Embed Size (px)
Citation preview

Diciembre 2011Volumen 4 | Número 136
¿UNA DÉCADA DE BIOLOGÍA DE SISTEMAS?
Visión parcial de una ilustración creada por Raúl Montañez Martínez para representar simbólicamente el enfoque que adopta la Biología de
Sistemas para estudiar la complejidad de la vida

Vol.4 | Nº 136
Director:Salvador [email protected]ía Celular -Neurobiología
Co-Editores:José María Pérez [email protected]ía del desarrollo y cardiovascularMiguel Ángel Medina [email protected]ía Molecular y de Sistemas-Biofísica-Bioquímica
Comité editorial:Alberto Martí[email protected]ón AmbientalE. Profesional para el EmpleoAlejandro Pérez Garcí[email protected]ía, Interacción planta-patógenoAlicia [email protected]íaEnfermedades neurodegenerativasAna [email protected]ética-Virología, Patogénesis viralesAntonio Dié[email protected]ía de la Ciencia
Enrique Moreno [email protected]ía- Limnología
Enrique [email protected]ética- GenómicaFélix López [email protected]ía-Fotobiología, Cambio climáticoFernando Ojeda Barceló[email protected]ón AmbientalEducacion SecundariaEmpleo de T.I.C. en docenciaFrancisco Cá[email protected]ía Molecular Vegetal, Bioquímica y Biología MolecularJesús [email protected]íaBiodiversidad animalJosé Carlos Dá[email protected]ía Celular -NeurobiologíaJuan Antonio Pérez [email protected]ía
Juan Carlos [email protected]ímica-Biología Molecular, Energética de procesos biológicosJuan Carlos [email protected]íaEducación Secundaria
Margarita Pérez Martí[email protected]ía AnimalNeurogénesisMaría del Carmen [email protected]ía de aguasPatología vírica de pecesMaría Jesús García Sá[email protected]ía VegetalNutrición mineralMaría Jesús Perlé[email protected]ía, Riesgos medioambientalesM. Gonzalo [email protected]ímica-Biología Molecular y BioinformáticaRaquel [email protected]!siologíaBiorremediaciónTrinidad Carrió[email protected] de la SaludE-Salud
Diseño:Raúl Montañez Martínez ([email protected])
Coordinador de la edición electrónica
(www.encuentros.uma.es):Ramón Muñoz-Chápuli
Correspondencia a:Miguel Ángel Medina Torres
Departamento de Biología Molecular y BioquímicaFacultad de Ciencias
Universidad de Málaga29071 Málaga
Editado con la !nanciación del Vicerrectorado de Investigación de la Universidad de Málaga
Depósito Legal: MA-1.133/94ISSN: 1134-8496
Imprenta: Imagraf
ÍndiceEditorial 57
La imagen comentadaNota. Figuras acompañantes
5758
¿Qué es la Biología de Sistemas? 59
La Biología de Sistemas: ¿Un desarrollo normal de la Biología?
61
¿El aniversario de la Biología de Sistemas? 62
La “materia oscura” de los sistemas biológicos 64
Existo, luego soy una red 65
Cinco preguntas sobre Biología Sintética 67
A debate: Biología de Sistemas... 68
El equipo editorial de esta publicación no se hace responsable de las opiniones vertidas por los autores colaboradores.
Diciembre 2011
Vol.4 | Nº 136
El presente número de Encuentros en la Biología con el que concluimos el año 2011 es muy especial, al estar dedicado íntegramente y de forma monográ�ca a la Biología de Sistemas. Desde aquí, queremos dejar constancia del agradecimiento del equipo editorial de Encuentros en la Biología a todos cuantos lo han hecho posible. Este monográ�co consta de nada menos que siete contribuciones en forma de artículos de divulgación que suman cerca de quince mil palabras escritas. Este inusual volumen de texto escrito en un mismo número de Encuentros en la Biología nos ha obligado a una ajustada labor de maquetación, incrementando el tamaño de las cajas de texto y disminuyendo un punto el t a m a ñ o d e l o s t i p o s d e l e t ra s empleados (y de forma proporcional la separación entre líneas). Además, las
simbolizan lo que los enfoques sistémicos pretenden representar en nuestra comprensión de la complejidad biológica. La imagen de portada es la reproducción de un dibujo alegórico hecho con lápiz de gra�to. Un f r a g m e n t o c i r c u l a r d e l m i s m o reproducido a menor tamaño ha servido de “marcador” del �nal de cada artículo, cuyo comienzo ha venido indicado a su vez por una reproducción a pequeño tamaño de una imagen digital creada con un collage que m u e s t r a d i f e r e n t e s n i v e l e s d e integración en Biología, desde el gen a los ecosistemas, pasando por las proteínas, funciones, células, tejidos y organismos.
Los co-editores
�guras acompañantes de algunas de estas contribuciones no han podido ser reproducidas al tamaño que hubiesen merecido. Estos inconvenientes, impuestos por las limitaciones de formato de la revista, no impiden que los textos sean diáfanamente legibles. Los artículos han sido escritos por investigadores de las Universidades de La Laguna, Lleida, Murcia y Pompeu Fabra (Barcelona), así como de la Universidad de Málaga (desde donde esta revista se edita). Dentro de la temática de este número monográ�co, la sección La imagen comentada presenta una red biológica. Finalmente, y n o m e n o s i m p o r t a nte, e s e l agradecimiento que debemos al Dr. R aúl Montañez Mar t ínez (CSIC-Universidad Pompeu Fabra) que nos h aya p e r m i t i d o re p ro d u c i r d o s imágenes creadas por él y que
EDITORIAL
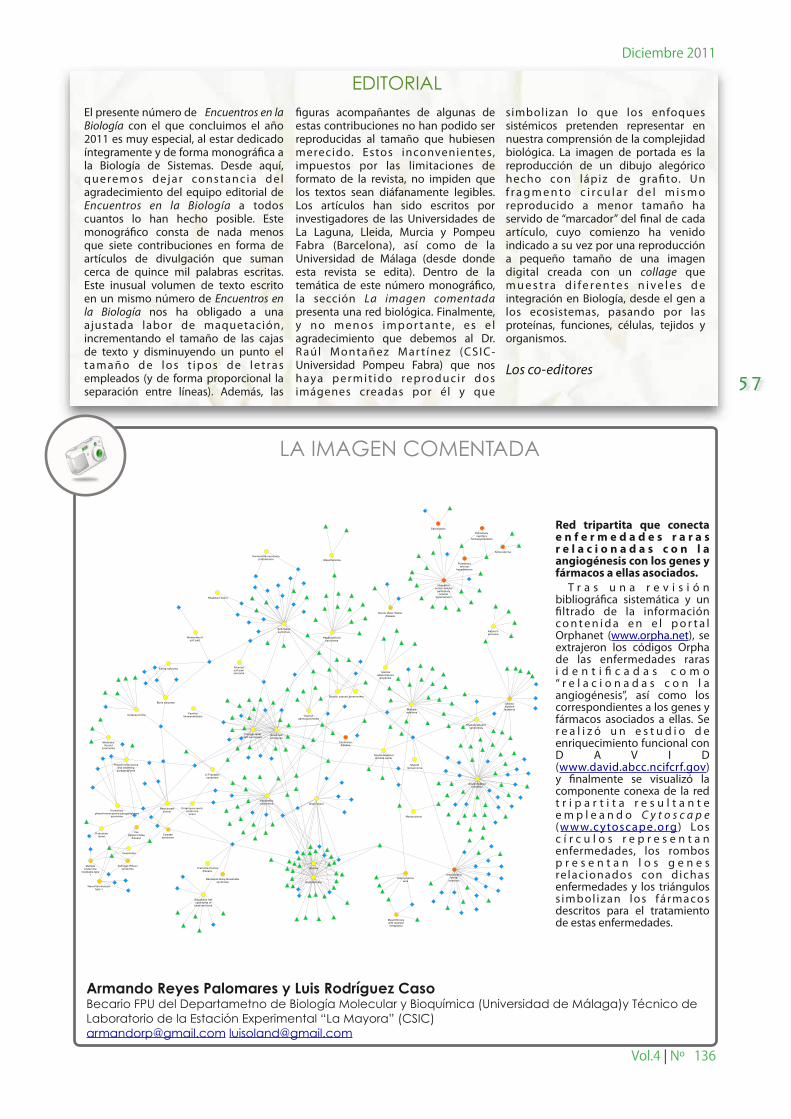
Red tripartita que conecta e n f e r m e d a d e s r a r a s r e l a c i o n a d a s c o n l a angiogénesis con los genes y fármacos a ellas asociados. T r a s u n a r e v i s i ó n bibliográ�ca sistemática y un �ltrado de la información c o n t e n i d a e n e l p o r t a l Orphanet (www.orpha.net), se extrajeron los códigos Orpha de las enfermedades raras i d e n t i � c a d a s c o m o “ r e l a c i o n a d a s c o n l a angiogénesis”, así como los correspondientes a los genes y fármacos asociados a ellas. Se r e a l i z ó u n e s t u d i o d e enriquecimiento funcional con D A V I D (www.david.abcc.ncifcrf.gov) y �nalmente se visualizó la componente conexa de la red t r i p a r t i t a r e s u l t a n t e e m p l e a n d o C y t o s c a p e (w w w.c ytoscape.org) Los c í r c u l o s r e p r e s e n t a n enfermedades, los rombos p r e s e n t a n l o s g e n e s re lac ionados con dichas enfermedades y los triángulos s imbol izan los fármacos descritos para el tratamiento de estas enfermedades.
LA IMAGEN COMENTADA
Armando Reyes Palomares y Luis Rodríguez CasoBecario FPU del Departametno de Biología Molecular y Bioquímica (Universidad de Málaga)y Técnico de Laboratorio de la Estación Experimental “La Mayora” (CSIC)[email protected] [email protected]
57
Chronic myeloid
leukemia
Bannayan-Riley-Ruvalcaba syndrome
Glioma
Myelofibrosis with myeloid metaplasia
Amyotrophic lateral
sclerosis Polycythemia
vera Glioblastoma
Myelodysplastic syndromes
Gastrointestinal stromal tumor
Myxoid liposarcoma
Mastocytosis
Multiple myeloma
Acute myeloid leukemia
Li-Fraumeni syndrome
Medullary thyroid
carcinoma
Rare breast cancer
Enteropancreatic endocrine
tumor Hereditary
pheochromocytoma-paraganglioma syndrome
Von Hippel-Lindau
disease Cowden
syndrome
Ewing sarcoma
Bone sarcoma
Familial leiomyomatosis
Pheochromocytoma and secreting
paraganglioma
Squamous cell carcinoma of
head and neck
Lhermitte-Duclos disease
Multiple endocrine
neoplasia type 1
Endocrine tumor
Osteosarcoma
Zollinger-Ellison syndrome
Insulinoma
Neurofibromatosis type 1
Ovarian adenocarcinoma
Familial renal cell carcinoma
Castleman disease
Pancreatic carcinoma
Renal cell carcinoma
LiposarcomaGastric cancer
Brain tumor
Rhabdoid tumor
Rendu-Osler-Weber disease
Familial adenomatous
polyposis
Soft tissue sarcomas
Hepatocellular carcinoma
Melanoma of soft part
Alveolar soft-part sarcoma
Dermatofibrosarcoma protuberans
Kaposi's sarcoma
MesotheliomaPulmonary
arterial hypertension
Pulmonary capillary
hemangiomatosis
Sarcoidosis
Idiopathic and/or familial
pulmonary arterial
hypertension
Scleroderma

Vol.4 | Nº 136
58
NOTA DE LOS EDITORES: Por necesidades de maquetación, adjuntamos aquí una �gura que complementa los contenidos del artículo de Salvador Durán “Cinco preguntas sobre Biología Sintética” (páginas 67 y 68 de este número de Encuentros en la Biología), así como las �guras acompañantes al artículo de la sección “A Debate” (páginas 68-70).Como nota de actualidad, debe añadirse que la iGEM 2011 (competición a la que se alude en el artículo de Durán) reunió del 5 al 7 de Noviembre en el MIT a los 160 equipos seleccionados como �nalistas. Resultó vencedor un equipo de estudiantes de la University of Washington por: 1) crear una cepa de E. coli capaz de biosintetizar alcanos, coponentes fundamentales de los combustibles diésel; 2) modi�car una enzima para aumentar su capacidad de degradar gluten; y 3) crear una cepa de E. coli magnética.
De “Cinco preguntas sobre Biología Sintética”
De “Biología de Sistemas... ¿qué biología de sistemas?”
Figura acompañante al artículo “Cinco preguntas sobre Biología Sintética” (páginas 67-68) reelaborada a partir de otra publicada en la referencia (7) donde se ponen de manifiesto los distintos hitos de la biología sintética según sus autores
Implementación del oscilador, el toggle switch y
la expresión con poco ruido en E. coli. También empiezan a realizarse
trabajos sobre la comunicación celular.
!"""# #!""!# #!""$# #!""%# #!""&# #!""'# #!""(# #!"))#
Trabajos notables en la expresión estocástica y en la evolución dirigida
de circuitos genéticos.
Se constituye el Registry of Standard Biological Parts.
Tiene lugar la primera competición del iGEM, con sólo cinco equipos.
Se celebra el primer congreso de biología sintética (SB1.0).
Primeros experimentos de formación de
patrones espaciales y en comunicación artificial en celulas de levadura.
Alteran células de levadura para que expresen un
precursor de la artemisina.
E. coli expresando Invasina bajo condiciones de hipoxia se usa para combatir tumores in vitro.
Uso de RNA de interferencia para
crear circuitos arbitrariamente
complejos basados en lógica booleana.
Se sintetiza, clona y ensambla el primer genoma bacteriano.
Última competición iGEM hasta la fecha, con 160 equipos
de todos los continentes.
Se sincroniza un oscilador en E. coli usando los sistemas de
quorum sensing naturales.
Figura 1
Figura 2: Diferentes formas de visualizar el enfoque de la Biología de Sistemas
Figura 3: Científicos que contribuyeron a una comprensión sistémica de la Biología antes del “advenimiento” de la Biología de Sistemas
Figura 4: La creciente brecha entre la cantidad de datos disponibles y el conocimiento generado. Adaptado de una imagen original de Dieter Mart.

