Embed Size (px)
Citation preview

TEMA 14. Oxidación de ácidos grasos. Obtención de la energía de los lípidos. Digestión, movilización y transporte extracelular de los triacilgliceroles del adipocito Transporte de los ácidos grasos al adipocito. Transporte de los ácidos grasos al interior de la mitocondria. β-oxidación de los ácidos grasos. Regulación de la degradación de ácidos grasos y triacilgliceroles. Metabolismo de los cuerpos cetónicos.
José C Rodríguez ReyDepartamento de Biología Molecular
2009-2010
GlucosaTAG
Glucosa 6P Ribosa 5 PGlucógenoAcidos grasos
Piruvato Acetil CoA
Lactato
OAA
C KREBSOAA

Los lípidos son la principal forma de almacenamiento de energía en el organismo
Combustible almacenado Tejido Gramos Kilocalorías*
el organismo
Combustible almacenado Tejido Gramos Kilocalorías*
Glucógeno Hígado 70 280Glucógeno Músculo 120 480Glucógeno Músculo 120 480Glucosa Fluidos
Corporales 20 80
Grasa Adiposo 15.000 135.000Proteína Muscular 6.000 24.000
Datos para un sujeto normal de 70 kilos de peso. Calculado a partir de 4 kcal/g para glúcido y proteína y 9 kcal/g para grasa. Tomado de Stryer. Bioquímica. Ed Reverté
* 1 Kcal= 4,18 KJulios,
Algunos datos importantes
1 L lí d l l d E ( l l d 1 00 l/dí d 1. Los lípidos son la principal reserva de E (calculando 1700 Kcal/día pueden proporcionar energía para unos 80 días)
2. Tienen un rendimiento energético mayor (9 vs 4 Kcal/g)3 No llevan agua de hidratación: constituyen una forma ligera de 3. No llevan agua de hidratación: constituyen una forma ligera de
almacenamiento de energía.4. Son el combustible favorito de muchos tejidos (músculo por ejemplo)
PEROPERO:
1. No pueden utilizarse de forma anaerobia. No pueden ser utilizados por las células que no poseen mitocondriascélulas que no poseen mitocondrias.
2. Algunos (ácidos grasos de cadena larga) no atraviesan la barrera hemato-encefálica: Es necesario buscar soluciones alternativas.
3. Por su insolubilidad deben ser transportados en forma de partículas complejas.

Los lípidos son moléculas insolubles y se transportan en forma de li t í s
l í
lipoproteínas
Apolipoproteínas
Fosfolípidos
TAGs y ésteres de Colesterol
ycolesterol
De Nelson et al. Principles of Biochemistry. 4th Ed. Freeman
PRINCIPALES RUTAS DEL METABOLISMO LIPIDICO
TAGTAGCuerpos cetónicos
FOSFOLIPIDOS
Acidos grasos Acetil CoA Colesteroly derivados
Derivados de ácidos grasos
CO2Ciclo de Krebs
g(prostaglandinas, leucotrienos)

Origen de los ácidos grasos para la oxidación
TAG Glicerol + AG
AG-AlbúminaT adiposoLPL
AG- FABPLPL
Lipoproteínas ricas en TAG
Intestino MUSCULO *
ricas en TAG
Hígado
* El hígado es otro tejido importante en la utilización de los ácidos grasos para producir energía
FASES DE LA OXIDACION DE TRIACIL GLICEROLES
Glicerol + Acido grasoTAG1
Acido graso
2
3
Acido graso
Tejido adiposo
Acil graso CoA1. Hidrólisis de TAGs2. Transporte
Tejido que utiliza la E de los ácidos
Acil raso CoA
4p
3. Activación de A Grasos4. Paso a la mitocondria5. β- oxidación
grasos
Acil graso CoA
5
Acetil CoA
CO2

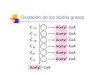
La degradación de TAGs produce ácidos grasos y glicerol
T GTAG
LSH
DAGAG
LSH
MAGAG
LSH
MAG lipasa
AG + Glicerol
El Glicerol obtenido por degradación de los TAGs del T adiposo se transporta al hígado en donde puede reciclarse
Glicerol
Glicerol quinasa (no en el tejido adiposo)
L- Glicerol 3- fosfato
SINTESIS DE TAGs
Glicerol 3 fosfato DH
GLUCOLISIS
DHAP
Triosa –fosfato GLUCOLISISGLUCONEOGENESIS
D-Gliceraldehido
Triosa fosfato isomerasa
D Gliceraldehido3-fosfato

