Embed Size (px)
Citation preview

Tạp chíKH-CN Nghệ An
SỐ 9/2016 [16]
NGHIÊN CỨU - TRAO ĐỔI
Ở sinh vật nhân thực, trong quá trình phânbào nguyên nhiễm, gọi tắt là nguyên phân,nhiễm sắc thể (NST) của tế bào mẹ được phânchia chính xác cho các tế bào con. Kết quả làtừ một tế bào mẹ tạo ra hai tế bào con có bộNST giống hệt nhau và giống hệt tế bào mẹ.Đặc điểm của quá trình nguyên phân đãđược làm rõ ở mức độ tế bào từ những nămcuối thế kỷ 20 và cũng có nhiều nghiên cứuphần nào đề cập đến quá trình phân chia ởmức độ phân tử. Cho đến nay, có nhiềunghiên cứu ở mức độ phân tử đã làm sáng tỏhơn một số chi tiết của quá trình nguyênphân. Trong khuôn khổ bài viết này, nhữngvấn đề liên quan đến thoi phân bào và sự dichuyển của NST trong quá trình nguyên phânsẽ được đề cập ở mức độ phân tử.
THOI PHÂN BÀO VÀ SỰ DI CHUYỂN CỦA NHIỄM SẮC THỂ TRONG QUÁ TRÌNH NGUYÊN PHÂN Ở TẾ BÀO SINH VẬT NHÂN THỰC DƯỚI GÓC ĐỘ PHÂN TỬ
n Nguyễn Thị ThảoKhoa Sinh học, Trường Đại học Vinh
1. Thoi phân bào và sự di chuyển của NSTỞ sinh vật nhân thực đơn bào hay đa bào thì sự sinh
sản của tế bào là một quá trình sinh trưởng, phân chia vàphân bào mang tính chu kỳ gọi là chu trình tế bào. Trongđó, pha M là giai đoạn nguyên phân giúp một tế bào banđầu phân chia thành hai tế bào con giống hệt nhau vềmặt di truyền. Một trong những đặc điểm quan trọnggiúp tế bào hoàn thành quá trình nguyên phân một cáchbình thường chính là sự di chuyển của NST về hai cựccủa tế bào nhờ thoi phân bào lưỡng cực. Thông thường,ở tế bào động vật, thoi phân bào được hình thành gầnnhân, từ các vi ống, trung thể (trung tâm tổ chức thoiphân bào) và thể sao cùng với các protein motor[4]. Ởtế bào thực vật, thoi phân bào cũng tương tự thoi phânbào ở tế bào động vật nhưng không có trung thể và viống thể sao. Nhưng thay vào đó, những loài thực vật bậc
cao lại có các dải preprophase ở hai đầu mỗitế bào đóng vai trò như trung thể[12]. Ở nấm,thoi phân bào cũng tương tự ở tế bào thực vật,nhưng trung tâm tổ chức thoi phân bào là thểcực. Đặc biệt, ở một số loài nấm, thoi phânbào hình thành trong nhân diễn ra quá trìnhnguyên phân kín (ví dụ, Saccharomyces ce-revisiae và Schizzosaccharomyces pombe)hoặc nửa mở (ví dụ, Aspergillus nidulans vàSchizosaccharomyces japonicus) khác vớinguyên phân mở ở tế bào động vật và tế bàothực vật (Hình 1)[10].
