Embed Size (px)
Citation preview

University of Groningen
In Bacillus subtilis LutR is part of the global complex regulatory network governing theadaptation to the transition from exponential growth to stationary phaseİrigül-Sönmez, Öykü; Köroğlu, Türkan E.; Öztürk, Büşra; Kovács, Ákos T.; Kuipers, Oscar;Yazgan-Karataş, Ayten; Zuber, P.Published in:Microbiology
DOI:10.1099/mic.0.064675-0
IMPORTANT NOTE: You are advised to consult the publisher's version (publisher's PDF) if you wish to cite fromit. Please check the document version below.
Document VersionPublisher's PDF, also known as Version of record
Publication date:2014
Link to publication in University of Groningen/UMCG research database
Citation for published version (APA):İrigül-Sönmez, Ö., Köroğlu, T. E., Öztürk, B., Kovács, Á. T., Kuipers, O. P., Yazgan-Karataş, A., & Zuber,P. (Ed.) (2014). In Bacillus subtilis LutR is part of the global complex regulatory network governing theadaptation to the transition from exponential growth to stationary phase. Microbiology, 160(2), 243-260.DOI: 10.1099/mic.0.064675-0
CopyrightOther than for strictly personal use, it is not permitted to download or to forward/distribute the text or part of it without the consent of theauthor(s) and/or copyright holder(s), unless the work is under an open content license (like Creative Commons).
Take-down policyIf you believe that this document breaches copyright please contact us providing details, and we will remove access to the work immediatelyand investigate your claim.
Downloaded from the University of Groningen/UMCG research database (Pure): http://www.rug.nl/research/portal. For technical reasons thenumber of authors shown on this cover page is limited to 10 maximum.
Download date: 10-02-2018

