Embed Size (px)
Citation preview

TIDAL EFFECTS ON ESTUARINE ZOOPLANKTON 173
SCI. MAR., 61 (2): 173-188 SCIENTIA MARINA 1997
Tidal influence on zonation and occurrence of residentand temporary zooplankton in a shallow system
(Estuary of Mundaka, Bay of Biscay)
F. VILLATE
Landare-Biologia eta Ekologia Saila, Ekologi Laborategia, Zientzi Fakultatea, Euskal Herriko Unibertsitatea,644 P.K. E-48080 Bilbao, Spain.
SUMMARY: Tide-induced variability in the zooplankton community was analysed during 2 tidal series of 14 h duringspring tides and neap tides in the shallow estuary of Mundaka by relating short-term changes in composition and spatial dis-tribution of populations to tide-associated parameters (water level, current velocity and direction, mixing and stratification,and salinity). A strong tidal influence was found in the zooplankton composition and distribution on a day-week time scale(spring-neap tidal cycles) and at hour time scale (high-low tidal cycle). During spring tides, suspended meiofauna showshigh densities at flood and ebb tides; the neritic zooplankton can reach the inner estuary at high water, and the autochtho-nous zooplankton move from the inner zone, at high water, to the outer zone, at low water, over distances of more than onethird of the estuary length. During neap tides, autochthonous populations remain around the low middle estuary, the pene-tration of neritic zooplankton is feeble, and the occurrence of benthic forms in the water column is negligible. The observedeffects on the abundance and zonation of resident and temporary zooplankton indicate that tides are a key factor in the estu-ary of Mundaka by regulating the stock of neritic and autochthonous zooplankton within the estuary, the reproductive suc-cess of benthic populations which have planktonic larval development, and meiofauna dispersion and availability as food inthe water column.
Key words: Zooplankton, tychoplankton, tidal cycle, transport, estuaries.
RESUMEN: INFLUENCIA DE LAS MAREAS EN LA ZONACIÓN Y PRESENCIA DEL ZOOPLANCTON EN UN SISTEMA COSTERO (ESTUARIODE MUNDAKA, BAHIA DE VIZCAYA). – Los cambios en la comunidad zooplanctónica por efecto de las mareas fueron analiza-dos durante 2 series mareales de 14 h en mareas vivas y muertas en el estuario somero de Mundaka, relacionando los cam-bios a pequeña escala temporal de la composición y distribución espacial de las poblaciones con parámetros ligados a lasmareas (nivel del agua, velocidad y dirección de corriente, mezcla y estratificación y salinidad). El efecto de la marea sobrela composición y la distribución del zooplancton fue muy notorio tanto a una escala temporal de dias-semanas (ciclos demarea vivas-muertas) como a una escala de tiempo horaria (ciclos pleamar-bajamar). En mareas vivas, la densidad de meio-fauna resuspendida es elevada durante el ascenso y el descenso de la marea, el zooplancton nerítico alcanza el tramo interi-or del estuario en pleamar y el zooplancton autóctono es transportado desde el tramo alto del estuario, en pleamar, hasta eltramo exterior, en bajamar, sobre distancias superiores a un tercio de la longitud total del estuario. En mareas muertas, laspoblaciones autóctonas permanecen alrededor del tramo medio bajo del estuario, la penetración del zooplancton nerítico esescasa y la aparición de formas bentónicas en la columna de agua es irrelevante. El efecto observado sobre la abundancia ydistribución del zooplancton residente y temporal indica que en el estuario de Mundaka las mareas son un factor determi-nante en la regulación de los efectivos de zooplancton nerítico y autóctono en el estuario, los sucesos reproductivos de laspoblaciones bentónicas con desarrollo larvario planctónico, y la dispersión y disponibilidad como alimento de la meiofau-na en la columna de agua.
Palabras clave: Zooplancton, ticoplancton, ciclos de marea, transporte, estuarios.
*Received August 23, 1996. Accepted February 10, 1997.
INTRODUCTION
Tides play a major role in the functioning ofmany coastal systems, being responsible for notice-able mid-term (spring-neap cycles) and short-term(low-high water cycles) variations in the abiotic andbiotic characteristics of these systems. This role hasinspired several authors to analyse tidal control ofcommunities and processes in different systems.Examples include: tide-induced phytoplankton andmicrophytobenthos exchanges (Riaux-Gobin,1987); tidal influence on bacteria, microphytoplank-ton and microzooplankton abundance (Morales-Zamorano et al., 1991); tidal control of seston quan-tity and quality (Flegey et al., 1992); tidal export ofparticulate organic matter (Boto and Bunt, 1981);tidal redistribution of macrodetrital aggregates
(Whitfield, 1988); and tidal influence on sedimenttransport and settling (Uncles et al., 1992; TenBrinke & Dronkers, 1993; Ten Brinke, 1994; Smithand FitzGerald, 1994).
Regarding zooplankton, several papers havepointed to the importance of tidal dynamics induc-ing spatial variability in zooplankton distributionwithin estuaries (e.g. Lee and McAlice, 1979; All-dredge and Hammer, 1980; Gagnon and Lacroix,1981, 1982). However, tides are not only responsi-ble for distributional patterns, but also for dynamicevents and survival of endemic populations byexporting and reducing their standing stocks. Thiseffect of tidal exchange in regulating estuarine zoo-plankton may be especially relevant in small sys-tems where strong river discharges can occur (Ken-nish, 1990).
174 F. VILLATE
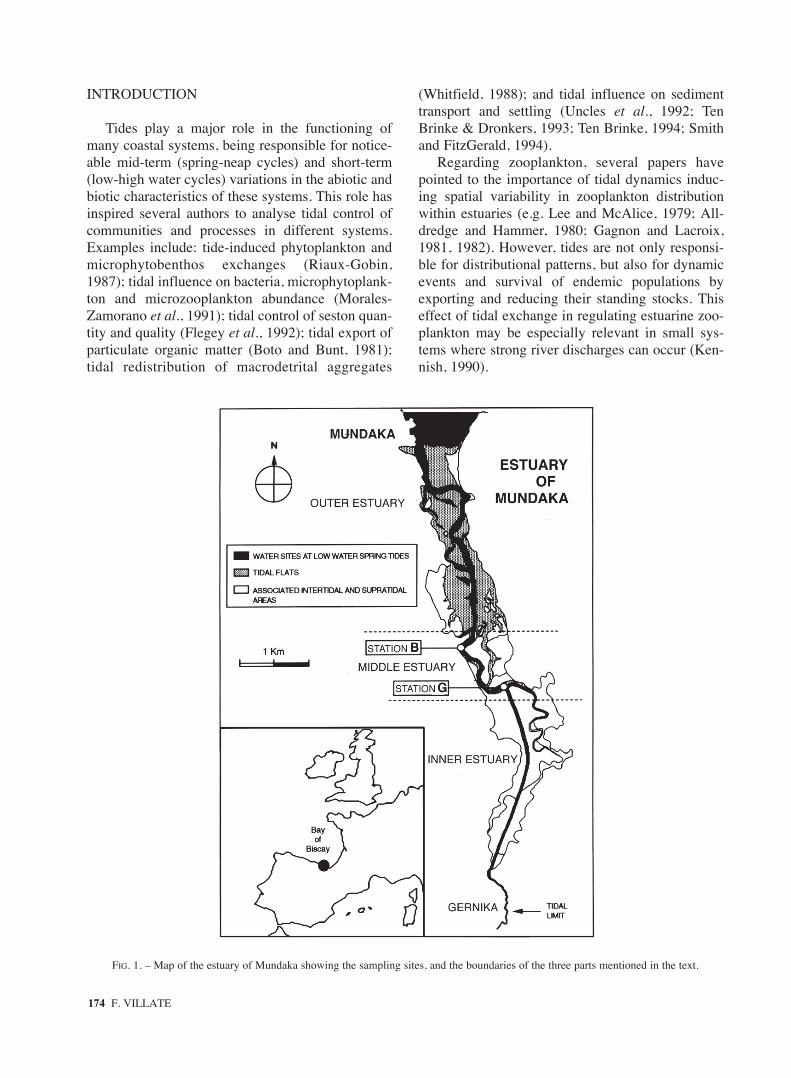
FIG. 1. – Map of the estuary of Mundaka showing the sampling sites, and the boundaries of the three parts mentioned in the text.
This report examines the effects of tides on mid-term and short-term variability of suspended livingorganisms larger than 200 µm in a shallow tidal sys-tem (the estuary of Mundaka). The aim of the studywas to evaluate the importance of changes in zoo-plankton composition, abundance and zonationinduced by tides in this type of estuary.
MATERIAL AND METHODS
Study area
The estuary of Mundaka is a shallow meso-tidalsystem located in the south-east of the Bay of Bis-cay, on the Basque Coast (Fig. 1). It forms the cen-tral axis of the Urdaibai Biosphere Reserve, is
around 13 km long, and is characterized by threeclearly distinguishable physiographic parts. Theouter part is a broad area open to the sea and filledby large intertidal flats, the middle part is occupiedmainly by emerging salt-marshes, and the inner partcomprises a narrow man-made channel that meetsthe meandering channel of the river in the upperestuary and its surrounding wetlands.
