Embed Size (px)
Citation preview

METANÁLISIS DE LOS EFECTOS DE LA ADICIÓN
DE FUENTES VEGETALES DE GRASA A LA DIETA
DE CABRAS LECHERAS SOBRE LA PRODUCCIÓN Y
LA COMPOSICIÓN DE LA LECHE
Roberto Pavón Vijande
Córdoba, año 2012.

METANÁLISIS DE LOS EFECTOS DE LA ADICIÓN DE FUENTES
VEGETALES DE GRASA A LA DIETA DE CABRAS LECHERAS SOBRE LA
PRODUCCIÓN Y LA COMPOSICIÓN DE LA LECHE
Trabajo Fin de Máster realizado por
Roberto Pavón Vijande
y dirigido por
Dr. Andrés L. Martínez Marín
para la superación del
Máster de Zootecnia y Gestión Sostenible: Ganadería Ecológica e Integrada
de la Universidad de Córdoba.
Córdoba, 12 de julio de 2012.

1
Metanálisis de los efectos de la adición de fuentes vegetales de grasa a la dieta
de cabras lecheras sobre la producción y la composi ción de la leche 1
Roberto Pavón Vijande
RESUMEN
El objetivo del presente trabajo fue realizar un metanálisis del efecto de la adición de
fuentes vegetales de grasa ricas en ácidos grasos insaturados a la dieta de cabras
lecheras sobre la producción y la composición de la leche. Un total de 21 experimentos
que incluyeron 74 tratamientos fueron utilizados en el estudio. Los tratamientos se
agruparon en: dietas sin grasa añadida (NOLIP) y dietas que incluyeron fuentes de
grasa ricas en ácido oleico (GO), ácido linoleico (GL) o ácido α-linolénico (GAL). Los
resultados productivos no se afectaron (P > 0,05) por la adición de grasa a la dieta
excepto el porcentaje de grasa láctea, que aumentó (P < 0,05) en GL, y la producción
de grasa láctea que aumentó (P < 0,05) en GO y GL respecto a NOLIP. Las tres clases
de grasa redujeron (P < 0,05) el contenido de ácidos grasos saturados de cadena
media de la grasa láctea y aumentaron (P < 0,05) el de C18:0 y C18:1-c9 en
comparación con NOLIP. Los tratamientos GL y GAL aumentaron (P < 0,05) el
contenido de C18:1-t11 y C18:2-c9,t11 en comparación con NOLIP y GO aunque el
efecto fue mayor (P < 0,05) en GL. En comparación con NOLIP, el contenido de C18:1-
t10 y la ratio C18:2-c9,c12/C18:3-c9,c12,c15 aumentaron (P < 0,05) en GL mientras
que GAL no afectó (P > 0,05) al contenido de C18:1-t10 y redujo (P < 0,05) la ratio
C18:2-c9,c12/C18:3-c9,c12,c15. Mediante ecuaciones de regresión lineal pudo
cuantificarse la respuesta de las variables estudiadas al consumo de cada una de las
tres clases de grasa. Los resultados obtenidos contribuyen al conocimiento de los
efectos de las grasas insaturadas incluidas en la dieta de las cabras lecheras sobre los
resultados productivos y el contenido de ácidos grasos de la grasa láctea y aportan
estimaciones cuantitativas de los mismos. Los resultados son consistentes con el
conocimiento del metabolismo ruminal y mamario de los ácidos grasos. Se confirmó
que las grasas ricas en ácido α-linolénico promueven los cambios más favorables de
los ácidos grasos de la grasa láctea desde el punto de vista de la salud humana de
acuerdo con los conocimientos actuales.
Palabras clave: cabras, leche, grasa, ácidos grasos.
1Un manuscrito derivado de este trabajo ha sido enviado a una revista de la categoría “Agriculture, Dairy & Animal Science” para su evaluación.

2
INTRODUCCIÓN
Los resultados de un experimento clásico permiten demostrar causas y efectos y
probar mecanismos y modos de acción pero las condiciones experimentales son
necesariamente limitadas y los resultados no pueden servir de base para extraer
conclusiones generalizadas. A menudo, los experimentos son repetidos por otros
autores para verificar la generalidad y la repetibilidad de las observaciones previas, lo
que resulta en la publicación de numerosos trabajos y la presentación de abundante
información sobre un mismo asunto. Clásicamente, la síntesis del conocimiento
científico sobre un tema en particular se ha circunscrito a la revisión cualitativa de los
resultados publicados. La principal limitación de este procedimiento es la subjetividad
ya que la opinión de los autores influye en la importancia concedida a los trabajos
revisados y los criterios para la inclusión de los mismos están pobremente definidos.
Por el contrario, el metanálisis es una herramienta estadística que permite la revisión
objetiva de la información procedente de trabajos de investigación aunque presenten
diferencias en su metodología (Sauvant et al., 2008).
El uso del metanálisis ha permitido extraer valiosas conclusiones cuando se ha
aplicado a experimentos de Nutrición Animal. Recientemente, Glasser et al. (2008)
publicaron un metanálisis del efecto de la inclusión de semillas oleaginosas en las
dietas de vacas sobre el contenido de ácidos grasos de la grasa láctea. No nos consta
que estudios similares se hayan realizado en cabras. La bibliografía indica que los
efectos observados en vacas no son trasladables a las cabras, ambas especies
muestran respuestas diferentes cuando se añaden fuentes de grasa similares a la dieta
(Chilliard et al., 2007).
El efecto de la grasa añadida a la dieta sobre la composición de la leche de las cabras
tiene relevancia económica para los ganaderos ya que perciben un precio por el litro de
leche entregada a la industria que es directamente proporcional a su contenido en
grasa y proteína, factores ambos relacionados con el rendimiento quesero (MARM,
2010). Por otro lado, la modificación de las proporciones de los ácidos grasos de la
grasa láctea tiene relevancia en cuanto a la creciente preocupación de los
consumidores por las cualidades saludables de los productos incluidos en su dieta. Es
bien conocido que la inclusión de fuentes vegetales de grasa en la dieta modifica el
contenido de ácidos grasos de la grasa láctea de las cabras (Chilliard et al., 2003). Los
cambios pueden ser considerados favorables desde el punto de vista de la salud
humana como la reducción del contenido de ácidos grasos saturados (AGS) de cadena

3
media (Ulbricht y Southgate, 1991) o de la relación entre los ácidos grasos
poliinsaturados (AGPI) de las series n-6 y n-3 (Simopoulos, 2008), y el aumento de los
ácidos vaccénico (C18:1-t11; Field et al., 2009) y ruménico (C18:2-c9,t11; Benjamin y
Spener, 2009). Sin embargo, también pueden ocurrir cambios indeseables como el
aumento del isómero C18:1-t10 cuya presencia en las plaquetas se ha asociado con el
riesgo de enfermedad cardiovascular (Hodgson et al., 1996). Los efectos observados
parecen ser dependientes del ácido graso mayoritario en la fuente de grasa (Martínez
Marín et al., 2012a) y del tipo y cantidad de forraje de la dieta (Mele et al., 2008;
Bernard et al., 2009b). El metanálisis de los trabajos experimentales disponibles
permitiría esclarecer la respuesta a los factores mencionados y cuantificar su efecto.
El objetivo de este trabajo fue investigar los efectos de la adición de fuentes vegetales
de grasa ricas en AGI a la dieta de cabras lecheras sobre la producción y composición
de la leche y el contenido de ácidos grasos de la grasa láctea mediante metanálisis.
MATERIAL Y MÉTODOS
Creación de la base de datos
Los trabajos de investigación referidos a la inclusión de fuentes vegetales de grasa en
la dieta de cabras lecheras se buscaron en bases de datos bibliográficas (ISI Web of
Science, Scopus, Pubmed, Google Scholar) utilizando como palabras clave “grasas
vegetales”, “cabras”, “leche”, “grasa” y “ácidos grasos” en los idiomas español, inglés,
francés y portugués. Se seleccionaron solamente aquellos trabajos en los que un
tratamiento control con una dieta sin grasa añadida se comparó con uno o más
tratamientos en los que la dieta fue enriquecida con una única fuente de grasa vegetal
no protegida (semilla o aceite) y rica en ácidos grasos insaturados (AGI). Se
encontraron 18 trabajos de investigación con 21 experimentos y un total de 74
tratamientos (Tabla I). La información referente a los experimentos y sus resultados se
almacenó en una hoja de cálculo de Excel (Microsoft Corp., Redmon, WA).
Codificación de los factores experimentales
Numerosos datos de cada experimento fueron recogidos en la hoja de cálculo (Tabla I).
Los tratamientos se codificaron como NOLIP, cuando la dieta no incluyó ninguna fuente
de grasa añadida, y GO, GL o GAL cuando la dieta incluyó fuentes de grasa ricas en
ácido oleico, ácido linoleico o ácido α-linolénico, respectivamente. Se distinguieron dos

