrucosum 0,0204 0,0397 0,0312 0,0284 0,0278 0,0267 0,0272 0,0243 0,0328 0,0272 0,0266 69 Siphonaria alternata 0,0176 0,0375 0,0284 0,0239 0,0249 0,0238 0,0232 0,0187 0,0283 0,0226 0,0232 70 Cepaea hortensis 0,0249 0,0425 0,0335 0,0323 0,0295 0,0289 0,0289 0,0255 0,0334 0,0300 0,0283 71 Lymnaea stagnalis 0,0324 0,0454 0,0347 0,0370 0,0358 0,0335 0,0335 0,0340 0,0408 0,0346 0,0357 72 Cassidula nucleus 0,0233 0,0403 0,0319 0,0291 0,0273 0,0273 0,0233 0,0227 0,0301 0,0272 0,0256 73 Melampus luteus 0,0238 0,0409 0,0324 0,0290 0,0267 0,0267 0,0267 0,0244 0,0335 0,0272 0,0266 74 Auriculinella bidentata 0,0261 0,0358 0,0296 0,0274 0,0262 0,0250 0,0239 0,0227 0,0313 0,0250 0,0261 75 Ovatella firminii 0,0221 0,0392 0,0307 0,0279 0,0262 0,0261 0,0222 0,0216 0,0290 0,0261 0,0244 76 Pythia scarabaeus 0,0199 0,0386 0,0296 0,0256 0,0250 0,0256 0,0216 0,0210 0,0284 0,0255 0,0244 77 Pyramidella sulcata 0,0278 0,0391 0,0313 0,0250 0,0261 0,0250 0,0255 0,0221 0,0323 0,0261 0,0266 78 Odostomia sp. 0,0267 0,0380 0,0313 0,0256 0,0261 0,0250 0,0255 0,0221 0,0323 0,0249 0,0261 79 Littoraria undulata 0,0805 0,0911 0,0867 0,0869 0,0872 0,0872 0,0854 0,0838 0,0924 0,0833 0,0838 80 Nodilittorina punctata 0,0799 0,0898 0,0854 0,0874 0,0871 0,0871 0,0847 0,0825 0,0923 0,0820 0,0831 81 Littorina striata 0,0759 0,0859 0,0821 0,0834 0,0826 0,0826 0,0814 0,0792 0,0889 0,0787 0,0780 82 Littorina saxatilis 0,0764 0,0864 0,0849 0,0857 0,0837 0,0855 0,0842 0,0808 0,0912 0,0797 0,0814 83 Littorina littorea 0,0801 0,0901 0,0863 0,0870 0,0857 0,0868 0,0856 0,0822 0,0925 0,0823 0,0828 84 Melaraphe neritoides 0,0800 0,0900 0,0850 0,0864 0,0855 0,0867 0,0855 0,0821 0,0918 0,0816 0,0832 85 Crepidula adunca 0,0792 0,0886 0,0848 0,0849 0,0836 0,0848 0,0847 0,0812 0,0916 0,0819 0,0813 86 Fasciolaria lignaria 0,0792 0,0905 0,0866 0,0857 0,0855 0,0866 0,0854 0,0820 0,0924 0,0820 0,0803 87 Pisania striata 0,0801 0,0924 0,0880 0,0870 0,0868 0,0880 0,0867 0,0833 0,0937 0,0834 0,0833 88 Bursa rana 0,0783 0,0912 0,0856 0,0841 0,0850 0,0874 0,0838 0,0804 0,0907 0,0817 0,0810 89 Truncatella guerinii 0,0799 0,0882 0,0866 0,0856 0,0848 0,0872 0,0865 0,0825 0,0923 0,0814 0,0819 90 Viviparus georgianus 0,0833 0,0945 0,0912 0,0885 0,0895 0,0906 0,0888 0,0848 0,0918 0,0860 0,0866 91 Balcis eburnea 0,0810 0,0925 0,0886 0,0893 0,0891 0,0902 0,0890 0,0855 0,0931 0,0845 0,0838
67 68 69 70 71 72 73 74 75 76 77
1 Bathydoris clavigera 0,1290 0,1321 0,1312 0,1399 0,1443 0,1310 0,1319 0,1358 0,1298 0,1300 0,1308 2 Chromodoris krohni 0,1220 0,1221 0,1207 0,1284 0,1386 0,1218 0,1210 0,1285 0,1207 0,1237 0,1315 3 Dendrodoris fumata 0,1078 0,1107 0,1123 0,1188 0,1331 0,1087 0,1121 0,1195 0,1097 0,1117 0,1223 4 Hypselodoris villafranca 0,1197 0,1189 0,1175 0,1251 0,1343 0,1177 0,1216 0,1266 0,1166 0,1205 0,1271 5 Dendronotus dalli 0,4727 0,4706 0,4681 0,4839 0,5071 0,4823 0,4770 0,4761 0,4793 0,4791 0,5031 6 Cuthona nana 0,4285 0,4275 0,4291 0,4478 0,4592 0,4446 0,4378 0,4444 0,4418 0,4401 0,4521 7 Flabellina babai 0,4101 0,4091 0,4194 0,4295 0,4558 0,4345 0,4205 0,4188 0,4318 0,4261 0,4390 8 Pleurobranchus peroni 0,1036 0,1043 0,0995 0,1070 0,1227 0,1033 0,1012 0,1055 0,1009 0,1021 0,1037 9 Berthellina citrina 0,1002 0,1000 0,0944 0,1017 0,1203 0,1005 0,0949 0,0997 0,0982 0,0994 0,1039
10 Bathyberthella antarctica 0,1122 0,1120 0,1033 0,1103 0,1299 0,1096 0,1073 0,1097 0,1073 0,1098 0,1115 11 Euselenops luniceps 0,1059 0,1046 0,0971 0,1045 0,1198 0,1058 0,1038 0,1065 0,1034 0,1056 0,1054 12 Tompthompsonia antarctica 0,1048 0,1044 0,0958 0,1046 0,1215 0,1013 0,1025 0,1053 0,0990 0,1014 0,1042 13 Berthella stellata 0,3483 0,3461 0,3389 0,3544 0,3604 0,3497 0,3466 0,3461 0,3444 0,3484 0,3555 14 Berthella platei 0,1659 0,1671 0,1605 0,1681 0,1860 0,1692 0,1649 0,1679 0,1688 0,1699 0,1652 15 Pupa solidula 0,0489 0,0504 0,0443 0,0519 0,0695 0,0495 0,0477 0,0503 0,0496 0,0482 0,0498 16 Hydatina physis 0,0450 0,0456 0,0418 0,0438 0,0587 0,0422 0,0437 0,0431 0,0438 0,0425 0,0446 17 Bullina lineata 0,0443 0,0424 0,0373 0,0422 0,0570 0,0415 0,0405 0,0407 0,0415 0,0401 0,0402 18 Acteon sp. 0,0358 0,0320 0,0277 0,0304 0,0499 0,0333 0,0316 0,0376 0,0325 0,0325 0,0399 19 Haminoea hydatis 0,0366 0,0328 0,0289 0,0306 0,0500 0,0348 0,0360 0,0348 0,0340 0,0339 0,0400 20 Cylichna gelida 0,0279 0,0237 0,0222 0,0209 0,0404 0,0269 0,0268 0,0276 0,0262 0,0248 0,0318 21 Scaphander lignarius 0,0300 0,0257 0,0229 0,0222 0,0426 0,0283 0,0282 0,0304 0,0275 0,0262 0,0346 22 cf. Retusa sp. 0,0357 0,0312 0,0304 0,0284 0,0492 0,0354 0,0346 0,0333 0,0346 0,0339 0,0362

255
67 68 69 70 71 72 73 74 75 76 77 23 Bulla cf. striata 0,0292 0,0249 0,0229 0,0248 0,0426 0,0290 0,0263 0,0263 0,0283 0,0269 0,0284 24 Smaragdinella sp. 0,0322 0,0278 0,0249 0,0256 0,0466 0,0305 0,0296 0,0299 0,0297 0,0283 0,0348 25 Phanerophthalmus smaragdinus 0,0315 0,0291 0,0243 0,0257 0,0491 0,0306 0,0303 0,0320 0,0298 0,0291 0,0357 26 Bullacta exarata 0,0299 0,0237 0,0228 0,0229 0,0421 0,0290 0,0260 0,0270 0,0283 0,0269 0,0333 27 Atys semistriatus 0,0314 0,0271 0,0255 0,0236 0,0458 0,0298 0,0303 0,0305 0,0290 0,0276 0,0355 28 Odontoglaja guamensis 0,0336 0,0298 0,0256 0,0277 0,0473 0,0276 0,0302 0,0326 0,0268 0,0267 0,0355 29 Navanax inermis 0,0320 0,0296 0,0254 0,0287 0,0470 0,0303 0,0280 0,0344 0,0295 0,0294 0,0366 30 Chelidonura inornata 0,0341 0,0297 0,0242 0,0288 0,0494 0,0310 0,0282 0,0345 0,0302 0,0288 0,0333 31 Philinopsis pilsbryi 0,0312 0,0296 0,0254 0,0261 0,0470 0,0302 0,0267 0,0323 0,0295 0,0281 0,0359 32 Philinoglossa praelongata 0,0389 0,0329 0,0307 0,0306 0,0475 0,0322 0,0334 0,0327 0,0314 0,0306 0,0356 33 Sagaminopteron psychedelicum 0,0327 0,0310 0,0304 0,0326 0,0496 0,0363 0,0325 0,0326 0,0354 0,0333 0,0405 34 Toledonia globosa 0,0271 0,0209 0,0182 0,0228 0,0381 0,0269 0,0262 0,0250 0,0261 0,0248 0,0298 35 Diaphana cf. inflata 0,0301 0,0278 0,0255 0,0243 0,0468 0,0325 0,0297 0,0319 0,0317 0,0304 0,0339 36 Newnesia antarctica 0,0272 0,0222 0,0201 0,0228 0,0410 0,0269 0,0261 0,0236 0,0262 0,0248 0,0318 37 Colpodaspis thomsoni 0,0451 0,0410 0,0390 0,0420 0,0580 0,0482 0,0425 0,0403 0,0472 0,0457 0,0456 38 Runcina adriatica 0,0287 0,0245 0,0258 0,0310 0,0406 0,0310 0,0323 0,0287 0,0317 0,0303 0,0359 39 Unela glandulifera 0,0182 0,0163 0,0138 0,0187 0,0329 0,0119 0,0150 0,0168 0,0106 0,0100 0,0194 40 Pontohedyle milatchevitchi 0,0268 0,0221 0,0201 0,0269 0,0413 0,0209 0,0222 0,0248 0,0209 0,0228 0,0305 41 Hedylopsis spiculifera 0,0182 0,0137 0,0100 0,0162 0,0295 0,0132 0,0138 0,0142 0,0119 0,0112 0,0156 42 Parahedyle cryptophthalma 0,0182 0,0162 0,0138 0,0187 0,0329 0,0119 0,0150 0,0168 0,0106 0,0100 0,0194 43 Umbraculum mediterraneum 0,0252 0,0245 0,0253 0,0272 0,0387 0,0244 0,0231 0,0217 0,0243 0,0210 0,0224 44 Tylodina perversa 0,0265 0,0265 0,0245 0,0271 0,0365 0,0258 0,0258 0,0230 0,0257 0,0237 0,0224 45 Akera bullata 0,0195 0,0131 0,0137 0,0227 0,0282 0,0228 0,0195 0,0202 0,0215 0,0182 0,0243 46 Aplysia californica 0,0195 0,0132 0,0137 0,0195 0,0276 0,0215 0,0201 0,0195 0,0201 0,0169 0,0223 47 Dolabella auricularia 0,0189 0,0138 0,0131 0,0207 0,0310 0,0209 0,0182 0,0183 0,0195 0,0163 0,0224 48 Siphonota geographica 0,0210 0,0145 0,0150 0,0209 0,0305 0,0229 0,0215 0,0216 0,0215 0,0183 0,0244 49 Bursatella leachii 0,0163 0,0113 0,0113 0,0188 0,0312 0,0196 0,0176 0,0210 0,0196 0,0164 0,0217 50 Petalifera petalifera 0,0164 0,0119 0,0100 0,0181 0,0296 0,0182 0,0170 0,0189 0,0182 0,0151 0,0210 51 Dolabrifera brazieri 0,0157 0,0107 0,0094 0,0188 0,0297 0,0183 0,0163 0,0196 0,0183 0,0151 0,0211 52 Stylocheilus longicauda 0,0177 0,0132 0,0113 0,0194 0,0310 0,0195 0,0169 0,0189 0,0195 0,0163 0,0223 53 Clio pyramidata 0,0418 0,0344 0,0336 0,0387 0,0557 0,0403 0,0381 0,0402 0,0395 0,0359 0,0448 54 Diacria quadridentata 0,0485 0,0406 0,0374 0,0480 0,0557 0,0468 0,0443 0,0468 0,0459 0,0422 0,0526 55 Creseis acicula 0,0719 0,0638 0,0610 0,0683 0,0776 0,0702 0,0684 0,0680 0,0691 0,0666 0,0739 56 Spongiobranchaea australis 0,0274 0,0224 0,0191 0,0278 0,0379 0,0258 0,0265 0,0291 0,0243 0,0217 0,0315 57 Calliopaea bellula 0,0533 0,0477 0,0445 0,0517 0,0566 0,0487 0,0494 0,0421 0,0470 0,0462 0,0469 58 Limapontia nigra 0,0414 0,0362 0,0324 0,0392 0,0411 0,0369 0,0378 0,0337 0,0354 0,0339 0,0362 59 Thuridilla bayeri 0,0367 0,0323 0,0267 0,0374 0,0443 0,0331 