The river inflow is normally low (monthly meanabout 1 m3 s-1 from the main tributary) in relation tothe volume of the estuarine basin (mean volume of32.9 105 m3), such that the estuary is mainly polyha-line and euhaline. However, strong increases infreshwater inputs are frequent due to the torrentialregime of tributary streams. As a result, the resi-dence time is highly variable, and calculated flush-ing times (21 to 581 days) at moderate and low river
TIDAL EFFECTS ON ESTUARINE ZOOPLANKTON 175
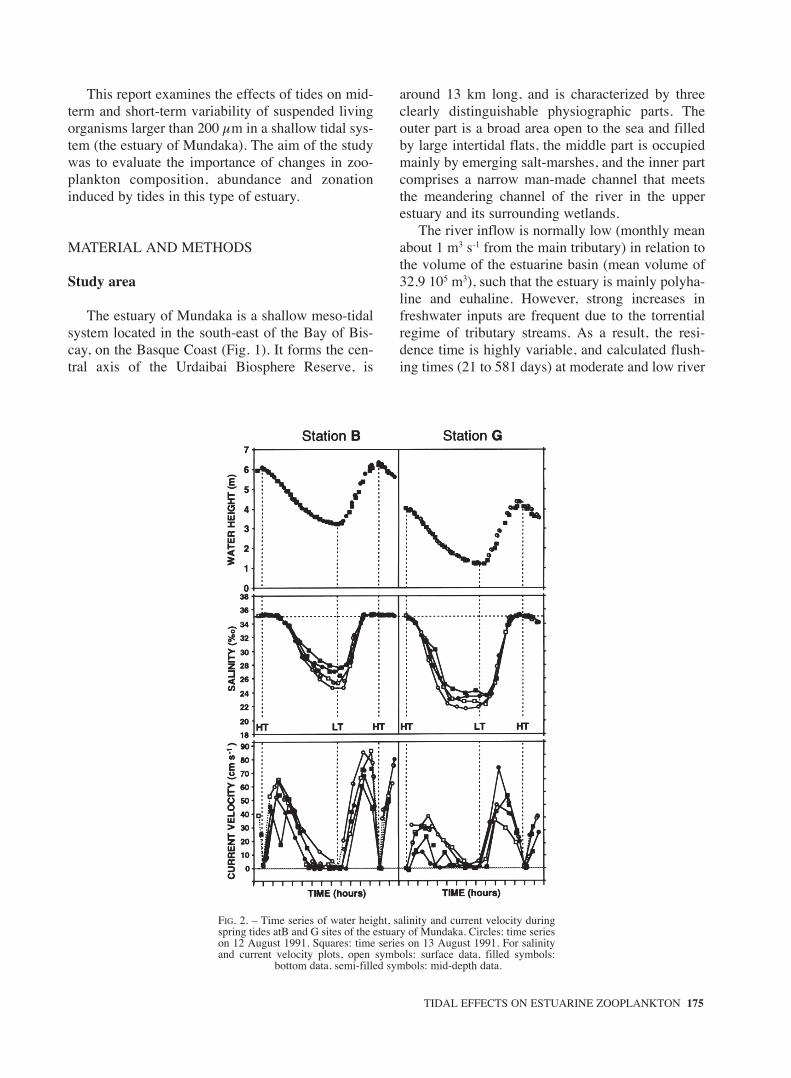
FIG. 2. – Time series of water height, salinity and current velocity duringspring tides atB and G sites of the estuary of Mundaka. Circles: time serieson 12 August 1991. Squares: time series on 13 August 1991. For salinityand current velocity plots, open symbols: surface data, filled symbols:
bottom data, semi-filled symbols: mid-depth data.
discharges differ largely (Villate et al., 1989). Tidesare semidiurnal, and at spring tide the high water isnot significantly delayed between the outer estuaryand the upper reaches. The tidal exchange is high,with the mean tidal prism-mean basin volume ratiogreater than 1 (Villate et al., 1989).
Previous research on mesozooplankton (Vil-late, 1989-90; 1991; Villate et al., 1993) showedthat true estuarine holoplankters such as Eury-temora species cannot develop in the estuary ofMundaka. The endemic mesozooplankton is dom-inated by the estuarine-marine species Acartiabifilosa, which reach the highest density in sum-mer. The meroplankton contributes largely to thetotal zooplankton, and is dominated by barnacleand polychaete spionid larvae in spring and bygastropod larvae in summer.
Sampling and data
Data presented here were obtained from analy-ses of samples collected during a study on the tidaleffects on seston quantity and quality in the estuaryof Mundaka. The study was carried out in summer(August 1991) when disturbances induced by river-runoff fluctuations are smaller and the develop-ment of endemic zooplankton reaches the annualmaximum.
Sampling was performed at two sites in themain channel during two spring tidal cycles on 12-13 August and two neap tidal cycles on 18-19August. Sampling sites were located at the lowerpart (station B) and at the upper part (station G) ofthe middle estuary (Fig. 1). These sites wereselected to obtain the most accurate estimation of
176 F. VILLATE
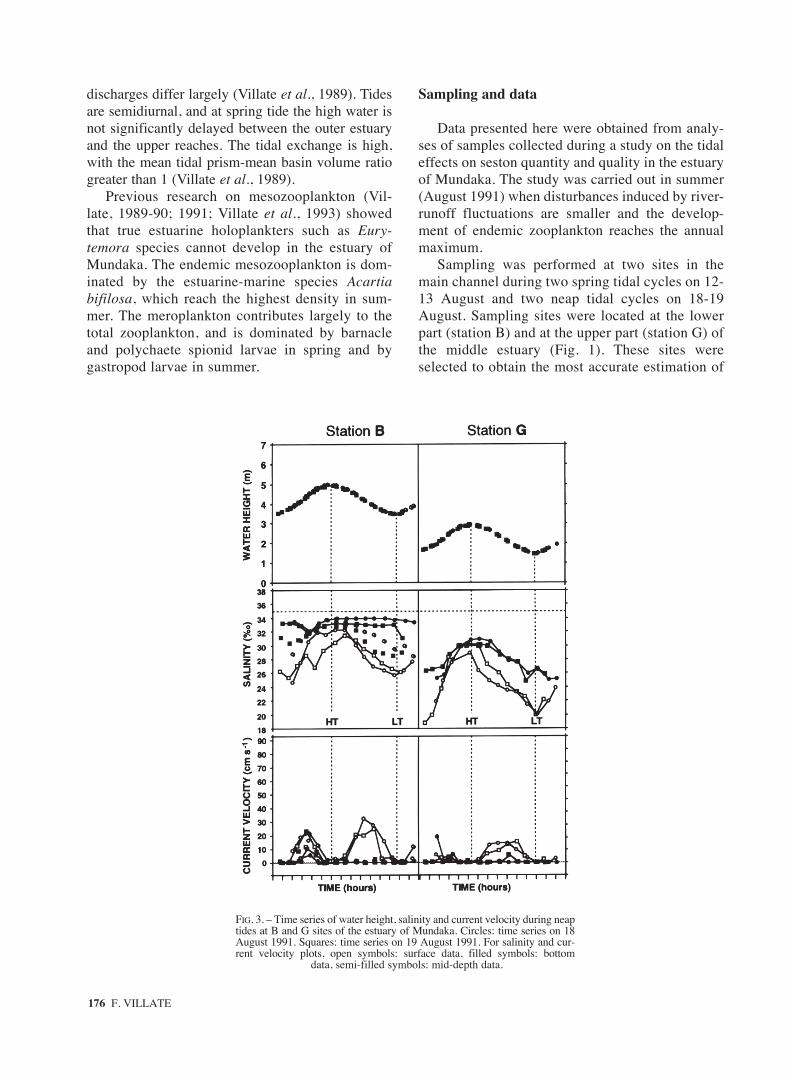
FIG. 3. – Time series of water height, salinity and current velocity during neaptides at B and G sites of the estuary of Mundaka. Circles: time series on 18August 1991. Squares: time series on 19 August 1991. For salinity and cur-rent velocity plots, open symbols: surface data, filled symbols: bottom
data, semi-filled symbols: mid-depth data.
populations moving throughout the estuary duringthe tidal cycle. Seaward to the B site, data record-ed at a single point may be less suitable becausethe basin is wider and the water circulationbecomes more complex. Upward to the G site,although most the water volume moves into theartificial channel, estuarine waters and popula-tions are also derived from the original channel.When tides fall, waters coming from both chan-nels meet just up from the G site.
Samples were taken from a central point at eachsite over a 14-hour period, beginning at high tide inspring tide series and at low tide in neap tide series.
Zooplankton samples were taken by pump at 2-hour intervals from surface water (within theupper 0.5 m layer), 0.5 m above the bottom, and atmid depth (equi-distance from the surface and bot-tom samples). Mid-depth samples were not takenwhen the water column was less than 2 m deep.Zooplankton samples were obtained by filtering30 l of the pumped water through a 200 µm sieve,preserved in 5% formalin and stained with RoseBengal.
Hourly profiles of salinity were recorded with afield salinometer WTW 196, and discrete mea-surements of the current velocity (at the samedepth as zooplankton samples), were also made by
using a hydraulic flow meter General Oceanics.The tidal height was determined by recording thewater level on graduated stakes located at eachsampling site. Measurements were made justbefore and after sampling.
For zooplankton analysis, the entire sample wasexamined under a stereoscopic microscope. All indi-viduals were counted, and keyed to different taxo-nomic categories. Most of the holoplanktonic formswere identified to species level, while meroplank-tonic larvae and benthic or littoral forms were usu-ally keyed to family, order or class categories. Afteridentification, and for this study, all taxa weregrouped into wider categories with ecological meanin the estuarine ecosystem.