LA HIDRÓLISIS DE LOS TRIGLICERIDOS DEL TEJIDO ADIPOSO ESTA REGULADA POR HORMONAS
GlucagónAdrenalina
ATP AMPc LSH
LSH
PKA activada
ACIDOS GRASOS unidos a albúmina
Origen de los ácidos grasos para la oxidación
TAG Glicerol + AG
AG-AlbúminaT adiposoLPL
AG- FABPLPL
Lipoproteínas ricas en TAG
Intestino Tejidos que utilizan
ácidos grasosricas en TAG
Hígado

ACTIVACION DE ACIDOS GRASOS
ATPAdenosina
Acido graso unido a FABP
Acil- graso CoA sintasa
Adenosina
Intermedio Acil-adenilato unido al Enzima
Pirofosfato
Acil- graso CoA sintasaPirofosfatasa
adenilato unido al Enzima
Acil graso –CoA
De Nelson et al. Principles of Biochemistry. 4th Ed. Freeman
La hidrólisis de Pirofosfato proporciona la energía suficiente para la activación de ácidos grasos
∆G0’ = -32,3 + 31,5 - 33,6 = - 34,4 KJ/mol
Enlace fosfato del ATP Formación
d l l
Hidrólisis de pirofosfato
del enlace acil- CoA

La lanzadera de Carnitina transporta los ácidos grasos al interior de la mitocondria
O- Acilcarnitina
L-CarnitinaAcil Carnitina
L-Carnitina
l
Espacio intermembranoso Translocasaintermembranoso
CAT I
CAT IIMatriz mitocondrial
CAT II
O- Acil-carnitinaA il C nitin
CAT I – Carnitina acil transferasa ICATII - Carnitina acil transferasa II
L-CarnitinaL-CarnitinaAcil Carnitina
Adaptado de Garret & GrishamBiochemistry (2nd Ed.) Saunders.
Ruta de β-oxidación de los ácidos grasos
Palmitil CoAAcil CoA DH
VLCAD 12-18 CMCAD 4-14 CSCAD 4 8 C
Trans ∆2
SCAD 4-8 C
Trans ∆Enoil CoAEnoil –CoA
hidratasa
L- β- hidroxi-Acil-CoAβ- hidroxi-Enzima Trifuncional para cadenas
β- Ceto-acil-CoA
Acil-CoA DHp
de más de 12 atomos de carbono
βAcil- CoAAcetiltransferasa(Tiolasa)
Acetil CoA

Los electrones liberados por oxidación de los ácidos grasos entran en la cadena de transporte electrónico vía CoQ
Espaciointermembranoso Glicerol 3P DH
Glicerol 3 P
Succinato
ETF- Q oxidorreductasa
Matriz
De Nelson et al Principles of
ACIL GRASO CoA DH
De Nelson et al. Principles of Biochemistry. 4th Ed. Freeman
Acil graso CoA
Rendimiento energético de la oxidación de una molécula de Palmitil CoA
Enzima NADH /FADH2 formados ATPs
Acil CoA DH 7 FADH2 10,5 Β id ióAcil CoA DH 7 FADH2 10,5 β- hidroxi-Acil-CoA DH 7 NADH 17,5
I it t DH 8 NADH 20
Β-oxidación
Isocitrato DH 8 NADH 20Α- Cetoglutarato DH 8 NADH 20Succinil-CoA Sintasa 8 (GTP)Succinato DH 8 FADH2 12
C KrebsSuccinato DH 8 FADH2 12 Malato DH 8 NADH 20
TOTAL 108 ª
ª En el caso de partir del ácido palmítico hay que considerar que su activación necesita dos enlaces fosfato

