Embed Size (px)
Citation preview

UNIVERSIDAD DE CHILE Facultad de Ciencias Químicas y Farmacéuticas
Modificaciones postraduccionales e interacción con Calreticulina de la proteína viral Tax en
linfocitos infectados con HTLV-I y su relación con la evolución de la Paraparesia Espástica
Tropical
Tesis para optar al grado académico de Magíster en Bioquímica, área de especialización en Bioquímica de Proteínas y Biotecnología,
y Memoria para optar al Título Profesional de Bioquímico
Por Fernando Edmundo Medina Ferrer
Directores de tesis:
Dra María Antonieta Valenzuela P. Dr Javier Puente P.
Santiago, Chile
2010

UNIVERSIDAD DE CHILE
FACULTAD DE CIENCIAS QUÍMICAS Y FARMACÉUTICAS
INFORME DE APROBACIÓN
TESIS DE MAGÍSTER Y TÍTULO DE BIOQUÍMICO
Se informa a la dirección de Postgrado de la Facultad de Ciencias Químicas y
Farmacéuticas que la Tesis de Magíster presentada por el candidato:
FERNANDO EDMUNDO MEDINA FERRER
Ha sido aprobada por la Comisión Informante de Tesis como requisito para optar al
Grado de Magíster en Bioquímica, área de especialización Bioquímica de Proteínas y
Biotecnología, y al Título de Bioquímico, en el examen de Defensa de Tesis rendido el
día ____/______________/________.
Directores de Tesis:
Dra. María Antonieta Valenzuela P. _______________________
Dr. Javier Puente P. _______________________
Comisión Informante de Tesis:
Dra. María Mercedes Zaldívar (Presidenta). _______________________
Dr. Arturo Ferreira V. _______________________
Dra. Ana María Jabalquinto. _______________________

Con el presente trabajo se originaron las siguientes presentaciones a congresos y publicaciones:
Medina F, Alberti C, Pando ME, Barriga A, Maldonado H, Kettlun AM, Collados L, Puente J, Cartier L, Ramirez E and Valenzuela MA: “Análisis de modificaciones postraduccionales de la proteína Viral Tax en células infectadas con HTLV-I”. Trabajo presentado en las X Jornadas Investigación en Ciencia y Tecnología, Facultad de Ciencias Químicas y Farmacéuticas, Universidad de Chile. Santiago, Chile. Abril 2009. Premiado como el mejor trabajo científico presentado del Departamento de Bioquímica y Biología Molecular.
Medina F, Alberti C, Barriga A, Ramírez E, Cartier L, Puente J, Valenzuela MA: “Ubiquitinación y Sumoilación de la proteína viral Tax en linfocitos infectados con HTLV-I”. Trabajo presentado en la XXXII Reunión Anual Sociedad de Bioquímica y Biología Molecular de Chile. Termas de Chillán, Chile. Septiembre 2009.
Medina F, Alberti C, Cartier L, Ramírez E, Puente J, Barriga A, Kettlun AM, Tanaka Y, Valenzuela MA. Tax protein from MT2 cells and human PBMC of HTLV-I infected patients: Postraslational modifications and calreticulin interactions (manuscrito en preparación).

i
“Alpinista es quién conduce su cuerpo allá dónde un día sus ojos lo soñaron”
Gaston Rébuffat

ii
AGRADECIMIENTOS
Mi familia está primero que todo. Fue el trabajo previo necesario para completar
esta tesis y en especial, esta etapa de mi vida. Agradezco a mis padres, mi hermana, y
mi tía Carmen. Sus decisiones sobre mí me tienen escribiendo ahora. Gracias por su
apoyo incondicional.
Quiero agradecer a quienes participaron en mi formación básica, en particular los
gratos recuerdos que me llevé desde “el primer foco de luz de la nación”, la huella que
dejaron en mí varios profesores, junto con el peso de la historia y en especial de mis
amigos Institutanos que aún están presentes en mi vida.
Agradezco sin duda la formación de la Universidad de Chile, la carrera de
Bioquímica y la gente que la compone, en especial a mis profesores del Laboratorio de
Bioquímica; la Dra María Antonieta, Dra Ana María, mi profesor Andrés, y la profe
Lucía. Muy gratos recuerdos me llevo de aquellos académicos que contribuyeron
enormemente con mi formación profesional, académica y personal. También quiero
agradecer a la comisión revisora de este trabajo por aportar con sugerencias y mejoras
para la tesis.
Agradezco a mis amigos de la Carrera, en particular al “Núcleo de Estudios
Científicos” Se’P; Iván, Ernesto, Juan, Yerko, Pato, Javier y César. Por la ciencia,
paciencia, amistad, deporte, recreación, música y compañía para hacer más grata mi
experiencia en la U y de aquí en más, para el resto de la vida. Agradezco también a mis
amigos del laboratorio; Mae, Javy, Caro, Ares, Seba. Y por supuesto a la Paty y a la
Yeka, por hacer más grato el tiempo en el lab.
Quiero agradecer el apoyo de todos quienes estuvieron presentes durante esta etapa,
inclusive a mis amigos de la Rama de Montaña de la U, quienes me enseñaron nuevas y

iii
variadas formas de observar el mundo, lo cual influye fuertemente en mi vida como
científico.
Por último, quiero agradecer enormemente a Ivonne por su paciencia y apoyo
incondicional durante la Carrera y desarrollo de mi tesis. Su perseverancia como
ejemplo me permite hoy enfrentar sin titubeos mis nuevos desafíos personales y
profesionales. A ella, mi familia y mi sobrinita que está en camino dedico esta memoria.

iv
FINANCIAMIENTO.
Esta tesis se realizó en el Laboratorio de Bioquímica General Dr Osvaldo Cori del
Departamento de Bioquímica y Biología Molecular de la Facultad de Ciencias Químicas
y Farmacéuticas de la Universidad de Chile, bajo la dirección de la Dra María Antonieta
Valenzuela Pedevila y el Dr Javier Puente Piccardo.
El trabajo de investigación fue financiado por el proyecto FONDECYT 1080396,
titulado “PAPEL DE LA PROTEINA VIRAL TAX EN LA PROGRESION DE LA
PARAPARESIA LIGADA A HTLV-I Y SU ACCION EXTRACELULAR EN UN
MODELO DE CULTIVO NEURONAL”.

v
TABLA DE CONTENIDOS
Pág.INDICE GENERAL................................................................................................. v INDICE DE FIGURAS............................................................................................ vii INDICE DE TABLAS.............................................................................................. ix ABREVIATURAS.................................................................................................... x RESUMEN................................................................................................................ xi ABSTRACT.............................................................................................................. xiii 1. INTRODUCCION................................................................................................ 1
1.1. Virus Linfotrópico Humano Tipo I o HTLV-I............................................... 11.2. Mielopatía Asociada al HTLV-I/Paraparesia Espástica Tropical o HAM/TSP……………………………………………………………………….. 51.3. Proteína Viral Tax.......................................................................................... 71.4. Modificaciones postraduccionales de Tax..................................................... 91.5. Contribución de la Calreticulina sobre el intercambio núcleo-citoplasmático de Tax............................................................................................ 121.6. Planteamiento del problema........................................................................... 13
2. HIPOTESIS........................................................................................................... 16 3. OBJETIVOS......................................................................................................... 17
3.1. Objetivo general............................................................................................. 173.2. Objetivos Específicos..................................................................................... 17
3.2.1. Analizar las modificaciones postraduccionales de Tax proveniente de la línea linfocitaria MT-2........................................................................... 173.2.2. Cuantificar los niveles de Calreticulina y establecer su interacción con Tax en células MT-2................................................................................ 173.2.3. Analizar las modificaciones postraduccionales de Tax proveniente de PBMC aislados desde sangre periférica de pacientes HAM/TSP.............. 183.2.4. Cuantificar los niveles de Calreticulina y establecer su interacción con Tax en PBMC aislados desde sangre periférica de pacientes HAM/TSP. 18
4. MATERIALES Y METODOS............................................................................. 19
4.1. Reactivos........................................................................................................ 19

vi
4.2. Material biológico.......................................................................................... 214.3. Metodología................................................................................................... 22
4.3.1. Cultivos celulares................................................................................. 224.3.2. Aislamiento de PBMC desde Sangre Periférica................................... 224.3.3. Lisados Celulares.................................................................................. 23
4.3.3.1. Lisado celular total................................................................... 234.3.3.2. Lisado y fraccionamiento celular............................................. 23
4.3.4. SDS-PAGE y Western Blot.................................................................. 244.3.5. Inmunoprecipitación............................................................................. 254.3.6. Inmunofluorescencia Indirecta y Análisis de Imágenes....................... 264.3.7. Citometría de Flujo............................................................................... 274.3.8. Aislamiento de la fracción de Linfocitos T CD4+............................... 274.3.9. Espectrometría de Masas...................................................................... 28
4.3.9.1. Digestión de Proteínas en Gel.................................................. 284.3.9.2. Análisis Peptídico mediante MALDI-TOF.............................. 294.3.9.3. Análisis Peptídico mediante HPLC-ESI-IT............................. 30
4.3.10. Análisis Estadístico............................................................................. 32 5. RESULTADOS..................................................................................................... 33
5.1. Analizar las modificaciones postraduccionales de Tax proveniente de la línea linfocitaria MT-2.......................................................................................... 335.2. Cuantificar los niveles de Calreticulina y establecer su interacción con Tax en células MT-2..................................................................................................... 415.3. Analizar las modificaciones postraduccionales de Tax proveniente de PBMC aislados desde sangre periférica de pacientes HAM/TSP......................... 455.4. Cuantificar los niveles de Calreticulina y establecer su interacción con Tax en PBMC aislados desde sangre periférica de pacientes HAM/TSP.................... 50
6. DISCUSION......................................................................................................... 54 7. CONCLUSIONES................................................................................................ 63 8. REFERENCIAS.................................................................................................... 65

vii
INDICE DE FIGURAS
Pág. Figura 1: Representación esquemática del genoma del HTLV-I............................. 3 Figura 2: Representación esquemática de la regulación por retroalimentación de la expresión genética del HTLV-I mediada por Tax y Rex..................................... 4 Figura 3: Mecanismos de replicación del HTLV-I.................................................. 5 Figura 4: Dominios funcionales y señales de localización intracelular de Tax....... 10 Figura 5: Modelo de la distribución subcelular de Tax, las señales que la rigen y sus efectos en linfocitos infectados por HTLV-I..................................................... 14 Figura 6: Identificación de la proteína viral Tax en células MT-2.......................... 34 Figura 7: Determinación de las modificaciones postraduccionales de Tax en células MT-2............................................................................................................ 35 Figura 8: Identificación de la N-glicosilación de la banda a 71 kDa en células MT-2........................................................................................................................ 37 Figura 9: Inmunoprecipitación y espectro de masas de la proteína Tax de células MT-2........................................................................................................................ 39 Figura 10: Identificación de Tax y sus modificaciones postraduccionales en los medios de cultivos de células MT-2........................................................................ 41 Figura 11: Cuantificación de los niveles de CRT en células MT-2 y células K562......................................................................................................................... 42 Figura 12: Determinación de colocalización entre Tax y CRT en células MT-2.... 43 Figura 13: Determinación de la expresión de Tax en células MT-2 por citometría de flujo..................................................................................................................... 44 Figura 14: Determinación de la interacción entre Tax y CRT en células MT-2 mediante ensayos de co-inmunoprecipitación.......................................................... 44 Figura 15: Determinación de la expresión de Tax en células de pacientes HAM/TSP mediante citometría de flujo.................................................................. 46

viii
Figura 16: Identificación y cuantificación de Tax y sus modificaciones en medios de cultivo de PBMC de pacientes infectados con HTLV-I...................................... 49 Figura 17: Inmunoprecipitación de la proteína Tax de medio de cultivo de PBMC de un paciente HAM/TSP............................................................................ 50 Figura 18: Identificación de CRT en medios de cultivo de PBMC aislados de pacientes con HAM/TSP.......................................................................................... 51 Figura 19: Correlación entre las cantidades de Tax extracelular y la forma de CRT extracelular de 56 kDa en pacientes HAM/TSP.............................................. 52 Figura 20: Determinación de la interacción entre Tax y CRT en Linfocitos T CD4+ mediante microscopía confocal..................................................................... 53 Figura 21: Modelo propuesto de co-secreción y efectos extracelulares de Tax y CRT.......................................................................................................................... 62

ix
INDICE DE TABLAS
Pág.Tabla I: Dilución de anticuerpos empleados............................................................. 25 Tabla II: Datos clínicos y resultados de los pacientes analizados mediante citometría de flujo (% linfocitos CD4+/Tax+) y western blot del medio de cultivo linfocitario (Unidades arbitrarias, UA) para la proteína Tax.................................... 47

x
ABREVIATURAS
ATL: Leucemia de Células T del Adulto BCA: Acido bicinconínico CHCA: Acido α-ciano-4-hidroxicinámico CID: Disociación inducida por colisión CRT: Calreticulina DNA: Acido desoxirribonucleico DMSO: Dimetilsulfóxido DTT: Ditiotreitol ESI-IT: Ionización por electrospray-Trampa iónica FITC: Isotiocianato de fluoresceína GAPDH: Gliceraldehído-3-fostato deshidrogenasa h: Horas HPLC: Cromatografía líquida de alta resolución HTLV-I: Virus Linfotrópico Humano tipo I HAM/TSP: Mielopatía Asociada al HTLV-I/Paraparesia Espástica Tropical IgG: Inmunoglobulina G IP: Inmunoprecipitación kDa: Kilodalton LCR: Líquido cefalorraquídeo LTR: Secuencia terminal repetida larga MALDI-TOF: Desorción/ionización láser asistida por matriz-Tiempo de vuelo min: Minutos mRNA: RNA mensajero NES: Señal de exportación nuclear NLS: Señal de localización nuclear PBMC: Células Mononucleares de Sangre Periférica PBS: Tampón salino de fosfato PMSF: Fenilmetilsulfonil fluoruro RE: Retículo endoplásmico RNA: Acido ribonucleico SDS: Dodecil sulfato de sodio SDS-PAGE: Electroforesis en geles de poliacrilamida en presencia de SDS SEM: Error estándar de la media SNC: Sistema nervioso central SUMO: Modificador pequeño tipo Ubiquitina TBS-T: Tampón salino de Tris con Tween-20 TEMED: Tetrametiletiléndiamina TFIIB: Factor de transcripción IIB TRE: Elementos de respuesta a Tax WB: Western Blot

xi
RESUMEN
“Modificaciones postraduccionales e interacción con Calreticulina de la proteína
viral Tax en linfocitos infectados con HTLV-I y su relación con la evolución de la
Paraparesia Espástica Tropical”
El virus linfotrópico humano tipo I o HTLV-I es un retrovirus humano que infecta
principalmente Linfocitos T CD4+ y es capaz de provocar dos patologías; la Leucemia
de Células T en el Adulto (ATL), y la Mielopatía Asociada al HTLV-I/Paraparesia
Espástica Tropical (HAM/TSP), una enfermedad neurodegenerativa progresiva que se
caracteriza por ser una axonopatía de carácter central originada por un trastorno en el
transporte axoplásmico. A pesar de conocerse bastante sobre las características del
HTLV-I, aún no se conocen los mecanismos moleculares involucrados en la patogénesis
de HAM/TSP y en la evolución de la infección durante la enfermedad.
En HAM/TSP, el daño está asociado a la presencia del HTLV-I en linfocitos
infiltrantes en el sistema nervioso central, en donde la proteína viral Tax, principal
transactivador de genes virales y celulares, jugaría un papel relevante en el trastorno
funcional de estas células, y posiblemente sobre las neuronas afectadas en la patología.
Se postula que Tax secretada desde linfocitos infectados altera el transporte axonal en
las neuronas motoras y con ello podría dar cuenta de la patogénesis de HAM/TSP.
Se ha descrito recientemente una modulación de la actividad transactivadora de Tax
mediada por modificaciones postraduccionales en ella, las cuales incluyen
Ubiquitinación y Sumoilación. Principalmente estas modificaciones regulan la
localización subcelular de Tax. Se postula hasta el momento que su ubiquitinación es
señal de localización citoplasmática y su sumoilación, de localización nuclear. Esto
último podría dar cuenta de la progresión en la ATL, pero aún no se han relacionado
con la evolución de la infección viral en HAM/TSP. También se ha descrito

xii
recientemente la interacción de Tax con Calreticulina, lo que podría aumentar la
exportación nuclear de la primera y su secreción al medio extracelular, pero hasta el
momento no se ha relacionado directamente esta interacción con la secreción de Tax, lo
que traería consecuencias directas sobre la etiología y progresión de HAM/TSP.
En este trabajo se identifican las modificaciones postraduccionales en la proteína
viral Tax mediante western blot a partir de la línea linfocitaria MT-2, la cual contiene el
genoma del HTLV-I en la forma de provirus, y de Células Mononucleares de Sangre
Periférica (PBMC) de individuos con HAM/TSP. Estos estudios tienen como objeto
identificar las modificaciones postraduccionales de Tax en linfocitos infectados para
poder relacionarlas con el nivel funcional de pacientes HAM/TSP. En células MT-2 se
encontró una forma de 71 kDa de Tax, de peso molecular mayor al teórico de 40 kDa, la
cual no se encuentra modificada por las proteínas pequeñas Ubiquitina y SUMO, pero si
se encuentra N-glicosilada y probablemente corresponde a una proteína de fusión viral
gp21-Tax. Al analizar Tax desde PBMC de pacientes infectados, sólo se pudo
identificar la presencia de una banda de 58 kDa correspondiente a Tax presente en los
medios de cultivos de las células. Esta proteína secretada se encuentra Ubiquitinada,
pero no se encontraron diferencias significativas en los niveles de Tax modificada entre
los distintos grupos de pacientes con diversa evolución de HAM/TSP estudiados.
Se cuantificaron también los niveles de Calreticulina en células MT-2 y PBMC de
individuos infectados, y se estableció la interacción entre Tax y Calreticulina en células
MT-2. En los medios de cultivo de PBMC se encontró un aumento significativo de los
niveles de una especie de Calreticulina a 56 kDa de los individuos infectados con
HTLV-I. Se estableció una relación directa entre las cantidades de Tax y Calreticulina
de 56 kDa secretadas al medio de cultivo. Esto último puede tener especial relevancia
en los mecanismos de secreción de ambas proteínas y en los efectos extracelulares de
las mismas.

xiii
ABSTRACT
“Tax postranslational modifications and Calreticulin interaction in HTLV-I
infected lymphocytes and its relationship with Tropical Spastic Paraparesis
evolution”
HTLV-I (Human T-cell Lymphotropic Virus Type-I) is a human retrovirus whose
target cells are CD4+ T lymphocytes. It is the etiologic agent of two human pathologies;
the Adult T-cell Leukemia (ATL) and HTLV-I Associated Myelopathy/Tropical Spastic
Paraparesis (HAM/TSP), a progressive neurological disease characterized as a central
axonopathy due to an axoplasmic transport disorder. In spite of the increased knowledge
about HTLV-I, the molecular mechanisms in HAM/TSP and the evolution of infection
during the disease are still unknown.
In HAM/TSP, the injury is associated with the presence of infected lymphocytes in
the central nervous system, where the viral Tax protein, main cellular and viral gene
transactivator, plays a relevant role in the functional disorder of these cells, and could
have an effect on affected neurons in the disease. It has been postulated that secreted
forms of Tax released from infected lymphocytes disturb the axonal transport in motor
neurons and may originate HAM/TSP.
Recently, it has been found a modulation of Tax transcriptional transactivation
through postraslational modifications; Ubiquitination and Sumoylation. These
modifications regulate Tax subcellular distribution. It is proposed that ubiquitination is a
cytoplasmic retention signal and sumoylation a nuclear retention signal. These
modifications and their effects on Tax activities explain ATL progression, but they have
not yet been related with HAM/TSP viral infection. Also, Tax and Calreticulin
interaction have been found, which could promote Tax nuclear export and Tax secretion
into the extracellular environment, but these interactions have not been related directly

xiv
with Tax secretion yet. This relationship would have direct consequences on HAM/TSP
evolution and progression.
In this work, Tax postranslational modifications are identified through western blot
analysis of MT-2 lymphocyte cell line (which contains HTLV-I genome like a provirus)
and Peripheral Blood Mononuclear Cells (PBMC) of HAM/TSP patients. The central
objective of this research was to determine Tax postranslational modifications in
infected lymphocytes and their relation with the functional level of HAM/TSP patients.
In MT-2 cell we found an intracelluar 71 kDa Tax form, rather than the theorical 40
kDa molecular weight. The 71 kDa Tax form was neither ubiquitinated nor sumoylated,
but was N-glycosylated and probably corresponds to a gp21-Tax fusion viral protein.
Tax postranslational modification analysis from PBMC of infected patients were done
with the cellular culture media, where Tax was a 58 kDa protein. This secreted protein
was ubiquitinated, and no significant difference in the secreted modified Tax was found
between the groups of patients with different HAM/TSP evolution.
The levels of Calreticulin were quantified in MT-2 cells and infected PBMC, and an
interaction between Tax and Calreticulin in MT-2 cells was established. In PBMC cell
culture media, significant increase in 56 kDa Calreticulin levels of HTLV-I infected
patients was found. A direct relationship between Tax and the 56 kDa secreted
Calreticulin levels in cell culture media was found. This could give an insight of the
secretion mechanisms of both proteins and its extracellular effects.

