Embed Size (px)
Citation preview

s t e r o i d s 7 1 ( 2 0 0 6 ) 298–303
avai lab le at www.sc iencedi rec t .com
journa l homepage: www.e lsev ier .com/ locate /s tero ids
S-palmitoylation modulates estrogen receptor �
localization and functions
Maria Marinoa,∗, Paolo Ascenzia,b, Filippo Acconciaa,1
a Department of Biology, University ‘Roma Tre’, Viale Guglielmo Marconi 446, 00146 Roma, Italyb National Institute for Infectious Diseases I.R.C.C.S. ‘Lazzaro Spallanzani’,Via Portuense 292, 00149 Roma, Italy
a r t i c l e i n f o
Article history:
Published on line 7 November 2005
a b s t r a c t
17�-Estradiol (E2) acts as a chemical messenger in target tissues inducing both slow nuclear
and rapid extra-nuclear responses. E2 binds to its cognate nuclear receptors (ER) resulting
Keywords:
17�-Estradiol
Estrogen receptor �
S-palmitoylation
Rapid non-genomic actions
Plasma membrane localization
Abbreviations:
2-Br, 2-bromo-hexadecanoic acid;
E2, 17�-estradiol; ER, estrogen
receptor; ERK, extracellular
regulated kinase; HE-14,
A-to-D-deleted ER� mutant; HE-15,
E-to-F-deleted ER� mutant; IP3,
inositol trisphosphate; MAPK,
mitogen-activated protein kinase;
MNAR, modulator of non-genomic
activity of ER; PAT, palmitoyl
acyl-transferase; PI3K,
phosphatidyl-inositol-3-kinase;
PKC�, protein kinase C�; PLC,
phospholipase C
in the activation of target gene transcription in the nucleus. In addition to these genomic
effects, E2 modulates cell functions through rapid non-genomic actions. Stimulation of G-
proteins, Ca2+ influx, inositol phosphate generation as well as phospholipase C, ERK/MAPK,
and PI3K/AKT activation all occur within seconds to minutes after E2 binding to a small pop-
ulation of ER� located at the plasma membrane. The great impact of these rapid signals on
cell physiology renders central the knowledge of the structural bases and mechanisms that
mediate extra-nuclear signaling by E2. Several laboratories, including our own, have recently
elucidated the structural requirements for localization and function of plasma membrane
ER�. This review summarizes the molecular mechanisms of E2-induced rapid non-genomic
actions relevant for cell functions, highlighting the role of lipid modification (i.e., palmitoy-
lation) in the ER� localization to and residence at the plasma membrane.
© 2005 Elsevier Inc. All rights reserved.
1. Introduction
The action of 17�-estradiol (E2) in living cells is mediated bya complex signaling network rather than a single uniform
∗ Corresponding author. Tel.: +39 06 55176345; fax: +39 06 55176321.E-mail address: [email protected] (M. Marino).
1 Present address: Department of Molecular and Cellular Oncology, Anderson Cancer Center, 1515 Holcombe Boulevard, Texas MedicalCenter, Houston, 77030 TX, USA.
mechanism. In fact, in addition to its role as a regulator ofgene transcription [1,2], sustained by the classical nuclearestrogen receptors (ER), E2 also shows rapid non-genomicactions [3,4]. Treatment with E2 generates intracellular sec-
0039-128X/$ – see front matter © 2005 Elsevier Inc. All rights reserved.doi:10.1016/j.steroids.2005.09.011

s t e r o i d s 7 1 ( 2 0 0 6 ) 298–303 299
ond messengers (i.e., Ca2+ influx, inositol phosphate, andcyclic AMP) [5–9] which, in turn, activate or inhibit membranestarting signal cascades in all the E2-target cells [3–5,7,10–16].
The physiological relevance of rapid non-genomic path-ways is starting to be clarified in both non-reproductiveand reproductive E2 target cells as well as in tissues. E2influences neural functions (e.g., cognition, behavior, stressresponses, and reproduction) [14,17,18] as well as bone form-ing and resorption [19–21]. Moreover, rapid E2-induced sig-nals [i.e., phospholipase C (PLC)/protein kinase C� (PKC�)]are deeply linked to the expression of low-density lipopro-tein receptor in the liver and to the decreased plasmaticlevel of cholesterol low-density lipoproteins [22,23]. Fur-thermore, vascular protection by E2 in ischemia/reperfusioninjury requires E2-induced activation of endothelial nitricoxide synthase, as mediated by the phosphatidylinositol3 kinase (PI3K)/AKT pathway [24,25]. Finally, the capabil-ity of rapid E2-induced signal transduction pathways toimpact the classic genomic mechanisms has been reported[3,26–28].
