Embed Size (px)
Citation preview

PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS CARRERA DE MICROBIOLOGIA INDUSTRIAL
APLICACIÓN DE LEVADURA Candida spp. COMO UNA ALTERNATIVA VIABLE PARA LA RETARDACIÓN EN LA PUDRICIÓN DEL BANANO
(Musa acuminata).
JONATHAN BUITRAGO ESTRADA ANGÉLICA MARIA ESCOBAR ROMERO
TRABAJO DE GRADO
Presentado como requisito parcial Para optar al título de
MICROBIÓLOGO INDUSTRIAL
Bogotá D. C. Enero 2009

NOTA DE ADVERTENCIA
“Los conceptos y opiniones emitidos en este trabajo son responsabilidad del autor y no comprometen en nada a la
Pontificia Universidad Javeriana”. Artículo 23 de la Resolución No 13 de julio de 1946.

APLICACIÓN DE LEVADURA Candida spp. COMO UNA ALTERNATIVA VIABLE PARA LA RETARDACIÓN EN LA PUDRICIÓN DEL BANANO
(Musa acuminata).
JONATHAN BUITRAGO ESTRADA
ANGÉLICA MARÍA ESCOBAR ROMERO
APROBADO
_________________________________________________ Gerardo Moreno
Director
___________________ __________________
Jurado 1 Jurado 2

APLICACIÓN DE LEVADURA Candida spp. COMO UNA ALTERNATIVA VIABLE PARA LA RETARDACIÓN EN LA PUDRICIÓN DEL BANANO
(Musa acuminata).
JONATHAN BUITRAGO ESTRADA
ANGÉLICA MARÍA ESCOBAR ROMERO
__________________________ _________________________ JANETH ARIAS PALACIOS INGRID SCHULER Directora Carreras de Microbiología Decana Académica facultad de ciencias

Agradecemos a nuestros padres y hermanos Alba, Luis Freddy, Johanna, María
Mercedes, José, Sindy, Duley y Gaby, por su colaboración, comprensión y apoyo
incondicional
A nuestro Director Gerardo Moreno por sus conocimientos, apoyo y entrega.
A nuestros amigos y compañeros por su colaboración y a todos los que nos han
dejado grandes enseñanzas.

TABLA DE CONTENIDO
1. Introducción
2. Marco teórico y revisión de literatura
2.1 Banano (Musa acuminata)
2.1.1 Clasificación taxonómica
2.2 Frutos climatéricos
2.2.1 Fisiología de la maduración.
2.2.2 Cambios durante la maduración.
2.2.2.1 Respiración
2.2.2.2 Producción de etileno
2.2.2.3 Enzimas pécticas
2.2.2.4 Color
2.2.2.5 Peso y tamaño
2.2.2.6 Contenido de sólidos solubles totales
2.2.2.7 Firmeza de la pulpa
2.2.2.8 pH y acidez titulable
2.2.2.9 Tasa de producción de CO2
2.2.2.10 Agua
2.2.2.11 Carbohidratos
2.2.2.12 Almidón.
2.2.2.13 Lípidos
2.2.2.14 Ácidos orgánicos

2.2.2.15 proteínas, fenoles y otros compuestos nitrogenados
2.3 Maduración comercial del banano
2.4 Levadura
2.4.1 Genero Cándida sp.
2.4.1.1 Características Generales
2.4.1.2 Fermentación de azúcares
2.4.1.3 Fisiología y crecimiento
2.4.1.4 Necesidades nutricionales
2.4.2 Cinética de Crecimiento
2.4.3 Efecto de la concentración de azúcares sobre la levadura.
2.4.4 Levaduras en frutos.
3. Formulación del problema
3.1 Justificación de la investigación
4. Objetivos
4.1 Objetivo general
4.2 objetivos específicos
5. Materiales y métodos
5.1 Ensayo preliminar y experimental: Aislamiento y recuperación de la levadura
5.2 Descripción macroscópica y microscópica de la levadura
5.3 Identificación bioquímica API 20C AUX
5.4 Prueba de patogenicidad
5.5 Determinación de biomasa por peso seco

5.6 Curva de crecimiento
5.7 Actividad Enzimática cualitativa
5.8 Crecimiento en agar banano.
5.9 Aplicación en el fruto.
5.9.1 Determinación fisiológica de peso PFP
5.9.2 Determinación de pH
5.9.3 Determinación de Grados Brix
6. Análisis Estadístico
7. Resultados y discusión
8. Conclusiones
9. Recomendaciones
10. Referencias
11. Anexos

LISTA DE TABLAS
1. Composición aproximada de un banano maduro.
2. Caracteres utilizados en la clasificación taxonómica del banano.
3. Número de bananos para cada día de estudio.
4. Resultados de asimilación de azucares por Candida sp.
5. Datos obtenidos a partir de curva de crecimiento C. guillermondii.
6. Resultados de la probabilidad por prueba T para SST con un intervalo de
confianza del 95%.
7. Promedio de pesos de las replicas de los frutos control a través de los días.
8. Promedio de pesos de las replicas de los frutos con aplicación de la
concentración 1.
9. Promedio de pesos de las replicas de los frutos con aplicación de la
concentración 2.

LISTA DE FIGURAS
1. Estadios en el desarrollo y maduración de frutos que experimentan incremento climatérico.
2. Curva de crecimiento típica de una levadura.
3. Clasificación de bananos para posterior aplicación. 4. Bananos después de aplicación. 4. A. Macerado de banano. 4. B. Refractómetro portátil 6. Características microscópicas de Candida sp.
7. Características macroscópicas de Candida sp.
8. Resultado programa informático Api web TM API 20 AUX.
9. Crecimiento de C. guillermondii en agar banano.
10. Curva de peso seco de C. guillermondii.
11. Curva de crecimiento de C. guillermondii.
12. Fase exponencial de C. guillermondii.
13. Comportamiento del pH durante la curva de crecimiento.
14. Ln de la concentración de C. guillermondii en función del tiempo, para determinar tiempo de duplicación y velocidad especifica de crecimiento.
15. Comportamiento del pH durante la maduración del fruto.
16. comportamiento de los SST en tratamientos y control.
17. Efecto de la levadura a concentración 1 sobre el fruto a los 18 días de estudio.
18. Perdida de agua y firmeza en el fruto.
19. Perdida lineal de los frutos control a través del tiempo.
20. Perdida lineal de los frutos con aplicación de la concentración 1 a través del tiempo.

21. Perdida lineal de los frutos con aplicación de la concentración 2 a través del tiempo.
22. Relación de pérdida lineal de peso, entre los frutos control y las concentraciones. 23. Porcentaje de pérdida de peso de los frutos a través del tiempo.
24. Estado de maduración del fruto control frente al fruto con levadura.

LISTA DE ANEXOS
1. Taxonomía, clasificación y nomenclatura de los bananos cultivados en
Colombia
2. Guía de Color del banano (Musa acuminata)
3. Sistema de identificación de levaduras API 20C AUX.
4. Resultados API 20 C AUX para Levadura.
5. Fotografía Confocal de Candida spp.

RESUMEN
El proceso de pudrición de frutos como el banano, ha llevado a la Industria bananera
a pérdidas económicas demasiado altas, ya que el proceso de maduración en este
fruto se ve afectado por la temperatura, la humedad, el transporte y el
almacenamiento.
En este estudio se evaluó el efecto de Candida spp como una alternativa viable para
retardar la pudrición que presenta el fruto del banano, la levadura fue aislada de frutos
de uchuva e identificada, se determinó el comportamiento en cuanto cinética de
crecimiento de la cual se determinaron dos concentraciones dentro de su fase
exponencial para ser aplicadas en los frutos y comparadas contra un control el cual no
fue tratado con levadura. El Estudio se llevó a cabo por 45 días, durante este tiempo
se determinaron variables tales como Sólidos solubles totales (grados Brix), pH,
pérdida de peso a través de los días, además de observar los cambios organolépticos
del fruto.
Los datos obtenidos se analizaron estadísticamente para conocer su comportamiento y
a la vez determinar la concentración óptima de aplicación que tenga efecto de
retardación sobre el fruto del banano.

1. INTRODUCCIÓN
Desde hace algunos años, entre las especies cultivadas de mayor trascendencia
económica en el mundo se encuentra el banano, teniendo un gran potencial en los
mercados de Norteamérica, Europa y últimamente en el continente asiático, siendo
los principales destinos de exportación en los últimos años Bélgica 37%, Estados
Unidos 36% y Alemania 11% según datos reportados por Bancoldex. La importancia
de este cultivo está en el hecho de que forma parte fundamental en la dieta de los
habitantes de los países productores y países importadores, al suministrar gran parte
de calorías. Las exportaciones de banano han registrado un incremento por la
creciente demanda en el exterior, generando ingresos cercanos a los 242 millones de
dólares anuales, según datos reportados por el DANE.
El cultivo de este fruto en Colombia es de gran importancia socio económica por ser
un producto fundamental de la canasta familiar, convirtiéndolo en una fuente de
ingreso de divisas para el país, a la vez que permite el ingreso a nuevos mercados. En
el cultivo y cosecha del banano se deben tener cuidados especiales su manejo, ya que
al ser un fruto climatérico, su rápida maduración juega un papel fundamental en la
obtención de un producto de calidad, es así como este factor natural del fruto se
convierte en un problema para los comercializadores de banano, provocando pérdidas
postcosecha del producto, afectando la economía del sector bananero.
Es necesario conocer los procesos fisiológicos de la maduración para el mejoramiento
de la calidad y la prevención de grandes pérdidas, durante la producción,
almacenamiento, industrialización y mercadeo de los frutos, buscando nuevas
alternativas para controlar este proceso que ha medida del tiempo conlleva a la
pudrición del fruto.
Este estudio forma parte de un proyecto integrado en el que se han evaluado las
características, propiedades, forma de conservación, antagonismo frente a

microorganismos del medio ambiente y fitopatógenos y su respuesta a aplicaciones en
campo de la Levadura Candida spp.
A partir del año 2000 se han realizado estudios que demuestran el efecto de Candida
spp. sobre la retardación en la pudrición de frutos. Por tal razón este estudio pretende
demostrar que la utilización de levadura representa una posibilidad para encontrar el
camino hacia la obtención de un producto con una vida útil más prolongada.

2. MARCO TEORICO
2.1 EL BANANO
El banano es una fruta de agradable sabor, alto contenido de vitaminas y minerales y
fácil digestión. Por cada 100 gramos de banano se tienen 460 calorías. Como pocas
frutas el banano permanece aséptico dentro de su envoltura natural, jamás tiene
gusanos, ni corazón, ni semillas. Es un alimento altamente energético, con hidratos de
carbono fácilmente asimilables, pero pobre en proteínas y lípidos. (Sierra, L. 1993)
Tabla 1. Composición aproximada de un banano maduro.
Humedad Azúcares totales
Dextrosa Levulosa Sucrosa Almidón
Fibra cruda Proteína Grasa
Cenizas
74.8 % 21.2
4.8 % 3.7 12.7 1.2 0.6 1.2 0.2 0.8
Vitaminas en 100 gramos
A Tiamina (B1)
Riboflavina (B2) Acido Ascórbico Piroxidina (B6)
430.0 UI 0.1 Mgr
0.05 Mgr 10.0 Mrg 0.52 Mgr
Tomado de: (Sierra, L. 1993).
El banano posee alrededor del 75% de agua. Mas o menos la cuarta parte corresponde
a hidratos de carbono. No engorda porque la cantidad de azúcar pasa rápidamente a la
sangre. El banano maduro contiene ciertos componentes químicos muy importantes:
serotonina, dopamina y la norafetamina. La serotonina ayuda en el retraso mental y
en las depresiones nerviosas, aumenta la actividad locomotora y reduce la apatía y los
retraimientos. El banano es uno de los alimentos favoritos dentro de las nuevas

tendencias de consumo en los países desarrollados que se inclinan por productos
frescos y saludables. (Sierra, L. 1993)
El banano ofrece enormes posibilidades para la elaboración de productos
transformados e industrializados, en forma de bananos pasos o deshidratados, cremas,
pastas, pulpas, compotas, mermeladas en conserva, frutas confitadas y congeladas,
alimento para ganado, abonos a base de desechos orgánicos, fibras y papel a base de
pseudotallos, alcohol, vinagre de la fermentación de la fruta, etc. (Sierra, L. 1993)
2.1.1 CLASIFICACIÓN TAXONÓMICA
Todas las especies de bananos y plátanos comerciales pertenecen a la familia
musácea. La familia musácea se encuentra subdividida en tres subfamilias:
Musoideae, Strelitroideae y Heliconoideae. La subfamilia Musoideae se divide en los
géneros: Ensete y Musa. El primero agrupa a las hierbas monocarpicas con numero
cromosómico de nueve y no producen frutos comestibles, el segundo, el genero Musa
está constituido por cuatro secciones: Australimusa, Callimusa, Rhodochlamys y
Eumusa (ANEXO 1). La sección Eumusa es la de mayor importancia económica, ya
que la mayor parte de los cultivares de banano y plátano son producto de esta
evolución del género Musa. sus cultivares tuvieron su origen es dos especies
silvestres, Musa acuminata y Musa balbisiana, dando origen por poliploidía e
hibridación a los plátanos y bananos cultivados. (Sierra, L. 1993)
Musa acuminata es una planta de porte bajo, con pseudotallos delgados y un sistema
foliar reducido. Presentan una coloración parduzca que se convierte en manchas
heterogéneas claramente definidas en las láminas de las hojas y una coloración vino
uniforme en las vainas internas del pseudotallo. Los racimos crecen horizontalmente,
son pequeños con dedos muy delgados que producen semillas. (Sierra, L. 1993)
Tabla 2. Caracteres utilizados en la clasificación taxonómica de banano.(Musa
acuminata)

Caracteres Musa acuminata
Color del pseudotallo
Canal peciolar
Más o menso densamente marcado con manchas de color pardo o negro Margen erecto o dilatado con alas escoraseas por debajo, sin abrazar el psudotallo.
Pedúnculo Pedicelos
Por lo general pubescente o piloso Cortos
Rudimentos seminales Dos hileras regulares en cada lóculo Hombro de bráctea
Enrollamiento de la bráctea Por lo general alto La bráctea se repliega y enrolla hacia atrás después de abrirse
Forma de la bráctea
Ápice la bráctea
Lanceolada o estrechamente aovada aguzándose abruptamente a partir del hombro Agudo
Color de la bráctea
Atenuamiento del color
Rojo, purpura mato o amarillo, en la parte exterior rosado, purpura mate o amarillo dentro. El color interno de la bráctea se atenúa hasta llegar al amarillo en dirección a la base.
Sépalo libre de la flor
Color de la flor Color del estigma
Corrugado en forma variable, por debajo de la punta. Banano cremoso Anaranjado o amarillo intenso
Tomado de Sierra, L. 1993
2.2 FRUTOS CLIMATÉRICOS
Los cambios en la respiración de las frutas a través del tiempo desde su crecimiento
hasta su senescencia marcan la característica de los dos tipos de frutos los llamados
climatéricos y no climatéricos. Los frutos climatéricos durante la división celular
tiene una actividad respiratoria muy alta, cuando termina esta etapa, la fruta pasa a un
estado de maduración, cuando esta etapa esta terminando se inicia dentro ella la
maduración final (organoléptica o sensorial) que coincide con un aumento en la

actividad respiratoria hasta completar la maduración, la que disminuye con el
envejecimiento del fruto, este incremento en la respiración, se denomina pico
climatérico o sencillamente climaterio. En los productos climatéricos como respuesta
a diferentes concentraciones ascendentes de (C2H4) se activa prematuramente la
maduración. Los productos climatéricos requieren de un manejo especial con el fin de
evitar que el climaterio se active y lo lleve a la senescencia rápidamente perdiendo
gran parte o la totalidad de su valor comercial. (Gallo, F. 1997)
2.2.1 Fisiología de la maduración.
La maduración es un proceso fisiológico que ocurre en un periodo de tiempo como
parte del crecimiento y desarrollo de una fruta, es una secuencia de hechos naturales.
Se produce sin que haya en muchos casos, un crecimiento en tamaño; es una
transformación interna de la fruta, que constituye uno de los más extraordinarios
fenómenos fisiológicos. El fruto se transforma totalmente en pocos días, pasando de
ser simple, sin atractivo, no comestible a uno atractivo, de un alto sabor, aroma y
deseable para ser consumido. Siendo la maduración un proceso de cambio
irreversible que proporciona las características óptimas para el consumo de una fruta.
(Cayón, D. et al. 2000).
Los procesos fisiológicos de la maduración ocurren a nivel celular y cuando terminan
las transformaciones se inician los procesos de degradación o desintegración de
sustancias como la clorofila, aromas, sabores, etc. y organelos iniciando por los
ribosomas y plastos, terminando con el núcleo y el plasmalema causando finalmente
la muerte de la célula. Estas últimas etapas de la maduración son los periodos de la
desorganización de tejidos o senescencia y la destrucción final. (Gallo, F. 1997). La
senescencia es la fase final de la vida útil de un órgano en el cual se presentan
alteraciones irreversibles que conducen al desorden y muertes celulares por el
aumento de la actividad de enzimas hidrolíticas. (Cayón, D. et al. 2000).

