Embed Size (px)
Citation preview

South African Journal of Botany 106 (2016) 8–16
Contents lists available at ScienceDirect
South African Journal of Botany
j ourna l homepage: www.e lsev ie r .com/ locate /sa jb
Combined effect of foliar-applied salicylic acid and deficit irrigation onphysiological–anatomical responses, and yield of squash plants undersaline soil
T.A. Abd El-Mageed a,⁎, W.M. Semida b, G.F. Mohamed c, M.M. Rady c
a Soil and Water Department, Faculty of Agriculture, Fayoum University, 63514 Fayoum, Egyptb Horticulture Department, Faculty of Agriculture, Fayoum University, 63514 Fayoum, Egyptc Botany Department, Faculty of Agriculture, Fayoum University, 63514 Fayoum,Egypt
⁎ Corresponding author. Tel.: +20 1067436208.E-mail address: [email protected] (T.A. Abd El-Ma
http://dx.doi.org/10.1016/j.sajb.2016.05.0050254-6299/© 2016 SAAB. Published by Elsevier B.V. All ri
a b s t r a c t
a r t i c l e i n f oArticle history:Received 21 September 2015Received in revised form 17 January 2016Accepted 13 May 2016Available online xxxx
Edited by L Sebastiani
Salicylic acid (SA) controls plant growth and induces water deficit tolerance in plants. Summer and fall seasonexperiments were conducted in 2013 to study the effect of 1mMSA on growth, anatomy, yield, chlorophyll fluo-rescence, osmoprotectants and water use efficiency (WUE) of squash plants under three levels of irrigation(I100 = 100%, I80 = 80% and I60 = 60% of crop evapotranspiration. Growth, chlorophyll fluorescence, leaf anato-my, leaf photosynthetic pigments, total soluble sugars (TSS), proline, harvest index (HI), yield andWUEwere sig-nificantly affected by both deficit irrigation (DI) and the combined DI + exogenously applied SA treatment. Thecombined DI+ SA treatmentmodified the adverse effects of DI and enhanced all aforesaid parameters, while de-creased proline concentrations. The combined treatment of I60 or I80 + SA produced plants having yields andWUE as produced with the plants generated under full irrigation conditions without SA treatment. From theseresults, we conclude thatwith exogenously applied 1mMSA, the I60 or I80 strategy studied here could be success-fully applied during summer and fall seasons for the production of commercial squash allowing water savings of20%–40% without any detrimental effect on plant growth or yield.
© 2016 SAAB. Published by Elsevier B.V. All rights reserved.
Keywords:Deficit irrigationSAOsmoprotectantsChlorophyll fluorescenceSquash yieldWater-use efficiency
1. Introduction
As one of the most important vegetable crops, squash (Cucurbitapepo L.) is grown in many temperate and sub-tropical regions, and ishighly ranked in economic importance worldwide. One-third of theworld's production of squash is from Turkey, Italy and Egypt (Paris,1996). In recent years, the agricultural sector began to face a problemof water shortage across the globe. The rapid population growth andgreater incidence of drought caused by climate change and differenthuman activities have caused this problem (World Bank, 2006). Betteragricultural practices and enhanced understandings ofwater productiv-ity are some of themeans that could lead to the successful managementof a limited amount of water available for agricultural uses (Howell,2001; Jones, 2004). Among the water management practices for in-creasing water use efficiency (WUE), particularly at field scale, com-bined practices of deficit irrigation (DI) and providing crop plants withsome antioxidants, including the use of salicylic acid for seed soakingand/or as foliar sprays, appears to be very promising.
Irrigation with water below the optimum crop requirements is astrategy for water-saving by which crops are subjected to a certain
geed).
ghts reserved.
level of water stress either during a particular period or throughoutthe whole growing season (Pereira et al., 2002). The increase in theWUE by reducing the amount of water applied and/or by reducing thenumber of irrigation events is the main objective of applying the DIstrategy (Kirda, 2002). Effects of DI on growth and productivity ofmany vegetables and field crops have been intensively investigated(Fereres and Soriano, 2007; Abd El-Mageed and Semida 2015b).Where the effects of DI are crop-specific, it is necessary to evaluate theimpact of DI strategies with multi-year open field experiments beforegeneralizing the most appropriate irrigation scheduling method to beadapted in a specific location for a given crop (Scholberg et al., 2000;Igbadun et al., 2008; Abd El-Mageed and Semida 2015a). It has beenshown that DI is successful in increasingwater productivity for differentcrops without causing severe yield reduction (Geerts and Raes, 2009).
Undoubtedly, water deficit (WD) is one of the most common andself-evident observations. It is well known that WD stress increasesthe production of reactive oxygen species (ROS; superoxide, H2O2, sin-glet oxygen and hydroxyl radical) in cellular organelles such as chloro-plasts, peroxisomes and mitochondria, negatively affecting variousprocesses such as transpiration, photosynthesis, stomatal conductanceand growth (Alscher et al., 1997; Batra et al., 2014). Extensive cellulardamage and deathwill occur ifWD stress is prolonged because ROS pro-duction will defeat the scavenging action of the antioxidant system

9T.A. Abd El-Mageed et al. / South African Journal of Botany 106 (2016) 8–16
(Cruz de Carvalho, 2008). One of the physiological processes that occurwhen plant is subjected to WD stress is the stomatal closure in order toavoid water losses through transpiration (Ache et al., 2010). Whenadopting deficit irrigation strategies, WD stress applied could havesome negative impacts on growth and productivity as reported byEl-Dewiny (2011) on summer squash, and applying the stressed plantswith some antioxidants such as salicylic acid will help crop plants totolerate WD stress.
Salicylic acid (SA) is classified as a phenolic growth regulator, knownas an antioxidant compound that can regulate plant growth, prohibitingthe activity of ROS (Amanullah et al., 2010; Hayat et al., 2010). It isalso recognized as a signal molecule, and has been intensively investi-gated for its role in plant adaptation to the adverse effects that occurin growing environments. It is found to improve plant tolerance tovarious environmental stresses (Manaa et al., 2014; Semida et al.,2015), including water stress (Singh and Usha, 2003; Hesami et al.,2013). It is well known that SA ameliorates the impairments that arisefromWD in plants (Hussain et al., 2009). It is involved in different pro-cesses in plants such as stomatal conductance, membrane permeability,plant water relations and nutrient uptake and transport (Gunes et al.,2005; Hayat and Ahmad, 2007; Hayat et al., 2010). The effect of SA onplant physiological processes varies depending on species, develop-mental stage, SA concentration and environmental conditions (Shraiyand Hegazi, 2009). Water use efficiency, rate of transpiration and inter-nal CO2 increased after supplementation with SA (Kumar et al., 2000).Different studies showed that plant growth, productivity and biochem-ical attributes in various crops have been improved by SA applicationunder water deficit stress (Khan et al., 2003; Shirani Bidabadi et al.,2012; Hesami et al., 2013).
The current study aims to assess the efficacy of foliar-applied SAunder deficit irrigation strategies on water use efficiency, growth andproductivity of squash as a model plant in different growing seasons.
2. Materials and methods
2.1. Experimental site
Two experiments were conducted in both growing summer season(SS) and growing fall season (FS) of 2013, at farmer's field located inEl-Fayoum province, which occupies a depression west of the Nile at90 km southwest of Cairo, Egypt between latitudes 29°02′ and 29°35′Nand longitudes 30°23′ and 31°05′E. Supplementary Table 1 shows theclimatic data of El-Fayoum during the months of the study. Accordingto the aridity index (Ponce et al., 2000), the area is located under ahyper-arid climate. Some initial physical and chemical properties of theirrigation water and the experimental site soil were analyzed accordingto Klute (1986) and Page et al. (1982) and are given in SupplementaryTables 2–4. According to Ayers and Wesctcot (1985), the scale used forthe irrigation water lies within the second categories for salinity andsodicity levels (C2S1, ECiw = 0.75–3.00 dS m−1 and SAR b 6.0).
2.2. Irrigation water applied (IWA)
The squash plants were irrigated at 2-day intervals with differentamounts of irrigation water. IWA was specified as a percentage ofthe crop evapotranspiration (ETc) representing one of the followingthree treatments: I100 = 100%, I80 = 80% and I60 = 60% of ETc. Thedaily ETo was computed according to Eq. (1) (Allen et al., 1998) asfollows:
ETo ¼0:408 Δ Rn−Gð Þ þ γ
900Tmean þ 273
u2 es−eað ÞΔþ γ 1þ 0:34 u2ð Þ ð1Þ
where ETo is the reference evapotranspiration (mm day−1), Δ isthe slope of the saturation vapor pressure curve at air temperature
(kPa C−1), Rn is the net radiation at the crop surface (MJ m−2 d−1),G is soil heat flux density (MJ m−2 d−1), γ is the psychometricconstant = (0.665 × 10−3 × P) kPa C−1 (Allen et al., 1998), P is the at-mospheric pressure (kPa), u2 is thewind speed at 2mheight (m s−1), esis the saturation vapor pressure (kPa), ea. is the actual vapor pressure(kPa), (es − ea) is the saturation vapor pressure deficit (kPa), andTmean is the mean daily air temperature at 2 m height (°C).