There is growing evidence that the rapid actions activatedupon E2 binding to ER� are important for cell proliferation.The E2 treatment of mammary-derived MCF-7 cells triggersassociation of ER� with Src and p85, the regulatory subunitof PI3K, leading to DNA synthesis [29]. Moreover, multipleand parallel membrane starting pathways are rapidly acti-vm(p[
cmhEtlp
2p
TcesbshffabpEbm
domain(s) in ER� is not yet clarified (see [3,42] and Bender JR,personal communication).
Current evidences indicate that the small population ofER� localized at the plasma membrane exists within cave-olar rafts [25,40,41,44]. This membrane ER� is tethered tothe inner face of the plasma membrane through bindingto specific membrane proteins such as caveolin-1 [40], Src[29], Shc [45], the p85 subunit of PI3K [24], receptor tyro-sine kinases (i.e., EGFR and IGF-1R) [3,42], as well as twoisoforms of G-protein (i.e., Gs� and Gq�) [7,43]. Interestingly,a novel protein termed “modulator of non-genomic activ-ity of ER” (MNAR) has been identified to act as an adapterbetween ER and Src, providing a mechanism for coupling E2and ER with the rapid signaling cascade [46,47]. New associ-ation between ER� and cytosolic or membrane proteins arerecognized monthly, but all interactions are E2-dependentbeing evident 5–15 min after hormone stimulation [42]. Bycontrast, ER� localizes at the plasma membrane also in un-stimulated cells [7,37–39,42] suggesting that the ER�–protein(s)interaction(s) is not the sole mode for ER� association to theplasma membrane, post-translational modifications may be inorder.
In 1999, Levin [48] speculated that ER post-translationalmodifications (e.g., lipid addition) could occur in the endo-plasmatic reticulum facilitating ER� movement to and inser-tion into the plasma membrane. The most diffused pro-
ated by the ER�–E2 complex in HepG2 cells [8,13,16]. Further-ore, the blockade of PLC/PKC, extracellular regulated kinase
ERK)/mitogen-activated protein kinase (MAPK), and PI3K/AKTathways completely prevents the E2-induced DNA synthesis
13,16,30].Due to the great impact of rapid non-genomic signals on
ell physiology, the understanding of the structural bases andechanisms that mediate extra-nuclear signaling by E2 is a
ot issue. This review reports the molecular mechanisms of2-induced rapid non-genomic actions relevant for cell func-ions, highlighting the role of lipid modification (i.e., palmitoy-ation) in the ER� localization to and in its maintenance at thelasma membrane.
. Estrogen receptor � localization at thelasma membrane: biochemical aspects
he existence of a membrane-associated ER(s) unrelated tolassical nuclear ER� has been long postulated [31–36]. How-ver, immunocytochemical studies revealed the presence of aignificant fraction of nuclear ER(s) also at the plasma mem-rane [37–42]. Moreover, a single mRNA originates a similarized nuclear and membrane ER in ER�-transfected Chineseamster ovary and HeLa cells [7,13,16]. Furthermore, cells
rom the DERKO mouse (ER� and ER� double knock-out mice)ail to show endogenous membrane or nuclear ER� or ER�
nd E2 binding [43]. On these bases, ER� and ER� have toe considered as a population of proteins which localize atlasma membrane, in the cytosol, and in the nucleus [3]. TheR� association to plasma membrane seems to be mediatedy association with membrane proteins and/or by ER� lipidodification(s) since the presence of intrinsic transmembrane
tein acylation process occurs either through amide link-age of myristate to the N-terminal residue (N-acylation) orof palmitate to the S� atom of Cys residues (S-acylation).Also S-prenylation occurs at the Cys S� atom. Protein N-acylation and S-prenylation occur only on strict consensussequences [49,50]. By contrast, no consensus sequence(s) hasbeen reported for S-palmitoylation [51]. Note that no consen-sus sequences for N-acylation and S-prenylation have beenfound in ER� [52].