1
1. INTRODUCCION
1.1. Virus Linfotrópico Humano Tipo I o HTLV-I
HTLV-I (del inglés “Human T-cell Lymphotropic Virus Type-I”) o virus
linfotrópico humano tipo I fue el primer retrovirus humano descubierto, comenzado
ya los años ochenta (Poiesz y cols., 1980; Yoshida y cols., 1982; Gallo, 2005). Desde
entonces, el estudio de los retrovirus y su impacto sobre la salud humana ha sido un
área de intensa investigación. Perteneciente a los retrovirus que poseen tropismo por
los linfocitos T, se ha demostrado que es el agente etiológico de al menos dos
patologías: la Leucemia de Células T del Adulto o ATL (del inglés “Adult T-cell
Leukemia”), una enfermedad de características neoplásicas muy agresivas, y de una
patología neurodegenerativa denominada Mielopatía Asociada al HTLV-
I/Paraparesia Espástica Tropical o HAM/TSP (del inglés “HTLV-I-Associated
Myelopathy/Tropical Spastic Paraparesis”) (Nagai y Osame, 2003; Matsuoka y cols.,
2003). Aún se desconoce cómo el virus causa estas enfermedades, y cuáles son los
mecanismos involucrados en la progresión de ellas, lo cual actualmente dirige una
intensa búsqueda focalizada en la comprensión de la biología molecular y
patogénesis del HTLV-I.
El blanco principal del HTLV-I son los Linfocitos T CD4+, y en un menor grado
los Linfocitos T CD8+, aunque in vitro se ha demostrado que puede infectar también
linfocitos B, macrófagos y astrocitos (Franchini, 1995; Méndez y cols 1997). Dichas
células infectadas constituyen el vehículo de transmisión viral vía sexual, vertical y
parenteral (Nagai y Osame, 2003; Yoshida, 2005), requiriendo del contacto célula-
célula facilitadas por la interacción con dos glicoproteínas de superficie viral
ubicadas en la membrana plasmática de las células infectadas: la proteína de
envoltura gp46 y la proteína transmembrana gp21. Estas glicoproteínas se requieren
para la infectividad del HTLV-I, puesto que median la formación de una sinapsis

2
virológica entre linfocitos infectados y linfocitos libres del virus (Derse y Heidecker,
2003; Igakura y cols., 2003; Pais-Correia y cols., 2010). El mecanismo de entrada del
virus desde la célula infectada a la célula no infectada aún es incierto, pero se ha
demostrado que el virus se transmite por medio del contacto célula-célula (entre
linfocitos T) seguido de la transferencia del genoma viral a la célula receptora. Por
otra parte, se considera que la contribución de los viriones o partículas virales en el
proceso infectivo es mínima, puesto que su producción por parte de los linfocitos
infectados es bajísima, y de éstos, sólo una millonésima parte llega a ser infectivo
(Igakura y cols., 2003).
El material genético del HTLV-I consta de dos hebras de RNA de cadena simple,
idénticas entre si y de polaridad positiva, las cuales se incorporan al DNA celular
posterior al proceso de transcripción reversa, quedando entonces como provirus en el
núcleo celular. El genoma viral contiene los genes enzimáticos y estructurales típicos
de los retrovirus (gag, pol y env) flanqueados por secuencias terminales repetidas
largas (LTRs) (Boxus y Willems, 2009). La región gag codifica para las proteínas
estructurales de la partícula viral: p19, p24 y p15. La región pol codifica para una
transcriptasa reversa (DNA polimerasa RNA dependiente, p62) y una integrasa (p32).
Entre gag y pol existe una pequeña región que codifica para una proteasa (p14), la
cual proteolisa los productos proteicos inmaduros originados a partir de gag, pol y
env. Esta última, la región env, codifica para las proteínas de envoltura gp46 y gp21
(Figura 1). El genoma viral posee además una región adicional llamada pX,
localizada entre el gen env y el LTR-3´. Esta región contiene al menos cuatro marcos
de lectura superpuestos que codifican dos proteínas accesorias (p12, p13/p30), así
como también dos proteínas reguladoras: Tax y Rex (Yoshida, 2005). Estas últimas
controlan la replicación y expresión de proteínas virales, y en particular, Tax ejerce
un papel esencial tanto en la transcripción viral como en la transformación celular
(Figura 2).

3
Figura 1: Representación esquemática del genoma del HTLV-I. En verde se representa el material genético en la forma de provirus. En azul se representan las proteínas originadas de las regiones genómicas de gag, pol y env. En naranjo se representan las proteínas reguladoras originadas de la región pX: Tax y Rex (Modificado de Boxus y Willems, 2009).
La transcripción del genoma viral es regulada tanto por factores transcripcionales
celulares que unen directamente al DNA en el 5´-LTR, así como por Tax, la cual
aumenta la transcripción uniendo los TREs (“Tax Response Elements”) ubicados en
las secuencias U3 del LTR (Paskalis y cols., 1986; Shimotohno y cols., 1986; Brady
y cols., 1987; Mogensen y Paludan, 2001). Tax además actúa como transactivador de
diversos genes celulares que codifican variadas citoquinas y los receptores para ellas,
así como también aumenta la expresión de factores de transcripción y proto-
oncogenes (Barmak y cols., 2003; Marriott y Semmes, 2005).

4
Figura 2: Representación esquemática de la regulación por retroalimentación de la expresión genética del HTLV-I mediada por Tax y Rex. Luego de un doble “splicing” del transcrito primario del genoma viral se origina el mRNA del cual se traduce Tax, la cual posteriormente activa la transcripción viral (1) y por ende las cantidades del transcrito primario para Tax, el cual posee un inicio de traducción alternativo que origina la proteína Rex. Esta última suprime el “splicing” del mRNA viral (2), de forma tal que se acumulan transcritos primarios y secundarios de los cuales se traducen las proteínas Gag, Pol y Env (3), seguido de una disminución en la expresión de Tax/Rex y por ende de la transcripción. (Modificado de Yoshida, 2005).
Una vez que el virus se integra en el DNA de un linfocito hospedero, éste puede
replicarse en forma activa durante las primeras etapas de la infección e infectar otras
células del hospedero mediante contacto célula-célula. Luego, al desarrollarse la
respuesta inmune específica contra el virus, éste puede permanecer silente en la
forma de provirus dentro de la célula y favorecer ahora su replicación por un
mecanismo distinto, que involucra la expansión clonal de las células infectadas.
Puesto que el DNA del virus se encuentra inserto en el DNA de la célula infectada, el
virus promueve la mitosis y la sobrevida de la célula infectada por medio de la acción
de la proteína regulatoria Tax, para así promover también su propia reproducción y
sobrevida (Matsuoka y Jeang, 2007) (Figura 3).

5
Figura 3: Mecanismos de replicación del HTLV-I. Durante etapas tempranas de la infección, los linfocitos T CD4+ infectados por HTLV-I (+) infectan otros linfocitos mediante contacto célula-célula durante la sinapsis virológica. Posteriormente, la expresión de Tax promueve la expansión clonal de las células infectadas. Después de un prolongado período de 20 a 40 años, el virus puede desencadenar enfermedades como ATL y/o HAM/TSP (en menos de un 5 % de los infectados). (Modificado de Matsuoka y Jeang, 2007).
1.2. Mielopatía Asociada al HTLV-I/Paraparesia Espástica Tropical o
HAM/TSP
Se estima que el HTLV-I infecta aproximadamente entre 10 a 20 millones de
personas en el mundo (Nagai y Osame, 2003; Yoshida, 2005), y Chile se considera
como uno de los países con más alta seroprevalencia del virus, encontrándose en un
rango de 0,75 a 2 % de la población (Cartier y cols., 1992, 1997). A diferencia de lo
ocurrido con otros retrovirus como el virus de inmunodeficiencia humana tipo I, la
mayoría de los pacientes infectados con HTLV-I son clínicamente asintomáticos:
menos del 5 % de los individuos infectados desarrollan HAM/TSP. Esta enfermedad,
que normalmente comienza a partir de la cuarta década de vida, se caracteriza
clínicamente por debilidad muscular, hiperreflexia, espasticidad en las extremidades

6
inferiores y vejiga neurogénica, produciendo una pérdida lenta, progresiva y crónica
en la movilidad de las extremidades inferiores. Dicha sintomatología se correlaciona
con la neurodegeneración y desmielinación axonal selectiva y progresiva observada
en los haces corticoespinales de los pacientes, lo que origina una lesión de la vía
motora en los segmentos lumbares y dorsales de la médula espinal (Cartier y cols.,
1997). Histopatológicamente, esta enfermedad es una axonopatía central originada
por una alteración en el transporte axoplásmico que provoca un proceso degenerativo
axonal sin compromiso del cuerpo neuronal (Cartier y cols., 2007). Además, la
paraparesia espástica asociada al HTLV-I presenta semejanzas clínicas e histológicas
con las paraparesias espásticas familiares o hereditarias (Evans y cols., 2006; Park y
cols, 2010), en las cuales su origen se asocia a mutaciones en distintas proteínas que
perturban el transporte axoplásmico, sugiriendo también una patogenia de esta índole
en pacientes HAM/TSP.
Sin embargo, la etiología de la degeneración axonal en HAM/TSP se desconoce,
principalmente porque no se ha demostrado la infección de neuronas con HTLV-I in
vivo, aunque sí se relacionaría con la infiltración de los linfocitos infectados en el
sistema nervioso central (SNC). Se postula que la respuesta inmune contra el virus en
el SNC origina una reacción inflamatoria con efectos deletéreos sobre los axones
neuronales (Nagai y Osame, 2003). Pero esta suposición no explica
convincentemente la degeneración selectiva observada en HAM/TSP. Para explicar
este efecto, emergen otras teorías. Una de ellas sugiere una enfermedad de tipo
autoinmune, en donde anticuerpos contra proteínas virales presentan reactividad
cruzada con proteínas neuronales (Lee y cols., 2005). Otra teoría que es más reciente
y que es apoyada por nuestro grupo de investigación, es la de una acción indirecta de
los productos secretados por linfocitos infectados, entre ellos Tax, sobre las vías
intracelulares que regulan el transporte axonal (Alefantis y cols., 2005a; Maldonado
y cols., 2008; Alberti y cols., 2009).

7
Aproximadamente un 60 % de pacientes HAM/TSP son seropositivos para
HTLV-I (Cartier y cols., 1990; Ramírez y cols., 2003). Sin embargo, en el resto de
los pacientes a pesar de ser seronegativos para el virus, se puede detectar la presencia
tanto del gen como de la proteína Tax (Cartier y cols., 1990; Cartier y Ramírez,
2005), presentando además las mismas manifestaciones clínicas que los pacientes
seropositivos (Cartier y cols., 1999). En este caso en que no se detectan anticuerpos
contra proteínas virales de superficie, se considera que se ha insertado un provirus
defectuoso (Ramírez y cols., 2003). En vista de que estos individuos seronegativos
presentan HAM/TSP, pierden fuerzas las hipótesis sobre la patogenia de la
enfermedad concerniente a una degeneración de tipo inflamatoria en virtud de la
respuesta inmune generada contra los linfocitos infectados, y de una degeneración
por una respuesta autoinmune hacia proteínas neuronales homólogas a proteínas
virales. Por otro lado, la detección de la proteína Tax soluble en líquido
cefalorraquídeo (LCR) de pacientes con paraparesias seronegativas (Cartier y
Ramírez, 2005) acrecienta la relación de esta proteína con la patogenia de la
enfermedad. Esta observación ha hecho postular que la degeneración selectiva de los
axones largos del haz córtico-espinal es producto de la acción indirecta de Tax
extracelular (secretada por linfocitos infectados inclusive con un provirus defectuoso,
con presencia del gen tax) a través de vías transduccionales neuronales que
finalmente perturban el transporte axoplásmico. Esta última conclusión sugiere que el
transactivador viral Tax es el centro de las respuestas concernientes a la etiología de
la paraparesia asociada al HTLV-I (Alefantis y cols., 2005a, 2005b; Cartier y
Ramírez, 2005).
1.3. Proteína Viral Tax
Entre las proteínas codificadas por el HTLV-I, la oncoproteína Tax ejerce un
papel esencial en la transcripción viral y en la transformación celular, promoviendo

8
tanto la sobrevida como la patogénesis del virus en la población humana (Alefantis y
cols., 2005a; Boxus y cols., 2008; Boxus y Willems, 2009). Existen estudios que
responsabilizan a Tax de la transformación de los leucocitos y de las características
oncogénicas del virus (Grassmann y Fleckenstein, 1989; Akagi y cols., 1993;
Grossman y cols., 1995; Yoshida, 2010). La expresión del gen tax es necesaria y
suficiente para la inmortalización de linfocitos primarios humanos, confirmando la
importancia de esta proteína en la etiología de la ATL. Por estas razones, la proteína
Tax se estudia en relación a su papel en la transactivación transcripcional tanto de
genes virales como de una variedad de genes celulares durante el proceso de
transformación oncogénica en la patogenia del ATL (Nagai y Osame, 2003; Yoshida,
2005). Igualmente es importante su estudio durante la progresión de la infección viral
en HAM/TSP. Estos procesos son dependientes de la localización de Tax en el
núcleo de la célula infectada, en donde ejerce su papel como transactivador génico al
interactuar con una variedad de factores transcripcionales.
También existe importante evidencia que apoya un reclutamiento de Tax en el
citoplasma a modo de regular su acción sobre la transactivación transcripcional
(Alefantis y cols., 2005a). La colocalización de Tax citoplasmática con variados
organelos involucrados en las vías celulares secretoras (Alefantis y cols., 2005b; Jain
y cols., 2007), junto con la secreción de la proteína en células transfectadas con el
gen tax y en células infectadas con el virus, sugieren que la presencia de Tax en el
LCR de pacientes con HAM/TSP deriva de la secreción regulada desde linfocitos
infectados. Esto último conlleva una especial relevancia de este proceso sobre la
etiología de HAM/TSP (Cartier y Ramírez, 2005).
Tax promueve directamente la unión de factores transcripcionales tanto al LTR
viral como a promotores de varios genes celulares que incrementan la tasa de síntesis
de mRNA viral y celular. Una de las características más estudiadas de Tax como
promotor de sobrevida celular es su capacidad de activar constitutivamente el factor
transcripcional NF-κB al interactuar con la subunidad regulatoria de la quinasa que

9
fosforila (IKKγ) al inhibidor de NF-κB (IκB), permitiendo la translocación nuclear
del factor transcripcional. Por otro lado, en el núcleo, Tax interactúa con diversas
proteínas y factores transcripcionales, permitiendo así la transactivación
transcripcional de diversos genes celulares y virales. En el contexto del ciclo viral,
Tax nuclear interactúa con la maquinaria transcripcional del hospedero,
especialmente con la vía ATF/CREB para facilitar la unión de factores
transcripcionales al LTR, aumentando así la transcripción del mRNA viral.
Con todos estos antecedentes se observa que la regulación de la localización
subcelular de Tax y el conocimiento de los factores determinantes en dichos procesos
resulta fundamental para comprender cómo la expresión de genes celulares y virales
es controlada durante la infección viral y la progresión de las patologías asociadas al
HTLV-I.
1.4. Modificaciones postraduccionales de Tax
La localización subcelular de Tax en las células infectadas está en parte regulada
por la presencia de secuencias de localización nuclear (NLS) y de señales de
exportación nuclear (NES) en la proteína (Alefantis y cols., 2005a). Además, ésta
posee secuencias que le permiten interactuar con las vías formales de secreción en el
retículo endoplásmico (RE) y el complejo de Golgi (Alefantis y cols., 2005b; Jain y
cols., 2007) (Figura 4). La localización de la proteína Tax depende también de
interacciones con otras proteínas celulares, tales como proteínas pertenecientes al
complejo de poro nuclear. Sin embargo, los mecanismos implicados en el
intercambio núcleo-citoplasmático de Tax no han sido totalmente caracterizados
(Alefantis y cols., 2005a). Últimamente se ha descrito un nuevo mecanismo que
regula la localización de Tax al interior de la célula, relacionado directamente con su
efecto activador de genes y dependiente de modificaciones postraduccionales
sufridas por la proteína Tax, las cuales comprenden ubiquitinación y sumoilación

10
(Chiari y cols., 2004; Hilgarth y cols., 2004; Peloponese y cols., 2004; Lamsoul y
cols., 2005; Nasr y cols., 2006).
Figura 4: Dominios funcionales y señales de localización intracelular de Tax. Se representan distintos sitios de interacción de la proteína Tax, los sitios de localización (NLS) y exportación nuclear (NES), y las secuencias que le permiten interactuar con complejos requeridos en las vías de secreción (DHE y YTNI). (Modificado de Jain y cols., 2007).
Al investigar los mecanismos responsables del recambio proteico de Tax con la
finalidad de comprender la persistente infección con el virus, se encontró la presencia
de modificaciones postraduccionales tales como ubiquitinación y sumoilación (Chiari
y cols., 2004; Peloponese y cols., 2004; Lamsoul y cols., 2005; Nasr y cols., 2006).
La poliubiquitinación de Tax permite su degradación vía proteosoma. No obstante la
monoubiquitación y diferentes grados de poliubiquitinación se han relacionado con
diversos mecanismos en vez de la clásica degradación proteosomal, los cuales
incluyen la modulación de la interacción con otras proteínas y la localización
subcelular entre otras (Kerscher y cols., 2006). Al respecto, Tax no es la excepción,
describiéndose la mono y poliubiquitinación en residuos de lisina del extremo C-