The average of daily ETo in El-Fayoum was 10.16, 10.74, 10.66,9.90, 8.64, 6.61, 4.63 and 3.49 mm day−1 in May, June, July, August,September, October, November and December, respectively. The cropwater requirements (ETc) were estimated using the crop coefficient ac-cording to Eq. (2):
ETc ¼ ETo� Kc ð2Þ
where ETc is the cropwater requirement (mmday−1) andKc is the cropcoefficient. The duration of the different crop growth stageswere 25, 35,25 and 15 days for initial, crop development, mid-season and lateseason stages, respectively, and the crop coefficients (Kc) of initial, midand end stages were 0.60, 1.00 and 0.75, respectively, according toAllen et al. (1998).
The amount of IWA to each treatment during the irrigation regimewas determined by using Eq. (3) as follows:
IWA ¼ A� ETc� Ii� KrEa� 1000� 1−LRð Þ ð3Þ
where IWA is the irrigation water applied (m3), A is the plot area (m2),ETc is the crop water requirements (mm day−1), Ii is the irrigation in-tervals (day), Ea is the application efficiency (%) (Ea= 85), Kr is the cov-ering factor, and to calculate (Kr), the Decroix and Cecroix method wasused (Eq. (4); Vermeiren and Jobling, 1980):
Kr ¼ 0:10þ GCð Þ≤1 ð4Þ
where GC is the ground cover. In addition, LR is the leaching require-ments, which was estimated according to Eq. (5):
LR ¼ ECw
2MaxECeð5Þ
where ECw is the electrical conductivity of the irrigationwater (dS.m−1)andMaxECe is themaximumelectrical conductivity of the soil saturationextract for a given crop (see the table shown according to Doorenbosand Pruitt (1984) and Keller and Bliesner (1990).
2.3. Salicylic acid (SA) treatments
From a preliminary pot study (data not shown), 1 mM SA and threetime applications (i.e., at 21 and 30 days after sowing) were selected.These concentration and application times generated the best growthof squash plants grown under 20% and 40% water depletion. For themain study, there were 6 treatments including SA that was applied in3 treatments. These treatments were as follows: I100 (irrigation with100% of ETc), I80 (irrigation with 80% of ETc), I60 (irrigation with 60%of ETc), I100 + 1 mM SA, I80 + 1 mM SA and I60 + 1 mM SA.
2.4. Experimental design
The experiments were conducted in a complete randomized blockdesign. In either summer or fall season, the 6 tested treatments werereplicated three times for each, making a total of 18 plots. The experi-mental plot area was 13.2 m2; 12 m length × 1.10 m row width, and0.5 m spacing between plants within rows.

10 T.A. Abd El-Mageed et al. / South African Journal of Botany 106 (2016) 8–16
2.5. Plant management and measurements
Squash hybrid Hi Tech® seeds were sown 0.5 m apart in each bedabout 5 cm away from the drip line at a depth of 4 cm, drip irrigatedwith one line and one dripper per plant giving 4.0 L h−1. Seeds wereplanted on 9 May and 13 September, and terminated on 4 August and7 December in the 2013 summer and fall growing seasons, respectively.All treatments were separated as surrounded by 1m non-irrigated area.Plants were adequately watered during the first irrigation. Irrigationtreatments were initiated one week after full germination.
Chemical fertilization was practiced at the recommended ratefor squash production in this area, 150 kg N ha−1, 60 kg P ha−1 and70 K kg ha−1. The cultural, disease and pest management practiceswere the same as local commercial crop production.
At the end of both summer and fall experiments, 3 individual plantswere randomly chosen from each experimental plot to evaluate growthcharacteristics and another group of 3 plants to determine chemical at-tributes. Shoot lengths were measured from the cotyledonary node tothe terminal bud of the stem, using a meter scale. Number of leaveswas counted per plant, and leaf area per plant was measured usingDigital Planimeter (Planix 7). Shoots of plants were weighed to recordtheir fresh weights, and then placed in an oven at 70 ± 2 °C till a con-stant weight to measure their dry weights.
Membrane stability index (MSI) was determined using duplicate0.2 g samples of fully expanded leaf tissue (Rady, 2011). The leaf samplewas placed in a test tube containing 10ml of double-distilledwater. Thecontent of the test tube was heated to 40 °C in a water bath for 30 min,and the electrical conductivity (C1) of the solution was recorded using aconductivity bridge. A second sample was boiled at 100 °C for 10 min,and the conductivity was measured (C2). The MSI was calculatedusing the formula:
MSI %ð Þ ¼ 1− C1=C2ð Þ½ � � 100
Total leakage of inorganic ions from fully expanded leaves was de-termined using the method of Sullivan and Ross (1979) with somemodifications. Twenty leaf disks (2 cm in diameter) were placed in aboiling tube containing 10 ml deionized water and the electrical con-ductivity (EC1) was recorded. The contents were then heated to 50 °Cin a water bath for 30 min and the electrical conductivity (EC2) was re-corded. The samplewas then boiled at 100 °C for 10min and the electri-cal conductivity (EC3) was recorded. Electrolyte leakage (EL) wascalculated using the formula:
EL %ð Þ ¼ EC2–EC1ð Þ=EC3½ � � 100
Relative water content (RWC) was estimated using 2-cm-diameterfully expanded leaf disks (Hayat et al., 2007). The disks were weighed(fresh mass; FM) and immediately floated on double-distilled water inPetri dishes for 24 h, in the dark, to saturate them with water. Any ad-hering water was blotted dry and the turgid mass (TM) was measured.The dry mass (DM) was recorded after dehydrating the disks at 70 °Cuntil the constant weight. The RWC was then calculated using the fol-lowing formula:
RWC %ð Þ ¼ FM−DMð Þ= TM−DMð Þ½ � � 100
Chlorophyll ‘a’, chlorophyll ‘b’ and carotenoid concentrations wereextracted and determined (in mg g−1 FW) following the proceduregiven by Arnon (1949). Fresh leaf samples (0.2 g) were homogenizedin 50 ml 80% (v/v) acetone, and then centrifuged at 10,000×g for10 min. The absorbance of the acetone extract was measured at 663,645, and 470 nm using a UV-160A UV–visible recording spectrometer(Shimadzu, Kyoto, Japan).
On two different sunny days, chlorophyll fluorescence was mea-sured using a portable fluorometer (Handy PEA, Hansatech Instruments
Ltd., Kings Lynn, UK). One leaf (at the same age)was chosen per plant toconduct the fluorescence measurements. Maximum quantum yield ofPS II Fv/Fm was calculated using the formula: Fv/Fm = (Fm − F0)/Fm(Maxwell and Johnson, 2000). Fv/F0 reflects the efficiency of electrondonation to the PSII RCs and the rate of photosynthetic quantum con-version at PSII RCs. Fv/F0 was calculated using the formula: Fv/F0 =(Fm − F0)/F0 (Spoustová et al., 2013). Performance index of photosyn-thesis based on the equal absorption (PIABS) was calculated as reportedby Clark et al. (2000).
Free proline concentrations were determined using the rapidcolourimetric method outlined by Bates et al. (1973). Dry leaf samples(0.5 g) were extracted by grinding in 10 ml 3% (v/v) sulphosalicylicacid. The mixtures were then centrifuged at 10,000×g for 10 min.Two milliliters of the supernatant was placed in a test tube, to which2 ml of a freshly prepared acidninhydrin solution was added. Thetubes were incubated in a water bath at 90 °C for 30 min and the reac-tionwas terminated in an ice bath. Each reactionmixture was extractedwith 5ml toluene and vortex-mixed for 15 s. The tubeswere allowed tostand for at least 20 min in the dark, at room temperature, to allow sep-aration of the toluene and aqueous phases. Each toluene phasewas thencarefully collected into a clean test tube and its absorbance was read at520 nm. The free proline content in each samplewas determined from astandard curve prepared using analytical grade proline.
Total soluble sugar concentrationwas assessed according to Irigoyenet al., 1992, using a UV-160A UV Visible Recording Spectrometer,Shimadzu, Japan. Cumulative fruits were quantified by harvestingevery 5 days and weighing the fruits on six randomly selected plantsfrom each plot. At each harvest, number of fruits plant−1, fresh fruitweight plant−1, were recorded for each replicate. The first and last har-vest dateswere on the7th of June and 4th of August for SS and the 9th ofOctober and 7th of December for FS, respectively.
2.6. Yields, harvest index (HI) and water use efficiency (WUE)
All plants in each experiment were used to measure the averagenumber of fruits per plant and total yield per hectare. HI was deter-mined as a ratio of fruit yield to total biomass production on a drymass basis.WUEvalues as kg fruits yieldm−3 of appliedwaterwere cal-culated for different treatments after harvest according to the followingequation (Jensen, 1983):
WUE ¼Fuit yield kg:ha‐1
� �
water applied m3:ha‐1� �
2.7. Anatomical study
To study the anatomy of squash leaf, samples were taken from themiddle of the fifth leaf from the apex. Samples were killed and fixed inFAA solution (50 ml 95% ethyl alcohol + 10 ml formalin + 5 ml glacialacetic acid+35ml distilledwater) for 48 h. Sampleswere thenwashedin 50% ethyl alcohol, dehydrated and cleared in tertiary butyl alcohol se-ries, embedded in paraffinwaxof 54 °C–56 °Cm.p. Using a rotarymicro-tome, cross-sections, 20 μ thick, were cut and adhered with Haupt'sadhesive and stained with the crystal violet–erythrosine combination(Sass, 1961). Then, samples were cleared in carbol xylene andmountedin Canada balsam. Using an upright light microscope (AxioPlan, Zeiss,Jena, Germany), the sectionswere observed and documented.Measure-ments were done using a micrometer eyepiece and an average of fivereadings were calculated.
