Embed Size (px)
Citation preview

UNIVERSIDAD DE CHILE
FACULTAD DE CIENCIAS AGRONÓMICAS
ESCUELA DE AGRONOMÍA
MEMORIA DE TÍTULO
TAXONOMÍA Y OBSERVACIONES BIOLÓGICAS DEL CHANCHITO BLANCO DE
LOS FRUTALES, Pseudococcus viburni (Signoret).
(HEMIPTERA: PSEUDOCOCCIDAE)
MARÍA SOLEDAD OYARZÚN IRACHETA
SANTIAGO, CHILE. 2004

1
UNIVERSIDAD DE CHILE
FACULTAD DE CIENCIAS AGRONÓMICAS
ESCUELA DE AGRONOMÍA
TAXONOMÍA Y OBSERVACIONES BIOLÓGICAS DEL CHANCHITO BLANCO DE
LOS FRUTALES, Pseudococcus viburni (Signoret).
(HEMIPTERA: PSEUDOCOCCIDAE)
Memoria para optar al Título Profesional
de Ingeniero Agrónomo Mención: Sanidad Vegetal.
MARÍA SOLEDAD OYARZÚN IRACHETA
PROFESOR GUÍA
Sr. Roberto H. González R. Ingeniero Agrónomo, M.S. Ph.D.
Calificaciones
7.0
PROFESORES CONSEJEROS : Sra. M. Angélica Guerrero S. Profesor Biología y Ciencias. Sr. Luis Sazo R. Ingeniero Agrónomo.
6.8
6.3
SANTIAGO, CHILE. 2004

2
TABLA DE CONTENIDOS
RESUMEN 4
SUMMARY 6
INTRODUCCIÓN 8
Objetivos 10
REVISIÓN BIBLIOGRÁFICA 11
Características de la familia Pseudococcidae 11
Sistemática e importancia del género Pseudococcus 12
Posición y clasificación sistemática del género 12
Sinopsis de Pseudococcus en Chile 14
Distribución geográfica y hospederos del género Pseudococcus 14
Importancia económica del género Pseudococcus en la fruticultura nacional 15
Pseudococcus viburni, el chanchito blanco de los frutales 18
Sinonimia y distribución mundial de la especie 19
Identificación 19
Biología y hábitos de la plaga 21
Dispersión del insecto 22
Hospederos de P. viburni 22
Enemigos naturales 24
Asociación con la hormiga argentina 25
Importancia económica y tipo de daño 26
MATERIALES Y MÉTODO 27
Lugar de estudio 27
Materiales 27
Materiales de laboratorio 27
Equipos ópticos y generales 27
Materiales de vidrio 27
Productos químicos 27

3
Instrumental y material auxiliar 28
Materiales generales de crianza 28
Material entomológico 28
Sustrato de crianza 28
Método 28
Metodología de crianza de los chanchitos blancos en laboratorio 28
Parámetros biológicos observados durante la crianza 29
Preparación para identificación y descripción de la especie 30
Identificación taxonómica 31
Análisis estadístico y diseño experimental 31
RESULTADOS 32
Identificación de la especie 32
Estudios de laboratorio 33
Aspectos taxonómicos 33
Descripción de la hembra (Fig.8) 36
Descripción del tercer estado ninfal (Fig.9) 41
Descripción del segundo estado ninfal (Fig.10) 45
Descripción del primer estado ninfal (Fig.11) 48
Descripción de la prepupa del macho (Fig.12) 51
Descripción de la pseudopupa del macho (Fig.13) 54
Descripción del macho (Fig.14) 57
Observaciones biológicas de la especie 58
Características macroscópicas e identificación a nivel de campo de los estados de
desarrollo de Pseudococcus viburni
CONCLUSIONES 70
LITERATURA CITADA 72
APÉNDICES 76

4
RESUMEN
Los hospederos atacados por Pseudocóccidos en Chile son numerosos, con la
característica que generalmente son infestados por más de una especie, generando un
complejo de plagas que pueden coexistir en un vegetal, lo cual agrega confusión para su
identificación y control. Es por ello que no existe todavía un adecuado método de
protección para evitar importantes pérdidas producidas por rechazos de fruta de
exportación.
Desde 1996, Estados Unidos declaró plagas restrictivas a este complejo de plagas por
la posible existencia de dos especies nuevas de Pseudococcus, pertenecientes al complejo
maritimus, las cuales aún no tienen estatus taxonómico sin embargo, y debido a que todavía
no han sido descritas, carecen de existencia taxonómica. Esta situación restrictiva se ha
ampliado prácticamente a todos los componentes de las poblaciones detectadas, toda vez
que no es posible hacer un diagnóstico específico, lo que afecta a cualquier componente del
género Pseudococcus, sea o no de carácter cuarentenario. La situación se ha agravado a
partir del 2003, con la incorporación específica de P. viburni (Signoret) como especie
cuarentenaria para México, afectando las exportaciones de uva, pomáceas y frutales de
carozo destinados a ese país.
Con esta medida, Pseudococcus viburni, el común chanchito blanco de los frutales, ha
resultado muy afectado, toda vez que se confunde con otras especies del complejo
ocasionando rechazos en la fruta de exportación. La deficiente identificación específica y el
desconocimiento que existe sobre su biología son los objetivos básicos del presente estudio.
El cual contribuye a la identificación y descripción microscópica de la especie en cada uno
sus estados de desarrollo; a través de una matriz de identificación basada en un conjunto de
caracteres taxonómicos, logrando así separar a P. viburni del complejo asociado.

5
Los caracteres diferenciales incluyen: número de poros discoidales en la órbita de los
ojos, número de poros traslúcidos en fémur y tibia del tercer par de patas, ausencia del
ducto tubular anillado (oral rim) entre el cerario 15 y 16, número de ductos anillados (oral
rims) en el dorso del abdomen y número de ductos tubulares (oral-collars) asociados a los
cerarios 10 y 11.
Los componentes de esta matriz han sido por primera vez determinados para la
población de P. viburni presente en Chile.
Esta investigación también permite conocer el desarrollo de una generación completa
de P. viburni, bajo condiciones controladas de laboratorio (temperatura, fotoperiodo, etc.)
en poblaciones mantenidas en brotes de papas. Así se determinó el periodo de desarrollo de
cada estadio ninfal, con su correspondiente descripción morfológica.
Palabras claves
Pseudocóccidos
Pseudococcus
Chanchitos blancos
Matriz taxonómica

6
SUMMARY
In Chile, mealybugs (Pseudococcidae) are general plant sucking insects occurring on a
large number of hosts. The latter are commonly infested by more than one mealybug
species, a feeding habit which leads to confusion in their identification and management.
For that reason, it is difficult to come to the development of adequate control programs to
cope with quarantine restrictions at present affecting fresh fruit for export.
In 1996, the USDA- APHIS restricted the entrance of this pest complex due to the
finding of two apparently new species of Pseudococcus possibly belonging to the
maritimus complex; however, since they have not yet been named, they do not have a legal
taxonomic existence. This quarantine restriction has been expanded to practically all
components of detected populations due to difficulty to attempting an specific diagnoses.
Accordingly, all members of Pseudococcus genus group could be rejected, either being or
not of quarantine concern. Since 2003, the above mentioned situation has been worsening
following Mexico’s quarantine restriction of Pseudococcus viburni (Signoret), affecting
exports of table grapes, pome and stone fruits exports to that country.
In that sense, the mealybug in common fruit trees, P. viburni, has become a particularly
subject to exclusion from fruit shipments. Its rather weak specific identification and the
lack of biological information have prompted to conduct the present research to establish an
adequate taxonomic matrix with all proper microscopic morphological characters.

7
The above mentioned differential characters include: number of disc pores around the
eyes, number of translucent pores in femora and tibiae of third pair of legs, lack of tubular
oral rim between cerario 15-16, numbers of dorsal oral rims ducts and number of tubular
oral-collars associated to cerario 10 and 11.
The above matrix components have been, for the first time, determined for the local
population existing in Chilean orchards.
In addition, this work provides biological information on the life-cycle of viburni
reared in potato sprouts under controlled laboratory conditions. The length of each
individual instar to the completion of full generation is given, as well as the morphological
description of each nymph instar.
Key words
Pseudococcids Pseudococcus Mealybugs Taxonomic matrix

8
INTRODUCCIÓN
Los chanchitos blancos constituyen actualmente un complejo de plagas integrado por 3
ó 4 especies que pueden coexistir en un vegetal, cuya separación taxonómica y métodos de
control resultan particularmente difíciles, no existiendo todavía un adecuado método de
protección para evitar importantes pérdidas producidas por rechazos de fruta de
exportación.
Aparte de los relativamente bajos niveles de daño directo, que ocurren por acción
trófica ejercida sobre la madera (floema), hojas y frutos, la acción más importante es
causada por su presencia, incluso en niveles subeconómicos de población, los que podrían
provocar rechazos cuarentenarios.
Respecto al comercio con los Estados Unidos, las especies de Pseudococcus,
estuvieron por décadas excluidas de “Acción Cuarentenaria” por el Departamento de
Agricultura de los Estados Unidos; sin embargo a partir de 1996, declaró plagas restrictivas
a este grupo por la posible existencia de dos nuevas especies pertenecientes al complejo
maritimus, las cuales por carecer de estatus taxonómico (y por tanto no tienen existencia
legal en el dominio entomológico), han ampliado esta situación restrictiva a todas las
poblaciones de este grupo de insectos.
Consecuentemente en las últimas temporadas se ha producido una importante escalada
de rechazos, principalmente por falta de capacidad de identificación inmediata durante la
inspección de aquellas especies afectas a la cuarentena (González, 2003). Con ésta medida,
la especie más común en Chile, P. viburni, ha resultado particularmente muy afectada,

9
generando en las últimas temporadas de exportación, valores de rechazo superiores al 50 %
del total nacional1.
Una reciente disposición cuarentenaria, reiterada por el gobierno de México, desde
marzo del 2003 prohibe el ingreso de fruta chilena por presencia del común chanchito
blanco de los frutales Pseudococcus viburni, no obstante esa especie se encuentra
distribuida en ese país (Miller et al, 1984 como P. affinis) (Gimpel & Miller, 1996).
La preocupación que significa el riesgo de que la cosecha para fines de exportación
esté infestada con estos insectos, se traduce en aplicaciones repetidas de insecticidas, lo
cual además de aumentar los costos, posterga la participación efectiva de enemigos
naturales.
La especie aquí tratada, P. viburni, el chanchito blanco de los frutales, considerada
como plaga clave en uva de mesa y ocasional en ciruelos, perales y manzanos, es la especie
de mayor distribución en Chile en huertos frutales, extendiéndose desde la primera a la
novena Región e Isla de Pascua (González et al, 2001).
El alto nivel de incidencia de Pseudococcus, la deficiente identificación específica del
grupo y el desconocimiento que existe sobre la biología de P. viburni hacen necesario su
estudio para proporcionar elementos de diagnóstico tanto al sector estatal que controla las
exportaciones, como para los productores y exportadores de productos hortofrutícolas.
1ASOEX, Asociación de Exportadores de Chile, 2004 (comunicación personal).

10
Objetivos
1. Describir taxonómicamente todos los estados de desarrollo de P. viburni para permitir
el adecuado reconocimiento de la especie.
2. Separar taxonómicamente a P. viburni del complejo asociado viburni-maritimus,
estableciendo los caracteres específicos propios de la especie.
3. Estudiar bajo condiciones de laboratorio, la biología reproductiva y estados de
desarrollo que caracterizan a una generación de Pseudococcus viburni.
4. Establecer principales características macroscópicas de la especie para su identificación
preliminar en el campo.

11
REVISIÓN BIBLIOGRÁFICA
Características de la familia Pseudococcidae
La familia Pseudococcidae es la más numerosa de los Coccoideos y la más importante
desde el punto de vista económico, conformada en Chile por un gran número de especies
polífagas, incluídos los géneros Pseudococcus, Planococcus y Phenacoccus los cuales se
encuentran asociados a especies de frutales de hoja caduca, frutales de hoja persistente
(cítricos en particular) y malezas de hoja ancha.
Las hembras se caracterizan por tener el cuerpo blando, ovalado, parcialmente
deprimido, con los metámeros muy marcados, y de tamaño variable entre 2 a 4 mm de
largo. El cuerpo se encuentra cubierto por un polvo ceroso de color blanco. Posiblemente la
función principal de la cera es mantener alejada la mielecilla de su cuerpo (Ripa y
Rodríguez, 1999), o bien proporcionarle un carácter hidrófobo que le permite soportar
condiciones de alta humedad, especialmente en el subsuelo, así como reducir sus riesgos
por aspersiones foliares de pesticidas.
Se caracterizan por poseer de 0 a 30 pares de cerarios, en concordancia con los
filamentos marginales, estas proyecciones tienen una forma y longitud característica en las
diferentes especies. La cabeza esta fusionada con el tórax, las antenas pueden estar
reducidas o presentes con cinco a nueve segmentos, normalmente un par de ojos simples,
aparato bucal dispuesto en forma de lazo, formado por 3 filamentos, usualmente tres pares
de patas bien desarrolladas (Williams & Granara de Willink, 1992).

12
En el extremo posterior se observan los lóbulos anales, ubicados lateralmente al anillo
anal, este último usualmente con 6 a 8 cerdas filamentosas. La vulva esta ubicada
ventralmente en el 8º segmento abdominal mientras que el ano ocupa posición dorsal en el
9º segmento (González, 1983).
Los machos son generalmente alados, de cuerpo más frágil y alargado que las hembras,
con dos largos filamentos cerosos en el ápice caudal. Poseen entre uno a tres y, a veces más
pares de ojos simples. Entre los machos hay formas ápteras y aladas (Artigas, 1994).
La gran mayoría de los chanchitos blancos, se reproducen ovíparamente (depositan
huevos). Los huevos protegidos por filamentos lanosos blancos son depositados bajo la
corteza, sobre los frutos, en las raíces y/o suelo y muy raramente en el follaje de las plantas.
Como otros insectos que se alimentan de savia, el aparato bucal de los chanchitos es
especializado y en su sistema digestivo poseen un órgano que filtra el contenido de lo que
succionan, excretando carbohidratos en forma de mielecilla que expelen por dos pares de
ostíolos ubicados dorsalmente.
Sistemática e importancia del género Pseudococcus
Posición y clasificación sistemática del género
El género Pseudococcus Westwood (del griego, Pseudo= falso y Kokkos = semilla), es
uno de los más abundantes y numerosos de la familia Pseudococcidae la cual agrupa las
especies fitófagas más importantes.

13
Su difícil caracterización morfológica, hace que su identificación haya sido compleja,
produciendo innumerables sinónimos para cada especie.
Según Yudelevich (1950), la sinonimia del género Pseudococcus es la siguiente:
1839 Pseudococcus Westwood
1843 Trechocorys Curt.
1867 Dactylopius Targ.
1875 ″ Signoret
1881 ″ Comst.
1881 Oudablis Signoret
1887 Dactylopius Maskell
1891 ″ Targ.
1895 ″ Cockerell
1902 Pseudococcus Cockerell
1903 ″ Fernald
1910 ″ Smith
1913 ″ Porter
La posición sistemática, según Storer y Usinger (1962), es la siguiente:
Phylum : Arthropoda
Clase : Insecta
Subclase : Hemimetabola
Orden : Homoptera (actualmente Hemiptera)
Suborden : Sternorryncha
Familia : Pseudococcidae
Género : Pseudococcus

14
Sinopsis de los Pseudococcus en Chile
Según Yudelevich (1950), en 1895 ya existía en Chile un problema económico causado
por Pseudocóccidos, desconociendo que especie lo originaba. Tampoco se conocía con
exactitud la forma como fueron introducidas al país, ya que las referencias eran escasas y
confusas.
Los autores Yudelevich (1950) y Campos (1953) mencionan a cuatro especies de
Pseudocóccidos establecidas en el país: P. fragilis Brain (actual P.calceolariae),
Planococcus citri (Risso), P. longispinus (Targ.) (antes P. adonidum) y P. maritimus
(Ehrhorn). Posteriormente en el Catálogo de las Plagas Agrícolas de Chile (González, et al,
1973), se establece la presencia de siete especies de Pseudocóccidos, que actualmente se
mantienen vigentes: Dysmicoccus brevipes (Ckll.), Planococcus citri, P. elisae (Borsch), P.
fragilis, (actual calceolariae) P. longispinus, P. maritimus y P. obscurus Essig. (actual P.
viburni) (González, 1989).
Distribución geográfica y hospederos del género Pseudococcus
No es posible precisar con exactitud la distribución o área geográfica que abarcan los
Pseudocóccidos en Chile, su extremada polifagia sumado a una serie de microclimas
regionales, los hace difíciles de ubicar. Sin especificar su intensidad de ataque, abundancia,
ni perjuicios, éstos han sido detectados globalmente desde la I región de Tarapacá hasta la
IX región de la Araucanía (Yudelevich, 1950).