11
terminal como una señal de retención citoplasmática (Lamsoul y cols., 2005; Nasr y
cols., 2006), modulándose así su actividad transcripcional.
Por otro lado, la sumoilación, modificación postraduccional bioquímicamente
similar pero funcionalmente distinta a la ubiquitinación, comprende la conjugación
de la proteína blanco con la proteína SUMO (“Small Ubiquitin-like Modifier”). Esta
modificación tiene un papel importante sobre la localización subcelular, en particular
mediando la destinación y/o retención nuclear y subnuclear de la proteína
modificada. También se asocia con la regulación de la interacción con otras
proteínas, así como la represión de la actividad de factores transcripcionales
(Kerscher y cols., 2006).
En el caso de Tax, se ha demostrado que su sumoilación es indispensable en la
retención nuclear, lo que permite ejercer su efecto activador en la transcripción de los
genes dependientes de NF-κB, pero no de los dependientes de CREB (Nasr y cols.,
2006). Se ha propuesto que tanto la ubiquitinación como la sumoilación de Tax son
indispensables para la activación de NF-κB en las células infectadas con el virus. La
localización citoplasmática de Tax mediada por su ubiquitinación permite la unión de
dicha proteína sobre IKKγ con la consecuente fosforilación y degradación de IκB,
mientras que en el núcleo, Tax sumoilada (en los mismos residuos de lisina en donde
se ubiquitina) media la formación de cuerpos nucleares en donde se asocia la
maquinaria de transcripción dependiente de NF-κB (Lamsoul y cols., 2005; Nasr y
cols., 2006). En consecuencia, la sumoilación y ubiquitinación regulan el intercambio
núcleo-citoplasmático de Tax, y con ello modulan la actividad transactivadora de la
proteína y por ende la evolución de la infección viral en las patologías asociadas al
HTLV-I. Sin embargo, los estudios sobre las modificaciones postraduccionales de
Tax sólo se han realizado en modelos de líneas celulares transfectadas con el gen tax
(Lamsoul y cols., 2005; Nasr y cols., 2006), y en ningún caso se han realizado en
muestras celulares de individuos infectados. Por esta razón, hasta el momento no se
ha relacionado el grado de modificación de la proteína Tax con la evolución de las

12
enfermedades asociadas al virus. Tampoco se han investigado las modificaciones
postraduccionales de Tax con la participación en las vías de secreción y consecuente
liberación de la proteína al medio extracelular, lo que conlleva especial relevancia en
el contexto de pacientes con HAM/TSP.
1.5. Contribución de Calreticulina sobre el intercambio núcleo-citoplasmático
de Tax
Además de sus funciones intracelulares, Tax extracelular también causaría una
variedad de efectos sobre otros tipos celulares, particularmente en el contexto de
HAM/TSP. La presencia de Tax secretada al LCR de pacientes con HAM/TSP
(Cartier y Ramírez, 2005), y la presencia de anticuerpos contra Tax en la mayoría de
los pacientes infectados con HTLV-I, demuestran que Tax secretada está disponible
para el reconocimiento y acción del sistema inmune y para mediar efectos
extracelulares deletéreos sobre otros tipos celulares no infectados (Alefantis y cols.,
2005a; Maldonado y cols., 2008). Además, se ha demostrado la secreción de Tax en
células transfectadas con el gen tax (Alefantis y cols., 2005b). Para que Tax
experimente una secreción controlada desde células infectadas, ésta debe localizarse
previamente en compartimientos definidos de la vía secretoria citoplasmática
(Alefantis y cols., 2005b, 2007), de manera que el conocimiento de los mecanismos
implicados en la exportación nuclear de Tax hacia las vías secretorias resultan
críticos a la hora de establecer el contexto global de las patogenias asociadas al
HTLV-I.
Tradicionalmente la exportación nuclear de proteínas que contienen NLS ricas en
leucina, como es el caso para Tax, es mediada por la proteína de exportación nuclear
CRM-1 a través del complejo de poro nuclear (Holaska y cols., 2001; Alefantis y
cols., 2007). Sin embargo, tal como se ha observado con otras proteínas, Tax no
experimenta exportación nuclear exclusivamente vía CRM-1, sino que también por

13
medio de la interacción con la proteína Calreticulina (CRT) como receptor de
exportación nuclear en el complejo de poro nuclear (Alefantis y cols., 2007),
añadiendo un nuevo nivel de complejidad en la exportación nuclear de Tax (Figura
5). De hecho, la distribución subcelular de Tax difiere según el tipo celular
transfectado con el gen tax, y se ha relacionado un mayor nivel de Tax citoplasmático
con una mayor expresión de CRT. También se ha encontrado una mayor expresión de
CRT en células infectadas con HTLV-I (Alefantis y cols., 2007), lo que se
correlaciona con la inducción de la expresión observada en esta proteína debido a
infecciones virales (Michalak y cols., 1999). Estos hechos dan cuenta de diferentes
eficiencias en la actividad transactivadora de Tax, replicación viral y patogénesis
viral entre las distintas poblaciones celulares. En particular, un aumento en los
niveles de CRT mediaría una mayor cantidad de Tax citoplasmática a costa de Tax
nuclear y con ello un aumento en la disponibilidad de Tax secretable, lo que trae
implicancias directas sobre la patogénesis de HAM/TSP. Sin embargo, la interacción
entre Tax y CRT y las implicancias que ésta conlleva a las enfermedades asociadas al
HTLV-I aún no se han estudiado en linfocitos infectados por el virus.
1.6. Planteamiento del problema
Hasta el momento no existen datos consistentes como para establecer los factores
predictivos en la evolución de la paraparesia asociada al HTLV-I, pero aparentemente
no se asocia ni con la carga proviral, ni con la cantidad de Tax presente en linfocitos
infectados.

14
Figura 5: Modelo de la distribución subcelular de Tax, las señales que la rigen y sus efectos en linfocitos infectados por HTLV-I. El modelo actualmente propuesto sugiere tres posibilidades de localización de Tax dentro de la célula, cada una con distintos efectos en la función de Tax. Primero, Tax se localiza en el núcleo gracias a su secuencia NLS y a su modificación por sumoilación, en donde actúa como transactivador transcripcional de diversos genes celulares y el LTR viral. Segundo, Tax se exporta hacia el citoplasma gracias a la secuencia de exportación nuclear y a la interacción con CRT en el complejo de poro nuclear. En el citoplasma, la ubiquitinación de Tax constituye una señal de retención citoplasmática, permitiéndole activar al factor transcripcional NF-κB. Tercero, Tax entra en las vías secretorias gracias a su secuencia de interacción con el complejo COPII en el RE y a su secuencia de destinación trans-Golgi. En el medio extracelular, Tax podría ocasionar HAM/TSP. (Modificado de Alefantis y cols., 2005a y de Lamsoul y cols., 2005).
Por otro lado, se ha demostrado que tanto las modificaciones postraduccionales de
Tax como su interacción con CRT en las células infectadas regulan el intercambio
núcleo-citoplasmático de Tax y con ello su actividad sobre la transactivación
transcripcional. Posiblemente estas interacciones también determinen los
mecanismos de secreción de la proteína y por ende la progresión de HAM/TSP
(Lamsoul y cols., 2005; Alefantis y cols., 2007). Pero ningún estudio ha relacionado
estas características en la población de células infectadas por el virus con la

15
evolución de la enfermedad y la susceptibilidad de desarrollar HAM/TSP en
individuos infectados con HTLV-I, de manera que surgen las siguientes
interrogantes:
¿Se correlacionan los niveles de ubiquitinación y sumoilación en Tax con la
severidad y progresión de la enfermedad?
¿La fracción de Tax ubiquitinada versus la sumoilada permitiría establecer los
factores predictivos en la evolución de HAM/TSP?
¿Participa CRT en los eventos de secreción de Tax desde linfocitos infectados?

16
2. HIPOTESIS
Como los datos sobre los niveles de Tax en linfocitos infectados no se
correlacionan con la severidad y evolución de la enfermedad, la base de este trabajo
es estudiar las modificaciones postraduccionales que experimenta Tax y su
interacción con Calreticulina en linfocitos infectados con el virus, para poder
relacionarlas con el nivel funcional de pacientes HAM/TSP, lo cual lleva a plantear la
siguiente hipótesis:
“Modificaciones postraduccionales en Tax y su interacción con Calreticulina en
PBMC de pacientes infectados con HTLV-I se relacionan con el nivel funcional
de pacientes HAM/TSP”.
El confirmar esta hipótesis permitiría a futuro utilizar las modificaciones y/o el nivel de
Calreticulina como marcadores de la progresión de HAM/TSP.

17
3. OBJETIVOS
3.1. Objetivo general
Analizar las formas modificadas de Tax y su interacción con Calreticulina en
linfocitos infectados con HTLV-I y relacionarlos con el nivel funcional de pacientes
HAM/TSP.
3.2. Objetivos específicos
3.2.1. Analizar las modificaciones postraduccionales de Tax proveniente de la
línea linfocitaria MT-2
Identificar las modificaciones postraduccionales presentes en la proteína Tax
obtenida de la línea linfocitaria MT-2 (células empleadas como modelo de estudio al
poseer el provirus del HTLV-I inserto en su genoma), utilizando Western Blot y
Espectrometría de Masas.
3.2.2. Cuantificar los niveles de Calreticulina y establecer su interacción con Tax
en células MT-2
Cuantificar mediante Western Blot los niveles de Calreticulina presentes en células
MT-2 e identificar la interacción entre Tax y CRT por ensayos de co-
inmunoprecipitación y colocalización mediante microscopía confocal.

18
3.2.3. Analizar las modificaciones postraduccionales de Tax proveniente de
PBMC aislados desde sangre periférica de pacientes HAM/TSP
Aislar mediante centrifugación en gradiente de ficoll los PBMC de individuos
controles, individuos portadores asintomáticos del virus y pacientes HAM/TSP con
distintos grados de marcha. A fin de concentrar y obtener la mayor cantidad de proteína
Tax desde las muestras pertinentes, los linfocitos se cultivan in vitro bajo condiciones
descritas para aumentar la expresión de Tax. El análisis de las formas modificadas de
Tax se realiza mediante Western Blot.
3.2.4. Cuantificar los niveles de Calreticulina y establecer su interacción con Tax
en PBMC aislados desde sangre periférica de pacientes HAM/TSP
Se efectúa igual que para las células MT-2 en el objetivo número 3.2.2,
considerando los PBMC aislados en 3.2.3.

19
4. MATERIALES Y MÉTODOS
4.1. Reactivos
Abcam Inc, Cambridge, MA, USA: Anticuerpos monoclonales contra SUMO-2/3
(Cat. Ab3742).
Amersham Pharmacia, Piscataway, NJ, USA: Ficoll-Hypaque.
Anticuerpos policlonales contra CRT, donados gentilmente por el Dr Arturo Ferreira
y Carolina Hager Ribeiro, Programa Disciplinario de Inmunología, ICBM, Facultad de
Medicina, Universidad de Chile, Independencia 1027, Santiago, Chile.
Anticuerpos contra Tax conjugados con Cy5, donados gentilmente por el Dr Yuetzu
Tanaka, Division of Immunology, Faculty of Medicine, University of Ryukyus,
Nishihara, Japón.
Beckton Dickinson Biosciences, San Jose, CA, USA: Anticuerpo monoclonal contra
CD4 humano conjugado con FITC (isotiocianato de fluoresceína) (Cat. 347413).
Biorad, Hércules, CA, USA: Membrana de nitrocelulosa “Trans-Blot® Transfer
Medium Pure Nitrocelulose Membrane (0,45 μm)” (Cat. 162-0115).
Bruker Daltonik GmbH, Alemania: Acido α-ciano-4-hidroxicinámico (CHCA).
Chemicon Internacional, Temecula, CA, USA: solución para bloqueo de membranas
de nitrocelulosa “Quick blocker” (Cat. B2080).
Calbiochem, La Jolla, CA, USA: EDTA.
Covalab, Villeurbanne, France: Anticuerpo monoclonal contra HTLV-1 Tax (Cat.
mab0022).

20
eBioscience Inc, San Diego, CA, USA: Tampón de fijación/permeabilización
“Fixation/Permeabilization, Concentrate” (Cat. 00-5123-43), Tampón de dilución para
fijación y permeabilización “Fixation & Permeabilization Diluent” (Cat. 00-5223-56).
Fermentas, Burlington, Ontario, Canadá: Estándar de pesos moleculares “Page
Ruler™ Prestained Protein Ladder” (Cat. SM0671).
Invitrogen Eugene, Oregon, USA: Anticuerpo contra IgG de conejo conjugado con
Alexa Fluor 594 “Alexa Fluor 594 goat anti-Rabbit IgG” (Cat. A11012), Anticuerpo
contra IgG de ratón conjugado con FitC “Rat anti-Mouse IgG1-FITC” (Cat. 04-6111),
Suero Fetal Bovino (Cat. 1600-044), Kit de aislamiento de linfocitos T reguladores
“Dynabeads® Regulatory CD4+CD25+ T Cell Kit” (Cat. 113.63D), medio de cultivo
“RPMI 1640 + GlutaMAX™-I” (Cat. 72400).
Merck, Darmstadt, Alemania: Acetona, Acetonitrilo, Acido acético, Acido
clorhídrico, Acido fórmico, Cloruro de Sodio, Desoxicolato de sodio, Etanol,
Isopropanol, Metanol, Persulfato de Amonio.
NIH, AIDS Reagent Program, Germantown, MD, USA: Anticuerpos policlonales
contra Tax “Antiserum to HTLV-I Tax” (Cat. 712), Anticuerpos monoclonales contra
Tax obtenidos directamente del medio de cultivo del Hibridoma: “HTLV-I Tax Hyb
168A51-2” (Cat. 1316).
PancReac, Barcelona, España: DMSO.
Pierce, Rockfort, IL, USA:, Anticuerpos contra conejo conjugados con peroxidasa
“InmunoPure Goat® Anti-Rabbit IgG (H+L), Peroxidase Conjugated” (Cat. 31460),
Anticuerpos contra ratón conjugados con peroxidasa “InmunoPure Goat® Anti-Mouse
IgG (H+L), Peroxidase Conjugated” (Cat. 31430), Kit para cuantificación de proteínas
“Micro BCA Protein Assay Kit™” (Cat. 23235), Matriz de inmunoprecipitación
“AminoLink Plus® Immobilization Kit” (Cat. 44894), Película para exposición “CL-
XPosure Film®” (Cat. 34090), Reactivo de quimioluminiscencia “Super Signal West

21
Femto® Maximum Sensivity Substrate” (Cat. 34095), Solución de tinción para geles
“Gelcode™ Blue safe protein stain” (Cat. 1860957).
Promega, Madison, WI, USA: Tripsina grado espectrometría de masas para secuenciar
“Sequencing Grade Modified Trypsin” (Cat. V5111).
Riedel-De Hain AG, Seelze-Hannover, Germany: Azul de Bromofenol.
Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA: Anticuerpos policlonales
contra TFIIB “TFIIB (C-18) Antibody” (Cat. sc-225).
Sigma-Adrich Inc., St Louis, MO, USA: Acrilamida, Anticuerpos monoclonales
contra GAPDH (Cat. G8795), Bicarbonato de sodio, Bisacrilamida, Ditiotreitol (DTT),
Fenilmetilsulfonil fluoruro (PMSF), Glicina, Iodoacetamida, Medio de montaje para
inmunofluorescencia “Gel mount™ Aqueous mounting medium” (Cat. G0918), N-
etilmaleimida, Dodecil sulfato de sodio (SDS), Trizma Base, β-Mercaptoetanol, Triton
X-100, Nonidet P-40, TEMED.
Upstate, Lake Placid, NY, USA: Anticuerpos monoclonales contra Ubiquitina (Cat.
04-263), Anticuerpos monoclonales contra SUMO-1 (Cat. 04-453).
US Biological, Swampscott, MA, USA: Tween 20 (Cat. 9005-64-6).
4.2. Material Biológico
El material biológico empleado consistió en:
- Líneas celulares MT-2 y K562 cedidas gentilmente por el Dr Eugenio Ramírez (ISP,
Santiago, Chile).
- Muestras sanguíneas de individuos controles y asintomáticos portadores del HTLV-I
obtenidas por el Dr Eugenio Ramírez en el Subdepartamento de Virología del ISP y

22
muestras de pacientes HAM/TSP con diversos grados de marcha que correspondieron a
pacientes diagnosticados clínicamente por el Dr Luis Cartier en el Hospital El Salvador,
Santiago, Chile. El procedimiento de obtención de estas muestras de pacientes y
controles se efectuó según el protocolo aprobado por la Comisión de Ética de la
Facultad de Medicina de la Universidad de Chile.
4.3. Metodología
4.3.1. Cultivos celulares
Las líneas celulares se cultivaron en medio “RPMI 1640 + GlutaMAX™-I”
suplementado con 10 % de suero fetal bovino y se cultivaron a 37 ºC y 5 % de CO2.
Para el cultivo de cada muestra de PBMC, un total de 2x106 PBMC fueron
cultivados durante 24 horas a 37 ºC y 5 % de CO2 en 1 mL de medio “RPMI 1640 +
GlutaMAX™-I” suplementado con 10 % de suero fetal bovino en presencia de 20 nM de
Concanamicina A (para inhibir perforinas) con el objetivo de detectar la proteína Tax.
Sin un cultivo previo de los PBMC, es imposible observar la proteína Tax mediante
Citometría de Flujo (Hanon y cols., 2000).
4.3.2. Aislamiento de PBMC desde sangre periférica
El aislamiento de PBMC desde sangre periférica se realizó mediante una
centrifugación en gradiente de densidad con Ficoll-Hypaque. En una campana de
extracción, para cada muestra a aislar se mezclaron dos mL de sangre periférica fresca
(tomada en tubos con anticoagulante) con dos mL de PBS (Phosphate Buffer Saline;
137 mM NaCl, 2,7 mM KCl, 100 mM Na2HPO4, 2 mM KH2PO4, pH 7,2) estéril y se
vertieron en un tubo de centrífuga estéril que contiene 3 mL de Ficoll-Hypaque (sin
mezclar la sangre con Ficoll-Hypaque). Se centrifugó a 400 g durante 30 min a

23
temperatura ambiente. Posteriormente se eliminó la capa superior correspondiente al
plasma sanguíneo, y se recolectó la capa siguiente (entre el plasma y el Ficoll-Hypaque)
que corresponde a la fracción de PBMC.
4.3.3. Lisados celulares
Previo al lisado, los linfocitos en suspensión se lavaron cinco veces con PBS. Los
lisados celulares se efectuaron siguiendo dos metodologías distintas:
4.3.3.1. Lisado celular total
Se obtuvo agregando a la pella celular 500 µL cada 2x107 de linfocitos de un
tampón de lisis 50 mM Tris-HCl pH 7,5, 150 mM NaCl, 1 % Tritón X-100, 0,5 %
Nonidet P-40, 10 mM N-etilmaleimida, 0,2 mM Na3VO4, 0,1 mM PMSF. Se incubó en
hielo durante 30 min agitando suave y constantemente y se centrifugó a 4 ºC durante 5
min a 16.000 g para eliminar los debris celulares. El sobrenadante corresponde al lisado
celular total.
4.3.3.2. Lisado y fraccionamiento celular
Se siguió el protocolo descrito por Kfoury y cols., 2008 para lisar linfocitos y
obtener tres fracciones distintas enriquecidas en componentes citoplasmáticos,
membranosos/organelares y nucleares. Las células (2x107 linfocitos) fueron lisadas en
hielo durante 10 min agregando a la pella celular 300 µL de una solución 10 mM
HEPES pH 7,9, 10 mM KCl, 1,5 mM MgCl2, 1mM DTT, 10 mM N-etilmaleimida, 0,2
mM Na3VO4, 0,1 mM PMSF. Se centrifuga a 4 ºC con 400 g durante 10 min para
obtener el sobrenadante, el que corresponde a la fracción citoplasmática. Posteriormente