4
grupos de fuentes de grasa según la forma de presentación -semilla o aceite-,
especificando su origen botánico. Los factores relacionados con los animales que se
codificaron fueron la raza -Saanen, Alpina, Murciano, Malagueña y cruces-, el peso
vivo (PV), la fase de la lactación -inicio <90 d, mitad 90 a 180 d y final >180 d- y el
número de partos -primíparas o multíparas-. También se anotó el diseño experimental,
especificando la duración de los tratamientos y el número de animales utilizados, y la
composición de las dietas (porcentaje de forraje, proteína bruta y fibra
neutrodetergente -FND- expresado sobre materia seca -MS-). Por último se incluyeron
los porcentajes de grasa añadida en cada experimento y su contenido de ácidos grasos
mayoritarios.
Tras un análisis preliminar de los datos disponibles se decidió que la investigación del
efecto de la grasa añadida a la dieta debería hacerse teniendo en cuenta el ácido
graso mayoritario presente en la misma por ser este el factor determinante de los
efectos observados, especialmente sobre el contenido de ácidos grasos de la grasa
láctea (Chilliard y Ferlay, 2004). Por otro lado, se observó que no existía suficiente
número de datos para el estudio de la influencia de otros factores.
Las variables estudiadas fueron el consumo de materia seca (CMS), la producción y la
composición de la leche, y el contenido de ácidos grasos relevantes de la grasa láctea.
Debido a las diferencias en la presentación de los resultados referidos a estos últimos,
los valores utilizados de los ácidos oleico (C18:1-c9), C18:1-t10, vaccénico, linoleico
(C18:2-c9,c12), ruménico y α-linolénico (C18:3-c9,c12,c15) correspondieron
exclusivamente a los trabajos experimentales en los que se ofreció un perfil detallado
de los ácidos grasos identificados o, en caso de tablas resumidas, la técnica de
cromatografía de gases empleada (longitud de la columna capilar y método de análisis
de los ésteres metílicos) se consideró de resolución suficiente para la separación de
los diferentes isómeros de cada grupo.
Análisis estadístico
Los análisis estadísticos se hicieron con el programa SAS 9.1 (SAS Institute Inc.,
2004). En todos los análisis se utilizó el procedimiento MIXED. Los datos se
ponderaron utilizando la raíz cuadrada del número de animales usados en cada
tratamiento mediante la declaración WEIGHT. El nivel de significación se estableció en
P < 0,05.

5
Primero se analizaron las características de las dietas de cada tratamiento y el
consumo de grasa añadida para descartar estos factores como fuentes de
interferencia. A continuación se analizó el efecto de cada clase de grasa añadida a la
dieta sobre las variables consideradas en el estudio. Mediante la declaración
CONTRAST se compararon las medias de mínimos cuadrados del tratamiento NOLIP
con las de los tratamientos GO, GL y GAL, y las de estos entre sí. El modelo mixto
lineal utilizado fue (Sauvant et al., 2008):
Yijk = µ + Ѕi + Тj + ЅТij + еijk
Donde:
Yijk = variable dependiente
µ = media general
Ѕi = efecto aleatorio del i-ésimo trabajo experimental
Тj = efecto fijo del j-ésimo nivel del tratamiento
ЅТij = interacción aleatoria entre el i-ésimo trabajo experimental y el j-ésimo nivel
del tratamiento
еijk = error residual
En segundo lugar se realizaron análisis de regresión (metarregresión) con objeto de
investigar la relación entre las variables y el consumo de grasa añadida. Para poder
separar los efectos debidos al ácido graso mayoritario en la grasa extra, los
experimentos fueron agrupados por la clase de grasa. El análisis gráfico de los
datos (dispersión de los valores en un gráfico de coordenadas) mostró que, de
existir, las únicas relaciones entre las variables dependientes y el consumo de
grasa eran lineales, por lo que solamente este tipo de relación fue investigada. El
trabajo experimental se incluyó en la declaración CLASS y no contuvo ninguna
información cuantitativa. El residual estudentizado y la distancia de Cook de cada
observación se utilizaron para identificar los valores anómalos. Para cada ecuación
obtenida se calculó la raíz cuadrada del error cuadrático medio (RECM) y el
coeficiente de determinación (R2). El modelo mixto lineal utilizado fue (St-Pierre,
2001):
Yij = В0 + Ѕi + В1Хij + biХij + еij

6
Tabla I. Resumen de los experimentos incluidos en el metanálisis.
Experimentos Clase de grasa
Fuente de grasa
Grasa añadida
Raza Fase de lactación
Número de partos
Diseño experimental
Días de tratamiento
Número de animales
Forraje % MS
Proteína % MS
FND % MS
Autor
1
NOLIP 0,0
Alpina Final Primíparas Cuadrado
latino (4x4)
28 4
51,1 15,0 35,3
Mir et al. (1999) & Okine et al.
(2003) GO A. colza
1,1 51,1 15,0 35,3
2,2 51,1 15,0 35,3
3,2 51,1 15,0 35,3
2
NOLIP 0,0
Saanen Inicio Multíparas Cuadrado
latino (4x4)
14 4
50,0 16,0 32,6
Mouro et al. (2002) &
Maia et al. (2006)
GO A. arroz 5,1 50,0 16,0 30,8
GO A. colza 5,1 50,0 16,0 30,8
GL A. soja 5,1 50,0 16,0 31,1
3
NOLIP 0,0
Alpina Inicio - Cuadrado
latino (3x3)
21 14
54,0 15,0 46,5
Bernard et al. (2005)
GAL S. lino 4,3 52,1 16,7 45,5
GO A. girasol alto oleico
3,6 55,4 15,7 46,0
4 NOLIP 0,0
Alpina - Multíparas Cuadrado
latino (4x4)
18 16 67,1 11,1 28,5
Lana et al. (2005) GL A. soja 4,0 67,2 11,9 27,9
5
NOLIP 0,0
Saanen &
Alpina Mitad Multíparas 6 grupos
(3x2) 91 6
51,0 18,4 42,5
Schmidely et al. (2005)
GL S. soja
extrusionada
2,1 51,0 18,4 41,5
4,3 51,0 19,1 39,9
NOLIP 0,0 50,0 18,2 42,1
GL S. soja
extrusionada
2,1 50,0 18,2 41,1
4,3 50,0 18,9 39,5
NOLIP, GO, GL y GAL: dietas sin grasa añadida o enriquecidas con ácido oleico, ácido linoleico y ácido α-linolénico, respectivamente. A., aceite; FND, fibra neutrodetergente; MS, materia seca; S., semilla.

7
Tabla I (continuación). Resumen de los experimentos incluidos en el metanálisis.
NOLIP, GO, GL y GAL: dietas sin grasa añadida o enriquecidas con ácido oleico, ácido linoleico y ácido α-linolénico, respectivamente. A., aceite; FND, fibra neutrodetergente; MS, materia seca; S., semilla.
Experimentos Clase de grasa
Fuente de grasa
Grasa añadida Raza Fase de
lactación Número de
partos Diseño
experimental Días de
tratamiento Número de animales
Forraje % MS
Proteína % MS
FND % MS Autor
6
NOLIP 0,0 Alpina
x Sarda
Mitad - 3 grupos 21 10
40,4 17,5 38,5 Nudda et al.
(2006) GAL S. lino
extrusionada
0,8 40,4 17,9 38,6
1,7 40,4 18,3 38,7
7
NOLIP 0,0
Saanen &
Alpina Inicio Multíparas 4 grupos
(2x2) 84 8
55,0 12,6 44,0
Andrade y Schmidely
(2006a)
GO S. colza aplastada 1,8 55,0 12,5 44,7
NOLIP 0,0 35,0 13,4 38,4
GO S. colza aplastada
3,7 35,0 13,3 38,1
8
NOLIP 0,0
-
Inicio Multíparas 4 grupos
(2x2) 77 2
55,0 12,4 50,0
Andrade y Schmidely
(2006b)
GO S. colza aplastada
1,8 55,0 12,4 47,0
NOLIP 0,0 35,0 13,3 37,6
GO S. colza aplastada
3,7 35,0 13,4 41,9
9
NOLIP 0,0
- Inicio - 4 grupos 86 6
40,8 17,9 43,6
Silva (2005) GL A. soja 4,5 40,8 16,9 42,8
GL S. soja 4,7 40,8 16,7 45,7
10 NOLIP 0,0
Murciana Inicio Multíparas
& primíparas
Doble reverso 28 24 58,0 17,3 36,5 Bouattour et al.
(2008) GL A. soja 2,5 58,0 17,5 35,4

8
Tabla I (continuación). Resumen de los experimentos incluidos en el metanálisis.
Experimentos Clase
de grasa
Fuente de grasa
Grasa añadida Raza Fase de
lactación Número de
partos Diseño
experimental Días de
tratamiento Número de animales
Forraje % MS
Proteína % MS
FND % MS Autor
11
NOLIP 0,0
Saanen Inicio Multíparas Cuadrado
latino (4x4)
21 12
63,0 15,9 43,1
Mele et al. (2008)
GL A. soja 4,0 63,0 15,5 42,5
NOLIP 0,0 35,0 16,0 37,7
GL A. soja 4,0 35,0 15,7 37,2
12
NOLIP 0,0
Saanen Inicio Multíparas Cuadrado
latino (4x4)
28 4
40,0 17,8 34,0
Almeida (2008) GL A. soja 2,0 40,0 17,8 34,0
GAL A. lino 2,0 40,0 17,8 34,0
13
NOLIP 0,0
Alpina Inicio Multíparas Cuadrado
latino (3x3)
28 13
44,4 16,7 42,1
Bernard et al. (2009b)
GL A. girasol 5,5 48,4 18,3 40,7
GAL A. lino 5,5 48,6 18,3 40,6
14
NOLIP 0,0
Alpina Inicio Multíparas Cuadrado
latino (3x3)
28 14
38,8 14,0 31,0
Bernard et al. (2009b)
GL A. girasol 6,1 45,5 17,1 29,5
GAL A. lino 6,2 45,2 17,1 29,4
15 NOLIP 0,0
Saanen Final Multíparas Cuadrado
latino (4x4)
14 4 20,0 14,6 34,1 Li et al.
(2009) GL A. soja 4,0 20,0 14,0 33,3
16
NOLIP 0,0
Alpina Inicio Multíparas Cuadrado
latino (4x4)
28 16
63,9 19,2 37,8
Ollier et al. (2009)
GO S. colza 6,5 74,4 20,7 42,0
NOLIP 0,0 42,8 16,9 30,2
GL A. girasol 4,4 48,7 18,5 32,4
NOLIP, GO, GL y GAL: dietas sin grasa añadida o enriquecidas con ácido oleico, ácido linoleico y ácido α-linolénico, respectivamente. A., aceite; FND, fibra neutrodetergente; MS, materia seca; S., semilla.