0,0331 0,0307 0,0316 0,0288 0,0281 60 Elysia viridis 0,0342 0,0313 0,0278 0,0334 0,0425 0,0308 0,0300 0,0291 0,0293 0,0279 0,0290 61 Elysiella pusilla 0,0341 0,0297 0,0264 0,0326 0,0395 0,0307 0,0299 0,0276 0,0292 0,0285 0,0276 62 Bosellia mimetica 0,0349 0,0306 0,0258 0,0328 0,0395 0,0259 0,0301 0,0264 0,0244 0,0238 0,0284 63 Cyerce nigricans 0,0320 0,0271 0,0204 0,0285 0,0400 0,0252 0,0273 0,0249 0,0238 0,0231 0,0244 64 Placobranchus ocellatus 0,0426 0,0381 0,0323 0,0389 0,0499 0,0346 0,0391 0,0358 0,0331 0,0323 0,0373 65 Placida cremoniana 0,0357 0,0306 0,0250 0,0343 0,0410 0,0307 0,0306 0,0276 0,0292 0,0285 0,0294 66 Polybranchia orientalis 0,0320 0,0300 0,0259 0,0322 0,0428 0,0289 0,0302 0,0293 0,0274 0,0274 0,0299 67 Onchidella floridana 0,0081 0,0131 0,0195 0,0361 0,0163 0,0151 0,0182 0,0176 0,0144 0,0235 68 Onchidium verrucosum 0,0079 0,0094 0,0187 0,0312 0,0163 0,0144 0,0169 0,0163 0,0132 0,0209 69 Siphonaria alternata 0,0125 0,0091 0,0149 0,0289 0,0131 0,0100 0,0162 0,0131 0,0112 0,0184 70 Cepaea hortensis 0,0181 0,0175 0,0142 0,0352 0,0175 0,0181 0,0188 0,0175 0,0162 0,0234 71 Lymnaea stagnalis 0,0313 0,0273 0,0256 0,0307 0,0324 0,0378 0,0344 0,0309 0,0295 0,0339 72 Cassidula nucleus 0,0153 0,0153 0,0125 0,0165 0,0284 0,0113 0,0143 0,0011 0,0034 0,0201 73 Melampus luteus 0,0142 0,0136 0,0096 0,0170 0,0324 0,0108 0,0170 0,0113 0,0101 0,0223 74 Auriculinella bidentata 0,0170 0,0159 0,0153 0,0176 0,0301 0,0136 0,0158 0,0131 0,0118 0,0174 75 Ovatella firminii 0,0165 0,0153 0,0125 0,0165 0,0273 0,0011 0,0108 0,0124 0,0023 0,0188 76 Pythia scarabaeus 0,0136 0,0125 0,0108 0,0153 0,0261 0,0034 0,0096 0,0113 0,0023 0,0175 77 Pyramidella sulcata 0,0215 0,0193 0,0170 0,0215 0,0295 0,0187 0,0204 0,0164 0,0176 0,0164 78 Odostomia sp. 0,0215 0,0187 0,0159 0,0221 0,0289 0,0181 0,0209 0,0170 0,0170 0,0164 0,0056 79 Littoraria undulata 0,0792 0,0769 0,0752 0,0805 0,0837 0,0776 0,0793 0,0753 0,0776 0,0781 0,0769 80 Nodilittorina punctata 0,0797 0,0774 0,0762 0,0797 0,0831 0,0781 0,0786 0,0746 0,0775 0,0780 0,0768 81 Littorina striata 0,0768 0,0745 0,0740 0,0764 0,0797 0,0752 0,0752 0,0718 0,0741 0,0746 0,0728 82 Littorina saxatilis 0,0762 0,0738 0,0739 0,0762 0,0807 0,0763 0,0757 0,0734 0,0757 0,0751 0,0744 83 Littorina littorea 0,0781 0,0758 0,0759 0,0782 0,0833 0,0783 0,0777 0,0760 0,0777 0,0771 0,0770 84 Melaraphe neritoides 0,0792 0,0780 0,0764 0,0787 0,0839 0,0794 0,0781 0,0741 0,0782 0,0782 0,0769 85 Crepidula adunca 0,0784 0,0772 0,0761 0,0779 0,0836 0,0785 0,0773 0,0739 0,0774 0,0773 0,0766 86 Fasciolaria lignaria 0,0791 0,0767 0,0757 0,0786 0,0842 0,0792 0,0780 0,0757 0,0781 0,0775 0,0768 87 Pisania striata 0,0805 0,0781 0,0770 0,0800 0,0862 0,0807 0,0788 0,0772 0,0795 0,0789 0,0781 88 Bursa rana 0,0781 0,0758 0,0741 0,0777 0,0816 0,0765 0,0776 0,0736 0,0754 0,0753 0,0746 89 Truncatella guerinii 0,0802 0,0779 0,0773 0,0808 0,0837 0,0798 0,0803 0,0763 0,0787 0,0780 0,0785 90 Viviparus georgianus 0,0837 0,0814 0,0797 0,0843 0,0871 0,0838 0,0849 0,0809 0,0827 0,0820 0,0842 91 Balcis eburnea 0,0832 0,0809 0,0803 0,0816 0,0868 0,0823 0,0822 0,0787 0,0811 0,0810 0,0815
78 79 80 81 82 83 84 85 86 87 88
1 Bathydoris clavigera 0,1316 0,2151 0,2123 0,2095 0,2059 0,2114 0,2206 0,2059 0,2127 0,2115 0,2062 2 Chromodoris krohni 0,1302 0,2043 0,2031 0,1973 0,2058 0,2062 0,2114 0,1981 0,2043 0,1981 0,1957 3 Dendrodoris fumata 0,1226 0,2057 0,2036 0,1998 0,2033 0,2052 0,2131 0,1957 0,2033 0,1968 0,1975 4 Hypselodoris villafranca 0,1259 0,2060 0,2031 0,2000 0,2068 0,2072 0,2166 0,2007 0,2081 0,2019 0,1980 5 Dendronotus dalli 0,4974 0,5809 0,5737 0,5403 0,5832 0,5770 0,5789 0,5390 0,5652 0,5478 0,5647

256
78 79 80 81 82 83 84 85 86 87 88 6 Cuthona nana 0,4563 0,5541 0,5322 0,5126 0,5323 0,5399 0,5513 0,5156 0,5320 0,5306 0,5153 7 Flabellina babai 0,4431 0,5093 0,4893 0,4792 0,5064 0,5026 0,5066 0,4850 0,5077 0,4991 0,4900 8 Pleurobranchus peroni 0,1064 0,2017 0,2017 0,1849 0,1935 0,1928 0,2034 0,1836 0,1956 0,2003 0,1869 9 Berthellina citrina 0,1042 0,1949 0,1948 0,1761 0,1868 0,1886 0,1953 0,1829 0,1873 0,1905 0,1803
10 Bathyberthella antarctica 0,1118 0,2125 0,2167 0,1966 0,2081 0,2075 0,2170 0,1961 0,2050 0,2093 0,1970 11 Euselenops luniceps 0,1056 0,2031 0,2015 0,1839 0,1969 0,1977 0,2085 0,1848 0,1978 0,2013 0,1879 12 Tompthompsonia antarctica 0,1044 0,1979 0,2016 0,1825 0,1935 0,1946 0,2061 0,1873 0,1952 0,2001 0,1867 13 Berthella stellata 0,3572 0,4544 0,4371 0,4198 0,4471 0,4444 0,4447 0,4182 0,4373 0,4260 0,4224 14 Berthella platei 0,1668 0,2620 0,2574 0,2389 0,2611 0,2626 0,2668 0,2515 0,2561 0,2635 0,2506 15 Pupa solidula 0,0537 0,1384 0,1343 0,1305 0,1306 0,1345 0,1261 0,1265 0,1303 0,1328 0,1268 16 Hydatina physis 0,0476 0,1318 0,1303 0,1279 0,1240 0,1277 0,1222 0,1249 0,1258 0,1268 0,1216 17 Bullina lineata 0,0440 0,1292 0,1277 0,1253 0,1214 0,1251 0,1196 0,1214 0,1232 0,1242 0,1190 18 Acteon sp. 0,0354 0,1265 0,1246 0,1139 0,1113 0,1145 0,1220 0,1176 0,1215 0,1241 0,1231 19 Haminoea hydatis 0,0367 0,1256 0,1226 0,1155 0,1151 0,1196 0,1143 0,1164 0,1213 0,1242 0,1225 20 Cylichna gelida 0,0275 0,1184 0,1172 0,1096 0,1091 0,1137 0,1136 0,1106 0,1154 0,1183 0,1171 21 Scaphander lignarius 0,0303 0,1201 0,1188 0,1091 0,1070 0,1105 0,1151 0,1099 0,1128 0,1157 0,1167 22 cf. Retusa sp. 0,0344 0,1253 0,1253 0,1137 0,1155 0,1215 0,1187 0,1183 0,1222 0,1262 0,1271 23 Bulla cf. striata 0,0262 0,1127 0,1112 0,1040 0,1020 0,1073 0,1070 0,1040 0,1105 0,1115 0,1124 24 Smaragdinella sp. 0,0304 0,1212 0,1194 0,1111 0,1104 0,1138 0,1148 0,1107 0,1155 0,1173 0,1193 25 Phanerophthalmus smaragdinus 0,0327 0,1230 0,1187 0,1139 0,1059 0,1124 0,1139 0,1123 0,1152 0,1201 0,1201 26 Bullacta exarata 0,0290 0,1162 0,1145 0,1074 0,1049 0,1114 0,1140 0,1100 0,1148 0,1177 0,1167 27 Atys semistriatus 0,0310 0,1187 0,1169 0,1117 0,1088 0,1132 0,1132 0,1101 0,1149 0,1178 0,1188 28 Odontoglaja guamensis 0,0318 0,1218 0,1202 0,1139 0,1126 0,1182 0,1208 0,1177 0,1216 0,1254 0,1212 29 Navanax inermis 0,0329 0,1258 0,1259 0,1178 0,1139 0,1185 0,1210 0,1179 0,1218 0,1268 0,1258 30 Chelidonura inornata 0,0310 0,1245 0,1240 0,1165 0,1110 0,1166 0,1192 0,1166 0,1215 0,1255 0,1255 31 Philinopsis pilsbryi 0,0323 0,1262 0,1238 0,1163 0,1107 0,1164 0,1189 0,1164 0,1213 0,1252 0,1275 32 Philinoglossa praelongata 0,0341 0,1219 0,1218 0,1155 0,1150 0,1170 0,1197 0,1166 0,1215 0,1237 0,1232 33 Sagaminopteron psychedelicum 0,0359 0,1171 0,1131 0,1084 0,1071 0,1105 0,1118 0,1081 0,1136 0,1118 0,1146 34 Toledonia globosa 0,0256 0,1157 0,1144 0,1069 0,1049 0,1114 0,1156 0,1099 0,1117 0,1134 0,1102 35 Diaphana cf. inflata 0,0324 0,1173 0,1153 0,1066 0,1019 0,1062 0,1122 0,1066 0,1114 0,1133 0,1152 36 Newnesia antarctica 0,0289 0,1174 0,1163 0,1095 0,1027 0,1142 0,1175 0,1137 0,1135 0,1172 0,1143 37 Colpodaspis thomsoni 0,0442 0,1232 0,1275 0,1223 0,1235 0,1268 0,1307 0,1280 0,1292 0,1271 0,1310 38 Runcina adriatica 0,0352 0,1136 0,1127 0,1050 0,1053 0,1125 0,1142 0,1154 0,1158 0,1179 0,1129 39 Unela glandulifera 0,0201 0,1082 0,1088 0,1008 0,1007 0,1079 0,1112 0,1088 0,1086 0,1082 0,1063 40 Pontohedyle milatchevitchi 0,0304 0,1112 0,1077 0,1022 0,0989 0,1036 0,1092 0,1033 0,1061 0,1118 0,1080 41 Hedylopsis spiculifera 0,0168 0,1059 0,1064 0,1003 0,1012 0,1060 0,1071 0,1047 0,1033 0,1077 0,1032 42 Parahedyle cryptophthalma 0,0201 0,1081 0,1087 0,1007 0,1006 0,1078 0,1111 0,1087 0,1085 0,1081 0,1062 43 Umbraculum mediterraneum 0,0244 0,1181 0,1186 0,1103 0,1099 0,1186 0,1161 0,1143 0,1188 0,1190 0,1182 44 Tylodina perversa 0,0258 0,1182 0,1177 0,1095 0,1073 0,1169 0,1185 0,1190 0,1177 0,1172 0,1130 45 Akera bullata 0,0229 0,1017 0,1021 0,0945 0,0943 0,1022 0,1008 0,1012 0,1015 0,1027 0,1007 46 Aplysia californica 0,0215 0,1066 0,1070 0,0992 0,0969 0,1062 0,1057 0,1027 0,1034 0,1052 0,1025 47 Dolabella auricularia 0,0209 0,1034 0,1039 0,0961 0,0971 0,1041 0,1026 0,1013 0,1012 0,1005 0,1024 48 Siphonota geographica 0,0230 0,1052 0,1061 0,0959 0,0995 0,1041 0,1061 0,1018 0,1025 0,1033 0,1016 49 Bursatella leachii 0,0210 0,1042 0,1061 0,0992 0,0961 0,1041 0,1057 0,1052 0,1055 0,1058 0,1026 50 Petalifera petalifera 0,0195 0,1029 0,1048 0,0980 0,0963 0,1044 0,1036 0,1028 0,1027 0,1044 0,1013 51 Dolabrifera brazieri 0,0196 0,1041 0,1061 0,0992 0,0975 0,1056 0,1048 0,1041 0,1039 