Embed Size (px)
Citation preview

1
SI Appendix
Small genome symbiont underlies cuticle hardness in beetles Hisashi Anbutsua,b,1,2, Minoru Moriyamaa,1, Naruo Nikohc,1, Takahiro Hosokawaa,d, Ryo Futahashia,
Masahiko Tanahashia, Xian-Ying Menga, Takashi Kuriwadae,f, Naoki Morig, Kenshiro Oshimah,
Masahira Hattorih,i, Manabu Fujiej, Noriyuki Satohk, Taro Maedal, Shuji Shigenobul, Ryuichi Kogaa,
and Takema Fukatsua,m,n,2
aBioproduction Research Institute, National Institute of Advanced Industrial Science and
Technology, Tsukuba 305-8566, Japan; bComputational Bio Big-Data Open Innovation Laboratory,
National Institute of Advanced Industrial Science and Technology, Tokyo 169-8555, Japan; cDepartment of Liberal Arts, The Open University of Japan, Chiba 261-8586, Japan; dFaculty of
Science, Kyushu University, Fukuoka 819-0395, Japan; eNational Agriculture and Food Research
Organization, Kyushu Okinawa Agricultural Research Center, Okinawa 901-0336, Japan; fFaculty
of Education, Kagoshima University, Kagoshima 890-0065, Japan; gDivision of Applied Life
Sciences, Graduate School of Agriculture, Kyoto University, Kyoto 606-8502, Japan; hGraduate
School of Frontier Sciences, University of Tokyo, Chiba 277-8561, Japan; iGraduate School of
Advanced Science and Engineering, Waseda University, Tokyo 169-8555, Japan; jDNA
Sequencing Section, Okinawa Institute of Science and Technology Graduate University, Okinawa
904-0495, Japan; kMarine Genomics Unit, Okinawa Institute of Science and Technology Graduate
University, Okinawa 904-0495, Japan; lNational Institute for Basic Biology, Okazaki 444-8585,
Japan; mDepartment of Biological Sciences, Graduate School of Science, University of Tokyo,
Tokyo 113-0033, Japan; and nGraduate School of Life and Environmental Sciences, University of
Tsukuba, Tsukuba 305-8572, Japan
1H.A., M.M. and N.N contributed equally to this work. 2To whom correspondence should be addressed.
E-mail: [email protected], [email protected]