24
la fracción membranosa/organelar se obtiene lavando la pella restante tres veces con
100 µL con un tampón 20 mM Tris-HCl pH 7,4, 100 mM NaCl, 1 % Tritón X-100, 0,5
% Desoxicolato de Sodio, 10 mM N-etilmaleimida, 0,2 mM Na3VO4, 0,1 mM PMSF.
Cada vez se centrifuga a 4 ºC con 400 g durante 10 min. La pella restante se lisa en
hielo incubando por 30 min con 300 µL de una solución 50 mM Tris-HCl pH 7,5, 150
mM NaCl, 0,1 % SDS, 0,5 % Tritón X-100, 0,5 % Nonidet P-40, 10 mM N-
etilmaleimida, 0,2 mM Na3VO4, 0,1 mM PMSF. Finalmente todas las fracciones se
limpiaron centrifugando a 12.000 g durante 10 min.
4.3.4. SDS-PAGE y Western Blot
Se separaron muestras de lisados celulares conteniendo 50 µg totales de proteínas o
muestras de 20 µL de medio de cultivo de PBMC mediante geles de poliacrilamida al
10 % en condiciones desnaturantes y reductoras, SDS-PAGE (Laemmli, 1970) de 0,75
mm de espesor, para luego ser electrotransferidas a una membrana de nitrocelulosa de
tamaño de poro 0,45 μm (Towbin y cols., 1979) utilizando una solución 25 mM Tris,
190 mM glicina y 20 % de metanol y aplicando una amperaje total de 600 mA. Luego
del tiempo necesario de electrotransferencia, la membrana se bloqueó durante 20 min a
temperatura ambiente en una solución de bloqueo al 5 % p/v de “Quick blocker”
disuelta en amortiguador de TBS-T (“Tris Buffer Saline with Tween”: Tris 20mM,
NaCl 137 mM, 0,1 % Tween-20, pH 7,6). Posteriormente la membrana se incubó con el
correspondiente anticuerpo primario en la dilución determinada en TBS-T (Tabla I)
durante toda la noche a 4 ºC o durante 2 h a temperatura ambiente.
Luego de la incubación, la membrana se lavó sucesivamente con TBS-T (5 lavados
de 5 min), para luego incubarla con un segundo anticuerpo conjugado a peroxidasa
contra IgG de la especie del primario, diluído 1:500.000. Posteriormente la membrana
se lavó 5 veces durante 5 min cada lavado con TBS-T, se identificó la proteína de
interés mediante la reacción de quimioluminiscencia empleando el kit “Super Signal

25
West Femto® Maximum Sensivity Substrate” de Pierce. La membrana fue entonces
expuesta a una placa fotográfica durante tiempos variables, y los resultados se
obtuvieron al revelarlas. Para el análisis de datos, las bandas obtenidas se cuantificaron
mediante el programa “Uni-Scan-It Automated Digitizing System”.
Para los casos en que se requirió deshibridizar el anticuerpo unido a las proteínas de
la membrana para repetir el western blot empleando otro anticuerpo, se lavó la
membrana dos veces durante 10 min cada lavado con una solución deshibridizadora 200
mM Glicina, 3,5 mM SDS y 10 % Tween-20 a pH 2,2.
4.3.5. Inmunoprecipitación
Para las inmunoprecipitaciones se utilizó el kit “AminoLink Plus® Immobilization
Kit” (Pierce) siguiendo las instrucciones del fabricante, pero utilizando el equivalente a
250 µL de matriz seca para cada inmunoprecipitado y durante el protocolo de
acoplamiento a pH 7,2 se escalaron todos los volúmenes de las soluciones empleadas al
volumen equivalente de matriz empleada. Los anticuerpos acoplados a la matriz
correspondieron a 30 µL de líquido ascítico contra Tax (Covalab) y 20 µL de suero

26
inmune contra CRT. En cada reacción se inmunoprecipitaron las proteínas desde 500 µg
totales de lisados celulares o 500 µL totales de medio de cultivo de PBMC.
Para observar la co-inmunoprecipitación, se inmunoprecipitó manteniendo la
incubación y los lavados de la matriz a 4 ºC para impedir la disociación por temperatura
de los complejos proteicos.
4.3.6. Inmunofluorescencia Indirecta y Análisis de Imágenes
104 células MT-2, K562 o linfocitos CD4+ aislados de PBMC previamente lavados
4 veces con PBS a 37 ºC (Phosphate Buffer Saline; 137 mM NaCl, 2,7 mM KCl, 100
mM Na2HPO4, 2 mM KH2PO4, pH 7,2) se depositaron en un portaobjeto como una
suspensión celular de 10 µL, y se dejó secar a temperatura ambiente (aproximadamente
2 a 3 h, necesario para la adhesión de las células al portaobjeto) para luego ser fijadas y
permeabilizadas con acetona a 4 ºC durante 8 min. Posteriormente las células fijadas se
incubaron con 25 µL de la dilución del anticuerpo primario respectivo [1:50 para el
anticuerpo monoclonal contra Tax (Covalab) y 1:200 para el anticuerpo policlonal
contra CRT] durante 40 min a 37 ºC. Luego se efectuaron tres lavados sucesivos con
250 µL de PBS y se incubaron las células en oscuridad por 40 min a 37 ºC con 25 µL de
una dilución de los anticuerpos secundarios respectivos (1:200 para el anticuerpo
conjugado con FitC y 1:400 para el anticuerpo conjugado con Alexa Fluor 594),
incluyendo en ella el marcador nuclear TO-PRO diluido 1:400. Para el caso de las
tinciones dobles, una misma solución contiene la dilución respectiva de ambos
anticuerpos y del TO-PRO. A continuación se lavaron las células tres veces con 250 µL
de PBS y una vez con agua bidestilada, se dejó secar a temperatura ambiente (en
oscuridad), se montó con un cubreobjeto utilizando 2 µL de medio de montaje acuoso y
se sellaron las orillas de los cubreobjetos con esmalte de uñas transparente. Las
inmunofluorescencias posteriormente se observaron en un microscopio de fluorescencia
confocal “Carl Zeiss LSM 510 Meta”. Las imágenes se obtuvieron con el programa

27
“LSM 510 Image Browser” y se analizó la colocalización con el programa “ImageJ,
plugin Colocalization Finder”.
4.3.7. Citometría de Flujo
Las células (2x106) fueron lavadas tres veces con PBS y luego incubadas 30 min a 4
ºC con el anticuerpo de superficie anti CD4 humano conjugado con FITC diluido 1:50.
Posteriormente se efectúan 2 lavados de 100 µL con PBS, centrifugando a 2000 g por
dos minutos cada uno en una centrífuga de placas. Luego se fijan las células con 100 µL
de tampón de fijación/permeabilización (eBioscience) durante 20 min a temperatura
ambiente. Una vez terminada la incubación, se lava con 100 µL de tampón de dilución
para fijar y permeabilizar (eBioscience) dos veces y centrifugando durante 2 min cada
vez. Luego se permeabiliza con 100 µL de tampón de fijación/permeabilización durante
15 min a 4 ºC y se lava de nuevo dos veces con 100 µL de tampón de dilución
centrifugando cada vez a 2000 g por 2 min. Posteriormente se incuba 30 min a 4 ºC con
el anticuerpo intracelular; anticuerpos contra Tax conjugados con Cy5 (cedidos
gentilmente por el Dr Yuetzu Tanaka) diluido 1:65 en tampón de dilución. Después se
lava 2 veces con 100 µL de tampón de dilución y se centrifuga cada vez a 2000 g por 2
min. Finalmente las células se resuspenden en 100 µL de PBS con 3 % de suero fetal
bovino y se analizaron las poblaciones en un citómetro de flujo Beckton Dickinson,
modelo FACS CANTO II.
4.3.8. Aislamiento de la fracción de Linfocitos T CD4+
Se aisló la población de linfocitos T CD4+ a partir de PBMC mediante el kit de
separación por esferas inmunomagnéticas de linfocitos T CD4+CD25+ “Dynabeads®
Regulatory CD4+CD25+ T Cell Kit”. El protocolo de aislamiento consistió en la

28
separación descrita por el kit hasta la primera separación celular correspondiente a la
purificación negativa de linfocitos T CD4+.
4.3.9. Espectrometría de Masas
4.3.9.1. Digestión de Proteínas en Gel
Luego de realizar la electroforesis en SDS-PAGE se cortaron las bandas requeridas,
transfirieron a un tubo de microcentrífuga y se lavaron 3 veces con 500 µL de agua
nanopura durante 15 min a temperatura ambiente. Luego se destiñeron las bandas con
150 µL de bicarbonato de amonio 200 mM preparado en acetonitrilo 50 % v/v durante
30 minutos a 30 ºC. Se lavó con esta solución hasta que la banda se vio opaca. Se
centrifugó y se removió el exceso de solución. Luego se agregaron 50 µL de DTT 10
mM preparado en bicarbonato de amonio 200 mM y se incubó 20 min a 37 ºC.
Posteriormente se removió el sobrenadante y se agregó 50 µL de iodoacetamida 55 mM
preparada en bicarbonato de amonio 200 mM. Se incubó a 37 ºC durante 20 min en
oscuridad. Se removió el exceso de iodoacetamida y se agregó 100 µL de acetonitrilo
100 %, incubando 5 min a 37 ºC. A continuación se agregó 50 µL de acetonitrilo 100 %
y se incubó a 37 ºC durante 5 min. Luego se secó a temperatura ambiente durante 10
min y se dejó en hielo. Se agregó 8 µL de amortiguador de digestión bicarbonato de
amonio 50 mM/acetonitrilo 10 % v/v que contenía 0,3 µg de tripsina, y se incubó en
hielo durante 45 min. Posteriormente se agrega 20-40 µL de bicarbonato de amonio 50
mM/acetonitrilo 10 % v/v de modo de cubrir completamente la banda, y se incubó toda
la noche a 37 ºC. Al día siguiente (12-18 h) se recuperó el sobrenadante y se transfirió a
un nuevo tubo de microcentrífuga. La banda se lavó dos veces con 20 µL de acetonitrilo
60 % v/v / ácido fórmico 0,1 % v/v y se mezcló durante 2 min en vórtex, y el
sobrenadante se combinó con el sobrenadante inicial, los cuales finalmente se
concentraron mediante centrifugación al vacío a temperatura ambiente, y la pella se
almacenó a -20 ºC hasta su análisis.

29
4.3.9.2. Análisis Peptídico mediante MALDI-TOF
Luego de ser concentrados hasta sequedad, los péptidos trípticos se resuspendieron
en 10 μL de ácido fórmico 0,1 % v/v. Se preparó una solución stock de matriz de ácido
α-ciano-4-hidroxicinámico (CHCA) 10 mg/mL en acetonitrilo/ácido fórmico 0,1 % v/v
1:1. Se colocaron porciones de 1 μL de una mezcla 1:1 o 1:10 de la muestra y la
solución de la matriz sobre una placa porta muestra “micro scout” (Bruker Daltonics,
Inc. MA-USA). Los espectros de masa se obtuvieron en un equipo MALDI-TOF
Microflex (Bruker Daltonics, Inc. MA-USA) en modo ion positivo mediante detección
por reflexión. La ionización se realizó mediante la utilización de pulsos de láser
ultravioleta para lo cual se utilizó un láser de nitrógeno de longitud de onda de 337 nm
con una energía de pulso de 150 μJ y frecuencia de 20 Hz. Se trabajó bajo los siguientes
parámetros: voltaje de fuente de iones 1, 19,0 kV; voltaje de fuente de iones 2, 16,3 kV;
voltaje de lentes, 9,75 kV; voltaje de reflector, 20,0 kV y tiempo de extracción pulsada
de iones, 100 ns. Para la supresión de señales de la matriz se utilizó modo de deflexión
de hasta 500 Da. Previo a la obtención de los espectros se realizó una calibración del
equipo con un estándar externo correspondiente a una mezcla de péptidos de masas
1.000-3.000 Da (Bruker Daltonics, Inc. MA-USA). Los espectros finales corresponden
a espectros sumados correspondientes a 15 barridos de 30 pulsos de láser aplicados en
diferentes puntos tomados al azar a lo largo de cada placa para cada muestra.
Para el análisis de los espectros, se buscaron los picos mediante el programa
FlexAnalysis versión 2.2 (Bruker Daltonik GmbH, Alemania). Para la detección de la
señales m/z se utilizó el algoritmo SNAP (sophisticated numerical annotation
procedure) considerando los parámetros por defecto. Una vez generadas las listas de
señales m/z se realizó una calibración interna utilizando las señales de la autoproteólisis
de la tripsina, las que se removieron de la lista y luego las señales restantes se
exportaron a Excel. Adicionalmente, las listas de masas exportadas de cada muestra se
examinaron por medio del programa PeakErazor versión 2.0.1 (Lighthouse data,
Odense, Denmark) en búsqueda de señales provenientes de la contaminación con

30
queratina las que también se removieron de las listas. Finalmente, las listas exportadas y
filtradas se utilizaron para su identificación en tres bases de datos disponibles en línea:
Mascot (www.matrixscience.com), MS-Fit (http://prospector.ucsf.edu/prospector/cgi-
bin/msform.cgi?form=msfitstandard) y de PeptideMap (PROWL The Rockefeller
University, http://prowl.rockefeller.edu), considerando para ambas 3 cortes no
realizados por la tripsina, la base de datos de Swissprot, carbamidometil en cisteína
como modificación fija, oxidación en metionina como modificación variable, y una
tolerancia de 0,1 Da.
4.3.9.3. Análisis Peptídico mediante HPLC-ESI-IT
Luego de ser concentrados hasta sequedad, los péptidos trípticos se resuspendieron
en 10 μL de ácido fórmico 0,1 % v/v. La muestra se examinó en un sistema LC-MS que
consiste del cromatógrafo HPLC Agilent 1100 (Agilent Technologies Inc.) acoplado al
espectrómetro de masas de tipo electrospray-trampa iónica ESI-TRAP Esquire 4000
(Bruker Daltonik GmbH, Alemania). Para el control del cromatógrafo HPLC se utilizó
el programa ChemStation for LC 3D Rev. A.10.02 (Agilent Technologies Inc., CA-
USA) y para el control del espectrómetro se utilizó el programa EsquireControl 5.2
(Bruker Daltonik GmbH, Alemania). Para la separación HPLC se utilizó una columna
C18 de 150×1,0 mm, 4 μm y 90 Å (Jupiter-Proteo, Phenomenex Inc., CA-USA), a la
salida de la cual se conectó directamente al espectrómetro de masas.
La separación de la muestra (30 μL totales de muestra inyectada, que consiste en
una dilución 1:6 de la muestra original resuspendida en ácido fórmico) en el HPLC se
realizó a temperatura ambiente utilizando ácido fórmico 0,1 % v/v en agua (Fase A) y
ácido fórmico 0,085 % v/v en acetonitrilo 80 % v/v (Fase B) bajo el siguiente programa:
0-10 min 2 % B, 10-70 min 2-100 % B, 70-80 min 100 % B a un flujo de 50 μL/min.

31
El proceso de ionización mediante electrospray se realizó utilizando nitrógeno a una
temperatura de nebulización de 325 ºC, presión de nebulización de 20 psi y flujo de gas
nebulizador de 6 L/min.
Las condiciones espectrométricas para la adquisición de datos en polaridad positiva
fueron: voltaje de ionización, 3000 V; offset de placa terminal (end plate), -500 V;
skimmer 1, 30,0 V; skimmer 2, 6,0 V; offset de salida de capilar, 60,0 V; salida de
capilar, 90,0 V; diferencia de octupolos, 2,40 V; trap drive, 55,0; voltaje de lente 1, -5,0
V; voltaje de lente 2, -60,0 V. La adquisición de espectros se realizó para el intervalo
20-2200 m/z, utilizando scan estándar y resolución normal (velocidad de scan 10300
m/z/s; ancho de peak 0.6 FWHM/m/z). La trampa de iones se utilizó bajo modo de
control de carga de iones (ICC activado), un tiempo máximo de acumulación de 200 ms.
La disociación inducida por colisión (CID) se efectuó mediante helio presente en la
trampa. La fragmentación se realizó utilizando los siguientes parámetros: SmartFrag,
30-200 %; amplitud de fragmentación, 1,00 V; tiempo de fragmentación, 40 ms; ancho
de aislamiento MS(n), 4 m/z; promedios MS(n), 5; número de iones precursores
AutoMS(2), 5 señales; y la intensidad umbral AutoMS(2) se fijó previo a cada
inyección. Adicionalmente, para la selección de los iones precursores se utilizó modo de
exclusión activa (análisis dependiente de datos) bajo los siguientes parámetros:
exclusión, 2 espectros; tiempo de exclusión, 1 min.
Para el análisis de los cromatogramas y espectros se utilizó el programa
DataAnalysis versión 3.2 (Bruker Daltonik GmbH, Alemania). Para la generación de
datos se utilizó la opción Compounds-AutoMSn que permitió la detección de señales
m/z y sus correspondientes fragmentaciones. Los datos generados fueron procesados
mediante desconvolución para la determinación de sus correspondientes cargas.
Finalmente, los datos procesados se exportaron como un archivo mgf (Mascot generic
format) para su análisis de identificación. Para el análisis de identificación se utilizó la
base de datos Mascot por medio de la opción MS/MS Ion Search
(http://www.matrixscience.com/cgi/search_form.pl?FORMVER=2&SEARCH=MIS).

32
Para la identificación se consideraron las siguientes restricciones: modificación por
iodoacetamida (modificación fija), oxidación de metionina (modificación variable),
ESI-TRAP (instrumento), tolerancia de masas de 1,0 Da y una tolerancia de
fragmentación de 0,5 Da. Las búsquedas no incluyeron restricción por taxonomía.
Alternativamente, se construyó una base de datos local en Mascot (Matrix Science,
UK) con las secuencias peptídicas de Tax, Ubiquitina, SUMO-1, SUMO-2, SUMO-3 y
gp68 obtenidas de GenBank con los números de acceso BAB18052.1, CAA44911.1,
AAC50996.1, P61956.1, P55854.2 y AAU04944.1 respectivamente. El análisis
consideró los siguientes parámetros: enzima de proteólisis, tripsina; pérdida de cortes, 3;
modificación fija, iodoacetamida (carbamidometilación de cisteínas); modificación
variable, oxidación de metionina; tolerancia de masas, 0,8 Da; tolerancia de
fragmentación, 0,3 Da; valores de masa, MH+ y monoisotópicos.
4.3.10. Análisis Estadístico
Los datos obtenidos de los western blots se analizaron estadísticamente utilizando
los programas “GraphPad Prism” y “SPSS” (Statistical Package for the Social Science).
Las diferencias entre los grupos control, asintomáticos y TSP/HAM se evaluaron
usando el test de Mann-Whitney. La correlación entre los grupos de datos se consideró
significativa según el coeficiente de correlación de Spearman. La significancia
estadística fue definida con un p < 0,05.