ER� contains 13 cysteines; eight out of the nine Cysresidues present in the ER� DNA binding domain are involvedin two zinc-thiolate centers (Cys185, Cys188, Cys202, Cys205,and Cys221, Cys227, Cys237, Cys240, respectively), whileCys245 is completely buried in the protein matrix [53]. TheCys381, Cys417, Cys447, and Cys530 residues present inthe ER� ligand binding domain may be solvent exposed[53,54] and then may undergo S-acylation. Interestingly,the amino acid sequence encompassing the Cys447 residueof ER� is highly homologous to that surrounding the S-palmitoylated Cys132 residue of human caveolin-1 [52,55,56].Endogenous (HepG2 and MCF7 cells) or expressed (HeLacells) ER� undergoes palmitate incorporation quickly; S-palmitoylation remains constant over 240 min. Remarkably,palmitate incorporation in ER� is not a spontaneous reaction,being catalyzed by palmitoyl acyl-transferase (PAT). In fact,a significant decrease in ER� S-palmitoylation occurs in cellspre-treated with the PAT inhibitor 2-bromo-hexadecanoicacid (2-Br) [57]. Only wild-type ER� and the A-to-D-deletedER� mutant (HE14) undergo palmitate incorporation whenexpressed in HeLa cells. By contrast, no palmitoylationoccurs in HeLa cells expressing the E-to-F-deleted ER�
mutant (HE15) or the Cys447Ala ER� mutant [57]. As a whole,the Cys447 residue represents the ER� S-palmitoylationsite.

300 s t e r o i d s 7 1 ( 2 0 0 6 ) 298–303
3. S-palmitoylation modulates ER�localization and activities
A weak cytosolic staining and no membrane-bound recep-tor has been detected in cells expressing the ER� Cys447Alamutant, whereas a major population of membrane-associatedER� receptor is clearly present in cells expressing wild-typeER� [58]. Therefore, the Cys447 residue is important for ER�
plasma membrane localization [58]. Moreover, no associationbetween caveolin-1 and the ER� Cys447Ala mutant occurs intransfected HeLa cells [58]. As a whole, these data are consis-tent with the role of S-palmitoylation in ER� localization at theplasma membrane and in the ER� association with caveolin-1. S-palmitoylation as a possible mechanism for membraneER localization was also suggested from previous studies inendothelial cells [59] and has been recently confirmed by ER�
expression in other cancer cell lines [60].S-palmitoylation is necessary for the E2-induced non-
genomic events. In fact, ER� Cys447Ala mutation or inhibitionof palmitoylation by 2-Br in wild-type ER� does not allow E2-induced proliferative signaling via ERK/MAPK and PI3K/AKTpathways in human cancer cells [58]. Similar results havebeen recently reported in other tumor cell lines [60]. Moreover,neither the cyclin D1 promoter activity nor thymidine incor-poration into DNA were affected by E2 administration to theER� Cys447Ala mutant-transfected cells. In addition, the pre-
ing [62,63]. Remarkably, preliminary data obtained in our lab-oratory indicate that naringenin stimulation in cancer cellsrapidly induces (1 min) the decrease of ER� palmitoylationand ER�-caveolin-1 disassociation (Marino M, unpublisheddata). Interestingly, naringenin induces the ER� rapid de-palmitoylation and decouples the ER� action mechanismsimpairing the activation of the ERK/MAPK and PI3K/AKT sig-nal transduction pathways [63]. The different ER� activitiesinduced by the different ligands (i.e., E2 and naringenin) rein-force the view that ER� is an allosteric protein. Thus, theligand-dependendent conformational changes of ER� may beon the root of the dynamic migration of ER� from plasmamembrane to the cytosol [39].
The ligand-binding domain of ER� (E domain) appears to bestrongly involved in the ligand-induced allosteric modulationof ER�. The E domain appears to be the minimum structurerequired for E2 signaling responses [3,13]. The E domain dele-tion prevents ER� S-palmitoylation, dimerization, and signal-ing via ERK/MAPK, PI3K, and cyclic AMP [3,13,58,64]. However,the A, B, and C domains of ER� contribute to the E2-induceddown-regulation of ER� S-palmitoylation and ER� associationto caveolin-1. In fact, E2 induces ∼40% increase of ER� S-palmitoylation and ER� association to caveolin-1 in HeLa cellstransfected with the ER� deletion mutant HE14 [58]. Thus, theA, B, and C domains of ER� may impair the E2-dependent S-palmitoylation and dissociation from caveolin-1 of the plasmamembrane-bound ER�. As a consequence, the A, B, and C
treatment of HeLa cells with the PAT inhibitor 2-Br preventednon-genomic E2-evoked effects [58].
Remarkably, the E2-induced gene transactivation is signif-icantly reduced (by ∼30%) in cells transfected with the ER�
Cys447Ala mutant or pre-treated with the PAT inhibitor 2-Br[57]. These data support the idea that the E2-induced rapid sig-nals cross-talk with genomic events to preserve the pleiotropichormone effects into the cells.
As a whole, S-palmitoylation allows the extra-nuclearlocalization of ER�; this promotes downstream signaling forE2-mediated proliferation and survival of cancer cells offeringa new target for the anti-tumor therapy.