0,1056 0,1026 52 Stylocheilus longicauda 0,0209 0,1031 0,1050 0,0982 0,0932 0,1012 0,1028 0,1011 0,1010 0,1027 0,1018 53 Clio pyramidata 0,0439 0,1111 0,1125 0,1079 0,1077 0,1123 0,1161 0,1122 0,1149 0,1189 0,1169 54 Diacria quadridentata 0,0506 0,1235 0,1221 0,1159 0,1167 0,1199 0,1257 0,1196 0,1210 0,1241 0,1218 55 Creseis acicula 0,0707 0,1457 0,1379 0,1313 0,1323 0,1326 0,1405 0,1289 0,1342 0,1376 0,1392 56 Spongiobranchaea australis 0,0300 0,1138 0,1131 0,1051 0,1049 0,1133 0,1117 0,1111 0,1110 0,1127 0,1095 57 Calliopaea bellula 0,0452 0,1397 0,1376 0,1282 0,1283 0,1375 0,1358 0,1326 0,1359 0,1405 0,1384 58 Limapontia nigra 0,0360 0,1278 0,1265 0,1191 0,1232 0,1273 0,1230 0,1233 0,1258 0,1293 0,1246 59 Thuridilla bayeri 0,0288 0,1289 0,1306 0,1221 0,1253 0,1294 0,1268 0,1244 0,1249 0,1282 0,1222 60 Elysia viridis 0,0291 0,1293 0,1300 0,1198 0,1210 0,1260 0,1251 0,1211 0,1235 0,1269 0,1236 61 Elysiella pusilla 0,0277 0,1285 0,1292 0,1192 0,1232 0,1273 0,1264 0,1224 0,1248 0,1281 0,1272 62 Bosellia mimetica 0,0285 0,1258 0,1255 0,1182 0,1222 0,1263 0,1247 0,1234 0,1238 0,1272 0,1212 63 Cyerce nigricans 0,0244 0,1221 0,1208 0,1136 0,1157 0,1197 0,1181 0,1168 0,1173 0,1205 0,1147 64 Placobranchus ocellatus 0,0374 0,1403 0,1410 0,1332 0,1367 0,1410 0,1378 0,1379 0,1383 0,1418 0,1355 65 Placida cremoniana 0,0280 0,1217 0,1204 0,1132 0,1143 0,1202 0,1177 0,1183 0,1179 0,1211 0,1175 66 Polybranchia orientalis 0,0292 0,1229 0,1226 0,1118 0,1168 0,1207 0,1202 0,1169 0,1142 0,1206 0,1164 67 Onchidella floridana 0,0236 0,1112 0,1133 0,1074 0,1051 0,1099 0,1110 0,1102 0,1111 0,1139 0,1091 68 Onchidium verrucosum 0,0202 0,1071 0,1091 0,1033 0,1011 0,1058 0,1088 0,1079 0,1070 0,1097 0,1050 69 Siphonaria alternata 0,0170 0,1045 0,1077 0,1032 0,1019 0,1066 0,1062 0,1068 0,1054 0,1085 0,1025 70 Cepaea hortensis 0,0240 0,1140 0,1143 0,1072 0,1058 0,1107 0,1108 0,1098 0,1103 0,1135 0,1089 71 Lymnaea stagnalis 0,0333 0,1238 0,1227 0,1161 0,1164 0,1228 0,1223 0,1225 0,1235 0,1279 0,1180 72 Cassidula nucleus 0,0195 0,1092 0,1115 0,1058 0,1063 0,1112 0,1122 0,1114 0,1118 0,1151 0,1071 73 Melampus luteus 0,0229 0,1126 0,1124 0,1053 0,1048 0,1097 0,1096 0,1089 0,1093 0,1116 0,1092 74 Auriculinella bidentata 0,0181 0,1052 0,1049 0,0995 0,1009 0,1066 0,1028 0,1030 0,1053 0,1085 0,1018 75 Ovatella firminii 0,0181 0,1094 0,1101 0,1035 0,1050 0,1098 0,1098 0,1090 0,1094 0,1126 0,1048 76 Pythia scarabaeus 0,0175 0,1105 0,1113 0,1047 0,1039 0,1087 0,1100 0,1092 0,1087 0,1119 0,1050 77 Pyramidella sulcata 0,0058 0,1086 0,1094 0,1014 0,1029 0,1086 0,1081 0,1077 0,1073 0,1104 0,1041 78 Odostomia sp. 0,1053 0,1060 0,0973 0,1019 0,1057 0,1058 0,1035 0,1031 0,1062 0,0999 79 Littoraria undulata 0,0751 0,0152 0,0178 0,0287 0,0288 0,0298 0,0317 0,0321 0,0355 0,0264 80 Nodilittorina punctata 0,0750 0,0142 0,0121 0,0211 0,0204 0,0216 0,0245 0,0251 0,0291 0,0210 81 Littorina striata 0,0705 0,0165 0,0113 0,0192 0,0191 0,0210 0,0204 0,0224 0,0249 0,0190
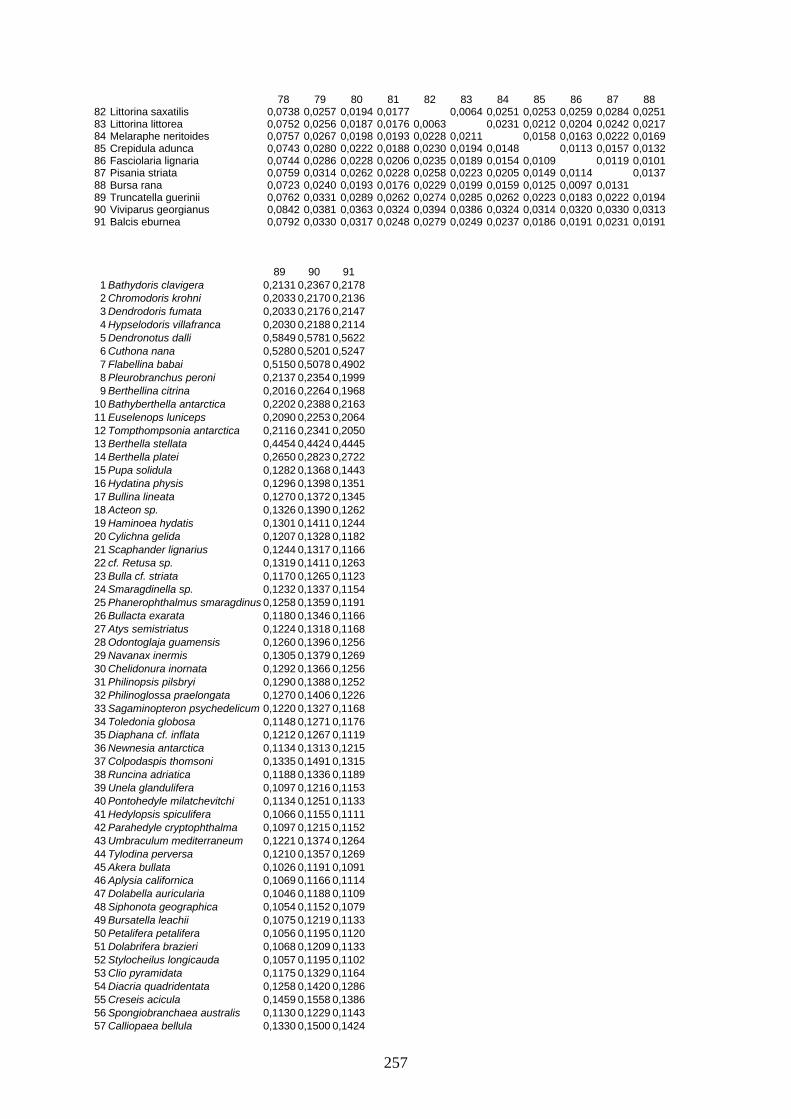
257
78 79 80 81 82 83 84 85 86 87 88 82 Littorina saxatilis 0,0738 0,0257 0,0194 0,0177 0,0064 0,0251 0,0253 0,0259 0,0284 0,0251 83 Littorina littorea 0,0752 0,0256 0,0187 0,0176 0,0063 0,0231 0,0212 0,0204 0,0242 0,0217 84 Melaraphe neritoides 0,0757 0,0267 0,0198 0,0193 0,0228 0,0211 0,0158 0,0163 0,0222 0,0169 85 Crepidula adunca 0,0743 0,0280 0,0222 0,0188 0,0230 0,0194 0,0148 0,0113 0,0157 0,0132 86 Fasciolaria lignaria 0,0744 0,0286 0,0228 0,0206 0,0235 0,0189 0,0154 0,0109 0,0119 0,0101 87 Pisania striata 0,0759 0,0314 0,0262 0,0228 0,0258 0,0223 0,0205 0,0149 0,0114 0,0137 88 Bursa rana 0,0723 0,0240 0,0193 0,0176 0,0229 0,0199 0,0159 0,0125 0,0097 0,0131 89 Truncatella guerinii 0,0762 0,0331 0,0289 0,0262 0,0274 0,0285 0,0262 0,0223 0,0183 0,0222 0,0194 90 Viviparus georgianus 0,0842 0,0381 0,0363 0,0324 0,0394 0,0386 0,0324 0,0314 0,0320 0,0330 0,0313 91 Balcis eburnea 0,0792 0,0330 0,0317 0,0248 0,0279 0,0249 0,0237 0,0186 0,0191 0,0231 0,0191
89 90 91 1 Bathydoris clavigera 0,2131 0,2367 0,2178 2 Chromodoris krohni 0,2033 0,2170 0,2136 3 Dendrodoris fumata 0,2033 0,2176 0,2147 4 Hypselodoris villafranca 0,2030 0,2188 0,2114 5 Dendronotus dalli 0,5849 0,5781 0,5622 6 Cuthona nana 0,5280 0,5201 0,5247 7 Flabellina babai 0,5150 0,5078 0,4902 8 Pleurobranchus peroni 0,2137 0,2354 0,1999 9 Berthellina citrina 0,2016 0,2264 0,1968
10 Bathyberthella antarctica 0,2202 0,2388 0,2163 11 Euselenops luniceps 0,2090 0,2253 0,2064 12 Tompthompsonia antarctica 0,2116 0,2341 0,2050 13 Berthella stellata 0,4454 0,4424 0,4445 14 Berthella platei 0,2650 0,2823 0,2722 15 Pupa solidula 0,1282 0,1368 0,1443 16 Hydatina physis 0,1296 0,1398 0,1351 17 Bullina lineata 0,1270 0,1372 0,1345 18 Acteon sp. 0,1326 0,1390 0,1262 19 Haminoea hydatis 0,1301 0,1411 0,1244 20 Cylichna gelida 0,1207 0,1328 0,1182 21 Scaphander lignarius 0,1244 0,1317 0,1166 22 cf. Retusa sp. 0,1319 0,1411 0,1263 23 Bulla cf. striata 0,1170 0,1265 0,1123 24 Smaragdinella sp. 0,1232 0,1337 0,1154 25 Phanerophthalmus smaragdinus 0,1258 0,1359 0,1191 26 Bullacta exarata 0,1180 0,1346 0,1166 27 Atys semistriatus 0,1224 0,1318 0,1168 28 Odontoglaja guamensis 0,1260 0,1396 0,1256 29 Navanax inermis 0,1305 0,1379 0,1269 30 Chelidonura inornata 0,1292 0,1366 0,1256 31 Philinopsis pilsbryi 0,1290 0,1388 0,1252 32 Philinoglossa praelongata 0,1270 0,1406 0,1226 33 Sagaminopteron psychedelicum 0,1220 0,1327 0,1168 34 Toledonia globosa 0,1148 0,1271 0,1176 35 Diaphana cf. inflata 0,1212 0,1267 0,1119 36 Newnesia antarctica 0,1134 0,1313 0,1215 37 Colpodaspis thomsoni 0,1335 0,1491 0,1315 38 Runcina adriatica 0,1188 0,1336 0,1189 39 Unela glandulifera 0,1097 0,1216 0,1153 40 Pontohedyle milatchevitchi 0,1134 0,1251 0,1133 41 Hedylopsis spiculifera 0,1066 0,1155 0,1111 42 Parahedyle cryptophthalma 0,1097 0,1215 0,1152 43 Umbraculum mediterraneum 0,1221 0,1374 0,1264 44 Tylodina perversa 0,1210 0,1357 0,1269 45 Akera bullata 0,1026 0,1191 0,1091 46 Aplysia californica 0,1069 0,1166 0,1114 47 Dolabella auricularia 0,1046 0,1188 0,1109 48 Siphonota geographica 0,1054 0,1152 0,1079 49 Bursatella leachii 0,1075 0,1219 0,1133 50 Petalifera petalifera 0,1056 0,1195 0,1120 51 Dolabrifera brazieri 0,1068 0,1209 0,1133 52 Stylocheilus longicauda 0,1057 0,1195 0,1102 53 Clio pyramidata 0,1175 0,1329 0,1164 54 Diacria quadridentata 0,1258 0,1420 0,1286 55 Creseis acicula 0,1459 0,1558 0,1386 56 Spongiobranchaea australis 0,1130 0,1229 0,1143 57 Calliopaea bellula 0,1330 0,1500 0,1424

258
89 90 91 58 Limapontia nigra 0,1282 0,1403 0,1317 59 Thuridilla bayeri 0,1272 0,1353 0,1340 60 Elysia viridis 0,1248 0,1367 0,1325 61 Elysiella pusilla 0,1281 0,1382 0,1338 62 Bosellia mimetica 0,1281 0,1352 0,1328 63 Cyerce nigricans 0,1204 0,1273 0,1259 64 Placobranchus ocellatus 0,1405 0,1411 0,1415 65 Placida cremoniana 0,1189 0,1300 0,1245 66 Polybranchia orientalis 0,1195 0,1312 0,1227 67 Onchidella floridana 0,1143 0,1241 0,1192 68 Onchidium verrucosum 0,1101 0,1197 0,1149 69 Siphonaria alternata 0,1096 0,1166 0,1149 70 Cepaea hortensis 0,1155 0,1255 0,1171 71 Lymnaea stagnalis 0,1224 0,1321 0,1291 72 Cassidula nucleus 0,1146 0,1246 0,1184 73 Melampus luteus 0,1150 0,1268 0,1181 74 Auriculinella bidentata 0,1080 0,1188 0,1116 75 Ovatella firminii 0,1122 0,1220 0,1159 76 Pythia scarabaeus 0,1111 0,1209 0,1161 77 Pyramidella sulcata 0,1123 0,1255 0,1170 78 Odostomia sp. 0,1076 0,1256 0,1126 79 Littoraria undulata 0,0380 0,0460 0,0378 80 Nodilittorina punctata 0,0328 0,0435 0,0364 81 Littorina striata 0,0293 0,0383 0,0274 82 Littorina saxatilis 0,0309 0,0479 0,0312 83 Littorina littorea 0,0322 0,0472 0,0276 84 Melaraphe neritoides 0,0291 0,0381 0,0260 85 Crepidula adunca 0,0245 0,0368 0,0200 86 Fasciolaria lignaria 0,0197 0,0372 0,0206 87 Pisania striata 0,0243 0,0389 0,0252 88 Bursa rana 0,0209 0,0363 0,0205 89 Truncatella guerinii 0,0416 0,0301 90 Viviparus georgianus 0,0355 0,0429 91 Balcis eburnea 0,0271 0,0362
Opisthobranchia-Datensatz (16S rDNA) Dargestellt sind die beobachteten (unten) und korrigierten (oben) Distanzen der 16S rDNA-Sequenzen. Die Parameter des für die Korrektur der beobachteten Distanzen verwendeten Sequenzevolutionsmodells wurden mit Hilfe hierarchischer likelihood ratio tests (Modeltest, Posada & Crandall 1998) ermittelt.