2.8. Statistical analysis
Statistical analysis of the experimental data was carried out usingANOVA procedures in GenStat statistical package (version 11) (VSN
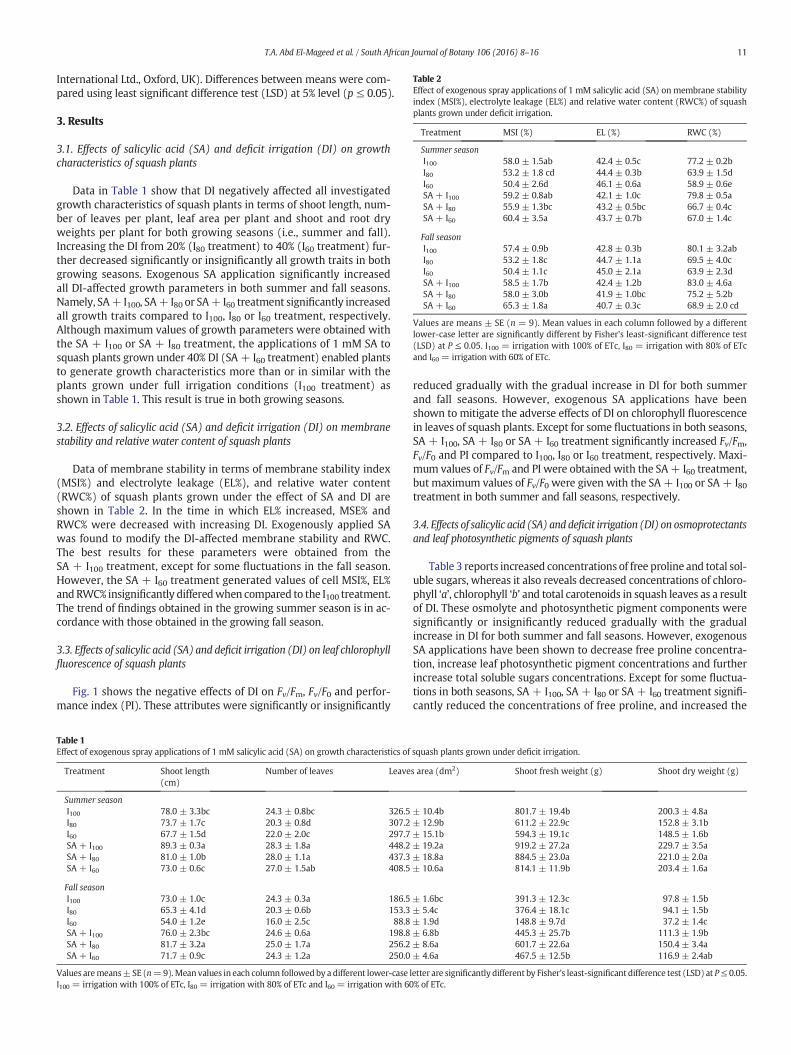
Table 2Effect of exogenous spray applications of 1 mM salicylic acid (SA) on membrane stabilityindex (MSI%), electrolyte leakage (EL%) and relative water content (RWC%) of squashplants grown under deficit irrigation.
Treatment MSI (%) EL (%) RWC (%)
Summer seasonI100 58.0 ± 1.5ab 42.4 ± 0.5c 77.2 ± 0.2bI80 53.2 ± 1.8 cd 44.4 ± 0.3b 63.9 ± 1.5dI60 50.4 ± 2.6d 46.1 ± 0.6a 58.9 ± 0.6eSA + I100 59.2 ± 0.8ab 42.1 ± 1.0c 79.8 ± 0.5aSA + I80 55.9 ± 1.3bc 43.2 ± 0.5bc 66.7 ± 0.4cSA + I60 60.4 ± 3.5a 43.7 ± 0.7b 67.0 ± 1.4c
Fall seasonI100 57.4 ± 0.9b 42.8 ± 0.3b 80.1 ± 3.2abI80 53.2 ± 1.8c 44.7 ± 1.1a 69.5 ± 4.0cI60 50.4 ± 1.1c 45.0 ± 2.1a 63.9 ± 2.3dSA + I100 58.5 ± 1.7b 42.4 ± 1.2b 83.0 ± 4.6aSA + I80 58.0 ± 3.0b 41.9 ± 1.0bc 75.2 ± 5.2bSA + I60 65.3 ± 1.8a 40.7 ± 0.3c 68.9 ± 2.0 cd
Values are means ± SE (n = 9). Mean values in each column followed by a differentlower-case letter are significantly different by Fisher's least-significant difference test(LSD) at P ≤ 0.05. I100 = irrigation with 100% of ETc, I80 = irrigation with 80% of ETcand I60 = irrigation with 60% of ETc.
11T.A. Abd El-Mageed et al. / South African Journal of Botany 106 (2016) 8–16
International Ltd., Oxford, UK). Differences between means were com-pared using least significant difference test (LSD) at 5% level (p ≤ 0.05).
3. Results
3.1. Effects of salicylic acid (SA) and deficit irrigation (DI) on growthcharacteristics of squash plants
Data in Table 1 show that DI negatively affected all investigatedgrowth characteristics of squash plants in terms of shoot length, num-ber of leaves per plant, leaf area per plant and shoot and root dryweights per plant for both growing seasons (i.e., summer and fall).Increasing the DI from 20% (I80 treatment) to 40% (I60 treatment) fur-ther decreased significantly or insignificantly all growth traits in bothgrowing seasons. Exogenous SA application significantly increasedall DI-affected growth parameters in both summer and fall seasons.Namely, SA+ I100, SA+ I80 or SA+ I60 treatment significantly increasedall growth traits compared to I100, I80 or I60 treatment, respectively.Although maximum values of growth parameters were obtained withthe SA + I100 or SA + I80 treatment, the applications of 1 mM SA tosquash plants grown under 40% DI (SA + I60 treatment) enabled plantsto generate growth characteristics more than or in similar with theplants grown under full irrigation conditions (I100 treatment) asshown in Table 1. This result is true in both growing seasons.
3.2. Effects of salicylic acid (SA) and deficit irrigation (DI) on membranestability and relative water content of squash plants
Data of membrane stability in terms of membrane stability index(MSI%) and electrolyte leakage (EL%), and relative water content(RWC%) of squash plants grown under the effect of SA and DI areshown in Table 2. In the time in which EL% increased, MSE% andRWC% were decreased with increasing DI. Exogenously applied SAwas found to modify the DI-affected membrane stability and RWC.The best results for these parameters were obtained from theSA + I100 treatment, except for some fluctuations in the fall season.However, the SA + I60 treatment generated values of cell MSI%, EL%and RWC% insignificantly differedwhen compared to the I100 treatment.The trend of findings obtained in the growing summer season is in ac-cordance with those obtained in the growing fall season.
3.3. Effects of salicylic acid (SA) and deficit irrigation (DI) on leaf chlorophyllfluorescence of squash plants
Fig. 1 shows the negative effects of DI on Fv/Fm, Fv/F0 and perfor-mance index (PI). These attributes were significantly or insignificantly
Table 1Effect of exogenous spray applications of 1 mM salicylic acid (SA) on growth characteristics of
Treatment Shoot length(cm)
Number of leaves Leave
Summer seasonI100 78.0 ± 3.3bc 24.3 ± 0.8bc 326.5I80 73.7 ± 1.7c 20.3 ± 0.8d 307.2I60 67.7 ± 1.5d 22.0 ± 2.0c 297.7SA + I100 89.3 ± 0.3a 28.3 ± 1.8a 448.2SA + I80 81.0 ± 1.0b 28.0 ± 1.1a 437.3SA + I60 73.0 ± 0.6c 27.0 ± 1.5ab 408.5
Fall seasonI100 73.0 ± 1.0c 24.3 ± 0.3a 186.5I80 65.3 ± 4.1d 20.3 ± 0.6b 153.3I60 54.0 ± 1.2e 16.0 ± 2.5c 88.8SA + I100 76.0 ± 2.3bc 24.6 ± 0.6a 198.8SA + I80 81.7 ± 3.2a 25.0 ± 1.7a 256.2SA + I60 71.7 ± 0.9c 24.3 ± 1.2a 250.0
Values aremeans±SE (n=9).Mean values in each column followedby a different lower-case lI100 = irrigation with 100% of ETc, I80 = irrigation with 80% of ETc and I60 = irrigation with 60
reduced gradually with the gradual increase in DI for both summerand fall seasons. However, exogenous SA applications have beenshown to mitigate the adverse effects of DI on chlorophyll fluorescencein leaves of squash plants. Except for some fluctuations in both seasons,SA + I100, SA + I80 or SA + I60 treatment significantly increased Fv/Fm,Fv/F0 and PI compared to I100, I80 or I60 treatment, respectively. Maxi-mum values of Fv/Fm and PI were obtained with the SA+ I60 treatment,but maximum values of Fv/F0 were given with the SA + I100 or SA + I80treatment in both summer and fall seasons, respectively.