Abundance of zooplankton taxa is expressed asnumber of individuals per litre of water, and as apercentage of the total zooplankton in each samplingseries. Short-term changes in zooplankton areexpressed in relative abundance, and data from con-secutive days are plotted together in figures, aftersettling high-tide times, in an attempt to obtain amore accurate estimate and to show the generaltrends of zooplankton populations at both spring andneap tides. Organism abundance in relation to tidalphases is expressed as a percentage of the total foreach taxon and sampling series.
TIDAL EFFECTS ON ESTUARINE ZOOPLANKTON 177
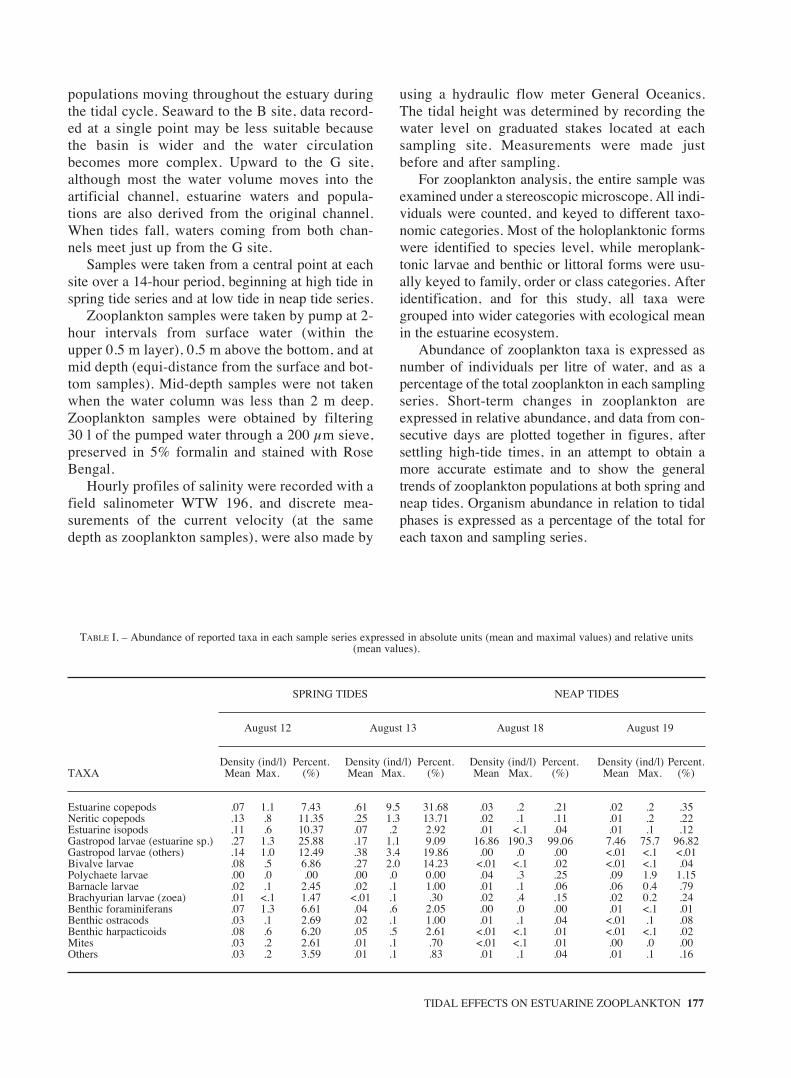
TABLE I. – Abundance of reported taxa in each sample series expressed in absolute units (mean and maximal values) and relative units(mean values).
SPRING TIDES NEAP TIDES
August 12 August 13 August 18 August 19
Density (ind/l) Percent. Density (ind/l) Percent. Density (ind/l) Percent. Density (ind/l) Percent.TAXA Mean Max. (%) Mean Max. (%) Mean Max. (%) Mean Max. (%)
Estuarine copepods .07 1.1 7.43 .61 9.5 31.68 .03 .2 .21 .02 .2 .35Neritic copepods .13 .8 11.35 .25 1.3 13.71 .02 .1 .11 .01 .2 .22Estuarine isopods .11 .6 10.37 .07 .2 2.92 .01 <.1 .04 .01 .1 .12Gastropod larvae (estuarine sp.) .27 1.3 25.88 .17 1.1 9.09 16.86 190.3 99.06 7.46 75.7 96.82Gastropod larvae (others) .14 1.0 12.49 .38 3.4 19.86 .00 .0 .00 <.01 <.1 <.01Bivalve larvae .08 .5 6.86 .27 2.0 14.23 <.01 <.1 .02 <.01 <.1 .04Polychaete larvae .00 .0 .00 .00 .0 0.00 .04 .3 .25 .09 1.9 1.15Barnacle larvae .02 .1 2.45 .02 .1 1.00 .01 .1 .06 .06 0.4 .79Brachyurian larvae (zoea) .01 <.1 1.47 <.01 .1 .30 .02 .4 .15 .02 0.2 .24Benthic foraminiferans .07 1.3 6.61 .04 .6 2.05 .00 .0 .00 .01 <.1 .01Benthic ostracods .03 .1 2.69 .02 .1 1.00 .01 .1 .04 <.01 .1 .08Benthic harpacticoids .08 .6 6.20 .05 .5 2.61 <.01 <.1 .01 <.01 <.1 .02Mites .03 .2 2.61 .01 .1 .70 <.01 <.1 .01 .00 .0 .00Others .03 .2 3.59 .01 .1 .83 .01 .1 .04 .01 .1 .16
RESULTS
Hydrodynamic features
At spring tides (Fig. 2), tidal amplitude rangedfrom 3.13 m (12 August) to 2.94 m (13 August), andthe deep-channel water depth at low tide wasapproximately 3.25 m at station B and 1.25 m at sta-tion G. The water column appeared homogenousmost of the time, although a weak stratificationdeveloped in both stations at low water. Salinityabove 35‰ was found in both stations at high water,while values fell to 29-25‰ at station B and to 25-22‰ at station G at low water. Current-speed mea-surements showed that a sudden rise followed by amore gradual decrease of the ebb-tide velocityoccurred after the high tide slack, and an extendedperiod of slow currents began before low water,
which occurred around seven and a half hours afterhigh water. The flooding period was shorter than theebb, and the step from maximum flood to maximumebb occurred in less than three hours, with only abrief period of slack currents around high water. Themain differences between surface and bottom cur-rent velocities occurred during ebb at the inner sta-tion (G), and was lower at near-bed velocity.
During neap tides (Fig. 3), tidal amplitude wasbetween 1.48 m (18 August) and 1.39 m (19August), but depths at low water were similar tothose measured during spring tides (3.5 m at stationB and 1.5 at station G). Salinity stratification wasnotable at both stations, being maximal around thelow tide slack. Surface salinity ranged between24‰ and 33‰ at the outer station, and between18‰ and 30‰ at the inner station. Bottom salinitywas of 30-34‰ and 24-32‰ at the outer and inner
178 F. VILLATE
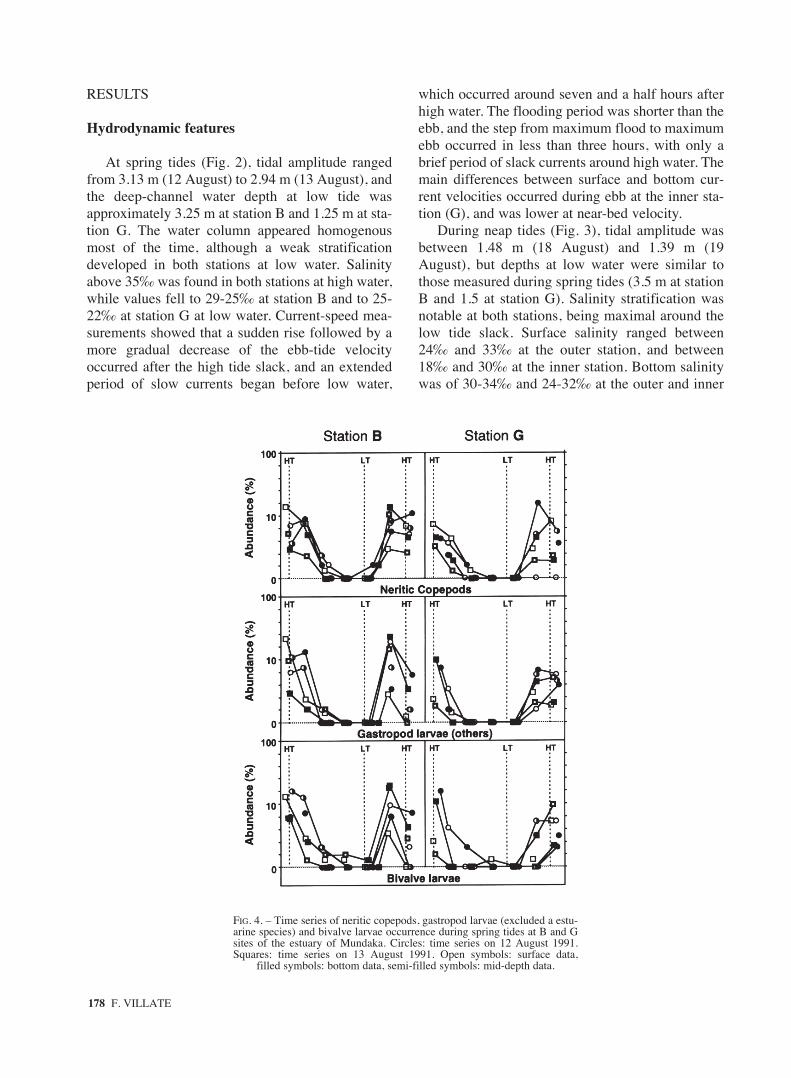
FIG. 4. – Time series of neritic copepods, gastropod larvae (excluded a estu-arine species) and bivalve larvae occurrence during spring tides at B and Gsites of the estuary of Mundaka. Circles: time series on 12 August 1991.Squares: time series on 13 August 1991. Open symbols: surface data,
filled symbols: bottom data, semi-filled symbols: mid-depth data.