9
Tabla I (final). Resumen de los experimentos incluidos en el metanálisis.
Experimentos Clase
de grasa
Fuente de grasa
Grasa añadida
Raza Fase de lactación
Número de partos
Diseño experimental
Días de tratamiento
Número de animales
Forraje % MS
Proteína % MS
FND % MS
Autor
17
NOLIP 0,0
Malagueña Inicio Multíparas Cuadrado
latino (4x4)
21 16
30,0 18,4 26,9
Martínez Marín et al. (2011a)
GO A. girasol alto oleico
3,0 30,0 17,4 25,8
GL A. girasol 3,0 30,0 17,5 25,5
GAL A. lino 3,0 30,0 17,7 25,5
18
NOLIP 0,0
Malagueña Inicio Multíparas 4 grupos 15 3
30,0 16,7 28,1
Martínez Marín et al. (2012a) GAL A. lino
1,8 30,0 16,3 26,9
2,9 30,0 16,3 26,9
3,9 30,0 15,9 26,1
19
NOLIP 0,0
Malagueña Inicio Multíparas 4 grupos 15 3
30,0 16,7 28,1
Martínez Marín et al. (2012a) GO A. girasol
alto oleico
1,8 30,0 16,2 27,5
2,9 30,0 15,9 27,5
3,9 30,0 16,0 27,1
20
NOLIP 0,0
Malagueña Mitad Multíparas 4 grupos 15 3
30,0 16,7 28,1
Martínez Marín et al. (2012a) GL A. girasol
1,8 30,0 16,3 27,7
2,9 30,0 16,2 27,1
3,9 30,0 16,0 27,2
21
NOLIP 0,0
Malagueña Inicio Multíparas 4 grupos 21 3
30,0 17,0 28,2
Martínez Marín et al. (2012b,c)
GO A. girasol alto oleico 2,9 30,0 16,4 27,5
GL A. girasol 2,9 30,0 16,4 27,0
GAL A. lino 2,9 30,0 16,5 26,9
NOLIP, GO, GL y GAL: dietas sin grasa añadida o enriquecidas con ácido oleico, ácido linoleico y ácido α-linolénico, respectivamente. A., aceite; FND, fibra neutrodetergente; MS, materia seca.

10
Donde:
Yij = variable dependiente
В0 = intersección total entre tratamientos
Ѕi = efecto aleatorio del i-ésimo trabajo experimental
В1 = coeficiente de regresión lineal de Y sobre X
Хij = variable predictora continua (consumo de grasa, efecto fijo)
bi = efecto aleatorio del i-ésimo trabajo experimental en el coeficiente de regresión
de Y sobre X
еij = error residual
La sintaxis de los programas SAS utilizados en cada uno de los modelos se muestra en
el Apéndice.
RESULTADOS
Fuentes de grasa y dietas
En la Tabla II se muestra la composición de las fuentes de grasa utilizadas en los
experimentos. De los 74 tratamientos, 26 correspondieron a dietas sin grasa añadida,
16 a dietas enriquecidas en ácido oleico, 21 a dietas enriquecidas en ácido linoleico y
11 a dietas enriquecidas en ácido α-linolénico. Las fuentes de grasa consistieron en
aceites (35 tratamientos de los que 16 aportaron ácido linoleico, 11 aportaron ácido
oleico y 8 aportaron ácido α-linolénico) y semillas (13 tratamientos de los que 5
aportaron ácido linoleico, 5 aportaron ácido oleico y 3 aportaron ácido α-linolénico).
Tabla II. Contenido en ácidos grasos mayoritarios (media ± desviación típica y rango, en porcentaje de los ácidos grasos totales) de las fuentes de grasa utilizadas en los experimentos.
Ácidos grasos
Fuentes de grasa
GO GL GAL
C18:1-c9 67,3 ± 14,1
[43-85] 23,6 ± 3,3
[18-31] 19,5 ± 2,3
[14-21]
C18:2-c9,c12 16,4 ± 8,8
[5-35] 56,9 ± 4,8
[51-70] 17,0 ±2,5 [15-24]
C18:3-c9,c12,c15 5,0 ± 4,3
[0-9] 4,1 ± 3,7
[0-8] 51,1 ± 4,3
[42-59]
GO, GL y GAL, fuentes de grasa ricas en ácido oleico, ácido linoleico, y ácido α-linolénico, respectivamente.

11
La Tabla III muestra la composición de las dietas. No hubo diferencias significativas (P
> 0,05) en el contenido de forraje, proteína bruta y FND entre los tratamientos.
Tampoco hubo diferencias en el contenido de grasa añadida entre GO, GL y GAL.
Tabla III. Composición de las dietas (% de la materia seca) incluidas en el metanálisis.
Componentes
Tratamientos
EEM P NOLIP GO GL GAL
Forraje 42,80 46,49 42,74 42,91 1,44 0,571
Proteína bruta 15,98 16,20 16,18 16,64 0,23 0,312
Fibra neutrodetergente 35,32 36,22 34,26 34,49 0,78 0,201
Grasa añadida - 3,61 3,48 3,53 0,21 0,956
NOLIP, GO, GL y GAL: dietas sin grasa añadida o enriquecidas con ácido oleico, ácido linoleico y ácido α-linolénico, respectivamente.
Consumo de materia seca y producción y composición de la leche
En la Tabla IV se muestran los efectos de la adición de grasa extra a la dieta sobre el
CMS y la producción y la composición de la leche. La adición de grasa a la dieta no
tuvo efectos (P > 0,05) sobre el CMS, la producción de leche ni el contenido y la
producción de proteína láctea. El porcentaje de grasa fue mayor (P < 0,05) en GL en
comparación con NOLIP pero no hubo diferencias (P > 0,05) entre los tratamientos con
grasa añadida. Los tratamientos GO y GL aumentaron (P < 0,05) la producción de
grasa en comparación con NOLIP pero no hubo diferencias (P > 0,05) entre los
tratamientos con grasa añadida
Contenido de ácidos grasos de la grasa láctea
Los efectos de la adición de grasa extra a la dieta sobre el contenido de ácidos grasos
de la grasa láctea se muestran en la Tabla IV. Los tratamientos con grasa añadida
tuvieron los mismos efectos sobre el contenido de los AGS de cadena corta (4 a 8
carbonos) y media (10 a 16 carbonos) de la grasa láctea en comparación con NOLIP:
aumento (P < 0,05) de C4:0, ninguna modificación (P > 0,05) de C6:0 y C8:0, y
disminución (P < 0,05) de C10:0, C12:0, C14:0 y C16:0. La suma de los contenidos de
los AGS de cadena media en los tratamientos con grasa añadida se redujo un 24%
respecto al valor de NOLIP.

12
Tabla IV. Efectos de la adición de grasa extra a la dieta sobre la producción y composición de la leche y el contenido de ácidos grasos de la grasa láctea de cabras.
Variables n
Tratamientos
EEM P NOLIP GO GL GAL
Consumo de materia seca, g/d 74 2110 2089 2050 2069 71 0,363
Producción de leche, g/d 74 2335 2387 2355 2314 111 0,861
Grasa, % 70 4,00 4,24 4,42A 4,22 0,12 0,003
Grasa, g/d 70 87 97A 96A 91 4 0,019
Proteína, % 74 3,24 3,19 3,30 3,27 0,04 0,212
Proteína, g/d 74 74 73 75 73 3 0,770
Ácidos grasos, % ácidos grasos totales
C4:0 55 2,098 2,248A 2,233A 2,291A 0,109 <0,001
C6:0 61 2,297 2,261 2,256 2,332 0,097 0,730
C8:0 61 2,744 2,559 2,568 2,765 0,114 0,127
C10:0 67 10,068 8,267A 8,120A 8,807A 0,227 <0,001
C12:0 67 5,169 3,563A 3,473A 3,851A 0,134 <0,001
C14:0 67 11,304 8,813A 8,611A 8,799A 0,214 <0,001
C16:0 67 30,889 23,053A 23,389A 22,897A 0,648 <0,001
C18:0 67 6,952 12,587Aa 11,124Ab 10,208Ab 0,414 <0,001
C18:1-c9 56 15,003 21,821Aa 17,624Ab 17,240Ab 0,407 <0,001
C18:1-t10 45 0,293 0,562b 1,106Aa 0,491b 0,093 0,009
C18:1-t11 45 0,940 1,732c 6,068Aa 4,113Ab 0,391 <0,001
C18:2-c9,c12 67 2,371 2,055Ab 2,978Aa 2,198b 0,090 <0,001
C18:2-c9,t11 50 0,593 0,756c 2,699Aa 1,746Ab 0,155 <0,001
C18:3-c9,c12,c15 67 0,469 0,468b 0,420b 1,047Aa 0,047 <0,001
C18:2-c9,c12/C18:3-c9,c12,c15 67 6,945 6,111b 10,650Aa 1,520Ac 0,596 <0,001
NOLIP, GO, GL y GAL: dietas sin grasa añadida o enriquecidas con ácido oleico, ácido linoleico y ácido α-linolénico, respectivamente. ADentro de una fila, las medias de los tratamientos GO, GL y GAL con el superíndice son diferentes (P < 0,05) de la media del tratamiento NOLIP. a,b,cDentro de una fila, las medias de los tratamientos GO, GL y GAL que no tienen un superíndice común son diferentes (P < 0,05) entre sí.

