Embed Size (px)
Citation preview

DELL ASIA CONTINENTAI AMERICANEAFRICANE 1111111911111111111~1POPOLAZIONI CAUCASOIDI
SINO-TIBETANE COREANE ESCHIMO-ALEUTINE AMERINDEFAMIGLIA DI LINGUE INDOEUROPEE KHOISAN NILO-SAHARIANE NIGER-CONGOLESI AUSTRONESIANE INDO-PACIFICHE AUSTRALIANE
Numero 281Gennaio 1992Anno XXVVolume XLVIII
O
Itre quarant'anni fa, nel labora-torio di Sir Ronald A. Fisherall'Università di Cambridge,
dove stavo studiando genetica batterica,l'atmosfera era satura di modelli mate-matici. Non sorprende, dunque, cheproprio allora cominciassi a pensare a unprogetto tanto ambizioso da apparirequasi folle: la ricostruzione del luogod'origine delle popolazioni umane e del-le vie lungo le quali esse si sono poi dif-fuse nel mondo. Secondo me. questa im-presa poteva essere portata a compimen-to misurando quanto stretto fosse il gra-do di affinità delle popolazioni viventi ededucendo da questa informazione unalbero genealogico completo.
Il traguardo è ormai a portata di ma-no. Un'analisi esauriente dei dati digenetica umana raccolti negli ultimi cm-
Gruppi etnici e lingue
LE SCIENZE' SCIENTIFICAMERICAN
di Luigi Luca Cavalli-Sforza
quant'anni e dei nuovi dati ottenuti contecniche messe a punto di recente hapermesso ai miei colleghi e a me di car-tografare la distribuzione di centinaia digeni su scala mondiale. Da questa map-pa abbiamo potuto dedurre le linee filo-genetiche delle popolazioni del mondo.Il nostro albero genealogico concordacon un altro albero che comprende unnumero minore di popolazioni, costruitoa partire da dati genetici fondamental-mente diversi. Inoltre la nostra ricostru-zione trova parallelismi sorprendenti inuna recente classificazione delle lingue.Pertanto i geni, le popolazioni e le linguesi sono irradiati parallelamente median-te una serie di migrazioni che hanno avu-to origine, a quanto pare, in Africa e sisono poi diffuse attraverso l'Asia in Eu-ropa, nel Nuovo Mondo e nel Pacifico.
Il concetto di albero genealogico haun'importanza fondamentale per collo-care tali eventi in una sequenza cronolo-gica. Se gli altri fattori sono uguali,quanto più lungo è il periodo di tempotrascorso dalla separazione di due popo-lazioni, tanto maggiore dovrebbe esserela differenza (o distanza) genetica traqueste ultime. L'analisi potrebbe al-lora essere utilizzata anche per ricostrui-re una storia più complessa, riguardantetre o più popolazioni.
Talvolta per le popolazioni umane siricorre alla denominazione di gruppi et-nici o «razze», anche se il razzismo hareso il secondo termine alquanto dete-stabile. Le popolazioni umane sono dif-ficilmente definibili in un modo che sianel contempo rigoroso e utile perché gliesseri umani si raggruppano in una sba-lorditiva varietà di sistemi - alcuni dei
quali si sovrappongono parzialmente -tutti in continua trasformazione. Le lin-gue ci danno, però, qualche aiuto.
La specie umana, per buona parte del-la sua storia e in alcuni casi ancora oggi,è stata organizzata in tribù di piccole di-mensioni, gruppi di persone con un le-game di parentela piuttosto stretto. Inol-tre vi è spesso una corrispondenza biu-nivoca tra lingua e tribù. Le lingue offro-no così una guida sommaria alla tribù diappartenenza e questa, quando la si co-nosce, permette una classificazione agrandi linee delle popolazioni.
Dato che la situazione è molto piùcomplicata nelle società metropolitane,abbiamo limitato i problemi focalizzan-do il nostro studio sulle popolazioni abo-rigene, cioè su quelle popolazioni cheoccupavano i loro territori attuali già pri-ma delle grandi ondate migratorie avve-nute in seguito ai viaggi di scoperta delRinascimento. Le distanze genetiche traquesti gruppi aborigeni non si possonoricavare dalla presenza o dall'assenza diun unico carattere ereditario, o del geneche lo esprime, perché ogni gruppo con-tiene in pratica tutti i geni umani ancoraesistenti; ciò che varia è invece la fre-quenza con cui si manifestano i geni.
Un buon esempio viene fornito dal va-sto insieme di dati sul fattore Rh,
un antigene del sangue umano, che vieneereditato come Rh positivo o negativo inmodo semplice e diagnosticabile in labo-ratorio. Per la sua importanza in medi-cina, esso è stato studiato in migliaia dipopolazioni. I medici devono identifica-re le gestanti Rh-negative il cui feto siaRh-positivo, e sottoporle subito dopo ilparto a un trattamento immunologicoche impedisca loro di produrre anticorpiche potrebbero danneggiare i figli con-cepiti successivamente. I geni per il fat-tore Rh-negativo sono frequenti in Eu-ropa, rari in Africa e nell'Asia occiden-tale e pressoché assenti nell'Asia orien-tale e tra le popolazioni aborigene del-l'America e dell'Australia (si veda l'illu-strazione a pagina 20).
Si può stimare l'affinità genetica sot-traendo, per esempio, la percentuale di
individui Rh-negativi tra gli inglesi (16per cento) dalla stessa percentuale tra ibaschi (25 per cento), trovando una dif-ferenza di nove punti percentuali, ma tragli inglesi e gli abitanti dell'Asia orienta-le la differenza diventa di 16 punti, unadistanza maggiore che forse sottintendeuna separazione più antica dei due grup-pi. Non vi è così nulla di eccezionale nelconcetto di distanza genetica.