15
Los hospederos afectados por los Pseudoccocus en Chile son numerosos, con la
característica que generalmente son atacados por más de una especie, lo que agrega
confusión en su identificación.
Importancia económica del género en la fruticultura nacional
Más de 20 especies frutales son atacadas por Pseudococcus, entre las cuales se
incluyen las más importantes de la fruticultura nacional (cuadro 1). No en todas las
especies mencionadas el ataque asume la misma intensidad de daño. Varía notablemente de
una especie a otra, lo que demuestra cierto tipo de preferencia alimentaria, edad y el tipo
de hábitat para determinadas especies. Mas aún, dentro de una misma especie se presenta
el caso de ciertas variedades más afectadas que otras (Yudelevich, 1950).
Su importancia económica ha aumentado en razón a la amplia capacidad de este grupo
de invadir frutos y formar colonias que se establecen en lugares protegidos por ejemplo: en
frutos de carozo, la infestación es pedicelar o en alguna sutura o grieta lateral, en peras y
nísperos la ubicación es calicinal, en pera asiática y manzanas se ubican en las cavidades
pedicelar y calicinal, pudiendo desde esas posiciones invadir la región carpelar. En caquis y
kiwis en cambio, la infestación ocurre bajo los tépalos del fruto (González et al, 2001).

16
Cuadro 1. Hospederos frutales atacados por Pseudococcus (Yudelevich 1950).
Nombre común Nombre científico
Arándano : Vaccinium corymbosum
Caqui : Diospyrus caqui
Chirimoya : Anonna cherimola
Ciruelo : Prunus domestica
Mandarina clementina : Citrus reticulata
Damasco : Prunus armeniaca
Duraznero –Nectarin : Prunus persica
Frambueso : Rubus idaeus
Granado : Punica granatum
Kiwi : Actinidia deliciosa
Limonero : Citrus limon
Manzano : Malus domestica
Naranjo : Citrus sinensis
Palto : Persea americana
Peral : Pyrus communis
Peral asiático : Pyrus pyrifolia
Pomelo : Citrus paradisi
Uva de mesa : Vitis vinifera
Zarzaparrilla : Ribes sp.
Los chanchitos blancos son insectos floemáticos que se alimentan directamente de
madera, hojas y frutos. El daño lo ocasionan por succionar savia e inyectar toxinas y por su
presencia, en forma de colonias y destacada por su coloración blanca harinosa. Sus
deyecciones contienen azúcares que son aprovechados por las hormigas y sirven de
sustrato para el desarrollo de hongos como el hongo negro, hollín o fumagina. La secreción

17
azucarada se dispersa por hojas, ramillas y frutos, estos órganos se ensucian creando
problemas para una adecuada fotosíntesis de las hojas y la comercialización de los frutos,
los cuales deben ser lavados antes de llegar al mercado (Artigas, 1994).
Además del daño directo al cultivo, debe agregarse el problema de las restricciones
cuarentenarias. En la temporada 2002-2003 los Pseudocóccidos, fueron responsables del
mayor volumen de rechazo de fruta de exportación, con un porcentaje superior al 50 % del
total nacional de rechazos SAG-USDA.2
Con respecto a las exportaciones a EE.UU., el USDA-APHIS ha señalado que
posiblemente en Chile existen dos especies no descritas, las cuales motivan la restricción, el
resto de las especies comunes en nuestros frutales se encuentran también representadas en
ese país, y por lo tanto no ofrecen riesgo cuarentenario. Las posibles nuevas especies
durante la inspección, no pueden ser correctamente identificadas, especialmente si se
encuentran como formas inmaduras o por falta de capacidad de identificación en ese
momento.
Actualmente las cuarentenas oficiales que Chile debe cumplir, proceden de Estados
Unidos desde 1997, contra especies innominadas de Pseudococcus “1 y 2” y de México,
desde marzo 2003, específicamente contra P. viburni. Adicionalmente, otros mercados,
como Perú, Brasil y Costa Rica, se han plegado a estas disposiciones sin comunicarlo
oficialmente (González, 2003).
En fruta fresca, la secuencia de rechazos por Pseudoccoccidae comienza en mayo con
limones, luego en septiembre-octubre con chirimoyas; en noviembre con nísperos y uva de
la Tercera y Cuarta Región, en diciembre con frutos de carozo; duraznos, nectarines
damascos y ciruelas, posteriormente de enero a marzo con ciruelas, peras, duraznos,
2 ASOEX, Asociación de Exportadores de Chile, 2004 (comunicación personal).

18
nectarines y uva de mesa; hacia fines de marzo continúan con caquis, kiwis y manzanas
(González, 1994).
Dentro de las frutas rechazadas en la temporada 2002-2003, las manzanas representan
el mayor porcentaje de rechazo, el 36 % se debió a problemas con Pseudocóccidos. Los
principales causales de rechazos incluyen: huevos, estados inmaduros no identificados y la
especie P. viburni que alcanzó valores cercanos al 30 % del total nacional de rechazos3.
Por la mayor extensión e intensidad de ataque y la reciente disposición cuarentenaria
por parte del gobierno mexicano, provocado por P. viburni, hace necesario referirse con
más detalle a esta especie.
Pseudococcus viburni, el chanchito blanco de los frutales
El chanchito blanco de los frutales es actualmente considerado plaga clave en uva de
mesa y ocasional en ciruelos, perales y manzanos. Está repartido en las zonas de climas
templado del H. Norte, S. Africa, Australia y N. Zelanda, así como desde México a
Argentina. En Chile es la especie de mayor distribución, extendiéndose desde la primera a
la novena Región e Isla de Pascua (González et al, 2001).
Esta especie, se alimenta directamente del floema, ya sea de la raíz o la porción aérea
de la planta, es notoriamente cortical en su ubicación en el hospedero, desde allí se
moviliza en distintos períodos directamente a los frutos, prácticamente sin detenerse en el
follaje (González, et al 2001).
3 ASOEX, Asociación de Exportadores de Chile, 2004 (comunicación personal).

19
Sinonimia y distribución geográfica de la especie
P. viburni ha recibido distintas denominaciones especificas que conforman su
sinonimia:
Período previo a 1962, recibió los nombres científicos actualizados hasta esa fecha por
Mckenzie (1962) como Pseudococcus obscurus Essig.
Período 1962-1984, sinonimia actualizada por Miller et al,1984 como Pseudococcus
affinis (Maskell).
Período reciente, con el aporte de Gimpel & Miller (1996), nombre actual de la especie
Pseudococcus viburni (Signoret), con distribución en: Argentina, Australia, Azores,
Bélgica, Brasil, Canadá, Chile, Costa Rica, Cuba, Dinamarca, Ecuador, Corea, México,
Perú, Venezuela, Uruguay, España, Italia, Inglaterra (invernaderos), Holanda, N. Zelanda,
etc. Por lo tanto, se considera especie cosmopolita. Identificación
Las hembras de P. viburni poseen 16 a 17 pares de cerarios, en correspondencia con
los filamentos marginales; el par caudal presenta un mayor tamaño, le sigue en longitud el
par paracaudal y luego el resto los filamentos marginales.
Los caracteres morfológicos más importantes de la especie son: número de poros
discoidales en la periferia basal del ojo, variable de 1 a 3; rango de poros translúcidos
presentes en las patas posteriores de 38 a 162 en las tibias y de 15 a 150 en el fémur;

20
número y distribución de oral rims presentes en el dorso del abdomen, etc. (Gimpel &
Miller, 1996). Otros autores consideran igualmente importante el número de poros
multiloculares en el vientre del tórax, el largo de las setas de la cabeza y la forma de la tibia
(Miller et al, 1984).
Cuerpo ovalado, cubierto por una cera pulverulenta blanca, bajo la cual se aprecia el
color ligeramente rosado del cuerpo. El color de la hemolinfa secretada por los poros
caudales es de color blanca perlada (González, 2003).
En relación a las hembras pasan por tres estadíos ninfales, los machos sólo presentan
dos estadíos, para luego encapullarse, donde ocurre el estado prepupoide y pupoide, dando
origen a un individuo alado, pequeño y de corta vida. (González, 1983).
Los machos son de color gris, con alas trasparentes cubiertas por un extracto de cera,
se distinguen de los demás Pseudococcidae por la presencia de una extremidad abdominal
(style) mas largo y redondeado. (Ciampolini et al, 2003)
Los huevos son ovalados de color amarillo brillante y están unidos por una masa
algodonosa suelta e uniforme.

21
Biología y hábitos de la plaga
El chanchito blanco de los frutales, es una especie gregaria y forma grupos en lugares
muy protegidos de la planta huésped. Es factible encontrarlas en el envés de la hoja, sobre
todo junto a la nervadura central, en la base del pecíolo y bajo el cáliz de los frutos.
Inverna de preferencia al estado de huevo, a pesar de que diferentes autores han
observado todos los estados de la plaga atribuible a la presencia de más de una especie de
chanchito blanco, esto último también sujeto a la verificación taxonómica (González,
1991).
Una vez que se inicia el crecimiento de la planta, en primavera, comienza la eclosión
de los huevos invernantes, las ninfas neonatas permanecen en el interior del saco por varios
días, luego los individuos ascienden por el tronco, estableciéndose en la unión de la madera
del año anterior con el brote tierno. Durante la segunda mitad de noviembre las hembras
regresan bajo la corteza a oviponer, originando la segunda generación y fase más
importante de ataque de la plaga. Durante esta postura, las hembras oviponen masas de
hasta 200 huevos. Recién puestos estos son de color amarillo, tornándose ligeramente
anaranjados a medida que maduran, siendo incubados en un periodo variable según la
temperatura por 8 a 10 días. Las ninfas recién emergidas, permanecen junto a la lanosidad
por algunos días, comenzando el ascenso hacia partes verdes y frutos a fines de noviembre
(González, 1983).
Entre el estado de huevo y adulto hay tres estados ninfales con una duración variable
según la generación, por lo general, la primera generación demora casi 50 días en alcanzar
el estado adulto. La segunda generación, en cambio es más rápida, alcanzando su estado
adulto en 30 a 40 días, momento en el cual las hembras comienzan a oviponer activamente,

22
por lo que en las cosechas de febrero se pueden observar lanosidades y masas de huevos en
los frutos.
En la vid, respecto al ataque de racimos, prefieren aquellos en contacto con la madera,
colonizan el racimo cuando éste se llena, lo que les da una mayor protección.
Posteriormente a la cosecha son colonizados los pequeños racimos remanentes, ubicándose
abundantes masas de huevos (Ripa y Rojas, 1990). Luego parte de la población desciende
al tronco (bajo la corteza) o sectores protegidos para seguir alimentándose y realizar
posturas de otoño, la otra parte de la población permanece en cargadores y racimos,
especialmente durante inviernos benignos (González, 1983). Dispersión del insecto
Las ninfas al emerger de los sacos ovígeros son muy activas y se dispersan por medios
propios, por la acción de hormigas o por el viento que arrastra a las pequeñas ninfas de los
primeros estados hacia plantas cercanas (González, 1991). El grado de infestación con
malezas de hoja ancha también ayuda en la diseminación de la plaga. Hospederos de P.viburni
Se ha observado que Pseudococcus viburni es cosmopolita y polífaga, con capacidad
de establecerse sobre un amplio número de hospederos primarios y secundarios. Dentro de
los hospederos primarios destacan alrededor de 16 especies cultivadas en nuestro país
(Cuadro 2). En cuanto a los hospederos secundarios, se han identificado un gran número de
especies, encontrándose especialmente a nivel radicular durante el invierno, es así como los
hospederos secundarios se convierten en la principal fuente de inoculo de la plaga para la
temporada siguiente (Ripa, et al 1992; Ripa y Rojas, 1990) (Cuadro 3).

23
Cuadro 2. Especies hospederas primarias detectadas en Chile para P. viburni (Prado,
1991).
Nombre común Nombre científico
Alfalfa : Medicago sativa
Cerezo : Prunus avium
Frambueso : Rubus idaeus
Garbanzo : Cicer arietinum
Jacarandá : Jacaranda angustifolia
Laurel de flor : Nerium oleander
Manzano : Malus domestica
Mora : Rubus sp.
Naranjo : Citrus sinensis
Níspero : Eriobotrya japonica
Papa : Solanum tuberosum
Pepino dulce : Solanum muricatum
Peral : Pyrus communis
Rábano : Raphanus sativus
Vid : Vitis vinifera
Zarzaparrilla : Ribes sp.

24
Cuadro 3. Especies hospederas secundarias detectadas en Chile para P. viburni (Ripa y
Rojas, 1990).
Nombre común Nombre científico
Alfilerillo : Erodium sp.
Amor seco : Bidens pilosa
Correhuela : Convolvulus arvensis
Hinojo : Foeniculum vulgare
Hualputra : Medicago sp.
Diente de león : Taraxacum officinale
Malva : Malva sp.
Natri : Solanum tomatillo
Ñilhue : Sonchus sp.
Palqui : Cestrum parqui
Quingüilla : Chenopodium sp.
Quilo : Mühelenbeckia hastulata
Senecio : Senecio sp.
Tomatillo : Solanum sp.
Enemigos naturales
Los chanchitos blancos presentan una gran diversidad de asociaciones con sus
enemigos naturales. En los cuadros 4 y 5, se ofrecen citas de las asociaciones con
parasitoides y depredadores respectivamente.

25
Cuadro 4. Parasitoides de chanchitos blancos (Ripa y Rodríguez, 1999 y Prado, 1991)
Especie Familia Orden Leptomastix epona
Pseudaphycus angelicus
Pseudaphycus flavidulus *
Tetracnemoidea brevicornis
Encyrtidae Hymenoptera
Cuadro 5. Depredadores de chanchitos blancos (Ripa y Rodríguez, 1999 y Prado, 1991)
Especie Familia Orden Cryptolaemus montrouzieri *
Hyperaspis funesta
Scymnus nitidus
Coccinellidae
″
″
Coleoptera
Leucopis sp Chamaemyiidae Diptera
Ocyptamus confusus Syrphidae Diptera
Chrysoperla sp * Chrysopidae Neuroptera
Sympherobius maculipennis Hemerobiidae Neuroptera
* Enemigos naturales más efectivos, según observación durante varios años (Ripa y Rojas, 1990).
Asociación con la hormiga argentina
La hormiga argentina Linepithema humile (Mayr), vive en colonias de hasta 3000
individuos, entre larvas, pupas, obreras, machos y reinas. Su mayor fuente de carbohidratos
son las secreciones azucaradas que secretan algunos insectos y el néctar de las plantas. Una
de las primeras citas que mencionan la asociación con homópteros fue Essig (1917), quien
observó a ésta especie construir en los vértices de ramillas, una protección sobre las
colonias de chanchitos blancos, usando detrito y moho de hojas. Tal como se relaciona con

26
gran número de insectos chupadores, la hormiga argentina se encuentra frecuentemente
alimentándose de la mielecilla secretada por los chanchitos blancos (Ripa y Rojas 1990).
La hormiga argentina no produce daño directo sobre las especies hospederas, porque
no es un insecto fitófago, sin embargo, interfiere con el control biológico que ejercen los
enemigos naturales. Este efecto se produce porque se establece una relación de beneficio
mutuo, donde la hormiga obtiene alimento, y la plaga protección de sus enemigos naturales
que son alejados por las hormigas. La abundancia de hormigas se considera un indicativo
que se está frente a una población importante de chanchitos blancos (Ripa y Rodríguez,
1999).
Importancia económica y tipo de daño
El chanchito blanco de los frutales provoca tres tipos de daños sobre su hospedero, uno
de tipo trófico considerado de escasa importancia fisiológica; referido especialmente a los
estados ninfales que se alimentan de la savia del huésped; el segundo de naturaleza
cosmético o de calidad, debido a la presencia de manchas de aspecto aceitoso oscuro en
ramas, ramillas y hojas derivado de la secreción azucarada que constituye un sustrato para
la colonización de hongos oscuros y polvo (Artigas, 1994). El tercer tipo de daño y más
importante es el cuarentenario; los insectos al ubicarse en los frutos y depositar en ellos
lanosidades y masas de huevos, se corre riesgo de rechazo por el hallazgo de ejemplares
vivos sobre el hospedero (Ripa, et al 1992), Actualmente existe tolerancia cero en
inspecciones de frutas de exportación.
P. viburni es considerada una especie cosmopolita. Sin embargo, el único país que ha
impuesto restricciones cuarentenarias especificas para esta especie, es México. En relación
a la cuarentena norteamericana, atribuida a especies aún no descritas, los estados ninfales y
huevos de P. viburni quedan también involucradas como responsables de rechazos por la
incapacidad de identificación taxonómica durante las inspecciones.

27
MATERIALES Y MÉTODO
Lugar de estudio. Las observaciones biológicas del chanchito blanco, Pseudococcus viburni
en laboratorio, se realizó en el año 2003, en el Museo Entomológico Luis Peña,
Departamento de Sanidad Vegetal de la Facultad de Ciencias Agronómicas de la
Universidad de Chile.
Materiales
Materiales de laboratorio
Equipos ópticos. Incluyeron microscopios binoculares corriente y de fase contrastada,
estereomicroscopio para observación de las características macroscópicas y realizar
observaciones biológicas in vivo de los ejemplares. Cámara de crianza con temperatura y
fotoperíodo regulable de 7 pies cúbicos.
Material de vidrio. Discos petri y frascos para guardar ejemplares en alcohol, crisol de
porcelana para maceración, porta y cubre objetos para montaje permanente.
Productos químicos. Solución de KOH al 10 % para remover la grasa corporal de los
especímenes, ácido acético para neutralizar, fucsina ácida para tinción y reactivo xilol. El
montaje se realizó en un portaobjeto con bálsamo del Canadá, diluido con xilol.