13
Los tratamientos con grasa añadida aumentaron (P < 0,05) el contenido de C18:0 y
C18:1-c9 en comparación con NOLIP pero el aumento fue mayor (P < 0,05) en GO
comparado con GL y GAL, que fueron iguales entre sí.
El tratamiento GL aumentó el contenido de C18:1-t10 en comparación con los demás
tratamientos hasta un valor que fue 277% mayor que el de NOLIP. El contenido de
C18:1-t11 fue mayor (P < 0,05) en GL y GAL en comparación con NOLIP, que fue igual
(P > 0,05) a GO. Dentro de los tratamientos con grasa añadida el contenido de C18:1-
t11 fue un 47% mayor (P < 0,05) en GL comparado con GAL.
El contenido de C18:2-c9,c12 fue mayor (P < 0,05) en GL, menor (P < 0,05) en GO y
no cambió (P > 0,05) en GAL en comparación con NOLIP. Además GL resultó en el
mayor contenido (P < 0,05) de C18:2-c9,c12 entre los tratamientos con grasa añadida y
no hubo diferencias (P > 0,05) entre GO y GAL. El contenido de C18:2-c9,t11 mostró
unos resultados paralelos a los observados en C18:1-t11: el contenido fue mayor en
GL y GAL en comparación con NOLIP, que fue igual (P > 0,05) a GO, a su vez, el
contenido fue un 55% mayor (P < 0,05) en GL comparado con GAL.
El contenido de C18:3-c9,c12,c15 de la grasa láctea únicamente aumentó (P < 0,05) en
GAL. Este aumento y los cambios observados en el contenido de C18:2-c9,c12
tuvieron como resultado un aumento (P < 0,05) de la ratio de ambos ácidos grasos en
GL y una disminución (P < 0,05) en GAL en comparación con NOLIP y GO, que fueron
iguales entre sí (P > 0,05). El valor de la ratio fue un 53% mayor y un 78% menor en
GL y GAL, respectivamente, respecto a NOLIP.
Metarregresiones
El consumo de grasa añadida fue similar en GO, GL y GAL (media ± EEM y rango, en
g/d): 83 ± 13 [29-207], 79 ± 8 [28-132] y 63 ± 12 [15-126], respectivamente. Las
estimaciones de los parámetros de las ecuaciones obtenidas se muestran en las
Tablas V, VI y VII. No se observaron relaciones lineales significativas entre el CMS, la
producción de leche y el contenido y la producción de proteína láctea, y el consumo de
ninguna de las clases de grasa. El porcentaje de grasa láctea aumentó linealmente (P
< 0,05) en respuesta al consumo de grasa en GO y GL, mientras que la producción de
grasa únicamente lo hizo (P < 0,05) en GO y GAL.

14
El efecto de los tratamientos con grasa añadida sobre el porcentaje de los AGS de
cadena corta y media de la grasa láctea fue similar. El consumo de grasa aumentó
linealmente (P < 0,05) el contenido de C4:0, no afectó (P > 0,05) al contenido de C6:0
y redujo de forma lineal (P < 0,05) el contenido de C10:0, C12:0, C14:0 y C16:0.
Únicamente el consumo de grasa en GL tuvo un efecto lineal negativo (P < 0,05) sobre
el contenido de C8:0 (Tabla VI). El valor de la pendiente de las ecuaciones obtenidas
para C16:0 fue más del doble de los valores de las pendientes de las ecuaciones
obtenidas para los restantes AGS de cadena media.
El consumo de grasa en GO tuvo un efecto lineal positivo (P < 0,05) sobre el contenido
de C18:0, C18:1-c9 y C18:1-t11 (Tabla V). El consumo de grasa en GL aumentó de
manera lineal (P < 0,05) el contenido de C18:0, C18:1-c9, C18:1-t10, C18:1-t11, C18:2-
c9,c12 y C18:2-c9,t11 y la ratio C18:2-c9,c12/C18:3-c9,c12,c15 (Tabla VI). El consumo
de grasa en GAL aumentó linealmente (P < 0,05) el contenido de C18:0, C18:1-t11,
C18:2-c9,t11, C18:3-c9,c12,c15 y redujo linealmente (P < 0,05) la ratio C18:2-
c9,c12/C18:3-c9,c12,c15 (Tabla VII). Las pendientes de las ecuaciones obtenidas en
GO para los ácidos esteárico y oleico tuvieron valores que fueron aproximadamente
50% y 90% mayores que las obtenidas en GL y GAL. Hubo un 12% de diferencia entre
las pendientes de las ecuaciones obtenidas en GL y GAL para el ácido vaccénico. La
pendiente de la ecuación obtenida para el ácido ruménico fue un 43% mayor en GL
que en GAL.
DISCUSIÓN
Metadiseño
El número de experimentos realizados con cabras para investigar el efecto de la
adición de fuentes vegetales de grasa ricas en AGI a la dieta sobre la producción y la
composición de la leche y el contenido de ácidos grasos de la grasa láctea es limitado
en comparación con los realizados con vacas. En el presente trabajo hubo más datos
disponibles para el análisis de la producción y la composición de la leche que para el
del contenido de ácidos grasos relevantes de la grasa láctea. Además, el número de
datos disponibles para este último fue muy diferente según el ácido graso investigado.
En particular el número de experimentos que reportaron el contenido de C18:1-t10,
C18:1-t11 y C18:2-c9,t11 de la grasa láctea fue muy limitado. No obstante, los trabajos
disponibles permitieron que todos los análisis incluyeran diez datos como mínimo.

15
Tabla V. Ecuaciones de regresión lineal del efecto del consumo de grasa rica en ácido oleico añadida a la dieta (expresado en g/d) sobre la producción y la composición de la leche y el contenido de ácidos grasos de la grasa láctea de cabras.
Variables n
Estimaciones de los parámetros
Intersección EEM P Pendiente EEM P RECM R2
Consumo de materia seca, g/d 27 2304 261 <0,001 0,1 0,6 0,841 245 0,96
Producción de leche, g/d 27 2523 322 <0,001 1,0 1,5 0,527 486 0,90
Grasa, % 24 4,38 0,40 <0,001 0,003 0,001 0,029 0,54 0,90
Grasa, g/d 24 98 8 <0,001 0,2 0,1 0,022 14 0,89
Proteína, % 27 3,26 0,12 <0,001 -0,0003 0,001 0,543 0,23 0,82
Proteína, g/d 27 81 10 <0,001 -0,02 0,001 0,584 13 0,92
Ácidos grasos, % de los ácidos grasos totales
C4:0 27 1,866 0,260 <0,001 0,0013 0,0005 0,016 0,20 0,96
C6:0 27 2,271 0,219 <0,001 -0,0002 0,0006 0,696 0,23 0,93
C8:0 27 2,589 0,178 <0,001 -0,0016 0,0008 0,063 0,33 0,83
C10:0 27 10,150 0,411 <0,001 -0,0209 0,0048 0,003 0,95 0,88
C12:0 27 5,230 0,246 <0,001 -0,0207 0,0033 <0,001 0,56 0,90
C14:0 27 11,364 0,351 <0,001 -0,0339 0,0056 <0,001 0,67 0,94
C16:0 27 30,061 0,762 <0,001 -0,0945 0,0157 <0,001 3,40 0,79
C18:0 27 6,891 0,438 <0,001 0,0771 0,0147 0,001 1,45 0,95
C18:1-c9 20 15,221 0,450 <0,001 0,0909 0,0193 0,002 2,29 0,85
C18:1-t10 18 0,244 0,032 0,001 0,0042 0,0017 0,056 0,09 0,92
C18:1-t11 18 0,780 0,112 <0,001 0,0159 0,0052 0,022 0,53 0,82
C18:2-c9,c12 27 2,262 0,233 0,001 -0,0025 0,0012 0,064 0,18 0,81
C18:2-c9,t11 20 0,511 0,074 <0,001 0,0034 0,0015 0,206 0,34 0,89
C18:3-c9,c12,c15 27 0,459 0,096 0,001 0,0002 0,0006 0,714 0,16 0,87
C18:2-c9,c12/C18:3-c9,c12,c15 27 7,189 1,502 0,001 -0,0062 0,0042 0,160 1,70 0,94
RECM, raíz cuadrada del error cuadrático medio; R2, coeficiente de determinación.