2
Contents
SI Materials and Methods Insects, Rearing, and Fitness Measurement …………………………………………………… 4
Histology …………………………………………………………………………………………… 4
Genome Analysis ………………………………………………………………………………… 5
Transcriptome Analysis ………………………………………………………………………… 5
Symbiont Quantification ………………………………………………………………………… 5
Tissue Culture and 15N-Labeling ………………………………………………………………… 5
Analysis of Amino Acids ………………………………………………………………………… 6
Evaluation of Elytra Color and Hardness ……………………………………………………… 6
Characterization, Expression and Functional Analysis of GOT1, GOT2 and TAT Genes …… 7
Statistics …………………………………………………………………………………………… 7 SI References …………………………………………………………………………………… 7 SI Figures Fig. S1. Transmission electron microscopic images of Nardonella in larval bacteriomes ……… 9
Fig. S2. Rearing systems for P. infernalis ……………………………………………………… 10
Fig. S3. Effects of antibiotic treatment on fitness parameters of larvae, pupae and adults of P.
infernalis reared on the artificial diets ………………………………………………………… 12
Fig. S4. Adult insects of P. infernalis subjected to elytra color analysis ……………………… 13
Fig. S5. Trimmed and processed elytra images of adult P. infernalis subjected to elytra color
analysis ………………………………………………………………………………………… 14
Fig. S6. A schematic diagram showing how tyrosine and L-DOPA are involved in beetle’s cuticular
pigmentation and hardening ……………………………………………………………………… 15
Fig. S7. A schematic diagram of tyrosine synthesis pathway presumably operating in the
Nardonella-weevil endosymbiotic system ……………………………………………………… 16
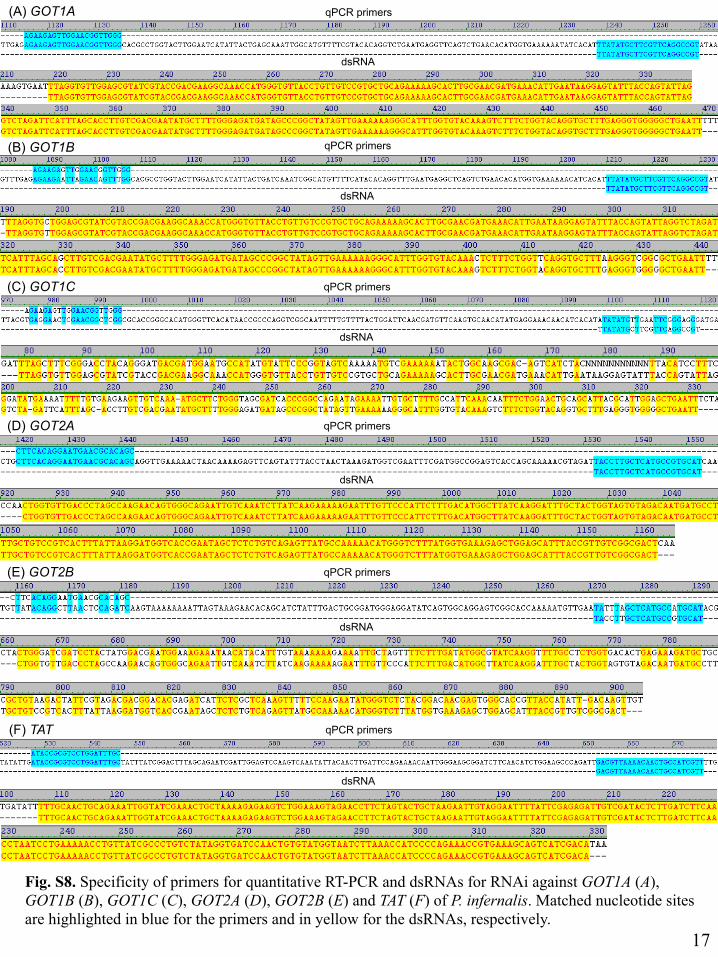
Fig. S8. Specificity of primers for quantitative RT-PCR and dsRNAs for RNAi …………………17
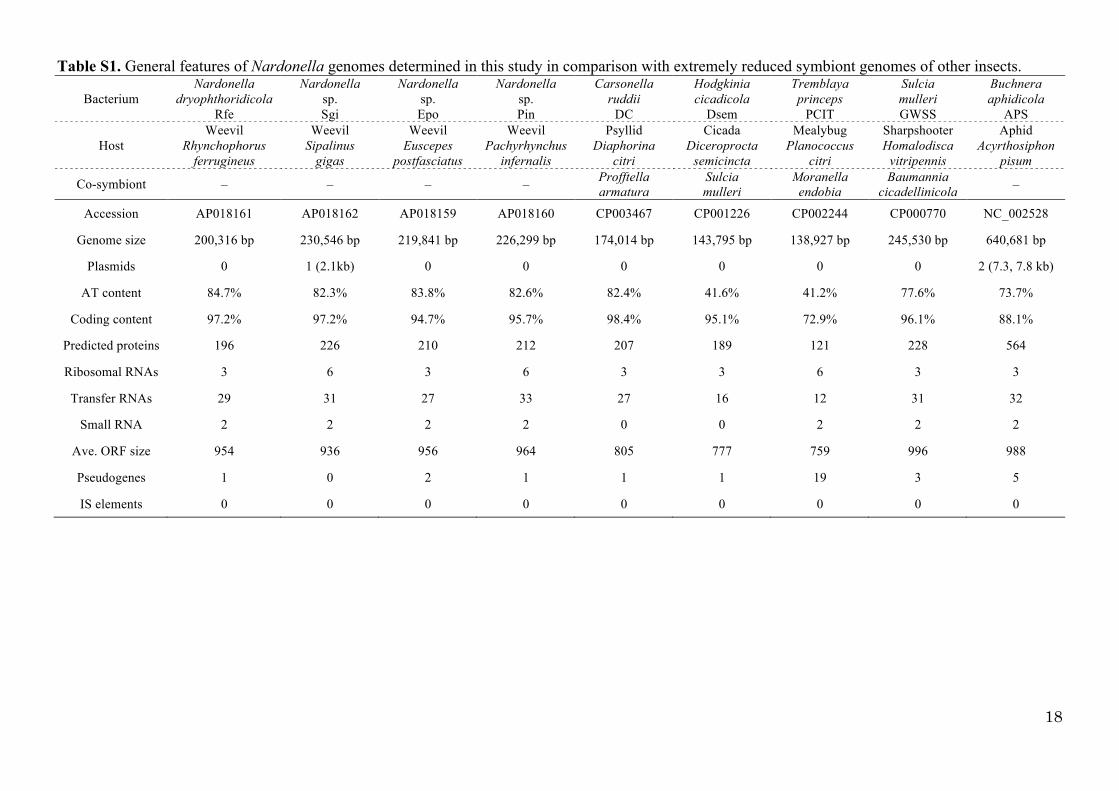
SI Tables Table S1. General features of Nardonella genomes determined in this study in comparison with
extremely reduced symbiont genomes of other insects …………………………………………… 18
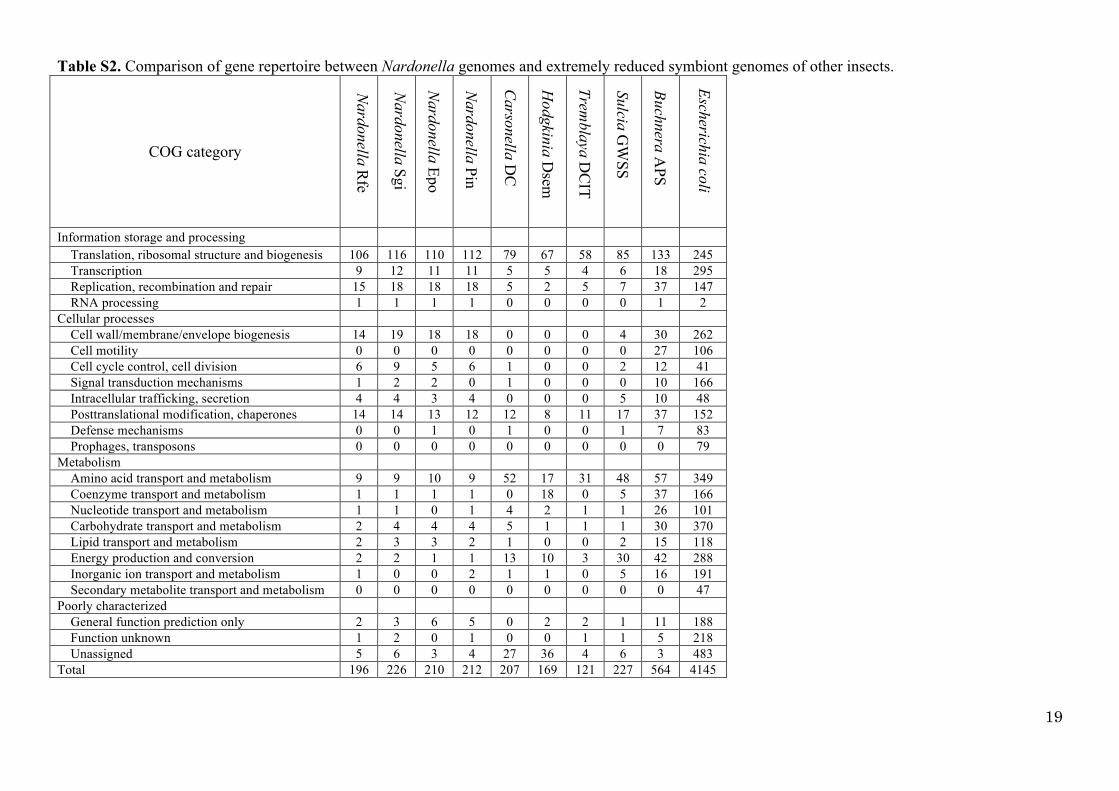
Table S2. Comparison of gene repertoire between Nardonella genomes and extremely reduced
symbiont genomes of other insects ……………………………………………………………… 19
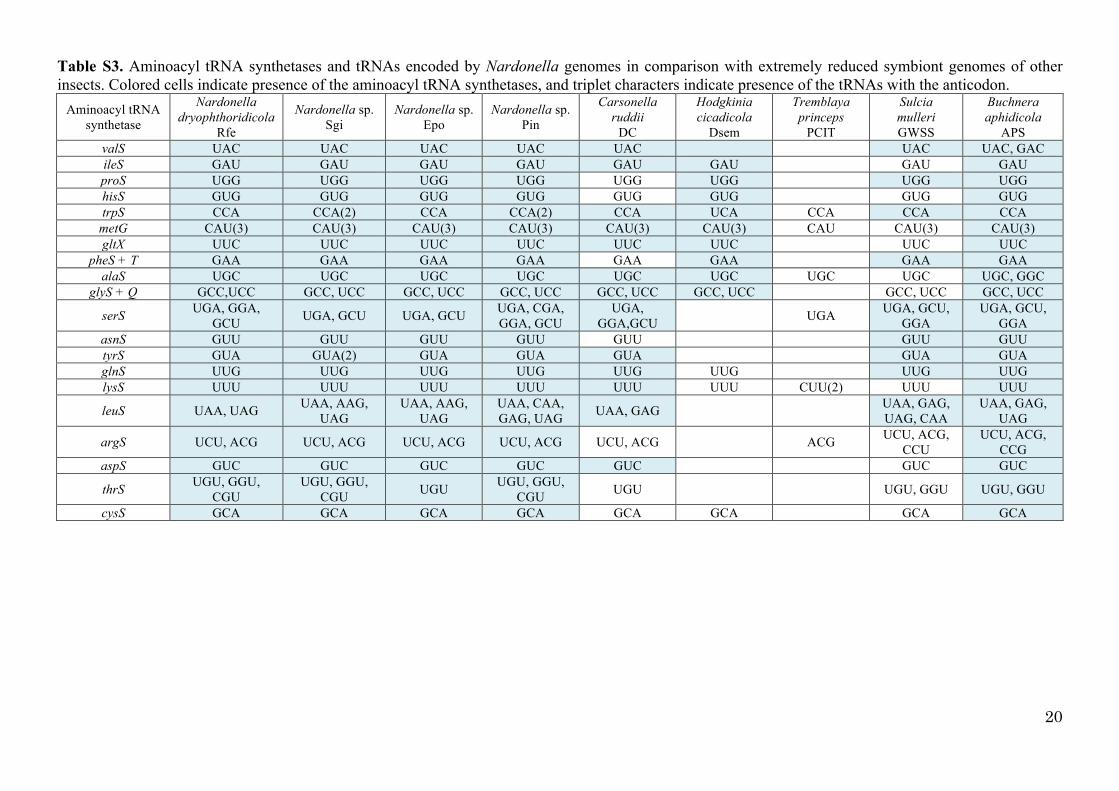
Table S3. Aminoacyl tRNA synthetases and tRNAs encoded by Nardonella genomes in

3
comparison with extremely reduced symbiont genomes of other insects ………………………… 20
Table S4. Stage-specific survival rates of P. infernalis on control artificial diet and
antibiotic-supplemented artificial diet …………………………………………………………… 21
Table S5. Overall survival rates of P. infernalis on control artificial diet and
antibiotic-supplemented artificial diet …………………………………………………………… 22
Table S6. Summary of RNA sequencing data of bacteriome and midgut samples dissected from
control and antibiotic-treated mature larvae of P. infernalis …………………………………… 23
Table S7. Weevil samples subjected to symbiont genome sequencing ………………………… 24
Table S8. Probes for in situ hybridization, primers for quantitative PCR, and primers for synthesis
of dsRNAs for RNAi used in this study ………………………………………………………… 25