3.4. Effects of salicylic acid (SA) and deficit irrigation (DI) on osmoprotectantsand leaf photosynthetic pigments of squash plants
Table 3 reports increased concentrations of free proline and total sol-uble sugars, whereas it also reveals decreased concentrations of chloro-phyll ‘a’, chlorophyll ‘b’ and total carotenoids in squash leaves as a resultof DI. These osmolyte and photosynthetic pigment components weresignificantly or insignificantly reduced gradually with the gradualincrease in DI for both summer and fall seasons. However, exogenousSA applications have been shown to decrease free proline concentra-tion, increase leaf photosynthetic pigment concentrations and furtherincrease total soluble sugars concentrations. Except for some fluctua-tions in both seasons, SA + I100, SA + I80 or SA + I60 treatment signifi-cantly reduced the concentrations of free proline, and increased the
squash plants grown under deficit irrigation.
s area (dm2) Shoot fresh weight (g) Shoot dry weight (g)
± 10.4b 801.7 ± 19.4b 200.3 ± 4.8a± 12.9b 611.2 ± 22.9c 152.8 ± 3.1b± 15.1b 594.3 ± 19.1c 148.5 ± 1.6b± 19.2a 919.2 ± 27.2a 229.7 ± 3.5a± 18.8a 884.5 ± 23.0a 221.0 ± 2.0a± 10.6a 814.1 ± 11.9b 203.4 ± 1.6a
± 1.6bc 391.3 ± 12.3c 97.8 ± 1.5b± 5.4c 376.4 ± 18.1c 94.1 ± 1.5b± 1.9d 148.8 ± 9.7d 37.2 ± 1.4c± 6.8b 445.3 ± 25.7b 111.3 ± 1.9b± 8.6a 601.7 ± 22.6a 150.4 ± 3.4a± 4.6a 467.5 ± 12.5b 116.9 ± 2.4ab
etter are significantly different by Fisher's least-significant difference test (LSD) at P≤ 0.05.% of ETc.
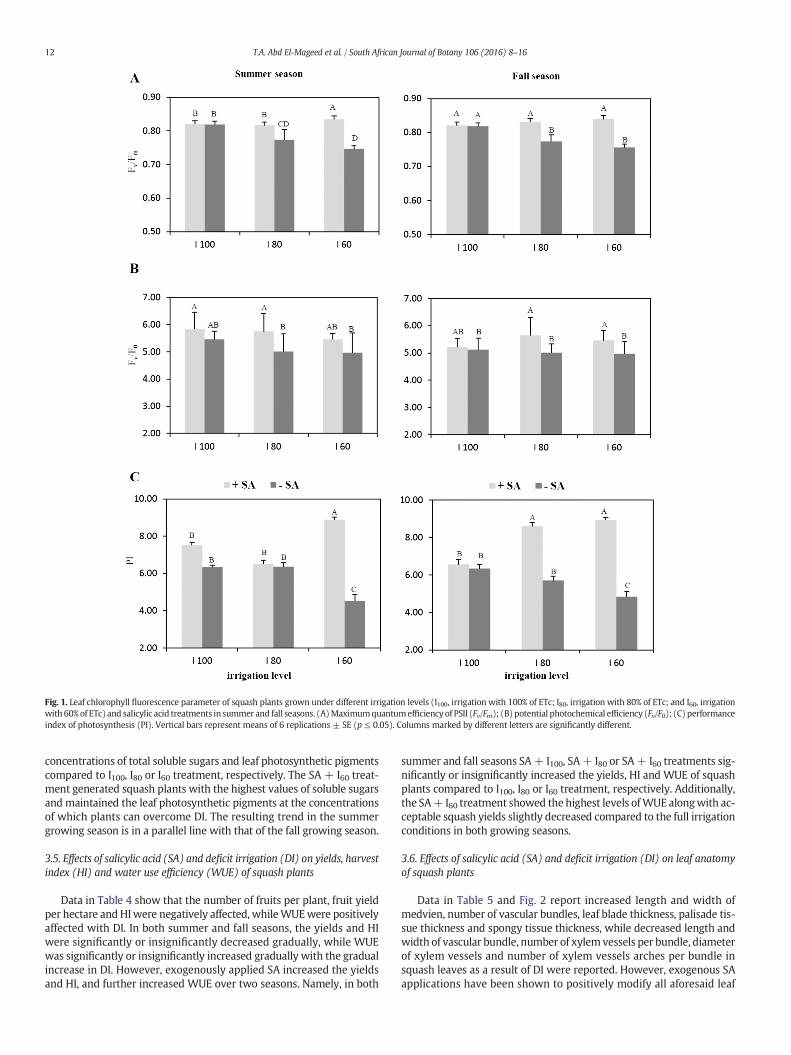
Fig. 1. Leaf chlorophyll fluorescence parameter of squash plants grown under different irrigation levels (I100, irrigation with 100% of ETc; I80, irrigation with 80% of ETc; and I60, irrigationwith 60% of ETc) and salicylic acid treatments in summer and fall seasons. (A)Maximumquantumefficiency of PSII (Fv/Fm); (B) potential photochemical efficiency (Fv/F0); (C) performanceindex of photosynthesis (PI). Vertical bars represent means of 6 replications ± SE (p ≤ 0.05). Columns marked by different letters are significantly different.
12 T.A. Abd El-Mageed et al. / South African Journal of Botany 106 (2016) 8–16
concentrations of total soluble sugars and leaf photosynthetic pigmentscompared to I100, I80 or I60 treatment, respectively. The SA + I60 treat-ment generated squash plants with the highest values of soluble sugarsand maintained the leaf photosynthetic pigments at the concentrationsof which plants can overcome DI. The resulting trend in the summergrowing season is in a parallel line with that of the fall growing season.
3.5. Effects of salicylic acid (SA) and deficit irrigation (DI) on yields, harvestindex (HI) and water use efficiency (WUE) of squash plants
Data in Table 4 show that the number of fruits per plant, fruit yieldper hectare and HIwere negatively affected, whileWUEwere positivelyaffected with DI. In both summer and fall seasons, the yields and HIwere significantly or insignificantly decreased gradually, while WUEwas significantly or insignificantly increased gradually with the gradualincrease in DI. However, exogenously applied SA increased the yieldsand HI, and further increased WUE over two seasons. Namely, in both
summer and fall seasons SA+ I100, SA+ I80 or SA+ I60 treatments sig-nificantly or insignificantly increased the yields, HI and WUE of squashplants compared to I100, I80 or I60 treatment, respectively. Additionally,the SA+ I60 treatment showed the highest levels ofWUE alongwith ac-ceptable squash yields slightly decreased compared to the full irrigationconditions in both growing seasons.
3.6. Effects of salicylic acid (SA) and deficit irrigation (DI) on leaf anatomyof squash plants
Data in Table 5 and Fig. 2 report increased length and width ofmedvien, number of vascular bundles, leaf blade thickness, palisade tis-sue thickness and spongy tissue thickness, while decreased length andwidth of vascular bundle, number of xylem vessels per bundle, diameterof xylem vessels and number of xylem vessels arches per bundle insquash leaves as a result of DI were reported. However, exogenous SAapplications have been shown to positively modify all aforesaid leaf
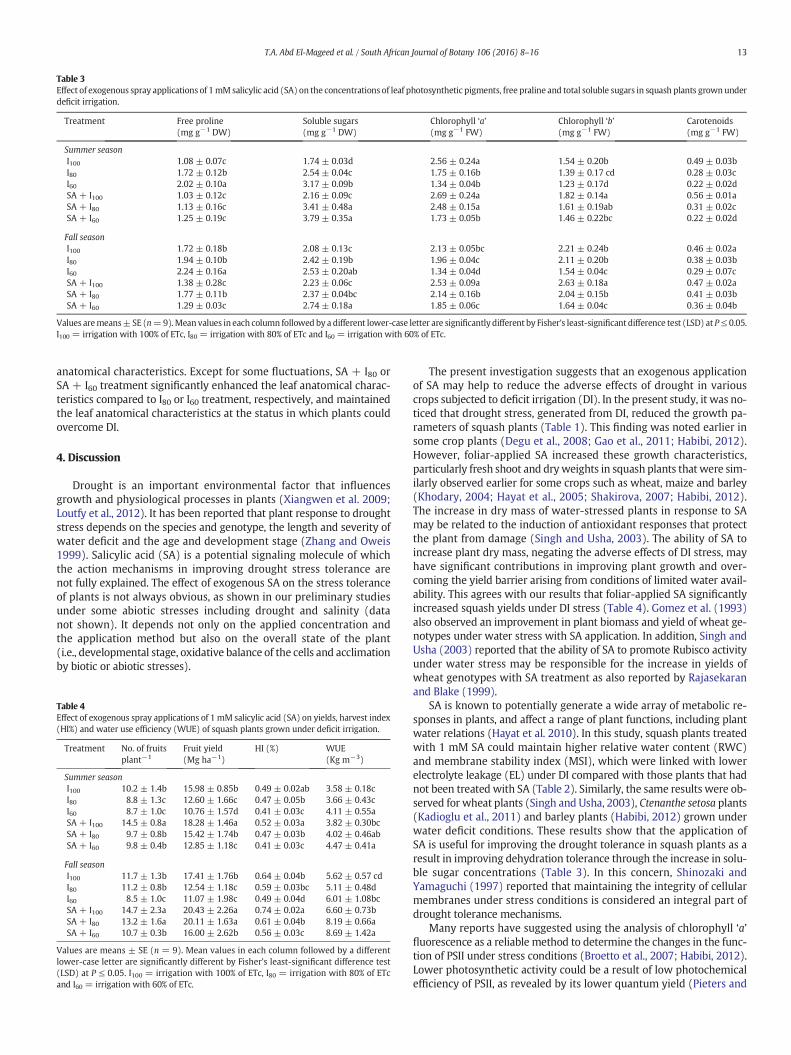
Table 3Effect of exogenous spray applications of 1mM salicylic acid (SA) on the concentrations of leaf photosynthetic pigments, free praline and total soluble sugars in squash plants grownunderdeficit irrigation.