1 2 3 4 5 6 7 8 9 10 11 1 Bathydoris clavigera 0,4214 0,2610 0,3756 0,2716 0,4561 0,9461 0,2895 0,2762 0,2748 0,4338 2 Archidoris pseudoargus 0,2156 0,1861 0,2055 0,1856 0,2602 1,0E+08 0,5265 0,5019 0,4415 0,5843 3 Goniodoris nodosa 0,1642 0,1239 0,1899 0,1288 0,3041 0,8036 0,3105 0,2858 0,2824 0,3956 4 Chromodoris krohni 0,1997 0,1372 0,1298 0,2097 0,2439 0,8538 0,4588 0,4109 0,3165 0,5270 5 Discodoris concinna 0,1666 0,1260 0,0992 0,1403 0,2801 0,8780 0,3981 0,3541 0,3517 0,4723 6 Hypselodoris villafranca 0,2266 0,1604 0,1814 0,1593 0,1737 0,9822 0,5357 0,5412 0,4896 0,5737 7 Dendrodoris nigra 0,3068 0,3163 0,2840 0,2987 0,2974 0,3050 1,1E+08 1,1E+08 1,1E+08 1,0E+08 8 Eubranchus exiguus 0,1799 0,2542 0,1899 0,2333 0,2194 0,2611 0,3264 0,0792 0,1626 0,2892 9 Flabellina pedata 0,1732 0,2418 0,1777 0,2147 0,2034 0,2611 0,3267 0,0709 0,1222 0,3018
10 Dendronotus dalli 0,1761 0,2287 0,1772 0,1893 0,2026 0,2382 0,3253 0,1234 0,0986 0,2989 11 Berthella sideralis 0,2225 0,2566 0,2144 0,2449 0,2332 0,2540 0,3037 0,1800 0,1874 0,1833 12 Berthella platei 0,2129 0,2524 0,2138 0,2413 0,2324 0,2578 0,2989 0,1834 0,1932 0,1868 0,0034 13 Pleurobranchus peroni 0,2163 0,2338 0,2008 0,2317 0,2101 0,2387 0,2986 0,1895 0,1713 0,1773 0,0742 14 Tomthompsonia antarctica 0,2091 0,2211 0,2166 0,2288 0,2031 0,2260 0,3045 0,2019 0,2052 0,2016 0,0987 15 Umbraculum umbraculum 0,2079 0,2345 0,1948 0,2352 0,2019 0,2532 0,2772 0,2461 0,2248 0,2274 0,2195 16 Aplysia oculifera 0,2090 0,2326 0,1958 0,2399 0,2122 0,2477 0,2852 0,2511 0,2419 0,2316 0,2161 17 Dolabella 0,2175 0,2498 0,2039 0,2414 0,2081 0,2493 0,2965 0,2680 0,2463 0,2328 0,2051 18 Stylocheilus longicauda 0,1914 0,2530 0,2074 0,2448 0,2145 0,2590 0,2775 0,2282 0,2193 0,2179 0,1989

259
1 2 3 4 5 6 7 8 9 10 11 19 Bursatella leachii 0,2142 0,2439 0,2074 0,2419 0,2050 0,2528 0,2842 0,2436 0,2346 0,2333 0,2160 20 Dolabrifera dolabrifera 0,2147 0,2471 0,2109 0,2231 0,2084 0,2499 0,2842 0,2438 0,2285 0,2273 0,1991 21 Phyllaplysia taylori 0,2123 0,2415 0,2052 0,2268 0,2057 0,2537 0,2917 0,2476 0,2261 0,2278 0,2101 22 Petalifera ramosa 0,2125 0,2449 0,2051 0,2427 0,2161 0,2703 0,3020 0,2508 0,2417 0,2370 0,2141 23 Notarchus indicus 0,2130 0,2367 0,2061 0,2281 0,2167 0,2589 0,2922 0,2549 0,2427 0,2386 0,2104 24 Akera bullata 0,2116 0,2321 0,1857 0,2140 0,1922 0,2345 0,2961 0,2406 0,2162 0,2123 0,1947 25 Cavolinia sp, 0,3224 0,3510 0,3141 0,3262 0,3375 0,3326 0,3225 0,3352 0,3268 0,3283 0,2865 26 Clione limacina 0,2996 0,3138 0,3069 0,2907 0,2969 0,3156 0,3493 0,3317 0,3221 0,3015 0,3177 27 Diaphana minuta 0,2163 0,2432 0,2061 0,2280 0,2000 0,2496 0,3100 0,2359 0,2195 0,2322 0,2299 28 Haminoea cymbalum 0,2077 0,2581 0,2189 0,2291 0,2142 0,2790 0,3444 0,1962 0,2103 0,2331 0,2271 29 Smaragdinella sp, 0,2107 0,2666 0,2195 0,2296 0,2406 0,2701 0,3316 0,2119 0,2055 0,2496 0,2269 30 Bulla gouldiana 0,2314 0,2539 0,2200 0,2396 0,2241 0,2766 0,3336 0,2091 0,2127 0,2406 0,2121 31 Philine aperta 0,2207 0,2319 0,1937 0,2297 0,1975 0,2250 0,3207 0,2150 0,2247 0,2244 0,2286 32 Scaphander punctostriatus 0,2663 0,2556 0,2290 0,2639 0,2231 0,2694 0,3309 0,2672 0,2672 0,2622 0,2320 33 Elysia viridis 0,2037 0,2573 0,2243 0,2424 0,2230 0,2588 0,2936 0,2514 0,2413 0,2334 0,2652 34 Thuridilla ratna 0,2042 0,2644 0,2356 0,2471 0,2436 0,2514 0,2778 0,2427 0,2225 0,2318 0,2487 35 Placida dendritica 0,2146 0,2418 0,2303 0,2546 0,2288 0,2605 0,3241 0,2430 0,2429 0,2230 0,2653 36 Acteon tornatilis 0,2040 0,2401 0,1936 0,2244 0,1991 0,2511 0,3077 0,2245 0,2122 0,2341 0,2316 37 Pupa solidula 0,2256 0,2925 0,2650 0,2826 0,2544 0,2762 0,3187 0,2424 0,2245 0,2107 0,2331 38 Hydatina physis 0,2070 0,2423 0,2231 0,2447 0,2169 0,2610 0,2896 0,2506 0,2323 0,2485 0,2365 39 Pontohedyle milatchevitchi 0,2798 0,3142 0,2826 0,2906 0,2948 0,3330 0,3239 0,3037 0,3181 0,2901 0,2871 40 Parahedyle cryptophthalma 0,2524 0,2524 0,2380 0,2215 0,2535 0,2774 0,2941 0,2844 0,2759 0,2591 0,2333 41 Unela glandulifera 0,2524 0,2525 0,2381 0,2215 0,2534 0,2774 0,2975 0,2845 0,2760 0,2592 0,2330 42 Pythia scarabaeus 0,2520 0,3155 0,2739 0,2628 0,2732 0,2886 0,3080 0,2590 0,2605 0,2500 0,2604 43 Odostomia sp, 0,3268 0,3289 0,3107 0,3151 0,3270 0,3346 0,3171 0,3224 0,3074 0,3158 0,3150 44 Littorina scutulata 0,3097 0,3113 0,2810 0,2964 0,2995 0,3116 0,3496 0,3239 0,3091 0,3180 0,3084 45 Tectarius viviparus 0,3039 0,2910 0,3031 0,3249 0,2800 0,3261 0,3452 0,3268 0,3056 0,3233 0,3295
12 13 14 15 16 17 18 19 20 21 22
1 Bathydoris clavigera 0,3968 0,4030 0,3680 0,3743 0,4191 0,4456 0,3510 0,4235 0,4310 0,4296 0,4209 2 Archidoris pseudoargus 0,5637 0,4996 0,4591 0,4701 0,5137 0,5758 0,5741 0,5395 0,5503 0,5424 0,5454 3 Goniodoris nodosa 0,3887 0,3628 0,4028 0,3510 0,3886 0,4026 0,4078 0,4060 0,4212 0,4116 0,4022 4 Chromodoris krohni 0,5027 0,4768 0,4675 0,4755 0,5734 0,5535 0,5638 0,5558 0,4926 0,5164 0,5644 5 Discodoris concinna 0,4732 0,4005 0,3778 0,3665 0,4375 0,4118 0,4388 0,3882 0,4033 0,4049 0,4279 6 Hypselodoris villafranca 0,5958 0,5125 0,4636 0,5644 0,5666 0,5831 0,6190 0,5819 0,5813 0,5855 0,6567 7 Dendrodoris nigra 0,9443 0,9611 0,9964 0,7897 0,8747 0,9435 0,8249 0,8548 0,8529 0,8796 0,9614 8 Eubranchus exiguus 0,2991 0,3187 0,3332 0,4912 0,5376 0,6067 0,4555 0,4966 0,5077 0,5082 0,5300 9 Flabellina pedata 0,3158 0,2721 0,3513 0,4400 0,5195 0,5373 0,4243 0,4724 0,4645 0,4487 0,5065
10 Dendronotus dalli 0,3027 0,2895 0,3406 0,4204 0,4857 0,4812 0,4102 0,4712 0,4468 0,4558 0,4801 11 Berthella sideralis 0,0033 0,0881 0,1262 0,4374 0,4521 0,4018 0,3736 0,4383 0,3832 0,4163 0,4265 12 Berthella platei 0,0791 0,1172 0,4403 0,4633 0,4116 0,3875 0,4479 0,3877 0,4182 0,4277 13 Pleurobranchus peroni 0,0679 0,1083 0,4099 0,4323 0,4037 0,3778 0,3709 0,3804 0,4038 0,4111 14 Tomthompsonia antarctica 0,0933 0,0868 0,3952 0,4136 0,3683 0,3903 0,3652 0,3367 0,3743 0,3892 15 Umbraculum umbraculum 0,2191 0,2102 0,2068 0,1955 0,1743 0,1789 0,2146 0,1969 0,2068 0,1975 16 Aplysia oculifera 0,2172 0,2076 0,2080 0,1245 0,0631 0,0895 0,0907 0,0813 0,0718 0,0714 17 Dolabella 0,2061 0,2063 0,1970 0,1153 0,0557 0,0943 0,0860 0,0693 0,0822 0,0571 18 Stylocheilus longicauda 0,2038 0,2040 0,2103 0,1214 0,0745 0,0805 0,0652 0,0644 0,0614 0,0603 19 Bursatella leachii 0,2197 0,1979 0,2010 0,1371 0,0744 0,0743 0,0588 0,0563 0,0502 0,0603 20 Dolabrifera dolabrifera 0,2006 0,2007 0,1883 0,1277 0,0683 0,0619 0,0588 0,0526 0,0188 0,0315 21 Phyllaplysia taylori 0,2108 0,2077 0,2015 0,1311 0,0624 0,0715 0,0559 0,0465 0,0186 0,0285 22 Petalifera ramosa 0,2139 0,2106 0,2078 0,1279 0,0624 0,0528 0,0560 0,0560 0,0311 0,0280 23 Notarchus indicus 0,2084 0,2118 0,2055 0,1315 0,0628 0,0625 0,0873 0,0935 0,0624 0,0657 0,0534 24 Akera bullata 0,1947 0,2010 0,2106 0,1313 0,0841 0,0836 0,1056 0,1087 0,0870 0,0843 0,0875 25 Cavolinia sp, 0,2903 0,2999 0,3257 0,2986 0,2886 0,2979 0,2821 0,2949 0,2881 0,2888 0,3018 26 Clione limacina 0,3154 0,3016 0,3067 0,2429 0,2652 0,2604 0,2617 0,2615 0,2652 0,2693 0,2599 27 Diaphana minuta 0,2298 0,2201 0,2166 0,1352 0,1697 0,1636 0,1563 0,1630 0,1698 0,1733 0,1798 28 Haminoea cymbalum 0,2233 0,2437 0,2398 0,1864 0,2117 0,2019 0,2081 0,2219 0,2091 0,2166 0,2130 29 Smaragdinella sp, 0,2291 0,2417 0,2479 0,1856 0,2068 0,1915 0,1945 0,2102 0,1975 0,2044 0,2016
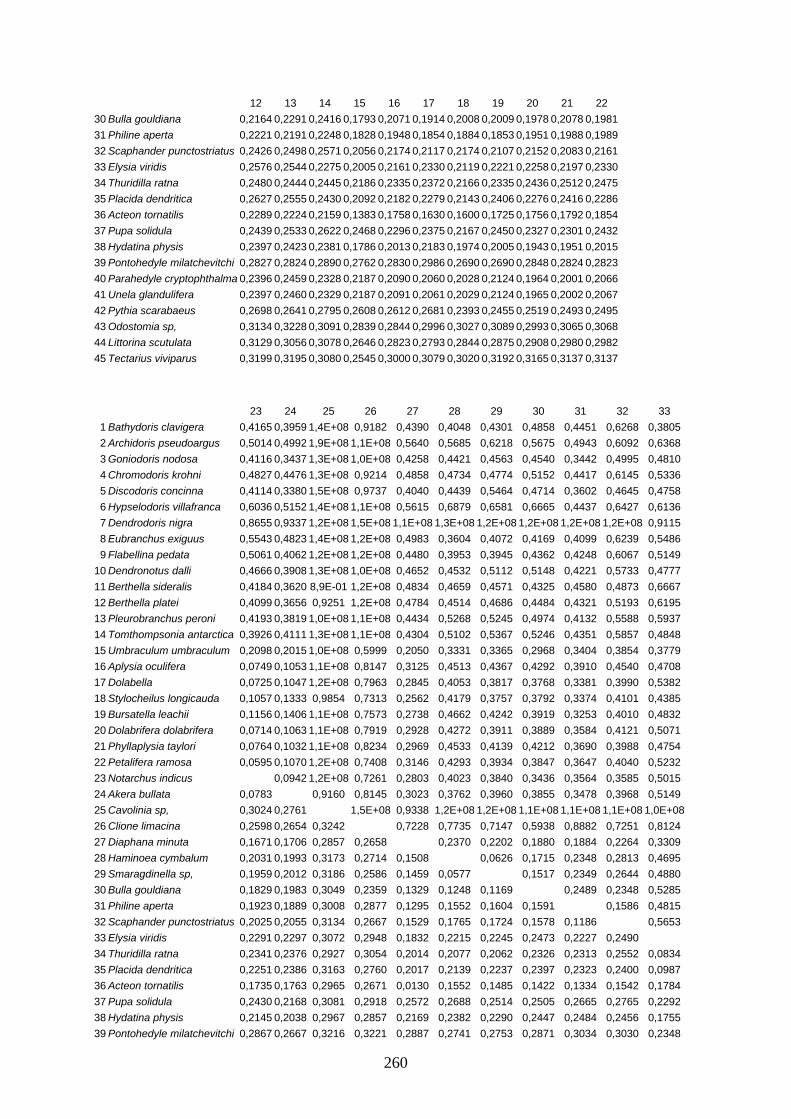
260
