Embed Size (px)
Citation preview

1
UNIVERSIDADE CEUMA - UNICEUMA PRÓ-REITORIA DE PÓS-GRADUAÇÃO, PESQUISA E EXTENSÃO
PROGRAMA DE PÓS-GRADUAÇÃO EM BIOLOGIA PARASITÁRIA
Propriedades de Adesão e Atividade Inibitória de Espécies de
Lactobacillus sobre a Interação de Escherichia coli Diarreiogênicas em
Células Eucarióticas
São Luís
2014

2
FRANCISCA MARIA FERREIRA NORONHA
Propriedades de Adesão e Atividade Inibitória de Espécies de
Lactobacillus sobre a Interação de Escherichia coli Diarreiogênicas em
Células Eucarióticas
Dissertação apresentada ao Programa de Mestrado em
Biologia Parasitária da Universidade CEUMA, linha
de pesquisa em Patogenicidade Bacteriana, como
requisito para obtenção do título de Mestre.
Orientador:
Prof.. Dr. Valério Monteiro Neto
São Luís
2014

3
FRANCISCA MARIA FERREIRA NORONHA
Noronha, Francisca Maria Ferreira
Propriedades de Adesão e Atividade Inibitória de Espécies de Lactobacillus sobre a Interação de Escherichia coli Diarreiogênicas em Células Eucarióticas / Francisca Maria Ferreira Noronha. – São Luís: UNICEUMA, 2014.
75 f.il. Impresso por computador (Fotocópia) Dissertação (Mestrado) – Universidade CEUMA, Programa de
Mestrado em Biologia Parasitária, 2014.
1. Probióticos. 2. Lactobacillus. 3. Escherichia coli diarreiogênica. I.Monteiro Neto, Valério. (Orientador). II. Título
CDU 576.8
N852p

4
FRANCISCA MARIA FERREIRA NORONHA
Propriedades de Adesão e Atividade Inibitória de Espécies de
Lactobacillus sobre a Interação de Escherichia coli Diarreiogênicas em
Células Eucarióticas
A comissão julgadora de Defesa do Trabalho Final de Mestrado em Biologia
Parasitária, em sessão pública realizada no dia | | , considerou a candidata.
( )APROVADA ( )REPROVADA
1) Examinador: ______________________________________________________________
Profa. Dra. Elizabeth Soares Fernandes
2) Examinador: ______________________________________________________________
Profa. Dra. Maria Rosa Quaresma Bomfim
3) Examinador: ______________________________________________________________
Profa. Rosemary de Jesus Gomes Turri
4) Presidente (Orientador):_____________________________________________________
Prof. Dr. Valério Monteiro Neto

5
Aos meus queridos pais, José Ferreira e Maria José,
exemplos de vida e responsáveis por minha caminhada. Ao
meu grande amor, minha filha, Maria Fernanda.

6
AGRADECIMENTOS
A Deus, pela força e direcionamento nos momentos difíceis dessa longa caminhada.
Aos meus pais e familiares pela compreensão, incentivo e apoio.
Ao Professor Dr. Valério Monteiro Neto, pela orientação, paciência, amizade e valiosa
oportunidade.
À Professora Drª Maria Rosa Quaresma Bomfim pela amizade, apoio e atenção.
À Professora Drª Ana Lúcia Alves por ter disponibilizado o microscópio para fotografia
dos experimentos.
Ao Professor Dr. Jorge A. Girón pela doação das amostras de referência das Escherichia
coli.
Ao Professor Dr. Álvaro Cantini Nunes pela doação das amostras de Lactobacillus
isolados de fezes de cães.
A Universidade Ceuma – UNICEUMA, em especial ao Mestrado em Biologia Parasitária,
pela oportunidade de realizar este Curso de Mestrado.
Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico e à Fundação de
Amparo à Pesquisa e ao Desenvolvimento Científico e Tecnológico do Maranhão pela
concessão do financiamento da pesquisa.
Às Professoras Drª Maria Rosa Quaresma Bomfim e Drª Priscila Sabbadini pelas sugestões
na banca de qualificação.
À Professora Mekaele Frota do Vale pela revisão do trabalho e sugestões.
As amigas queridas, Cianna Rodrigues, Márcia Cristina Machado e Marinete Diniz pelo
apoio e carinho nessa trajetória.
Aos companheiros da pesquisa, especialmente: Monique Santos do Carmo, Thiago
Azevedo Feitosa Ferro, Ennio Patrezzi da Silva Costa e Soraia Silveira Beserra, pela
atenção, amizade, paciência, ensinamentos e grande participação no estudo.

7
Aos amigos de laboratório, Mariana, Marinaldo, Margareth, Ronildson e Stanley pela
amizade e contribuição na pesquisa.
Às minhas amigas e companheiras de trabalho, Amparo, Elenilde, Helde, Patrícia e Samila
pela paciência e apoio.
À Margarida Leite pela amizade e apoio ao longo da pesquisa.
A todos os Professores do Programa do Mestrado em Biologia Parasitária da Universidade
Ceuma - UNICEUMA pela atenção, ensinamentos e contribuição.

8
LISTA DE FIGURAS
Figura 01. Padrão de amplificação por PCR multiplex que identificam e diferenciam os
tipos de Escherichia coli........................................................................................................40
Figura 02. Aspectos Microscópicos das espécies de Lactobacillus após coloração de
Gram...................................................................................................................................41
Figura 03. Análise comparativa do perfil de hidrofobicidade de superfície celular, das
diferentes amostras de Lactobacillus de cães, de iogurtes e de cepas de referência na
presença de clorofórmio...................................................................................................45
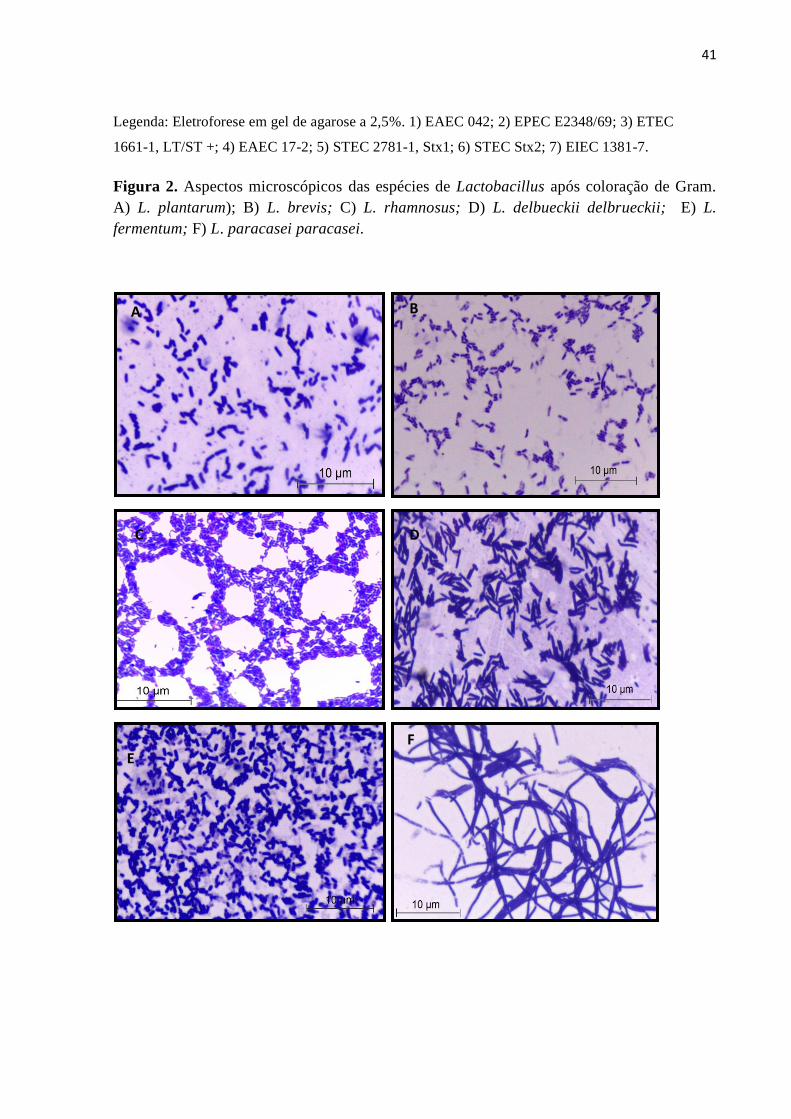
Figura 04. Avaliação da atividade inibitória de Lactobacillus sobre a adesão das
diferentes categorias diarreiogênicas de E. coli em células eucarióticas..........................50
Figura 05. Adesão em células HeLa de isolados de Lactobacillus plantarum e
Escherichia coli EIEC – 1381/7......................................................................................51
Figura 06. Adesão em células HT-29 de isolados de Lactobacillus plantarum e
Escherichia coli 042.........................................................................................................52
Figura 07. Adesão em células HT-29 de isolados de Lactobacillus brevis e Escherichia
coli 042..................................................................................................................................53
Figura 08. Adesão em células HeLa de isolados de Lactobacillus fermentum e
Escherichia coli ETEC LT/ST+.......................................................................................54
Figura 09. Correlação entre escore de coagregação entre cepas de E. coli e Lactobacillus
e a adesão em células HeLa e HT- 29 de Escherichia coli após competição com cepas de
Lactobacillus.....................................................................................................................55

9
LISTA DE TABELAS
Tabela 01. Análise microscópica da adesão das espécies de Lactobacillus de
referência, isolados de cães e de iogurtes em células HT-29.....................................42
Tabela 02. Quantificação da adesão das espécies de Lactobacillus de referência em
células HT-29............................................................................................................43
Tabela 03. Distribuição comparativa da hidrofobicidade da superfície celular das
espécies de Lactobacillus de referência (ATCC), isolados de cães e de iogurtes.....44
Tabela 04. Distribuição comparativa da produção de biofilme pelas amostras de
Lactobacillus de referência (ATCC), isolados de cães e de iogurtes........................46
Tabela 05. Ensaio de coagregação entre espécies de Lactobacillus e E. coli
diarreiogênicas............................................................................................................48

10
LISTA DE ABREVIATURAS E SIGLAS
ATCC - American Type Culture Collection
BHI – Brain Heart Infusion
DMEM – Dulbecco´s Modified Eagle Medium
D.O.c. – Densidade óptica do controle
D.O.i. – Densidade óptica do isolado
EAEC – Escherichia coli enteroagregativa
EDTA – Ethylenediamine tetraacetic acid
EIEC – Escherichia coli enteroinvasora
ELISA - Enzyme-linked Immunosorbent Assay
EPEC – Escherichia coli enteropatogênica
ETEC – Escherichia coli enterotoxigênica
FAO/WHO – Food and Agriculture Organization/ World Health Organization
LB – meio Luria-Bertani
MRS – meio de Man, Rogosa e Sharp (1960)
MH – agar Mueller-Hinton
NaOH – Hidróxido de sódio
OMS – Organização Mundial da Saúde
PBS – Phosphate-Buffered Saline
PCR - Polymerase Chain Reaction
STEC – Escherichia coli produtora de toxina de Shiga
TGI – Trato gastrointestinal
TSB – Tryptic Soy Broth
rpm - rotações por minuto
UFC/g – unidades formadoras de Colônias por grama

11
RESUMO
O trato gastrointestinal é constituído por uma microbiota considerada o ecossistema mais
complexo do corpo humano. Alguns membros da microbiota têm sido empregados como
probióticos. O uso de alimentos contendo bactérias probióticas é considerado uma
possibilidade de prevenir a instalação de um processo infeccioso. Desta forma, o presente
trabalho objetivou investigar as propriedades relacionadas à adesão, bem como a ação de
Lactobacillus na inibição da interação das espécies de Escherichia coli diarreiogênicas em
células intestinais. Foram avaliadas 16 espécies de Lactobacillus (linhagens de referência
(ATCC) e isolados de cães e de iogurtes) e sete amostras de Escherichia coli diarreiogênicas
quanto às propriedades de adesão e de inibição da adesão em células eucarióticas. O ensaio de
adesão evidenciou que todas as seis espécies de referência de Lactobacillus foram aderentes
em células HT-29 e HeLa, sendo que quatro espécies apresentaram melhor capacidade de
adesão. Nos isolados de cães, três amostras aderiram, enquanto que, entre os isolados de
iogurte, o L. casei foi o que apresentou a melhor capacidade de adesão. Na avaliação da
hidrofobicidade da superfície celular, constatou-se que quatro espécies de referência, quatro
espécies isoladas de cães e quatro linhagens isoladas de iogurtes eram hidrofóbicas. Das 16
espécies bacterianas, 13 (81,2%) isolados produziram biofilme em microplacas. No ensaio de
coagregação entre espécies de Lactobacillus e enteropatógenos foi verificado que a espécie
com maior capacidade de coagregação foi o L. plantarum. Constatou-se ainda que L.
fermentum e L. plantarum apresentaram atividade de coagregação contra todas as linhagens
de E. coli diarreiogênicas. A adesão de E. coli enterotoxigênica (ETEC) não foi inibida por
nenhuma espécie de Lactobacillus, no entanto, todas as espécies de Lactobacillus
conseguiram inibir a interação da E. coli enteroinvasora (EIEC) nas células HeLa e HT-29
(p<0.001). Na metodologia empregada, nenhuma espécie de Lactobacillus apresentou
atividade antimicrobiana do sobrenadante contra as espécies de E. coli diarreiogênicas
testadas. Os resultados sugerem que algumas espécies de Lactobacillus empregadas nesse
estudo possuem potencial probiótico, considerando-se a capacidade inibitória in vitro da
interação das categorias de E. coli diarreiogênica com células intestinais, contribuindo para a
implementação de futuras medidas de intervenção na doença diarréica, baseadas no uso de
alimentos com bactérias probióticas.
Palavras-chave: Probióticos. Lactobacillus. Escherichia coli diarreiogênica.

12
ABSTRACT
The gastrointestinal microflora is constituted by a complex ecosystem of the human body.
Some members of the microbiota have been used as probiotics. The use of food products
containing probiotic bacteria is considered a possibility to prevent the onset of an infections.
Thus, the present study aimed to investigate the properties related to adhesion, as well as the
action of Lactobacillus in the inhibition of the interaction of species of diarrheagenic
Escherichia coli in intestinal cells. 16 species of Lactobacillus (reference strains (ATCC) and
isolated from dogs and yogurts) and seven Escherichia coli diarrheagenic properties regarding
adhesion and adhesion inhibition in eukaryotic cells were evaluated. Results show that all six
species of Lactobacillus reference were adherent in HT-29 and HeLa cells, four species
showed better adhesion ability. In isolated dog, joined three samples, whereas among isolates
of yogurt, L. casei showed the best adhesion ability. In assessing the hydrophobicity of the
cell surface, it was found that four species of reference, four species isolated from dogs and
four strains isolated from yogurts were hydrophobic. Of the 16 bacterial species, 13 (81.2%)
isolates produced biofilms in microplates. In the assay of coaggregation among Lactobacillus
species and enteropathogens was found that showed the greatest ability to coaggregation was
L. plantarum. It was also found that L. fermentum and L. plantarum showed coaggregation
activity against all strains of diarrheagenic E. coli. The adhesion of enterotoxigenic E. coli
(ETEC) was not inhibited by any species of Lactobacillus, however, all Lactobacillus species
able to inhibit the interaction of enteroinvasive E.coli (EIEC) in HeLa cells and HT-29 (p
<0.001 cells ). In methodology, no Lactobacillus species showed antimicrobial activity
against species of the supernatant of E. coli diarrheagenic tested. The results suggest that
some species of Lactobacillus probiotic used in this study have potential, considering the
inhibitory capacity in vitro interaction of the categories of diarrheagenic E. coli with intestinal
cells, contributing to the implementation of future intervention measures on diarrheal disease,
based the use of probiotic bacteria in food.
Keywords: Probiotics. Lactobacillus. Diarrheagenic Escherichia coli.