28
Instrumental y material auxiliar. Bandejas de plástico, pocillos, pinzas, minucias, mesa
iluminada de dibujo y cámara digital Sony, modelo Digital Still Camera DSC-P31.
Materiales generales de crianza
Material entomológico. El material vivo de Pseudococcus sp fue colectado en parronales
caseros con escasa o ninguna intervención, ubicados en la Región Metropolitana.
Sustrato de crianza. La crianza de chanchitos blancos en laboratorio se realizó sobre brotes
etiolados de papa de la variedad Desirée, de tamaño y brotación uniforme.
Método
Metodología de Crianza de los chanchitos blancos en laboratorio
La población se obtuvo a partir de hembras adultas, previamente identificadas por
claves taxonómicas, material que fue inoculado en el hospedero para su posterior
observación y desarrollo en el laboratorio.
La crianza se realizó sobre brotes etiolados de papas, de acuerdo con la metodología
indicada por Fisher 1963, (citada por Martini 1992.)

29
Para esto, se colocaron papas-semillas en cajones con arena húmeda previamente
tratada con agua hirviendo. Después de brotados los tubérculos, se infestaron con hembras
adultas fertilizadas. Posteriormente las bandejas fueron almacenadas a 25º C en la cámara
de crianza, con un fotoperíodo de 12 horas.
Los criterios de selección de las hembras fueron: hembras fertilizadas (más de 5 días
en contacto con machos adultos) de tamaño homogéneo, promedio de 3 mm y apariencia
activa, sin presencia de daño físico.
Los tubérculos brotados se distribuyeron en 8 bandejas con 5 pocillos cada una, con
sólo una hembra fertilizada de manera de poder observar el desarrollo individual de la
especie; se obtuvo una población inicial de 40 hembras, que fue representativa para
determinar los parámetros observados durante toda una generación.
Parámetros biológicos observados durante la crianza
a) Duración del período de postura del saco ovígero, desde el inicio del proceso hasta la
mortalidad de la hembra, todo el proceso verificado a través de fotografías digitales.
b) Número promedio de huevos por ovisaco.
c) Período de desarrollo de las ninfas en cada estado, desde neonatas hasta adultas.
d) Respecto a los machos se observó: hábitos de las ninfas en encapullarse en las fases
prepupoide y pseudopupa, longevidad del insecto, proporción de hembras/machos y
eficiencia reproductiva. Para determinar la eficiencia reproductiva de los machos se
instaló otro ensayo con 4 tratamientos y 4 repeticiones cada uno, donde se aisló machos,
con distinto número de hembras vírgenes, durante un número variable de días (según la
mortalidad del macho).

30
e) Mediciones taxonómicas macroscópicas por estado de desarrollo, como por ejemplo:
longitud de los filamentos caudales, largo de las antenas u otros apéndices del cuerpo
Preparación para identificación y descripción de la especie
Parte del material criado en los brotes de papas, se utilizó para realizar preparaciones
microscópicas de tipo permanente, con el que se realizaron las determinaciones por estado
de desarrollo de ninfas, hembras y machos.
El montaje para las hembras se realizó mediante la técnica descrita por Etcheverry y
Herrera (1972), cuyos pasos son los siguientes:
1. Perforar al insecto con aguja fina en la mitad anterior del cuerpo estando aún vivo;
2. Calentar en KOH al 10 % y hervir durante un minuto;
3. Presionar el cuerpo con el objeto de eliminar su contenido interno;
4. Hervir nuevamente en KOH al 10 %
5. Lavar en agua destilada, posteriormente en ácido acético, hasta vaciar la grasa corporal
completamente (2-3 minutos);
6. Teñir con fucsina ácida (20 minutos), y luego pasar a alcohol 75 % (5 minutos), alcohol
96 % (10 minutos), xilol (5 minutos);
7. Finalmente, el montaje permanente se hace en un portaobjeto con bálsamo del Canadá,
diluido con xilol.
Los ejemplares machos, huevos y primeros estados ninfales, se montaron directamente
en medio de Hoyer.

31
Identificación taxonómica
La identificación taxonómica de los ejemplares adultos, se hizo en base a claves
taxonómicas establecidas por Miller et al (1984), Williams & Granara de Willink (1992) y
Gimpel & Miller (1996).
La caracterización taxonómica de los diferentes estadíos se realizó a través de dibujos
macroscópicos y fotografías digitales originales del autor.
Para la caracterización taxonómica y las observaciones por parámetro biológico a
observar, se tomó una muestra de la población de 100 ejemplares.
Análisis estadístico y diseño experimental
Los resultados, se analizaron mediante una estadística descriptiva: a través de
promedios y rango de variación obtenidos con instrumentos de medición microscópica de
las observaciones biológicas realizadas durante el período de ensayo.

32
Figura 1. Hembras adultas de P. viburni, criadas en brotes de papa.
RESULTADOS
Identificación de la especie
El material vivo de Pseudococcus sp. fue colectado en la temporada 2002-2003 desde
parronales ubicados en la Región Metropolitana. Las poblaciones se obtuvieron a partir de
hembras adultas.
Mediante claves taxonómicas de hembras adultas establecidas por Miller et al (1984),
Williams & Granara de Willink (1992) y (Gimpel & Miller, 1996). Se determinó que la
especie estudiada correspondía a Pseudococcus viburni (Signoret), material que se inoculó
sobre brotes de papas para su posterior observación y desarrollo en el laboratorio (figura 1).

33
Estudios de laboratorio
Los estudios de laboratorio fueron conducidos sobre colonias de poblaciones puras de
la especie mantenidas durante meses sobre brotes de papa, para realizar los estudios
taxonómicos, observaciones biológicas y características macroscópicas de los distintos
estados de desarrollo de la especie.
Aspectos taxonómicos
Debido a la compleja identificación taxonómica de la especie, la descripción de cada
estado de desarrollo, se realizó observando un conjunto de caracteres morfológicos que
conforman una matriz de identificación. Esta matriz presenta caracteres comunes para
varias especies, como por ejemplo: número de cerarios, de antenitos, ubicación de circulus
etc. y caracteres específicos los que taxonómicamente difieren entre las especies y permiten
separar a P. viburni de otros Pseudococcus del complejo maritimus (Miller et al 1984) y
(Gimpel & Miller, 1996).
P. viburni presenta 3 estados ninfales en ejemplares femeninos y dos estadios
adicionales para el caso de los machos. Los estudios taxonómicos y su posterior descripción
para cada fase de desarrollo, se realizó sobre preparaciones permanentes, registrándose los
componentes taxonómicos de las superficies dorsales y ventrales para cada individuo. Se ha
elaborado esquemas de todos los estados de desarrollo, destacando los caracteres
específicos (cuadro 6), y un glosario de términos y abreviaturas empleadas (Anexo I).
La figura 8 muestra una hembra adulta con la ubicación de los caracteres referenciales
que se mencionan mas adelante.
34
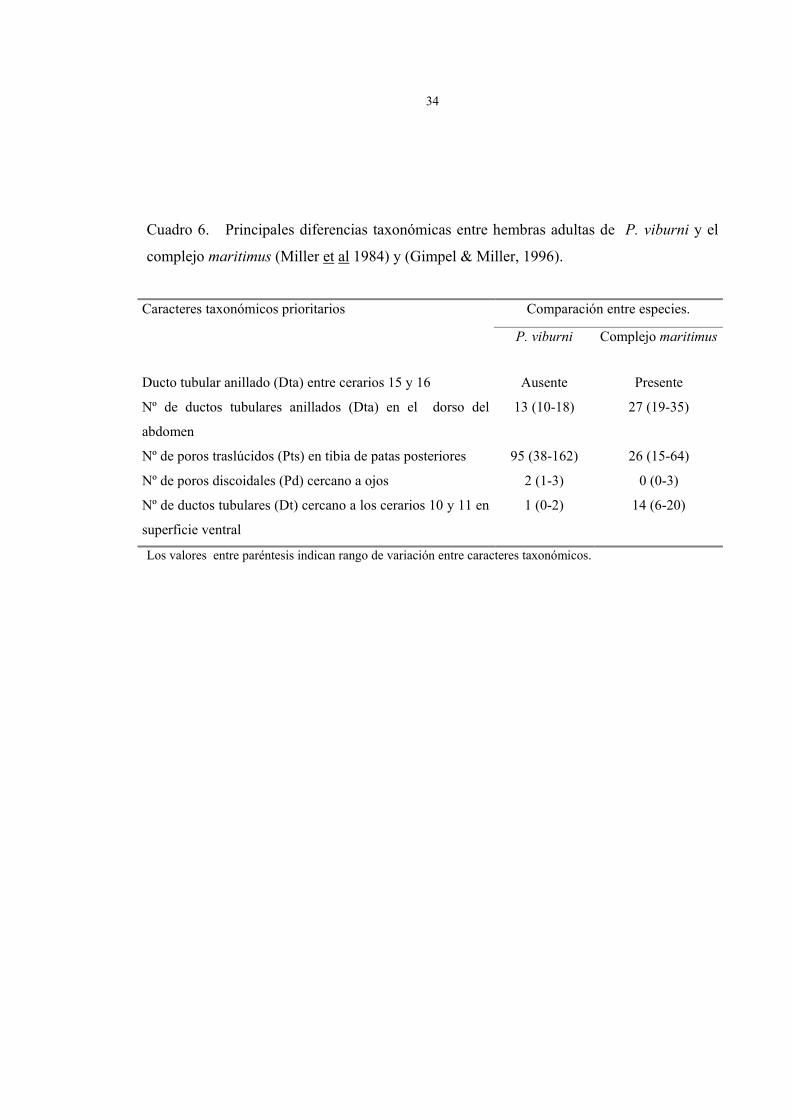
Cuadro 6. Principales diferencias taxonómicas entre hembras adultas de P. viburni y el
complejo maritimus (Miller et al 1984) y (Gimpel & Miller, 1996).
Comparación entre especies. Caracteres taxonómicos prioritarios
P. viburni Complejo maritimus
Ducto tubular anillado (Dta) entre cerarios 15 y 16 Ausente Presente
Nº de ductos tubulares anillados (Dta) en el dorso del
abdomen
13 (10-18) 27 (19-35)
Nº de poros traslúcidos (Pts) en tibia de patas posteriores 95 (38-162) 26 (15-64)
Nº de poros discoidales (Pd) cercano a ojos 2 (1-3) 0 (0-3)
Nº de ductos tubulares (Dt) cercano a los cerarios 10 y 11 en
superficie ventral
1 (0-2) 14 (6-20)
Los valores entre paréntesis indican rango de variación entre caracteres taxonómicos.
35
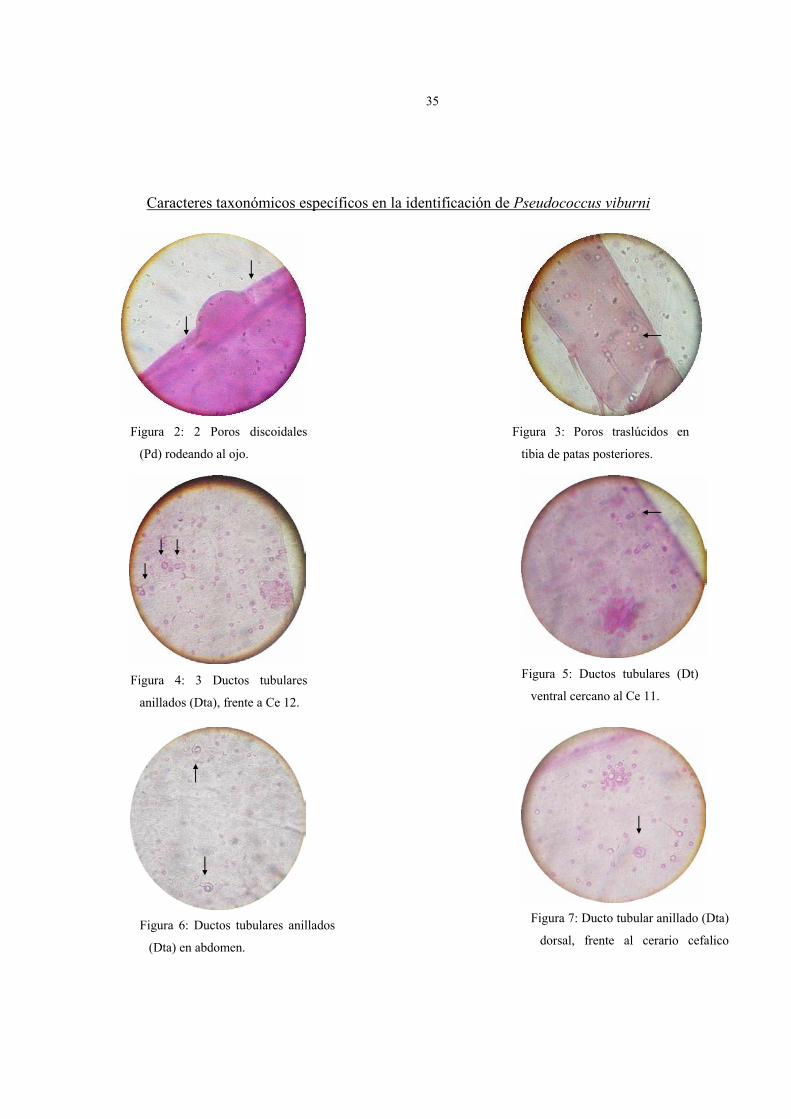
Caracteres taxonómicos específicos en la identificación de Pseudococcus viburni
Figura 2: 2 Poros discoidales
(Pd) rodeando al ojo.
Figura 3: Poros traslúcidos en
tibia de patas posteriores.
Figura 7: Ducto tubular anillado (Dta)
dorsal, frente al cerario cefalico
Figura 5: Ductos tubulares (Dt)
ventral cercano al Ce 11. Figura 4: 3 Ductos tubulares
anillados (Dta), frente a Ce 12.
Figura 6: Ductos tubulares anillados
(Dta) en abdomen.

36
Descripción de la hembra (Fig.8)
Cuerpo de forma oval de 2 a 4 mm de largo por 1 a 2 mm de ancho, antenas de 8
segmentos, 17 (16) pares de cerarios, abundantes poros y setas distribuidas en todo el
cuerpo, poros traslúcidos en patas posteriores, circulus y vulva presentes.
En la figura 8 están representados los caracteres taxonómicos de la hembra adulta en
vista dorsal y ventral fijado en preparaciones permanentes. Los parámetros observados que
caracterizan a la hembra son los siguientes:
Vista dorsal:
1. En general, 17 pares de cerarios; cerario caudal (Ce1) ligeramente esclerosado y
con mayor número de poros triloculares (Pt) rodeando las setas cónicas.
2. El número de setas cónicas (Sc), setas auxiliares (Sa) y poros triloculares (Pt) que
acompañan a cada cerario, varia según la siguiente matriz:
Ce1: 2 setas cónicas, acompañado de 2 setas auxiliares, rodeado de 40 a 50 poros
triloculares.
Ce2 al Ce9: 2 setas cónicas, acompañado de 1 a 2 setas auxiliares, rodeado de 20 a
25 poros triloculares.
Ce10: 2 setas cónicas, setas auxiliares de 1 a 0, generalmente ausente y 10 a 15
poros triloculares rodeando las setas.

37
Ce11: 2 setas cónicas acompañado de 1 a 2 setas auxiliares, rodeado de 15 a 25
poros triloculares.
Ce12: 3 setas cónicas acompañado de 3 a 4 setas auxiliares y 20 a 25 poros
triloculares.
Ce13-Ce14: 2 setas cónicas acompañado de 1 a 2 setas auxiliares, rodeado de 15 a
25 poros triloculares.
Ce15-Ce16-Ce17: Cerarios cefálicos, de 3 a 4 setas cónicas acompañados de 1 a 2
setas auxiliares, rodeado de 20 a 25 poros triloculares.
3. Los ductos tubulares anillados (Dta) (oral rim), están asociados a un poro
discoidal y sin seta auxiliar. La mayoría se ubica frente a los cerarios; en el cerario 12
se pueden encontrar de 1 a 3 Dta y en la parte media del tórax de 3 a 4. Los más
importantes desde el punto de vista taxonómico, considerado como carácter
específico de la especie, son los ubicados en el dorso del abdomen cuyo número varía
de 10 a 18, a diferencia del resto de los Pseudococcus del complejo maritimus que
presentan un mayor número. Otra característica relevante para P. viburni es la
ausencia de un Dta entre el Cerario 15 y el 16; en cambio en otros individuos del
complejo estos ductos tubulares anillados existen.
4. Ductos tubulares (Dt), oral-collars en los submárgenes entre cada cerario,
disminuyendo hacia la región cefálica.
5. Existen abundantes poros triloculares dispersos sobre el dorso, disminuyendo
hacia los márgenes.

38
6. Abundantes poros discoidales (Pd) de pequeño tamaño, ubicados en toda la
superficie, frente a cada cerario y asociados a cada ducto tubular anillado.
7. Ausencia de poros multiloculares.
8. Setas corporales numerosas y de diversos tamaños: tres pares de setas largas se
ubican en el anillo anal; le siguen en longitud las del abdomen y luego las de la
región toráxica del insecto.
Vista ventral:
1. Antenas de 8 segmentos, cada segmento acompañado de 3 a 5 setas, el último
segmento de mayor longitud y con mayor número de setas.
2. Circulus (Cir., Fig.8) presente entre los segmentos III y IV del abdomen.
3. Poros translúcidos (Pts) presentes en un rango de 38 a 162 en tibia y de 15 a 150
en el fémur del tercer par de patas. El número de poros es considerado un carácter
específico y propio de la especie.
4. Poros multiloculares (Pm) entre segmentos IV y VIII, abundantes rodeando la
vulva y generalmente ausentes en el tórax.
5. Abundantes poros triloculares dispersos sobre toda la superficie y concentrados en
un número de 100 a 200 en el segmento VI del abdomen.
6. Escasos poros discoidales dispersos en el vientre, ubicados en el lóbulo anal, en
el tórax bajo espiráculos y de 1 a 3 alrededor de cada ojo.