33
5. RESULTADOS
5.1. Analizar las modificaciones postraduccionales de Tax proveniente de la línea
linfocitaria MT-2
Dependiendo de la localización subcelular, la proteína viral Tax podrá ejercer
diferentes acciones sobre los linfocitos T infectados. En el núcleo, Tax puede favorecer
el ciclo replicativo del virus y la expresión de proteínas celulares que promuevan la
transformación celular, mientras que en el citoplasma Tax puede afectar la sobrevida de
la célula y reclutarse en vesículas secretorias. De tal manera, la ubicación y las
modificaciones postraduccionales que modulan esta localización en la proteína Tax
influyen en la regulación y el control del ciclo viral durante la infección, y
probablemente en la progresión de las patologías asociadas al HTLV-I. Es por esta
razón que en primera instancia se espera relacionar la localización subcelular de la
proteína viral Tax en células MT-2 con la presencia de modificaciones
postraduccionales en ella, para lo cual se efectuó un protocolo de lisis celular de
linfocitos que permite obtener tres diferentes fracciones, cada una enriquecidas en
componentes citoplasmáticos, componentes membranosos/organelares y componentes
nucleares (Kfoury y cols., 2008).
Las líneas celulares MT-2 y K562 sometidas a este fraccionamiento subcelular
muestra por western blot los resultados presentados en la Figura 6, en donde se observó
la presencia de Tax (peso molecular teórico de 40 kDa) utilizando tres distintos
anticuerpos contra ella. Se emplearon como marcadores de las distintas fracciones
anticuerpos contra gliceraldehído 3-fosfato deshidrogenasa (GAPDH) para la fracción
citoplasmática, anticuerpos contra CRT para la fracción membranosa/organelar y
anticuerpos contra el factor transcripcional II B (TFIIB) como marcador nuclear
(Figura 6d).

34
Figura 6: Identificación de la proteína viral Tax en células MT-2. Western blot de las fracciones citoplasmáticas (1), membranosas/organelares (2) y nucleares (3) en células MT-2 y K562, utilizando (a) anticuerpos monoclonales contra Tax (Covalab, Villeurbanne, France), (b) anticuerpos policlonales contra Tax (NIH, AIDS Reagent Program), (c) anticuerpos monoclonales contra Tax (NIH, AIDS Reagent Program, HTLV-I Tax Hyb 168A51-2), (d) anticuerpos contra GAPDH, CRT y TFIIB como marcadores citoplasmático, membranoso/organelar y nuclear, respectivamente. En cada carril se cargó un total de 50 µg de proteína. A la izquierda de cada western blot se representa el peso molecular aparente en kDa. Las flechas rojas indican la presencia de la proteína Tax expresada en células MT-2 tanto a 40 como a 71 kDa.
Todos los anticuerpos diferentes contra Tax reconocen fuertemente una banda que
migra a 71 kDa en lisados de células MT-2, lo cual indica que se trata de la proteína
viral Tax. También se observa la presencia de una pequeña banda a 40 kDa que
concuerda con el peso molecular teórico de la proteína sin modificaciones
postraduccionales. Los anticuerpos empleados como marcadores de las distintas
fracciones celulares indican que la proteína Tax en la línea celular MT-2 se sitúa
predominantemente en la fracción membranosa/organelar, donde deben estar presentes
vesículas secretoras y complejos organelares del RE y del Aparato de Golgi.
Debido a la discrepancia del peso molecular teórico de la proteína Tax con el peso
molecular experimental de 71 kDa observado en la Figura 6, se propone que esta

35
especie se encuentra modificada con proteínas pequeñas, como SUMO o Ubiquitina,
que den cuenta del retardo electroforético observado.
Para estudiar la presencia de modificaciones postraduccionales en la proteína Tax, se
realizaron western blot de las fracciones de lisis de células MT-2 y K562 utilizando
anticuerpos contra Ubiquitina, SUMO-1, SUMO-2/3 y se identificó la presencia de
inmunorreactividad a 71 kDa (Figura 7).
Figura 7: Determinación de las modificaciones postraduccionales de Tax en células MT-2. Western blot de las fracciones citoplasmáticas (1), membranosas/organelares (2) y nucleares (3) en células MT-2 y K562, utilizando anticuerpos contra las proteínas pequeñas Ubiquitina (Upstate, Lake Placid, NY, USA), SUMO-1 (Upstate, Lake Placid, NY, USA), SUMO-2/3 (Abcam Inc, Cambridge, MA, USA). En cada carril se cargó un total de 50 μg de proteína. A la izquierda de cada western blot se representa el peso molecular aparente en kDa. La flecha roja indica la presencia de inmnorreactividad a 71 kDa.
Como se puede observar en la Figura 7, la presencia de señales al utilizar
anticuerpos contra Ubiquitina dificulta el análisis al no distinguirse bandas discretas. Al
emplear anticuerpos contra SUMO-1 no se detecta ninguna banda a 71 kDa que pueda
corresponder a una modificación de Tax. Sólo al utilizar anticuerpos contra SUMO-2/3
se distingue una inmunorreacción a 71 kDa presente en la fracción nuclear de las células
MT-2 (flecha roja en la Figura 7).
Para confirmar y/o descartar la presencia de modificaciones postraduccionales y
relacionarlas con la especie de Tax a 71 kDa, a partir de la muestra del lisado celular de

36
MT-2 se inmunoprecipitó la proteína Tax empleando anticuerpos monoclonales
(Covalab, Villeurbanne, France) y posteriormente se analizó por western blot, revelando
la completa ausencia de inmunorreactividad para Ubiquitina, SUMO-1 y SUMO-2/3
(datos no mostrados). Estos resultados indican que la proteína Tax presente en células
MT-2 no se encuentra modificada con las proteínas pequeñas estudiadas.
En la literatura se reconoce la especie de Tax a 71 kDa expresada en células MT-2,
pero no se relaciona este aumento en el peso molecular con la presencia de proteínas
pequeñas conjugadas a Tax, sino que más bien se cree que corresponde a una proteína
de fusión viral entre Tax y Env o Gag (Miwa y cols., 1984; Jeang y cols., 1997). De
estas dos proteínas, Env se describe N-glicosilada. Esta proteína corresponde a una
especie inmadura, la cual se convierte en funcional al sufrir un procesamiento
proteolítico que da origen a las proteínas gp46 y gp21. Para determinar la presencia de
N-glicosilaciones en estas dos bandas Tax positivas de 40 y 71 kDa, se obtuvo un lisado
de células MT-2 cultivadas en presencia de tunicamicina (inhibidor de N-glicosilación)
y se observó mediante western blot si había cambio en la migración electroforética de la
proteína Tax (Figura 8). En presencia de tunicamicina se encontró una banda de 60 kDa
en vez de la banda a 71 kDa, lo cual indica que esta última especie está N-glicosilada.
Como se esperaba, no se observó cambios en el peso molecular de la otra banda de Tax
a 40 kDa (Tax sin modificaciones).
Sin embargo, aún existe un retardo electroforético importante entre las especies a 60
y 40 kDa. Esta vez puede explicarse por la proteína de fusión sugerida en la literatura
(Miwa y cols., 1984). En la línea celular MT-2 se han descrito al menos 7 formas del
virus HTLV-I las cuales se encuentran alteradas en su composición genómica dentro de
la célula y que corresponden a versiones defectuosas del virus (Kobayashi y cols.,
1984).

37
Figura 8: Identificación de la N-glicosilación de la banda a 71 kDa en células MT-2. Western blot de lisados totales de células MT-2 y K562 utilizando anticuerpos monoclonales contra Tax (NIH, AIDS Reagent Program, HTLV-I Tax Hyb 168A51-2). El carril 1 representa un control de 106 células incubadas durante 24 h en medio RPMI 1640 (Invitrogen Eugene, Oregon, USA) suplementado con 10 % de suero fetal bovino, mientras que el carril 2 corresponde a la misma cantidad de células incubadas en las mismas condiciones y en presencia de 40 ng/mL de tunicamicina. En cada carril se cargó un total de 50 µg de proteína. A la izquierda se representa el peso molecular aparente en kDa.
Es probable que alguna de estas formas presente una deleción entre los marcos de
lectura abiertos para las proteínas Env y Tax, de tal forma que no exista un codón de
término entre ambos. De ser así, la polimerasa del hospedero transcribirá continuamente
los genes en las regiones env y pX, dando como resultado una proteína quimera Env-
Tax. Posteriormente la proteasa viral cortará la proteína Env, como lo haría en un
proceso de replicación viral normal, dando como resultado de la proteólisis la proteína
gp46 y gp21-Tax.
La proteína gp21 tiene un peso molecular de 21 kDa, de modo que la fusión de ésta
con Tax pesaría alrededor de 60 kDa, justamente lo observado en la Figura 8 para las
células incubadas en presencia de tunicamicina. También, gp21 se encuentra glicosilada
(Sugamura y cols., 1984), de modo que la presencia de glicosilaciones en la especie de
71 kDa observada en células MT-2 puede atribuírsele a la fracción gp21 de la quimera,
ya que la proteína Tax no se ha descrito N-glicosilada, ni tampoco posee sitios
potenciales de N-glicosilación. El hecho que la proteína gp21 sea una proteína
transmembrana que se localiza en la membrana de la célula infectada, podría explicar la
presencia predominante de la especie a 71 kDa en las fracciones
membranosas/organelares de los lisados diferenciales realizados en los experimentos de
la Figura 6. Por último, de transcribirse continuamente los marcos de lectura en env y
pX, la transcripción de la región pX tendría una menor regulación negativa, ya que
estaría regida por el promotor de env, lo cual explicaría la gran cantidad de proteína

38
expresada de 71 kDa con respecto a la de 40 kDa observada en los lisados de células
MT-2 (Figuras 6 y 8). Estos resultados sugieren que la especie de 71 kDa presente en
los lisados de células MT-2 corresponde a una proteína de fusión glicosilada de Tax y
gp21, la cual no se encuentra modificada con proteínas pequeñas.
Para confirmar la presencia de Tax en la banda a 71 kDa, ésta se inmunoprecipitó
empleando anticuerpos monoclonales (Covalab, Villeurbanne, France) a partir de
lisados totales de células MT-2, se concentró y proteolisó con tripsina para
posteriormente analizar los péptidos trípticos mediante espectrometría de masas (Figura
9). En la Figura 9a se observa un gel de SDS-PAGE teñido con Azul de Coomassie G-
250 donde se muestra el producto de la inmunoprecipitación obtenido a partir de lisados
de células MT-2, y que corresponde a una banda de proteínas de aproximadamente 71
kDa, la cuál es reconocida por anticuerpos monoclonales contra Tax (Figura 4b). La
banda se cortó del gel y las proteínas en ella fueron sometidas a una proteólisis en gel
utilizando tripsina como proteasa, y iodoacetamida como agente alquilante.
Posteriormente se obtuvieron los espectros de masas mediante un equipo MALDI-
TOF (Figura 9c). Se compararon los patrones peptídicos obtenidos experimentalmente
con distintas bases de datos utilizando la herramienta “on-line” www.matrixscience.com
encontrándose que un gran número de señales correspondían a albúmina de ratón. Estos
resultados indicaron que la muestra estaba contaminada con esta proteína proveniente
del líquido ascítico rico en anticuerpos contra Tax que fue utilizado para
inmunoprecipitar la proteína. Por esta razón, se procedió a eliminar del espectro las
señales que coinciden con las masas teóricas de los péptidos trípticos para albúmina de
ratón. Los péptidos experimentales restantes se compararon con los deducidos de la
proteólisis in silico para Tax, utilizando el programa “on-line” PeptideMap (PROWL
The Rockefeller University, http://prowl.rockefeller.edu). Al comparar los datos con
una tolerancia de 0.1 Da se encontró un péptido experimental que coincide con la masa
teórica de un péptido tríptico para Tax, el cual se representa en las Figuras 9c y 9d
(secuencia de color rojo). Al emplear el mismo rango de tolerancia, no se encuentran
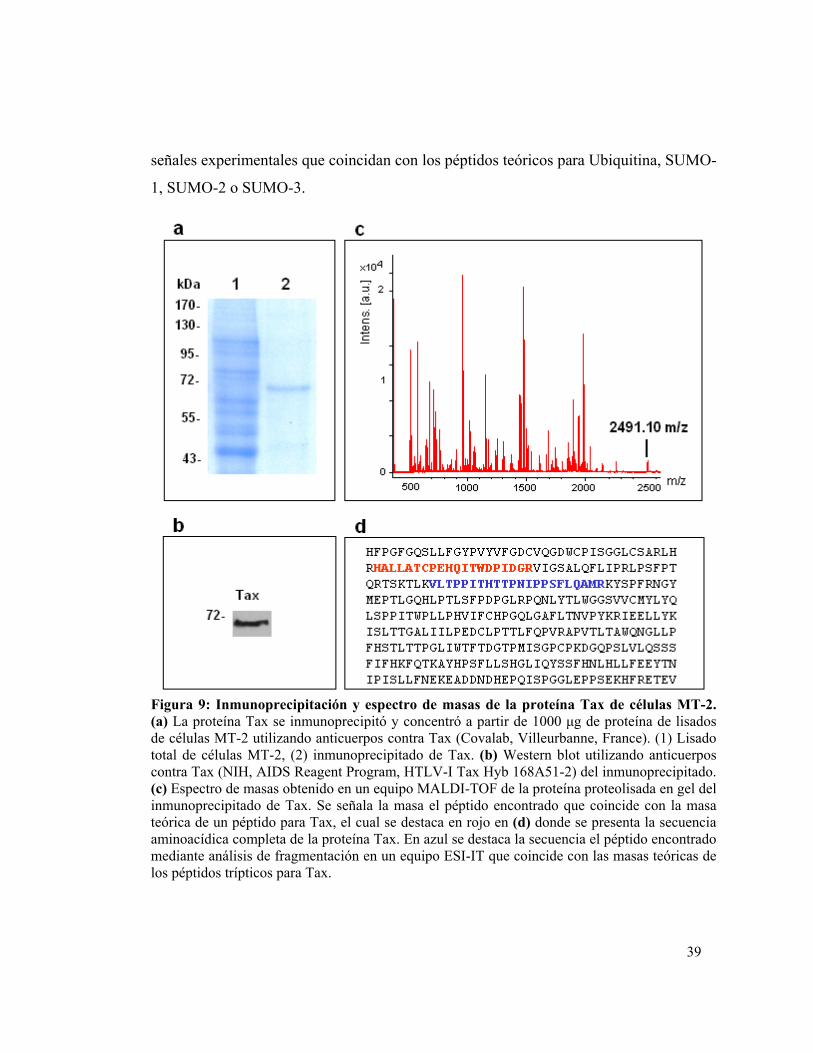
39
señales experimentales que coincidan con los péptidos teóricos para Ubiquitina, SUMO-
1, SUMO-2 o SUMO-3.
Figura 9: Inmunoprecipitación y espectro de masas de la proteína Tax de células MT-2. (a) La proteína Tax se inmunoprecipitó y concentró a partir de 1000 μg de proteína de lisados de células MT-2 utilizando anticuerpos contra Tax (Covalab, Villeurbanne, France). (1) Lisado total de células MT-2, (2) inmunoprecipitado de Tax. (b) Western blot utilizando anticuerpos contra Tax (NIH, AIDS Reagent Program, HTLV-I Tax Hyb 168A51-2) del inmunoprecipitado. (c) Espectro de masas obtenido en un equipo MALDI-TOF de la proteína proteolisada en gel del inmunoprecipitado de Tax. Se señala la masa el péptido encontrado que coincide con la masa teórica de un péptido para Tax, el cual se destaca en rojo en (d) donde se presenta la secuencia aminoacídica completa de la proteína Tax. En azul se destaca la secuencia el péptido encontrado mediante análisis de fragmentación en un equipo ESI-IT que coincide con las masas teóricas de los péptidos trípticos para Tax.

40
La misma muestra analizada mediante un equipo de espectrometría de masas HPLC-
ESI-IT (el cual permite hacer un análisis de fragmentación de los péptidos detectados),
arrojó la presencia de un péptido para Tax que coincide con la fragmentación teórica, el
cual se representa en la Figura 9d (secuencia de color azul). Para coincidir con las
masas teóricas, ambos péptidos deben estar fosforilados. El primero, encontrado por
MALDI-TOF (destacado con rojo en la Figura 9d) debe poseer 2 grupos fosfatos en
alguno de sus residuos de Ser o Thr, mientras el que segundo péptido (ilustrado en azul
en la Figura 9d) debe poseer grupos fosfato en T9, T10 y S16 (considerando el primer
residuo del péptido como el aminoácido 1).
En la misma muestra se encontró también mediante HPLC-ESI-IT dos péptidos que
coinciden con las fragmentaciones teóricas para péptidos trípticos originados de la
proteína gp21. Estos comprenden los péptidos entre los aminoácidos 145-169 y 173-192
de la proteína gp21 (datos no mostrados).
Estos resultados concuerdan con que en la banda de 71 kDa debe encontrarse la
proteína Tax como una quimera con gp21, la cual no se encuentra modificada con las
proteínas Ubiquitina ni SUMO.
Puesto que la proteína viral Tax se ha encontrado preferentemente en fracciones
dentro de las cuales podrían corresponder a las vías formales de secreción de proteínas
al medio extracelular, también se evaluó la presencia de Tax en el medio de cultivo
utilizado en las líneas celulares MT-2 y K562 (Figura 10). Se encontraron dos especies
de Tax de 60 y 65 kDa, la primera banda presenta inmunorreactividad al utilizar
anticuerpos contra SUMO-2/3. El papel fisiológico de esta modificación en una proteína
secretada es desconocido, siendo éste el primer reporte de esta naturaleza. En estas
muestras de medio de cultivo tampoco se detectó la presencia de Ubiquitina.

41
Figura 10: Identificación de Tax y sus modificaciones postraduccionales en los medios de cultivos de células MT-2. Western blot de los medios de cultivos de células MT-2 (1) y K562 (2) contra Tax (NIH, AIDS Reagent Program, HTLV-I Tax Hyb 168A51-2), Ubiquitina (Upstate, Lake Placid, NY, USA), SUMO-1 (Upstate, Lake Placid, NY, USA) y SUMO-2/3 (Abcam Inc, Cambridge, MA, USA). A la izquierda se representa el peso molecular aparente en kDa.
5.2. Cuantificar los niveles de Calreticulina y establecer su interacción con Tax en
células MT-2
Además de las modificaciones postraduccionales de Tax con proteínas pequeñas,
existen otras formas por la cual la localización subcelular de Tax es regulada dentro de
la célula. Una de ellas sugiere que la proteína Calreticulina actúa como receptor de
exportación nuclear de Tax, promoviendo la localización citoplasmática de la proteína
(Alefantis y cols., 2007), influyendo entonces en la regulación y progresión de las
patologías asociadas al virus. Es por esta razón que se busca identificar una interacción
entre Tax y CRT en linfocitos infectados con HTLV-I (células MT-2), y comparar los
niveles de CRT en células infectadas respecto a células no infectadas.
En la Figura 11 se muestran los niveles relativos de CRT obtenidos mediante
western blot en muestras de lisados celulares (Figura 11a) y medios de cultivo (Figura
11b) tanto de células MT-2 como K-562. No se encontraron diferencias significativas
en los niveles de la proteína intracelular, pero si se observó un aumento significativo en
la banda de CRT a 75 kDa encontrada en el medio de cultivo.