Treatment Free proline(mg g−1 DW)
Soluble sugars(mg g−1 DW)
Chlorophyll ‘a’(mg g−1 FW)
Chlorophyll ‘b’(mg g−1 FW)
Carotenoids(mg g−1 FW)
Summer seasonI100 1.08 ± 0.07c 1.74 ± 0.03d 2.56 ± 0.24a 1.54 ± 0.20b 0.49 ± 0.03bI80 1.72 ± 0.12b 2.54 ± 0.04c 1.75 ± 0.16b 1.39 ± 0.17 cd 0.28 ± 0.03cI60 2.02 ± 0.10a 3.17 ± 0.09b 1.34 ± 0.04b 1.23 ± 0.17d 0.22 ± 0.02dSA + I100 1.03 ± 0.12c 2.16 ± 0.09c 2.69 ± 0.24a 1.82 ± 0.14a 0.56 ± 0.01aSA + I80 1.13 ± 0.16c 3.41 ± 0.48a 2.48 ± 0.15a 1.61 ± 0.19ab 0.31 ± 0.02cSA + I60 1.25 ± 0.19c 3.79 ± 0.35a 1.73 ± 0.05b 1.46 ± 0.22bc 0.22 ± 0.02d
Fall seasonI100 1.72 ± 0.18b 2.08 ± 0.13c 2.13 ± 0.05bc 2.21 ± 0.24b 0.46 ± 0.02aI80 1.94 ± 0.10b 2.42 ± 0.19b 1.96 ± 0.04c 2.11 ± 0.20b 0.38 ± 0.03bI60 2.24 ± 0.16a 2.53 ± 0.20ab 1.34 ± 0.04d 1.54 ± 0.04c 0.29 ± 0.07cSA + I100 1.38 ± 0.28c 2.23 ± 0.06c 2.53 ± 0.09a 2.63 ± 0.18a 0.47 ± 0.02aSA + I80 1.77 ± 0.11b 2.37 ± 0.04bc 2.14 ± 0.16b 2.04 ± 0.15b 0.41 ± 0.03bSA + I60 1.29 ± 0.03c 2.74 ± 0.18a 1.85 ± 0.06c 1.64 ± 0.04c 0.36 ± 0.04b
Values aremeans±SE (n=9).Mean values in each column followed by a different lower-case letter are significantly different by Fisher's least-significant difference test (LSD) at P≤ 0.05.I100 = irrigation with 100% of ETc, I80 = irrigation with 80% of ETc and I60 = irrigation with 60% of ETc.
13T.A. Abd El-Mageed et al. / South African Journal of Botany 106 (2016) 8–16
anatomical characteristics. Except for some fluctuations, SA + I80 orSA + I60 treatment significantly enhanced the leaf anatomical charac-teristics compared to I80 or I60 treatment, respectively, and maintainedthe leaf anatomical characteristics at the status in which plants couldovercome DI.
4. Discussion
Drought is an important environmental factor that influencesgrowth and physiological processes in plants (Xiangwen et al. 2009;Loutfy et al., 2012). It has been reported that plant response to droughtstress depends on the species and genotype, the length and severity ofwater deficit and the age and development stage (Zhang and Oweis1999). Salicylic acid (SA) is a potential signaling molecule of whichthe action mechanisms in improving drought stress tolerance arenot fully explained. The effect of exogenous SA on the stress toleranceof plants is not always obvious, as shown in our preliminary studiesunder some abiotic stresses including drought and salinity (datanot shown). It depends not only on the applied concentration andthe application method but also on the overall state of the plant(i.e., developmental stage, oxidative balance of the cells and acclimationby biotic or abiotic stresses).
Table 4Effect of exogenous spray applications of 1 mM salicylic acid (SA) on yields, harvest index(HI%) and water use efficiency (WUE) of squash plants grown under deficit irrigation.
Treatment No. of fruitsplant−1
Fruit yield(Mg ha−1)
HI (%) WUE(Kg m−3)
Summer seasonI100 10.2 ± 1.4b 15.98 ± 0.85b 0.49 ± 0.02ab 3.58 ± 0.18cI80 8.8 ± 1.3c 12.60 ± 1.66c 0.47 ± 0.05b 3.66 ± 0.43cI60 8.7 ± 1.0c 10.76 ± 1.57d 0.41 ± 0.03c 4.11 ± 0.55aSA + I100 14.5 ± 0.8a 18.28 ± 1.46a 0.52 ± 0.03a 3.82 ± 0.30bcSA + I80 9.7 ± 0.8b 15.42 ± 1.74b 0.47 ± 0.03b 4.02 ± 0.46abSA + I60 9.8 ± 0.4b 12.85 ± 1.18c 0.41 ± 0.03c 4.47 ± 0.41a
Fall seasonI100 11.7 ± 1.3b 17.41 ± 1.76b 0.64 ± 0.04b 5.62 ± 0.57 cdI80 11.2 ± 0.8b 12.54 ± 1.18c 0.59 ± 0.03bc 5.11 ± 0.48dI60 8.5 ± 1.0c 11.07 ± 1.98c 0.49 ± 0.04d 6.01 ± 1.08bcSA + I100 14.7 ± 2.3a 20.43 ± 2.26a 0.74 ± 0.02a 6.60 ± 0.73bSA + I80 13.2 ± 1.6a 20.11 ± 1.63a 0.61 ± 0.04b 8.19 ± 0.66aSA + I60 10.7 ± 0.3b 16.00 ± 2.62b 0.56 ± 0.03c 8.69 ± 1.42a
Values are means ± SE (n = 9). Mean values in each column followed by a differentlower-case letter are significantly different by Fisher's least-significant difference test(LSD) at P ≤ 0.05. I100 = irrigation with 100% of ETc, I80 = irrigation with 80% of ETcand I60 = irrigation with 60% of ETc.
The present investigation suggests that an exogenous applicationof SA may help to reduce the adverse effects of drought in variouscrops subjected to deficit irrigation (DI). In the present study, it was no-ticed that drought stress, generated from DI, reduced the growth pa-rameters of squash plants (Table 1). This finding was noted earlier insome crop plants (Degu et al., 2008; Gao et al., 2011; Habibi, 2012).However, foliar-applied SA increased these growth characteristics,particularly fresh shoot and dryweights in squash plants that were sim-ilarly observed earlier for some crops such as wheat, maize and barley(Khodary, 2004; Hayat et al., 2005; Shakirova, 2007; Habibi, 2012).The increase in dry mass of water-stressed plants in response to SAmay be related to the induction of antioxidant responses that protectthe plant from damage (Singh and Usha, 2003). The ability of SA toincrease plant dry mass, negating the adverse effects of DI stress, mayhave significant contributions in improving plant growth and over-coming the yield barrier arising from conditions of limited water avail-ability. This agrees with our results that foliar-applied SA significantlyincreased squash yields under DI stress (Table 4). Gomez et al. (1993)also observed an improvement in plant biomass and yield of wheat ge-notypes under water stress with SA application. In addition, Singh andUsha (2003) reported that the ability of SA to promote Rubisco activityunder water stress may be responsible for the increase in yields ofwheat genotypes with SA treatment as also reported by Rajasekaranand Blake (1999).
SA is known to potentially generate a wide array of metabolic re-sponses in plants, and affect a range of plant functions, including plantwater relations (Hayat et al. 2010). In this study, squash plants treatedwith 1 mM SA could maintain higher relative water content (RWC)and membrane stability index (MSI), which were linked with lowerelectrolyte leakage (EL) under DI compared with those plants that hadnot been treated with SA (Table 2). Similarly, the same results were ob-served for wheat plants (Singh and Usha, 2003), Ctenanthe setosa plants(Kadioglu et al., 2011) and barley plants (Habibi, 2012) grown underwater deficit conditions. These results show that the application ofSA is useful for improving the drought tolerance in squash plants as aresult in improving dehydration tolerance through the increase in solu-ble sugar concentrations (Table 3). In this concern, Shinozaki andYamaguchi (1997) reported that maintaining the integrity of cellularmembranes under stress conditions is considered an integral part ofdrought tolerance mechanisms.