39
7. Ducto tubular anillado con 1 a 2 poros discoidales y una seta asociada cercano al
cerario 10.
8. Ductos tubulares asociados a bandas de poros multiloculares en los segmentos IV
al VII y abundantes en el submargen de los mismos segmentos, generalmente
ausentes en la cabeza, masas de 5 a 15 en cerario 12 y 13 y de 0 a 2 cercano al cerario
11.
9. Setas más largas que las del dorso, las de mayor tamaño son las del lóbulo anal y
las del trocánter, luego las setas del abdomen y las que se ubican en la región cefálica
entre y posterior a las antenas.
10. El número de setas en las patas es de 54 a 80 y se distribuyen como sigue: 6 a 8
en la coxa, 2 a 6 en el trocánter, 18 a 25 en el fémur 20 a 30 en la tibia y 8 a 12 en el
tarso.
11. La relación de tamaño tibia/tarso es de 2,2 a 2,7.
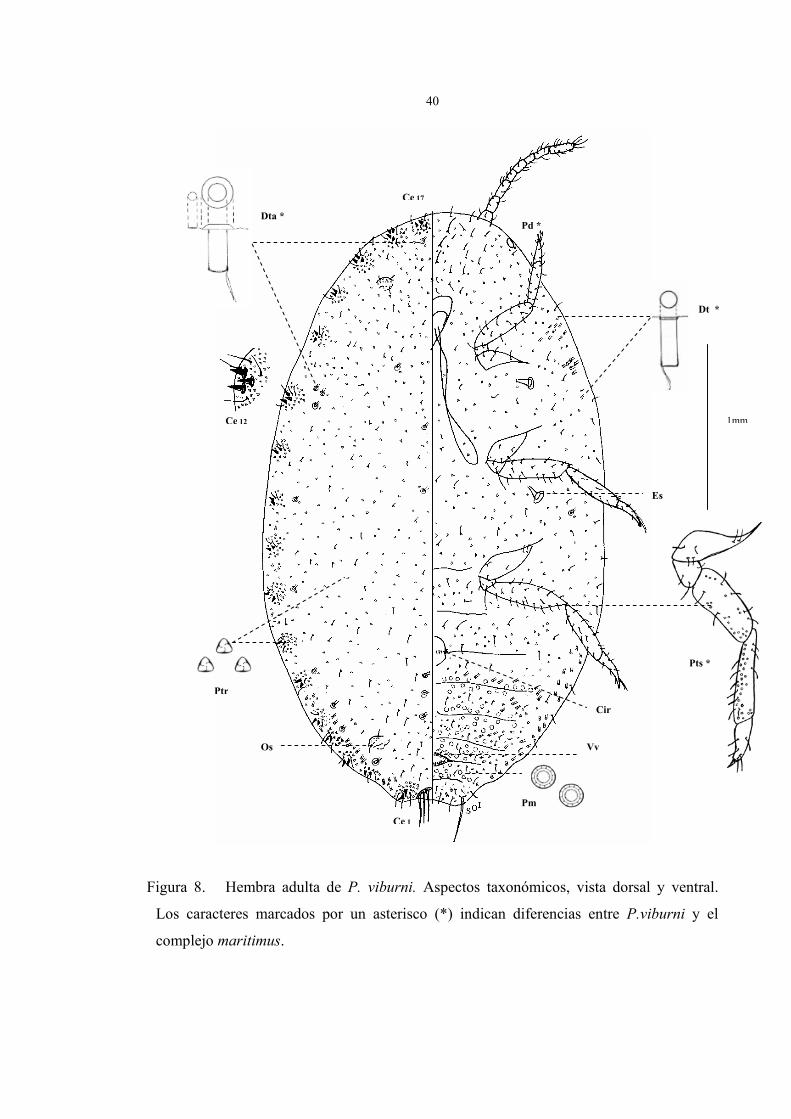
40
Figura 8. Hembra adulta de P. viburni. Aspectos taxonómicos, vista dorsal y ventral.
Los caracteres marcados por un asterisco (*) indican diferencias entre P.viburni y el
complejo maritimus.
1mm
Dt *
Dta *
Ce 17
Ce 1
Ce 12
Es
Os Vv
Pm
Cir
Pts *
Ptr
Pd *

41
Descripción del tercer estado ninfal (Fig.9)
Cuerpo de forma oval de 1,5 a 2 mm de largo por 0,7 a 1 mm de ancho, antenas de 7
segmentos, 17 (16) pares de cerarios, escasos poros y setas en comparación con la hembra
adulta, vulva aún no formada y sin poros traslúcidos en patas.
En la figura 9 están representados los caracteres taxonómicos de la ninfa de tercer
estado en vista dorsal y ventral fijado en preparaciones permanentes. Los parámetros
observados que caracterizan al estado son los siguientes:
Vista dorsal:
1. En general, 17 pares de cerarios; cerario caudal (Ce1), ligeramente esclerosado y
con mayor número de poros triloculares (Pt) rodeando las setas cónicas.
2. El número de setas cónicas (Sc), setas auxiliares (Sa) y poros triloculares (Pt) que
acompaña a cada cerario, varia según la siguiente matriz:
Ce1: 2 setas cónicas, acompañado de 2 setas auxiliares, rodeado de 25 a 30 poros
triloculares.
Ce2 al Ce9: 2 setas cónicas, acompañado de 1 a 2 setas auxiliares, rodeado de 8 a 12
poros triloculares.
Ce10: 2 setas cónicas, setas auxiliares ausentes y 2 a 5 poros triloculares rodeando
las setas.
Ce11: 2 setas cónicas acompañado de 1 a 2 setas auxiliares, rodeado de 6 a 8 poros
triloculares.

42
Ce12: 3 setas cónicas acompañado de 2 a 3 setas auxiliares, rodeado de 8 a 12
poros triloculares.
Ce13-Ce14: 2 setas cónicas acompañado de 1 a 2 setas auxiliares, rodeado de 6 a 8
poros triloculares.
Ce15-Ce16-Ce17: Cerarios cefálicos, de 3 setas cónicas acompañados de 1 a 2 setas
auxiliares, rodeado de 6 a 8 poros triloculares.
3. Los ductos tubulares anillados (Dta) (oral rim), están asociados a un poro
discoidal y sin seta auxiliar, pueden variar de 10 a 14 y se ubican frente a los cerarios
4,5,6,8,12,17 en algunas ocasiones se encuentra un Dta frente al cerario 10, en la
parte media del tórax. Al igual que el estado adulto no existe Dta entre el Cerario 15 y
el 16, a diferencia del complejo maritimus.
4. Existen abundantes poros triloculares dispersos sobre el dorso.
5. Escasos y pequeños poros discoidales (Pd), ubicados frente a cada cerario y
asociados a cada ducto tubular anillado.
6. Ausencia de poros multiloculares (Pm).
7. Setas corporales numerosas y de diversos tamaños: tres pares de setas largas se
ubican en el anillo anal; le siguen en longitud las del abdomen y las de la región
toráxica del insecto.
8. Ductos tubulares (Dt) oral-collars ausentes.

43
Vista ventral:
1. Antenas de 7 segmentos, cada segmento acompañado de 3 a 4 setas, el último
segmento de mayor longitud y mayor número de setas.
2. Circulus presente entre el segmento III y IV del abdomen.
3. Poros translúcidos (Pts) de patas posteriores ausentes.
4. Poros multiloculares (Pm) ausentes.
5. Abundantes poros triloculares dispersos sobre toda la superficie y concentrados en
un número de 22 a 70 en el segmento VI del abdomen.
6. Escasos poros discoidales dispersos en el vientre, ubicados en el lóbulo anal, en el
tórax bajo espiráculos y de 1 a 3 alrededor de cada ojo.
7. Ductos tubulares anillado y ductos tubulares oral-collars generalmente ausentes.
8. Setas más largas que las del dorso, las de mayor tamaño son las del lóbulo anal y
las del trocánter, luego las setas del abdomen y las que se ubican en la región cefálica
entre y posterior a las antenas.
9. El número de setas en las patas es de: 28 a 40 y se distribuyen de la siguiente
forma: 6 a 8 en la coxa, 2 a 4 en el trocánter, 8 a 12 en el fémur 10 a 12 en la tibia y 2
a 4 en el tarso.
10. La relación tibia/tarso es de 1,2 a 1,4.
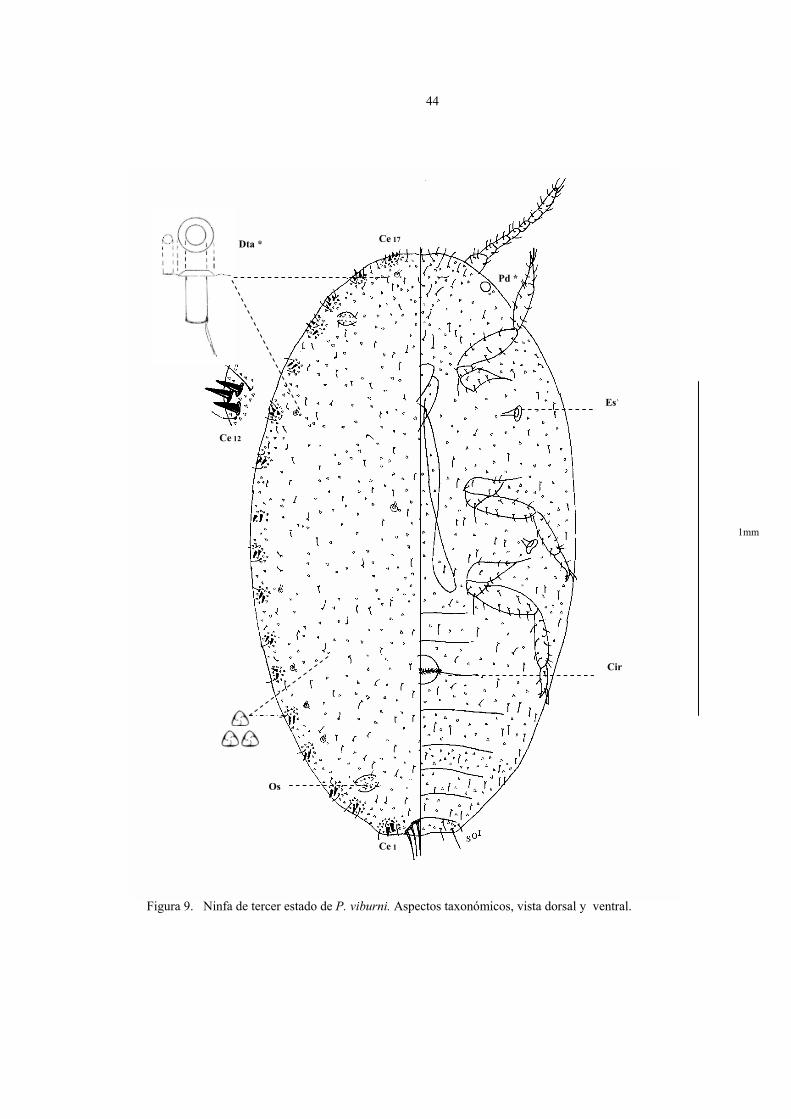
44
Figura 9. Ninfa de tercer estado de P. viburni. Aspectos taxonómicos, vista dorsal y ventral.
1mm
Pd *
Ce 17
Ce 12
Ce 1
Dta *
Es
Cir
Os

45
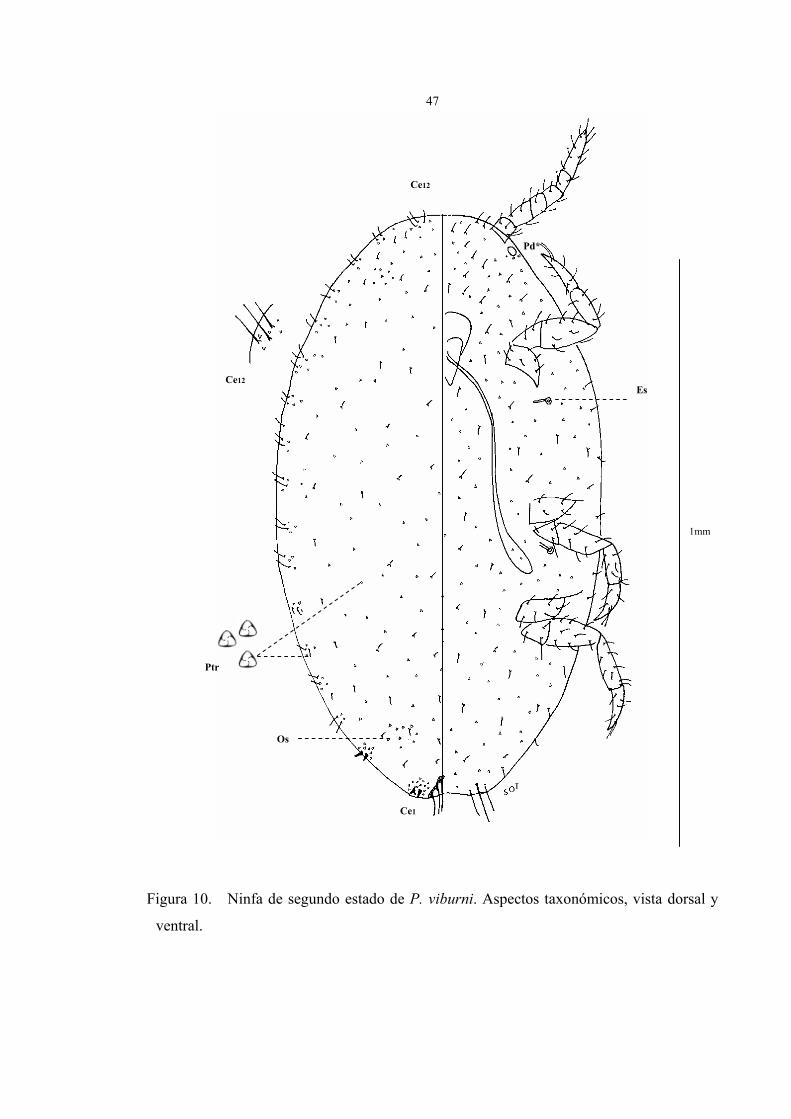
Descripción del segundo estado ninfal (Fig. 10)
Cuerpo angostamente ovalado de 1 a 1,5 mm de largo y 0,5 a 0,7 mm de ancho,
antenas robustas de 6 segmentos, espiráculos presentes y ostíolos en formación.
En la figura 10 están representados los caracteres taxonómicos de la ninfa de segundo
estado en vista dorsal y ventral fijado en preparaciones permanentes. Debe advertirse que,
no obstante se han seleccionado los caracteres que existen en esta fase ninfal de P. viburni,
no se puede asegurar su validez taxonómica para caracterizar la especie. Los parámetros
observados fueron:
Vista dorsal:
1. Los cerarios no están totalmente desarrollados, únicamente se observan como tal
el cerario caudal y el cerario paracaudal, varían según la siguiente matriz:
El cerario caudal (Ce1), presenta 2 setas cónicas rodeado de 16 a 18 poros
triloculares, sin setas auxiliares.
El cerario paracaudal (Ce2), presenta 2 setas cónicas rodeado de 6 a 8 poros
triloculares, sin setas auxiliares.
El resto de los cerarios sólo presentan 2 setas suaves y largas de 2 a 4 y poros
triloculares.
2. Ausencia de ductos tubulares anillados (Dta) y ductos tubulares collar-oral(Dt).
3. Abundantes poros triloculares y setas en toda la superficie.

