Embed Size (px)
Citation preview

of July 13, 2018.This information is current as
ModelTolerance in a Kidney Transplantation Relationship Between Chimerism and
and David H. SachsAkihiko Yasumoto, Tokihiko Sawada, Christene H. Huang Yasushi Fuchimoto, Kazuhiko Yamada, Akira Shimizu,
http://www.jimmunol.org/content/162/10/57041999; 162:5704-5711; ;J Immunol
Referenceshttp://www.jimmunol.org/content/162/10/5704.full#ref-list-1
, 7 of which you can access for free at: cites 37 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 1999 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Relationship Between Chimerism and Tolerance in a KidneyTransplantation Model1
Yasushi Fuchimoto,2 Kazuhiko Yamada, Akira Shimizu, Akihiko Yasumoto,3
Tokihiko Sawada,4 Christene H. Huang, and David H. Sachs5
The persistence of donor leukocytes in recipients of organ allografts has been associated with long-term graft acceptance. However,it remains unclear whether this peripheral donor cell microchimerism plays an active role in graft acceptance or is simply aconsequence of the maintenance of sufficient immunosuppression to avoid rejection. A model of kidney transplantation betweenswine leukocyte Ag (SLA)-matched miniature swine, in which tolerance can be established with or without immunosuppressivetreatment, has been used to study the correlation between donor leukocyte chimerism and kidney graft acceptance. SLA-identicalkidney transplants were performed from animals positive for an allelic pig leukocyte Ag to animals negative for this marker.SLA-identical kidney transplant recipients given a 12-day course of cyclosporine (CyA) (n 5 3) became tolerant, showing stableserum creatinine levels (1–2 mg/dl) after cessation of CyA treatment. Donor cell chimerism (0.2–0.7%) was present by FACS inall three animals with peak levels detected at 3 wk. Two control animals receiving SLA-identical kidney grafts without CyA alsoshowed stable serum creatinine levels and became tolerant. However, in neither of these animals could donor leukocytes bedetected in the peripheral blood beyond 1 wk following transplantation. In one additional control animal, ureteral obstructionoccurred at day 10, and was associated with additional peripheral chimerism, presumably related to inflammation rather than toimmune status. These results indicate that the persistence of donor cell chimerism is not a requirement for the maintenance oftolerance to organ allografts in this model. The Journal of Immunology,1999, 162: 5704–5711.
T ransplantation tolerance, the long-term acceptance ofgrafted tissue in the absence of continuous immunosup-pression, remains an important goal in human transplan-
tation. Proposed mechanisms influencing tolerance include periph-eral anergy or clonal deletion of donor-reactive cytotoxic T cellsand suppression of alloreactive clonotypes. With the reports inrecent years of small percentages of donor-derived dendritic orhematopoietic cells detected in various tissues in long-term kidney(1), liver (2–4), and heart recipients (4), a novel theory to elucidategraft tolerance has been proposed. The theory contends that long-lived donor-derived cells play an important role in generatinglong-term graft acceptance (5, 6). However, consensus has notemerged regarding the causal relationship between such chimerismand tolerance.
Current limitations in clinical transplantation technology neces-sitate the use of continuous posttransplant immunosuppression inallograft recipients to prevent graft rejection. Thus, the persistenceof donor hematopoietic cells in recipient tissues may be caused by
the inability of an immunosuppressed host to effectively eliminatethese cells. Indeed, rather than inducing tolerance itself, donor chi-merism may be an epiphenomenon accompanying immunosup-pression-induced graft acceptance.
Using the partially inbred miniature swine developed in thislaboratory as a large animal model of organ transplantation (7), wehave studied the possible role of chimerism in transplantation ac-ceptance and the factors that influence this chimerism. Earlier stud-ies in this laboratory have demonstrated that tolerance can be es-tablished in swine leukocyte Ag (SLA)6-matched kidney grafts,with or without immunosuppression (8, 9). This model has allowedus to assess the potential role of chimerism in graft acceptance,independent of immunosuppression. Using an allelic nonhistocom-patibility marker, pig allelic Ag (PAA), previously described inthis laboratory,7 we have been able to identify the presence ofchimeric cells in kidney graft recipients.
We performed SLA-identical kidney transplants using donor an-imals positive for PAA and recipient animals negative for thismarker, with and without immunosuppression. We also performedan SLA-mismatched kidney transplant with immunosuppression.We evaluated the relationship between donor chimerism, immu-nologic response, and graft acceptance in these animals.
Materials and MethodsAnimals
Transplant donors and recipients were selected from our herd of partiallyinbred miniature swine at 5–7 mo of age. The immunogenetic character-istics of this herd and intra-MHC recombinant haplotypes have previouslybeen described (7, 10).