Diciembre 2011
Vol.4 | Nº 136
¿QUÉ ES LA BIOLOGÍA DE SISTEMAS?
59
Néstor Torres Darias Catedrático de Bioquímica y Biología Molecular. Departamento de Bioquímica y Biología Molecular
Universidad de La Laguna [email protected]
explicita, pero forma parte del fondo implícito de otras muchas aproxi-maciones.
El carácter dinámico de los sistemas biológicos está estrecha-mente conectado con las relaciones que estos mantienen con su entor-no, cambiante a su vez. Un sistema vivo, desde la célula al tejido, desde el órgano al organismo completo y desde este al ecosistema se mueve, actúa, reacciona y responde en función de las condiciones ambientales sobre el que el sistema, a su vez, in�uye. El concepto de sistema propio de la Biología de Sistemas implica, además de la dimensión temporal, una estrecha relación e in�uencia con y sobre el entorno.
De lo anterior se in�ere una distinción que es crítica en Biología de Sistemas, la que debe hacerse entre las dinámicas internas de cada nivel de organización (célula, tejido, organismo), responsables del fun-cionamiento de cada uno de ellos, y la dinámica entre los niveles, que determinan la función de estos en un todo mayor. La búsqueda de los principios generales de organización de la dinámica interna de cada nivel organizativo y los que operan entre distintos niveles de organización, en el contexto de la función de cada uno de estos, son objetos de investigación que de�nen la Biología de Sistemas.
Los seres vivos son complejos sistemas de sistemas complejos. Los sistemas biológicos muestran distintos niveles de organización; se nos presentan como sistemas de (sub)sistemas dotados de principios or-ganizativos propios que rigen la estructura y función de la totalidad. En el sistema cada subsistema mantiene su identidad (su estructura y su fun-ción) como resultado de leyes y principios especí�cos que operan en cada uno. Pero además, las interacciones propias de los sistemas bioló-gicos (no sólo las que se dan en cada (sub)sistema sino también las que se dan entre (sub)sistemas son generalmente no lineales. Es la riqueza y variedad de las interacciones y su no linealidad la que hace imposible que se pueda explicar el comportamiento y las propiedades de un sis-tema con referencia sólo a sus partes componentes.
Los sistemas biológicos mani�estan propiedades emergentes. En los sistemas biológicos las relaciones entre sus partes y el todo gene-ra nuevas propiedades; propiedades que se describen en términos de emergencia. Dicho de otra manera, el comportamiento del conjunto "emerge" del funcionamiento de las partes y de las interacciones entre ellas.
El concepto de emergencia alude a cómo el comportamiento del todo se ve afectado no sólo por los cambios entre las partes, sino por las interacciones entre ellas. Su explicación requiere conocer las relaciones que se dan entre los distintos niveles. En cada nivel de organización de un sistema biológico (e.g. un tejido) el biólogo sistémico distingue entre el todo (el tejido) y sus partes (las células) y reconoce la in�uencia de-terminante que existe entre ambos niveles. En esta representación el todo es el producto de las partes y las partes a su vez dependen del todo en su funcionamiento y existencia. Un tejido es por tanto a la vez causa y efecto de sí mismo. Es esta relación circular entre las interacciones de las células y su entorno lo que distingue a la Biología de Sistemas de las aproximaciones reduccionistas propias de la Biología clásica.
Hacia una de�nición de Biología de Sistemas: el papel de la modelización
La Biología de Sistemas es, en gran medida, modelización mate-mática. Si el objetivo de la Biología de Sistemas es comprender el com-portamiento dinámico de los sistemas vivos un elemento central de su metodología propia es la modelización matemática.
La modelación cientí�ca es un arte. Se trata de una actividad creativa y rigurosa que implica la integración de descubrimientos e hipótesis y que requiere intuición, imaginación e independencia de pensamiento. Modelar en ciencia es el arte de hacer las preguntas co-rrectas, de elegir el marco conceptual adecuado para formular y testar hipótesis y de hacer suposiciones y simpli�caciones acertadas.
La modelización no es una opción, sino una necesidad en el estudio de los sistemas biológicos. La primera razón que justi�ca la necesidad instrumental del modelo en la Biología es la complejidad inherente a los seres vivos a la que ya nos referimos antes. Llamamos aquí la atención sobre el hecho de que complejidad no es sinónimo de abundancia de elementos. La complejidad, por el contrario, tiene que ver con el número de diferentes tipos de componentes antes que con el número de componentes. De hecho, tener muchas moléculas o células que interactúan no es un problema como tal, especialmente si podemos describir el conjunto como un promedio de los elementos o si actúan coordinadamente. El reto y la di�cultad para el modelador reside en la variedad de clases de elementos.
Hay una escena en el Génesis (2, 19-20) en la que Dios concede a Adán la capacidad de poner nombre a los seres y las cosas, lo que es interpretado al mismo tiempo como un poder y una responsabilidad que la deidad atribuye al ser humano. De�nir no es fácil tampoco, hasta el punto de que algunos han proclamado su odio a las de�niciones (Benjamin Disraeli) mientras que otros nos han advertido (e.g. Erasmo de Rotterdam) que toda de�nición es peligrosa. Si nos referimos al tema que nos ocupa, la Biología de Sistemas, la evidencia de la di�cultad y del peligro que representa dar una de�nición aparece pronto. Cualquiera que se haya iniciado en el estudio de la Biología de Sistemas habrá po-dido constatar que no abundan las de�niciones generales y comprensi-vas de la misma, lo que constituye un buen un indicador de la di�cultad y el riego que implica la respuesta a la pregunta que da título a este artículo. Pero aceptaremos el reto, conscientes del riesgo que asumimos y de la di�cultad que conlleva. Con�emos en no salir malparados del intento…
La inspección de muchas de las de�niciones de Biología de Sis-temas que se han presentado (algunos ejemplos signi�cativos se pue-den encontrar en Kitano, 2001; Fundación Genoma España, 2007; Liu and Lau�enburger, 2010; Klip et al, 2005) pone de mani�esto que a pesar de la diversidad de las de�niciones en contenidos y planteamien-tos todas comparten algunos elementos, si bien no todas ponen el énfa-sis en los mismos. Ante situaciones como esta, que no son nuevas en Ciencia, un recurso del que se ha hecho uso en ocasiones es acudir a una de�nición operativa, consistente en de�nir el qué a través del cómo. No caeré en la tentación de de�nir la Biología de Sistemas diciendo que es lo que hacen aquellos que se llaman a sí mismos biólogos de sistemas. Pero sí me aproximaré a una de�nición a través de las actividades que le son propias.
En lo que sigue plantearé pues lo que, desde mi punto de vista, distingue a la Biología de Sistemas, y por tanto lo que la diferencia de lo que no lo es. De la exposición de sus rasgos distintivos se derivará una propuesta metodológica, con características diferenciadas, que permi-ten identi�car a un proyecto de investigación como de Biología de Sis-temas.
Hacia una de�nición de Biología de Sistemas: el contextoUna condición necesaria para aproximarse satisfactoriamente a
la de�nición de una disciplina es la formulación de sus objetivos. Por tanto ¿cuáles son las preguntas que pretende responder la Biología de Sistemas? ¿Qué aspectos del mundo biológico son los que interesan desde la óptica sistémica?
El rasgo distintivo de la Biología de Sistemas es su preocupación por la organización y la función biológica. La Biología de Sistemas inves-tiga i) las relaciones que se dan entre los componentes estructurales del sistema biológico (e.g. la célula) y su función y, ii) las características de las interacciones que se dan entre los distintos sistemas (las células) que permite al conjunto desarrollar y mantener niveles superiores de organi-zación estructural y funcional.
Estas preguntas, para poder ser entendidas cabalmente, deben situarse en su contexto correcto, es decir deben de�nirse las “condiciones de contorno” en las que deben ser formuladas para ser pertinentes. Di-chas condiciones de contorno forman parte constitutiva, ineludible, del fondo en el que la Biología de Sistemas se desenvuelve.
Los seres vivos son sistemas abiertos alejados del equilibrio termodinámico. Los sistemas biológicos son sistemas abiertos que intercambian materiales con su entorno. Dentro de sus límites tienen lugar reacciones químicas, organizadas en redes, en las que los distintos componentes se consumen y regeneran continuamente de tal manera que la composición se mantiene aproximadamente constante. El carác-ter abierto, junto con la existencia de gradientes termodinámicos (prin-cipalmente electroquímicos) posibilitan que, sin entrar en contradicción con el Segundo Principio de la Termodinámica, generen orden y estruc-tura espacio temporales.
Los seres vivos son sistemas dinámicos. Desde la perspectiva de la Biología de Sistemas un ser vivo es un sistema en el que sus elementos constituyentes están relacionados entre sí a través de interacciones dinámi-cas. Esta de�nición de sistema enfatiza el hecho de que los sistemas naturales no sólo son multicompuestos, sino que se relacionan entre sí, cambian, se adaptan y evolucionan. Aquí radica uno de los retos que afronta la Biología de Sistemas y que en gran medida ha sido ignorado por otros enfoques no sistémicos: sólo tiene sentido estudiar a los seres vivos si se los considera entes cambiantes, dinámicos y fuertemente interrelacionados. Queda excluido por tanto cualquier planteamiento fundamentado en una concepción estacionaria e invariante de los sis-temas biológicos. Esta concepción estacionaria normalmente no se

Vol.4 | Nº 136
La necesidad de los modelos surge también del hecho de que el sistema y su entorno son indisociables: no es posible entender al prime-ro sin una referencia al segundo. Las interacciones entre las partes y por tanto el comportamiento de las variables del sistema depende del esta-do del sistema completo. Por ello cualquier proyecto de modelización debe combinar dos modos complementarios. Por una parte se requiere reducir el sistema en subsistemas más simples, (rutas metabólicas, redes, módulos) y sus modelos mecanísticos correspondientes. Por otra, se precisan planteamientos basados en enfoques más abstractos que per-mitan estudiar la organización funcional en distintos niveles. Esta se-gunda dimensión de la modelización re�eja el hecho de que, si bien el comportamiento del conjunto (el todo) está determinado por las pro-piedades de sus componentes (causalidad ascendente), el comporta-miento de sus componentes (las partes) también se ve determinado por las propiedades del conjunto (causalidad descendente). En un tejido el comportamiento de una célula está controlado tanto por las propieda-des de sus macromoléculas constituyentes como por las propiedades del tejido.
La modelización es simpli�cación. Aunque a primera vista la complejidad inherente de los sistemas vivos podría sugerir modelos complejos, lo cierto es justo lo contrario: en un modelo se busca delibe-radamente simpli�car la realidad a través de la abstracción y la reducción de elementos. Los grados de libertad en el ejercicio simpli�cador son amplios, con la condición de que el modelo simpli�cado re�eje la esen-cia del sistema real. Las simpli�caciones son, además, necesarias para que los modelos sean viables y manipulables.
Puestos a simpli�car nos encontramos con distintos tipos de simpli�caciones. Una simpli�cación inevitable es la asociada a la delimi-tación física del sistema. Esta pasa por establecer una frontera entre el sistema y su entorno; por seleccionar las magnitudes susceptibles de ser observadas (medidas) y la clasi�cación de estas en variables, parámetros y constantes. Por otra parte, tal como se comentó antes también, cual-quier sistema es a la vez, un subsistema. Todo nivel organizativo tiene siempre un nivel inferior y otro superior, por lo que toda descripción estará necesariamente referida a un nivel y contexto determinados. Por tanto, el entorno en el que se desenvuelve el sistema es una cuestión de elección; algunas podrán ser más apropiadas que otras. Pero no hay motivo para escandalizarse por las elecciones a las que se ve abocado el modelador. Las simpli�caciones del modelador son las propias de cual-quier práctica cientí�ca. El mundo, tal como nos lo muestra la ciencia, es sólo una versión más abstracta y sistemática de la que nos revela la ex-periencia directa del mismo.
Los modelos organizan el conocimiento en todos coherentes. El proceso de modelización de un sistema, a través del cual se integra in-formación de los distintos niveles de organización (genética, epigenéti-ca, celular, tejido) y de las distintas escalas temporales, constituye en sí mismo una propuesta metodológica para integrar información e hipóte-sis y diseñar los experimentos que permitan comprobarlas. Vistos así, los modelos son útiles incluso como proyecto. Los modelos no son tampoco la medida del éxito de un proyecto de investigación ni por lo tanto el objetivo �nal del mismo. Los modelos cumplen la función de suministrar el marco conceptual necesario para la formulación y evaluación cuanti-tativa de las hipótesis. Los modelos son sólo (¡y nada menos!) que herra-mientas para mejorar nuestra comprensión de los sistemas biológicos.
La �nalidad de los modelos no es la predicción sino la compren-sión del sistema. La generación de predicciones es sólo una manera de comprobar si la comprensión alcanzada es consistente con el compor-tamiento del sistema que estudiamos. Es la discusión a la que abocan los resultados de un modelo lo que le da o no validez y lo que determina en última instancia lo que el modelo aporta a la comprensión del fenóme-no.
Construir modelos no es fácil. La modelización matemática tiene menos que ver con ecuaciones que con conceptos, ideas y patrones de comportamiento. Y es aquí en donde residen muchas de las di�cultades para el modelador. Los investigadores que pierden de vista la visión sistémica al enfrentarse con la complejidad inherente de los sistemas biológicos no ven el paisaje, sólo los detalles. Resulta así más fácil dar con nuevos resultados. El modelador por el contrario se esfuerza por superar los obstáculos que nos pone la intuición y nuestros propios perjuicios cientí�cos. El modelador, como cualquier artista, se enfrenta a la creación y esto es un proceso arduo y doloroso. No hay fórmulas senci-llas para construir modelos, no existe un algoritmo que estructure y organice la información y lo transforme en conocimiento. Por eso con frecuencia la interacción entre los modeladores y los que no lo son es difícil; los modeladores tienen a destruir seguridades, enfatizar la duda y promocionar las preguntas antes que las respuestas.
La Biología de Sistemas y su Programa de InvestigaciónA la vista de lo anterior, la Biología de Sistemas se nos presenta
como una aproximación interdisciplinar al estudio de las interacciones y las dinámicas propias de los (complejos) sistemas biológicos. Aproxima-ción que no centra su atención en las propiedades de los elementos constituyentes, sino que persigue descifrar las propiedades y compor-
tamientos emergentes de los sistemas vivos. La complejidad de los sis-temas biológicos y sus funciones surgen de las interacciones no lineales que se dan entre millares de componentes de distinta naturaleza y fe-nómenos espaciotemporales: la Biología de Sistemas pretende identi�-car y analizar los principios, leyes y mecanismos subyacentes al compor-tamiento de los sistemas vivos.
Los objetivos de la Biología de Sistemas son al mismo tiempo un programa de fusión de las disciplinas (transdisciplinariedad) que descri-ben los elementos que forman parte de los sistemas (la Biología Molecu-lar; la Biología Celular; la Fisiología, la Ecología, Bioinformática, etc.) con la Teoría de Sistemas Dinámicos (el Análisis Matemático, la Investigación Operativa, las Ingenierías, la Ciencias de la Computación, etc.)
La Biología de Sistemas suministra su propia versión del método experimental para la comprensión de los sistemas a través de sus com-ponentes, las interacciones entre éstos y de los mismos con su entorno. Se trata de la iteración de un ciclo básico: observación seguida de mode-lización matemática; simulación, teoría y vuelta a la observación. En esta estrategia de investigación, en la metodología que emana de sus postu-lados, el modelo tiene un papel central. Una vez elegido el objeto de investigación y antes de plantear y llevar a cabo cualquier experimento, el genuino biólogo de sistemas debe re�exionar sobre el modelo que va a construir. A partir de la información disponible y de hipótesis bien fundamentadas construiría una primera versión del modelo en el que se presenta la selección inicial de las variables consideradas clave, los pro-cesos y las interacciones relevantes. De esta propuesta, generalmente acompañada de su complemento en forma de modelo mecanístico, se derivaría la primera formulación matemática del modelo. Llegar a este punto habrá supuesto un ejercicio de integración de información e hipótesis, y por tanto haber alcanzado ya un grado de conocimiento sobre el problema que no se tenía antes. Como resultado de estas dos fases se hace casi directamente evidente la información que se necesita para la re�nar el modelo y por tanto los diseños experimentales necesa-rios: qué experimentos realizar, la condiciones de los mismos, la tempo-ralización, los procesos a valorar, etc. El proceso de construcción del modelo y el modelo en sí mismo marcan pues el camino hacia una ma-yor comprensión del sistema y señala qué diseños experimentales son relevantes. En este proceso los experimentos son subsidiarios del mode-lo y se justi�can sólo en función de su utilidad para construir o veri�car el modelo y las hipótesis en las que se basa.
En Biología de Sistemas las habilidades matemáticas son esencia-les aunque no las únicas necesarias. El análisis de los datos, tanto direc-tamente a partir de los resultados experimentales como el que se puede realizar con la información contenida en las bases de datos y la literatura cientí�ca juega un papel crucial. La construcción de cualquier modelo requiere información sobre los elementos que participan y sus propie-dades. Esta información puede obtenerse en las bases de datos, utili-zando métodos y herramientas de la Bioinformática, realizando experi-mentos ad hoc o, más frecuentemente, por una combinación de ambos. Técnicas matemáticas y minería de datos se complementan.
El desarrollo de la Biología de Sistemas debe correr paralelo al desarrollo de metodologías teóricas dirigidas a la identi�cación de sis-temas, de conceptos teóricos para el diseño de experimentos, de méto-dos para testar hipótesis, de marcos teóricos que permitan el acopla-miento de procesos que tienen lugar en diferentes escalas temporales y de algoritmos e�caces para resolver problemas computacionales com-plejos.
ConclusiónAunque todavía hoy la Biologías de Sistemas se presenta como
una nueva disciplina en el ámbito de las Ciencias Biológicas, realmente sus planteamientos y propuestas tienen importantes antecedentes. De hecho la aproximación sistémica es un tema recurrente en Biología. Norbert Wiener y su propuesta de la cibernética (1948) fue uno de sus precursores, como lo fue también Ludwig von Bertalan�y y su Teoría General de los Sistemas (1968). La diferencia entre estas propuestas y la Biología de Sistemas actual no es tanto de naturaleza cualitativa o de planteamientos, sino más bien cuantitativa, en términos de cantidad: de información y de recursos de computación, análisis y comunicación. Las tecnologías disponibles hoy día permiten no sólo acceder a enormes cantidades de información sobre la composición, estructura y dinámica de los sistemas en sus niveles molecular y genético sino que ponen a nuestro alcance inmensos recursos en términos de potencia de cálculo y métodos de análisis.
Los retos que limitan pues el desarrollo de la aproximación sis-témica, no son técnicos sino intelectuales. Sólo en la medida de que la forma de pensar sistémica se generalice se garantizara su desarrollo y podremos explotar todo su enorme potencial de conocimiento y aplicaciones.
60