46
4. Ausencia de poros discoidales y poros multiloculares.
5. Ostíolos en formación, se distingue una pequeña agrupación de poros.
Vista ventral:
1. Antenas gruesas de 6 segmentos, cada segmento acompañado de 3 a 5 setas, él
último segmento presenta la mayor longitud y el mayor número de setas.
2. Circulus no visible.
3. Ausencia de poros translúcidos y multiloculares
4. Poros discoidales de 1 a 3 alrededor de cada ojo.
5. Ausencia de ductos tubulares anillados (Dta) y ductos tubulares oral-collars (Dt).
6. Presencia de poros triloculares y setas en toda la superficie.
7. Setas más largas que las del dorso, las de mayor tamaño son las del lóbulo anal de
casi el doble de tamaño que las del resto del cuerpo, le siguen en longitud las setas del
trocánter y luego las del resto del cuerpo.
8. El número de setas en las patas es de: 30 a 42 y se distribuyen de la siguiente
forma: 4 a 6 en la coxa, 2 a 4 en trocánter, 8 a 12 en el fémur, 8 a 10 en la tibia y 8 a
10 en el tarso.
9. Los tarsos y las tibias de las patas, presentan un tamaño similar, por lo que la
relación tibia/tarso es de1.
47
Figura 10. Ninfa de segundo estado de P. viburni. Aspectos taxonómicos, vista dorsal y
ventral.
Ce12
Ce12
Ce1
Ptr
Es
Pd*
Os
1mm

48
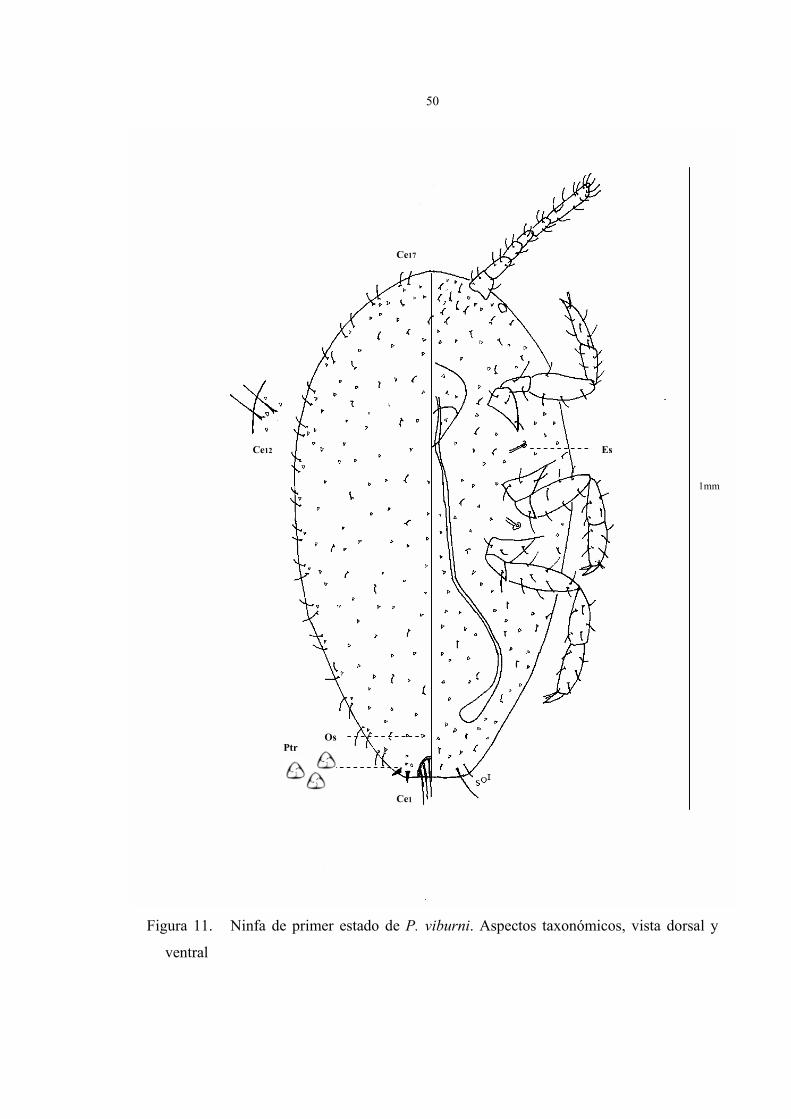
Descripción del primer estado ninfal (Fig.11)
Cuerpo oval y angosto de 0,5 a 1mm de largo y 0,25 a 0,5 mm de ancho, antenas
robustas de 6 segmentos, espiráculos muy pequeños, ostíolos y cerarios en formación.
En la figura 11 están representados los caracteres taxonómicos de la ninfa de primer
estado en vista dorsal y ventral fijado en preparaciones permanentes. Debe advertirse, al
igual que las ninfas de segundo estado, no obstante se han seleccionado los caracteres tipo
que existe en esta fase ninfal de P. viburni, no se puede asegurar su validez para
caracterizar taxonómicamente a la especie. Los parámetros observados fueron:
Vista dorsal:
Los cerarios no están totalmente desarrollados, únicamente se observa el cerario
caudal (Ce1); presenta dos setas cónicas y 3 o 4 poros triloculares.
El resto de los cerarios sólo presentan algunas setas suaves y largas de 1 a 2 y poros
triloculares.
1. Ausencia de ductos tubulares anillados (Dta) y ductos tubulares oral-collars (Dt).
2. Abundantes poros triloculares y setas en toda la superficie.
3. Ausencia de poros discoidales y poros multiloculares.
4. Ostíolos en formación, se distingue una pequeña agrupación de poros.

49
Vista ventral:
1. Antenas gruesas de 6 segmentos, cada segmento acompañado de 3 a 5 setas, el
último segmento presenta la mayor longitud y el mayor número de setas.
2. Circulus no visible.
3. Ausencia de poros: translúcidos, multiloculares y discoidales.
4. Ausencia de ductos tubulares anillados (Dta) y ductos tubulares oral-collars (Dt).
5. Presencia de poros triloculares y setas en toda la superficie.
6. Setas más largas que las del dorso, las de mayor tamaño son las del lóbulo anal
de casi el doble de tamaño que las del resto del cuerpo, le siguen en longitud las setas
del trocánter y luego las del resto del cuerpo.
7. El número de setas en las patas es de: 21 a 26 y se distribuyen de la siguiente
forma: 3 a 4 en la coxa, 1 a 2 en trocánter, 5 a 6 en el fémur, 5 a 6 en la tibia y 7 a 8
en el tarso.
8. Los tarsos son de mayor tamaño que las tibias, presentan una relación tibia/tarso
de: 0,7 a 0,8.
50
Figura 11. Ninfa de primer estado de P. viburni. Aspectos taxonómicos, vista dorsal y
ventral
Ce17
Ce12
Ce1
Es
Ptr Os
1mm

51
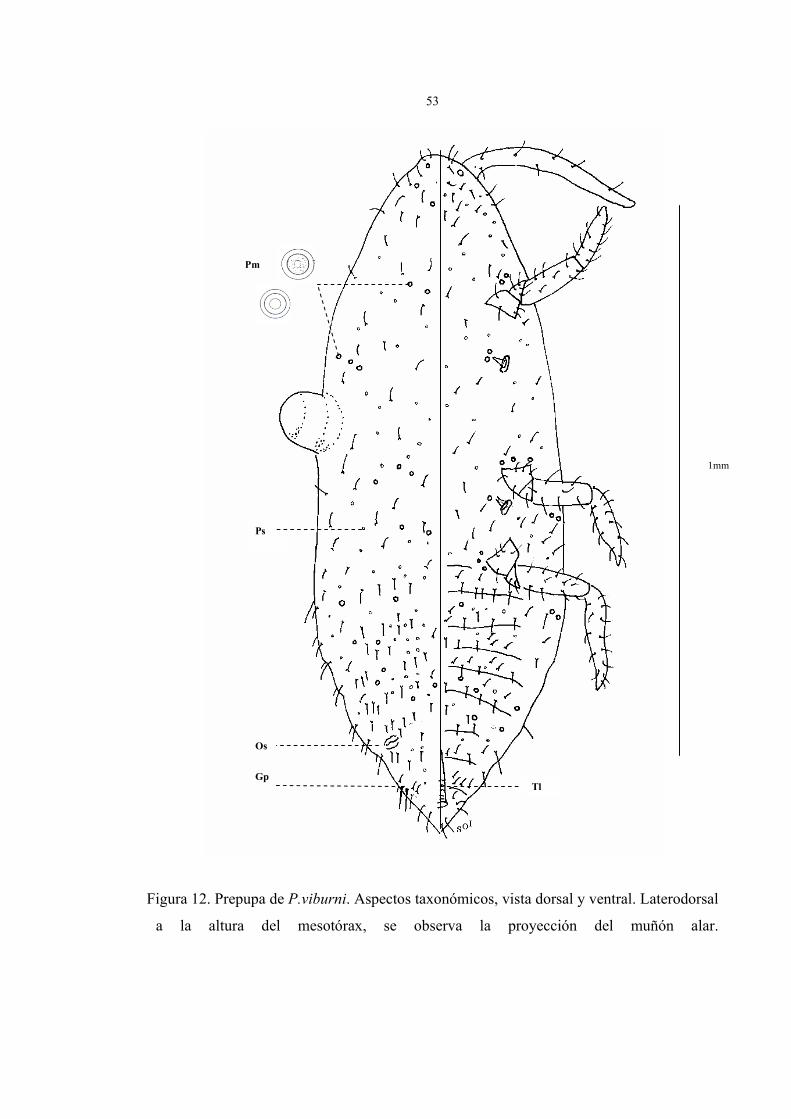
Descripción de la prepupa del macho (Fig.12)
Cuerpo alargado, estrechándose gradualmente hacia el abdomen, su tamaño fluctúa de
1 a 1,5 mm de largo por 0,2 a 0,3 mm de ancho, la cabeza no está separada del protórax,
antenas robustas, sin segmentos distinguibles, espiráculos presentes en la superficie ventral
del cuerpo.
En la figura 12 están representados los caracteres taxonómicos de la prepupa del
macho en vista dorsal y ventral fijado en preparaciones permanentes. Los parámetros
observados fueron:
Vista dorsal:
1. Pequeños muñones de alas en el tórax del insecto.
2. Bolsa glandular (Gp) en el VIII segmento, acompañada de poros, desde donde
nacen dos setas largas y gruesas en comparación al resto de las setas del cuerpo.
3. Presencia de poros simples (Ps) y poros multiloculares (Pm) sobre el dorso.
4. Abundantes setas ubicadas en cabeza, abdomen y en la región pleural del insecto,
escasas en el tórax.
5. Ausencia de ductos y ostíolos.

52
Vista ventral:
1. Antenas sin segmentos distinguibles, envuelta en una antenoteca o estuche antenal
gruesa, con 8 a 12 setas totales en la superficie.
2. 1 par de ojos simples cubiertos por setas.
3. Abdomen con 9 segmentos muy marcados, estrechándose hacia el noveno
segmento; segmentos II hasta el VII presentan abundantes setas.
4. Segmento abdominal con zona genital anchamente triangular, provisto de finas
setas y un tubo longitudinal que nace del VII segmento y se extiende hasta el último.
5. Presencia de poros simples y de mayor tamaño, con varios lóculos (Pm) en la
superficie, los poros de mayor tamaño se ubican cercano a patas, rodeando
espiráculos y uniformemente sobre el segmento II al VII del abdomen.
6. Presencia de setas en cabeza, tórax y abdomen, abundantes en cabeza y abdomen
y escasas en el tórax.
7. Dos pares de espiráculos de igual tamaño ubicados en el tórax del insecto.
8. Patas con tibia y tarso fusionados, uña ausente y setas que varían de 28 a 40.
53
Figura 12. Prepupa de P.viburni. Aspectos taxonómicos, vista dorsal y ventral. Laterodorsal
a la altura del mesotórax, se observa la proyección del muñón alar.
Es
1mm
Ps
Gp Tl
Os
Pm

54
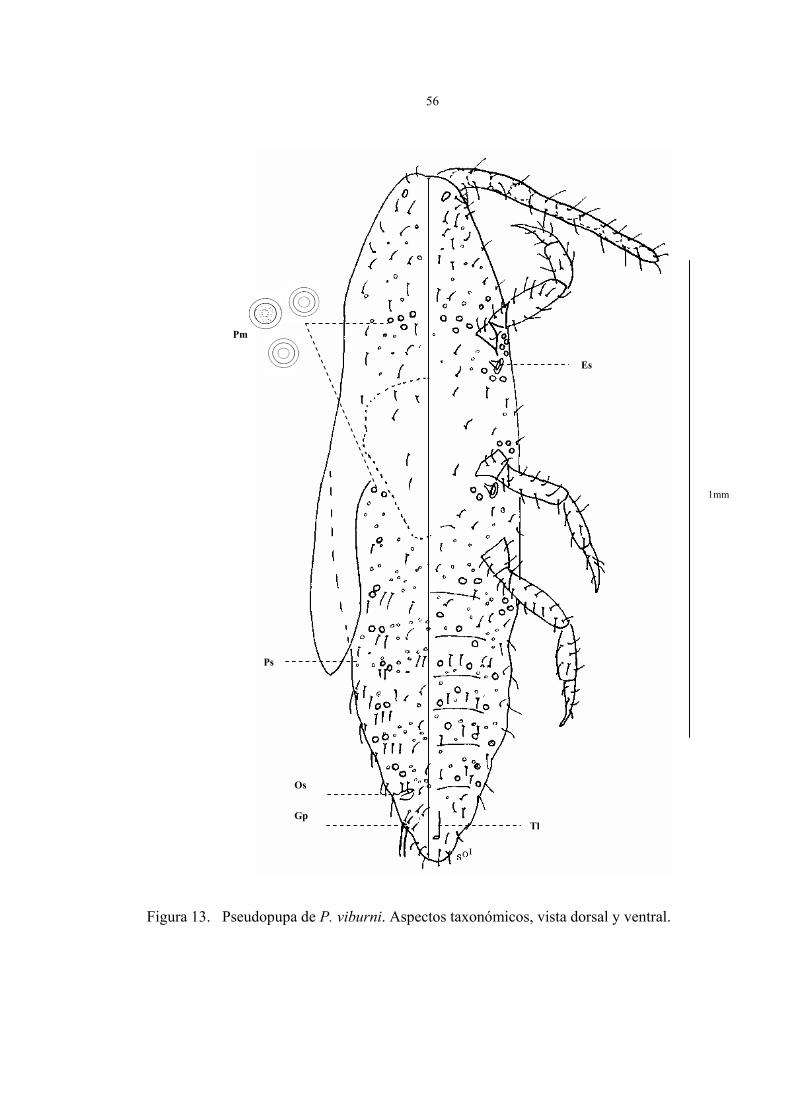
Descripción de la pseudopupa del macho (Fig.13)
Cuerpo alargado, estrechándose gradualmente hacia el abdomen, su tamaño fluctúa de
1,2 a 1,5 mm de largo por 0,2 a 0,3 mm de ancho, la cabeza no esta claramente separada
del protórax, antenas robustas dentro de un saco, espiráculos presentes en la superficie
ventral del cuerpo.
En la figura 13 están representados los caracteres taxonómicos de la pseudopupa del
macho en vista dorsal y ventral fijado en preparaciones permanentes. Los parámetros
observados fueron:
Vista dorsal:
1. 1 par de ojos simples en la superficie dorsal y ocelos laterales a ambos lados del
cuerpo, ambos rodeados de delgadas y largas setas.
2. Muñones de alas extendidos que nacen del tórax y terminan aproximadamente en
el III segmento abdominal
3. Bolsa glandular (Gp) en el VIII segmento, acompañada de poros, desde donde
nacen dos setas largas.
4. Presencia de poros simples (Ps) y poros multiloculares (Pm) sobre el dorso.
5. Abundantes setas ubicadas en cabeza, abdomen y en la región pleural del insecto,
escasas en el tórax.
6. Ausencia de ductos y ostíolos.

55
Vista ventral:
1. Antenas en antenoteca, distinguiéndose 8 segmentos, con 18 a 25 setas totales
sobre la superficie.
2. 1 par de ojos simples rodeado de setas sobre la superficie ventral del insecto.
3. Abdomen angosto con 9 segmentos muy marcados, del II hasta al VII segmento
presenta abundantes setas.
4. Segmento abdominal con zona genital anchamente triangular, similar a prepupa,
con finas setas y un tubo longitudinal que nace del VII segmento y termina en el
último.
5. Presencia de poros simples, y poros multiloculares de varios lóculos, cuyo tamaño
es notablemente superior al resto de los poros, estos últimos se ubican cercanos a las
patas, rodeando espiráculos y uniformemente sobre el segmento II al VII del
abdomen.
6. Abundantes setas en cabeza y abdomen, escasas en el tórax.
7. Dos pares de espiráculos de igual tamaño ubicados en el tórax del insecto.
8. Patas con tibia y tarso diferenciados, uña ausente y setas que varían de 32 a 40.
56
Figura 13. Pseudopupa de P. viburni. Aspectos taxonómicos, vista dorsal y ventral.
1mm
Ps
Gp
Os
Tl
Es
Pm

