Embed Size (px)
Citation preview

(CANCER RESEARCH 53. 2970-2975. July 1. 19931
Sugar Chains of Human Cord Serum a-Fetoprotein: Characteristics of AMinked
Sugar Chains of Glycoproteins Produced in Human Liver and HepatocellularCarcinomas1
Katsuko Yamashita,2 Kazuhisa Taketa, Shinzo Nishi, Keiko Fukushima, and Takashi Ohkura
Department af Biochemistry. Sasaki Institute. Tokyo 101 ¡K.Y.. K. F.. T. O./: Department of Public Health. Okayama University Medical Se/mol. Okayama 700 ¡K.T.¡:and FirstDepartment of Biochemistry Hokkaido University School of Medicine. Sapporo 060 ¡S./V./. Japan
ABSTRACT
Human serum a-fetoprotein (AFP) is elevated in not only hepatocellu-
lar carcinoma (HCC) but also benign liver diseases. AFP produced in HCCand benign liver diseases was separated into several isoforms corresponding to different sugar chain structures by several types of lectin affinityelectrophoresis, and the HCC-specific AFP isoform was discriminatedfrom those of benign liver diseases. Because a small amount of HCC-
specific AFP isoform was detected in cord serum AFP, the whole sugarchain structures of human cord serum AFP were determined, as follows:
Neu5Aca2-»6Galßl-»4GlcNAcßl-»2Manal-»6(Neu5Aca2—¿�6Galßl-'4GlcNAcßl-(2Manal-»3)Manßl-»4R1and R2,
Galßl-*4GlcNAcßl->2Manal-'6(Neu5Aca2->6Galßl-»4GlcNAcßl-»2Manal-»3)Manßl-»4R,and R2, and
Neu5Ac«2—3Galßl—4GlcNAcßl—2Manal-»6(Neu5Aca2-6Galßl-»4GlcNAcßl—2Manal —¿�3)Manßl—4R, and R2
in the ratio of 81.6:8.9:9.5. R, and R, denote GlcNAcßl->4GlcNAcor{subscript OT represents an NaBJH4-reduced oligosaccharide) andGlcNAcßl~>4(Fucal~>6)GlcNACor. respectively, and the ratio between
RI and K. in the respective fractions was approximately 19:1. In contrast,
the sugar chain structure of HCC highly specific AFP isoform was foundto comprise a monosialyl-biantcnnary sugar chain with additional fuco-sylation of the proximal iV-acetylglucosamine. Fucosylation of AFP pro
duced in fetal liver increased in inverse proportion to the gestation period,in weeks, indicating that fucosylation of AFP in HCC may be related to thededifferentiation of human hepatocytes through malignant transformation.
INTRODUCTIONAFP' is the most reliable tumor marker of hepatocellular carcino
mas, being an oncofetal glycoprotein with one asparagine iV-linked
oligosaccharide ( I ). The serum level of AFP sometimes also increasesin LC and CH (2). In order to increase the diagnostic value of thistumor marker, it is important to discriminate AFP which is producedin benign liver diseases, including LC and CH. from that produced inHCC.
Lectin affinity electrophoresis in combination with the antibody-
affinity blotting method opened the way for resolution of this problem(3, 4). AFP produced in HCC and benign liver diseases was separated
Received 3/4/93; accepted 4/27/93.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore he hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1A part of this work was supported by a Granl-in-Aid for Cancer Research from the
Ministry of Education. Science, and Culture of Japan.2 To whom requests for reprints should he addressed, at Sasaki Institute. 2-2 Kanda-
Surugadai. Chiyoda-Ku. Tokyo 101. Japan.1The abbreviations used are: AFP. a-fetoprotein; Neu? Ac. N-acetylneuraminic acid;
Gal. galactose; GlcNAc, A'-acetylglucosamine; Man. mannose; Fuc. i--fucose; HCC. hepa
tocellular carcinoma; LC. liver cirrhosis; CH. chronic hepatitis; Con A. concunavalin A;LCA. lentil leclin; E4-PHA. phytohemagglulinin E4; TJA-I. Trichosanthes japónica ag-glutinin-1; AAL. Alt'iiria anranlia lectin; subscript OT. NaB'H4-reduced oiigosaccharidc;subscript OD. NaB;H4-reduced sugar; TBS-NaN,. K) mvt Tris-HCI buffer. pH 7.4. con
taining 0.02* NaN, and 0.15 M NaCI.
into several isoforms using Con A-, LCA-, and E4-PHA-affinity elec
trophoresis (5). The respective AFP isoforms should correspond todifferent sugar chain structures, because the AFP produced in thehuman body has one asparagine iV-linked oligosaccharide exhibiting
marked alteration under malignant conditions. Among the isoforms ofAFP separated by lectin-affinity electrophoresis, an E4-PHA-reactive
AFP isoform has been shown to have the highest sensitivity. 88%,with regard to the discrimination between benign liver diseases andhepatocellular carcinomas, followed by LCA-reactive AFP with asensitivity of 78% under identical conditions (5). E4-PHA-reactiveAFP becomes positive earlier than LCA-reactive AFP during thefollow-up with diagnostic imaging of cirrhotic patients to monitor the
evolution of hepatocellular carcinomas (6). However, the sugar chainstructures of E4-PHA-reactive AFP have not been elucidated. Since asmall amount of E4-PHA-reactive AFP is present in cord serum, as can
be seen in Fig. 1, the whole sugar chain structure of cord serum AFPwas analyzed in the present study to find oligosaccharides with highaffinities to E4-PHA and with high specificity toward hepatocellular
carcinomas.