Transplantation Biology Research Center, Massachusetts General Hospital/HarvardMedical School, Boston, MA 02129
Received for publication November 5, 1998. Accepted for publication February17, 1999.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was sponsored in part by National Institutes of Health Grants 5RO1AI37692-03, 5PO1 AI39755-03, and by a sponsored research agreement betweenMassachusetts General Hospital and BioTransplant, Inc.2 Y.F. received partial support from Uehara Memorial Foundation.3 Current address: Department of Surgery, Chiba University School of Medicine,Chiba-city, Japan.4 Current address: Kidney Center, Tokyo Women’s Medical College, Tokyo, Japan.5 Address correspondence and reprint requests to Dr. David H. Sachs, TransplantationBiology Research Center, Massachusetts General Hospital, MGH-East, Building 149-9019, 13th Street, Boston, MA 02129. E-mail address: [email protected]
6 Abbreviations used in this paper: CyA, cyclosporine; GIL, graft infiltrating leuko-cytes; PAA, pig allelic Ag; SLA, swine leukocyte Ag.7 Y. Fuchimoto, C. Huang, A. Shimizu, J. Seebach, S. Arn, and D. H. Sachs. Anallelic non-histocompatibility antigen with wide tissue distribution as a marker forchimerism in pigs.Submitted for publication.
Copyright © 1999 by The American Association of Immunologists 0022-1767/99/$02.00
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Surgery
The details of the surgical procedures have been previously described (8,11). Placement of an indwelling central venous silastic catheter into anexternal jugular vein facilitated cyclosporine (CyA) administration and fre-quent blood sampling for monitoring of renal function (blood urea nitro-gen, serum creatinine) and whole blood CyA levels.
Immunosuppression
An i.v. preparation of CyA was generously provided by Novartis Phar-maceuticals (Hanover, NJ). CyA was given each morning as a singledaily infusion at a dose of 10–13 mg/kg (adjusted according to blood
levels) for 12 consecutive days posttransplant. The first dose was admin-istered preoperatively, before the unclamping of vessels to the kidneyallograft.
CyA levels
Whole blood trough levels were determined by a monoclonal radioimmu-noassay technique. Daily trough levels between 500 and 800 ng/ml wereachieved by adjusting the dose between 10 and 13 mg/kg, since these bloodlevels have previously been shown to be effective for induction of toleranceto renal allografts across selective SLA class I and minor Ag disparities(12–14).
FIGURE 1. Clinical course of recipients of an SLA-matched kidney allograft: with CyA (A) and without CyA (B).
FIGURE 2. Chimerism in peripheral blood in re-cipients of SLA-identical kidney allografts with CyAtreatment (A), and without CyA treatment (B), shownas percent donor cells in PBMC. Percent donor cellswas defined as: 1003 (PAA-positive PBMCs/totalPBMCs) by FACS.
5705The Journal of Immunology
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Histology
Sequential wedge kidney biopsies were performed on postoperative days 8,11, 18, 30, and 60 through a flank incision. Tissues were stained usinghematoxylin and eosin and periodic acid-Schiff, and coded slides wereexamined.
Isolation of PBMC
Freshly drawn, heparinized whole blood was diluted with HBSS (LifeTechnologies/BRL, Grand Island, NY), and mononuclear cells were ob-tained by gradient centrifugation using lymphocyte separation medium(Organon Teknika, Durham, NC). The mononuclear cells were washedonce with HBSS, and contaminating red cells were lysed with ACK buffer(B&B Research Laboratories, Fiskeville, RI). Cells were washed a secondtime with HBSS and resuspended in tissue culture cell-mediated lympholy-sis medium. Cell suspensions were kept at 4°C until used in cellular assays.
Preparation of renal cell suspensions
Kidney biopsies (100–500 mg) were minced and then suspended in HBSSbuffer, using the flat end of a syringe plunger. The cell suspension was thenfiltered on a coarse mesh and centrifuged at 1600 rpm for 10 min at 4°C.Cells were resuspended in 1 ml of flow cytometry analysis medium (HBSSbuffer, 0.1% of BSA, and 0.1% sodium azide).
Identification of graft-infiltrating leukocytes (GIL) within renalcell suspensions
Purification of GIL from small wedge biopsies of kidney has been difficultto achieve because GIL represent,3% of total renal cells at any given timepoint. Selective gating of these cells was therefore performed as previouslydescribed (15).