stations respectively. Flood and ebb times weresimilar, but the velocity structure in the water col-umn, and the salinity vertical pattern differedbetween flood and ebb tides. Higher flood currentsin surface and near-bed occurred in a short period atthe middle flood, when the water column showed agreater saline homogeneity at the same time. As thetide fell, downstream currents were only evident atthe surface, where salinity decreased more drasti-cally. Near-bed salinity also decreased at the innerstation, but the highest saline waters (33-34‰)remained in depth at the outer station until lowwater. Salinity also differed in sampled sites fromthe first day to the next. Waters of 34‰ and 31‰reached in depth the outer and the inner stationsrespectively the first day, but the next day the high-er salinities recorded were around 33‰ and 30‰respectively.
Both flood and ebb currents were alwaysstronger at the outer station. During the neap tide,maximal current velocities were around three timessmaller than those of spring tides at the surface, andfour to five times smaller near the bottom.
Zooplankton composition and abundance
As shown in Table I, gastropod larvae, holoplank-tonic estuarine and neritic copepods, bivalve larvae andestuarine isopods were the most abundant taxa in sam-ples taken at the spring tide series. Among gastropodlarvae one species inhabiting brackish waters (estuar-ine species) was distinguished from species with highsalinity preferences. These estuarine larvae probablybelong to the mud snail Hydrobia ulvae, which is veryabundant in the intertidal sediments of the estuary ofMundaka, although we found that the planktonic phase
TIDAL EFFECTS ON ESTUARINE ZOOPLANKTON 179
FIG. 5. – Time series of estuarine isopods, estuarine copepods and estuar-ine gastropod larvae occurrence during spring tides at B and G sites of theestuary of Mundaka. Circles: time series on 12 August 1991. Squares: timeseries on 13 August 1991. Open symbols: surface data, filled symbols:
bottom data, semi-filled symbols: mid-depth data.
time of these larvae was at least two weeks (Villate etal., 1993) while the planktonic larval phase reportedfrom other locations is very short (e.g., Armonies andHartke, 1995). Endemic copepods (grouped in theestuarine copepod category) were dominated by Acar-tia bifilosa. Low densities of Acartia grani, and someindividuals of Calanipeda aquae-dulcis were alsofound. Among neritic copepods, Paracalanus parvus,Oncaea media and Euterpina acutifrons were the more
representative species. Isopods were mainly of theestuarine species Paragnathia formica. Benthic cope-pods (mainly harpacticoids), foraminiferans and ostra-cods, and mites were also found in zooplankton sam-ples taken during spring tides.
Gastropod larvae of the estuarine species domi-nated zooplankton largely in the middle estuary atneap tides. A maximal density of 190 ind l-1 wasrecorded from a sample taken on August 18. These
180 F. VILLATE
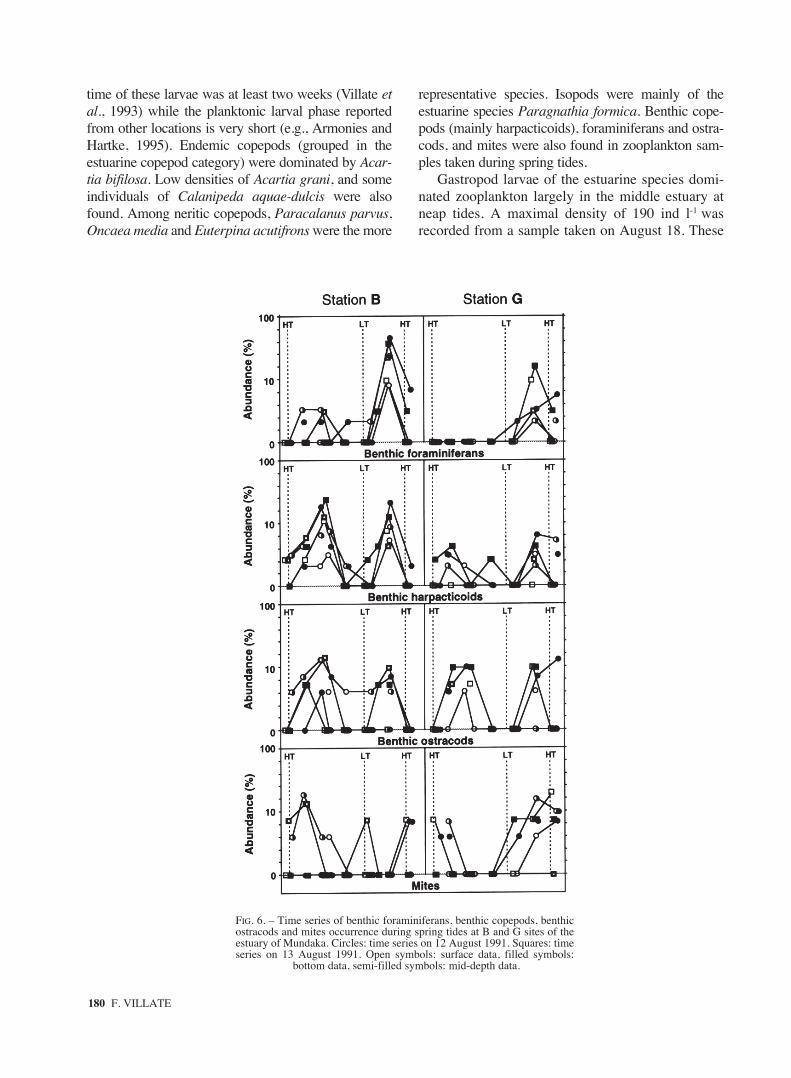
FIG. 6. – Time series of benthic foraminiferans, benthic copepods, benthicostracods and mites occurrence during spring tides at B and G sites of theestuary of Mundaka. Circles: time series on 12 August 1991. Squares: timeseries on 13 August 1991. Open symbols: surface data, filled symbols:
bottom data, semi-filled symbols: mid-depth data.
high densities of gastropod larvae constituted morethan 95% of the total zooplankton sampled in bothneap-tide series. Polychaete, barnacle and brachyu-ran larvae, and holoplanktonic estuarine and neriticcopepods were abundant in some samples, but theircontribution to the total zooplankton was very small.The occurrence and contribution of benthic forms,however, was negligible. Copepod species composi-tion did not differ from that observed at spring tides:A. bifilosa dominated in the autochthonous assem-blage and P. parvus, O. media and E. acutifrons inthe neritic assemblage.
The densities of estuarine larvae of gastropodspolychaetes barnacles and brachyurans increasedfrom the spring to the neap tide period, while thedensities of estuarine copepods and isopods, andneritic populations decreased.
Zooplankton dynamics throughout the high-lowwater cycles
Spring tides
Taxa of neritic origin showed higher densitiesaround high water, and were almost absent at lowwater at both stations (Fig. 4). Their maximaldensities, however, were found moving upstreamacross the outer station at flood-tide, just afterthe sudden flood rise occurred and waters above35‰ reached the middle estuary. They movedupstream across the outer station mainly in thesurface on August 12, but mainly at depth onAugust 13.
Among autochthonous taxa (Fig. 5), the highestabundance of estuarine isopods (P. formica)
TIDAL EFFECTS ON ESTUARINE ZOOPLANKTON 181
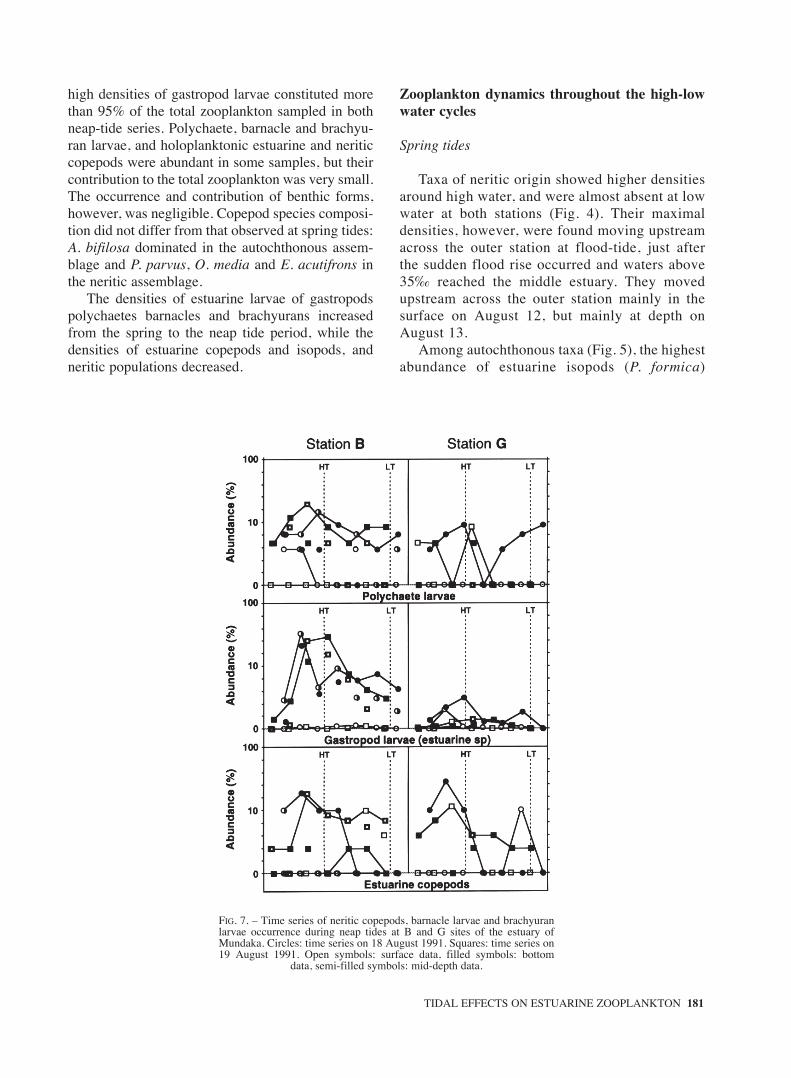
FIG. 7. – Time series of neritic copepods, barnacle larvae and brachyuranlarvae occurrence during neap tides at B and G sites of the estuary ofMundaka. Circles: time series on 18 August 1991. Squares: time series on19 August 1991. Open symbols: surface data, filled symbols: bottom
data, semi-filled symbols: mid-depth data.
