Embed Size (px)
Citation preview


Izoenzirni ALP 0. Sokolit i I. t e ~ e l a k
ekspresijom u osleoblastima, hepatocitima, bubre2nim stanicama i placenti. Gen koji je odgovoran za nastajanje ovoga izoenzima nalazi se na kraju kratkoga kraka kromozoma 1 (bands p36.1-p34). Ovaj genski lokus jc heterogen i po pitanju elektroforeze, kao i organskoga porijekla. Cini se, medutim, kako je heterogenost rczultat i postranslacijskih modifikacija, poglavito sadrgaja ugljikohidrata;
- intestinalni lokus na kromozomu 2 (bands q34-q37) odreduje tzv. intestinalni izoenzim ALP, koji se normalno nalazi u crijevnoj sluznici (posebice u mikrovilima epilelnih stanica crijeva) fetusa i odraslih osoba. U novije se vrijeme raspravlja i o znatajnijoj ekspresiji ovoga izoenzima u S3 segmentu proksimalnih bubrehih tubula;
- sinteza placentne ALP kodirana je jednim lokusom za koji je poznato nekoliko aIealnih varijanti. Boyer je jog 1961. godine elektroforezom na Skrobnom gelu iden- tificirao tri placentne linije ALP: tip A, koji se obitho nalazi u bijelih k n a ; tip B koji je odgovoran za povedanje aktivnosti ukupne ALP u Zena tijekom trudnote, i lip D koji se nalazi u crnkinja;
- tzv. "germ ceI1" lokus odreduje izoenzim sl ihn, ali ne i identican placentnom izoenzimu ALP, nacienom u malim koliCinama u testisima i timusu zdravih osoba (tzv. testikulni izoenzim).
Posljednja tri tkivno spec i fha gena nalaze se na kraju dugoga kraka kromozoma 2 i medusobno su vrlo sliCni po genomskoj strukturi. B k o se npr. placentna i "germ cell" ALP razlikuju samo u 7-10 aminokiselinskih ostataka, odnosno njihova homolognost je 98%, homolognost placentne i crijevne je 86.5%, te crijevne i tkivno nespecifiCne je 56.6%.
b) NOMENKLATURA IZOENZMA I VISESTRUKIH OBLIKA ALP
Izoenzime u serumu klasificiramo na osnovi njihove elektroforetske pokretljivos- ti. Oznabvaju se imenom ili skraCenicorn i brojem: najbrii izoenzim, dakle koji putuje najviSe anodno, je osteohepatski izoenzim ili ALP 1: subfrakcija ALP l a oznatava hepatski izoenzim, a ALP l b koStani izoenzim. Izoenzimi sa sporijom pokretljivoSCu se oznakavaju s veCim brojevirna: ALP 2 je placentni, a ALP 3 crijevni izoenzim. Oblici negenetskog podrijetla, viSestruki oblici koji se pojavljuju u patoloSkim stanjima, zovu se atipieni oblici i oznahvaju se dijagnostieki znahjnim imenima. Thko se oblik koji tijekom elektroforeze na celogel vrpcama putuje brie od osteohepatskoga izoenzirna naziva 2uCni oblik, oblik visoke molekularne mase ili brzi jetreni oblik ALP. Ovaj oblik se nalazi u serumu bolesnika s opstrukcijskom bolesti jetre, sadr2i lipidni dio koji mo2e biti prisutan kao rnembranski mjehurie, odnosno vezikula. Iako se ovaj oblik nikada ne nalazi u serumu djece, Schoneau i sur. su opisali prisutnost oblika visoke molekularne mase ("high Mr ALP") u serumu djece s dijagnozom jetrene disfunkcije u cistiCnoj fibrozi. Oblik ALP u serumu bolesnika s neoplazmom zove se onkofetalni oblik. U elektroforetskom polju putuje tako Sto se preklapa s placentnim izoenzimom (ALP 3), iako su fizikalno kemijski raz1iCiti. Oblici sa sporo izra2enom rnigsacijom su uglavnom enzim-imunoglobulin kompleksi (prema nekima, EIC-ALP) ili enzim-lipoprotein X kompleksi, odnosno makroenzimi. U literaturi se takoder spominje i tzv. mezenhimni oblik negenetskog podrijetla, kao multipli oblik crijevnoga izoenzima ALP.