12 13 14 15 16 17 18 19 20 21 22 30 Bulla gouldiana 0,2164 0,2291 0,2416 0,1793 0,2071 0,1914 0,2008 0,2009 0,1978 0,2078 0,1981 31 Philine aperta 0,2221 0,2191 0,2248 0,1828 0,1948 0,1854 0,1884 0,1853 0,1951 0,1988 0,1989 32 Scaphander punctostriatus 0,2426 0,2498 0,2571 0,2056 0,2174 0,2117 0,2174 0,2107 0,2152 0,2083 0,2161 33 Elysia viridis 0,2576 0,2544 0,2275 0,2005 0,2161 0,2330 0,2119 0,2221 0,2258 0,2197 0,2330 34 Thuridilla ratna 0,2480 0,2444 0,2445 0,2186 0,2335 0,2372 0,2166 0,2335 0,2436 0,2512 0,2475 35 Placida dendritica 0,2627 0,2555 0,2430 0,2092 0,2182 0,2279 0,2143 0,2406 0,2276 0,2416 0,2286 36 Acteon tornatilis 0,2289 0,2224 0,2159 0,1383 0,1758 0,1630 0,1600 0,1725 0,1756 0,1792 0,1854 37 Pupa solidula 0,2439 0,2533 0,2622 0,2468 0,2296 0,2375 0,2167 0,2450 0,2327 0,2301 0,2432 38 Hydatina physis 0,2397 0,2423 0,2381 0,1786 0,2013 0,2183 0,1974 0,2005 0,1943 0,1951 0,2015 39 Pontohedyle milatchevitchi 0,2827 0,2824 0,2890 0,2762 0,2830 0,2986 0,2690 0,2690 0,2848 0,2824 0,2823 40 Parahedyle cryptophthalma 0,2396 0,2459 0,2328 0,2187 0,2090 0,2060 0,2028 0,2124 0,1964 0,2001 0,2066 41 Unela glandulifera 0,2397 0,2460 0,2329 0,2187 0,2091 0,2061 0,2029 0,2124 0,1965 0,2002 0,2067 42 Pythia scarabaeus 0,2698 0,2641 0,2795 0,2608 0,2612 0,2681 0,2393 0,2455 0,2519 0,2493 0,2495 43 Odostomia sp, 0,3134 0,3228 0,3091 0,2839 0,2844 0,2996 0,3027 0,3089 0,2993 0,3065 0,3068 44 Littorina scutulata 0,3129 0,3056 0,3078 0,2646 0,2823 0,2793 0,2844 0,2875 0,2908 0,2980 0,2982 45 Tectarius viviparus 0,3199 0,3195 0,3080 0,2545 0,3000 0,3079 0,3020 0,3192 0,3165 0,3137 0,3137
23 24 25 26 27 28 29 30 31 32 33 1 Bathydoris clavigera 0,4165 0,3959 1,4E+08 0,9182 0,4390 0,4048 0,4301 0,4858 0,4451 0,6268 0,3805 2 Archidoris pseudoargus 0,5014 0,4992 1,9E+08 1,1E+08 0,5640 0,5685 0,6218 0,5675 0,4943 0,6092 0,6368 3 Goniodoris nodosa 0,4116 0,3437 1,3E+08 1,0E+08 0,4258 0,4421 0,4563 0,4540 0,3442 0,4995 0,4810 4 Chromodoris krohni 0,4827 0,4476 1,3E+08 0,9214 0,4858 0,4734 0,4774 0,5152 0,4417 0,6145 0,5336 5 Discodoris concinna 0,4114 0,3380 1,5E+08 0,9737 0,4040 0,4439 0,5464 0,4714 0,3602 0,4645 0,4758 6 Hypselodoris villafranca 0,6036 0,5152 1,4E+08 1,1E+08 0,5615 0,6879 0,6581 0,6665 0,4437 0,6427 0,6136 7 Dendrodoris nigra 0,8655 0,9337 1,2E+08 1,5E+08 1,1E+08 1,3E+08 1,2E+08 1,2E+08 1,2E+08 1,2E+08 0,9115 8 Eubranchus exiguus 0,5543 0,4823 1,4E+08 1,2E+08 0,4983 0,3604 0,4072 0,4169 0,4099 0,6239 0,5486 9 Flabellina pedata 0,5061 0,4062 1,2E+08 1,2E+08 0,4480 0,3953 0,3945 0,4362 0,4248 0,6067 0,5149
10 Dendronotus dalli 0,4666 0,3908 1,3E+08 1,0E+08 0,4652 0,4532 0,5112 0,5148 0,4221 0,5733 0,4777 11 Berthella sideralis 0,4184 0,3620 8,9E-01 1,2E+08 0,4834 0,4659 0,4571 0,4325 0,4580 0,4873 0,6667 12 Berthella platei 0,4099 0,3656 0,9251 1,2E+08 0,4784 0,4514 0,4686 0,4484 0,4321 0,5193 0,6195 13 Pleurobranchus peroni 0,4193 0,3819 1,0E+08 1,1E+08 0,4434 0,5268 0,5245 0,4974 0,4132 0,5588 0,5937 14 Tomthompsonia antarctica 0,3926 0,4111 1,3E+08 1,1E+08 0,4304 0,5102 0,5367 0,5246 0,4351 0,5857 0,4848 15 Umbraculum umbraculum 0,2098 0,2015 1,0E+08 0,5999 0,2050 0,3331 0,3365 0,2968 0,3404 0,3854 0,3779 16 Aplysia oculifera 0,0749 0,1053 1,1E+08 0,8147 0,3125 0,4513 0,4367 0,4292 0,3910 0,4540 0,4708 17 Dolabella 0,0725 0,1047 1,2E+08 0,7963 0,2845 0,4053 0,3817 0,3768 0,3381 0,3990 0,5382 18 Stylocheilus longicauda 0,1057 0,1333 0,9854 0,7313 0,2562 0,4179 0,3757 0,3792 0,3374 0,4101 0,4385 19 Bursatella leachii 0,1156 0,1406 1,1E+08 0,7573 0,2738 0,4662 0,4242 0,3919 0,3253 0,4010 0,4832 20 Dolabrifera dolabrifera 0,0714 0,1063 1,1E+08 0,7919 0,2928 0,4272 0,3911 0,3889 0,3584 0,4121 0,5071 21 Phyllaplysia taylori 0,0764 0,1032 1,1E+08 0,8234 0,2969 0,4533 0,4139 0,4212 0,3690 0,3988 0,4754 22 Petalifera ramosa 0,0595 0,1070 1,2E+08 0,7408 0,3146 0,4293 0,3934 0,3847 0,3647 0,4040 0,5232 23 Notarchus indicus 0,0942 1,2E+08 0,7261 0,2803 0,4023 0,3840 0,3436 0,3564 0,3585 0,5015 24 Akera bullata 0,0783 0,9160 0,8145 0,3023 0,3762 0,3960 0,3855 0,3478 0,3968 0,5149 25 Cavolinia sp, 0,3024 0,2761 1,5E+08 0,9338 1,2E+08 1,2E+08 1,1E+08 1,1E+08 1,1E+08 1,0E+08 26 Clione limacina 0,2598 0,2654 0,3242 0,7228 0,7735 0,7147 0,5938 0,8882 0,7251 0,8124 27 Diaphana minuta 0,1671 0,1706 0,2857 0,2658 0,2370 0,2202 0,1880 0,1884 0,2264 0,3309 28 Haminoea cymbalum 0,2031 0,1993 0,3173 0,2714 0,1508 0,0626 0,1715 0,2348 0,2813 0,4695 29 Smaragdinella sp, 0,1959 0,2012 0,3186 0,2586 0,1459 0,0577 0,1517 0,2349 0,2644 0,4880 30 Bulla gouldiana 0,1829 0,1983 0,3049 0,2359 0,1329 0,1248 0,1169 0,2489 0,2348 0,5285 31 Philine aperta 0,1923 0,1889 0,3008 0,2877 0,1295 0,1552 0,1604 0,1591 0,1586 0,4815 32 Scaphander punctostriatus 0,2025 0,2055 0,3134 0,2667 0,1529 0,1765 0,1724 0,1578 0,1186 0,5653 33 Elysia viridis 0,2291 0,2297 0,3072 0,2948 0,1832 0,2215 0,2245 0,2473 0,2227 0,2490 34 Thuridilla ratna 0,2341 0,2376 0,2927 0,3054 0,2014 0,2077 0,2062 0,2326 0,2313 0,2552 0,0834 35 Placida dendritica 0,2251 0,2386 0,3163 0,2760 0,2017 0,2139 0,2237 0,2397 0,2323 0,2400 0,0987 36 Acteon tornatilis 0,1735 0,1763 0,2965 0,2671 0,0130 0,1552 0,1485 0,1422 0,1334 0,1542 0,1784 37 Pupa solidula 0,2430 0,2168 0,3081 0,2918 0,2572 0,2688 0,2514 0,2505 0,2665 0,2765 0,2292 38 Hydatina physis 0,2145 0,2038 0,2967 0,2857 0,2169 0,2382 0,2290 0,2447 0,2484 0,2456 0,1755 39 Pontohedyle milatchevitchi 0,2867 0,2667 0,3216 0,3221 0,2887 0,2741 0,2753 0,2871 0,3034 0,3030 0,2348
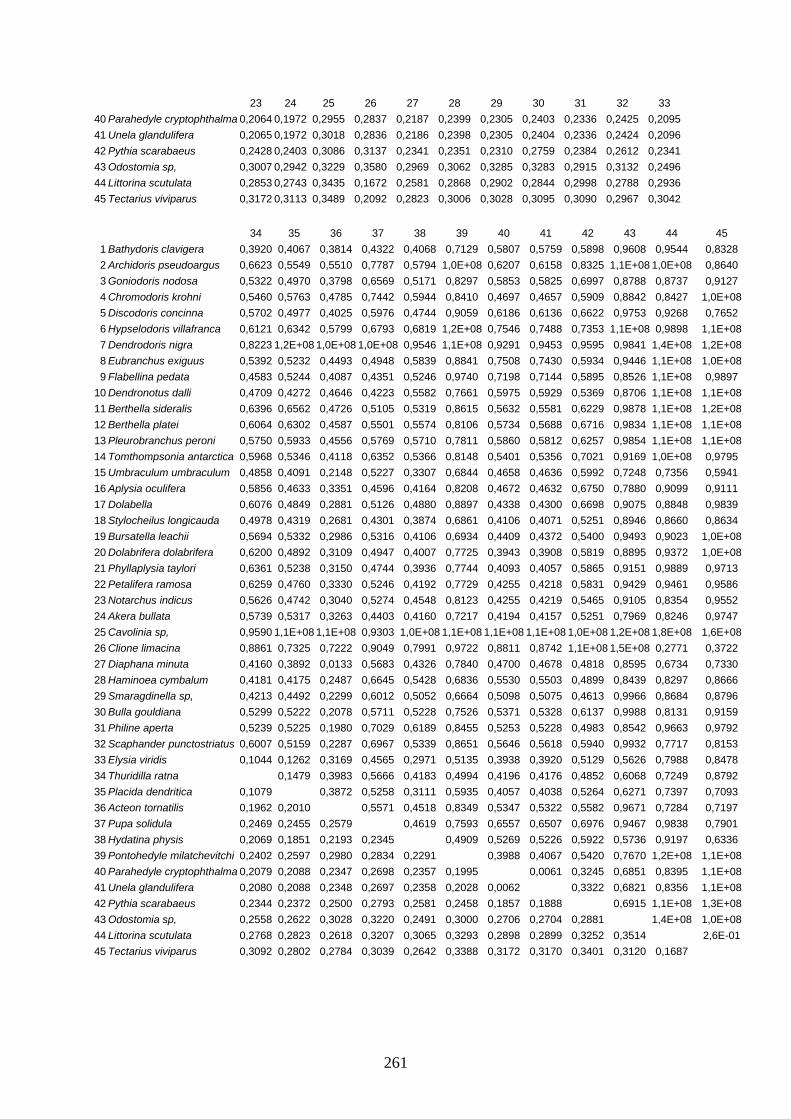
261
23 24 25 26 27 28 29 30 31 32 33 40 Parahedyle cryptophthalma 0,2064 0,1972 0,2955 0,2837 0,2187 0,2399 0,2305 0,2403 0,2336 0,2425 0,2095 41 Unela glandulifera 0,2065 0,1972 0,3018 0,2836 0,2186 0,2398 0,2305 0,2404 0,2336 0,2424 0,2096 42 Pythia scarabaeus 0,2428 0,2403 0,3086 0,3137 0,2341 0,2351 0,2310 0,2759 0,2384 0,2612 0,2341 43 Odostomia sp, 0,3007 0,2942 0,3229 0,3580 0,2969 0,3062 0,3285 0,3283 0,2915 0,3132 0,2496 44 Littorina scutulata 0,2853 0,2743 0,3435 0,1672 0,2581 0,2868 0,2902 0,2844 0,2998 0,2788 0,2936 45 Tectarius viviparus 0,3172 0,3113 0,3489 0,2092 0,2823 0,3006 0,3028 0,3095 0,3090 0,2967 0,3042
34 35 36 37 38 39 40 41 42 43 44 45
1 Bathydoris clavigera 0,3920 0,4067 0,3814 0,4322 0,4068 0,7129 0,5807 0,5759 0,5898 0,9608 0,9544 0,8328 2 Archidoris pseudoargus 0,6623 0,5549 0,5510 0,7787 0,5794 1,0E+08 0,6207 0,6158 0,8325 1,1E+08 1,0E+08 0,8640 3 Goniodoris nodosa 0,5322 0,4970 0,3798 0,6569 0,5171 0,8297 0,5853 0,5825 0,6997 0,8788 0,8737 0,9127 4 Chromodoris krohni 0,5460 0,5763 0,4785 0,7442 0,5944 0,8410 0,4697 0,4657 0,5909 0,8842 0,8427 1,0E+08 5 Discodoris concinna 0,5702 0,4977 0,4025 0,5976 0,4744 0,9059 0,6186 0,6136 0,6622 0,9753 0,9268 0,7652 6 Hypselodoris villafranca 0,6121 0,6342 0,5799 0,6793 0,6819 1,2E+08 0,7546 0,7488 0,7353 1,1E+08 0,9898 1,1E+08 7 Dendrodoris nigra 0,8223 1,2E+08 1,0E+08 1,0E+08 0,9546 1,1E+08 0,9291 0,9453 0,9595 0,9841 1,4E+08 1,2E+08 8 Eubranchus exiguus 0,5392 0,5232 0,4493 0,4948 0,5839 0,8841 0,7508 0,7430 0,5934 0,9446 1,1E+08 1,0E+08 9 Flabellina pedata 0,4583 0,5244 0,4087 0,4351 0,5246 0,9740 0,7198 0,7144 0,5895 0,8526 1,1E+08 0,9897