Flow cytometry as a measure of chimerism
To determine the level of peripheral donor cell chimerism, 13 106 PBMCwere incubated with FITC-conjugated 1038H-10-9 (IgM mAb) specific forswine PAA. FITC-conjugated 12-2-2 was used, as an IgM isotype-matchedcontrol. Staining was performed using FACS medium consisting of HBSSwith Ca11, 0.1% BSA, and 0.1% NaN3. Flow cytometry was performedusing a Becton Dickinson (Sunnyvale, CA) FACScan II. Flow cytometrydata were analyzed using Winlist mode software (Verity Software House,Topsham, ME).
For two-color analysis, incubation with biotinylated mouse mAbs spe-cific for porcine CD2 (MSA4), CD3 (2-6-15), CD4 (74-12-4), or CD8(76-2-11) (16–18) were added, followed by staining with PE-conjugatedstreptavidin (Becton Dickinson). Because the levels of chimerism in theperipheral blood were low, we used a statistical analysis to assure thepresence of chimeric cells. The mean percent chimerism among all fourtwo-color tubes from experimental animals was calculated and comparedwith the mean percent background staining for the two-color tubes of naivePAA-negative animals. The statistical significance of the mean donor cellchimerism for each experimental animal was determined relative to themean background staining of the naive PAA-negative control animals. Thebiotinylated anti-mouse-class I Ab 36.7.5 (IgG2a) served as a negativeisotype-matched control. Propidium iodide (PI) was added immediatelybefore acquisition, and dead cells were excluded from analysis based on PIstaining.
ResultsEffects of CyA on renal function and renal allograft survival
Three SLA-matched, PAA-mismatched allografts were performedwith CyA treatment, and three were performed without CyA treat-ment. Serum creatinine levels were measured to assess kidneyfunction. Results are shown in Fig. 1. All three animals with CyAtreatment maintained stable serum creatinine levels (1–2 mg/dl),even after cessation of CyA treatment, and two of three animalswithout CyA treatment maintained stable serum creatinine levels(1–2 mg/dl). One animal showed a transient increase in creatininedue to ureteral obstruction (day 10), but levels rapidly returned tonormal after relieving the obstruction, and the kidney allograft wasaccepted long-term (shown below). All three animals survivedlong-term (.2 yr) with normal renal function and normal kidneyhistology. Thus, normal kidney function and long-term graft ac-ceptance were established with or without transient CyA immu-nosuppression.
Effects of CyA on donor cell chimerism in peripheral blood andgraft
Peripheral chimerism.Two-color FACS analysis of PBMC wasperformed at periodic intervals posttransplant to evaluate donorcell chimerism in peripheral blood. Donor chimerism (0.2–0.7%,p , 0.005 relative to naive controls) was detected in all threeanimals with CyA treatment, with peak levels around 3–4 wk anda gradual decrease thereafter. Chimerism disappeared by day40–60 (Fig. 2A), except in one animal (no. 11870, shown later). Intwo of the three animals without CyA treatment, donor chimerismcould not be detected in the peripheral blood beyond 1 wk follow-ing transplantation (Fig. 2B).Intragraft chimerism. In all three animals with CyA treatment,donor chimerism (20–30%) was present in the graft at 2–3 wk and,thereafter, gradually disappeared (Fig. 3A). In two of the threeanimals without CyA treatment, donor chimerism could not bedetected in the graft beyond 2 wk following transplantation(Fig. 3B).Representative animals.Peripheral and intragraft chimerism inrepresentative animals is shown in Fig. 4. On day 18 after kidneytransplantation, CyA-treated animal no. 11784 showed 0.51%( p , 0.005) donor chimerism in peripheral blood (Fig. 4A) and20% chimerism in graft (Fig. 4C). On the other hand, CyA-un-treated animal no. 11892 showed no significant chimerism in pe-ripheral blood (Fig. 4B) and only 0.4% chimerism in graft (Fig.4D). Thus, there was a positive correlation between CyA treatmentand transient donor chimerism in both PBMC and graft.
Histology
Serial kidney biopsies were performed on days 18, 30, and 60. Athistologic examination, all three CyA-treated pigs showed a min-imal focal mononuclear interstitial infiltration, without tubulitisand endothelialitis on days 18 and 60 (Fig. 5,A andC). In com-parison, despite the stability of graft function, CyA-untreated pigsshowed a patchy and mild mononuclear cell infiltrate with focaltubulitis on day 18 (Fig. 5B). The mononuclear cell infiltratedecreased spontaneously and remained only minimal on day 60(Fig. 5D).
Effect of ureteral obstruction
One CyA-untreated animal showed a transient increase of creati-nine around day 10 due to ureteral obstruction. PBMC-chimerism
FIGURE 3. Chimerism in graft in recipients of SLA-identical kidneyallografts with CyA treatment (A), and without CyA treatment (B), shownas percent donor cells in the graft.