Después de cosechados, los frutos climatéricos como el banano (Musa acuminata)
pasan por cuatro estadios de desarrollo fisiológico: preclimaterio, climaterio,
maduración, maduración de consumo y senescencia. El preclimaterio representa el
periodo desde la cosecha hasta la iniciación de la respiración climatérica; durante esta
fase los frutos son verdes, de textura rígida y la actividad metabólica es baja, siendo
el objetivo comercial prolongar al máximo esta fase. El climaterio se caracteriza por
un incremento rápido en la respiración denominado “respiración climatérico” que
generalmente ocurre cuando se completa el del proceso de maduración del fruto. El
máximo climaterio puede ocurrir antes o después que el fruto es removido de la
planta, dependiendo del tipo del fruto y los procedimientos de cosecha. Durante el
estadio de maduración el cambio más notable es la pérdida paulatina del color verde y
amarillento de la cáscara como resultado de la degradación de la clorofila,
permitiendo que la pigmentación debida a los carotenos y xantofilas se torne visible;
al mismo tiempo, la pulpa comienza a ablandarse y el almidón es convertido
rápidamente a sacarosa, glucosa y fructosa. (Cayón, D. et al. 2000).
2.2.2 Cambios durante la maduración.
2.2.2.1 Respiración En todos los frutos, la tasa respiratoria es elevada cuando son
jóvenes mientras las células aun se dividen y crecen con rapidez. Después la tasa
declina de manera gradual, incluso si el fruto se corta. Sin embargo, en muchas
especies como el banano la disminución gradual en la respiración se revierte en un
marcado incremento conocido como climatérico (Figura 1), este suele coincidir con la
maduración plena y la adquisición del sabor del fruto que se acelera por la producción
celular de etileno que estimula la maduración. Una acumulación adicional provoca
senescencia y disminución en la respiración. (Salisbury, F. Ross, C., 1994)

Tomado de Salisbury y Ross, 1994
Figura 1. Estadios en el desarrollo y maduración de frutos que experimentan
incremento climatérico en la respiración y de frutos que no la experimentan.
Los bananos al pertenecer al grupo de las frutas climatéricas, siguen madurando
después de la cosecha. En fruta inmadura de tipo climatérico, la aplicación de un
tratamiento con etileno exógeno acelera el comienzo de los cambios de maduración,
sin alterar, generalmente, el patrón de los cambios de respiración; una vez que ha
entrado en esta fase y se ha iniciado la elevación climatérica, la maduración es un
proceso irreversible que puede ser atrasado pero no detenido por factores externos. El
banano es una fruta que muestra aumentos en la respiración y producción de etileno
al comienzo de la maduración. La respiración se incrementa como consecuencia de
los daños mecánicos y del estado de sanidad del banano; para que la respiración no se
acelere el producto debe colocarse en un lugar fresco, sin corrientes fuertes de aire,
evitando el exceso de manipulación y los daños causados por éste. (Ovalle, J. Et.al.
1999).
La respiración es un proceso global que consiste en una oxido-reducción en la que
algunos compuestos se oxidan a CO2 y el oxigeno que se absorbe se reduce para

formar agua. El almidón, sacarosa, azucares simples, grasas y ácidos orgánicos
pueden servir como sustratos respiratorios. La respiración es una serie de 50
reacciones o más que se complementan entre sí, cada una catalizada por una enzima
distinta. Es una oxidación que se efectúa en un medio acuoso, a un pH casi neutro y a
temperatura moderada. Esta degradación paulatina de moléculas grandes es un medio
para convertir energía en ATP. Además a medida que se efectúa la degradación se
obtienen intermediarios útiles para un gran número de otros productos vegetales
esenciales. Entre estos productos se incluyen aminoácidos para proteínas, nucleótidos
para ácidos nucleicos y precursores carbonados para pigmentos porfirínicos (clorofila
y citocromos), así como otros compuestos aromáticos (lignina). (Salisbury, F. Ross,
C., 1994).
2.2.2.2 Producción de etileno Además de mostrar un incremento en la respiración,
la fruta climatérica produce también etileno, que aumenta durante la maduración. El
etileno ha sido identificado como una hormona natural en las plantas; ejerce mayor
influencia sobre los aspectos del crecimiento, el desarrollo y la senescencia,
incluyendo la iniciación de la maduración. El banano emite etileno entre 0.1 y 1.0 µl
C2H4 Kg h (microlitros de etileno hora por cada kilo de fruta). Su sensibilidad por
este gas es grande, por lo que en contacto con algunas frutas que lo produzcan, como
maracuyá, chirimoya o uchuva, se acelera su maduración. (Ovalle, J. Et.al. 1999).
Los tejidos vegetales sintetizan etileno a través de una ruta metabólica que utiliza
como precursor el aminoácido metionina. El etileno difiere de todas las fitohormonas
conocidas en su extrema volatilidad, característica gaseosa, que permite que el etileno
producido por una planta o fruto influya sobre el crecimiento y desarrollo de otros
cercanos. (Cayón, D. Et al. 2000). La sensibilidad de los frutos al etileno se reduce
durante el almacenamiento a bajas temperaturas, con el incremento de los niveles de
CO2 (inhibiendo la respiración) o disminuyendo la presión parcial de oxigeno que
inhibe la respiración y la síntesis de etileno. (Gallo, F. 1997)

2.2.2.3 Enzimas pécticas Juegan un papel importante en los cambios de la textura
durante el proceso de maduración. El ablandamiento se ha atribuido a la
solubilización de pectina por efecto de enzimas pécticas como la poligalacturonasa
(PG) (E.C.3.2.1.1.11), que cataliza la hidrólisis de los enlaces glicosidicos α1-4 de las
sustancias pécticas generando una disminución de la firmeza de los tejidos. Así
mismo la pectinestereasa (E.C.3.1.1.11) hidroliza los grupos metil éster del ácido
pectico y de la pectina produciendo grupos carboxilo libres. (Gallo, F. 1997)
La actividad de la PG se ha cuantificado mediante la formación de grupos reductores
por los métodos de Samogy-Nelson (1944), Luschinger and Cornesky (1962),
también se utiliza el método de Gross et. al., 1982, el cual es más fácil y sensible que
los otros métodos debido a que cuantifica nanomoles de azúcares reductores. (Gallo,
F. 1997)
2.2.2.4 Color Es el cambio más notorio en muchas frutas durante su maduración y
con frecuencia es el criterio más utilizado para decidir sobre la madurez de esta. La
transformación más importante es la degradación del color verde. La perdida de color
verde es consecuencia de la degradación de la clorofila se debe a uno o varios
procesos secuenciales, los más relevantes son el cambio de pH, los procesos
oxidativos y la acción de las clorofilazas. Esta desaparición de color verde esta
asociado con la síntesis o el desenmascaramiento de pigmentos cuyos colores oscilan
entre amarillo y el rojo. Los pigmentos responsables son los carotenoides,
hidrocarburos no saturados de cuarenta carbonos generalmente y cuyas moléculas
pueden contener una o mas funciones oxigenadas, las antocianinas que son
hidrosolubles, producen colores muy fuertes que pueden en ocasiones enmascarar a
los carotenoides y a la clorofila. (Gallo, F. 1997). Los cambios de color en el banano
son debidos a una degradación de la clorofila. Con la maduración a temperatura alta
(arriba de 27°C) no hay degradación de la clorofila; lo cual resulta en fruta madura,
pero con cáscara verde. (Ovalle, J. Et.al. 1999).

A medida que transcurre la maduración del fruto, se sintetizan los azucares totales
encontrándose muy bajos contenidos en frutos en estado verde y valores alrededor del
35 % en frutos maduros la síntesis de azúcares es lenta después de cosechado el fruto
y hasta que este alcanza el estado verde claro, correspondiente a la fase de
preclimaterio; el aumento en azucares ocurre cuando el fruto empieza a tornarse
amarillo acelerándose la síntesis de azucares hasta alcanzar la coloración totalmente
amarilla. (Cayón, D. Et al. 2000).
2.2.2.5 Peso y tamaño La propiedades físicas del fruto cambian durante el proceso
natural de maduración disminuyendo significativamente el peso promedio y la
humedad del fruto, el peso de la pulpa aumenta y el de la cáscara disminuye,
incrementándose la relación pulpa-cáscara. La reducción del peso fresco es típica del
comportamiento metabólico de los frutos durante la postcosecha, siendo más
pronunciada en los frutos que se desarrollan en las zonas cálidas y secas. (Cayón, D.
Et al. 2000).
2.2.2.6 Contenido de sólidos solubles totales. Durante la maduración de los
bananos, el contenido de sólidos solubles totales aumenta. En algunos híbridos, el
contenido aumenta hasta un pico y luego disminuye (la caída puede deberse a la
conversión del azúcar de la pulpa en alcohol), mientras que en otros, los sólidos
solubles continúan su aumento con la maduración. (Chang-Yuen, Et.al 2005)
2.2.2.7 Firmeza de la pulpa. La fruta crujiente, dura, y verde se convierte en una
fruta amarilla, con pulpa tierna y suave en la etapa óptima de madurez, y se torna
blanda a medida que avanza hacia la senescencia. La firmeza de la pulpa a menudo
está relacionada con la maduración, implicando que al progresar la maduración, la
firmeza de la pulpa disminuye. La pérdida de firmeza de la pulpa se asocia a 3
procesos: 1. Degradación del almidón para formar azúcar; 2. Degradación de las
paredes celulares o reducción en la cohesión de la lamela media debido a la
solubilización de las sustancias pécticas; 3. El movimiento del agua desde la cáscara
hacia la pulpa debido al proceso de ósmosis. (Chang-Yuen, Et.al 2005)

2.2.2.8 pH y acidez titulable. En la mayoría de los híbridos y cultivares existe una
rápida disminución del pH de la pulpa, en respuesta a un aumento en la madurez (la
magnitud de la disminución depende del cultivar). La acidez titulable de los tejidos de
la pulpa, de la mayoría de los cultivares e híbridos de banano, muestra grandes
aumentos durante la maduración. (Chang-Yuen, Et.al 2005)
2.2.2.9 Tasa de producción de CO2. Durante la maduración de los bananos existe un
gran aumento en la cantidad del etileno producido, usualmente acompañado por un
aumento en la tasa de respiración de la fruta (climaterio). El control del proceso de
maduración de frutos climatéricos es uno de los aspectos clave para la
comercialización internacional de este tipo de fruta; esto se debe a que el proceso está
íntimamente relacionado con la capacidad de esa fruta para soportar períodos de
transporte y almacenamiento prolongados hasta alcanzar los mercados de destino.
(Chang-Yuen, Et.al 2005)
2.2.2.10 Agua El agua es el mayor componente del fruto confiriéndole fragilidad a
los tejidos, la pulpa esta compuesta esencialmente de agua y carbohidratos, mientras
que los contenidos de grasas y proteínas son bajos. Las condiciones del trópico
favorecen la perdida rápida de agua de los frutos de plátano y se ha demostrado que
esta perdida de agua acelera la maduración, reduciendo la duración de la vida verde
pre climatérica del fruto, además de las condiciones ambientales, los cambios físicos
a los frutos también influyen en la maduración. El fruto verde, inmediatamente
después de cortado, muestra una baja intensidad transpiratoria que luego se estabiliza
a un nivel continuo que depende de la temperatura y la humedad: en el climaterio
ocurre un drástico aumento de la transpiración, a medida que el fruto madura se
mantiene un nuevo estado constante mayor que el de el preclimaterio y, finalmente se
registra una caída en pérdida de agua. El contenido de humedad de la cáscara
disminuye durante la maduración mientras en la pulpa aumenta, el porcentaje de agua
en la pulpa aumenta durante la maduración, no solo por la hidrólisis del almidón sino
también por el movimiento osmótico de agua desde la cáscara hacia la pulpa. Un
incremento en el peso de la pulpa afectando la relación pulpa: cáscara, parece ser una

consecuencia de la salida osmótica de la humedad de la cáscara. Una diferencia
marcada de la presión osmótica entre la pulpa: cáscara se desarrolla durante la
maduración ya que la concentración de azucares se incrementa más rápidamente en la
pulpa que en la cáscara. La cáscara del plátano aunque no es usada para la
alimentación humana ejerce una función reguladora entre la pulpa comestible y las
condiciones ambientales externas, además de ser particularmente importante para
proteger la pulpa contra daños físicos y desecación. (Cayón, D. Et al. 2000).
2.2.2.11 Carbohidratos presentan el cambio más importante de los frutos
climatéricos, el almidón es convertido casi en su totalidad en azúcares. Esta
transformación altera el sabor, la textura y consistencia del fruto; haciéndolo mas
dulce y con mayor aceptabilidad. La degradación de sustratos poliméricos,
especialmente de las sustancias pécticas y hemicelulosa, debilita las paredes celulares
y las fuerzas cohesivas que mantienen las células unidas. Las sustancias pécticas
proceden de un precursor insoluble en agua llamado protopectina; es un
macropolimero; ligado por enlaces cruzados con otros polímeros, a través de puentes
de calcio, unido con otros azucares y otros derivados fosforilados de los mismos
dando una molécula de tamaño excesivo. Durante la maduración la protopectina va
degradándose a fracciones de peso molecular mas bajo, las cuales son más solubles
en el agua; la velocidad de esta degradación y de las sustancias pécticas induce la
velocidad de ablandamiento de la fruta. (Gallo, F. 1997).
La sacarosa necesaria para la síntesis del almidón en la pulpa de los frutos en
formación provienen, en su mayoría, de la fotosíntesis foliar y muy poco de la
fotosíntesis realizada por la cáscara, ya que la actividad fotosintética de las hojas del
plátano es mayor a la de la cáscara de los frutos en formación como consecuencia de
la muy baja densidad estomática de la cáscara comparada con la de las hojas. (Cayón,
D. et al. 2000).
2.2.2.12 Almidón. Es el carbohidrato predominante en el fruto del banano, cuyo
porcentaje es del 48% de la materia seca o del 12.7 % del peso fresco. La pulpa fresca