13
SUMÁRIO
1 INTRODUÇÃO ................................................................................................................. 15
2 REFERENCIAL TEÓRICO ............................................................................................. 17
2.1 Probióticos ......................................................................................................................... 18
2.1.1 Lactobacillus spp ............................................................................................................. 19
2.1.2 Mecanismo de ação dos probióticos ................................................................................. 20
2.2 Características das Escherichia coli diarreiogênicas ...................................................... 24
2.3 Aspectos tecnológicos-industriais ..................................................................................... 26
3 OBJETIVOS ....................................................................................................................... 27
3.1 Geral ................................................................................................................................... 27
3.2 Específicos .......................................................................................................................... 27
4 MATERIAL E MÉTODOS ............................................................................................... 28
4.1 Amostras bacterianas ........................................................................................................ 28
4.1.1 Identificação dos tipos de Escherichia coli por reação em cadeia de polimerase em
multiplex ........................................................................................................................................ 29
4.2 Isolamento das bactérias de iorgute ....................................................................................30
4.3 Isolamento de bactérias láticas a partir das fezes de cães lactentes ................................ 30
4.4 Cultura e preparo de células HT-29 e HeLa ...................................................................... 31
4.5 Ensaio de adesão em células HT-29 e HeLa ....................................................................... 31
4.5.1 Preparo das amostras bacterianas para ensaio de adesão .................................................... 31
4.5.2 Ensaio de adesão bacteriana........................... ...................................................................... 32
4.5.3 Quantificação da adesão ....................................................................................................... 33
4.5.3.1 Quantificação do inóculo das amostras bacterianas ....................................................... 33
4.5.3.2 Quantificação bacteriana após ensaio de adesão ............................................................ 34
4.5.3.3 Quantificação bacteriana após ensaio de competição .................................................... 35
4.6 Hidrofobicidade da superfície celular ................................................................................. 35
4.7 Ensaio de produção de biofilme dos Lactobacillus ............................................................ 37
4.8 Ensaio de coagregação .......................................................................................................... 37
4.9 Análise da atividade do sobrenadante ................................................................................ 38
4.10 Análises estatísticas ............................................................................................................. 39
5 RESULTADOS ....................................................................................................................... 40
5.1 Confirmação dos enteropatógenos e das linhagens dos Lactobacillus ............................. 40

14
5.2 Ensaio de adesão em células HeLa e HT-29 ........................................................................ 42
5.3 Hidrofobicidade da superfície celular de Lactobacillus spp ............................................. 43
5.4 Produção de biofilme .............................................................................................................46
5.5 Ensaio de coagregação ........................................................................................................... 47
5.6 Analise quantitativa de atividade inibitória de Lactobacillus spp sobre a adesão de
E.coli...................... ........................................................................................................................ 49
5.7 Análise da atividade do sobrenadante ................................................................................. 56
6 DISCUSSÃO............................................................................................................................ 57
7 CONCLUSÃO ......................................................................................................................... 61
REFERÊNCIAS ....................................................................................................................... 63

15
1 INTRODUÇÃO
O trato gastrointestinal é constituído por um ecossistema muito complexo, cujo papel
essencial é digerir e absorver nutrientes importantes para as necessidades metabólicas do
organismo. Ao longo do tubo digestivo, existe uma diversidade na quantidade e nas espécies
que compõem a microbiota intestinal e esta população bacteriana funciona como uma defesa
protetora do indivíduo contra agentes patogênicos (BACKHED et al., 2005; ZOETENDAL et
al., 2006). Algumas situações decorrentes de fatores específicos, como estresse, uso excessivo
de antibióticos, alimentação inadequada, dentre outros, podem levar a alterações no aparelho
digestivo, tornando-o um hospedeiro suscetível à invasão por enteropatógenos (COLLIER-
HYAMS; NEISH, 2005; KELLY et al.,2005).
Algumas espécies de micro-organismos apresentam uma capacidade de aderência
relevante no trato gastrointestinal e conferem benefícios ao hospedeiro, sendo estas definidas
como probióticos (FAO/WHO, 2002). Os probióticos são micro-organismos que
desempenham papel importante na modulação da microbiota intestinal, proporcionando
reações benéficas ao organismo humano (VARAVALLO, THOMÉ, TESHIMA, 2008).
Os micro-organismos, para preencherem os pré-requisitos que os caracterizam como
probióticos, devem ser efetivos e seguros, não patogênicos, assim como resistentes contra a
digestão por enzimas pancreáticas ou entéricas, ao ácido gástrico e suco biliar, ter habilidade
para prevenir a aderência à mucosa intestinal do patógeno, capacidade de persistir no trato
gastrointestinal e de influenciar atividade metabólica local, além de evitar o estabelecimento
e/ou a replicação de patógenos no trato gastrintestinal (SAAVEDRA, 1994).
As bactérias probióticas, geralmente estudadas, pertencem ao gênero Lactobacillus e
Bifidobacterium, que apresentam um histórico de segurança no uso. Umas das principais
funções dos probióticos é a competição por sítios de adesão com os micro-organismos
enteropatogênicos, compondo a primeira linha de defesa intestinal impedindo que patógenos
colonizem o epitélio intestinal. Neste contexto, a indústria de laticínios encontrou nos
probióticos uma ferramenta para o desenvolvimento de novos produtos. Os iogurtes,
entretanto, em meio aos vários produtos alimentícios, têm sido muito utilizados como
veículos mais populares para a incorporação desses micro-organismos (KOMATSU, BURITI,

16
SAAD, 2008).
Considerando-se que a doença diarreica é responsável por elevadas taxas de
morbidade e mortalidade - em particular, com o envolvimento das categorias diarreiogênicas
de Escherichia coli, e que os Lactobacillus têm sido descritos como micro-organismos com
elevado potencial probiótico, este estudo teve por objetivo investigar se algumas espécies de
Lactobacillus de referência, isolados de iogurte e de animais apresentam características
probióticas, bem como a capacidade de interferir na adesão das espécies de Escherichia coli
diarreiogênicas.

17
2 REFERENCIAL TEÓRICO
A microbiota intestinal humana começa a ser formada no momento do nascimento,
por meio do contato do recém-nascido com o canal vaginal, com a pele da mãe, com as
mãos das enfermeiras e com o ar do ambiente. Trata-se de um sistema complexo que
apresenta uma grande influência na saúde humana, sendo conhecido por contribuir para a
maturação do intestino, nutrição do hospedeiro, resistência a patógenos e manutenção da
saúde do hospedeiro (MACKIE et al., 1999; KLINGER et al., 2007).
O trato gastrointestinal humano pode sofrer uma série de patologias, principalmente
de natureza infecciosa. Dentre estas, a doença diarreica permanece como a principal causa
de morte, especialmente entre crianças menores de cinco anos em países em
desenvolvimento (BRYCE et al., 2005). Apesar da redução significativa nos índices de
mortalidade infantil por diarreia, reduzindo de 4,6 milhões por ano para aproximadamente
2,5 milhões em todo o mundo, tais valores ainda são considerados altos, demonstrando que
os índices de morbidade ainda permanecem tão elevados quanto os observados há 30
anos (KOSEK et al., 2003). Altas taxas de morbidade por doença diarreica são preocupantes
porque a diarreia infantil pode deixar sequelas que se manifestarão em longo prazo sobre o
desenvolvimento físico e cognitivo da criança (BLACK et al., 1984; GUERRANT et al.,
1999).
A diarreia pode ser causada por vários micro-organismos infecciosos, incluindo:
vírus, bactérias, protozoários e helmintos, tendo como a principal via de transmissão a rota
fecal-oral. Estes patógenos diferem no seu modo de transmissão e na dose necessária para
causar infecção. Entre as bactérias, a capacidade de sobreviver ao ácido estomacal
juntamente ao tamanho do inóculo é condição crucial para causar doença (JAMISON et al.,
2006).
As linhagens diarreiogênicas da bactéria Escherichia coli são categorizadas segundo
suas propriedades de virulência, manifestações clínicas e aspectos epidemiológicos. As
propriedades ou fatores de virulência são definidos como estruturas, produtos ou estratégias
que contribuem para que o micro-organismo aumente sua capacidade de causar infecção.
Entre estas propriedades, destacam-se aquelas que permitem à bactéria reconhecer e colonizar

18
a superfície das células do hospedeiro (adesinas fimbriais e não fimbriais); a produção de
toxinas e hemolisinas; expressão de sistemas de captação de ferro (sideróforos); e
resistência a antibióticos e ao sistema imune (BUERIS et al., 2007).
A alteração da microbiota intestinal vem sendo reconhecida nos últimos anos como
fonte de doenças alérgicas e autoimunes, bem como de infecções agudas e crônicas
(BENGMARCK; ORTIZ DE URBINA, 2005). Além disso, estudos in vitro e in vivo têm
demonstrado que o ecossistema bacteriano intestinal normal representa uma barreira
extremamente efetiva em oposição a micro-organismos patogênicos e oportunistas (FULLER,
1989).
2.1 Probióticos
O termo probiótico recebeu diferentes acepções ao longo do tempo, desde substâncias
secretadas por protozoários e suplementos alimentares para animais até se chegar ao conceito
mais aceito, de que probióticos são produtos ou culturas de micro-organismos viáveis que
podem se implantar ou colonizar em um tecido do hospedeiro, trazendo benefícios à sua saúde
(OYETANO; OYETANO, 2005; COPPOLA, TURNES, 2004; TANNOCK, 2004).
Candidatos a probióticos devem ter certas propriedades, tais como a tolerância às
condições gastrointestinais (ácido e bile), alta capacidade de aderir à mucosa gastrointestinal e
exclusão competitiva de patógenos (BHATIA, KOCHAR, ABRAHAM, 1989;
OUWEHAND, SALMINEN, ISOLAURI, 2002; SAARELA et al., 2000). Tem sido relatado
que a adesão à mucosa intestinal, seguida por colonização transitória, é necessária para que os
micro-organismos com propriedades probióticas exerçam seus efeitos benéficos à saúde do
hospedeiro, pois prolongará o tempo de contato com o hospedeiro. Estes efeitos sobre a saúde
incluem a modulação do sistema imune e antagonismo contra patógenos, competindo pelos
sítios de ligação e de nutrientes na mucosa intestinal (HIRANO et al., 2003).
As pesquisas com probióticos têm progredido consideravelmente e avanços
significativos foram feitos na seleção e caracterização de culturas probióticas para uso
específico (FAO/WHO, 2002). A terminologia “probiótico” deriva do grego e significa “pró –

19
vida” e foi, inicialmente, introduzida na literatura médica por Lilly e Stillwel (1965 apud
MORAIS; JACOB, 2006). Em particular, as espécies pertencentes aos gêneros
Bifidobacterium e Lactobacillus predominam na microbiota gastrointestinal, sendo as
bactérias probióticas mais amplamente utilizadas, e estão incluídas em muitos alimentos
funcionais e suplementos alimentares (GUARNER; MALGELADA, 2003; MACPHERSON;
HARRIS, 2004; FRICK et al., 2007).
Os micro-organismos utilizados como probióticos devem ser avaliados para uso geral e,
portanto, seguros para os consumidores. São usualmente componentes não patogênicos da
microbiota normal, tais como as bactérias acido lácticas do gênero Lactobacillus,
Bifidobacterium (GROSSO; TRINDADE, 2004), Lactococcus (DESMOND et al., 2004),
Leuconostoc, Enterococcus, Streptococcus e leveduras como a Saccharomyces boulardii,
entre outras (PRIDMORE et al., 2004). Segundo a literatura, os gêneros Bifidobacterium e
Lactobacillus são altamente relevantes para a prevenção da invasão dos tecidos por
enteropatógenos (FOOKS; GIBSON, 2002).
No processo de seleção de um micro-organismo probiótico, devem ser considerados: I)
origem da estirpe; II) capacidade de aderir à mucosa intestinal; III) resistência aos sais
biliares; IV) resistência ao suco gástrico; V) capacidade de produzir substâncias
antimicrobianas; VI) antagonismo a patógenos; VII) ausência de patogenicidade; VIII)
ausência de genes de resistência a antibióticos; IX) ausência de virulência; X) incapacidade de
produzir metabólitos tóxicos ao homem; XI) incapacidade de translocar ou induzir a
translocação de micro-organismos patogênicos (FAO/WHO, 2002).
Os probióticos são utilizados como reguladores da microbiota intestinal, agindo como
agentes profiláticos em desordens intestinais em geral (DUC et al., 2004) e terapêuticos na
recuperação das comunidades microbianas endógenas da mucosa gastrointestinal e vaginal
(NAVA; DAVILA, 2004).
2.1.1 Lactobacillus spp.
O gênero bacteriano Lactobacillus é o mais estudado devido a suas propriedades
probióticas. Eles habitam o trato gastrointestinal e o genital feminino de mulheres saudáveis,

20
constituindo a maior parte da microbiota endógena vaginal e endocervical (ZHOU et al.,
2007).
O gênero Lactobacillus foi descrito, pela primeira vez, por Beijerinck em 1901. As
espécies deste gênero foram agrupadas com base em características fenotípicas, de acordo
com a temperatura para um bom crescimento e a fermentação de hexoses (ORLA-JENSEN,
1919) e, posteriormente, como homofermentativos obrigatórios, ou seja, fermentadores de
glicose exclusivamente em ácido láctico, e como heterofermentativos facultativos, os quais
fermentam hexoses em ácido láctico e podem produzir gás a partir de gliconato, mas não
através da glicose. Os heterofermentativos obrigatórios incluem os Lactobacillus que
fermentam hexoses em ácido láctico, ácido acético e ou etanol e dióxido de carbono, sendo a
produção de gás a partir da glicose (KANDLLER; WEISS, 1986).
De acordo com a classificação taxonômica dos procariotos, o gênero Lactobacillus
pertence ao filo Firmicutes, classe Bacilli, ordem Lactobacillales, família Lactobacillaceae
(HAMMES; HERTEL, 2003). No momento, o gênero Lactobacillus inclui 106 espécies
descritas de forma válida, e são, portanto, os mais numerosos gêneros da ordem dos
Lactobacillales. Essas bactérias são encontradas em ambientes onde os carboidratos estão
disponíveis, tais como produtos alimentares (lácteos, carne fermentada, massas ácidas,
vegetais, frutas, bebidas), no trato respiratório, gastrointestinal e genital e em esgotos (FELIS;
DELLAGLIO, 2007). As bactérias lácticas inibem o crescimento de vários micro-organismos
indesejáveis e patogênicos nos alimentos. Além de lhes conferir características sensoriais
desejáveis, estendem a sua vida útil e melhoram a sua qualidade sanitária. Isto se deve aos
metabólitos produzidos por estes micro-organismos, principalmente os ácidos orgânicos e as
bacteriocinas (PRADO et al., 2000).
2.1.2 Mecanismos de ação dos probióticos
Existem vários produtos probióticos disponíveis comercialmente, com características
distintas. Embora o uso de probióticos pareça promissor, é preciso cautela para escolher o melhor
probiótico para uma determinada situação (NAVA; DAVILA, 2004). Um probiótico deve se
enquadrar em pelo menos uma das seguintes categorias: I) inibidor da proliferação de

21
patógenos e restaurador da homeostase da microbiota através de interações micróbio-
micróbio; II) promotor da função de barreira epitelial; ou III) modulador da resposta imune do
hospedeiro (LEBEER et al., 2008). Embora existam diversos estudos sobre o efeito imuno-
modulador dos micro-organismos considerados probióticos, as formas de ação ainda estão
pouco esclarecidas, sugerindo a necessidade de um aprofundamento científico nesta área
(COPPOLA; TURNES, 2004; KANAMINOGAWA; NANNO, 2004).
A atividade antagonista dos probióticos pode acontecer de diversas maneiras:
competição por nutrientes, produção de antimicrobianos e exclusão competitiva. As
linhagens de lactobacilos mais competitivas apresentam melhores chances de adesão ao
epitélio do intestino em detrimento de patógenos invasivos, promovendo a eliminação
destes. Existem pesquisas avaliando os Lactobacillus que possuem sítios de ligação em
comum na mucosa intestinal com patógenos bacterianos como E. coli tipo 1 que se ligam a
sítios receptores de oligossacarídeos (LEBEER et al., 2008).
A atividade antimicrobiana dá-se pela produção de ácido láctico e outros ácidos
orgânicos, peróxido de hidrogênio e bacteriocinas. A síntese de ácido láctico pela linhagem
GG de L. rhamnosus apresenta grande atividade antimicrobiana contra Salmonella
Typhimurium, atuando no crescimento do patógeno, bem como sobre os fatores de
virulência deste (DE KEERSMAECKER et al., 2006).
A produção de metabólitos, como ácidos orgânicos, incluindo ácido láctico e ácido
acético, de peróxido de hidrogênio e de bacteriocinas confere efeito protetor ao ambiente
intestinal do hospedeiro (JACK et al., 1995). Bacteriocinas são proteínas ou peptídeos
antimicrobianos, secretadas por bactérias, sendo ativos contra bactérias patogênicas
(KLAENHAMMER, 1993; KAISER; MONTVILLE, 1996; CLEVELAND et al., 2001). A
atividade das bacteriocinas pode ser ainda por exclusão competitiva ou pela promoção da
função de barreira epitelial, ocorrendo pela competição entre as linhagens patogênicas e
aquelas com propriedades probióticas pelos mesmos sítios de adesão na mucosa intestinal
ou por nutrientes (LEBEER et al., 2008).
Estudos demonstraram que certas linhagens de Lactobacillus são capazes de induzir
a produção de mucinas (MACK et al., 2003) e estimular a adesão célula a célula (SETH et
al., 2008) e prevenir a apoptose pelas células intestinais (YAN et al., 2007), contribuindo e
promovendo a manutenção da barreira epitelial. A adesão é considerada uma etapa crítica do

22
processo infeccioso. Caso o patógeno não consiga aderir às células epiteliais, pode
simplesmente ser eliminado pelos mecanismos de defesa do hospedeiro, tais como compostos
orgânicos ou inorgânicos com atividade antimicrobiana, variações de pH, dentre outros
(SPURBECK; ARVIDSON, 2010).
Segundo Jacobsen et al. (1999) o maior benefício do uso de probióticos está
relacionado à aderência dos micro-organismos ao muco e ao epitélio intestinal. Algumas
espécies bacterianas com propriedades probióticas ao colonizarem a mucosa intestinal,
aderem a esse tecido, estabilizando a microbiota local dos animais durante as fases de
desenvolvimento e do organismo adulto (PENÃ et al., 2004). A presença de micro-
organismos com propriedades probióticas no intestino é de grande relevância, considerando
que estes compõem um ecossistema, impedindo a colonização por espécies patogênicas
como Listeria monocytogenes, E. coli, Salmonella spp, entre outras, através de um efeito
competitivo ou produzindo compostos inibitórios (GRANATO et al., 2004).
Dificuldades nas pesquisas sobre a adesão bacteriana in vivo, especialmente em
humanos, têm despertado o interesse em se desenvolver modelos in vitro para definição de
linhagens potencialmente aderentes (MAYRA-MAKINEN et al., 1983; CONWAY;
KJELLEBERG, 1989; KIMOTO et al., 1999; LAPARRA; SANZ, 2009; DUARY et al.,
2011). Desse modo, é necessário caracterizar a adesão bacteriana com modelos eficientes in
vitro (VAN DEN ABBEELE et al., 2009).
Os diferentes métodos no estudo da adesão bacteriana in vitro podem fornecer
informações mais completas em relação à habilidade de aderência de diferentes bactérias,
além de ajudar a elucidar os tipos de interações e moléculas que medeiam à relação micro-
organismo/hospedeiro. A propriedade de exclusão ou redução da aderência de
enteropatógenos já foi demonstrada para vários probióticos, provavelmente sendo
consequência do bloqueio de receptores que seriam utilizados por estes. A importância
dessa propriedade é óbvia, pois ela contribui para o efeito protetor dos probióticos contra
infecções intestinais (LAPARRA; SANZ, 2009).
Os Lactobacillus apresentam dois mecanismos de defesa contra a ação dos micro-
organismos patogênicos: 1- competição por receptores de células epiteliais e coagregação com
os patógenos, podendo levar à formação de biofilme; 2- produção de componentes
antimicrobianos (LEPARGNEUR; ROUSSEAU, 2002; ANTONIO et al., 2005).