5706 CHIMERISM AND TOLERANCE IN SWINE
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
in this animal rose simultaneously with the creatinine increase,and, likewise, dropped rapidly with the creatinine decrease uponrelieving the ureteral obstruction. The chimerism gradually disap-peared, with a peak level at around 1.8% at day 11 (Fig. 6,A andB). This level was around four times the average of the CyA-treated model.
Class I-mismatched, PAA-mismatched kidney graft
Unlike SLA-matched grafts, which are accepted with or withoutCyA treatment, class I-mismatched grafts require immunosuppres-sion for 12 days posttransplant to induce tolerance (12). In thisexperiment, one animal received a class I-mismatched renal
FIGURE 4. Chimerism in peripheral blood andgraft in representative CyA-treated (A andC) anduntreated (Band D) animals that received SLA-identical kidney allografts on day 18 is shown byFACS analysis. Thex-axis shows PAA (1038H-10-9) staining;y-axis shows pig CD2 (MSA4)staining.
FIGURE 5. Renal biopsies taken on day 18 and day 60 from representative CyA-treated (Aand C) and untreated (Band D) animals that receivedSLA-identical kidney allografts (hematoxylin and eosin).
5707The Journal of Immunology
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
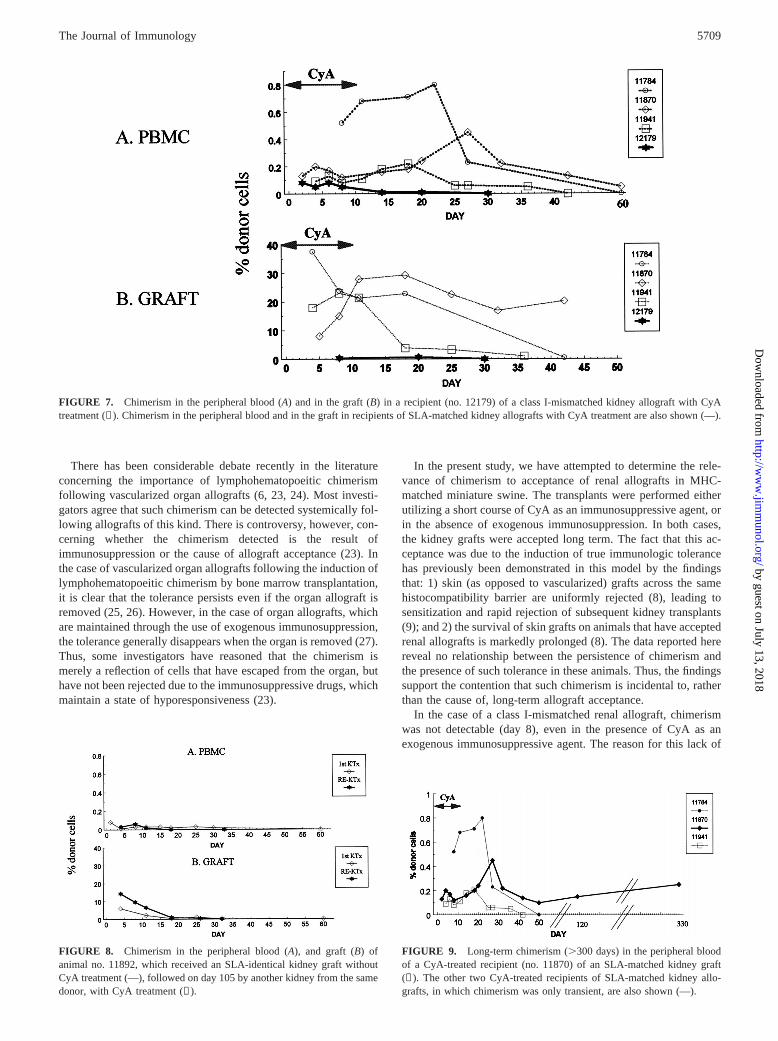
allograft with CyA treatment. Despite the use of immunosuppres-sion, donor chimerism could not be detected in the peripheralblood and in the graft beyond day 10 after kidney transplantation(Fig. 7). However, the animal demonstrated normal kidney func-tions and no signs of rejection. Similar studies attempting to detectchimerism have been performed in two additional class I-mis-matched animals, with the same results.
Donor chimerism in the rekidney transplantation model
Animal no. 11892 received an SLA-identical kidney graft withoutCyA treatment, and, on day 105, received a second kidney graftfrom the same donor, this time with CyA treatment. Surprisingly,after the second kidney graft, even with CyA treatment, no donorchimerism could be detected in the peripheral blood and nonecould be detected in the graft beyond 2 wk (Fig. 8).