114 BIOCHEMLA MEDICA god 4, bl: 4, 1994.

B. Sokolie i I. b p e l a k Izoenzimi ALP
C) DIJAGNOSTICKO ZNACENJE IZOENZIMA I VISESTRUKIH OBLIKA ALP U SERUMU
Jetrene bolesti
Roberts je joS u svojim prvim radovima 1930. godine opisao glavne varijacije ALP u bolestima jetre i ZuCnih putova, koje vrijede joS i danas. Primjerice, katalititka aktivnost ALP je umjereno povekana u serumu bolesnika s infektivnim hepatitisom, ali znabjno u serumu bolesnika s opstrukcijskom Zuticom i malignim bolestima jetre.
Prisustvo abnormalnoga ZuCnog oblika u serumu je v a h a znaCajka bolesnika s opstrukcijskom jetrenom bolesti, pri Cemu pojava ovoga oblika korelira s tezinom opstrukcije. NajveCe povekanje jetrene ALP, ukljuCujuCi i 2uCnu, biljegi se u serumu bolesnika s potpunom opstrukcijom 2uCnih putova, npr. u sluCaju karcinoma 2uc'ovoda ili glave pankreasa, primarnoga karcinoma jetre i primarne bilijarne ciroze. SliCni oblici opazaju se i u bolesnika sa sekundarnim jetrenim metastazama. ZnaCajno je da u ranom stadiju jetrene infiltracije povehna aktivnost jetrenoga izoenzima mo2e biti jedini patoloSki nalaz.
Humani serum redovito sadr2i jetreni izoenzim, elektroforetski identiCan tkivnoj (citozol-topivi oblik) ALP, Radi se o glavnom jetrenom izoenzimu ili serum-topivoj ALP. Naime, ALP u serumu moZe biti prisutna u najmanje tri razIiCita oblika: a) "slobodni" topivi oblik; b) kao membranski vezani oblik ("membrane like") ili koinozim; i c) kao oblik ALP vezan s LP-X.
MoguCi pojedinaCni ili mijeSani mehanizmi sekrecije jetrene ALP u cirkulaciju, ovisno o patoloSkom stanju su slijededi: a) izravno oslobadanje sa sinusoidne membrane; b) transcitoza; i c) regurgitacija iz 2uCi kroz "tight junction",
Glavna jetrena ALP u serumu ima relativno malu molekulu (Mr otprilike 180000), sliCnu kao citozolska ALP, dok patoloSka 2uCna ALP ima veliku molekular- nu masu (najmanje lo6). Ovaj oblik je termolabilniji, a ispitivanja su pokazala kako odgovara serum-topivoj jetrenoj ALP priCvrStenoj za membranu, odnosno kako je "membrane like" fragment ALP. U nekim slutajevima kolestaze prisutan je jasan oblik ALP vezan s lipoproteinom X (ALP-LP-X kompleks).
Oslobactanje membranski vezanoga enzima omoguteno je putom nekoliko mehanizama, koji ukljutuju aktivnost 2uCnih kiselina, proteolizu, bromelain i membransku fragrnentaciju. Noviji radovi posebice naglaSavaju ulogu endogenih fosfatidilinozitol specifiCnih fosfolipaza u stvaranju neagregirajueih multiplih oblika ALP, Svi navedeni mehanizmi u razliCitirn situacijama utjeCu na oslobactanje ALP iz jetrenih stanica. Sinusoidna membrana jc glavni izvor jetrene ALP u serumu boles- nika s jetrenim boleslima. Medutim, za vrijeme kolestaze mo2e takoder doki do "iskrivljenja" Cvrste kanikulne veze (tight junction), tako da ALP iz kanikuinoga podruCja, povezana sa 2uCnim lipoproteinima, npr. LP-X, utazi u cirkulaciju. U serumu bolesnika s kroniCnim jetrenim bolestirna, poglavito cirozom, mo2e biti povekna aktivnost i crijevnoga izaenzima ALP. U tih se bolesnika moZe razviti melabolitka bolest kostiju s povedanjem koStanog enzima, koji k m o prepoznati s vise razliCitih postupaka separacije.