10 Dendronotus dalli 0,4709 0,4272 0,4646 0,4223 0,5582 0,7661 0,5975 0,5929 0,5369 0,8706 1,1E+08 1,1E+08 11 Berthella sideralis 0,6396 0,6562 0,4726 0,5105 0,5319 0,8615 0,5632 0,5581 0,6229 0,9878 1,1E+08 1,2E+08 12 Berthella platei 0,6064 0,6302 0,4587 0,5501 0,5574 0,8106 0,5734 0,5688 0,6716 0,9834 1,1E+08 1,1E+08 13 Pleurobranchus peroni 0,5750 0,5933 0,4556 0,5769 0,5710 0,7811 0,5860 0,5812 0,6257 0,9854 1,1E+08 1,1E+08 14 Tomthompsonia antarctica 0,5968 0,5346 0,4118 0,6352 0,5366 0,8148 0,5401 0,5356 0,7021 0,9169 1,0E+08 0,9795 15 Umbraculum umbraculum 0,4858 0,4091 0,2148 0,5227 0,3307 0,6844 0,4658 0,4636 0,5992 0,7248 0,7356 0,5941 16 Aplysia oculifera 0,5856 0,4633 0,3351 0,4596 0,4164 0,8208 0,4672 0,4632 0,6750 0,7880 0,9099 0,9111 17 Dolabella 0,6076 0,4849 0,2881 0,5126 0,4880 0,8897 0,4338 0,4300 0,6698 0,9075 0,8848 0,9839 18 Stylocheilus longicauda 0,4978 0,4319 0,2681 0,4301 0,3874 0,6861 0,4106 0,4071 0,5251 0,8946 0,8660 0,8634 19 Bursatella leachii 0,5694 0,5332 0,2986 0,5316 0,4106 0,6934 0,4409 0,4372 0,5400 0,9493 0,9023 1,0E+08 20 Dolabrifera dolabrifera 0,6200 0,4892 0,3109 0,4947 0,4007 0,7725 0,3943 0,3908 0,5819 0,8895 0,9372 1,0E+08 21 Phyllaplysia taylori 0,6361 0,5238 0,3150 0,4744 0,3936 0,7744 0,4093 0,4057 0,5865 0,9151 0,9889 0,9713 22 Petalifera ramosa 0,6259 0,4760 0,3330 0,5246 0,4192 0,7729 0,4255 0,4218 0,5831 0,9429 0,9461 0,9586 23 Notarchus indicus 0,5626 0,4742 0,3040 0,5274 0,4548 0,8123 0,4255 0,4219 0,5465 0,9105 0,8354 0,9552 24 Akera bullata 0,5739 0,5317 0,3263 0,4403 0,4160 0,7217 0,4194 0,4157 0,5251 0,7969 0,8246 0,9747 25 Cavolinia sp, 0,9590 1,1E+08 1,1E+08 0,9303 1,0E+08 1,1E+08 1,1E+08 1,1E+08 1,0E+08 1,2E+08 1,8E+08 1,6E+08 26 Clione limacina 0,8861 0,7325 0,7222 0,9049 0,7991 0,9722 0,8811 0,8742 1,1E+08 1,5E+08 0,2771 0,3722 27 Diaphana minuta 0,4160 0,3892 0,0133 0,5683 0,4326 0,7840 0,4700 0,4678 0,4818 0,8595 0,6734 0,7330 28 Haminoea cymbalum 0,4181 0,4175 0,2487 0,6645 0,5428 0,6836 0,5530 0,5503 0,4899 0,8439 0,8297 0,8666 29 Smaragdinella sp, 0,4213 0,4492 0,2299 0,6012 0,5052 0,6664 0,5098 0,5075 0,4613 0,9966 0,8684 0,8796 30 Bulla gouldiana 0,5299 0,5222 0,2078 0,5711 0,5228 0,7526 0,5371 0,5328 0,6137 0,9988 0,8131 0,9159 31 Philine aperta 0,5239 0,5225 0,1980 0,7029 0,6189 0,8455 0,5253 0,5228 0,4983 0,8542 0,9663 0,9792 32 Scaphander punctostriatus 0,6007 0,5159 0,2287 0,6967 0,5339 0,8651 0,5646 0,5618 0,5940 0,9932 0,7717 0,8153 33 Elysia viridis 0,1044 0,1262 0,3169 0,4565 0,2971 0,5135 0,3938 0,3920 0,5129 0,5626 0,7988 0,8478 34 Thuridilla ratna 0,1479 0,3983 0,5666 0,4183 0,4994 0,4196 0,4176 0,4852 0,6068 0,7249 0,8792 35 Placida dendritica 0,1079 0,3872 0,5258 0,3111 0,5935 0,4057 0,4038 0,5264 0,6271 0,7397 0,7093 36 Acteon tornatilis 0,1962 0,2010 0,5571 0,4518 0,8349 0,5347 0,5322 0,5582 0,9671 0,7284 0,7197 37 Pupa solidula 0,2469 0,2455 0,2579 0,4619 0,7593 0,6557 0,6507 0,6976 0,9467 0,9838 0,7901 38 Hydatina physis 0,2069 0,1851 0,2193 0,2345 0,4909 0,5269 0,5226 0,5922 0,5736 0,9197 0,6336 39 Pontohedyle milatchevitchi 0,2402 0,2597 0,2980 0,2834 0,2291 0,3988 0,4067 0,5420 0,7670 1,2E+08 1,1E+08 40 Parahedyle cryptophthalma 0,2079 0,2088 0,2347 0,2698 0,2357 0,1995 0,0061 0,3245 0,6851 0,8395 1,1E+08 41 Unela glandulifera 0,2080 0,2088 0,2348 0,2697 0,2358 0,2028 0,0062 0,3322 0,6821 0,8356 1,1E+08 42 Pythia scarabaeus 0,2344 0,2372 0,2500 0,2793 0,2581 0,2458 0,1857 0,1888 0,6915 1,1E+08 1,3E+08 43 Odostomia sp, 0,2558 0,2622 0,3028 0,3220 0,2491 0,3000 0,2706 0,2704 0,2881 1,4E+08 1,0E+08 44 Littorina scutulata 0,2768 0,2823 0,2618 0,3207 0,3065 0,3293 0,2898 0,2899 0,3252 0,3514 2,6E-01 45 Tectarius viviparus 0,3092 0,2802 0,2784 0,3039 0,2642 0,3388 0,3172 0,3170 0,3401 0,3120 0,1687

262
Opisthobranchia-Datensatz (28S rDNA) Dargestellt sind die beobachteten (unten) und korrigierten (oben) Distanzen der 28S rDNA-Sequenzen. Die Parameter des für die Korrektur der beobachteten Distanzen verwendeten Sequenzevolutionsmodells wurden mit Hilfe hierarchischer likelihood ratio tests (Modeltest, Posada & Crandall 1998) ermittelt.
1 2 3 4 5 6 7 8 9 10 11 1 Bathydoris clavigera 0,1744 0,1702 0,1886 0,6407 0,6244 0,5522 0,3897 0,4638 0,4129 0,8694 2 Dendrodoris fumata 0,1133 0,0686 0,0749 0,5031 0,5263 0,4758 0,3603 0,3790 0,3475 0,7622 3 Chromodoris krohni 0,1069 0,0543 0,0358 0,5219 0,5112 0,4773 0,3684 0,4137 0,3429 0,7385 4 Hypselodoris villafranca 0,1158 0,0583 0,0302 0,4839 0,5188 0,5087 0,3801 0,4109 0,3450 0,7528 5 Dendronotus dalli 0,2422 0,2112 0,2140 0,2074 0,2334 0,1867 0,6310 0,6859 0,6549 0,8637 6 Flabellina babai 0,2298 0,2117 0,2062 0,2058 0,1293 0,1395 0,5764 0,5907 0,5301 0,8453 7 Cuthona nana 0,2231 0,2076 0,2054 0,2102 0,1157 0,0946 0,5222 0,5488 0,5440 0,8056 8 Euselenops luniceps 0,1793 0,1701 0,1705 0,1732 0,2338 0,2200 0,2153 0,0733 0,0580 0,3886 9 Pleurobranchus peroni 0,1977 0,1764 0,1840 0,1824 0,2434 0,2282 0,2259 0,0577 0,0683 0,4584
10 Berthellina citrina 0,1838 0,1679 0,1639 0,1657 0,2369 0,2148 0,2205 0,0471 0,0549 0,4045 11 Berthella stellata 0,2603 0,2436 0,2378 0,2411 0,2653 0,2638 0,2635 0,1754 0,1945 0,1795 12 Bathyberthella antarctica 0,1895 0,1717 0,1752 0,1770 0,2374 0,2251 0,2225 0,0427 0,0574 0,0428 0,1795 13 Tompthompsonia antarctica 0,1815 0,1639 0,1640 0,1723 0,2461 0,2371 0,2344 0,0941 0,0987 0,0889 0,1896 14 Tylodina perversa 0,1833 0,1659 0,1717 0,1783 0,2731 0,2507 0,2523 0,1513 0,1640 0,1502 0,2276 15 Umbraculum mediterraneum 0,1929 0,1655 0,1702 0,1747 0,2638 0,2551 0,2458 0,1531 0,1660 0,1534 0,2381 16 Calliopaea bellula 0,2094 0,1912 0,1973 0,2009 0,2599 0,2628 0,2541 0,1837 0,1888 0,1768 0,2566 17 Limapontia nigra 0,1910 0,1702 0,1754 0,1822 0,2570 0,2483 0,2331 0,1625 0,1712 0,1580 0,2381 18 Thuridilla bayeri 0,1792 0,1658 0,1675 0,1743 0,2573 0,2385 0,2338 0,1614 0,1756 0,1588 0,2490 19 Bosellia mimetica 0,1803 0,1657 0,1642 0,1701 0,2537 0,2385 0,2412 0,1592 0,1734 0,1590 0,2414 20 Cyerce nigricans 0,1838 0,1682 0,1671 0,1725 0,2538 0,2360 0,2365 0,1634 0,1730 0,1583 0,2405 21 Elysia viridis 0,1973 0,1768 0,1755 0,1846 0,2575 0,2390 0,2431 0,1638 0,1700 0,1612 0,2523 22 Placobranchus ocellatus 0,1856 0,1701 0,1676 0,1733 0,2552 0,2295 0,2366 0,1644 0,1756 0,1588 0,2479 23 Haminoea hydatis 0,1875 0,1680 0,1760 0,1797 0,2710 0,2471 0,2424 0,1588 0,1669 0,1621 0,2451 24 Phanerophthalmus smaragdinus 0,1900 0,1650 0,1729 0,1819 0,2645 0,2460 0,2333 0,1552 0,1691 0,1612 0,2460 25 Atys semistriatus 0,1826 0,1607 0,1666 0,1703 0,2677 0,2423 0,2364 0,1509 0,1674 0,1546 0,2428 26 cf. Retusa sp. 0,1880 0,1763 0,1741 0,1822 0,2709 0,2541 0,2407 0,1643 0,1784 0,1642 0,2457 27 Philinopsis pilsbryi 0,1851 0,1631 0,1701 0,1791 0,2681 0,2506 0,2435 0,1675 0,1822 0,1718 0,2459 28 Chelidonura inornata 0,1905 0,1750 0,1796 0,1844 0,2705 0,2494 0,2468 0,1726 0,1821 0,1748 0,2501 29 Odontoglaja guamensis 0,1894 0,1728 0,1795 0,1854 0,2725 0,2548 0,2486 0,1712 0,1819 0,1746 0,2565 30 Navanax inermis 0,1916 0,1654 0,1689 0,1770 0,2688 0,2517 0,2430 0,1684 0,1824 0,1696 0,2441 31 Philinoglossa praelongata 0,1797 0,1593 0,1582 0,1695 0,2569 0,2467 0,2363 0,1608 0,1714 0,1611 0,2416 32 Bulla cf. striata 0,1954 0,1655 0,1626 0,1696 0,2465 0,2417 0,2372 0,1600 0,1753 0,1591 0,2405 33 Runcina adriatica 0,2033 0,1893 0,1945 0,1980 0,2844 0,2592 0,2535 0,1720 0,1843 0,1757 0,2508 34 Sagaminopteron psychedelicum 0,1929 0,1644 0,1688 0,1757 0,2644 0,2588 0,2469 0,1697 0,1750 0,1686 0,2477 35 Pupa solidula 0,1722 0,1539 0,1536 0,1540 0,2576 0,2494 0,2453 0,1677 0,1718 0,1652 0,2358 36 Hydatina physis 0,1763 0,1602 0,1578 0,1571 0,2614 0,2473 0,2464 0,1663 0,1724 0,1654 0,2380 37 Aplysia californica 0,1904 0,1615 0,1642 0,1687 0,2641 0,2393 0,2362 0,1620 0,1800 0,1651 0,2450 38 Dolabella auricularia 0,1809 0,1556 0,1637 0,1640 