Diciembre 2011
Vol.4 | Nº 136
La Biología de Sistemas: ¿Un desarrollo normal de la Biología?
61
Manuel Cánovas Díaz Catedrático de Bioquímica y Biología Molecular Departamento de Bioquímica y Biología Molecular (B) e
Inmunología. Facultad de Química. Universidad de Murcia. [email protected]
del ser vivo a un nivel completo, generándose un nuevo conocimiento que va más allá de los propios componentes celulares. Las “ómicas” es-tán permitiendo establecer el conocimiento jerárquico de los distintos niveles, genes, productos de ellos (mRNAs), circuitos genéticos, proteí-nas, moléculas señalizadoras y metabolitos (Fig. 1) [3]. La función celular depende de la coordinación existente entre ellos y el estudio de esa
acción coordinadora es el objeto de estudio de la BS. La aproximación al estudio de un organismo desde el punto de
vista de la BS, consiste en primer lugar en realizar la anotación génica, identi�cando todos los marcos abiertos de de lectura (ORF) de su DNA, su localización y sus secuencias reguladoras. Se puede asignar función a cada ORF del DNA mediante la búsqueda de homología de los genes desconocidos; esto sería una anotación monodimensional. No obstante, si lo que se pretende es tener la máxima información, es necesario hacer una anotación bidimensional en la cual se pueda incluir la matriz de relaciones de estados, las relaciones entre ellos y la estequiometría que hay entre los componentes que se están transformando. La tabla que se genera constituye la matriz estequiométrica a escala genómica comple-ta y su uso está consiguiendo que se reconstruyan las rutas metabólicas a partir del genoma (Fig. 2). No sólo eso, sino que además se están de�-niendo las rutas de señalización, las reguladoras transcripcionales y post-transcripcionales y su acción a nivel metabólico.
Cada circuito génico realiza una función y los genes implicados pueden incluso participar en múltiples circuitos génicos. De hecho la vida de un organismo es el resultado de múltiples circuitos génicos que
En la actualidad a partir de la secuenciación y anotación del genoma de un organismo estamos en condiciones de conocer la organi-zación de dicho genoma. No obstante, es necesario representar y des-cribir (con modelos fáciles de comprender) cómo se expresa la informa-ción génica de un organismo, cómo se regula, cómo se conduce y tradu-ce en un metabolismo concreto y cómo dicho metabolismo se regula, permitiéndole adaptaciones múltiples a diferentes entornos de creci-miento, e incluso optimizando la expresión génica en procesos adapta-tivos en aquellos casos necesarios (perturbaciones genéticas, mutacio-nes, diferentes entornos de crecimiento, etc)[2]. La Biología de Sistemas (BS) fue de�nida por primera vez por Leroy Hood, cofundador del Institut of Systems Biology (Seattle, USA), en 1999, y aunque se ha de�nido de varias formas, en general se acepta que es la parte de la ciencia que se encarga del estudio de los mecanismos que describen procesos biológi-cos complejos en un organismo, representándolos como sistemas inte-grados de múltiples componentes interactuando entre sí. La BS implica i) la obtención de grandes conjuntos de datos experimentales, ii) el desarrollo de modelos matemáticos que puedan representar al menos algunos de los aspectos signi�cativos procedentes de estos conjuntos de datos, iii) la obtención de soluciones computacionales de las ecua-ciones matemáticas para obtener predicciones numéricas, y iv) la de-terminación de la calidad del modelo mediante la comparación de las simulaciones numéricas con los datos experimentales.
Desde esta primera de�nición, se han acuñado diferentes de�ni-ciones. Algunos cientí�cos han manifestado que la BS no es nada nuevo, sino más bien la ciencia biológica de siempre con un nuevo toque, una buena aproximación al proceso �siológico. Otros opinan que es la apro-ximación de los ingenieros y matemáticos a la descripción de cómo funcionan los sistemas bio/�siológicos [1]. Lo cierto es que el proyecto Genoma Humano demostró que sólo se estaba al inicio del conocimien-to del funcionamiento del ser completo. Y que la opinión de algunos cientí�cos, basada en que al conocer los componentes del ser vivo com-prenderíamos lo que es la vida, no era más que una falacia, puesto que solo presentaba la línea de salida en la carrera por el conocimiento del ser vivo completo.
Existe un acuerdo general de que la fuerza motriz de la BS ha sido el advenimiento de las “ómicas” (genómica, transcriptómica, pro-teómica, señalómica y metabolómica), las llamadas técnicas de alto rendimiento (high throughput techniques) generadoras de datos masi-vos, que están permitiendo enumerar los componentes celulares y estu-diar su evolución temporal. El conjunto de interacciones físico/químicas entre esos componentes da lugar a la reconstrucción de las rutas de reacciones bioquímicas que representan las distintas funciones de los seres vivos. Además, la BS no se centra en los propios componentes celulares, sino en la naturaleza de las uniones que los conecta y los esta-dos funcionales de las rutas que resultan del ensamblaje de estas unio-nes.
Durante la última parte del siglo pasado imperó la aproximación biológica reduccionista, basada en el estudio de los componentes celu-lares, su composición química, estructura y función [4]. El advenimiento de la genómica aceleró el proceso y desde entonces se ha anotado el genoma de un gran número de organismos, aunque todavía no se co-noce la asignación funcional de un gran número de genes y muy poco sobre moléculas pequeñas e iones que presentan una gran importancia en el funcionamiento celular (AMPc, ppGpp, cationes, aniones, etc.). En este siglo, el conocimiento se está generando de forma masiva y rápida, de forma que se ha abordado el sistema completo de forma integradora. De las rutas metabólicas se pasó a comprender que era necesario cono-cer los distintos niveles representados por el transcriptoma, el proteoma, el metaboloma y el señaloma, entre otros. Además, el desarrollo de las ómicas mostró la necesidad vital de conocer los �ujos de información en el ser vivo, así como la posibilidad de establecer relaciones matemáticas entre los distintos niveles celulares (Fig. 1). Para la descripción del ser vivo con este enfoque se hacen necesarios los distintos niveles de in-formación y las relaciones existentes entre ellos. Por otra parte, se vis-lumbró la posibilidad de optimizar y controlar el funcionamiento celular para su uso en bioindustrias y biomedicina, si se conocía mejor el orga-nismo completo.
Así las cosas, la pregunta que surge es, ¿qué se puede hacer con toda la información que se genera sobre los componentes de una célula o un organismo? O incluso: ¿cómo se llega a las propiedades biológicas y a los comportamientos del ser vivo a partir de esa ingente cantidad de componentes celulares? Desde luego, el manejo de esa información y el análisis integrador que se debe realizar para entender el sistema com-pleto necesita de métodos de análisis de sistemas y de la Bioinformática [2]. Por ello, en los próximos años se va a desarrollar una nueva metodo-logía de estudio de los sistemas celulares, de los tejidos, de los órganos y
Figura 1: Distintos niveles de información celular
Figura 2: Matriz bidimensional de unión e interacción entre componentes

Vol.4 | Nº 136
funcionan de forma coordinada (Fig. 3). Estos circuitos génicos se carac-terizan por ser complejos, autónomos, robustos, ejecutar funciones �siológicas, tomar decisiones y ajustarse para que el ser vivo se adapte (homeostasis). Su estudio necesita de la bioinfomática, que puede com-parar circuitos génicos de varios seres vivos y clasi�carlos por la función que realizan. Más aún, los circuitos génicos son los motores de la evolu-ción, dirigen y coordinan los procesos celulares que son responsables de la transferencia de la información génica, la producción de energía, cre-cimiento, división, movimiento, etc (Fig. 3). Por ello, surge la necesidad de analizar al ser vivo en primer lugar a escala genómica, lo que conduce a ver el genoma como un sistema generador de información y dinámico en su expresión.
A resultas de todo lo mencionado, se establecen y reconstruyen las rutas metabólicas, se establecen los modelos matemáticos que las describen, estos modelos se usan para predecir el funcionamiento me-tabólico completo o una función metabólica determinada, y a posteriori se utiliza la predicción generada para conocer mejor el sistema y diseñar nuevos experimentos que permitan avanzar en el conocimiento del mismo. Todo ello supone una mejora del modelo elaborado in silico en un proceso iterativo. El procedimiento establecido a nivel celular, sigue a nivel de tejido y órgano y �naliza a nivel de ser completo.
El siguiente paso en la descripción del sistema completo es con-siderar que todos los componentes biológicos tienen un recambio. Los metabolitos tiene un recambio de menos de un minuto, los mRNA po-seen vidas medias del orden de minutos (bacterias) a horas (eucariotas), los componentes de una célula poseen diferentes vidas medias. Más aún, en un ser multicelular las células de los tejidos y los órganos poseen vidas medias diferentes y sincronizadas (por ejemplo las células intesti-nales poseen un recambio que implica el cambio del 3-6% del total diario). Una célula que hoy vemos contiene elementos que poseen dife-rente vida media y diferente edad en la misma. No obstante, lo que permanece es la interrelación entre los componentes celulares. De he-cho, esas relaciones son las verdaderas características que perduran y dan sentido al proceso vital. Este es en de�nitiva el sentido de la nueva Biología o Biología de Sistemas, el conocimiento del ser completo a través de sus componentes, cómo se relacionan y recambian. De ahí que la BS, se pueda considerar el desarrollo normal de la Biología clásica gracias a los avances tecnológicos que se están produciendo, que per-miten observar cómo evolucionan los distintos niveles celulares y sus relaciones y así entender al organismo completo.
En 2006 se creó en España la Red Española de Biología de Sistemas (REBS, http://www.sysbiol.net/) durante el Ist International Systems Bio-logy Symposium: From Genome to Bioproducts and Back (ISBN 84-611-1135-4), celebrado en Murcia. A continuación y gracias a ayudas conce-didas para el mantenimiento de la REBS por la Fundación Séneca (CARM, Murcia), Genoma España, Generalitat Catalana y Ministerio de Educación y Ciencia/MICINN, se han realizado reuniones cientí�cas con participan-tes nacionales e internacionales. Las reuniones de mantenimiento de la REBS se han realizado en Murcia (2), Valencia (2), Madrid y Barcelona. Además, en la Universidad de Murcia se organizó el 1er Curso de Inicia-ción en Biología de Sistemas en 2010. En las reuniones, se han identi�ca-do necesidades futuras y aspectos nuevos que están surgiendo del uso de este nuevo campo, así como se ha contribuido a crear grupos de trabajo que están permitiendo optimizar los recursos cientí�cos y hu-manos en nuestro país.
62
Figura 3: Circuitos genéticos y manifestación de vida en un organismo
Bibliografía citada:1. Alberghina L, Westerho� HV (2005). Systems Biology. De�nitions and perspectives. Springer. ISBN 13 978 3 540 22968 1. 2. Maly VI (2009). Systems Biology. Humana Press. Springer Protocols. Methods in Molecular Biology. ISBN 978-1-934115-64-0.3. Tomita M, Nishioka T (2005). Metabolomics. The frontier of Systems Biology. Springer-Verlag Tokyo. ISBN 4-431-25121-9 .4. Voit EO (2000). Computational Analysis of Biochemical Systems. A practical guide for biochemyst and molecular biologysts. Cambridge
University Press. ISBN 0 521 78087 X.
¿El aniversario de la Biología de Sistemas? Rui Alves, Baldiri Salvado, Ester Vilaprinyo & Albert Sorribas
Departament de Ciències Mèdiques Bàsiques, Universitat de Lleida & IRBLleida [email protected]
¿La Biología Molecular clásica no tiene el mismo objetivo?Sin duda éste es el objetivo de la Biología Molecular clásica. Sin
embargo, el camino hacia ese objetivo fue tradicionalmente muy dife-rente de lo que propone la Biología de Sistemas. El éxito de la Biología Molecular se ha debido en gran parte a la aparición de una corriente cientí�ca REDUCCIONISTA que defendió que para entender cómo fun-cionan los seres vivos y sus células sería necesario y su�ciente catalogar sus componentes moleculares individuales y medir el comportamiento de estos componentes en un tubo de ensayo. Esta aproximación reduc-cionista llevó a descubrimientos tan importantes como la penicilina, la teoría cromosómica de la herencia o el código genético.
Si es así, ¿por qué necesitamos el concepto de análisis y medición integrada propuesto por la Biología de Sistemas?
Con el tiempo y a pesar de todos sus éxitos, se veri�có que la aproximación reduccionista no permitiría entender todos los misterios de la Biología a nivel molecular por un motivo: el comportamiento de las
Este número de la revista Encuentros en la Biología nos honró con su invitación a escribir un artículo de divulgación para celebrar el déci-mo aniversario de la Biología de Sistemas. Sin embargo, como malos invitados, nos toca discordar de nuestro huésped. Aunque cada año la Biología de Sistemas cumpla un año más, esta rama de la Biología no está entrando en la preadolescencia, sino que se encuentra en plena madurez y cumple ya varias décadas de vida. ¿Por qué lo decimos? Para aclararlo hace falta primero que nos entendamos sobre qué es la Biolo-gía de Sistemas.
¿Qué es la Biología de Sistemas?La respuesta a esta pregunta no es consensual. Sin embargo,
todos sus practicantes concuerdan seguramente en que es una forma de estudiar sistemas biológicos en la cual se pretende entender cómo funciona el sistema como un todo (célula), a partir de un análisis inte-grado del comportamiento de los componentes (moléculas).