4
SI Materials and Methods Insects, Rearing, and Fitness Measurement. Weevil samples subjected to bacteriome dissection
and symbiont genome sequencing are as listed (SI Appendix, Table S7). Adult insects of P.
infernalis were repeatedly collected from such trees as Mallotus japonicus, Macaranga tanarius
and Bauhinia sp. at Ishigaki Isl., Okinawa, Japan, and their offspring were maintained in the
laboratory at 25oC under a long day condition of 16 h light and 8 h dark. The adult insects were fed
with fresh cut vegetables like carrots, apples and aubergines in plastic containers (SI Appendix, Fig.
S2J). Eggs laid into grooves carved on the surface of the vegetables were carefully collected and
placed on wet filter paper in petri dishes to obtain newborn larvae (SI Appendix, Fig. S2K and L).
For rearing on sweet potatoes, which was mainly for maintenance and supply of experimental
insects, the newborn larvae were introduced into small holes bored on fresh sweet potatoes (SI
Appendix, Fig. S2A). The larvae fed on, pupated in, and emerged as adults from the sweet potatoes
(SI Appendix, Fig. S2B and C). For rearing on artificial diet plates, which was mainly for
manipulative experiments, the newborn larvae were placed in 3.5 cm plastic petri dishes poured
with 4% agar-based diet containing sweet potato powder, sucrose, soybean protein powder, yeast,
cellulose powder, vitamins and other ingredients (S1, S2) (SI Appendix, Fig. S2D). For preparing
antibiotic-supplemented artificial diets, melted artificial diets were cooled down to 40oC, to which
an aliquot of supplement solution was added to the final concentration of 0.003% rifampicin. From
3 months after the hatching, head capsule width of larvae was measured at an interval of 1 month,
and fully-grown larvae whose head capsule width attained 2.7 mm or more were transferred to
plastic sandboxes (89 mm x 57 mm x 19 mm), in which the larvae dug into the sand, made pupal
chambers, and pupated (SI Appendix, Fig. S2G-I). Newly-emerged adult insects were reared as
described above (see SI Appendix, Fig. S2J). During the rearing process, some insects were sampled
for a variety of experiments, while other insects were examined for the following fitness
parameters: survival rate, body weight, and head capsule width of larvae; survival rate, body weight,
and period of pupae; and survival rate, body weight, and elytra length of adults (see SI Appendix,
Fig. S3; Table S4; Table S5).
Histology. Dissected insect tissues were fixed in Carnoy’s solution (60% ethanol, 30% chloroform
and 10% acetic acid) or phosphate-buffered 4% paraformaldehyde, treated with 6%
H2O2-containing ethanol for a week to quench autofluorescence of the tissues, subjected to whole
mount fluorescence in situ hybridization as described previously (S3, S4) with the oligonucleotide
probes specifically targeting Nardonella’s 16S rRNA of P. infernalis (SI Appendix, Table S8), and
observed under an epifluorescence microscope or a laser scanning confocal microscope. For
transmission electron microscopy, dissected bacteriomes were prefixed in phosphate-buffered 2.5%
glutaraldehyde, postfixed in 2% OsO4, embedded in epoxy resin, processed into ultrathin sections

5
(80 nm) using a ultramicrotome, mounted on copper meshes, stained with uranyl acetate and lead
citrate, and observed under a transmission electron microscope. Semi-ultrathin sections (0.5 µm) of the resin-embedded samples were mounted on glass slides, stained with toluidine blue, and
observed under a light microscope.
Genome Analysis. Dissected larval bacteriomes were subjected to DNA extraction and symbiont
genome sequencing. The genomes of Nardonella Rfe and Sgi were determined by the conventional
random shotgun library construction and Sanger sequencing, whereas the genomes of Nardonella
Epo and Pin were determined by random shotgun sequencing using 454 GS FLX (Roche) and
HiSeq 2000 (Illumina). Genome assembly, gene prediction and annotation were conducted as
described previously (S5-S8). Contaminated host genes were automatically removed in the process
of symbiont genome assembly.
Transcriptome Analysis. Total RNA samples were individually extracted from freshly-dissected
bacteriomes and midguts using NucleoSpin RNA II Kit (Macherey-Nagel). Using 1 µg of total
RNA per sample as template, cDNA libraries were constructed using TruSeq RNA Sample
Preparation Kit v2 (Illumina) and sequenced by HiSeq2000 (Illumina). The raw reads were
subjected to de novo assembling by using the Trinity program (S9) implemented in the MASER
pipeline (http://cell-innovation.nig.ac.jp/). After automatic assembling, we checked and manually
corrected each of the GOT, TAT and Elongation Factor 1α (EF1α) gene sequences using the Integrative Genomics Viewer (S10). After revising the sequences, sequence read mapping was
performed by using the bowtie2-RSEM program (S11, S12), whereby transcript expression levels
were estimated in terms of transcripts per million reads (TPM) values.
Symbiont Quantification. Each whole insect or dissected tissue was subjected to DNA extraction
using QIAamp DNA mini Kit (Qiagen). Quantitative PCR of Nardonella’s groEL gene, and also
host’s EF1α gene for density normalization, were performed as described previously (S13) using the specific primers (SI Appendix, Table S8) under a temperature profile of 95oC for 10 min
followed by 40 cycles of 95oC for 30 sec, 60oC for 30 sec and 72oC for 40 sec.
Tissue Culture and 15N-Labeling. Final instar larvae of P. infernalis, which were 70-100 days old
at 25oC and 40-70 days old at 30oC, respectively, were dissected and subjected to in vitro
bacteriome culture. The culture medium was originally developed for in vitro tracer experiments
with aphid bacteriocytes (buffer A in S14), which was modified on account of sugar and amino acid
compositions in the hemolymph of P. infernalis. The filter-sterilized basal medium consists of 25
mM KCl, 10 mM MgCl, 250 mM trehalose and 35 mM

6
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (pH 7.4), to which [2-15N] glutamine
(Cambridge Isotope Laboratories) was added at a final concentration of 10 mM. Larval bacteriomes
were dissected in sterilized phosphate-buffered saline, individually placed on each well of a 24-well
glass slide (Fig. 4A and B), washed three times with the basal medium without 15N-glutamine, and
floated in 15 µl of the basal medium. The glass slide was hung upside down on the lid of a moistened plastic petri dish (Fig. 4C), and pre-incubated for 30 min at 25°C. Then, the medium was
replaced by 15 µl of the 15N-glutamine-supplemented medium. After 2 h incubation, the medium
was recovered and stored at –80°C for amino acid analysis.
Analysis of Amino Acids. From each insect sample, 2 µl of hemolymph was collected, suspended
in 200 µl of 80% methanol, and stored at −80°C until use. After purification through a solid phase
extraction column (GL-tip SDB, GL Science), amino acids and L-DOPA in the samples were
subjected to propyl-chloroformate derivatization (S15), which were separated by a high
performance liquid chromatography system (Prominence, Shimadzu) with a Shim-pack FC-ODS
column (150 mm x 2 mm i.d., Shimadzu) and detected by an electrospray ionization mass
spectrometry system (LCQduo, Thermo). The 15N-labeled portion of each amino acid was
calculated by subtracting the natural isotope abundance.
Evaluation of Elytra Color and Hardness. Adult insects were placed on white paper and
photographed by a digital camera (IXY 430F, Canon) under the same lighting condition (SI
Appendix, Fig. S4). Image processing operations and measurements were conducted on the software
Natsumushi version 0.99 (https://sites.google.com/site/mtahashilucanid/program/natsumushi). From
each of dorsal images of adult insects, a square area of maximal size was extracted. On the square
image, the pixels whose brightness was either over top 10% or below bottom 10% were masked in
blue and excluded from the analysis to minimize the effects of highlights and shadows (Fig. 6E-G; SI Appendix, Fig. S5). Then, 8-bit integer RGB values (0 ≦ R, G, B ≦ 255) for all (= n) pixels were
measured and averaged to obtain the redness index by Σ (R – mean [R, G, B])/n. An automatic dynamic viscoelastometer (Rheovibron DDV-25FP, A&D) (Fig. 6H) was used to perform
mechanical measurements. Adult insects were sacrificed by exposure to –80°C, from which elytra
were removed by tweezers and preserved in acetone. Prior to testing, each sample elytron was
mounted with epoxy glue on polycarbonate strips and was placed in a desiccator at room
temperature for at least 24 h prior to measurement. Once the sample was set on the instrument grips
by clamping the strips (Fig. 6I), elastic modulus of the elytra was measured with a frequency sweep
from 0.1 to 100 Hz, a vibration amplitude of ± 8 µm, and a static tension of 10 gf. Elytra thickness
was measured on the center of the sample elytron under an optical microscope with a digital
micrometer (MDC-25MX, Mitutoyo).