appeared around low water at the outer station, butonly occurred occasionally at this station aroundhigh water. At the inner station they were foundover all tidal phases, although the highest densitiesin the water column were observed around lowwater. On August 12, the highest densities of A.bifilosa were recorded at the outer station movingdownstream at the middle ebb, in waters of 31-32‰, but this peak was not found movingupstream at flood. The next day, however, thehighest densities were only recorded movingupstream at the first flooding period, in waters of29-30‰. Although maximal abundance observedat the inner station was always lower, we can sup-pose that the population patch moved across thisstation during unsampled flood and ebb times. Thegastropod larvae of the estuarine species showed asimilar variation pattern in relation to the tidal
cycle. They moved downstream and upstream atthe middle ebb and flood respectively, but theirhigher densities skewed towards higher salinitywaters.
Among the tychoplankton (Fig. 6), organismsinoculated periodically into the plankton by bot-tom currents (Kennish, 1990), the density of ben-thic foraminiferans in the water column differedgreathy between the outflow and the inflow peri-ods. They peaked mainly in the water columncoinciding with the stronger flood currents, andmoved upstream in waters above 35‰. In con-trast, benthic harpacticoids appeared in similarnumbers at ebb and flood phases, but ebb andflood peaks occurred at different current veloci-ties, and in waters of different salinity. At ebb,harpacticoids peaked in waters of 31-32‰, andafter maximal currents occurred. At flood, how-
182 F. VILLATE
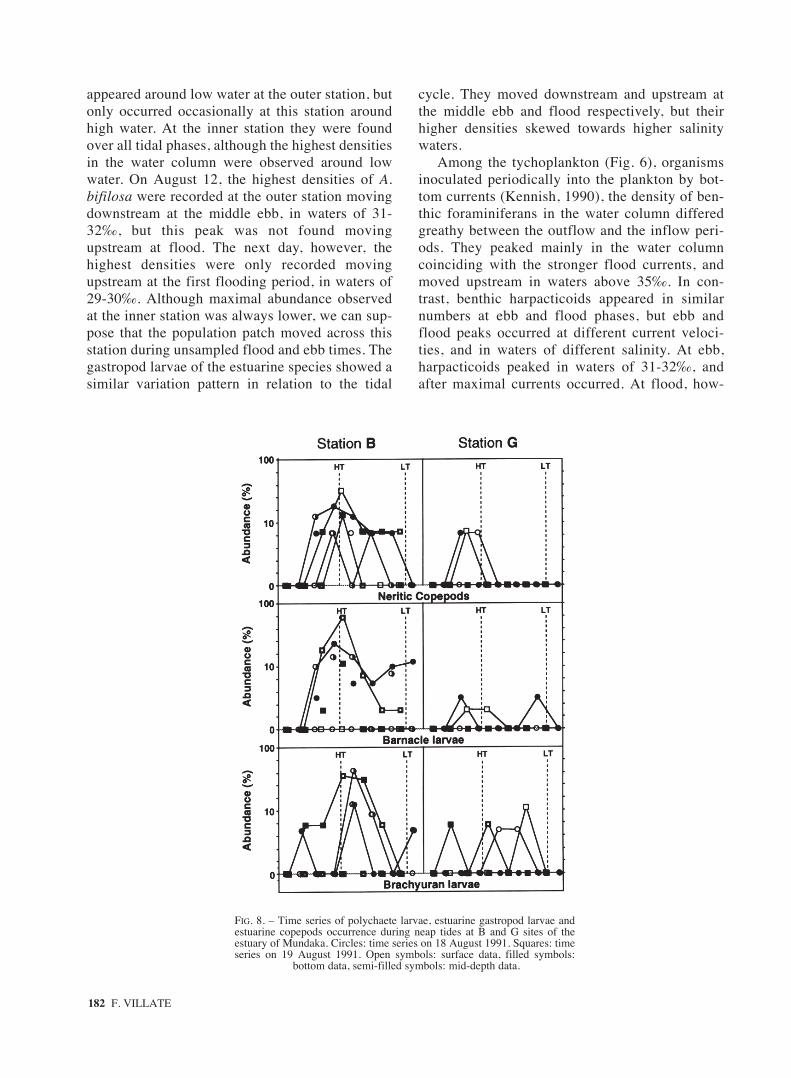
FIG. 8. – Time series of polychaete larvae, estuarine gastropod larvae andestuarine copepods occurrence during neap tides at B and G sites of theestuary of Mundaka. Circles: time series on 18 August 1991. Squares: timeseries on 19 August 1991. Open symbols: surface data, filled symbols:
bottom data, semi-filled symbols: mid-depth data.
ever, they peaked in waters around 35‰ duringthe maximum flooding current. Benthic ostracodsshowed an occurrence pattern similar to that ofharpacticoids. However, the higher densities ofostracods were recorded at mid depth while thehigher densities of foraminiferans and harpacti-coids appeared at the bottom. Mites were moreabundant in surface and mid depth around highwater at the inner station, and just after the highwater slack at the outer station.
Neap tides
As shown in Fig. 7, neritic copepods onlyoccurred in noticeable numbers at high water inthe outer station. At this site they were presentlonger during ebb, and in waters of higher salini-ty remained near the bottom. Barnacle larvae also
peaked during high water at the outer station, buton August 18 their abundance also increased dur-ing low water at this site. The brachyuran larvaewere mainly found at high water and the first halfof the ebb. All these groups only appeared spo-radically at the inner station.
The higher densities of polychaete larvae, gas-tropod larvae and autochthonous copepods (Fig. 8)occurred at the outer site around high tide or in thelate flood period, just following a sudden rise ofthe flood-tide velocity and a decrease of the salineheterogeneity in the water column. Over the ebbtide they remained at this site in greater abundancethan that found in the early flood. Polychaete andgastropod larvae decreased more drastically thanestuarine copepods toward the inner station, wheretheir temporal trends in relation to the tidal cyclewere not so clear.
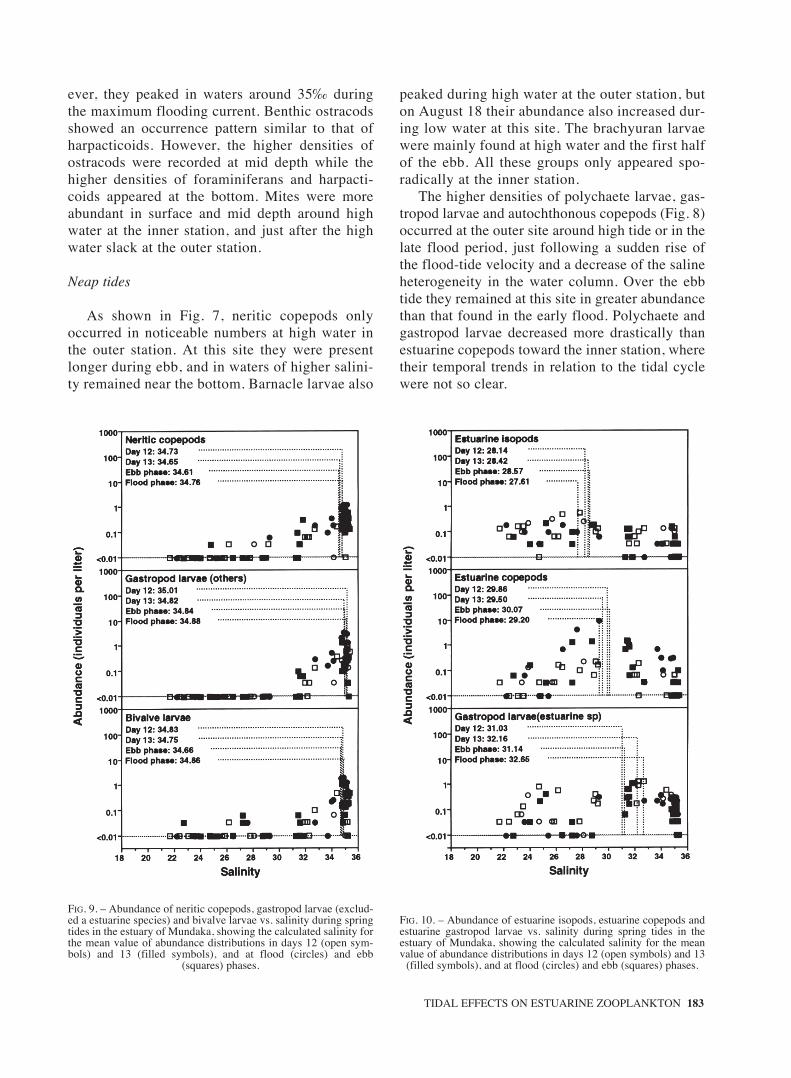
TIDAL EFFECTS ON ESTUARINE ZOOPLANKTON 183
FIG. 9. – Abundance of neritic copepods, gastropod larvae (exclud-ed a estuarine species) and bivalve larvae vs. salinity during springtides in the estuary of Mundaka, showing the calculated salinity forthe mean value of abundance distributions in days 12 (open sym-bols) and 13 (filled symbols), and at flood (circles) and ebb
(squares) phases.