del fruto verde contiene 62.4% de agua, 23.3% de almidón y 2.1% de glucosa, los
tejidos de la cáscara contienen almidón alrededor del 3% del peso fresco. Los frutos
acumulan almidón durante su desarrollo y se transforman o degradan en azucares mas
sencillos antes o durante la maduración por acción de las enzimas. (Cayón, D. et al.
2000).
La mayoría de los pasos de la degradación de almidón a glucosa pueden ser
catalizados por tres enzimas distintas, si bien hay otras más que se necesitan para
completar el proceso. Las primeras tres enzimas son-. Una alfa amilasa, una beta
amilasa y la almidón fosforilasa. De estas al parecer solo la alfa amilasa puede atacar
gránulos de almidón intactos, por lo que cuando participa la beta amilasa y la almidón
fosforilasa, es probable que actúe sobre los primeros productos liberados por alfa
amilasa. La alfa amilasa ataca de manera aleatoria enlaces 1-4 en las moléculas de
amilosa y amilopectina, al principio creando huecos al azar en los gránulos de
almidón y liberando productos que aun son grandes. En cadenas de amilosa no
ramificadas, el ataque repetido por la alfa amilasa produce maltosa, un disacárido que
contiene dos unidades de glucosa. Sin embargo la alfa amilasa no puede atacar los
enlaces 1-6 localizados en los puntos de ramificación de la amilopectina, por lo que la
digestión de amilopectina cesa cuando aun quedan dextrinas ramificadas con cadena
de longitud corta. Muchas alfa amilasas son activadas por Calcio siendo este un
elemento esencial. (Salisbury, F.; Ross, C.; 1994).
La beta amilasa hidroliza el almidón en beta maltosa; la enzima actúa primero sobre
los extremos no reductores. La beta maltosa cambia con rapidez, por muta rotación,
para formar las mezclas naturales de isómeros alfa y beta. La hidrólisis de amilosa
por la beta amilasa es casi completa, pero la degradación de amilopectina es
incompleta porque no son atacados los enlaces de los puntos de ramificación
quedando nuevamente dextrinas ramificadas. La actividad de ambas amilasas implica
la incorporación de una molécula de agua por cada enlace roto por lo que son enzimas
hidrolasas, las reacciones hidrolíticas no son reversibles, de modo que no se puede
detectar síntesis de almidón por amilasas. La almidón fosforilasa degrada el almidón

empezando por un extremo no reductor, esta degradación no se realiza incorporando
agua en los productos como lo hacen las amilasas, si no incorporando fosfatos.
(Salisbury, F.; Ross, C.; 1994).
La amilopectina solo es degradada parcialmente por la acción de la almidón
fosforilasa. La reacción procede de manera consecutiva a partir del extremo no
reductor de cada cadena principal o cadena ramificada, hasta unos pocos residuos de
glucosa de las uniones alfa 1-6 de las ramificaciones. Las uniones 1-6 de las
ramificaciones de la amilopectina o las dextrinas ramificadas que no son atacadas por
ninguna de las enzimas mencionadas, son hidrolizadas por diversas enzimas
desramificantes. Las plantas contienen tres tipos principales dependiendo de los tipos
de polisacáridos que atacan: una pululanasa, una isoamilasa y una dextrinasa limite.
La acción de estas enzimas sobre cadenas de almidón ramificadas, generan grupos
terminales adicionales sobre los que actúan la amilasa o la almidón fosforilasa.
(Salisbury, F. Ross, C., 1994).
2.2.2.13 Lípidos El contenido de lípidos en la pulpa de los frutos de banano es bajo y
permanecen constantes durante el proceso de maduración. El 45% de los ácidos
grasos de los lípidos son saturados, siendo el ácido linolénico el principal
componente. La concentración total de lípidos en la pulpa fresca es de 0.2 a 0.5 % y
en la cáscara fresca de 1.0 %. (Cayón, D. Et al. 2000).
2.2.2.14 Ácidos orgánicos Los ácidos orgánicos son esenciales para el
mantenimiento del balance azúcar: ácido que confiere a los frutos del plátano y
banano un sabor agradable durante la maduración y pueden ser considerados como
una reserva energética de los frutos ya que normalmente son degradados y
convertidos a azucares durante la maduración. Se sabe que la mayoría de frutos tienen
concentraciones elevadas de ácidos orgánicos relacionados con el ciclo de los ácidos
tricarboxilicos (ciclo de krebs) y otras rutas metabólicas, cuyo exceso suele
almacenarse en las vacuolas. (Cayón, D. Et al. 2000).

Los principales ácidos orgánicos en la pulpa del fruto son: málico, cítrico y oxálico,
cuyos niveles se incrementan durante la maduración. En el fruto verde la
concentración de ácido oxálico es mayor, mientras que en el fruto maduro es el ácido
málico. La pulpa del fruto contiene en estado verde 0.7% de ácido málico y 1.5% en
estado maduro. La cáscara en estado verde contiene 1.0% de ácido málico y 1.4% en
estado maduro. (Cayón, D. Et al. 2000).
2.2.2.15 Proteínas, fenoles y otros compuestos nitrogenados Durante la
maduración se producen cambios el la actividad enzimática, que alteran las
estructuras subcelulares provocando cambios en la actividad mitocondrial,
desintegración interna de las mitocondrias y perdida de la eficiencia respiratoria, la
cantidad total de compuestos nitrogenados permanece constante una vez que el fruto
es arrancado de la planta, pero se presenta un ligero incremento neto en la proteína
durante la maduración de algunos frutos. Algunos cambios en las rutas metabólicas
observadas durante la maduración, entre ellos el aumento de la actividad de la enzima
málica y la carboxilasa pirúvica, pueden explicar el aumento climatérico de la
producción de CO2 que ocurre en los frutos del plátano.
La pulpa de plátano, como muchos otros frutos, es susceptible al pardeamiento
cuando es cortado o tajado, fenómeno directamente relacionado con niveles de ácido
ascórbico, contenido de polifenoles, actividad de la enzima polifenol oxidasa (PPO) o
a una combinación de estos factores, el grado de pardeamiento de la pulpa esta
correlacionado positivamente con la concentración total de polifenoles y la actividad
de la PPO. (Cayón, D. Et al. 2000).
Durante el periodo climatérico puede producirse un descenso en la cantidad de los
aminoácidos libres, atribuible a la síntesis proteica y durante la senescencia se
presenta un incremento en el contenido de los aminoácidos libres, consecuencia de la
degradación de enzimas. (Gallo, F. 1997).
2.3 MADURACIÓN COMERCIAL DEL BANANO

El inicio de la maduración organoléptica natural en los frutos climatéricos viene
acompañado de un incremento en la producción de etileno (C2H4); el tratamiento con
etileno exógeno en el período preclimatérico acelera el comienzo del proceso de
maduración. En la práctica comercial se aprovecha esta respuesta al etileno exógeno
para controlar la maduración del banano.
En los depósitos de almacenamiento se puede acumular el etileno generado por otros
frutos o el que provenga de otras fuentes, y puede alcanzar niveles que desencadenen
un proceso de maduración no deseado en otras frutas ¨verdes¨ presentes en ese mismo
depósito. Hay etapas de la cadena comercial en que es bastante frecuente que se
almacenen juntas diversas frutas y hortalizas; en tales condiciones, el etileno
desprendido por unas puede afectar negativamente a otras.
El banano suele cosecharse antes de que haya adquirido su grado óptimo de fruta
comestible (banano hecho verde); en ese estado se transporta a distancias
considerables hasta las áreas de consumo; donde se deja madurar bajo condiciones
controladas de temperatura y humedad relativa y por acción del etileno (tanto natural
como exógeno); en algunos casos, se da la acción del acetileno exógeno (C2H2),
liberado del carburo de calcio CaC2 en contacto con el agua. (Ovalle, J. Et.al. 1999).
El banano madura, generalmente a una temperatura de 18 a 21 °C, con una humedad
relativa de 90% a 95% y en 1000 ppm de etileno durante 24 horas; al cabo de 5 días,
se obtienen esas condiciones un banano en estado 5. (Anexo 2 Tomado de Turbana).
2.4 LEVADURA
Las levaduras son un grupo de organismos unicelulares, muchas de las cuales son
dimorfitas, es decir que son capaces de cambiar desde una fase de crecimiento
unicelular a una fase micelial. Sus células son ovales o cilíndricas las cuales se
reproducen de manera asexual por gemación; este es un proceso en el cual se forma
una pequeña protuberancia (yema) que con el tiempo se separa de la célula

progenitora (Port 1990). Las levadura también se reproducen de manera asexual por
fisión y en forma sexual por medio de un proceso llamado apareamiento
(combinación), en el cual dos células se fusionan, dando origen a un cigoto dentro del
cual se producen ascosporas; las células de levadura son mucho más grandes que las
bacterianas y pueden distinguirse no solo por su tamaño sino por la presencia obvia
de elementos intracelulares tales como el núcleo. Además, la célula de levadura
contiene citoplasma, pared celular, membrana citoplasmática y se encuentra rodeada
por una capa de celulosa. El núcleo, no presenta membrana de separación por lo
tanto se encuentra incluido en el citoplasma, en este último puede haber una o más
vacuolas, las cuales son especies de bolsas con materiales de reserva (azúcares,
grasas, etc.) o con productos de desecho del metabolismo celular (Biely 1987). La
membrana citoplasmática es semipermeable, dejando pasar los elementos nutritivos
que necesita la célula y permitiendo la salida de desechos de la misma. (Vargas,L
2002)
La mayoría de levaduras crecen mejor en medios en los que se dispone de agua en
gran cantidad, y cabe anotar que casi todas requieren más agua que los hongos. El Aw
de las levaduras normales necesaria para su crecimiento se encuentra entre 0.88 y
0.94 (Stewart & Russel 1991); sin embargo, las levaduras osmofílicas, es decir las
que crecen en concentraciones de soluto altas, crecen escasamente en medios con Aw
más baja que 0.78, por lo tanto, cada levadura tiene un Aw óptimo y un intervalo de
Aw para su crecimiento; los valores de Aw varían cuando lo hacen las propiedades
nutritivas del sustrato, pH, temperatura, disponibilidad de oxígeno y presencia de
sustancias inhibitorias. El intervalo de temperaturas de crecimiento de las levaduras,
es en general semejante al de los hongos, con un óptimo alrededor de 25°C a 30°C y
un máximo de aproximadamente 35-47°C (Port 1990). El crecimiento de la mayoría
de levaduras se favorece con un pH ácido próximo a 4-4.5 y no se desarrollan bien en
medio alcalino a menos que se hayan adaptado al mismo; por otro lado, las levaduras
crecen mejor en condiciones oxigénicas, pero las fermentativas pueden hacerlo
lentamente en condiciones anoxigénicas. Usualmente, los azúcares son los mejores

alimentos energéticos de las levaduras, aunque las oxidativas, como lo son las
formadoras de película, oxidan ácidos orgánicos y alcoholes (Boulton & Ratledge
1984).
2.4.1 Género Candida spp.
Este género fue propuesto por Berkhout en el año 1923 y desde entonces ha
experimentado algunas modificaciones en cuanto a su definición y composición. Este
género pertenece a la clase Blastomycetes que comprende las levaduras imperfectas
(asexuales). Se considera un taxón heterogéneo que puede dividirse en 40 secciones
que comprenden tres grupos principales, basados principalmente en la composición
de sus ácidos grasos y en el cariotipado electroforético. El nombre del género
significa “blanco radiante” y de ahí que sus células no contengan pigmentos
carotenoides. Forman hifas verdaderas o falsas con abundantes células en gemación o
blastosporas y pude formar clamidosporas (Pelczar 1994).
Las especies de Candida son clasificadas como levaduras, que frecuentemente se
confunden con los hongos, pero estas poseen un predominante modo unicelular de
desarrollo. El genero Candida abarca más de 160 especies, de las cuales se considera
que sólo 18 son patógenas, cuya mas común característica es la ausencia de una
forma sexual. (Odds, F. 1988)
El tubo germinal es una extensión filamentosa de la levadura, sin estrechamiento en
su origen, cuyo ancho suele ser la mitad de la célula progenitora y su longitud tres o
cuatro veces mayor que la célula madre. Sólo C. albicans es capaz de producir
verdaderos tubos germinales; sin embargo, otras especies como C. tropicalis pueden
producir pseudohifas precoces de aspecto similar a los tubos germinales pero con una
zona de constricción característica adyacente a la célula madre, por lo que esta
prueba es útil para diferenciar C. albicans del resto de las especies de Candida,
aunque no está exenta de falsos negativos. (Linares, M; Solís, F. 2001).

Estas levaduras de forma muy variada se reproducen vegetativamente, mediante
gemación. Tienen metabolismo oxidativo, igualmente como fermentativo, en ciertas
condiciones especiales de cultivo forman pseudomicelio. (Romero, J. 2002)
Las especies de Candida pueden secretar una variedad de sustancias durante el
crecimiento, la mayoria de estos son ácidos y alcoholes, todas las formas de acetato y
piruvato como metabolitos comunes. Propionato, succinato, lactato isovalerato y
formato son encontrados en cultivos de C. albicans y otras especies de Candida spp.
La mayoria de especies de cepas patógenas producen acetoína y otras poseen
aminotransferasas que son capaces de formar derivados de piruvato y desde lactato y
alcoholes se da el catabolismo de aminoácidos aromáticos. (Odds, F. 1988)
El potencial patógeno de las levaduras varía en forma considerable, siendo el
microorganismo más virulento Candida albicans, especie del género capaz de generar
con mayor frecuencia enfermedad mortal en seres humanos. C. tropicalis es la
segunda levadura de importancia en cuanto a su patogenicidad. C. parapsilosis
también aparece con cierto grado de virulencia, asociada a micosis oportunistas. Por
otra parte, no todas las cepas de una misma especie presentan igual capacidad
patogénica. Otras especies de Candida causan algunas infecciones, pero la debilidad
del huésped debe ser muy marcada para permitir que estos microorganismos menos
virulentos lo invadan. (López, C. et al. 2005) En el sentido más estricto de la palabra,
no existen levaduras patógenas por naturaleza; las que están relacionadas con
enfermedad en el hombre o animales, son incapaces de producir infección en un
individuo sano. Se deben presentar algunas alteraciones en las defensas celulares del
huésped, en la fisiología, o en la composición de la flora normal para que pueda
producirse la colonización, infección y la enfermedad por levaduras.
Tradicionalmente, la taxonomía de las levaduras se ha llevado a cabo en base a
estudios morfológicos y fisiológicos, pero éstos dependen de las condiciones de
cultivo de las cepas. (López, C. et al. 2005)

