Embed Size (px)
Citation preview

UNIVERSIDAD DE CIENCIAS Y ARTES DE CHIAPAS
ESCUELA DE BIOLOGÍA
DIVERSIDAD DE RATONES EN DOS TIPOS DE CULTIVO DE
MAIZ, EN LA COMUNIDAD EMILIANO ZAPATA, MUNICIPIO DE OCOSINGO, CHIAPAS.
T E S I S
QUE PARA OBTENER EL TITULO DE:
LICENCIADO EN BIOLOGIA
PRESENTA:
JORGE ALBERTO NÁJERA TORRES.
DIRECTOR DE TESIS:
M. en C. ANNA HORVÁTH EL COLEGIO DE LA FRONTERA SUR, UNIDAD SAN
CRISTÓBAL DE LAS CASAS
ASESORES:
DR. EDUARDO E. ESPINOSA MEDINILLA. UNIVERSIDAD DE CIENCIAS Y ARTES DE CHIAPAS Y EL
COLEGIO DE LA FRONTERA SUR, UNIDAD SAN CRISTÓBAL DE LAS CASAS
BIOL. ROBERTO VIDAL LÓPEZ INSTITUTO DE HISTORIA NATURAL Y ECOLOGÍA
TUXTLA GUTIÉRREZ, CHIAPAS. MARZO DE 2008

CONTENIDO
DEDICATORIA
AGRADECIMIENTOS
INDICE DE CUADROS
INDICE DE FIGURAS
INDICE DE ANEXOS
RESUMEN
1. INTRODUCCIÓN
2. JUSTIFICACIÓN
3. ANTECEDENTES
Diversidad biológica
Comunidad
Diversidad
Abundancia relativa
Estudios sobre la alteración del hábitat y su efecto sobre
la diversidad de comunidades de roedores
Trabajos que se han realizados en el Estado de Chiapas sobre roedores en
sistemas agrícolas
3.5. Estudios de roedores en la Reserva de la Biosfera Montes Azules, Chiapas
4. OBJETIVO
4.1 General
4.2 Particulares
5. MARCO DE REFERENCIA
5.1 Ubicación
5.2 Clima

5.3 Vegetación.
5.4 Geología
5.5 Suelo
5.6 Hidrografía
5.6.1. Subcuenca del Santo Domingo
5.6.2. Subcuenca del Lacantún
6. METODOS
6.1 Selección de sitos de muestreo
6.2 Muestreo de ratones
6.3 Análisis de datos
6.3.1 Riqueza
6.3.2 Abundancia Relativa
6.3.3 Índice de diversidad de Shannon
6.3.4 Equidad de Pielou (J)
6.3.5 Método de Hutcheson
7. RESULTADOS
7.1 Éxito de captura y diversidad de ratones
7.2 Sexo, Edad y Estado Reproductivo
7.2.1 Sexo
7.2.2 Edad
7.2.3 Estado reproductivo
7.3 Temporalidad
8. DISCUSIÓN.
9. COCLUSIONES
10. LITERATURA CITADA
11. ANEXOS

INDICE DE CUADROS
Cuadro 1. Riqueza, abundancia y diversidad de ratones
Cuadro 2. Total de abundancia relativa de captura y recaptura
Cuadro 3. Número de individuos de hembras y machos por especies
Cuadro 4. Patrón temporal de la distribución de sexos y edad
INDICE DE FIGURAS
Figura 1. Localización del área de estudio y la ubicación de los sitios de muestreos
Figura 2. Diagrama umbrotérmico, estación San Quintín
Figura 3. Índice de diversidad de ratones por tipos de hábitat
Figura 4. Porcentaje de individuos de hembra y macho total y por especie
Figura 5. Porcentaje de edad de ratones en el área total y por especie
Figura 6. Condición reproductiva de los ratones
Figura 7. Patrón temporal de la abundancia de ratones
Figura 8. Patrón temporal de la abundancia de las tres especies
Figura 9. Patrón temporal de la determinación de sexo
Figura 10. Patrón temporal de la etapa reproductiva de los ratones

INDICE DE ANEXOS
Anexo 1. Características de los sitios de estudio
Anexo 2. Hoja de registro de especies y datos de campo
Anexo 3. Marcaje permanente en las orejas

RESUMEN
El trabajo se realizó en la comunidad de Emiliano Zapata, en el Municipio de Ocosingo,
Chiapas, con el objetivo de conocer la diversidad de ratones en dos tipos de cultivo de
maíz: cultivos tradicionales y cultivos con fríjol abono. Se elaboró un listado de
especies y se determinó la riqueza en los dos tipos de cultivo de maíz. Se estimó la
abundancia y se comparó la riqueza y abundancia de ratones en los dos tipos de cultivo.
Para el muestreo, se seleccionaron seis parcelas, tres por cada tipo de hábitat en
cuestión: milpa tradicional con solo maíz y milpa con fríjol abono. La definición de los
tipos de hábitat se realizó según su tipo de hábitat; contar con el permiso del propietario;
tener una distancia al menos de 500 metros entre cada una de las parcelas, evitar que las
parcelas estén demasiado cerca al poblado y accesibilidad a la parcela. Además de estas
características se tomaron otros datos del hábitat, que se consideraron importantes para
los ratones. Los datos de las comunidades de ratones se obtuvieron a través de captura-
marcaje-recaptura entre los meses de mayo, junio, agosto, septiembre y diciembre del
2005. Se aplicó un esfuerzo de captura total de 2058 noches-trampa, se obtuvieron 92
capturas de 84 individuos en total de tres especies. En los sitios de estudio se
identificaron tres especies de ratones, correspondientes a dos familias y tres géneros:
Heteromys desmarestianus (Rodentia: Geomyidae), Sigmodon hispidus y Oligoryzomys
fulvescens (Rodentia: Muridae). En las parcelas de milpa con fríjol abono se
encontraron las tres especies, mientras en las parcelas de milpa tradicional se
encontraron dos: S. hispidus y H. desmarestianus. La especie mas abundante fue S.
hispidus, seguido por H. desmarestianus. La mayor riqueza de especies se encontró en
la milpa con fríjol abono, mientras en la milpa tradicional con solo maíz se registró la
mayor abundancia total. La abundancia de ratones en la milpa tradicional con solo maíz
mostró una marcada temporalidad, teniendo un pico de abundancia en agosto-
septiembre. Al mismo tiempo, el número de individuos de ratones en la milpa con fríjol
abono se mantuvo bajo y relativamente estable durante todo el ciclo de muestreo. El
cultivo con fríjol abono juega un papel importante en el control de plagas de ratones y
favorece la biodiversidad regional.

1. INTRODUCCIÓN
México, es un país en el que la diversidad de formas de vida alcanza una
magnitud extraordinaria. Del total de especies vegetales, animales y de
microorganismos conocidos en el planeta, alrededor del 10 por ciento existen en
México. Más aun, muchos de ellos viven exclusivamente en México (Mittermeier,
1988; Ceballos y Ecardi, 1996; Mittermeier et al., 1997). Es el tercer país de mayor
diversidad biológica después de Brasil e Indonesia (Ceballos y Ecardi, 1996; Ceballos y
Brown, 1995; Mittermeier, 1988; Mittermeier et al., 1997). Ocupa el primer lugar en
reptiles y anfibios con más de 1100 especies, el tercero en mamíferos (529 especies), el
cuarto en plantas vasculares (22000 especies) y el onceavo en aves (1050 especies).
Además, nuestro país tiene el primer lugar mundial en cactáceas y tan solo en el Golfo
de California habitan 28 especies de mamíferos marinos, lo que representa el 35 por
ciento de las especies existentes a nivel mundial. (Ceballos y Oliva, 2005). Los roedores
constituyen el orden de mamíferos más diverso en el mundo, y también así es en el nivel
del país. México cuenta con 221 especies de roedores, es decir, casi la mitad de la
riqueza de mamíferos a nivel nacional, y esta diversidad significa el cinco por ciento de
la riqueza de mamíferos a nivel mundial (Retana y Lorenzo, 2002). En Chiapas, que se
encuentra en el cinturón tropical, es una de las zonas de mayor diversidad biológica en
el mundo. Debido a su localización geográfica, historia biogeográfica y las
características fisiográficas y climáticas de esta zona, existen mamíferos tanto de la
Región Neártica como de la Neotropical (Aranda y March, 1987; Álvarez del Toro,
1993). De los mamíferos de Chiapas se reporta la presencia de 38 especies de roedores
de las familias Geomyidae y Muridae. Seis especies pertenecen a la familia Geomyidae;
de esta, Heteromys nelsoni (subfamilia Heteromyinae, tribu Heteromyini) es endémica,
monotípica, continental, rara y en protección especial. Para la familia Muridae se
registran 32 especies en dos subfamilias: Arvicolinae con una especie y Sigmodontinae
con 31 especies. De esta última subfamilia, tres especies: Peromyscus zarhynchus (tribu
Peromyscini), Tylomys bullaris y Tylomys tumbalensis (tribu Tylomyini), son
endémicas, monotípicas, continental, en el caso de T. bullaris es una especie
amenazada. P. zarhynchus y T. tumbalensis son especies raras y sujetas a protección
especial (Flores-Villela y Gerez, 1994; Ramírez-Pulido et al; 2005).

Sin embargo la pérdida de diversidad biológica en México al igual que en otros
países megadiversos, está ocurriendo rápidamente y en grandes dimensiones, debido a
las diferentes actividades humanas que inducen el cambio de uso del suelo. Las
actividades humanas incluyen aspectos económicos (la ganancia esperada por usos
alternativos del suelo), demográficos (como la presión por tierra para agricultura,
ganadería y nuevas áreas urbanas debido al acelerado crecimiento poblacional),
políticos (las normas que regulan la tenencia de la tierra) y técnicos (como la aptitud de
los terrenos para diferentes usos; SARH, 1984). Una de las causas de la pérdida de la
diversidad biológica en Chiapas es la agricultura. La mayor parte es de tipo de roza-
tumba-quema también denominada agricultura migratoria. Es un sistema mediante la
cual se realizan actividades agrícolas durante un período aproximado de dos o tres años
seguidos por uno de descanso, mayor que el periodo de cultivo (Flores et al., 1971).
Esta técnica de cultivo implica la disminución de la biodiversidad y alcanza una forma
extrema en los monocultivos. El resultado final es una producción artificial que requiere
de una constante intervención humana. En la mayoría de los casos, esta intervención
ocurre en la forma de insumos de agroquímicos, los cuales, además de aumentar los
rendimientos y al mismo tiempo los gastos de producción, resultan en una cantidad de
costos ambientales y sociales indeseables (Altieri, 1987). Una alternativa utilizada por
los campesinos en Chiapas es la práctica de cultivo de maíz combinado con el fríjol
terciopelo (Mucuna pruriens), conocido también como fríjol abono, el cual reduce los
costos de mano de obra al controlar la maleza y aumenta los rendimientos de maíz al
proporcionar nutrimentos cuando son mas necesarios. La combinación maíz-fríjol
terciopelo representa un desvío radical de las técnicas tradicionales de agricultura de
roza-tumba-quema (Wood, 1994). El uso del fríjol abono es una práctica accesible para
la mayoría de agricultores de países en desarrollo debido a sus bajos costos de
establecimiento y manejo (Flores, 1991).
Los sistemas agrícolas por lo general son de monocultivo y más frecuentemente
de variedades vegetales genéticamente similares. Las plantas, los herbívoros y los
predadores de los cultivos agrícolas no constituyen un sistema de coevolución, de modo
que no tienen lugar los fenómenos normales de integración evolutiva. El número de
alteraciones es mucho mayor en los sistemas agrícolas que en los naturales, lo cual