MATERIALS AND METHODS
Purification of a-Fetoprotein. AFP was purified from pooled cord serum
at term after a normal pregnancy by affinity chromatography on a column ofan immunoadsorbent coupled with a monoclonal antibody to human AFP (7).
Chemicals, Lectin, and Enzymes. NaB'H4 (490 mCi/mmol) was pur
chased from New England Nuclear (Boston. MA). NaB2H4 (98%) was ob
tained from Merck (Darmstadt. Federal Republic of Germany). ConA-Sepharose was obtained from Pharmacia Biotechnology Inc. (Uppsala.Sweden). Methyl a-i>-glucopyranoside, lactose, L-t'ucose. and Anhmbacter
ureafaciens sialidase were purchased from Nacalai Tesque Inc. (Kyoto, Japan).Bio-Gel P-4 (minus 400 mesh) was purchased from Bio-Rad Laboratories(La Jolla, CA). Diplococcal ß-galactosidase and ß-A'-acetylhexosaminidase
were purified from the culture fluid of Diplococcus pneumonia according tothe method of Glasgow et ill. (8). Salmonella typhimiirium LT2 sialidase waspurchased from Takara Shu«iCo.. Ltd. (Kyoto. Japan). E4-PHA and LCA werepurchased from HöhnenOil (Tokyo. Japan). AAL-Sepharose (7 mg/ml gel) (9)was kindly provided by Dr. Kochibe of Gunma University. TJA-I-Sepharose(3 mg/ml gel) and E4-PHA-Sepharose (4.8 mg/ml gel) were prepared according
to the methods described in the previous papers (10. II ).Oligosaccharides. Neu5Aca2^6Galßl— 4GlcNAcßl^2Manal—
6<Neu5Aca2^6Galßl^4GlcNAcßl —¿�2Manal^3)Manßl— 4GlcNAcßl—4GlcN Ac()T(Neu5 Acn •¿�GaU •¿�GlcN Ac2 •¿�Man, •¿�GlcNAc •¿�GlcNAc„T), Galßl —¿�
3>Manßl^4GlcNAcßl^4GlcNAc„T (Neu5Aca2^6GalvGlcNAcvMan,-GIcNAc-GIcNAcoT), and Galßl^4GlcNAcßl^2Manal->6(Galßl-*4Glc-
NAcßl—¿�2Manal^3) Manßl^4GlcNAcßl^4GlcNAcOT(Gal,-GlcNAcv
ManvGlcNAc-GIcNAcor) were derived from transferrin by hydrazinoly-sis. followed by reduction with NaB'H4 (12). Neu5Ac«2^3Galßl^
4GlcNAcßl -•2Manal -•6(Galßl—¿�4GlcNAcßl -•2Manal —¿�3)Manßl-»4GlcNAcßl—4(Fucal-*6)GlcNAC(,T(Neu5Aca2— 3GalvGlcNAc:-Man,-
GIcNAc-Fuc-GlcNAcor) was derived from recombinant human lymphotoxinproduced by Chinese hamster ovary cells (13). Galßl~~>4GlcNAcßl^>2Manal ^6(Galßl ^4GlcNAcßl ^2Manal -»3)Manßl^4GlcNAcßl —¿�4(F'ucal—'6)GlcNAcOT was derived from urinary ribonuclease L (14).
2970
Research. on December 30, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

SUiAR CHAINS OF CORD SERUM c.-hKTOI'KOH IN
3) Manß|-»4GlcNAcßl—4GlcNAc(),(Man,-GlcNAc-GlcNAcor) and Manal^6(Manal^3)Manßl ^4GlcNAcßl-»4(Fucal^
6)GlcNAcoT (ManyGlcNAc-Fuc'GlcNAcar) were prepared from Gal2-
GIcNAc2-ManyGlcNAc-GlcNAcoT and Gal2-GlcNAc2-Man.,-GlcNAc-Fuc-
GlcNAcoT by digestion with the mixture of diplococcal ß-galactosidase and
ß-/V-acetylhexosaminidase, respectively.Release of Asparagine-linked Sugar Chains from Cord Serum a-Feto-
protein. a-Fetoprotein (20 mg) was hydra/.inoly/ed at I(X)°Cfor 10 h (15).
After iV-acetylation, a quarter of the released oligosaccharides was reducedwith NaB'Hj, and the remaining three quarters was reduced with NaB2H4 for
methylation analysis. The total yield of radioactive oligosaccharides was 3 X10" dpm.