Rutas especiales de degradación de ácidos grasos
1. Acidos grasos de cadena muy larga2. Acidos grasos insaturados3. Acidos grasos ramificados3. Ac dos grasos ram f cados4. Acidos grasos de número impar de átomos de carbono
OXIDACION PEROXISOMAL (5- 10% del total)
1 3Acil CoA(cadena l )
Acil CoA Enoil CoA AcilCoAAcetilCoA
AcilCarnitinaAcetil CarnitinaAcetilCoA
1 2 3
FAD FADH2 NADHlarga)
O2 H2O2
2 NADH
e-
1. Transportador independiente de carnitina2 Oxidasa2. Oxidasa3. Transportador dependiente de carnitina

Oxidación de un ácido graso monoinsaturado
Tres ciclos de β-
d ó3 Acetil- CoA
Oleil- CoA (18:1 ∆9)
oxidación
Δ3Δ2- Enoil- CoA isomerasa
Cis ∆3-Dodecenil- CoAUna isomerasa transforma un enlace cis ∆3 en un trans ∆2Una isomerasa transforma un enlace cis ∆3 en un trans ∆2
Cinco ciclos de β-oxidación
Trans-∆2-Dodecenil-CoA
6 Acetil CoA De Nelson et al. Principles of Biochemistry. 4th Ed. Freeman
Oxidación de ácidos grasos poliinsaturados
Linoleil CoA (cis-∆9 cis-∆12)
3 ciclos de β oxidación
Cis ∆3, cis ∆6
3 Acetil CoA
transforma un enlace cis ∆3 en un trans ∆ 2Enoil CoA isomerasa
Cis ∆ , cis ∆
Trans ∆2 cis ∆61 ciclo completo y1ª reacción del siguiente
Trans ∆2, cis ∆6
Acetil CoA
La hidrogenación de carbonos 2 y 5 convierte dos enlaces conjugados (trans ∆2 cis ∆4) en un doble enlace trans ∆3
2,4 dienoil-CoA reductasa
Trans ∆2, cis ∆4
(trans ∆ cis ∆4) en un doble enlace trans ∆3
Enoil CoA Trans ∆3
Cambio del doble enlace trans ∆3 en un trans ∆ 2isomerasa
4 nuevos ciclos Trans ∆2
De Nelson et al. Principles of Biochemistry. 4th Ed. Freeman
4 nuevos ciclos de β oxidación
5 Acetil CoA

Etapa final de la oxidación de los ácidos grasos de número impar de átomos de carbono
P i il C APropionil CoA
Propionil- CoA Propionil- CoA Carboxilasa (Biotina)
D-metil-malonil CoA
Metil-malonil CoA epimerasa Metil-malonil CoA
mutasa (Coenzima B12)
De Nelson et al. Principles of Biochemistry. 4th Ed. Freeman
L-metil-malonil CoA Succinil CoA
Estructura del Coenzima B12 (desoxiadenosil cobalamina)
desoxiadenosina
Anillo de corrina
Dimetil benzimidazol
De Nelson et al. Principles of Biochemistry. 4th Ed. Freeman

Mecanismo de reacción de la Metil-malonil mutasa
1. Formación del radical desoxi-adenosilo a partir del Coenzima B12 (el cobalto pasa de estado de oxidación +3 a +2)
2. El radical adenosilo extrae un átomo de H del sustrato
3. El carbono reactivo del nuevo radical arranca el grupo (X) del carbono adyacente originando un nuevo radical librelibre.
4. El nuevo carbono reactivo extrae un atomo de hidrógeno de la desoxiadenosina convirtiéndola de nuevo en un radical desoxiadenosilonuevo en un radical desoxiadenosilo.
5. El radical desoxiadenosilo regenera el Coenzima B12 formando de nuevo un enalcae de coordinación con el cobalto del anillo de Corrina (el cobalto pasa de estado de oxidación +2 a + 3)
De Nelson et al. Principles of Biochemistry. 4th Ed. Freeman
Beta oxidación de ácidos grasos saturados
RESUMEN DE LOS PROCESOS DE OXIDACION DE ACIDOS GRASOS
g
1. Mitocondrial a. Intervienen DH específicas dependiendo del tamaño : VLCAD, MCAD y SCAD. Existe una LCAD
pero se cree que está involucrada principalmente en la degradación de ácidos grasos de cadena p p pramificada
b. En los ácidos grasos de cadena larga las tres actividades enzimáticas de la β oxidación (enoil CoA hidratasa, 3 hidroxi acil CoA DH y Tiolasa) están en una proteína trifuncional (TFP) ligada a membrana. Para los de cadena media estos enzimas se encuentran solubles en la matriz
2. Peroxisomala. Se digieren los ácidos grasos de más de 18 átomos de carbonob. La DH cede los electrones al oxígeno para la producción de peróxido de hidrógeno.c. La Enoil hidratasa y la β- hidroxiacil CoA DH forman parte de un enzima bi- funcional
Beta oxidación de ácidos grasos insaturados
1. Monoinsaturados. Requiere un enzima especial: ∆3∆2- Enoil- CoA isomerasa.2. Polinsaturados: Requiere además otro enzima: 2-4- dienoilCoA reductasa (PEROXISOMAL)
Alfa oxidación
1. En el caso de los ácidos grasos ramificados tiene lugar en los peroxisomas. Pero también puede producirse en mitocondria y en retículo endoplasmático
óOmega oxidación
Se lleva a cabo en el retículo endoplasmático (isoenzima P450)