Diciembre 2011
Vol.4 | Nº 136
63
O sea, ¿la Biología de Sistemas hoy se reduce a medir abundancias, �ujos y actividades de todas las especies molecu-lares que componen las células?
La verdad es que no. Hoy en día una gran parte de la investiga-ción en Biología de Sistemas está dedicada a caracterizar el comporta-miento dinámico de los componentes moleculares de la célula de un modo simultáneo y a utilizar esta información para crear modelos que permitan predecir que pasará en el sistema en condiciones diferentes de las utilizadas en las mediciones. Sin embargo, la Biología de Sistemas no se reduce a esto.
¿Qué más hay?Si para cada sistema y/u organismo que nos interesara tuviéra-
mos que medir todas las interacciones entre todos sus componentes moleculares, no acabaríamos nunca. Sin embargo la ciencia tiene como objetivo encontrar leyes generales que permitan describir el universo de la forma más sencilla y general posible. Así, la Biología de Sistemas tam-bién tiene la aspiración de encontrar principios organizativos o de dise-ño biológico que sean válidos y aplicables a una gran cantidad de orga-nismos y de circuitos moleculares y que permitan explicar cómo y por qué estos organismos o circuitos han evolucionado para funcionar de la manera que funcionan.
¿Circuitos? ¿Pero el “Sistema” no es la célula o el organismo? Los organismos han evolucionado de una forma que, en muchos
casos es modular. Como consecuencia de esto, parecen existir módulos o conjuntos de proteínas o genes que en conjunto se especializaron en efectuar una determinada función o respuesta biológica. Si estamos interesados en estudiar la respuesta del organismo a determinados estímulos y esta respuesta es mediada por un grupo de proteínas, en-tonces podemos estudiar sólo el circuito biológico formado por estas proteínas y sus interacciones.
¿Y qué es un principio organizativo o de diseño?Puede ser muchas cosas, pero la de�nición más genérica es la
siguiente. Un principio organizativo o de diseño biológico es una carac-terística de un sistema o circuito biológico que hace que el sistema efec-túe una determinada función de un modo más e�caz que si no tuviera esa característica. Por ejemplo, las vías que sintetizan aminoácidos en las células tienen una regulación inhibitoria de la primera reacción de la vía por el aminoácido producido. Se ha demostrado que esta regulación permite que la célula ajuste de un modo más e�caz y rápido la velocidad de producción de aminoácido a las necesidades que tiene la célula. La ausencia de esta regulación crea circuitos que funcionalmente no son tan e�cientes y por eso crea células que no pueden competir de igual a igual con los organismos que presentan esta regulación.
¿Se conocen muchos principios de diseño?Sí, bastantes y a diferentes niveles. A diario se identi�can nuevos
principios organizativos y de diseño biológico en ámbitos que van des-de la regulación de la expresión genética a la manera cómo funcionan algunos circuitos metabólicos o de transducción de señal. En las lecturas recomendadas se encuentra una revisión sobre el tema.
¿Y ya está? ¿Hay algo más que deba saber sobre estos principios organizativos?
Sí. Además de permitir entender por qué funcionan los organis-mos y cómo funcionan, estos principios nos permiten establecer una biblioteca de diseños que pueden ser útiles en la construcción de circui-tos biológicos arti�ciales, lo que se llama hoy Biología Sintética. Éste se considera un nuevo campo de investigación y es uno de los más excitan-tes y revolucionarios de la Biología hoy en día. Sin embargo, los ingenie-ros genéticos os podrán contar por qué ellos no creen que sea un nuevo campo, del mismo modo que esperamos haberos convencido de lo mismo respecto a la Biología de Sistemas…
diferentes moléculas biológicas medido en el tubo de ensayo di�ere del que se observa en la célula. Esto pasa porque pueden existir otros com-ponentes celulares no identi�cados que interaccionan con las moléculas conocidas y/o porque las condiciones del medio celular son diferentes de las de un tubo de ensayo y porque las interacciones entre las diferen-tes moléculas biológicas son muchas veces no lineales. Esta no-lineali-dad hace que cuando se juntan dos o más tipos de moléculas, su com-portamiento sea diferente del que tienen cada una de ellas aisladas y que, además, este cambio de comportamiento sea difícil de predecir.
Esta noción fue claramente presentada por Ludwig von Berta-lan�y en su Teoría General de Sistemas, inicialmente esbozada en 1937 y desarrollada en los 50 y 60. Sin embargo, el desarrollo tecnológico no permitía efectuar los experimentos adecuados para aplicar el concepto en la práctica. La única técnica que estaba disponible para conseguir este grado de integración era la creación y análisis de modelos matemá-ticos que, utilizando la información determinada individualmente para cada tipo de molécula, integrase esa información de forma no-linear permitiendo predecir cómo funcionaba el sistema reconstruido. Por eso, durante muchas décadas, la Biología de Sistemas fue sinónimo de Biolo-gía Matemática. Sin embargo, sólo unos pocos biólogos y/o matemáti-cos tenían los conocimientos su�cientes para poder aplicar estas herra-mientas. Por este motivo, la Biología de Sistemas en su faceta más ma-temática no era ni muy conocida ni muy utilizada dentro de la Biología Molecular. Solo en la década de 1960 empiezan a aparecer, en universi-dades anglosajonas, los primeros departamentos que incorporan Biolo-gía de Sistemas en su nombre.
¿Y por qué cambió este panorama?Porque al �nal, para poder entender el comportamiento de una
célula a nivel molecular, se requiere conocer todos los componentes que la célula utiliza para ejecutar ese comportamiento. Es necesario identi�-car todos los componentes de las células para poder realmente recons-truir el sistema como un todo. Esta situación llevó a la necesidad de desarrollar métodos que permitieran esta identi�cación, además de la medición simultánea del comportamiento de cada componente en su medio normal. De aquí vino la motivación para los primeros proyectos de secuenciar genomas completos, y entre ellos el genoma humano en particular. Entrada la década de los 90 se demostró que el desarrollo técnico ya era su�ciente para empezar a secuenciar todo el genoma humano aplicando las técnicas utilizadas en la secuenciación de geno-mas de organismos más sencillos. El año 2002, �nalmente, se publicó la primera versión de la secuencia del genoma humano. Por esto, lo que en realidad se cumple es el décimo aniversario de la secuenciación del genoma humano.
Si es así, ¿por qué asociar esta fecha con el aniversario de la Biología de Sistemas?
La secuenciación del genoma humano y, anteriormente, de otros genomas más pequeños llevó al desarrollo de nuevas técnicas que per-miten la medición efectiva y simultanea de las cantidades y actividad de una fracción muy grande de las diferentes moléculas que constituyen las células. Se desarrollaron métodos que permiten medir simultáneamente como cambia la expresión genética de todos los genes de un organis-mo. En seguida se desarrollaron métodos que permiten medir simultá-neamente como cambia la cantidad y/o actividad de una fracción im-portante de las proteínas codi�cadas en el genoma de un organismo. Además, también se desarrollaron métodos que permiten medir simul-táneamente los cambios en las cantidades de pequeños metabolitos celulares. Todas estas técnicas han proporcionado globalmente a la comunidad de la Biología Molecular, por primera vez, la sensación de que todos podrían estudiar su organismo de interés de un modo sisté-mico, global e integrado. Así el nombre de Biología de Sistemas es real-mente adecuado para describir todos los estudios efectuados con estas metodologías y fue adoptado, ahora sí, por la comunidad de la Biología Molecular en general. Eso hizo que la gente que no conocía los trabajos anteriores creyera que el origen de la Biología de Sistemas coincide con la secuenciación del genoma humano y que simbólicamente asociaran los dos aniversarios.
Lecturas recomendadas para saber más:Salvado B, Karathia H, Chimenos AU, Vilaprinyo E, Omholt S, Sorri-bas A, Alves R. Methods for and results from the study of design principles in molecular systems. Math Biosci. 231:3-18, 2011.Savageau MS. Biochemical Systems Analysis: A Study of Function and Design in Molecular Biology. CreateSpace, 2010.Alon U. An Introduction to Systems Biology: Design Principles of Biological Circuits. Chapman and Hall/CRC, 2006.

Vol.4 | Nº 136
La materia oscura es un concepto surgido en la Astrofísica para de�nir aquella masa del universo que no podemos ver con los medios técnicos actuales, pero cuya existencia predecimos por la in�uencia indirecta que ejerce en el devenir de estrellas y galaxias.
En la era de la biología de sistemas hemos comenzado nuestra an-dadura en la investigación del cosmos de interacciones moleculares que articulan los sistemas vivos. En este contexto, y de forma análoga a lo ocurrido en la exploración del universo, los cientí�cos comenzamos a intuir que grandes espacios del intrincado mundo de interrelaciones moleculares subyacentes a la vida aún permanecen oscuros a nuestro conocimiento.
La irrupción de la “revolución industrial” en los experimentos de biología molecular nos ha hecho creer que por �n contamos con los medios técnicos necesarios para la exploración de estos vastos espacios desconocidos. Pero la gran paradoja es que a pesar de que contamos con toneladas de datos experimentales no somos capaces de enunciar modelos sistémicos consistentes que expliquen la diversidad funcional de los sistemas biológicos complejos. Ante esta paradoja, cabría preguntarse ¿Cuál es la razón o razones que, en biología de sistemas, limi-tan tan signi�cativamente la transformación de la información en conocimiento?
De forma general, la comunidad cientí�-ca ha apuntado como principal sospechoso de dicha frustración a la falta de un mayor desarrollo formal y conceptual en una disci-plina incipiente como es la Biología de Sis-temas. No obstante, y sin menoscabo de que esta primera razón sea cierta, cabría también cuestionarse si con el conocimiento acumu-lado hasta la fecha, más las técnicas experi-mentales de exploración actuales, estamos en el buen camino para llegar a comprender como funcionan los sistemas moleculares que estudiamos. En resumen, podríamos decir que mientras la primera razón tendría que ver con una insu�ciencia del desarrollo teórico de la biología de sistemas, la segun-da apuntaría a un desarrollo ine�ciente de la exploración experimental del espacio de interacciones, generándose así modelos incompletos difíciles de interpretar.
Adivinar qué progreso teórico futuro terminará por salvar las limitaciones actuales en este área de conocimiento se presenta como una tarea más especulativa que cientí�ca. En cambio, con los medios computacionales actuales sí es factible estimar el efecto que nuestras técnicas experimentales ejercen en el modelado de ciertos sistemas moleculares importantes, como por ejemplo lo son las redes de interacción entre proteínas. Esto es posible porque, al igual que en Astronomía la materia oscura se predice por su in�uencia indirecta en otros cuerpos celestes, muchas interacciones de las que aún no dispo-nemos de evidencia experimental son producto de largos procesos de co-dependencia evolutiva entre proteínas que dejan su impronta en la estructura y función de los genomas, señales que son detectables me-diante distintos métodos bio-computacionales.
Con el objeto de estudiar los males y virtudes de nuestras técnicas experimentales de exploración y estimar la su�ciencia de los datos acumulados por la comunidad cientí�ca durante décadas, los grupos de la Profesora Christine Orengo, en la University College London, y nuestro grupo, en la Universidad de Málaga, colaboran en un estudio compara-tivo entre la red de interacciones de proteínas basada en el compendio de todas las evidencias experimentales disponibles en bases de datos públicas, y la red de interacciones predichas mediante métodos bio-computacionales [1]. Para dicho estudio utilizamos como objetos de investigación los proteomas completos de dos especies eucariotas muy diferentes, como son el hombre y la levadura S. cerevisiae. Especies que se consideran entre las mejor caracterizadas desde el punto de vista de la Biología Molecular entre los eucariotas, y a la vez lo su�cientemente alejadas en el árbol de la vida como para poder ofrecernos la posibilidad de encontrar patrones universales en nuestros estudios. De esta forma
tenemos dos modelos independientes de la red de interacciones de proteínas que comparar en humano y levadura, por un lado una red compendio del conocimiento derivado de datos experimentales (red experimental), y por otro, la red de interacciones predicha mediante técnicas bio-computacionales (red de predicciones).
Con la comparación y estudio de ambas redes tratamos de dar res-puestas a las siguientes preguntas: ¿Cuánto espacio de las redes de interacciones de proteínas permanece aún oscuro a nuestro conoci-miento experimental? ¿Este espacio oscuro aún por explorar encierra componentes funcionalmente claves necesarios para comprender el funcionamiento del sistema? ¿Nuestras técnicas actuales de exploración experimental nos permitirán con el tiempo caracterizar todos los rinco-nes del espacio de interacciones, o presentan limitaciones de muestreo que están dejando en la oscuridad importantes regiones funcionales?
El estudio de solapamiento entre ambas redes [1] nos desveló que una proporción muy signi�cativa de las redes de interacción entre pro-teínas predichas por medios bio-computacionales no ha sido aún carac-
terizada experimentalmente (el 82% en levadura y el 98% en humanos). A este espacio de interacción inexplorado lo denominamos la “materia oscura” de las redes de interacción. Aunque un alto porcentaje de materia oscura fue observado en ambos organismos, en la levadura el porcentaje de predicciones de las que sí tenemos evidencia experimental (18%) se estimó del orden de 14 veces supe-rior que en humanos (1,34%), indicándonos que al menos desde el punto de vista de la su�ciencia de los datos experimentales ac-tuales, probablemente nos encontramos mucho más cerca de comprender el funcio-namiento de los sistemas en S. cerevisiae de lo que estamos en el ser humano.En la búsqueda de componentes funcional-mente claves en estos espacios no caracteri-zados experimentalmente, encontramos un alto número de proteínas hubs o nodos cen-trales de conexión, claves en la arquitectura de relaciones de la red. Al estudiar el papel funcional de estas proteínas que denomina-mos “hubs ocultos”, descubrimos que en la levadura correspondían principalmente a proteínas de membrana, mientras que en humanos estaban relacionados con funcio-
nes regulatorias en vías de señalización, como las señales mediadas por proteínas quinasas mediante interacciones transitorias y que están implicadas en multitud de procesos de transmi-sión de señales y regulación celulares fundamentales de los organismos eucariotas.
Los resultados obtenidos en estos experimentos ponen en cuestión la manera en la que exploramos experimentalmente las redes de inte-racción molecular, así como nuestras posibilidades de llegar a modelar y comprender su funcionamiento. La estimación de la materia oscura nos indica que incluso para los organismos mejor estudiados como son el hombre y la levadura, en estos momentos estamos muy lejos de dispo-ner de un catálogo su�ciente de componentes y relaciones que nos permita generar modelos completos. Además, no solo nos falta mucha información, sino que desconocemos muchos componentes clave en el funcionamiento de dichas redes, como lo demuestra la existencia de nodos centrales o hubs ocultos. Por tanto, con datos incompletos e información determinante no disponible, es obvio concluir que nos encontramos muy lejos de poder generar modelos consistentes que nos permitan reproducir y entender el funcionamiento de dichos sistemas complejos [1,2].
La insu�ciencia de datos no sería en sí demasiado preocupante si supiéramos que es sólo cuestión de tiempo que la aplicación recurrente de las técnicas experimentales actuales termine la caracterización de dichos espacios oscuros. Pero desafortunadamente, el análisis funcional de la materia oscura nos indica que nuestra exploración experimental de este espacio es sesgada, ignorando de forma sistemática regiones funcionalmente fundamentales, sin las cuales difícilmente alcanzaremos
La “materia oscura” de los sistemas biológicos
Juan Antonio García RaneaInvestigador Ramón y Cajal. Departamento de Biología Molecular y
Bioquímica. Universidad de Málaga. [email protected]
64