23
A hidrofobicidade é uma força atrativa que ocorre entre as moléculas hidrofóbicas da
superfície das células bacterianas e superfícies inertes ou bióticas, formando uma interação
atrativa reversível (JONES et al., 1996). A hidrofobicidade da superfície celular microbiana,
apesar de ser uma interação inespecífica, é um dos fatores determinantes da adesão
bacteriana em superfícies (VAN LOOSDRECHT et al., 1987).
A adesão bacteriana depende, em parte, de interações reversíveis ou irreversíveis. O
estágio inicial e reversível é mediado por um complexo de interações físico-químicas,
incluindo hidrofobicidade e cargas, que não são consideradas específicas, mas, propriedade
importante (PELLETIER, 1997). A aderência é aceita como o primeiro passo no processo
de colonização de superfícies inertes e nas células epiteliais do hospedeiro e as propriedades
físico-químicas da superfície podem exercer uma forte influência sobre a adesão dos micro-
organismos, os quais aderem com maior facilidade em superfícies hidrofóbicas do que nas
hidrofílicas (BUSSCHER et al., 1990).
Estudos têm demonstrado que os Lactobacillus são capazes de formar biofilmes, que
são definidos como sistemas biológicos estruturalmente complexos e dinâmicos de bactérias
aderidas em superfícies (WHITTAKER, KLIER, KOLENBRANDER, 1996). A tendência
das bactérias aderirem e formarem biofilmes em diversos ambientes está relacionada às
características adquiridas através de bilhões de anos de adaptação evolucionária. A
formação do biofilme é um componente integrante do ciclo de vida dos procariotos,
apareceu no período fóssil (3,25 bilhões de anos atrás) em ambientes hidrotermais da África
do Sul e da Austrália (STOODLEY, COSTERTON, STOODLEY, 2004). Com a formação
do biofilme, as bactérias podem colonizar praticamente todos os tipos de materiais e se
desenvolverem em quase todos os tipos de superfície sem a necessidade de um local com
água em abundância. Desta forma, representam um modo protegido de crescimento que
permite que as células sobrevivam em ambientes hostis (CASTONGUAY et al., 2006).
A capacidade de coagregação demonstra a interação entre diferentes micro-
organismos. Experiências com coagregação mostraram que a capacidade de Lactobacillus
para se ligar a um agente patogênico varia de acordo com a espécie de Lactobacillus e o
agente patogênico implicado. A coagregação pode ser um importante fator na manutenção e
estabilidade da flora vaginal, devido ao fato de que Lactobacillus spp. podem gerar um
microambiente em torno do patógeno onde a concentração de substâncias inibitórias
produzidas é grande (MASTROMARINO et al., 2002).

24
2.2 Características das Escherichia coli diarreiogênicas
A bactéria E. coli é um bacilo Gram - negativo, que não forma esporo, usualmente
móvel, devido à presença de flagelos peritríquios, o qual frequentemente apresenta fímbrias
e cápsula. É um anaeróbio facultativo da família Enterobacteriaceae que habita o cólon
intestinal, desempenhando um papel fundamental na fisiologia e manutenção da microbiota,
capaz de fermentar glicose com produção de ácido e gás. As linhagens envolvidas em
processos patogênicos são morfológicas e bioquimicamente indistinguíveis das não
patogênicas (DRASAR, HILL, 1974; KAPER, NATARO, MOBELY, 2004).
Algumas linhagens podem proporcionar benefícios ao hospedeiro. Estas, geralmente,
fazem parte da microbiota normal do trato intestinal e ajudam a prevenir possíveis
infecções por bactérias patogênicas, além de sintetizarem vitaminas úteis ao bom
funcionamento do organismo. Entretanto, algumas linhagens patogênicas podem provocar
diversos cenários clínicos, como: meningites, infecção do trato urinário, diarreia e
sepse (FERRIERES, HANCOCK, KLEMM, 2007; SHARMA, YADAV, CHAUDHARY,
2009).
E. coli diarreiogênica é classificada com base em seus fatores de virulência e
manifestações clínicas. Dentre elas, duas linhagens destacam-se como enteropatógenos: a
enteropatogênica (EPEC) e a enteroagregativa (EAEC). A EPEC é o principal agente
causador de diarreia em crianças menores de doze meses de idade. Este patógeno causa
lesão histopatológica conhecida como attaching and effacement (lesão A/E). Todas as
linhagens apresentam uma ilha de patogenicidade cromossômica, denominada locus of
enterocyte effacement (região LEE) (BUERIS et al., 2007).
A EAEC tem emergido como um importante patógeno em vários cenários clínicos,
incluindo diarreia do viajante e diarreia persistente. É definida como a linhagem que não
secreta as toxinas termoestáveis (ST) e termolábil (LT) da E. coli enterotoxigênica (ETEC)
e caracterizada pelo padrão de aderência agregativa a culturas de células HEp-2 (NATARO;
KAPER, 1998).
Devido ao seu padrão de virulência heterogêneo, a EAEC pode causar uma ampla
variedade de danos ao hospedeiro: encurtamento das microvilosidades intestinais, necrose

25
hemorrágica das pontas das vilosidades, resposta inflamatória média com edema e
infiltração mononuclear da submucosa (NATARO, STEINER, GUERRANT, 1998). A
EAEC é capaz de colonizar superfícies através de sua adesão e consequente
desenvolvimento de biofilme, que pode resultar em maior persistência no ambiente
(CASTONGUAY et al., 2006).
A ETEC produz sintomatologia similar à doença cólera, caracterizada por diarreia
aquosa, desidratação e vômito. O período de incubação varia de 8 a 44 horas, com média de
26 horas, sendo a duração da doença curta, de aproximadamente 24 a 30 horas. Produz uma
ou mais toxinas, que vão agir no intestino delgado. Não ocorre invasão nem dano à camada
epitelial do intestino delgado, apenas ocorre colonização. Para a ocorrência de diarreia, o
micro-organismo deve ser toxigênico. Pode haver produção de um ou dois tipos de toxinas,
dependendo do plasmídio contido (DOYLE; CLIVER, 1990).
A E. coli enteroinvasora (EIEC) apresenta características bioquímicas, genéticas e
patogênicas muito semelhante a Shigella spp. O período de incubação varia de 8 a 24 horas,
com média de 11 horas e a duração da doença é usualmente de vários dias. Os sintomas são:
calafrios, febre, dores abdominais e disenteria. O micro-organismo invade células do epitélio
intestinal, provocando a lise do vacúolo endocítico; multiplica-se dentro destas células e
invade células epiteliais adjacentes, provocando ulcerações do cólon, resultando, finalmente,
em diarreia com sangue. A EIEC produz enterotoxinas secretoras que podem desempenhar
um importante papel na patogênese da diarreia (NATARO; KAPER, 1998). A EIEC pode
provocar uma forma leve de diarreia ou disenteria, que se caracteriza pelo aparecimento de
sangue e muco nas fezes de indivíduos infectados ou por colite. Até 10 % da disenteria bacilar
causada por EIEC ocorre entre 12 e 72 h após a ingestão de alimentos contaminados
(WANGER et al.,1988).
A Síndrome hemolítico-urêmica (SHU) é a principal causa de insuficiência renal
aguda em crianças, e tem sido fortemente associada a E. coli produtora da toxina de
Shiga (STEC). Cerca de 3 dias após a ingestão de alimentos ou de água contaminados, vai
induzir uma diarreia que pode ser seguida por colite hemorrágica e síndrome hemolítico-
urêmica. A toxina produzida pela bactéria pode atravessar a barreira intestinal, entrando na
circulação sistêmica e chegando ao rim e a outros órgãos (RITCHIE et al., 2003).

26
2.3 Aspectos tecnológico-industriais
Os micro-organismos probióticos vêm sendo empregados na medicina humana para a
prevenção e o tratamento das disfunções da microbiota intestinal, distúrbios de
metabolismo, para imunomodulação e como anticarcinogênicos (COPPOLA; TURNES,
2004).
Um estudo da Probiotics Market (2009 – 2014), desenvolvido pela empresa de
consultoria Markets and Markets, relatou que o mercado mundial de probióticos alcançou
ganhos de 15,9 bilhões de dólares no ano de 2008 e tende a alcançar 32 bilhões de dólares
até 2014, oferecendo oportunidades para o desenvolvimento de alimentos funcionais para
humanos e animais, terapias e prevenção de doenças. As principais empresas que trabalham
com bactérias probióticas, na atualidade, são a Danisco, a Morinaga e a Biogaia, que
fornecem produtos para várias outras grandes companhias alimentícias, como a Nestlé. No
Brasil, a tendência é semelhante, pois a Danone e a Actimel deverão ter um crescimento
anual em torno de 8,7% (INPI, 2011). As bactérias lácticas inibem o crescimento de vários
micro-organismos indesejáveis e patogênicos nos alimentos. Além de lhes conferir
características sensoriais desejáveis, estendem a sua vida útil e melhoram a sua qualidade
sanitária. Isto se deve aos metabólitos produzidos por estes micro-organismos,
principalmente os ácidos orgânicos e as bacteriocinas (PRADO et al., 2000).
Os aspectos tecnológicos constituem critérios importantes a serem avaliados, uma
vez que as bactérias probióticas presentes no alimento carreador devem resistir às condições
de processamento e estocagem, não promovendo alterações na textura e nas características
sensoriais do produto, além de se manterem ativas e viáveis para exercer seus benefícios no
hospedeiro (HUIS IN’TVELD; SHORTT, 1996; FERREIRA, 2003; VASILJEVIC; SHAH,
2008).
Aspectos de segurança, como origem da estirpe, ausência de histórico de associação
com doenças, ausência de genes plasmidiais de resistência a antibióticos, não promover a
degradação de muco intestinal, não translocar ou induzir translocação de micro-organismos
patogênicos para sítios extraintestinais e ausência de fatores de patogenicidade, como a
produção de hemolisina e gelatinase, devem ser avaliados (SAARELA, 2000; FAO/WHO,
2002).

27
3. OBJETIVOS
3.1 Objetivo Geral
Investigar as propriedades relacionadas à adesão e a ação dos Lactobacillus na
inibição da interação da Escherichia coli diarreiogênicas em células intestinais in vitro.
3.2 Objetivos Específicos
Analisar as propriedades de adesão de diferentes espécies de Lactobacillus,
incluindo às células HT-29 e HeLa, a capacidade de produção de biofilme e características de
hidrofobicidade;
Verificar a capacidade de diferentes Lactobacillus inibirem a adesão de E. coli
diarreiogênicas em células intestinais HT-29 e HeLa in vitro;
Investigar se os Lactobacillus têm a capacidade de coagregar com E. coli
diarreiogênicas e se há associação dessa propriedade com a inibição da adesão às células HT-
29 e HeLa;
Determinar se os sobrenadantes das culturas das linhagens de Lactobacillus
estudadas possuem capacidade de inibir o crescimento das E. coli diarreiogênicas.

28
4 MATERIAL E MÉTODOS
4.1 Amostras bacterianas
Neste estudo, foram utilizadas as seguintes espécies de referência pertencentes ao
gênero Lactobacillus: L. brevis (ATCC 367), L. delbrueckii subsp. delbrueckii (ATCC 9649),
L. fermentum (ATCC 23271), L. paracasei subsp. paracasei (ATCC 335), L. plantarum
(ATCC 8014) e L. rhamnosus (ATCC 9595) que foram cultivadas em ágar e caldo De Man,
Rogosa e Sharpe (MRS) (Difco). Adicionalmente, foram analisadas as bactérias envolvidas na
produção comercial de iogurtes das seguintes marcas: Actimel Defensis – Danone (L. casei
defensis), Chamyto – Nestlé (L. vivos paracasei), Danoninho – Danone (L. casei), Ninho
Soleil – Nestlé (L. paracasei) e Pense Bio Fibras – Batavo (L. acidophillus). Foram analisadas
linhagens de Lactobacillus isolados de fezes de cães recém-nascidos, doadas pelo Prof. Dr.
Álvaro Cantini Nunes do Departamento de Biologia Geral da Universidade Federal de Minas
Gerais: L. casei 4F-4, L. johnsonii 1F-36, L. paraplantarum A2/3, L. reuteri 6F-9 e o L.
salivarius 2F-38. As bactérias enteropatogênicas que foram utilizadas no estudo incluem
amostras de referência das categorias de E. coli diarreiogênicas, gentilmente cedidas pelo
Prof. Dr. Jorge A. Girón, da University of Florida, e incluíram: E. coli enteropatogênica
(EPEC, amostra E - 2348/69); E. coli enterotoxigênica (ETEC, amostra LT/ST + 1661-1); E.
coli enteroinvasora (EIEC, amostra 1381/7 LAC -); E. coli enteroagregativa (EAEC, amostra
042); E. coli enteroagregativa (EAEC, amostra 17-2); E. coli produtora da toxina de Shiga
(STEC Stx1, amostra 2781-8); E. coli produtora da toxina de Shiga (STEC Stx2). Os
lactobacilos foram mantidos em anaerobiose através do uso de geradores Anaerobac (Probac)
a 37ºC por 24 h. As bactérias enteropatogênicas foram cultivadas em caldo LB – meio Luria-
Bertani (Difco) e em caldo BHI - Infusão de cérebro e coração (Difco) a 37ºC por 24h. As
linhagens bacterianas foram estocadas em tubos criogênicos em solução do caldo BHI (Difco)
com glicerol (20%) (Amresco) a -86ºC.

29
4.1.1 Identificação dos tipos de Escherichia coli por reação em cadeia de polimerase em
multiplex
Para detecção de genes de virulência em E. coli, o DNA genômico dos isolados foi
extraído pelo método da fervura. Para tal, um alíquota de 2 mL de cada cultura de 24h de
E.coli crescida em meio BHI, foi centrifugado 10.000 rpm por 5 minutos, o sobrenadante foi
descartado e o sedimento bacteriano foi lavado em 700 ul de solução salina tamponada (PBS)
pH 7.5. Novamente foi centrifugado 10.000 rpm por 5 minutos e o PBS descartado. O
sedimento bacteriano foi ressuspenso em 400ul de Tris-EDTA (TE) 1 x, pH.75. Os tubos
contendo as amostras foram colocados em fervura a 100ºC durante 15 minutos e centrifugado
por 1 minuto a 10.000 rpm e o sobrenadante foi recolhido e guardado a -20ºC até o uso. A
verificação da qualidade/quantificação relativa do DNA extraído foi feita em gel de agarose
0,8%, contendo um marcador de massa molecular (New England Biolabs). Os ensaios de
mPCR foram feitos com iniciadores específicos, para a identificação dos grupos de E. coli
(Sk1, Sk2, VT com U, VT com D, LTL, LTR, AggRKS1, AggRSK2, Ipa III, Ipa IV). Os
ensaios de mPCR foram feitos em termociclador Amplitherm (Eppendorf) num volume final
de 25 uL contendo 15 picomoles de cada iniciador, 12.5 µl do Master Mix (PROMEGA ®,
Madison, USA), mais 100 ng do DNA bacteriano mais água livre de nuclease para completar
o volume final de 25 µl. O programa de amplificação consistiu de um passo inicial de
desnaturação de 94ºC por 5 minutos, seguidos de 30 ciclos de 94ºC por 1minuto, 52ºC por 1
minuto e 72ºC por 1 minuto e uma extensão final a 72ºC por 10 minutos.
O tamanho dos fragmentos amplificados foi aferido por eletroforese em gel de agarose
2%, que foi submetido à eletroforese em tampão TBE 0,5X (Tris-borato-EDTA) [100mM
Tris-base; 2,0 mM de solução 0,5 EDTA (pH 8,0) e 50 mM ácido bórico] a 85 mV durante 2
horas (SAMBROOK; FRITSCH; MANIATIS, 1989). Após a eletroforese, os géis foram
corados em uma solução de brometo de etídio (1:1000), durante 20 minutos e observados em
transiluminador ultravioleta. Para a determinação do tamanho dos produtos da mPCR foi
incluído em cada corrida um marcador de tamanho molecular 100pb DNA ladder
(PROMEGA, EUA).