In realtà i genetisti usano formule unpoco più complicate della semplice sot-trazione, in modo che le distanze possa-no fornire il più alto contenuto di infor-mazioni circa la storia evolutiva umana.Per esempio, se un'unica popolazione siframmentasse in varie parti completa-mente separate l'una dall'altra, tali sot-topopolazioni si differenzierebbero an-che in assenza di mutazioni e di selezionenaturale (si veda l'articolo La geneticadelle popolazioni umane di L. L. Caval-li-Sforza in «Le Scienze» n. 79, marzo1975). Il semplice caso può modificare lerispettive frequenze geniche di questesottopopolazioni nel processo chiamato«deriva genetica».
Qualora gli altri fattori evolutivi (di-versi dalla deriva genetica) si mantenes-sero immutati, la distanza genetica au-menterebbe in modo semplice e regolarecon il passare del tempo. Quanto piùtempo è trascorso dalla separazione didue popolazioni, tanto maggiore è la di-stanza genetica tra di esse. Pertanto ladistanza genetica potrebbe fungere daorologio con cui datare la storia evoluti-va. Considerazioni statistiche, tuttavia,mettono in evidenza l'impossibilità daparte di un singolo gene, come quelloper il fattore Rh, di fornire una cronolo-gia precisa. Nel calcolo delle distanze ge-netiche e, idealmente, nella successivaverifica delle conclusioni con differentigruppi di geni è essenziale servirsi dellemedie di molti geni. Per fortuna i geninoti sono migliaia, anche se soltanto unapiccola percentuale è stata esaminata indiverse popolazioni.
La ricostruzione degli alberi genealo-gici a partire dalle distanze genetiche sibasa su numerosi principi. Un esempioè fornito da un albero genealogico che
considera 15 popolazioni, pubblicato 27anni fa da Anthony W. F. Edwards, oggiall'Università di Cambridge, e da me.Esso è stato ricavato a partire da distan-ze calcolate dalle informazioni geneticheallora disponibili, secondo la formula diEdwards del «cammino genetico mini-mo». Essenzialmente, questo metodopermette di ricavare l'albero che hala minima lunghezza totale dei rami.Quando quest'albero viene proiettato suuna mappa del globo, in modo che i pun-ti di ramificazione coincidano con le areeabitate oggi dalle popolazioni in esame,lo schema che ne emerge coincide grossomodo con le ricostruzioni antropologi-che di antiche migrazioni (si veda l'illu-strazione a pagina 21 in alto).
Purtroppo non disponiamo di alcunaprova incontrovertibile che la formuladel cammino genetico minimo offra ilmodo migliore per adattare un alberofilogenetico ai dati. Altri metodi di co-struzione degli alberi possono essere piùsoddisfacenti nel correlare la lunghezzadei rami al tempo trascorso e nel trovareuna «radice» databile dell'albero stesso(si veda l'illustrazione a pagina 21 in bas-so). Quando è possibile, la radice mettein rapporto le popolazioni a un gruppoesterno, per esempio gli scimpanzé chesi sarebbero distaccati dalla linea evolu-tiva che ha condotto agli esseri umani tracinque e sette milioni di anni fa. Se siassume che la velocità del cambiamentoevolutivo sia costante lungo tutti i rami,si possono equiparare le lunghezze deirami al tempo trascorso dal momento incui si sono formati. Tuttavia, questi al-beri con «radice» (punto di origine) pos-sono dare indicazioni fuorvianti se in al-cuni rami il cambiamento evolutivo èstato più rapido che in altri.
Le tecniche matematiche della geneti-ca delle popolazioni possono ridurre alminimo le distorsioni, permettendo diprevedere con esattezza le velocità dievoluzione. Il modello evolutivo che ab-biamo scelto è il più semplice. Prevedeche i rami evolvano con uguale velocità,ammesso che la deriva genetica sia lacausa principale del cambiamento e chele varie popolazioni abbiano, in media,
Geni, popolazioni e lingueL'albero genealogico delle popolazioni umane corrisponde all'alberodelle lingue parlate nel mondo: entrambi presuppongono migrazioni che,a quanto indicano i dati biologici, avrebbero avuto origine in Africa
MEDITERRANEI NORDEUROPEI
INDIANI
JU/HUA
MASAI
PIGMEI
CINESI
TIBETAN COREANI
INUIT
AZTECHI
YANOMAMI POLINESIANI
MAORI
MELANESIANI AUSTRALIANIBOSCIMANI
18 LE SCIENZE n. 281, gennaio 1992
LE SCIENZE n. 281, gennaio 1992 19

La mappa genica mostra che il fattore Rh negativo è assai comune tra i baschi e meno co-mune andando verso est. Tali dati indicano che i baschi conservano tracce di una primitivapopolazione europea successivamente mescolatasi con gruppi etnici provenienti dall'Asia.
Geni e reperti archeologici narrano storie simili. Il più antico alberogenetico (in rosso) è proiettato su una mappa: le estremità dei ramisono poste nelle patrie attuali (dischi rossi). Ricerche genetiche più
recenti indicano due possibili vie migratorie dall'Africa verso l'Asia(in giallo); i particolari di tali vie sono ipotetici. I numeri rap-presentano le stime archeologiche sulle date dei primi insediamenti.