Diciembre 2011
Vol.4 | Nº 136
a comprender el funcionamiento de los sistemas moleculares. Todos conocemos las limitaciones y el bajo rendimiento que distintas técnicas experimentales presentan en la caracterización, por ejemplo, de proteí-nas de membrana, las cuales están subrepresentadas en las bases de datos actuales, o las limitaciones que métodos como el sistema de doble híbrido en levadura presentan en la detección de interacciones transito-rias como, por ejemplo, las que se producen en vías de señalización y cascadas de regulación protagonizadas por las proteínas quinasas tan abundantes en los proteomas de eucariotas complejos [3].
A la luz de los resultados actuales, podemos concluir que el avance del conocimiento en biología de sistemas no solo depende del desarro-llo de nuevos conceptos, sino también de nuevos diseños experimenta-les de alto rendimiento que nos permitan corregir y compensar el sesgo de las técnica aplicadas actualmente, así como completar la caracteriza-ción de las relaciones funcionales que permanecen oscuras y de las que por ahora sólo podemos tener evidencias indirectas.
La cosmovisión del hombre ha sufrido cambios dramáticos a medida que su conocimiento del universo se ha hecho más preciso, pasando de
un modelo geocéntrico a otro heliocéntrico, y posteriormente a la com-probación de la humilde ubicación de nuestro Sistema Solar en un brazo lateral de la galaxia Vía Lactea. Es probable que, de igual forma, se tenga que vencer la resistencia cultural a revisar alguno de los paradigmas y dogmas más queridos por parte de comunidades y grupos cientí�cos bien establecidos en el ámbito de la Biología, lo que implicaría aceptar la idea de que probablemente las proteínas que se piensan centrales en muchos de los modelos experimentales actuales deban compartir su protagonismo con otras proteínas cuyo papel central no ha sido descu-bierto todavía. E incluso, en algunos casos, proteínas ahora consideradas claves deban dejar su papel protagonista a otras en modelos posteriores más precisos a medida que nos adentremos en la “materia oscura” de los organismos vivos.
Bibliografía citada:1. Ranea JA, Morilla I, Lees JG, Reid AJ, Yeats C, Clegg AB, Sanchez-Jimenez F, Orengo C (2010). Finding the "dark matter" in human and yeast protein net-
work prediction and modelling. PLoS Comput Biol. 6: pii: e1000945.2. Wass MN, David A, Sternberg MJ (2011). Challenges for the prediction of macromolecular interactions. Curr Opin Struct Biol. 21: 382-90. 3. Russell RB, Aloy P (2008). Targeting and tinkering with interaction networks. Nat Chem Biol 4: 666–673.
65
!"#$%&'()*+,&($&-(*./(0+1( Albert Sorribas, Ester Vilaprinyó, Rui Alves
Departament de Ciències Mèdiques Bàsiques Institut de Recerca Biomèdica de Lleida (IRBLLEIDA) Universitat de Lleida
23+0%*0/!"#$#%&%#$'('"#$#")$*+,*-%),%$ (-.&"/*/+%#$ 0%#0%$ %+$ .1),"$ 0%$ '(#,*$ 0%$ #1$2"-.+%3(0*04$5($ 2")#(0%&*-"#$,"0"#$+"#$.&"2%#"#$ 61%$ *2"),%2%)$ %)$1)*$
#(-.+%$ 27+1+*8$ %#$0(92(+$(-*:()*&$2;-"$<*$ ++%:*0"$*$ #%&$."#(/+%4$!*$ &%#.1%#,*$%#$+*$ %'"+12(;)$/("+;:(2*$-%0(*),%$#%+%22(;)$)*,1&*+$=$ ",&"#$.&"2%#"#$*>)%#4$?")$ %++"8$ +*$%@.+(2*2(;)$*$ %#,%$ *.*&%),%$ -(#,%&("$%#,A$ #%&'(0*8$ *1)61%$ ,"0*'B*$ 0%/%-"#$%-.+%*&)"#$*$ C")0"$.*&*$ 2"-.&%)0%&$ +"#$-%2*)(#-"#$(-.+(2*D0"#$=$#1#$2")#%21%)2(*#4$E%#0%$ %#,%$.1),"$0%$ '(#,*8$ +*$ F("+":B*$ 0%$ 5(#,%-*#8$ 61%$#%$2*&*2,%&(G*$ ."&$1)*$'(#(;)$#(#,7-(2*$ 2"-"$-*&2"$2")2%.,1*+$.*&*$ +*$2"-.&%)#(;)$0%$ +"#$.&"2%#"#$/("+;:(2"#8$ #1$%'"+12(;)$=$."#(/+%$-*)(.1+*2(;)$/(",%2)"+;:(2*8$ #1&:(;$2"-"$1)*$)%2%#(0*0$*212(*),%$ ,&*#$%+$ 0%#*&&"++"$0%$,%2)"+":B*#$61%$*/&(%&")$1)$()#"#.%2<*0"$-*&$0%$)1%'"#$0*,"#$=$0%,*++%#$*$,"0"#$+"#$)('%+%#$H:%);-(2"8$%#,&12,1&*+8$-%,*/;+(2"8$%,24I4
J"&$ +"$,*),"8$%)$+*$/("+":B*$ 0%$ #(#,%-*#$2")K1=%)$+"#$#(:1(%),%#$.&"/+%-*#L$HMI$N)*$%)"&-%$ 2*.*2(0*0$%@.%&(-%),*+$61%$ .%&-(,%$ 2*&*2,%&(G*&$ #(-1+D,A)%*-%),%$ -12<"#$2"-.")%),%#$0%$ 0(#O),"#$)('%+%#$0%$2"-.+%3(0*08$ HPI$!*$)%2%#(0*0$0%$.&"2%#*&$ %#,*$()C"&-*2(;)8$ HQI$R+$&%,"$0%$ (),%&.&%,*&$ +*$2"-D.+%3(0*08$ %#$ 0%2(&8$ +*$ )%2%#(0*0$ 0%$ :%)%&*&$ 0%#2&(.2(")%#$ 2")2%.,1*+%#$61%$ .%&-(,*)$%),%)0%&$ 0(2<*$ 2"-.+%3(0*04$!*$ /("+":B*$ 0%$ #(#,%-*#$2")++%'*$ 1)$2*-/("$0%$ %#2*+*$ -1=$ (-."&,*),%$%)$+*$:%)%&*2(;)$0%$ 0*,"#$%@.%&(-%),*+%#$=$%+$ &%2")"2(-(%),"$0%$ +*$ (-."&,*)2(*$ 0%$ +*#$<%&&*-(%),*#$2"-.1,*2(")*D+%#8$ 0%#0%$ +*$ )%2%#(0*0$ 0%$ /1%)*#$/*#%#$0%$ 0*,"#$=$ *)",*2(")%#8$ <*#,*$ +*$#(-1+*2(;)$ 0%$ -"0%+"#8$ .*#*)0"$."&$ +*$ &%2")#,&122(;)$0%$ %#,&12,1&*#$%$ (),%D&*22(")%#$-"+%21+*&%#4
R)$%#,%$ 2"&,"$%)#*="$ )"#$2%),&*&%-"#$%)$1)*$ (0%*$ 2%),&*+$61%$ #1/=*2%$%)$+*$ -*="&B*$ 0%$ 21%#O")%#$0%$ +*$ F("+":B*$ 0%$ 5(#,%-*#L$R+$ 2")2%.,"$0%$ &%0$0%$(),%&*22(")%#$H!"#$%&'I$0%$-12<"#$%+%-%),"#4$
!)(4&.4+5%&(1+(0+1(1+(#.%+0/44#6.N)"$0%$ +"#$*'*)2%#$(-."&,*),%#$&%+*2(")*0"#$2")$ +*$ /("+":B*$ 0%$ #(#,%-*#$%#$%+$0%#*&&"++"$0%+$*)A+(#(#$:%);-(2"$*$ :&*)$%#2*+*$ 61%$ <*$.%&-(O0"$2")D
>&-*&8$ 2"-"$ E*&S()$0%3;$2+*&"8$ 61%$ %+$ <"-/&%$ )"$ O%)%$ 1)$ +1:*&$ %#.%2(*+$ %)$ +*$ %'"+12(;)4$T1%#,&*$ %#.%2(%$ O%)%$ 1)$)U-%&"$0%$ :%)%#8$ %),&%$ PVWWW$=$QWWWW8$ #(-(+*&$ *+$&*,;)$=$*$",&*#$%#.%2(%#4$R)$.+*),*#8$%#,%$ )U-%&"$.1%0%8$CA2(+-%),%8$ ,&(.+(2*&#%4$J"&$",&*$ .*&,%8$ +*$ -*="&B*$0%$)1%#,&"#$:%)%#$#")$(07)DO2"#$%)$2<(-.*)27#4
R#,"#$&%#1+,*0"#$0%$ +*$ :%);-(2*$#*210%)$*+:1)*#$(0%*#$.&%2")2%/(0*#$*2%&2*$ 0%+$ #(:)(>2*0"$0%$ +*$ 2"-.+%3(0*0$0%$ +*#$C"&-*#$0%$ '(0*4$X?;-"$.1%0%$#%&$ 61%$ )"$,%):*-"#$-12<"#$-A#$:%)%#$61%$ 1)$&*,;)Y$ !*$ &%#.1%#,*$ O%)%$ 61%$ '%&$ 2")$+*$&%0$0%$ (),%&*22(")%#4$T"$%#$ +"$-(#-"$21*,&"$:%)%#$.&"012(%)D0"$2*0*$ 1)"$."&$ #1$21%),*$ 1)*$ .&",%B)*8$ 61%$21*,&"$.&",%B)*#$(),%&*22(")*0"$0()A-(2*-%),%$ %)$%+$ %#.*2("$=$%+$ O%-."$2%+1+*&$ H0%#*&&"++"I4$!*$ (),%&*2D2(;)$ 0%$ 0('%&#"#$%+%-%),"#8$ 2"-"$ #12%0%$ %)$1)*$ #"2(%0*0$<1-*)*8$ .%&-(,%$ +*$ %-%&:%)2(*$ 0%$ )1%'*#$ .&".(%0*0%#$61%$ )"$%#,A)$.&%#%),%#8$ )($ #")$ *+D2*)G*/+%#8$ ."&$ +"#$%+%-%),"#$ ()0('(01*+%#4$ !*#$ ,72)(2*#$%@.%&(-%),*+%#$<*)$.%&-(O0"$ 2*&*2,%&(G*&$ 1)*$ .*&,%$ 0%$ +*$ 2"-.+%3(0*0$ 0%$ +*#$ (),%&*22(")%#$ *$-12<"#$)('%+%#$=$.%&-(,%$%),%)0%&$61%$%+$)U-%&"$0%$:%)%#$)"$#%*$,*)$(-."&,*),%4$
7869&(5&1+9&$(+.%+.1+0()/$(0+1+$(4&95)+:/$;E%#2&(/(&$ 1)*$ &%0$0%$ (),%&*22(")%#$ %#8$ <*#,*$ 2(%&,"$ .1),"8$ 1)$,&*/*3"$&1O)*&("$."&61%$ 0(#.")%-"#$0%$ ,72)(2*#$%@.%&(-%),*+%#$=$ 2"-.1,*2(")*+%#$
61%$ )"#$*=10*)$ *$ .&"2%#*&$ +*$ ()C"&-*2(;)$.*&*$ "/,%)%&$ 1)*$ */#,&*22(;)$ UO+4$Z#B8$ ."&$ %3%-.+"8$ ."0%-"#$&%.&%#%),*&$ +*#$&%+*2(")%#$ (0%)O>2*0*#$%),&%$