origina una disminución en la diversidad de las especies y los hace diferentes de las
comunidades naturales (Krebs, 1985).
La gran diversidad de formas de vida y adaptaciones de los roedores, les ha
permitido ocupar un gran número de nichos ecológicos. Así, los encontramos terrestres,
semiacuáticos, fosoriales, arborícolas e incluso planeadores. Así mismo han
desarrollado una gran variedad de estrategias alimentarías ya que encontramos
frugívoros, herbívoros, insectívoros, granívoros, omnívoros (Espinosa-Salvatierra y
Canela-Zarate, 1996). Su distribución geográfica alcanza casi todos los ecosistemas
terrestres en los continentes, con la excepción de Nueva Zelanda y algún grupo insular
oceánico (Ceballos y Galindo, 1984).
De las familias de roedores más comunes que habitan lugares perturbados,
incluidas las zonas agrícolas; tenemos a las tuzas (Geomyidae), que tienen un área de
distribución que se extiende desde el sur de Canadá hasta América Central y el sur de
Panamá. Estos roedores son animales muy voraces y comen cualquier tipo de materia
vegetal que puedan encontrar bajo tierra: raíces, bulbos, tallos y partes subterráneas de
las plantas. En algunas zonas se han convertido en plagas, pues destruyen árboles y
cultivos, sin embargo, su actividad excavadora ayuda a mejorar la calidad del suelo,
pues lo ahuecan y entierran materia orgánica (Ceballos y Galindo, 1984). Los ratones
espinosos de abazones (Geomyidae: Heteromyinae), viven en regiones áridas o
semiáridas del oeste de Estados Unidos hasta el sur de Ecuador, aunque algunas
especies habitan en zonas más húmedas. Su dieta consiste en semillas, plantas e insectos
(Álvarez del Toro, 1977). De las ratas y ratones de la familia Muridae, las especies mas
familiares son las del viejo mundo, ratas o ratones de casa, pertenecientes a los géneros
Mus y Rattus. Dos especies Mus musculus y Rattus rattus son los cuales están
estrechamente asociados con los humanos y con ayuda del humano llegaron a
distribuirse en todos los rincones del mundo, introduciéndose ya en casi todas las islas
también. Estas especies son omnívoras, como lo son muchos muridos. Otros son mas
especializados en su dieta y pueden ser herbívoros, insectívoros, especialistas en frutas y
semillas o inclusos carnívoros (Reid, 1997).
A pesar de su gran diversidad de formas de vida, este grupo de mamíferos es
notablemente uniforme en caracteres estructurales, cuya característica principal son los
cuatro incisivos muy desarrollados y la carencia de caninos, lo que deja en las

mandíbulas un espacio entre los incisivos y los molares, llamado diastema (Álvarez del
Toro, 1977). Ellos exhiben un rango en el tamaño del cuerpo y forma, con patas cortas y
colas largas, sin embargo, algunas especies tienen patas largas y delgadas y colas cortas.
La mayoría de los roedores tienen cuatro dedos en las patas delanteras y cinco en las
traseras. Tienen una formula dentaria: I 1/1, C 0/0, P 0-2/0-2, M 2-3/2-3 (Ceballos y
Galindo, 1984; Reid, 1997). Los roedores tienen importantes funciones en el ecosistema
como consumidores de diferentes partes de plantas, insectos, hongos y líquenes. Son
importantes dispersores de semillas, diseminadores de hongos ectomicorríticos y
polinizadores de la vegetación. Además son presas potenciales de muchas especies de
vertebrados como reptiles, aves y otros mamíferos. La mayoría de ellos tienen
poblaciones abundantes, ciclo de vida corto y alto potencial reproductivo (Carey y
Jonson, 1995). Un punto importante es que las poblaciones de pequeños mamíferos
pueden ser indicadores de la calidad del hábitat, ya que la dinámica poblacional cambia
en respuesta a disturbios (Ceballos y Galindo, 1984).
2. JUSTIFICACIÓN
En Chiapas, durante la última década casi el 60% de la vegetación natural se ha
transformado en tierras agropecuarias, o bien en zonas perturbadas y áreas urbanas. Las
consecuencias de la destrucción de la capa arbórea constituyen un problema grave con
consecuencias múltiples. Los principales efectos de la deforestación son: la reducción
de la productividad general de la zona, el suelo se convierte más susceptible a la
erosión, se altera el ciclo hidrológico, disminuye en buena medida la biodiversidad y se
reducen las reservas de nutrientes y biomasa antes almacenadas en los restos de árboles
y hojas (SEMARNAP, 1998).
Chiapas es el estado con el mayor número de ejidos en el país y con una de las
tasas más altas de apertura a nuevas tierras para usos agropecuarios (Flores-Villela y
Gerez, 1994; SEMARNAP, 1998). Las prácticas agrícolas intensivas han generado
nuevos hábitats disponibles para algunas especies de los roedores (Wood, 1994). Los
monocultivos en muchos casos magnifican las fluctuaciones poblacionales de los

roedores, alterando tanto la amplitud de los periodos reproductivos como la elevación
de la densidad poblacional (Altieri, 1987).
En la comunidad Emiliano Zapata, Municipio de Ocosingo, Chiapas,
Conservación Internacional (CI) en años anteriores llevó a cabo un programa para
fomentar la técnica de cultivo orgánico utilizando el fríjol abono como cultivo de
cobertura y abono verde en las milpas con maíz; con el propósito de que los agricultores
de esta comunidad no siguieran practicando la técnica tradicional de roza-tumba-quema
que provocaba accidentalmente incendios forestales y que por medio de esta leguminosa
los agricultores no utilizarían mas fertilizantes y mejorarían la calidad de sus tierras. Se
eligió estudiar los cultivos de maíz porque es el cultivo mas importante de México, es el
núcleo de la economía campesina, base de la dieta popular, y el cereal de mayor
consumo.
En el presente trabajo se estudió la diversidad de roedores pequeños en dos tipos
de hábitat agrícola: cultivo de maíz con fríjol abono y el cultivo de maíz tradicional de
tumba-quema-roza. Se identificó el impacto de estos dos tipos de cultivo de maíz sobre
la diversidad de ratones. Se determinaron las especies de ratones y sus abundancias en
los dos tipos de cultivo y se identificaron las posibles especies que puedan llegar a ser
plagas. Así también, se analizó la dinámica poblacional y permanencia de los ratones en
los cultivos, para conocer si existe alguna tendencia de temporalidad con relación al
ciclo de cultivo y/o ciclo natural de la época de lluvia y seca.

3. ANTECEDENTES
3.1. Diversidad biológica
La biodiversidad se define como la variabilidad entre los organismos vivientes
de todas las fuentes, incluyendo, entre otros, los organismos terrestres, marinos y de
otros ecosistemas acuáticos, así como los complejos ecológicos de los que forman parte;
esto incluye diversidad dentro de las especies, entre especies y de ecosistemas (UNEP,
1992). El término comprende, por tanto, diferentes escalas biológicas: desde la
variabilidad en el contenido genético de los individuos y las poblaciones, el conjunto de
especies que integran grupos funcionales y comunidades completas, hasta el conjunto de
comunidades de un paisaje o región (Solbrig, 1991; Halffter y Ezcurra, 1992; Heywood,
1994).
3.2. Comunidad
Como comunidad ecológica, entendemos un conjunto de poblaciones de
especies diferentes que interactúan entre si, localizadas en un área geográfica específica
(Krebs, 1985; Maffe y Carroll, 1997). Las comunidades se han definido con diferentes
enunciados, en ocasiones establecidos arbitrariamente, dependiendo de lo que se quiere
estudiar y las preguntas que se pretendan contestar. Aunque los ecólogos clasifican las
comunidades de diferentes maneras, todas las comunidades poseen ciertas
características que definen su estructura física y biológica. Estas características varían
tanto en el espacio como en el tiempo (Smith y Smith, 2001). La ecología de
comunidades intenta, por lo tanto, entender los patrones o interacciones que se dan no a
nivel de cada población que la compone, sino a nivel de todo el ecosistema. Las plantas,
animales y microorganismos que componen una comunidad son afectados unos a otros
por las relaciones de alimentación, competencia y otras interacciones.
Las interrelaciones dentro de la comunidad determinan sus atributos funcionales,
el flujo de energía y ciclos de los elementos dentro del ecosistema. A su vez, estas
interrelaciones determinan no solo la abundancia de los individuos de las poblaciones
que la componen, sino también su distribución, su densidad y la evolución y

coevolución de las especies que coexisten dentro de la misma (Begon et al., 1996;
Brown y Harney, 1993; Kelt, 1995; Krebs, 1985; Morton et al., 1994; Vázquez et al.,
2000).
3.2.1. Diversidad
La diversidad de especies es uno de los temas centrales en la ecología de
comunidades y es la variedad de especies existentes en una región determinada. Esta
diversidad puede medirse de muchas maneras. El número de especies de una región es
decir su riqueza, es una medida que a menudo se utiliza, así como el grado de
dominancia y la abundancia relativa de cada especie en la comunidad (Brown, 1995;
Halffter 1998; Magurran, 1988).
La diversidad de especies se puede medir en tres niveles espaciales y temporales;
alfa, beta y gama (Whittaker, 1972), los cuales son útiles para medir y monitorear los
efectos de las actividades humanas (Halffter, 1998). La diversidad alfa es la riqueza de
especies de una comunidad particular a la que consideramos homogénea, es decir el
número de especies presentes en un área determinada. La diversidad beta es el grado de
cambio en la composición de especies entre diferentes ensambles en un paisaje, y la
diversidad gamma es la riqueza de especies del conjunto de comunidades que integran
un paisaje, resultante tanto de las diversidades alfa como la de las diversidades beta
(Moreno, 2001; Whittaker, 1972).
3.2.2. Abundancia relativa
La abundancia relativa de una especie en una comunidad se refiere a la fracción
con la que contribuye dicha especie, a la abundancia total. La abundancia de una
especie, en un sentido absoluto, puede evaluarse en términos del número de individuos,
biomasa por unidad de área, cobertura, o alguna otra unidad de significado funcional
(Franco et al., 1989; Smith, y Smith, 2001.).

3.3. Estudios sobre la alteración del hábitat y su efecto sobre la diversidad de
comunidades de roedores.
Existen estudios sobre el orden Rodentia en sistemas agrícolas en México, la
mayor parte de los trabajos se han realizado en el centro de la República. Entre los
trabajos importantes realizados tenemos los de:
Mellink (1991), estudió la riqueza y la abundancia de la comunidad de roedores
en tres agrosistemas en San Luis Potosí y encontró menor número de especies en los
sistemas agrícolas áridos más simples en comparación con las áreas no cultivadas, en
contraste, en los agrosistemas mas diversos no hubo diferencias.
Mellink (1995), realizó un estudio sobre las preferencias de hábitat, dinámica
poblacional y estacionalidad reproductiva de roedores en asociación con tres sistemas
agrícolas áridos, sus bordes y sus alrededores no cultivados, en el Altiplano de San Luis
Potosí, México, en donde documenta que la cubierta herbácea es la característica más
importante del hábitat. Los roedores presentaron dos patrones estacionales diferentes: la
mayoría tuvo el tamaño poblacional más alto durante el verano, aunque algunos lo
presentaron en invierno. Todos los roedores tuvieron su pico reproductivo durante el
verano, relacionado en alguna forma con la época de lluvias.
Gallina et al. (1996), estudiaron la relación que tiene la estructura de la
vegetación de los cultivos de café con la diversidad de la comunidad de mamíferos
medianos. En este estudio registraron 24 especies y encontraron una relación directa
entre la riqueza y diversidad con la complejidad estructural de los cafetales. Basándose
a sus resultados se recomienda mantener una alta diversidad en el estrato arbóreo en los
cafetales, ya que de esta forma la mayoría de los mamíferos tienen mayor disponibilidad
de recursos alimentarios y protección lo cual puede promover su conservación.
Cuarón (2000), llevó a cabo un estudio sobre los efectos de los cambios en la
cobertura del suelo sobre la disponibilidad de hábitat para los mamíferos de un área del
sureste de México y norte de Guatemala. Realizó un análisis para identificar las
coberturas de suelo presentes en esa área y de las 12 clases de cobertura que el
consideró, cuatro de ellas dominaron el área: pastizales, vegetación tropical secundaria,
bosque tropical húmedo y humedales. Encontró una relación muy fuerte entre estos
tipos de cobertura y los mamíferos, por lo que la presencia de estos tipos de cobertura

para la transición dentro y entre ellas de las especies de mamíferos es crítica para
mantener su presencia en la región.
González-Romero et a. (2000), realizaron un estudio en el desierto
Chihuahuense, en el que comparan los cambios en la estructura de la comunidad de
roedores entre dos tipos de hábitat: matorral y pastizal. Encontraron una mayor
diversidad y densidad en el matorral que en el pastizal, así mismo el matorral presentó
tres especies exclusivas mientras que en el pastizal fueron dos, siendo estas últimas
características de hábitat alterados y áreas de cultivo. Mientras tanto, las tres especies
características del matorral son especies que necesitan una mayor productividad y
complejidad estructural que le son proporcionados por este hábitat.
Vázquez et al. (2000), analizaron las comunidades de roedores en un hábitat
conservado y otro alterado en un bosque lluvioso en la sierra de Manantlán y
encontraron que la composición de las comunidades varió temporalmente entre los
hábitat. La menor riqueza de especies se presentó durante la mitad de la estación seca y
la mayor al final del periodo húmedo.
3.4. Trabajos que se han realizados en el Estado de Chiapas sobre roedores
en sistemas agrícolas.
Muñoz et al. (2000), realizaron un estudio sobre el efecto de la fragmentación
del hábitat sobre la biodiversidad, en donde analizaron como afecta la alteración y
fragmentación del hábitat sobre la composición y la riqueza de anfibios, reptiles y
roedores.
Horváth et al. (2001), realizaron un estudio en Montebello, Chiapas, donde
evaluaron el efecto del cambio del uso de suelo sobre la diversidad de ratones. Reportan
que la composición de la comunidad cambió de forma descendente del bosque al
ecotono y cultivo que fueron los hábitat muestreados. Así mismo se señala que el
bosque presentó el mayor índice de diversidad en comparación con los otros tipos de
hábitat de mayor alteración, además de que el bosque albergó una comunidad de ratones
particular para el área de estudio por lo tanto juega un papel importante en la
conservación de la biodiversidad local.