I NUMERI SI RIFERISCONO AL TEMPO TRASCORSO.IN ANNI, DAL PRIMO INSEDIAMENTO
1-4 4-9 9-16 16-25 25+I NUMERI SI RIFERISCONO ALLEPERCENTUALI DI INDIVIDUI Rh-NEGATIVI
0-1
POPOLAZIONE 1
POPOLAZIONE 2
I i i i 0,20 0,15 0,10 0,05
DISTANZA GENETICA
POPOLAZIONI
DELLA NUOVA GUINEA E DELL'AUSTRALIA
DELLE ISOLE DEL PACIFICO
DELL'ASIA SUDORIENTALE
DELL'ASIA NORDORIENTALE
DELL'ASIA NORDORIENTALE ARTICA
AMERINDE
EUROPEE
CAUCASOIDI NON EUROPEE
AFRICANE
0,00
TEMPO (ANNI)
0 500 1000 1500 2000 2500
0 10 20 30 40 50 60 70 80 90 100
TEMPO (GENERAZIONI)
uJ
D 100
(S 80ccuJ
o2u i 40
20LL,
O O
CZ
60
Cambiamenti nei geni producono, al passare del tempo, una diffe-renziazione genetica come quella che si rispecchia nell'albero fi-logenetico rappresentato qui sopra. Un modello della deriva ge-netica (a destra), che è uno dei meccanismi evolutivi secon-
do cui avviene il cambiamento, può essere ottenuto al calcolatore.Quando due metà di una popolazione si separano, i geni portati daognuna hanno frequenze in un primo momento uguali, ma il tempoe il caso possono finire per farle evolvere in direzioni opposte.
la stessa dimensione. Prove indipenden-ti confermano la prima affermazione,mentre una selezione ragionata delle po-polazioni rende probabile la seconda.Quando le popolazioni sono grandi e vi-vono in territori che occupano intericontinenti e per periodi di tempo che siestendono fino all'insediamento origina-rio, è verosimile che le velocità di evolu-zione siano costanti.
Con i colleghi Paolo Menozzi e Alber-to Piazza, rispettivamente delle Univer-sità di Parma e di Torino, ho messo apunto un protocollo analitico comunenell'ambito del quale si possono studiarela storia e la geografia dei geni umani.In un progetto realizzato nell'arco di 12anni, abbiamo studiato la mole di infor-mazioni genetiche che si è accumulatanegli ultimi cinquant'anni: più di 100 dif-ferenti caratteri ereditari ricavati da cir-ca 3000 campioni prelevati da 1800 po-polazioni. La maggior parte dei campio-ni includeva centinaia, se non addiritturamigliaia di individui. Questo insieme didati, cui abbiamo dato il nome di insie-me «classico», è derivato indirettamentedalle proteine prodotte dai geni.
Inoltre abbiamo di recente sviluppatoun secondo insieme, totalmente nuovo:si tratta di dati molecolari studiati diret-tamente nelle sequenze codificate delDNA contenuto nel nucleo cellulare. Lamaggior parte di questi dati da noi uti-lizzati è stata raccolta in uno studio set-tennale, realizzato in collaborazione trail mio gruppo di ricerca alla StanfordUniversity e quello di Kenneth K. e Ju-dith R. Kidd del Dipartimento di gene-tica della Yale University. Anche se que-sti dati sono, sotto molti aspetti, di qua-lità superiore rispetto a quelli che si ba-sano sui prodotti genici, finora essi ri-guardano soltanto un centesimo dellepopolazioni studiate con il metodo pre-cedente. Tuttavia, in ogni confronto cheabbiamo effettuato finora, i dati mole-colari concordano in modo eccellentecon i dati classici.
I l nostro primo risultato suffraga una conclusione emersa da studi su reperti
fossili umani e su manufatti: l'origineafricana della nostra specie. Abbiamotrovato che le distanze genetiche tra afri-cani e non africani superano quelle os-servate in altri confronti fra popolazionidi continenti diversi. Questo risultato èesattamente quello che ci si aspetterebbese, nell'albero filogenetico umano, la se-parazione dell'Africa fosse la prima e lapiù antica.
La distanza genetica tra africani e nonafricani è, in prima approssimazione, ildoppio di quella tra australiani e asiaticie quest'ultima è più del doppio di quellatra europei e asiatici. I tempi di separa-zione corrispondenti indicati dalla pa-leoantropologia hanno analoghi rappor-ti: 100 000 anni per la separazione traafricani e asiatici, circa 50 000 anni perquella tra asiatici e australiani e da35 000 a 40 000 anni per quella tra asia-
tici ed europei. In questi casi, almeno, lenostre distanze genetiche servono comeun orologio di discreta qualità.
Un orologio diverso e raffinato erastato escogitato da altri ricercatori chestudiavano dati genetici di tipo fonda-mentalmente diverso rispetto ai nostri. Iloro risultati più interessanti divennerodisponibili solo quando avevamo quasicompletato la nostra analisi, ma essi han-no confermato le nostre conclusioni intutti i punti essenziali. Il sistema geneti-co da loro adottato riguarda il numerorelativamente piccolo di geni codificatinel DNA dei mitocondri, organelli cel-lulari che presiedono alla produzionedell'energia necessaria alla cellula. An-che noi avevamo studiato inizialmentequesti geni, ma il compianto Allan C.Wilson e collaboratori dell'Universitàdella California a Berkeley hanno svi-luppato metodi dotati di una migliore ri-soluzione. (Posso qui ricordare solo al-cuni importanti contributi di Wilson nelcampo dell'evoluzione molecolare. Egliè morto di leucemia acuta nel luglio1991, all'età di 56 anni.)