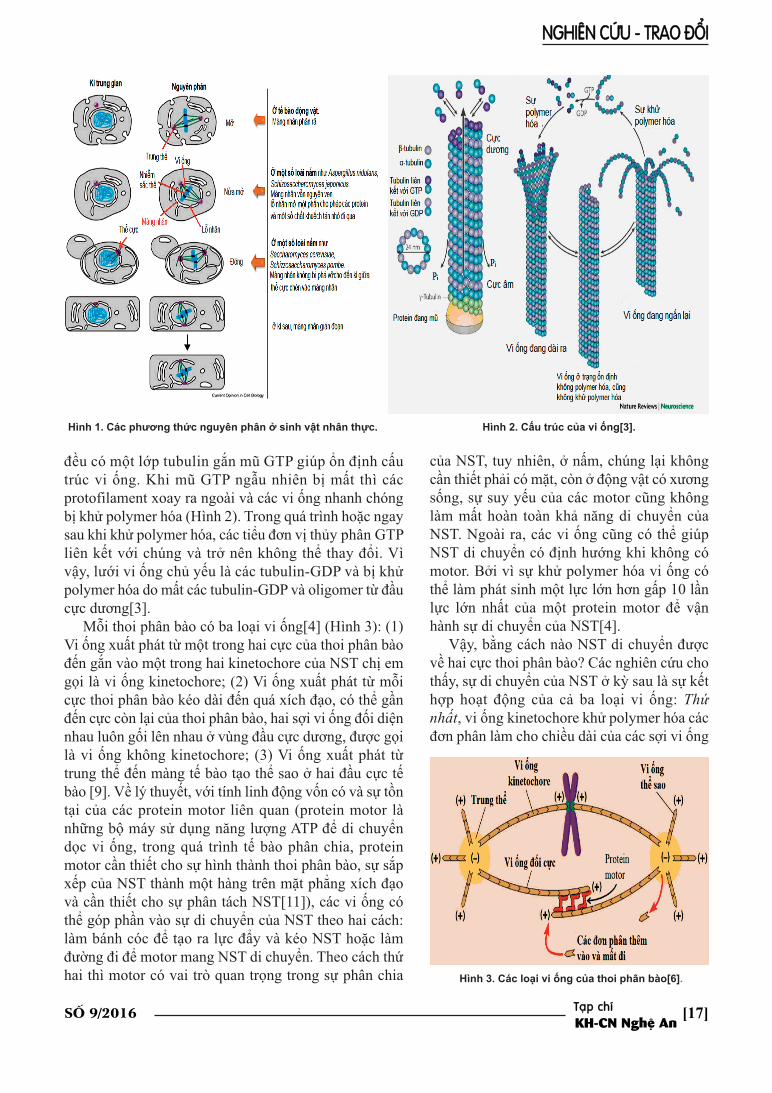
Vi ống là polymer protein có ở tất cả các tếbào sinh vật nhân thực, có đường kính khoảng24mm, bao gồm hàng nghìn đơn phân (dimertubulin gồm α-tubulin và β-tubulin) sắp xếp lạivới nhau tạo thành vách dạng lưới có từ 10-15 protofilament (ở động vật có vú là 13)[3](Hình 2). Bởi vì tính định hướng của dimertubulin, hai đầu của vi ống có phần khácnhau. Đầu có thể thêm vào hoặc loại bỏ dimertubulin với tốc độ nhanh hơn nhiều so với đầucòn lại (đầu cực âm) gọi là đầu cực dương[3].Trong quá trình vi ống lắp ráp thành thoi phânbào, cực âm ở tại cực thoi phân bào, còn cựcdương ở xa so với cực của thoi phân bào (Hình3)[6]. Một đặc điểm nổi bật giúp vi ống luônthực hiện được vai trò quan trọng của mình làtính linh động. Tính linh động của vi ống thểhiện ở khả năng thêm vào các đơn phân làmvi ống dài ra (gọi là polymer hóa) hoặc loại bỏcác đơn phân làm vi ống ngắn đi (gọi là khửpolymer hóa) từ hai đầu của vi ống. Vi ống cóthể chuyển từ trạng thái polymer hóa sangtrạng thái khử polymer hóa hoặc ngược lại.Thông thường, đầu cực dương của mỗi vi ống
Tạp chíKH-CN Nghệ An
SỐ 9/2016 [17]
NGHIÊN CỨU - TRAO ĐỔI
của NST, tuy nhiên, ở nấm, chúng lại khôngcần thiết phải có mặt, còn ở động vật có xươngsống, sự suy yếu của các motor cũng khônglàm mất hoàn toàn khả năng di chuyển củaNST. Ngoài ra, các vi ống cũng có thể giúpNST di chuyển có định hướng khi không cómotor. Bởi vì sự khử polymer hóa vi ống cóthể làm phát sinh một lực lớn hơn gấp 10 lầnlực lớn nhất của một protein motor để vậnhành sự di chuyển của NST[4].