Supplementary Figure Legends
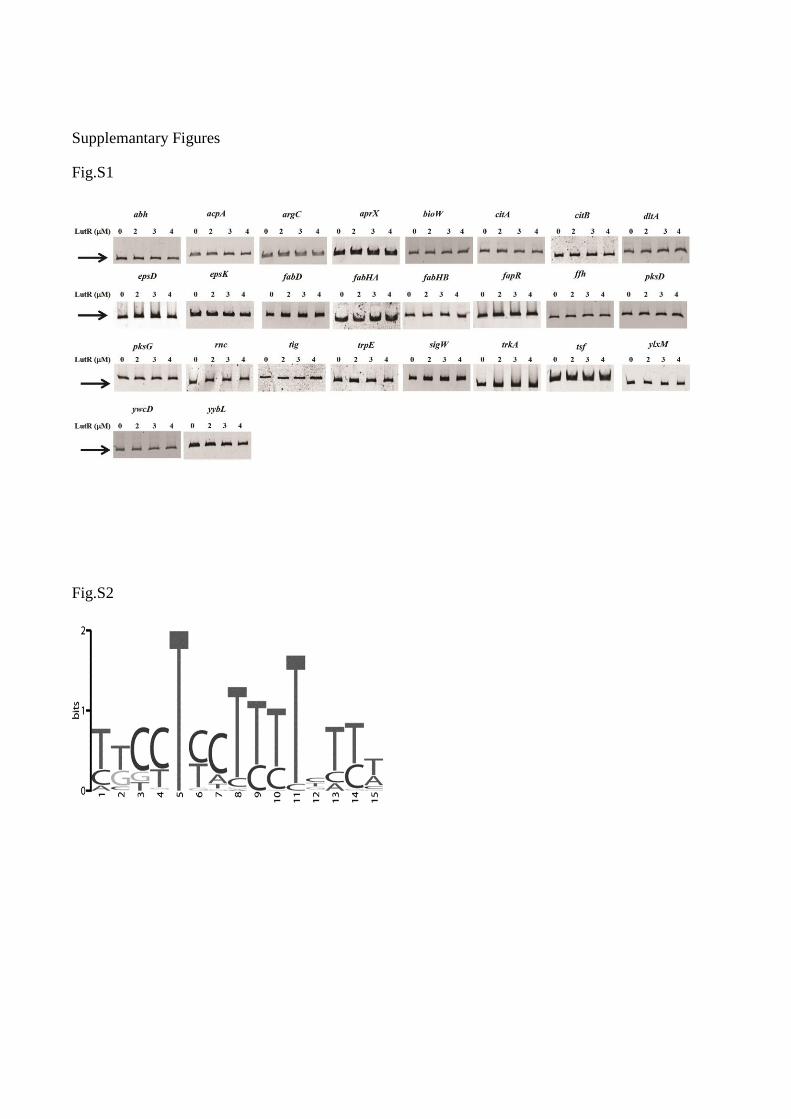
Fig. S1. Regulatory regions of genes with no gel- shift observed. Electrophoretic Mobility Shift
Assays were performed in the presence of putative regulatory regions of genes under the control of
LutR and purified LutR-His6 at the indicated concentrations. In each assay, 25 l total reaction
mixture supplemented with competitor DNA polydIdC (1 g/l), BSA (1 mg/ml). Promoter region
of the unrelated gene ywbH was used as negative control. Positive and negative controls were run
together with each EMSA employed. For detection, gels were treated with SYBR Green I Nucleic
Acid Gel Stain (1/10.000, v/v) (Roche) and visualized with a UV transilluminator. Each gel-shift
assay was repeated at least two times
Fig. S2. Putative LutR binding motif is displayed as sequence LOGO. Each stack represents a
position in the sequence. The height of the individual letters in a stack is the probability of the letter
at that position multiplied by the total information content of the stack
Fig. S3. Effect of lutR mutation on the expression of spoIIE. Cells were grown in DSM medium at
37oC and spollE-directed β-galactosidase activity was determined at the indicated times. Time zero
denotes the end of exponential growth thus initiation of sporulation in DSM. β-galactosidase activity
of spoIIE::lacZ fusion (squares) and its isogenic derivative lutR mutant (spoIIE::lacZ::cat,
lutR::Tn10::spc) (triangle). Error bars indicate the standard deviation of the mean of three
independent experiments (n=3).
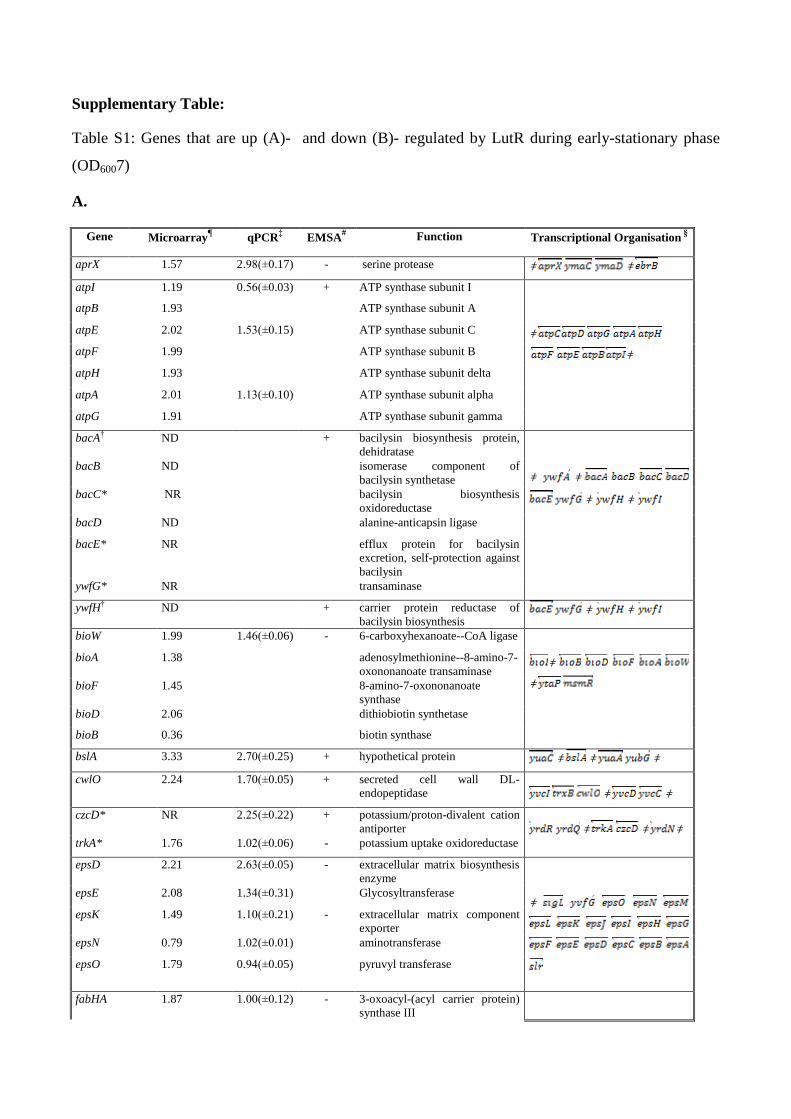
Supplementary Table:
Table S1: Genes that are up (A)- and down (B)- regulated by LutR during early-stationary phase
(OD6007)
A.
Gene
Microarray
¶ qPCR
‡ EMSA
# Function Transcriptional Organisation §
aprX 1.57 2.98(±0.17) - serine protease ǂ ǂ
atpI 1.19 0.56(±0.03) + ATP synthase subunit I
atpB 1.93 ATP synthase subunit A ǂ
ǂ
atpE 2.02 1.53(±0.15) ATP synthase subunit C
atpF 1.99 ATP synthase subunit B
atpH 1.93 ATP synthase subunit delta
atpA 2.01 1.13(±0.10) ATP synthase subunit alpha
atpG 1.91 ATP synthase subunit gamma
bacA† ND + bacilysin biosynthesis protein,
dehidratase
ǂ ǂ
ǂ ǂ
bacB ND isomerase component of
bacilysin synthetase
bacC* NR bacilysin biosynthesis
oxidoreductase
bacD ND alanine-anticapsin ligase
bacE* NR efflux protein for bacilysin
excretion, self-protection against
bacilysin
ywfG* NR transaminase
ywfH† ND + carrier protein reductase of
bacilysin biosynthesis ǂ ǂ
bioW 1.99 1.46(±0.06) - 6-carboxyhexanoate--CoA ligase
ǂ
ǂ
bioA 1.38 adenosylmethionine--8-amino-7-
oxononanoate transaminase
bioF 1.45 8-amino-7-oxononanoate
synthase
bioD 2.06 dithiobiotin synthetase
bioB 0.36 biotin synthase
bslA 3.33 2.70(±0.25) + hypothetical protein ǂ ǂ ǂ
cwlO 2.24 1.70(±0.05) + secreted cell wall DL-
endopeptidase
ǂ ǂ
czcD* NR 2.25(±0.22) + potassium/proton-divalent cation
antiporter
ǂ ǂ ǂ trkA* 1.76 1.02(±0.06) - potassium uptake oxidoreductase
epsD 2.21 2.63(±0.05) - extracellular matrix biosynthesis
enzyme
ǂ
epsE 2.08 1.34(±0.31) Glycosyltransferase
epsK 1.49 1.10(±0.21) - extracellular matrix component
exporter
epsN 0.79 1.02(±0.01) aminotransferase
epsO 1.79 0.94(±0.05) pyruvyl transferase
fabHA 1.87 1.00(±0.12) - 3-oxoacyl-(acyl carrier protein)
synthase III
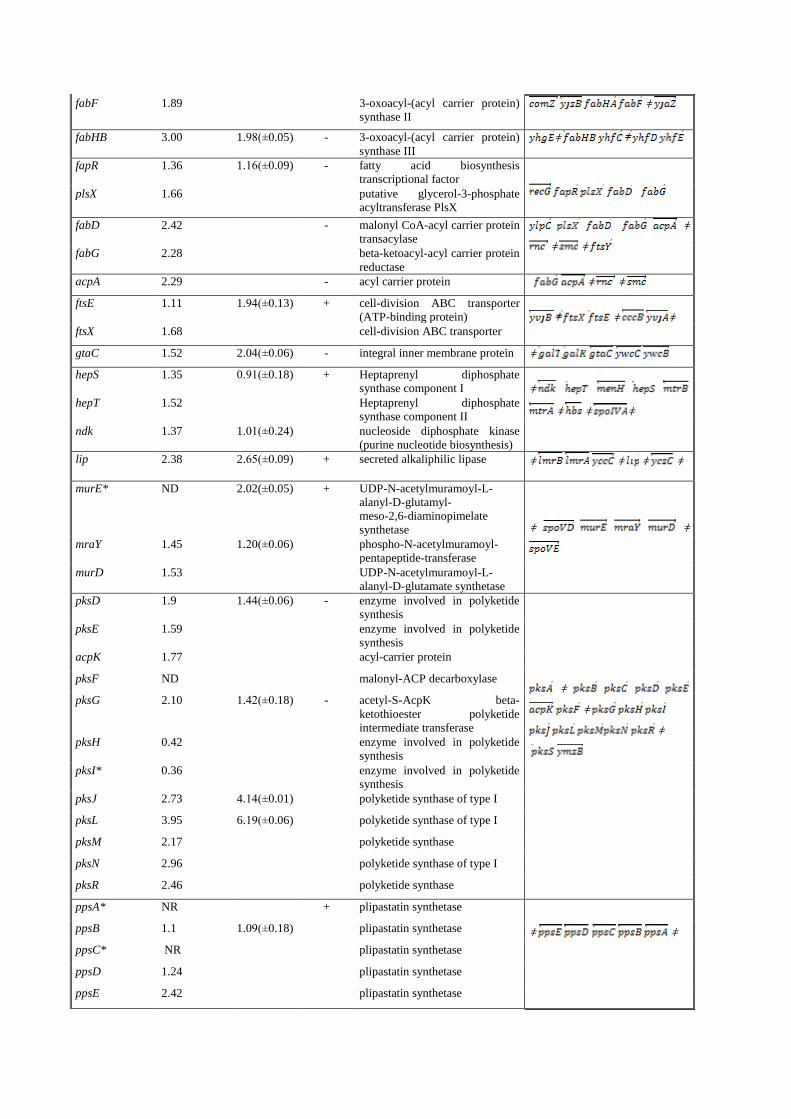
fabF 1.89 3-oxoacyl-(acyl carrier protein)
synthase II ǂ
fabHB 3.00 1.98(±0.05) - 3-oxoacyl-(acyl carrier protein)
synthase III ǂ
fapR 1.36 1.16(±0.09) - fatty acid biosynthesis
transcriptional factor
plsX 1.66 putative glycerol-3-phosphate
acyltransferase PlsX
fabD 2.42 - malonyl CoA-acyl carrier protein
transacylase ǂ
ǂ ǂ
fabG 2.28 beta-ketoacyl-acyl carrier protein
reductase
acpA 2.29 - acyl carrier protein ǂ ǂ
ftsE 1.11 1.94(±0.13) + cell-division ABC transporter
(ATP-binding protein)
ǂ ǂ
ftsX 1.68 cell-division ABC transporter
gtaC 1.52 2.04(±0.06) - integral inner membrane protein ǂ
hepS 1.35 0.91(±0.18) + Heptaprenyl diphosphate
synthase component I
ǂ
ǂ ǂ ǂ
hepT 1.52 Heptaprenyl diphosphate
synthase component II
ndk 1.37 1.01(±0.24) nucleoside diphosphate kinase
(purine nucleotide biosynthesis)
lip 2.38 2.65(±0.09) + secreted alkaliphilic lipase ǂ ǂ ǂ ǂ
murE* ND 2.02(±0.05) + UDP-N-acetylmuramoyl-L-
alanyl-D-glutamyl-
meso-2,6-diaminopimelate
synthetase
ǂ ǂ
mraY 1.45 1.20(±0.06) phospho-N-acetylmuramoyl-
pentapeptide-transferase
murD 1.53 UDP-N-acetylmuramoyl-L-
alanyl-D-glutamate synthetase
pksD 1.9 1.44(±0.06) - enzyme involved in polyketide
synthesis
ǂ
ǂ
ǂ
pksE 1.59 enzyme involved in polyketide
synthesis
acpK 1.77 acyl-carrier protein
pksF ND malonyl-ACP decarboxylase
pksG 2.10 1.42(±0.18) - acetyl-S-AcpK beta-
ketothioester polyketide
intermediate transferase
pksH 0.42 enzyme involved in polyketide
synthesis
pksI* 0.36 enzyme involved in polyketide
synthesis
pksJ 2.73 4.14(±0.01) polyketide synthase of type I
pksL 3.95 6.19(±0.06) polyketide synthase of type I
pksM 2.17 polyketide synthase
pksN 2.96 polyketide synthase of type I
pksR 2.46 polyketide synthase
ppsA* NR + plipastatin synthetase ǂ ǂ
ppsB 1.1 1.09(±0.18) plipastatin synthetase
ppsC* NR plipastatin synthetase
ppsD 1.24 plipastatin synthetase
ppsE 2.42 plipastatin synthetase
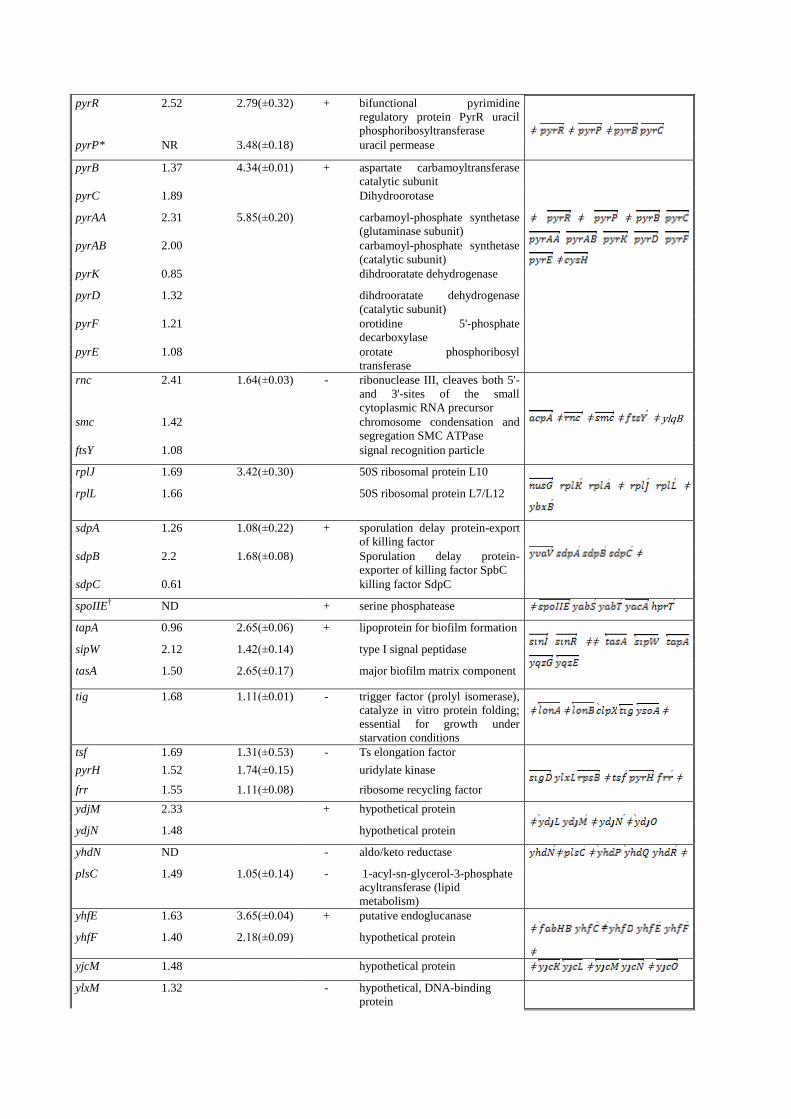
pyrR 2.52 2.79(±0.32) + bifunctional pyrimidine
regulatory protein PyrR uracil
phosphoribosyltransferase
ǂ ǂ ǂ
pyrP* NR 3.48(±0.18) uracil permease
pyrB 1.37 4.34(±0.01) + aspartate carbamoyltransferase
catalytic subunit
ǂ ǂ ǂ
ǂ
pyrC 1.89 Dihydroorotase
pyrAA 2.31 5.85(±0.20) carbamoyl-phosphate synthetase
(glutaminase subunit)
pyrAB 2.00 carbamoyl-phosphate synthetase
(catalytic subunit)
pyrK 0.85 dihdrooratate dehydrogenase
pyrD 1.32 dihdrooratate dehydrogenase
(catalytic subunit)
pyrF 1.21 orotidine 5'-phosphate
decarboxylase
pyrE 1.08 orotate phosphoribosyl
transferase
rnc 2.41
1.64(±0.03) - ribonuclease III, cleaves both 5'-
and 3'-sites of the small
cytoplasmic RNA precursor
ǂ ǂ ǂ ǂ ylqB smc
1.42 chromosome condensation and
segregation SMC ATPase
ftsY 1.08 signal recognition particle
rplJ 1.69 3.42(±0.30) 50S ribosomal protein L10
ǂ ǂ
rplL 1.66 50S ribosomal protein L7/L12
sdpA 1.26 1.08(±0.22) + sporulation delay protein-export
of killing factor
ǂ
sdpB 2.2 1.68(±0.08) Sporulation delay protein-
exporter of killing factor SpbC
sdpC 0.61 killing factor SdpC
spoIIE† ND + serine phosphatease ǂ
tapA 0.96 2.65(±0.06) + lipoprotein for biofilm formation
ǂǂ
sipW 2.12 1.42(±0.14) type I signal peptidase
tasA 1.50 2.65(±0.17) major biofilm matrix component
tig 1.68 1.11(±0.01) - trigger factor (prolyl isomerase),
catalyze in vitro protein folding;
essential for growth under
starvation conditions
ǂ ǂ ǂ
tsf 1.69 1.31(±0.53) - Ts elongation factor
ǂ ǂ pyrH 1.52 1.74(±0.15) uridylate kinase
frr 1.55 1.11(±0.08) ribosome recycling factor
ydjM 2.33 + hypothetical protein ǂ ǂ ǂ
ydjN 1.48 hypothetical protein
yhdN ND - aldo/keto reductase ǂ ǂ ǂ
plsC 1.49 1.05(±0.14) - 1-acyl-sn-glycerol-3-phosphate
acyltransferase (lipid
metabolism)
yhfE 1.63 3.65(±0.04) + putative endoglucanase ǂ
ǂ
yhfF 1.40 2.18(±0.09) hypothetical protein
yjcM 1.48 hypothetical protein ǂ ǂ ǂ
ylxM 1.32 - hypothetical, DNA-binding
protein
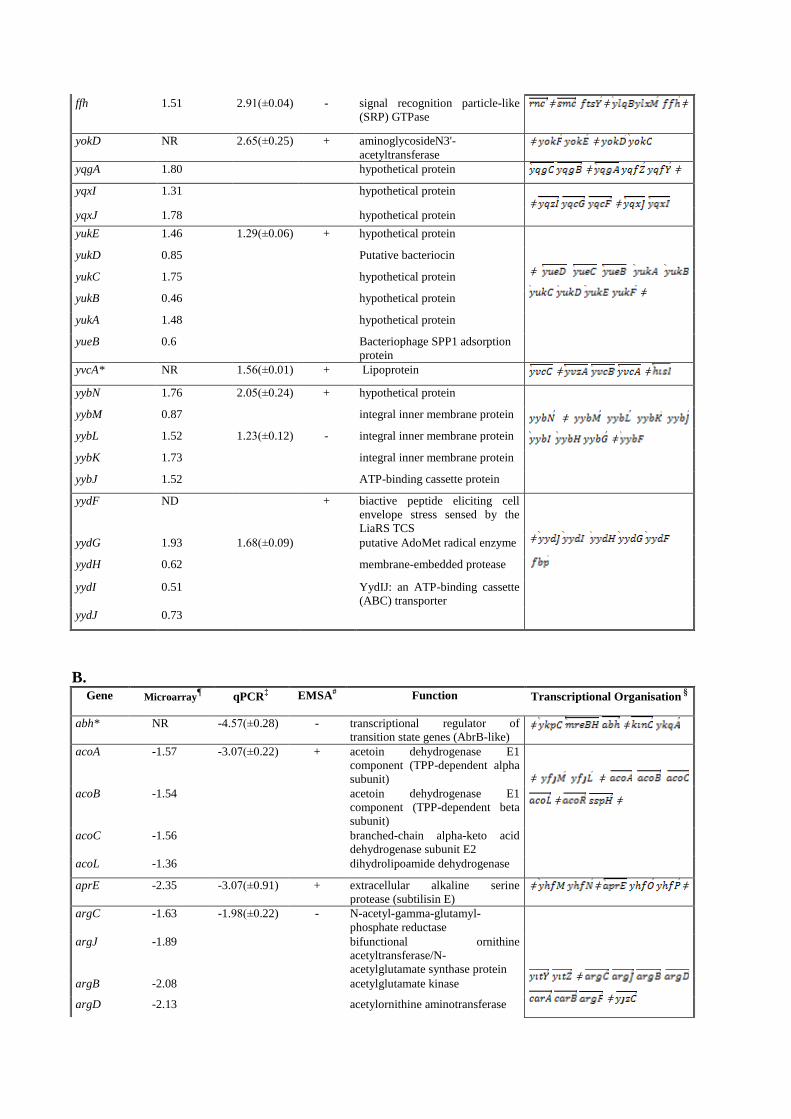
ffh 1.51 2.91(±0.04) - signal recognition particle-like
(SRP) GTPase ǂ ǂ ǂ
yokD NR 2.65(±0.25) + aminoglycosideN3'-
acetyltransferase ǂ ǂ
yqgA 1.80 hypothetical protein ǂ ǂ
yqxI 1.31 hypothetical protein ǂ ǂ
yqxJ 1.78 hypothetical protein
yukE 1.46 1.29(±0.06) + hypothetical protein
ǂ
ǂ
yukD 0.85 Putative bacteriocin
yukC 1.75 hypothetical protein
yukB 0.46 hypothetical protein
yukA 1.48 hypothetical protein
yueB 0.6 Bacteriophage SPP1 adsorption
protein
yvcA* NR 1.56(±0.01) + Lipoprotein ǂ ǂ
yybN 1.76 2.05(±0.24) + hypothetical protein
ǂ
ǂ
yybM 0.87 integral inner membrane protein
yybL 1.52 1.23(±0.12) - integral inner membrane protein
yybK 1.73 integral inner membrane protein
yybJ 1.52 ATP-binding cassette protein
yydF ND + biactive peptide eliciting cell
envelope stress sensed by the
LiaRS TCS
ǂ
yydG 1.93 1.68(±0.09) putative AdoMet radical enzyme
yydH 0.62 membrane-embedded protease
yydI 0.51 YydIJ: an ATP-binding cassette
(ABC) transporter
yydJ 0.73
B. Gene
Microarray
¶ qPCR
‡ EMSA# Function Transcriptional Organisation §
abh* NR -4.57(±0.28) - transcriptional regulator of
transition state genes (AbrB-like) ǂ ǂ
acoA -1.57 -3.07(±0.22) + acetoin dehydrogenase E1
component (TPP-dependent alpha
subunit)
ǂ ǂ
ǂ ǂ
acoB -1.54 acetoin dehydrogenase E1
component (TPP-dependent beta
subunit)
acoC -1.56 branched-chain alpha-keto acid
dehydrogenase subunit E2
acoL -1.36 dihydrolipoamide dehydrogenase
aprE -2.35 -3.07(±0.91) + extracellular alkaline serine
protease (subtilisin E) ǂ ǂ ǂ
argC -1.63 -1.98(±0.22) - N-acetyl-gamma-glutamyl-
phosphate reductase
ǂ
ǂ
argJ -1.89 bifunctional ornithine
acetyltransferase/N-
acetylglutamate synthase protein
argB -2.08 acetylglutamate kinase
argD -2.13 acetylornithine aminotransferase
carA -2.22 carbamoyl phosphate synthase
small subunit
carB -2.15 carbamoyl phosphate synthase
large subunit
argF -2.16 ornithine carbamoyltransferase
argG -1.68 + argininosuccinate synthase ǂ ǂ ǂ ǂ
ǂ argH -2.04 argininosuccinate lyase
bceA ND -2.68(±0.32) + bacitracin ABC efflux transporter
ATP-binding protein
ǂ ǂ
ǂ bceB ND -3.77(±0.06) bacitracin export permease protein
BceB
citA -1.50 -2.20(±0.30) - citrate synthase 1 ǂ ǂ ǂ ǂ
citB -1.38 -2.21(±0.21) - aconitate hydratase ǂ ǂ
glnR -1.31 -0.73(±0.10) + transcriptional regulator (nitrogen
metabolism)
ǂ ǂ
glnA -1.19 glutamine synthase
gltA -1.31 -2.04(±0) + glutamate synthase (large subunit) ǂ ǂ ǂ
gltB -1.45 Glutamate synthase [NADPH]
small chain
ispA -2.01 -1.68(±0.22) + intracellular serine protease ǂ ǂ ǂ
liaI -1.65 -1(±0.05) + putative transmembrane protein
ǂ
ǂ
liaH -1.78 similar to phage-shock protein
A(PspA) of E. coli
liaG -1.31 putative membrane-anchored
hypothetical protein
liaF -1.05 membrane protein
liaS -1.02 LiaRS: two component regulatory
system
liaR -0.85
msmR -1.48 -1.43(±0.27) + transcriptional regulator (LacI
family) ǂ
ǂ msmE -1.51 multiple sugar-binding lipoprotein
pbpE ND -3.44(±1.13) + penicillin-binding protein 4*
(spore cortex)
ǂ ǂ ǂ racX ND amino acid racemase
rapI*† -1.84(NR)
+ response regulator aspartate
phosphatase
ǂ ǂ
phrI* NR phosphatase RapI regulator
sigW ND -4.31(±0.01) - RNA polymerase sigma factor
sigW
trnSL-Gln2 ǂ ǂ
ybbM ND hypothetical protein
trpE -1.9 -1.26(±0.05) - anthranilate synthase component I
ǂ
trpD -2.1 anthranilate
phosphoribosyltransferase
trpC -2.62 indole-3-glycerol-phosphate
synthase
trpF -2.22 N-(5'-phosphoribosyl) anthranilate
isomerase
trpB -2.00 tryptophan synthase subunit beta
trpA -1.04 tryptophan synthase subunit alpha
ybfO -1.76 -1.08(±0.06) + hypothetical protein; similar to
erythromycin esterase
ǂ ǂ ǂ
ybyB -1.48 hypothetical protein ǂ ǂ ǂ
yceC -1.00 -0.88(±0.06) + putative stress adaptation protein
yceD -1.44 putative stress adaptation protein ǂ ǂ
ǂ
yceE -1.39 putative stress adaptation protein
yceF -1.73 -1.38(±0.08) putative stress adaptation
transporter
yceH -1.43 putative stress adaptation protein
yneN -1.41 -1.04(±0.19) + putative membrane-bound protein
with a thioredoxin-like domain ǂ ǂ
yotH -1.80 hypothetical protein
yuaF -1.72 -1.12(±0.09) + membreane integrity integral inner
membrane protein
ǂ ǂ
ǂ
floT -1.46 putative flotillin-like protein
yuaI -1.62 putative acetyl-transferase
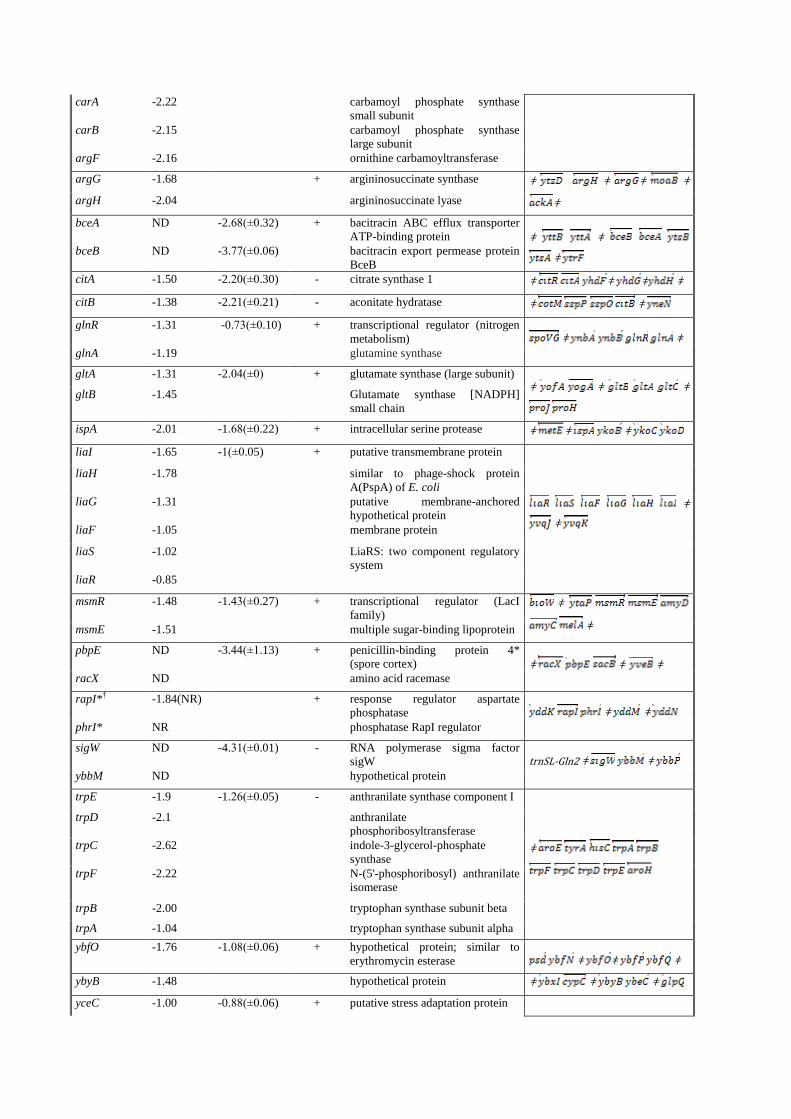
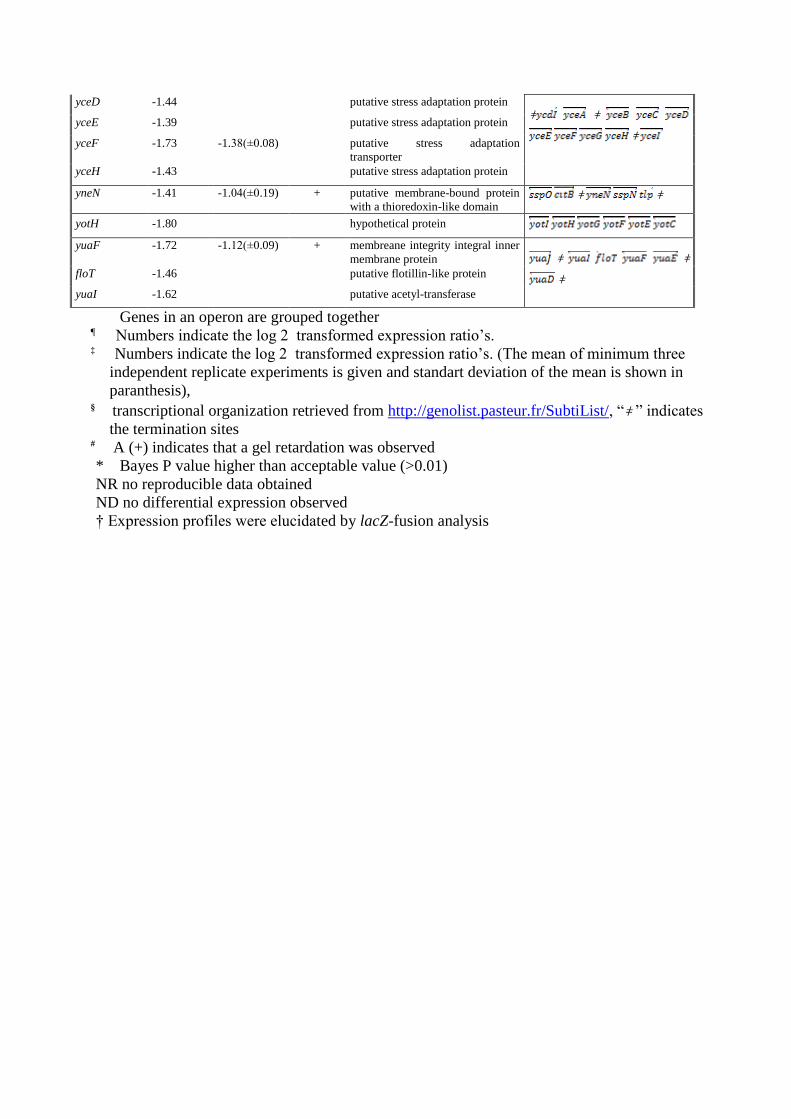
Genes in an operon are grouped together ¶ Numbers indicate the log 2 transformed expression ratio’s. ‡ Numbers indicate the log 2 transformed expression ratio’s. (The mean of minimum three
independent replicate experiments is given and standart deviation of the mean is shown in
paranthesis), § transcriptional organization retrieved from http://genolist.pasteur.fr/SubtiList/, “ǂ ” indicates
the termination sites # A (+) indicates that a gel retardation was observed
* Bayes P value higher than acceptable value (>0.01)
NR no reproducible data obtained
ND no differential expression observed
† Expression profiles were elucidated by lacZ-fusion analysis
Supplemantary Figures
Fig.S1
Fig.S2
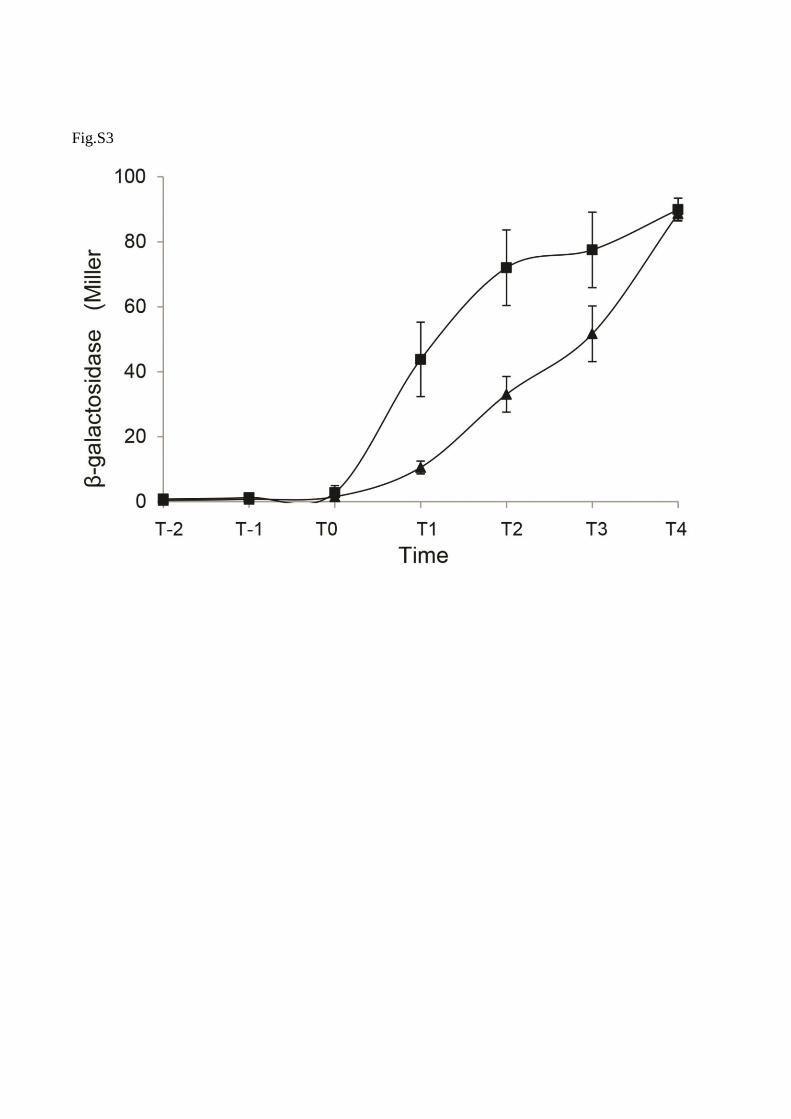
Fig.S3