I geni mitocondriali differiscono daquelli presenti nel nucleo per aspetti fon-damentali. I geni del nucleo derivano inmaniera uguale dal padre e dalla madre,mentre quelli mitocondriali vengono tra-smessi alla prole quasi esclusivamentedalla madre. Questa forma semplice dieredità rende i geni mitocondriali moltoconvenienti per poter stimare le distanzegenetiche. Essi hanno anche velocità dimutazione più elevate dei geni del nu-cleo, sicché si può in parte modificare la
determinazione statistica delle distanzegenetiche calcolandole non a partire dal-le frequenze geniche, ma dalle mutazio-ni nei geni stessi.
L'orologio mitocondriale si basa sulnumero di mutazioni accumulate anzi-ché sui cambiamenti nelle frequenze ge-niche. Mentre, secondo la nostra ipote-si, le frequenze geniche si sarebbero mo-dificate a velocità costanti, il gruppo diWilson postulò che fossero stati i genimitocondriali a mutare a velocità costan-ti. Grazie alla natura di questi dati, è piùfacile determinare la radice di un alberomitocondriale che quella di uno nuclea-re. Basta confrontare il proprio alberocon un campione di riferimento (il grup-po di Wilson faceva ricorso agli scimpan-zé) di cui sia nota la data, o l'intervallodi date, di divergenza dall'insieme dellepopolazioni in esame.
Sulla base di queste distanze il gruppodi Wilson ha ricavato un albero genea-logico che mostra in Africa una differen-ziazione maggiore rispetto a qualsiasi al-tra parte del mondo. Questo risultato hamesso in evidenza che il DNA mitocon-driale umano si è evoluto per la maggiorparte del tempo in Africa, cioè esso puòessere fatto risalire a un'unica donnaafricana. Inoltre è stato possibile datarei punti di ramificazione dell'albero con-frontando i DNA degli esseri umani edegli scimpanzé, le cui linee di discen-denza si sono distaccate l'una dall'altracirca cinque milioni di anni fa. Avendocalibrato in questo modo il loro albero,i ricercatori del gruppo di Wilson hannopotuto stimare le date delle ramificazio-
ni successive e, risultato della massimaimportanza, sono stati in grado di valu-tare che la donna africana «capostipite»visse tra 150 000 e 200 000 anni fa, con-fermando così le nostre conclusioni conmezzi del tutto indipendenti.
Di recente gli studiosi hanno spostatoun poco più avanti la data stimata, ma laloro donna africana è ancora precedentela data che noi attribuiamo alla divergen-za delle popolazioni africane e asiatiche.E, in effetti, ella dovrebbe essere più an-tica. Le due date si riferiscono a eventidiversi: la nascita di una singola donna e
la frammentazione di una popolazionealla quale quella donna apparteneva. Imass media hanno confuso le cose facen-do abbondantemente circolare, a propo-sito di questa donna, il nome di «Eva».Di fatto, non abbiamo alcuna prova chevi sia mai stato un tempo in cui una soladonna vivesse sulla Terra. Molte altredonne potrebbero essere vissute nel me-desimo lasso di tempo, ma le loro lineedi discendenza mitocondriali si sarebbe-ro semplicemente estinte.
Alcune di queste conclusioni riman-gono controverse. Anche se i paleo-
antropologi concordano che il genereHomo si sia originato in Africa circa 2,5milioni di anni fa e che la testimonianzafossile dell'Homo sapiens anatomica-mente moderno risalga solo intorno a100 000 anni fa in Africa o nelle zonecircostanti, non tutti accettano la teoriadella «diffusione dall'Africa». Alcuni ri-cercatori sostengono che esseri umani ditipo moderno comparvero in epoca mol-to più antica e contemporaneamente indiverse popolazioni del Vecchio Mondo(si veda l'articolo La comparsa dell'uo-mo moderno di Christopher B. Stringer
20 LE SCIENZE n. 281, gennaio 1992
LE SCIENZE n. 281, gennaio 1992 21
Correlazione tra popolazioni e lingue
GENETICA POPOLAZIONI FAMIGLIE LINGUISTICHE
PIGMEI MBUTI Maar
AFRICANI OCCIDENTALI Wffiffilni,BANTU IMMIliff"""
NILOTICI 111IVLIIIIIIIIIIW I
SAN (BOSCIMANI) XXXXXXXXXXXXXXX XX
ETIOPICI
BERBERI. NORDAFRICANI 'IMMIII MIIMlSIATICI SUDOCCIDENTALI rffffmnf I
(LINGUA ORIGINARIA SCONOSCIUTA)
NIGER-KORDOFANIANA
NILO-SAHARIANA
KHOISAN
AFROASIATICA
EUROPEI
SARDI
INDIANI EVVWWW0
U.M,\nn,\IRANIANI
0111WIM11LWMI‘ INDOEUROPEA
INDIANI SUDORIENTALI XXXXXXXXXXXXXXXXX
rì
AMERINDI MERIDIONALI
AMERINDI CENTRALI
MERINDI SETTENTRIONALI
ERINDI NORDOCCIDENTALIIIMAIIMMILM
DRAVIDICA
<(DO
SINO-TIBETANA CCCL Hu_1(130_0D ZCr)
ALTAICA
ESCHIMO-ALEUTINA
CHUKCHI-KAMCHADAL
AMERINDA •
NA-DENE
LAPPONI mumemiSAMOIEDI
MONGOLI
TIBETANI
COREANI
GIAPPONESI
AINU
SIBERIANI
ESCHIMESI
CHUKCHI
Immuneummumi
II III 1/4
'1111I
1:«0:4:070:0:0:0:01
URALICA
CITALIc)9,
\5)•-• \AS3LARIcp
(155-
CINESI MERIDIONALI
MON KHMER
THAILANDESI
INDONESIANI
MALAYSIANI
FILIPPINI
POLINESIANI
MICRONESIANI
MELANESIANI
PAPUANI
AUSTRALIANI WDISTANZA GENETICA
AUSTRALIANA
Classificazione linguistica da Merritt Ruhlen, A Guide to the World's Languages0,030 0,024 0,018 0,012 0,006
C.)