4. Ligand-dependent ER� S-palmitoylation
Physiological E2 concentration (10 nM) reduces by half theamount of palmitate incorporated in ER� within about 60 minwithout any change in the ER level [58]. Accordingly, E2 inducesa ∼90% reduction in the ability of ER� to form a complexwith caveolin-1 [58]. Consistent with this, Katzenellenbogenand coworkers [61] generated MDA MB-231 breast cancer celllines that have stably integrated the membrane-targeted ER(MT-ER). MT-ER lacks the nuclear localization sequence andis dually acylated with a myristoylation sequence at the N-terminus and a palmitoylation sequence at the C-terminus.In the absence of E2, MT-ER is localized primarily to the cellmembrane with some cytoplasmic localization. After E2 treat-ment, MT-ER reduced its localization at the plasma membraneshowing a punctuate pattern with localization in the mem-brane and cytoplasm.
We recently reported that the flavonoid naringenin exertsanti-proliferative effects by altering the ER�-mediated signal-
domains, rendering the membrane-bound ER� competent tohormone regulation, may account for the coexistence of bothmembrane-bound and soluble isoforms of ER�.
5. Conclusion and perspectives
The mechanism(s) underlying the mitogenic role played by E2in different target tissues is now better understood based onthe studies reported by different laboratories, including ourown. These studies designate a strict relationship betweenthe E2-induced non-genomic functions and cell proliferation.In particular, the activation of ERK/MAPK, PI3K/AKT and pro-tein kinase C, rapidly generated after E2 binding to ER�, are alldefined as necessary and sufficient for E2-induced cell cycle-regulating genes (e.g., cyclin D1) and the G1-to-S phase pro-gression in different cell lines [7,8,12,13,16,29,30,65–67]. Theseactions are thought to require a plasma membrane ER� whichis dependent on S-palmitoylation.
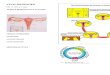
Unliganded ER� at steady state is localized at the plasmamembrane caveolae, cytosol, and nucleus. S-palmitoylationof the Cys447 residue at the E domain of ER� (Fig. 1, toppanel) sequentially guarantees receptor maintenance at theplasma membrane and association with caveolin-1. E2 bind-ing to the membrane receptor, enacting ER� structural mod-ifications [68–70], could impair the PAT action (e.g., PAT-ER�
recognition) decreasing the ER�–caveolin-1 complex forma-tion. This event allows the interaction of ER� with other adap-tors and/or partner proteins (e.g., Shc/IGF-1R, Src/p85, andMNAR) [29,42,45,46,71]. As a consequence, the non-genomicsignals could be generated (i.e., ERK/MAPK, PI3K/AKT) (Fig. 1,bottom panel). Besides its role in receptor shuttling, E2 plays acrucial role also in modifying the ER expression in the whole

s t e r o i d s 7 1 ( 2 0 0 6 ) 298–303 301
Fig. 1 – Schematic model illustrating the receptorlocalization at and maintenance to the plasma membranecaveolae (top panel) and the E2-dependent ER�
re-localization (bottom panel). The relationship betweenrapid and long-term actions of E2 on target cells is alsoshowed (bottom panel). E2, 17�-estradiol; ER�, estrogenreceptor �; MNAR, modulator of non-genomic activity of ER;PA, palmitic acid; PAT, palmitoyl acyl-transferase; p85,active sub-unit of phosphatidyl-inositol-3-kinase. Fordetails, see text.
cells. In fact, 5 h after exposure of MCF7 cells to E2 the nuclearlevel of ER� increases, the ER gene transcription is repressed,and proteasome-mediated ER� degradation occurs [72–74].Notably, pre-treating MCF7 cells with the PAT inhibitor 2-Br theagonist-induced ER� breakdown fastens (2 h) while 2-Br alonedoes not affect the ER level (Marino M, unpublished data).Thus, the presence of an ER� membrane population seems tobe necessary to maintain the receptor half-life indispensablefor the E2-induced cell functions.
Covalent lipid modifications anchor numerous signallingproteins to the cytoplasmic face of the plasma membrane.In particular, protein S-palmitoylation, due to its reversiblenature, is particularly important for modulating protein func-tion during cycles of activation and deactivation [49]. In fact,it is well accepted that S-palmitoylation can affect protein
binding to membranes, protein subcellular localization, andprotein–protein interactions [49,50,75,76]. Our data indicatethat ER� activities depend on S-palmitoylation. Consideringthat all classes of steroid hormones possess receptors ableto generate rapid responses [18,67,70], a new area of inves-tigation focusing the role played by chemical modification(s)on steroid receptor activity and localization could be opened.Intriguingly, several members of the steroid hormone receptorfamily display an amino acid sequence highly homologous tothat encompassing the Cys447 residue of ER� which under-goes S-palmitoylaton (Marino M, unpublished data). Thesestudies will aid the development of specific agonist and antag-onists with well-characterized effects able to modify discreteE2-induced cellular functions.