7
Characterization, Expression and Functional Analysis of GOT1, GOT2 and TAT Genes. The
assembled RNA sequencing data were subjected to tBLASTn searches using GOT1, GOT2, TAT (=
CG1461) and EF1α protein sequences of Drosophila melanogaster as queries, by which GOT1A,
GOT1B, GOT1C, GOT2A, GOT2B, TAT and EF1α gene sequences of P. infernalis were identified
(accession numbers LC260175-LC260180, LC260491). Neighbor-joining and maximum-likelihood
phylogenies were constructed using the program MEGA7 (S16) with 1,000 bootstrap resamplings.
Quantitative RT-PCR of GOT1A, GOT1B, TAT and EF1α transcripts was performed as described
previously (S17) by designing the specific primers as depicted (SI Appendix, Fig. S8; Table S8)
under a temperature profile of 95oC for 10 min followed by 40 cycles of 95oC for 15 sec, 60oC for
20 sec and 72oC for 20 sec. RNAi of GOT1, GOT2 and TAT gene expression was performed as
described previously (S18): dsRNAs targeting GOT1A, GOT2A, TAT and Green Fluorescent
Protein (GFP) transcripts were synthesized from PCR products of the genes using MEGAscript
RNAi kit (Ambion) wherein fragment amplification was conducted using the specific primers (SI
Appendix, Table S8) attached to T7 promoter sequence, and the dsRNAs were injected into the
abdomen of mature larvae using a thin glass capillary needle connected to an air pump at a dose of
1 µg each dsRNA dissolved in 1.5 µl (for 1 gene) or 4.5 µl (for 3 genes) water. It should be noted that, although the dsRNA for GOT1 was designed to specifically suppress GOT1A, it may also act
on GOT1B and GOT1C on account of the sequence similarities, so was the dsRNA for GOT2
specific to GOT2A but potentially also acting on GOT2B (see SI Appendix, Fig. S8).
Statistics. All statistical analyses were performed using R version 3.3.2 (S19). Considering data
dispersion, we used a Wilcoxon rank sum test for elytra redness, a Welch's T-test for body size, wet
weight, pupal period, elytra elastic modulus, elytra thickness, and hemolymphal tyrosine titer and
L-DOPA titer, or a generalized linear model (GLM) assuming a Gamma error structure for bacterial
titer, gene expression level, and tyrosine synthesis.
SI References S1. Urasaki K, Ohno S, Haraguchi D, Kohama T (2009) Effect of simplifying the preparation of
larval artificial diet on survival and development of the West Indian sweet potato weevil
Euscepes postfasciatus (Fairmaire) (Coleoptera: Curculionidae). Jpn J Appl Entomol Zool
53:1-6.
S2. Kuriwada T, et al. (2010) Biological role of Nardonella endosymbiont in its weevil host. PLoS
One 5:e13101.
S3. Koga R, Tsuchida T, Fukatsu T (2009) Quenching autofluorescence of insect tissues for in situ
detecton of endosymbionts. Appl Entomol Zool 44:281-291.

8
S4. Hosokawa T, et al. (2015) Nardonella endosymbionts of Japanese pest and non-pest weevils
(Coleoptera: Curculionidae). Appl Entomol Zool 50: 223-229.
S5. Nikoh N, Hosokawa T, Oshima K, Hattori M, Fukatsu T (2011) Reductive evolution of
bacterial genome in insect gut environment. Genome Biol Evol 3:702-714.
S6. Nikoh N, et al. (2014) Evolutionary origin of insect-Wolbachia nutritional mutualism. Proc
Natl Acad Sci USA 111:10257-10262.
S7. Kaiwa N, et al. (2014) Symbiont-supplemented maternal investment underpinning host's
ecological adaptation. Curr Biol 24:2465-2470.
S8. Hosokawa T, et al. (2016) Obligate bacterial mutualists evolving from environmental bacteria
in natural insect populations. Nat Microbiol 1:15011.
S9. Grabherr MG, et al. (2011) Full-length transcriptome assembly from RNA-Seq data without a
reference genome. Nat Biotechnol 29:644-652.
S10. Thorvaldsdóttir H, Robinson JT, Mesirov JP (2013) Integrative Genomics Viewer (IGV):
high-performance genomics data visualization and exploration. Brief Bioinformatics
14:178-192.
S11. Li B, Dewey CN (2011) RSEM: accurate transcript quantification from RNA-Seq data with or
without a reference genome. BMC Bioinformatics 12:323.
S12. Langmead B, Salzberg SL (2012) Fast gapped-read alignment with Bowtie 2. Nat Methods
9:357-359.
S13. Harumoto T, Anbutsu H, Fukatsu T (2014) Male-killing Spiroplasma induces sex-specific cell
death via host apoptotic pathway. PLoS Pathog 10:e1003956.
S14. Sasaki T, Ishikawa H (1995) Production of essential amino acids from glutamate by
mycetocyte symbionts of the pea aphid, Acyrthosiphon pisum. J Insect Physiol 41:41-46.
S15. Uutela P, Ketola RA, Piepponen P, & Kostiainen R (2009) Comparison of different amino
acid derivatives and analysis of rat brain microdialysates by liquid chromatography tandem
mass spectrometry. Anal Chim Acta 633:223-231.
S16. Kumar S, Stecher G, Tamura K (2016) MEGA7: Molecular Evolutionary Genetics Analysis
Version 7.0 for bigger datasets. Mol Biol Evol 33:1870-1874.
S17. Moriyama M, et al. (2012) Comparative transcriptomics of the bacteriome and the spermalege
of the bedbug Cimex lectularius (Hemiptera: Cimicidae). Appl Entomol Zool 47:233-243.
S18. Moriyama M, Hosokawa T, Tanahashi M, Nikoh N, Fukatsu T (2016) Suppression of bedbug's
reproduction by RNA interference of vitellogenin. PLoS One 11:e0153984.
S19. R Core Team (2016). R: A language and environment for statistical computing. R Foundation
for Statistical Computing, Vienna, Austria. URL https://www.R-project.org/.
1 µm
B C
E F
10 µm
10 µm 1 µm
1 cm
1 cm
A
D
H IG
1 µm10 µm1 cm
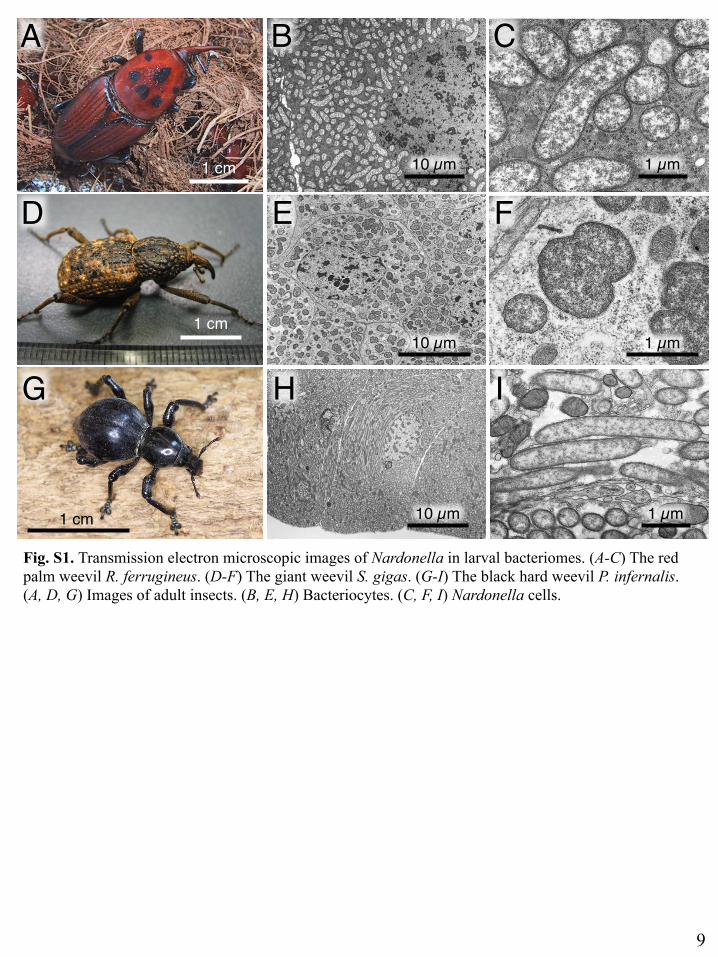
Fig. S1. Transmission electron microscopic images of Nardonella in larval bacteriomes. (A-C) The red palm weevil R. ferrugineus. (D-F) The giant weevil S. gigas. (G-I) The black hard weevil P. infernalis. (A, D, G) Images of adult insects. (B, E, H) Bacteriocytes. (C, F, I) Nardonella cells.
9