co pP p
=-1r - INDO-PACIFICA
SINO-TIBETANA
AUSTROASIATICA
DAICA
AUSTRONESIANA
in «Le Scienze» n. 270, febbraio 1991).Non solo abbiamo localizzato i primis-
simi uomini moderni in Africa, ma ab-biamo anche individuato le tracce di unaserie di ondate migratorie. Tale quadroci fornisce molte informazioni sull'ori-gine delle popolazioni attuali. Inoltre ilcollegamento tra il nostro lavoro e quel-lo dei linguisti e degli archeologi promet-te di rivelare informazioni ancora piùdettagliate.
In generale, i processi migratori riflet-tono cambiamenti che possono essere vi-sti nel contempo come pressioni e comeopportunità. A più riprese gli esseriumani e gli ominidi loro progenitori han-no subito un notevole incremento nume-rico e hanno potuto così espandersi geo-graficamente. Questo successo demo-grafico sarebbe soprattutto il risultato disviluppi culturali che, per il periodo pre-istorico, devono potersi dedurre dalladocumentazione archeologica. Questafonte, costituita per la maggior parte daossa e da utensili in pietra, indica chel'Africa è stata effettivamente la terrad'origine degli ominidi. Dall'Africa, lemigrazioni hanno proceduto verso l'Asiaattraverso l'istmo di Suez e, in seguito,dall'Asia in Europa. In queste regioni gliominidi si insediarono forse un milionedi anni fa.
Lo stadio successivo è più difficile dadocumentare perché dipende dall'epocain cui si pensa che gli uomini moderni sisiano separati dagli ominidi. Comunqueè chiaro che questo distacco si era giàverificato quando gli esseri umani si dif-fusero dall'Asia alle Americhe, un even-to che aveva dovuto attendere il prosciu-gamento dello Stretto di Bering e un cli-ma sufficientemente mite da permettereil passaggio via terra. Anche l'insedia-mento in Australia e nelle isole del Pa-cifico deve essersi realizzato solo di re-cente, una volta acquisita una certa pa-dronanza della navigazione marittima.
Sembra che l'Australia sia stata occu-pata da popolazioni provenienti dall'A-sia sudorientale perlomeno 40 000 annifa e forse anche da 10 000 a 20 000 anniprima di questa data. Gli archeologi so-no, invece, discordi sulla data del primoingresso nelle Americhe. Finora i primidocumenti pienamente convincenti del-la presenza di esseri umani in Alaska ri-salgono a circa 15 000 anni fa, ma sem-bra che alcuni siti in America Meridio-nale risalgano a epoche precedenti. Per-tanto le stime vanno da 15 000 a 35 000anni fa. I nostri dati relativi ai geni pre-senti nel nucleo cellulare fanno ipotizza-re un'inizio dell'insediamento attorno a30 000 anni fa.
L'Europa è stata attraversata da molteondate migratorie, che però non hannocancellato le vestigia dei primissimi inse-diamenti. Una chiave di questo enigmaè stata proposta nel 1954 da Arthur E.Mourant, uno dei primi studiosi di «geo-grafia dei geni», allora al Population Ge-netics Laboratory del Medical ResearchCouncil a Londra. Secondo la sua ipote-
si, i baschi (popolazione della Spagnasettentrionale e della Francia sudocci-dentale), i più antichi abitanti dell'Euro-pa, avrebbero conservato parte della co-stituzione genetica originaria malgrado icontatti successivi. Questa teoria è soste-nuta da dati sul gene per il fattore Rhnegativo: nei baschi questo gene ha unaincidenza superiore a quella di qualsiasialtra popolazione al mondo. Ricerchecompiute su molti altri geni confermanol'ipotesi, così come gli studi sulle profon-de differenze tra la lingua dei baschi equelle delle popolazioni vicine.
Un'analisi recente delle variazioni ge-niche nelle diverse regioni d'Europa hapermesso di proporre un modello di co-me sarebbero avvenuti gli insediamentiin tutta Europa. In base a esso, i primicoltivatori del Neolitico avrebbero por-tato i loro geni, la loro cultura e le lingueindoeuropee dal Medio Oriente all'Eu-ropa in un processo di lenta espansione(si veda l'articolo Le origini delle lingueindoeuropee di Colin Renfrew in «LeScienze» n. 256, dicembre 1989). Datoche gli antenati dei baschi vivevano nellaregione più lontana dall'origine di que-sto movimento di espansione, essi si so-no con tutta probabilità mescolati mini-mamente con i coltivatori.
Si può soltanto sperare di identificareinsediamenti che riflettano le migrazioniche hanno avuto successo. Potrebberoesservi state anche migrazioni genetica-mente insignificanti. Nelle Americhe,per citare un esempio molto recente,i vichinghi stabilirono insediamenti dibreve durata, ma il contributo che po-trebbero aver dato al patrimonio genicolocale non è valutabile.