Supplementary Methods:
Detailed information about the results of EMSA analysis performed in the presence of both
LutR and SinR
It has been previously suggested that LutR and SinR seem to act cooperatively to repress lutABC
(Chai et al., 2009). To analyze the overlap between LutR and SinR DNA targets, we performed
EMSA with the addition of either or both proteins to given promoters. As a first step, we analyzed
the interaction in the case of the lutA promoter. As shown in Fig. 5, an apparent mobility shift was
observed only at the highest LutR concentration tested (4 µM): neither 2 nor 3 µM LutR alone was
able to produce a detectable mobility shift, but the presence of 3 µM SinR, which by itself had not
shifted the DNA probe, greatly stimulated the binding capacity of LutR, almost all of the DNA probe
was shifted even at 2 µM LutR. Consistently, only 4 µM SinR alone was able to produced a
detectable shifted DNA complex, but its binding was stimulated by the presence of 3 µM LutR,
almost all of DNA probe was shifted at 2 µM SinR, which itself had not shifted the DNA probe. On
the other hand, neither LutR nor SinR alone, nor a mixture of them was able to shift the control DNA
probe (the regulatory region of the unrelated ywhB), at all of the protein concentrations tested. These
results elucidated not only the specific interactions of SinR and LutR with the PlutA but also the
cooperative nature of their interaction. Besides the lutABC operon, tapA operon and aprE gene were
previously known to be under direct control of SinR and were found in our study to be directly
regulated by LutR. Under light of these findings, we then wondered whether SinR would interact
with the regulatory regions of all or only some LutR-target genes and it could stimulate the LutR
binding capacity for those genes. For this, EMSA analysis described above were performed with the
regulatory regions of all of the LutR-target genes identified in our study. Interestingly, as shown in
Fig. 5, SinR was capable to interact with the regulatory regions of all of the LutR-target genes tested,
but they exhibited variations in the nature of their interactions. Both LutR and SinR apparently
stimulate each other’s binding to the regulatory regions of lip and bslA. As an example, only 4 µM
SinR alone could shift the regulatory regions of lip, but in the presence of 3 µM LutR which itself
had not shifted the DNA probe, almost all of the DNA probe was shifted even at 2 µM SinR, which
itself had not shifted the DNA probe SinR. Similarly, in the presence of 3 µM SinR, even 2 µM
LutR, which itself had not shifted the DNA probe, shifted all of the DNA probe. Besides to
cooperative interactions, they exhibited additive or simultaneous binding to the regulatory regions of
acoA, aprE, atpI, bacA, bceA, czcD, cwlO, ftsE, glnR, gltA, hepS, ispA, liaA, msmR, mraY, pbpE,
ppsA, pyrB, pyrR, rapI, sdpA, tapA, ybfO, yceC, ydjM, yneN, yhfE, ywfH, yuaF, yukE, yvcA, yokD,
yydF, yybN. In these cases, both LutR and SinR alone was able to produce a mobility shift with the