16
El contenido de los ácidos oleico, linoleico y α-linolénico de las tres clases de grasa fue
claramente diferente entre ellas (Tabla II) mientras que el porcentaje medio de grasa
añadida en los tratamientos GO, GL y GAL (Tabla IV) y su consumo (1,32 ± 0,72 1,31 ±
0,72 y 1,05 ± 0,60 g/kg PV en GO, GL y GAL, respectivamente) fue similar. Esto
probablemente contribuyó a las diferencias observadas entre tratamientos debidas al
ácido graso mayoritario en cada uno de ellos y redujo la influencia de la cantidad de
grasa consumida sobre los resultados obtenidos.
Debido al insuficiente número de datos dentro de cada clase, al realizar los análisis no
fue posible tener en cuenta la influencia de los factores propios de la fuente de grasa,
como su presentación y procesado, y de los factores dependientes del animal, como la
fase de lactación, o de la dieta, como el contenido de forraje o FND. No obstante, la
interferencia debida a la composición de la dieta en los resultados obtenidos, si existió,
debió ser mínima ya que no se observaron diferencias entre los tratamientos (Tabla III).
Consumo de materia seca y producción y composición de la leche
El hecho de que la adición de grasa extra a la dieta no tuviera efecto negativo sobre el
CMS (Tabla IV) indica que no ocurrió ralentización del vaciado ruminal por alteración
de la digestión ruminal de las paredes vegetales o por efectos metabólicos (Chilliard et
al., 1993) dentro del rango de consumo de grasa estudiado. De acuerdo con Martínez
Marín et al. (2011b), el efecto negativo de la adición de grasa extra a la dieta sobre la
digestión de la FND es más probable cuando el nivel de inclusión es superior a 4% MS.
En el presente trabajo, 14 tratamientos contenían más de 4% de grasa añadida y
solamente en 6 de ellos se encontraron diferencias significativas del CMS respecto a
los correspondientes controles (Bernard et al., 2005; Silva, 2005; Bernard et al., 2009b,
experimento 2). En cuanto al efecto metabólico de la grasa añadida sobre el CMS, los
trabajos de Teh et al. (1994) y Brown-Crowder et al. (2001) sugieren que dicho efecto
comienza a manifestarse cuando el consumo de grasa es superior a 2,5 g/kg PV, valor
que solo se alcanzó en un tratamiento (Ollier et al., 2009).
Los efectos observados sobre la producción y la composición de la leche (Tabla IV)
coinciden con lo señalado por Chilliard et al. (2003). Las causas por las cuales la
mayor concentración energética de las dietas de los tratamientos con grasa añadida
(0,69 ± 0,30 MJ energía neta de lactación/kg MS, respecto a NOLIP en este trabajo) no
repercutió positivamente sobre la producción de leche en ausencia de reducción del

17
Tabla VI. Ecuaciones de regresión lineal del efecto del consumo de grasa rica en ácido linoleico añadida a la dieta (expresado en g/d) sobre la producción y la composición de la leche y el contenido de ácidos grasos de la grasa láctea de cabras.
Variables n
Estimaciones de los parámetros
Intersección EEM P Pendiente EEM P RECM R2
Consumo de materia seca, g/d 37 1984 136 <0,001 -0,4 0,3 0,224 126 0,98
Producción de leche, g/d 37 2381 238 <0,001 -0,2 0,9 0,845 206 0,98
Grasa, % 35 4,04 0,31 <0,001 0,004 0,001 0,008 0,58 0,89
Grasa, g/d 35 87 8 <0,001 0,1 0,05 0,060 16 0,91
Proteína, % 37 3,26 0,10 <0,001 0,0004 0,0004 0,361 0,20 0,85
Proteína, g/d 37 74 7 <0,001 0,01 0,03 0,635 7 0,98
Ácidos grasos, % de los ácidos grasos totales
C4:0 22 2,251 0,309 <0,001 0,0013 0,0005 0,018 0,19 0,99
C6:0 25 2,369 0,298 <0,001 -0,0010 0,0007 0,153 0,26 0,97
C8:0 25 2,918 0,328 <0,001 -0,0030 0,0011 0,017 0,44 0,95
C10:0 31 10,024 0,492 <0,001 -0,0235 0,0033 <0,001 1,18 0,88
C12:0 31 4,950 0,240 <0,001 -0,0173 0,0020 <0,001 0,88 0,80
C14:0 31 11,096 0,352 <0,001 -0,0340 0,0044 <0,001 0,89 0,92
C16:0 31 29,576 1,039 <0,001 -0,0780 0,0084 <0,001 3,71 0,84
C18:0 31 7,383 0,592 <0,001 0,0527 0,0091 <0,001 1,46 0,94
C18:1-c9 27 15,285 0,616 <0,001 0,0490 0,0170 0,019 1,50 0,94
C18:1-t10 18 0,318 0,096 0,007 0,0112 0,0046 0,045 0,50 0,88
C18:1-t11 18 1,159 0,255 <0,001 0,0553 0,0040 <0,001 1,39 0,93
C18:2-c9,c12 31 2,445 0,168 <0,001 0,0069 0,0020 0,007 0,56 0,79
C18:2-c9,t11 20 0,697 0,096 <0,001 0,0244 0,0017 <0,001 0,57 0,94
C18:3-c9,c12,c15 31 0,440 0,084 <0,001 -0,0006 0,0007 0,435 0,31 0,56
C18:2-c9,c12/C18:3-c9,c12,c15 31 8,086 1,605 <0,001 0,0331 0,0094 0,031 4,05 0,79
RECM, raíz cuadrada del error cuadrático medio; R2, coeficiente de determinación.

18
CMS no están aclaradas. Tampoco se conocen los motivos por los que la adición de
grasa extra a las dietas de cabras no afecta al contenido de proteína láctea en
contraposición a lo observado en vacas y ovejas (Chilliard et al., 2003).
Por otro lado, el efecto positivo del consumo de cualquiera de las tres clases de grasa
sobre el porcentaje de grasa láctea difiere de lo observado en vacas por Chilliard et al.
(1993). Una posible explicación sería que la mayor velocidad de tránsito ruminal
característica de los pequeños rumiantes bien atenúa el efecto negativo de las fuentes
de grasa incluidas en la dieta sobre la digestión ruminal de los carbohidratos fibrosos y
la producción de precursores para la síntesis de novo en la ubre (Sanz Sampelayo et
al., 2007), o bien reduce la producción ruminal de isómeros del ácido linoleico
conjugado que se conoce que afectan negativamente a la lipogénesis mamaria como p.
ej. C18:2-t10,c12 (Chilliard et al., 2003). También es posible que existan diferencias
interespecíficas en la respuesta de la ubre a un mayor aporte de dichos isómeros pues
los efectos observados en vacas (Shingfield y Griinari, 2007) y ovejas (Lock et al.,
2006) no se han observado en cabras (Andrade y Schmidely, 2006b).
Contenido de ácidos grasos de la grasa láctea
Ácidos grasos saturados de cadena corta y media
Los resultados obtenidos (Tablas IV, V, VI y VII) confirman observaciones previas de
nuestro laboratorio (Martínez Marín et al., 2012d) según las cuales un consumo de
grasa añadida comprendido entre 1 y 3 g/kg PV no disminuye el contenido de C4:0,
C6:0 y C8:0 de la grasa láctea y que los efectos negativos sobre los ácidos grasos de
la leche sintetizados de novo empiezan claramente solo a partir de C12:0 en la mayoría
de los experimentos realizados con cabras, a diferencia de lo observado en
experimentos realizados con vacas y ovejas donde es más frecuente que se observen
efectos negativos desde C6:0 y C8:0.
A partir de datos obtenidos con vacas, Glasser et al. (2007) sugirieron que la síntesis
de novo podría estar limitada cuando los animales consumen dietas con alto porcentaje
de grasa extra (3-6%) debido al efecto combinado de una menor disponibilidad de
ácidos grasos volátiles, derivada de la menor digestión ruminal de la FND, y de una
mayor captación de ácidos grasos de cadena larga por la ubre. El aumento de los
ácidos grasos de cadena larga captados por la glándula mamaria podría reducir las
actividades de las enzimas que participan en las rutas de síntesis de ácidos grasos en