La caracterización de levaduras hasta el nivel de especie es de relevancia desde el
punto de vista industrial, debido a que muchos grupos forman parte de la microflora
natural de alimentos y bebidas fermentadas y/o participan en el proceso de obtención
de éstos. De ahí la necesidad de poseer métodos de identificación rápidos, precisos y
fáciles, que puedan ser aplicados al control de la calidad en la industria, con el fin de
asegurar que la cepa de partida, la que conduce el proceso y la que rinde el producto
final, sea la misma. (Orberá, T. 2004)
2.4.1.1 Características Bioquímicas y Fisiológicas
Mientras que las características morfológicas y sexuales permiten generalmente
identificar el género, las características bioquímicas permiten definir la especie de la
levadura. Los principales caracteres son la utilización de compuestos carbonados y
nitrogenados, el crecimiento a 37°C, el crecimiento en medio sin vitaminas y la
resistencia a la ciclohexamida (Beech et al. 1980)
2.4.1.2 Fermentación de los azúcares
Los azúcares probados habitualmente en fermentación son la glucosa, la galactosa, la
sacarosa, la maltosa, la lactosa y la rafinosa. Pueden igualmente probarse otros como
la trehalosa, la melobiosa, y polisacáridos como la inulina o el almidón (Gancedo &
Serrano 1989). La solución base que se utiliza para la fermentación de los azúcares es
el medio agua de levadura (extracto de levadura al 0.5% en agua), provista de una
campana de Durham. Antes de la siembra, en estos tubos se añaden soluciones
estériles de los azúcares a ensayar. La concentración final del azúcar debe ser del 2%
para todos los azúcares salvo para la rafinosa que debe ser del 4%. Los cultivos se
incuban a 25-28°C durante un tiempo que va de 48 horas a tres semanas. Un azúcar
es fermentado cuando hay presencia de gas en la campana (Fiechter et al. 1981).
2.4.1.3 Fisiología del crecimiento de la levadura

El crecimiento de un microorganismo puede considerarse como un serie de
interacciones entre las células y el entorno, aportando el medio los elementos
necesarios para el crecimiento y siendo el mismo modificado por el metabolismo de
las células. Además del aporte de elementos nutritivos, el medio crea alrededor de las
células un entorno más o menos favorable en función de su humedad, de su
temperatura, de su pH o por la presencia de sustancias antimicrobianas (Gancedo &
Serrano 1989).
2.4.1.4 Necesidades nutricionales
El medio de cultivo debe aportar todos los elementos necesarios para la síntesis
celular y para cubrir las necesidades energéticas de las levaduras. Se han dedicado a
este tema trabajos de revisión, entre los cuales se destacan los requerimientos
nutricionales para el crecimiento y la fermentación con levaduras, utilizando
diferentes oligoelementos y sales minerales (Jones & Greenfields 1984, Jones et al.
1981).
Carbono
El carbono es el compuesto mayoritario de la célula de levadura: alrededor del 50%
del peso seco; los compuestos carbonados son utilizados por las levaduras a la vez
como fuente de energía y como fuente de carbono (Fiechter et al. 1981). Según el tipo
de levadura, la capacidad de utilizar ciertos compuestos carbonados varía; algunas
levaduras pueden utilizar una amplia gama de compuestos pero otras asimilan
solamente un número pequeño de ellos (Harrison & Rose 1983).
Entre las fuentes de carbono, los carbohidratos son los más frecuentemente utilizados:
los carbohidratos simples como las hexosas, los disacáridos y los trisacáridos son
metabolizados por un gran número de levaduras (Jones & Greenfields 1984). Durante
los últimos diez años, se han realizado trabajos sobre los carbohidratos abundantes y
poco costosos: las pentosas y los polisacáridos. S. cerevisiae es incapaz de utilizar

pentosas, pero algunas especies de Candida, Metschnikowia y Pichia pueden
convertir la D-xilosa en etanol (Schneider et al. 1998). Además de los carbohidratos,
pueden ser utilizados otros compuestos carbonados que incluyen alcoholes, ácidos o
compuestos menos oxigenados como hidrocarburos. Ha sido efectuada por Levi et al.
(1980) una revisión de las levaduras que crecen sobre alcanos, entre las cuales se
destaca principalmente el género Candida. Ninguna levadura puede oxidar metano,
pero varias especies pueden utilizar el metanol y han sido cultivadas como fuente de
proteína; los principales géneros involucrados son Candida, Hansenula, Pichia y
Torulopsis (Van Unden & Buckley 1982).
Nitrógeno
El nitrógeno es cuantitativamente el segundo constituyente aportado por el medio de
cultivo. Es utilizado por las células en los aminoácidos, los nucleótidos y algunas
vitaminas. Pueden ser utilizadas numerosas fuentes de nitrógeno. Todas las levaduras
son capaces de utilizar el nitrógeno en forma de ion amonio; los iones amonio pueden
ser aportados en el medio por el cloruro amónico, el nitrato amónico, el fosfato
amónico y sobre todo el sulfato amónico, que es el mejor compuesto pues aporta al
mismo tiempo azufre necesario para la síntesis de ciertos aminoácidos (Cooper 1982).
Algunas levaduras son capaces de utilizar los nitratos y los nitritos, en particular las
de los géneros Hansenula, Pachysolen, Citeromyces y ciertas especies de Candida y
de Trichosporon; esta capacidad se utiliza en taxonomía (Kreger Van Rij 1984). En
los medios de cultivo, los nitritos forman a pH inferior a seis ácido nitroso que es
tóxico para las células.
Fósforo
El fósforo se halla incluido en los ácidos nucleicos y los nucleósidos di y trifosfato.
Se encuentra también en forma de polímeros lineales polifosfatos que juegan un papel
importante en la regulación del metabolismo celular (Kulaev & Vagabov 1983). La

concentración de iones fosfato (PO4-3) regula la síntesis de lípidos y carbohidratos. El
fósforo es asimilado por la célula en forma de iones orto fosfato (H2PO4). Las fuentes
de fósforo en el medio de cultivo deben estar constituidas por dihidrogenofosfato de
potasio (KH2PO4) o por el dihidrogenofosfato disódico (Na2 H PO4). En condición
limitante de fósforo en el medio, la célula sintetiza una fosfatasa a partir de los ésteres
fosfóricos (Schurr & Yagil 1981).
Otros compuestos
Pueden añadirse otros compuestos al medio de cultivo como potasio, magnesio,
calcio, zinc, manganeso, y ciertos iones, las necesidades varían cuantitativamente y
según las cepas (Beech et al. 1980). Algunos compuestos son factores de crecimiento
como el inositol y el pantotenato. El inositol juega un papel importante en la síntesis
de los lípidos de las membranas (Henry 1996), pero sólo puede ser utilizada la forma
“meso” (Nikawa et al. 1998). Normalmente el inositol puede ser sintetizado por las
levaduras. El pantotenato se incorpora a las coenzimas A implicadas en las reacciones
de oxidación de los ácidos cetónicos y en el metabolismo de los ácidos grasos. La
vitamina B6 o piridoxina es trasformada en fosfato de piridoxal y en piridoxamina,
coenzimas implicadas en la desaminación y la descarboxilación de los aminoácidos
(Stryer 1981); juega igualmente un papel en el metabolismo de los carbohidratos y de
los ácidos nucleicos (cantidad necesaria: 6.25 mg.1-1 en forma de piridoxina – HCl).
En la formulación de un medio de cultivo o en un medio industrial complejo, es
necesario tener en cuenta las interacciones entre los diferentes constituyentes;
Nagamune et al. (1991) mostraron por ejemplo, el efecto de la interacción de las
concentraciones en (NH4)2SO4 y KH2PO4 sobre la tasa de crecimiento. Por otro lado,
ciertos elementos, a concentraciones demasiado altas, pueden llegar a ser tóxicos.
2.4.2 Cinética de Crecimiento
Para preparar un medio de cultivo para el crecimiento de levaduras, es necesario
determinar que componentes químicos son fuente potencial de alimento para estos

organismos, debido a que algunas sustancias son más aceptables con respecto a otras,
generando en algunas ocasiones efectos antagonistas resultado del crecimiento de
levaduras y otro tipo de organismos debido a la presencia de una mezcla de
componentes no definidos en el medio de cultivo (Haettn 1986). Variaciones en las
condiciones físicas del medio de cultivo, como cambios en la temperatura, pH y
aireación, afectan el crecimiento de las levaduras; por lo tanto, este tipo de factores
deben ser controlados, generando una constante en la población de levaduras y la
evaluación de la cinética de crecimiento en las fases de esporulación, formación de
pseudomicelio, además de la estandarización de un proceso de criopreservación
efectivo para este tipo de microorganismos (Kirsop 1987).
Curva de crecimiento
La cinética de crecimiento de una levadura, puede ser evidenciada en un medio de
cultivo que proporcione todos los requerimientos nutricionales para el
microorganismo, además de poseer las condiciones óptimas de temperatura, pH,
aireación y agitación (Graham & Horman 1990). Las fases conocidas de una curva de
crecimiento típica de una levadura, incluyen la fase de adaptación, fase de
crecimiento exponencial, fase estacionaria y fase de decaimiento (figura 2).
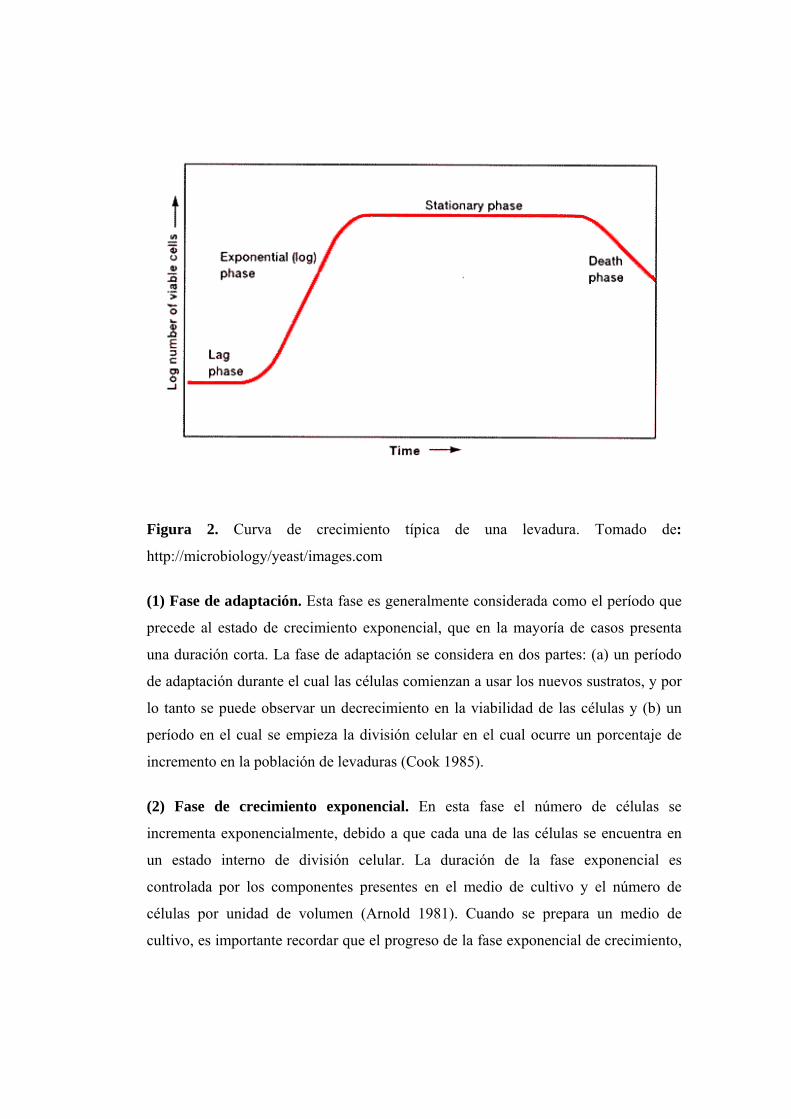
Figura 2. Curva de crecimiento típica de una levadura. Tomado de:
http://microbiology/yeast/images.com
(1) Fase de adaptación. Esta fase es generalmente considerada como el período que
precede al estado de crecimiento exponencial, que en la mayoría de casos presenta
una duración corta. La fase de adaptación se considera en dos partes: (a) un período
de adaptación durante el cual las células comienzan a usar los nuevos sustratos, y por
lo tanto se puede observar un decrecimiento en la viabilidad de las células y (b) un
período en el cual se empieza la división celular en el cual ocurre un porcentaje de
incremento en la población de levaduras (Cook 1985).
(2) Fase de crecimiento exponencial. En esta fase el número de células se
incrementa exponencialmente, debido a que cada una de las células se encuentra en
un estado interno de división celular. La duración de la fase exponencial es
controlada por los componentes presentes en el medio de cultivo y el número de
células por unidad de volumen (Arnold 1981). Cuando se prepara un medio de
cultivo, es importante recordar que el progreso de la fase exponencial de crecimiento,

está dado por la rápida utilización de los nutrientes, seguida de un incremento del
número de células y una acumulación de productos metabólicos finales, los cuales
pueden causar daños a las células. El medio de cultivo se debe mantener en continua
agitación, permitiendo el intercambio de gases entre el medio y la atmósfera interior;
además, es importante adicionar al medio una sustancia buffer durante la fase
exponencial, la cual protege a las células de los continuos cambios de pH, los cuales
son originados por la acumulación de productos metabólicos en su gran mayoría
ácidos (Beech et al. 1980). El tamaño del inóculo no influye en el porcentaje de
crecimiento durante la fase exponencial, sin embargo, un inóculo pequeño puede
retardar la iniciación del crecimiento de la población (Rhan 1991). Es pertinente
mencionar, que todas las determinaciones en el porcentaje de crecimiento de la
población de levaduras, pueden ser modificadas si no se ejerce un rígido control de la
temperatura, debido a que una variación en este factor, es generalmente reflejada en
un cambio sustancial en el porcentaje de crecimiento (Arnold 1981).
(3) Fase estacionaria. En este período el porcentaje de crecimiento comienza a
decrecer y eventualmente el número de células comienza a ser constante. En este
caso, el número de células que mueren es comparativamente igual, al número de
células que se forman, lo cual establece un estado de equilibrio entre las dos
condiciones. Existen diferentes factores que ayudan a la reducción del porcentaje de
reproducción en esta fase, entre los cuales se encuentran los asociados al descenso de
los nutrientes o a la acumulación de productos metabólicos tóxicos (Cook 1985). El
efecto de la disminución de algún nutriente en particular en el medio de cultivo,
supone que el porcentaje de crecimiento es dependiente de la función de ese nutriente
en el metabolismo celular (Berry & Brown 1987).
(4) Fase de decaimiento. Generalmente, el número de células que mueren exceden el
número de células que se producen y el cultivo experimenta una fase de declive
(Cook 1985).
2.4.3 Efecto de la concentración de azucares sobre la levadura