FIG. 10. – Abundance of estuarine isopods, estuarine copepods andestuarine gastropod larvae vs. salinity during spring tides in theestuary of Mundaka, showing the calculated salinity for the meanvalue of abundance distributions in days 12 (open symbols) and 13
(filled symbols), and at flood (circles) and ebb (squares) phases.
Variations in relation to water masses
Spring tides
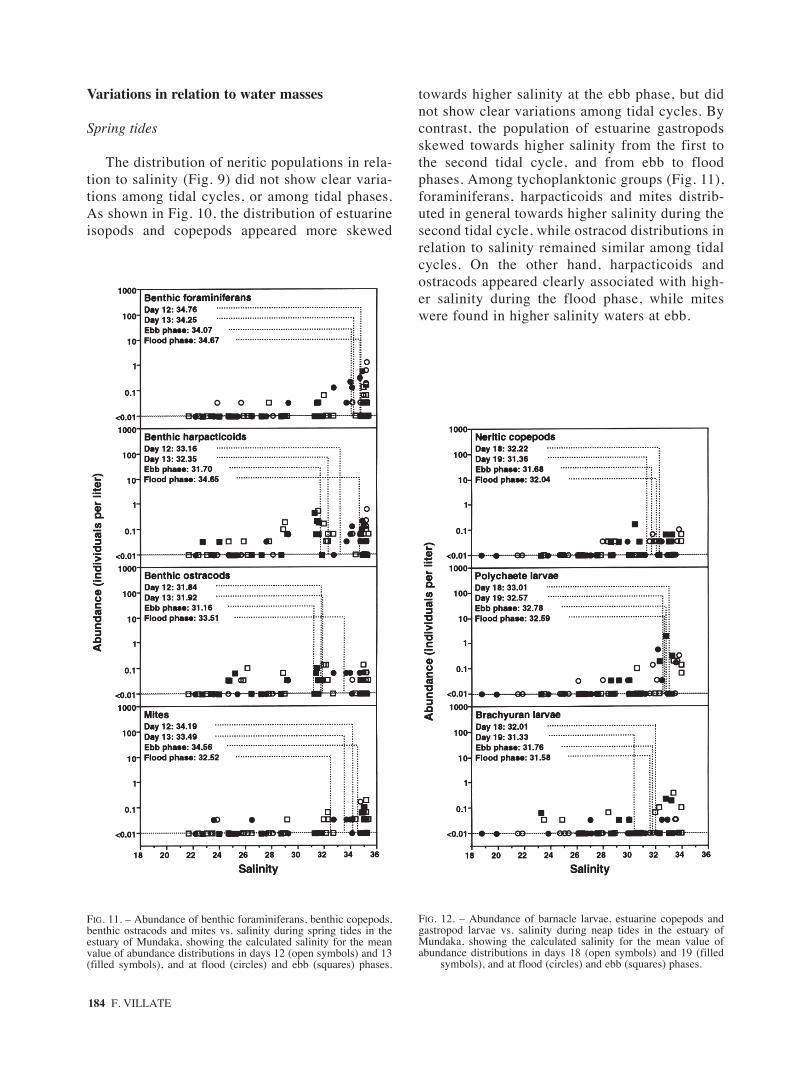
The distribution of neritic populations in rela-tion to salinity (Fig. 9) did not show clear varia-tions among tidal cycles, or among tidal phases.As shown in Fig. 10, the distribution of estuarineisopods and copepods appeared more skewed
towards higher salinity at the ebb phase, but didnot show clear variations among tidal cycles. Bycontrast, the population of estuarine gastropodsskewed towards higher salinity from the first tothe second tidal cycle, and from ebb to floodphases. Among tychoplanktonic groups (Fig. 11),foraminiferans, harpacticoids and mites distrib-uted in general towards higher salinity during thesecond tidal cycle, while ostracod distributions inrelation to salinity remained similar among tidalcycles. On the other hand, harpacticoids andostracods appeared clearly associated with high-er salinity during the flood phase, while miteswere found in higher salinity waters at ebb.
184 F. VILLATE
FIG. 11. – Abundance of benthic foraminiferans, benthic copepods,benthic ostracods and mites vs. salinity during spring tides in theestuary of Mundaka, showing the calculated salinity for the meanvalue of abundance distributions in days 12 (open symbols) and 13(filled symbols), and at flood (circles) and ebb (squares) phases.
FIG. 12. – Abundance of barnacle larvae, estuarine copepods andgastropod larvae vs. salinity during neap tides in the estuary ofMundaka, showing the calculated salinity for the mean value ofabundance distributions in days 18 (open symbols) and 19 (filled
symbols), and at flood (circles) and ebb (squares) phases.
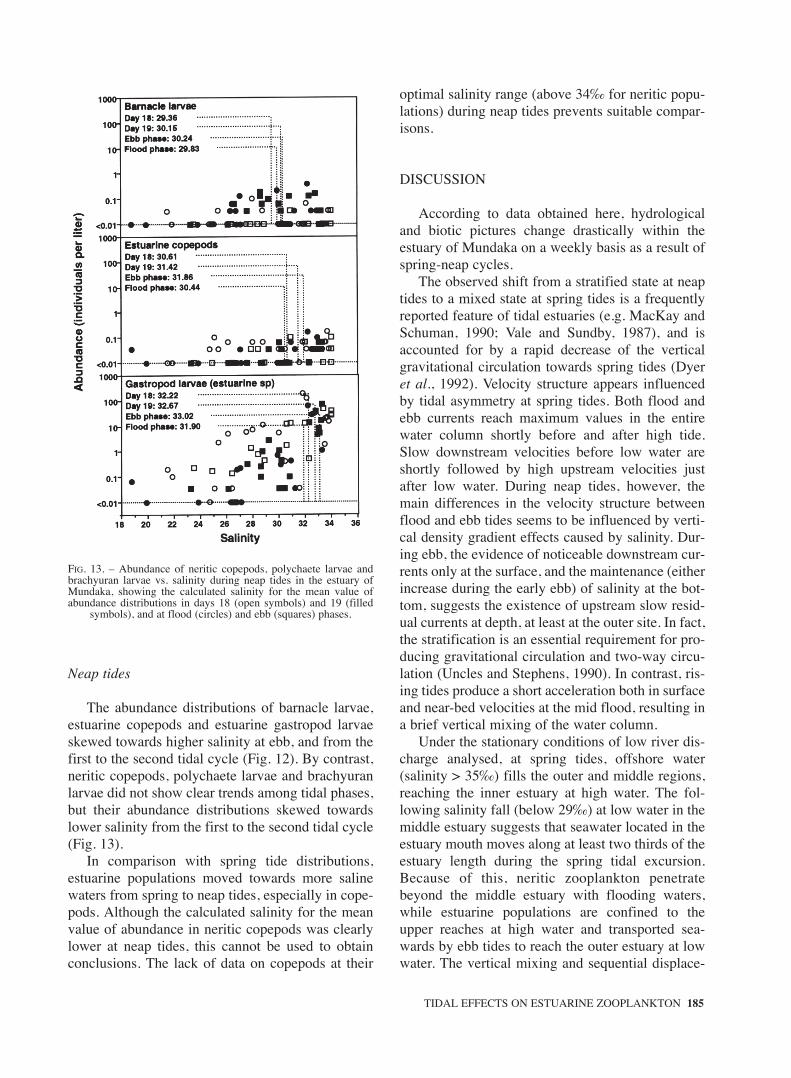
Neap tides
The abundance distributions of barnacle larvae,estuarine copepods and estuarine gastropod larvaeskewed towards higher salinity at ebb, and from thefirst to the second tidal cycle (Fig. 12). By contrast,neritic copepods, polychaete larvae and brachyuranlarvae did not show clear trends among tidal phases,but their abundance distributions skewed towardslower salinity from the first to the second tidal cycle(Fig. 13).
In comparison with spring tide distributions,estuarine populations moved towards more salinewaters from spring to neap tides, especially in cope-pods. Although the calculated salinity for the meanvalue of abundance in neritic copepods was clearlylower at neap tides, this cannot be used to obtainconclusions. The lack of data on copepods at their
optimal salinity range (above 34‰ for neritic popu-lations) during neap tides prevents suitable compar-isons.
DISCUSSION
According to data obtained here, hydrologicaland biotic pictures change drastically within theestuary of Mundaka on a weekly basis as a result ofspring-neap cycles.