Vậy, bằng cách nào NST di chuyển đượcvề hai cực thoi phân bào? Các nghiên cứu chothấy, sự di chuyển của NST ở kỳ sau là sự kếthợp hoạt động của cả ba loại vi ống: Thứnhất, vi ống kinetochore khử polymer hóa cácđơn phân làm cho chiều dài của các sợi vi ống
đều có một lớp tubulin gắn mũ GTP giúp ổn định cấutrúc vi ống. Khi mũ GTP ngẫu nhiên bị mất thì cácprotofilament xoay ra ngoài và các vi ống nhanh chóngbị khử polymer hóa (Hình 2). Trong quá trình hoặc ngaysau khi khử polymer hóa, các tiểu đơn vị thủy phân GTPliên kết với chúng và trở nên không thể thay đổi. Vìvậy, lưới vi ống chủ yếu là các tubulin-GDP và bị khửpolymer hóa do mất các tubulin-GDP và oligomer từ đầucực dương[3].
Mỗi thoi phân bào có ba loại vi ống[4] (Hình 3): (1)Vi ống xuất phát từ một trong hai cực của thoi phân bàođến gắn vào một trong hai kinetochore của NST chị emgọi là vi ống kinetochore; (2) Vi ống xuất phát từ mỗicực thoi phân bào kéo dài đến quá xích đạo, có thể gầnđến cực còn lại của thoi phân bào, hai sợi vi ống đối diệnnhau luôn gối lên nhau ở vùng đầu cực dương, được gọilà vi ống không kinetochore; (3) Vi ống xuất phát từtrung thể đến màng tế bào tạo thể sao ở hai đầu cực tếbào [9]. Về lý thuyết, với tính linh động vốn có và sự tồntại của các protein motor liên quan (protein motor lànhững bộ máy sử dụng năng lượng ATP để di chuyểndọc vi ống, trong quá trình tế bào phân chia, proteinmotor cần thiết cho sự hình thành thoi phân bào, sự sắpxếp của NST thành một hàng trên mặt phẳng xích đạovà cần thiết cho sự phân tách NST[11]), các vi ống cóthể góp phần vào sự di chuyển của NST theo hai cách:làm bánh cóc để tạo ra lực đẩy và kéo NST hoặc làmđường đi để motor mang NST di chuyển. Theo cách thứhai thì motor có vai trò quan trọng trong sự phân chia
Hình 1. Các phương thức nguyên phân ở sinh vật nhân thực. Hình 2. Cấu trúc của vi ống[3].
Hình 3. Các loại vi ống của thoi phân bào[6].

Tạp chíKH-CN Nghệ An
SỐ 9/2016 [18]
NGHIÊN CỨU - TRAO ĐỔI
mút hai vi ống kinetochore[7]. Số lượng vi ống gắnvới kinetochore ở các loài khác nhau là khác nhau.Để kinetochore gắn vững chắc vào đầu mút cựcdương của mỗi vi ống, trước hết, kinetochore “bị bắtgiữ” vào phần mép bên của vi ống để có diện tíchtiếp xúc lớn giúp việc gắn hiệu quả, sau đó vi ốngloại bỏ dần các đơn phân ở đầu cực dương đến vị trícó kinetochore thì kinetochore chuyển sang gắn vàođầu mút của vi ống (Hình 6)[1]. Khi đã gắn vào đầumút vi ống, NST bắt đầu di chuyển hướng về cựcmà vi ống có thể dài ra được. Tuy nhiên, sự dichuyển này bị kiểm soát ngay khi vi ống ở cực đốidiện gắn được với kinetochore còn lại. Tiếp theo,NST di chuyển dao động giữa hai hướng giống nhưmột “cuộc kéo co” và kết thúc bằng “một trận hòa”là NST nằm ở giữa (Hình 6). Ở kỳ giữa, các tâmđộng của tất cả NST kép đều nằm trên mặt phẳngxích đạo của thoi phân bào[4].