42
Figura 11: Cuantificación de los niveles de CRT en células MT-2 y células K562. Western blot y cuantificación densitométrica de las bandas inmunorreactivas totales de CRT reveladas empleando anticuerpos policlonales contra CRT. (a) Lisados de las líneas celulares; (b) proteínas secretadas en el medio de cultivo, en donde se muestran dos revelados representativos de las mismas muestras que difieren en el tiempo de exposición. Al costado de las bandas se representa el peso molecular aparente en kDa. Los análisis se realizaron por triplicado utilizando el test Mann-Whitney y los resultados se expresan como promedio ± SEM. UA: Unidades arbitrarias obtenidas del análisis densitométrico expresadas en pixeles. *p<0,05.
Para determinar la participación de CRT en la localización subcelular de Tax, se
procedió a investigar la interacción de ambas en células MT-2 mediante ensayos de
colocalización por microscopía de fluorescencia confocal y co-inmunoprecipitación.
Los resultados de microscopía confocal indican colocalización de ambas proteínas en
el citoplasma de las células MT-2 y no en el núcleo (Figura 12).

43
Figura 12: Determinación de colocalización entre Tax y CRT en células MT-2. Inmunofluorescencia indirecta utilizando anticuerpos primarios monoclonales contra Tax y policlonales contra CRT. Los anticuerpos secundarios conjugados con FITC (verde) marcaron Tax, y Alexa Fluor 594 (rojo) marcó CRT. El núcleo se tiñó con TO-PRO (pseudo color azul). Las células se observaron en un microscopio de fluorescencia confocal. Las flechas blancas indican sitios representativos de co-localización de Tax y CRT observados al utilizar el programa computacional “ImageJ”.

44
De un total de 480 células MT-2 visualizadas, el 57 % expresa la proteína Tax, y en
un 42 % se observa colocalización de Tax con CRT. Esto concuerda con resultados
obtenidos por citometría de flujo, en donde un 50,3 % de las células MT-2 expresa la
proteína Tax (Figura 13).
Figura 13: Determinación de la expresión de Tax en células MT-2 por citometría de flujo. Se utilizaron anticuerpos contra Tax conjugados con Cy5 para marcar 2x106 células MT-2 y posteriormente se analizaron en un citómetro de flujo. El gráfico de puntos muestra las células analizadas, en donde se representa en el eje X la población positiva para Tax (un 50,3 % de la población total).
En la Figura 14 se observan los resultados obtenidos en los ensayos de co-
inmunoprecipitación donde al inmunoprecipitar con anticuerpos contra cualquiera de las
dos proteínas, siempre se observa la presencia de ambas (Tax de 71 kDa y CRT de 63
kDa). Esto confirma que la colocalización observada mediante microscopía de
fluorescencia confocal en la Figura 12 se debe a interacción entre ambas proteínas.
Figura 14: Determinación de la interacción entre Tax y CRT en células MT-2 mediante ensayos de co-inmunoprecipitación. Se inmunoprecipitaron (IP) las proteínas Tax (utilizando anticuerpos monoclonales contra Tax, Covalab) y CRT (empleando anticuerpos policlonales contra CRT) y se reveló mediante western blot (WB) la presencia de ambas en cada inmunoprecipitado. A la derecha se indican los pesos moleculares aparentes en kDa.

45
5.3. Analizar las modificaciones postraduccionales de Tax proveniente de PBMC
aislados desde sangre periférica de pacientes HAM/TSP
La progresión de HAM/TSP en los pacientes infectados con HTLV-I no se ha podido
relacionar con la carga proviral ni con las cantidades de Tax expresadas en los linfocitos
infectados (Ramírez y cols., 2007). Sin embargo, las modificaciones postraduccionales
y el efecto que éstas ejercen sobre la localización intracelular de Tax podrían explicar
las diferencias en la progresión de HAM/TSP observadas en los pacientes. Para
establecer la severidad de la enfermedad, se identifica el estado de la marcha del
paciente en una escala de 1 a 5 (niveles de marcha), el cual corresponde al parámetro a
identificar y relacionar con las modificaciones de Tax en este estudio. Estos niveles
corresponden a: (1) marcha sin defecto, síndrome piramidal; (2) marcha con
movimientos espásticos sin necesidad de apoyo al caminar (a: eficiente; b: ineficiente);
(3) marcha con movimientos espásticos con necesidad de apoyo al caminar, (a:
eficiente; b: ineficiente); (4) capaz de estar de pie, sin caminar: (5) paciente postrado (a:
incapaz de mantenerse sólo; b: discapacidad global).
Para relacionar las modificaciones postraduccionales de Tax con el nivel de marcha
de pacientes HAM/TSP, se aislaron los PBMC (mediante centrifugación en gradiente de
Ficoll-Hypaque de 14 pacientes con distintos grados de marcha, 3 portadores
asintomáticos del virus y 6 individuos controles. Se cultivaron in vitro 2x106 células
durante 24 h en medio RPMI 1640 suplementado con 10 % de suero fetal bovino, se
lisaron, y mediante western blot se determinó la presencia de Tax en los lisados
celulares. No fue posible detectar la presencia de Tax en estos lisados, posiblemente
debido a que la concentración celular de la proteína es muy baja para la sensibilidad de
la técnica. Se intentó concentrando las muestras (dentro de la disponibilidad de ellas),
pero aún así no se obtuvieron resultados reproducibles.
Sin embargo, al emplear citometría de flujo como técnica alternativa para detectar
Tax en PBMC de los pacientes, se pudo establecer la presencia de la proteína viral en

46
estas células (Figura 15 y Tabla II). La Tabla II muestra los resultados de citometría
de flujo expresados como porcentaje de linfocitos T CD4+/Tax+, tal como se ha
descrito en la literatura (Hanon y cols., 2000). También muestra la presencia de Tax
determinada como Unidades Arbitrarias (UA) a partir de los datos densitométricos del
western blot contra Tax en el medio de cultivo, la seropositividad contra antígenos
virales, los datos clínicos sobre severidad de la enfermedad expresado con el grado de
marcha junto con los años de evolución de la enfermedad.
Para la mayoría de los casos, se encontró que menos de un 2 % de la población de
linfocitos T CD4+ en los PBMC expresó la proteína Tax. La muestra que presentó una
mayor cantidad de Tax expresada en linfocitos T CD4+, sólo registró un 4,9 % de la
población total. Esto resultados indican que en la muestra de lisados celulares de estos
PBMC, hay un bajo porcentaje de células que contribuye con la proteína Tax, de modo
que es concordante que para la mayoría de los casos se obtenga una muestra con muy
poca concentración de Tax, dificultando el análisis de la proteína mediante western blot.
Figura 15: Determinación de la expresión de Tax en células de pacientes HAM/TSP mediante citometría de flujo. Gráfico de puntos representativo del análisis de expresión de Tax en 2x106 PBMC de pacientes HAM/TSP incubados durante 24 h en medio RPMI 1640 suplementado con 10 % de suero fetal bovino. Las células se marcaron con anticuerpos contra Tax conjugado con Cy5 y anticuerpos contra CD4 conjugados con FITC. En el eje X se representa la población positiva para CD4 y en el eje Y la población positiva para Tax. El total de células doblemente positivas (CD4+/Tax+) corresponde a un 1,4 % de la población total.

47
Tabla II: Datos clínicos y resultados de los pacientes analizados mediante citometría de flujo (% linfocitos CD4+/Tax+) y western blot del medio de cultivo linfocitario (Unidades arbitrarias, UA) para la proteína Tax. Los grados de marcha comprenden: 2a: marcha eficiente con movimientos espásticos sin necesidad de apoyo al caminar; 2b: marcha ineficiente con movimientos espásticos sin necesidad de apoyo al caminar; 3a: marcha eficiente espástica con apoyo 4: en silla de ruedas pero capaz de mantenerse de pie.
Nº % Linfocitos Western Blot (UA Tax) Seropositividad HAM/TSP Años de muestra CD4+/Tax+ Medio de cultivo HTLV-I (Grado de marcha) evolución
1 0,9 39 + No - 2 0,9 34 + No - 3 0,1 0 + No - 4 2,1 15 + Si (3a) 10
5 4,9 100 + Si (2b) 3 6 3,2 4 + Si (2b) 2 7 2,4 29 + Si (2a) 20 8 2,8 3 + Si (2b) 5 9 1,4 28 + Si (2a) 4 10 1,1 23 + Si (2b) 12 11 1,7 24 + Si (2b) 5 12 0,2 125 + Si (2b) 14 13 1,8 83 + Si (3a) 3 14 0,2 42 + Si (3a) 10 15 1,1 103 + Si (4) 30 16 1,6 234 + Si (2b) 3 17 0,5 22 + Si (2b) 9 18 No detectado No detectado - No - 19 No detectado No detectado - No - 20 No detectado No detectado - No - 21 No detectado No detectado - No - 22 No detectado No detectado - No - 23 No detectado No detectado - No -
Lamentablemente, mediante citometría de flujo no puede establecerse la presencia de
modificaciones postraduccionales en la proteína Tax presente en las células estudiadas,
puesto que la señal de la modificación postraduccional en una célula dada no puede
vincularse con la proteína Tax.
Como Tax es una proteína viral secretable, como se confirmó con las células MT-2,
se estudió además su presencia en los medios de cultivo de los PBMC. El análisis de la
presencia de Tax mediante western blot de los medios de cultivo de PBMC mostró una

48
banda con un peso molecular aparente de aproximadamente 55 kDa inmunorreactiva
para Tax al utilizar dos anticuerpos monoclonales diferentes (Covalab, Villeurbanne,
France y NIH, AIDS Reagent Program, HTLV-I Tax Hyb 168A51-2) (Figura 16a y
Tabla II). La identificación de Tax en estos medios de cultivo permitió el estudio de las
modificaciones postraduccionales en ella. Se observó al mismo peso molecular de 55
kDa inmunorreactividad para Ubiquina, SUMO-1 y SUMO-2/3 (Figura 16a). Esta
forma de Tax a 55 kDa probablemente se encuentra modificada por proteínas pequeñas
como SUMO o Ubiquitina.
Al cuantificar las bandas obtenidas en el Western Blot, no se encontraron diferencias
significativas entre las razones de cada tipo de modificación postraduccional/proteína
Tax total al comparar los distintos grados de marcha de los pacientes HAM/TSP
(Figura 16b). Tampoco se observó algún tipo de correlación entre la severidad de la
enfermedad y las cantidades de Tax o las modificaciones postraduccionales de la
proteína viral secretada in vivo.
Para corroborar la presencia de estas modificaciones postraduccionales en la proteína
Tax secretada al medio de cultivo de los PBMC, se inmunoprecipitó Tax a partir del
medio de cultivo del paciente número 6 indicado en la Tabla II, y se comprobó la
presencia de modificaciones en la proteína inmunoprecipitada mediante western blot
(Figura 17). Tax inmunoprecipitada esta vez presenta un peso molecular aparente de 58
kDa. El aumento en el peso molecular se debe a que la muestra no presenta proteínas
séricas de los medios de cultivos de PBMC provenientes del suero fetal bovino, las
cuales ejercen un efecto de empuje de las proteínas de menor peso molecular a éstas, ya
que al añadir suero al inmunoprecipitado y revelar Tax se observa nuevamente la banda
a 55 kDa (datos no mostrados).
Se detectó inmunorreactividad para Ubiquitina, pero no de SUMO-1 y SUMO-2/3
para la banda de 58 kDa correspondiente a Tax inmunoprecipitada. El peso molecular
de 58 kDa coincide con el peso molecular teórico de Tax (40 kDa) más dos unidades de

49
Ubiquitina (8,5 kDa cada una). Estos resultados indican que la proteína Tax
inmunoprecipitada del medio de cultivo del paciente número 6 se encuentra modificada
con dos unidades de Ubiquitina, y no se encuentra modificada con SUMO.
Figura 16: Identificación y cuantificación de Tax y sus modificaciones en medios de cultivo de PBMC de pacientes infectados con HTLV-I. (a) Western blot representativos contra Tax, Ubiquitina, SUMO-1 y SUMO-2/3 de individuos controles, portadores asintomáticos y pacientes HAM/TSP. En la parte superior se indica el paciente analizado según la Tabla II, y en la parte inferior se indica el grado de marcha del paciente o si corresponde a un control no infectado (C) o bien a un portador asintomático (A). A la izquierda se representa el peso molecular aparente en kDa y la proteína revelada en cada caso. (b) Cuantificación densitométrica de las bandas a 55 kDa obtenidas en los western blot. Cada gráfico expresa una razón entre los pixeles de la modificación postraduccional respectiva y Tax total. Se compararon los resultados de los distintos grupos analizados según su grado de marcha. Los análisis se realizaron utilizando el test de Mann-Whitney y los resultados se expresan como promedio ± SEM. UA: Unidades arbitrarias obtenidas del análisis densitométrico expresadas en pixeles.

50
Figura 17: Inmunoprecipitación de la proteína Tax de medio de cultivo de PBMC de un paciente HAM/TSP. Western blot contra Tax, Ubiquitina, SUMO-1 y SUMO-2/3 del medio de cultivo del paciente número 6 indicado en la Tabla I (1) y del inmunoprecipitado de Tax a partir de este medio de cultivo (2). Al costado de las bandas se indica el peso molecular aparente en kDa.
5.4. Cuantificar los niveles de Calreticulina y establecer su interacción con Tax
en PBMC aislados desde sangre periférica de pacientes HAM/TSP
Para relacionar los niveles de expresión de CRT con la progresión de HAM/TSP, y
en particular su relación con la proteína Tax, mediante Western Blot se cuantificaron las
cantidades de ésta proveniente de los mismos medios de cultivos de PBMC aislados de
pacientes con distintos niveles de marcha, de portadores asintomáticos y de individuos
controles que se enumeran en la Tabla II (Figura 18). Los resultados indican la
presencia de dos bandas, una que abarca desde 87 hasta 70 kDa (banda gruesa,
característica de proteínas glicosiladas o proteínas con distintos grados de degradación),
y una banda discreta a 56 kDa. Los resultados al comparar las cantidades relativas de
cada especie con los niveles de marcha de los pacientes HAM/TSP indican que no
existen diferencias significativas. Sólo existen diferencias significativas entre los
individuos controles y los portadores asintomáticos del virus para la banda a 87-70 kDa
(p<0,05), y entre los controles y todos los demás grupos (portadores asintomáticos y
pacientes HAM/TSP) para la banda a 56 kDa (p<0,01), encontrándose un aumento
significativo de la especie de CRT a 56 kDa en individuos infectados por el virus.

51
Figura 18: Identificación de CRT en medios de cultivo de PBMC aislados de pacientes con HAM/TSP. (a) Western blot representativo contra CRT de muestras de pacientes controles, portadores asintomáticos y pacientes con diversos grados de marcha. Los números en la parte superior indican el paciente analizado según la Tabla II, y en la parte inferior se indica el grado de marcha del paciente, si corresponde a una muestra control no infectado por el virus (C) o un portador asintomático del virus (A). A la izquierda se representa el peso molecular aparente en kDa. (b) cuantificación de las bandas obtenidas por western blot. Cada gráfico expresa la banda de CRT analizada, y en ellos se comparan los distintos niveles de ella según las características clínicas de los pacientes. *p<0,05. **p<0,01. Los análisis se realizaron utilizando el test Mann-Whitney y los resultados se expresan como promedio ± SEM. UA: Unidades arbitrarias obtenidas del análisis densitométrico expresadas en pixeles.
Es importante hacer notar que las cantidades de la banda de CRT de 56 kDa presenta
una relación directa con las cantidades de la proteína viral Tax secretada en los medios

52
de cultivo de los PBMC de pacientes infectados con el virus, independiente de la
situación clínica de cada paciente (Figura 19). Es decir, mientras mayor es la cantidad
de Tax secretada al medio de cultivo, mayor es la cantidad de CRT de 56 kDa secretada
al mismo medio. Este hecho puede deberse a la función de chaperona de CRT y podría
estar involucrado directamente con la secreción de Tax, lo cual repercutiría en el daño al
sistema nervioso central observado en los pacientes HAM/TSP. Este corresponde al
primer reporte en donde se relacionan las cantidades de CRT con las cantidades de Tax
en los medios de cultivos de PBMC obtenidos de pacientes HAM/TSP.
Figura 19: Correlación entre las cantidades de Tax extracelular y la forma de CRT extracelular de 56 kDa en pacientes HAM/TSP. Gráfico de las Unidades Arbitrarias (UA) de Tax versus UA de la banda a 56 kDa de CRT expresadas en pixeles. La relación encontrada es significativa con un coeficiente de correlación de Spearman de 0,012.
Para determinar la interacción entre CRT y Tax en los linfocitos de pacientes
infectados con HTLV-I mediante microscopía confocal, se obtuvieron los PBMC de un
paciente con HAM/TSP (muestra número 17 en la Tabla II), se aisló la fracción de
linfocitos T CD4+ mediante esferas inmunomagnéticas, y se observó colocalización de
ambas proteínas mediante microscopía confocal (Figura 20). De aproximadamente 500
células íntegras observadas, se identificó la presencia de una célula que expresa la

53
proteína Tax en zonas definidas dentro del núcleo, en donde parte de la proteína Tax
interacciona con CRT en estas zonas.
Figura 20: Determinación de la interacción
entre Tax y CRT en Linfocitos T CD4+
mediante microscopía confocal.
Inmunofluorescencia indirecta utilizando
anticuerpos monoclonales contra Tax y
anticuerpos policlonales contra CRT en
Linfocitos T CD4+ aislados mediante esferas
inmunomagnéticas desde PBMC de un
paciente HAM/TSP. Como anticuerpos
secundarios se emplearon conjugados con el
fluoróforo FITC (verde), para marcar el
anticuerpo contra Tax, y conjugados con
Alexa 594 (rojo) para marcar el anticuerpo
contra CRT. El núcleo se tiñó con el marcador
nuclear TO-PRO (pseudo color azul). A la
izquierda se observa una célula que expresa
Tax, mientras que en el mismo campo, a la
derecha, una célula que no la expresa. El
núcleo se tiñó con el marcador nuclear TO-
PRO. La flecha blanca indica sitios
representativos de colocalización entre Tax y
CRT en la célula que expresa la proteína Tax
observada al utilizar el programa “ImageJ”.