30
4.2 Isolamento das bactérias de iogurte
Uma alíquota de cada iogurte foi semeada, com auxílio de alça de platina, em agar
MRS (Difco) para obtenção das bactérias presentes nos iogurtes constantes no item 4.1. A
incubação foi realizada a 37ºC por 24h ou 48h, em condições de microaerofilia em 5% de
CO2. Após incubação, as colônias esbranquiçadas ou cremosas, de tamanho médio a grande,
foram submetidas à coloração de Gram-positivos. As colônias formadas por bacilos Gram-
positivos foram congeladas a -86ºC para uso posterior.
4.3 Isolamento de bactérias láticas a partir das fezes de cães lactentes
Amostras de fezes foram coletadas de cães lactentes, dois da raça Crista Chinês e
quatro da raça York Shire, com idade entre 15 e 20 dias, certificando-se que eles e as mães
não tenham sido tratados com antimicrobianos. As fezes dos cães foram usadas como fonte de
lactobacilos. Cada amostra foi suspensa e homogeneizada com bastão de vidro em função do
peso, numa diluição 10-2
em salina. A partir desta diluição foram feitas novas diluições
decimais em salina. Para o isolamento, inoculou-se 1,0 mL das diluições (10-5
, 10-6
, 10-7
, 10-
8), pelo método pour plate, em duas repetições de placas de Petri, estéreis, com o meio ágar
De Man Rogosa e Sharpe (MRS, Difco, Sparks, EUA), fundido e vertido sobre os inóculos,
em temperatura aproximada de 45ºC. O conteúdo das placas foi bem homogeneizado e após
solidificarem, foram invertidas e incubadas à 37ºC, em câmara de anaerobiose (Forma
Scientific Company, Marrietta, USA), contendo uma atmosfera de 85%N2, 10%H2 e 5%CO2,
por 48h.
As placas que apresentaram visualmente, um número de unidades formadoras de
colônias (UFC), próximo a 100 foram escolhidas para contagem e observação macroscópica
dos morfotipos coloniais. Colônias com aspectos diferentes entre si foram isoladas em placas
de Petri contendo ágar MRS, pelo método do estriamento. Essas placas foram invertidas e
incubadas em câmara de anaerobiose à 37ºC por 24h.
Para identificação do gênero, os isolados foram submetidos a uma observação
microscópica após coloração de Gram e teste de catalase. As bactérias Gram-positivas com
morfologia meio ou muito alongada e semelhante a um bacilo foram selecionadas e repicadas

31
em tubos tipo Falcon contendo 3,0 mL do meio MRS líquido e novamente incubadas por 24h
nas condições já descritas. Do crescimento em meio líquido, foi retirada assepticamente, uma
pequena alíquota para o teste de produção de catalase, realizado com a adição de uma gota de
peróxido de hidrogênio a 30% para observar a ocorrência ou não da formação de pequenas
bolhas. Lactobacilos não produzem catalase, assim duas alíquotas de 1,0 mL de cada amostra
com resultado negativo foram transferidas para tubos Eppendorf, aos quais foram adicionados
30% de glicerol a 20% v/v, previamente esterilizado. As amostras foram armazenadas em
freezer a -80ºC até o uso.
4.4 Cultura e preparo de células HT-29 e células HeLa
Alíquotas das células HT-29 (carcinoma de intestino) e HeLa (carcinoma do colo do
útero) que eram mantidas em freezer em solução de dimetilsulfóxido (DMSO) (5%) e soro
fetal bovino (95%) a -86ºC (KAEWNOPPARAT et al., 2013), foram descongeladas e
transferidas para garrafa de cultura (TPP®) de células, contendo o meio Dulbecco´s Modified
Eagle Medium (DMEM) com GlutaMAXTM
(Gibco®), suplementado com 1% da solução de
antibiótico/antimicótico (10.000 unidades/mL de penicilina, 10.000 μg/mL de estreptomicina
e 25 μg/mL de anfotericina B) (Gibco®) e com 10% de soro fetal bovino (Gibco®). Os
frascos de cultura foram incubados em atmosfera úmida de 5% de CO2 a 37°C por 48 h
(SPURBECK; ARVIDSON, 2010), até a formação do tapete celular. O repique das
monocamadas celulares era realizado a cada 72h, ou após a confluência total da monocamada.
4.5 Ensaio de adesão em células HT-29 e células HeLa
4.5.1 Preparo das microplacas para os ensaios de adesão
A fim de se obter a monocamada celular para os ensaios de adesão bacteriana foram
preparadas microplacas de 24 cavidades (TPP®). Inicialmente, em cada poço foi colocado
uma lamínula de vidro redonda, com diâmetro de 13 mm, previamente desengorduradas com

32
álcool a 70% e devidamente esterilizadas. Após distribuição das lamínulas, deixou-se a luz
UV incidir sobre as mesmas por 15 minutos.
Em seguida, foi realizada a tripsinização, com 1 mL de tripsina, da monocamada de
uma das linhagens celulares previamente mantidas em garrafa de cultivo. Após a detaxação
das células elas foram ressuspensas em um volume de 25 mL de meio DMEM contendo 10%
SFB e 1% de antibiótico. Foram então distribuídos individualmente, 1 mL por poço da
suspensão celular, utilizando um tipo celular para cada microplaca. As microplacas foram
incubadas a 37º C em atmosfera de 5% de CO2 (CRAVIOTO et al., 1979; CANDELA et al.,
2008) a 37°C por 48 h ou até que a monocamada atingisse a semiconfluência..
Para a triagem das espécies de Lactobacillus que interagiam com células HeLa e HT-
29 em cultura de tecido, um dia após o repique celular, as linhagens de Lactobacillus spp.,
foram incubadas em 3 mL de caldo MRS (em triplicata) a 37ºC por 24 h em condições de
microaerofilia em 5% de CO2 e as amostras de E. coli foram incubadas em 3 mL do meio LB
(em triplicata) a 37ºC por 24 h.
4.5.2 Ensaio de adesão bacteriana
Após o período de incubação das culturas bacterianas crescidas em 6 mL de caldo,
foram transferidas para tubos de microcentrífuga de 2 mL. As espécies de Lactobacillus foram
centrifugadas a 9.000 rpm por 10 minutos e os sedimentos bacterianos foram lavados três
vezes com 1 mL de tampão salina fosfato, posteriormente, foi descartado o sobrenadante. Em
seguida, o sedimento foi ressuspenso em 300 µL de DMEM.
Paralelamente, cada poço das placas, previamente preparada 48 h antes, onde as
monocamadas foram semiconfluentes, com variação entre 80 – 90% de confluência, foi
lavado por três vezes, com solução salina tamponada. Em seguida adicionou-se a cada poço,
950 µL de DMEM suplementado com 5% de SFB; 1% de glutamina; 1% de manose (sem
antibiótico) e inoculado 50 µL do Lactobacillus (ensaio de exclusão). A placa foi incubada na
estufa de CO2 por 2 h. Decorrido o período, a placa foi lavada três vezes com PBS e realizou-
se nova troca do meio de cultura e as monocamadas foram infectadas com 40µL de E. coli
(ensaio de exclusão: 960 µL do meio para inoculação de 40µL do patógeno). As placas foram
re-incubadas na estufa de CO2 por 3 horas.

33
Em seguida, as monocamadas foram lavadas três vezes com PBS (pH 7,4) para
remoção das bactérias não aderidas e foi acrescentado 1 mL de metanol (LAFAN) em cada
poço para fixação (10-20 minutos). A placa foi lavada com PBS 1x e realizou-se a coloração
pelo método de Gram. Após secagem, foi realizada a montagem das lâminas e feita à leitura
do ensaio através da microscopia óptica (Leica ICC50 HD) (aumento de 1000x)
(KAEWNOPPARAT et al., 2013).
4.5.3 Quantificação da adesão
Para a quantificação da adesão e da inibição da adesão bacteriana às células intestinais,
foram utilizados poços sem lamínulas, tratados com 1 mL de Triton X-100 (Sigma) a 0,1%
por 5 minutos. Após a lise, os micro-organismos foram quantificados por plaqueamento de
alíquotas de 0,1 mL de diluições seriadas decimais em Ágar MRS e MacConkey, seguido de
incubação a 37oC por 24h. As quantificações foram realizadas em triplicata. Os resultados do
número de bactérias (UFC/mL) foram expressos em porcentagem de redução em relação ao
ensaio realizado com o enteropatógeno, mas na ausência de células de Lactobacillus. O
controle consistiu no ensaio com o enteropatógeno na ausência das espécies de Lactobacillus
(CORTÉS et al., 2007). Após inoculação dos micro-organismos na microplaca de 24 poços,
foi realizada diluição da suspensão bacteriana em PBS para semeadura em placa para
contagem do número de colônias que foram inoculadas. Para o ensaio qualitativo, os poços
foram revestidos com lamínulas que, após o período de adesão, foram submetidas à fixação
em chama, colagem em lâminas, coloração de Gram e visualização em microscópio óptico
(Leica ICC50 HD) (aumento de 1000x) (CANDELA et al., 2008).
4.5.3.1 Quantificação do inóculo das amostras bacterianas
Para o controle do número de micro-organismos que estavam sendo inoculados, foram
realizadas diluições seriadas decimais e semeadura em ágar. Após inoculação na microplaca
de 24 poços (TPP®), foi aspirada e transferida uma alíquota de 50 μL da suspensão bacteriana
para tubo de microcentrífuga com 1 mL de PBS (diluição 1:20). Em seguida, foi realizada
homogeneização em agitador de tubos e aspiração de uma nova alíquota de 5 μL, que foi

34
transferida para tubo contendo 495 μL de PBS (diluição 1:2.000). Após a homogeneização,
foi retirada uma alíquota de 50 μL para tubo contendo 450 μL de PBS (diluição 1:20.000). A
suspensão foi semeada sobre o meio de cultura MacConkey por esgotamento com alça
bacteriológica de 1 μL (F = 2 x 107) e incubada em aerobiose a 37ºC por 24 horas e, após o
período, realizou-se contagem das colônias de E. coli. Para a quantificação das espécies de
Lactobacillus, além das etapas descritas acima, foi realizada mais uma diluição através de
nova alíquota de 50 μL que foi retirada do tubo contendo 450 μL de PBS 1x (diluição
1:20.000) e transferida para tubo com 450 μL de PBS (diluição 1:200.000). Utilizaram-se
diluições seriadas associadas ao uso de alça bacteriológica calibrada de 1 μL (F = 2x108). As
placas foram incubadas a 37ºC por 24h em condições de microaerofilia em 5% de CO2. Após
a incubação, realizou-se a contagem das unidades formadoras de colônias por mL (UFC/mL)
(CANDELA et al., 2008).
4.5.3.2 Quantificação bacteriana após ensaio de adesão
Para a contagem do número de bactérias aderidas às monocamadas celulares, realizou-
se uma nova diluição para semeadura em placa. A placa foi lavada 3x com PBS, adicionando-
se 1 mL de Triton X-100/ a 1% em PBS por 5 minutos, e, posteriormente, o fundo do poço foi
raspado com ponteira de 1 mL. Uma alíquota de 5 μL da suspensão bacteriana foi transferida
para tubo com 495 μL de PBS (diluição 1:100) e, após homogeneização, 50 μL da suspensão
foi transferidos para tubo com 450 μL de PBS (diluição 1:1.000). Utilizou-se alça
bacteriológica de 1 μL (F = 106) para semeadura por esgotamento da suspensão em placas
contendo meio de cultura MacConkey. As placas foram incubadas em aerobiose a 37ºC por
24h e realizada a contagem das colônias de E. coli. Para a quantificação dos Lactobacillus,
além das etapas já descritas, foi realizada mais uma diluição através de nova alíquota de 50
μL a qual foi retirada do tubo contendo 450 μL de PBS (diluição 1:1.000) e transferida para
tubo com 450 μL de PBS (diluição 1:10.000). Utilizou-se alça bacteriológica de 1 μL (F =
107) para semeadura por esgotamento da suspensão em placas contendo o meio de cultura
MRS. As placas foram incubadas a 37ºC por 24h em condições de microaerofilia em 5% de
CO2. Após a incubação, realizou-se a contagem das unidades formadoras de colônias por mL
(UFC/mL) (CANDELA et al., 2008).

35
4.5.3.3 Quantificação bacteriana após ensaio de competição
Para o ensaio sem lamínulas de E. coli, realizaram-se diluições para semeadura em
placa e contagem do número de colônias que aderiram após a competição. A microplaca foi
lavada, sendo, em seguida, adicionado 1 mL do detergente Triton X-100/PBS 0,1% no fundo
das cavidades por 5 minutos. Posteriormente, foi realizada a raspagem do fundo do poço com
ponteira de 1 mL, e transferida uma alíquota de 50 μL da suspensão bacteriana para tubo com
450 μL de PBS (diluição 1:10) e outra alíquota de 5 μL para tubo com 495 μL de PBS
(diluição 1:100). Após a homogeneização da primeira diluição em 450 μL de PBS, utilizou-se
alça bacteriológica de 10 μL (x1000 (F = 103)) para semeadura em placas de petri e outra alça
de 1 μL (x10.000 (F = 104)) para inoculação em agar MacConkey. Da segunda diluição em
495 μl de PBS, utilizou-se alça bacteriológica de 1 μL (x100.000 (F = 105)) para inoculação
em agar MacConkey. As placas foram incubadas em aerobiose a 37º por 24h e realizada a
contagem de colônias de E. coli.
Para o ensaio sem lamínulas das linhagens de Lactobacillus, realizou-se uma diluição
para semeadura em placa e contagem do número de bactérias que aderiram após a
competição. A microplaca foi lavada, sendo, em seguida, adicionado 1 mL do detergente
Triton X-100/PBS 0,1% por 5 minutos. Posteriormente, foi raspado o fundo do poço com
ponteira de 1 mL, e transferida uma alíquota de 50 μL da suspensão bacteriana para tubo com
450 μL de PBS (diluição 1:10) e outra alíquota de 5 μL para tubo com 495 μL de PBS
(diluição 1:100). Após homogeneização da primeira diluição em 450 μL de PBS, utilizou-se
alça bacteriológica de 1 μL (x1000 (F = 104)) para semeadura em placas de petri contendo o
meio MRS. Da segunda diluição em 495 μL de PBS, utilizou-se uma alça de 1 μL (x100.000
(F = 105)) para inoculação da suspensão em Agar MRS. As placas foram incubadas a 37ºC
por 24h em condições de microaerofilia em 5% de CO2. Após a incubação, realizou-se a
contagem das colônias de Lactobacillus (CANDELA et al., 2008).
4.6 Hidrofobicidade da superfície celular
Os ensaios de hidrofobicidade da superfície celular foram realizados segundo as
metodologias descritas por Rosenberg (1983), Pelletier (1997) e Pérez (1998) com

36
modificações de Barbosa et al., (2005). Os isolados de Lactobacillus foram inoculados para
um tubo de ensaio com 3 mL de caldo MRS e incubadas a 37ºC por 20h. Após esse período,
foi realizado o segundo repique do cultivo dos Lactobacillus para 8 tubos de ensaio contendo
4 mL de caldo MRS para cada espécie e foram reincubadas a 37º C por 20h. Após a
incubação, o cultivo foi centrifugado a 3.500 rpm durante 15 minutos. Posteriormente, as
células foram lavadas duas vezes com solução tampão fosfato (PBS, 50 mM, pH 7,0), e
ressuspensas em 4 mL de nitrato de potássio (KNO3, 0,1 M, pH 6,2) e homogeneizadas em
agitador de tubos. A suspensão bacteriana foi repassada para tubos de ensaio estéreis no
volume de 5 mL para amostra A e 4 mL para amostra B. Sendo que, na amostra B, utilizaram-
se 3 tubos de ensaio por cada bactéria devido ao ensaio ser feito com três solventes: Xilol
(solvente apolar), Clorofórmio (solvente ácido) e Acetato de Etila (solvente básico).
Para a amostra B, foi adicionado 1mL de cada solvente nos tubos de ensaio,
totalizando uma solução de 5 mL. As amostras ficaram em repouso por 5 minutos e foram
homogeneizadas por agitação durante 2 minutos. Os tubos foram mantidos em repouso por 40
minutos aproximadamente, para que ocorresse a separação das fases. Após esse período, foi
realizada a leitura da absorbância da fase aquosa a 600 nm em espectrofotômetro. Todos os
testes foram realizados em duplicata.
AMOSTRA A: DOA ⇒ Tubo A = 5 mL de suspensão celular em solução de KNO3
Branco = 5 mL de solução de KNO3
AMOSTRA B: DOB ⇒ Tubo B = 4 mL de suspensão celular em solução de KNO3 + 1 mL
dos solventes em cada tubo.
Branco = 4 mL de solução de KNO3 + 1 mL dos solventes em cada
tubo.
A porcentagem de hidrofobicidade, de acordo com o solvente utilizado, foi calculada
com a seguinte fórmula:
% Hidrofobicidade = (DOA – DOB) x 100
--------------------------------------------------------
DOA