Attimi) Chromatography on Immobilized I rrlin Columns. Columnscontaining 1 ml of AAL-Sepharose, TJA-I-Sepharose, E4-PHA-Sepharose, andCon A-Sepharose were equilibrated with TBS-NaN,. Tritium-labeled oligosac
charides dissolved in 100 ¿ilof TBS-NaN, were applied to the columns, whichwere then allowed to stand at 4°Cfor 15 min. Elution from the E4-PHA column
was performed with 15 ml of TBS-NaN., at 4°C.Elution from the AAL, TJA-I,
and Con A columns was performed with 10 ml of TBS-NaN,, followed by 5
ml of TBS-NaN, containing 5 min L-fucose (AAL column). O.I M lactose(TJA-I column), and 5 ITIMmethyl a-u-glucopyranoside (Con A column),
respectively.Glycosidase Digestion. Radioactive oligosaccharides (0.1-10 nmol) were
digested in one of the following ways at 37°C for 17 h: digestion with
Anhmhcicter sialidase, 100 munits of enzyme in sodium acetate buffer, pH 5.0(40 /il); digestion with a mixture of diplococcal ß-galactosidase and ß-N-
acetylhexosaminidase. 2 munits of each en/.yme in 0.2 M citrate phosphatebuffer. pH 6.0 (20 fj.1). Salmonella typhimurium LT2 sialidase digestion was
performed in a mixture comprising 10 munits of en/.yme in 0.2 M citratephosphate buffer, pH 6.0 (20 /xl), at 37"C for 2 h. The other en/yme digestions
were performed according to the procedures described in the previous paper(16).
Lectin Affinity Electrophoresis. Sera from patients with CH. LC, andHCC and spent culture media of fetal livers were analy/.ed for AFP isoformsby lectin-affinity electrophoresis and antibody-affinity blotting as describedpreviously (4). The percentages of lectin-reactive isoforms of AFP were de
termined by densitometry of the stained bands.Other Analytical Methods. Methylation analysis of oligosaccharides was
performed as reported in the preceding paper (13. 17). High-voltage paper
electrophoresis was performed with pyridine/acetate buffer, pH 5.4 (pyridine/acetic acid/water, 3:1:387), at a potential of 73 V/cm for 90 min. Radiochro-
matoscanning was performed with a Raytest radiochromatogram scanner,model RITA-90. Bio-Gel P-4 (25-45 f¿m)column chromatography (2 cm i.d.
x 1.25 m long) was performed as reported previously (18). Radioactivitywas determined with a Beckman liquid scintillation spectrometer, modelLS-6000LL.
RESULTS AND DISCUSSION
Lectin-Affinity Electrophoretography of Serum AFP from Pa
tients with Liver Diseases. Typical electrophoretic patterns of serumAFP in patients with HCC, LC, and CH and cord serum AFP separatedwith the lectin-affinity electrophoresis are shown in Fig. 1. The major
isoform of serum AFP in patients with HCC is denoted as ConA'LCA1 "r 'E4-PHAJ, using the abbreviated names of the three lectins
with the numbers of the corresponding bands as superscripts. On theother hand, the major isoform of serum AFP in patients with LC andCH and cord serum was Con A2LCA'E4-PHA2. Thus, Con A2LCA'or 3E4-PHA4 is an AFP isoform specific to HCC, which can be dis
criminated from the AFP isoforms of CH and LC. Because a smallamount of the Con A2LCA' "r 3E4-PHA4 AFP isoform was detected in