BIOCHEMLA MEDICA gad. 4, 61: 4, 1994.

Izoenzimi ALP B. Sokolik i I. C e ~ e l a k
KoStane bolesti
KoStani izoenzim ALP nalazi se u membranama osteoblasta, a dokazana je korelacija izmedu povckanja koStanog izoenzima i aktivnosti osteoblasta, $to posebice vrijedi za djecu u vrijeme rasta kostiju.
NajkSka patoloSka stanja kostiju u kojima se povedava katali tiCka aktivnost ALP su Pagetova bolest, rahitis, primarni i sekundarni tumori kostiju i primarni hiper- paratiroidizam.
Najvek poveknje koStane ALP u serumu biljegi se u bolesnika s Pagetovom bolesti. Lijeknje ti h bolesnika s citotoksiCnirn lijekovima, kalcitoninom ili bifos- fonatima rezultira smanjenjem katalitieke aktivnosti.
Poveeana a ktivnos t koStanoga izoenzima u serurnu bolesnika s osteomalacijom i rahitisom posljedica je rnalnutricije, malapsorpcije uslijed gastrektomijc, kroniCne jetrene bolesti, trovanja teSkim metalima ili rezistencije na vitamin D. Zato lijeknje osteomalacije s vitaminom D ili njegovim analozima poCetno rezultira povetanjem serumske ALP, zbog povehne osteoblatske aktivnosti, a kako se stanje popravlja, ta aktivnost se smanjuje.
Mhligne bolesti
Neki radovi ukazuju na povezanost ekspresije tkivno-specifihih ALP i malignos- ti. Tbmore koji stvaraju takve oblike ALP opkni to dijelimo u dvije skupine : a) tumori s pojahnim stvaranjem izoenzim ALP, koji se normalno stvaraju u tkivu, tm. eutopiCna ekspresija i b) tumori koji pokazuju ekspresiju jednoga ili vige izoenzima i viSestrukih oblika koji se ne nalaze u tkivu zdrave osobe, tzv. ektopitna produkcija. Mnogi tumori pokazuju i simultanu ekspresiju dvaju ili vise razlititih izoenzima ili viSestrukih ablika ALP.
Ektopiha sinteza placentne ALP zabilje2ena je kod karcinoma p l u k (Nagao i Regan oblik), jajnika, maternice i gastrointestinalnog trakta. "Germ cell" ALP se ektopiCno stvara u velikim koliCinama u tumorima testisa, karcinomu jajnika, kao i u tirnusu.
Crijevna ALP se primarno stvara u hepatomu, premda je nadena i u nekim drugim tumorima.
Pretpostavlja se i raspravlja o nekoliko mehanizama za ekspresiju izoenzima ALP u tumorskim stanicama, kao Sto su: funkcijsko ukljueivanje izoenzima ALP u tumorogenezu, po drugom je ekspresija ALP samo jedan od Cinitelja u multifaktor- noj etiogenezi, a po t r ekmu postoji uska povezanost gena ALP s genima odgovornim za bolest.
Ostala patologka stanja
RazliCiti poremekji mogu rezultirati povehnjem katalitieke aktivnosti ALP bez jasnih znakova ukljuknja odredenog organa. Tmko bolesnici s reurnatskim artritisom mogu imati poveknu aktivnost ukupne ALP, pretegito jetrenoga podrijetla, dok bolesnici s ankiloznim spondilitisom imaju povedanu ALP na raCun koStanoga izoen- zima.
U serumu dojenCadi i djece moguh je izrazito povetana aktivnost ALP ( i do 10 puta od gornje granice normalnih vrijednosti), ali bez prisustva jetrenih i koStanih bolesti. NajCeSCe se radi o benignoj prolaznoj hiperfosfatazemiji joS uvijek nepoznate
BIOCHEMU MEDICA god. 4, bl: 4,1994.

B. Sokolik i I. Cepelak Izoenzimi ALP
etiologije. Obuhvateni su uglavnom jetreni i koStani izoenzim, a aktivnost se spon- tan0 v r a k na normalu unutar 4 mjeseca. R i su pretpostavljena mehanizma odgovor- na za povecanje ALP: povehna sinteza ili oslobadanje iz tkiva, aktiviranje normalnih koliCina cirkulirajutega enzima i smanjeno odstranjivanje enzima iz cirkulacije.