regulatory regions. Addition of increased amount of LutR in the presence of fixed amount of SinR or
addition of increased amount of SinR in the presence of fixed amount of LutR resulted in an
additional retardation in the electrophoretic mobility of SinR-bound or LutR bound-DNA probes,
suggesting a co-binding to these regions without promoting each other`s binding activity.
Interestingly, in the case of the spoIIE gene, LutR apparently stimulated the binding capacity of SinR
while SinR did not affect the binding capacity of LutR which itself had produced a mobility shift at
all concentrations tested in a concentration dependent manner. SinR alone was not capable to
produce a mobility shift at 2 or 3 µM protein concentrations even it produced at 4 µM concentration,
but in the presence of 3 µM LutR, resulted in an additional retardation in the electrophoretic mobility
of LutR bound-DNA probes. Specific inhibition of sporulation stage II genes: spoIIA, spoIIE and
spoIIG by SinR had been reported previously (Mandic-Mulec et al., 1995) that provides an in vivo
support for the in vitro interaction of SinR with the spoIIE regulatory region detected in this study.
Mandic-Mulec, I., Doukhan, L. & Smith, I. (1995). The Bacillus subtilis SinR protein is a
repressor of the key sporulation gene spo0A. J Bacteriol 177, 4619-4627.
Detailed information on the DNA-microarray analysis
Total RNA isolat ed from all samples using the Macaloid/Roche protocol (van Hijum et al., 2005).
10-20 µg of total RNA was first reverse transcribed into cDNA with the Superscript III Reverse
Transcriptase kit (Invitrogen) and labeled with a Cy3- or Cy5-monoreactive dye (GE Healthcare,
Amersham). Purification of CyDye labeled cDNA was executed using NucleoSpin Extract II
columns (Machery Nagel). Concentrations of Cy3 and Cy5 were expected to be at least 0.5 pmol/µl
in a total volume of 50 μl. The labeled and purified cDNA samples were hybridized to
oligonucleotide microarrays in Ambion Slidehyb #1 buffer (Ambion Europe Ltd) at 48 °C for more
than 16 hours. The arrays were constructed as described elsewhere (van Hijum et al., 2003a).
Furthermore, slide spotting, slide treatment after spotting and also slide quality control were done as
before (van Hijum et al., 2005). Following hybridization, slides were washed for 5 min in 2x SSC
with 0.5% SDS, 2 times 5 min in 1x SSC with 0.25% SDS, 5 min in 1x SSC 0.1% SDS for the
removal of hybridization buffer and unbound cDNAs. Finally slides were dried by centrifugation (2
min at 2000 rpm) and scanning of the slides was performed in a GenePix 4200AL Microarray
Scanner (Axon Instruments, CA, USA). Fluorescent signals were quantified using ArrayPro 4.5
(Media Cybernetics Inc., Silver Spring, MD). Following the acquisition of expression data,