cord serum AFP, the whole sugar chain structure of AFP purified fromcord serum was elucidated.
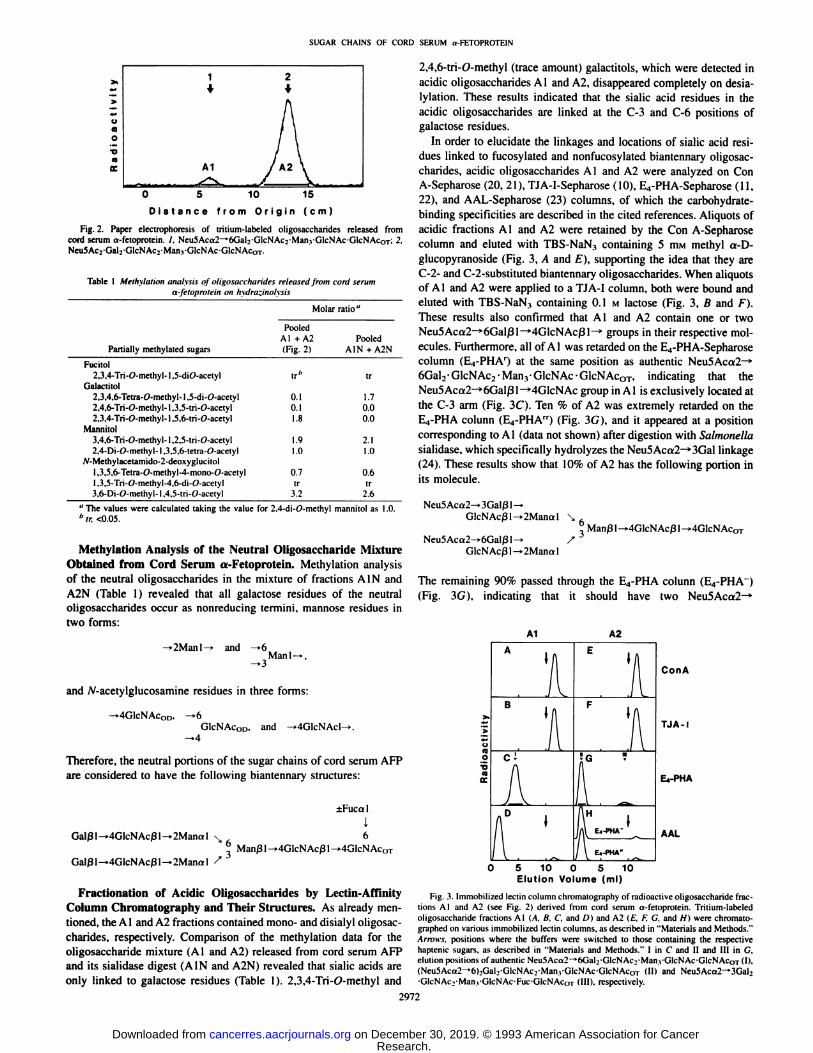
Paper Electrophoresis of the Radioactive Oligosaccharides Released from Cord Serum a-Fetoprotein. When the tritium-labeled
oligosaccharide fraction released from cord serum AFP was subjectedto paper electrophoresis, it was separated into two acidic fractions (Aland A2) as shown in Fig. 2. The % molar ratio of A1 and A2 wascalculated to be 1:9 on the basis of their radioactivities. Acidic oligosaccharides A1 and A2 were converted to neutral oligosaccharides(AIN and A2N, respectively) by exhaustive Arthmbacler sialidasedigestion, and the oligosaccharides in fractions Al and A2 were confirmed to be monosialyl and disialyl derivatives by determining thenumber of acidic fractions produced from each fraction on partialdesialylation (16) (data not shown). An aliquot of the radioactiveneutral oligosaccharide mixture (AIN and A2N) was digested, with amixture of diplococcal ß-galactosidase and ß-/V-acetylhexosamini-dase which specifically hydrolyzes the Gal/3 l—^GIcNAcßl—»2
group (8, 19), and then analyzed by Bio-Gel P-4 column chromatog
raphy (18). All of them were converted to two radioactive oligosaccharides with the same mobilities as authentic Man,-GlcNAc-Fuc•¿�GlcNAcoiand Man,-GlcNAc-GlcNAcOT through the release of2 mol of the Galßl >4GlcNAc group (data not shown). The structuresof these two oligosaccharides were confirmed to be Manal—»6(Manal^3)Manßl^4GlcNAcß|->4(Fucal^6)GlcNAc„T andManal^6(Manal~>3)Manßl »4GlcNAcßl^4GlcNAcOI by se
quential exoglycosidase digestion, as described in the preceding papers ( 14, 16). These results indicated that the oligosaccharides in bothfractions are biantennary ones with either fucosylated or nonfucosyl-
ated trimannosyl cores.
Con A
2 1l i
LCA
Hepatocellularcarcinoma
Fig. 1. Patterns of serum AFP separated by af- i :..arUnity electrophoresis with three lectins. LIVCl
Chronichepatitis
Cord Serum
2971
Research. on December 30, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from
SUGAR CHAINS OF CORD SERUM a-FETOPROTEIN
A1
0 5 10 15
Distance from Origin (cm)
Fig. 2. Paper electrophoresis of tritium-labeled oligosaccharides released fromcord serum o-fetoprolein. /. Neu5Aca2-t6Gal;-GlcNAc2-Man,-GlcNAc-GlcNAcOT; 2,NeuSAcvGalvGlcNAcrManyGlcNAc-GicNAcoT.
Table I Methylaliun analysis of oligosaccharides released from coni serina-fetopmiein on hydrazinolysis
Molarratio"Partially
methylatedsugarsFucitol2.3.4-Tri-O-methyl-
1,5-diO-acetylGalactitol2,3,4.6-Tetra-O-methyl-
1,5-di-O-acetyl2,4,6-Tri-O-methyl-1.3.5-tri-O-acetyl2.3.4-Tri-O-methyl-1.5.6-tri-O-acetylMannitol3,4,6-Tri-O-methyl-
1,2,5-tri-O-acetyl2.4-Di-O-methyl-1,3.5.6-tetra-O-acetylW-Methylacetamido-2-deoxyglucitoll,3,5.6-Tetra-O-methyl-4-mono-O-acetvll.3,5-Tri-O-methyl-4.6-di-O-acetyl3.6-Di-O-
methyl- 1.4.5-tri-O-acetylPooledAl
+A2(Fig.2)tr*0.10.11.81.91.00.7tr3.2PooledA1N+A2Ntr1.70.00.02.11.00.6tr2.6
value for 2,4-di-O-methyl mannitol as 1.0.