Sin embargo, según sea la severidad del estrés osmótico, el citoesqueleto puede llegar
a colapsar, conllevando a una despolarización de los parches de actina (Willem et al.,
2002), siendo el citoesqueleto una red de fibras proteicas, cuya función es modelar y
organizar el citoplasma, donde muchas de las moléculas individuales de éste se
adhieren al citoesqueleto (Audesirk, 1997). La respuesta primaria de las células de
levadura frente al continuo estrés osmótico, consiste en varios eventos moleculares
que son activados por los cambios celulares repentinos. En primera instancia, la
célula temporalmente detiene el crecimiento, pero hasta el momento no es claro en
qué punto específico del ciclo celular se detiene. Sin embargo, este bloqueo ha sido
reportado a nivel de la G1 (primera fase del ciclo celular en la que existe crecimiento
celular con síntesis de proteínas y de ARN), probablemente por una baja transcripción
de genes que codifican para ciclinas y quinasas dependientes de ciclinas que son
sintetizadas para fosforilar las serinas y treoninas de proteínas diana y desencadenar
así los procesos celulares necesarios para la continuación del ciclo (Willem et al.,
2002).
No es claro hasta el momento por qué las células detienen el crecimiento durante el
ciclo, pero probablemente es parte de un mecanismo estratégico de reajuste, requerido
para un apropiado acondicionamiento a las nuevas condiciones en las que se
encuentran (Willem et al., 2002). En segunda instancia, con el fin de buscar una
recuperación en la turgencia celular, las células producen glicerol como soluto
compatible (Erasmus et al., 2002), el cual es formado como un subproducto durante
la fermentación de etanol por muchas levaduras en concentraciones entre 2.5 y 3.6%
de la producción total de etanol (Petrovska et al., 1999). Por otro lado, el
metabolismo del glicerol cumple un papel muy importante en el balance redox al
igual que en la biosíntesis de fosfolípidos (Willem et al., 2002). En tercera instancia,
la ruta HOG MAP Kinasa (Glicerol en respuesta a la osmolaridad) es activada
resultando en una rápida fosforilación y en una translocación nuclear de la MAP
kinasa Hog1p, encargada de regular tanto la represión como la activación de genes

tales como el gen GPD1, el cual codifica para la expresión de la enzima glicerol 3-
fosfato deshidrogenasa (Willem et al., 2002).
2.4.4 Levaduras en las frutas
Las levaduras mas importantes encontradas dentro de los tejidos de algunas frutas
son; Torulopsis albida, en uva, Rhodotorula glutinis en cereza, junto con Candida
humícola. También se ha encontrado gran variedad de levaduras en el jugo de
naranja, manzana y uva. (Romero, J. 2002)
En los jugos cítricos se ha encontrado Candida krusei, C. parapsilosis,
Hanseniospora melligeri, Pichia fermentaus y Oidum lactis. Donde las propiedades
biológicas de las levaduras determinan la participación en diferentes frutas, siendo
igualmente importante los factores como la temperatura, el pH y la humedad.
(Romero, J. 2002)
Estudios realizados por Ortegón y Ramírez, 2001; Romero, 2002, y Vargas, 2002,
demostraron la presencia de una levadura del género Candida en tomates
heteroinjertados, los cuales fueron estudiados por que presentan un fenómeno de
momificación, ya que se conserva durante más de cuatro meses y medio sin presentar
putrefacción, y además, presenta efecto antagónico frente a ciertos patógenos.
Este fenómeno de momificación en tomates heteroinjertados se ha observado en
frutos de uchuva recolectados de un biocultivo en condiciones de almacenamiento,
por lo que se ha llevado a investigar la posible presencia de esta misma levadura u
otro género como responsable de tal efecto. (Romero, J. 2002)

3. JUSTIFICACION
Colombia es el tercer productor mundial de banano con un área cultivada aproximada
de 30.000 Ha, de las cuales aproximadamente 500.000 toneladas son de banano para
el consumo nacional. Las pérdidas durante la cosecha y postcosecha de banano se han
estimado en 50.000 toneladas/año, equivalentes a más del 10% de la producción
nacional, lo que afecta seriamente la economía del país. El control del proceso de
maduración de frutos climatéricos es uno de los aspectos claves para la
comercialización de este tipo de fruta; esto se debe a que el proceso está íntimamente
relacionado con la baja capacidad de este fruto para soportar períodos de transporte y
almacenamiento prolongados hasta alcanzar los mercados de destino, encontrándose
el producto afectado por factores físicos, biológicos y químicos. Debido a que el
banano se comercializa en diferentes estados de maduración y se almacena bajo
diferentes condiciones se quiere evaluar la eficacia de la aplicación de levadura in
vivo a diferentes concentraciones sobre banano del género Musa especie acuminata,
con el fin de obtener la retardación de la pudrición del fruto y adicionalmente
observar el comportamiento a lo largo del tiempo, siendo ésta una alternativa
biotecnológica, que conlleve a un beneficio potencial a nivel tecnológico, económico
y social. Los bananos son susceptibles de daño mecánico durante la cosecha,
transporte, almacenamiento o procesamiento; dichos cambios causan estrés físico que
afecta los tejidos de la planta y altera el metabolismo fenólico. Las enzimas son
activadas al momento de la cosecha, por lo tanto, durante la maduración se producen
cambios en la actividad enzimática que alteran las estructuras subcelulares. En
ensayos previos realizados en la PUJ se estudio el efecto de una levadura
perteneciente al género Candida spp, aislada inicialmente de tomates HIB y luego
conservada en frutos de uchuva HIB, los resultados obtenidos mostraron que la
levadura disminuía la actividad de la poligalacturonasa PG encargada del proceso de
maduración en postcosecha y de la producción de etileno, siendo capaz de retardar la
pudrición del fruto aproximadamente por 30 días. Es por esto que se pretende evaluar

el efecto de diferentes concentraciones de levadura sobre la retardación de la
maduración del fruto de banano.

4. OBJETIVOS
4.1 Objetivo general
• Aplicación de levadura Cándida sp. como una alternativa viable para la
retardación en la pudrición del banano (Musa acuminata)
4.2 Objetivos específicos
• Aislar de frutos de uchuva HIB, multiplicar y evaluar la cinética de
crecimiento de Candida sp.
• Establecer la concentración más apropiada de inóculo para ser aplicado en el
fruto del banano en un estado de maduración definido.
• Evaluar el efecto de la levadura sobre la maduración de los frutos de banano.

5. MATERIALES Y METODOS
Este proyecto de investigación, se desarrolló en las instalaciones del laboratorio de
Microbiología Ambiental de la Pontificia Universidad Javeriana.
5.Material Vegetal
Se seleccionaron frutos de uchuva HIB provenientes de la Estación Experimental de
la Pontificia Universidad Javeriana que se encontraran en buen estado fitosanitario.
5.1 Ensayo Experimental: Aislamiento y recuperación de la levadura
Al llegar la muestra al laboratorio se procedió a retirar las posibles impurezas
(Guerrero, 2003). Se pesaron 10 gramos de las uchuvas seleccionadas y se lavaron
con agua destilada estéril. En un mortero se macero la fruta, empleando técnicas
asépticas con el fin de evitar contaminación cruzada. (Romero, 2002). El resultado de
la uchuva macerada (extracto) fue llevado a un frasco de vidrio con 90ml de agua
peptonada estéril al 0.1% (Vargas, 2002; Ortiz y Pedraza, 1999). Se realizaron
diluciones en base 10, de 10-1 a 10-3, siendo el frasco de vidrio la primera dilución
(10-1). Se sembró en superficie por duplicado 0.1ml de cada una de las diluciones en
Agar Extracto de malta. Posteriormente se incubo a 25°C +/- 2 durante 5 días. Se
realizaron repiques en Agar YGC el cual se incubo a 25°C durante 4 días (Merck,
2006).
5.2 Descripción macroscópica y microscópica de la levadura
Se realizo una observación macroscópica de la morfología de las colonias teniendo
en cuenta: a) aspecto superficial: liso, arrugado o membranoso, b) color: mate o
brillante; c) consistencia (Casas, 1989). Para la observación microscópica se utilizo la
coloración de Gram y la coloración con azul de lactofenol.
5.3 Identificación bioquímica API 20C AUX (Protocolo de Biomerieux)

A partir de un cultivo joven de la levadura, se realizo una suspensión en 2 ml de agua
destilada estéril hasta obtener una turbidez igual al tubo 2 de McFarland, a partir de
este se transfirieron 100 μl (2 gotas) de la suspensión del Kit API 20C AUX de la
casa comercial Biomerieux a una ampolla de C Medium y se homogenizo evitando la
formación de burbujas. Las cúpulas fueron llenadas con la suspensión anterior
evitando la formación de burbujas y creando un nivel horizontal para generar
resultados correctos, se llevo a incubación a 30 °C durante 48 y 72 h respectivamente.
La lectura de estas reacciones se hizo por comparación con un control de crecimiento
y la identificación fue determinada utilizando el programa Apiweb TM. (ANEXO 3).
5.4 Prueba de patogenicidad
Siguiendo el protocolo expuesto por Linares, F.; Solis, M. 2001 Se emulsiono una
colonia aislada en 0,5 ml de suero humano, se incubo a 35°C durante 2 h. y se
visualizó por coloración de Gram a objetivo 100x.
5.5 Determinación de biomasa por peso seco en función del tiempo
Para realizar la curva patrón de peso seco, se preparó una suspensión concentrada de
la levadura, fue lavada dos veces con solución salina 0.85% (p/v) por centrifugación a
2250 g por 20 minutos. El pellet se resuspendió al volumen inicial. Posteriormente,
fueron adicionados en 10 tubos previamente pesados 10 mL de la solución de células
a cada uno, igualmente 10 mL de solución salina en 10 tubos que sirvieron como
blanco. Estos fueron llevados al horno a 105°C por 24 horas, terminado el tiempo de
evaporación del líquido se pasaron al desecador para ser pesados nuevamente y
determinar el promedio del peso de los 10 tubos que contenían las células y el de los
10 tubos con solución salina, a partir de esto se calculó la diferencia de peso entre los
dos promedios para expresar la concentración en gramos de biomasa seca por litro de
solución (g/L). A partir de la solución concentrada de células se realizaron diluciones
por triplicado para ajustar la absorbancia entre 0.2 – 0.9 y calcular las
concentraciones finales para cada dilución. Esto permitió hallar la ecuación

bcmAbs += * al graficar la absorbancia promedio en función de la concentración
celular (g/L). Se realizaron diluciones 1/1.5, 1/2, 1/3, 1/4, 1/5, 1/6, 1/7, 1/8, 1/9. Y se
registro la absorbancia de cada réplica a 620 nm.
5.6 Curva de crecimiento
Preparación de Inoculo
La cepa de levadura fue cultivada en agar YGC cuya composición es en g/L: extracto
de levadura 5, D-glucosa 20, cloramfenicol 0.1 y agar-agar 14.9, pH final 6.6 (Merck,
2000), bajo condiciones controladas de 25ºC por 4 días. Posteriormente, se realizó un
raspado de las colonias para preparar una suspensión de 5 ml en solución salina
0.85%(p/v), a una concentración de 108 células/ml, igualando al tubo 3 de McFarland.
y realizando recuento en cámara de Neubauer para obtener la misma concentración,
esta suspensión fue adicionada a 45 ml de caldo YGC el cual se incubó por 24 horas a
25° C y 120 RPM.
Fermentación.
En un erlenmeyer de 1000 ml se adicionaron 450 ml de caldo YGC estéril y
posteriormente se agregó el 10% de inóculo (50 ml) y las condiciones de operación
fueron controladas: temperatura 25°C, agitación de 120 rpm (Noor, et al., 2003).
Luego de ser inoculado el erlenmeyer se retiraron 3 ml de muestra, que correspondió
a la hora cero (0) y posteriormente se realizaron cada dos horas hasta completar26
horas; a estas muestras se les determinó el pH, la Absorbancia a 620 nm y se realizo
recuento en Cámara de Neubauer.
Recuento en placa
A partir de las muestras de cada uno de los tiempos de fermentación, se tomó 1 ml del
cultivo y se realizaron diluciones seriadas de 10 -1 a 10 -9 y se sembró 50 microlitros

mediante la técnica de microgota en medio YGC. Las cajas fueron incubadas a
temperatura óptima de 25°C por 72 horas. Transcurrido el tiempo de incubación se
realizó recuento de UFC/ml por triplicado.
5.7 Actividad Enzimática Cualitativa
Se realizó un pre enriquecimiento de la levadura en caldos específicos para inducir las
diferentes actividades enzimáticas; por lo tanto, se inoculó una colonia de levadura en
200 ml de medio, distribuido en erlenmeyer de 500 ml y se dejó en agitación a 200
rpm durante 48 horas. Los medios utilizados en este procedimiento incluyeron: caldo
celulosa, caldo pectina y caldo almidón. (Barrera 2000)
5.8 Crecimiento en Agar Banano
Con el fin de evidenciar la capacidad de la levadura por consumir los sustratos que le
proporciona el banano, se procedió a realizar un agar cuya composición fue: 20 g/L
de pulpa de banano en estado de maduración 3 según la tabla de Turbana y 15 g/L de
agar agar, no se agrego ningún inhibidor con el fin de evidenciar si se presentaba
alguna relación de tipo antagonista con otros microorganismos nativos provenientes
del banano, se realizo el mismo procedimiento pero utilizando banano en estado de
maduración 5 según la tabla de Turbana (Anexo 2) con el fin de proporcionar mayor
cantidad de azúcares disponibles para el crecimiento de la levadura.
5.9 Aplicación
Para la aplicación de la levadura se utilizaron bananos baby (Musa acuminata) en
estado de maduración número 5 según la tabla de colores de Turbana (Anexo 2), se
realizaron 3 aplicaciones con 3 replicas para controles, concentración #1 y
concentración #2 respectivamente. (Tabla 3).

Dias control concentracion 1 concentracion 2 0 3 3 33 3 3 36 3 3 39 3 3 3
12 3 3 315 3 3 318 3 3 327 3 3 336 3 3 345 3 3 3
30 30 30Total 90
Numero de bananos
Tabla 3. Número de bananos para cada día de estudio
Los bananos a utilizar en la aplicación y como controles fueron agrupados de
acuerdo a su peso y condiciones físicas Figura 3.
Figura 3. Clasificación y agrupación de bananos para el estudio.
La forma de aplicación utilizada en este estudio fue inmersión por 2 min la cual ha
sido utilizada en estudios previos de deshidratación de frutos por Lemos, L. et al,
2006. El ensayo fue realizado en el laboratorio bajo condiciones ambientales de

temperatura entre 19°C y 20 °C, y una humedad relativa de 70%. Fueron
determinadas variables para evaluar el desarrollo de la maduración de los frutos cada
3 días. Las variables evaluadas fueron:
5.9.1 Pérdida Fisiológica de Peso (PFP)
Los frutos fueron agrupados según el tratamiento y pesados cada 3 días en una
balanza analítica con el fin de determinar su peso en gramos y evaluar el
comportamiento. Figura 4
Figura 4. Bananos después de aplicación.
5.9.2 Determinación de pH:
Se preparo una mezcla de pulpa/agua 1:1 en agua destilada, se homogenizo y se
midió el pH siguiendo el protocolo descrito por Arrieta, A et al. 2006
5.9.3 Determinación de Grados Brix: Debido a la dificultad de obtener liquido a
partir del fruto, 30g de fruta fueron macerados en 90ml de agua destilada, (Figura 5.
A) Esta mezcla fue filtrada y una gota de jugo se colocó sobre el cristal del
refractómetro y se observo a la luz el ocular del instrumento observando la escala,

(Foto 5 B) teniendo en cuenta la temperatura de medición, la determinación se hizo
por triplicado para cada tratamiento. (Arrieta, A.; et al 2006)
Figura 5-A: Macerado de banano Figura 5.B: Refractómetro Portatil
6. ANALISIS ESTADISTICO
Los datos obtenidos a partir de los promedios de los pesos del fruto con respecto al
tiempo, en los controles y en las concentraciones de estudio, se analizaron
estadisticamente por medio de anova, la prueba T student y regresión lineal, con el fin
de evaluar el comportamiento de los datos y poder comparar las concentraciones
utilizadas, para conocer la acción de la levadura en el fruto.
Hipotesis general
Ho = No hay diferencia significativa entre los datos.
Fexp< Ftabla; acepta la Ho. Fexp>Ftabla; rechaza la Ho. Hi = Hay diferencia significativa entre las concentraciones.
Fexp>Ftabla; acepta la Hi. Fexp<Ftabla; rechaza la Hi. Ho= No hay diferencia significativa entre los grados brix entre concentraciones, para p<0.05 Si p<0.05: se acepta Ho Si p>0.05: se rechaza Ho
Ho=No hay diferencia significativa entre los valores de pH entre las

concentraciones, para p <0.05 Si p<0.05: se acepta Ho Si p>0.05: se rechaza Ho
7. RESULTADOS Y DISCUSIÓN
7.1 Descripción Macroscópica y Microscópica
A partir de los frutos de uchuva recolectados, se realizo el aislamiento de la
Levadura en medio YGC, La coloración de Gram mostró células, ovaladas, con
diámetro considerable, en estado de gemación y Gram positivas, debido a la presencia
de una pared celular gruesa, que permite la retención del complejo cristal violeta-
yodo, confirmando así la presencia de estructuras levaduriformes (Figura 6).
Figura 6. Características microscópicas de Cándida sp. obtenidas por coloración de
Gram.
Las colonias de Candida spp, en este medio se visualizan como colonias blancas
pequeñas, redondas, lisas, brillantes, las cuales a través del tiempo toman una forma
puntiforme (Figura 7). El agar YGC favoreció el crecimiento de esta levadura por su
alto contenido de glucosa como fuente de carbono y extracto de levadura como fuente
de nitrógeno