57
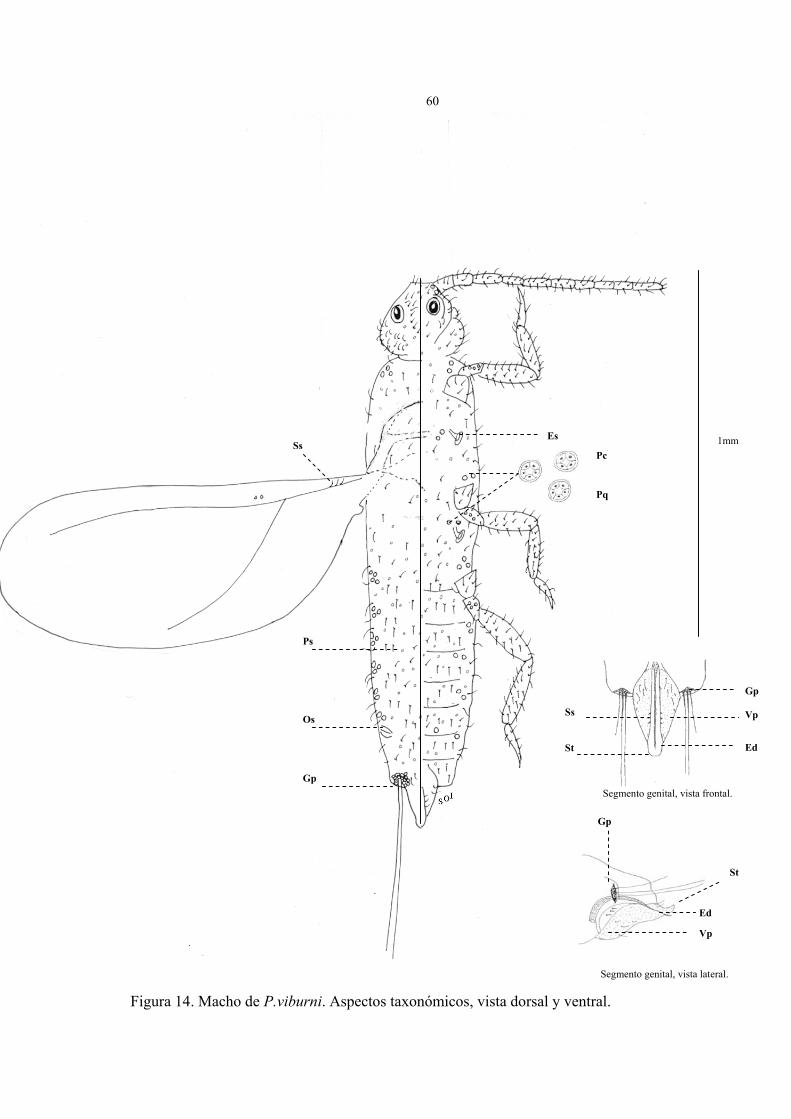
Descripción del macho (Fig. 14)
Cuerpo alargado y estrecho, su tamaño fluctúa de 1,2 a 1,5 mm de largo por 0,2 a 0,3
mm de ancho, la cabeza se encuentra separada del protórax, antenas largas de 10
segmentos, alas extendidas de aproximadamente 1,5 mm de largo, espiráculos presentes en
la superficie ventral y un par de ostíolos en el abdomen del insecto.
En la figura 14 están representados los caracteres taxonómicos del macho adulto en
vista dorsal y ventral fijado en preparaciones permanentes. Los parámetros observados que
caracterizan al estado son los siguientes:
Vista dorsal:
1. 1 par de ojos simples en la superficie dorsal y ocelos laterales, ambos rodeados
de delgadas y largas setas.
2. Alas extendidas cuyo tamaño varía de 1,5 a 2 mm de largo, su venación es muy
reducida y rudimentaria, con una vena subcostal poco evidente y corta; presentan un
lóbulo alar, 3 setas sensitivas y un par poros sensoriales.
3. Tórax esclerosado, con abundantes setas y poros simples.
4. Bolsa glandular (Gp) en el VIII segmento abdominal, con dos setas largas y
gruesas, rodeado de 15 a 30 poros quinqueloculares.
5. Presencia de poros simples, cuadriculares y quinqueloculares distribuidos sobre el
dorso, los de mayor tamaño se agrupan en la región pleural del insecto, en números
de 3 a 4 acompañados de 1 a 2 setas largas y finas.

58
6. Abundantes setas ubicadas en cabeza, tórax y abdomen. Las setas de la cabeza se
ubican entre los ojos dorsales, rodeando a ocelos y en ambas genas del insecto.
7. Presencia de 1 par de ostíolos frente al VII segmento abdominal.
8. Ausencia de ductos.
Vista ventral:
1. Antenas filiformes, delgadas de 10 segmentos, cada segmento acompañados de 6
a 14 setas. En la base de las antenas es posible observar de 2 a 3 poros
quinqueloculares.
2. Depresión preocular y 1 par de ojos simples rodeado de setas sobre la superficie
ventral del insecto.
3. Abdomen angosto con 10 segmentos muy marcados, del II al VII segmento
presenta abundantes setas.
4. Segmento genital ubicado al final del abdomen, de forma triangular y curvado
(vista lateral), vaina que envuelve el edeago acompañado de diminutas setas
sensitivas.
5. Presencia de poros simples, poros cuadriculares y quinqueloculares, en cabeza,
tórax y abdomen, los poros de mayor tamaño se ubican cercano a patas, rodeando
espiráculos y uniformemente sobre el segmento II al VII del abdomen.

59
6. Escasas setas en el tórax y abundantes en cabeza, y abdomen. Las setas de la
cabeza forman bandas transversales a ambos lados de los ojos ventrales.
7. Dos pares de espiráculos de igual tamaño ubicados en el tórax del insecto.
8. Patas diferenciadas, uña presente, 2 a 3 poros quinqueloculares en trocánter de
patas, el número de setas en las patas es de 70 a 100 y se distribuyen de la siguiente
forma: 8 a 10 en la coxa, 3 a 6 en el trocánter, 22 a 30 en el fémur 30 a 40 en la tibia
y 15 a 20 en el tarso.
9. Ausencia de ductos.
60
Ps
Gp
Os
Ss
Pq
Pc
Es 1mm
Gp Vp Ed
Ss St
Segmento genital, vista frontal.
Segmento genital, vista lateral.
Ed
St
Gp
Vp
Figura 14. Macho de P.viburni. Aspectos taxonómicos, vista dorsal y ventral.

58
Observaciones biológicas de la especie
Las observaciones que se presentan a continuación, corresponde a poblaciones de
P.viburni criadas sobre brotes de papas, en condiciones de laboratorio, con un
fotoperíodo de 12 horas y temperatura variable de 24 a 26ºC en la cámara de crianza.
La ubicación de la especie preferentemente fue en tallos, hojuelos y ocasionalmente
sobre el tubérculo.
• P. viburni, es una especie ovípara, que pasa por tres estados ninfales hasta
alcanzar el estado adulto, para el caso de las hembras y dos estadios adicionales
para los machos.
• La población inicial fue de 40 hembras adultas fertilizadas, de las cuales el 90 %
de ellas realizó su postura normalmente, el otro 10 % corresponde a: 5 % de
hembras que murieron antes de finalizar su período de postura y 5 % de falsas
posturas (la hembra en condiciones de estrés o por falta de fertilización, pone
varios huevos infértiles que se deshidratan rápidamente).
• El período de pre-postura, es decir desde que las hembras fueron fertilizadas hasta
que comenzara su proceso de postura, duró entre 3 a 21 días, con un promedio de
10 días (cuadro 7). En esos días las hembras aprovecharon de alimentarse para
posteriormente fijarse en un lugar protegido e iniciar el proceso de postura,
período en el cual deja de alimentarse.
• El proceso de postura comienza lentamente, formando un saco ovígero de fibras
blancas, algodonosas, el cual se expande hacia atrás a medida que progresa la
postura, demorándose entre 7 a 25 días, con un promedio de 17 días. Las
dimensiones de los ovisacos variaron entre 5 a 9 mm de largo, con un ancho
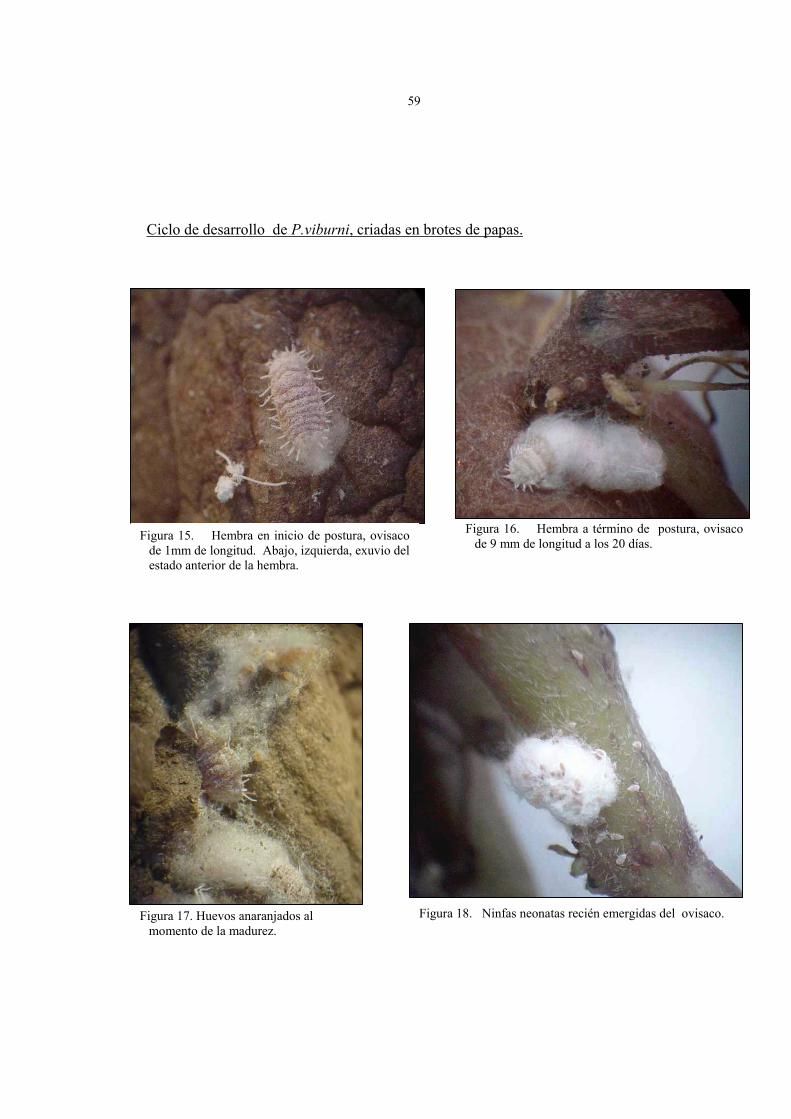
aproximado de 2 mm o igual al ancho del cuerpo de la hembra (figura 15 y 16).
59
Ciclo de desarrollo de P.viburni, criadas en brotes de papas.
Figura 15. Hembra en inicio de postura, ovisaco de 1mm de longitud. Abajo, izquierda, exuvio del estado anterior de la hembra.
Figura 16. Hembra a término de postura, ovisaco de 9 mm de longitud a los 20 días.
Figura 18. Ninfas neonatas recién emergidas del ovisaco. Figura 17. Huevos anaranjados al momento de la madurez.

60
• El período de eclosión de los huevos, es un proceso gradual, que ocurre
paulatinamente desde el extremo posterior del ovisaco hacia el cuerpo de la
hembra, el proceso demora entre 15 a 20 días, según la ubicación de los huevos.
• El número de huevos por ovisaco fluctúo entre 70 a 300, con un promedio de 200
huevos por ovisaco. Al comienzo, son de color amarillo pálido para tornarse
anaranjados al momento de la madurez (figura 17).
• Una vez eclosados los huevos, las ninfas neonatas permanecen en el saco
ovígero, junto a la hembra aún viva, entre 3 a 7 días, con un promedio de 4 días,
posteriormente emergen del saco, dispersándose masivamente por un par de días,
con un notable fototropismo positivo (figura 18).
• Una vez terminada su dispersión y elegido su lugar de fijación, las ninfas
neonatas se fijan preferentemente a lo largo de los brotes y comienzan a
alimentarse. Aproximadamente a los 10 días de nacidas, comienzan a secretar una
cera pulverulenta blanca, que cubre completamente su cuerpo y que da por
finalizado el primer estado de desarrollo (figura 19). Al término de la fase se
produce la ecdisis liberándose del exuvio por la parte posterior del cuerpo.
• El segundo estado ninfal dura aproximadamente entre 4 a 10 días, con un
promedio de 7 días y es de gran importancia, pues ocurre la diferenciación de los
sexos. En efecto al avanzar en el segundo estadio se aprecia una notable
diferencia de color entre las ninfas hembra y las ninfas macho, estas últimas
presentan una coloración blanca grisácea, mientras que las ninfas femeninas
presentan un ligero color rosado (Figura 20).

61
• Dada la marcada diferencia de color de las ninfas, es posible cuantificar la
relación de hembras por macho, que bajo las condiciones del ensayo alcanzó
valores de 2 : 1.
• Posteriormente las ninfas hembra pasan a un tercer estado que en promedio dura
10 días (figura 21). En esta fase se observan con mayor precisión los exuvios muy
blancos y de forma triangular que quedan abandonados a lo largo de la superficie
del brote.
• Las hembras alcanzan su estado adulto luego de unos 30 días de desarrollo; los
machos aparecen casi en igual fecha, lo que demuestra cierto tipo de
sincronización biológica entre los sexos (figura 22).
Cuadro 7. Duración promedio de los estados de desarrollo, de P. viburni, bajo
condiciones de laboratorio a 25ºC y fotoperíodo de 12 horas.
Estados de desarrollo Duración promedio(días)
Hembra Macho
Hembra prepostura 10 (3-21) 10 (3-21)
Hembra en postura 17 (10-24) 17 (10-24)
Ninfas de primer estado 10 (8-12) 10 (8-12)
Ninfas de segundo estado 7 (4-10) 7 (4-10)
Ninfas de tercer estado 10 (7-11) ----
Prepupa de machos ---- 7 (4-10)
Pseudopupa de machos ---- 7 (7-11)
Duración acumulada hasta alcanzar el estado adulto 27 (19-33) 31 (23-43)
Duración Ciclo completo de la hembra: 54 (29-79)
Los valores entre paréntesis indican rango de variación de los distintos estados de desarrollo.

62
Estados de desarrollo de hembra P. viburni, criadas en brotes de papas.
Figura 19. Ninfas de 1º estado. Figura 20. Ninfas de 2º estado.
Figura 21. Ninfas de 3º estado. Figura 22. Hembras adultas.

63
• Las ninfas macho una vez terminado su segundo estado, se encapullan formando
grupos, en un cilindro lanoso de unos 4 mm de longitud, durante unos 11 a 20
días, en el interior del capullo, pueden observarse dos estadios: prepupoide y
pseudopupa, con duración promedio de 7 días cada uno (figura 23, 24 y 25).
• El proceso de fecundación de las hembras se lleva a cabo rápidamente dada la
corta vida del macho. (figura 26)
• La longevidad de los machos es variable de 2 a 6 días, con un promedio de 3 días,
en los cuales demuestra una notable eficiencia reproductiva.
• Respecto al ensayo de eficiencia reproductiva de los machos, en todos los
tratamientos, la totalidad de las hembras fue fecundada y el macho alcanzó un
valor promedio de fecundación de 6 hembras/día. (cuadro 8).
Cuadro 8. Ensayo de eficiencia reproductiva del macho, bajo condiciones de
laboratorio.
Tratamiento Nº de machos Nº de hembras Duración
(días)
Nº Promedio de hembras
fecundadas/día
Eficiencia
%
T1 1 10 2 5 100
T2 1 20 4 5 100
T3 1 30 4 7,5 100
T4 1 40 6 6,6 100

64
Estados de desarrollo de macho P.viburni, criadas en brotes de papas.
Figura 25. Macho adulto alado. Tamaño aprox. es de 1,5 mm
Figura 23. Capullos de machos, junto a ninfas de 2º estado.
Figura 24. Prepupas y pseudopupas de macho en alcohol.
Figura 26. Acoplamiento sexual de P. viburni

65
Características macroscópicas e identificación a nivel de campo de los estados de
desarrollo de Pseudococcus viburni
Huevos: Los huevos son depositados en sacos algodonosos blancos y presentan un
tamaño de 0,36 a 0,42 mm de largo por 0,23 a 0,26 mm de ancho con un color
amarillo variando a anaranjado al momento de la madurez (Cuadro 9 y 10).
Ninfa neonatas: Las ninfas recién eclosadas, miden aproximadamente 0,5 mm de
longitud, presentan una coloración rosada, sin pulverulencia blanca y corresponden al
inicio del primer estado.
Primer estado ninfal: La longitud total del cuerpo varia de 0,5 a 1mm de largo y 0,25
a 0,5 mm de ancho, la longitud de antenas es variable en relación al tamaño del
cuerpo desde 30 hasta un 15 %. Respecto a los filamentos marginales (que van en
correspondencia a los cerarios) solo es posible distinguir a simple vista el par caudal
(correspondiente al Ce 1) de 0,1 a 0,3 mm de largo. La coloración del cuerpo es
ligeramente rosado, cubierto por una secreción pulverulenta de color blanco.
Segundo estado ninfal: Su tamaño varía de 1 a 1,5 mm de largo por 0,5 a 0,7 mm de
ancho, las antenas presentan una longitud menor a los 0,3 mm. A simple vista es
posible distinguir: el par de filamentos paracaudales (segundo par de atrás hacia
delante) con un rango de 0,2 a 0,3 mm de largo y el par caudal cercano a los 0,5 mm
de largo, el resto de los filamentos marginales no se logra apreciar. El cuerpo esta
cubierto por una secreción pulverulenta blanca, bajo ella se aprecia el color del
cuerpo, rosado para las ninfas hembras y ligeramente gris para las ninfas machos.