Methylation Analysis of the Neutral Oligosaccharide MixtureObtained from Cord Serum a-Fetoprotein. Methylation analysis
of the neutral oligosaccharides in the mixture of fractions A IN andA2N (Table I) revealed that all galactose residues of the neutraloligosaccharides occur as nonreducing termini, mannose residues intwo forms:
•¿�2Manl-»andManl-
»3
and /V-acetylglucosamine residues in three forms:
—¿�4GlcNAcoD. ->6
-»4and -»4GlcNAcl-
Therefore, the neutral portions of the sugar chains of cord serum AFPare considered to have the following biantennary structures:
GalßI^4GlcNAcßI-»2Manal
Galßl^4GlcNAcßl—¿�2Manal
±Fucal
I6
Manßl -»4GlcNAcßl ->4GlcN Ac()T
Fractionation of Acidic Oligosaccharides by Lectin-Affinity
Column Chromatography and Their Structures. As already mentioned, the A1 and A2 fractions contained mono- and disialyl oligosac
charides, respectively. Comparison of the methylation data for theoligosaccharide mixture (Al and A2) released from cord serum AFPand its sialidase digest (AIN and A2N) revealed that sialic acids areonly linked to galactose residues (Table 1). 2,3,4-Tri-O-methyl and
2,4,6-tri-O-methyl (trace amount) galactitols. which were detected in
acidic oligosaccharides Al and A2. disappeared completely on desia-lylation. These results indicated that the sialic acid residues in theacidic oligosaccharides are linked at the C-3 and C-6 positions of
galactose residues.In order to elucidate the linkages and locations of sialic acid resi
dues linked to fucosylated and nonfucosylated biantennary oligosaccharides, acidic oligosaccharides Al and A2 were analyzed on ConA-Sepharose (20, 21), TJA-I-Sepharose (10), E4-PHA-Sepharose (11,22), and AAL-Sepharose (23) columns, of which the carbohydrate-
binding specificities are described in the cited references. Aliquots ofacidic fractions Al and A2 were retained by the Con A-Sepharosecolumn and eluted with TBS-NaN, containing 5 HIMmethyl a-D-
glucopyranoside (Fig. 3, A and £),supporting the idea that they areC-2- and C-2-substituted biantennary oligosaccharides. When aliquotsof Al and A2 were applied to a TJA-I column, both were bound andeluted with TBS-NaN, containing 0.1 M lactose (Fig. 3. B and F).
These results also confirmed that Al and A2 contain one or twoNeu5Aca2—»6Galßl-»4GlcNAcßl—* groups in their respective mol
ecules. Furthermore, all of AI was retarded on the E4-PHA-Sepharosecolumn (E4-PHAr) at the same position as authentic Neu5Aca2^6Gal2-GlcNAc2-Man,-GlcNAc-GlcNAcOT, indicating that theNeu5Aca2^6Galß|->4GlcNAc group in Al is exclusively located at
the C-3 arm (Fig. 3C). Ten % of A2 was extremely retarded on theEj-PHA colunn (E4-PHArr) (Fig. 3G), and it appeared at a position
corresponding to Al (data not shown) after digestion with Salmonellasialidase, which specifically hydrolyzes the Neu5Aca2—»3Gallinkage
(24). These results show that 10% of A2 has the following portion inits molecule.
Manßl^4GlcNAcßl^4GlcNAcOTGlcNAcßl^2Manal \,
GlcNAcß!-»2Manal
The remaining 90% passed through the E4-PHA colunn (E4-PHA~)
(Fig. 3G). indicating that it should have two Neu5Aca2~ *
A1 A2
E.-PHA"
ConA
TJA-I
E«-PHA
AAL
0 5 10 0 5 10Elution Volume (ml)
Fig. 3. Immobilized lectin column chromatography of radioactive oligosaccharide fractions Al and A2 (see Fig. 2) derived from cord serum a-fetoprotein. Tritium-labeledoligosaccharide fractions Al (A. B, C, and £>)and A2 (E, F, G. and H} were chromato-graphed on various immobilized lectin columns, as described in "Materials and Methods."
Arrows, positions where the buffers were switched to those containing the respectivehaptenic sugars, as described in "Materials and Methods." I in C and II and III in G.elution positions of authentic Neu5Aca2 -"oGaN'GlcNALvManvGlcNAc-GlcNAcoT (I),(Neu5Aca2^6)2Ga]2-GlcNAcyManvGlcNAc:GlcNAcoT (II)' and Neu5Aca2^3Gal2
•¿�GlcNAcyManyGlcNAc-Fuc-GlcNAcoT (HD. respectively.