Many reports have suggested using the analysis of chlorophyll ‘a’fluorescence as a reliable method to determine the changes in the func-tion of PSII under stress conditions (Broetto et al., 2007; Habibi, 2012).Lower photosynthetic activity could be a result of low photochemicalefficiency of PSII, as revealed by its lower quantum yield (Pieters and
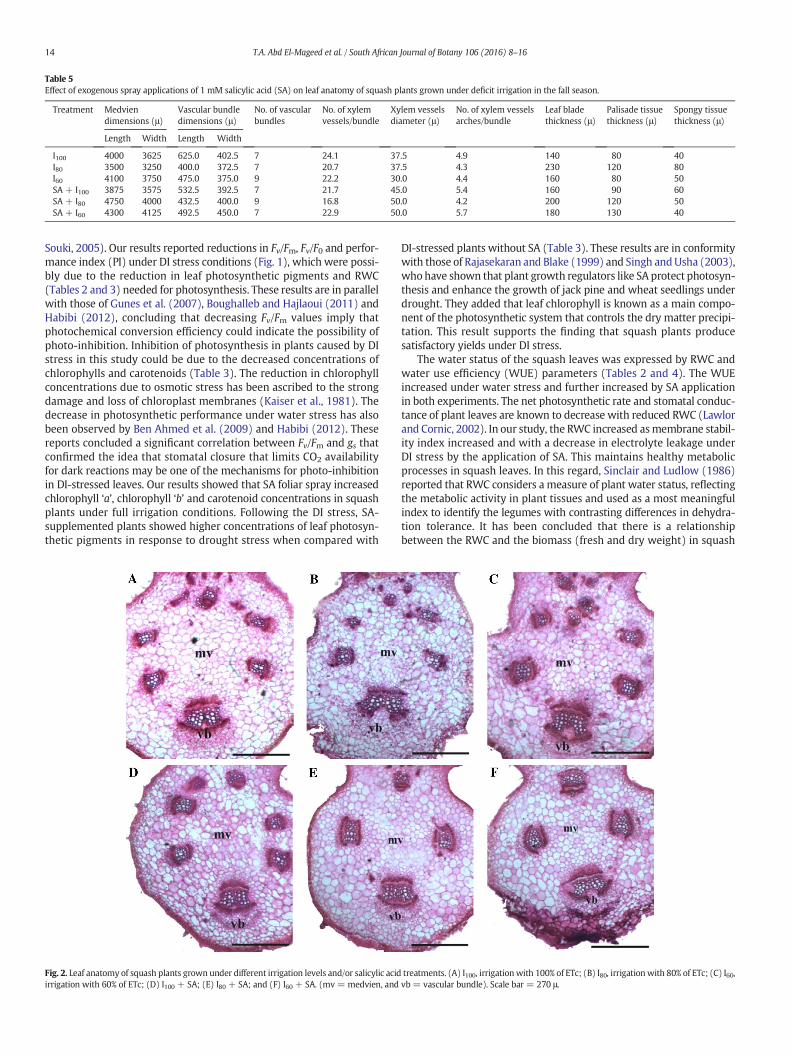
Table 5Effect of exogenous spray applications of 1 mM salicylic acid (SA) on leaf anatomy of squash plants grown under deficit irrigation in the fall season.
Treatment Medviendimensions (μ)
Vascular bundledimensions (μ)
No. of vascularbundles
No. of xylemvessels/bundle
Xylem vesselsdiameter (μ)
No. of xylem vesselsarches/bundle
Leaf bladethickness (μ)
Palisade tissuethickness (μ)
Spongy tissuethickness (μ)
Length Width Length Width
I100 4000 3625 625.0 402.5 7 24.1 37.5 4.9 140 80 40I80 3500 3250 400.0 372.5 7 20.7 37.5 4.3 230 120 80I60 4100 3750 475.0 375.0 9 22.2 30.0 4.4 160 80 50SA + I100 3875 3575 532.5 392.5 7 21.7 45.0 5.4 160 90 60SA + I80 4750 4000 432.5 400.0 9 16.8 50.0 4.2 200 120 50SA + I60 4300 4125 492.5 450.0 7 22.9 50.0 5.7 180 130 40
14 T.A. Abd El-Mageed et al. / South African Journal of Botany 106 (2016) 8–16
Souki, 2005). Our results reported reductions in Fv/Fm, Fv/F0 and perfor-mance index (PI) under DI stress conditions (Fig. 1), which were possi-bly due to the reduction in leaf photosynthetic pigments and RWC(Tables 2 and 3) needed for photosynthesis. These results are in parallelwith those of Gunes et al. (2007), Boughalleb and Hajlaoui (2011) andHabibi (2012), concluding that decreasing Fv/Fm values imply thatphotochemical conversion efficiency could indicate the possibility ofphoto-inhibition. Inhibition of photosynthesis in plants caused by DIstress in this study could be due to the decreased concentrations ofchlorophylls and carotenoids (Table 3). The reduction in chlorophyllconcentrations due to osmotic stress has been ascribed to the strongdamage and loss of chloroplast membranes (Kaiser et al., 1981). Thedecrease in photosynthetic performance under water stress has alsobeen observed by Ben Ahmed et al. (2009) and Habibi (2012). Thesereports concluded a significant correlation between Fv/Fm and gs thatconfirmed the idea that stomatal closure that limits CO2 availabilityfor dark reactions may be one of the mechanisms for photo-inhibitionin DI-stressed leaves. Our results showed that SA foliar spray increasedchlorophyll ‘a’, chlorophyll ‘b’ and carotenoid concentrations in squashplants under full irrigation conditions. Following the DI stress, SA-supplemented plants showed higher concentrations of leaf photosyn-thetic pigments in response to drought stress when compared with
Fig. 2. Leaf anatomy of squash plants grown under different irrigation levels and/or salicylic acidirrigation with 60% of ETc; (D) I100 + SA; (E) I80 + SA; and (F) I60 + SA. (mv= medvien, and
DI-stressed plants without SA (Table 3). These results are in conformitywith those of Rajasekaran and Blake (1999) and Singh andUsha (2003),whohave shown that plant growth regulators like SA protect photosyn-thesis and enhance the growth of jack pine and wheat seedlings underdrought. They added that leaf chlorophyll is known as a main compo-nent of the photosynthetic system that controls the dry matter precipi-tation. This result supports the finding that squash plants producesatisfactory yields under DI stress.
The water status of the squash leaves was expressed by RWC andwater use efficiency (WUE) parameters (Tables 2 and 4). The WUEincreased under water stress and further increased by SA applicationin both experiments. The net photosynthetic rate and stomatal conduc-tance of plant leaves are known to decrease with reduced RWC (Lawlorand Cornic, 2002). In our study, the RWC increased asmembrane stabil-ity index increased and with a decrease in electrolyte leakage underDI stress by the application of SA. This maintains healthy metabolicprocesses in squash leaves. In this regard, Sinclair and Ludlow (1986)reported that RWC considers a measure of plant water status, reflectingthe metabolic activity in plant tissues and used as a most meaningfulindex to identify the legumes with contrasting differences in dehydra-tion tolerance. It has been concluded that there is a relationshipbetween the RWC and the biomass (fresh and dry weight) in squash
treatments. (A) I100, irrigationwith 100% of ETc; (B) I80, irrigationwith 80% of ETc; (C) I60,vb = vascular bundle). Scale bar = 270 μ.

15T.A. Abd El-Mageed et al. / South African Journal of Botany 106 (2016) 8–16
under the interaction effect of DI stress and SA application, indicatingthat the water status in squash leaves is basically dependent on the re-spective shoot biomass. This also suggests that squash plantswith great-er biomass can maintain higher water content in their leaves, and thuscan bemore tolerant to drought. This parameter (RWCvs. plant biomassor growth) could be used as a convenient evidence to distinguish thespecific and non-specific traits for drought tolerance in squash plants.
SA significantly increased the total concentrations of soluble sugars,while it decreased proline concentrations in squash leaves under DIstress (Table 3). Thus, it was concluded that the SA-increased concen-trations of soluble sugars might contribute as a solute for the osmoticregulation and/or a substrate for the protein and polysaccharide synthe-ses in roots, and thereby for the growth of whole plants (Loutfy et al.,2012). SA stimulated the various parameters in leaves and consequentlyin shoots. These changes can be explained as the result of SA-enhancedgrowth or biomass of shoots. From these results, we conclude that SAmay be effective for growth and related attributes under drought, andhence promote growth and leaf function in shoots. Loutfy et al. (2012)reported that no direct evidence was found for ROS generation inwheat roots; however, SA has been reported to induce a rapid and tran-sient generation of ROS and then elevate the cytosolic Ca concentrationby activating Ca channel on membranes in tobacco cells (Kawano et al.1998).Mori et al. (2001) reported on Vicia faba that ROS generation andelevation of Ca levels in leaves are also linked to SA-induced stomatalclosure induced by ABA. In addition, Loutfy et al. (2012) hypothesizedthat drastic Ca level changes in wheat roots could be involved in SA-induced signaling and response in leaves, including stomatal closurethrough its transport from root to shoot. These results signified therole of SA in regulating the drought response of squash and as a poten-tial growth regulator for improving plant growth under DI stress.
As a crucial mechanism, osmotic adjustment is included in plant ad-aptations to various stresses. Proline and soluble sugars are major com-ponents of osmoregulation in the expanded leaves of many species(Mostajeran and Rahimi-Eichi 2009). In the present study, we foundthat their concentrations of soluble sugars significantly increased,while proline concentrationswere reduced in response to the combinedDI and SA treatments (Table 3). Changes in total solute contents supportthe idea that SA more actively stimulates soluble sugar accumulation inroots under drought stress (Loutfy et al., 2012). In contrast to SA appli-cation that reduced them, drought stress greatly increased the prolineconcentrations in squash leaves. From this result, we conclude that SAmay not be involved in the up-regulation of proline biosynthesis forits promoting mechanism of drought tolerance in squash. This findingsuggests that RWC might be dependent on water status of shoots butnot directly on the proline levels in this organ. The increase in solublesugar concentrations under DI stress and their further increase undercombined SA and DI treatments positively correlated with the increasein RWC, contributing to the maintenance of the water status in leavesof squash plants.
It has been concluded that endogenous SA levels increase during thedrought period, which may be associated with oxidative stress indrought-stressedplants (Kadioglu et al., 2011). Increases in endogenousSA levels occur as a response to drought stress (Senaratna et al. 2000;Munne´-Bosch and Penuelas 2003; Kadioglu et al., 2011). Thus, SA treat-ment had a key role in providing tolerance to plants exposed to droughtstress by decreasing thewater loss as a result of the increase in RWC andthe concentrations of soluble sugars.