The observed shift from a stratified state at neaptides to a mixed state at spring tides is a frequentlyreported feature of tidal estuaries (e.g. MacKay andSchuman, 1990; Vale and Sundby, 1987), and isaccounted for by a rapid decrease of the verticalgravitational circulation towards spring tides (Dyeret al., 1992). Velocity structure appears influencedby tidal asymmetry at spring tides. Both flood andebb currents reach maximum values in the entirewater column shortly before and after high tide.Slow downstream velocities before low water areshortly followed by high upstream velocities justafter low water. During neap tides, however, themain differences in the velocity structure betweenflood and ebb tides seems to be influenced by verti-cal density gradient effects caused by salinity. Dur-ing ebb, the evidence of noticeable downstream cur-rents only at the surface, and the maintenance (eitherincrease during the early ebb) of salinity at the bot-tom, suggests the existence of upstream slow resid-ual currents at depth, at least at the outer site. In fact,the stratification is an essential requirement for pro-ducing gravitational circulation and two-way circu-lation (Uncles and Stephens, 1990). In contrast, ris-ing tides produce a short acceleration both in surfaceand near-bed velocities at the mid flood, resulting ina brief vertical mixing of the water column.
Under the stationary conditions of low river dis-charge analysed, at spring tides, offshore water(salinity > 35‰) fills the outer and middle regions,reaching the inner estuary at high water. The fol-lowing salinity fall (below 29‰) at low water in themiddle estuary suggests that seawater located in theestuary mouth moves along at least two thirds of theestuary length during the spring tidal excursion.Because of this, neritic zooplankton penetratebeyond the middle estuary with flooding waters,while estuarine populations are confined to theupper reaches at high water and transported sea-wards by ebb tides to reach the outer estuary at lowwater. The vertical mixing and sequential displace-
TIDAL EFFECTS ON ESTUARINE ZOOPLANKTON 185
FIG. 13. – Abundance of neritic copepods, polychaete larvae andbrachyuran larvae vs. salinity during neap tides in the estuary ofMundaka, showing the calculated salinity for the mean value ofabundance distributions in days 18 (open symbols) and 19 (filled
symbols), and at flood (circles) and ebb (squares) phases.

ment of mixohaline water bodies over the entirewater column prevent the success of a behaviouralvertical migration to enable the estuarine popula-tions to maintain their location, as has been reportedin larger estuaries with a two-way circulation pattern(e.g. Wooldridge and Erasmus, 1980). In spite ofthis, results indicate that under low river dischargeconditions the net seaward transport of estuarinepopulations during spring tides seems slow (estuar-ine gastropod larvae apparently moved toward high-er salinity waters from one day to the next) or negli-gible (estuarine copepods did not show clear varia-tion in relation to salinity during consecutive days).However, taking into account that outgoing waterdisplaces estuarine populations to the outer estuary,their maintenance could become critical when river-ine discharges increase. In deeper estuarine systems,population abundance can be unaffected by dis-charges by migrating from the seaward surface cur-rent to the deep landward compensation current(Kaartvedt and Nordby, 1992; Kaartvedt and Svend-sen, 1995). In the shallow estuary of Mundaka thecoupled effect of strong river inflows and springtides could bring estuarine populations out of theestuary.
At neap tides, waters above 35‰ do not reachthe middle estuary even at high tide. As a result,estuarine populations remain around the low middlepart of the estuary, while the penetration of neriticforms is very poor. A previous recording (Villate,1991) on zooplankton in the outer and middle zonesof this estuary, conducted at high tides, showed thatseasonal changes in neritic populations within theestuary did not agree at all with their seasonaldynamics in neritic waters, but were related to vari-ations in tidal amplitude. The differences in theabundance of estuarine populations between springand neap tides can be linked in part to behaviouralresponses to spring-neap cycles. Although anenhancement of copepod populations under strati-fied conditions is suggested in coastal marine sys-tems (Sullivan, 1993), the tides regulated stratifica-tion in estuarine systems should have differenteffects since autochthonous copepods (mainly A.bifilosa) clearly decreased from the well-mixed con-ditions at spring tides to the highly stratified condi-tions at neap tides. In the estuary of Mundaka, shortterm changes in the A. bifilosa population have notbeen related to tidal rhythms. Conversely therelease, development and settlement of gastropodlarvae seem to follow a semilunar rhythmicity:smaller individuals are found in larger number dur-
ing spring tides in the microzooplancton fractionwhile larger individuals peak during neap tides inthe mesozooplankton fraction (Villate et al. 1993).Settlement around neap tides agrees with modelsthat predict that little settlement will occur duringperiods of stronger flow (Gross et al., 1992). Reten-tion of these larvae prior to settlement could befavored by vertical active migration in the highlystratified water column that develops at neap tides inthe middle estuary. In this study, the highest densi-ties of gastropod larvae entering the middle estuarywith the rising tide appeared at middle depth, whilethe higher densities were found deeper as the tidefell. This suggests that larvae might concentrate atthe bottom to mitigate seaward transport because ofthe upward flow of water in deeper zones. Larvalretention due to the interaction between residual cur-rents and vertical distribution of larvae is one of themore frequently observed larval transport patterns inestuaries (Lagadeuc, 1992), where the interaction oflarval transport processes with larval behavior isbetter understood in general terms (Le Fèvre andBourget, 1992). On the other hand, the clear avoid-ance of the surface layer by larval groups at neaptides and their more homogenous distribution in thewater column at spring tides corroborate that thevertical distribution of larvae is a function of verti-cal migration, which results in concentration of lar-vae at top or bottom layers, and turbulence, whichtends to produce vertically uniform concentrations(Smith and Stoner, 1993). In this sense, the skeweddistribution of estuarine populations (both copepodsand gastropod larvae) towards higher salinity atneap tides, if compared with spring tides, is account-ed for by a stronger aggregation in depth within thehighly stratified water column rather than for a netseaward displacement along the estuary.
Tychoplankton were only abundant during springtides. Although the behavioural responses of organ-isms are implied, the occurrence of benthic forms inthe water column of the estuary of Mundaka appearsmodulated mainly by spring-neap variations andvelocity asymmetries in the tidal currents, in thesame way that is described for total suspended solidconcentrations in other tidal systems (Leonard et al.,1995). If under natural conditions particle suspen-sion starts at current velocities of about 10 cm s-1
(Jonge and Bergs, 1987), this would account for thescarcity of benthic forms at neap tides. The quanti-tative importance of tychoplankton at spring tides isevidenced by the high density of benthic organismsrecorded at flood and ebb tide, which sometimes
186 F. VILLATE

was similar to that of true planktonic forms. Thiscorroborates the finding observed in other shallowsystems where meiobenthos is found to contributelargely to the mesozooplankton biomass over theentire tidal cycle (Armonies, 1989), and indicatesthat the cyclic inputs of meiofauna into the watercolumn might be relevant in the diet patterns ofpelagic planktophages and filter feeding benthicpopulations.
The occurrence patterns of benthic forms in rela-tion to the tidal currents, however, differed amongtaxa and denoted differences between passive andactively swimming organisms. The significant high-er densities of benthic foraminiferans, which wereaccompanied by high amounts of sand, during therising tide suggest a net upward transport of thesebiotic and abiotic materials within the estuary ofMundaka. In fact, low river flow spring tide periodscan lead to upstream storage of materials under tidalasymmetric conditions (Burton et al., 1995), andbenthic foraminiferal tests are used to state the nettransport of sands within tidal systems (Gao andCollins, 1995).
Moving forms such as benthic harpacticoids andostracods not only peaked at maximal flood but alsoat the mid-ebb, after strongest downstream currentsoccurred. In addition, these peaks were associatedwith waters of different salinity at flood and ebbperiods, denoting behavioural responses associatedwith tides. Active emergence is a common behav-ioural activity of moving meiofauna (Palmer, 1988;Armonies 1988 and 1989; Walters and Bell, 1994),but responses to tidal currents seem to differ amongspecies, and in relation to current intensity. Someestuarine meiobenthic harpacticoids are supposed tomigrate into the water column to be transportedwithin the estuary (Warwick and Gee, 1984). How-ever, it has also been observed that some benthicharpacticoids burrow deeper into the sediment inresponse to increasing flow to avoid suspension inthe water column (Foy and Thistle, 1991). In ourcase, although a species-level identification ofmeiobenthic forms was not made, compositionaldifferences were observed between the harpacticoidassemblage moving upstream at flood tide and thosemoving downstream at ebb tide. In addition, erosioneffects were evident in the flooding peak by theoccurrence of sand, but not so clear in the ebbingpeak. This finding indicates that passive erosionmay be important in meiobenthos resuspension,although behavioural factors, may also be important(Hicks, 1992). More detailed studies would be
required to evaluate the overall significance of thetidal suspension on meiofauna in our study area.
The higher abundance of mites around the highwater slack permits us to suppose that these organ-isms are mainly flushed to the estuary channel whenwater overflows upper intertidal areas and salt-marshes of the estuary shores. Mites are mainly dis-tributed in the littoral zone, where some supralit-toral-terrestrial invader forms are found togetherwith mites from the phytal meiofaunal communities(Somerfiel and Jeal, 1995).
In conclusion, results show that tidal hydrody-namism has strong effects on the composition andzonation of permanently and temporarily residentorganisms in the pelagic habitat of the estuary ofMundaka. That, in combination with river dis-charge, will have implications in the regulation ofplanktonic and benthic populations by translocationof individuals from one part to another within theestuary, or even outside the estuary.