Regulación de la degradación de los ácidos grasos
TAG+Glucagón
Adrenalina LSH
Acidos grasos
Adrenalina LSH
Acil graso CoA-CPT I
Malonil CoAAcil graso carnitina
CPT I
INSULINA+
-Acil graso CoA
CPT II
NADHβ-OH acil DH
Acetil CoA-
Tiolasa
Acetil CoA
La disminución de los niveles de glucosa, la preparación para el ejercicio o las situaciones de estrés provocan la degradación de TAGs y ácidos grasos
TAGAdrenalinaGlucagónCortisol TAG
Acidos grasos
Cortisol
Acetil CoAAcetil CoA
GLUCOSA PIRUVATO
NADHCi l d K bs
OAA
NADHFADH2
Ciclo de Krebs

En condiciones de ayuno prolongado el acetil CoA se deriva hacia la producción de cuerpos cetónicos
TAGGlucagón TAG
Acidos grasos
Acetil CoA Cuerpos cetónicosAcetil CoA
GLUCOSA PIRUVATO
Cuerpos cetón cos
Ci l d K bs
OAA
NADHCiclo de Krebs NADHFADH2
Estructura de los cuerpos cetónicos
Acetona
Acetoacetato
D-β- Hidroxibutirato

Síntesis hepática de los cuerpos cetónicos
2 Acetil CoA
TiolasaTiolasa
Acetoacetil CoA
HMG CoA sintasa
β hidroxi β metil-l t il C Aglutaril CoA
De Nelson et al. Principles of Biochemistry. 4th Ed. Freeman
Síntesis hepática de los cuerpos cetónicos (cont.)
β hidroxi β metil- glutaril CoA
HMG CoA LiasaAcetil CoA
Acetil acetato
Acetoacetato descarboxilasa D- β- hidroxibutirato
deshidrogenasadeshidrogenasa
De Nelson et al. Principles of Biochemistry. 4th Ed. Freeman
AcetonaD- β- hidroxibutirato

Los cuerpos cetónicos se utilizan en tejidos extrahepáticos
D- β- hidroxibutirato
D- β- hidroxibutirato deshidrogenasa
Músculo cardíaco, léti b Β-cetoacilCoA
Acetil acetato
Succinil CoAesquelético y cerebroNO EN HIGADO
Β cetoacilCoA transferasa
Acetoacetil CoA
Succ n l CoASuccinato
Acetoacetil CoA
Tiolasa
2 A til C A2 Acetil CoA
Niveles de sustratos y hormonas en un individuo sometido a inanición
Después de una A las 12 h A los 3 días A las 5 semanascomida
Insulina (U/ml) 40 15 8 6Insulina (U/ml) 40 15 8 6Glucagón (pg/ml) 80 100 150 120Relación Ins/Glucagón 0,5 0,15 0,05 0,05Glucosa (mM) 6,1 4,8 3,8 3,6( )Ac grasos (mM) 0,14 0,6 1,2 1,4Acetoacetato (mM) 0,04 0,05 0,4 1,3 hidroxibutirato (mM) 0,03 0,10 1,4 6,0