All enhanced parameters (i.e., growth traits, plant water relations,concentrations of osmoprotectants, WUE and final yields) by the foliarapplication of 1 mM SA were accompanied with the improved leaf ana-tomical characteristics that enabled squash plants to growwell under DIstress. Exposure of squash plants to DI reduced the leaf anatomical char-acteristics, whichmay be due to the inhibition of cellular expansion andelongation (Table 5 and Fig. 2). The main anatomical changes observedin the leaves of plants submitted to DI stress include a thinner leaf bladeresulting from smaller palisade parenchyma cells and a decrease in the
number and size of the spongy parenchyma cells (Gratặo et al., 2009).One of the mechanisms of DI stress effects on cell elongation might bechanges in the cell wall plasticity, which have been reported earlier(Barcelo et al., 1986). In the present study, exogenous application of1 mM SA to squash plants under DI stress enhanced the leaf anatomicalcharacteristics and alleviated the adverse effects of DI stress. The bene-ficial effect of SA on leaf structure may be due to the crucial role in celldivision and cell expansion as shown in Table 5 and Fig. 2.
5. Conclusion
Exposure of squash plants to DI resulted in decreases in plantgrowth, RWC, MSI, leaf photosynthetic pigments, harvest index andyields and increases in the concentrations of soluble sugars and proline,electrolyte leakage and WUE. Overall, the present study revealed thatexogenous SA application could overcome the adverse effects of DIstress by further accumulation of soluble sugars and increasing RWC,acting as osmotic and metabolic regulators or substrates and in a partas cell component stabilizers. These results indicated that SA applicationis useful for improving the growth, anatomical characteristics and yieldof squash crops under DI by 20%–40%.
Appendix A. Supplementary data
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.sajb.2016.05.005.
References
Abd El-Mageed, T.A., Semida, W.M., 2015a. Effect of deficit irrigation and growing seasonson plant water status, fruit yield and water use efficiency of squashunder saline soil.Scientia Horticulturae 186, 89–100.
Abd El-Mageed, T.A., Semida, W.M., 2015b. Organo mineral fertilizer can mitigate waterstress for cucumber production (Cucumis sativus L.). Agricultural Water Management159, 1–10.
Ache, P., Bauer, H., Kollist, H., Al-Rasheid, K.A., Lautner, S., Hartung, W., Hedrich, R., 2010.Stomatal action directly feeds back on leaf turgor: new insights into the regulation ofthe plant water status from non-invasive pressure probe measurements. The PlantJournal 62, 1072–1082.
Allen, R.G., Pereira, L.S., Raes, D., Smith, M., 1998. Crop Evapotranspiration Guidelines forComputing Crop Water Requirements. Irrigation and Drainage Paper 56. FAO, Rome,p. 300.
Alscher, R.G., Donahue, J.L., Cramer, C., 1997. Reactive oxygen species and antioxidants:relationships in green cells. Physiologia Plantarum 100, 224–233.
Amanullah, M.M., Sekar, S., Vincent, S., 2010. Plant growth substances in crop production.Asian Journal of Plant Sciences 9, 215–222.
Arnon, D.I., 1949. Copper enzymes in isolated chloroplasts. Polyphenol-oxidase in Betavulgaris L. Plant Physiology 24, 1–5.
Ayers, R.S., Wesctcot, D.W., 1985. Water Quality for Agriculture, Irrigation and Drainage.Paper No. 29. FAO, Rome, Italy.
Barcelo, J., Poschenrieder, C.H., Andren, I., Gunse, B., 1986. Cadmium induced decrease ofwater stress resistance in bush bean plants (Phaseolus vulgaris cv. Contender). PlantPhysiology 125, 17–25.
Bates, L., Waldren, R.P., Teare, I.D., 1973. Rapid determination of free proline for water-stress studies. Plant and Soil 39, 205–207.
Batra, N.G., Sharma, V., Kumari, N., 2014. Drought-induced changes in chlorophyll fluores-cence, photosynthetic pigments, and thylakoid membrane proteins of Vigna radiate.Journal of Plant Interactions 1, 712–721.
Ben Ahmed, C., Ben Rouinab, B., Sensoyc, S., Boukhrisa, M., Ben Abdallah, F., 2009. Changesin gas exchange, proline accumulation and antioxidative enzyme activities in threeolive cultivars under contrasting water availability regimes. Environmental and Ex-perimental Botany 67, 345–352.
Boughalleb, F., Hajlaoui, H., 2011. Physiological and anatomical changes induced bydrought in two olive cultivars (cv Zalmati and Chemlali). Acta Physiologiae Plantarum33, 53–65.
Broetto, F., Duarte, H.M., Lüttge, Q., 2007. Responses of chlorophyll fluorescence parame-ters of the facultative halophyte and C3-CAM intermediate species Mesembryanthe-mum crystallinum to salinity and high irradiance stress. Journal of Plant Physiology164, 904–912.
Clark, A.J., Landolt, W., Bucher, J.B., Strasser, R.J., 2000. Beech (Fagus sylvatica) response toozone exposure assessed with a chlorophyll a fluorescence performance index. Envi-ronmental Pollution 109, 501–507.
Cruz de Carvalho, M.H., 2008. Drought stress and reactive oxygen species: production,scavenging and signaling. Plant Signaling & Behavior 3, 156–165.
Degu, H.D., Ohta, M., Fujimura, T., 2008. Drought tolerance of Eragrostis tef and develop-ment of roots. International Journal of Plant Sciences 169, 768–775.

16 T.A. Abd El-Mageed et al. / South African Journal of Botany 106 (2016) 8–16
Doorenbos, J., Pruitt, W.O., 1984. Guidelines for Predicting Crop Water Requirements.Irrigation and Drainage Paper 24. FAO, Rome, p. 348.
El-Dewiny, C.Y., 2011. Water and fertilizer use efficiency by squash grown under stress onsandy soil treated with acrylamide hydrogels. Journal of Applied Sciences Research 7,1828–1833.
Fereres, E., Soriano, M.A., 2007. Deficit irrigation for reducing agricultural water use. Spe-cial issue on ‘integrated approaches to sustain and improve plant production underdrought stress’. Journal of Experimental Botany 58, 147–159.
Gao, D., Cai, K., Chen, J., Luo, S., Zeng, R., Yang, J., Zhu, X., 2011. Silicon enhances photo-chemical efficiency and adjusts mineral nutrient absorption in Magnaporthe oryzaeinfected rice plants. Acta Physiologiae Plantarum 33, 675–682.
Geerts, S., Raes, D., 2009. Deficit irrigation as an on-farm strategy to maximize crop waterproductivity in dry areas. Agricultural Water Management 96, 1275–1284.
Gomez, L., Blanca, L., Antonio, C.S., 1993. Evidence of the Beneficent Action of the AcetylSalicylic Acid on Wheat Genotypes Yield under Restricted Irrigation. Proc. ScientificMeeting on Forestry, Livestock and Agriculture Mexico, p. 112.
Gratặo, P.L., Monteiroa, C.C., Rossi, M.B., Martinelli, A.P., Peres, L.E.P., Medici, L.O., Lea, P.J.,Azevedo, R.A., 2009. Differential ultrastructural changes in tomato hormonal mutantsexposed to cadmium. Environmental and Experimental Botany 67, 387–394.
Gunes, A., Inal, A., Alpaslan, M., Cicek, N., Guneri, E., Eraslan, F., Guzelordu, T., 2005. Effectsof exogenously applied salicylic acid on the induction of multiple stress tolerance andmineral nutrition in maize (Zea mays L.). Archives of Agronomy and Soil Science 51,687–695.
Gunes, A., Inal, A., Bagci, E.G., Coban, S., 2007. Silicon mediated changes on some physio-logical and enzymatic parameters symptomatic of oxidative stress in barley grown insodic-B toxic soil. Journal of Plant Physiology 164, 807–811.
Habibi, G., 2012. Exogenous salicylic acid alleviates oxidative damage of barley plantsunder drought stress. Acta Biologica Szegediensis 56, 57–63.
Hayat, S., Ahmad, H., 2007. Salicylic Acid: A Plant Hormone. Springer, Dordrecht, TheNetherlands.
Hayat, S., Fariduddin, Q., Ali, B., Ahmad, A., 2005. Effect of salicylic acid on growth andenzyme activities of wheat seedlings. Acta Agronomica Hungarica 53, 433–437.
Hayat, S., Ali, B., Hasan, S.A., Ahmad, A., 2007. Brassinosteroid enhanced the level of anti-oxidants under cadmium stress in Brassica juncea. Environmental and ExperimentalBotany 60, 33–41.
Hayat, Q., Hayat, S., Irfan, M., Ahmad, A., 2010. Effect of exogenous salicylic acid underchanging environment: a review. Environmental and Experimental Botany 68,14–25.
Hesami, S., Rokhzadi, A., Rahimi, A.R., Hesami, G., Kamangar, H., 2013. Coriander responseto foliar application of salicylic acid and irrigation intervals. International Journal ofBiosciences 3, 35–40.
Howell, T.A., 2001. Enhancing water use efficiency in irrigated agriculture. AgronomyJournal 93, 281–289.
Hussain, M., Malik, M.A., Farooq, M., Khan, M.B., Akram, M., Saleem,M.F., 2009. Exogenousglycinebetaine and salicylic acid application improves water relations, allometry andquality of hybrid sunflower under water deficit conditions. Journal of Agronomy andCrop Science 195, 98–109.
Igbadun, H.E., Salim, B.A., Tarimo, A.K.P.R., Mahoo, H.F., 2008. Effects of deficit irrigationscheduling on yields and soil water balance of irrigated maize. Irrigation Science27, 11–23.