19
la ubre (Bernard et al., 2009a). De hecho se ha observado in vitro que la reducción de la
ratio de las enzimas acetil-CoA carboxilasa y ácido graso sintetasa en las células
mamarias modifica el patrón de los ácidos grasos sintetizados de novo hacia los de
cadena más corta (Bauman y Davies, 1974). En este sentido, los valores de las
pendientes de las ecuaciones de regresión obtenidas para C16:0 sugieren que la
síntesis de este ácido graso se afectó en mayor medida al aumentar el consumo de
grasa añadida en comparación con los restantes AGS de cadena media (Figura 1).
Figura 1. Evolución del contenido de los ácidos grasos saturados de cadena media de la grasa láctea de cabras con el consumo de grasa añadida a la dieta. Elaborado a partir de los valores medios obtenidos con las ecuaciones de regresión de los tratamientos con grasa añadida.
Ácidos esteárico y oleico
El mayor contenido de C18:0 y C18:1-c9 de la grasa láctea en los tratamientos con
grasa añadida (Tabla IV) fue debido al mayor aporte de ácidos grasos de 18 carbonos
con las dietas de dichos tratamientos en comparación con NOLIP. Cualquier AGI de 18
carbonos presente en el rumen puede ser una fuente de ácido esteárico por
biohidrogenación (BH) ruminal. A su vez, el contenido de C18:1-c9 de la grasa láctea
puede aumentar por una mayor captación mamaria de ácido oleico preformado
procedente de la dieta o, en su caso, de la movilización de reservas corporales de
grasa, o por ∆-9 desaturación mamaria del ácido esteárico procedente a su vez de la
BH de los AGI o de la grasa corporal movilizada.
0,0
5,0
10,0
15,0
20,0
25,0
30,0
35,0
0 50 100
% d
e lo
s ác
ido
s g
raso
s to
tale
s
Consumo de grasa, g/d
C10:0
C12:0
C14:0
C16:0
20
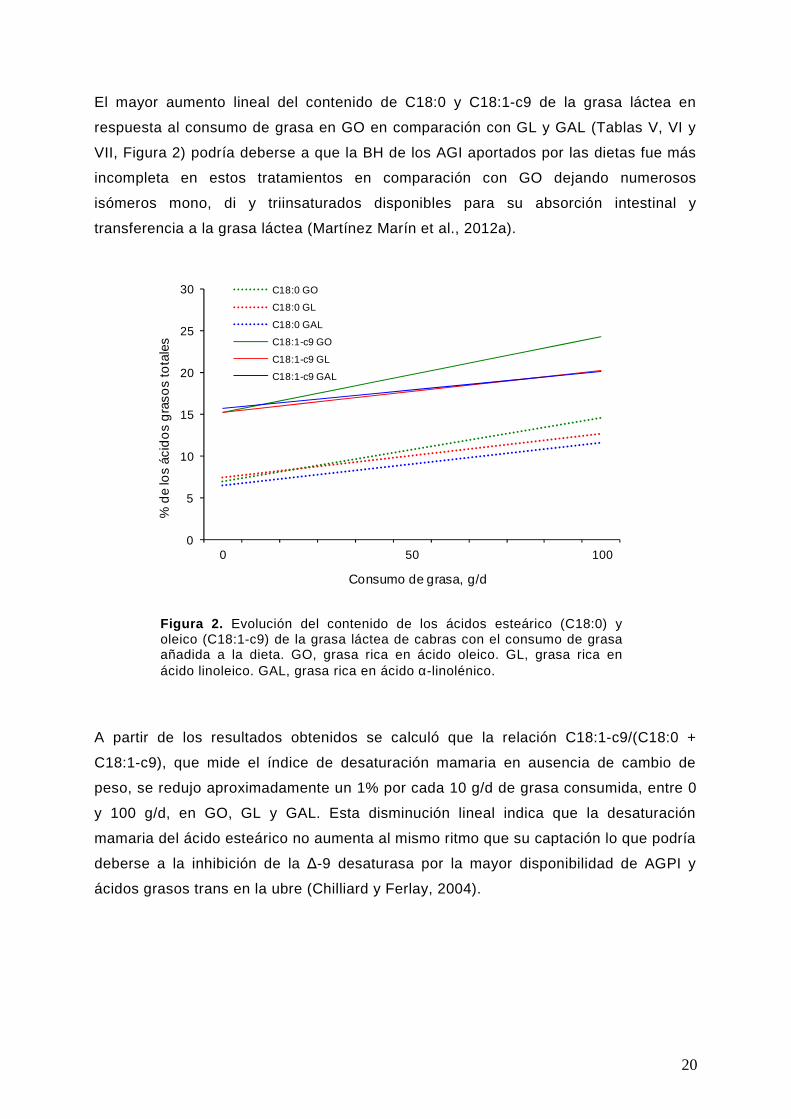
El mayor aumento lineal del contenido de C18:0 y C18:1-c9 de la grasa láctea en
respuesta al consumo de grasa en GO en comparación con GL y GAL (Tablas V, VI y
VII, Figura 2) podría deberse a que la BH de los AGI aportados por las dietas fue más
incompleta en estos tratamientos en comparación con GO dejando numerosos
isómeros mono, di y triinsaturados disponibles para su absorción intestinal y
transferencia a la grasa láctea (Martínez Marín et al., 2012a).
Figura 2. Evolución del contenido de los ácidos esteárico (C18:0) y oleico (C18:1-c9) de la grasa láctea de cabras con el consumo de grasa añadida a la dieta. GO, grasa rica en ácido oleico. GL, grasa rica en ácido linoleico. GAL, grasa rica en ácido α-linolénico.
A partir de los resultados obtenidos se calculó que la relación C18:1-c9/(C18:0 +
C18:1-c9), que mide el índice de desaturación mamaria en ausencia de cambio de
peso, se redujo aproximadamente un 1% por cada 10 g/d de grasa consumida, entre 0
y 100 g/d, en GO, GL y GAL. Esta disminución lineal indica que la desaturación
mamaria del ácido esteárico no aumenta al mismo ritmo que su captación lo que podría
deberse a la inhibición de la ∆-9 desaturasa por la mayor disponibilidad de AGPI y
ácidos grasos trans en la ubre (Chilliard y Ferlay, 2004).
0
5
10
15
20
25
30
0 50 100
% d
e lo
s ác
ido
s g
raso
s to
tale
s
Consumo de grasa, g/d
C18:0 GO
C18:0 GL
C18:0 GAL
C18:1-c9 GO
C18:1-c9 GL
C18:1-c9 GAL

21
Tabla VII. Ecuaciones de regresión lineal del efecto del consumo de grasa rica en ácido α-linolénico añadida a la dieta (expresado en g/d) sobre la producción y la composición de la leche y el contenido de ácidos grasos de la grasa láctea de cabras.
Variables n
Estimaciones de los parámetros
Intersección EEM P Pendiente EEM P RECM R2
Consumo de materia seca, g/d 19 1844 113 <0,001 -0,8 0,5 0,180 149 0,92
Producción de leche, g/d 19 2166 310 <0,001 0,1 0,8 0,930 238 0,98
Grasa, % 19 4,17 0,44 <0,001 0,003 0,002 0,241 0,66 0,90
Grasa, g/d 19 82 8 <0,001 0,1 0,04 0,037 11 0,95
Proteína, % 19 3,12 0,13 <0,001 0,001 0,0004 0,172 0,13 0,96
Proteína, g/d 19 65 9 <0,001 0,01 0,02 0,631 6 0,98
Ácidos grasos, % de los ácidos grasos totales
C4:0 14 2,494 0,066 <0,001 0,0025 0,0006 0,002 0,15 0,75
C6:0 17 2,465 0,225 <0,001 -0,0002 0,0010 0,866 0,27 0,92
C8:0 17 2,794 0,231 <0,001 -0,0012 0,0015 0,438 0,42 0,82
C10:0 17 10,320 0,562 <0,001 -0,0209 0,0028 <0,001 0,79 0,93
C12:0 17 4,974 0,211 <0,001 -0,0188 0,0018 0,002 0,35 0,95
C14:0 17 10,923 0,371 <0,001 -0,0350 0,0020 <0,001 0,30 0,99
C16:0 17 30,267 0,939 <0,001 -0,1079 0,0130 <0,001 3,80 0,85
C18:0 17 6,515 0,436 <0,001 0,0503 0,0122 0,007 1,39 0,88
C18:1-c9 17 15,664 0,665 <0,001 0,0450 0,0225 0,095 0,56 0,99
C18:1-t10 15 0,309 0,042 <0,001 0,0023 0,0016 0,212 0,07 0,99
C18:1-t11 15 1,064 0,128 <0,001 0,0494 0,0076 0,002 0,40 0,99
C18:2-c9,c12 17 2,053 0,116 <0,001 -0,0021 0,0016 0,419 0,25 0,82
C18:2-c9,t11 17 0,704 0,053 <0,001 0,0171 0,0029 <0,001 0,26 0,97
C18:3-c9,c12,c15 17 0,458 0,147 0,021 0,0105 0,0024 0,005 0,07 0,99
C18:2-c9,c12/C18:3-c9,c12,c15 17 6,326 1,198 <0,001 -0,0498 0,0167 0,012 4,90 0,47
RECM, raíz cuadrada del error cuadrático medio; R2, coeficiente de determinación.

22
Ácido C18:1-t10
El aumento del contenido de C18:1-t10 de la grasa láctea en GL (Tabla IV) puede
explicarse porque las dietas ricas en C18:2-c9,c12 modifican las rutas de BH de este
ácido graso, resultando en mayor producción de C18:1-t10 vía C18:2-t10,c12 (McKain
et al., 2010). Por el contrario, la BH de C18:3-c9,c12,c15 es menos propensa a
promover la formación de C18:1-t10 que la del ácido linoleico (AbuGhazaleh y
Jacobson, 2007; Jouany et al., 2007). La BH de C18:1-c9 hasta C18:0 tiene como paso
previo la producción de numerosos isómeros trans monoinsaturados (Mosley et al.,
2002; AbuGhazaleh et al., 2005) pero es proporcionalmente más completa que la de
los AGPI (Martínez Marín et al., 2012b).
Ácidos vaccénico y ruménico
El aumento del contenido de C18:1-t11 de la grasa láctea en GL y GAL (Tabla IV) se
debió a que este ácido graso es el paso previo común a la producción de ácido
esteárico en las rutas de BH de los ácidos linoleico y α-linolénico (Bauman et al., 1999)
y es conocido que su concentración ruminal aumenta cuando se incrementa el aporte
de los propios ácidos grasos que sirven de sustrato (Harvatine y Allen, 2006;
Troegeler-Meynadier et al., 2006), lo que explicaría el aumento lineal de su contenido
observado en respuesta al aumento del consumo de grasa añadida en GL y GAL
(Tablas VI y VII, Figura 3). En pruebas realizadas in vitro se ha observado que el
ácido vaccénico no es el isómero mayoritario en la BH del ácido oleico hasta esteárico
(Mosley et al., 2002; AbuGhazaleh et al., 2005). Sin embargo, la respuesta lineal del
contenido de C18:1-t11 observada en GO (Tabla V) sugiere que el consumo de
cantidades crecientes de ácido oleico modifica las proporciones entre los isómeros
trans monoinsaturados producidos en el rumen a favor de aquel. Los resultados
presentados en el experimento 2 de Martínez Marín et al. (2012a) apoyarían esta
hipótesis. Estos autores observaron que el aumento lineal del contenido de C18:1-t11
de la grasa láctea en respuesta al consumo creciente de aceite de girasol alto oleico
fue más rápido que el de la suma de los contenidos de los restantes isómeros trans
monoinsaturados de 18 carbonos.
El menor aumento del contenido de ácido ruménico de la grasa láctea en GAL en
respuesta al consumo creciente de grasa en comparación con GL (Tablas VI y VII,
Figura 3) fue debido a que todo o la mayor parte del ácido ruménico de la leche provino