Như đã đề cập ở trên, sự di chuyển một cáchchính xác và có định hướng của NST nhờ phức hệkinetochore gắn vào vi ống linh động, có khả năngthay đổi chiều dài của thoi phân bào[1]. Tuy nhiên,kinetochore lại gắn vào đầu mút cực dương của viống, nơi diễn ra quá trình loại bỏ các đơn phân đểvi ống ngắn dần (Hình 6). Vậy, trong quá trình dichuyển của NST, bằng cách nào mà kinetochore vẫnduy trì được sự gắn kết với vi ống đang ngắn lại từđầu mút cực dương. Có hai mô hình được đưa ra: Môhình khuếch tán định hướng và mô hình sóng (Hình7). Ở mô hình khuếch tán định định hướng, kineto-chore có sự thay đổi lớn về cấu hình để gắn với đầumút của vi ống [1] bằng cách tạo nhiều tương tác diđộng và đa phụ trợ với vi ống làm tăng mỗi tương
ngắn dần kéo NST về cực của thoi phân bào, đồngthời làm phát sinh lực để hỗ trợ vi ống kinetochorekéo NST hướng cực. Thứ hai, ở vi ống không kine-tochore có lực phát sinh từ dòng vi ống. Dòng vi ốnglà sự di chuyển hướng cực của vi ống bằng cáchđồng thời xảy ra sự khử polymer hóa đầu cực âmvà sự polymer hóa đầu cực dương của cùng một viống (Hình 4). Thứ ba, protein motor neo giữ vi ốngthể sao với màng tế bào góp phần kéo các cực thoiphân bào ra xa nhau [4].
2. Sự gắn kết giữa NST và vi ốngCác nghiên cứu cho thấy, NST có thể gắn được
vào vi ống là nhờ một phức hệ protein gọi là kine-tochore. Cấu trúc của kinetochore gồm ba phần thểhiện ở Hình 4: Phần trong (tương tác với tâm độngcủa NST), phần ngoài (tương tác trực tiếp với viống) và phần giữa nối với phần ngoài và phần trong.Ở nấm men, phức hệ NDC80 chứa bốn protein, cócấu trúc dạng cuộn dây linh động với hai đầu hìnhcầu; phức hệ DASH (Dam1) chứa mười protein tạothành vòng tròn bao quanh vi ống, có thể duy trì gắnkết với vi ống đang dài ra hay ngắn lại, đồng thờităng cường sự ổn định gắn kết giữa NDC80 và viống [1] (Hình 5). Các protein kinetochore có khuynhhướng bảo thủ ở các loài khác nhau từ nấm đếnngười[2] (chỉ có một số trường hợp ngoại lệ, nhưngkhác nhau chủ yếu là phần trong của kinetochore)[4]. Kinetochore định vị ở tâm động của NST (Hình6, 7) [1][5]. Đối với mỗi chromatid trong cặp NSTchị em, kinetochore định vị ở tâm động của hai chro-matide ở hai hướng khác nhau và chúng gắn vào đầu
Hình 4. Ba vai trò của protein motor. (1) Protein motor liênkết với kinetochore kéo NST về phía cực thoi phân bào, (2) pro-tein motor liên kết phần gối nhau của hai vi ống không kineto-chore đối lập nhau và đưa chúng xa nhau trong khi thoi phânbào dài ra, (3) protein motor néo giữ các vi ống thể sao đangkhử polymer hóa ở đầu cực dương gần màng tế bào qua đógóp phần kéo dài thoi phân bào[6].
Hình 5. Mô hình cấu trúc của kinetochore ở nấm men[4].Phần trong kinetchore liên kết với tâm động NST, phần ngoàigắn kết với vi ống, phần giữa nối phần ngoài và phần trong.

Tạp chíKH-CN Nghệ An
SỐ 9/2016 [19]
NGHIÊN CỨU - TRAO ĐỔI
nấm men cho đến động vật có vú[8]. Các nghiên cứutiên phong ở nấm men cho thấy, bốn thành phần đógồm Smc1, Smc3, Scc1/Mcd1 và Scc3 (Hình 8).Những thành phần này sẽ lắp ráp lại để tạo nên cấutrúc mà có thể dính hai chromatid chị em với nhau.Điều đáng lưu ý là để dính được hai chromatid chịem thì cohesin phải liên kết với NST trước khi ADNnhân đôi. Nhiều mô hình khác nhau được đưa ra đểgiải thích bằng cách nào phức hệ cohesin có thể giữhai chromatid chị em với nhau và được chia thànhbốn nhóm chính, trong đó, mô hình một vòng trònđược thừa nhận rộng rãi nhất[8]. Ở mô hình này, bốntiểu đơn vị của cohesin lắp ráp lại thành một cấu trúchình vòng tròn, phân bố ở toàn bộ chromatid để giữhai chromatid chị em dính nhau (Hình 8). Chính co-hesin gắn kết hai chromatid chị em với nhau đã cungcấp lực đối trọng với lực vi ống và tạo ra sức cănglàm cấu trúc NST ổn định, tạo điều kiện thuận lợiđể các NST chị em gắn kết với vi ống và di chuyểnhiệu đến mặt phẳng xích đạo thoi phân bào.