A CB
D FE
G IH
J LK
1 mm 1 cm 1 cm
1 cm 1 cm 1 cm
1 cm1 cm
1 cm
1 cm5 cm 1 cm
10

Fig. S2. Rearing systems for P. infernalis. (A-C) Rearing on sweet potatoes. (A) A newborn larva introduced into a small hole bored on a sweet potato. (B) A mature larva in a sweet potato. (C) A pupa in a sweet potato. (D-F) Rearing on artificial diet plates. (D) A mature larva in a diet plate. (E) A diet plate contaminated with bacteria. Color and smell of the diet change, the surface of the diet become slimy and/or wet, and young larvae in such diet plates tend to suffer retarded growth and/or high mortality. This type of contamination is observed more frequently in control diet plates than in antibiotic-supplemented diet plates. (F) A diet plate contaminated with fungi. Larvae in such diet plates tend to suffer high mortality. This type of contamination is occasionally found in both control diet plates and antibiotic-supplemented diet plates. (G-I) Sandboxes for facilitating successful pupation. Transfer of fully-grown larvae on the artificial diet to the sandboxes triggers formation of a pupal chamber and pupation in the sand. (G) A prepupa. (H) A pupa. (I) A newly-eclosed, pale-colored adult. (J) Adult rearing and egg collection system. Water, apple and carrot are provided in a plastic cage for adult feeding. On the surface of the carrot, grooves are made with a knife, which serve as preferred oviposition sites for gravid females. (K) Eggs laid in a carrot groove. (L) Keeping of collected eggs on a sheet of wet filter paper in a Petri dish. The hatchling larvae are transferred to either sweet potatoes or artificial diet plates. In (A-D) and (G-J), yellow arrowheads indicate insects. In (K, L), yellow, red, and blue arrowheads point eggs, hatched eggshells, and newborn larvae, respectively.
11
●
●
●
●
●
6
7
8
9
C RTreatments
Ely
tra le
ngth
(mm
)
TreatmentsCR
●
●
●
●
100
200
300
400
C RTreatments
Wet
bod
y w
eigh
t (m
g)
TreatmentsCR
●
●
●
100
150
200
250
300
C RTreatments
Wet
bod
y w
eigh
t (m
g)
TreatmentsCR
●
●
●
●
●
20
40
60
C RTreatments
Dur
atio
n in
san
d (d
ays)
TreatmentsCR
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●
●●
●
●
●
●
●
●●
●
●
●
●●
●
1.0
1.5
2.0
2.5
3.0
C RTreatments
Hea
d ca
psul
e w
idth
(mm
)
TreatmentsCR
●
●
●●●
●●●0
100
200
300
400
C RTreatments
Wet
bod
y w
eigh
t (m
g)
TreatmentsCR
***
A Larval weight
B Larval body size
C Pupal weight
30 23616 602
610 602 71 60
**
D Pupal period
ns
E Adult weight
F Adult body size
134 119
ns
119 99
Wet
bod
y w
eigh
t (m
g)
Wet
bod
y w
eigh
t (m
g)
Wet
bod
y w
eigh
t (m
g)
Hea
d ca
psul
e w
idth
(mm
)
Dur
atio
n in
san
d (d
ays)
Ely
tra le
ngth
(mm
)
Control Antibiotic Control Antibiotic Control Antibiotic
ns
ns
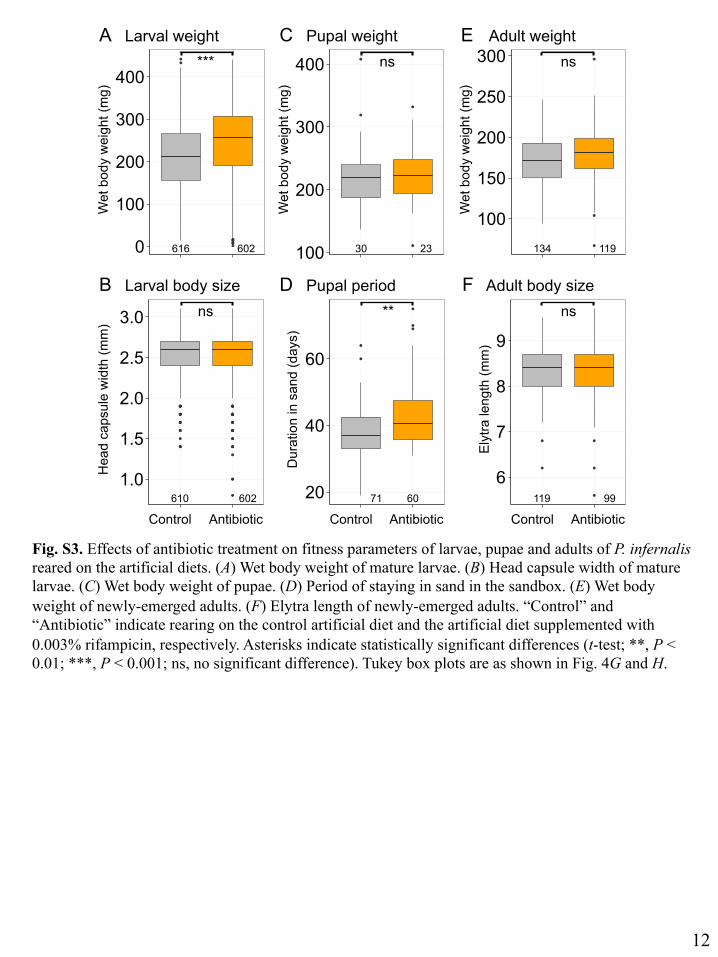
Fig. S3. Effects of antibiotic treatment on fitness parameters of larvae, pupae and adults of P. infernalis reared on the artificial diets. (A) Wet body weight of mature larvae. (B) Head capsule width of mature larvae. (C) Wet body weight of pupae. (D) Period of staying in sand in the sandbox. (E) Wet body weight of newly-emerged adults. (F) Elytra length of newly-emerged adults. “Control” and “Antibiotic” indicate rearing on the control artificial diet and the artificial diet supplemented with 0.003% rifampicin, respectively. Asterisks indicate statistically significant differences (t-test; **, P < 0.01; ***, P < 0.001; ns, no significant difference). Tukey box plots are as shown in Fig. 4G and H.
12

Control (0-day) N=69
Control (7-day) N=68
Control (35-day) N=45
Antibiotic (0-day) N=79
Antibiotic (7-day) N=77
Antibiotic (35-day) N=43
Fig. S4. Adult insects of P. infernalis subjected to elytra color analysis. In total 381 individuals were analyzed, of which 182 were reared on the control artificial diet (Control) and 199 were reared on the artificial diet supplemented with 0.003% rifampicin (Antibiotic).
13