23
de la ∆-9 desaturación del ácido vaccénico captado por la ubre ya que el ácido
ruménico no es un intermediario en la BH del ácido α-linolénico (Bauman et al., 1999).
El ácido ruménico tampoco es un intermediario en la BH del ácido oleico pero, a
diferencia de lo observado en GAL, la ∆-9 desaturación del ácido vaccénico, si ocurrió,
fue inapreciable en GO (Tabla IV).
Con las ecuaciones de las Tablas V, VI y VII se calculó que la ratio de los ácidos
ruménico y vaccénico de la grasa láctea fue lineal y tuvo un valor medio de 0,38. Este
valor fue 38% y 17% menor que los observados por Nudda et al. (2006) y Mele et al.
(2008), respectivamente, y 68% mayor que el observado por Luna et al. (2008).
Figura 3. Evolución del contenido de los ácidos vaccénico (C18:1-t11) y ruménico (C18:2-c9,t11) de la grasa láctea de cabras con el consumo de grasa añadida a la dieta. GL grasa rica en ácido linoleico. GAL, grasa rica en ácido α-linolénico.
Ácidos linoleico y α-linolénico
El aumento del contenido de los ácidos linoleico y α-linolénico de la grasa láctea en los
tratamientos GL y GAL, respectivamente, (Tabla IV) era esperado ya que las dietas que
incluyeron estos tratamientos fueron las únicas enriquecidas apreciablemente en dichos
ácidos grasos: 1,98% MS de ácido linoleico y 1,80% MS de ácido α-linolénico en GL y
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
5
5,5
6
6,5
7
7,5
0 50 100
% d
e lo
s á
cid
os g
raso
s to
tale
s
Consumo de grasa, g/d
C18:2-c9,t11 GL
C18:2-c9,t11 GAL
C18:1-t11 GL
C18:1-t11 GAL

24
GAL, respectivamente, a partir de las Tablas II y III. Estos cambios fueron los principales
responsables del aumento y la disminución de la ratio de ambos ácidos grasos en la grasa
láctea de GL y GAL, respectivamente (Figura 4).
Figura 4. Evolución de la ratio de los ácidos linoleico y α-linolénico de la grasa láctea de cabras con el consumo de grasa añadida a la dieta. GL, grasa rica en ácido linoleico. GAL, grasa rica en ácido α-linolénico.
Chilliard y Ferlay (2004) señalaron que las fuentes de grasa ricas en ácido linoleico
incluidas en la dieta tienen un efecto negativo sobre el contenido de ácido α-linolénico
de la grasa láctea mientras que las fuentes de grasa ricas en ácido α-linolénico tienen
un efecto negativo sobre el contenido de ácido linoleico de la grasa láctea, de lo que se
deduce que la incorporación de ambos ácidos grasos a los triglicéridos de la leche no es
independiente. El signo de las pendientes de las ecuaciones de regresión obtenidas para
dichos ácidos grasos apoya dicha observación (Tablas VI y VII).
El tratamiento GO no cambió los contenidos ni el valor de la ratio de los ácidos linoleico y α-
linolénico de la grasa láctea en comparación con NOLIP (Tabla IV) porque las dietas
incluidas en aquel tratamiento no contenían cantidades importantes de dichos ácidos grasos
(0,59 y 0,18% MS de ácido linoleico y α-linolénico, respectivamente, a partir de las Tablas II
y III).
0
2
4
6
8
10
12
0 50 100
Rat
io
Consumo de grasa, g/d
GL
GAL

25
CONCLUSIONES
Este trabajo presenta resultados sobre la respuesta cualitativa y cuantitativa de la
producción y la composición de la leche y el contenido de ácidos grasos de la grasa
láctea de cabras cuya dieta incluyó fuentes vegetales de grasa ricas en ácidos grasos
insaturados. A pesar del limitado número de trabajos experimentales disponibles para
el estudio, los resultados obtenidos son consistentes con el conocimiento actual del
metabolismo ruminal y mamario de los ácidos grasos.
La modificación del contenido de ácidos grasos saturados de cadena corta y media de
la grasa láctea fue similar entre los tratamientos con grasa añadida. Sin embargo, el
contenido de ácidos grasos de cadena larga mostró respuestas diferentes en función
del ácido graso mayoritario en la grasa añadida a la dieta. En conjunto, estos
resultados muestran la enorme capacidad de la ubre para modificar las proporciones
de los ácidos grasos incorporados a los triglicéridos en respuesta a los sustratos
disponibles.
Por otro lado, los resultados obtenidos confirman que las grasas ricas en ácido α-
linolénico promueven los cambios más favorables del perfil de ácidos grasos de la
grasa láctea desde el punto de vista de la salud humana de acuerdo con los
conocimientos actuales.
Por último, la publicación de más experimentos para investigar el efecto de la adición
de fuentes vegetales de grasa a la dieta de cabras lecheras que abarquen una amplia
diversidad de dietas y tratamientos y que provean una información completa de las
condiciones experimentales y de los resultados obtenidos permitirá estudiar más
factores e incluir un mayor número de variables en futuros metanálisis.
AGRADECIMIENTOS
A los profesores de Nutrición Animal del Departamento de Producción Animal de la
Universidad de Córdoba por su revisión crítica del manuscrito.

26
APÉNDICE
Sintaxis de los programas de SAS para los modelos e stadísticos utilizados
Efecto de la adición de grasa a la dieta
PROC MIXED DATA= metanalisis METHOD=REML COVTEST CL ALPHA=.05;
WEIGHT raiz_n_animales;
CLASS grasa experimento;
MODEL variable1 = grasa / HTYPE=3 DDFM=ST;
RANDOM experimento experimento*grasa;
LSMEANS grasa;
CONTRAST 'NOLIP vs GO' grasa -1 1 0 0;
CONTRAST 'NOLIP vs GL' grasa -1 0 1 0;
CONTRAST 'NOLIP vs GAL' grasa -1 0 0 1;
CONTRAST 'GO vs GL' grasa 0 -1 1 0;
CONTRAST 'GO vs GAL' grasa 0 -1 0 1;
CONTRAST 'GL vs GAL' grasa 0 0 -1 1;
run;
1Cualquiera de las variables estudiadas.
Efecto de la cantidad de grasa consumida
PROC MIXED DATA= metanalisis METHOD=REML COVTEST CL ALPHA=.05;
WEIGHT raiz_n_animales;
CLASS experimento;
MODEL variable1 = consumodegrasa/ INFLUENCE(ITER=0) HTYPE=3 DDFM=ST;
RANDOM experimento experimento*consumodegrasa;
run;
1Cualquiera de las variables estudiadas.

27
REFERENCIAS
AbuGhazaleh AA, Jacobson BN. 2007. The effect of pH and polyunsaturated C18 fatty
acid source on the production of vaccenic acid and conjugated linoleic acids in ruminal
cultures incubated with docosahexaenoic acid. Anim. Feed Sci. Technol., 136: 11-22.
AbuGhazaleh AA, Riley MB, Thies EE, Jenkins TC. 2005. Dilution rate and pH effects
on the conversion of oleic acid to trans C18:1 positional isomers in continuous culture.
J. Dairy Sci., 88: 4334-4341.
Almeida OC. 2008. Concentração arterial, retenção de metabólitos pela glândula
mamária e concentração de CLA no leite de cabras, em resposta à ingestão de fontes
de ácidos graxos poliinsaturados ou doses crescentes de óleo de soja. Tesis Doctoral.
Piracicaba, São Paulo, Brasil. Universidade de São Paulo.
Andrade PVD, Schmidely PH. 2006a. Influence of percentage of concentrate in
combination with rolled canola seeds on performance, rumen fermentation and milk
fatty acid composition in dairy goats. Livest. Sci., 104: 77-90.
Andrade PVD, Schmidely PH. 2006b. Effect of duodenal infusion of trans10,cis12-CLA
on milk performance and milk fatty acid profile in diary goats fed high or low
concentrate diet in combination with rolled canola seed. Reprod. Nutr. Dev., 46: 31-48.
Bauman DE, Davis CL. 1974. Biosynthesis of milk fat. Pages 31–75 in Lactation: A
Comprehensive Treatise. Volume 2. BL Larson and VR Smith, ed. Academic Press,
New York, NY.
Bauman DE, Baumgard LH, Corl BA, Griinari JM. 1999. Biosynthesis of conjugated
linoleic acid in ruminants. Proc. Am. Soc. Anim. Sci. Annu. Mtg. 1999.
http://www.asas.org/jas/symposia/proceedings/0937.pdf. Consultado: 28-02-2012.
Benjamin S, Spener F. 2009. Conjugated linoleic acids as functional food: an insight
into their health benefits. Nutr. Metabol., 6: 36-48.
Bernard L, Rouel J, Leroux C, Ferlay A, Faulconnier Y, Legrand P, Chilliard Y. 2005.
Mammary lipid metabolism and milk fatty acid secretion in alpine goats fed vegetable
lipids. J. Dairy. Sci., 88: 1478-1489.

28
Bernard L, Bonnet M, Leroux C, Shingfield KJ, Chilliard Y. 2009a. Effect of sunflower-
seed oil and linseed oil on tissue lipid metabolism, gene expression, and milk fatty acid
secretion in Alpine goats fed maize silage-based diets. J. Dairy. Sci., 92: 6083-6094.
Bernard L, Shingfield KJ, Rouel J, Ferlay A, Chilliard Y. 2009b. Effect of plant oils in the
diet on performance and milk fatty acid composition in goats fed diets based on grass
hay or maize silage. Br. J. Nutr., 101: 213-224.
Bouattour MA, Casals R, Albanell E, Such X, Caja G. 2008. Feeding soybean oil to
dairy goats increases conjugated linoleic acid in milk. J.Dairy. Sci., 91: 2399-2407.
Brown-Crowder IE, Hart SP, Cameron M, Sahlu T, Goetsch AL. 2001. Effects of dietary
tallow level on performance of Alpine does in early lactation. Small. Rumin. Res., 39:
233-241.
Chilliard Y, A Ferlay, J Rouel, G Lamberet. 2003. A review of nutritional and
physiological factors affecting goat milk lipid synthesis and lipolyisis. J. Dairy. Sci., 86:
1751-1770.
Chilliard Y, Doreau M, Gagliostro G, Elmeddah Y. 1993. Addition de lipides protégés
(encapsulés ou savons de calcium) à la ration de vaches laitières. Effets sur les
performances et la composition du lait. INRA Prod. Anim., 6: 139-150.
Chilliard Y, Ferlay A. 2004. Dietary lipids and forages interactions on cow and goat milk
fatty acid composition and sensory properties. Reprod. Nutr. Dev., 44: 467-492.
Chilliard Y, Glasser F, Ferlay A, Bernard L, Rouel J, Doreau M. 2007. Diet, rumen
biohydrogenation and nutritional quality of cow and goat milk fat. Eur. J. Lipid. Sci.
Tech., 109: 828-855.
Field CJ, Blewett HH, Proctor S, and Vine D. 2009. Human health benefits of vaccenic
acid. Appl. Physiol. Nutr. Metabol., 34: 979-991.
Glasser F, Doreau M, Ferlay A, Loor JJ, Chilliard Y. 2007. Milk fatty acids: mammary
synthesis could limit transfer from duodenum in cows. Eur. J. Lipid Technol., 109: 817-
827.