Long-term chimerism in peripheral blood after kidneytransplantation
Animal no. 11870 received an SLA-identical kidney allograft withCyA treatment. Donor chimerism in the peripheral blood was
present (0.1–0.5%;p , 0.05) with a peak level detected at around4 wk. Chimerism decreased to 0.1% at around 2 mo, however, itthen increased to 0.15% at 4 mo, and 0.25% at 11 mo after trans-plantation (Fig. 9). At this time, donor chimerism could be de-tected in the peripheral blood and in the graft, but not in the thymusor mesenteric lymph nodes (data not shown).
DiscussionWe have previously demonstrated that SLA-identical renal allo-grafts are accepted permanently in approximately two-thirds ofuntreated miniature swine (8, 19) and in 100% of animals treatedwith a 12-day course of CyA (20, 21). Similarly, class II-matchedrenal allografts are accepted in approximately one-third of casesacross a single haplotype class I-mismatch (22), but are uniformlyrejected across a two-haplotype class I-mismatch (12). In bothcases, 100% of such class I-mismatched transplants are accepted ifthe animals are treated with a 12-day course of CyA (12, 14). Thus,this model provides the possibility for studying tolerance inductionto renal allografts with or without the use of exogenous immuno-suppression.
FIGURE 6. Studies on animal no. 12082, recipient of an SLA-identical kidney allograft without CyA treatment, who suffered ureteral obstruction. Theclinical course was evaluated by serum creatinine levels (A), percent donor cells in peripheral blood (B), and percent donor cells in the graft (C). The arrowin A indicates the time of the surgical procedure to relieve obstruction.
5708 CHIMERISM AND TOLERANCE IN SWINE
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
There has been considerable debate recently in the literatureconcerning the importance of lymphohematopoeitic chimerismfollowing vascularized organ allografts (6, 23, 24). Most investi-gators agree that such chimerism can be detected systemically fol-lowing allografts of this kind. There is controversy, however, con-cerning whether the chimerism detected is the result ofimmunosuppression or the cause of allograft acceptance (23). Inthe case of vascularized organ allografts following the induction oflymphohematopoeitic chimerism by bone marrow transplantation,it is clear that the tolerance persists even if the organ allograft isremoved (25, 26). However, in the case of organ allografts, whichare maintained through the use of exogenous immunosuppression,the tolerance generally disappears when the organ is removed (27).Thus, some investigators have reasoned that the chimerism ismerely a reflection of cells that have escaped from the organ, buthave not been rejected due to the immunosuppressive drugs, whichmaintain a state of hyporesponsiveness (23).
In the present study, we have attempted to determine the rele-vance of chimerism to acceptance of renal allografts in MHC-matched miniature swine. The transplants were performed eitherutilizing a short course of CyA as an immunosuppressive agent, orin the absence of exogenous immunosuppression. In both cases,the kidney grafts were accepted long term. The fact that this ac-ceptance was due to the induction of true immunologic tolerancehas previously been demonstrated in this model by the findingsthat: 1) skin (as opposed to vascularized) grafts across the samehistocompatibility barrier are uniformly rejected (8), leading tosensitization and rapid rejection of subsequent kidney transplants(9); and 2) the survival of skin grafts on animals that have acceptedrenal allografts is markedly prolonged (8). The data reported herereveal no relationship between the persistence of chimerism andthe presence of such tolerance in these animals. Thus, the findingssupport the contention that such chimerism is incidental to, ratherthan the cause of, long-term allograft acceptance.
In the case of a class I-mismatched renal allograft, chimerismwas not detectable (day 8), even in the presence of CyA as anexogenous immunosuppressive agent. The reason for this lack of
FIGURE 7. Chimerism in the peripheral blood (A) and in the graft (B) in a recipient (no. 12179) of a class I-mismatched kidney allograft with CyAtreatment (✡). Chimerism in the peripheral blood and in the graft in recipients of SLA-matched kidney allografts with CyA treatment are also shown (—).
FIGURE 8. Chimerism in the peripheral blood (A), and graft (B) ofanimal no. 11892, which received an SLA-identical kidney graft withoutCyA treatment (—), followed on day 105 by another kidney from the samedonor, with CyA treatment (✡).
FIGURE 9. Long-term chimerism (.300 days) in the peripheral bloodof a CyA-treated recipient (no. 11870) of an SLA-matched kidney graft(✡). The other two CyA-treated recipients of SLA-matched kidney allo-grafts, in which chimerism was only transient, are also shown (—).