expression levels were processed and normalized with Micro-Prep which is a helpful software tool
that enables scientists to handle microarray data and transform raw data into processeable Excel data
through avoidance of inconsistencies (van Hijum et al., 2003b). Then the expression difference ratio
of the lutR mutant and wild type PY79 strains during stationary growth were further processed using
the Cyber-T tool, which is a software program that works on a t-test variant combined with a
Bayesian statistical framework (Baldi & Long, 2001). Parameter use in the Cyber-T tool was
followed as recommended, giving 2 as ‘minimum non-zero replicates’, a ‘sliding window’ of 101
and a confidence value of 10. Accepted Cyber-T (Bayes) p value was set to 0.01.
van Hijum, S. A., de Jong, A., Baerends, R. J., Karsens, H. A., Kramer NE, Larsen, R., den
Hengst, C. D., Albers, C. J., Kok, J. & Kuipers, O. P. (2005). A generally applicable validation
scheme for the assessment of factors involved in reproducibility and quality of DNA-microarray
data. BMC Genomics 6, 77.
van Hijum, S. A., de Jong, A., Buist, G., Kok, J. & Kuipers, O. P. (2003a). UniFrag and
GenomePrimer: selection of primers for genome-wide production of unique
amplicons. Bioinformatics 19, 1580-1582.
van Hijum, S. A., Garcia de la Nava, J., Trelles, O., Kok, J. & Kuipers, O. P. (2003b). MicroPreP: a cDNA microarray data pre-processing framework. Appl Bioinformatics 2, 241-244.

Primers used in this study
abh forward 5’ gat gaa tta ggc cgc att 3’
abh reverse 5’ ggc ttg aat ttc ttc gag 3’
abh EMSA forward 5’ gtc aat ccc gat ttc agt tga 3’
abh EMSA reverse 5’ gac aat gcg gcc taa ttc atc 3’
acoA EMSA.forward 5’ggc ggt att gga tat gtc aaa 3’
acoA EMSA.reverse 5’gta cat cca cag cgc ttt ttc 3’
acoA forward 5’ ctg gag atc agg ggc ttt 3’
acoE reverse 5’ aca gcc ttt ggc gat aca 3’
acpA EMSA forward 5’ cgg ttt aac caa atc ttc tgc 3'
acpA EMSA reverse 5’tac gat gat ttt cgt tac acg 3’
aprE EMSA forward 5’atc cat tgt tct cac gga agc 3’
aprE EMSA reverse 5’cgc aaa caa caa gct gat cca 3’
aprE Forward 5’ gcg ttc agc aac atg tct 3’
aprE Reverse 5’cac ata tgc aac gct cgg 3’
aprX EMSA forward 5’ gcc ggatcc ctg tct cac aat aat cgt tct 3’
aprX EMSA Reverse 5’ cgg gaa ttc cca atc tag ctt gtg agc att 3’
aprX F 5’ tgg act cct tgc ttc ttg 3’
aprX R 5’ tgc tga agg agt aac ctc 3’
argC EMSA forward 5’ggc ttt ctc gct gac ttt ttc 3’
argC EMSA reverse 5’tcc tgt agc acc tac aat tcc 3'
argC forward 5’ agc gag ggt tat cct cat 3’
argC reverse 5’ cag atc acc tga cag atc 3’
argG EMSA forward 5’caa tca tgt cga gag caa ctg 3’
argG EMSA reverse 5’acc tcc tga gta tgc taa tac 3’
argG forward 5’ gag ggc aaa gat ttg gca 3’

argG reverse 5’ acg aac ctg gtc att tcc 3’
atpA reverse 5’acc aac agg aac ctc cat 3’
atpA forward 5’gtc atc caa gtc ggt gac 3’
atpE reverse 5’gaa tgc gat aac gac agc
atpE forward 5’gca gct gcg att gca att 3’
atpI EMSA forward 5’- gcc ggatcc tca tgt gtc ttg tag cag cgc-3’
atpI EMSA reverse 5’- cgg gaattc cat tct tct gac gag cag taa-3’
atpI forward 5’ ttg gca gtg tat gta ctg ggt-3’
atpI reverse 5’ cgg gaa ttc cat tct tct gac gag cag taa-3’
bioW EMSA forward 5’aca ggg agc tgc tga ttg cta 3’
bioW EMSA reverse 5’cat tga agc cct cat tct gac 3’
bioW forward 5’gtc aga atg agg gct tca 3’
bioW reverse 5’ cac agg caa tgg ctg aat 3’
citA EMSA forward 5’gcc ttc atc agt cag ctg gat 3’
citA EMSA reverse 5’aca tgt aat tcc ctt taa tcc 3’
citA forward 5’ agc ttt gaa gaa gcg gct 3’
citA reverse 5’cgg atg gaa tgt gta cgt 3’
citB EMSA forward 5’cgt ttt caa tat agc cgg cgc 3’
citB EMSA reverse 5’cgc ttg gaa aac gtc ttt tga 3’
citB forward 5’ aag gtt tcg aag ctt cct 3’
citB reverse 5’ cag tga agc cag atc tac 3’
czcD forward 5’ tct gat gca ggc cat atg ctg 3’
czcD reverse 5’ aag cat gcc ggt tgt tgc tac 3’
czcD EMSA forward 5’ttg tag ttg aat act acc ctc 3’
czcD EMSA reverse 5’tgc tcc ttc att atg att gtg 3’
fabD EMSA forward 5’ctg gaa ggc tct gcg ttg tca 3’

fabD EMSA reverse 5’tga tcc ctg acc cgg gaa taa 3’
fabHA EMSA forward 5’cgg gct tga ttt gat caa gag 3’
fabHA EMSA reverse 5’acg tcc aac acc aag tat tcc 3’
fabHA forward 5’ gac gag tgg att cgt aca 3’
fabHA reverse 5’ gcc gag ttg ttc ttg aat 3’
fabHB EMSA forward 5’gca gta cat aaa cat gaa cgc 3’
fabHB EMSA reverse 5’ata ggt gcc gat agc tgt aat 3’
fabHB forward 5’ gaa tgg atc gtt cag cgc 3’
fabHB reverse 5’ gct ttc cca gcc gaa ata 3’
ffh EMSA forward 5’tat cag tcg ttg ttg acg tca 3’
ffh EMSA reverse 5’tcg gtc ggc taa tcc ttc aaa 3’
ffh forward 5’ atg atg cgt gag gtc cgt ctt-3’
ffh reverse 5’ aac ttt aat gac ctg ctg gcc-3’
frr forward 5’ tat tac gga gcg cag aca 3’
frr reverse 3’ccg tct ctc ttc tgt tag 3’
ftsE EMSA forward 5’taa aaa cgg ccg cgg caa tat 3’
ftsE EMSA reverse 5’gcc gtt cgg ata ggc ttt ata 3’
ftsE forward 5’ tat gtt gtt ggt ccg agc 3’
ftsE reverse 5’cac ttc aag ggc aaa tgc 3’
glnR forward 5’ cca gcc aga agt gaa gga 3’
glnR reverse 5’ tct cag ttc gtc atc gga 3’
glnR EMSA forward 5’tacatgcctggatacgaggat 3’
glnR EMSA reverse 5’tgg aaa taa agg cat tga gcg 3’
gltA EMSA forward 5’gat att tct ccc ttc acg ctc 3’
gltA EMSA reverse 5’acg gta gag acc ttg agc ttt 3’
gltA forward 5’ atc ggc cta tat gca cac 3’