37
4.7 Ensaio de produção de biofilme dos Lactobacillus
A quantificação da produção de biofilme em superfície de poliestireno foi realizada em
microplacas com 96 cavidades (TPP®) com fundo em “U”. Suspensões bacterianas de
Lactobacillus cultivadas em caldo MRS a 37°C por 24h, padronizadas por espectrofotometria
para a densidade óptica igual (Do) 0.7, foram diluídas 1:40 em meio MRS. Posteriormente,
alíquotas de 200 μL foram adicionadas nas cavidades. Caldo MRS sem crescimento bacteriano
foi utilizado como controle. As placas foram incubadas a 37°C por 24h. Após incubação, as
placas foram lavadas com 1mL de PBS (pH 7,4) por três vezes e deixadas secar em
temperatura ambiente. Adicionou-se aos poços 200 μL de metanol para fixação do biofilme
bacteriano. Transcorridos 15 minutos de incubação, o metanol foi descartado e a microplaca
foi submetida à secagem em temperatura ambiente. Foram adicionados 200 μL de cristal
violeta a 2% em cada poço e o biofilme foi corado por 15 minutos. Posteriormente, a placa foi
lavada três vezes com água destilada e seca em temperatura ambiente. O corante impregnado
no biofilme foi extraído com 160 μL de ácido acético glacial a 33%, sendo, em seguida,
submetida à espectrofotometria com filtro de 540 nm para aferir as respectivas absorbâncias
(Ab) de cada cavidade em leitor de ELISA (Biotek). Baseando-se nas densidades ópticas
produzidas pelos isolados (D.O.i) e tomando-se por base o controle negativo (D.O.c), os
isolados foram classificados nas seguintes categorias (STEPANOVIC et al., 2004):
Não produtor: D.O.i < D.O.c
Produtor fraco: D.O.c < D.O.i ≤ (2 X D.O.c)
Produtor moderado: (2x D.O.c) < D.O.i ≤ (4 X D.O.c)
Produtor forte: (4 X D.O.c) < D.O.i
4.8 Ensaio de coagregação
O ensaio de coagregação foi realizado para verificar se as linhagens de Lactobacillus
spp. coagregavam com os patógenos intestinais. Inicialmente, tanto os Lactobacillus quanto
as E. coli foram ajustados em PBS (pH 7,4) na concentração de 1,2 x 109 UFC/mL (escala de
Mac Farland Nº 4). Em seguida, 500 µl das suspensões bacterianas foram homogeneizados
em agitador de tubos por 10 segundos e, posteriormente, inoculados em placas de 24 poços,

38
que foram incubadas a 37ºC por 4h sob agitação em orbital (banho-maria). Em seguida, as
suspensões foram observadas micro e macroscopicamente com auxílio de um microscópio de
luz invertida (XDS – 1B) para a avaliação do grau de coagregação, que foi classificado em
scores que variavam de 0 a 4. Sendo: 0 = não visualização microscópica de coagregados de
bactérias; 1 = pequenos agregados de bactérias visíveis microscopicamente; 2 = agregados
visíveis microscopicamente de um maior número de bactérias depositados no centro do poço;
3 = formação de grumos visíveis macroscopicamente de um número considerável de bactérias
depositadas no centro do poço; 4 = escore máximo para descrever os maiores grumos de
agregação microbiana, visíveis macroscopicamente. Além disso, as amostras foram
visualizadas pela coloração de Gram (REID et al., 1990).
4.9 Análise da atividade do sobrenadante
A análise da atividade do sobrenadante dos Lactobacillus foi realizada para verificação
da produção de substâncias antimicrobianas. As linhagens de Lactobacillus foram incubadas
em 3 ml de caldo MRS a 37ºC por 24h em condições de microaerofilia em 5% de CO2 e as E.
coli foram incubadas em 3 ml do meio BHI a 37ºC por 24h. Para o ensaio, o sobrenadante dos
Lactobacillus foi obtido a partir da centrifugação a 9.000 rpm por 10 minutos e foram,
posteriormente, neutralizados com hidróxido de sódio (NaOH) a 1N e filtrado em microfiltro
de seringa (0,22 μm, Millipore®). Suspensões das E. coli foram preparadas em PBS
ajustando a turbidez segundo a escala 0,5 de Mac Farland, que corresponde à 1,5 x 108
UFC/mL. Em seguida, as suspensões de E.coli foram semeadas sobre a superfície do ágar MH
contendo escavações de 5,0 mm de diâmetro. Após a centrifugação da cultura de
Lactobacillus, alíquotas de 20 µL dos sobrenadantes de cada espécie de Lactobacillus foram
inoculadas nos poços. Os poços foram demarcados a uma distância de 25 mm entre si e de 20
mm em relação à borda da placa, a fim de se evitar a sobreposição de halos. Posteriormente,
as placas foram incubadas a 37ºC por 24h e observou-se a formação de halos de inibição, de
acordo com a metodologia proposta por Mastromarino et al., (2002) com modificações.

39
4.10 Análises estatísticas
As análises estatísticas foram realizadas através dos testes de Correlação Linear de
Pearson, Kruskal Wallis e ANOVA (01 critério; com medidas repetidas post hoc com teste de
Bonferroni) na análise de dados de quantificação expressos em percentual das contagens de
UFC/mL. A quantificação expressa em porcentagem foi analisada pelo teste do 2
(SCHLESSELMANN, 1982). Em todos os testes de hipóteses, adotou-se o nível de
significância de 0,05 ou 5% (p < 0,05). No processamento eletrônico dos dados, foi utilizado
o software GraphPad Prism (versão 5).

40
5 RESULTADOS
5.1 Confirmação dos enteropatógenos e das linhagens dos Lactobacillus
Neste estudo, foram utilizadas amostras de bactérias enteropatogênicas de referência
das categorias de E. coli diarreiogênicas: E. coli enteropatogênica (EPEC, amostra E -
2348/69); E. coli enterotoxigênica (ETEC, amostra LT/ST + 1661-1); E. coli enteroinvasora
(EIEC, amostra 1381/7 LAC -); E. coli enteroagregativa (EAEC, amostra 042); E. coli
enteroagregativa (EAEC, amostra 17.2); E. coli produtora de toxina de Shiga (STEC Stx1,
amostra 2781-8); E. coli produtora de toxina de Shiga (STEC Stx2). Os isolados foram
confirmados através de PCR multiplex: aggR (EAEC 042), eae (EPEC E2348/69), elt (ETEC
1661-1, LT/ST), aggR (EAEC 17-2), stx (STEC 2781-1, Stx1), stx (STEC Stx2), ipaH(
EIEC 1381-7) para detectar os genes de virulência específicos das seguintes categorias
(Figura 1).
As amostras bacterianas probióticas de referência: L. brevis (ATCC 367), L.
delbrueckii subsp. delbrueckii (ATCC 9649), L. fermentum (ATCC 23271), L. paracasei
subsp. paracasei (ATCC 335) e L. plantarum (ATCC 8014) e L. rhamnosus (ATCC 9595),
foram isoladas em meio seletivo para Lactobacillus e as colônias confirmadas por meio da
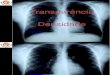
coloração de Gram para se comprovar a pureza das culturas (Figura 2).
Figura 1. Padrão de amplificação por PCR multiplex que identificam e diferenciam os tipos
de Escherichia coli.
41
Legenda: Eletroforese em gel de agarose a 2,5%. 1) EAEC 042; 2) EPEC E2348/69; 3) ETEC
1661-1, LT/ST +; 4) EAEC 17-2; 5) STEC 2781-1, Stx1; 6) STEC Stx2; 7) EIEC 1381-7.
Figura 2. Aspectos microscópicos das espécies de Lactobacillus após coloração de Gram.
A) L. plantarum); B) L. brevis; C) L. rhamnosus; D) L. delbueckii delbrueckii; E) L.
fermentum; F) L. paracasei paracasei.
A B
C D
E
F
i
g
u
r
a
1
F

42
5.2 Ensaio de adesão em células HeLa e HT-29
O ensaio de adesão evidenciou que todas as seis espécies de referência são aderentes
às células HeLa e HT-29, sendo que quatro espécies (L. brevis, L. delbrueckii delbrueckii, L.
plantarum e L. fermentum) apresentam melhor capacidade de adesão. Nos isolados de cães,
três amostras aderiram (L. casei, L. johnsonii e L. paraplantarum). O único isolado que não
apresentou adesão foi o do iogurte Chamyto (L. paracasei), enquanto que o isolado do iogurte
Actimel (L. casei) apresentou melhor capacidade de adesão em células eucarióticas em
relação aos demais iogurtes (Tabela 1). Não houve diferença no perfil de adesão entre as duas
linhagens celulares avaliadas.
Tabela 1. Análise microscópica da adesão das espécies de Lactobacillus de referência,
isolados de cães e de iogurtes em células HT-29.
Legenda: * - negativo (ausência de bactérias); + (1 - 10 bactérias), ++ (11 - 20 bactérias), +++ (20 – 30 bactérias); ++++ (> 30 bactérias).
Origem dos Lactobacillus Adesão*
ATCC L. brevis (ATCC 367) ++++
L. d. delbrueckii (ATCC 9645) ++++
L. fermentum (ATCC 23271) +++
L. paracasei (ATCC 335) ++
L. plantarum (ATCC 8014) ++++
L. rhamnosus (ATCC 9595) +
Cães L. casei ++
L. johnsonii +
L. paraplantarum ++
L. reuteri -
L. salivarius -
Iogurte Actimel/Danone (L. casei) ++
Chamyto/Nestlé (L. vivos paracasei) -
Danoninho/Danone (L.casei) +
Ninho/Soleil/Netlé (L. paracasei) +
Pense Biofibras/Batavo (L. acidophilus e Bifidobacterium lactis) +

43
Na tabela 2 constam os resultados dos testes de quantificação da adesão em células
HT-29 e em células HeLa. Os isolados com maior capacidade de adesão (Lactobacillus de
referência), fenômeno avaliado por microscopia, foram submetidos à quantificação de
bactérias viáveis aderidas às células HT-29 por plaqueamento de diluições decimais em agar
MRS. As espécies selecionadas (L. brevis, L. delbrueckii, L. fermentum e L. plantarum)
apresentaram contagens bacterianas entre 2 e 8,5 x 107UFC/mL, confirmando a análise
anterior.
Tabela 2. Quantificação da adesão das espécies de Lactobacillus de referência em células
HT-29.
Espécies de Lactobacillus Adesão (UFC/µl X107)*
L. brevis (ATCC 367) 8.5 ± 2.1
L. d. delbrueckii (ATCC 9645) 8.4 ± 0
L. fermentum (ATCC 23271) 2 ± 0
L. plantarum (ATCC 8014) 3 ± 1.4
* Os ensaios foram realizados em triplicata
5.3 Hidrofobicidade da superfície celular de Lactobacillus spp.
Na avaliação da hidrofobicidade da superfície celular das linhagens de Lactobacillus,
sob presença dos solventes clorofórmio, xilol e acetato de etila, foi verificado que, dos seis
isolados de Lactobacillus de referência, apenas 2 (33%) espécies (L. delbrueckii delbrueckii e
L. paracasei) foram hidrofílicas, sendo as outras 4 (67%) hidrofóbicas (L. brevis; L.
fermentum; L. plantarum e L. rhamnosus). Nos isolados de cães, apenas 1 (20%) foi
classificado como hidrofílico (L. johnsonii). Dos cinco isolados de iogurtes avaliados, 1
(20%) foi considerado hidrofílico (L. acidophilus e Bifidobacterium lactis) e 4 (80%)
hidrofóbicos, conforme demonstrado na Tabela 3.

44
Tabela 3. Distribuição comparativa da hidrofobicidade de superfície celular das linhagens de
Lactobacillus de referência (ATCC’s), isolados de cães e de iogurtes.
Origem dos Lactobacillus Hidrofobicidade*
ATCC
L. brevis (ATCC 367) +
L. d. delbrueckii (ATCC 9645) -
L. fermentum (ATCC 23271) +
L. paracasei (ATCC 8014) -
L. plantarum (ATCC 8014) +
L. rhamnosus (ATCC 9595) +
Cães
L. casei +
L. johnsonii -
L. paraplantarum +
L. reuteri +
L. salivarius +
Iogurte
Actimel/Danone (L. casei) +
Chamyto/Nestlé (L. vivos paracasei) +
Danoninho/Danone (L.casei) +
Ninho/Soleil/ Nestlé(L. paracasei) +
Pense Biofibras/Batavo (L. acidophilus + B. lactis) -
Legenda: * Foram consideradas espécies com superfície hidrofóbicas aquelas que aderiram predominantemente
no clorofórmio. + = isolados hidrofóbicos; - = isolados hidrofílicos.

45
A análise do ensaio de hidrofobicidade de superfície celular na presença do solvente
clorofórmio das linhagens de Lactobacillus isolados de cães, demonstrou que L.
paraplantarum, L. reuteri e L. casei foram mais hidrofóbicos que o L. johnsonii (Figura 3).
Nos isolados de iogurtes, o Ninho Soleil e o Chamyto foram mais hidrofóbicos que o iogurte
Pense Bio Fibras. Entre as amostras de Lactobacillus de referência, observou-se que as
espécies de Lactobacillus fermentum, brevis e plantarum foram mais hidrofóbicas que as
espécies de L. delbrueckii delbrueckii e L. paracasei. A comparação entre todos os isolados
de Lactobacillus revelou que o L. reuteri, L. paraplantarum e L. casei de cães; Ninho soleil
da Nestlé; L. plantarum, L. fermentum e L. brevis (ATCC) não apresentaram diferença no
perfil de hidrofobicidade, sendo os mais hidrofóbicos em comparação aos demais isolados (p
< 0.001).
Figura 3. Análise comparativa do perfil de hidrofobicidade da superfície celular de
Lactobacillus isolados de cães ou de iogurtes e de espécies de referência, na presença de
clorofórmio.
Teste Estatístico: ANOVA 01 critério.

46
5.4 Produção de biofilme
Em relação ao teste quantitativo em microplaca com os 16 Lactobacillus, foi
constatado que 13 eram produtoras de biofilme, sendo que, destas: 3 (18,7%) foram
classificadas como produtoras moderadas; 10 (62,6%) como fracas produtoras e, por fim; 3
(18,7%) não produziram biofilme. Após análises das diversas amostras, verificou-se que não
houve significância estatística frente às diferentes espécies bacterianas na produção de
biofilme (Tabela 4).
Tabela 4. Distribuição comparativa da produção de biofilme pelas amostras de Lactobacillus
de referência (ATCC’s), isolados de cães e de iogurtes.
Classificação*
Origem dos Lactobacillus
**H Valor de p n (%)
Iogurte Cães ATCC
n= 5 n= 5 n= 6
Não produtor 2 (40) 1 (20) - 0.35 0.83
Fraco 2 (40) 3 (60) 5 (83.3)
Moderado*** 1 (20) 1 (20) 1 (16.7)
* (D.O.i): Densidade óptica isolados; (D.O.c): Densidade óptica controle negativo; Não produtor: D.O.i <
D.O.c; Produtor fraco: D.O.c < D.O.i ≤ (2 X D.O.c); Produtor moderado: (2x D.O.c) < D.O.i ≤ (4 X D.O.c);
Produtor forte: (4 X D.O.c) < D.O.i (STEPANOVIC et al., 2004).
** Teste Estatístico: Kruskal-Wallis.
*** Produção moderada de biofilme: (Iogurte: Danoninho (L. casei); Cães: L. casei; ATCC: L. fermentum).

47
5.5 Ensaio de Coagregação
Considerando-se que as linhagens de referências: L. brevis (ATCC 367), L. delbrueckii
subsp. delbrueckii (ATCC 9649), L. fermentum (ATCC 23271), L. paracasei subsp. paracasei
(ATCC 335) e L. plantarum (ATCC 8014) e L. rhamnosus (ATCC 9595) foram as que
apresentaram a melhor capacidade de adesão às células eucarióticas, elas foram selecionadas
para os testes de coagregação, cujos resultados foram descritos na Tabela 5. Foi demonstrado
que o Lactobacillus plantarum foi a espécie com maior capacidade de coagregação. Cinco
espécies de referência apresentaram atividade de coagregação com escores que variaram de 1
a 4, com pelo menos dois enteropatógenos. Duas espécies (L. fermentum e L. plantarum)
apresentaram atividade contra todas as espécies de E. coli diarreiogênicas analisadas. A
espécie de L. brevis não apresentou atividade apenas contra a ETEC, enquanto que o L.
paracasei foi a única espécie de Lactobacillus que não apresentou capacidade de coagregação
com nenhum dos enteropatógenos analisados (Tabela 5).