5709The Journal of Immunology
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

detectable chimerism is not clear, but could reflect the increasedstrength of the immune reactivity to this class I disparity beforetolerance induction, or could be the effect of NK cell activity,which would only be detectable in the face of a class I mismatch(28–30).
Second renal allografts showed no detectable chimerism, al-though they were accepted long term. The most plausible expla-nation for the failure to detect such chimerism following the sec-ond transplant would be sensitization of the immune system to Agspresent on the lymphohematopoeitic cells, which escape from thegraft. This hypothesis would require that there be Ags, in additionto those expressed on the kidney, which are expressed only on thesurface of such lymphohematopoietic cells. Such differential Agexpression would not be unique, since tissue- and organ-specificAgs have previously been described (31–34).
One surprising finding in these studies was the large increase inchimeric cells detectable following an incidental ureteral obstruc-tion during the postoperative course of one animal. The fact thatthis chimerism disappeared promptly after the ureteral obstructionwas relieved surgically suggests that the chimerism was incidentalto inflammation and/or another physiologic disturbance in the kid-ney, but once again, not related to the immune status.
It is interesting to note that one CyA-treated kidney recipient didexhibit persistent, long-term donor chimerism in the peripheralblood. The clinical course of this animal was unremarkable for thefirst few months after transplant, as the chimeric cell levels fol-lowed the same trends as other CyA-treated animals. However,while chimerism disappeared in the other animals at 40–60 days,it began to increase steadily in this animal, to a maximum of0.25%. This result suggests that among the passenger leukocytes inthe kidney graft was a population of hematopoietic stem cells,which apparently survived and proliferated in the immunosup-pressed animal (35, 36). In addition, the increase and persistence ofthese cells even after discontinuation of the immunosuppressivetreatment indicates that the animal was tolerized, not only to thekidney graft, but also to any tissue-specific Ags expressed on thehematopoietic stem cells.
Previous examples of such long-term chimerism in graft recip-ients are well-documented (1, 2, 37) and have been used as evi-dence that chimerism causes the induction of graft tolerance. Ourdata would indicate that while long-term chimerism can exist inorgan transplant recipients, it is not essential for graft survival.
In summary, these data support the likelihood that chimerismmay be associated with long-term organ allograft survival, but thatit is not essential for the induction or maintenance of the tolerantstate. It is, of course, impossible to ever prove the absence of cellsbelow a detectable level. Nevertheless, the clear presence of suchcells in animals prepared by bone marrow transplantation (25, 38),and in animals on immunosuppression, and the absence of suchcells in animals that become tolerant without immunosuppressionprovides no evidence for a relationship between such chimerismand graft acceptance.
AcknowledgmentsWe thank Drs. David K. C. Cooper and Yongguang Yang for their criticalreview of the manuscript, Mr. Joseph Ambroz for expert animal care, Jes-sica Sachs for editorial assistance, Lisa A. Bernardo for help in manuscriptpreparation, and Dr. Hiroshi Kitamura for help with production of histol-ogy photographs. The authors would also like to acknowledge Lloyd, Inc.for generously providing Xylazine, Baxter Healthcare Corporation for pro-viding Albumin 5%, and Abbott Laboratories for providing Hetastarch 6%.
References1. Starzl, T. E., A. J. Demetris, M. Trucco, A. Zeevi, H. Ramos, P. Terasaki,
W. A. Rudert, M. Kocova, C. Ricordi, S. Ildstad, and N. Murase. 1993. Chimer-ism and donor-specific nonreactivity 27 to 29 years after kidney allotransplanta-tion. Transplantation 55:1272.
2. Starzl, T. E., A. J. Demetris, M. Trucco, H. Ramos, A. Zeevi, W. A. Rudert,M. Kocova, C. Ricordi, S. Ildstad, and N. Murase. 1992. Systemic chimerism inhuman female recipients of male livers.Lancet 340:876.
3. Starzl, T. E., A. J. Demetris, M. Trucco, C. Ricordi, S. Ildstad, P. I. Terasaki,N. Murase, R. S. Kendall, M. Kocova, W. A. Rudert, A. Zeevi, and D. Van Thiel.1993. Chimerism after liver transplantation for type IV glycogen storage diseaseand type 1 Gaucher’s disease.N. Engl. J. Med. 328:745.
4. Hisanaga, M., J. Hundrieser, K. Boker, K. Uthoff, G. Raddatz, T. Wahlers,K. Wonigeit, R. Pichlmayr, and H. J. Schlitt. 1996. Development, stability, andclinical correlations of allogeneic microchimerism after solid organ transplanta-tion. Transplantation 61:40.