Tuy nhiên, bắt đầu từ giai đoạn kỳ đầu, tùy thuộcvào từng loài, cohesin cần phải loại bỏ khỏi chro-
tác gắn kết với vi ống. Nghiên cứu ở nấm men chothấy, có nhiều bản sao của các yếu tố kinetochorengoài giúp kinetochore có khả năng hình thành đagắn kết với vi ống. Hơn nữa, sự tái tổ hợp phức hợpNDC80 và DASH (Dam1) phân tán nhanh chóngdọc theo vi ống giúp kinetochore gắn vào vi ốngđang ngắn dần. Ở mô hình sóng, trong quá trình khửpolymer hóa, các protofilament của vi ống xoay rangoài (Hình 2) về phía cực âm sẽ đẩy kinetochoredần dần về phía cực âm khi các đơn phân tách rakhỏi vi ống và cứ như thế sẽ kéo kinetochore dọctheo vi ống[4].
3. Sự gắn kết các NST chị em với nhauỞ pha S của chu kỳ tế bào, mỗi NST tiến hành
nhân đôi tạo thành hai chromatid giống hệt nhau(gọi là hai chromatide chị em) và dính nhau thànhmột NST kép cho đến khi kết thúc kỳ giữa thì chúngmới phân tách nhau ra. Sự dính được với nhau củahai chromatid chị em là nhờ phức hệ protein gọi làcohesin [8]. Phức hệ cohesin gồm bốn thành phầnbảo thủ ở các loài sinh vật nhân thực khác khau từ
Hình 7. Hai mô hình khác nhau về sự liên kết giữa vi ốngđang ngắn dần và NST[4].
Hình 8. Mô hình cấu trúc dạng một vòng tròn của phức hệ co-hesin ở nấm men. Cả hai NST chị em được giữ lại với nhau trongmột vòng tròn cohesin[9].
Hình 6. Sự “bắt giữ” NST vào vi ống trong quá trìnhnguyên phân của tế bào. Khởi đầu, kinetochore tiếp xúc vàgắn vào mép bên của vi ống, sau đó chuyển sang gắn vào đầumút của của vi ống. Kinetochore luôn duy trì gắn kết với đầumút của vi ống khi vi ống dài ra và ngắn lại. Khi một NST képđược định hướng đúng - có một kinetochore gắn vào vi ốngbên trái của thoi phân bào và kinetochore còn lại gắn vào viống bên phải của thoi phân bào tạo lực kéo đối lập cân bằngcủa hai NST. Các NST đã được gắn vào vi ống dao động quanhxích đạo của thoi phân bào cùng với sự dài ra và ngắn đi củahai vi ống đối lập nhau. Ở kỳ sau, sau khi sự gắn kết giữa cácchromatid chị em mất đi, các vi ống của các chromatid chị emloại bỏ dần các đơn phân dẫn đến vi ống ngắn dần đồng thờikéo các NST về các cực của thoi phân bào[1].

Tạp chíKH-CN Nghệ An
SỐ 9/2016 [20]
NGHIÊN CỨU - TRAO ĐỔI
nhất, là điều kiện tiên quyết cho sự đóng xoắncủa NST ở kỳ đầu và kỳ giữa; thứ hai, cải thiệnsự phân cắt các chromatid chị em ở những kỳsau đó; thứ ba, tránh cho cohesin bị phá hủybởi enzyme phân cắt ở các kỳ sau thay vì đượcmở ra như ở kỳ đầu. Kết quả là, ở động vật cóxương sống, tế bào thoát khỏi quá trìnhnguyên phân mà hầu như các phức hệ cohesinvẫn còn nguyên vẹn và có thể tiếp tục chuyểnsang trạng thái hoạt động ở tế bào con. Conđường kỳ đầu này có ý nghĩa sinh lý tuyệt vời,bởi vì những nghiên cứu gần đây cho thấy co-hesion có những ảnh hưởng quan trọng đến sựđiều hòa gen ở pha G1 trước khi sự kết dínhcủa các chromatid chị em được thiết lập./.