Acknowledgments
The authors wish to thank past and present members oftheir laboratories who contributed to the ideas presentedhere through data and discussions. This work was supportedby grants from the Ministry of Education, University, andResearch of Italy (PRIN-COFIN 2004 to M.M.).
r e f e r e n c e s
[1] Nilsson S, Makela S, Treuter E, Tujague M, Thomsen J,Andersson G, et al. Mechanisms of estrogen action.Physiol Rev 2001;81:1535–65.
[2] Acevedo ML, Kraus WL. Transcriptional activation bynuclear receptors. In: McEwan IJ, editor. Essays inbiochemistry: the nuclear receptor superfamily. Oxford:Portland Press Ltd.; 2004. p. 73–88.
[3] Levin ER. Integration of the extranuclear and nuclearactions of estrogen. Mol Endocrinol 2005;19:1951–9.
[4] Marino M, Acconcia F, Ascenzi P. Estrogen receptorsignalling: bases for drug actions. Curr Drug TargetsImmune Endocr Metabol Disord 2005;5:305–14.
[5] Morley P, Whitfield JF, Vanderhyden BC, Tsang BK,Schwartz JL. A new nongenomic estrogen action: the rapidrelease of intracellular calcium. Endocrinology1992;131:1305–12.
[6] Farhat MY, Abi-Younes S, Dingaan B, Vargas R, RamwellPW. Estradiol increases cyclic adenosine monophosphatein rat pulmonary vascular smooth muscle cells by anongenomic mechanism. J Pharmacol Exp Ther1996;276:652–7.
[7] Razandi M, Pedram A, Greene GL, Levin ER. Cellmembrane and nuclear estrogen receptors (ERs) originatefrom a single transcript: studies of ER� and ER� expressedin Chinese hamster ovary cells. Mol Endocrinol1999;13:307–19.
[8] Marino M, Pallottini V, Trentalance A. Estrogens causerapid activation of IP3-PKC-� signal transduction pathwayin HEPG2 cells. Biochem Biophys Res Commun1998;245:254–8.
[9] Perret S, Dockery P, Harvey BJ. 17�-Oestradiol stimulatescapacitative Ca2+ entry in human endometrial cells. MolCell Endocrinol 2001;176:77–84.
[10] Watters JJ, Campbell JS, Cunningham MJ, Krebs EG, DorsaDM. Rapid membrane effects of steroids in neuroblastomacells: effects of estrogen on mitogen activated proteinkinase signalling cascade and c-fos immediate early genetranscription. Endocrinology 1997;138:4030–3.

302 s t e r o i d s 7 1 ( 2 0 0 6 ) 298–303
[11] Picotto G, Vazquez G, Boland R. 17�-Oestradiol increasesintracellular Ca2+ concentration in rat enterocytes.Potential role of phospholipase C-dependentstore-operated Ca2+ influx. Biochem J 1999;339:71–7.
[12] Marino M, Distefano E, Pallottini V, Caporali S, Ceracchi G,Trentalance A. �-Estradiol stimulation of DNA synthesisrequires different PKC isoforms in HepG2 and MCF7 cells. JCell Physiol 2001;188:170–7.
[13] Marino M, Acconcia F, Bresciani F, Weisz A, Trentalance A.Distinct nongenomic signal transduction pathwayscontrolled by 17�-estradiol regulate DNA synthesis andcyclin D1 gene transcription in HepG2 cells. Mol Biol Cell2002;13:3720–9.
[14] Farach-Carson MC, Davis PJ. Steroid hormone interactionswith target cells: cross talk between membrane andnuclear pathways. J Pharmacol Exp Ther 2003;30:839–45.
[15] Incerpi S, D’Arezzo S, Marino M, Musanti R, Pallottini V,Pascolini A, et al. Short-term activation by low17�-estradiol concentrations of the Na+/H+ exchanger inrat aortic smooth muscle cells: physiopathologicalimplications. Endocrinology 2003;144:4315–24.
[16] Marino M, Acconcia F, Trentalance A. Biphasic estradiolinduced AKT-phosphorylation is modulated by PTEN viaMAP kinase in HepG2 cells. Mol Biol Cell 2003;14:2583–91.
[17] Wise PM, Dubal DB, Wilson ME, Rau SW, Bottner M.Neuroprotective effects of estrogen-new insights intomechanisms of action. Endocrinology 2001;142:969–73.