2972
Research. on December 30, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

SUGAR CHAINS OF CORD SERUM o-FETOPROTEIN
6Galßl—"4GlcNAcgroups in its molecule. These three fractions ob- can be concluded to comprise a monosialylated biantennary sugartained with E4-PHA-Sepharose columns (E4-PHA~, E4-PHAr, E4- chain with additional fucosylation of the proximal W-acetylglu-PHArr) were separated into an AAL passed-through fraction (AAL~) cosamine. Interestingly, the structure of the Con A+AAL+ "r ~E4-
and an AAL bound and eluted with 5 mM L-fucose fraction (AAL+) in PHAr fraction from cord serum AFP is equal to the major sugar chain
the ratio of 95:5, respectively, by AAL-Sepharose column chroma- structure from ascites fluid AFP from a patient with HCC reported bytography (Fig. 3, D and //). Yoshima et al. (25). Accordingly, the Con A2LCA' "r 'Ej-PHA4 AFP
From the results described thus far, the sugar chain structures re- isoform may become a useful tumor marker of HCC. These resultsleased from AFP were determined to be as shown in Table 2. Because may indicate that Galßl—*4GlcNAc-specific «2—>6sialyltransferase
human AFP has one asparagine AMinked oligosaccharide per mole- activity decreases by malignant transformation; however, there is nocule, sugar chain structures released from AFP should directly corre- enzymatic evidence that two different a2—>6sialyltransferases spe-spond to the sugar chain structures of AFP isoforms, which were cific to the Galßl—^GlcNAcßl—^Manal—»6group and theseparated by the three types of lectin-affmity electrophoresis, as Galßl-^4GlcNAcßl—>2Manal->3 group exist,
shown in parentheses in Table 2. Accordingly, the sugar chain struc- Fucosylation of a-Fetoprotein Produced in Fetal Liver. Theture of the Con A2LCA' or 3E4-PHA4 AFP isoform specific to HCC sugar chains of most plasma glycoproteins, including transferrin (26),
Table 2 Proposed structures of sugar chains released from cord serum a-fetoprotein and possible sugar chain structures of a-fetoprotein isoforms separatedon lectin affinity electrophoresis
Fraction structures" % molar ratio
Con A»AAL-E4-PHA-TJA-I*(Con A;LCA'E4-PHA:)
Neu5Aca2^6Galßl-^4GlcN Acß1—¿�2Mano 1,, Manßl^4GlcN AcßI-^4GlcNAc 732
Neu5Aca2->6Galßl-4GlcN AcßI-2Mane»I
Con A'AAL'E,-PHA-TJA-I' (Con A:LCA'E4-PHA:>
Fuco INeu5Aca2-6Galßl-4GlcNAcßl^2Manal i
fi 6°Manßl-4GlcNAcßl^4GlcNAc
Neu5Aca2^6Galßl->4GlcNAcßl->2Mancrl ^ 3-4
Con A*AAL-E4-PHA"TJA-I- (Con A:LCA'E4-PHA'(
Neu5Aca2—3Galßl—4GlcNAcß1—¿�2Mana I
3 ManßI-4G1CN AcßI-»4GlcNAc 9.0
Neu5Aca2-6Galßl-^4GlcNAcßl—¿�2Manalx*
Con A*AAL-Er-PHATJA-I* (Con A;LCA'E4-PHA')
Fucal
N 63 Manßl-^4GlcN AcßI-MGlcNAc 0.5
Neu5Aca2^6Galßl^4GlcNAcßl-*2Manal 7"
Con A'AAL-E4-PHATJA-I* (Con A;LCA'E4-PHA4)
Galßl-^4GlcNAcßl-.2Manal s
6
Neu5Aca2^6Galßl-«4GlcNAcß1-*2Mana I /
Con A*AAL'E,-PHATJA-I' (Con A'LCA'E4-PHA4)
FucalGalßl—»4GlcNAcßl—*2Manal |
\ft°Manßl^4GlcN Acßl-*4G1cNAc 0.4
NeuSAca2-.6Galßl-»4GlcNAcßl-.2Mann I /
' Oligosaccharide fractions obtained on lectin columns. Those in parentheses are AFP isoforms separated by lectin-affmity electrophoresis.
2973
Research. on December 30, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

SUIAR CHAINS OF CORD SERUM a-l;l:T()PR()TI:IN
fibrinogen (27), prothrombin (28), amithrombin III (29), ceruloplas-min (16), a,-antitrypsin (30, 31), and a,-acid glycoprotein (32, 33),
are not fucosylated, as follows:
{Neu5Aca2--.6(3)Galßl-*4GlcNAcßl-»},
f Mana I
Manul
>4GlcNAcßl->4GlcNAc
On the other hand, intracellular glycoproteins in parenchymal cells ofhuman liver are fucosylated as shown by the sugar chains of ribonu-cleases (34), saposins A, B. C. and D (35), and y-glutamyltranspep-
tidase (36), as follows:
{Neu5Aca2—6(3)Galßl—¿�4GlcNAcßl—),4
Man«I
Manul
±FUCQ1
4'6 6
-, Manßl-»4GlcNAcßl—4GlcNAc3
However, it has not yet been elucidated whether the fucosylation ofglycoproteins produced in normal human hepatotes is related to asecretion or nonsecretion trafficking mechanism.
When malignant transformation of hepatocytes occurs, the sugarchains of membrane-bound y-glutamyltranspeptidase became highly
branched, although the degree of fucosylation did not change (36).These results may indicate that the increase in fucosylation of AFP isrelated not to an increase in GlcNAc:al^>6 fucosyltransferase activity
but to the depolarization mechanism of malignant hepatocytes, because parenchymal cells of human liver are quite well-differentiated
epithelial cells, which are polarized into basolateral and apical sides,and the tumorigenesis of hepatocytes is a kind of dedifferemiation.
There is a general concept that fetal hepatocytes become morededifferentiated in inverse proportion to the gestation period, inweeks. Tissues obtained from fetal liver at 16, 20, and 23 weeks ofgestation were cultured for 2 days, and the AFPs secreted into therespective medium were subjected to LCA affinity electrophoresis (4).As summarized in Fig. 4, the c/c molar ratio of fucosylated AFP
increased in inverse proportion to the gestation period, in weeks.These results support our hypothesis that the fucosylation of AFP maybe related to dedifferentiation of human hepatocytes through tumorigenesis.
40
20
20
Weeks Gestation
40
Fig. 4. % molar ratio of fucosylated AFP in the culture media of fetal liver tissuesobtained at the time of legal abortion was calculated from the relative intensities of AFPbands obtained on LCA-afftnity electrophoresis.