Sánchez-Hernández et al. (2001), compararon la diversidad de mamíferos en
cuatro sitios del sureste de México con diferente grado de alteración, en donde señalan
que la riqueza de especies de mamíferos pequeños en la selva alta perennifolia fue
mayor que en los otros sitios mas alterados sugiriendo que la mayor diversidad de
especies tanto de murciélagos como de roedores, se localizan en áreas donde existe
mayor diversidad vegetal, así mismo concluyeron que la alteración afecta tanto la
diversidad de especies como la diversidad de los recursos disponibles para las especies.
Ramírez-Lozano (2002), realizó un estudio en la Reserva de la Biosfera El
Triunfo donde evaluó el efecto de la alteración de los tipos de hábitat sobre los roedores,
encontrando que el hábitat conservado presentó la mayor riqueza y abundancia de
especies. Así mismo efectuó un análisis de correlación que indicó una asociación
positiva entre la diversidad de ratones y la complejidad y cantidad de los elementos
estructurales del hábitat.
Pérez (2005), determinó la diversidad de mamíferos en el área de El Pozo,
municipio de Berriozabal, Chiapas, México. Comparó la diversidad entre cuatro tipos de
hábitat con diferente grado de alteración: Boque mesófilo perturbado, Acahual, Cafetal
con sombra de Inga sp y Maizal. Registró un total de 46 especies de mamíferos en el
área de estudio, distribuidas en 18 familias y 38 géneros, destacando los Quirópteros,
Roedores y los Carnívoros como los ordenes más diversos. El bosque presentó la mayor
riqueza y abundancia. El maíz tuvo la menor riqueza y el cafetal tuvo la abundancia mas
baja.
3.5. Estudios de roedores en la Reserva de la Biosfera Montes Azules,
Chiapas.
En la Reserva de la Biosfera Montes Azules se llevaron a cabo varios trabajos
sobre aspectos de diversidad y estructura de la comunidad de mamíferos, entre ellos de
roedores.
Medellín (1992), estudió la comunidad de mamíferos pequeños en la Selva
Lacandona y encontró que la riqueza de especies no fue diferente entre las selvas y los
sitios alterados, aunque señalo que la diversidad de los mamíferos pequeños estuvo

correlacionada con la complejidad estructural del hábitat, con la riqueza de especies
vegetales y con la densidad de los árboles mayores de 25 metros.
Medellín (1994), realizó un estudio en la Reserva de la Biosfera Montes Azules
en la Selva Lacandona sobre la diversidad y conservación de mamíferos, donde
menciona el estatus de conservación de cada una de las especies registradas así como la
importancia de la reserva para proteger los mamíferos mexicanos que se encuentran
amenazadas, registrando 112 especies para el área de la Reserva.
Zarza (2001), realizó un estudio sobre la estructura de la comunidad de pequeños
mamíferos en diversos hábitat en la Selva Lacandona, Chiapas, donde observó que la
riqueza y diversidad de roedores y murciélagos disminuye conforme se incrementan los
niveles de perturbación del hábitat, además encontró una correlación entre la
abundancia de los roedores con la cobertura del dosel y la profundidad de hojarasca.
Cruz-Lara (2002), realizó un estudio en la región de las Cañadas en la Selva
Lacandona, sobre la diversidad de mamíferos en cafetales y selva mediana, donde
registró 54 especies en ambos tipos de hábitat. Observó que no hubo diferencias
significativas de la riqueza, diversidad, abundancia relativa y similitud de especies entre
cafetal y selva mediana pero si en la estacionalidad en cuanto a la abundancia relativa y
riqueza de mamíferos medianos. Debido a que en este estudio la riqueza y diversidad de
especies son muy similares a las de la selva mediana, el autor señala que los cafetales
con sombra son importantes para mantener la diversidad de mamíferos en ecosistemas
transformados.
Horváth et al. (2006), realizaron un estudio sobre la diversidad de roedores en
cultivos tradicionales de maíz y cultivos donde se utilizan el fríjol abono como una
opción a la fertilización del suelo en la comunidad Emiliano Zapata, en el Municipio de
Ocosingo, Chiapas. Observaron que el tipo de hábitat con mayor riqueza de especies de
ratones fue la milpa con fríjol abono, mientras en la milpa tradicional con solo maíz se
registró la mayor abundancia total, además consideraron que la utilización de los abonos
verdes es recomendable para obtener beneficios adicionales en el control de plagas de
roedores y favorecer la biodiversidad regional.

4. OBJETIVO
4.1 General
Conocer la diversidad de ratones en dos tipos de cultivo de maíz: cultivos
tradicionales y cultivos con fríjol abono, en la comunidad Emiliano Zapata,
municipio de Ocosingo, Chiapas.
4.2 Particulares
Elaborar un listado y determinar la riqueza de ratones en los dos tipos de cultivo
de maíz.
Estimar la abundancia relativa de ratones en los diferentes cultivos.
Comparar la riqueza y abundancia de ratones en los dos tipos de cultivo del área
de estudio.
5. MARCO DE REFERENCIA
5.1 Ubicación
La comunidad Emiliano Zapata, se encuentra ubicado al Oeste de la Reserva de
la Biosfera Montes Azules (RIBMA), en el municipio de Ocosingo en el Estado de
Chiapas. Se ubica entre las coordenadas 16° 23´46´´ N 91° 20´29´´ W a una altitud 220
metros sobre el nivel del mar (Figura 1).

Figura 1. Localización del área de estudio y la ubicación de los sitios de muestreos, en
la Comunidad Emiliano Zapata, Municipio de Ocosingo, Chiapas.

5.2 Clima
La Comunidad Emiliano Zapata se sitúa en una zona tropical lluviosa, con un
clima del tipo Af(m) w(i) g, cálido húmedo según la clasificación de Köppen
modificado por Garcia (1973), con una temperatura media anual de 22 a 26 ºC,
presentando temperaturas máximas que van de 30 a 34 ºC y temperaturas mínimas de 12
a 18 ºC. La precipitación máxima en 24 horas es de 200 a 400 mm, con una humedad
relativa mayor al 76% y una precipitación media anual de 2500 a 3000 mm, presentando
de 100 a 150 días nublados y el número de días de lluvias va de 120 a 140 (Mata-
Sánchez, 1979; Figura 2).
Figura 2. Diagrama umbrotérmico, estación San Quintín, municipio de Ocosingo,
Chiapas. Fuente: Mata-Sánchez 1979.

5.3 Vegetación
El tipo de vegetación encontrada en la zona de la comunidad Emiliano Zapata es
de selva alta siempre verde (Miranda, 1951), esta asociación vegetal también se le
conoce como: “rain forest”, “tropical evergreen forest” (Leopold, 1950), “pluveesilvae”
(Ruebel, 1930), “rain forest”, “evergreen seasonal forest” (Beard, 1955), selva alta
perennifolia (Miranda y Hernández, 1963), bosque tropical perennifolio (Rzedowski,
1978) y selva alta perennifolia (Flores et al., 1971).
El bosque tropical perennifolio se desarrolla comúnmente en México en altitudes
entre 0 y 1000 m, aunque en algunas partes de Chiapas asciende hacia 1500 metros
sobre el nivel del mar (Rzedowski, 1978). La selva alta siempre verde, que en Chiapas
suele llamarse “montaña alta” o simplemente “montaña”, se caracteriza por la gran
altura media de los árboles que la forman, que va de 30 metros hasta unos 60 metros, y
por que permanece siempre verde (Miranda, 1951).
5.4 Geología
El área donde se sitúa la comunidad Emiliano Zapata, tiene su origen geológico
durante la Era mesozoica en el período del cretácico inferior y durante el cuaternario.
Está formada de roca sedimentaria de tipo caliza, carbonatada, generalmente plegadas,
se puede distinguir la unidad fisiográfica conformada por planicie intermontaña con una
altitud que va desde los 100 hasta los 1000 metros sobre el nivel del mar, se encuentra
circundada por laderas de pendientes abruptas y lomeríos presentando una fuerte
actividad cárstica (Zúñiga, 1996).
5.5 Suelo
Las características del origen y evolución de los suelos que presenta la
comunidad Emiliano Zapata se encuentran determinadas por el tipo de materiales
geológicos del que derivan, por la acción de las condiciones climáticas, por las

particularidades del relieve y por las condiciones de las comunidades vegetales que
sustentan (García-Gil et al., 1988).
En general, existen dos conjuntos de unidades edáficas de acuerdo a la
clasificación FAO-UNESCO de suelos y son las siguientes:
IEL= Litosoles en mayor proporción, Rendzinas en mediana proporción y Luvisol en
proporciones bajas.
NAI= Nitosol en mayor proporción, Acrisol en mediana proporción y Litosol en
proporciones bajas.
5.6 Hidrografía
La comunidad Emiliano Zapata forma parte de la región hidrológica denominada
Grijalva-Usumacinta, según De Vos (1996). Según C.F.E. (1979) este sistema fluvial es
el más importante de México, y ocupa el séptimo lugar en importancia a nivel mundial y
pertenece a la vertiente oriental aportando las siguientes subcuencas que pasan por la
comunidad:
5.6.1. Subcuenca del Santo Domingo
El Río Santo Domingo-Río Perlas nace en el norte de Margaritas, antes de
confluir al Jataté, recibe aportaciones en su margen izquierdo de los ríos Dolores,
Caliente y Euseba y en su margen derecho al Patarario Seco de Guatemala.
5.6.2. Subcuenca del Lacantún
El río Lacantún nace en el cerro de Tenejapa, al Sureste de San Cristóbal de las
Casas, con el nombre de Huixtán, penetra en la selva como Tzaconejá y su primer
aporte importante lo constituye el Jataté continuando con este nombre. La unión del
Jataté con el Río Santo Domingo-Río Perlas conforman el Río Lacantún, el cual en su
recorrido tiene aportaciones de tributarios originados dentro de la Reserva Integral de la
Biosfera Montes Azules como el Tzendales, San Pedro, Miranda y Lacanja (De Vos
1996).

6. METODOS
6.1 Selección de sitos de muestreo
Se llevó a cabo un reconocimiento del área de estudio para la identificación y
selección de tres sitios de muestreo por cada tipo de hábitat: cultivos tradicionales (C/T)
y cultivo con fríjol abono (F/A), seis en total. Las parcelas para establecer los sitios de
muestreo se seleccionaron tomando en cuenta los siguientes criterios: 1) Tipo de hábitat;
2) Contar con el permiso del propietario; 3) Al menos una hectárea de extensión; 4)
Tener una distancia al menos de 500 metros entre cada una de las parcelas, para
disminuir lo máximo posible la probabilidad de que el mismo individuo de los ratones
pueda ser capturado en mas de una de las parcelas; 5) Evitar que las parcelas estén
demasiado cerca al poblado, para que las trampas no fueran dañadas por personas o
animales domésticos; y 6) Accesibilidad a la parcela. Además, se consideró que las
parcelas de muestreo con fríjol abono deberían tener un período de al menos 2 o 3 años
sin quemarse y cultivándose el fríjol abono, así como las parcelas de muestreo con
cultivo de maíz tradicional hayan sido sujetas a la quema durante al menos el mismo
período. Las parcelas de muestreo se ubicaron geográficamente con GPS y se llevó a
cabo una descripción general, tomando en cuenta el tipo de hábitat (F/A y C/T), los
tipos de hábitat circunvecinos y la cercanía a ellos, así como si era posible, información
sobre la historia de la parcela. Las características de los sitios elegidos se resumen en el
Anexo 1.
6.2 Muestreo de ratones
Se realizaron un total de 45 días de trabajo de campo y se aplicó un esfuerzo de
captura total de 2058 noches-trampa durante los meses de Mayo, Junio, Agosto,
Septiembre y Diciembre de 2005. Las salidas a campo tuvieron como mínimo una
duración de una semana y como máximo doce días en promedio. Para la captura de los
ratones, se realizaron series de trampeos de tres días por cada parcela y sección de
muestreo. Se capturaron los ejemplares con trampas tipo Sherman cebadas con hojuelas