0,2547 0,2429 0,2383 0,1577 0,1746 0,1591 0,2336 39 Akera bullata 0,1908 0,1756 0,1737 0,1817 0,2639 0,2477 0,2353 0,1668 0,1818 0,1633 0,2443 40 Unela glandulifera 0,1868 0,1744 0,1762 0,1850 0,2694 0,2594 0,2472 0,1625 0,1777 0,1626 0,2424 41 Parahedyle cryptophthalma 0,1868 0,1744 0,1762 0,1850 0,2694 0,2594 0,2472 0,1625 0,1777 0,1626 0,2424 42 Pontohedyle milatchevitchi 0,1922 0,1766 0,1803 0,1839 0,2730 0,2552 0,2397 0,1658 0,1768 0,1597 0,2436 43 Hedylopsis spiculifera 0,1761 0,1680 0,1688 0,1745 0,2601 0,2464 0,2404 0,1493 0,1601 0,1518 0,2391 44 Siphonaria alternata 0,1837 0,1777 0,1731 0,1809 0,2680 0,2533 0,2324 0,1621 0,1740 0,1615 0,2327 45 Onchidella floridana 0,1840 0,1741 0,1787 0,1798 0,2732 0,2592 0,2463 0,1647 0,1818 0,1601 0,2422 46 Onchidium verrucosum 0,1810 0,1741 0,1718 0,1784 0,2693 0,2545 0,2454 0,1632 0,1777 0,1569 0,2514 47 Odostomia sp. 0,1811 0,1591 0,1630 0,1709 0,2650 0,2512 0,2379 0,1506 0,1719 0,1490 0,2275 48 Cepaea hortensis 0,2121 0,2013 0,1984 0,2057 0,2846 0,2668 0,2557 0,1803 0,1964 0,1760 0,2633 49 Lymnaea stagnalis 0,1873 0,1764 0,1780 0,1842 0,2587 0,2542 0,2362 0,1659 0,1796 0,1723 0,2405

263
1 2 3 4 5 6 7 8 9 10 11 50 Nodilittorina unifasciata 0,1815 0,1687 0,1796 0,1782 0,2638 0,2414 0,2381 0,1718 0,1810 0,1698 0,2410 51 Tectarius striatus 0,1811 0,1683 0,1760 0,1768 0,2542 0,2382 0,2390 0,1779 0,1892 0,1724 0,2430 52 Littorina littorea 0,1842 0,1653 0,1773 0,1792 0,2533 0,2427 0,2449 0,1703 0,1802 0,1691 0,2452 53 Littoraria undulata 0,1794 0,1666 0,1722 0,1740 0,2565 0,2428 0,2413 0,1771 0,1886 0,1771 0,2473 54 Melarhaphe neritoides 0,1899 0,1708 0,1774 0,1847 0,2551 0,2497 0,2416 0,1724 0,1806 0,1626 0,2439
12 13 14 15 16 17 18 19 20 21 22 1 Bathydoris clavigera 0,4333 0,4111 0,3838 0,4107 0,4694 0,4043 0,3680 0,3658 0,3765 0,4199 0,3857 2 Dendrodoris fumata 0,3636 0,3346 0,3231 0,3242 0,4073 0,3444 0,3307 0,3277 0,3364 0,3608 0,3399 3 Chromodoris krohni 0,3846 0,3425 0,3514 0,3501 0,4390 0,3630 0,3410 0,3305 0,3371 0,3654 0,3395 4 Hypselodoris villafranca 0,3857 0,3652 0,3701 0,3634 0,4436 0,3807 0,3625 0,3490 0,3560 0,3929 0,3593 5 Dendronotus dalli 0,6560 0,7133 0,8353 0,7686 0,7210 0,7218 0,7365 0,7211 0,7368 0,7221 0,7344 6 Flabellina babai 0,5907 0,6507 0,7080 0,7568 0,7476 0,6833 0,6496 0,6522 0,6404 0,6451 0,6058 7 Cuthona nana 0,5545 0,6176 0,7013 0,6717 0,6577 0,5672 0,6081 0,6326 0,6157 0,6364 0,6155 8 Euselenops luniceps 0,0505 0,1420 0,2871 0,2925 0,3745 0,3182 0,3222 0,3095 0,3276 0,3171 0,3330 9 Pleurobranchus peroni 0,0742 0,1507 0,3356 0,3425 0,4092 0,3512 0,3814 0,3663 0,3711 0,3505 0,3838
10 Berthellina citrina 0,0511 0,1294 0,2787 0,2932 0,3610 0,3058 0,3137 0,3058 0,3087 0,3048 0,3146 11 Berthella stellata 0,3966 0,4445 0,6376 0,6978 0,7827 0,7000 0,7667 0,7223 0,7253 0,7670 0,7730 12 Bathyberthella antarctica 0,1216 0,3251 0,3293 0,3916 0,3432 0,3592 0,3430 0,3518 0,3273 0,3656 13 Tompthompsonia antarctica 0,0842 0,2821 0,3046 0,3898 0,3316 0,3611 0,3390 0,3532 0,3309 0,3643 14 Tylodina perversa 0,1658 0,1507 0,0965 0,2194 0,1823 0,1721 0,1612 0,1701 0,2046 0,1725 15 Umbraculum mediterraneum 0,1653 0,1572 0,0722 0,2009 0,1723 0,1559 0,1460 0,1472 0,1741 0,1586 16 Calliopaea bellula 0,1857 0,1852 0,1344 0,1249 0,0741 0,1036 0,0914 0,1071 0,1193 0,1083 17 Limapontia nigra 0,1692 0,1681 0,1171 0,1122 0,0586 0,0646 0,0546 0,0705 0,0847 0,0698 18 Thuridilla bayeri 0,1723 0,1755 0,1116 0,1024 0,0780 0,0524 0,0241 0,0376 0,0591 0,0294 19 Bosellia mimetica 0,1692 0,1704 0,1065 0,0982 0,0707 0,0451 0,0219 0,0298 0,0493 0,0282 20 Cyerce nigricans 0,1692 0,1740 0,1099 0,0975 0,0791 0,0556 0,0324 0,0261 0,0693 0,0340 21 Elysia viridis 0,1662 0,1682 0,1265 0,1122 0,0862 0,0651 0,0490 0,0417 0,0556 0,0597 22 Placobranchus ocellatus 0,1722 0,1757 0,1108 0,1035 0,0801 0,0556 0,0260 0,0250 0,0293 0,0490 23 Haminoea hydatis 0,1683 0,1647 0,1160 0,1063 0,1356 0,1111 0,1047 0,0985 0,1061 0,1049 0,1078 24 Phanerophthalmus smaragdinus 0,1659 0,1612 0,1106 0,1020 0,1333 0,1109 0,1066 0,1025 0,1049 0,1099 0,1077 25 Atys semistriatus 0,1670 0,1573 0,1068 0,0948 0,1252 0,1038 0,0920 0,0899 0,0913 0,0996 0,0941 26 cf. Retusa sp. 0,1747 0,1666 0,1094 0,0976 0,1322 0,1086 0,1086 0,1003 0,1027 0,1078 0,1106 27 Philinopsis pilsbryi 0,1766 0,1590 0,1175 0,1086 0,1418 0,1132 0,1174 0,1121 0,1189 0,1206 0,1183 28 Chelidonura inornata 0,1831 0,1655 0,1149 0,1030 0,1439 0,1196 0,1141 0,1120 0,1187 0,1196 0,1129 29 Odontoglaja guamensis 0,1829 0,1656 0,1181 0,1060 0,1512 0,1269 0,1203 0,1193 0,1240 0,1248 0,1192 30 Navanax inermis 0,1768 0,1625 0,1203 0,1052 0,1420 0,1156 0,1078 0,1037 0,1136 0,1122 0,1109 31 Philinoglossa praelongata 0,1723 0,1568 0,1214 0,1126 0,1387 0,1132 0,1129 0,1077 0,1156 0,1069 0,1129 32 Bulla cf. striata 0,1695 0,1637 0,1130 0,1011 0,1401 0,1124 0,1080 0,1037 0,1050 0,1082 0,1069 33 Runcina adriatica 0,1822 0,1643 0,1368 0,1189 0,1476 0,1307 0,1378 0,1367 0,1333 0,1420 0,1376 34 Sagaminopteron psychedelicum 0,1765 0,1654 0,1195 0,1000 0,1293 0,1123 0,1079 0,1037 0,1041 0,1059 0,1089 35 Pupa solidula 0,1720 0,1618 0,1169 0,1122 0,1383 0,1139 0,1095 0,1063 0,1111 0,1234 0,1128 36 Hydatina physis 0,1681 0,1653 0,1241 0,1108 0,1342 0,1141 0,1119 0,1087 0,1113 0,1236 0,1172 37 Aplysia californica 0,1772 0,1703 0,1040 0,0866 0,1322 0,1070 0,0939 0,0972 0,1050 0,1163 0,1013 38 Dolabella auricularia 0,1695 0,1550 0,1086 0,0905 0,1297 0,1053 0,0998 0,1019 0,1066 0,1201 0,1062 39 Akera bullata 0,1725 0,1601 0,1087 0,0979 0,1361 0,1117 0,1074 0,1064 0,1143 0,1195 0,1095 40 Unela glandulifera 0,1730 0,1610 0,0932 0,0889 0,1150 0,0916 0,0905 0,0884 0,0972 0,1045 0,0968 41 Parahedyle cryptophthalma 0,1730 0,1610 0,0932 0,0889 0,1150 0,0916 0,0905 0,0884 0,0972 0,1045 0,0968 42 Pontohedyle milatchevitchi 0,1734 0,1614 0,1009 0,0996 0,1213 0,1033 0,0968 0,0926 0,1036 0,1076 0,0979 43 Hedylopsis spiculifera 0,1612 0,1512 0,0912 0,0848 0,1064 0,0844 0,0852 0,0809 0,0900 0,0928 0,0893 44 Siphonaria alternata 0,1750 0,1663 0,0796 0,0743 0,1087 0,0855 0,0779 0,0789 0,0824 0,0960 0,0842 45 Onchidella floridana 0,1740 0,1722 0,1044 0,0820 0,1163 0,0974 0,0888 0,0889 0,0903 0,1007 0,0963 46 Onchidium verrucosum 0,1742 0,1638 0,0928 0,0821 0,1187 0,0986 0,0919 0,0899 0,0945 0,0995 0,0909 47 Odostomia sp. 0,1617 0,1611 0,0957 0,0962 0,1313 0,1018 0,0951 0,0950 0,0975 0,1113 0,0971 48 Cepaea hortensis 0,1895 0,1922 0,1129 0,1161 0,1345 0,1111 0,1090 0,1068 0,1079 0,1132 0,1079 49 Lymnaea stagnalis 0,1780 0,1712 0,1064 0,0990 0,1220 0,0974 0,0985 0,0974 0,1028 0,1049 0,1016 50 Nodilittorina unifasciata 0,1766 0,1700 0,1521 0,1522 0,1651 0,1486 0,1440 0,1450 0,1431 0,1550 0,1462

264
12 13 14 15 16 17 18 19 20 21 22 51 Tectarius striatus 0,1817 0,1773 0,1507 0,1500 0,1659 0,1516 0,1437 0,1437 0,1417 0,1515 0,1437 52 Littorina littorea 0,1763 0,1710 0,1494 0,1540 0,1720 0,1557 0,1490 0,1457 0,1524 0,1556 0,1501 53 Littoraria undulata 0,1820 0,1710 0,1561 0,1542 0,1691 0,1484 0,1437 0,1426 0,1438 0,1536 0,1480 54 Melarhaphe neritoides 0,1764 0,1722 0,1529 0,1564 0,1626 0,1463 0,1427 0,1459 0,1408 0,1526 0,1439
Anaspidea-Datensatz (18S rDNA) Dargestellt sind die beobachteten (unten) und korrigierten (oben) Distanzen der 18S rDNA-Sequenzen. Die Parameter des für die Korrektur der beobachteten Distanzen verwendeten Sequenzevolutionsmodells wurden mit Hilfe hierarchischer likelihood ratio tests (Modeltest, Posada & Crandall 1998) ermittelt.