Finally, the AFP isoform comprising a monosialyl-biantennarysugar chain with additional fucosylation of the proximal /V-acetylglu-cosamine was separated by using Con A-, LCA-, E4-PHA-affinityelectrophoresis as Con A2LCA'E4-PHA"* pattern, and it was found
that the AFP isoform specific to HCC is a useful tumor marker todifferentiate benign liver diseases, including LC and CH.
REFERENCES
i.
10.
16.
20.
21.
23.
24.
25.
Morinaga, T., Sakai, M.. Wegmann. T. G.. and Tamaoki, T. Primary structures ofhuman a-fetoprotein and its mRNA. Proc. Nail. Acad. Sci. USA, 80: 4604-1608.
1983.Taketa, K. a-Fetoprotein: réévaluationin hepatology. Hepatology, 12: 1420-1432.
1990.Breborowicz. J.. Mackiewicz. A., and Breborowicz, D. Microheterogeneity of a-fe-toprotein in patient serum as demonstrated by lectin affino-electrophoresis. Scand. J.Immunol., 14: 15-20. 1981.Miyazaki. J.. Endo, Y.. and Oda. T. Lectin affinities of u-fetoprotein in liver cirrhosis,hepatocellular carcinoma and metastatic liver tumor. Acta Hepatol. Jpn.. 22: 1559-1568. 1981.Taketa. K., Sekiya, C.. Namiki. M.. Akamat.su. K.. Ohta. Y.. Endo, Y., and Kosaka. K.Lectin-reactive profiles of u-fetoprotein characterizing hepatocellular carcinoma andrelated conditions. Gastroenterology. W: 508-518. 1990.
Endo. Y. Taga. H.. and Mirai. H. Analysis of patients wilh high risk of hepatocellularcarcinoma by lectin binding technique of AFP. Physico-Chemicul Biology, .f.V 205-208. 1991.Nishi, S.. and Mirai. H. Purification of human, dog and rabbit «-fetoprotein byimmunoadsorbents of Sepharose coupled wilh anti-human «-fetoprotein. Biochim.Biophys. Acta. 278: 293-298. 1972.Glasgow. L. R., Paulson, J. C., and Hill, R. L. Systematic purification of fiveglycosidases from Slreptucoccus (Dipltxwcus) [meumtmiat'. J. Biol. Chem., 252;8615-8623. 1977.Kochibe, N., and Furukawa. K. Purification and properties of a novel fucose-specifichemagglutinin of Ali'urUi (turiinliu. Biochemistry. 19: 2841-2846, 1980.
Yamashita, K., Umetsu, K.. Suzuki. T., and Ohkura. T. Purification and characterization of a Neu5Aca2 -6GalßI MGlcNAc and HSO, '6Galßl MGlcNAc specificlectin in tuberous roots of Triclwsanlhes jnjiÃniÃi'ü.Biochemistry, M : \ 1647-11650,
1992.Kobata. A., and Yamashita. K. Extenlion and limit of affinity chromatography ofoligosaccharides on an E-PHA-aizarose column. Methods Enzymol., /79: 46-54,1989.Yamashita. K., Koide, N., Endo, T., Iwaki, Y. and Kobata, A. Altered glycosylationof serum transferrin of patients wilh hepatocellular carcinoma. J. Biol. Chem.. 264:2415-2423, 1989.Fukushima. K.. Waianabe. H.. Takeo, K.. Nomura. M.. Asahi, T., and Yamashila. K.¿V-linkedsugar chain structures of recombinant human lymphotoxin produced byCHO cells: the functional role of carbohydrates on the lectin-like character andclearance velocity. Arch. Biochem. Biophys., in press.Hitoi, A., Yamashita, K., Niwala, Y. trie, M., Kochibe, N., and Kobata. A. Thecarbohydrate moieties of human urinary ribonuclease U. J. Biochem. (Tokyo), KKÌ:29-11. 1987.Takasaki, S.. Mizuochi. T.. and Kobata. A. Hydrazinolysis of asparaginc-linked sugarchains to produce free oligosaccharides. Methtxls Enzymol.. <S'.?;263-268. 1982.
Yama.shita. K.. Liang. C-J.. Funakoshi, S., and Kobata, A. Structural studies ofasparagine-linked sugar chains of human ceruloplasmin: structural characteristics of
the triantennary complex type sugar chains of human plasma glycoproteins. J. Biol.Chem.. 256: 1283-1289. 1981.
Yamashita. K., Hitoi, A.. Matsuda, Y. Tsuji. A., Katunuma, N., and Kobata, A.Structural studies of carbohydrate moiety of rat kidney -y-glutamyltranspeptidase: anextremely heterogeneous pattern enriched with nonreducing terminal -V-acetylglu-cosamine residues. J. Biol. Chem.. 25«:1098-1107, 1983.
Yamashita. K.. Mizuochi. T.. and Kobata. A. Analysis of oligosaccharides by gelfiltration. Methods Enzymol.. «J: 105-126. 1982.
Yamashita. K.. Ohkura. T., Yoshima. H.. and Kobata. A. Substrate specificities ofdiplococcal ß-iV-acetylhexosaminidase. a useful enzyme for the structural studies ofcomplex type asparagine-linked sugar chains. Biochem. Biophys. Res. Commun..100: 226-232. 1981.