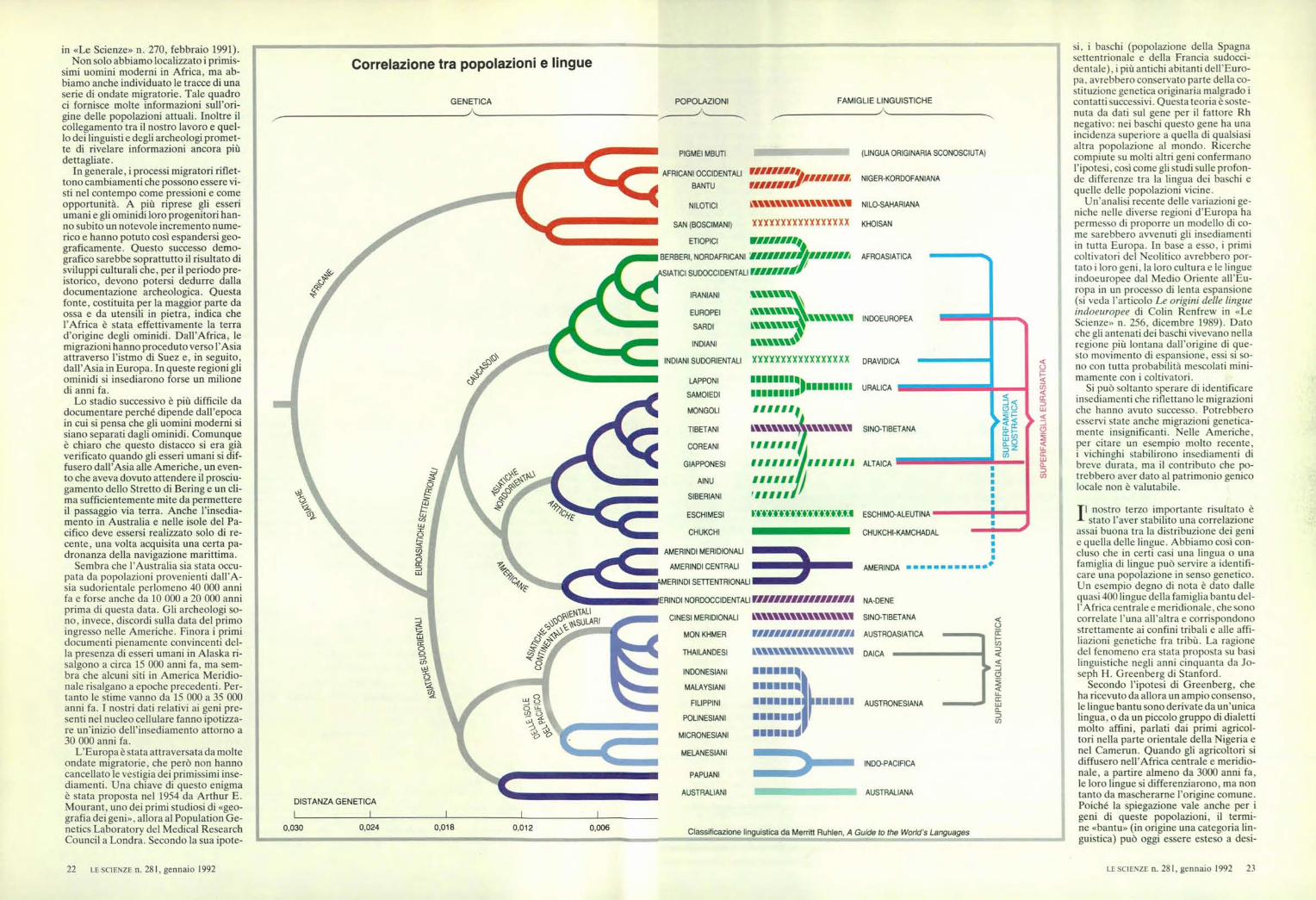
Il nostro terzo importante risultato èstato l'aver stabilito una correlazione
assai buona tra la distribuzione dei genie quella delle lingue. Abbiamo così con-cluso che in certi casi una lingua o unafamiglia di lingue può servire a identifi-care una popolazione in senso genetico.Un esempio degno di nota è dato dallequasi 400 lingue della famiglia bantu del-l'Africa centrale e meridionale, che sonocorrelate l'una all'altra e corrispondonostrettamente ai confini tribali e alle affi-liazioni genetiche fra tribù. La ragionedel fenomeno era stata proposta su basilinguistiche negli anni cinquanta da Jo-seph H. Greenberg di Stanford.
Secondo l'ipotesi di Greenberg, cheha ricevuto da allora un ampio consenso,le lingue bantu sono derivate da un'unicalingua, o da un piccolo gruppo di dialettimolto affini, parlati dai primi agricol-tori nella parte orientale della Nigeria enel Camerun. Quando gli agricoltori sidiffusero nell'Africa centrale e meridio-nale, a partire almeno da 3000 anni fa,le loro lingue si differenziarono, ma nontanto da mascherarne l'origine comune.Poiché la spiegazione vale anche per igeni di queste popolazioni, il termi-ne «bantu» (in origine una categoria lin-guistica) può oggi essere esteso a desi-
22 LE SCIENZE n. 281, gennaio 1992 LE SCIENZE n. 281, gennaio 1992 23

SONO DISPONIBILII RACCOGLITORI PER IL
1992DELLA RIVISTA
LE SCIENZEQuesti raccoglitori corrispondono ai volumi XLVIII e XLIX
de LE SCIENZE e rispettivamente ai fascicoli da gennaio (n. 281)
a giugno (n. 286) e da luglio (n. 287) a dicembre (n. 292).
Sono ancora disponibili i raccoglitori dal Vol. XL al XLVII e
raccoglitori appositamente non numerati per sostituire quelli esauriti.
I raccoglitori si possono richiedere direttamente all'editore usando
l'apposita cartolina allegata alla rivista e unendo il relativo importo;
gli ordini vengono evasi solo a pagamento avvenuto.