66
Tercer estado ninfal (solo para hembras): Longitud total del cuerpo de 1,5 a 2 mm de
largo y 0,7 a 1 mm de ancho; las antenas miden entre 0,3 a 0,5 mm de largo. Todos
los filamentos marginales se pueden apreciar a simple vista: el par paracaudal
presenta un tamaño variable de 0,3 a 0,5 mm de longitud, el par caudal alcanza
valores cercanos a 1 mm de longitud y los restantes 15 pares de filamentos
marginales miden aproximadamente 0,2 mm. La coloración del cuerpo es ligeramente
rosado, cubierto por secreción pulverulenta blanca.
Hembra adulta: El cuerpo de la hembra es largamente ovalado y aplastado
dorsoventralmente, fluctuando entre 2 a 4 mm de largo, y 1 a 2 mm de ancho,
dependiendo de su alimentación. La longitud de las antenas es equivalente al 25 %
del cuerpo, aproximadamente 0,5 mm. Respecto a los filamentos marginales es
posible distinguir a simple vista los 17 pares: el par paracaudal mide de 0,6 a 0,7
mm, el par caudal mide de 1 a 2 mm, equivalentes aproximadamente al 50% del
cuerpo, los restantes 15 pares marginales pueden variar de 0,2 a 0,4 mm de longitud.
La coloración del cuerpo es ligeramente rosado cubierto por secreción pulverulenta
blanca.
Machos: Terminado el segundo estado ninfal, las ninfas se encapullan para formar el
estado de prepupa, donde deja su apariencia de ninfa pasando a un estado alargado y
segmentado, de aproximadamente 1,2 mm de longitud, patas y antenas cortas y
pequeños muñones de alas. Posteriormente, dentro del capullo también, se desarrolla
un estado de pseudopupa, cuyo cuerpo es mas alargado y segmentado que el estado
anterior de 1,2 a 1,5 mm de longitud, antenas gruesas y largas envueltas en un exuvio
y largos muñones de alas. En el interior del capullo prosigue el desarrollo de las alas
y filamentos caudales, dando origen a un insecto alado de color café grisáceo de 1,2
a 1,5 mm de largo por 0,2 a 0,3 mm de ancho, un tamaño de antenas cercano a los
0,5 mm y un par de filamentos caudales superior al 50 % del largo total del cuerpo.
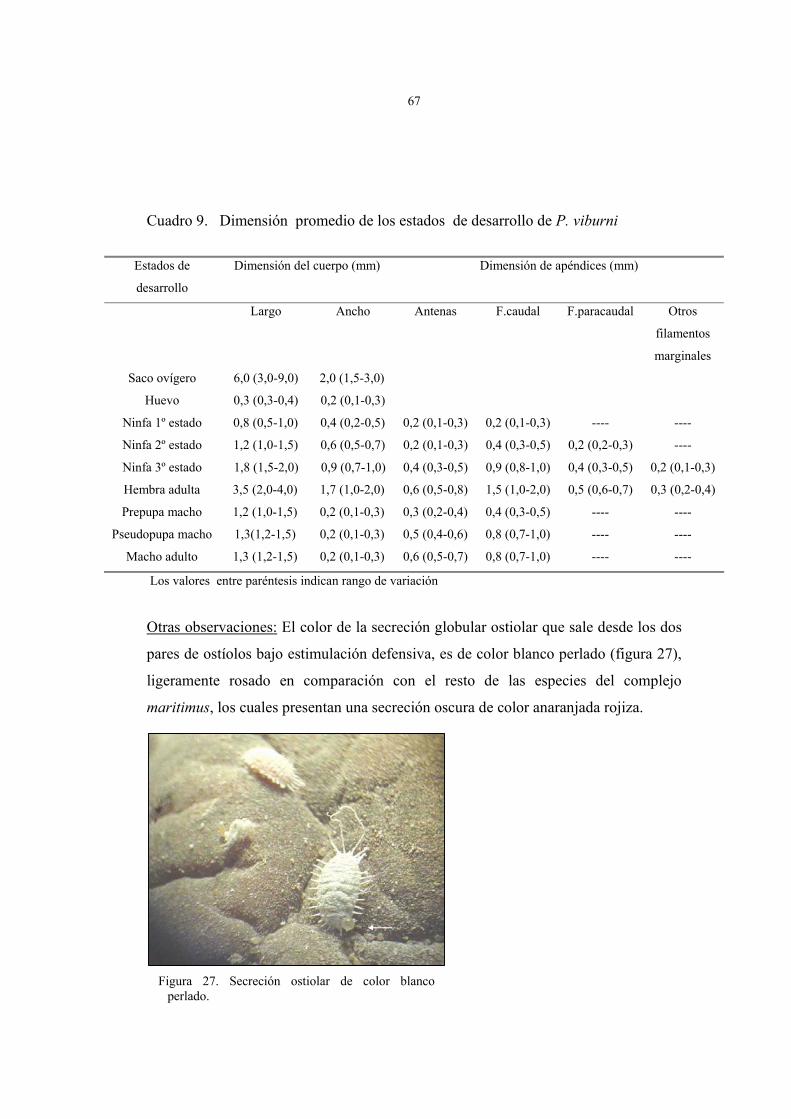
67
Cuadro 9. Dimensión promedio de los estados de desarrollo de P. viburni
Estados de
desarrollo
Dimensión del cuerpo (mm)
Dimensión de apéndices (mm)
Largo Ancho Antenas F.caudal F.paracaudal Otros
filamentos
marginales
Saco ovígero 6,0 (3,0-9,0) 2,0 (1,5-3,0)
Huevo 0,3 (0,3-0,4) 0,2 (0,1-0,3)
Ninfa 1º estado 0,8 (0,5-1,0) 0,4 (0,2-0,5) 0,2 (0,1-0,3) 0,2 (0,1-0,3) ---- ----
Ninfa 2º estado 1,2 (1,0-1,5) 0,6 (0,5-0,7) 0,2 (0,1-0,3) 0,4 (0,3-0,5) 0,2 (0,2-0,3) ----
Ninfa 3º estado 1,8 (1,5-2,0) 0,9 (0,7-1,0) 0,4 (0,3-0,5) 0,9 (0,8-1,0) 0,4 (0,3-0,5) 0,2 (0,1-0,3)
Hembra adulta 3,5 (2,0-4,0) 1,7 (1,0-2,0) 0,6 (0,5-0,8) 1,5 (1,0-2,0) 0,5 (0,6-0,7) 0,3 (0,2-0,4)
Prepupa macho 1,2 (1,0-1,5) 0,2 (0,1-0,3) 0,3 (0,2-0,4) 0,4 (0,3-0,5) ---- ----
Pseudopupa macho 1,3(1,2-1,5) 0,2 (0,1-0,3) 0,5 (0,4-0,6) 0,8 (0,7-1,0) ---- ----
Macho adulto 1,3 (1,2-1,5) 0,2 (0,1-0,3) 0,6 (0,5-0,7) 0,8 (0,7-1,0) ---- ----
Los valores entre paréntesis indican rango de variación
Otras observaciones: El color de la secreción globular ostiolar que sale desde los dos
pares de ostíolos bajo estimulación defensiva, es de color blanco perlado (figura 27),
ligeramente rosado en comparación con el resto de las especies del complejo
maritimus, los cuales presentan una secreción oscura de color anaranjada rojiza.
Figura 27. Secreción ostiolar de color blanco perlado.

68
Cuadro 10: Principales características macroscópicas para identificación preliminar
de campo por estado de desarrollo de P. viburni
Estado de desarrollo Principales características
Huevos Saco lanoso de color blanco, con huevos amarillos
variando a anaranjados.
Ninfa neonata ≤ 0,5 mm, de color rosado, sin secreción pulverulenta
blanca sobre el cuerpo.
Ninfa 1º estado ≤ 1,0 mm, con secreción pulverulenta blanca sobre su
cuerpo ligeramente rosado, solo con filamentos caudales
visibles a simple vista.
Ninfa 2 º estado hembra ≤ 1,5 mm, con secreción pulverulenta blanca sobre su
cuerpo ligeramente rosado, con filamentos caudales y
paracaudales visibles a simple vista.
Ninfa 2 º estado macho ≤ 1,5 mm, con secreción pulverulenta blanca sobre su
cuerpo grisáceo, con filamentos caudales y paracaudales
visibles a simple vista.
Ninfa de 3º estado ≤ 2,0 mm, cuerpo ligeramente rosado cubierto por una
secreción pulverulenta blanca, con todos los filamentos
visibles a simple vista.
Hembra adulta ≥ 2,0 mm cuerpo ligeramente rosado cubierto por una
secreción pulverulenta blanca, con todos los filamentos
visibles a simple vista. Al apretar el cuerpo sale una
secreción de color blanca perlada.
Prepupas, Pseudopupas y Macho adulto Raramente visibles en el campo.
Nota: Para las mediciones taxonómicas micro y macroscópicas se evalúo un número
de 100 ejemplares por cada estado de desarrollo de la especie. La relación relativa del
tamaño de los estados de desarrollo se presenta en la figura 28 y 29.
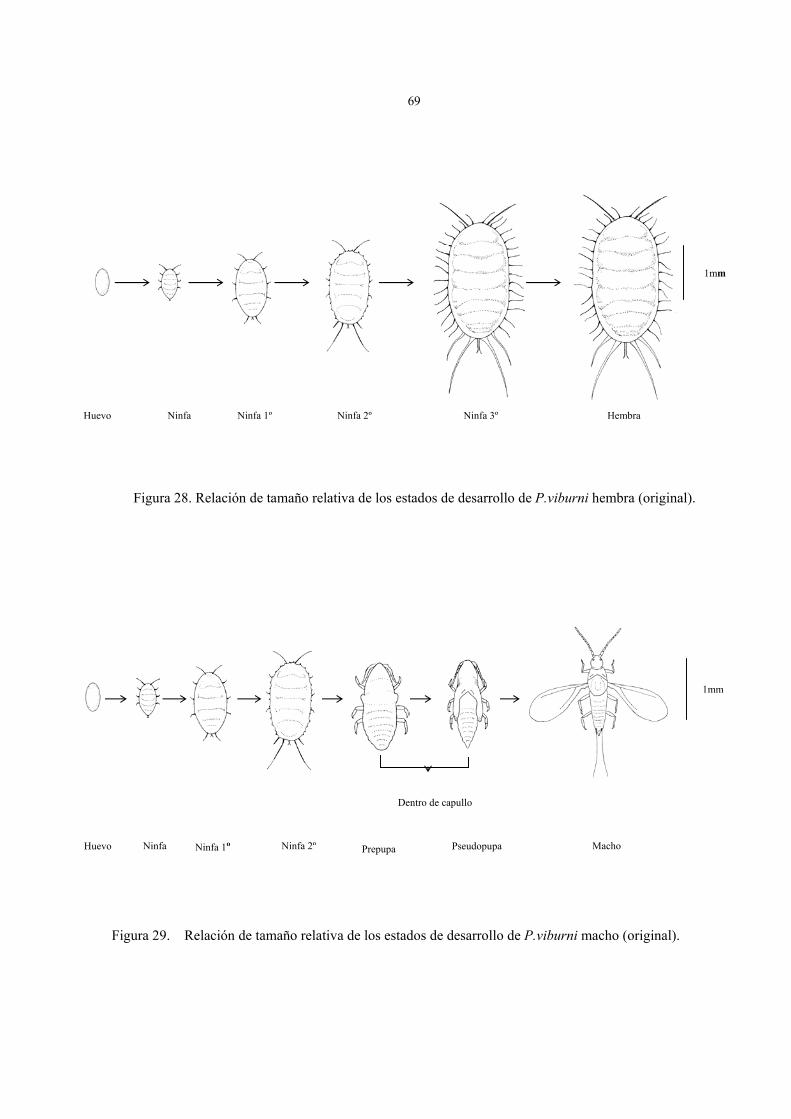
69
Huevo Ninfa Ninfa 1º Ninfa 2º Ninfa 3º Hembra
1mm
Figura 28. Relación de tamaño relativa de los estados de desarrollo de P.viburni hembra (original).
Huevo Ninfa Ninfa 1º Ninfa 2º Prepupa Pseudopupa Macho
Dentro de capullo
1mm
Figura 29. Relación de tamaño relativa de los estados de desarrollo de P.viburni macho (original).

70
CONCLUSIONES
• Para describir taxonómicamente todos los estados de desarrollo de P.viburni y
separarlos del complejo maritimus asociado, se seleccionó un conjunto de
caracteres específicos, que conforman una matriz de identificación.
• Los componentes de esta matriz fueron por primera vez utilizados para la
población de P. viburni presente en Chile e incluyen parámetros básicos
considerados en la identificación microscópica de los estados ninfales de la
especie.
• Los estados que permiten una adecuada identificación taxonómica de la especie se
refieren a ninfas de tercer estado y hembras adultas, las cuales presentan un
importante número de caracteres específicos que permitieron elaborar claves de
reconocimiento.
• La identificación de ninfas de primer y segundo estado, si bien debe ser conocida
para compararlas con otras especies, no permiten por si solas caracterizar
taxonómicamente a la especie P. viburni, fundamentalmente debido al escaso
número de caracteres presentes en dichos estados.
• Respecto a la identificación taxonómica de prepupa, pseudopupa y macho adulto,
tampoco permiten reconocer por si solas a la especie, dada su gran complejidad
taxonómica y la falta de claves adecuadas de identificación.
• Con relación a las observaciones biológicas sobre el insecto, se determinó la
duración media del ciclo, número de huevos promedio por ovisaco, secuencia de
nacimientos de neonatas, período de desarrollo de cada estadio ninfal, etc.

71
• Específicamente respecto a la secuencia en que ocurre la eclosión de los huevos y
posterior emergencia de neonatas, se observó un desfase aproximado de 20 días
entre estados de desarrollo que generó un traslape generacional.
• Se observó diversos aspectos de la biología del macho, bajo condiciones de
laboratorio, se cuantificó una alta población de machos viables en todos sus
estados de desarrollo y altamente eficaces en su reproducción, se sugiere verificar
y comparar resultados a nivel de campo.
• Respecto a la caracterización macroscópica de la especie, sólo la hembra adulta
permite establecer una identificación preliminar en el campo, basada en el color
del cuerpo, longitud de los filamentos marginales y color de la secreción ostiolar,
no obstante deba complementarse con estudios microscópicos.

72
LITERATURA CITADA
AFIFI, S. 1968. Morphology and taxonomy of the adult males of families
Pseudococcidae and Eriococcidae (Homoptera: Coccoidea). Bulletin of the British
Museum (Natural History) Entomology. Supplement 13, London, 210 p.
ARTIGAS, J.N. 1994. Entomología Económica. Insectos de interés agrícola,
forestal, médico y veterinario (nativos, introducidos y susceptibles de ser
introducidos), Vol. 1. Ediciones Universidad de Concepción, Concepción, 1129 p.
ASOCIACION DE EXPORTADORES DE CHILE A.G. 2000. Documento
estadístico temporada 1999-2000. Santiago, Chile. 166 p.
CAMPOS, L. 1953. Plagas Entomológicas de la agricultura en Chile. Cart. 49, Dept.
San. Veg. Min. Agric. Chile.16 p.
CIAMPOLINI, M., LUNGHINI, D. y MOCETTI, G. 2003. Insidioso nemico della
frutticoltura: Pseudococcus viburni. L’ Inf. Agrario. 1/ 2003: 57-60.
ETCHEVERRY, M. y HERRERA, J. 1972. Curso teórico práctico de Entomología.
Ed Universitaria. Santiago, 356 p.
GIMPEL, W.F. & MILLER, D.R. 1996. Systematic analysis of the mealybugs in the
Pseudococcus maritimus complex (Homoptera: Pseudococcidae), Contrib. Entom.
Internac. 2 (1), 163 p.

73
GONZALEZ, R.H. 1983a. El chanchito blanco de la uva de mesa. Revista Frutícola
4(1): 3-7.
GONZALEZ, R.H. 1983b. Manejo de plagas de la vid. Santiago. Facultad de
Ciencias Agrarias y Forestales, Universidad de Chile. Publicaciones Ciencias
Agrícolas Nº 13.
115 p.
GONZALEZ, R.H. 1989. Insectos y ácaros de importancia agrícola y cuarentenaria
en Chile. Edit. Ograma, Santiago, Chile.310 p
GONZALEZ, R.H. 1991. Chanchitos Blancos (Homoptera: Pseudoccoccidae). Una
nueva plaga de ciruelos en Chile. Revista Frutícola. 12(1): 3-7.
GONZALEZ, R.H. 1994. Cuarentena Internacional y exportaciones Chilenas
Frutícolas. Revista Frutícola. 15 (2): 53-57.
GONZALEZ R.H. 2003a. Chanchitos blancos de importancia agrícola y
cuarentenaria en huertos frutales de Chile (Hemiptera: Pseudococcidae). Revista
Frutícola 24(1): 5-17.
GONZALEZ R.H. 2003b. Manejo cuarentenario de chanchitos blancos en
pomáceas en Chile (Hemiptera: Pseudococcidae). Revista Frutícola 24(3): 89-98.
GONZALEZ R.H. 2003c. Programa de control del chanchito blanco de frutales,
Pseudococcus viburni (Signoret) en uva de mesa y pomáceas de exportación a
México. Programa ASOEX. Santiago, 2003.