matid chị em. Ở nấm men, tất cả các phức hợp cohesindính các chromatid chị em cho đến kỳ cuối. Hoạt độngphân tách của NST chỉ xảy ra ở kỳ sau và tất cả cohesincó mặt trên NST đều được loại bỏ bởi enzyme separasetrong một bước (Hình 9B). Nhưng, ở tế bào động vật cóxương sống, trong quá trình nguyên phân, tế bào thựchiện con đường kỳ đầu. Trong con đường này, ở kỳ đầu,cohesin ở hai vai NST chuyển từ trạng thái gắn kết NST- dạng vòng tròn sang trạng thái không gắn kết NST -dạng mở mà không bị phá hủy nhờ các enzyme và cácprotein khác nhau. Nhưng ở tâm động, cohesin vẫn đượcduy trì để giữ hai chromatid chị em với nhau cho đến kỳsau do ở vị trí này cohesin được bảo vệ khỏi sự tác độngcủa các enzyme và protein trên (Hình 9A). Sự loại bỏcohesin ở kỳ đầu trong trường hợp này có thể do: thứ
Hình 9. Sự loại bỏ các phức hệ cohesin trên NST trong quá trình nguyên phân. A) Ở động vật có xương sống:phức hệ Sgo1/PP2A bảo vệ cohesin ở tâm động không bị loại bỏ cho đến kỳ sau; Sự tương tác của Wapl và Pds5chuyển cohesin từ trạng thái vòng - kết dính hai chromatid chị em sang trạng thái mở - không còn khả năng kếtdính hai chromatid chị em (Smc-Scc1 không còn tương tác với nhau nữa). B) Ở nấm men, cohesin dính 2 chromatidchị em với nhau cho đến kỳ sau. Ở kỳ sau tất cả các phức hệ cohesin dính chromatid chị em đều bị loại bỏ.
Tài liệu tham khảo:1. Asbury C.L., Tien J.F. and Davis T.N. (2011), “Kinetochores’ Gripping Feat: Conformational Wave or Biased Diffusion?”, Trends
in cell biology, 21(1)38-46.2. Ciferri C., Musacchio A. and Petrovic A. (2007), “The Ndc80 complex: Hub of kinetochore activity”, FEBS Letters, 581(15)2862–2869.3. Conde C. and Cáceres A. (2009), “Microtubule assembly, organization and dynamics in axons and dendrites”, Nature Reviews
Neuroscience, 10(5)319-332.4. Duro E. and Marston A.L. (2015), “From equator to pole: splitting chromosomes in mitosis and meiosis”, Genes & Development, 29(2)109–122.[5]. Joglekar A.P., Bouck D.C., Molk J.N., Bloom K.S. and Salmon E.D. (2006), “Molecular architecture of a kinetochore–micro-
tubule attachment site”, Nature cell biology, 8(6)581–585.6. Lodish H., Berk A., Zipursky S., Matsudaira P., Baltimore D. and Darnell J. (2000), Molecular Cell Biology. W. H. Freeman.7. Maresca T.J. and Salmon E.D. (2010), “Welcome to a new kind of tension: translating kinetochore mechanics into a wait-anaphase
signal”, J Cell Sci, 123(6)825–835.8. Mehta G.D., Rizvi S.M.A. and Ghosh S.K. (2012), “Cohesin: A guardian of genome integrity”, Biochimica et Biophysica Acta
(BBA) - Molecular Cell Research, 1823(8)1324–1342.9. Rankin K.E. and Wordeman L. (2010), “Long astral microtubules uncouple mitotic spindles from the cytokinetic furrow”, The
Journal of Cell Biology, 190(1)35-43.10. Smoyer C.J. and Jaspersen S.L. (2014), “Breaking down the wall: the nuclear envelope during mitosis”, Current Opinion in
Cell Biology, 261-9.11. Titus J. and Wadsworth P. (2001), Mitotic Spindle Assembly: The Role of Motor Proteins. eLS. John Wiley & Sons, Ltd.12. Zhang H. and Dawe R.K. (2011), “Mechanisms of plant spindle formation”, Chromosome Research: An International Journal
on the Molecular, Supramolecular and Evolutionary Aspects of Chromosome Biology, 19(3)335-344.




