Control (0-day) N=69
Control (7-day) N=68
Control (35-day) N=45
Antibiotic (0-day) N=79
Antibiotic (7-day) N=77
Antibiotic (35-day) N=43
Fig. S5. Trimmed and processed elytra images of adult P. infernalis subjected to elytra color analysis. In total 381 individuals were analyzed, of which 182 were reared on the control artificial diet (Control) and 199 were reared on the artificial diet supplemented with 0.003% rifampicin (Antibiotic).
14
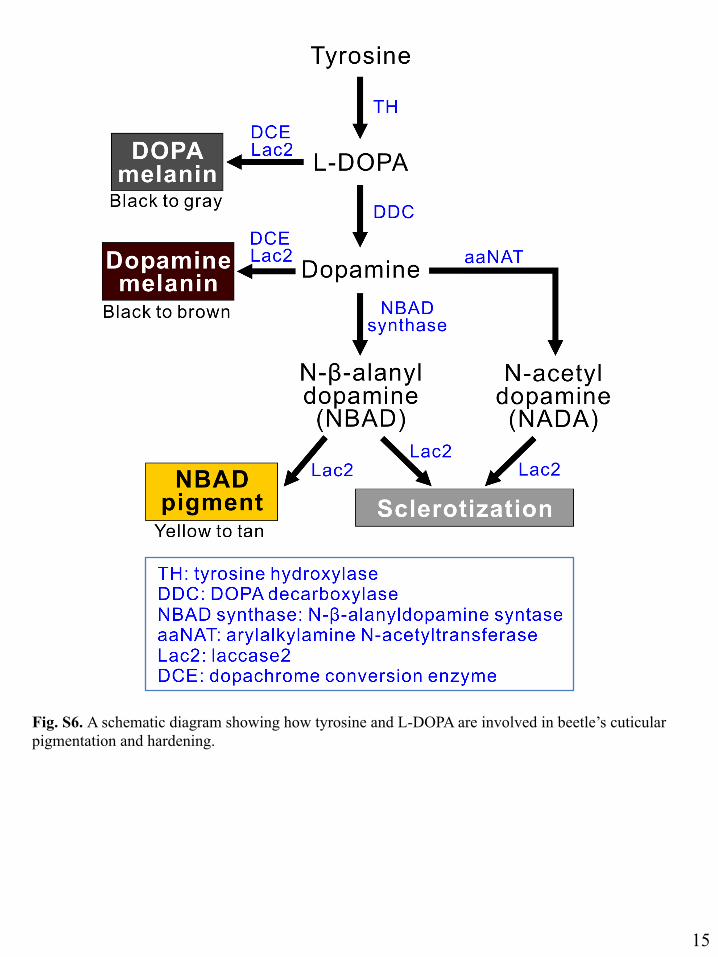
Fig. S6. A schematic diagram showing how tyrosine and L-DOPA are involved in beetle’s cuticular pigmentation and hardening.
15
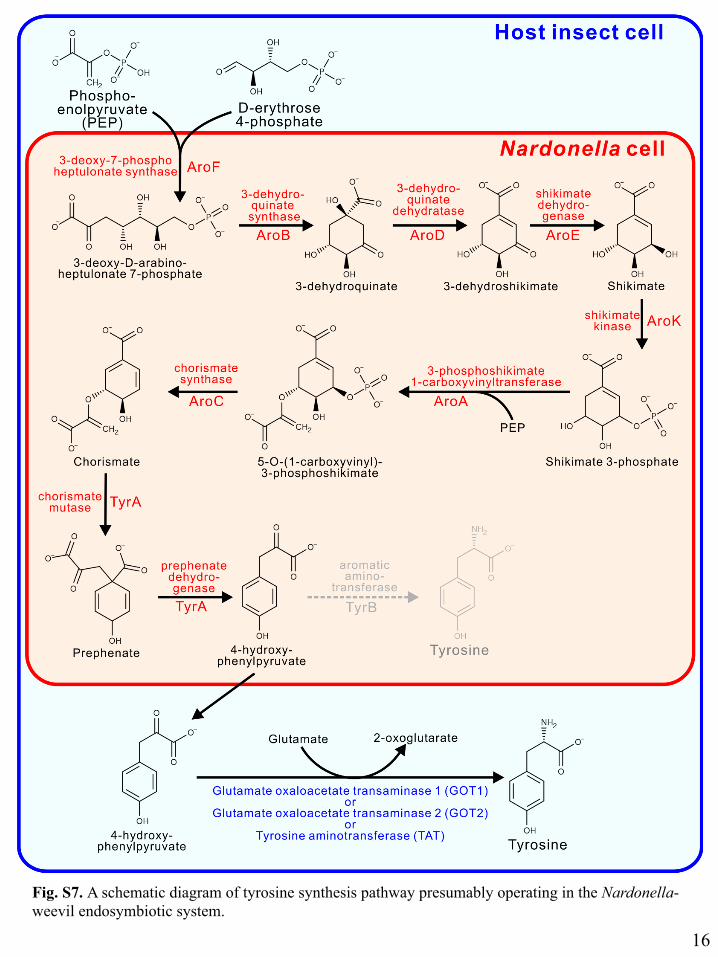
Fig. S7. A schematic diagram of tyrosine synthesis pathway presumably operating in the Nardonella-weevil endosymbiotic system.
16
(A) GOT1A qPCR primers
dsRNA
(B) GOT1B qPCR primers
dsRNA
(C) GOT1C qPCR primers
dsRNA
(D) GOT2A qPCR primers
dsRNA
(E) GOT2B qPCR primers
dsRNA
(F) TAT qPCR primers
dsRNA
Fig. S8. Specificity of primers for quantitative RT-PCR and dsRNAs for RNAi against GOT1A (A), GOT1B (B), GOT1C (C), GOT2A (D), GOT2B (E) and TAT (F) of P. infernalis. Matched nucleotide sites are highlighted in blue for the primers and in yellow for the dsRNAs, respectively.
17
18
Table S1. General features of Nardonella genomes determined in this study in comparison with extremely reduced symbiont genomes of other insects.
Bacterium Nardonella
dryophthoridicola Rfe
Nardonella sp. Sgi
Nardonella sp. Epo
Nardonella sp. Pin
Carsonella ruddii
DC
Hodgkinia cicadicola
Dsem
Tremblaya princeps
PCIT
Sulcia mulleri GWSS
Buchnera aphidicola
APS
Host Weevil
Rhynchophorus ferrugineus
Weevil Sipalinus
gigas
Weevil Euscepes
postfasciatus
Weevil Pachyrhynchus
infernalis
Psyllid Diaphorina
citri
Cicada Diceroprocta
semicincta
Mealybug Planococcus
citri
Sharpshooter Homalodisca
vitripennis
Aphid Acyrthosiphon
pisum
Co-symbiont – – – – Profftella armatura
Sulcia mulleri
Moranella endobia
Baumannia cicadellinicola –
Accession AP018161 AP018162 AP018159 AP018160 CP003467 CP001226 CP002244 CP000770 NC_002528
Genome size 200,316 bp 230,546 bp 219,841 bp 226,299 bp 174,014 bp 143,795 bp 138,927 bp 245,530 bp 640,681 bp
Plasmids 0 1 (2.1kb) 0 0 0 0 0 0 2 (7.3, 7.8 kb)
AT content 84.7% 82.3% 83.8% 82.6% 82.4% 41.6% 41.2% 77.6% 73.7%
Coding content 97.2% 97.2% 94.7% 95.7% 98.4% 95.1% 72.9% 96.1% 88.1%
Predicted proteins 196 226 210 212 207 189 121 228 564
Ribosomal RNAs 3 6 3 6 3 3 6 3 3
Transfer RNAs 29 31 27 33 27 16 12 31 32
Small RNA 2 2 2 2 0 0 2 2 2
Ave. ORF size 954 936 956 964 805 777 759 996 988
Pseudogenes 1 0 2 1 1 1 19 3 5
IS elements 0 0 0 0 0 0 0 0 0
19
Table S2. Comparison of gene repertoire between Nardonella genomes and extremely reduced symbiont genomes of other insects.
COG category
Information storage and processing Translation, ribosomal structure and biogenesis 106 116 110 112 79 67 58 85 133 245 Transcription 9 12 11 11 5 5 4 6 18 295 Replication, recombination and repair 15 18 18 18 5 2 5 7 37 147 RNA processing 1 1 1 1 0 0 0 0 1 2 Cellular processes Cell wall/membrane/envelope biogenesis 14 19 18 18 0 0 0 4 30 262 Cell motility 0 0 0 0 0 0 0 0 27 106 Cell cycle control, cell division 6 9 5 6 1 0 0 2 12 41 Signal transduction mechanisms 1 2 2 0 1 0 0 0 10 166 Intracellular trafficking, secretion 4 4 3 4 0 0 0 5 10 48 Posttranslational modification, chaperones 14 14 13 12 12 8 11 17 37 152 Defense mechanisms 0 0 1 0 1 0 0 1 7 83 Prophages, transposons 0 0 0 0 0 0 0 0 0 79 Metabolism Amino acid transport and metabolism 9 9 10 9 52 17 31 48 57 349 Coenzyme transport and metabolism 1 1 1 1 0 18 0 5 37 166 Nucleotide transport and metabolism 1 1 0 1 4 2 1 1 26 101 Carbohydrate transport and metabolism 2 4 4 4 5 1 1 1 30 370 Lipid transport and metabolism 2 3 3 2 1 0 0 2 15 118 Energy production and conversion 2 2 1 1 13 10 3 30 42 288 Inorganic ion transport and metabolism 1 0 0 2 1 1 0 5 16 191 Secondary metabolite transport and metabolism 0 0 0 0 0 0 0 0 0 47 Poorly characterized General function prediction only 2 3 6 5 0 2 2 1 11 188 Function unknown 1 2 0 1 0 0 1 1 5 218 Unassigned 5 6 3 4 27 36 4 6 3 483 Total 196 226 210 212 207 169 121 227 564 4145
Nardonella Rfe
Nardonella Sgi
Nardonella Epo
Nardonella Pin
Carsonella DC
Hodgkinia D
sem
Tremblaya D
CIT
Sulcia GW
SS
Buchnera APS
Escherichia coli
20
Table S3. Aminoacyl tRNA synthetases and tRNAs encoded by Nardonella genomes in comparison with extremely reduced symbiont genomes of other insects. Colored cells indicate presence of the aminoacyl tRNA synthetases, and triplet characters indicate presence of the tRNAs with the anticodon.
Aminoacyl tRNA synthetase
Nardonella dryophthoridicola
Rfe
Nardonella sp. Sgi
Nardonella sp. Epo
Nardonella sp. Pin
Carsonella ruddii
DC
Hodgkinia cicadicola
Dsem
Tremblaya princeps
PCIT
Sulcia mulleri GWSS
Buchnera aphidicola
APS valS UAC UAC UAC UAC UAC UAC UAC, GAC ileS GAU GAU GAU GAU GAU GAU GAU GAU proS UGG UGG UGG UGG UGG UGG UGG UGG hisS GUG GUG GUG GUG GUG GUG GUG GUG trpS CCA CCA(2) CCA CCA(2) CCA UCA CCA CCA CCA
metG CAU(3) CAU(3) CAU(3) CAU(3) CAU(3) CAU(3) CAU CAU(3) CAU(3) gltX UUC UUC UUC UUC UUC UUC UUC UUC
pheS + T GAA GAA GAA GAA GAA GAA GAA GAA alaS UGC UGC UGC UGC UGC UGC UGC UGC UGC, GGC
glyS + Q GCC,UCC GCC, UCC GCC, UCC GCC, UCC GCC, UCC GCC, UCC GCC, UCC GCC, UCC
serS UGA, GGA, GCU UGA, GCU UGA, GCU UGA, CGA,
GGA, GCU UGA,
GGA,GCU UGA UGA, GCU, GGA
UGA, GCU, GGA
asnS GUU GUU GUU GUU GUU GUU GUU tyrS GUA GUA(2) GUA GUA GUA GUA GUA glnS UUG UUG UUG UUG UUG UUG UUG UUG lysS UUU UUU UUU UUU UUU UUU CUU(2) UUU UUU
leuS UAA, UAG UAA, AAG, UAG
UAA, AAG, UAG
UAA, CAA, GAG, UAG UAA, GAG UAA, GAG,
UAG, CAA UAA, GAG,
UAG
argS UCU, ACG UCU, ACG UCU, ACG UCU, ACG UCU, ACG ACG UCU, ACG, CCU
UCU, ACG, CCG
aspS GUC GUC GUC GUC GUC GUC GUC
thrS UGU, GGU, CGU
UGU, GGU, CGU UGU UGU, GGU,
CGU UGU UGU, GGU UGU, GGU
cysS GCA GCA GCA GCA GCA GCA GCA GCA