Vol.4 | Nº 136
66
,"0"#$ +"#$:%)%#$61%8$ 0%$ 1)*$ -*)%&*$ 1$ ",&*8$ #%$ &%+*2(")*)$2")$1)$2(%&,"$O."$0%$ 2A)2%&4$[$/(%)8$ ."0%-"#$ &%.&%#%),*&$ ,"0*#$ +*#$ &%+*2(")%#$61%$ #%$ 0*)$%),&%$+"#$-%,*/"+(,"#$=$.&",%B)*#$(0%)O>2*0*#$%)$1)$2(%&,"$O."$0%$27+1+*#4$X\$*<"&*$617Y
!*$ &%#.1%#,*8$ 2"-"$ %)$-12<"#$",&"#$2*#"#8$ 0%.%)0%$ %)$:&*)$-*)%&*$ 0%+$ O."$ 0%$ .&"/+%-*$ 61%$ 61%&*-"#$ &%#"+'%&4$J"&$ %3%-.+"8$ ."0%-"#$.+*)D,%*&)"#$21%#O")%#$*2%&2*$ 0%$ +*$ #(-(+(,10$ 0%$ +*$ %#,&12,1&*$ 0%$ 0('%&#*#$&%0%#$*$0(#O),"#$ )('%+%#4$XJ"0%-"#$ (0%)O>2*&$ .&".(%0*0%#$ /A#(2*#$2"-1)%#Y$ !*$&%#.1%#,*$ %#$61%$ #B$ =$ 61%$ &%0%#$,*)$0(#O),*#$2"-"$%+$ -%,*/"+(#-"8$ (),%&)%,8$ "$+*#$&%+*2(")%#$#"2(*+%#8$.*&%2%)$2"-.*&O&$ 1)$O."$0%$ "&:*)(G*2(;)$61%$ #%$2*&*2,%&(G*$$."&$-12<"#$)"0"#$2")$."2*#$2")%@(")%#$(),%&*22(")*0"$2")$*+:1)"#$)"0"#$*+,*-%),%$2")%2,*0"#4
!*$ #(,1*2(;)$%#$0(#O),*$ 21*)0"8$ 1)*$ '%G$(0%)O>2*0"#$+"#$2"-.")%),%$ #(:)(>2*O'"#$0%$1)*$ &%08$)"#$.+*),%*-"#$&%#"+'%&$21%#O")%#$2"-"L$X."&$617$1)*$ 0%,%&-()*0*$.%&#")*$ 0%#*&&"++*$1)$O."$0%$%)C%&-%0*0$ =$",&*8$ *.*&%),%-%),%$ #(-(+*&8$ )"Y$[8$ X."&$ 617$ 1)$O."$0%$ ,&*,*-(%),"$ %#$%C%2O'"$%)$ 1)*$.%&#")*$=$)"$%)$",&*Y
</$(9/%+9=%#4/$(/)(0+$4/%+Z1)61%$ *$*+:1)"#$.1%0*$#"&.&%)0%&"#8$ .*&,%$ 0%$ +*$ &%#.1%#,*$%#,A$ %)$ +*#$ -*,%-AO2*#4$R+$ *)A+(#(#$ (),1(O'"$0%$ #(,1*2(")%#$2"-.+%3*#$=$ +*$ .&%0(22(;)$
0%$ 2*-/("#$0()A-(2"#$C&1,"$0%$ +*$ (),%&*22(;)$0%$ -12<"#$%+%-%),"#$)"$%#8$ %)$ :%)%&*+8$ ."#(/+%$ #()$%+$ 2")21&#"$0%$ +*#$ -*,%-AO2*#4$R+$ &*G")*-(%),"$ ()D,1(O'"$=$+()%*+$0%+$O."L$5($Z$#%$ *2O'*8$ #%$ *2O'*$ F4$Z+$ (),%&*22(")*&$2")$F8$ ?$&%:1+*$+*$ %@.&%#(;)$0%]8$ O%)%$1)$+B-(,%$ ."&61%$ +*#$(),%&*22(")%#$#")$)"D+()%*D+%#4$Z#B8$ %),%)0%&$ +*$.&":&%#(;)$0()A-(2*$0%$ +"#$.&"2%#"#$2%+1+*&%#$=$ .&%0%2(&$2;-"$ #%$ 2"-."&,*&A$+*$ 27+1+*$ &%61(%&%$ %+$ 1#"$0%$ <%&&*-(%),*#$-*,%-AOD2*#4
!"#$-"0%+"#$-*,%-AO2"#$)"#$*=10*)$%)$-12<*#$C*2%,*#$.&A2O2*#8$ ()2+1=%)0"$%+$ 0(#%^"$0%$ 2"2<%#8$%+$ &%)0(-(%),"$0%."&O'"8$ %+$*)A+(#(#$%2");-(2"8$%,24$!*$ /("+":B*$ 0%$ #(#,%-*#$ <*$ #(,1*0"8$ %),&%$ ",&"#$*'*)2%#8$ *$ +"#$ -"0%+"#$-*,%-AO2"#$ %)$%+$ 2%),&"$ 0%$ +*#$<%&&*-(%),*#$)%2%#*&(*#$.*&*$ %),%)0%&$ +*$/("+":B*4
>&1+)&$?(*./(5/$/0+)/(5/0/(+.%+.1+0()/(4&95)+:#1/1?1*)0"$%+$.&"/+%-*$ %#$2"-.+%3"8$ )%2%#(,*-"#$2*.,*&$ +*$ %#%)2(*$0%+$-(#-"$=$ &%.&%#%),*&+"$%)$1)$-"0%+"$2")2%.,1*+$H-*,%-AO2"I$UO+$ 61%$ .%&-(,*$
#1$ #"+12(;)4$R)$ %#,%$ .1),"8$ +*$ /("+":B*$ .&%#%),*$ -12<"#$.&"/+%-*#$ -%,"0"+;:(2"#4$J"&$ 1)*$ .*&,%8$ %+$ )U-%&"$ 0%$ %+%-%),"#$0%$ +*$ C"&-*$ 0%$ '(0*$ -A#$#(-.+%$ %#$%)"&-%4$T%2%#(,*-"#$&%.&%#%),*&$ #1#$(),%&*22(")%#$H61%$ %)$:&*)$.*&,%$ #")$0%#2")"2(0*#I$0%$ 1)*$ C"&-*$61%$ ."0*-"#$.&%0%2(&$ 617$ .*#*&A$ %)$&%#.1%#,*$ *$2(%&,"#$%#_-1+"#4$Z0%-A#$0%/%&B*-"#$#%&$2*.*2%#$0%$ (0%)O>2*&$ 617$ .&()2(.("#$0%$ "&:*)(G*2(;)$#")$2"-1)%#$*$ 0(C%&%),%#$&%0%#8$ C&1,"$0%$ +*$%'"+12(;)4
Z2,1*+-%),%8$ +"#$-"0%+"#$-*,%-AO2"#$#")$C1)0*-%),*+%#$%)$F("+":B*$ 0%$ 5(#,%-*#4$J"&$%3%-.+"8$ +"#$ -"0%+"#$0%$ 2(2+"$2%+1+*&$ <*)$.%&-(O0"$2"-D.&%)0%&$ -%3"&$%+$2"),&"+$ 0%$ +*#$0(#O),*#$ C*#%#$0%+$ 2(2+"4$!*$ (-."&,*)2(*$0%$ %#,%$ 2")"2(-(%),"$.*&*$ %),%)0%&$ +"#$.&"2%#"#$2*)2%&"#"#$%#$%'(0%),%$ =$.%&D-(,%$ *.&%2(*&$ +*$ (-."&,*),%$ 2"),&(/12(;)$0%$ %#,*$ ,72)(2*$ %)$ +*$ &%*+(0*0$0%$ +*$ ()'%#O:*2(;)$*.+(2*0*4$Z$ ",&"$)('%+8$ #%$ %#,A)$&%*+(G*)0"$:&*)0%#$%#C1%&G"#$.*&*$ -"0%+*&$ ;&:*)"#$<1-*)"#8$."&$ %3%-.+"$%+$<B:*0"8$2"-"$1)$.*#"$C1)0*-%),*+$ .*&*$0(#.")%&$ 0%$ 1)$+*/"&*,"&("$/*&*,"$0")0%$ .&"/*&$ )1%'*#$,%&*D.(*#4$R)$%#,%$ #%)O0"8$ %+$1#"$0%$ -"0%+"#$-*,%-AO2"#$.1%0%$#%&$1)$C*2,"&$-1=$ (-."&,*),%$ 61%$ 2"),&(/1=*$*$ &%012(&$%+$2"#,%$ 0%$ 0%#*&&"++*&$ )1%'"#$,&*D,*-(%),"#8$/%)%>2(*)0"$*$+"#$.*2(%),%#$2")$1)$-%)"&$2"#,%$0%$2"-%&2(*+(G*2(;)4
R+$ 21%++"$ 0%$ /",%++*$ %)$,"0*$ %#,*$ *2O'(0*0$ %#$+*$ "/,%)2(;)$ 0%$ 0*,"#$>*/+%#$ *2%&2*$ 0%$ +"#$ 2"-.")%),%#$ 2%+1+*&%#$61%$ .%&-(,*)$1)*$ -"0%+(G*2(;)$&%*+(#,*4$!*#$21%#O")%#$ ,72)(2*#$ &%+*2(")*0*#$2")$ +*$ #(-1+*2(;)$."&$ "&0%)*0"&8$ +*$ *.+(2*2(;)$ 0%$ ,72)(2*#$0%$ ".O-(G*2(;)$ 0%$ .&"2%#"#$.*&*$ %'*+1*&$ %+$%C%2,"$ 0%$ 2*-/("#$%)$%+$ #(#,%-*8$ %,248$ .1%0%)$0*&#%$ ."&$ &%#1%+,*#4$!*$2*.*2(0*0$*2,1*+$ 0%$2A+21+"$0%$ +"#$"&0%)*0"&%#$.%&-(,%$ *C&"),*&$ :&*)0%#$.&"=%2D,"#$#()$0%-*#(*0"#$.&"/+%-*#4$R#$0%$%#.%&*&$61%$ %)$1)$C1,1&"$.&;@(-"8$ +*#$,72)(2*#$%@.%&(-%),*+%#$0%)$1)$#*+,"$21*+(,*O'"$=$.%&-(,*)$+*$ "/,%)2(;)$0%$0(2<"#$0*,"#$2")$+"#$61%$.+*),%*&$+*$#"+12(;)$0%$.&"/+%-*#$0%$:&*)$(),%&7#$-70(2"4
7@*A(50&9+$/$(5&1+9&$(4*95)#0(+.(*.(B*%*0&(506"#9&;R)$+"#$.&(-%&"#$O%-."#$0%$ +*$*2%.,*2(;)$:%)%&*+$0%+$ 2")2%.,"$0%$ F("+":B*$0%$ 5(#,%-*#8$<*2%$ 1)"#$0(%G$*^"#8$ #%$ .&"013"$1)$2(%&,"$%#,*++(0"$0%$ .&%D
0(22(")%#4$R),&%$ %++*#8$ +*$ ."#(/(+(0*0$0%$2*&*2,%&(G*&$ %+$ :%)"-*$ 0%$ 2*0*$.%&#")*$ */&%$ :&*)0%#$."#(/(+(0*0%#$ -70(2*#$ *+$.%&-(O&$ (0%)O>2*&$ :%)%#$*+,%&*D0"#4$R#$%'(0%),%$ 61%$ +*$*1#%)2(*$ "$0(#C1)2(;)$0%$1)$:%)$0%,%&-()*0"$.1%0%$ C*2(+(,*&$%+$0%#*&&"++"$0%$ 1)*$.*,"+":B*8$2"-"$%#$%+$2*#"$0%+$ :%)$F`?ZM$%)$%+$2A)2%&$ 0%$ -*-*4$5()$%-/*&:"8$ %)$-12<"#$2*#"#$ 0(2<*#$ *+,%&*2(")%#$ .&"'"2*)$0(#C1)2(")%#$61%$ .1%0%)$2"-.%)#*&#%8$ %)$.*&,%8$ ."&$ %+$ ()2&%-%),"$ 0%$C1)2(;)$0%$",&"#$:%)%#$"$.&"2%#"#4$R#,"$0(>21+,*$&%+*2(")*&$+*$.*,"+":B*$HC%)"O."I$2")$+*$:%)7O2*$0%$2*0*$()0('(01"4
Z1)61%$ +"#$%#C1%&G"#$ 0%$ ()'%#O:*2(;)$#")$-1=$ (-."&,*),%#8$ +*$ 2"-.&%)#(;)$0%+$ 2")2%.,"$ 0%$ &%0$ )"#$ <*2%$ #%&$ .&10%),%#$ <*2(*$ +*$ ."#(/(+(0*0$0%$0%#*&&"++*&8$ %)$1)$C1,1&"$ ()-%0(*,"8$ 1)*$ -%0(2()*$ '%&0*0%&*-%),%$ .%&#")*+(G*0*8$ ."&$ C*+,*$0%$ ()C"&-*2(;)$*2%&2*$0%$ +"#$.&"2%#"#$2%+1+*&%#$(-.+(2*0"#$%)$1)*$ %)C%&-%0*04$5()$ %-/*&:"8$ 0(#.")%-"#$0%$ *+:"$'*+("#"4$5*/%-"#$+"$61%$ )"#$C*+,*4$J"&$ +"$,*),"8$ #%$ ,&*,*$ 0%$ .")%&$ -*)"#$*$ +*$ "/&*$ =$ 0%$ #%&$ "#*D0"#4$?1*)0"$#%$ .&".1#"$%+$ .&"=%2,"$ :%)"-*$ <1-*)"$ -12<"#$0(3%&")$61%$ %&*$ 1)*$ ,*&%*$ %@2%#('*-%),%$ 2"-.+%3*4$Z+$>)*+8$ %+$ .&"=%2,"$#%$ 2"-.+%,;$ %)$1)$O%-."$&72"&08$0*)0"$.*#"$*+$(),%&7#$:%)%&*+$."&$+*$F("+":B*$0%$#5#,%-*#4
C+()/(1+$40#54#6.(1+()/(D#1/(/()/(40+/4#6.(1+(.*+D&$(&0,/.#$9&$N)"$ 0%$ +"#$ *#.%2,"#$.&A2O2"#$(-."&,*),%#$0%$ +*$ /("+":B*$ 0%$ #(#,%-*#$%#$+*$ ."#(/(+(0*0$0%$ -"0(>2*&$ "&:*)(#-"#$H/(",%2)"+":B*I$ "$0%$ 2&%*&$ )1%'"#$
"&:*)(#-"#$ =$ (),&"012(&$ 2(&21(,"#$-%,*/;+(2"#$0(#%^*0"#$ .*&*$ &%*+(G*&$ 0%,%&-()*0*#$ C1)2(")%#$ HF("+":B*$ 5(),7O2*I4$R)$*-/"#$2*#"#8$ )"#$%)2"),&*-"#$*),%$ )1%'*#$."#(/(+(0*0%#$61%8$ 2"-"$-12<"#$2*#"#$%)$2(%)2(*8$ */&%)$:&*)0%#$(),%&&":*),%#$7O2"#$*1)61%$.1%0%)$.&"."&2(")*&$ :&*)0%#$*'*)2%#$.&A2DO2"#4$!*$/(",%2)"+":B*$/*#*0*$%)$1)$*)A+(#(#$&*2(")*+$0%$ +*#$.&".(%0*0%#$0%$ +"#$.&"2%#"#$2%+1+*&%#$H():%)(%&B*$ -%,*/;+(2*I$0%/%$ .%&-(O&$+*$ "/,%)2(;)$0%$"&:*)(#-"#$-"0(>2*0"#$61%$ ()2&%-%),%)8$ ."&$ %3%-.+"8$ +*$.&"0122(;)$0%$ 0%,%&-()*0"#$.&"012,"#$0%$ (),%&7#$ .&A2O2"4$R+$ *<"&&"$%)$%@.%&(-%),"#8$*)(D-*+%#$0%$ +*/"&*,"&("8$%,248$ .1%0%$ #%&$-1=$ (-."&,*),%4$R)$%#,%$ #%)O0"8$ +*$/(",%2)"+":B*$%#$=*$ 1)*$ 0%$ +*#$*2O'(0*0%#$%2");-(2*#$-A#$0()A-(2*#$=$ 2")$1)$C1,1&"$-1=$(),%&%#*),%4
>+.$/:+!*$/("+":B*$0%$#(#,%-*#$O%)%$=*$&%#1+,*0"#$%#.%2,*21+*&%#$%)$0('%&#"# $A-/(,"#8$.%&"$.+*),%*$&%,"#$,*)$*-.+("#$61%$#%$ )%2%#(,*&A)$
*^"#$.*&*$ +":&*&+"#4$5($"#$#%012%$ +*$ (0%*$ 0%$,&*/*3*&$%)$%#,%$ A-/(,"8$ (),%&%#*&"#$."&$ +*$/("+":B*8$+*$ 2"-.1,*2(;)8$ +*#$-*,%-AO2*#8444$ a=$#%0$-1=$21&("#"#b
Lecturas recomendadas para saber más: Guimerà R, Sales-Pardo M (2009) Missing and spurious interactions and the reconstruction of complex networks. Proceedings of the National Academy of Sciences USA
106(52):22073-22078. Koonin EV (2012) The logic of chance: The nature and origin of biological evolution. FT Press Science. New Jersey, USA. Nalylor S, Chen JY (2010) Unraveling human complexity and disease with systems biology and personalized medicine. Personalized Medicine 7(3):275-289. Scott Turner J (2007) The tinkerer’s accomplice: How design emerges from life itself. Harvard University Press. Cambridge, Massachussets, USA.

Diciembre 2011
Vol.4 | Nº 136
Cinco preguntas sobre Biología Sintética
67
Salvador Durán Estudiante de doctorado de la Universitat Pompeu Fabra (IBE-UPF) [email protected]
contiene hoy más de 17000 piezas únicas llamadas “biobricks” (promotores, termina-dores, secuencias codi-�cantes completas o dominios de proteínas, RBS, plásmidos, etc.), estandarizadas de tal forma que permiten generar constructos arbitrariamente gran-des usando sólo cuatro enzimas de restricción distintas. Además de poder depositar y con-seguir piezas nuevas para seguir investigan-
do, el Registry permite anotar características, funciones e incluso incompatibilidades con otras secuencias a la vez que sus contribuidores ofrecen conocimientos técni-cos variados.
Vale la pena mencionar que ligada a la mencionada iniciativa está la iGEM Competition (International Genetically Engineered Machine Competition), en la que estudiantes aún por licenciarse de todo el mun-do, compiten por construir la “maquina biológica” más ingeniosa. Con ayuda de profesores y tutores, los alumnos dedican un verano a diseñar las redes de genes, construirlas ensamblando o clonando las piezas pertinentes y probar los constructos resultantes, partiendo siempre de un conjunto de piezas estandarizadas comunes, enviadas desde el Regis-try en el MIT.
¿Qué se ha logrado? Si bien, los artículos antes mencionados son los pilares teóricos
sobre los que se funda la Biología Sintética, es mucho más interesante hablar de sus aplicaciones y resultados prácticos, ya que, potencialmen-te, son muchos los procesos que pueden bene�ciarse del uso de ele-mentos biológicos programados. Características como una elevada especi�cidad o simplemente la capacidad autoreplicativa de estos sis-temas hacen que la sustitución de algunos procesos �sicoquímicos por otros puramente biológicos sea, a parecer de muchos, un tema de cuándo o cómo antes que de por qué.
Algunas de las áreas que han recibido especial atención por parte de la comunidad de biólogos sintéticos son la biotecnología, combustibles y energía, la ciencia de materiales, la bioremediación, la medicina y también el rediseño o creación de novo de sistemas biológi-cos.
En el caso de la biotecnología sobresale el artículo de Jay D. Keasling y colaboradores (5), en el que proponen una alternativa semi-sintética a la artemisina extraída directamente de la planta A. annua para el tratamiento de la malaria. Esto se consigue alterando la ruta metabó-lica del mevalonato endógena en S. cerevisiae a la vez que añadiendo una enzima de la propia A. annua para aumentar la productividad de la cepa hasta niveles no logrados anteriormente.
En el área de la bioremediación existen gran cantidad de iniciati-vas en las áreas de �jación de metales pesados, biodetección de conta-minantes varios o biodegradación mediante cepas modi�cadas genéti-camente. Pero la parte fascinante ocurre cuando ese conocimiento se transforma en aplicaciones reales, aunque casi de ciencia �cción, como es el caso de las plantas modi�cadas genéticamente por Medford y colaboradores (6). En su articulo, publicado recientemente en la revista PLoS ONE, se explica cómo su equipo consiguió rediseñar una vía de señalización tal que, una vez alterada, provoca que las plantas cambien su pigmentación verde a una más clara si se exponen a explosivos.
El área de Biomedicina (sobretodo en las materias de cáncer e infecciones), también ha visto el uso de enfoques muy creativos para el tratamiento de enfermedades. Un ejemplo podría ser la estrategia pro-puesta por la universidad de Ljubljana en Eslovenia para combatir la infección por VIH (8), presentada en el contexto de la competición iGEM de 2007. Esta estrategia está basada en la colocalización de los correcep-tores del VIH CD4 y CCR5 en la super�cie de las células que están siendo infectadas para complementar en la cara intracelular de la membrana dos dominios de la proteasa TEV. De esta forma se transmite la señal de infección mediante la proteólisis de sustratos arti�ciales hasta las estra-tegias antivíricas.
¿Qué es la Biología Sintética?La Biología Sintética es una disciplina relativamente nueva para
la cual existen una gran cantidad de de�niciones, aunque prácticamente todas ellas incorporan las ideas de biología molecular, genética y inge-niería. Para muchos cientí�cos se trata de reordenar secuencias de ADN naturales, generando constructos o redes arti�ciales de genes que ten-gan un comportamiento no existente en la naturaleza y de carácter aplicado. Por �losofía se parece mucho a la Biología de Sistemas, ya que también intenta integrar en un marco matemático la biología descripti-va usando modelos y simulaciones.
Está claro dónde participan la Biología Molecular, de Sistemas y laGgenética en todo esto, pero ¿y la Ingeniería? Conceptos y problemas tradicionalmente tratados desde la ingeniería constituyen el sustrato teórico para entender y modelizar muchos procesos biológicos, sean naturales o arti�ciales. La modularidad, la escalabilidad, el tratamiento del ruido y la señal o la propia lógica digital de los circuitos electrónicos son ejemplos de ello. Un caso práctico de este paralelismo entre lo bio-lógico y lo electrónico es el detector de contornos (1), un problema no trivial en el campo de la computación y análisis de imagen que se re-suelve brillantemente en el artículo referenciado mediante Escherichia coli que albergan pequeñas redes génicas arti�ciales.
¿Cuál es su origen? ¿Por qué no existió la Biología Sintética desde los años 80 o 90?
La verdad es que no se trata sólo de modi�car, sino también de la exten-sión y la intención de dicha modi�cación. Mayoritariamente, las altera-ciones de la biología natural que se hacían antes de la biología sintética eran perturbaciones con la intención de ver cómo reaccionaba el siste-ma de estudio o expresiones de una/s pocas proteínas para recuperar una actividad perdida por las células (por ejemplo gran cantidad de modi�caciones en terapia génica). Sin embargo, el cambio de concep-ción está en que se busca introducir no sólo una expresión de una o varias características como se haría en una la aproximación biotecnoló-gica, sino un comportamiento en sí. En otras palabras, si hay que intro-ducir una variación fenotípica en una célula se intenta controlar cuándo, cómo y durante cuánto tiempo tendrá lugar esa alteración. De esta forma, incluyendo “sencillas” redes de reguladores de la transcripción se pueden conseguir muy variadas o complejas dinámicas: constantes y poco sensibles al ruido (2), oscilantes en el tiempo (3), o incluso múlti-ples estados fenotípicos distintos excluyentes entre sí (4).
Una de las razones de que el “boom” de esta disciplina haya teni-do lugar justamente ahora es el esfuerzo por parte de distintas institu-ciones, como el MIT (Massachusetts Institute of Tecnology), en la creación de una comunidad de biólogos sintéticos, con canales de comunicación propios (congresos, revistas, etc.) y repositorios gratuitos de secuencias de ADN como el Registry of Standard Biological Parts. Dicho repositorio
Figura 1: Esquema del detector de contornos. Los circuitos genéticos reescritos en forma de puertas lógicas son simulados numéricamente (derecha) construidos y testados en E. coli (izquierda). Imagen reelaborada a partir de una incluida en la referencia (1).
Figura 2: Figura extraída de la referencia (6) –con licencia CopyLeft Creative Commons- en la que se observa cómo plantas individuales cambian de color (sólo las dos de arriba) a medida que aumenta el tiempo de exposición (horas, en el eje horizontal) a tierra con TNT.