[18] Losel RM, Falkenstein E, Feuring M, Schultz A, TillmannHC, Rossol-Haseroth K, et al. Nongenomic steroid action:controversies, questions, and answers. Physiol Rev2003;83:965–1016.
[28] Acconcia F, Marino M. Synergism between genomic andnon-genomic estrogen action mechanisms. IUBMB Life2003;55:145–50.
[29] Castoria G, Migliaccio A, Bilancio A, Di Domenico M, deFalco A, Lombardi M, et al. PI3-kinase in concert with Srcpromotes the S-phase entry of oestradiol-stimulatedMCF-7 cells. EMBO J 2001;20:6050–9.
[30] Castoria G, Barone MV, Di Domenico M, Bilancio A,Ametrano D, Migliaccio A, et al. Non-trascriptional actionof oestradiol and progestin triggers DNA synthesis. EMBO J1999;18:2500–10.
[31] Filardo EJ, Quinn JA, Bland KI, Frackelton Jr AR.Estrogen-induced activation of Erk-1 and Erk-2 requiresthe G protein-coupled receptor homolog, GPR30, andoccurs via trans-activation of the epidermal growth factorreceptor through release of HB-EGF. Mol Endocrinol2000;14:1649–60.
[32] Ropero AB, Soria B, Nadal A. A nonclassical estrogenmembrane receptor triggers rapid differential actions inthe endocrine pancreas. Mol Endocrinol 2002;16:497–505.
[33] Toran-Allerand CD, Guan X, MacLusky NJ, Horvath TL,Diano S, Singh M, et al. ER-X: a novel, plasmamembrane-associated, putative estrogen receptor that isregulated during development and after ischemic braininjury. J Neurosci 2002;22:8391–401.
[34] Deecher DC, Swiggard P, Frail DE, O’Connor LT.Characterization of a membrane-associated estrogenreceptor in a rat hypothalamic cell line (D12). Endocrine2003;22:211–23.
[35] Maggiolini M, Vivacqua A, Fasanella G, Recchia AG, Sisci D,Pezzi V, et al. The G protein-coupled receptor GPR30
[19] Kousteni S, Chen JR, Bellido T, Han L, Ali AA, O’Brien CA,et al. Manolagas SC reversal of bone loss in mice bynongenotropic signaling of sex steroids. Science2002;298:843–6.
[20] Manolagas SC, Kousteni S, Jilka RL. Sex steroids and bone.Recent Prog Horm Res 2002;57:385–409.
[21] Kousteni S, Han L, Chen JR, Almeida M, Plotkin LI, BellidoT, et al. Kinase-mediated regulation of commontranscription factors accounts for the bone-protectiveeffects of sex steroids. J Clin Invest 2003;111:1651–64.
[22] Marino M, Distefano E, Pallottini V, Caporali S, BruscalupiG, Trentalance A. Activation of IP3-protein kinase C-�
signal transduction pathway precedes the changes ofplasma cholesterol, hepatic lipid metabolism andinduction of low-density lipoprotein receptor expression in17-�-oestradiol-treated rats. Exp Physiol 2001;86:39–45.
[23] Distefano E, Marino M, Gillette JA, Hanstein B, Pallottini V,Bruning J, et al. Role of tyrosine kinase signaling inestrogen-induced LDL receptor gene expression in HepG2cells. Biochim Biophys Acta 2002;1580:145–9.
[24] Simoncini T, Hafezi-Moghadam A, Brazil DP, Ley K, ChinWW, Liao JK. Interaction of oestrogen receptor with theregulatory subunit of phosphatidylinositol-3-OH kinase.Nature 2000;407:538–41.
[25] Chambliss KL, Shaul PW. Rapid activation of endothelialNO synthase by estrogen: evidence for a steroid receptorfast-action complex (SRFC) in caveolae. Steroids2002;67:413–9.
[26] Bjornstrom L, Sjoberg M. Signal transducers and activatorsof transcription as downstream targets of nongenomicestrogen receptor actions. Mol Endocrinol 2002;16:2202–14.
[27] Pedram A, Razandi M, Aitkenhead M, Hughes CC, LevinER. Integration of the non-genomic and genomic actionsof estrogen. Membrane-initiated signaling by steroid totranscription and cell biology. J Biol Chem2002;277:50768–75.
mediates c-fos up-regulation by 17�-estradiol andphytoestrogens in breast cancer cells. J Biol Chem2004;279:27008–16.
[36] Thomas P, Pang Y, Filardo EJ, Dong J. Identity of anestrogen membrane receptor coupled to a G protein inhuman breast cancer cells. Endocrinology 2005;146:624–32.