gltA reverse 5’tcc tac ccc gta acg ttc 3’
hepS EMSA forward 5’gcc gtt gag gac gga gtg aat 3’
hepS EMSA reverse 5’gtt cag att ggc taa agt tcc 3’
hepS forward 5’ gcg aag cat att tct gcg 3’
hepS reverse 5’ gag aag agt caa ttg gcg 3’
ispA forward 5’ aag gcg cca gaa atg tgg 3’
ispA reverse 5’ agc tgc aat tgt tcc ggc 3’
ispA EMSA forward 5’act gtt gca tta tgt agg gcg 3’
ispA EMSA reverse 5’cac ata cgg gat caa gcg gat 3’
lip EMSA forward 5’aat ggt gtc gtc aca cca aac 3’
lip EMSA reverse 5’tac aag tgc aat gat cct tct 3’
lip forward 5’ gtc gtt atg gtt cac ggt 3’
lip reverse 5’ cat gct gtg agc gac aat 3’
murE.EMSA.forward 5’gat acg cgt gta ttt gac tga3’
murE.EMSA.reverse 5’ggt tgt taa gta tgt aag cag 3’
murE.forward 5’ gtc gat gtg aat gtt cct 3’
murE.reverse 5’ agg aag tgt ttc gtc tcc3’
mraY forward 5’gga ccg aaa tca cat cag 3’
mraY reverse 5’gcg ctt cat gac aac ctt 3’
msmR EMSA forward 5’aat gtc tca aca ctg att ggc 3’
msmR EMSA reverse 5’gga aac ttt agc ttt caa ggc 3’
msmR forward 5’ ctt tct gtt gcg ggc gaa 3’
msmR reverse 5’ ccc ttt ccg aat gga aga 3’
ndk forward 5’ gtc caa cgt cag ctc att 3’
ndk reverse 5’ cag ctg tct cgt cac ttc 3’
pbpE EMSA forward 5’gat tgc aac tgg tct att ttc 3’

pbpE EMSA reverse 3’taa cgt ctg aag atg ctt tct 3’
pbpE forward 5’- cag ttt aac ggg acg gtt-3’
pbpE reverse 5’-ctg ata cgg aaa acc ggg-3’
pksD EMSA forward 5’aac cat cgc tcc cac atc tag 3’
pksD EMSA reverse 5’ata gta ttg gga acc ttg ccc 3’
pksD forward 5’ cga atc ggc aca tcc att 3’
pksD reverse 5’ cca gac tga tcc caa tac 3’
pksG EMSA forward 5’gaa tta aag gcg ctc cgt gca 3’
pksG EMSA reverse 5’att cat cgc ttc tat tcc ggc 3’
pksG forward 5’ ctt gat gtc atg gag ctg 3’
pksG reverse 5’ gtt gcg gtt gag acc taa 3’
pksJ forward 5’ ccg gct cca tta gcc gtt 3’
pksJ reverse 5’ tcc cgt act gcc tga agt 3’
pksL EMSA forward 5’agc gcc gct ttg tcc cat ttt 3’
pksL EMSA reverse 5’ctt ttt cac gtt aga cct cca 3’
pksL forward 5’ aag gct gac atg cac gca 3’
pksL reverse 5’ caa atg att ggc acc cac 3’
ppsA EMSA forward 5’caa tta gaa gaa tga tgc aca 3’
ppsA EMSA reverse 5’ttg ggc atg ggt taa aga ata 3’
ppsB forward 5’ aat cta tgc gtc gac tcg 3’
ppsB reverse 5’agc atc aat cag cgt ctg 3’
pyrAA forward 5’tct tac tgc gga cag atc 3’
pyrAA reverse 5’ atc aat tcc ctg gag tcc 3’
pyrB EMSA forward 5’cag tta tcc ttg tca tcg gtg 3’
pyrB EMSA reverse 5’agt gct aag ttc act cat cgt 3’
pyrB forward 5’ttc gaa ccg agc acg aga 3’

pyrB reverse 5’ ctg gct gac aag ctc ttc 3’
pyrH forward 5’atc gct gag ctt gaa gtc gaa 3’
pyrH reverse 5’gga tgt ttg cac tct gga 3’
pyrP forward 5’gtc gga atg agt cct gct 3’
pyrP reverse 5’ caa taa gga aat cag ccc 3’
pyrR EMSA forward 5’ctt taa tgg cca acc gct tca 3’
pyrR EMSA reverse 5’cag cgc ccg tct aat tgc ctg 3’
pyrR forward 5’agg att gct cac gaa atg 3’
pyrR reverse 5’ tac cgg aat atc tgc acc 3’
rapI EMSA forward 5-gca gtc tgg att gtt tgg gta-3’
rapI reverse 5’-cgt gac taa gtc gta cgg aat-3’
rapI BamHI reverse 5’ cgg gga tcc ttc agc tat tcg ata agc 3’
rapI HindIII forward 5’ gcc aag ctt ttg cgg ggt gtt ttc tta 3’
rnc EMSA forward 5’taa cga aaa tca tcg tag atc 3’
rnc EMSA reverse 5’ttt atc ttt ata atg tga gtg 3’
rnc forward 5’-caa gaa cgg att tcg gtt cac-3’
rnc reverse 5’- cct ttc att atc ttc ata cgg-3’
rplJ forward 5’-cgc gga ctt aac gtt tct gaa-3’
rplJ reverse 5’-aag ctc agc ttg ttc aac cgc-3’
sigW forward 5’- gcg gac atc gta gat att-3’
sigW reverse 5’-aga ata cat ggt caa gcc-3’
sipW forward 5’- tca gtt ctg tca ggt tcg atg-3’
sipW reverse 5’-aac aat tct gtg ggt gac cgc-3’
spoIIE EMSA forward 5’tac ggt tca tac ccg tga ggt 3’
spoIIE EMSA reverse 5’tgg ccc gtt cac tct tct ttc 3’
tasA forward 5’-aag ccg gga gat aag ttg aca -3’

tasA reverse 5’-ctg gct gag gaa atc ttc tgg-3’
tig EMSA forward 5’tta cga gat gca cgg gat tga 3’
tig EMSA reverse 5’cgt taa aac gcc ttc gtt gcc 3’
tig forward 5’- caa gtt tca att cct gga ttc-3’
tig reverse 5’- agg gta ttc tac agg aag agg-3’
trkA(czcO) EMSA forward 5’tgg tct atc aca agc gga ttg 3’
trkA(czcO) EMSA reverse 5’ttg acc agc ccc gat tac tat 3’
trkA(czcO) forward 5’ ata gta atc ggg gct ggt 3’
trkA(czcO) reverse 5’ tcc ttc aag atg cat tcc 3’
trpE EMSA forward 5’gac aga tgt ccc tca gga tca 3’
trpE EMSA reverse 5’gct gtc ctc taa aaa tgc gga 3’
trpE forward 5’ gag aag ctt gac agg gag 3’
trpE reverse 5’ agg aat gcc aag ctc agg 3’
tsf EMSA.forward 5’cat ccc tat cat cgg tat cgt 3’
tsf EMSA reverse 5’cat gcc cgc gcc agt ttt ttc 3’
tsf forward 5’ gcg tta act gaa act gac gga 3’
tsf reverse 5’ tgc aag aag gtg gtc agc taa 3’
ybfO EMSA forward 5’cga gaa gtg tgc tgg ccg atc 3’
ybfO EMSA reverse 5’cag cgg cag ggt cat tct cac 3’
ybfO forward 5’ gaa cga cat gca cag cct 3’
ybfO reverse 5’ atc agg aaa gcc gga ttc 3’
yceC EMSA forward 5’tgc tta aat gaa tca aaa ggc 3’
yceC EMSA reverse 5’atc aat tcg ttg acc ttt ttc 3’
yceC forward 5’ aaa ttg atg gtc ggt ctc 3’
yceC reverse 5’ cag gtt gtc gcc tgt atg 3’
yceF forward 5’ ggt ttg atc ggt tcc ctt 3’

yceF reverse 5’cac ctt gat cca cca gaa 3’
ydjM EMSA forward 5’aaa ggg tct cgc aca ctc ttt 3’
ydjM EMSA reverse 5’tcc tac taa gat aaa agc ggc 3’
yhdN EMSA forward 5’gct gtg atc ata gga gtg aat 3’
yhdN EMSA reverse 5’ggc ttc tat tcc tgt atc tgc 3’
yhdO forward 5’ atc gcg tgt aca cat tcc 3’
yhdO reverse 5’ cgg cgt ttt aat act gct 3’
yhfE reverse 5'-tcc cga ggc ggt ttc aat ttg-3’
yhfE forward 5’-gaa acg gtt cgg aac cac aag -3’
yhfE EMSA forward 5’ ggg tca tca gaa tat ttc tga 3’
yhfE EMSA reverse 5’aat gag ctc cat cgt ttt acg 3’
yhfF forward 5’- ctg att cag cag atc ctt gcc-3’
yhfF reverse 5’-aat tcg gtc tcc att gcc ctc-3’
ylpC EMSA forward 5’aaa caa agc gga atg ccg gaa 3’
ylpC EMSA reverse 5’ttc ctg gcg ttc tct ctt att 3’
ylpC forward 5’ gat gaa gaa cta gcg ggt 3’
ylpC reverse 5’ aat gga tat cgc ctg atc 3’
ylxM EMSA forward 5’ tgt ccg taa ttg att tta ccg 3’
ylxM EMSA reverse 5’ cag ata att cat tct cgt tgt 3’
yneN EMSA forward 5’tgc gga tac act cgg ctt aac 3’
yneN EMSA reverse 5’cat gat aag cag gat ccc tgc 3’
yneN forward 5’ gtc ggt tat acg gga tgg 3’
yneN reverse 5’ ttg cag ctt ttc cat cgc 3’
yokD forward 5’aat ggt gga gct gtt gc 3’
yokD reverse 5’ ggc tgg cat act ttc tct 3’
yokD EMSA reverse 5’tgg aaa agt tgt act ttc aac-3’