Irigoyen, J.J., Emerich, D.W., Sanchez-Diaz, M., 1992. Water stress induced changes in theconcentrations of proline and total soluble sugars in nodulated alfalfa (Medicagosativa) plants. Plant Physiology 8, 455–460.
Jensen, M.E., 1983. Design and Operation of Farm Irrigation Systems. ASAE, Michigan,USA, p. 827.
Jones, H.G., 2004.What IsWater Use Efficiency? In: Bacon,M.A. (Ed.),Water Use Efficiencyin Plant Biology. Blackwell Publishing, Oxford, UK, pp. 27–41
Kadioglu, A., Saruhan, N., Sağlam, A., Terzi, R., Acet, T., 2011. Exogenous salicylic acid alle-viates effects of long term drought stress and delays leaf rolling by inducing antioxi-dant system. Plant Growth Regulation 64, 27–37.
Kaiser, W.M., Kaiser, G., Schöner, S., Neimanis, S., 1981. Photosynthesis under osmoticstress. Differential recovery of photosynthetic activities of stroma enzymes, intactchloroplasts, and leaf slices after exposure to high solute concentrations. Planta153, 430–435.
Kawano, T., Sahashi, N., Takahashi, K., Uzumi, N., Muto, S., 1998. Salicylic acid induces ex-tracellular superoxide generation followed by an increase in cytosolic calcium ion intobacco suspension culture: the earliest events in salicylic acid signal transduction.Plant and Cell Physiology 39, 721–730.
Keller, J., Bliesner, R.D., 1990. Sprinkle and Trickle Irrigation. An Avi Book. Chapman&Hall,NewYork, p. 629.
Khan, W., Prithiviraj, B., Smith, D.L., 2003. Photosynthetic responses of corn and soybeanto foliar application of salicylates. Journal of Plant Physiology 160, 485–492.
Khodary, S.F.A., 2004. Effect of salicylic acid on the growth, photosynthesis and carbohy-drate metabolism in salt stressed maize plants. International Journal of Agricultureand Biology 6, 5–8.
Kirda, C., 2002. Deficit Irrigation Scheduling Based on Plant Growth Stages ShowingWater Stress Tolerance. Deficit Irrigation Practices. FAO Corp. Doc. Rep. 22, pp. 3–10(Rome)
Klute, A., 1986. Methods of Soil Analysis. Part 1: Physical and Mineralogical Methods.second ed. American Society of Agronomy Madison, Wisconsin, USA.
Kumar, P., Lakshmi, N.J., Mani, V.P., 2000. Interactive effects of salicylic acid and phytohor-mones on photosynthesis and grain yield of soybean (Glycine max L.). Physiologicaland Molecular Biology of Plants 6, 179–186.
Lawlor, D.W., Cornic, G., 2002. Photosynthetic carbon assimilation and associated metab-olism in relation to water deficits in higher plants. Plant, Cell & Environment 25,275–294.
Loutfy, N., El-Tayeb, M.A., Hassanen, A.M., Moustafa, M.F.M., Sakuma, Y., Inouhe, M., 2012.Changes in the water status and osmotic solute contents in response to droughtand salicylic acid treatments in four different cultivars of wheat (Triticum aestivum).Journal of Plant Research 125, 173–184.
Manaa, A., Gharbi, E., Mimouni, H., Wasti, S., Aschi-Smiti, S., Lutts, S., Ben Ahmed, H., 2014.Simultaneous application of salicylic acid and calcium improves salt tolerance in twocontrasting tomato (Solanum lycopersicum) cultivars. South African Journal of Botany95, 32–39.
Maxwell, K., Johnson, G.N., 2000. Chlorophyll fluorescence—a practical guide. Journal ofExperimental Botany 51, 659–668.
Mori, I.C., Pinontoan, R., Kawano, T., Muto, S., 2001. Involvement of superoxide generationin salicylic acid-induced stomatal closure in Vicia faba. Plant and Cell Physiology 42,1383–1388.
Mostajeran, A., Rahimi-Eichi, V., 2009. Effects of drought on growth and yield of rice(Oryza sativa L) cultivars and accumulation of proline and soluble sugars in sheathand blades of their different age leaves. American-Eurasian Journal of Agricultural &Environmental Sciences 5, 264–272.
Munne´-Bosch, S., Penuelas, J., 2003. Photo and antioxidative protection, and a role forsalicylic acid during drought and recovery in field grown Phillyrea angustifolia plants.Planta 217, 758–766.
Page, A.I., Miller, R.H., Keeney, D.R., 1982. Methods of Soil Analysis. Part 2: Chemicaland Microbiological Properties. second ed. American Society of Agronomy Madison,Wisconsin, USA.
Paris, H.S., 1996. Summer squash. History, diversity, and distribution. Horticulture Tech-nology 6, 6–13.
Pereira, L.S., Oweis, T., Zairi, A., 2002. Irrigation management under water scarcity.Agricultural Water Management 57, 175–206.
Pieters, A.J., Souki, S.E., 2005. Effects of drought during grain filling on PSII activity in rice.Journal of Plant Physiology 62, 903–911.
Ponce, V.M., Pandey, R.P., Ercan, S., 2000. Characterization of drought across the climatespectrum. Journal of Hydrologic Engineering, ASCE 5, 222–224.
Rady, M.M., 2011. Effect of 24-epibrassinolide on growth, yield, antioxidant system andcadmium content of bean (Phaseolus vulgaris L.) plants under salinity and cadmiumstress. Scientia Horticulturae 129, 232–237.
Rajasekaran, L.R., Blake, T.J., 1999. New plant growth regulators protect photosynthesisand enhance growth under drought of jack pine seedlings. Journal of Plant GrowthRegulation 18, 175–181.
Sass, J.A., 1961. Botanical Microtechnique. third ed. The Iowa State Univ. Press, Ames,Iowa, USA.
Scholberg, J., McNeal, B.L., Jones, J.W., Boote, K.J., Stanley, C.D., Obreza, T.A., 2000. Growthand canopy characteristics of field-grown tomato. Agronomy Journal 92, 152–159.
Semida, W.M., Rady, M.M., Abd El-Mageed, T.A., Howladar, S.M., Abdel Hamid, M.T., 2015.Alleviation of cadmium toxicity in common bean (Phaseolus vulgaris L.) plants by theexogenous application of salicylic acid. Journal of Horticultural Science & Biotechnol-ogy 90, 83–91.
Senaratna, T., Touvhell, D., Bunn, E., Dixon, K., 2000. Acetyl salicylic acid (aspirin) andsalicylic acid inducemultiple stress tolerance in bean and tomato plant. Plant GrowthRegulation 30, 157–161.
Shakirova, F.M., 2007. Role of Hormonal System in theManifestation of Growth Promotingand Antistress Action of Salicylic Acid. In: Hayat, S., Ahmad, A. (Eds.), Salicylic Acid:a Plant Hormone. Springer, Dordrecht, pp. 69–89.
Shinozaki, K., Yamaguchi, K., 1997. Gene expression and signal transduction in waterstress response. Plant Physiology 115, 327–334.
Shirani Bidabadi, S., Mahmood, M., Baninasab, B., Ghobadi, C., 2012. Influence of salicylicacid on morphological and physiological responses of banana (Musa acuminata cv.‘Berangan’, AAA) shoot tips to in vitro water stress induced by polyethylene glycol.Plant Omics Journal 5, 33–39.
Shraiy, A.M.E., Hegazi, A.M., 2009. Effect of acetylsalicylic acid, indole-3-bytric acid andgibberellic acid on plant growth and yield of pea (Pisum sativum L.). Australian Journalof Basic and Applied Sciences 3, 3514–3523.
Sinclair, T.R., Ludlow,M.M., 1986. Influence of soilwater supply on the plantwater balanceof four tropical grain legumes. Australian Journal of Plant Physiology 13, 329–341.
Singh, B., Usha, K., 2003. Salicylic acid induced physiological and biochemical changes inwheat seedlings under water stress. Plant Growth Regulation 39, 137–141.
Spoustová, P., Synková, H., Valcke, R., Čeřovská, N., 2013. Chlorophyll a fluorescence as atool for a study of the potato virus Y effects on photosynthesis of nontransgenic andtransgenic Pssu-ipt tobacco. Photosynthetica 51, 191–201.
Sullivan, C.Y., Ross, W.M., 1979. Selecting the Drought and Heat Resistance in GrainSorghum. In: Mussel, H., Staples, R.C. (Eds.), Stress Physiology in Crop Plants. JohnWiley & Sons, New York, NY, USA, pp. 263–281.
Vermeiren, L., Jobling, G.A., 1980. Localized Irrigation: Design, Installation, Operation andEvaluation. Irrigation and Drainage Paper No.36. FAO, Roma, Italy.
World Bank, 2006. Directions in Development. Reengaging in Agricultural WaterManagement: Challenges and Options. The International Bank for Reconstructionand Development/The World Bank, The World Bank, Washington, DC, p. 218.
Xiangwen, X., Fan, Y., Sheng, Z., Helena, K., Chunyang, L., 2009. Physiological and proteo-mic responses of two contrasting Populus cathayana populations to drought stress.Physiologia Plantarum 136, 150–168.
Zhang, H., Oweis, T., 1999. Water-yield relations and optimal irrigation scheduling ofwheat in the Mediterranean region. Agricultural Water Management 38, 195–211.





