[37] Pappas TC, Gametchu B, Watson CS. Membrane estrogenreceptors identified by multiple antibody labeling andimpeded-ligand binding. FASEB J 1995;9:404–10.
[38] Norfleet AM, Thomas ML, Gametchu B, Watson CS.Estrogen receptor-� detected on the plasma membrane ofaldehyde-fixed GH3/B6/F10 rat pituitary tumor cells byenzyme-linked immunocytochemistry. Endocrinology1999;140:3805–14.
[39] Dan P, Cheung JC, Scriven DR, Moore ED. Epitopedependent localization of estrogen receptor-�, but not �,in en face arterial endothelium. Am J Physiol2003;284:H1295–306.
[40] Razandi M, Alton G, Pedram A, Ghonshani S, Webb P, LevinER. Identification of a structural determinant necessary forthe localization and function of estrogen receptor � at theplasma membrane. Mol Cell Biol 2003;23:1633–46.
[41] Arvanitis DN, Wang H, Bagshaw RD, Callahan JW, BoggsJM. Membrane-associated estrogen receptor and caveolin-1are present in central nervous system myelin andoligodendrocyte plasma membranes. J Neurosci Res2004;75:603–13.
[42] Song RXD, Barnes CJ, Zhang Z, Bao Y, Kumar R, Santen RJ.The role of Shc and insulin-like growth factor 1 receptorin mediating the translocation of estrogen receptor � tothe plasma membrane. Proc Natl Acad Sci USA2004;101:2076–81.
[43] Razandi M, Pedram A, Merchenthaler I, Greene GL, LevinER. Plasma membrane estrogen receptors exist andfunctions as dimers. Mol Endocrinol 2004;18:2854–65.
[44] Razandi M, Oh P, Pedram A, Schnitzer J, Levin ER. ERsassociate with and regulate the production of caveolin:

s t e r o i d s 7 1 ( 2 0 0 6 ) 298–303 303
implications for signaling and cellular actions. MolEndocrinol 2002;16:100–15.
[45] Song RXD, McPherson RA, Adam L, Bao Y, Shupnik M,Kumar R, et al. Linkage of rapid estrogen action to MAPKactivation by ER�-Shc association and Shc pathwayactivation. Mol Endocrinol 2002;16:116–27.
[46] Wong C-W, McNally C, Nickbarg E, Komm BS, Cheskis BJ.Estrogen receptor-interacting protein that modulates itsnongenomic activity-crosstalk with Src/Erkphosphorylation cascade. Proc Natl Acad Sci USA2002;99:14783–8.
[47] Barletta F, Wong CW, McNally C, Komm BS,Katzenellenbogen BS, Cheskis BJ. Characterization of theinteractions of estrogen receptor and MNAR in theactivation of cSrc. Mol Endocrinol 2004;18:1096–108.
[48] Levin ER. Cellular functions of the plasma membraneestrogen receptor. Trends Endocrinol Metab 1999;10:374–7.
[49] Dunphy JT, Linder ME. Signalling functions of proteinpalmitoylation. Biochim Biophys Acta 1998;1436:245–61.
[50] Resh MD. Fatty acylation of proteins: new insights intomembrane targeting of myristoylated and palmitoylatedproteins. Biochim Biophys Acta 1999;1451:1–16.
[51] Bijlmakers MJ, Marsh M. The on-off story of proteinpalmitoylation. Trends Cell Biol 2003;13:32–42.
[52] Acconcia F, Bocedi A, Ascenzi P, Marino M. Doespalmitoylation target estrogen receptors to plasmamembrane caveolae? IUBMB Life 2003;55:33–5.
[53] Marino M, Ficca R, Ascenzi P, Trentalance A. Nitric oxideinhibits selectively the 17�-estradiol-induced geneexpression without affecting nongenomic events in HeLacells. Biochem Biophys Res Commun 2001;286:529–33.
[62] Virgili F, Acconcia F, Ambra R, Rinna A, Totta P, Marino M.Nutritional flavonoids modulate estrogen receptor �
signaling. IUBMB Life 2004;56:145–51.[63] Totta P, Acconcia F, Leone S, Cardillo I, Marino M.
Mechanisms of naringenin-induced apoptotic cascade incancer cells: involvement of estrogen receptor � and �
signalling. IUBMB Life 2004;56:491–9.[64] Kousteni S, Bellido T, Plotkin LI, O’Brien CA, Bodenner DL,
Han L, et al. Nongenotropic, sex-nonspecific signalingthrough the estrogen or androgenreceptors: dissociationfrom transcriptional activity. Cell 2001;104:719–30.