48
Tabela 5. Ensaio de coagregação entre espécies de Lactobacillus e E. coli diarreiogênicas.
Cepas
Escores de Coagregação*
EAEC EAEC EPEC EIEC STEC ETEC
(17.2) (042) (2348/69) (1381-7) (STX1 2781-8) (LT+ ST+ 1661-1)
L. brevis (ATCC 367) 1 1 2 2 1 -
L. delbrueckii (ATCC 9645) - - - 1 1 -
L. fermentum (ATCC 23271) 2 1 2 3 1 1
L. paracasei (ATCC 335) - - - - - -
L. plantarum (ATCC 8014) 2 3 2 4 2 1
L. rhamnosus (ATCC 9595) 1 - 2 1 2 - Legenda: * - = ausência de coagregação; 1 = pequenos agregados de bactérias visíveis microscopicamente; 2 = agregados visíveis microscopicamente de um maior número de
bactérias depositadas no centro do poço; 3 = formação de grumos visíveis macroscopicamente de um número considerável de bactérias depositadas no centro do poço; 4 =
escore máximo para descrever os maiores grumos de agregação microbiana visível macroscopicamente e depositada no centro do poço (REID et al., 1990).

49
5.6 Análise quantitativa de atividade inibitória de Lactobacillus spp., sobre a adesão
de E. coli
A análise preliminar, por microscopia de campo claro, do ensaio de inibição da
adesão, demonstrou que o L. delbrueckii não apresenta atividade inibitória sobre a interação
das cepas de E. coli às células em cultura de tecido. Portanto, ela foi excluída do ensaio de
quantificação da inibição. As espécies L. brevis, L. fermentum e L. plantarum foram as que
inibiram significantemente a adesão de EPEC, EAEC e EIEC (p < 0,05). A EAEC 17-2 foi a
única categoria que não foi inibida pelo L. brevis. Os percentuais de inibição variaram de
65.2% a 77.5% para EPEC, de 44.7% a 73.8% para EAEC 17-2, de 63.4% a 69.5% para
EAEC 042, de 4.7% a 48.9% para ETEC e de 97.8% a 99.1% para EIEC (Figura 4). As
Figuras 5 a 7 ilustram a atividade inibitória de algumas espécies de Lactobacillus sobre EIEC,
EAEC 17-2 e EAEC 042, enquanto a Figura 8 ilustra a ausência de inibição da adesão da
ETEC.
Na tentativa de se explicar a inibição da adesão, foi realizada uma análise de
correlação linear de Pearson. De modo geral, foi constatado que, quanto maior a capacidade
de coagregação dos Lactobacillus com as E. coli, maior a capacidade de inibição da adesão
das cepas de E. coli nas células humanas (Figura 9). A análise estatística revelou que somente
a propriedade de coagregação corresponde a 63% da capacidade dos Lactobacillus de inibir a
adesão da E. coli.
50
Figura 4. Avaliação da atividade inibitória de Lactobacillus de referência sobre a adesão das
diferentes categorias de E. coli diarreiogênicas em células eucarióticas.
* p < 0,05; ** p < 0,01; *** p < 0,0001. Teste ANOVA com medidas repetidas post hoc com teste de
Bonferroni.

51
Figura 5. Adesão em células HeLa dos isolados: A) L. plantarum; B) E. coli EIEC – 1381/7;
C) Inibição da EIEC pelo L. plantarum.
Figura 07. Microscopia óptica da adesão em células HT-29 de isolados de cepas de: A)
A
B
C

52
Figura 6. Adesão em células HT-29 dos isolados: A) L. plantarum; B) EAEC – 042; C)
Inibição da EAEC 042 pelo L. plantarum.
A
B
C
A
B
C

53
Figura 7. Adesão em células HT-29 dos isolados: A) L. brevis; B) Escherichia coli EAEC –
042; C) Inibição da EAEC 042 pelo L. brevis.
A
B
C
A
B
C
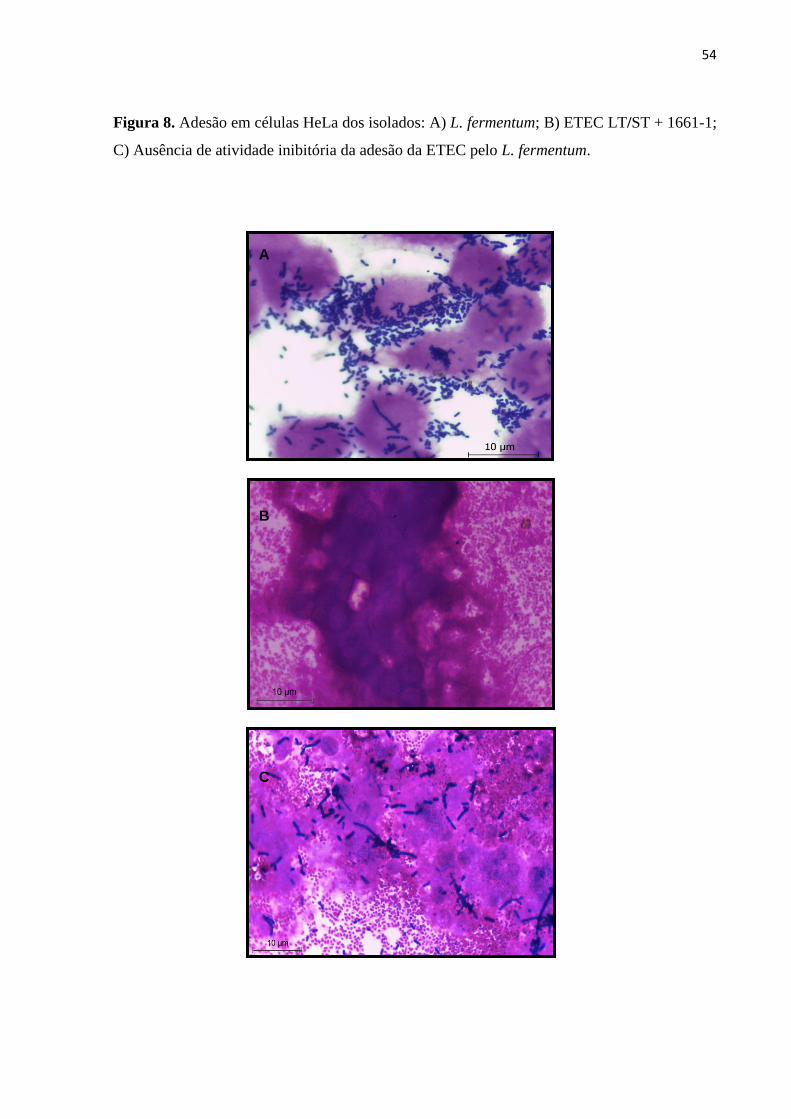
54
Figura 8. Adesão em células HeLa dos isolados: A) L. fermentum; B) ETEC LT/ST + 1661-1;
C) Ausência de atividade inibitória da adesão da ETEC pelo L. fermentum.
5.6 Análise da atividade do sobrenadante
B
C
A
B
C

55
Figura 9. Correlação entre escore de coagregação entre cepas de E. coli e Lactabacillus e a
capacidade de adesão das cepas de Escherichia coli em células HT-29 e HeLa após
competição com cepas de Lactobacillus. r = - 0.72, p=0.0004.
0 20 40 60 800
1
2
3
4
5
Capacidade de adesão
Sco
re d
e C
oag
reg
ação

56
5.7 Análise da atividade do sobrenadante
Um ensaio adicional, realizado com as linhagens de referência para explicar a inibição
da adesão, consistiu em se empregar o sobrenadante das culturas de Lactobacillus em um teste
de inibição do crescimento bacteriano. No entanto, nenhuma espécie de Lactobacillus
apresentou atividade antimicrobiana contra as espécies de E. coli diarreiogênicas testadas com
a metodologia citada.

57
6 DISCUSSÃO
Várias espécies de Lactobacillus têm sido avaliadas quanto ao potencial probiótico
(ZHANG et al., 2013). No entanto, ainda existem lacunas no conhecimento com relação à
aplicação de determinadas linhagens para essa finalidade. No presente estudo, verificou-se
que algumas espécies de coleção de culturas de referência da ATCC apresentam capacidade
de inibir a interação de categorias de E. coli diarreiogênicas às células eucarióticas in vitro.
Neste contexto, verificou-se que as espécies L. plantarum, L. brevis e L. fermentum inibiram a
adesão de EPEC, EAEC e EIEC sobre as células HT-29 e HeLa. A ETEC foi à única
categoria que não foi inibida por nenhuma espécie de referência de Lactobacillus testada.
Uma característica desejada para um micro-organismo probiótico é a capacidade de
aderência, permitindo a sua permanência e atuação em um determinado ecossistema. Essa
característica depende, em parte, das propriedades físico-químicas da parede celular
bacteriana, incluindo a hidrofobicidade, entre outros fatores. Estudos têm demonstrado
relações entre cargas superficiais, hidrofobicidade e a composição elementar da superfície
celular. Eles indicam que a presença de glicoproteínas na superfície celular resultará em alta
hidrofobicidade e que a baixa hidrofobicidade está associada à presença de polissacarídeos
(PELLETIER, 1997; TRABULSI; SAMPAIO, 2000). Segundo Kos et al. (2003), os micro-
organismos que têm a parede celular do tipo hidrofóbica possuem uma maior variedade de
glicoproteínas de superfície, e apresentam, dessa forma, uma maior probabilidade de aderir às
matrizes celulares.
O processo de adesão dos micro-organismos às superfícies celulares é considerado
multifatorial, sendo relacionado às diferentes interações específicas e não específicas. Os
mecanismos não específicos envolvem as interações eletrostáticas e hidrofóbicas de superfície
celular. Já os mecanismos específicos estão relacionados às diversas moléculas de superfície
celular, tais como as proteínas da camada S (FRECE et al., 2005; CHEN et al., 2007; MENG
et al., 2014) exopolissacarídeos (LEBEER et al., 2010), peptídeoglicanos (VAN TASSEL;
MILLER, 2011), glicosaminoglicanos (MARTIN et al., 2013) e as fímbrias (SINGH et al.,
2013).
No presente estudo, a capacidade de adesão foi notória, principalmente, para as
espécies de Lactobacillus de referência avaliadas, apesar dessas bactérias não apresentarem

58
capacidade de produção de biofilme significante. No entanto, não foi observada uma
associação entre hidrofobicidade e a capacidade de aderir às células HT-29 e HeLa, uma vez
que a maioria dos isolados de lactobacilos das fezes de cães e de iogurtes que não aderiram às
células eucarióticas apresentaram características de hidrofobicidade em suas superfícies
celulares. Isso sugere que a interação das linhagens padrão dos Lactobacillus às células
poderia estar relacionada à produção de adesinas específicas. Segundo Conway e Kjelleberg
(1989), existem estruturas de natureza proteica que podem mediar a adesão dessas bactérias.
De fato, há evidências do envolvimento de adesinas em Lactobacillus. Granato et al. (2004),
descobriram que a proteína de membrana SLAP, de 50 kDa, podia possibilitar a adesão às
células e à mucina. Essa proteína têm a capacidade de inibir a adesão da E. coli
enterohemorrágica em uma membrana basal sintética, provavelmente por competição pelos
mesmos receptores.
A capacidade dos Lactobacillus de inibir a adesão dos enteropatógenos é relevante,
principalmente porque a adesão ao epitélio do hospedeiro é a etapa inicial e essencial para o
estabelecimento de uma infecção bacteriana (FINLAY, FALKOW, 1997; XU et al., 2009).
Desta forma, seria possível prevenir um processo infeccioso bacteriano em uma mucosa,
como no caso das infecções intestinais, simplesmente pela ingestão ou outra forma de
administração de um probiótico. De fato, estudos têm demonstrado essa possibilidade em
diferentes sítios anatômicos. No caso do trato urogenital, foi demonstrado que Lactobacillus
representam uma importante barreira microbiológica a infecções por patógenos como:
Escherichia coli, Gardnerella vaginalis, Candida albicans, Pseudomonas aeruginosa,
Klebsiella pneumoniae, Staphylococcus aureus e Streptococcus agalactiae (OZTURK et al.,
2009). O estudo de Chan et al. (1985) demonstrou a capacidade de exclusão por competição
de uropatógenos de células uroepitelias humanas pela ação dos Lactobacillus. Já a capacidade
dos Lactobacillus de inibir o crescimento de uropatógenos e bloquear a adesão em células
epitelias vaginais foi evidenciada por Osset et al. (2001).
No caso de infecções intestinais, os estudos têm se limitado a empregar algumas
espécies de probióticos contra poucos enteropatógenos, mas a atividade inibitória tem sido
observada. Recentemente, Sánchez e Urdaci (2012) relataram a interferência da espécie L.
plantarum sobre a adesão à mucina da E. coli LMG2092 e da Salmonella enterica subsp.
enterica LMG15860. Liu et al. (2013) relataram atividade inibitória da invasão da EIEC por
várias espécies de Lactobacillus, incluindo: L. bulgaricus, L. casei, L. helveticus, L.
plantarum, L. rhamnosus e L. salivarius. Essa atividade foi mais significante para o L.

59
plantarum, em que, além da inibição da capacidade invasora da EIEC em até 34,7%, foi
demonstrada a presença de atividade antimicrobiana (LIU et al., 2013). Essa mesma espécie
também apresentou atividade inibitória da adesão de EPEC (LIU et al., 2011). Frick et al.
(2007), demonstraram que o Lactobacillus fermentum foi capaz de atenuar o efeito pró-
inflamatório de Yersina enterocolitica em células epitelias humanas. O estudo de Hirano et
al. (2003), evidenciou a capacidade do Lactobacillus rhamnosus de amenizar infecção por
Escherichia coli enterohemorágica (EHEC) em células intestinais humanas in vitro. Jacobsen
et al. (1999), relataram que a presença de micro-organismos com propriedades probióticas
impedem a colonização por espécies patogênicas como Listeria monocytogenes, Escherichia
coli e Salmonella spp através de efeito competitivo ou produção de compostos inibitórios.
A inibição da adesão de um patógeno por probiótico pode estar relacionada a um ou
mais dos seguintes mecanismos, os quais incluem: competição por receptores na célula alvo,
produção de um ambiente tóxico, produção de moléculas com atividade antimicrobiana e
também através do mecanismo de coagregação (OELSCHLAEGER, 2010). Com relação aos
mecanismos de inibição da adesão, neste estudo, constatou-se uma correlação significante
entre os Lactobacillus com propriedade de coagregação e a inibição da adesão sobre as
diferentes espécies de E. coli diarreiogênicas, mostrando a importância desse mecanismo na
prevenção da colonização de um tecido por este patógeno. Por outro lado, não foi observada a
produção de substâncias com atividade antimicrobiana no sobrenadante das culturas e a
competição por receptores não foi investigada. Segundo Falagas et al. (2007), a produção de
H2O2 pode representar uma característica muito relevante na seleção de bactérias com
potencial probiótico. Estudos relatam que mulheres que apresentam espécies de Lactobacillus
produtores de H2O2 são menos susceptíveis a desenvolver infecções no trato genital em
relação àquelas que não os possuem (ESCHENBACH et al., 1998; CHERPES et al., 2008).
A importância do fenômeno da coagregação tem sido relatada por outros
pesquisadores. Em ensaios com uropatógenos, tais como E. coli, C. albicans e G. vaginalis,
espécies de Lactobacillus podem realizar a coagregação e ajudar a removê-los do sítio de
infecção (BORIS et al., 1998; OSSET, BARTOLOME et al., 2001). Desta forma, a
coagregação pode ser considerada um importante processo de interação, pois, através dela, os
lactobacilos formam um microambiente em torno dos invasores e depositam substâncias
antimicrobianas de forma localizada, dificultando a colonização dos patógenos
(MASTROMARINO et al., 2002; XU et al., 2009). O mecanismo envolvido na coagregação

60
não está bem esclarecido, mas, aparentemente, as características físico-químicas da superfície
celular, tais como hidrofobicidade, podem interferir na coagregação (OZTURK et al., 2009).
Resultados similares foram descritos para a espécie L. fermentum e a sua capacidade
de coagregar com E. coli (BARBERIS et al., 2010). Coincidentemente, verificou-se, no
presente estudo, que essa espécie foi uma das que demonstrou considerável capacidade de
coagregação, uma vez que foram encontrados resultados positivos contra a maioria das E. coli
diarreiogênicas testadas. Observou-se que esta foi a espécie que mais inibiu a adesão da EIEC
às células eucarióticas. Portanto, os dados da literatura e os encontrados neste estudo
reforçam a ideia de que as espécies de Lactobacillus que apresentam os melhores resultados
no ensaio de coagregação podem se constituir como potenciais micro-organismos em termos
de competição contra enteropatógenos, tais como as E. coli diarreiogênicas.
O estudo de Kaewnopparat et al. (2013), demonstrou que uma estirpe do L. fermentum
isolada do trato genital feminino, denominada de SK 5, foi classificada como uma potencial
candidata probiótica, considerando-se que o micro-organismo produziu compostos que
inibiram o crescimento de E. coli e G. vaginalis, coagregou com esses patógenos, produziu
biofilme, apresentou parede hidrofóbica, aderiu às células HeLa, HT-29 e Caco-2 e inibiu a
adesão dos patógenos às monocamadas epiteliais. Outros trabalhos apresentaram resultados
semelhantes para outros isolados dessa espécie (ANUKAM; REID, 2007; KAUR et al.,
2013).
Um aspecto interessante do uso dos probióticos é que diferentes espécies podem
apresentar propriedades distintas. Isso tem possibilitado o uso conjunto de linhagens
promissoras sob ponto de vista probiótico. O uso combinado de várias espécies de
Lactobacillus constitui-se uma importante ferramenta que pode ser fundamental no processo
de eliminação de uma grande gama de patógenos, haja vista que cada micro-organismo
apresenta suas peculiaridades para competir com diferentes espécies de invasores
(MASTROMARINO et al., 2002; TALLON et al., 2007 MARTIN et al., 2012).
Outras pesquisas que possam evidenciar a interação de L. brevis, L. fermentum e L.
plantarum com enteropatógenos são necessárias, além de estudos in vivo para avaliação do
comportamento dessas espécies bacterianas, objetivando uma melhor compreensão dos
diversos mecanismos de interação micro-organismo-hospedeiro e o fornecimento de subsídios
que possam propor probióticos à base dos lactobacilos estudados.