ACKNOWLEDGEMENTS
This study has been supported financially by theUniversity of the Basque Country (UPV 118.310-E054/90). Many thanks to Astilleros de MuruetaS.L. for providing a place for field work.
REFERENCES
Alldredge, A.L. and W.M. Hammer. – 1980. Recurring aggregationof zooplankton by a tidal current. Estuar. Coast. Mar. Sci., 10:31-37.
Armonies, W. – 1988. Physical factors influencing active emer-gence of meiofauna from boreal intertidal sediment. Mar.Ecol.-Prog. Ser., 49: 277-286.
Armonies, W. – 1989. Meiofaunal emergence from intertidal sedi-ment measured in the field: significant contribution to noctur-nal planktonic biomass in shallow waters. Helgoländer Meere-sunt., 43: 29-43.
Armonies, W. and D. Hartke. – 1995. Floating of mud snails Hydro-bia ulvae in tidal waters of the Wadden Sea, and its implica-tions in distribution patterns. Helgoländer Meeresunt., 49: 529-538.
Boto, K.G. and J.S. Bunt. – 1981. Tidal Export of ParticulateOrganic Matter from a Northern Australian Mangrove System.Estuar. Coast. .Shelf Sci., 13: 247-255.
Burton, D.J., J.R. West, R.W. Horsington and K. Randle. – 1995.Modelling transport processes in the Ribble Estuary. Environ-ment International, 21: 131-141.
Dyer, K.R., W.K. Gong and J.E. Ong. – 1992. The Cross SectionalSalt Balance in a Tropical Estuary During A Lunar Tide and aDischarge Event. Estuar. Coast. Shelf Sci., 34: 579-591.
Fegley, S.R., B.A. MacDonald and T.R. Jacobsen. – 1992. Short-term Variation in the Quantity and Quality of Seston Availableto Benthic Suspension Feeders. Estuar. Coast. Shelf Sci.,34:393-421.
Foy, M.S. and D. Thistle. – 1991. On the vertical distribution of abenthic harpacticoid copepod: field, laboratory, and flumeresults. J. Exp. Mar. Biol. Ecol., 153: 153-163.
TIDAL EFFECTS ON ESTUARINE ZOOPLANKTON 187

Gao, S. and M. Collins. – 1995. Net Sand Transport Direction in aTidal Inlet, using Foraminiferal Tests as Natural Tracers. Estu-ar. Coast. Shelf Sci., 40: 681-697.
Gagnon, M. and G. Lacroix. – 1981. Zooplankton sample variabil-ity in a tidal estuary: An interpretative model. Limnol.Oceanogr., 26: 401-413.
Gagnon, M. and G. Lacroix. – 1982. The effects of tidal advectionand mixing on the statistical dispersion of zooplankton. J. Exp.Mar. Biol. Ecol., 56: 9-22.
Gross, T.F., F.E. Werner and J.E. Eckman. – 1992. Numerical mod-eling of larval settlement in turbulent bottom boundary layers.J. Mar. Res., 50: 611-642.
Hicks, G.R.F. – 1992. Tidal and diel fluctuations in abundance ofmeiobenthic copepods on an intertidal estuarine sandbank.Mar. Ecol.-Progr. Ser., 87: 15-21.
Jonge, V.N. de and J. van den Bergs. – 1987. Experiments on theResuspension of Estuarine Sediments Cointaining BenthicDiatoms. Estuar. Coast. Shelf Sci., 24: 725-740.
Kaartvedt, S. and E. Nordby. – 1992. Impact of a controlled fresh-water discharge on zooplankton distribution in a Norwegianfjord. J. Exp. Mar. Biol. Ecol., 162: 279-293
Kaartvedt, S. and H. Svendsen. – 1995. Effect of freshwater dis-charge, intrusions of coastal water, and bathymetry on zoo-plankton distribution in a Norwegian fjord system. J. Plank.Res., 17: 493-511.
Kennish, M.J. – 1990. Ecology of estuaries, Volume II BiologicalAspects. CRC Press, Boca Raton, Florida, 391 pp.
Langadeuc, Y. – 1992. Transport larvaire en Manche. Exemple dePectinaria koreni (Malmgren), annélide polychète, en baie deSeine. Oceanol. Acta, 15: 383-395.
Le Fèvre, J. and E. Bourget. – 1992. Hydrodynamics and Behav-iour: Transport Processes in Marine Invertebrate Larvae. TREE, 7: 288-289.
Lee, W.Y. and B.J. McAlice. – 1979. Sampling variability ofmarine zooplankton in a tidal estuary. Estuar. Coast. Shelf Sci.,8: 565-582
Leonard, L.A., A.C. Hine, M.E. Luther, R.P. Stumpf and E.E.Wright. – 1995. Sediment Transport Processes in a West-cen-tral Florida Open Marine Marsh Tidal Creek; the Role of Tidesand Extra-tropical Storms. Estuar. Coast. Shelf Sci., 41: 225-248.
MacKay, H.M. and E.H. Schumann. – 1990. Mixing and Circula-tion in the Sundays River Estuary, South Africa. Estuar. Coast.Shelf Sci., 31: 203-216.
Morales-Zamorano, L.A., R. Cajal-Medrano, E. Orellana-Cepedaand L.C. Jiménez-Pérez. – 1991. Effect of tidal dynamics on aplanktonic community in a coastal lagoon of Baja California,Mexico. Mar. Ecol.-Progr. Ser., 78: 229-239.
Palmer, M.A. – 1988. Dispersal of marine meiofauna: a review andconceptual model explaining passive transport and active emer-gence with implications for recruitment. Mar. Ecol.-Progr.Ser., 48: 81-91.
Riaux-Gobin, C. – 1987. Phytoplankton, tripton et microphytoben-thos: échanges au cours de la mareé, dans un estuaire du Nord-Finistère. Cah. Biol. Mar., 28: 159-184.
Smith, J.B. and D.M. FitzGerald. – 1994. Sediment Transport Pat-terns at the Essex River Inlet Ebb-Tidal Delta, Massachusetts,U.S.A. J. Coastal Res., 10: 752-774.
Smith, N.P. and A.W. Stoner. – 1993. Computer Simulation of Lar-val Transport Trough Tidal Channels: Role of Vertical Migra-tion. Estuar. Coast. Shelf Sci., 37: 43-58.
Somerfield, P.J. and F. Jeal. 1995. Vertical distribution and sub-stratum association of Halacaridae (Acari: Prostigmata) onsheltered and exposed Irish shores. J. Nat. Hist., 29: 909-917.
Sullivan, B.K. – 1993. How does water column structure influencecopepod populations in coastal marine systems?. B. Mar. Sci.,53: 240-246.
Ten Brinke, W.B.M. – 1994. In situ aggregate size and settlingvelocity in the Oosterschelde tidal basin (The Netherlands).Neth. J. Sea Res., 32: 23-35.
Ten Brinke, W.B.M. and J. Dronkers. – 1993. Physical and bioticaspects of fine-sediment import in the Oosterschelde tidal basin(The Netherlands). Neth. J. Sea Res., 31: 19-36.
Uncles, R.J. and Stephens, J.A. 1990 The Structure of Vertical Cur-rent Profiles in a Macrotidal, Partly-Mixed Estuary. Estuaries,13: 349-361.
Uncles, R.J., J.A. Stephens and M.L. Barton. – 1992. Observationson fine-sediment concentrations and transport in the turbiditymaximun region of an estuary, p. 255-276. In D. Prandle (ed.),Dynamics and Exchanges in Estuaries and the Coastal Zone.Proceedings of the Gregynog Conference, Wales, UK, July1990. Coastal and Estuarine Studies, AGU, Washington, D.C.
Vale, C. and B. Sundby. – 1987. Suspended Sediment Fluctuationsin the Tagus Estuary on Semi-diurnal and Fortnightly TimesScales. Estuar. Coast. Shelf Sci., 25: 495-508.
Villate, F. – 1989-90. Zooplanktoni buruzko aurreikerketa koanti-tatiboa Gernika-Mundakako itsasadarrean. Munibe, 41: 3-30.
Villate, F. – 1991. Zooplankton assemblages in the shallow tidalestuary of Mundaka (Bay of Biscay). Cah. Biol. Mar., 32: 105-119.
Villate, F., J. Franco, A. Ruiz and E. Orive. – 1989. Caracterizacióngeomorfológica e hidrológica de cinco sistemas estuáricos delPaís Vasco. Kobie, 18: 157-170.
Villate, F., A. Ruiz and J. Franco. – 1993. Summer zonation anddevelopment of zooplankton populations within a shallowmesotidal system: the estuary of Mundaka. Cah. Biol. Mar., 34:131-143.
Walters, K. and S.S. Bell. – 1994. Significance of copepod emer-gence to benthic, pelagic, and phytal linkages in a subtidal sea-grass bed. Mar. Ecol.-Prog. Ser., 108: 237-249.
Warwick, R.M. and J.M. Gee. – 1984. Community structure of estu-arine meiobenthos. Mar. Ecol. Prog. Ser., 18: 97-111.
Whitfield, A.K. – 1988. The Role of Tides in RedistributingMacrodetrital Aggregates Within the Swartvlei Estuary. Estu-aries, 11: 152-159.
Wooldridge, T. and T. Erasmus. – 1980. Utilization of tidal currentsby estuarine zooplankton. Estuar. Coast. Shelf Sci., 12: 107-114.
Scient. ed.: M. Alcaraz
188 F. VILLATE





