yokD EMSA forward 5’gta gct ata cta aga gag caa-3’
yqxM forward 5’ gcc gca ata tgc tta caa-3’
yqxM reverse 5’ctt aag ttt ctc acc tgt-3’
yqxM EMSA forward 5’gtg cca aag acg aga aga gat-3’
yqxM EMSA reverse 5’ctt cgc ctt ttg ctg att gtg-3’
ytsC forward 5’- ggc gaa ttc gtc agt att-3’
ytsC reverse 5’-gat cga taa agg cag aag-3’
ytsC EMSA forward 5’tcc aaa aga tgt acc gcg tat-3’
ytsC EMSA reverse 5’ctg ttt att cag ctt gtt tcc-3’
ytsD forward 5’ gtc acg ctg cag tat gat-3’
ytsD reverse 5’ tac ccc gat cgc taa tga 3’
yuaB forward 5’- gca cct aca gct tct ttc-3’
yuaB reverse 5’-aaa tcc gct tgg caa tgt-3’
yuaB forward EMSA 5’-gat cag ctg gaa agc tct-3’
yuaB reverse EMSA 5’-ccc gag act taa tgc act-3’
yuaF EMSA forward 5’ aat ggt caa gaa ctt ccc gta 3’
yuaF EMSA reverse 5’ cat tgt ttg tat agg tac tcc 3’
yuaF forward 5’ ccg aca tta gtg ctc tca 3’
yuaF reverse 5’tcc tct gag atc atc ttc 3’
yukE EMSA forward 5’ttc taa gaa agt cat cgg agg 3’
yukE EMSA reverse 5’tcc tgc cat att cct cat tac 3’
yukE forward 5’atg gca cag gag gta atg 3’
yukE reverse 5 aag gtt tga gct gct cgt 3’
yvaW (sdpA) BamHI forward 5’ gcc gga tcc ttg atg cca aca ttg ccg aga
3'
yvaW (sdpA) EcoRI reverse 5’ cgg gaa ttc aat gtt ttc ttc tgt agg gct 3’

yvaW (sdpA) forward 5’ agc aat att tca cct cag aa 3’
yvaX (sdpB) forward 5’-aga agt tta ctt ggt ttc tca-3’
yvaX (sdpB) reverse 5’-agc ggt agg gat ata gac att-3’
yvcA reverse 5’ tgt ctt agg ttc att cgc-3’
yvcA forward 5’ gaa gag gaa cca gga tat-3’
yvcA EMSA forward 5’gct tgt cct gct cag aga-3’
yvcA EMSA reverse 5’ act gca tcc tcc tgt cag-3’
yvcE(cwlO) reverse 5’ gtt gct tgt atc aag cgc 3’
yvcE(cwlO) forward 5’ gca tcg gcg gaa aca tta 3’
yvcE EMSA forward 5’-cac gct taa gaa aaa tga caa-3’
yvcE EMSA reverse 3’-agc caa acc aag tgt aat taa-3’
yveN EMSA forward 5’aaa cct cat tca ttt gtc cgg 3’
yveN EMSA reverse 5’atc aac agt cgc gca aaa caa 3’
yveN forward 5’- aag ctg ccg tat gtg gat gag-3’
yveN reverse 5’- gtg cgc tgt gtc cag cac ctt-3’
yveO Forward 5’-tgc gat gat gcg tca aca-3’
yveO Reverse 5’-aac cac ctg ata gtg tcg gtg-3’
yvfA EMSA forward 5’att cta caa tca cta cgg ctt 3’
yvfA EMSA reverse 5’cgt gag att cgc gct gaa att 3’
yvfA forward 5’ ctc acg gct ttt ctc ttg tct 3’
yvfA reverse 5’ ata cgc att tgc ttt ctc ccg 3’
yvfE forward 5’gaa gaa cag ctg gca gaa cga 3’
yvfE reverse 5’ cat att cca cgt atc agg ctc 3’
yvfF forward 5’ cgc tgg aat cca gac aat ttc 3’
yvfF reverse 5’cgt ccg ctt gag gtt gtc ttt 3’
yvfI BamHI reverse 5’ cgg gga tcc aat atc ccg aaa gca cat 3’

yvfI HindIII forward 5’ gcc aag ctt atg aaa cag gga gaa ggc 3’
yvfqI EMSA forward 5’tat gcc gct aat gct ttc ggc 3’
yvqI EMSA reverse 5’gac gga gat ccc aaa tac aat 3’
yvqI forward 5’ atc ggg agc ttg atg aca 3’
yvqI reverse 5’ aga agc cgg ttc agg atc 3’
ywcD EMSA forward 5’cat gcc gac ata taa agt ctt 3’
ywcD EMSA reverse 5’cac aat ggt tgt gaa aac ccc 3’
ywcD forward 5’ gtt gcc gct tgg att ttg 3’
ywcD reverse 5’ ctg gcc gac aag aat gat 3’
ywfB EMSA reverse 5' gat cgc gga tcc tta tgc gta ctc act gct tgt 3'
ywfB EMSA forward 5' gct atg cag ctg tcg gat 3'
ywfH EMSA forward 5’ ccc ttg act ggc tcc cat aac 3’
ywfH EMSA reverse 5’ agc cac ggt tta atc ggc cgc 3’
ywfH HindIII Forward 5’ gca agc ttt ttt ccc tcg tca tta att 3’
ywfH BamHI Reverse 5’ cat ccc att cat cat act gtt tgt 3’
yybL EMSA forward 5’ata aca gcc ttt ata ctt tta 3’
yybL EMSA reverse 5’aca taa aat taa tag tag tct 3’
yybL forward 5’ ttc aac cag aac cac act 3’
yybL reverse 5’ acg ctt aac cat cct cat 3’
yybN forward 5’ gta cct tat ggc tat ggt 3’
yybN reverse 5’ tcc ctg gct att tgc atg 3’
yybN EMSA reverse 5’ tat gcc taa agc agc cgc aag 3’
yybN EMSA forward 5’ctc aat tga gca aat cgc cca 3’
yydF EMSA forward 5’atg ggg taa gca aga gct taa 3’
yydF EMSA reverse 5’ttc taa gtt ttt cac agt ctc 3’
yydG forward 5’gag tta gta act gaa ttt gca 3’

yydG reverse 5’act tat agt cag tgc tat cac 3’

Information of the B. subtilis PY79A variant used in the current study, i.e. presence of the
ICEBs1 region.
Previously, common B. subtilis PY79 strain was reported to lack the ICEBs1 region on its
chromosome (Auchtung et al., 2005). However, our microarray results suggested the rapI and phrI
genes described in the ICEBs1 region are highly upregulated in the lutR mutant strain (TEK1).
Subsequently, as a further confirmation, to construct rapI-lacZ transcriptional fusion strain, a 389-
bp-long rapI gene fragment was amplified from the genome of our lab strain PY79 by PCR with the
following specific primers: RapI F (5‘ -GCC AAGCTT TTG CGG GGT GTT TTC TTA- 3’) and
RapI R (5’ -CGG GGATCC TTC AGC TAT TCG ATA AGC- 3’). The resulting PCR product was
cloned into pGEMT vector for sequencing. As shown in Fig. S4, our sequencing results revealed
that our amplified region sequence were identical in sequence with the rapI gene present in B.
subtilis 168 genome. After sequence confirmation, the 389-bp-long rapI gene fragment was
subcloned in to pMUTIN T3 vector rapI-lacZ transcriptional fusion was created in our laboratory
strain PY79 and used in further experiments. As mentioned above this region is missing in other
PY79 strains examined before. This prompted us to examine our PY79 laboratory stock used in the
current study. The PY79 laboratory stock that was used in our study has been obtained in 1985 from
the laboratory of Arnold L. Demain (Fermentation Microbiology Laboratory, Department of Applied
Biological Sciences, Massachusetts Institute of Technology, Cambridge, Massachusetts) gifted by
Philipe Youngman. To describe our PY79 strain, we have characterized the strain using various
diagnostic PCR reactions. To confirm the presence of W23 island in panB and sacA regions in our
stock PY79 strain, we amplified a 396 bp DNA region stretching from 1082 bp to 1478 bp in panB
gene and the 524 bp DNA fragment between 1402 bp and 1926 bp of the sacA gene using PCR
reaction and the resulting fragments were cloned into pGEMT vector for sequencing. The sequenced
sacA and panB regions showed similar sequence to PY79 (Fig. S5 and Fig. S6).
Further, the 17 kb DNA region from ydzA to ydaQ was shown to be absent in the genome of strain
PY79 (Zeigler et al., 2008). Therefore, we tested for the presence of topB gene locus using PCR
reaction (arbitrary selected a gene located within the region between ydzA to ydaQ in B. subtilis 168).
The following primer set was used to amplify the topB gene: topB F (5’-
GTCGACGGCATGACGCTGACG-3’), topB R (5’-CAGACTGCTGAGAACGAGCGG-3’). Our
PCR assays indicated that our laboratory stock PY79 strain similar to 1A147 (PY79 BGSC) likely do
not contain ydzA to ydaQ region since the 480 bp fragment of the tapB gene could not be amplified
from their genomes while this region was successfully amplified from the genome of strain 168
(Fig. S7). Thus, these additional results supported that our stock strain is originated from PY79.

Whether our laboratory stock originates from a stock during the creation of PY79 before ICEBs1
was cured, or the ICEBs1 region originates from another laboratory strain that possesses this region
(i.e. ICEBs1 can be transferred between strains by conjugation) is unknown. Based on these
experiments, we described our laboratory stock strain as PY79(ICEBs1+), thus as PY79 variant that
contains the ICEBs1 region
Auchtung, J. M., Lee, C. A., Monson, R. E., Lehman, A. P. & Grossman, A. D. (2005).
Regulation of a Bacillus subtilis mobile genetic element by intercellular signaling and the global
DNA damage response. Proc Natl Acad Sci USA 102, 12554-12559.

Fig. S4 The DNA sequence chromatogram and blastn analysis of the 389 bp fragment of rapI gene
amplified from our PY79(ICEBs1+) strain. DNA sequencing was performed by using an automated
model 3730 DNA analyzer and BigDye Terminator kit (Applied Bipsystems, Inc). A 389-bp-long
rapI gene fragment was amplified from the genome of our lab strain primers RapI F and RapI R
(see sequence above), and labeled with a box on the chromatogram.
Target reference sequence used for blastn: Bacillus subtilis subsp. subtilis str. 168 complete genome
Sequence ID: emb|AL009126.3|
Query 565 TTGCGGGGTGTTTTCTTAGATAAAGATAAAATTCCGTACGACTTAGTCACGAAAAAGTTA 624
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 547306 TTGCGGGGTGTTTTCTTAGATAAAGATAAAATTCCGTACGACTTAGTCACGAAAAAGTTA 547365
Query 625 AATGAATGGTATACATCAATAAAAAATGATCAAGTTGAGCAAGCCGAGATTATAAAAACA 684
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 547366 AATGAATGGTATACATCAATAAAAAATGATCAAGTTGAGCAAGCCGAGATTATAAAAACA 547425
Query 685 GAAGTAGAGAAAGAATTGTTAAACATGGAAGAAAATCAAGATGCCCTGTTATATTATCAA 744
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 547426 GAAGTAGAGAAAGAATTGTTAAACATGGAAGAAAATCAAGATGCCCTGTTATATTATCAA 547485
Query 745 CTATTAGAATTTAGACATGAGATAATGCTGAGTTATATGAAATCTAAGGAAATAGAAGAT 804
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 547486 CTATTAGAATTTAGACATGAGATAATGCTGAGTTATATGAAATCTAAGGAAATAGAAGAT 547545
Query 805 CTCAATAATGCTTATGAGACTATAAAAGAAATTGAGAAGCAAGGGCAATTAACTGGCATG 864
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 547546 CTCAATAATGCTTATGAGACTATAAAAGAAATTGAGAAGCAAGGGCAATTAACTGGCATG 547605
Query 865 TTGGAATACTATTTTTACTTTTTTAAGGGTATGTACGAGTTTAGGCGTAAAGAATTAATT 924
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 547606 TTGGAATACTATTTTTACTTTTTTAAGGGTATGTACGAGTTTAGGCGTAAAGAATTAATT 547665