de avena y esencia de vainilla. El diseño de trampeo en cada sitio fue en forma de
cuadrantes que contenía 49 trampas, estableciendo una retícula de 7 x 7 trampas con 10
metros de espaciamiento entre cada una. Las trampas se revisaron al amanecer,
volviéndose a cebar diariamente.
Los animales capturados se manejaron con guantes de carnaza y con mucho
cuidado para no lesionarlos; se extrajeron de las trampas por medio de bolsas de tela o
bolsas de plástico. Posteriormente se identificaron a nivel de especie por sus rasgos
externos con la ayuda de la guía de campo de Reid (1997), se tomaron los datos
morfométricos usuales para mamíferos pequeños (longitud total, longitud de cola,
longitud de pata, longitud de oreja y peso), los datos biológicos: sexo, estado
reproductivo, y clase de edad (juvenil o adulto) así como datos adicionales (hábitat,
fecha, hora, estado de tiempo y microhábitat en donde se capturó) (Anexo 2). Los
primeros tres ejemplares de cada especie se colectaron y se prepararon como
especimenes científicos para formar parte de una colección de referencia del área de
estudio y se depositaron en la Colección Mastozoológica de ECOSUR (ECO-SC-M).
Los subsecuentes individuos capturados tuvieron un marcaje individual permanente en
las orejas (Horváth et al., 2001; Anexo 3) y después de la toma de datos se liberaron en
el lugar de la captura. Adicionalmente, se tomaron fotografías de algunos ejemplares
capturados.
Para determinar el sexo de los individuos entre Macho (M) y Hembra (H) se
observaron las diferencias en los genitales. En el caso de los individuos juveniles o no
escrotados para evitar confusión, se tomó en cuenta la distancia ano-genital, ya que esta
distancia es más corta en las hembras que en los machos.
En el caso de la Edad, se definieron dos categorías: Juvenil (J) y Adulto (A).
Como juveniles se determinaron aquellos individuos destetados que aún se encuentran
asociados con su madre o hermanos, todavía no ha alcanzado el tamaño y pelaje de
adulto ni la madurez sexual. Como adultos se determinaron aquellos individuos que ya
alcanzaron el tamaño promedio, coloración y características del pelaje propias de cada
especie y que alcanzaron la madurez sexual.
La condición reproductiva en las hembras se determinó como: Lactante (LAC),
aquellas con presencia de tetillas alargadas, glándulas mamarias llenas de leche (cuando
eran palpadas), enrojecimiento de las tetillas y no había presencia de embriones; como

Preñada (PRE), se consideró a la hembra que presentaba hinchamiento de tetillas y
presencia de embriones cuando se examinaba por medio del tacto; y Sin Actividad
(S/A) a hembras que no presentaron ninguna de las anteriores características. En los
machos la condición reproductiva se determinó como: Escrotado (ESC), en los
individuos que se observaron notoriamente el descenso de los testículos y Sin Actividad
(S/A), en los machos que no presentaron el descenso de los testículos (Kuns et al.,
1996).
6.3 Análisis de datos
Se diseñó y alimentó una base de datos computarizada en el programa MS
ACCESSR de todos los informes de campo.
6.3.1 Riqueza
Se determinó el número de especies de ratones (riqueza) en cada tipo de cultivo,
así como la riqueza total del área de estudio.
6.3.2 Abundancia Relativa
Para obtener la abundancia de cada especie, se contabilizó el número de
individuos diferentes capturados.
6.3.3 Índice de diversidad de Shannon
Con los datos de la riqueza y abundancia relativa se calculó el índice de
diversidad de Shannon (H´) para cada hábitat.
El índice de Shannon toma en cuenta los dos componentes de la diversidad:
número de especies y equitatividad o uniformidad de la distribución de número de
individuos en cada especie; de acuerdo con esto, un mayor número de especies
incrementa la diversidad y, además, una mayor uniformidad también lo hará (Franco et
al., 1989; Seber, 1986).
= - 2logpi pi

donde:
H´= diversidad (bits/individuo).
pi = proporción del número de individuos de la especie i con respecto al total (ni / NT).
Puede demostrarse que cuando pi = 1/S para toda pi, se alcanza la uniformidad
máxima.
H´ máx = log2 S
H´ máx = diversidad bajo condiciones de máxima equitatividad.
S = número de especies.
La equitatividad puede entonces definirse como:
E = H´/H´máx = H´/log2 S
6.3.4 Equidad de Pielou (J)
Además se calculó la uniformidad o equidad, utilizando el índice de Equidad
(Equitatividad) de Pielou, indica en que medida las especies son abundantes por igual, a
través de la siguiente formula:
J´= H´/ H´max
donde H’max = ln (S).
Este índice mide la proporción de la diversidad observada con relación a la
máxima diversidad esperada (Magurran, 1988).
6.3.5 Método de Hutcheson
Para determinar diferencias significativas en la diversidad de especies en cultivo
tradicional (C/T) y cultivo de maíz con fríjol abono (F/A), se utilizó la prueba de t
modificada por Hutcheson (Magurran, 1988):
t = (H´1 – H´2) / (Var H´1 + Var H´2)1/2
Donde H'1 = valor del índice de diversidad de Shannon para hábitat uno;
H'2 = valor del índice de diversidad de Shannon para hábitat dos;
H'1 = varianza del índice de diversidad de Shannon en hábitat uno;

H'2 = varianza del índice de diversidad de Shannon en hábitat dos; y los grados de
libertad (gl) utilizando:
gl = (Var H´1 + Var H´2)2 / ((VarH´1)
2 / N1) + ((VarH´2)
2 / N2)
La varianza de las H' de cada sistema se determina mediante la ecuación
VarH´= ∑pi(ln pi)2 – (∑pi ln pi)
2 / N – S-1 / 2N
2
7. RESULTADOS
7.1 Éxito de captura y diversidad de ratones
Durante el estudio se obtuvieron 92 capturas de 84 individuos en total. La tasa
de recaptura ha sido muy baja, solo se obtuvieron 8 recapturas. Esto significa un éxito
de captura de 4.08%.
Se identificaron en los sitios de estudio tres especies de ratones,
correspondientes a dos familias y tres géneros: Heteromys desmarestianus (Rodentia:
Geomyidae), Oligoryzomys fulvescens y Sigmodon hispidus (Rodentia: Muridae). En las
parcelas con fríjol abono se encontraron las tres especies, mientras en las parcelas con
milpa de sólo maíz se encontraron dos: Heteromys desmarestianus y Sigmodon
hispidus. La especie más abundante fue Sigmodon hispidus, seguida por Heteromys
desmarestianus. La especie Oligoryzomys fulvescens fue poco abundante, solo se
registró un individuo durante todo el estudio (Cuadro 1).
Las parcelas de cultivos tradicionales de maíz (C/T) obtuvieron 53 capturas
distribuidas en 2 familias y 2 géneros, representada en Sigmodon hispidus con 51
individuos y Heteromys desmarestianus con 2 individuos. También presento 7
recapturas de una familia y un género Sigmodon hispidus. En las parcelas de maíz con
fríjol abono (F/A) obtuvieron 31 capturas distribuidas en 2 familias y 3 géneros,
representada en Sigmodon hispidus con 21 individuos; Heteromys desmarestianus con 9
individuos y Oligoryzomys fulvescens con un individuo. Presentando una recaptura de
una familia y un género Sigmodon hispidus (Cuadro 2).
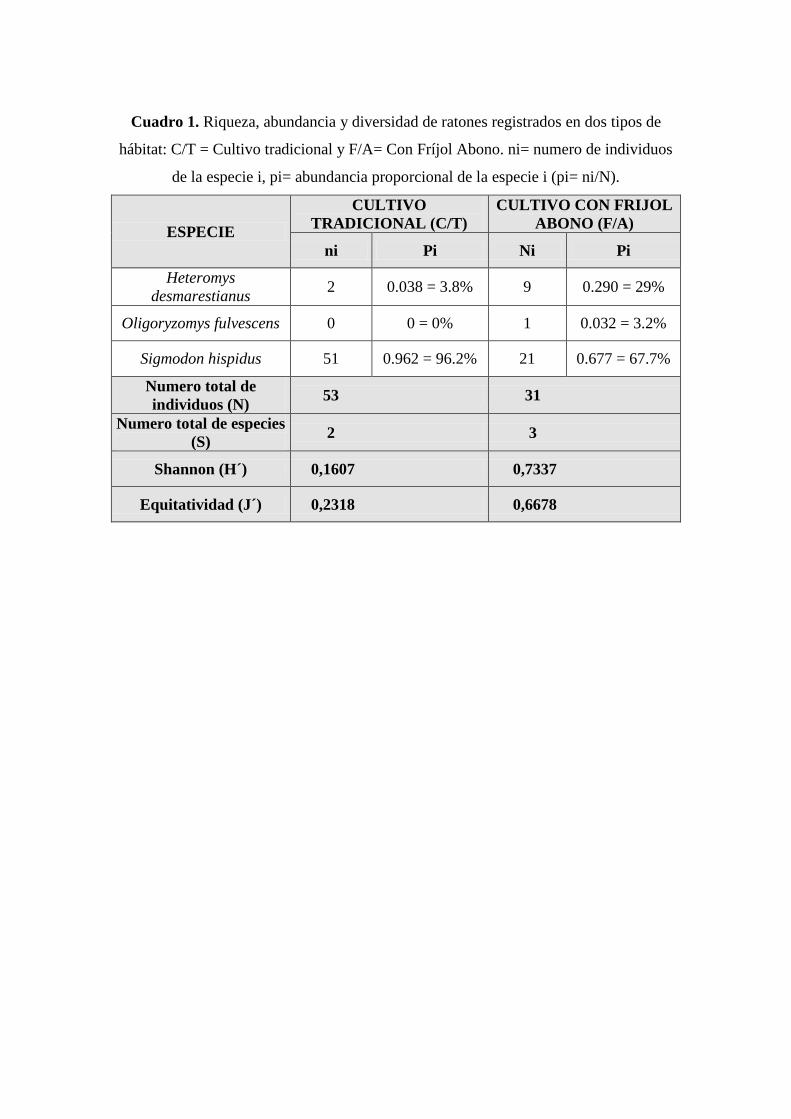
Cuadro 1. Riqueza, abundancia y diversidad de ratones registrados en dos tipos de
hábitat: C/T = Cultivo tradicional y F/A= Con Fríjol Abono. ni= numero de individuos
de la especie i, pi= abundancia proporcional de la especie i (pi= ni/N).
ESPECIE
CULTIVO
TRADICIONAL (C/T)
CULTIVO CON FRIJOL
ABONO (F/A)
ni Pi Ni Pi
Heteromys
desmarestianus 2 0.038 = 3.8% 9 0.290 = 29%
Oligoryzomys fulvescens 0 0 = 0% 1 0.032 = 3.2%
Sigmodon hispidus 51 0.962 = 96.2% 21 0.677 = 67.7%
Numero total de
individuos (N) 53 31
Numero total de especies
(S) 2 3
Shannon (H´) 0,1607 0,7337
Equitatividad (J´) 0,2318 0,6678

Cuadro 2. Total de abundancia relativa de captura y recaptura de ratones en dos tipos
de cultivo de maíz en los sitios de estudio, en la Comunidad Emiliano Zapata,
Municipio de Ocosingo. C/T = Cultivo tradicional, F/A= Con Fríjol Abono.
ESPECIES
CULTIVO TRADICIONAL CULTIVO CON FRIJOL ABONO
CAPTURA RECAPTURA CAPTURA RECAPTURA
Sigmodon hispidus 51 7 21 1
Heteromys
desmarestianus 2 0 9 0
Oligoryzomys
fulvescens 0 0 1 0
TOTAL 53 7 31 1
El índice de Shannon (H´) muestra que F/A presentó la mayor diversidad (H´=
0,7337). C/T presentó una diversidad mucho menor (H´= 0,1607). En cuanto al índice
de equitatividad (J´) F/A mostró la mayor uniformidad en la abundancia de las especies
(J´= 0.6678), mientras C/T presentó menor uniformidad de especies (J´= 0.2318;
Cuadro 1; Figura 3). Analizando los datos de ambos sitios de estudio con la prueba de t
modificada por Hutcheson, encontramos diferencia significativa entre F/A y C/T (t=
4.1427, g.l.= 65.1219; P<0.001).