Figura 7. Características macroscópicas de las colonias de Candida sp. en agar YGC
7.2 Prueba de patogenicidad
La presencia de Candida albicans fue descartada al realizar la prueba de tubo
germinal, donde el resultado fue negativo ya que solamente se observo la levadura
aislada, donde se evidencio la ausencia del tubo germinal (filamentación precoz)
7.3 Prueba de asimilación de azúcares (API 20 C AUX)
La identificación realizada mediante pruebas bioquímicas en caldo de hidratos de
carbono base rojo de fenol, permitió evidenciar la ausencia de CO2 en la campana de
Durham, lo que indica la ausencia de metabolismo fermentativo por parte de la
levadura. Los resultados en tubo fueron evaluados con base en el cambio de pH
(viraje de color) y crecimiento (turbidez).
Tabla 4. Resultados de asimilación de azucares por Candida spp.
Hidrato de carbono Resultado de la asimilación
Glucosa Positivo
Sacarosa Positivo

Lactosa Negativo
Xilosa Positivo
Fructosa Positivo
Galactosa Positivo
Celobiosa Positivo
La identificación de la Levadura fue llevada a cabo por el API 20C AUX, luego de
ser observados los resultados de las bioquímicas, para conocer la asimilación de
azucares por parte de la levadura, para contar con mayor exactitud en los resultados.
Luego de las 48 y 72 horas de incubación del API 20C AUX, el resultado se
evidencio al evaluar las cúpulas con respecto al control de crecimiento (ANEXO 4),
de esta manera los resultados fueron adicionados al programa informático Apiweb
TM, el perfil numérico arrojado por la levadura fue 6776377, identificándose de esta
forma la levadura Candida guilliermondii presentando un taxón significativo de
99.7%. (Figura 8).
Fig. 3 Resultado programa informático Apiweb TM API 20 C AUX.
7.4 Crecimiento en agar banano amarillo y verde

Para determinar si la levadura de estudio contaba con la capacidad enzimática para
tomar como sustrato los componentes del banano se realizó un agar a base del fruto
del banano donde la levadura fue sembrada a partir de colonias aisladas del agar
YGC, de esta manera, después de 5 días de crecimiento a temperatura ambiente se
observo la presencia de colonias mucosas, blancas, pequeñas (Figura 9). En el agar
banano maduro se evidencio que el tamaño de las colonias era más grande y el
crecimiento se presento en menos días, en agar banano verde se evidencio
crecimiento, sin embargo, un poco más limitado que en agar banano amarillo,
probablemente por la disponibilidad de nutrientes en los diferentes tipos de banano,
puesto que por factores fisiológicos de la maduración del banano, la concentración de
almidón es mucho mas alta al estar el banano en estado verde que en estado de mayor
maduración de color amarillo donde este ya ha sido convertido en azucares simples,
siendo mas fácil su asimilación por parte de la C. guillermondii.
Figura 9. Crecimiento de C. guillermondii en agar banano
7.5 Actividad enzimática cualitativa
El comportamiento presentado en el caldo enriquecido con almidón mostró la
ausencia de actividad amilolitica por parte de la levadura, es decir, carece de la
maquinaria enzimática propia para poder degradar la molécula de almidón en
moléculas simples de glucosa, este resultado demuestra el pobre crecimiento que
presenta la levadura aplicada en los frutos de banano verde en estado de maduración
1 y 2 según la tabla de colores de Turbana ya que su principal componente es el

almidón. Sharrock y Lusty (2000) encontraron que los niveles de almidón en banano
verde son del orden del 20%, y van disminuyendo hasta 1%-2% en banano
completamente maduro, por esta razón la aplicación se realizo en banano en estado
de maduración 3, donde la concentración de azucares disponibles es mayor para la
levadura y le permite colonizar rápidamente el fruto.
Por otra parte se evaluó el comportamiento celulolitico por parte de la levadura
evidenciándose un crecimiento favorable en este caldo, siendo este el principal
componente de la cascara del fruto, lo que sugiere la capacidad que tiene la levadura
de poder penetrar por las laminas de la cascara llegando a la pulpa. La actividad
pectinolitica se evidencio por crecimiento en caldo pectina, este componente es un
polisacárido constituido principalmente por la unión de moléculas de acido
galacturónico parcialmente metoxilado, siendo abundante en la pulpa del fruto, la
actividad que presento la levadura frente a este sustrato fue favorable demostrándose
su capacidad por consumir esta fuente de carbono para su crecimiento y consiguiente
colonización del fruto.
7.6 Determinación de parámetros cinéticos de crecimiento
En la curva de peso seco, no se tuvo en cuenta el peso de la sal, ya que esta es soluble
en agua y se retiro al dejar solo 2mL en los tubos para el secado, por esta razón los
tubos con solución salina presentaron mayor peso, por lo cual no fue posible
determinar la diferencia con los tubos de biomasa, de esta manera solo fueron
tomados los pesos de biomasa para establecer la curva de peso seco. (Figura 10)

y = 0.2584x + 0.0401R² = 0.9918
0.0000.1000.2000.3000.4000.5000.6000.7000.8000.9001.000
0.00 0.50 1.00 1.50 2.00 2.50 3.00 3.50 4.00
ABS
(620
nm)
Concentración (g/L)
CURVA DE PESO SECO
Figura 10. Curva de peso seco de C. guillermondii. Absorbancia en función de la
concentración de Biomasa (g/L)
Debido al optimo crecimiento por parte de la levadura en agar YGC, se llevo a cabo
la curva de crecimiento, en la cual se evidencio el comportamiento de C.
guilliermondii en función del tiempo. La curva de crecimiento fue llevada a cabo con
el fin de determinar la concentración de levadura para aplicar al fruto del banano,
conociendo la fase exponencial, para así asegurar que la aplicación cuenta con la
levadura en su momento máximo de duplicación. (Tabla 5)
Tabla 5. Datos de abs y concentración obtenidos a partir de la curva de crecimiento
de C. guillermondii

TIEMPO (Horas) Abs 620 nm CONCENTRACIÓN g/L CONCENTRACIÓN mg/L Ln Biomasa UFC/mL0 0,511 0,239 239 5,476 5,80E+082 0,713 0,32 320 5,768 7,00E+084 0,944 0,412 412 6,021 8,00E+08
6 1,304 0,556 556 6,321 9,80E+088 1,470 0,622 622 6,433 1,80E+0910 1,620 0,682 682 6,525 2,60E+0912 1,668 0,701 701 6,553 3,80E+09
14 1,753 0,736 736 6,601 4,50E+09
16 1,721 0,723 723 6,583 4,10E+0918 1,790 0,75 750 6,620 4,63E+0920 1,864 0,78 780 6,659 4,79E+09
22 1,932 0,807 807 6,693 4,96E+09
24 1,912 0,799 799 6,683 4,88E+0926 1,900 0,794 794 6,677 4,82E+09
En la curva de crecimiento al utilizar un pre inoculo de 24 horas correspondiente al
10% del volumen final de escalado, se pudo reducir la fase de latencia al inicio de la
curva de crecimiento evaluándose hasta la hora 26 (Figura 11).
0,000
0,500
1,000
1,500
2,000
2,500
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28
Abs 620
nm
Tiempo (horas)
Abs (nm) Vs Tiempo (horas)
Figura 11. Curva de crecimiento C. guillermondii en medio YGC.
La fase exponencial donde la levadura cumple con su mayor ciclo de duplicación se
evidencio desde la hora 0 hasta la hora 14 (Figura 12), aunque el mayor crecimiento
de la levadura fue presentado a la hora 22, este tiempo de crecimiento no fue tomado
en la fase exponencial de C. guillermondii, debido a la baja linealidad que se
presento entre la hora 14 a 22 mostrando un comportamiento correspondiente a la

fase estacionaria del crecimiento de la levadura, por tal razón luego de conocer la
curva de crecimiento, la levadura fue aplicada en el fruto a las 14 y 22 horas de
crecimiento; donde la biomasa de C. guillermondii estaba a una concentración de 4.7
x 109 UFC/mL a las 14 horas de crecimiento siendo esta la concentración 1, y a una
concentración de 5.17 x 109 UFC/mL, correspondiente a las 22 horas de crecimiento,
siendo esta ultima la concentración 2 de aplicación.
y = 0.0932x + 0.5955R² = 0.9459
0.000
0.500
1.000
1.500
2.000
0 2 4 6 8 10 12 14 16
Abs (6
20nm
)
Tiempo (horas)
Crecimiento Exponencial de Candida guillermondii
Figura 12. Fase exponencial de C. guillermondii. Abs 620 nm en función del tiempo
(horas)
El comportamiento del pH no se mantuvo estable a través del tiempo, debido al
crecimiento de C. guillermondii en el medio, el pH fue de 5.42 a la hora 0 de la curva
de crecimiento, a través del tiempo, el pH disminuyo hasta llegar a un valor de 4.73 a
las 10 horas, indicando la acidificación del medio debido al consumo de glucosa y
producción de ácidos como el succínico, acético, fórmico, propiónico y pirúvico por
parte de la levadura los cuales no fueron cuantificados, sin embargo varios autores lo
reportan, el pH ácido (4.5 – 5) que se evidencia durante la curva es una de las
condiciones óptimas para la levadura al incrementar la producción de biomasa (Yu y
Zhang, 2004; Martín, et al, 2005; Palmarola, et al, 2005). A partir de la hora 12 el pH
aumento debido al consumo del extracto de levadura; ya que se genera la liberación

de aminoácidos y otros productos nitrogenados (Buitrago. J; Tenjo, D. 2007.).
(Figura 13)
4.64.74.84.95
5.15.25.35.45.5
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28
pH
Tiempo (horas)
Comportamiento del pH en función de tiempo (horas)
Figura 13. Comportamiento del pH durante la curva de crecimiento
La cinética de crecimiento presentada por C. guillermondii a través de la curva de
peso seco, demostró la presencia de una prolongada etapa de crecimiento logarítmico,
sin embargo, debido posiblemente al tipo de sustrato la tasa reproductiva fue baja y el
tiempo de duplicación estuvo entre las 7 horas 37 min con una velocidad especifica
de crecimiento de 0.0908h-1. (Figura 14)
Td= Ln2 / μx Td= 0,693/0.0908hTd= 7.63h Td= 7h 37'
y = 0,0908x + 7,7289R² = 0,8833
7,000
7,500
8,000
8,500
9,000
9,500
0 2 4 6 8 10 12 14 16
Ln (Con
centración
mg/L)
Tiempo (horas)
Ln de la concentración (mg/L) en función del tiempo

Figura 14. Determinación de la velocidad específica de crecimiento por C. guillermondii
7.8 Comportamiento del pH en el fruto.
El pH ácido que se evidencia durante el proceso de maduración en el fruto (Figura
15) se reporta según literatura que es debido a la presencia de ácidos orgánicos en la
pulpa del fruto tales como: málico, cítrico y oxálico, cuyos niveles se incrementan
durante el proceso. En un fruto verde la concentración de ácido oxálico es mayor,
mientras que en el fruto maduro es el ácido málico. La pulpa del fruto contiene en
estado verde 0.7% de ácido málico y 1.5% en estado maduro. La cáscara en estado
verde contiene 1.0% de ácido málico y 1.4% en estado maduro. (Cayón, D. Et al.
2000).
Los resultados obtenidos con respecto al pH en los frutos de banano, mostraron un
incremento de la acidez a medida que aumentaba su tiempo de maduración tal y como
se reporta anteriormente, sin embargo a partir del día 18 aproximadamente en los
tratamientos se evidencia un comportamiento que tiende a aumentar el pH según
literatura esto puede deberse a que durante la senescencia se presenta un incremento
en el contenido de los aminoácidos libres, consecuencia de la degradación de
enzimas. (Gallo, F. 1997).
4
4,5
5
5,5
6
6,5
0 10 20 30 40 50
pH
Tiempo (dias)
Comportamiento del pH durante la maduración de Musa acuminata.
Conc. 1
control
conc 2

Figura 15. Comportamiento del pH durante la maduración del fruto Tratamientos y
control.
Con relación a los sólidos solubles totales (grados Brix), éstos se incrementaron en
función a las semanas de maduración de los frutos. Debido a la dificultad de extraer
pulpa para realizar el macerado, los SST fueron cuantificados hasta el día 25.
Según Hubbard et al. (1990) y Seymour et al. (1987), los grados Brix aumentan
durante la maduración debido a la rápida degradación del almidón, acumulándose
azúcares, principalmente glucosa, fructosa y sacarosa. Tanto en los dos tratamientos
como en el fruto control, el comportamiento de los SST fue similar debido a que el
proceso de maduración no es inherente en el fruto, sin embargo, en el fruto control a
los 20 días, se evidencia un incremento en los SST seguido de un periodo de
estabilidad esto puede deberse a que en la pulpa se presenta conversión del azúcar en
alcohol, proceso natural que se presenta durante la maduración del fruto. (Figura 16).
10121416182022242628
0 5 10 15 20 25
°Brix
Tiempo (días)
Comportamiento de los SST (° Brix) en función del tiempo (dias)
Control
Concentración 1
Concentracion 2
Figura 16. Comportamiento de los SST durante la maduración del banano en
tratamientos y control.