Ogni raccoglitore L. 6.500
I raccoglitori si trovano anche presso i seguenti punti vendita:
BOLOGNA- Libreria Parolini - Via U. Bassi 14FIRENZE - Libreria Marzocco - Via de' Martelli 22/R
GENOVA - Libreria Int. Di Stefano - Via R. Ceccardi 40/RMILANO - Le Scienze S.p.A. - Piazza della Repubblica 8TORINO - Libreria Zanaboni - C.so Vittorio Emanuele 41
NAPOLI - Libreria Guida A. - Via Port'Alba 20/21PADOVA- Libreria Cortina - Via F. Marzolo 4
PALERMO - Libreria Dante - Quattro Canti di CittàROMA- Libreria Feltrinelli - Via V. Emanuele Orlando
LE SCIENZESCIENTIFIC
ANERICAN
Un campione di sangue viene prelevato dall'autorea un membro della tribù Aka dei pigmei africani.
11F
gnare un gruppo di popolazioni che han-no una base comune sia genetica sialinguistica.
Nel 1988 i miei collaboratori e io ab-biamo pubblicato un albero geneticodelle origini evolutive di 42 popolazioni,assieme alle rispettive affiliazioni lingui-stiche, che dimostra come il raggrup-pamento genetico delle popolazioni nelmondo coincida strettamente con quellodelle lingue. Con pochissime eccezio-ni, sembra che le famiglie linguisticheabbiano, nel nostro albero geneti-co, un'origine abbastanza recente.Inoltre tentativi da poco compiutida due gruppi di linguisti di gene-rare categorie linguistiche di livellopiù elevato («superfamiglie») han-no dato risultati anch'essi in lineacon i dati genetici, del tutto indi-pendenti. E stato emozionante tro-vare che avevamo confermato un'i-potesi avanzata nientemeno che daun pioniere come Charles Darwin,il quale, nel capitolo 14 di L'originedelle specie per selezione naturale,sosteneva che, se si fosse conosciu-to l'albero dell'evoluzione geneti-ca, si sarebbe potuto prevederequello dell'evoluzione linguistica.
Perché l'evoluzione genetica el'evoluzione linguistica dovrebberoavere una così stretta corrispon-denza? La risposta sta non nel de-terminismo genetico, ma nella sto-ria: i geni non controllano la lingua,ma sono piuttosto le circostanzedella nascita a determinare le lin-gue alle quali l'individuo è esposto.Le differenze linguistiche possonogenerare o rafforzare le barrieregenetiche tra popolazioni, ma è impro-babile che siano la causa principale dellacorrelazione. L'evoluzione umana è co-stellata dalla frammentazione di popola-zioni in parti, alcune delle quali vanno ainsediarsi altrove. Ogni parte è soggettaa un'evoluzione linguistica e geneticache porta le tracce dei punti di ramifica-zione comuni. Pertanto è inevitabile checi sia qualche correlazione.
Si può obiettare che una separazionecompleta, come quella che si ha quandoun gruppo staccatosi da una popolazionemigra in un nuovo continente, deve es-sere un evento raro. Ma per isolare duepopolazioni non è necessaria la frappo-sizione di oceani o catene montuose: ba-sta la semplice distanza, come dimostra-no gli studi genetici compiuti su moltespecie animali. Dato che l'interscambioche si ha con le migrazioni è normalmen-te più grande a brevi distanze, ci si aspet-ta, e si trova, un grado tanto più elevatodi differenza genetica quanto più distan-ziate sono tra loro le popolazioni; lo stes-so accade per le lingue. Quando non visiano barriere particolari, le variazionisia genetiche sia linguistiche tendono aessere continue, mentre la discontinuitàcompare in entrambe quando esistanobarriere alla libera migrazione.
Si dovrebbe richiamare l'attenzione su
due tipi di eccezioni alla nostra regoladella corrispondenza tra geni e lingue: leeccezioni in cui vi è una sostituzione dilingua e quelle in cui vi è una sostituzionedi geni. Le prime si hanno quando ungruppo di persone sostituisce una nuovalingua alla propria lingua di origine, sot-to l'influsso, per esempio, di immigranti,o di conquistatori, o di una nuova éliteculturale. Tuttavia questo genere di so-stituzioni non avviene sempre ed è menoprobabile che accada quando la nuova
lingua deriva da una famiglia linguisticadiversa. La lingua basca è un caso estre-mo di relitto linguistico evidentemen-te sopravvissuto a millenni di continuisconvolgimenti linguistici nelle regioniconfinanti.
La sostituzione dei geni, generalmen-te parziale, avviene quando una popola-zione si mescola con un'altra. Il rimesco-lamento può essere assolutamente gra-duale e influenzare la frequenza relativadi tutti i geni in uguale proporzione.Questa gradualità distingue in manieranetta i geni dalle lingue che, per princi-pio, o sono sostituite o non lo sono af-fatto. Una lingua conserva l'integrità diorigine anche se adotta un gran numerodi parole di un'altra famiglia o sottofa-miglia linguistica. I linguisti concordano,per esempio, sul fatto che l'inglese ri-manga una lingua della sottofamigliagermanica, malgrado le parole di deriva-zione francese, greca e latina. Ciò cheimporta è che la struttura e il vocabo-lario di base conservino i caratteri dellafamiglia.
Questa differenza significa che, quan-do una ristretta minoranza impone lapropria lingua alla maggioranza che haconquistato, la sostituzione della linguaè quasi completa, mentre la sostituzionedei geni è proporzionale al rapporto de-
mografico. Per esempio gli ungheresiparlano una lingua originaria degli Urali(catena montuosa che divide l'Europadall'Asia) imposta dai conquistatori ma-giari nel Medioevo, mentre hanno unacostituzione genetica tipicamente euro-pea. Solo con qualche difficoltà è possi-bile scoprire tracce di geni magiari nellapopolazione ungherese contemporanea.