Vol.4 | Nº 136
¿Cuáles son los próximos desafíos?A mi parecer, el principal obstáculo que existe para la expansión
de la Biología Sintética es la legislación actual. Sin juzgar si es apropiada o no (ni estoy capacitado para emitir un juicio al respecto ni es el objeti-vo de este artículo), basta decir que limita el uso de los diseños sintéticos en áreas tan cruciales como el uso medioambiental o médico. Así pues, los biólogos sintéticos deberán hacer un especial esfuerzo por intentar convencer a la sociedad que, de la misma forma que los transgénicos tienen un momento y un lugar en el sistema, los diseños arti�ciales pue-den ser soluciones reales y deseables a trastornos, enfermedades o pro-blemas medioambientales. En esta línea, es muy probable que haya que hacer un esfuerzo cientí�co especial en mejorar la seguridad y control y reducir la mutabilidad de las células alteradas.
Por otra parte, los recientes avances técnicos en fabricación y ensamblaje de genomas (9) indican que quizás en el espacio de unos años podamos observar no la copia sino el diseño dirigido de nuevos genomas y nuevas especies, con todos los riesgos y bene�cios que esto incluya.
¿Dónde encontrar mas información?Para los lectores que quieran saber más respecto a los temas de
Bioética y históricos asociados con la Biología Sintética, la pagina web http://www.synbioproject.org/ los redirigirá a textos importantes del área, como por ejemplo el informe de la comisión presidencial de Esta-dos Unidos de América respecto al potencial destructivo de la Biología Sintética.
Aquellos que deseen conocer lo último en material educativo, conferencias y grandes proyectos de colaboración en el área deberían buscar en http://syntheticbiology.org/, desde el cual se organiza anual-mente el Synthetic Biology International Meeting.
Una web imprescindible es http://ung.igem.org/, donde se alma-cenan los resultados y las investigaciones de todos los equipos de todas las ediciones del International Genetically Engineered Machine competi-tion anteriormente mencionado.
68
Bibliografía citada:(1) Tabor J, Salis H, Simpson Z, Chevalier A, Levskaya A, Marcotte E, Voigt CA, Ellington AD. A Synthetic Genetic Edge Detection Program. Cell 137, 1272-1281, 2009. (2) Becskei A, Serrano L. Engineering stability in gene networks by autoregulation. Nature 405, 590-593, 2000.(3) Elowitz MB, Leibler S. A synthetic oscillatory network of transcriptional regulators. Nature 403, 335-8, 2000. (4) Gardner TS, Cantor CR, Collins JJ. Construction of a genetic toggle switch in Escherichia coli. Nature 403, 339-42, 2000.(5) Ro DK, Paradise EM, Ouellet M, Fisher KJ, Newman KL, Ndungu JM, Ho KA, Eachus RA, Ham TS, Kirby J, Chang MC, Withers ST, Shiba Y, Sarpong R, Keasling JD. Production of
the antimalarial drug precursor artemisinic acid in engineered yeast. Nature 440, 940-3, 2006.(6) Antunes MS, Morey KJ, Smith JJ, Albrecht KD, Bowen TA, Zdunek JK, Troupe JF, Cuneo MJ, Webb CT, Hellinga HW, Medford JI. Programmable Ligand Detection System in Plants
through a Synthetic Signal Transduction Pathway. PLoS One. 2011; 6(1): e16292.(7) Purnick PEM, Weiss R. The second wave of synthetic biology: from modules to systems. Nature Reviews Molecular Cell Biology 10, 410-422, 2009. (8) Consultar toda la información en la web: http://parts.mit.edu/igem07/index.php/Ljubljana(9) Gibson DG, Benders GA, Andrews-Pfannkoch C, Denisova EA, Baden-Tillson H, Zaveri J, Stockwell TB, Brownley A, Thomas DW, Algire MA, Merryman C, Young L, Noskov VN,
Glass JI, Venter JC, Hutchison CA 3rd, Smith HO. Complete chemical synthesis, assembly, and cloning of a Mycoplasma genitalium genome. Science 319, 1215-20, 2008.
Biología de Sistemas... ¿qué biología de sistemas?Miguel Ángel Medina Torres [email protected]
Catedrático del Departamento de Bioquímica y Biología Molecular, Universidad de Málaga
En diciembre de 2010 tuve el honor y la satisfacción de participar como ponente invitado en la Reunión Científica interna-cional Systems Biology: Bridging the Gaps between Disciplines organizada en Barcelona por la Red Española de Biología de Siste-mas (REBS). En concreto, se me comisionó impartir una ponencia a continuación de la impartida por el Dr. Adriano Henney dentro de una sesión dedicada a Medicina Personalizada. El tema que escogí para mi ponencia fue "Systems Biology Approaches to the Study of Emergent and Rare Disease" y lo que viene al caso de este número monográfico de Encuentros en la Biolo-gía es que me permití la libertad de concluir la ponencia con unas reflexiones personales alrededor del título que he escogi-do para el presente artículo. También hacia finales del año 2010 apareció publicada en Annual Reviews of Cell and Deve-lopmental Biology una revisión cofirmada por Trey Ideker (uno de los más reconocidos "biólogos de sistemas") y titulada "A Decade of Systems Biology". ¿Realmente una década? En todo caso, mi reflexión personal y esta revisión fueron los estímulos
que hicieron nacer en mí la idea de proponer al equipo editorial de Encuentros en la Biología la conveniencia de dedicar un número monográfico al tema. El proceso de preparación ha sido largo pero satisfactorio. Debo hacer constar que la revista ha recibido mensajes de apoyo de cuantos científicos nacionales (más alguno internacional) hice llegar la invitación para que cooperasen en este número, aunque no todos pudie-ron aceptar la invitación por motivos de agenda. En todo caso, en Encuentros en la Biología tenemos motivos para sentirnos orgullosos por haber consegui-do reunir una colección de excelentes contribuciones procedentes de investigadores de las Universidades de La Laguna, Lleida, Murcia y Pompeu Fabra, además de las provenientes de la Universidad de Málaga, donde esta publicación se edita. Sin duda faltan nombres en la nómina de autores, no están todos los que son, pero sí son todos los que están. Me congratula además poder constatar que los distintos enfoques aportados por las diferentes contri-buciones ayudan mucho a ofrecer una visión caleidoscópica de qué representa el tema y a testimoniar que hay distintas maneras de entender qué es la biología de sistemas. Al mismo tiempo, esos diversos y dispares enfoques arrojan luz acerca de aspectos esenciales en los que los convencidos de la nece-sidad de una biología de sistemas coinciden. Precisamente, un aspecto en el que están de acuerdo los interesantes artículos aportados por Alves et al., Cánovas y Torres es la existencia de múltiples definiciones de "Biología de Sistemas". La Figura 1 (pagina 58) muestra el ejemplo de dos definiciones: una extraída de la versión inglesa de Wikipedia y la otra como cita del Presidente del Institute for Systems Biology de Seatle (Estados Unidos). La Figura 2 (página 58) muestra un pequeño repertorio de diferentes formas de visualizar cómo distintos autores y grupos de investigación entienden qué es la biolo-gía de sistemas. Hay quienes, desde una perspectiva meramente instrumental, consideran la Biología de Sistema exclusivamente como una forma de adquisición masiva de información biológica y su manejo computacional. O incluso quienes estiman que el término "Biología de Sistemas" no es sino una "marca" de moda que "vende".
Alves et al., señalan acertadamente que la Biología de Sistemas no nació hace diez años, sino que -en realidad- desde muchos años antes ha habido científicos que han estado haciendo una Biología de Sistemas cuando ese nombre no había sido "institucionalizado". La Figura 3 (página 58) muestra algunos de los científicos con notables contribuciones a la comprensión sistémica de la biología, entre los que hay físicos, químicos, matemáticos, ecólo-gos, biólogos experimentalistas y biólogos teóricos. Una mirada a la historia pasada de la ciencia puede resultar muy esclarecedora.
Un poco de perspectiva históricaSin lugar a duda, el triunfo de la biología contemporánea se ha fundamentado en el empleo del método reduccionista para indagar hasta las últimas
bases moleculares de los fenómenos biológicos. El gran desarrollo de la Bioquímica y Biología Molecular y de la Genética durante el siglo XX posibilitó la emergencia de la gran revolución científico-tecnológica del DNA recombinante en el último cuarto del siglo pasado, que a su vez ha traído de la ma-no los grandes avances de los primeros años del presente siglo asociados a las "ómicas". Este comienzo de milenio ha contemplado cómo la biología ocupa su actual privilegiada posición de frontera. Sin embargo, las bases conceptuales de este triunfante enfoque de la ciencia se remonta, al menos, hasta el siglo XVII, con la exitosa formulación del racionalismo y del mecanicismo. Por una parte, el racionalismo cartesiano estableció un programa de investigación para las preguntas fundamentales del pensamiento protocientífico: descomponer los problemas "complejos" en otros más simples (análisis) para, una vez resueltos estos por separado, alcanzar una comprensión y/o resolución de los problemas "complejos" iniciales por combinación de los resultados parciales (síntesis). En esto consiste el reduccionismo. Por otra parte, la construcción de los primeros relojes fiables para una medición del tiempo con una precisión sin precedentes contribuyó a configurar una imagen mecaniscista del mundo según la cual los diversos sistemas naturales no serían sino mecanismos/maquinarias más o menos complejas. El espectacular triunfo y asentamiento de la primera ciencia moderna, la Física, prestigió
A DEBATE

Diciembre 2011
Vol.4 | Nº 136
69
el mecanicismo y el método reduccionista como la forma de "hacer" ciencia con éxito. Sin embargo, las emergentes ciencias naturales contemplaron hasta bien entrado el siglo XIX una pugna entre los dos enfoques complementarios de la reducción y del estudio sistémico. El primero, con un método y programa de investigación precisos, triunfó (al menos, cuantitativamente) sobre el segundo, que carecía por entonces de ellos. La Biología mecanicista llegó hasta sus últimas consecuencias en el famoso libro The Mechanistic Conception of Life (1912), en el que Jacque Loeb consideraba que los organismos no son sino unas máquinas particularmente complejas.
Pronto se formularon dos tipos de objeciones al reduccionismo biológico. Por una parte, algunos científicos utilizaban en sus argumentos el principio aristotélico de que el todo está siempre por encima de y es más que la suma de sus partes. En 1926, Jan Smuts acuñó el término "holismo" para refe-rirse a este principio. [Merece la pena remarcar aquí que, al igual que hubo practicantes de la Biología de Sistemas antes de su emergencia -tal como argumentan Alves et al. en su articulo-, también hubo científicos holistas antes de que se introdujera el término holismo. El científico ruso Vladimir Vernadsky -que aparece en la Figura 3-, con sus estudios sobre la biosfera (The Biosphere, 1926; versión original en ruso; hay una interesante edición de 1998 anotada y comentada en inglés), sería un destacado ejemplo de ello]. Este principio fundamenta que las propiedades de los sistemas no puedan ser reducidas a las de sus constituyentes. O, dicho de otra forma, la integración de diversas partes en un sistema hace emerger propiedades nuevas de éste. [Un sencillo ejemplo de la química elemental aclarará esta idea fundamental: aunque toda molécula de agua está formada por dos átomos de hidrógeno y uno de oxígeno, las propiedades físico-químicas del agua no pueden ser explicadas a partir de las propiedades del hidrógeno y del oxígeno]. Por otra parte, algunas sencillas investigaciones sobre el cerebro y sobre el desarrollo embrionario mostraron que la estructura de un sistema completo realmente orquestaba y constreñía el comportamiento de sus partes integrantes. El capítulo introductorio de un libro escrito por Roger Williams (1956) sobre la "individualidad bioquímica " en animales marcó el final de las posturas estrictamente mecaniscistas en Biología (aunque no el del reduccionismo, que aún no había alcanzado su actual edad dorada): demostró que el "individuo medio", la media estadística no tiene ninguna realidad biológica.
En este punto, puede resultar conveniente remarcar que, aunque frecuentemente se muestran al reduccionismo y al holismo como enfrentados, es posible encontrar vías de reconciliación que entiendan que, más bien, ambos enfoques son complementarios. De hecho, un preclaro ejemplo de biólogo reduccionista, el premio Nobel François Jacob, dejó escrito en su muy influyente libro "La lógica de lo viviente" (1974) que "todo objeto de estudio de la biología es un sistema de sistemas". [Torres parafrasea, matiza y enriquece esta cita en su excelente artículo cuando titula uno de sus subapartados "Los seres vivos son complejos sistemas de sistemas complejos"].
Hasta aquí la palabra sistema ha aparecido reiteradas veces sin haber quedado aún definida. Asumiendo las prevenciones que Torres expone frente a todo intento de definición, todo sistema puede ser entendido -en principio- como una red de componentes interconectados y mutua-mente dependientes que configuran un todo unificado. Es destacable que las propiedades de tales sistemas son el resultado de dos características importantes: los sistemas tienen una estructura jerárquica y esta estructura es mantenida a través de las interrelaciones que configuran su estructura de redes complejas (para más información sobre la importancia de las redes complejas y su papel en la Biología de Sistemas, me remito al excelente artículo de Sorribas et al.). Una vez clarificada la estructura jerárquica de los sistemas, una importante generalización fue esbozada (1937), sugerida y finalmente formalizada por Ludwig von Bertalanffy en su teoría general de sistemas (1950, 1968). Bertalanffy argumentó que todos los sistemas comparten la propiedad común de estar compuestos por componentes interconectados (tal como consta en la definición aportada un poco más arriba), lo cual posibilita que puedan compartir similaridades en su estructura detallada y en los principios de diseño y control. La estructura jerárquica de los sistemas implica la existencia de distintos niveles dentro de los mismos, así como la existencia de interacciones no sólo entre los elementos que configu-ran cada nivel, sino también entre los distintos niveles, generándose las denominadas causalidades ascendente y descendente comentadas en el artículo de Torres. Estas relaciones causales entre distintos niveles dentro de un sistema jerárquico fueron clarificadas inicialmente por Polyani (1968).
Pero los sistemas biológicos no son meras estructura reticulares estáticas. La Termodinámica del No Equilibrio desarrollada por el premio Nobel Ilya Prigogine permite lo más parecido a una contestación "objetiva" (desde luego, al menos utilitaria) a la recurrente pregunta "¿Qué es la vida?". Sea cual sea la química en que cualquier forma de vida se base, todo ser vivo (aquí y en cualquier otro lugar de nuestro universo) es un sistema termodiná-micamente abierto, en continuo intercambio de materia, energía e información con su entorno. Esos continuos intercambios suponen un continuo cam-bio, esto es, los sistemas biológicos son sistemas dinámicos. Y, además, esos cambios suponen respuestas concretas a los cambios del entorno, esto es, los sistemas biológicos tienen la capacidad de responder a estímulos externos e internos, y por tanto tienen la capacidad de adaptarse, de "aprender". Las teorías de sistemas dinámicos y las más recientes teorías de la complejidad aportan, pues, un marco de referencia para el estudio de siste-mas complejos adaptativos tales como los seres vivos.
Llegados a este punto, puede asumirse que el estudio integral de un sistema biológico como "un todo" supone: 1. El conocimiento de la propia es-tructura del sistema. 2. La comprensión de la dinámica del sistema. 3. La identificación de los principios de diseño que justifiquen la estructura y la dinámica del sistema. 4. La identificación de las reglas de control y regulación que rigen el comportamiento del sistema. Esto no es nada diferente a lo que uno de los reconocidos pioneros de la moderna biología de sistemas, Hiroaki Kitano, establecía en su influyente artículo "Systems Biology: a brief over-view" (Science 295: 1662-4, 2002) como los cuatro pilares fundamentales a partir de los cuales puede derivarse un comprensión sistémica de los sistemas biológicos.
Los signos de los tiemposEl previo recorrido histórico nos ha traído de vuelta al momento presente, a estos primeros años del siglo XXI que han contemplado la emergencia y
primer acelerado desarrollo de la moderna Biología de Sistemas. ¿Qué ha hecho que tal emergencia y desarrollo haya sucedido precisamente en estos tiempos? Los colaboradores del presente número monográfico de Encuentros en la Biología coinciden en apuntar la influencia que tuvieron los grandes proyectos genoma (y, en particular, el Proyecto Genoma Humano), lo cual (al menos simbólicamente) justificaría poner el hito de la publicación de los primeros borradores del genoma humano (febrero de 2001) como fecha de arranque de la Biología de Sistemas. ¿Celebramos, pues, en 2011, a fin de cuentas, una década de la Biología de Sistemas? No vayamos a enredarnos a esta altura: vuelvo a apelar al artículo de Alves et al. y también señalo el apartado anterior de este artículo para dejar definitivamente establecido que hubo científicos que hicieron biología de sistemas antes de que este término se impusiera. Sin embargo, resulta pertinente formular la pregunta: ¿Cuándo empezó a aparecer el término "Biología de Sistemas" en los títulos de la literatura científica? Una sencilla consulta a las bases de datos de bibliografía científica puede arrojar luz.
Según una consulta a Pubmed (www.pubmed.org), la primera aparición del término "systems biology" en un título se produjo en la revista Nature Biote-chnology en 1999, en un comentario/anuncio de la creación del Institut for Systems Biology de Seatle (primer centro de investigación del mundo dedicado a la Biología de Sistemas, que inició su exitosa andadura en 2000). A esta nota le siguen otros 18 trabajos en el periodo comprendido entre 2001 y 2002, de los cuales la mayoría son revisiones y comentarios.
Aunque ni en su título ni en el texto menciona explícitamente el término "systems biology", en mi opinión el artículo "From molecular to modular cell biology" (Hartwell et al., Nature 402: C47-C52, 1999) es un ensayo que tuvo gran influencia en el arranque de la "nueva" Biología de Sistemas (así como en la nueva Biología Sintética, término éste que sí menciona explícitamente). Los autores argumentan que los módulos funcionales deberían ser reconoci-dos como niveles críticos de organización biológica y afirman que principios de diseño generales (profundamente configurados por las restricciones impuestas por la evolución) gobiernan la estructura y función de dichos módulos. Estos términos claves (organización modular y principios de diseño de la organización en red de los módulos y de la regulación de sus funciones) son comunes a la Biología de Sistemas y a la Biología Sintética, que además han compartido una historia paralela de asentamiento y desarrollo en los años que han transcurrido de este siglo XXI. [Esos argumentos son una justifica-ción más que suficiente de la conveniencia de dedicar cierta atención a la Biología Sintética en un monográfico sobre Biología de Sistemas, función que en el presente número de Encuentros en la Biología cumple perfectamente el artículo de Durán].
Identificado como un "nuevo" enfoque para el estudio de los sistemas biológicos, en estos años no ha parado de crecer el número de trabajos científi-cos publicados que contienen el término "systems biology" en su título: alcanza casi los casi 1500 hoy día. Esto implica que una importante nómina de científicos ha tenido que incorporarse a la práctica de la Biología de Sistemas. ¿De qué campos procedían los que que eran investigadores en activos desde antes? Ciertamente, una parte de ellos eran científicos convencidos del interés y la necesidad de los enfoques sistémicos y que hacían biología de sistemas antes de la Biología de Sistemas. Pero había también científicos procedentes de los campos de la Física y las Ingenierías interesados en nuevos retos, como los que impone la complejidad de los sistemas biológicos. Y también se incorporaron muchos biólogos moleculares y practicantes de las tecnologías del DNA recombinante y de los enfoques "ómicos". ¡No nos engañemos! Los científicos procedentes del "núcleo duro" del reduccionismo son aplastante mayoría entre los actuales practicantes de la Biología de Sistemas.
La inflación de los datos y la "materia oscura" del conocimientoLos artículos de Alves et al. y de Cánovas coinciden en señalar el desarrollo tecnológico asociado a los proyectos genoma como motor esencial para el
arranque de la moderna Biología de Sistemas. Si bien es cierto que el Proyecto Genoma Humano sólo se pudo abordar cuando el desarrollo tecnológico lo permitió, no es menos cierto que -a su vez- promovió nuevos desarrollos tecnológicos. Por una parte, con la implantación de las tecnologías "ómicas", que permiten analizar múltiples datos (secuencias, genes, expresiones de mensajeros o proteínas, niveles de metabolitos, etc.) al mismo tiempo. Por otra parte, con el desarrollo de herramientas computacionales para el manejo y gestión de esos datos masivos, para la adecuada visualización de sus interac-ciones y para facilitar la construcción de modelos matemáticos que describan la estructura del sistema y sus respuestas a las perturbaciones. De esta forma queda completado el marco referencial que justifica la emergencia de la Biología de Sistemas en los comienzos del siglo XXI. En efecto, así queda recogido en una de las primeras definiciones escritas de "biología de sistemas", que aparece en una revisión publicada en 2001 y escrita por científicos del entonces recién constituido Institut for Systems Biology (Ideker et al.: A new approach to decoding life: Systems Biology. Annu Rev Genom Human Genet 2: 343-372, 2001). Los autores dicen que "la Biología de Sistemas estudia sistemas biológicos perturbándolos (biológica, genética o químicamen-te) de forma sistemática, monitorizando las respuestas génicas, proteicas y en las rutas de información, integrando estos datos y, finalmente, formulando modelos matemáticos que describan la estructura del sistema y su respuesta a las perturbaciones indivi-duales". Esta forma de definir la Biología de Sistemas justifica y, al mismo tiempo, viene determinada por las tres procedencias mencionadas más arri-ba desde las que se incorporaron científicos a este campo. Desde el principio, el modelado aparece como componente esencial de este enfoque sistémi-co de la Biología, tal como todos los contribuyentes a este monográfico expresan. Del mismo modo, se reconoce la necesidad de herramientas computa-