21
Table S4. Stage-specific survival rates of P. infernalis on control artificial diet and antibiotic-supplemented artificial diet.
Stage1 Treatment2 Survival rate3 [1] Eggs ~ final instar larvae (larval survival)4 Control 27.8% (396/1427) a
Antibiotic 34.3% (400/1167) b [2] Final instar larvae ~ pupae (prepupal survival)5 Control 70.2% (219/312) a
Antibiotic 57.1% (214/375) b [3] Pupae ~ adult eclosion (pupal survival)6 Control 84.8% (168/198) ns
Antibiotic 78.1% (150/192) [4] Adult eclosion ~ emergence (teneral adult survival)7
Control 82.8% (130/157) ns Antibiotic 74.6% (100/134)
1 For rearing systems, see Fig. S2. 2 Control, insects reared on the artificial diet without supplements; Antibiotic, insects reared on the artificial diet containing 0.003% (w/v) rifampicin. 3 Some of the final instar larvae, pupae, and newly-eclosed adults were used for other experiments at the end of the stages 1, 2 and 3, respectively. Different alphabetical letters (a, b) indicate statistically significant differences (Pearson’s Chi-squared test with Bonferroni correction, P < 0.05), while “ns” means no significant difference. 4 Eggs inoculated to artificial diet plates ~ mature final instar larvae showing feeding cessation. 5 Post-feeding final instar larvae transferred to sand boxes ~ successful pupation in pupal chambers. 6 Pupae in pupal chambers ~ newly-eclosed teneral adults in pupal chambers. 7 Teneral adults in pupal chambers ~ darkened adults emerging from pupal chambers.