0.7337
0.1607
0.6678
0.2318
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
cultivo con fríjol
abono (F/A)
Cultivo tradicional
(C/T)
HABITAT
IND
ICE
DE
DIV
ER
SID
AD
Shannon (H´)
Equitatividad (J´)
Figura 3. Índice de diversidad de ratones por tipos de hábitat en la comunidad Emiliano
Zapata, Municipio de Ocosingo, Chiapas. Cultivo con fríjol abono (F/A); Cultivo
tradicional (C/T).
7.2 Sexo, Edad y Estado Reproductivo
7.2.1 Sexo
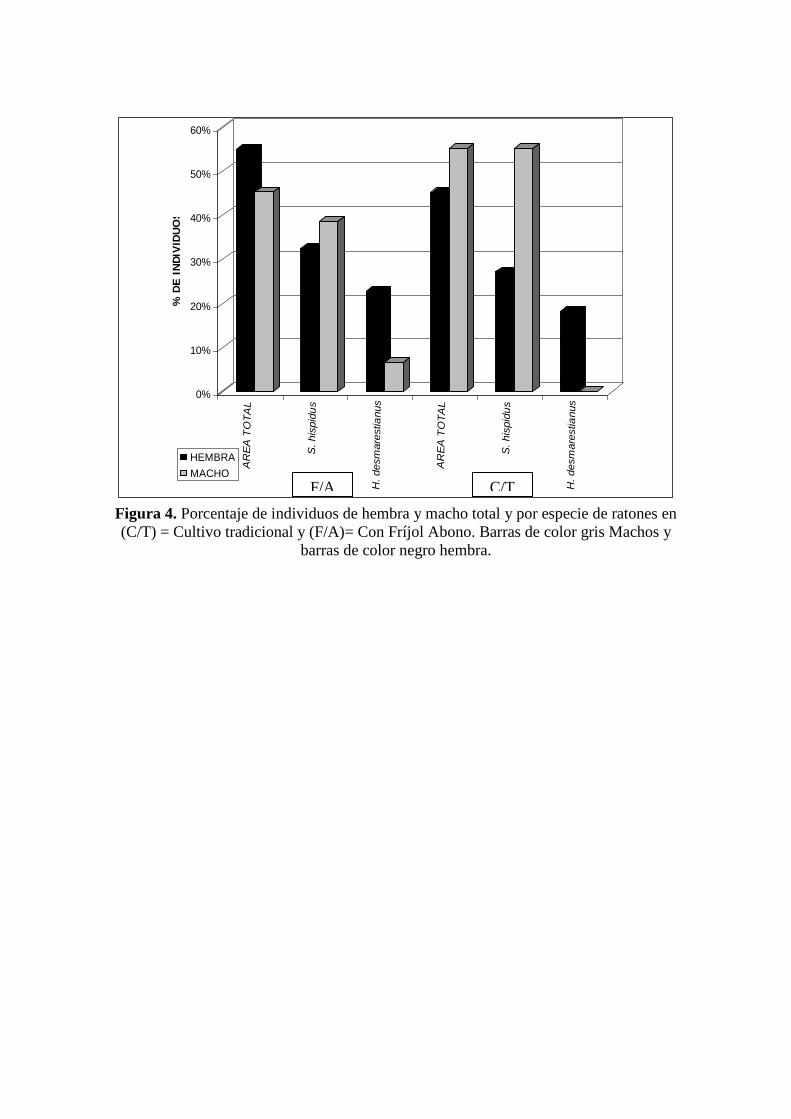
Los porcentajes de los sexos del área total del cultivo de maíz con fríjol abono
(F/A) se inclina a favor de las hembras con 55% (17) de individuos. Este porcentaje es
similar en el H. desmarestianus que presentó el mayor porcentaje de hembras con
22.65% (7) de individuos. Lo que respecta a Sigmodon hispidus, presentó la mayor
inclinación hacia los machos con 38.57% (12) de individuos. El único ejemplar
capturado de Oligoryzomys fulvescens fue macho. En el caso del cultivo de maíz
tradicional (C/T) la proporción de sexos del área total se inclina a favor de los machos
con 55% (37) de individuos. Este porcentaje corresponde en su totalidad a Sigmodon
hispidus. En el caso del H. desmarestianus, las hembras presentaron mayor porcentaje
con 16.43% (2) de individuos (Figura 4; Cuadro 3).
0%
10%
20%
30%
40%
50%
60%
% D
E I
ND
IVID
UO
S
AR
EA
TO
TA
L
S. his
pid
us
H. desm
are
stia
nus
AR
EA
TO
TA
L
S. his
pid
us
H. desm
are
stia
nus
HEMBRA
MACHO
Figura 4. Porcentaje de individuos de hembra y macho total y por especie de ratones en
(C/T) = Cultivo tradicional y (F/A)= Con Fríjol Abono. Barras de color gris Machos y
barras de color negro hembra.
C/T F/A

Cuadro 3. Número de individuos de hembras y machos por especies de ratones en dos
tipos de hábitat: C/T = Cultivo tradicional y F/A= Con Fríjol Abono.
ESPECIES
CULTIVO TRADICIONAL CULTIVO CON FRIJOL ABONO
HEMBRA MACHO
SIN
DETERMINA
CION
HEMBRA MACHO
SIN
DETERMINA
CION
Sigmodon
hispidus 18 37 3 10 12 0
Heteromys
desmarestian
us
2 0 0 7 2 0
Oligoryzomys
fulvescens 0 0 0 0 1 0
TOTAL 20 37 3 17 15 0
7.2.2 Edad
Para ambos tipos de hábitat se observó una inclinación a favor de la edad Adulta,
con 78 y 77% de individuos para F/A y C/T respectivamente. Sigmodon hispidus,
mostró mayor porcentaje de adultos con 73.42% de individuos en el hábitat C/T y F/A
con 49.92% de individuos. Mientras que en caso de H. desmarestianus la edad Adulta
fue mucho mayor en el hábitat F/A con 24.96% de individuos. Oligoryzomys fulvescens
el único ejemplar fue de la edad Adulta; presentándose en el hábitat F/A (Figura 5).

0%
10%
20%
30%
40%
50%
60%
70%
80%
% D
E I
ND
IVID
UO
S
AR
EA
TO
TA
L
S.
his
pid
us
H.
desm
are
stia
nus
O.
fulv
escens
AR
EA
TO
TA
L
S.
his
pid
us
H.
desm
are
stia
nus
JUVENIL
ADULTO
Figura 5. Porcentaje de edad de ratones en el área total y por especie en dos tipos de
hábitat. C/T = Cultivo tradicional y F/A= Con Fríjol Abono. Barra de color negro
Juvenil y Barra de color gris Adulto.
7.2.3 Estado reproductivo
Para la condición reproductiva, Sigmodon hispidus presentó la mayor
abundancia en C/T de individuos sin actividad (S/A) con 19; 28 escrotados (ESC); 5
preñadas (PRE); 3 lactantes (LAC) y 3 sin determinación (S/D). En caso de Heteromys
desmaretianus se encontraron más individuos con actividad reproductiva en F/A con 2
individuos sin actividad (S/A); 1 escrotado (ESC); 3 preñadas (PRE) y una lactante
(LAC). El único individuo Oligoryzomys fulvescens que fue capturado en F/A su estado
reproductivo fue sin actividad (S/A Figura 6).
F/A C/T

0
5
10
15
20
25
30
No.
DE
IN
DIV
IDU
OS
S.
his
pid
us
H.
desm
are
stia
nus
O.
fulv
escens
S.
his
pid
us
H.
desm
are
stia
nus
S/A
ESC
PRE
LAC
Figura 6. Condición reproductiva de los ratones en (C/T) = Cultivo tradicional y (F/A)=
Con Fríjol Abono. Barras negras Sin Actividad (S/A), Barra con franjas verticales
Escrotado (ESC), Barra con franjas horizontales Preñada (PRE), y Barra gris Lactante
(LAC).
7.3 Temporalidad
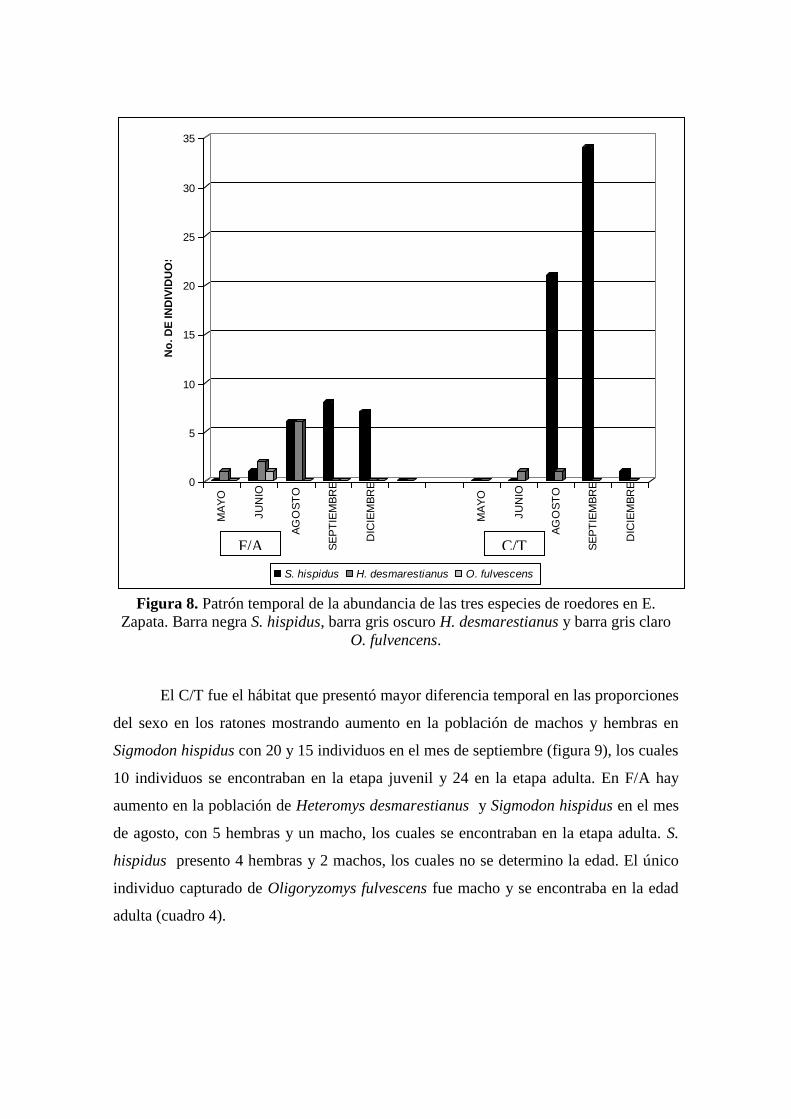
Claramente se observa una marcada temporalidad en el patrón de la abundancia
de ratones durante el estudio. Hay un pico en la abundancia total en el periodo de
Agosto-Septiembre, esto es porque se dispara la población de Sigmodon hispidus en
ambos cultivos, mientras que la abundancia de Heteromys desmarestianus se mantiene
estable y relativamente baja en los dos tipos de cultivo (Figura 7 y 8).
F/A C/T

0
5
10
15
20
25
30
35
40
45
50
MAYO
JUNIO
AGOSTO
SEP
TIEM
BRE
DIC
IEM
BRE
MESES DE MUESTREO
No.
DE
RA
TO
NE
S C
AP
TU
RA
DO
S
C/T= Cultivo Tradicional
F/A= Con Frijol Abono
TOTAL
Figura 7. Patrón temporal de la abundancia de ratones en los dos tipos de cultivo y el
total; en la comunidad Emiliano Zapata. Línea pentagonal negro Cultivo tradicional
(C/T), Línea cuadrada negra Cultivo con fríjol abono y Línea triangular negro Total.
0
5
10
15
20
25
30
35
No
. D
E IN
DIV
IDU
OS
MA
YO
JU
NIO
AG
OS
TO
SE
PT
IEM
BR
E
DIC
IEM
BR
E
MA
YO
JU
NIO
AG
OS
TO
SE
PT
IEM
BR
E
DIC
IEM
BR
E
S. hispidus H. desmarestianus O. fulvescens
Figura 8. Patrón temporal de la abundancia de las tres especies de roedores en E.
Zapata. Barra negra S. hispidus, barra gris oscuro H. desmarestianus y barra gris claro
O. fulvencens.
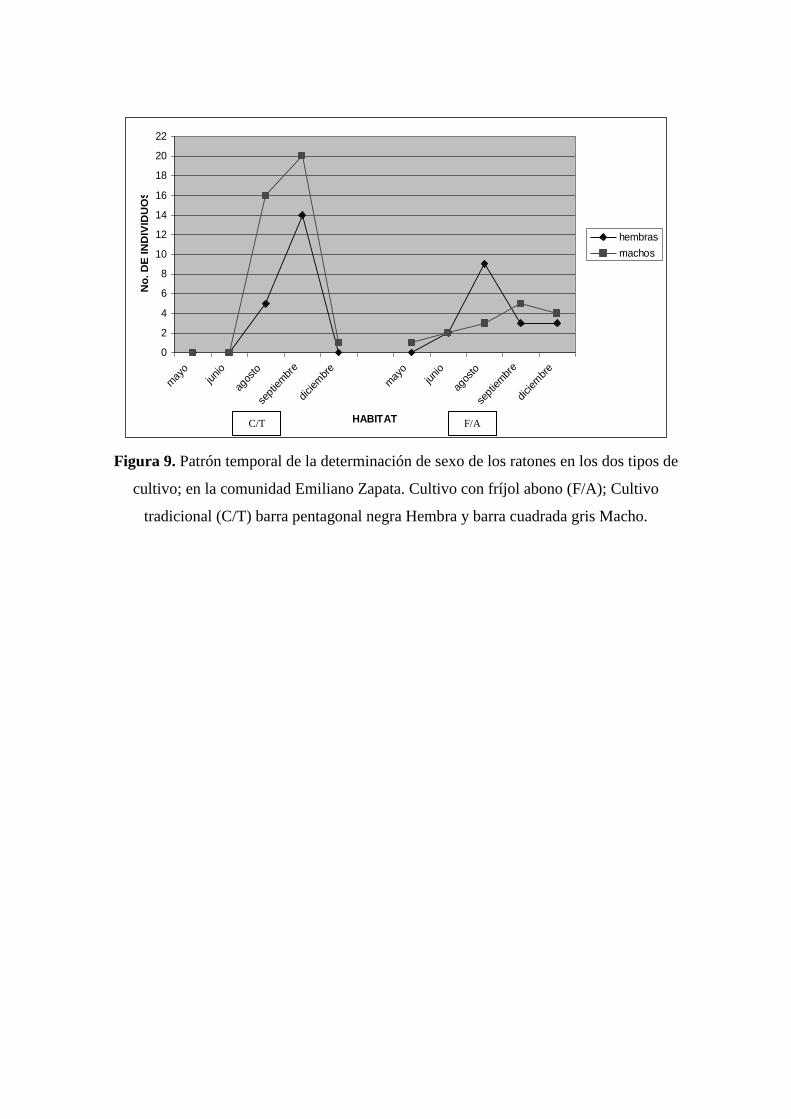
El C/T fue el hábitat que presentó mayor diferencia temporal en las proporciones
del sexo en los ratones mostrando aumento en la población de machos y hembras en
Sigmodon hispidus con 20 y 15 individuos en el mes de septiembre (figura 9), los cuales
10 individuos se encontraban en la etapa juvenil y 24 en la etapa adulta. En F/A hay
aumento en la población de Heteromys desmarestianus y Sigmodon hispidus en el mes
de agosto, con 5 hembras y un macho, los cuales se encontraban en la etapa adulta. S.
hispidus presento 4 hembras y 2 machos, los cuales no se determino la edad. El único
individuo capturado de Oligoryzomys fulvescens fue macho y se encontraba en la edad
adulta (cuadro 4).
F/A C/T
0
2
4
6
8
10
12
14
16
18
20
22
may
ojunio
agos
to
sept
iem
bre
diciem
bre
may
ojunio
agos
to
sept
iem
bre
diciem
bre
HABITAT
No
. D
E IN
DIV
IDU
OS
hembras
machos
Figura 9. Patrón temporal de la determinación de sexo de los ratones en los dos tipos de
cultivo; en la comunidad Emiliano Zapata. Cultivo con fríjol abono (F/A); Cultivo
tradicional (C/T) barra pentagonal negra Hembra y barra cuadrada gris Macho.
C/T F/A