29
Glasser F, Ferlay A, Chilliard Y. 2008. Oilseed lipid supplements and fatty acid
composition of cow milk: A Meta-Analysis. J. Dairy. Sci., 91: 4687-4703.
Harvatine KJ, Allen MS. 2006. Fat supplements affect fractional rates of ruminal fatty
acid biohydrogenation and passage in dairy cows. J. Nutr., 136: 677-685.
Hodgson JM, Wahlqvist ML, Boxall JA, Balazs ND. 1996. Platelet trans fatty acids in
relation to angiographically assessed coronary artery disease. Atherosclerosis, 120:
147-154.
Jouany J P, Lassalas B, Doreau M, Glasser F. 2007. Dynamic features of the rumen
metabolism of linoleic acid, linolenic acid and linseed oil measured in vitro. Lipids, 42:
351–360.
Lana RP, Camardelli MML, Queiroz AC, Rodrigues MT, Eifert EC, Miranda EN, Almeida
ICC. 2005. Soybean oil and propolis in the diets of dairy goats. Rev. Bras. Zootec., 34:
650-658.
Li XZ, Yan CG, Long RJ, Jin GL, Khuu JS, Ji BJ, Choi SH, Lee HG, Song MK. 2009.
Conjugated linoleic acid in rumen fluid and milk fat, and methane emission of lactating
goats fed a soybean oil-based diet supplemented with sodium bicarbonate and
monensin. Asian Australas. J. Anim. Sci., 22: 1521-1530.
Lock AL, Teles BM, Perfield JW, Bauman DE, Sinclair LA. 2006. A conjugated linoleic
acid supplement containing trans-10, cis-12 reduces milk fat synthesis in lactating
sheep. J. Dairy Sci., 89: 1525-1532.
Luna P, Bach A, Juárez M, De la Fuente MA. 2008. Effect of a diet enriched in whole
linseed and sunflower oil on goat milk fatty acid composition and conjugated linoleic
acid isomer profile. J. Dairy Sci., 91: 20-28.
Maia FJ, Branco AF, Mouro GF, Coneglian SM, Dos Santos GT, Minella TF, Guimarães
KC. 2006. Inclusão de fontes de óleo na dieta de cabras em lactação: produção,
composição e perfil dos ácidos graxos do leite. Rev. Bras. Zootec., 35: 1504-1513.
MARM, Ministerio de Medio Ambiente y Medio Rural y Marino. 2010. Plan de Acción
para la Leche de Cabra. Madrid.

30
Martínez Marín AL, Gómez-Cortés P, Gómez Castro G, Juárez M, Pérez Alba L, Pérez
Hernández M, De La Fuente MA. 2011a. Animal performance and milk fatty acid profile
of dairy goats fed diets with different unsaturated plant oils. J. Dairy Sci., 94: 5359-
5368.
Martínez Marín AL, Gómez-Cortés P, Gómez Castro G, Juárez M, Pérez Alba L, Pérez
Hernández M, De La Fuente MA. 2012a. Effects of feeding increasing dietary levels of
high oleic or regular sunflower or linseed oil on fatty acid profile of goat milk. J. Dairy
Sci., 95: 1942-1955.
Martínez Marín AL, Gómez-Cortés P, Gómez Castro G, Juarez M, Pérez Alba LM,
Pérez Hernández M, De La Fuente MA. 2012b. Adición de aceites vegetales a la dieta
de cabras lecheras: efecto sobre el contenido de ácidos grasos de la grasa láctea. Arch
Med. Vet., enviado para evaluación.
Martínez Marín AL, Pérez Hernández M, Pérez Alba LM, Carrión Pardo D, Gómez
Castro AG. 2012c. Adición de aceites vegetales a la dieta de cabras lecheras: efecto
sobre la digestibilidad y los resultados productivos. Arch. Med. Vet., 44: 21-28.
Martínez Marín AL, Pérez Hernández M, Pérez Alba LM, Gómez Castro G, Carrión
Pardo D. 2011b. Efecto de las fuentes de grasa sobre la digestión de la fibra en los
rumiantes. REDVET, 12: 1695-7504.
Martínez Marín AL, Pérez Hernández M, Pérez Alba LM, Gómez Castro G, Carrión
Pardo D, Garzón Sígler AI. 2012d. Efecto de la grasa vegetal añadida a la dieta sobre
los ácidos grasos de la grasa láctea de los rumiantes: revisión. Tec. Pecu. Mex,.
enviado para evaluación.
McKain N, Shingfield KJ, Wallace RJ. 2010. Metabolism of conjugated linoleic acids
and 18:1 fatty acids by ruminal bacteria: products and mechanisms. Microbiology, 156:
579–588.
Mele M, Serra A, Buccioni A, Conte G, Pollicardo A, Secchiari P. 2008. Effect of
soybean oil supplementation on milk fatty acid composition from Saanen goats fed diets
with different forage:concentrate ratios. Ital. J. Anim. Sci., 7: 297-311.

31
Mir Z, Goonewardene LA, Okine E, Jaegar S, Scheer HD. 1999. Effect of feeding
canola oil on constituents, conjugated linoleic acid (CLA) and long chain fatty acids in
goats milk. Small Rumin. Res., 33: 137-143.
Mosley EE, Powell GL, Riley MB, Jenkins TC. 2002. Microbial biohydrogenation of oleic
acid to trans isomers in vitro. J. Lipid. Res., 43: 290-296.
Mouro GF, Branco AF, Macedo FAF, Maia FJ, Coneglian SM, Guimarães KC, Santos
GT. 2002. Óleos vegetais em dietas de cabras Saanen em lactação: Produção e
composição do leite e ingestão de nutrientes. In: 39 Reunião Anual da Sociedade
Brasileira de Zootecnia, Recife, Brasil.
Nudda A, Battacone G, Usai MG, Fancellu S, Pulina G. 2006. Supplementation with
extruded linseed cake affects concentrations of conjugated linoleic acid and vaccenic
acid in goat milk. J. Dairy. Sci., 89: 277-282.
Okine EK, Goonewardene LA, Mir Z, Mir P, Wang Z, Chanmugam P S. 2003. Influence
of canola oil on the fatty acid profile in goats' milk. Can. J. Anim. Sci., 83: 323-325.
Ollier S, Leroux C, De la Foye A, Bernard L, Rouel J, Chilliard Y. 2009. Whole intact
rapeseeds or sunflower oil in high-forage or high-concentrate diets affects milk yield,
milk composition, and mammary gene expression profile in goats. J. Dairy. Sci., 92:
5544-5560.
Sanz Sampelayo MR, Chilliard Y, Schmidely P, Boza J. 2007. Influence of type of diet
on the fat constituents of goat and sheep milk. Small Rumin. Res., 68: 42-63.
SAS. 2004. SAS/STAT 9.1 User's Guide. SAS Institute Inc., Cary, NC.
Sauvant D, Schmidely P, Daudin JJ, St-Pierre NR. 2008. Meta-analyses of
experimental data in animal nutrition. Animal 2: 1203-1214.
Schmidely P, Morand-Fehr P, Sauvant D. 2005. Influence of extruded soybeans with or
without bicarbonate on milk performance and fatty acid composition of goat milk. J.
Dairy Sci., 88: 757-765.

32
Shingfield KJ, Griinari JM. 2007. Role of biohydrogenation intermediates in milk fat
depression. Eur. J. Lipid. Sci. Technol., 109: 799-816.
Silva MMC. 2005. Suplementaçao de lipídios em dietas para cabras leiteiras. Tesis
Doctoral. Viçosa, Minas Gerais, Brasil: Universidad Federal de Viçosa.
Simopoulos AP. 2008. The importance of the omega-6/omega-3 fatty acid ratio in
cardiovascular disease and other chronic diseases. Exp. Biol. Med., 233: 674-688.
St-Pierre NR. 2001. Invited review: integrating quantitative findings from multiple
studies using mixed model methodology. J. Dairy Sci., 84: 741-755.
Teh TH, Trung LT, Jia ZH, Gipson TA, Ogden KB, Sweeney TF. 1994. Varying amounts
of rumen-inert fat for high producing goats in early lactation. J. Dairy Sci., 77: 253-258.
Troegeler-Meynadier A, Bret-Bennis L, Enjalbert F. 2006. Rates and efficiencies of
reactions of ruminal biohydrogenation of linoleic acid according to pH and
polyunsaturated fatty acids concentrations. Reprod. Nutr. Dev., 46: 713-724.
Ulbricht TLV, Southgate DAT. 1991. Coronary heart disease: seven dietary factors.
Lancet., 338: 985-992.