22
Table S5. Overall survival rates of P. infernalis on control artificial diet and antibiotic-supplemented artificial diet.
Stage Treatment1 Survival rate2
[1] ~ Final instar larvae Control 27.8% Antibiotic 34.3% [2] ~ Pupae Control 19.5% Antibiotic 19.6% [3] ~ Adult eclosion Control 16.5% Antibiotic 15.3% [4] ~ Adult emergence Control 13.7% Antibiotic 11.4% 1 Control, insects reared on the artificial diet without supplements; Antibiotic, insects reared on the artificial diet containing 0.003% (w/v) rifampicin. 2 The overall survival rates were estimated on the basis of the stage-specific survival rates shown in Table S4.
23
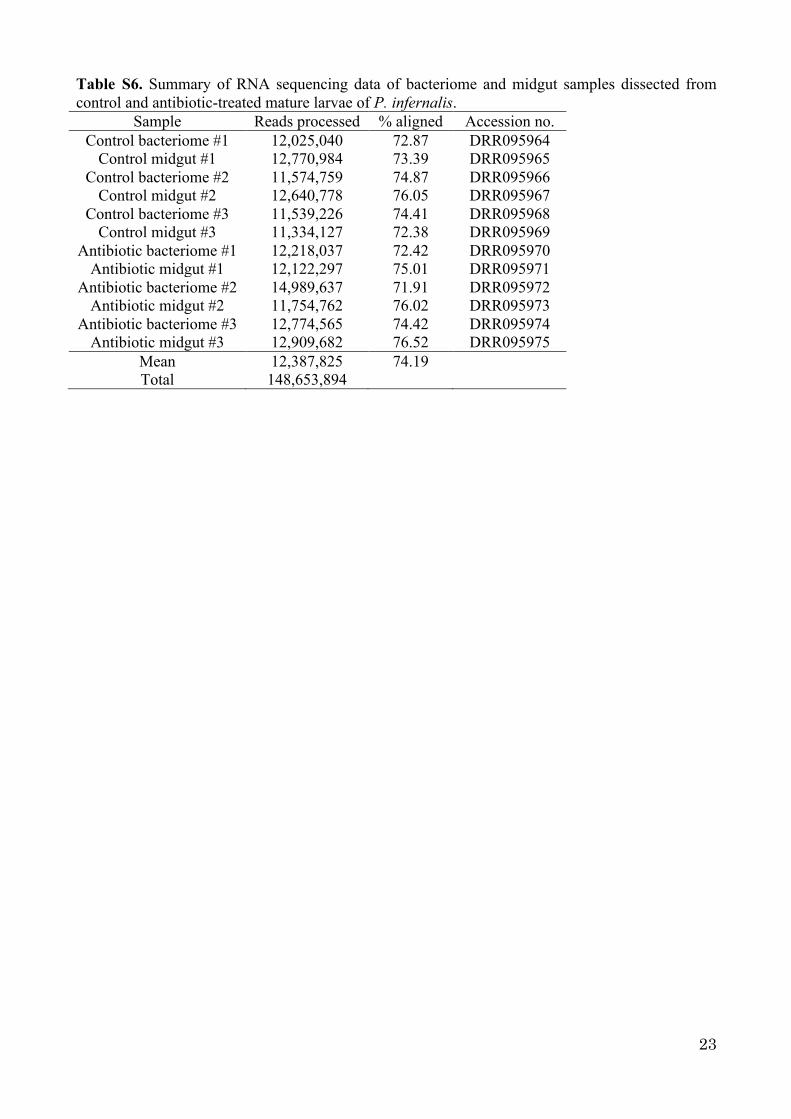
Table S6. Summary of RNA sequencing data of bacteriome and midgut samples dissected from control and antibiotic-treated mature larvae of P. infernalis.
Sample Reads processed % aligned Accession no. Control bacteriome #1 12,025,040 72.87 DRR095964
Control midgut #1 12,770,984 73.39 DRR095965 Control bacteriome #2 11,574,759 74.87 DRR095966
Control midgut #2 12,640,778 76.05 DRR095967 Control bacteriome #3 11,539,226 74.41 DRR095968
Control midgut #3 11,334,127 72.38 DRR095969 Antibiotic bacteriome #1 12,218,037 72.42 DRR095970
Antibiotic midgut #1 12,122,297 75.01 DRR095971 Antibiotic bacteriome #2 14,989,637 71.91 DRR095972
Antibiotic midgut #2 11,754,762 76.02 DRR095973 Antibiotic bacteriome #3 12,774,565 74.42 DRR095974
Antibiotic midgut #3 12,909,682 76.52 DRR095975 Mean 12,387,825 74.19 Total 148,653,894
24
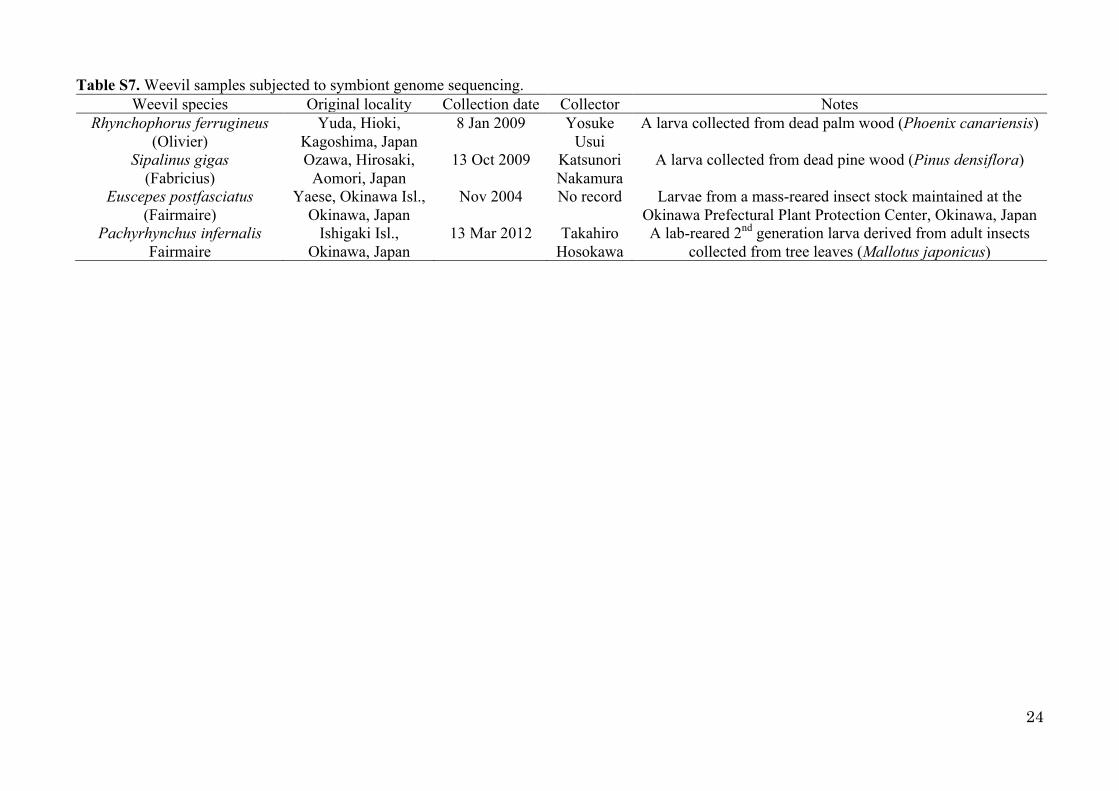
Table S7. Weevil samples subjected to symbiont genome sequencing. Weevil species Original locality Collection date Collector Notes
Rhynchophorus ferrugineus (Olivier)
Yuda, Hioki, Kagoshima, Japan
8 Jan 2009 Yosuke Usui
A larva collected from dead palm wood (Phoenix canariensis)
Sipalinus gigas (Fabricius)
Ozawa, Hirosaki, Aomori, Japan
13 Oct 2009 Katsunori Nakamura
A larva collected from dead pine wood (Pinus densiflora)
Euscepes postfasciatus (Fairmaire)
Yaese, Okinawa Isl., Okinawa, Japan
Nov 2004 No record Larvae from a mass-reared insect stock maintained at the Okinawa Prefectural Plant Protection Center, Okinawa, Japan
Pachyrhynchus infernalis Fairmaire
Ishigaki Isl., Okinawa, Japan
13 Mar 2012 Takahiro Hosokawa
A lab-reared 2nd generation larva derived from adult insects collected from tree leaves (Mallotus japonicus)

25
Table S8. Probes for in situ hybridization, primers for quantitative PCR, and primers for synthesis of dsRNAs for RNAi used in this study.
Target Name Sequence (5’-3’) For fluorescence in situ hybridization 16S rRNA NarPin_Al555-862R AlexaFluor555-AAT CTT GCG ATC GTA CTT CC
NarPin_Al555-1237R AlexaFluor555-TTC TTA CGA AAT AGC TTC TC For quantitative PCR
groEL NarPin_groEL_F CAG CAG GAG ATG GAA CTA CTA CAG C NarPin_groEL_R ACA TTT ACG GCT TGA ATA ACT GCT T
EF1α1 NarPin_EF1_625F CCT TGG TTC AAG GGA TGG AA NarPin_EF1_702R GGC ATC CAA AGC ATC AAT GA
For quantitative RT-PCR GOT1A Pin_GOT1A_993F AGA AGA GTT GGA ACG GTT GGG
Pin_GOT1A_1137R ACG GCC TGA ACG AAG CAT ATA A GOT2A Pin_GOT2A_1128F CTT CAC AGG AAT GAA CGC ACA GC
Pin_GOT2A_1260R ATG CAC GGC ATG AGC AAG GTA TAT Pin_TAT_460F ATA CCG CGT CCT GGA TTT GC
Pin_TAT_609R AAC GAT GGC AGT TGT TTT AAC GTC For synthesis of dsRNA for RNAi
GOT1 Pin_GOT1A_97F TTA GGT GTT GGA GCG TAT CGT ACC G Pin_GOT1A_347R AAT TCA GCC CCC ACC CTC AAA G
GOT2 Pin_GOT2A_632F CTG GTG TTG ACC CTA GCC AAG AAC A Pin_GOT2A_871R AGT CGC CGA CAA CGG TAA ATG CT
TAT Pin_TAT_40F TTT GCA ACT GCA GAA ATT GGT ATC G Pin_TAT_262R TGT CGA TGA CTG CTT TCA CGG TTT C
GFP2 Avi_GFP_10F GGA GAA GAA CTT TTC ACT GGA GT Avi_GFP_711R GTA TAG TTC ATC CAT GCC ATG TG
1Elongation Factor 1α: also used for quantitative RT-PCR. 2Green Fluorescence Protein: used for synthesizing control dsRNA.





