Cuadro 4. Patrón temporal de la distribución de sexos y edad de las tres especies de
ratones en dos tipos de cultivo; en la comunidad Emiliano Zapata. Cultivo con fríjol
abono (F/A); Cultivo tradicional (C/T). H=hembra; M=macho; J=juvenil y A=adulto.
ESPECIES
CULTIVO CON FRIJOL ABONO
MAY JUN. AGOS SEP DIC
H M J A H M J A H M J A H M J A H M J A
Heteromys
desmarestianus 0 1 1 0 2 0 0 2 5 1 0 5 0 0 0 0 0 0 0 0
Oligoryzomys
fulvescens 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0
Sigmodon
hispidus 0 0 0 0 0 1 0 1 4 2 0 0 3 5 0 0 3 4 0 0
TOTAL 0 1 1 0 2 2 0 4 9 3 0 11 3 5 2 6 3 4 4 3
ESPECIES
CULTIVO TRADICIONAL
MAY JUN. AGOS SEP DIC
H M J A H M J A H M J A H M J A H M J A
Heteromys
desmarestianus 0 0 0 0 0 0 0 0 1 0 0 1 1 0 0 1 0 0 0 0
Sigmodon
hispidus 0 0 0 0 0 0 0 0 5 16 3 18 14 20 10 24 0 1 0 1
TOTAL 0 0 0 0 0 0 0 0 6 16 3 19 15 20 10 25 0 1 0 1
Con respecto al estado reproductivo de los ratones en los dos tipos de hábitat,
encontramos que C/T en el mes de septiembre hay un aumento de población de
individuos sin actividad reproductiva (S/A) con 14 individuos, el aumento de individuos
escrotados (ESC) con 14, preñadas (PRE) con 3 y lactantes (LAC) con 4. Seguido por el
mes de agosto con 4 individuos sin actividad reproductiva (S/A), 14 escrotados (ESC) y
3 preñadas (PRE). El mes de diciembre solo se presento un individuo sin actividad
reproductiva(S/A). En el mes de mayo y junio no hubo captura de individuos. En el
cultivo con fríjol abono (F/A) el mes de agosto presento el mayor pico, debido al
aumento de población de individuos sin actividad reproductiva (S/A) con 2, escrotados
(ESC) con 3, preñadas (PRE) con 5 y lactantes (LAC) con 2; seguido por el mes de
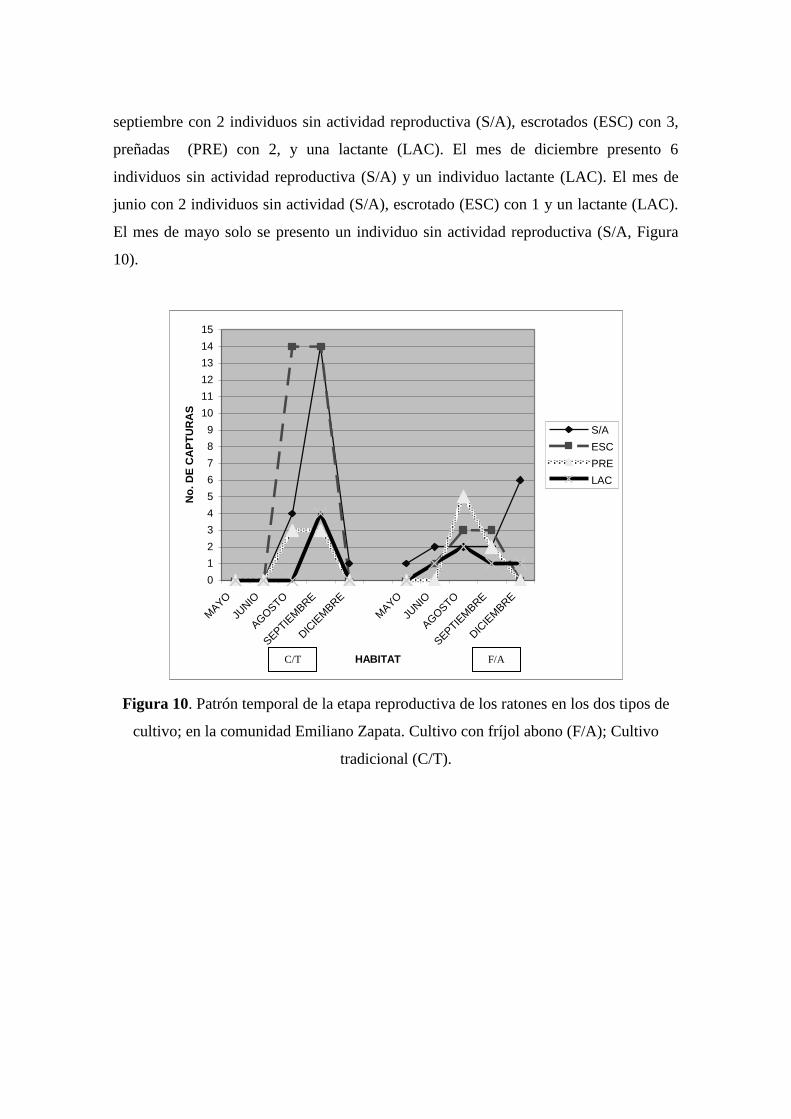
septiembre con 2 individuos sin actividad reproductiva (S/A), escrotados (ESC) con 3,
preñadas (PRE) con 2, y una lactante (LAC). El mes de diciembre presento 6
individuos sin actividad reproductiva (S/A) y un individuo lactante (LAC). El mes de
junio con 2 individuos sin actividad (S/A), escrotado (ESC) con 1 y un lactante (LAC).
El mes de mayo solo se presento un individuo sin actividad reproductiva (S/A, Figura
10).
0
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
MAYO
JUNIO
AGOSTO
SEP
TIEM
BRE
DIC
IEM
BRE
MAYO
JUNIO
AGOSTO
SEP
TIEM
BRE
DIC
IEM
BRE
HABITAT
No
. D
E C
AP
TU
RA
S
S/A
ESC
PRE
LAC
Figura 10. Patrón temporal de la etapa reproductiva de los ratones en los dos tipos de
cultivo; en la comunidad Emiliano Zapata. Cultivo con fríjol abono (F/A); Cultivo
tradicional (C/T).
C/T F/A

8. DISCUSIÓN
Los factores ecológicos más importantes que influyen en la sobrevivencia y
dispersión de las especies en los hábitat agrícolas, son la reducción de la diversidad de
los hábitat, los cambios en la longitud y la estructura de los bordes o ecotonos, la
degradación, fragmentación y la inestabilidad temporal del hábitat. Los paisajes
agrícolas, debido a los ciclos biológicos de las plantas cultivadas y de las actividades
humanas en estas áreas, están en un constante cambio. La transformación de los bosques
en pastizales y áreas de cultivos provoca una perdida en la riqueza y abundancia de las
especies (August, 1983). Esto podemos observarlo en los dos tipos hábitat muestreados
en el área de estudio, La composición de las comunidades de ratones en los sitios de
estudio, se caracteriza por una riqueza de especies baja junto con la fuerte dominancia
de una sola especie (Cuadro 1). El cultivo con fríjol abono (F/A) fue el que presentó la
mayor riqueza de especies de ratones, donde se notó que la especie característica y
dominante fue S. hispidus, seguida por H. desmarestianus y la menos abundante fue O.
fulvescens. Mientras en el cultivo tradicional (C/T) se encontraron dos especies, siendo
la dominante S. hispidus, seguido por H. desmarestianus. El éxito de captura en el área
de estudio es relativamente bajo pero aceptable, ya que en estudios de roedores en
diferentes áreas neotropicales se han reportado éxitos de captura entre 1% y 10.4 %,
pero en la mayoría de los casos no sobrepasa al 5% (August 1983, Bergallo 1994,
Horváth et al. 2001).
Los cultivos son hábitat muy dinámicos, pues a corto plazo están cambiando en
sus características físicas y biológicas. Por ello, no pueden mantener aquellas especies
que tienen requerimientos especiales o una baja tolerancia a la transformación del
hábitat, y favorecen a las especies generalistas o más tolerantes a un ambiente
cambiante. Las especies generalistas adquieren una ventaja competitiva en la ocupación
de los nichos ecológicos y muchas veces llegan a ser dominantes en los hábitat agrícolas
(Harris y Woolard, 1990). Tal es el caso de Sigmodon hispidus, que en el área de
estudio sus abundancias tuvieron dicho comportamiento en los dos tipos de hábitat. La
especie que dominó en ambos hábitat, S. hispidus, es una especie conocida desde el Sur
de los E.U.A. hasta Venezuela como plaga en los cultivos. Está adaptada a los hábitat

perturbados y al parecer sólo requiere cobertura herbácea con pastos, tanto para refugio
y nidos como para alimentos. Por su alta tasa de reproducción (corta duración de
gestación, desarrollo muy rápido de las crías y gran tamaño de camada (Cameron y
Spencer, 1981); logra compensar las desventajas derivadas de los cambios en el hábitat
y los riesgos más elevados de depredación que se presentan en los cultivos. Las otras
dos especies tienen requerimientos complejos y variados del hábitat; como se observó
en el cultivo con fríjol abono (F/A), las cuales al parecer se asocian a un mayor grado de
complejidad y a elementos estructurales para refugio y lugares con características
particulares para los nidos o madrigueras (Horváth et al., 2001). Probablemente su
alimentación esté fuertemente ligada a los recursos que se encuentran en este tipo de
hábitat, como semillas, hojas, tallos y raíces de las plantas y algunos insectos. Mientras
que en el cultivo tradicional (C/T) la captura de estas dos especies fue muy baja en el
caso del H. desmarestianus y ausente en O. fulvescens; esto se puede asociar a la falta
de estratos verticales que pudieron ser barreras para estas especies (August, 1983). Para
mamíferos pequeños, especialmente ratones, es importante la extensión de los parches
de hábitat; por lo anterior, parches relativamente pequeños con hábitat desfavorable que
se presenten en el área de estudio (cultivos, poca complejidad vertical, falta de
cobertura), pueden significar un impedimento para la movilidad de los individuos
(Ausgust, 1983; Andrén 1994; Horváth et al., 2001).
A pesar de que son casi las mismas especies que se encuentran en los dos tipos
de cultivo, las diferencias en los índices de diversidad (H´) y de equitatividad (J´) son
notables (Figura 3). La mayor diversidad y equitatividad que se encontró en el cultivo
de maíz con fríjol abono (F/A) es debido a que este tipo de cultivo es diversificado,
aunque no es un hábitat tan complejo como los bosques naturales pero el hecho de
integrar la leguminosa, afectan en menor grado la complejidad del hábitat original en
cuanto a los requerimientos de los ratones (alimento, refugio, lugares de anidación,
etc.). El cultivo tradicional (C/T) es un hábitat muy dinámico por lo que no pueden
mantener aquellas especies que tienen requerimientos específicos de hábitat pero si
favorece la ocupación de estos nichos ecológicos por especies generalistas.
La temporada lluviosa, en la mayoría de los casos se asocia con mayor
abundancia, y el período de reproducción por lo general también coincide con la mayor
disponibilidad de alimentos (Martínez-Gallardo y Sánchez-Cordero, 1993). Para la