Query 925 TCAGCGATAGGTGCTTATCGAATTGCTGAA 954
||||||||| ||||||||||||| ||||||
Sbjct 547666 TCAGCGATAAGTGCTTATCGAATAGCTGAA 547695

Fig. S5. The DNA sequence chromatogram and blastn analysis of the 524 bp long fragment of the
sacA gene amplified from our PY79(ICEBs1+) strain. DNA sequencing was performed using an
automated model 3730 DNA analyzer and BigDye Terminator (Applied Bipsystems, Inc). The 524
bp long sacA gene fragment was amplified from the genome of strain PY79(ICEBs1+) by PCR with
the following specific primers: sacA F (5’-TCACGCCATTCGGGTCATTCA-3’), sacA R (5’-
TCAATGGCGAATGTCGCACAG-3’) as labeled with a box on chromatogram.
Target reference sequence used for blastn: Bacillus subtilis strain PY79, sucrase-6-phosphate
hydrolase (sacA), PTS sucrose-specific enzyme IIBC component (sacP), putative formate/nitrite
transporter (ywcJ), sacPA operon antiterminator (sacT), and hypothetical protein YwcI (ywcI) genes,
complete cds; Sequence ID: gb|EU146093.1|
Query 64 TCACGCCATTCGGGTCATTCAGCAGCCCAACCGGCGGCATGATGTGAAAATGCTGGCGGT 123
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1402 TCACGCCATTCGGGTCATTCAGCAGCCCAACCGGCGGCATGATGTGAAAATGCTGGCGGT 1461
Query 124 GCGGATCGCTATTAGCAATAGGTTCATTTTTCTCCACTTCTTCATAAGCCCGGCGACGAA 183
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1462 GCGGATCGCTATTAGCAATAGGTTCATTTTTCTCCACTTCTTCATAAGCCCGGCGACGAA 1521
Query 184 GCTCCTGATCATGTGCTGTCATTTTTTTCTCTCCTCATCCTCATTAATCTTCATGATAAA 243
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1522 GCTCCTGATCATGTGCTGTCATTTTTTTCTCTCCTCATCCTCATTAATCTTCATGATAAA 1581
Query 244 CGCTGCAATGAATGCTGACACTGCCGCAATCGCCATTCCAATCAGATAGTTGACCAGGTT 303
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1582 CGCTGCAATGAATGCTGACACTGCCGCAATCGCCATTCCAATCAGATAGTTGACCAGGTT 1641

Query 304 GCCAAAGCCAAACGGCGCCGCAATAGCGATCATCGGAATACCCGTTAACCCGTAAGCGTT 363
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1642 GCCAAAGCCAAACGGCGCCGCAATAGCGATCATCGGAATACCCGTTAACCCGTAAGCGTT 1701
Query 364 TGCAGCAACATGTGTAAAGACGACATATGCACCGCCCAATGCGCCTCCGATCATCGCGGC 423
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1702 TGCAGCAACATGTGTAAAGACGACATATGCACCGCCCAATGCGCCTCCGATCATCGCGGC 1761
Query 424 GATAAACGGTTTTCGGTAGCGAAGATTGACTCCGAATATGACTGGCTCAGTAATGCCGAG 483
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1762 GATAAACGGTTTTCGGTAGCGAAGATTGACTCCGAATATGACTGGCTCAGTAATGCCGAG 1821
Query 484 AAAAGCGGAAAATGCAGCCGGAAGCGCGATTTCTTTTGTCTTTGCCTTCTTCGCCATAAA 543
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1822 AAAAGCGGAAAATGCAGCCGGAAGCGCGATTTCTTTTGTCTTTGCCTTCTTCGCCATAAA 1881
Query 544 GAAGACGGCAAGGCCGGCACCGCCCTGTGCGACATTCGCCATTGA 588
|||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1882 GAAGACGGCAAGGCCGGCACCGCCCTGTGCGACATTCGCCATTGA 1926

Fig. S6. The DNA sequence chromatogram (above) and blastn analysis (below) of a 396 bp long
fragment of the panB gene amplified from our PY79(ICEBs1+) strain. DNA sequencing was
performed by using an automated model 3730 DNA analyzer and BigDye Terminator (Applied
Bipsystems, Inc). The 396 bp long panB gene fragment was amplified from the genome of strain
PY79(ICEBs1+) by PCR with the following primer sets: panB F (5’-
TCATGTCCGCAACTGTCACAC-3’), panB R (5’-ATTGGGCAGTATCGCCTGCGA-3’) and
labeled with a box on chromatogram.
Target reference sequence used for blastn: Bacillus subtilis strain PY79 pantothenate synthetase
(panC) gene, partial cds; ketopantoate hydroxymethyltransferase (panB) gene, complete cds; and
transcriptional regulator and biotin acetyl-CoA-carboxylase synthetase (birA) gene, partial cds;
Sequence ID: gb|EF191505.1
Query 82 TCATGTCCGCAACTGTCACACCGACAGTTGAATCAAGGCCGAGGACGACCATTCCGAGTG 141
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1082 TCATGTCCGCAACTGTCACACCGACAGTTGAATCAAGGCCGAGGACGACCATTCCGAGTG 1141
Query 142 AATCACCGACTAAAATCATGTCAACTCCCGCTTGTTCAGCAAGCTTAGCTGCCGGATAAT 201
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1142 AATCACCGACTAAAATCATGTCAACTCCCGCTTGTTCAGCAAGCTTAGCTGCCGGATAAT 1201
Query 202 CATAAGCGGTCAGCATCACAATCGGTTCTTCATTCTCCTTCATTTTTAGAAAATCGAGTT 261
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1202 CATAAGCGGTCAGCATCACAATCGGTTCTTCATTCTCCTTCATTTTTAGAAAATCGAGTT 12
Query 262 TTGTTTTCATGTTTTCTCCTCCTCATGTTTAGAGGAGATGAGCTGCCGTTTTCTTATACA 321
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1262 TTGTTTTCATGTTTTCTCCTCCTCATGTTTAGAGGAGATGAGCTGCCGTTTTCTTATACA 1321
Query 322 AGCCAAAAAACCTTCCGTTTAACGAGAAAGATTCTTCACTTTCTAAAGTTCGGCGAGTTT 381
|||||||||||||||||||||||||||| |||||||||||||||||||||||||||||||
Sbjct 1322 AGCCAAAAAACCTTCCGTTTAACGAGAAGGATTCTTCACTTTCTAAAGTTCGGCGAGTTT 1381

Query 382 CATCCCTCTGTCCCAGTCCTTTTTTGGATCAAGGCAGACTGCTGCAATGTCTATCTATTT 441
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Sbjct 1382 CATCCCTCTGTCCCAGTCCTTTTTTGGATCAAGGCAGACTGCTGCAATGTCTATCTATTT 1441
Query 442 TTATAATAGGTGCAGTTCGCAGGCGATACTGCCCAAT 478
|||||||||||||||||||||||||||||||||||||
Sbjct 1442 TTATAATAGGTGCAGTTCGCAGGCGATACTGCCCAAT 1478
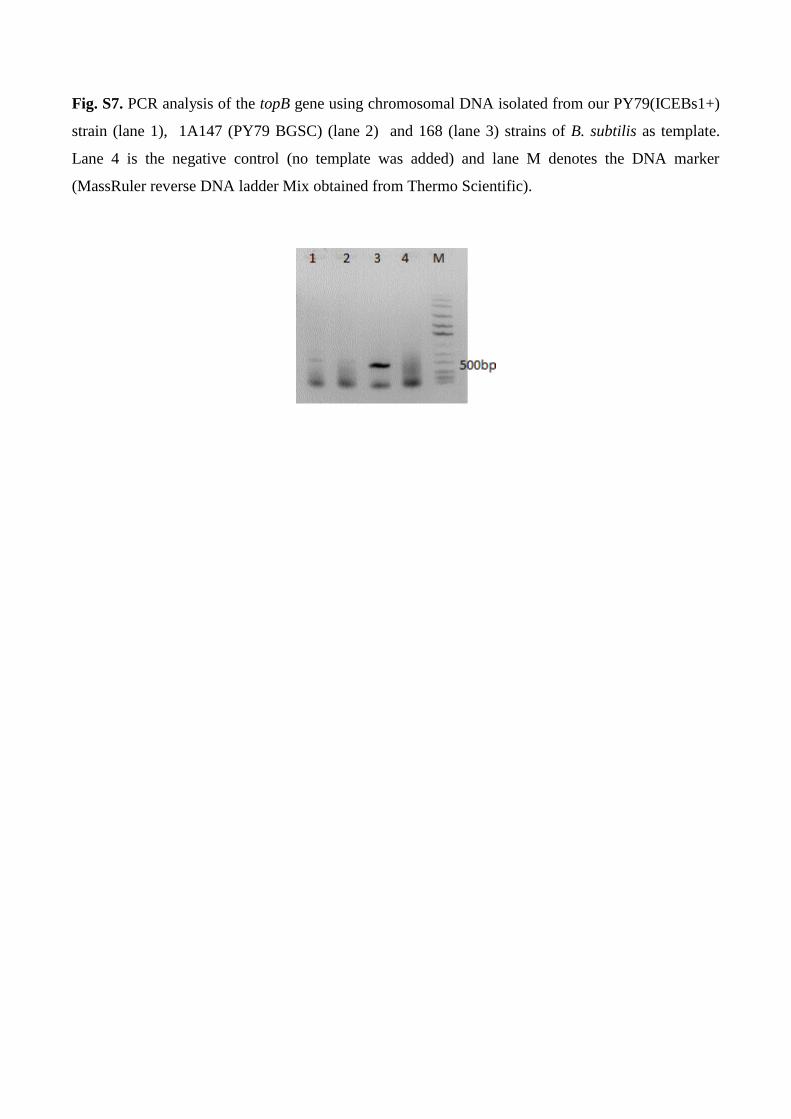
Fig. S7. PCR analysis of the topB gene using chromosomal DNA isolated from our PY79(ICEBs1+)
strain (lane 1), 1A147 (PY79 BGSC) (lane 2) and 168 (lane 3) strains of B. subtilis as template.
Lane 4 is the negative control (no template was added) and lane M denotes the DNA marker
(MassRuler reverse DNA ladder Mix obtained from Thermo Scientific).





