5. Starzl, T. E., A. J. Demetris, N. Murase, S. Ildstad, C. Ricordi, and M. Trucco.1992. Cell migration, chimerism, and graft acceptance [see comments].Lancet339:1579.
6. Starzl, T. E., A. J. Demetris, N. Murase, A. W. Thomson, M. Trucco, andC. Ricordi. 1993. Donor cell chimerism permitted by immunosuppressive drugs:a new view of organ transplantation.Immunol.Today 14:326.
7. Sachs, D. H., G. Leight, J. Cone, S. Schwartz, L. Stuart, and S. Rosenberg. 1976.Transplantation in miniature swine. I. Fixation of the major histocompatibilitycomplex.Transplantation 22:559.
8. Kirkman, R. L., M. W. Colvin, G. S. Flye, S. A. Leight, S. A. Rosenberg,G. M. Williams, and D. H. Sachs. 1979. Transplantation in miniature swine. VI.Factors influencing survival of renal allografts.Transplantation 28:18.
9. Kirkman, R. L., R. B. Colvin, M. W. Flye, G. M. Williams, and D. H. Sachs.1979. Transplantation in miniature swine. VII. Evidence for cellular immunemechanisms in hyperacute rejection of renal allografts.Transplantation 28:24.
10. Pennington, L. R., J. K. Lunney, and D. H. Sachs. 1981. Transplantation inminiature swine. VIII. Recombination within the major histocompatibility com-plex of miniature swine.Transplantation 31:66.
11. Kortz, E. O., K. Sakamoto, T. Suzuki, P. C. Guzzetta, C. H. Chester,J. K. Lunney, and D. H. Sachs. 1990. Mechanism of tolerance following class Idisparate renal allografts in miniature swine: cellular responses of tolerant ani-mals.Transplantation 49:1142.
12. Rosengard, B. R., C. A. Ojikutu, P. C. Guzzetta, C. V. Smith, T. M. Sundt III,K. Nakajima, S. M. Boorstein, G. S. Hill, J. M. Fishbein, and D. H. Sachs. 1992.Induction of specific tolerance to class I disparate renal allografts in miniatureswine with cyclosporine.Transplantation 54:490.
13. Gianello, P. R., G. Blancho, J. F. Fishbein, T. Lorf, V. Nickeleit, D. Vitiello, andD. H. Sachs. 1994. Mechanism of cyclosporin-induced tolerance to primarilyvascularized allografts in miniature swine. Effect of administration of exogenousIL-2. J. Immunol. 153:4788.
14. Gianello, P. R., J. F. Fishbein, B. R. Rosengard, T. Lorf, D. M. Vitiello, J. S. Arn,and D. H. Sachs. 1995. Tolerance to class I disparate renal allografts in miniatureswine: maintenance of tolerance despite induction of specific antidonor CTLresponses.Transplantation 59:772.
15. Giangrande, I., K. Yamada, S. Arn, T. Lorf, D. H. Sachs, and C. LeGuern. 1997.Selective increase in CD4-positive graft-infiltrating mononuclear cells among theinfiltrates in class I disparate kidney grafts undergoing rejection.Transplantation63:722.
16. Pescovitz, M. D., J. K. Lunney, and D. H. Sachs. 1984. Preparation and charac-terization of monoclonal antibodies reactive with porcine PBL.J. Immunol. 133:368.
17. Pescovitz, M. D., J. K. Lunney, and D. H. Sachs. 1985. Murine anti-swine T4 andT8 monoclonal antibodies: distribution and effects on proliferative and cytotoxicT cells.J. Immunol. 134:37.
18. Saalmuller, A., B. Aasted, A. Canals, J. Dominguez, T. Goldman, J. K. Lunney,S. Maurer, M. D. Pescovitz, R. Pospisil, and H. Salmon. 1994. Analyses of mAbreactive with porcine CD8.Vet. Immunol. Immunopathol. 43:249.
19. Rosengard, B. R., C. A. Ojikutu, J. Fishbein, E. O. Kortz, and D. H. Sachs. 1992.Selective breeding of miniature swine leads to an increased rate of acceptance ofMHC-identical, but not of class-I disparate, renal allografts.J. Immunol. 149:1099.
20. Gianello, P., J. M. Fishbein, and D. H. Sachs. 1993. Tolerance to primarily vas-cularized allografts in miniature swine.Immunol. Rev. 133:19.
21. Gianello, P. R., and D. H. Sachs. 1996. Effect of major histocompatibility com-plex matching on the development of tolerance to primarily vascularized renalallografts: a study in miniature swine.Hum. Immunol. 50:1.