comunidad Emiliano Zapata, todavía no se han realizado estudios sobre las temporadas
de mayor disponibilidad de alimentos para pequeños mamíferos, por lo que no se sabe si
realmente, exista una relación entre las temporadas y dicha disponibilidad, aunque se
sugiere que esta relación podría ser cierta. Los dos tipos de hábitat mostraron picos de
abundancia y mayor proporción de individuos con actividad reproductiva en agosto-
septiembre. Este comportamiento temporal en las actividades reproductivas, asociadas a
la temporada lluviosa, probablemente se explica por la mayor disponibilidad de
alimentos y sitios de anidación y cobertura que presentaron los dos tipos de hábitat
(Martínez-Gallardo y Sánchez-Cordero, 1993).
En los sistemas agrícolas, la biodiversidad cumple funciones que van más allá de
la producción de alimentos, fibra, combustible e ingresos. Algunas de éstas incluyen el
reciclaje de nutrientes, el control del microclima local, la regulación de procesos
hidrológicos locales, la regulación de la abundancia de organismos indeseables, y la
detoxificación de residuos químicos nocivos (Aguilar-Ortiz, 1982; Groombridge, 1992).
La cobertura de abono verde en la milpa, a través de brindar una mayor estabilidad en la
disponibilidad de los recursos de cobertura para desplazamientos más seguros, refugios
y alimentos, posiblemente funciona como un regulador natural de abundancia,
impidiendo los grandes crecimientos poblacionales de ratones. La utilización de los
abonos verdes es muy recomendable, se menciona que la contribución del fríjol abono
ha sido entre otros la fijación de nitrógeno atmosférico (hasta 200 kg/ha); aumento en la
cantidad de follaje verde que se agrega al suelo (hasta 35 Ton/ha); control de malezas;
protección del suelo contra la erosión y escorrentías; incremento o sostenimiento de los
rendimientos de maíz sin necesidad de fertilizantes químicos. El uso del fríjol abono es
una práctica accesible para la mayoría de agricultores de países en desarrollo debido a
sus bajos costos de establecimiento y manejo (Flores, 1991). También se obtienen
grandes beneficios adicionales al utilizar el fríjol abono en los cultivos en el control de
plagas de ratones y favorecer la biodiversidad regional.

9. CONCLUSIONES
En las parcelas estudiadas se obtuvieron 92 capturas de 84 individuos en total de
tres especies de ratones, correspondientes a dos familias y tres géneros:
Heteromys desmarestianus (Geomyidae), Oligoryzomys fulvescens y Sigmodon
hispidus (Muridae).
El cultivo con mayor riqueza de especies de ratones fue en las parcelas con fríjol
abono con las tres especies.
La parcela con milpa de solo maíz se registró la mayor abundancia, llegando casi
al doble del número de individuos que en los cultivos de maíz con fríjol abono.
Con una fuerte dominancia de S. hispidus.
La abundancia de ratones en la milpa tradicional con solo maíz muestra una
marcada temporalidad, teniendo un pico de abundancia en agosto-septiembre.
Este pico de abundancia es causado por un rápido crecimiento poblacional de S.
hispidus que coincide con una mayor disponibilidad de recursos por el
establecimiento e invasión de las malezas durante la época de pre-cosecha y la
tapisca de la milpa.
Al mismo tiempo, el número de individuos de ratones en la milpa con frijol
abono se mantiene bajo y relativamente estable durante todo el ciclo de
muestreo. Esto es debido a los patrones temporales de abundancia de H.
desmarestianus, que aprovecha los granos de frijol abono como alimento. El
frijol abono dificulta la invasión por las malezas gramíneas, y por consecuencia
no hay un marcado aumento poblacional de S. hispidus.
El uso del fríjol abono (Mucuna pruriens) en los cultivos de maíz, además de
reducir los costos de mano de obra al controlar la maleza y aumentar los
rendimientos de maíz al proporcionarle nutrimentos cuando son mas necesarios
es, muy recomendable también para obtener beneficios adicionales en el control
de plagas de ratones y favorecer la biodiversidad regional.

10. LITERATURA CITADA
Altieri, M.A. 1987. Agroecology: The scientific basis of alternative agriculture.
Westview Press, Boulder. 355 pp.
Aguilar-Ortíz, F. 1982. Estudio ecológico de las aves del cafetal. In: E.
Jiménez-Avila y Gómez-Pompa (eds.). Estudios ecológicos en el sistema cafetalero.
Instituto Nacional de Investigaciones sobre Recursos Bióticos, Xalapa, Ver. México.
Pp. 103-122.
Álvarez del Toro, M. 1977. Los Mamíferos de Chiapas. Universidad Autónoma
de Chiapas, México. 147 pp.
Alvarez del Toro, M. 1993. Introducción en : Álvarez del Toro, M., Palacios E.
E., Cabrera C. T. G., Guichard R. A. Ramírez V. A. y Cartas H. G. J. 1993. Chiapas y
su Biodiversidad. Gobierno del Estado de Chiapas, Tuxtla Gutiérrez, Chiapas. 152 pp.
Andrén, H. 1994. Effects of habitat fragmentation on birds and mammals in
landscapes with differnet proportions of suitable habitat: a review. Oikos 71:355-366.
Aranda, M. y March I. 1987. Guía de los Mamíferos Silvestres de Chiapas. 1ª
edición. Instituto Nacional de Investigaciones sobre Recursos Bióticos. México, D.F.
145 pp.
Ausgust, P.V. 1983. The role of habitat complexity and heterogeneity in
structuring tropical mammal communities. Ecology. 64:1495-1507.
Begon, M., Harper, J. L. y Townsend, C. R. 1996. Ecology. 3a Edición.
Blanckewell Sciencie Oxford. 1068 pp.
Beard, J. S. 1955. The Classification of tropical American Vegetation Types.
Ecology 36: 89-100 pp.
Bergallo, H.D. 1994. Ecology of a small mamad community in an Atlantic
forest area in southeastern Brazil. Studies on Neotropical Fauna and Environment
29(4):197-217.
Brown, J. H. 1995. Macroecolgy. The University of Chicago Press. Chicago.
269 pp.
Brown, J. H. y Harney, B. A. 1993. Population and community ecology of
heteromyid rodents in temperate habitats. Pp 618-650. En Biology of the Heteromydae

(H. H. Genoways y J. H. Brown, eds.). Special Publication. The American Society of
Mammalogists 10:1-719.
Cameron, G.N. y Spencer, S.R. 1981. Sigmodon hispidus. Mammalian Species
158: 1-9.
Carey, A. B. y Jonson, M. L.. 1995. Small mammals in managed, naturally
young, and old-growth forest. Ecological Applications. 5 (2):336-352.
Ceballos, G. y Brown J. H. 1995. Global patterns of mammalian diversiy,
endemism, and endangerment. Conservation Biology, 9:559-568.
Ceballos, G. y Ecardi F. 1996. Diversidad de fauna Mexicana. 2ª edición. Edit.
CEMEX. México, D.F. 1991 pp.
Ceballos, G. y Galindo . C. L. 1984. Mamíferos silvestres de la cuenca de
México, Editorial Limusa. México. 149 pp.
Ceballos, G. y Oliva, G., coordinadores 2005. Los mamíferos silvestres de
México. Comisión Nacional para el Conocimiento y Uso de la Biodiversidad y Fondo
de Cultura Económica. México, D. F. 986 pp.
C. F. E. 1979. Estudio de gran visión de la cuenca del Río Usumacinta;
Departamento de Ingeniería Civil, Gerencia General de Estudios e Ingeniería
Preliminar; México, D. F.
Cruz-Lara, L. 2002. Diversidad de mamíferos en cafetales y selva mediana de
las cañadas de la Selva Lacandona, Chiapas, México. Tesis de Maestría. El Colegio de
la Frontera Sur. San Cristóbal de las Casas, Chiapas. 66 pp.
Cuaron, A.D. 2000. Effects of Land-Cover Changes on Mammals in a
Neotrópical Region: a Modeling Appach. Conservation Biology 14:1676-1692.
De Vos, J. 1996. Semblanza del Río Usumacinta. En Arqueología Mexicana;
Vol. IV, No. 22; INAH; Edit. Raíces, México; 54-63 pp.
Espinosa-Salvatierra, A. y Canela-Zárate, V. 1996. Pequeños mamíferos
¿Inofensivos para el hombre? Divulgare. 14:4-7. Universidad Autónoma de Baja
California.
Flores, M. 1991. “Noticias Sobre Cultivos de Cobertura”, Boletín Trimestral
No.5, CIDICCO, Honduras.
Flores, M. G., Jiménez L. J., Madrigal X., Moncayo R. F. y Takaki F. 1971.
Memoria del mapa de tipos de vegetación de la República Mexicana. Secretaría de
Recursos Hidráulicos. México D. F. 59 pp.

Flores-Villela, O. A. y Gerez, P. 1994. Biodiversidad y conservación en
México: Síntesis sobre vertebrados terrestres, vegetación y uso del suelo. 2ª edición.
Universidad Nacional Autónoma de México. Comisión Nacional para el Conocimiento
y Uso de la Biodiversidad. México. 439 pp.
Franco, L. J., G. de la Cruz, A., Cruz, G. A., Rocha, R. A., Navarrete, S. N.,
Flores, M. G., Kato, M. E., Sánchez, C. S., Abarca, A. L. G., Bedia, S. C. M. 1989.
Manual de Ecología. 2ª edición. Editorial Trillas. México. Pp 128, 129, 130, 132.
Gallina, S. Mandujano, S. y González-Romero, A. 1996. Conservation of
mammalian biodiversity in coffee plantations of central Veracruz, México. Agroforestry
Systems. 33:13-27.
García, E. 1973. Modificaciones al sistema de clasificación climática de
Köppen (adaptado a las condiciones de la República Mexicana). 2ª edición. Edit.
Instituto de Geog. UNAM. México D. F. 246 pp.
García-Gil, G.S., Vargas, M.A., Vazquez y Domínguez, R. 1988. Normas de
Ordenamiento Ambiental por actividades y sectores productivos en la Selva Lacandona.
Instituto Nacional de Investigación sobre Recursos Bióticos. Dirección General de
Normatividad y Regulación Ecológica (SEDUE). México D.F. 121pp.
Groombridge, B. (ed). 1992. International Union for the Conservation of
Nature Red List of thereatened mammals. IUCN-The Nature Conservation Union,
Gland, Zwitzerland.
Halffter, G. 1998. A strategy for measuring landscape biodiversity. Biology
International, 36: 3-17.
Halffter, G. y Escurra, E. 1992. ¿Qué es la biodiversidad? En: La diversidad
biológica de Iberoamérica I, G. Halffter (Comp). Acta Zoológica Volúmen Especial.
CYTED-D, Instituto de Ecología, Secretaría de Desarrollo Social, México. pp. 3-24.
Harris, S. y Woolard, T. 1990. The dispersal of mammals in agricultural
habitats in Britain. Pages 159-168 en R.G.H. Bunce and D.C. Howard, editors. Species
dispersal in Agricultural habitats. Institute of Terrestrial Ecology, Belhaven Press,
London, New York.
Heywood, V. H. 1994. The measurement of biodiversity and the politics of
implementation. En: Systematics and conservation evaluation, P. L. Forey, C. J.
Humphries y R. I. Vane-Wright (Eds). Systematics Association Special Vol. 50,
Claredon Press, Oxford. Pp 15-22.