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
1 Akera bullata 0,0138 0,0138 0,0145 0,0145 0,0162 0,0131 0,0138 0,0159 0,0132 0,0153 0,0152 0,0167 0,0152 0,0798 0,0428 0,0474 0,0235 0,0406 0,0473
2 Aplysia gigantea 0,0123 0,0000 0,0006 0,0029 0,0059 0,0006 0,0000 0,0078 0,0078 0,0072 0,0072 0,0085 0,0072 0,0807 0,0355 0,0418 0,0211 0,0342 0,0417
3 Aplysia oculifera 0,0123 0,0000 0,0006 0,0029 0,0059 0,0006 0,0000 0,0078 0,0078 0,0072 0,0072 0,0085 0,0072 0,0807 0,0355 0,0418 0,0211 0,0342 0,0417
4 Aplysia sydneyensis 0,0129 0,0006 0,0006 0,0034 0,0053 0,0011 0,0006 0,0085 0,0084 0,0079 0,0079 0,0091 0,0079 0,0823 0,0365 0,0429 0,0220 0,0345 0,0419
5 Aplysia parvula 0,0129 0,0028 0,0028 0,0034 0,0072 0,0023 0,0029 0,0097 0,0090 0,0091 0,0091 0,0104 0,0091 0,0831 0,0357 0,0428 0,0219 0,0337 0,0435
6 Aplysia juliana 0,0140 0,0056 0,0056 0,0050 0,0067 0,0053 0,0059 0,0124 0,0139 0,0118 0,0118 0,0131 0,0118 0,0916 0,0432 0,0500 0,0269 0,0380 0,0457
7 Aplysia dactylomela 0,0118 0,0006 0,0006 0,0011 0,0022 0,0050 0,0006 0,0084 0,0084 0,0079 0,0078 0,0091 0,0078 0,0817 0,0363 0,0426 0,0218 0,0350 0,0425
8 Siphonota geographica 0,0123 0,0000 0,0000 0,0006 0,0028 0,0056 0,0006 0,0078 0,0078 0,0072 0,0072 0,0085 0,0072 0,0797 0,0355 0,0418 0,0211 0,0343 0,0417
9 Bursatella leachii 0,0141 0,0073 0,0073 0,0079 0,0090 0,0112 0,0079 0,0073 0,0084 0,0041 0,0041 0,0053 0,0041 0,0793 0,0324 0,0368 0,0245 0,0326 0,0342
10 Dolabella auricularia 0,0118 0,0073 0,0073 0,0079 0,0084 0,0123 0,0079 0,0073 0,0078 0,0065 0,0053 0,0065 0,0053 0,0773 0,0350 0,0421 0,0239 0,0353 0,0412
11 Petalifera petalifera 0,0135 0,0067 0,0067 0,0073 0,0084 0,0106 0,0073 0,0067 0,0039 0,0062 0,0011 0,0023 0,0011 0,0768 0,0315 0,0370 0,0222 0,0298 0,0347
12 Dolabrifera brazieri 0,0135 0,0067 0,0067 0,0073 0,0084 0,0106 0,0073 0,0067 0,0039 0,0050 0,0011 0,0023 0,0000 0,0764 0,0332 0,0368 0,0216 0,0307 0,0357
13 Stylocheilus longicauda 0,0146 0,0079 0,0079 0,0084 0,0095 0,0118 0,0084 0,0079 0,0050 0,0062 0,0022 0,0022 0,0023 0,0789 0,0312 0,0385 0,0222 0,0331 0,0362
14 Dolabrifera dolabrifera 0,0135 0,0067 0,0067 0,0073 0,0084 0,0106 0,0073 0,0067 0,0039 0,0050 0,0011 0,0000 0,0022 0,0764 0,0332 0,0368 0,0223 0,0315 0,0364
15 Creseis acicula 0,0514 0,0519 0,0519 0,0525 0,0530 0,0558 0,0525 0,0514 0,0513 0,0507 0,0501 0,0502 0,0513 0,0501 0,0753 0,0932 0,0975 0,1026 0,1105
16 Clio pyramidata 0,0321 0,0276 0,0276 0,0281 0,0276 0,0321 0,0281 0,0276 0,0259 0,0276 0,0253 0,0265 0,0253 0,0265 0,0485 0,0421 0,0494 0,0486 0,0583
17 Diacria quadridentata 0,0348 0,0314 0,0314 0,0320 0,0320 0,0359 0,0320 0,0314 0,0287 0,0320 0,0287 0,0287 0,0298 0,0287 0,0553 0,0314 0,0496 0,0574 0,0560
18 Spongiobranchaea australis 0,0197 0,0179 0,0179 0,0185 0,0185 0,0218 0,0185 0,0179 0,0202 0,0196 0,0185 0,0180 0,0185 0,0185 0,0576 0,0355 0,0353 0,0409 0,0483
19 Atys semistriatus 0,0315 0,0276 0,0276 0,0276 0,0270 0,0298 0,0281 0,0276 0,0264 0,0287 0,0248 0,0253 0,0270 0,0259 0,0616 0,0356 0,0405 0,0314 0,0199
20 Chelidonura inornata 0,0353 0,0325 0,0325 0,0326 0,0336 0,0347 0,0331 0,0325 0,0275 0,0325 0,0280 0,0286 0,0292 0,0292 0,0638 0,0421 0,0392 0,0358 0,0174
Anaspidea-Datensatz (ITS1-5.8S-IST2) Dargestellt sind die beobachteten (unten) und korrigierten (oben) Distanzen der ITS1-5.8S-ITS2 - Sequenzen. Die Parameter des für die Korrektur der beobachteten Distanzen verwendeten Sequenzevolutionsmodells wurden mit Hilfe hierarchischer likelihood ratio tests (Modeltest, Posada & Crandall 1998) ermittelt.
1 2 3 4 5 6 7 8 9 10 11
1 Bursatella leachii 0,2643 0,2839 0,3349 0,3296 0,3828 0,3529 0,4636 0,5230 0,6002 0,6113
2 Petalifera petalifera 0,1702 0,1011 0,1179 0,1769 0,2765 0,2356 0,3142 0,3638 0,4893 0,5445
3 Dolabrifera brazieri 0,1791 0,0829 0,0245 0,1720 0,2256 0,2072 0,2741 0,3274 0,4442 0,4500
4 Dolabrifera dolabrifera 0,1972 0,0934 0,0232 0,1687 0,2451 0,2169 0,2984 0,3400 0,4643 0,4689
5 Dolabella auricularia 0,1955 0,1285 0,1265 0,1258 0,1706 0,1689 0,2638 0,2976 0,4248 0,4450
6 Aplysia juliana 0,2102 0,1743 0,1530 0,1626 0,1259 0,1055 0,1473 0,2575 0,4271 0,4332
7 Aplysia gigantea 0,2012 0,1581 0,1461 0,1516 0,1236 0,0844 0,0625 0,2307 0,3740 0,4188
8 Aplysia oculifera 0,2382 0,1934 0,1775 0,1890 0,1712 0,1112 0,0545 0,2417 0,3962 0,4671
9 Akera bullata 0,2569 0,2104 0,1959 0,2022 0,1881 0,1696 0,1593 0,1648 0,3023 0,3570
10 Atys semistriatus 0,2780 0,2511 0,2349 0,2423 0,2320 0,2302 0,2125 0,2211 0,1868 0,2815
11 Chelidonura inornata 0,2814 0,2673 0,2374 0,2454 0,2385 0,2325 0,2291 0,2469 0,2096 0,1794

265
Aeolidoidea-Datensatz (18S rDNA) Dargestellt sind die beobachteten (unten) und korrigierten (oben) Distanzen der ITS1-5.8S-ITS2 - Sequenzen. Die Parameter des für die Korrektur der beobachteten Distanzen verwendeten Sequenzevolutionsmodells wurden mit Hilfe hierarchischer likelihood ratio tests (Modeltest, Posada & Crandall 1998) ermittelt.
1 2 3 4 5 6 7 8 9 10 11 12 13
1 Baeolidia nodosa 0,0509 0,0541 0,0498 0,0462 0,0462 0,0437 0,0443 0,0437 0,0458 0,0490 0,0462 3,53E+09
2 Godiva banyulensis 0,0355 0,0315 0,0371 0,0365 0,0365 0,0336 0,0350 0,0344 0,0363 0,0390 0,0359 3,53E+09
3 Cratena peregrina 0,0382 0,0252 0,0180 0,0163 0,0163 0,0250 0,0248 0,0263 0,0265 0,0280 0,0241 3,53E+09
4 Phidiana lynceus 0,0354 0,0284 0,0158 0,0146 0,0146 0,0200 0,0199 0,0200 0,0209 0,0228 0,0250 3,53E+09
5 Facelina bostoniensis 0,0334 0,0280 0,0142 0,0131 0,0000 0,0209 0,0195 0,0209 0,0217 0,0216 0,0266 3,53E+09
6 Facelina auriculata 0,0334 0,0280 0,0142 0,0131 0,0000 0,0209 0,0195 0,0209 0,0217 0,0216 0,0266 3,53E+09
7 Phyllodesmium briareum 0,0322 0,0263 0,0213 0,0174 0,0180 0,0180 0,0034 0,0011 0,0023 0,0041 0,0094 3,53E+09
8 Phyllodesmium crypticum 0,0327 0,0274 0,0213 0,0174 0,0170 0,0170 0,0033 0,0023 0,0034 0,0023 0,0094 3,53E+09
9 Phyllodesmium colemani 0,0322 0,0268 0,0224 0,0174 0,0180 0,0180 0,0011 0,0022 0,0011 0,0029 0,0082 3,53E+09
10 Phyllodesmium longicirrum 0,0333 0,0279 0,0224 0,0180 0,0186 0,0186 0,0022 0,0033 0,0011 0,0041 0,0095 3,53E+09
11 Phyllodesmium jakobsenae 0,0352 0,0297 0,0235 0,0195 0,0185 0,0185 0,0039 0,0022 0,0028 0,0039 0,0109 3,53E+09
12 Phyllodesmium cf. magnum 0,0339 0,0280 0,0208 0,0213 0,0225 0,0225 0,0087 0,0087 0,0076 0,0087 0,0100 3,53E+09
13 Embletonia sp. 0,2351 0,2278 0,2270 0,2291 0,2282 0,2282 0,2256 0,2258 0,2262 0,2257 0,2293 0,2243
14 Cuthona nana 0,2361 0,2326 0,2303 0,2312 0,2317 0,2317 0,2307 0,2292 0,2301 0,2297 0,2327 0,2295 0,0501
15 Cuthona caerulea 0,2372 0,2327 0,2310 0,2308 0,2312 0,2312 0,2308 0,2293 0,2302 0,2298 0,2328 0,2302 0,0487
16 Cuthona sibogae 0,2375 0,2357 0,2322 0,2320 0,2330 0,2330 0,2326 0,2311 0,2321 0,2316 0,2347 0,2314 0,0491
17 Eubranchus exiguus 0,2394 0,2338 0,2350 0,2325 0,2341 0,2341 0,2319 0,2304 0,2314 0,2303 0,2339 0,2336 0,0535
18 Eubranchus sp. 0,2414 0,2368 0,2356 0,2331 0,2341 0,2341 0,2337 0,2322 0,2332 0,2327 0,2358 0,2337 0,0518
19 Tergipes tergipes 0,2492 0,2423 0,2402 0,2400 0,2392 0,2392 0,2399 0,2384 0,2394 0,2389 0,2422 0,2400 0,0624
20 Flabellina babai 0,2380 0,2303 0,2310 0,2291 0,2307 0,2307 0,2298 0,2282 0,2292 0,2287 0,2317 0,2291 0,0378
21 Flabellina pedata 0,2415 0,2399 0,2362 0,2371 0,2381 0,2381 0,2365 0,2349 0,2359 0,2360 0,2386 0,2353 0,0515
22 Flabellina verrucosa 0,2348 0,2287 0,2282 0,2306 0,2303 0,2303 0,2284 0,2275 0,2284 0,2291 0,2309 0,2277 0,0484
23 Flabellinaspec 0,2489 0,2393 0,2385 0,2388 0,2387 0,2387 0,2362 0,2359 0,2369 0,2358 0,2396 0,2396 0,0951
24 Flabellina affinis 0,2540 0,2478 0,2504 0,2492 0,2494 0,2494 0,2491 0,2473 0,2479 0,2480 0,2512 0,2495 0,0804
25 Notaeolidia depressa 0,2336 0,2294 0,2295 0,2269 0,2285 0,2285 0,2281 0,2266 0,2276 0,2283 0,2301 0,2286 0,0372
14 15 16 17 18 19 20 21 22 23 24 25
1 Baeolidia nodosa 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09
2 Godiva banyulensis 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09
3 Cratena peregrina 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09
4 Phidiana lynceus 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09
5 Facelina bostoniensis 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09
6 Facelina auriculata 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09
7 Phyllodesmium briareum 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09
8 Phyllodesmium crypticum 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09
9 Phyllodesmium colemani 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09
10 Phyllodesmium longicirrum 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09
11 Phyllodesmium jakobsenae 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09
12 Phyllodesmium cf. magnum 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09 3,53E+09
13 Embletonia sp. 0,0754 0,0720 0,0718 0,0858 0,0848 0,1067 0,0516 0,0787 0,0710 0,2293 0,1623 0,0510
14 Cuthona nana 0,0141 0,0139 0,0346 0,0430 0,0511 0,0439 0,0353 0,0722 0,2115 0,1427 0,0593
15 Cuthona caerulea 0,0126 0,0165 0,0350 0,0419 0,0494 0,0410 0,0403 0,0703 0,2253 0,1462 0,0622

266
14 15 16 17 18 19 20 21 22 23 24 25
16 Cuthona sibogae 0,0125 0,0148 0,0340 0,0385 0,0554 0,0416 0,0389 0,0722 0,2094 0,1445 0,0566
17 Eubranchus exiguus 0,0279 0,0285 0,0279 0,0390 0,0645 0,0475 0,0483 0,0787 0,2034 0,1530 0,0569
18 Eubranchus sp. 0,0322 0,0322 0,0300 0,0301 0,0619 0,0506 0,0532 0,0836 0,2330 0,1528 0,0596
19 Tergipes tergipes 0,0377 0,0372 0,0405 0,0449 0,0426 0,0747 0,0722 0,1100 0,2593 0,1585 0,0915
20 Flabellina babai 0,0338 0,0324 0,0327 0,0362 0,0366 0,0493 0,0442 0,0621 0,1959 0,1168 0,0350
21 Flabellina pedata 0,0276 0,0315 0,0304 0,0360 0,0376 0,0485 0,0344 0,0787 0,2322 0,1402 0,0521
22 Flabellina verrucosa 0,0472 0,0474 0,0484 0,0501 0,0516 0,0639 0,0435 0,0500 0,1585 0,1176 0,0713
23 Flabellinaspec 0,0904 0,0945 0,0904 0,0883 0,0925 0,0994 0,0871 0,0933 0,0784 0,2167 0,1913
24 Flabellina affinis 0,0762 0,0767 0,0779 0,0798 0,0791 0,0794 0,0664 0,0746 0,0676 0,0936 0,1385
25 Notaeolidia depressa 0,0418 0,0442 0,0407 0,0403 0,0402 0,0564 0,0280 0,0381 0,0482 0,0871 0,0754
V Chromosomenzahlen der Heterobranchia Taxon Anzahl
Chromosomen im haploiden
Satz
Quelle
Opisthobranchia Nudibranchia 13 Inaba & Hirota 1958, Burch & Natarajan 1967,
Inaba & Saiki 1967 Pleurobranchoidea 12 Inaba 1959 Tylodinoidea - Sacoglossa 17 Mancino & Sordi 1964 Anaspidea 16-17 Inaba 1959, Burch & Natarajan 1967, Vitturi et
al. 1982 Cephalaspidea s.str. 17-18 Inaba 1959, Burch & Natarajan 1967 Acteonoidea 15 Natarajan 1970 Acochlidiacea - Thecosomata 10-18 Zarnick 1911, Thiriot-Quiévreux 1988 Gymnosomata 16 Thiriot-Quiévreux 1988 Pulmonata Onchidiidae 17-18 Inaba 1961, Natarajan 1959, Burch & Natarajan
1967 Siphonariidae 16 Natarajan & Burch 1966, Choudhury et al. 1992 Ellobiidae 17-19 Natarajan 1958, Choudhury et al. 1992 Lymnaeidae 16-18 Burch 1960 Lymnaea stagnalis 18 Burch 1965 Helicidae 22-30 Perrot 1938, Rainer 1967 Cepaea hortensis 22 Perrot 1938 Allogastropoda Pyramidellidae 17 Inaba (pers. Mitteilung, aus Patterson 1969) “Prosobranchia” Littorinidae 15-17 Nishikawa 1962





