La sostituzione genica su vasta scala èforse più rara, ma nella nostra coppiadi alberi complementari si può notare
almeno un caso probabile: quellodei lapponi, o Saame , della Scan-dinavia settentrionale. Anche la lo-ro lingua appartiene alla famigliauralica, mentre la loro struttura ge-netica fa pensare a un miscuglio trale popolazioni mongoloidi della Si-beria e gli scandinavi, che sono re-sponsabili della maggioranza deigeni. Il rimescolamento genico èevidente anche nei capelli e nel co-lorito dei lapponi, molto variabilidal chiaro allo scuro. Una situazio-ne non dissimile da quella dei lap-poni si ha tra gli etiopi, miscugliogenetico di africani e di caucasoidiprovenienti dall'Arabia, con pre-dominanza dei primi.
Anche un modesto flusso di genipuò produrre effetti rilevanti se siprolunga per un tempo sufficiente.Un esempio classico è quello degliafroamericani , il cui patrimonio ge-nico deriva oggi per il 30 per centoin media da individui di origine eu-ropea. Questo risultato si spieghe-rebbe se a ogni generazione il 5 percento di tutte le unioni dei neri fos-se avvenuto con europei fin dagli
albori della schiavitù in America e se tut-ti i discendenti di queste unioni fosserostati classificati come neri. Altri 1000 an-ni di questo processo lascerebbero benpoco dell'originario genoma africano.
può forse sorprendere che la prevista
correlazione tra lingue e geni per-manga nel tempo in misura così rilevantenonostante il rimescolamento causatodalle sostituzioni di geni o di lingue. Inparte ciò può essere dovuto al fatto checi siamo concentrati sulle popolazioniaborigene. In ogni caso, altre analisiconfermano oggi l'esistenza di questacorrelazione a livello microgeografico,talvolta in modo superiore alle attese.Forse l'esempio più sorprendente è datodalla stretta concordanza tra la nostraanalisi genetica degli indigeni americanie la recente classificazione in tre famiglieprincipali delle lingue del Nuovo Mondoa opera di Greenberg. I due studi hannoproceduto in maniera indipendente, uti-lizzando dati molto diversi; eppure l'unoe l'altro indicano con grande evidenzache nelle Americhe si è verificato un nu-mero esiguo di migrazioni distinte.
La spiegazione definitiva di questacorrelazione tra geni e cultura deve es-sere ricercata nei due meccanismi di tra-smissione: orizzontale e verticale. I geni,
trasmessi sempre dai genitori alla prole,descrivono una traiettoria verticale at-traverso le generazioni. La cultura puòa sua volta essere trasmessa di genera-zione in generazione, ma, diversamentedai geni, può esserlo anche orizzontal-mente, tra individui che non hanno rap-porti di parentela. Per esempio, l'altamoda è generalmente trasmessa da Pa-rigi al resto del mondo ogni stagione (an-che se pare che sia ora l'Italia ad assu-mere la guida). Nel mondo contempora-neo, la trasmissione orizzontale sta di-ventando sempre più importante. Ma lesocietà tradizionali sono così chiamateproprio perché le loro culture (e gene-ralmente le loro lingue) si conservano evengono trasmesse da una generazionea quella successiva. La trasmissione pre-valentemente verticale della cultura contutta probabilità rende queste societàmolto più conservatrici.
Le sostituzioni di geni e di lingue co-stituiscono qualcosa di più che sempli-ci e fastidiose eccezioni ai nostri schemiinterpretativi. Ogni eccezione opera se-condo norme proprie, che dovrebberodirci molto sull'evoluzione delle popola-zioni e delle lingue e quindi sullo svilup-po della cultura umana. Gli studi di que-ste sostituzioni potrebbero fornire infor-mazioni complementari a quelle che giàsono state offerte dal nostro lavoro. Lericerche antropologiche sul campo devo-no confrontarsi con questi strumenti diindagine e con dati che stanno rapida-mente svanendo. Documenti di incalco-labile valore ci sfuggono di mano con laperdita progressiva di identità da partedelle popolazioni aborigene. L'interessecrescente nel Progetto genoma umanopuò, d'altra parte, stimolare i ricercatoria raccogliere testimonianze della diver-sità genetica umana prima che questascompaia.
BIBLIOGRAFIA
CAVALLI-SFORZA L. L. e FELDMAN MARCW., Cultura! Transmission and Evolu-tion: A Quantitative Approach, Prince-ton University Press, 1981.
CAVALLI-SFORZA L. L., PIAZZA A., ME-NOZZI P. e MOUNTAIN J. L., Reconstruc-tion of Human Evolution: Bringing To-gether Genetic, Archaeological and Lin-guistic Data in «Proceedings of the Na-tional Academy of Sciences», 85, n. 16,agosto 1988.
BOWCOCK A. M., K1DD J. R., MOUNTAINJ. L., HEBERTJ. M., CAROTENUTO L., KIDDK. K. e CAVALLI-SFORZA L. L., Drift,Admixture and Selection in Human Evo-lution: A Study with DNA Polymor-phisms in «Proceedings of the NationalAcademy of Sciences», 88, n. 3, 1 feb-braio 1991.
CAVALLI-SFORZA L. L., MENOZZI RPIAZZA A., History and Geography ofHuman Genes (in stampa).
24 LE SCIENZE n. 281, gennaio 1992
LE SCIENZE n. 281, gennaio 1992 25





