Vol.4 | Nº 136
70
cionales tales como las que se vienen aportando desde la Bioinformática. Y todo ello en continuo diálogo con la Biología experimental en una forma de hacer ciencia conducida por la formulación de hipótesis a partir de los modelos y el contraste experimental de dichas hipótesis.
Pero no es sólo esto. Mi impresión es que el triunfante reduccionismo de la Biología Molecular hizo que ésta tuviera que afrontar el peligro de poder "morir de éxito". Toda la nueva y sofisticada tecnología de adquisición masiva de datos biológicos ha conducido a incrementar de forma acelerada la brecha existente entre la cantidad de datos disponibles y de información biológica procesada y más aún la brecha entre ésta y la cantidad real de nuevo conocimiento (Figura 4, en pagina 58). En el cambio del milenio, los biólogos se enfrentaban por primera vez al reto de qué hacer con los datos que estaban adquiriendo masivamente a una velocidad mucho mayor que la que requerían para su procesamiento. A pesar de los enormes progresos conse-guidos en estos diez últimos años, este problema dista de ser resuelto; es más, esas brechas no han dejado de incrementarse. Nos enfrentamos a lo que Ranea et al. ("Finding the 'dark matter' in human and yeast protein network prediction and modelling". Plos Comput Biol 6: e1000945, 2010) han descrito como "materia oscura" del conocimiento. Sólo una pequeña parte de los datos biológicos acumulados se ha integrado en nuevo conocimiento, que vendría a ser la punta del iceberg del conjunto total de nuevo conocimiento potencialmente extraíble de los datos ya existentes. Esta circunstancia justifica la incorpora-ción al cuerpo de la Biología de Sistemas de un enfoque que se centra en el desarrollo de herramientas predictivas capaces de "extraer" nueva informa-ción, de hacer emerger nuevo conocimiento a partir de esa creciente "materia oscura" (ver en este mismo número la contribución de Ranea).
Un mundo de redes mundo pequeñoSi los sistemas están integrados por componentes que interactúan no linealmente estructurándose en redes, parece natural que a su estudio se le
pueda aplicar el enfoque y los métodos de la moderna ciencia de las redes. Aunque también con una historia previa muy rica y con notables anteceden-tes en el mundo de las ciencias sociales, la ciencia de las redes entró con fuerza en el campo de la Biología a partir de los últimos años del pasado siglo. Su enfoque necesariamente sistémico permite considerar los estudios de las redes biológicas que aplican la teoría de redes como otra parte integrante de la Biología de Sistemas, aunque sus orígenes y primeros desarrollos fueran independientes. Aunque las redes biológicas exigirían y merecen el espacio de un monográfico centrado exclusivamente en ellas, no hemos querido renunciar a mencionarlas en el presente número de Encuentros en la Biología. La contribución de Sorribas et al. pone el foco en este asunto y la imagen comentada de este monográfico es una red biológica.
¿Un nuevo tipo de ciencia o una fusión de ciencias?Con todo lo dicho, no es de extrañar que la Biología de Sistemas que se ha venido desarrollando durante esta década sea -al menos, mayoritaria-
mente- una biología molecular de sistemas, tal como evidencia la exitosa publicación (con muy alto factor índice de impacto) Molecular Systems Biology del grupo editorial Nature. Incluso un destacado representante de los actuales "biólogos de sistemas" procedentes del campo del modelado y la biología teóri-ca (el Dr. Hans Westerhoff) aboga en una reciente revisión por una biología de sistemas cuyo objetivo sea estudiar "las bases biológico-moleculares de la función biológica" (Westherhoff et al.: Systems Biology: the elements and principles of life. FEBS Lett 583: 3882-3890, 2009). En esta interesante revisión, se argumenta que los dos grandes paradigmas pertinentes a las ciencias físicas representados por la navaja de Occam (principio filosófico introducido por el fraile franciscano Guillermo de Occam en el siglo XIV, según el cual las soluciones más sencillas a un problema son probablemente las más ciertas) y la preva-lencia de soluciones de mínimos energéticos o máximas eficiencias no son aplicables a la Biología de Sistemas. En su sustitución, defienden que es posi-ble enunciar unos "principios de la Biología de Sistemas", por lo que reivindican para ella el estatus de un nuevo tipo de ciencia con sus propios (y nue-vos) paradigmas. Finalmente, reconocen que a la Biología de Sistemas le hace todavía falta una clara formulación de su metodología.
Frente a esta reivindicación -aunque no necesariamente en completa oposición a ella- se sitúa el enfoque aportado por el artículo de Torres cuando señala que "los objetivos de la Biología de Sistemas son al mismo tiempo un programa de fusión de las disciplinas". Esto constituye una reivindicación de su carácter transdisciplinar por la que siento especial afinidad como componente del "oficioso" Grupo Transdisciplinar de la Universidad de Málaga. Cierta-mente, coincido con los planteamientos de quienes vienen reivindicando una necesaria reunificación del conocimiento. En el ámbito de la Biología, dos importantes contribuciones en esta línea de pensamiento se publicaron en el cambio de milenio: el libro Consilience de Edward Wilson (véase "Premios Fronteras del Conocimiento 2010" en Encuentros en la Biología 134) y un editorial en el número de diciembre de 1999 de BioEssays, que señalaba la reunifi-cación del conocimiento como uno de los tres grandes retos de la Biología del siglo XXI, junto con el reto de la comunicación y el reto de la compleji-dad. Una reivindicación de y una aspiración a la transdisciplinariedad son pertinentes en los tiempos que corren. El problema es que la inmensa mayo-ría de los científicos en activo o entienden mal qué representa la transdisciplanariedad o, simplemente, ignoran lo que el término significa. Esto queda patente en la diversidad de definiciones en las que -como en la confusa e imprecisa definición aportada en Wikipedia (Figura 1) o como en varias de las formas de representar visualmente el enfoque de la biología de sistemas (Figura 2)- se hace referencia al carácter pluri-, multi- o interdisciplinar de la Biología de Sistemas, haciéndome sospechar que ni siquiera se sabe diferenciar entre pluri-, multi- e interdisciplinariedad. Un segundo problema igual-mente importante es que falta por formular una clara metodología para la praxis transdisciplinar.
Biología de Sistemas... ¿qué biología de sistemas? A la larga, si la vida (y la Biología) es por naturaleza sistémica, puede resultar pertinente reformu-lar la pregunta: ¿por qué una biología de sistemas? En mi opinión, la justificación última que hace recomendable mantener el uso del término es que puede representar de forma natural el marco de trabajo óptimo para el desarrollo de aproximaciones auténticamente sistémicas al estudio de la complejidad biológica.
TendenciasUn conocido e influyente científico, actualmente identificado como uno de los "pesos pesados" de la Biología de Sistemas española (y años atrás
reconocido como uno de los más destacados biólogos moleculares españoles) hace unos años pronunció una conferencia en la que (no sin ciertas dosis de cinismo) reconocía su adscripción meramente "instrumental" a la Biología de Sistemas como un término que había estado de moda y del que podía sacarse réditos. Es más, en dicha conferencia se permitió emitir por adelantado el "certificado de defunción" de la Biología de Sistemas (entendida como "tendencia en boga") porque el término empezaba ya a quedar "fuera de moda". Para quienes tienen la habilidad de mantenerse "en la cresta de la ola" sabiendo saltar anticipadamente de una ola a la siguiente, este planteamiento es plenamente lógico y admisible. Sin embargo, la tozuda realidad es que años después de este "réquiem" adelantado la Biología de Sistemas sigue exhibiendo síntomas de gozar de una estupenda salud y de estar todavía en pleno proceso de crecimiento. En la revisión de 2010 "A Decade of Systems Biology" (mencionada al comienzo de este artículo), los autores identifican cua-tro aplicaciones emergentes que marcarán las tendencias del más inmediato futuro: 1. El reconocimiento de biomarcadores basados en estudios de rutas biológicas. 2. La generación de mapas de interacción genéticos globales. 3. Enfoques sistémicos para la identificación de genes asociados a enfermeda-des. Y 4. La biología de sistemas de células madres y pluripotenciales.
En relación con la tercera de las cuatro tendencias arriba apuntadas, conviene mencionar que Leroy Hood, presidente del Institut for Systems Biology, aboga en estos últimos años por la implantación de lo que denomina una Medicina P4, esto es, una medicina que sea realmente predictiva, preventiva, personalizada y participativa. El propio Hood ha apoyado la implantación en Seatle del primer P4 Medicine Institut con la participación de su instituto como socio (junto con Ohio State University). Por otra parte, en el número de Noviembre/Diciembre de 2011 de WIREs Systems Biology and Medicine el espe-cialista en procesos de bioseñalización Joseph Loscalzo y uno de los más activos y destacados representantes de la moderna ciencia de redes (Al-bert-Laszlo Barabasi) han publicado un artículo de opinión con el título "Systems biology and the future of medicine" que está destinado a marcar tendencias. En este artículo, Loscalzo y Barabasi reivindican que todas las enfermedades (incluso las asociadas a mutaciones en un único gen y describibles por genética mendeliana simple) son complejas, de forma que los patofenotipos reflejan la acción de una red molecular defectiva determinista dentro de un contexto ambiental esencialmente estocástico que modula la función de dicha red. Definidas de esta forma, las enfermedades encuentran acomodo en su estudio desde la perspectiva de la Biología de Sistema, ya que podrían ser interpretadas como el resultado de las interacciones efectivas entre nodos "ómicos" y nodos ambientales en una red modular compleja, que es lo que denominan "diseae-ome". Esto autores apuntan que hay tres representa-ciones diferentes posibles del disease-ome: 1. El formalismo de genes compartidos, que reconoce que las enfermedades que comparten uno o más genes tienen probablemente una base genética común. 2. El formalismo de las rutas metabólicas compartidas, que reconoce que los defectos enzimáticos que pueden afectar a una reacción en una ruta metabólica podrían también afectar a los flujos downstream (o sea, los flujos de materia en las reacciones metabólicas poste-riores a la directamente afectada en dicha ruta), conduciendo así a patofenotipos que se sabe están asociados con las reacciones posteriores a la afectada en la ruta. Y 3. El formalismo de comorbilidad de la enfermedad, que asocia enfermedades de acuerdo a su co-ocurrencia (si ésta no es explicable por las meras reglas del azar), lo cual lleva a la construcción de mapas de redes fenotípicas de enfermedades. Loscalzo y Barabasi concluyen su artículo identificando tres áreas de la biomedicina en la que los enfoque sistémicos probablemente demuestren su utilidad en el inmediato futuro: el desarrollo de fármacos, el estudio de la influencia de las conductas en la propensión a las enfermedades y los estudios metagenómicos de interacciones entre el genoma humano y su complejo microbioma comensal y/o simbionte. Este último punto pone de actualidad una de las continuas reivindicaciones de Lynn Margulis, una de las grandes biólogas de todos los tiempos, fallecida el 22 de noviembre de 2011, justo cuando el presente artículo se encontraba en fase de redacción. Sirvan estas palabras finales como un humilde pero sincero homenaje a una científica valiente, polémica y polemista que toda su vida como científica activa hizo biología sistémica antes de los tiempos de la 'nueva' Biología de Sistemas.





