Horváth, A., March, I.J. y Wolf, J.H.D. 2001. Rodent diversity and land use
in Montebello, Chiapas, México. Studies on Neotropical Fauna and Environment
36(3):169-176.
Horváth, A., Chávez, C., Nájera, J., Pérez, O., y Vidal, R. 2006. Evaluación
de la diversidad de roedores en cultivos manejados con abono verde en una localidad en
la Reserva de la Biosfera Montes Azules, Chiapas, México. Informe Técnico Final.
ECOSUR-Conservation International-USAID. San Cristóbal de las Casas, Chiapas.
México. 29 Pp.
Kelt, D. A. 1995. Ecology of small mammals across a strong environmental
gradient in southern Sounth America. Journal of Mammalogy, 77:205-219.
Krebs, C. 1985. Ecología: estudio de la distribución y abundancia. Editorial
Harla. México. Pp. 504, 505, 753.
Kuns, T. H., Wemmer CH., Hayssen V. 1996. Sex, Age and Reproductive
Condition in Mammals. En: Wilson, D. E., F. R. Cole, J. D. Nichols, R. Rudran y M. S.
Foster (Eds.). Measuring and Monitoring Biological Diversity. Standard Metrhods for
Mammals. Smithsonian Institution Press. Washington, USA. 405 pp.
Leopold, A. S. 1950. Vegetations zones of México Ecology. Pp 507-518.
Maffe, G. K. y Carroll, C. R. 1997. Principles of conservation biology. 2a
edición. Sinauer Associates, Inc. Publishers. Sunderland, Massachusetts, E. U. A. 729
pp.
Magurran, A. E. 1988. Ecological diversity and its measurement. Princeton
University Press, New Jersey, 179 pp.
Martínez-Gallardo, R. y Sánchez-Cordero, V. 1993. Dietary value of fruits
and seeds to Spiny Pocket Mice, Heteromys desmarestianus (Heteromyidae). Journal of
Mammalogy. 74(2): 436-442.
Mata-Sánchez, O. F. 1979. Consideraciones Climatológicas e Hidrológicas de
la Selva Lacandona. Facultad de Fisiología y Letras, Colegio de Geografía, UNAM.
México D. F. Tesis para obtener el titulo de Licenciado en Geografía. 104 pp.
Medellín, R. A. 1992. Community ecology and conservation of mammals in a
Mayan tropical rainforest and abandoned agricultural fields. Ph. D. dissertation,
University of Florida, Gainesuille, FL, USA. 333 pp.

Medellin, R.A. 1994. Mammals Diversity and conservation in the Selva
Lacandona, Chiapas, México. Conservation Biology. 8:3:780-799.
Mellink, E. 1991. Rodent communities associated with three traditional
agroecosystems in the San Luis Potosí Plateau, México. Agriculture, Ecosystems and
Enviroment, 33:363-375.
Mellink, E. 1995. Uso del hábitat, Dinámica poblacional y estacionalidad
reproductiva de roedores en el Altiplano Potosino, México. Revista Mexicana de
Mastozoología. 1:1-8.
Miranda, F. 1951. La vegetación de Chiapas. 1ª parte. 1ª edición. Edit.
Gobierno del Estado de Chiapas. Tuxtla Gutiérrez, Chiapas, México. 334 pp.
Miranda, F. y Hernández, X. 1963. Los tipos de vegetación de México y su
clasificación. Boletín Sociedad Mexicana de Botánica. México D. F. 28: 29-129 pp.
Mittermeier, R.A. 1988. Primate diversity and the tropical forest: case studies
of Brazil and Madagascar and the importance of megadiversity countries. Pp. 145-154
In: E.O. Wilson (ed). Biodiversity National Academic Press, Washington. 501 pp.
Mittermeier, R.A., Mittermeier y Robles G. P. 1997. Megadiversidad, los
paises biológicamente mas ricos del mundo. CEMEX, México. 501 pp.
Moreno, C. E. 2001. Método para medir la biodiversidad. CYTED-ORCYT-
Sociedad Entomológica Aragonesa. 82 pp.
Morton, S. R., Brown, J.H., Kelt, D. A. y Reid, J. R. W. 1994. Comparisons
of community structure among small mammals of North America and Australian
deserts. Australian Journal of Zoology 42:501-525.
Muñoz, A.A. Horvath, A. Vidal, R. Percino, D.R. Gozález, E. y Larrazaga,
V.S. 2000. Efectos de la fragmentacion del hábitat sobre la biodiversidad de la reserva
de la biosfera “El Triunfo”. Informe final ECOSUR-SIBEJ-TNC. San Cristóbal de las
Casas, Chiapas. México. 77 pp.
Pérez, C.R. 2005. Diversidad de Mamíferos en cuatro hábitat con diferente
grado de alteración en el área de el pozo, municipio de Berriozabal, Chiapas. Tesis de
Licenciatura. Universidad de Ciencias y Artes de Chiapas. 50 pp.
Ramírez-Lozano, M. 2002. Diversidad de Roedores en Cafetales de la Reserva
de la biosfera “El Triunfo” Chiapas, México. Tesis de Licenciatura. Benemérito
Universidad Autónoma de Puebla. 70 pp

Ramírez-Pulido, J.; Arroyo-Cabrales, J. y Castro- Campillo, A. 2005. Estado actual y relacion nomenclatural de los mamíferos terrestres de México. Acta
Zoológica Mexicana (n.s.) 21 (1): 21-82
Reid, F.A. 1997. A field guide to the mammals of central Americana of
southeast Mexico. New York Oxford. Oxford University Press. 334 pp.
Renata, O.G. y Lorenzo, C. 2002. Lista de los mamíferos terrestres de Chiapas:
Endemismo y Estado de Conservación. Acta Zoológica Mexicana. 85:25-49.
Ruebel, E. 1930. Pflanzangesellschaften der Erde Verlag. H. Huber. Bern,
Alemania. 464 pp.
Rzedowski, J 1978. Vegetación de México. Editorial Limusa. México.
Sánchez-Hernández, C., Romero-Almaraz, M.L., Colin-Martínez, H. y
García-Estrada, C. 2001. Mamíferos de cuatro áreas con diferente grado de alteración
en el sureste de México. Acta Zoológica Mexicana, (n.s.). 84:35-48.
SARH. Comisión del Plan Nacional Hidráulico 1984: Desarrollo Rural
Integral de la Selva Lacandona, México. México, D. F.
Seber, G.A.F. 1986. A review of estimating animal abundance. Biometrics
42:267-292.
Secretaria de Medio Ambiente, Recursos Naturales y Pesca. 1998. Campaña
nacional de restauración ecológica y contra el cambio del uso del suelo de áreas
afectadas por incendios forestales. México, D. F. 19 pp.
Smith, R. L. y Smith, T. M. 2001. Ecología. 4ª edición. Editorial Perarson
Educación, S. A. Madrid, España. Pp 304, 305, 306, 620.
Solbrig, O. T. 1991. From genes to ecosystems: a research agenda for
biodiversity. IUBSSCOPE-UNESCO, Cambridge, 124 pp.
UNEP. 1992. Convention on biological diversity. United Nations Environmental
Program, Environmental Law and Institutions Program Activity Centre. Nairobi.
Vázquez, L. B., Medillín, R. A. y Cameron, G. N. 2000. Population and
community ecology of small rodents in montane forest of western México. Journal of
Mammalogy, 81:77-85.
Whittaker, R. H. 1972. Evolution and measurement of species diversity. Taxon,
21(2/3) : 213-251.
Wood, B. J. 1994. Rodents in Agriculture and Forestry; in Rodent Pests and
Their Control, Buckle A.P. and R.H. Smith, Research and Development Manager
ZNECA public Healt U.K. 45 – 84 pp.

Zarza, H. 2001. Estructura de la comunidad de pequeños mamiferos en diversos
hábitat en la Selva Lacandona, Chiapas, México. Tesis de la Licenciatura. Universidad
Nacional Autónoma de México. 129 pp.
Zúñiga, M. J. 1996. Evaluación Rural Participativa como herramienta para
alcanzar el Desarrollo Sustentable en la Región Miramar en la Biosfera Montes Azules.
Tesis de Licenciatura, Escuela de Ciencias Agronómicas Campus V. UNACH. 53 pp.
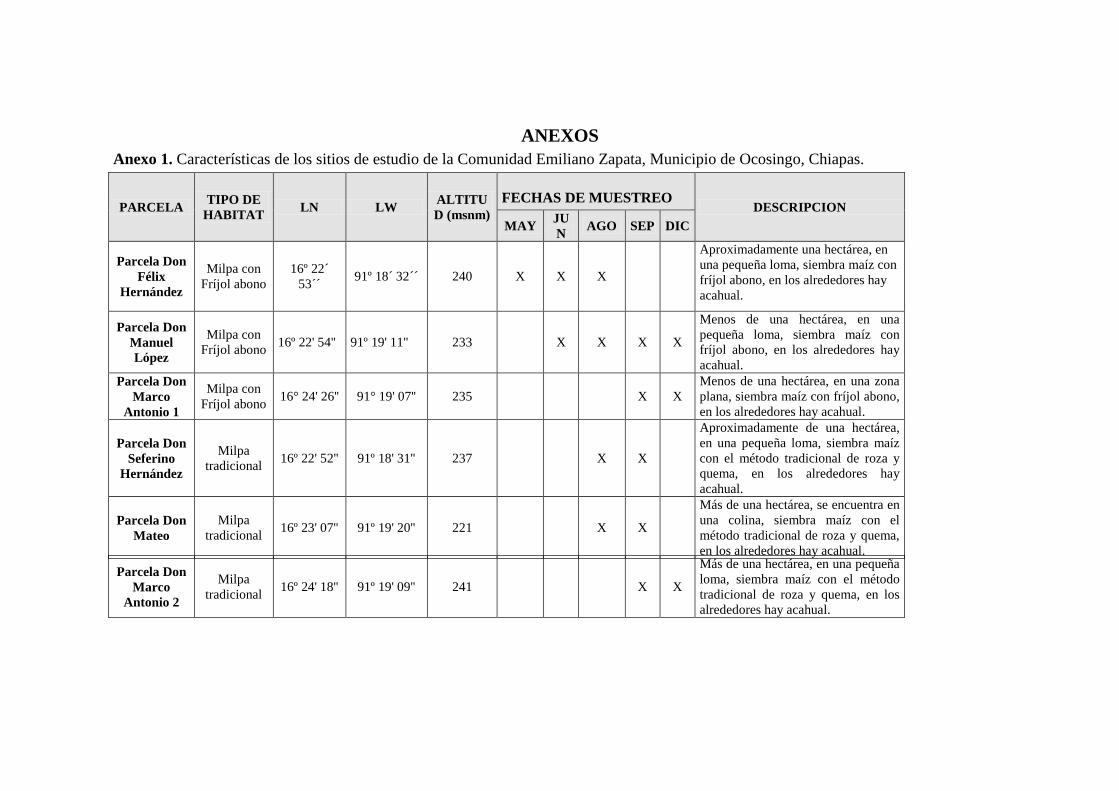
ANEXOS
Anexo 1. Características de los sitios de estudio de la Comunidad Emiliano Zapata, Municipio de Ocosingo, Chiapas.
PARCELA TIPO DE
HABITAT LN LW
ALTITU
D (msnm)
FECHAS DE MUESTREO DESCRIPCION
MAY JU
N AGO SEP DIC
Parcela Don
Félix
Hernández
Milpa con
Fríjol abono
16º 22´
53´´ 91º 18´ 32´´ 240 X X X
Aproximadamente una hectárea, en
una pequeña loma, siembra maíz con
fríjol abono, en los alrededores hay
acahual.
Parcela Don
Manuel
López
Milpa con
Fríjol abono 16º 22' 54'' 91º 19' 11'' 233 X X X X
Menos de una hectárea, en una
pequeña loma, siembra maíz con
fríjol abono, en los alrededores hay
acahual.
Parcela Don
Marco
Antonio 1
Milpa con
Fríjol abono 16° 24' 26'' 91° 19' 07'' 235 X X
Menos de una hectárea, en una zona
plana, siembra maíz con fríjol abono,
en los alrededores hay acahual.
Parcela Don
Seferino
Hernández
Milpa
tradicional 16º 22' 52'' 91º 18' 31'' 237 X X
Aproximadamente de una hectárea,
en una pequeña loma, siembra maíz
con el método tradicional de roza y
quema, en los alrededores hay
acahual.
Parcela Don
Mateo
Milpa
tradicional 16º 23' 07'' 91º 19' 20'' 221 X X
Más de una hectárea, se encuentra en
una colina, siembra maíz con el
método tradicional de roza y quema,
en los alrededores hay acahual.
Parcela Don
Marco
Antonio 2
Milpa
tradicional 16º 24' 18'' 91º 19' 09'' 241 X X
Más de una hectárea, en una pequeña
loma, siembra maíz con el método
tradicional de roza y quema, en los
alrededores hay acahual.

Anexo 2. Hoja de registro de especies y datos de campo.
HOJA DE REGISTRO
Fecha _______________ Localidad__________________ Sitio de Muestreo_____________________
Coordenada____________________________________ Estado de Tiempo_____________________
Lugar de Captura: Milpa con f/a ( ) Cultivo Tradicional ( )
No c/ind ca/re/co No. de
trampa Especie sexo edad Es/re LT LC LP LO peso obs

Anexo 3. Marcaje permanente en las orejas.