Analizando estadísticamente los datos de SST por la prueba T (Tabla 6), se demostró
que no hay diferencias estadísticamente significativas entre los datos obtenidos con
respecto a los controles y la concentración 1 y 2, corroborando que el
comportamiento de los SST es similar tanto en los frutos control como en los frutos
con levadura, lo que indica que el aumento en los SST es un proceso fisiológico
natural de la maduración y no se ve significativamente influenciado por la aplicación
de levadura.
Prueba T Ctr Vs Conc 1 0,44446287Prueba T Ctr Vs Conc 2 0,24868646Prueba T conc 1 Vs conc 2 0,68963099
Tabla 6. Resultados de la probabilidad por prueba T para SST con un intervalo de
confianza del 95 %.
7.9 Textura
Durante el tiempo de estudio se evidencio pérdida de firmeza de la pulpa del fruto
tanto en los controles como en los tratamientos este comportamiento se asocia a tres
procesos principalmente, 1. Degradación del almidón para formar azúcar; 2.
Degradación de las paredes celulares o reducción en la cohesión de la lamela media
debido a la solubilización de las sustancias pécticas; 3. El movimiento del agua desde
la cáscara hacia la pulpa debido al proceso de ósmosis, el cual favorece posiblemente
la movilidad de la levadura dentro del fruto permitiéndole invadir el fruto y ejercer
efecto sobre el. Este comportamiento de la textura es muy leve al inicio de la
maduración porque el fruto está verde, pero de ahí en adelante la estructura se debilita
por la maduración hasta llegar a la senescencia, efecto causado por la degradación de
la protopectina en pectina y en ácido galacturónico. (Chang-Yuen, Et.al 2005)
La perdida de firmeza y agua en el fruto posiblemente sea una de la causas por la
cuales aproximadamente entre el día 15 y 18 donde se evidencia deshidratación del
fruto (Figura 17) haya una disminución considerable en la población de la levadura

debido a la dificultad de movilizarse dentro de la pulpa, lo que le impide consumir el
sustrato presente en el medio, provocando su decaimiento.
Figura 17. Efecto de la levadura a concentración 1 sobre el fruto a los 18 días de
estudio.
Estudios realizados por Rubiano, A. 2008, han demostrado que la concentración de
levadura dentro del fruto se incrementa progresivamente, se estudio el crecimiento de
la levadura C. guillermondii en la Cáscara Total (CT), Pulpa Exterior (PE) y en la
Pulpa Centro (PC) del banano (Musa acuminata) en estado de maduración 5 leída a lo
largo de 17 días de experimento y sometida durante 5 min en una solución acuosa con
una concentración de 45*108 UFC/ml y 46*108 UFC/ml.
Los resultados encontrados por Rubiano, A. 2008, demuestran la presencia de la
levadura y su incremento progresivo en la cascara y en la pulpa del fruto. Se observa
que la levadura en un tratamiento de inmersión por 5 min, empezó a migrar en la
pulpa a partir del día 11 y se incremento hasta el día 14 con 200 UFC. Este
comportamiento se relaciona directamente con el comportamiento del pH en el fruto

(Figura 15) ya que aproximadamente a partir de este día se evidencia una fase de
estabilización del pH lo que no ocurre con el fruto control en el cual el pH sigue
disminuyendo progresivamente, comportamiento natural del fruto.
El comportamiento de la levadura en los dos tratamientos posiblemente sea escaso ya
que el tiempo de inmersión no fue el adecuado para permitir una completa
impregnación de la levadura en el fruto.
En un banano control (Figura 18) el contenido de humedad de la cáscara disminuye
durante la maduración mientras en la pulpa aumenta, el porcentaje de agua en la
pulpa aumenta durante la maduración, no solo por la hidrólisis del almidón sino
también por el movimiento osmótico de agua desde la cáscara hacia la pulpa. Un
incremento en el peso de la pulpa afectando la relación pulpa: cáscara, parece ser una
consecuencia de la salida osmótica de la humedad de la cáscara. (Chang-Yuen, Et.al
2005). Efecto similar en cuanto al contenido del agua de la cáscara se evidencia en
los dos tratamientos sin embargo a partir del día 15 se evidencia que el
comportamiento del peso se estabiliza, efecto contrario al fruto control, esto puede
deberse a que aunque la perdida de agua en el banano a lo largo del estudio es
notoria, su peso se mantiene debido a que aproximadamente en este día se encuentra
la mayor población de levadura dentro del fruto y esta a su vez se encarga de
consumir el agua presente en el fruto necesaria para su supervivencia, una hipótesis
planteada acerca de el tiempo que puede durar la levadura en el fruto se atribuye
igualmente a la cantidad de agua en el fruto ya que como se relaciona anteriormente
en los frutos tratados se evidencia un proceso de deshidratación que se prolonga
desde el día 15 y continua por los 45 días del estudio (Figura 18), la disminución de
agua en la pulpa del fruto impide que haya una textura suave en el fruto cerrándose
los canales entre las laminas que le permitían a la levadura movilizarse para consumir
alimento presente en el fruto, es por esto que la concentración de levadura tendería a
disminuir hasta que llegue el momento en que sea insignificante dentro del fruto.

Figura 18. Perdida de agua y firmeza en el fruto del banano después de la aplicación
de levadura (Musa acuminata).
De izquierda a derecha fruto control y tratamiento Concentración 1.
Con respecto al color de la pulpa se evidencio que en los frutos tratados después del
día 18 presentaron pardeamiento de la pulpa, fenómeno directamente relacionado
según literatura con los niveles de ácido ascórbico, el contenido de polifenoles, la
actividad de la enzima polifenol oxidasa (PPO) o una combinación de estos factores,
el grado de pardeamiento de la pulpa esta correlacionado positivamente con la
concentración total de polifenoles y la actividad de la PPO. (Cayón, D. Et al. 2000).
Otras de las hipótesis planteadas para argumentar la desaparición de la levadura en el
fruto es la concentración de glucosa a la que se encuentra expuesta en el fruto
maduro, esto puede ser una consecuencia de la respuesta primaria hacia el estrés
osmótico al que están expuestas continuamente las levaduras (Willem et al., 2002).
Resultados similares fueron obtenidos por Erasmus y colaboradores (2003), quienes
encontraron que el estrés osmótico redujo significativamente el crecimiento de
Saccharomyces cerevisiae en jugo de uva, el cual tenía una concentración de azúcar
del 40% (w/v).
Las levaduras tienen la capacidad de responder rápidamente al estrés provocado por
las concentraciones de azúcar en el medio en el que se encuentran mediante una serie
de mecanismos, entre los que se distinguen: 1) cambios celulares inmediatos, que
ocurren como consecuencia de las fuerzas físico-mecánicas que se presentan bajo
estas condiciones; 2) procesos de defensa primaria que buscan dar protección a la
célula, reparación y recuperación, al igual que 3) eventos adaptativos que permiten la
restauración de la homeostasis celular bajo las nuevas circunstancias (Willem et al.,
2002).
Se puede entonces establecer que factores como el pH, la temperatura y las altas
concentraciones de glucosa (esta última relacionada con el Aw y el estrés osmótico)
ejercen una gran influencia en el desarrollo y supervivencia de las levaduras.
7.10. ANÁLISIS ESTADÍSTICO.
Análisis estadístico.
Por medio de la herramienta estadística Anova normal, se determino la diferencia
significativa entre los datos de los pesos de los frutos control, frutos con aplicación de
concentración 1 y frutos con aplicación de concentración 2 de C. guillermondii,
presentándose un comportamiento normal de los datos obtenidos. (Tabla 7, 8, 9)
Tabla 7. Promedio de pesos de las replicas de los frutos control a través de los días.
Réplicas dia 0 dia 3 dia 6 dia 9 dia 12 dia 15 dia 18 dia 27 dia 36 dia 45 R 1 58.32 57.45 55.67 53.04 50.14 45.27 44.08 41.09 37.16 35.67R 2 58.44 56.87 53.72 52.42 51.84 49.65 45.17 42.74 38.29 36.49R 3 59.21 57.78 56.48 55.53 52.72 47.06 44.53 41.72 37.38 36.08
dia 0 dia 3 dia 6 dia 9 dia 12 dia 15 dia 18 dia 27 dia 36 dia 45 Promedio 58.66 57.37 55.29 53.66 51.57 47.33 44.59 41.85 37.61 36.08Desviacion 0.483 0.461 1.419 1.646 1.312 2.202 0.548 0.831 0.599 0.410
Decrecimiento 1.29 2.08 1.63 2.10 4.24 2.73 2.74 4.24 1.53Perdida de peso total 22.58Porcentaje de perdida 0 5.71 9.20 7.20 9.29 18.78 12.11 12.14 18.78 6.78
Tabla 8. Promedio de pesos de las replicas de los frutos con aplicación de la
concentración 1.

Réplicas dia 0 dia 3 dia 6 dia 9 dia 12 dia 15 dia 18 dia 27 dia 36 dia 45 R 1 51.79 50.92 49.39 47.18 44.68 40.17 38.96 37.08 36.96 35.98R 2 52.18 51.22 49.71 47.66 45.35 41.06 39.29 37.36 37.24 36.16R 3 51.90 50.57 49.36 47.10 44.44 39.13 38.34 36.59 36.52 35.68
dia 0 dia 3 dia 6 dia 9 dia 12 dia 15 dia 18 dia 27 dia 36 dia 45 Promedio 51.96 50.90 49.49 47.31 44.82 40.12 38.86 37.01 36.91 35.94Desviacion 0.201 0.325 0.194 0.303 0.472 0.966 0.482 0.390 0.363 0.242
Decrecimiento 1.05 1.42 2.17 2.49 4.70 1.26 1.85 0.10 0.97Perdida de peso total 16.02Porcentaje de perdida 0 6.58 8.84 13.57 15.55 29.37 7.85 11.57 0.65 6.04
Tabla 9. Promedio de pesos de las replicas de los frutos con aplicación de la
concentración 2.
Réplicas dia 0 dia 3 dia 6 dia 9 dia 12 dia 15 dia 18 dia 27 dia 36 dia 45 R 1 53.98 52.12 50.08 47.71 45.05 43.24 41.98 41.16 39.65 38.03R 2 53.88 51.89 50.21 48.14 45.58 43.52 42.48 41.45 39.89 38.34R 3 53.91 52.02 49.98 46.84 44.73 42.79 41.89 40.85 39.28 37.87
dia 0 dia 3 dia 6 dia 9 dia 12 dia 15 dia 18 dia 27 dia 36 dia 45 Promedio 53.92 52.01 50.09 47.56 45.12 43.18 42.12 41.15 39.61 38.08Desviacion 0.051 0.115 0.115 0.662 0.429 0.368 0.318 0.300 0.307 0.239
Decrecimiento 1.91 1.92 2.53 2.44 1.94 1.07 0.96 1.55 1.53Perdida de peso total 15.84Porcentaje de perdida 0 12.08 12.12 15.95 15.42 12.22 6.73 6.08 9.76 9.64
A partir de la regresión lineal de los datos de la pérdida de peso a medida del tiempo,
se determino la ecuación de la recta y = mx + b tanto de los controles, concentración
1 y concentración 2, la pendiente con tendencia negativa por su comportamiento
decreciente, determino que la pérdida de peso es directamente proporcional al
tiempo, de este modo se obtuvieron pendientes iguales a un valor de: -0.5403 para los
controles,-0.3856 para la concentración 1 y de -0.3399 para la concentración 2.
(Figura 19, 20, 21)
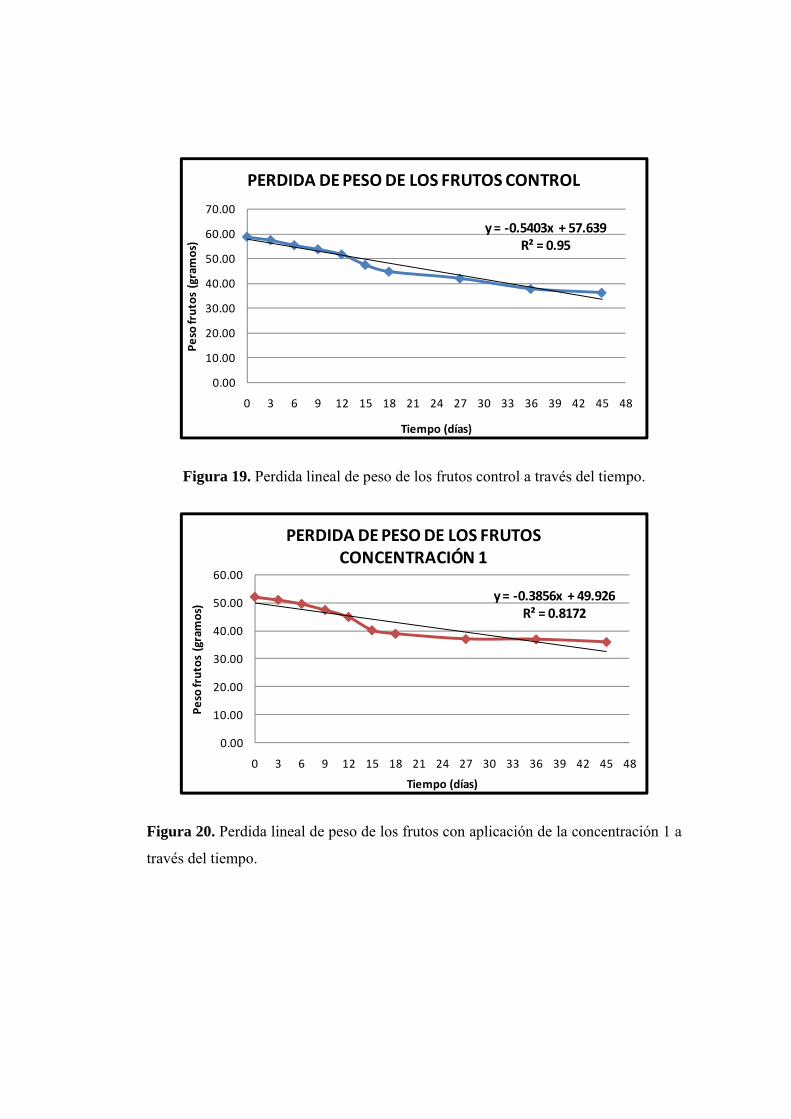
y = ‐0.5403x + 57.639R² = 0.95
0.00
10.00
20.00
30.00
40.00
50.00
60.00
70.00
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42 45 48
Peso frutos (gram
os)
Tiempo (días)
PERDIDA DE PESO DE LOS FRUTOS CONTROL
Figura 19. Perdida lineal de peso de los frutos control a través del tiempo.
y = ‐0.3856x + 49.926R² = 0.8172
0.00
10.00
20.00
30.00
40.00
50.00
60.00
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42 45 48
Peso frutos (gram
os)
Tiempo (días)
PERDIDA DE PESO DE LOS FRUTOS CONCENTRACIÓN 1
Figura 20. Perdida lineal de peso de los frutos con aplicación de la concentración 1 a
través del tiempo.

y = ‐0.3399x + 51.098R² = 0.8519
0.00
10.00
20.00
30.00
40.00
50.00
60.00
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42 45 48
Peso Frutos (gramos)
Tiempo (días)
PERDIDA DE PESO DE LOS FRUTOS CONCENTRACIÓN 2
Figura 21. Perdida lineal de peso de los frutos con aplicación de la concentración 2 a
través del tiempo.
La pendiente más cercana a 0, presenta una tendencia mucho más lineal que las
demás, por tal motivo, tanto los controles como las concentraciones utilizadas
presentaron este comportamiento, sin embargo los controles mostraron una linealidad
mayor a la de las concentraciones aplicadas en el fruto, (Figura 19) es decir que la
pérdida de peso de los frutos se ve afectada por las concentraciones de levadura
aplicadas, a través de los 45 días de estudio, en cada una de las replicas se presencio
la acción de C. guillermondii, de esta manera al ser comparadas las dos
concentraciones por medio de la linealidad, se determino que la levadura presenta
efectividad al ser aplicada a las concentraciones usadas. El estadístico de ANOVA
determinó que no existe diferencia significativa entre los pesos de cada una de las
replicas en cada uno de los días de estudio, lo que comprueba que los datos se
comportan normalmente. Según ANOVA normal existe una diferencia significativa
entre la pérdida de peso de los frutos control y los frutos con levadura, de igual
manera existe una diferencia significativa entre las dos concentraciones aplicadas en
los frutos, aunque la linealidad no mostro gran diferencia entre los tratamientos