54
6. DISCUSION
La proteína Tax hoy en día es el principal factor para comprender los mecanismos
de patogénesis y sobrevida del HTLV-I. Su papel multifuncional la cataloga en diversos
aspectos dentro y fuera de la célula, lo cual la transforma en la proteína más estudiada
del virus (Boxus y cols., 2008) e incluso en un blanco de interés terapéutico en las
patologías asociadas a éste (Boross y cols., 2009; Egbaria y cols., 2009). Sin embargo,
muchas de las funciones de esta proteína aún no se conocen.
Independiente de las hipótesis empleadas para explicar el desarrollo de HAM/TSP
en individuos infectados por el virus, la proteína Tax siempre se presenta como el actor
principal, fundamentalmente por la importancia de ésta en la sobrevida del virus (Boxus
y Willems, 2009). La presencia de provirus defectuosos que contienen sólo copias de la
región pX que codifica para Tax en pacientes HAM/TSP, pone en evidencia la
importancia de Tax en dicha patología (Ramírez y cols., 2003). No obstante, los
mecanismos por los cuales esta proteína origina HAM/TSP sigue siendo tema
controversial.
Recientemente se han descrito importantes resultados de ubiquitinación y
sumoilación de Tax en células 293T y HeLa transfectadas con vectores de expresión
para Tax, Ubiquitina y SUMO-1 (Peloponese y cols., 2004; Chiari y cols., 2004;
Lamsoul y cols., 2005; Nasr y cols., 2006), y la interacción entre Tax y CRT en células
BHK-21 transfectadas con vectores de expresión para ambas proteínas (Alefantis y
cols., 2007). Estos hallazgos abren un nuevo campo de estudio sobre los mecanismos
que regulan la funcionalidad de Tax, particularmente en el contexto de la localización
subcelular de esta proteína, y por ende, de la enfermedad HAM/TSP.
El presente trabajo corresponde al primero en donde se relaciona la ubiquitinación,
sumoilación e interacción con CRT de Tax con la evolución de HAM/TSP. La dificultad
de estos estudios radica en la baja expresión de Tax en linfocitos aislados de pacientes

55
con la enfermedad, debido a la estricta regulación de la expresión de proteínas virales
como mecanismo de escape inmunológico (Matsuoka 2003; Yoshida 2005). Como
resultado de ello, se han desarrollado estrategias que permiten incrementar la expresión
de Tax in vitro en linfocitos aislados de pacientes (Hanon y cols., 2000). A pesar de
lograr incrementar la cantidad de Tax intracelular sintetizada en PBMC mediante estas
metodologías, su expresión no logra ser suficiente para detectarla mediante western
blot. Esta última técnica es la adecuada para establecer un peso molecular aparente de la
proteína y vincular distintas modificaciones postraduccionales a la proteína en cuestión.
La actual relevancia que empieza a adquirir Tax en el medio extracelular y sus
implicancias en HAM/TSP (Alefantis y cols., 2005b; Alefantis y cols., 2007) condujo,
en la presente tesis, a analizar mediante western blot la presencia de Tax en los medios
de cultivo de PBMC de pacientes HAM/TSP. La presencia de Tax en el medio y no en
los lisados celulares indica que la proteína preferentemente se secreta desde los
linfocitos de pacientes infectados (Figura 16). Esta vez, la identificación de Tax
mediante western blot permitió establecer, por vez primera, la presencia de Ubiquitina
conjugada a Tax proveniente de linfocitos de pacientes infectados (Figura 17). Este
corresponde al primer reporte en donde se identifica mediante western blot la proteína
viral Tax proveniente de PBMC de pacientes con HAM/TSP. La presencia de proteína
Tax ubiquitinada secretada al medio de cultivo podría tener una estrecha vinculación
con el papel patológico que podría estar ocasionando en el desarrollo de HAM/TSP.
Para estudiar esta vinculación se identificó Tax ubiquitinada de pacientes HAM/TSP
con distintos grados de marcha. Se esperaba obtener una relación entre la severidad de
la enfermedad (expresada según los grados de marcha) y las cantidades de Tax
modificada, pero no se encontró una relación significativa como la planteada en la
hipótesis de este trabajo, de tal forma que ésta fue rechazada: “no existe relación entre el
nivel funcional de los pacientes y las modificaciones postraduccionales de Tax”.
La búsqueda de modificaciones postraduccionales en Tax como mecanismo de
regulación y consecuente participación en la patogénesis de las enfermedades asociadas

56
al HTLV-I ha sido realizada por distintos grupos de investigación al observar especies
inmunorreactivas para Tax a un peso molecular superior a 40 kDa en líneas celulares
infectadas con el virus (Peloponese y cols., 2004; Chiari y cols., 2004; Lamsoul y cols.,
2005; Nars y cols., 2006). Sin embargo, el estudio de Tax a 71 kDa en células MT-2
realizado en el presente trabajo requiere de una explicación distinta, puesto que se
encontró que esta especie no se encuentra modificada por las proteínas pequeñas
estudiadas (Figura 7). Si bien no se comprobó la existencia de la proteína quimera
gp21-Tax en células MT-2 que explique el peso molecular observado (Miwa y cols.,
1984; Jeang y cols., 1997), este trabajo entrega nuevas evidencias que apoyan esta teoría
mediante los ensayos realizados para detectar N-glicosilaciones y los análisis por
espectrometría de masas (Figuras 8 y 9). El estudio de Tax de 40 kDa se dificulta al
utilizar este modelo debido a la presencia más abundante de la proteína de fusión. Sin
embargo, utilizando este modelo se obtiene un dato fundamental sobre el interactoma de
Tax: su interacción con CRT.
Si bien se ha estudiado con anterioridad la interacción entre Tax y CRT (Alefantis y
cols., 2007), estos trabajos son poco claros, poco rigurosos, y han sido efectuados
utilizando como modelo la línea celular BHK-21 (no linfocitaria) transfectadas con
vectores de expresión para Tax y CRT. Los datos obtenidos en esta tesis apoyan la
noción de interacción entre estas dos proteínas, pero esta vez empleando un modelo más
cercano a la situación fisiológica (Figuras 12 y 14). Sin embargo, no fue posible
determinar con certeza la localización citoplasmática de la interacción, a diferencia del
trabajo de Alefantis y cols. (2007), en donde se observa que la interacción ocurre
principalmente los alrededores perinucleares. Esta localización sugiere a estos autores
postular que la razón de esta interacción se debe a la función como chaperona de
exportación nuclear de CRT. De esta forma, CRT promovería el transporte de Tax
desde el núcleo hacia el citoplasma. No obstante, aún se requieren estudios mayormente
detallados para comprender la naturaleza de esta interacción. Es probable que ésta
ocurra también en compartimientos del RE en donde CRT participa en el proceso de

57
plegamiento y control de calidad de las proteínas a secretarse (Michalak y cols., 1999).
De ahí la importancia de identificar la interacción entre Tax y CRT, ya que últimamente
se ha reportado la localización de la proteína Tax en compartimientos de las vías
canónicas de secreción dentro de la célula (RE y redes cis y trans del complejo de
Golgi) (Alefantis y cols., 2005b). También se han identificado las señales aminoacídicas
en Tax involucradas en la secreción de la proteína y la interacción de éstas con proteínas
componentes de la vía secretoria; entre ellas CRT como proteína del RE, βCOP y COP-
II como proteínas que permiten el tránsito desde el RE a la red cis del Aparato de Golgi,
y proteínas que permiten el movimiento desde trans-Golgi hacia la membrana
plasmática (SNAP23, SCAMP1 y SCAMP2) (Jain y cols., 2007). Toda esta evidencia
sugiere que Tax se secreta en forma controlada desde los linfocitos infectados, lo cual
puede adquirir especial relevancia en el desarrollo de HAM/TSP.
El presente estudio de CRT secretada en los medios de cultivos de PBMC aislados
de individuos infectados con HTLV-I complementa la información anterior. CRT
corresponde a una proteína originalmente descrita como residente del RE (Michalak y
cols., 1992), la cual posee una señal de retención reticular, y que por ende durante
varios años ha sido tema de controversia la presencia de CRT en lugares distintos al RE.
Sin embargo, últimamente se ha acumulado suficiente evidencia acerca de las funciones
fuera del RE de la CRT, y se ha llegado a aceptar la idea general de que CRT
corresponde más bien a una proteína multifuncional que puede ejercer diversas acciones
dependiendo de la localización donde se encuentre dentro y fuera de la célula (Gold y
cols., 2010). En el medio extracelular, CRT se describe como una proteína con
funciones muy diversas, las cuales varían desde promover la curación de heridas al
favorecer la migración de keratinocitos, fagocitosis y fibrinogénesis (Gold y cols., 2006)
hasta participar en la promoción de la respuesta inmune contra células cancerígenas, en
un proceso llamado muerte celular inmunogénica (Garg y cols., 2010). De hecho, se ha
estudiado un aumento en la inmunogenicidad al administrar CRT como adyuvante en la
quimioterapia contra el cáncer (Hsieh y cols., 2004; Cheng y cols., 2005).

58
A pesar de toda la evidencia acerca de las funciones extracelulares de CRT, poco se
conoce sobre los mecanismos de secreción de esta proteína, lo cual ha sido tema de
intensa investigación en los últimos años (Gold y cols., 2010). Puesto que CRT posee en
su extremo carboxilo terminal una señal aminoacídica de retención reticular (KDEL), es
aún más difícil explicar cómo es que CRT sale del RE y es liberada al medio
extracelular. Al respecto se ha propuesto la idea de que la señal de retención KDEL es
proteolisada, o bien enmascarada por la interacción de CRT con alguna otra proteína o
que se libere en respuesta a cambios en el pH o los niveles de calcio en el RE (Corbett y
cols., 2000; Gold y cols., 2010). Más si este fuera el caso, aún queda sin explicar cómo
ocurre la secreción de CRT al medio extracelular. Esta secreción puede ocurrir
principalmente por dos vías: secreción activa o por muerte celular.
Existe evidencia que sugiere una liberación de CRT durante la necrosis celular, la
cual podría corresponder a un mecanismo de liberación fisiológico durante una herida,
en donde CRT ejerce una función en el posterior proceso de reparación en el medio
extracelular (Basu y cols., 2000). Sin embargo, los mecanismos de secreción activa de
CRT desde las células plantea más interrogantes, puesto que esta proteína no posee las
señales clásicas de secreción necesarias para interactuar con los componentes presentes
en las vías secretorias. Estas señales incluyen el transporte desde el RE hasta cis-Golgi y
desde tras-Golgi hasta la fusión con la membrana plasmática y consecuente liberación
de los componentes vesiculares al medio extracelular. Debido a esto, se ha propuesto
que CRT participa en otra forma de secreción no clásica correspondiente a la formación
de exosomas, los cuales participan en la liberación al medio extracelular de proteínas
citosólicas (Février y Raposo, 2004). De esta manera CRT dependería de su capacidad
para retrotrasladarse desde el RE hasta el citoplasma (Afshar y cols., 2005). Sin
embargo, también se ha encontrado en numerosos reportes la presencia de CRT como
proteína transmembrana expuesta al medio extracelular (Madeo y cols., 2009), y además
la presencia de CRT en el complejo de Golgi, en donde se especula que posteriormente
se envía a la superficie celular por asociación con otras proteínas como α-integrinas y

59
moléculas del complejo MHC-I (Coopolino y cols., 1997; Howe y cols., 2009). Esto
sugiere que CRT también se secreta por la ruta clásica, aunque el mecanismo de tránsito
por la vía secretora sigue siendo un enigma (Gold y cols., 2010).
Los datos expuestos en la presente tesis sugieren un posible mecanismo por el cual
CRT es liberada al medio extracelular transitando por la vía clásica de secreción. Esta
proposición se basa, por una parte, en que los niveles extracelulares de la forma de CRT
de 56 kDa están significativamente aumentados en los medios de cultivo de PBMC de
individuos infectados por HTLV-I, independiente de la situación clínica en la que se
encuentren (Figura 18). Por otra parte, se observa una relación directa entre las
cantidades de esta especie de CRT y las cantidades de Tax en el medio de cultivo de
PBMC (Figura 19). Esto en su conjunto, constituye una fuerte evidencia de que por
efecto de la proteína viral Tax se secreta una mayor cantidad de CRT en las células
infectadas por el virus. Probablemente esta relación directa se deba a que CRT se co-
secreta con Tax desde las células infectadas, lo cual puede proceder por la interacción
entre Tax y CRT en los compartimientos de la vía secretora clásica. Es probable que
esta interacción enmascare la señal KDEL de retención reticular de CRT en el RE y que
posteriormente CRT acompañe el tránsito de secreción de Tax, la cual posee las señales
de interacción con los componentes de la vía formal de secreción que permiten el
tránsito desde RE hacia Golgi y a la membrana plasmática (Alefantis y cols., 2005b;
Jain y cols., 2007). Este constituiría un nuevo mecanismo por el cual CRT puede
liberarse de las células al medio extracelular, esta vez mediado gracias a la interacción
con una proteína viral.
Las razones por las cuales CRT es liberada, en este caso, pueden ser muy diversas y
por sobretodo aumentan las preguntas sobre los mecanismos de patogénesis y sobrevida
del HTLV-I. Se ha demostrado la habilidad de ciertos patógenos de “engañar” el
sistema inmune del hospedero, no sólo en evadir sus mecanismos sino que también
utilizarlos para favorecer su patogenicidad y sobrevida (Ferreira y cols., 2005; Mulhern
y cols., 2009; Diacovich y Gorvel, 2010). Al respecto, el HTLV-I no es la excepción.

60
Puesto que se considera un organismo experto en la evasión del sistema inmune y
sobrevida, el HTLV-I es capaz de mantener un equilibrio con su hospedero, hasta el
punto que la mayoría de los individuos infectados no presentan ninguna patología
clínica (sobre un 90 %), y de presentarse, ésta requiere de un período de décadas para
manifestarse (Matsuoka y Jeang, 2007). Esta observación concuerda con la estimación
de que HTLV-I corresponde a un patógeno ancestral que infectaba a los seres humanos
desde hace más de 1500 años, luego de encontrarse segmentos genómicos del provirus
en el DNA de momias chilenas (Li y cols., 1999). De hecho, los fragmentos genéticos
encontrados (correspondientes a la región pX) son similares a los de hoy en día en
individuos infectados de Japón y Chile, sugiriendo una conservación genética del virus
de hace más de 1500 años.
Esto indica que el HTLV-I ha evolucionado desde entonces, y probablemente
mucho antes, razón por la que se encuentra totalmente adaptado al “ambiente humano”,
para lo cual utiliza diversas y sofisticadas estrategias que aún no se conocen. Una de
aquellas estrategias podría corresponder al aumento en la secreción de CRT mediada
por Tax en los linfocitos infectados por el virus. Debido a las múltiples funciones que
ejerce la proteína Tax para facilitar la sobrevida del virus, no es sorprendente que el
virus emplee esta proteína multifuncional para favorecer la secreción de CRT, la cual a
su vez tendrá efectos benéficos sobre la salud del individuo infectado. Con esto, el virus
consigue aumentar la sobrevida del hospedero, lo que a su vez aumenta su propia
sobrevida. Se ha postulado que ciertos patógenos utilizan la exposición al medio
extracelular de CRT para promover la sobrevida del hospedero, como es el caso de
Trypanosoma cruzi, el agente etiológico de la enfermedad de Chagas (Molina y cols.,
2005). Se ha demostrado que la actividad antiangiogénica de CRT de T. cruzi, protege
al hospedero de la formación de tumores dependientes de vascularización, aumentando
la sobrevida de éste y por ende la del parásito. También hay trabajos que demuestran las
propiedades antiangiogénicas de CRT humana, dificultando el desarrollo de tumores

61
(Cheng y cols., 2001). Esto propone un papel protector de CRT liberada desde células
infectadas con HTLV-I (Figura 21).
Otra de las funciones de CRT en el medio extracelular corresponde a la unión del
péptido β-amiloide en líquido cefalorraquídeo, actuando como una proteína
transportadora que previene la agregación de depósitos αβ (Erickson y cols., 2005;
Duus y cols., 2008). Es interesante observar que en estudios de anatomía patológica de
individuos con HAM/TSP se han encontrado depósitos de proteína precursora de
amiloide en los segmentos dorso-lumbar y cervical de la médula espinal (Cartier y cols.,
2007). En este caso, la liberación de CRT fomentada por Tax en las células infectadas
podría evitar o retrasar la formación de agregados αβ (Figura 21). ¿Es posible que el
virus intente menguar sus efectos patológicos en individuos infectados al favorecer la
secreción de CRT? De utilizar un mecanismo similar, correspondería a uno altamente
sofisticado para favorecer su permanencia, sobrevida y equilibrio con el hospedero.
A futuro se requiere despejar estas dudas y dilucidar el mecanismo por el cual en
células infectadas por HTLV-I Tax y CRT se co-secretan durante el tránsito en conjunto
por las vías RE-Golgi-membrana plasmática, y el papel fisiológico de la ubiquitinación
de la proteína Tax secretada en el desarrollo de las enfermedades asociadas al HTLV-I.

62
Figura 21: Modelo propuesto de co-secreción y efectos extracelulares de Tax y CRT. La proteína Tax se exporta al citoplasma por acción de CRT. En el citoplasma podría trasladarse al RE donde interacciona con CRT. No es claro si Tax se ubiquitina en el citoplasma o en el RE. La presencia de señales en Tax que le permiten interactuar con la vía secretoria, permite también que CRT se secrete junto con ella. En el medio extracelular, Tax puede promover la evolución de HAM/TSP, mientras que CRT podría participar en mecanismos que prolonguen la vida del hospedero, y con ello contribuir a la sobrevida del HTLV-I.

63
7. CONCLUSIONES
- En la línea celular MT-2 existe una forma intracelular de Tax de 71 kDa, y en menor
proporción una de 40 kDa en las fracciones membranosa/organelar. De las
modificaciones postraduccionales estudiadas asociadas a la banda de Tax de 71 kDa no
se encontró la presencia de proteínas pequeñas Ubiquitina, SUMO-1 y SUMO-2/3, de
tal manera que la proteína Tax intracelular de células MT-2 no se encuentra modificada
por estas proteínas pequeñas.
- La especie de 71 kDa correspondiente a Tax se encuentra N-glicosilada. La
glicosilación puede atribuirse a la presencia de gp21 que podría estar fusionada con Tax.
- Tax secretada desde las células MT-2 se distribuye en dos bandas de 60 y 65 kDa,
mostrándose modificada con SUMO-2/3 la banda de 60 kDa.
- La proteína CRT se expresa en células MT-2 y K562 como una forma intracelular de
63 kDa y otra secretada al medio de cultivo de 81, 75, 67 y 55 kDa. Existe un aumento
significativo en los niveles de la especie a 75 kDa de CRT secretada al medio de cultivo
en células MT-2 comparado con células K562.
- CRT interacciona con Tax en el citoplasma de células MT-2.
- En pacientes con HAM/TSP y en individuos portadores, Tax se detecta en linfocitos T
CD4+ mediante citometría de flujo y en el medio de cultivo de los PBMC por western
blot en la forma de una proteína de 58 kDa que se encuentra ubiquitinada. El peso
molecular sugiere que Tax en el medio extracelular se encuentra modificada con dos
unidades de Ubiquitina.

64
- No se observan correlaciones entre la severidad de la enfermedad expresada con el
grado de marcha de los pacientes analizados y las cantidades de Tax intracelular o
extracelular, ni de sus formas modificadas, de tal manera que la hipótesis de este trabajo
se rechaza.
- CRT se expresa en PBMC en la forma de una banda gruesa de entre 87 a 70 kDa y una
banda discreta de 56 kDa. Esta última se encuentra significativamente elevada en
individuos infectados por el virus, independiente de la situación clínica en la cual se
encuentren.
- Al menos en una célula de una población de linfocitos T CD4+ de un paciente
HAM/TSP se observó colocalización entre Tax y CRT mediante microscopía confocal.
- Existe una relación directa entre los niveles de Tax y los niveles de CRT de 56 kDa en
el medio de cultivo de PBMC de individuos infectados por HTLV-I.