22. Pescovitz, M. D., J. R. Thistlethwaite, Jr., H. Auchincloss, Jr., S. T. Ildstad,T. G. Sharp, R. Terrill, and D. H. Sachs. 1984. Effect of class II antigen matchingon renal allograft survival in miniature swine.J. Exp. Med. 160:1495.
23. Wood, K., and D. H. Sachs. 1996. Chimerism and transplantation tolerance:cause and effect.Immunol. Today 17:584.
24. Shirwan, H., C. Wang, L. Barwari, and D. V. Cramer. 1997. Recipient-donormicrochimerism is not a prerequisite for the maintenance of allograft tolerance.Transplant. Proc. 29:1182.
25. Sharabi, Y., and D. H. Sachs. 1989. Mixed chimerism and permanent specifictransplantation tolerance induced by a nonlethal preparative regimen.J. Exp.Med. 169:493.
26. Sharabi, Y., V. S. Abraham, M. Sykes, and D. H. Sachs. 1992. Mixed allogeneicchimeras prepared by a non-myeloablative regimen: requirement for chimerismto maintain tolerance.Bone Marrow Transplant. 9:191.
5710 CHIMERISM AND TOLERANCE IN SWINE
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

27. Shizuru, J. A., K. B. Seydel, T. F. Flavin, A. P. Wu, C. C. Kong, E. G. Hoyt,N. Fujimoto, M. E. Billingham, V. A. Starnes, and C. G. Fathman. 1990. Induc-tion of donor-specific unresponsiveness to cardiac allografts in rats by pretrans-plant anti-CD4 monoclonal antibody therapy.Transplantation 50:366.
28. Murphy, W. J., V. Kumar, and M. Bennett. 1987. Rejection of bone marrowallografts by mice with severe combined immune deficiency (scid): evidence thatnatural killer cells can mediate the specificity of marrow graft rejection.J. Exp.Med. 165:1212.
29. Lanier, L. L., and J. H. Phillips. 1996. Inhibitory MHC class I receptors on NKcells and T cells.Immunol. Today 17:86.
30. Sentman, C. L., V. Kumar, and M. Bennett. 1991. Rejection of bone marrow cellallografts by natural killer cell subsets: 5E61 cell specificity for Hh-1 determinant2 shared by H-2d and H-2f. Eur. J. Immunol. 21:2821.
31. Steinmuller, D., and J. S. Lofgreen. 1974. Differential survival of skin and heartallografts in radiation chimaeras provides further evidence for Sk histocompati-bility antigen.Nature 248:796.
32. Warren, R. P., J. S. Lofgreen, and D. Steinmuller. 1973. Factors responsible forthe differential survival of heart and skin allografts in inbred rats.Transplantation16:458.
33. Rosengard, B. R., E. O. Kortz, C. A. Ojikutu, P. C. Guzzetta, T. M. Sundt,C. V. Smith, K. Nakajima, S. M. Boorstein, G. S. Hill, and D. H. Sachs. 1991.
The failure of skin grafting to break tolerance to class I disparate renal allograftsin miniature swine despite inducing marked anti-donor cellular immunity.Trans-plantation 52:1044.
34. Auchincloss, H., Jr., and D. H. Sachs. 1993. Transplantation and graft rejection.In Fundamental Immunology. 3rd Ed. W.E. Paul, ed. Raven Press, New York, p.1099.
35. Taniguchi, H., T. Toyoshima, K. Fukao, and H. Nakauchi. 1996. Presence ofhematopoietic stem cells in the adult liver.Nat. Med. 2:198.
36. Murase, N., T. E. Starzl, Q. Ye, A. Tsamandas, A. W. Thomson, A. S. Rao, andA. J. Demetris. 1996. Multilineage hematopoietic reconstitution of supralethallyirradiated rats by syngeneic whole organ transplantation. With particular refer-ence to the liver.Transplantation 61:1.
37. Burlingham, W. J., A. P. Grailer, J. H. J. Fechner, S. Kusaka, M. Trucco,M. Kocova, F. O. Belzer, and H. W. Sollinger. 1995. Microchimerism linked tocytotoxic T lymphocyte functional unresponsiveness (clonal anergy) in a tolerantrenal transplant recipient.Transplantation 59:1147.
38. Ildstad, S. T., and D. H. Sachs. 1984. Reconstitution with syngeneic plus allo-geneic or xenogeneic bone marrow leads to specific acceptance of allografts orxenografts.Nature 307:168.
5711The Journal of Immunology
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from





