74
GONZALEZ, R.H, ARRETZ, P. y CAMPOS, L. 1973. Catálogo de las Plagas
Agrícolas de Chile. Santiago, Univ. de Chile. Facultad de Ciencias Agrarias y
Forestales. Publicaciones Ciencias Agrícolas Nº 2. 68 p.
GONZALEZ, R.H, POBLETE, G., BARRIA, P. 2001. El chanchito blanco de los
frutales en Chile, Pseudococcus viburni (Signoret), (Homoptera: Pseudococcidae).
Revista Frutícola 22(1): 17-26.
MCKENZIE, H.L 1967. Mealybugs of California. Berkeley. University of
California. 526 p.
MARTINI B, A. 1989. Identidad, fenología y biología del chanchito blanco de la
vid en dos regiones de Chile Central. Memoria Ing.Agr. Universidad de Chile. Fac.
Ciencias Agr. y For. Santiago, 1986, 79 p.
MILLER, D.R., GILL, R. & D.J WILLIAMS. 1984. Taxonomic Analysis of
Pseudococcus affinis (Maskell), a seniors synonym of Pseudococcus obscurus Essig,
and comparisson with Pseudococcus maritimus (Ehrhorm) (Homoptera: Coccoidea:
Pseudococcidae) Proc.Ent.Soc.Wash. 86 (3): 703-713.
PRADO, E. 1991. Artrópodos y sus enemigos naturales asociados a plantas
cultivadas en Chile. Instituto de Investigaciones Agropecuarias. Serie Boletín Técnico
Nº 169. Publicaciones Estación Experimental “ La Platina”, Santiago. 207 p.
RIPA, R, ROJAS, S. 1990a. Chanchitos blancos en parronales: ¿Problema o
Manejo?. Investigación y Progreso Agropecuario. La Platina 61: 18-26.
RIPA, R, ROJAS, S. 1990b. Manejo y control biológico del chanchito blanco de la
vid. Revista Frutícola.11(3) p. 82-87

75
RIPA, R, ROJAS, S. 1993. Nuevos avances en el manejo del chanchito blanco de la
vid. Investigación y Progreso Agropecuario La Platina 76: 28-30.
RIPA, R., RODRIGUEZ, F; ROJAS, S. 1992a. Control biológico y químico del
chanchito blanco de la vid. Aconex 38: 17-22.
RIPA, R., RODRIGUEZ, F; ROJAS, S. 1992b. Chanchito blanco de la vid: Nuevos
antecedentes para su control. Investigación y Progreso Agropecuario La Platina 71: 8-
21.
RIPA, R., RODRIGUEZ, F. 1999. Plagas de cítricos, sus enemigos naturales y
manejo. Instituto de Investigaciones Agropecuarias. Colección libros INIA Nº 3. 151
p.
STORER, T.I, USINGER, R.L. 1962. Zoología General. Barcelona, Editorial
Omega.1003 p.
WILLIAMS, DJ & C. GRANARA DE WILLINK. 1992. Mealybugs of Central and
South America. U.K. CAB, Internacional, 635 p.
YUDELEVICH, M. 1950. Control biológico de los Pseudoccocus en Chile. Tesis
(Ing Agr) Universidad de Chile. Esc. de Agronomía, 126 p.

76
APÉNDICES

77
APÉNDICE I
Abreviaturas y glosario de términos usados en la matriz de caracterización taxonómica de
P. viburni.
Para facilitar la identificación de los caracteres taxonómicos utilizados en la
descripción de la especie, fue necesario confeccionar un glosario, basado en trabajos
anteriores diseñados para otras especies (Afifi, 1968, Miller et al, 1984, Williams &
Granara de Willink, 1992, Gimpel & Miller, 1996).
Dentro de la matriz, existen caracteres comunes y caracteres específicos (cuadro 6),
marcados por asterisco (*) en los esquemas de identificación (figuras 8 y 9).
Anillo anal (Aa): Area esclerosada semejando un anillo, provista de 6 setas largas en
comparación al resto de las setas del cuerpo.
Ano: Ubicado dorsalmente en el noveno segmento abdominal, delimitado por el anillo anal.
Bolsa glandular (glandular pouch) (Gp): Estructura glandular rodeada de poros, desde
donde se origina un par de setas de gran tamaño. Ubicado en el VIII segmento abdominal
de los estados de desarrollo del macho.
Cerarios (Ce): Diminutas aberturas u orificios microscópicos a través de los cuales se
excreta cera. También se les llama poros cerosos o poros glandulares. En concordancia a
los cerarios se desprenden los 17 pares de filamentos cerosos marginales que identifican el
género.

78
En las figuras, cada cerario está acompañado por un número, indicando su posición, a
partir del cerario caudal, el cual corresponde al Ce 1 y así sucesivamente por el margen
hacia delante hasta la zona frontal.
Circulus (Cir): Estructura rectangular u oval sobre el vientre, ubicada entre el segmento III
y el IV. Aparentemente es usada como sostén rápido para asirse al sustrato.
* Ducto tubular anillado (Dta) (Oral-rim tubular duct): Conductos con anillos que
vehiculizan la cera, generalmente se acompañan de un poro discoidal y/o una seta auxiliar.
Muy importantes desde el punto de vista taxonómico, representa un carácter especifico de
la especie, pues su número y ubicación, permiten separar a P.viburni del complejo
maritimus.
* Ducto tubular (Dt) (Oral- collar tubular duct): Conductos sin anillos que cumplen la
misma función que los ductos anteriores, también considerado como carácter especifico de
la especie y de importancia taxonómica.
Edeago (Ed): Órgano copulador del macho.
Espiráculos (Es): Orificios pares en correspondencia con las tráqueas, ubicados en el tórax
bajo el primer y el segundo par de patas respectivamente.
Lóbulo anal (Lob): Proyecciones como paletas situadas en el extremo caudal del abdomen
a cada lado del anillo anal. Generalmente presenta una seta de gran tamaño acompañada de
par de setas auxiliares.
Ocelos (Oc): Órgano visual unifacetado. Ojo simple. Presente en estados de desarrollo del
macho.

79
Oral-collar tubular duct: Ducto tubular.
Oral-rim tubular duct: Ducto tubular anillado.
Ostíolos (Os): Abertura de la cara dorsal del cuerpo, en forma de labio; en las hembras son
dos pares; el primer par se ubica entre la cabeza y el tórax, entre el cerario 14 y 15 y el
segundo par frente al cerario 3, entre los segmentos abdominales VI y VII. En los machos
existe solo un par abdominal. Al ser estimulados emite a través de ellos una secreción de
color variable, blanca perlada para el caso de P.viburni.
Poros : Abertura que produce secreción filamentosa del cuerpo existen: Poros simples (Ps)
y complejos. Dentro de estos últimos se encuentran:
Poros cuadriculares (Pc): Aberturas de 4 lóculos, presente en los estados de desarrollo del
macho.
* Poros discoidales (Pd): Abertura sin lóculos separados. Considerado un carácter
especifico de la especie, son taxonómicamente muy importantes aquellos que se ubican
rodeando el borde del ojo; junto con otros caracteres permiten la identificación de la
especie.
Poros multiloculares (Pm): Aberturas grandes con muchos lóculos
Poros trilocular (Ptr): Aberturas con 3 lóculos
* Poros translúcidos (Pts): Aberturas como manchas trasparentes, ubicadas en fémur y
tibias de patas posteriores, el rango o número de ellos permite la identificación de la
especie. En otras especies pueden existir en la región coxal.

80
Poros quinqueloculares (Pq): Aberturas de 5 lóculos, presente en los estados de desarrollo
del macho.
Setas: Estructuras implantadas en la cutícula, derivadas de células hipodermales especiales.
La disposición de las setas es característica para cada especie. Existen distintos tipos:
Setas cónicas (Sc), Setas auxiliares (Sa); ambas ubicadas junto a cada cerario, Setas
sensitivas (Ss) ubicadas en las alas y en la estructura genital del macho y setas comunes que
se distribuyen sobre ambas superficies del cuerpo.
Stylus (St): Terminalia de estructura genital del macho.
Tubo longitudinal (Tl): Ubicado en el segmento genital del macho, nace en el VII segmento
abdominal y termina en el último.
Vaina del pene (Vp): En la genitalia masculina es la estructura que cubre el pene.
Vulva (Vv): Abertura exterior del órgano sexual femenino. Se ubica entre el séptimo y
octavo segmento abdominal.

81
APÉNDICE II
Fotografías macro y microscópicas de los estados de desarrollo de P.viburni
Figura a. Hembras de P. viburni en alcohol.
Figura d. Restos de muda en sustrato de crianza. Figura c. Ninfas de tercer estado mudando a adulta.
Figura b. Secreción ostiolar blanca perlada de P. viburni.
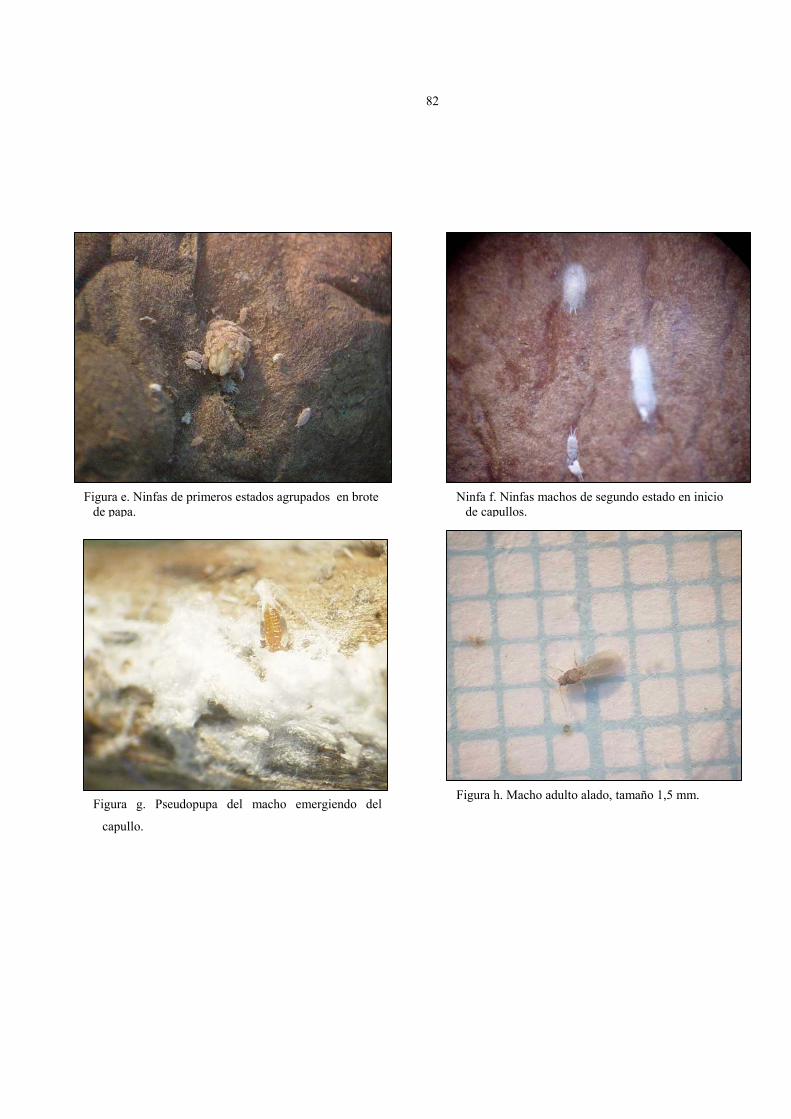
82
Figura g. Pseudopupa del macho emergiendo del
capullo.
Figura h. Macho adulto alado, tamaño 1,5 mm.
Figura e. Ninfas de primeros estados agrupados en brote de papa.
Ninfa f. Ninfas machos de segundo estado en inicio de capullos.
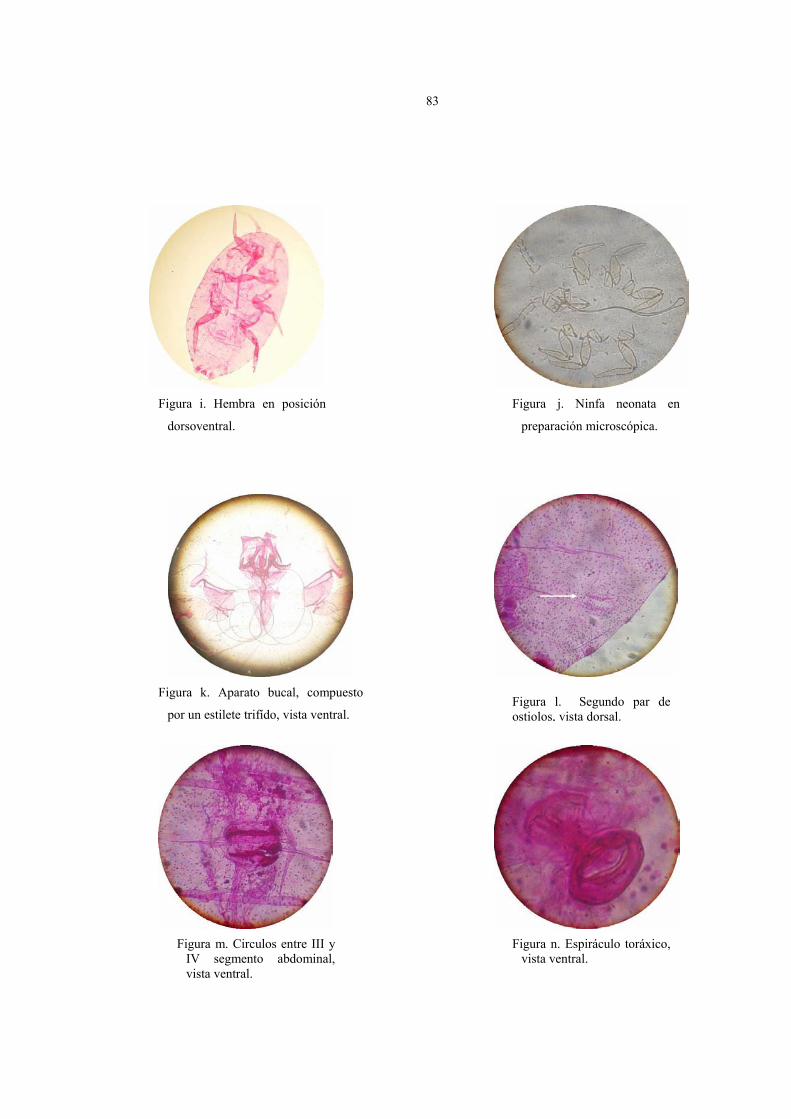
83
Figura i. Hembra en posición
dorsoventral.
Figura j. Ninfa neonata en
preparación microscópica.
Figura k. Aparato bucal, compuesto
por un estilete trifído, vista ventral. Figura l. Segundo par de ostiolos, vista dorsal.
Figura m. Circulos entre III y IV segmento abdominal, vista ventral.
Figura n. Espiráculo toráxico, vista ventral.
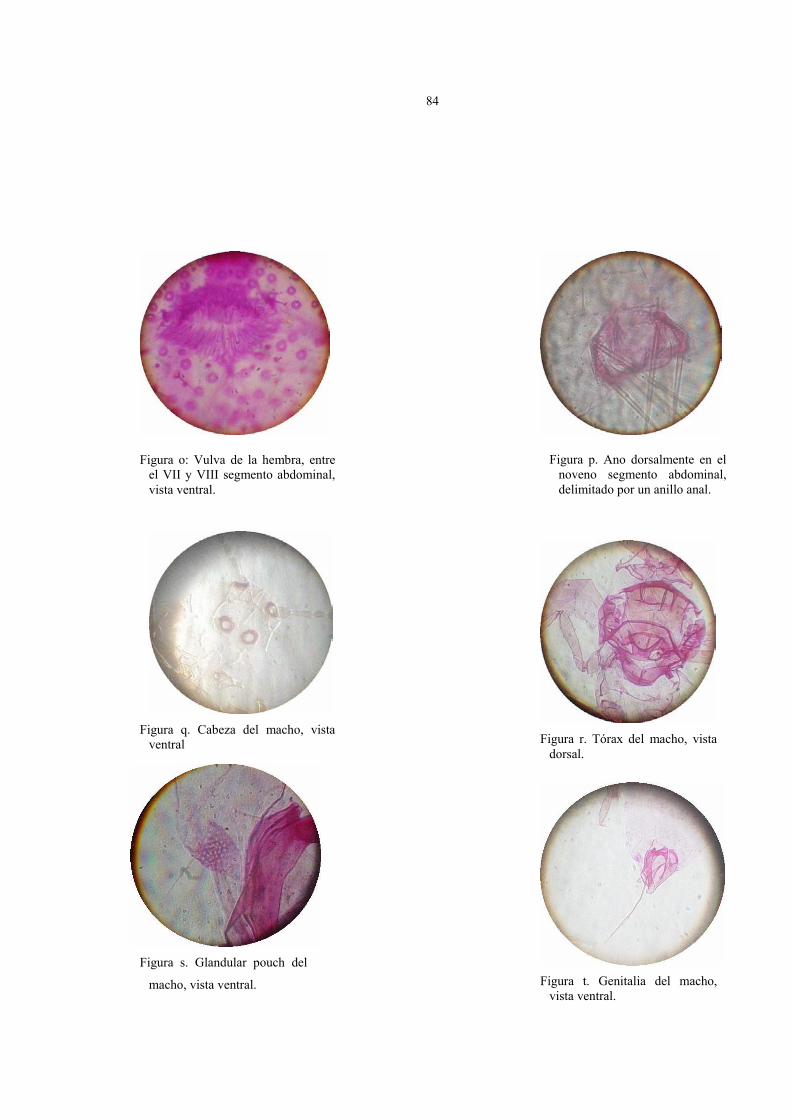
84
Figura o: Vulva de la hembra, entre el VII y VIII segmento abdominal, vista ventral.
Figura p. Ano dorsalmente en el noveno segmento abdominal, delimitado por un anillo anal.
Figura q. Cabeza del macho, vista ventral Figura r. Tórax del macho, vista
dorsal.
Figura s. Glandular pouch del
macho, vista ventral. Figura t. Genitalia del macho, vista ventral.

85





