65
8. REFERENCIAS
Afshar N, Black BE, Paschal BM. (2005) Retrotranslocation of the chaperone calreticulin from the endoplasmic reticulum lumen to the cytosol. Mol Cell Biol. 25:8844-53.
Akagi T, Shimotohno K. (1993) Proliferative response of Tax1-transduced primary human T cells to anti-CD3 antibody stimulation by an interleukin-2-independent pathway. J Virol. 67:1211-1217.
Alberti C, Gonzalez J, Maldonado H, Medina F, Barriga A, García L, Kettlun AM, Collados L, Puente J, Cartier L, Valenzuela MA. (2009) Comparative study of CSF neurofilaments in HTLV-1-associated myelopathy/tropical spastic paraparesis and other neurological disorders. AIDS Res Hum Retroviruses. 25:803-9.
Alefantis T, Jain P, Ahuja J, Mostoller K, Wigdahl B. (2005a) HTLV-1 Tax nucleocytoplasmic shuttling, interaction with the secretory pathway, extracellular signaling, and implications for neurologic disease. J Biomed Sci.12:961-74.
Alefantis T, Mostoller K, Jain P, Harhaj E, Grant C, Wigdahl B. (2005b) Secretion of the human T cell leukemia virus type I transactivator protein tax. J Biol Chem. 280:17353-62.
Alefantis T, Flaig KE, Wigdahl B, Jain P. (2007) Interaction of HTLV-I Tax protein with calreticulin: implications for Tax nuclear export and secretion. Biomed Pharmacother. 61:194-200.
Barmak, Harhaj E, Grant C, Alefantis T, Wigdahl B. (2003) Human T cell leukemia virus type-I induced disease: pathways to cancer and neurodegeneration. Virology. 308:1-12.
Basu S, Binder RJ, Suto R, Anderson KM, Srivastava PK. (2000) Necrotic but not apoptotic cell death releases heat shock proteins, which deliver a partial maturation signal to dendritic cells and activate the NF-kappa B pathway. Int Immunol. 12:1539-46.
Boross P, Bagossi P, Weber IT, Tözsér J. (2009) Drug targets in human T-lymphotropic virus type 1 (HTLV-1) infection. Infect Disord Drug Targets. 9:159-71.
Boxus M, Twizere JC, Legros S, Dewulf JF, Kettmann R, Willems L (2008). The HTLV-I Tax interactome. Retrovirology. 5:76. (doi: 10.1186/1742-4690-5-76).
Boxus M, Willems L. (2009) Mechanisms of HTLV-1 persistence and transformation. Br J Cancer. 101:1497-501.

66
Brady J, Jeang KT, Duvall J, Khoury G. (1987) Identification of p40x-responsive regulatory sequences within the human T-cell leukemia virus type I long terminal repeat. J Virol. 61:2175-81.
Cartier L, Araya F, Castillo JL, Verdugo R, Mora C, Gajdusek DC, Gibbs CJ. (1990) HTLV-I retrovirus in Chile: study on 140 neurological patients. Rev Med Chil 118: 622-8.
Cartier L, Araya F, Castillo JL, Ruiz F, Gormaz A, Tajima K. (1992) Progressive spastic paraparesis associated with human T-cell leukemia virus type I (HTLV-I). Intern Med. 31:1257-61.
Cartier L., Cea JG, Vergara C., Araya F., Born P. (1997) Clinical and neuropathological study of six patients with spastic paraparesis associated with HTLV-I: an axomyelinic degeneration of the central nervous system. J Neuropathol Exp Neurol 56:403-13.
Cartier L, Ramírez E, Galeno H. (1999) HTLV-I Tax gene on the etiological identification of tropical spastic paraparesis. A clinical, serological and polymerase chain reaction (PCR) study in 72 patients. Rev Med Chil 127:945-52.
Cartier L, Ramírez E. (2005) Presence of HTLV-I Tax protein in cerebrospinal fluid from HAM/TSP patients. Arch Virol. 4:743-53.
Cartier L, Vergara R C, Valenzuela MA. (2007) Immunohistochemistry of degenerative changes in the central nervous system in spastic paraparesis associated to human T lymphotropic virus type I (HTLV-I). Rev Med Chil. 135:1139-46.
Cheng WF, Hung CF, Chai CY, Hsu KF, He L, Ling M, Wu TC. (2001) Tumor-specific immunity and antiangiogenesis generated by a DNA vaccine encoding calreticulin linked to a tumor antigen. J Clin Invest. 108:669-78.
Cheng WF, Hung CF, Chen CA, Lee CN, Su YN, Chai CY, Boyd DA, Hsieh CY, Wu TC. (2005) Characterization of DNA vaccines encoding the domains of calreticulin for their ability to elicit tumor-specific immunity and antiangiogenesis. Vaccine. 23:3864-74.
Chiari E, Lamsoul I, Lodewick J, Chopin C, Bex F, Pique C. (2004) Stable Ubiquination of Human T-Cell Leukemia Virus Type 1 Tax is Required for Proteasome Binding. J Virol. 78:11823-32.
Coppolino MG, Woodside MJ, Demaurex N, Grinstein S, St-Arnaud R, Dedhar S. (1997) Calreticulin is essential for integrin-mediated calcium signalling and cell adhesion. Nature. 386:843-7.

67
Corbett EF, Michalak KM, Oikawa K, Johnson S, Campbell ID, Eggleton P, Kay C, Michalak M. (2000) The conformation of calreticulin is influenced by the endoplasmic reticulum luminal environment. J Biol Chem. 275:27177-85.
Derse D, Heidecker G. (2003) Forced entry--or does HTLV-I have the key? Science. 299:1670-1.
Diacovich L, Gorvel JP. (2010) Bacterial manipulation of innate immunity to promote infection. Nat Rev Microbiol. 8:117-28.
Duus K, Hansen PR, Houen G. (2008) Interaction of calreticulin with amyloid beta peptide 1-42. Protein Pept Lett. 15:103-7.
Egbaria M, Tabakin-Fix Y, Huleihel M. (2009) Dominant negative Tax double mutants as molecular inhibitors for w.t. Tax gene functions. Leuk Res. 33:974-9.
Erickson RR, Dunning LM, Olson DA, Cohen SJ, Davis AT, Wood WG, Kratzke RA, Holtzman JL. (2005) In cerebrospinal fluid ER chaperones ERp57 and calreticulin bind beta-amyloid. Biochem Biophys Res Commun. 332:50-7.
Evans K, Keller C, Pavur K, Glasgow K, Conn B, Lauring B. (2006) Interaction of two hereditary spastic paraplegia gene products, spastin and atlastin, suggests a common pathway for axonal maintenance. Proc Natl Acad Sci USA.103:10666-71.
Franchini G. (1995) Molecular mechanisms of human T-cell leukemia/lymphotropic virus type I infection. Blood. 86:3619-39.
Ferreira V, Molina MC, Schwaeble W, Lemus D, Ferreira A. (2005) Does Trypanosoma cruzi calreticulin modulate the complement system and angiogenesis? Trends Parasitol. 21:169-74.
Février B, Raposo G. (2004) Exosomes: endosomal-derived vesicles shipping extracellular messages. Curr Opin Cell Biol. 16:415-21.
Gallo RC. (2005) The discovery of the first human retrovirus: HTLV-1 and HTLV-2. Retrovirology. 2:17.
Garg AD, Nowis D, Golab J, Vandenabeele P, Krysko DV, Agostinis P. (2010) Immunogenic cell death, DAMPs and anticancer therapeutics: an emerging amalgamation. Biochim Biophys Acta. 1805:53-71.
Gold LI, Rahman M, Blechman KM, Greives MR, Churgin S, Michaels J, Callaghan MJ, Cardwell NL, Pollins AC, Michalak M, Siebert JW, Levine JP, Gurtner GC, Nanney LB, Galiano RD, Cadacio CL. (2006) Overview of the role for calreticulin in the enhancement of wound healing through multiple biological effects. J Investig Dermatol Symp Proc. 11:57-65.

68
Gold LI, Eggleton P, Sweetwyne MT, Van Duyn LB, Greives MR, Naylor SM, Michalak M, Murphy-Ullrich JE. (2010) Calreticulin: non-endoplasmic reticulum functions in physiology and disease. FASEB J. 24:665-83.
Grassmann R, Fleckenstein B. (1989) Selectable recombinant herpesvirus saimiri is capable of persisting in a human Tcell line. J Virol 63:1818-21.
Grossman WJ, Kimata JT, Wong FH, Zutter M, Ley TJ, Ratner L. (1995) Development of leukemia in mice transgenic for the tax gene of human T-cell leukemia virus type I. Proc Natl Acad Sci USA. 92:1057-61.
Hanon E, Hall S, Taylor GP, Saito M, Davis R, Tanaka Y, Usuku K, Osame M, Weber JN, Bangham CR. (2000) Abundant tax protein expression in CD4+ T cells infected with human T-cell lymphotropic virus type I (HTLV-I) is prevented by cytotoxic T lymphocytes. Blood. 95:1386-92.
Hilgarth RS, Murphy LA, Skaggs HS, Wilkerson DC, Xing H, Sarge KD. (2004) Regulation and function of SUMO modification. J Biol Chem. 279:53899-902.
Holaska JM, Black BE, Love DC, Hanover JA, Leszyk J, Paschal BM. (2001) Calreticulin is a receptor for nuclear export. J Cell Biol.152:127-40.
Howe C, Garstka M, Al-Balushi M, Ghanem E, Antoniou AN, Fritzsche S, Jankevicius G, Kontouli N, Schneeweiss C, Williams A, Elliott T, Springer S. (2009) Calreticulin-dependent recycling in the early secretory pathway mediates optimal peptide loading of MHC class I molecules. EMBO J. 28:3730-44.
Hsieh CJ, Kim TW, Hung CF, Juang J, Moniz M, Boyd DA, He L, Chen PJ, Chen CH, Wu TC. (2004) Enhancement of vaccinia vaccine potency by linkage of tumor antigen gene to gene encoding calreticulin. Vaccine. 22:3993-4001.
Igakura T, Stinchcombe JC, Goon PK, Taylor GP, Weber JN, Griffiths GM, Tanaka Y, Osame M, Bangham CR. (2003) Spread of HTLV-I between lymphocytes by virus-induced polarization of the cytoskeleton. Science. 299: 1713-6.
Jain P, Mostoller K, Flaig KE, Ahuja J, Lepoutre V, Alefantis T, Khan ZK, Wigdahl B. (2007) Identification of human T cell leukemia virus type 1 tax amino acid signals and cellular factors involved in secretion of the viral oncoprotein. J Biol Chem. 282:34581-93.
Jeang KT, Derse D, Matocha M, Sharma O. (1997) Expression status of Tax protein in human T-cell leukemia virus type 1-transformed MT4 cells: recall of MT4 cells distributed by the NIH AIDS Research and Reference Reagent Program. J Virol. 71:6277-8.

69
Kerscher O, Felberbaum R, Hochstrasser M. (2006). Modification of proteins by ubiquitin and ubiquitin-like proteins. Annu Rev Cell Dev Biol. 22:159-180.
Kfoury Y, Nasr R, Favre-Bonvin A, El-Sabban M, Renault N, Giron ML, Setterblad N, Hajj HE, Chiari E, Mikati AG, Hermine O, Saib A, de Thé H, Pique C, Bazarbachi A. (2008) Ubiquitylated Tax targets and binds the IKK signalosome at the centrosome. Oncogene. 13:1665-76.
Kobayashi N, Konishi H, Sabe H, Shigesada K, Noma T, Honjo T, Hatanaka M. (1984) Genomic structure of HTLV (human T-cell leukemia virus): detection of defective genome and its amplification in MT-2 cells. EMBO J. 3:1339-43.
Laemmli UK. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 227:680-5.
Lamsoul I, Lodewick J, Lebrun S, Brasseur R, Burny A, Gaynor RB, Bex F. (2005) Exclusive Ubiquination and Sumoylation on Overlapping Lysine Residues Mediate NF-kB Activation by the Human T-Cell Leukemia Virus Tax Oncoprotein. Mol Cell Biol. 25:10391-406.
Lee SM, Morcos Y, Jang H, Stuart JM, Levin MC. (2005) HTLV-1 induced molecular mimicry in neurological disease. Curr Top Microbiol Immunol. 296:125-36.
Li HC, Fujiyoshi T, Lou H, Yashiki S, Sonoda S, Cartier L, Nunez L, Munoz I, Horai S, Tajima K. (1999) The presence of ancient human T-cell lymphotropic virus type I provirus DNA in an Andean mummy. Nat Med. 5:1428-32.
Madeo F, Durchschlag M, Kepp O, Panaretakis T, Zitvogel L, Fröhlich KU, Kroemer G. (2009) Phylogenetic conservation of the preapoptotic calreticulin exposure pathway from yeast to mammals. Cell Cycle. 8:639-42.
Maldonado H, Ortiz-Riaño E, Krause B, Barriga A, Medina F, Pando ME, Alberti C, Kettlun AM, Collados L, García L, Cartier L, Valenzuela MA. (2008) Microtubule proteins and their post-translationally forms in cerebrospinal fluid of patients with paraparesis associated to HTLV-I infection and in SH-SY5Y cells, an in vitro model of HTLV-I-induced disease. Biol Res. 41:239-59.
Marriott S, Semmes O. (2005) Impact of HTLV-I Tax on cell cycle progression and the cellular damage repair response. Oncogene. 24: 5986-95.
Matsuoka M. (2003) Human T-cell leukemia virus type I and adult T-cell leukemia. Oncogene. 22: 5131-40.
Matsuoka M, Jeang KT. (2007) Human T-cell leukaemia virus type 1 (HTLV-1) infectivity and cellular transformation. Nat Rev Cancer. 7:270-80.

70
Méndez E, Kawanishi T, Clemens K, Siomi H, Soldan S, Calabresi P, Brady J, Jacobson S. (1997) Astrocyte-Specific Expresión of Human T-Cell Lymphotropic Virus Type 1 (HTLV-1) Tax: Induction of Tumor Necrosis Factor Alpha and Susceptibility to Lysis by CD8+ HTLV-1- Specific Cytotoxic T Cells. J Virol. 71:9143-9149.
Michalak M, Milner RE, Burns K, Opas M. (1992) Calreticulin. Biochem J. 285:681-92.
Michalak M, Corbett EF, Mesaeli N, Nakamura K, Opas M. (1999) Calreticulin: one protein, one gene, many functions. Biochem J. 344 Pt 2:281-92.
Miwa M, Shimotohno K, Hoshino H, Fujino M, Sugimura T. (1984) Detection of pX proteins in human T-cell leukemia virus (HTLV)-infected cells by using antibody against peptide deduced from sequences of X-IV DNA of HTLV-I and Xc DNA of HTLV-II proviruses. Gann. 75:752-5.
Mogensen TH, Paludan S. (2001) Molecular pathways in virus-induced cytokine production. Microbiol Mol Biol Rev. 65: 131-50.
Molina MC, Ferreira V, Valck C, Aguilar L, Orellana J, Rojas A, Ramirez G, Billetta R, Schwaeble W, Lemus D, Ferreira A. (2005) An in vivo role for Trypanosoma cruzi calreticulin in antiangiogenesis. Mol Biochem Parasitol. 140:133-40.
Mulhern O, Harrington B, Bowie AG. (2009) Modulation of innate immune signalling pathways by viral proteins. Adv Exp Med Biol. 666:49-63.
Nagai M, Osame M. (2003) Human T-cell lymphotropic virus type I and neurological diseases. J Neurovirol. 9:228-35.
Nasr R, Chiari E, El-Sabban M, Mahieux R, Kfoury Y, Abdulhay M, Yazbeck V, Hermine O, de Thé H, Pique C, Bazarbachi A. (2006) Tax ubiquitylation and sumoylation control critical cytoplasmic and nuclear steps of NF-kB activation. Blood. 107: 4021-9.
Pais-Correia AM, Sachse M, Guadagnini S, Robbiati V, Lasserre R, Gessain A, Gout O, Alcover A, Thoulouze MI. (2010) Biofilm-like extracellular viral assemblies mediate HTLV-1 cell-to-cell transmission at virological synapses. Nat Med. 16:83-9.
Park SH, Zhu PP, Parker RL, Blackstone C. (2010) Hereditary spastic paraplegia proteins REEP1, spastim, and atlansin-1 coordinate microtubule interactions with the tubular ER network. J Clin Invest. 120:1097-110.
Paskalis H, Felber BK, Pavlakis GN. (1986) Cis-acting sequences responsible for the transcriptional activation of human T-cell leukemia virus type I constitute a conditional enhancer. Proc Natl Acad Sci USA. 83: 6558-62.

71
Peloponese JM Jr, Iha H, Yedavalli VR, Miyazato A, Li Y, Haller K, Benkirane M, Jeang KT. (2004) Ubiquination of human T-Cell leukemia virus type 1 Tax modulates its cctivity. J Virol. 78: 11686-95.
Poiesz BJ, Ruscetti FW, Gazdar AF, Bunn PA, Minna JD, Gallo RC. (1980) Detection and isolation of type C retrovirus particles from fresh and cultured lymphocytes of a patient with cutaneous T-cell lymphoma. Proc Natl Acad Sci U S A. 77: 7415-9.
Ramírez E, Cartier L, Torres M, Barría M. (2007) Temporal dynamics of human T-lymphotropic virus type I tax mRNA and proviral DNA load in peripheral blood mononuclear cells of human T-lymphotropic virus type I-associated myelopathy patients. J Med Virol. 79:782-90.
Ramírez E, Fernández J, Cartier L, Villota C, Ríos M. (2003) Defective human T-cell lymphotropic virus type I (HTLV-I) provirus in seronegative tropical spastic paraparesis/HTLV-I-associated myelopathy (TSP/HAM) patients. Virus Res. 91: 231-9.
Shimotohno K, Takano M, Teruuchi T, Miwa M. (1986) Requirement of multiple copies of a 21-nucleotide sequence in the U3 regions of human T-cell leukemia virus type I and type II long terminal repeats for trans-acting activation of transcription. Proc Natl Acad Sci USA. 83: 8112-6.
Sugamura K, Fujii M, Ueda S, Hinuma Y. (1984) Identification of a glycoprotein, gp21, of adult T cell leukemia virus by monoclonal antibody. J Immunol. 132:3180-4.
Towbin H, Staehelin T, Gordon J. (1979) Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: procedure and some applications. Proc Natl Acad Sci USA. 76: 4350-4.
Yoshida M, Miyoshi I, Hinuma Y. (1982) Isolation and characterization of retrovirus from cell lines of human adult T-cell leukemia and its implication in the disease. Proc Natl Acad Sci U S A. 6:2031-5.
Yoshida M. (2005) Discovery of HTLV-I, the first human retrovirus, its unique regulatory mechanisms, and insights into pathogenesis. Oncogene. 24:5931-7.
Yoshida M. (2010) Molecular approach to human leukemia: isolation and characterization of the first human retrovirus HTLV-1 and its impact on tumorigenesis in adult T-cell leukemia. Proc Jpn Acad Ser B Phys Biol Sci. 86:117-30.
Labor Omnia Vincit











![MECANISMOS TRANSDUCCIONALES DE INSULINA EN EL ...repositorio.uchile.cl/tesis/uchile/2010/qf-contreras_ae/pdfAmont/qf... · 11.3 La primera fase del incremento del [Ca2+] i depende](https://img.pdfslide.tips/doc/110x75/5e3de3906b213d3075414b06/mecanismos-transduccionales-de-insulina-en-el-113-la-primera-fase-del-incremento.jpg)

