61
7 CONCLUSÃO
De acordo com os resultados obtidos no presente estudo, pode-se concluir que:
As espécies de Lactobacillus de referência apresentaram potencial de colonizar células
intestinais, considerando-se que elas aderiram significantemente em células HT-29 e HeLa
in vitro;
Entre as espécies de Lactobacillus de referência avaliadas e que são comumente utilizadas
comercialmente em alimentos com características probióticas, L. brevis (ATCC 367), L.
plantarum (ATCC 8014 e L. fermentum (ATCC 23271), realmente apresentam in vitro
capacidade inibitória sobre EIEC, EPEC, EAEC e ETEC, em diferentes níveis;
A capacidade de coagregação apresentada pelas diferentes espécies de Lactobacillus que
foram testadas é sugestiva de um provável mecanismo envolvido na inibição da adesão de
enteropatógenos às células eucarióticas, principalmente o observado nas espécies L.
plantarum e L. fermentum;
A capacidade de inibição da adesão observada nas linhagens de Lactobacillus sobre as
diferentes categorias de E. coli diarreiogênicas testadas não está relacionada à produção de
substâncias com atividade antimicrobiana do sobrenadante das culturas;
Das seis linhagens de Lactobacillus de referência analisadas, quatro possuem superfícies
celulares hidrofóbicas (L. brevis; L. fermentum; L. plantarum e L. rhamnosus) e duas
foram hidrofílicas (L. delbrueckii delbrueckii e L. paracasei) quando avaliadas in vitro
pela utilização de solventes orgânicos;
Nos isolados de cães, quatro foram classificados como hidrofóbicos (L. casei; L.
paraplantarum; L. reuteri e L. salivarius), com exceção de L. johnsonii. Dos cinco
isolados de iogurtes avaliados, quatro foram considerados hidrofóbicos ((L. casei; L. vivos
paracasei; L.casei e L. paracasei), com exceção de L. acidophilus.

62
Das dezesseis linhagens de Lactobacillus analisadas, treze foram produtoras de biofilme,
mas apenas três foram classificadas como produtoras moderadas (ATCC: L. fermentum;
Iogurte: L. casei; Cães: L. casei).

63
REFERÊNCIAS
ANTONIO, M. A.; RABE, L. K.; HILLIER, S. L. Colonization of the rectum by
Lactobacillus species and decreased risk of bacterial vaginosis. J Infect Dis, v. 192, n. 3, p.
394-8, 2005.
ANUKAM, K. C.; REID, G. Lactobacillus plantarum and Lactobacillus fermentum with
probiotic potentials isolated from the vagina of healthy Nigerian woman. Res J Microbiol, v.
2, p. 81-87, 2007.
AXELSSON, L. Lactic acid bactéria: classification and physiology. In: Salminen S, Von
Wright A, eds. Lactic acid bactéria: microbiology and functional aspects. 2nd ed. New York:
Marcel Dekker Inc, p. 1- 72, 1998.
BACKHED, F.; LEY, R. E.; SONNENBURG, J. L.; PETERSON, D. A.; GORDON, J. I.
Host/bacterial mutualism in the human intestine. Science, v. 307, p.1915-1920, 2005.
BARBOSA, F. H. F.; SILVA, A. M.; MARTINS, F. S.; NICOLI, J. R. Perfil de
hidrofobicidade da superfície celular de Bifidobacterium lactis Bb12 e Bifidobacterium
longum Bb46 em função do meio de cultura. Biologia e Ciências da Terra, v. 5, n.2, 2005.
BENGMARCK, S.; ORTIZ DE URBINA, J. J. Simbióticos: uma nueva estrategia em el
tratamiento de pacientes críticos. Nutr. Hosp, v. 2, n.20, p.147-156, 2005.
BERNARDEAU, M.; VERNOUX, J. P.; HENRI-DUBERNET S. AND GUGUEN, M. Safety
assessment of dairy microorganisms: The Lactobacillus genus. Int J Food Microbiol, v. 126,
p.278-285, 2006.
BHATIA, S. J.; KOCHAR, N.;ABRAHAM, P. Lactobacillus acidophilus inhibits growth of
Compylobacter in vitro. J Clin Microbiol, v. 27, p. 2328-2330,1989.
BLACK, R. E.; BROWN, K. H.; BECKER, S. Malnutrition is a determining factor in
diarrheal duration, but not incidence, among young children in a longitudinal study in rural
Bangladesh. Am. J Clin Nutr, v.39, n.1, Jan, p.87-94, 1984.
BORIS, S.; SUAREZ, J. E.; VAZQUEZ, F.; BARBES, C. Adherence of human vaginal
lactobacilli to vaginal epithelial cells and interaction with uropathogens. Infect Immun, v. 66,
n. 5, p. 1985-9, 1998.

64
BRYCE J, BLACK RE, WALKER N, BHUTTA ZA, LAWN JE, STEKETEE RW. Can
the world afford to save the lives of 6 million children each year? Lancet, 365:2193-
200, 2005.
BUERIS, V., et al. Detection of diarrheagenic Escherichia coli from children with and
without diarrhea in Salvador, Bahia, Brazil. Mem Inst Oswaldo Cruz, v.102, n.7, Nov,
p.839-44, 2007.
BUSSCHER, H. J. et al. Relative importance of surface free energy as a measure of
hydrophobicity in bacterial adhesion to solid surfaces. In: DOYLE, R. J.; ROSENBERG, M.
Microbial cell surfaces hydrophobicity. American Society for microbiology, Washington DC,
p. 335-361, 1990.
CANDELA, M.; PERNA, F.; CARNEVALI, P.; VITALI, B.; CIATI, R.; GIONCHETTI, P.;
RIZZELLO, F.; CAMPIERI, M.; BRIGIDI, P. Interaction of probiotic Lactobacillus and
Bifidobacterium strains with human intestinal epithelial cells: Adhesion properties,
competition against enteropathogens and modulation of IL-8 production. Int J Food
Microbiol, v.125, p. 286–292, 2008.
CASTONGUAY, M. H. et al. Biofilm formation by Escherichia coli is stimulated by
synergistic interactions and co-addhesion mechanisms with adherence-proficient bacteria.
Research in Microbiology, v.157, p. 471–478, 2006.
CHAN, R. C.; REID, G.; IRVIN, R. T.; BRUCE, A. W.; COSTERTON, J. W. Competitive
exclusion of uropathogens from human uroepithelial cells by Lactobacillus whole cells and
cells wall fragments. Infect Immun, v.47, n. 1, p. 84-89, 1985.
CHAN, X.; XU, J.; SHUAI, J.; CHEN, J.; ZHANG, Z.; FANG, W. The S-layer proteins of
Lactobacillus crispatus strains ZJ001 is responsible for competitive exclusion against
Escherichia coli O157: H7 and Samonella typhimurium. Int J Food Microbiol, v.115, n. 3, p.
307-312, 2007.
CHERPES, T. L.; HILLIER, S. L.; MEYN, L. A.; BUSCH, J. L.; KROHN, M. A. A delicate
balance: risk factors for acquisition of bacterial vaginosis include sexual activity, absence of
hydrogen peroxide-producing lactobacilli, black race, and positive herpes simplex virus type 2
serology. Sex Transm Dis, v.35, n. 1, p. 78-83, 2008.
CLEVELAND, J.; MONTVILLE, T. J.; NES, I. F.; CHIKINDAS, M.L. Bacteriocins: safe,
natural antimicrobials for food preservation. Int J Food Microbiol, v.71, p. 1–20, 2001.

65
COLLIER-HYAMS, L.S.; NEISH, A.S. Innate immune relationship between commensal
flora and the mammalian intestinal epithelium. Cellular and Molecular Life Science, v.62,
p.1339-1348, 2005.
CONWAY, P. L.; KJELLEBERG, S. Protein-mediated adhesion of Lactobacillus fermentum
strain 737 to mouse stomach squamous epithelium. J Gen Microbiol, v. 135, n. 5, p. 1175-86,
1989.
COPPOLA, M. M.; TURNES, C. G. Probióticos e resposta imune. Ciência Rural, v. 34, n. 4,
p. 1297-1303, 2004.
CORTÉS, J. X. et al. Intestinal adherence associated with type IV pili of enterohemorrhagic
Escherichia coli O157:H7. The Journal of Clinical Investigation, v. 117, n. 11, p. 3519-
3529, 2007.
CRAVIOTO, A. et al. An adhesive factor found in strains of Escherichia coli belonging to the
traditional infantile enteropathogenic serotypes. Curr Microbiol, v.3, p.95-99, 1979.
DE KEERSMAECKER, S. C. J. et al. Strong antimicrobial activity of Lactobacillus
rhamnosus GG against Salmonella typhimurium is due to accumulation of lactic acid. FEMS
Microbiology Letters, v. 259, n. 1, p. 89-96, 2006.
DESMOND, C.; FITZEGERALD, G. F.; STANTON, C.; ROSS, R. P. Improved stress
tolerance of GroESL – overproducing Lactococcus lactis and probiotic Lactobacillus
paracasei NFBC 338. Appl Environ Microbiol, v.70, n.10, p.5929-5936, 2004.
DOYLE, P. M; CLIVER, D. O. Foodborne Diseases London; New York: Academic Press, p.
209 – 2015, 1990.
DRASAR, B. S; HILL, M. J. Human Intestinal Flora. London; New York: Academic Press,
1974.
DUARY, R. K.; RAJPUT, Y. S.; BATISH, V. K.; GROVER, S. Assessing the adhesion of
putative indigenous probiotic lactobacilli to human colonic epithelial cells. Indian J Med
Res, v. 134, n. 5, p. 664-71, 2011.

66
DUC, L. H.; HONG, H. A.; BARBOSA, T. M.; HENRIQUES, A. O.; CUTTING, S. M.
Characterization of bacillus probiotics available for human use. Appl Environ Microbiol,
v.70, n.4, p.2161-2171, 2004.
ESCHENBACH, D. A.; DAVICK, P. R.; WILLIAMS, B. L.; KLEBANOFF, S. J.; YOUNG-
SMITH, K.; CRITCHLOW, C. M.; HOLMES, K. K. Prevalence of hydrogen peroxide-
producing Lactobacillus species in normal women and women with bacterial vaginosis. J
Clin Microbiol, v. 27, n.2, p.251-256, 1989.
FALAGAS, M.; BETSI, G. I.; ATHANASIOU, S. Probiotics for the treatment of women with
bacterial vaginosis. Clin Microbiol Infect, v.13, n. 7, p. 657-664, 2007.
FELIS, G. E.; DELLAGLIO, F. Taxonomy of lactobacilli and bifidobacteria. Current issues
in intestinal microbiology, v.8, n. 2, p. 44-61, 2007.
FERREIRA, C. L.L.F.; Grupo de bactérias láticas: caracterização e aplicação tecnológica de
bactérias láticas probióticas. In: Ferreira, CLLF, ed. Prebióticos e Probióticos: Atualização e
Prospecção. Suprema Gráfica e Editora, Rio Branco, MG. p. 7-26, 2003.
FERRIERES, L.V.; HANCOCK, P.; KLEMM. Specific selection for virulent urinary tract
infections Escherichia coli strains during catheter-associated biofilm formation. FEMS
Immunol Med Microbiol, v.51, n.1, Oct, p.212-219, 2007b.
FINLAY, B. B.; FALKOW, S. Common themes in microbial pathogenicity revisited.
Microbiol Mol Biol Rev, v.61, p.136–16, 1997.
FOOKS, L.J.; GIBSON, G.R. Probiotics as modulators of the gut flora. British Journal of
Nutrition., v.88, p.39-49, 2002.
FOOD AND AGRICULTURE ORGANIZATION OF THE UNITED NATIONS; WORLD
HEALTH ORGANIZATION. Guidelines for the evaluation of probiotics in food. Joint
FAO/WHO Working Group Report on Drafting London. Ontario, Canadá, April and
May, v.1, 2002.
FRECE, J.; KOS, B.; SVETEC, I. K.; ZGAGA, Z.; MRSA, V.; SUSKOVIC, J. Importance of
S-layer proteins in probiotic activity of Lactobacillus acidophilus M92. J Appl Microbiol,
v.98, n. 2, p.285-292, 2005.

67
FRICK, J.S. et al. Lactobacillus fermentum attenuates the proinflammatory effect of Yersinia
enterocolitica on human epithelial cells. Inflammatory Bowel Diseases, v.13, p.83-90, 2007.
FULLER, R. Probiotics in man and animals. J Appl bacteriol, Oxford, v.66, n.5, p.365 –
378,1989.
GRANATO, D.; BERGONZELLI, G. E.; PRIDMORE, D. R.; MARVIN, L.; ROUVET, M.;
THEULAZ, I. E. C. Cell surface-associated elongation factor tu mediates the attachament of
lactobacillus jhonsonii NCC533 (La1) to human intestinal cells and mucins. Infect Immun,
v. 72, n.4 , p. 2160-2169, 2004.
GROSSO, C. R. F.; TRINDADE, C. S. F. Stability of free and immobilized Lactobacillus
acidophilus and Bifidobacterium lactis in acidifield milk and of immobilized B.lactis in
yoghurt. Braz J Microbiol, v. 35, p. 151-156, 2004.
GUARNER, F., MALGELADA, J.R., 2003. Gut flora in health and disease. Lancet, v. 361,
p. 512-519, 2003.
GUERRANT, D. I. et al. Association of early childhood diarrhea and cryptosporidiosis with
impaired physical fitness and cognitive function four-seven years later in a poor urban
community in northeast Brazil. Am J Trop Med Hyg, v.61, n.5, Nov, p.707-713, 1999.
HAMMES, W.P; HERTEL, C. The Genera Lactobacillus and Carnobacterium. In: The
Prokaryotes Release 3.15, Editor Martin Dworkin, 2003.
HIRANO, J.; YOSHIDA, T.; SUGIYAMA, T.; KOIDE, N.; Mori, I.; YOKOCHI, T. The
effect of Lactobacillus rhamnosus on enterohemorrhagic Escherichia coli infection of human
intestinal cells in vitro. Microbiology and Immunology, v.47, p.405-409, 2003.
HUIS IN’TVELD, J. H. J.; SHORTT, C. Selection criteria for probiotic microorganisms. In:
Leeds, A. R.; Rowland, I. R. Ed. Gut flora and healthc – past, present and future.
Internacional Congress and Symposium 219, Royal Society of Medicine Press Limited,
p. 27-36, 1996.
Instituto Nacional de Propriedade Industrial (INPI). Endereço na Internet:
http://www.inpi.gov.br/menu-superior/imprensa/clipping/novembro-2007-1-old-version-
replaced 12112007-124018/22-11-2007#2.

68
JACK, R. W.; TAGG, J. R.; RAY, B. bacteriocins of Gram-positive bacteria. Microbiology
Reviews, v.59, p.171-200, 1995.
JACOBSEN, C. N.; NIELSEN, V. R.; HAYFORD, A. E.; MOLLER, P. L.; MICHAELSEN,
K. F,; PAERREGAARD, A.; SANDSTROM, B.; TVEDE, M.; JAKOBSEN, M. Screening of
probiotic activities of forty-seven strains of lactobacillus spp. By in vitro techniques and
evaluation of colonization ability of five selected strains in humans. Appl Environ
Microbiol, v.65, n.11, p.4949-4956, 1999.
JAMISON, D.T.; WORLD BANK; DISEASE CONTROL PRIORITIES PROJECT.
Disease control priorities in developing countries. 2nd. New York, Washington, DC:
Oxford University Press; World Bank, 2006.
JONES, D. S. et al. Standardisation and comparison of methods employed for microbial cell
surface hydrophobicity and change determination. International Journal of Pharmaceutics, v.
131, p.83-89, 1996.
KAISER, A. L.; MONTVILLE, T. J.; Purification of the bacteriocin MN and characterization
of its mode of action against Listeria monocytogenes scott a cells and lipids vesicles. Appl
Environ Microbiol, v.62, p.4529-4535, 1996.
KAEWNOPPARAT, S.; DANGMANEE, N.; KAEWNOPPARAT, N.; SRICHANA, T,;
CHULASIRI, M.; SETTHARAKSA, S. In vitro probiotic properties of Lactobacillus
fermentum SK5 isolated from vagina of a healthy woman. Anaerobe, v.22, p. 6-13, 2013.
KAMINOGAWA, S.; NANNO, M. Modulation of immune functions by foods. CAM. v.3,
p.241-250, 2004.
KANDLLER, O.; WEISS, N. Genus Lactobacillus. In: Sneath, P. H. A., Mair, N. S., Sharpe,
M. E., Holt, J. G. Bergey' s Manual of Systematic Bacteriology, v.2, Williams and Wilkins,
Baltimore M. D., USA, p.1209-1234, 1986.
KAPER, J. B.; NATARO, J. P.; MOBELY, H.L.T. Pathogenic Escherichia coli. Nat Rev
Microbiol, v.2, p.123-40, 2004.