Baenziger, J. U., and Fiele. D. Structural determinants of concanavalin A specificityfor oligosaccharides. J. Biol. Chem., 254: 24(X)-2407. 1979.
Ogata. S.. Muramatsu. T.. and Kobata. A. Fractionation of glycopeptides by affinitycolumn chromatography on concanavalin A-Sepharose. J. Biochem. (Tokyo), 7«:687-696. 1975.Yamashita. K., Hitoi, A., and Kobata, A. Structural determinants of studies of Pluueu-lu.'i w</£«n.verythroagglutinating lectin for oligosaccharides. J. Biol. Chem.. 25«:14753-14755. |983.Yamashita, K., Kochibe. N.. Ohkura. T. Ueda. I., and Kobata, A. Fractionation ofi.-fucose-containing oligosaccharides on immobilized Alcuna aiirwitia lectin. J. Biol.Chem.. 260: 4688-1693. 1985.Hoyer, L. L.. Roggentin, P.. Schauer. R., and Vimr, E. R. Purification and propertiesof cloned Salnumella lyphimurium LT2 sialidase with virus-typical kinetic preferencefor sialyl a2 »3linkages. J. Biochem. (Tokyo), 110: 462-167. 1991.Yoshima, H.. Mizuwhi, T.. Ishii, M.. and Kobata. A. Structure of the asparagine-linked sugar chains of a-fetoprotein purified from human ascites Huid. Cancer Res.,40: 4276-1281. 1980.
2974
Research. on December 30, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

Sl'CrAR CHAINS OF CORD Si-Rl'M «-H-.I OI'KO] I IN
26. Spik, G., Bayard. B.. Fournet. B.. Strecker. G.. Bouquelel. S., and Montreuil, J.Studies on glycoconjugates. LXIV. Complete structure of two carbohydrate units ofhuman serotransferrin. FEBS Lett., JO: 296-299. 1975.
27. Mizuochi, T.. Taniguchi. T., Asami, Y., Takamatsu. J.. Okude. M.. Iwanaga. S.. andKohata, A. Comparative studies on the structures of the carbohydrate moieties ofhuman fibrinogen and abnormal librinogen Nagoya. J. Biochem. (Tokyo). 92: 283-293. 1982.
28. Mizuochi. T., Fujii. J.. Kisiel. W., and Kobata. A. Studies on the structures of thecarbohydrate moiety of human prothrombin. J. Biochem. (Tokyo). 90: 1023-103!.1981.
29. Mizuochi. T.. Fujii, J., Kurachi, K., and Kobata. A. Structural studies of the carbohydrate moiety of human amithrombin III. Arch. Biochem. Biophys., 203: 458-465.
1980.30. Hodges, L. C.. Laine. R., and Chan. S. K. Structure of the oligosaccharide chains in
human a,-protease inhibitor. J. Biol. Chem.. 254: 8208-8212. 1979.31. Mega. T.. Lujan, E., and Yoshida. A. Studies on the oligosaccharide chains of human
a I-protease inhibitor. J. Biol. Chem.. 255: 4057-4061, 1980.32. Fournet. B., Monlreuil. J., Strecker. G., Dorland. L.. Haverkamp. J.. Vliegenthart. F.
G.. Binette. J. P.. and Schmid. K. Determination of the primary structures of 16asialo-carbohydrate units derived from human plasma a I-acid glycoprotcin by 360-MHZ 'H NMR. Biochemistry. 17: 5206-5214. 1978.
33. Yoshima. H.. Matsumoto. A.. Mizuochi. T.. Kawasaki. T.. and Kobata. A. Comparative study of the carbohydrate moieties of rat and human plasma a i-acid clycopro-teins. J. Biol. Chem.. 25o: 8476-8484. 1981.
34. Yamashita, K.. Hitoi. A.. Irie. M.. and Kobata. A. Fractionation by lectin affinitychromatography indicates that the glycosylation of most ribonucleases in humanviscera and body fluids is organ specific. Arch. Biochem. Biophys.. 250: 263-266,1986.
35. Yamashita. K.. Inui. K.. Totani. K.. Kochibe. N.. Furukawa. M., and Okada. S.Characteristics of asparagine-linked sugar chains of sphingolipid activator protein I
purified from normal human liver and GM1 gangliosidosis. type 1 liver. Biochemistry.29: 3030-3039. 1990.
36. Yamashita. K., Totani. K.. Iwaki. Y. Takamizawa. !.. Tateichi, N.. Higashi, T.. Sakamoto. Y., and Kobata. A. Comparative study of sugar chains of -y-glutamyltranspep-
tidases purified from human hepatocellular carcinoma and from human liver. J.Biochem. (Tokyo). 105: 728-735. 1989.
2975
Research. on December 30, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

1993;53:2970-2975. Cancer Res Katsuko Yamashita, Kazuhisa Taketa, Shinzo Nishi, et al. Produced in Human Liver and Hepatocellular Carcinomas
-linked Sugar Chains of GlycoproteinsNCharacteristics of -Fetoprotein:αSugar Chains of Human Cord Serum
Updated version
http://cancerres.aacrjournals.org/content/53/13/2970
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/53/13/2970To request permission to re-use all or part of this article, use this link
Research. on December 30, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from





