[65] Lobenhofer EK, Huper G, Iglehart JD, Marks JR. Inhibition ofmitogen-activated protein kinase and phosphatidylinositol3-kinase activity in MCF-7 cells prevents estrogen-inducedmitogenesis. Cell Growth Differ 2000;11:99–110.
[66] Razandi M, Pedram A, Levin ER. Plasma membraneestrogen receptors signal to antiapoptosis in breast cancer.Mol Endocrinol 2000;14:1434–47.
[67] Acconcia F, Totta P, Ogawa S, Cardillo I, Inoue S, Leone S,et al. Survival versus apoptotic 17�-estradiol effect: role ofER� and ER� activated non-genomic signalling. J CellPhysiol 2005;203:193–201.
[68] Brzozowski AM, Pike ACW, Dauter Z, Hubbard RE, Bonn T,Engstrom O, et al. Molecular basis of agonism andantagoism in the oestrogen receptor. Nature1997;389:753–8.
[69] Paige LA, Christensen DJ, Gron H, Norris JD, Gottlin EB,Padilla KM, et al. Estrogen receptor (ER) modulators eachinduce distinct conformational changes in ER � and ER �.Proc Natl Acad Sci USA 1999;96:3999–4004.
[70] Norman AW, Mizwicki MT, Norman DP. Steroid-hormone
[54] Greene GL, Gilna P, Waterfield M, Baker A, Hort Y, Shine J.Sequence and expression of human estrogen receptorcomplementary DNA. Science 1986;231:1150–4.
[55] Glenney Jr JR. The sequence of human caveolin revealsidentity with VIP21, a component of transport vesicles.FEBS Lett 1992;314:45–8.
[56] Dietzen DJ, Hastings WR, Lublin DM. Caveolin ispalmitoylated on multiple cysteine residues.Palmitoylation is not necessary for localization of caveolinto caveolae. J Biol Chem 1995;270:6838–42.
[57] Acconcia F, Ascenzi P, Fabozzi G, Visca P, Marino M.S-palmitoylation modulates human estrogen receptor-�
functions. Biochem Biophys Res Commun 2004;316:878–83.[58] Acconcia F, Ascenzi P, Bocedi A, Spisni E, Tomasi V,
Trentalance A, et al. Palmitoylation-dependent estrogenreceptor � membrane localization: regulation by17�-estradiol. Mol Biol Cell 2005;16:231–7.
[59] Li L, Haynes MP, Bender JR. Plasma membrane localizationand function of the estrogen receptor � variant (ER46) inhuman endothelial cells. Proc Natl Acad Sci USA2003;100:4807–12.
[60] Pietras RJ, Marquez DC, Chen H-W, Tsai E, Weinberg O,Fishbein M. Estrogen and growth factor receptorinteractions in human breast and non-small cell lungcancer cells. Steroids 2005;70:372–81.
[61] Rai D, Frolova A, Frasor J, Carpenter AE, KatzenellenbogenBS. Distinctive actions of membrane targeted versusnuclear localized estrogen receptors in breast cancer cells.Mol Endocrinol 2005;19:1606–17.
rapid actions, membrane receptors and a conformationalensemble model. Nat Rev Drug Discov 2004;3:27–41.
[71] Migliaccio A, Castoria G, Di Domenico M, de Falco A,Bilancio A, Auricchio F. Src is an initial target of sexsteroid hormone action. Ann NY Acad Sci 2002;963:185–90.
[72] Marsaud V, Gougelet A, Maillard S, Renoir JM. Variousphosphorylation pathways, depending on agonist andantagonist binding to endogenous estrogen receptor �
(ER�), differentially affect ER� extractability,proteasome-mediated stability, and transcriptional activityin human breast cancer cells. Mol Endocrinol2003;17:2013–27.
[73] Reid G, Hubner MR, Metivier R, Brand H, Denger S, ManuD, et al. Cyclic, proteasome-mediated turnover ofunliganded and liganded ER� on responsive promoters isan integral feature of estrogen signaling. Mol Cell2003;11:695–707.
[74] Nonclercq D, Journe F, Body JJ, Leclercq G, Laurent G.Ligand-independent and agonist-mediated degradation ofestrogen receptor-� in breast carcinoma cells: evidence fordistinct degradative pathways. Mol Cell Endocrinol2004;227:53–65.
[75] Han JM, Kim Y, Lee JS, Lee CS, Lee BD, Ohba M, et al.Localization of phospholipase D1 to caveolin enrichedmembrane via palmitoylation: implications for epidermalgrowth factor signalling. Mol Biol Cell 2002;13:3976–88.
[76] Patterson SI. Posttranslational protein S-palmitoylationand the compartmentalization of signaling molecules inneurons. Biol Res 2002;35:139–50.