(Figura 22), solo estadísticamente se pudo comprobar las diferencias significativas
entre estos.
0.00
10.00
20.00
30.00
40.00
50.00
60.00
70.00
0 3 6 9 12 15 18 21 24 27 30 33 36 39 42 45 48
Peso frutos (gram
os)
Tiempo(días)
PERDIDA DE PESO DE LOS FRUTOS EN FUNCIÓN DEL TIEMPO
CONCENTRACIÓN 1
CONCENTRACIÓN 2
CONTROLES
Figura 22. Relación de pérdida lineal de peso entre los frutos control y las
concentraciones.
Por medio de los resultados obtenidos estadísticamente se evidencio que tanto la
concentración 1 como la concentración 2 aplicada en el fruto, cumplen con el mismo
fin de retardar la maduración del fruto y de deshidratarlo a través del tiempo. Aunque,
por medio de la regresión lineal se evidencio que la concentración 1 presento una
linealidad menor, lo que muestra un mejor comportamiento en cuanto a la actividad
de la retardación del fruto por medio de C. guillermondii, ya que indica que luego de
los 18 días de aplicación de la levadura, se genera un mayor estado de reposo de la
perdida de peso del fruto, con respecto a los frutos control y los frutos en los cuales la
levadura fue aplicada a la concentración 2. Según el porcentaje de perdida de peso
del fruto a través del tiempo de estudio, los frutos control, y los frutos aplicados con
la levadura a concentración 1 perdieron mas peso a los 15 días de estudio, mientras
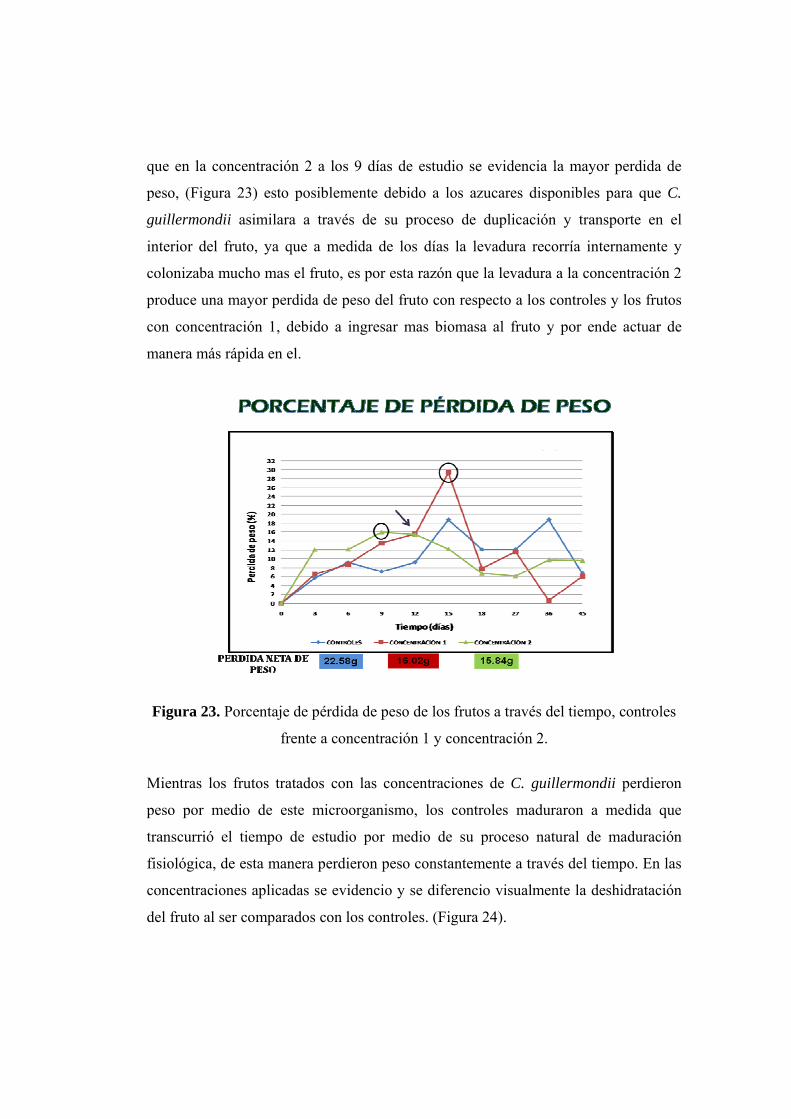
que en la concentración 2 a los 9 días de estudio se evidencia la mayor perdida de
peso, (Figura 23) esto posiblemente debido a los azucares disponibles para que C.
guillermondii asimilara a través de su proceso de duplicación y transporte en el
interior del fruto, ya que a medida de los días la levadura recorría internamente y
colonizaba mucho mas el fruto, es por esta razón que la levadura a la concentración 2
produce una mayor perdida de peso del fruto con respecto a los controles y los frutos
con concentración 1, debido a ingresar mas biomasa al fruto y por ende actuar de
manera más rápida en el.
Figura 23. Porcentaje de pérdida de peso de los frutos a través del tiempo, controles
frente a concentración 1 y concentración 2.
Mientras los frutos tratados con las concentraciones de C. guillermondii perdieron
peso por medio de este microorganismo, los controles maduraron a medida que
transcurrió el tiempo de estudio por medio de su proceso natural de maduración
fisiológica, de esta manera perdieron peso constantemente a través del tiempo. En las
concentraciones aplicadas se evidencio y se diferencio visualmente la deshidratación
del fruto al ser comparados con los controles. (Figura 24).

Figura 24. Estado de maduración del fruto control frente al fruto con levadura.

8. Conclusiones
• La cepa de levadura en estudio fue aislada de uchuva HIB cultivada en la
estación experimental de la Pontificia Universidad Javeriana.
• La levadura fue identificada mediante la técnica de API 20C AUX con un
99.7% de certeza de ser C. guillermondii
• Después de varias pruebas se descartó la presencia de tubo germinal
evidenciándose que no es una levadura patógena.
• La levadura C. guillermondii presenta su máximo crecimiento entre las 14
horas y 22 horas, su velocidad especifica de crecimiento es igual a 0.0908h-1,
y se duplica cada 7 horas y 37 minutos en caldo YGC.
• La actividad de la Levadura C1 (4.7 x 109 UFC/mL) y de la C2 (5.17 x 109
UFC/mL), generaron diferencias significativas, con respecto a la pérdida de
peso de los frutos durante el tiempo de estudio.
• Del estudio de las dos Concentraciones de levadura, se encontró que la C1
genera mayor pérdida de peso (16.02 %) produciendo un mayor efecto en la
retardación de la pudrición del fruto.
• C. guillermondii a partir de los 12 días de aplicación muestra la máxima
perdida de peso y el inicio de la deshidratación del fruto del banano.

9. Recomendaciones
• Buscar nuevas técnicas de identificación que confirmen que es C.
guillermondii aplicando técnicas o métodos de biología molecular.
• Realizar el estudio por un tiempo mayor a 45 días, para conocer más aspectos
importantes con respecto a la deshidratación del fruto.
• Cuantificar compuestos producidos por acción de la levadura en el fruto por
medio de cromatografía.
• Evaluar otras formas de aplicación de C. guillermondii diferentes a inmersión.
• Estudiar el efecto de la levadura en la retardación de la pudrición de frutos
diferentes al banano, al igual que el efecto de deshidratación.
• Estudiar el gradiente de pérdida de agua en el fruto durante el tiempo de
estudio

10. Referencias
Arnold, W.N. 1981. Yeast cell envelopes: biochemistry, biophysics and
ultrastructure. First Edition. Ed. CRC Press, Boca Raton, Floride. 129 p.
Barrera, R. 2000. Aislamiento e Identificación de Bacterias Proteolíticas y
Amilolíticas a partir de Aguas Residuales del Lavado del Café. Investigación en
Pedagogía DISTRALIA. 2:69-83
Beech, F.W., Davenport, R., Mossel, D., Dijkmann, K.E., Koopmans, M., Jong, J. &
Put, H. 1980. Media and methods for growing yeast: proceedings of a discussion
meeting. Second Edition. Ed. Academic Press, London. 259 p.
Berry, D.R. & Brown, C. 1987. Physiology of yeast growth. First edition. Ed. Allen
et Unwin, London. 159 p.
Biely, P. 1987. Changes in the rafe of synthesis of wall polysaccharides during the
cell cycle of yeast. Arch. Microbiol. 119: 213-236
Boulton, C. A. & Ratledge, C. 1984. The physiology of hydrocarbonutilizing
microorganisms. Top. Enz. Ferment. Biotech. 9: 11-39
Casas, G. 1989. Micología general. Ediciones de la biblioteca. Caracas. Pg 102-105.
Cayón, D. Giraldo, G. Arcila, M. 2000. Postcosecha y agroindustria del plátano en el
eje cafetero de Colombia. CORPOICA. Armenia, Quindio.
Chang-Yuen. 2005. Efecto del 1-metil-ciclopropeno (1-mcp) en la maduración de
banano. Agronomía Costarricense 29(3): 211-220. ISSN:0377-9424
Cook, A. 1985. The chemistry and biology of yeast. First Edition. Ed. Academic
Press, New York. 500 p.

Cooper, T. G. 1982. Nitrogen metabolism in S. cerevisiae. En: Strathern J., Jones, E.
W. & Broach, J. The molecular biology of the yeast Saccharomyces. First Edition.
Ed. Cold Spring Harbor Laboratories, New York. 399 p.
Erasmus D., Merwe G., Vuuren H. 2002. Genome-wide expresión analices metabolic
Adaptation of Saccharomyces cerevisiae to high sugar stress. FEMS yeast research.
3: 375-399.
Fiechter, A., Fuhrmann, G. F. & Kapelli, O. 1981. Regulation of glucose metabolism
in growing yeast cells. Adv. Microb. Physiol. 22: 123-140
Gancedo, C. & Serrano, R. 1989. The yeast: metabolism and physiology. Second
edition. Ed. Academic Press, London. 308 p.
Harrison, J.S. & Rose, A. 1983. The yeast. Second Edition. Ed. Academic Press,
London. 410 p.
Haettn, H. 1986. Bioquímica de las levaduras. Ed. Aguilar. Madrid, España. 250 p.
Henry, S. A. 1996. Membrane lipids of yeast. En: Strathern, J., Jones, E. & Broach,
R. The molecular biology of yeast Saccharomyces. Third edition. Ed. Cold Spring
Harbor, New York. 487 p.
Gallo, F. 1997. Manual de fisiología, patología postcosecha y control de calidad de
frutas y hortalizas. Convenio SENA-Reino Unido. Armenia, Colombia. Segunda
edición. 406 p.
Graham, F & Horman, J. 1990. Yeast, mould and plant protoplast. Ed. Academic
Press, London. 61p.
Jones, R. P. & Greenfields, P. E. 1981. A rewiew of yeast ionic nutrition. Process.
Biochem. 14: 48-52

Kirsop, B. 1987. Maintenance of yeast cultures. En: Berry, D. R., Russel, I. &
Stewart, G. G. Yeast Biotechnology. Second edition. Ed. Allen and Uwin, London.
250 p.
Kreger-Van Rij, N. 1984. The yeast: a taxonomic study. Third Edition. Ed. Elsevier
Science Publishers, Amsterdam. 1020 p.
Kulaev, I. S. & Vagabov, V. M. 1983. Polyphospate metabolism in micro-organisms.
Adv. Microbiol. Physiol. 24: 83-90
Lemos, L; Diaz, A. Rodríguez, A. 2006. Reutilización de soluciones concentradas
utilizadas en deshidratación-impregnación por inmersión (D.I.I.) de plátano maduro
(Musa paradisiaca). Universidad del valle. Dpto. de Ingeniería de Alimentos. Cali
Valle del Cauca.
Levi, D., Shennan, J. L.. & Ebbon, P. 1980. Biomass from liquid nalkanes. En: Rose,
A. Microbial Biomass. Second Edition. Ed. Academic Press, London. 93 p.
Linares, M. Solís, F. 2001. Identificación de Levaduras. Revista Iberoamericana de
Micología (En línea) http://www.guia.reviberoammicol.com/Capitulo11.pdf ISBN:
84-607-3050-6. [Consulta; 30 oct. 2007]
Lindegren, B. & Hadad, J. Kinetics of yeast. Ed. Standford-Wrisley. Liverpool,
England. 189 p.
López, C .Et al. 2005. Comparación de diferentes métodos para la identificación de
especies del género Candida. Revista Argentina de Microbiología 37: 16-21
Merck. 2000. Manual de Medios de Cultivo. Cuarta edición. Darmstadt, Alemania.
1010 p.

Nagamune, T., Indo, I. & Inone, I. 1981. The effect of medium composition on yeast
physiology activities for ethanol production. En: Moo-Young, M. Advances in
biotechnology. Third Edition. Ed. Pergamon Press, Oxford. 219 p.
Nikawa, J., Nagumo, I. & Yamashita, S. 1998. Inositol transport in Saccharomyces
cerevisiae. J. Bacteriol. 150: 441-449
Odds, F. 1988. Candida and candidosis. Segunda edición. Editorial Bailliere Tindall.
Londres. 468 p.
Orberá, T. 2004. Métodos moleculares de identificación de levaduras de interés
biotecnológico Centro de Estudios de Biotecnología Industrial, Santiago de Cuba.
Revista Iberoamericana de Micología; 21: 15-19 15
Ortegón, C. & Ramírez, C. 2001. Evaluación preliminar de los efectos de la
inoculación de una cepa de levadura extraída de tomates heteroinjertados sobre
variedades de tomates comerciales. Trabajo de grado (Bacteriólogo). Universidad
Javeriana. Facultad de Ciencias, Carrera de Bacteriología. Bogotá. 98 p.
Ovalle, J; Rueda, S. 1999. Manejo postcosecha del banano criollo (Musa spp.Grupo
AAA). Serie de paquetes de capacitación sobre maniejo postcosecha de frutas y
hortalizas No 2. Edición magnitud Ltda. Pereira. Segunda edición. 298 p.
Pelczar. 1994. Microbiología. Cuarta edición. Editorial Mc Graw Hill. México.
Port, G. 1990. Yeast. Third Edition. Ed. Press Pergamon. Michigan, USA. 510 p.
Rhan, M. 1991. Metabolism of yeast. Ed. Academic Press. London, England. 230 p.
Romero J. 2002. Efecto antagónico de C. Rugosa sobre microorganismos
contaminantes de la uchuva nativa Physalis peruviana. Microbiología Industrial.
Pontificia Universidad Javeriana. Facultad de Ciencias. Bogotá. 88 p.

Salisbury, Ross. 1994. Fisiología Vegetal. Ed. Iberoamérica. México D.C. 759 P.
Schneider, H., Wang, P. Y., Chan, Y. K. & Maleszka, R. 1982. Conversion of D-
xylose into ethanol by the yeast. Biotech. Lett. 3: 89-98
Schurr, A. & Yagil, E. 1981. Regulation and characterization of acid and alkaline
phosphatase in yeast. J. Gen. Microbiol. 65: 291-310
Sierra, L. 1993. Post cosecha y agroindustria del plátano en el eje cafetero de
Colombia. 145 p.
Stewart, G. G. & Russel, I. 1991. Biochemical and genetic control of sugar and
carbohydrate metabolism in yeast. First Edition. Ed. Allen and Uwin, London. 277 p
Stryer, L.1981.Biochemistry. First Edition. Ed. W. H. Freeman, San Francisco. 609 p.
Van Unden, N. & Buckley, A. Yeast. 1982. Fourth Edition. Ed. Prentice Hall, London. 1034 p.
Vargas, L. 2002. Caracterización morfológica y bioquímica de la levadura asociada a
tomates heteroinjertados. Microbiología Industrial. Pontificia Universidad Javeriana.
Facultad de Ciencias. Bogotá. 84 p.





