69
KAUR, B.; BALGIR, P.; MITTU, B.; CHAUHAN, A.; KUMAR, B. Purification and
physicochemical characterization of anti-gardnerella vaginalis bacteriocin hv6b produced by
Lactobacillus Fermentum isolate from human vaginal ecosystem. Am J Biochem mol Biol,
v.3 p.91-100, 2013.
KELLY, D., CONWAY, S., AMINOV, R. Commensal gut bacteria: mechanisms of immune
modulation. Trends in Immunology, v. 26, p. 326-333, 2005.
KIMOTO, H.; KURISAKI, J.; TSUJI, N. M.; OHMOMO, S.; OKAMOTO, T. Lactococci as
probiotic strains: adhesion to human enterocyte-like Caco-2 cells and tolerance to low pH and
bile. Lett Appl Microbiol, v. 29, n. 5, p. 313-6, 1999.
KLAENHAMMER, T. R. Genetics of bacteriocins produced by lactic acid bacteria. FEMS
Microbiology Reviews, v. 12, p. 39-85, 1993.
KLINGER, B.; HANAWAY, P.; COHRSSEN, A. Probiotics in children. Pediatric Clinics of
North America, v. 54, p. 949-967, 2007.
KOMATSU, T. R.; BURITI, F. C. A.; SAAD, S. M. I. Inovação, persistência e criatividade
superando barreiras no desenvolvimento de alimentos probióticos. Revista Brasileira de
Ciências Farmacêuticas, vol. 44, n.3, jul./set., 2008.
KOSEK, M.; BERN, C.; GUERRANT, R. L. The global burden of diarrhoeal disease, as
estimated from studies published between 1992 and 2000. Bull World Health Organ, v.81,
n.3, p.197-204, 2003.
KOS, B.; SUSKOVIC, J.; VUKOVI, S.; SIMPRAGA, M.; FRECE, J.; MATOSIC, S.
Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J Appl
Microbiol, v.94, n.6, p.981-987, 2003.
KRIEG, N. R. Identificacion of Bacteria. Bergey' s Manual of Systematic Bacteriology, V.
3, p. 1624-1626, 1984.
LAPARRA, J. M.; SANZ, Y. Comparison of in vitro models to study bacterial adhesion to the
intestinal epithelium. Lett Appl Microbiol, v. 49, n. 6, p. 695-701, 2009.
LEBEER, S.; VANDERLEYDEN, J.; DE KEERSMAECKER, S. C. Genes and Molecules of
Lactobacilli Supporting Probiotic Action. Microbiology and Molecular Biology Reviews, v.
72, n. 4, p. 728-764, 2008.

70
LEBEER, S.; VANDERLEYDEN, J.; DE KEERSMAECKER, S. C. Host interactions of
probiotic bacterial surface molecules: comparison with commensals and pathogens. Nat Rev
Microbiol, v. 8, n. 3, p. 171-184, 2010.
LEPARGNEUR, J. P.; ROUSSEAU, V. Protective role of the Doderlein flora. J Gynecol
Obstet Biol Reprod (Paris), v. 31, n. 5, p. 485-94, 2002.
LIU, Z.; SHEN, T.; ZHANG, P.; MA, Y.; QIN, H. Lactobacillus plantarum surface layer
adhesive protein protects intestinal epithelial cells against tight junction injury induced by
enteropathogenic Escherichia coli. Mol Biol Rep, v. 38, p. 3471-3480, 2011.
LIU, X.; LIU, W.; ZHANG, Q.; TIAN, F.; WANG, G.;; ZHANG, H.; CHEN, W. Screening
of lactobacilli with antagonistic activity against enteroinvasive Escherichia coli. J Food
Control, v. 30, p. 563-568, 2013.
MACK, D. R. et al. Extracellular MUC3 mucin secretion follows adherence of
Lactobacillus strains to intestinal epithelial cells in vitro. Gut, v. 52, n. 6, p. 827-833, Jun
2003.
MACKIE, R.I.; SGHIR, A.; GASKINS, H.R. Developmental microbial ecology of the
neonatal gastrointestinal tract. American Journal of Clinical Nutrition, v. 69, p.1035-1045,
1999.
MACPHERSON, A.J., HARRIS, N.L. Interactions between commensal intestinal bacteria
and the immune system. Nature Reviews Immunology, v. 4, p. 478-485, 2004.
MARTIN, R.; SANCHEZ, B.; SUAREZ, J. E.; URDACI, M. C. Characterization of the
adherence properties of human Lactobacilli strains to be used as vaginal probiotics. FEMS
Microbiol Lett, v. 328, n. 2, p. 166-73, 2012.
MARTIN, R.; MARTIN, C.; ESCOBEDO, S.; SUAREZ, J. E.; QUIRÓS, L. Surface
glycosaminoglycans mediate adherence between HeLa cells and Lactobacillus salivarius
Lv72. BMC Microbiol, v. 13, n. 210, p. 1-11, 2013.
MASTROMARINO, P.; BRIGIDI, P.; MACCHIA, S.; MAGGI, L.; PIROVANO, F.;
TRINCHIERI, V.; CONTE, U.; MATTEUZZI, D. Characterization and selection of vaginal
Lactobacillus strains for the preparation of vaginal tablets. J Appl Microbiol, v. 93, n. 5, p.
884-93, 2002.

71
MAYRA-MAKINEN, A.; MANNINEN, M.; GYLLENBERG, H. The adherence of lactic
acid bacteria to the columnar epithelial cells of pigs and calves. J Appl Bacteriol, v. 55, n. 2,
p. 241-5, 1983.
MENG, J.; ZHU, X.; GAO, S. M.; ZHANG, Q. X.; SUN, Z,; LU, R. R. Characterization of
surface layer proteins and its role in probiotic properties of three Lactobacillus strains. Int J
Biol Macromol, v. 65, p. 110-114, 2014.
MORAIS, M. B.; JACOB, C. M. A. The role of probiotics and prebiotics in pediatric practice.
Jornal de Pediatria, Sociadade Brasileira de Pediatria., vol. 82, n.5, 2006.
NATARO, J. P.; KAPER, J. B. Diarrheagenic Escherichia coli. Clin Microbiol Rev, v.11,
n.1, Jan, p.142-201, 1998.
NATARO, J. P.; STEINER, T.; GUERRANT, R. L. Enteroaggregative Escherichia coli.
Emerg Infect Dis, v.4, n.2, Apr-Jun, p.251- 261, 1998.
NAVA, G. M.; DAVILA, M. Nuevas perpectivas em La selection and evaluation of
probiotics. Rer Chil Nutr, v.21, p.184-185, 2004.
OELSCHLAEGER, T. A. Mechanisms of probiotic actions – A review. Int J Med
Microbiol, v.300, p.57-62, 2010.
ORLA-JENSEN, S. In: Orla-Jensen, S. (Ed.), The lactic acid bacteria. Host, Copenhagen., p.
1-196, 1919.
OSSET, J.; BARTOLOME, R. M.; GARCIA, E.; ANDREU, A. Assessment of the capacity
of Lactobacillus to inhibit the growth of uropathogens and block their adhesion to vaginal
epithelial cells. J Infect Dis, v. 183, n. 3, p. 485-91, 2001.
OYETAYO, V. O.; OYETAYO, F. L. Review: Potential of probiotics as biotherapeutic
agents targeting the innate immune system. Afr J Biotechnol, v. 4, n. 2, p. 123-127, 2005.

72
OUWEHAND, A. C.; SALMINEN, S.; ISOLAURI, E. Probiotics: an overview of beneficial
effects. Antonie Van Leeuwenhoek International Journal of General and Molecular
Microbiology, v. 82, p. 279-289, 2002.
OZTURK, S.; ASLIM, B.; EKMEKCI, H. Characterization of vaginal lactobacilli
coaggregation ability with Escherichia coli. Microbiol Immunol, v. 53, p. 59-65, 2009.
PELLETIER, C.; BOULEY, C.C.; BOUTTIER, S.; BOURLIOUX, P.; BELLON-
FONTAINE, M-N. Cell surface characteristics of Lactobacillus casei subsp. casei,
Lactobacillus paracasei subsp. paracasei, and Lactobacillus rhamnosus strains. Appl
Environ Microbiol, v. 63, p. 1725-1731, 1997.
PENÃ, J. A.; LI, S. Y.; WILSON, P. H.; THIBODEAU, S. A.; SZARY, A. J.;
VERSALOVIC, J. Genotypic and phenotypic studies of murine intestinal lactobacilli: species
differences in mice with and without colitis. Appl Environ Microbiol, v.70, n.1, p. 558-568,
2004.
PRADO, C. S.; SANTOS, W. L. M.; CARVALHO, C. R.; MOREIRA, E. C.; COSTA, J. O.
Antimicrobial activity of lactic acid bacteria isolated from Brasilian dry fermented sausages
against Listeria monocytogenes. Arq Bras Med Vet Zootec, v.52, n.4, p. 417-423, 2000.
PRIDMORE, R. D.; BERGER, B.; DESIERE, F.; VILANOVA, D.; BARRETTO, C.;
PITTET, A. C.; ZWAHLEN, M. C.; ROUVERT, M.; ALTERMANN, E.; BARRANGOU,
R.; MOLLET, B.; MERCENIER, A.; KLAENHAMMER, T.; ARIGONI, F.; SCHELL, M. A.
The genome sequence of the probiotic intestinal bacterium Lactobacillus johasonii NCC 533.
PNAS., v.101, n.8, p. 2512- 2517, 2004.
REID, G.; HEINEMANN, C.; VELRAEDS, M.; VAN DER MEI, H. C.; BUSSCHER, H. J.
Biosurfactants produced by Lactobacillus. Methods Enzymol, v. 310, p. 426-33, 1999.
REID, G. The scientifics basis for probiotic strains of Lactobacillus. Appl Environ
Microbial, v. 65, n.9, p. 3763-3766, 1999.
REID, G.; McGROARTY, J. A.; DOMINGUE, P. A. G.; CHOW, A. W.; BRUCE, A. W;
EISEN, A.; COSTERTON, J. W. Coaggregation of Urogenital Bacteria in Vitro and in
Vivo. Current Microbiology, v. 20, p. 47-52, 1990.
RITCHIE, J. M.; WAGNER, P. L.; ACHESON, D. W. K.; WALDOR, M. K. Comparison of
Shiga Toxin production by hemolytic-uremic syndrome – associated and bovine- associated

73
Shiga Toxin-producing Escherichia coli isolates. Appl Environ Microbiol, v. 69, n. 2,
p.1059-1066, 2003.
SAARELA, M.; MOGENSEN, G.; FONDEN, R.; MATTO, J.; MATTILA-SANDHOLM, T.
Probiotics bacteria: safety, functional and technological properties. J Biotechnol, v. 84,
p.197-215, 2000.
SAAVEDRA, J.M. et al. Feeding of Bifidobacterium bifidum and Streptococcus thermophilus
to infants in hospital for prevention of diarrhea and shedding of rotavirus. Lancet, v. 344,
p.1046-1049, 1994.
SAMBROOK, J.; FRITSCH, E. F.; MANIATIS, T. Molecular Cloning: a laboratory manual.
2nd
ed. N.Y., Cold Spring Harbor Laboratory, Cold Spring Harbor Laboratory Press, ISBN 0-
87969-309-6, 1659 p, 1989.
SÁNCHEZ, B.; URDACI, M. C. Extracellular proteins from Lactobacillus plantarum
BMCM12 prevent adhesion of Enteropathogens to mucin. Curr Microbiol, v. 64, p.592-596,
2012.
SCHLESSELMANN, J. J. Case-control studies Design conduct analysis, New York:
Oxford University Press, p.354, 1982.
SETH, A. et al. Probiotics ameliorate the hydrogen peroxide-induced epithelial barrier
disruption by a PKC- and MAP kinase-dependent mechanism. American Journal of
Physiology-Gastrointestinal and Liver Physiology, v. 294, n. 4, p. G1060-G1069, Apr
2008.
SHARMA, M.; YADAV, S.; CHAUDHARY, U. Biofilm production in uropathogenic
Escherichia coli. Indian J Pathol Microbiol, v.52, n.2, Apr-Jun, p.294, 2009.
SINGH, D.; VON OSSOWSKI, I.; PALVA, A.; KRISHNAN, V. Purification, crystallization
and preliminary crystallographic analysis of the SpaA backbone-pilin subunit from probiotic
Lactobacillus rhamnosus GG. Acta Crystallogr Sect F Struct Biol Cryst Commun, v.69,
n.2, p.1182-1185, 2013.

74
STOODLEY, L. H.; COSTERTON, J. W.; STOODLEY, P. Bacterial biofilms: from the
natural environment to infectious diseases. Nature Reviews Microbiology, v. 2, p. 95-108,
2004.
SPURBECK, R. R.; ARVIDSON, C. G. Lactobacillus jensenii surface-associated proteins
inhibit Neisseria gonorrhoeae adherence to epithelial cells. Infect Immun, v. 78, n. 7, p.
3103-3111, 2010.
STEPANOVIC, S., et al. Biofilm formation by Salmonella spp. and Listeria
monocytogenes on plastic surface. Lett Appl Microbiol, v.38, n.5, p.428-432, 2004.
TALLON, R.; ARIAS, S.; BRESSOLLIER, P.; URDACI, M. C. Strain-and matrix-dependent
adhesion of Lactobacillus plantarum is mediated by proteinaceous bacterial compounds. Appl
Microbiol, v. 102, n.2, p. 442-451, 2007.
TANNOCK, G. W. A special fondness for lactobacilli. Appl Environ Microbiol, v.70, n.6,
p. 3189-3194, 2004.
TRABULSI, L.R.; SAMPAIO, M.M.S.C. A composição e papel da microflora intestinal na
saúde e proteção do organismo. Os Probióticos e a Saúde Infantil, v. 1, Brasil: Nestlé
Ltda., p. 3-11, 2000.
VAN DEN ABBEELE, P.; GROOTAERT, C.; POSSEMIERS, S.; VERSTRAETE, W.;
VERBEKEN, K.; VAN DE WIELE, T. In vitro model to study the modulation of the mucin-
adhered bacterial community. Appl Microbiol Biotechnol, v. 83, n. 2, p. 349-59, 2009.
VAN LOOSDRECHT, M. C. M. et al. Electrophoretic mobility and hydrophobicity as a
measure to predict the initial steps of bacterial adhesion. Appl Environ Microbiol, v. 53, p.
1898-1901, 1987.
VAN TASSEL, M. L.; MILLER, M. J. Lactobacillus adhesion to mucus. Nutrients, v. 3, n. 5,
p. 613-636, 2011.
VARAVALLO, M. A.; THOMÉ, J. N.; TESHIMA, E. Aplicação de bactérias probióticas para
profilaxia e tratamento de doenças gastointestinais. Semina: Ciências Biológicas e da Saúde,
Londrina, vol. 29, n.1, p. 83-104, jan./jun. 2008.
VASILJEVIC, T.; SHAH, N. P. Probiotics – from metchnikoff to bioactives. Internacional
Dairy Journal, vol. 18, n.1, p. 714-728, 2008.

75
WANGER, R. A.; MURRAY, B. E., MATHEWSON, J. J.; DuPont, H. L. Enteroinvasive
Escherichia coli in travellers with diarrhea. Journal of Infectious Diseases, v.158, p. 640-
642, 1998.
WHITTAKER, C. J.; KLIER, C. M.; KOLENBRANDER, P. E. Mechanisms of adhesion by
oral bacteria. Annu, Rev Microbial, vol. 50, p. 513-552, 1996.
YAN, F. et al. Soluble proteins produced by probiotic bacteria regulate intestinal
epithelial cell survival and growth. Gastroenterology, v. 132, n. 2, p. 562-575, Feb 2007.
XU, H.; JEONG, H. S.; LEE, H. Y.; AHN, J. Assessment of cell surface properties and
adhesion potential of selected probiotic strains. Lett Appl Microbiol, v. 49, n. 4, p. 434-442,
2009.
ZHOU, X.; BROWN, C. J.; ABDO, Z.; DAVIS, C. C.; HANSMANN, M. A.; JOYCE, P.;
FOSTER, J. A.; FORNEY, L. J. Differences in the composition of vaginal microbial
communities found in healthy Caucasian and black women. ISME J, v. 1, n. 2, p. 121-33,
2007.
ZHANG, W.; WANG, H.; LIU, J.; ZHAO, Y.; GAO, K.; ZHANG, J. Adhesive ability means
inhibition activities for lactobacillus against pathogens and S-layer protein plays an important
role in adhesion. Anaerobe, v. 22, p. 97-103, 2013.
ZOETENDAL, E.G., Vaughan, E.E., De Vos, W.M. A microbial world within us. Molecular
Microbiology, v. 59, p.1639-1650, 2006.