Bubreina tubulna acidoza, kao i drugi tubulni poremehji, takoder su povezani s poveCanjem aktivnosti ALP u serumu, iako nije uvijek poznat tkivni izvor enzima. Odrect'ivanje crijevnoga izoenzima ALP (ekspresija u S3 segmentu proksimalnih tubula bubrega) u mokraCi, uz visoko specifiCna monoklonska protutijela ALP 250, imat Ce, Cini se, znahjnu ulogu u diferencijalnoj dijagnostici bubreinih bolesti.
Dva vrlo rijetka patoloSka stanja, bolest alfa-lanaca ili limfom tankoga crijeva i "graft versus host" bolest tankoga crijeva, su povezana s poveCanjem crijevnoga izoenzima ALP.
d) POSTUPCI ZA ODJEWIVANJE I RAZLIKOVANJE IZOENZIMA I MULTIPLIH OBLIKA ALP U SERUMU
Postupke za odjeljivanje izoenzima ALP dijelimo na one koji se osnivaju na promjenama u katalititkirn svojstvima enzima i one koji se osnivaju na svojstvima vezanima uz molekularnu strukturu izoenzima (Bblica 1).
Tab 1 i c a 1. - Svojstva za razlikovanje izoenzima i vigestrukih oblika ALP. T a b 1 e I. -- Traits to distinguishing isoensymes and multiple forms of ALP.
/ Svojs~va vezana ue rnolckulanru strukturu I KatalitiEka svojstva I - r e l a ~ i w a rnoIekularna masa i naboj
- antigena specifirnost
- srabilnost na toplinu, urea, kiseii pH
- specifiCnost prema supstratu
- osjetljivost na inhibitore
E 1 e k t r o f o r e z a se i danas smatra referentnim postupkom za razdvajanje izoenzima ALP, jer ornogukiva odjeljivanje i identifikaciju patoloSkih izoenzima, h k i kada je ukupna aktivnost ALP u granicama normalnih vrijednosti, a Sto se ne moie otkriti ni jednim drugim postupkom.
Vckina izoenzima se odjcljuje na temelju njihove elektroforetske pokretljivosti na nosaeima kao Sto su Skrobni gel, agar gel; agaroza, celuloza acetat te poliakrilamid gel. Medutim, ni na jednom od navedenih n o s a h nije moguk, bez dodatnih pos- tupaka, jasno odvojiti glavne serumske izoenzime ALP, jetreni i kostani.
Kada se kao nosat rabi agaroza i celuloza acetat, glavni jetreni izoenzim putuje kao kompaktna linija u alfa 2 globulinskom podruqu. KoStani izoenzim je odmah iza jetrenoga, doti& ga se i karakterizira ga viSe difuzno obojena linija. Zbog toga ih je vrlo teSko denzitometrijski kvantificirati, posebice ako se radi o malom poveCanju jednoga u prisutnosti velikoga povehnja drugoga izoenzima.
Na Skrobnom i poliakrilamid gelu, pokretljivost djelomiee ovisi o koncentraciji geiova, ali su navedeni izoenzimi takoder vrlo blizu.
BIUCHEMLA MELVCA god. 4, br: 4, 1994. 11 7
Izoenzinli ALP B. Sokolik i I. Cepelak
Ovim se postupcima mogu registrirati i drugi izoenzimi i vikstruki oblici. B k o se u bolesnika s opstrukcijskim bolestima pojavljuje tzv. drugi jetreni ZuCni oblik, koji putuje ispred glavnoga jetrenog izoenzima kada se kao nosaC rabi celuloza acetat ili agaroza gel; na Skrobnom ili poliakrilamid gelu ovaj izoenzim ostaje na startu ili je u blizini starta.
Crijevni izoenzim putuje sporije od jetrenoga i koStanogra na svim nosatima, dok fetalno crijevni putuje br2e od crijevnoga u odraslih.
Placentni izoenzim putuje ovisno o nosaCu, uglavnom kao diskretna linija vrlo blizu koStanoga izoenzima ili ga ovaj prekriva.
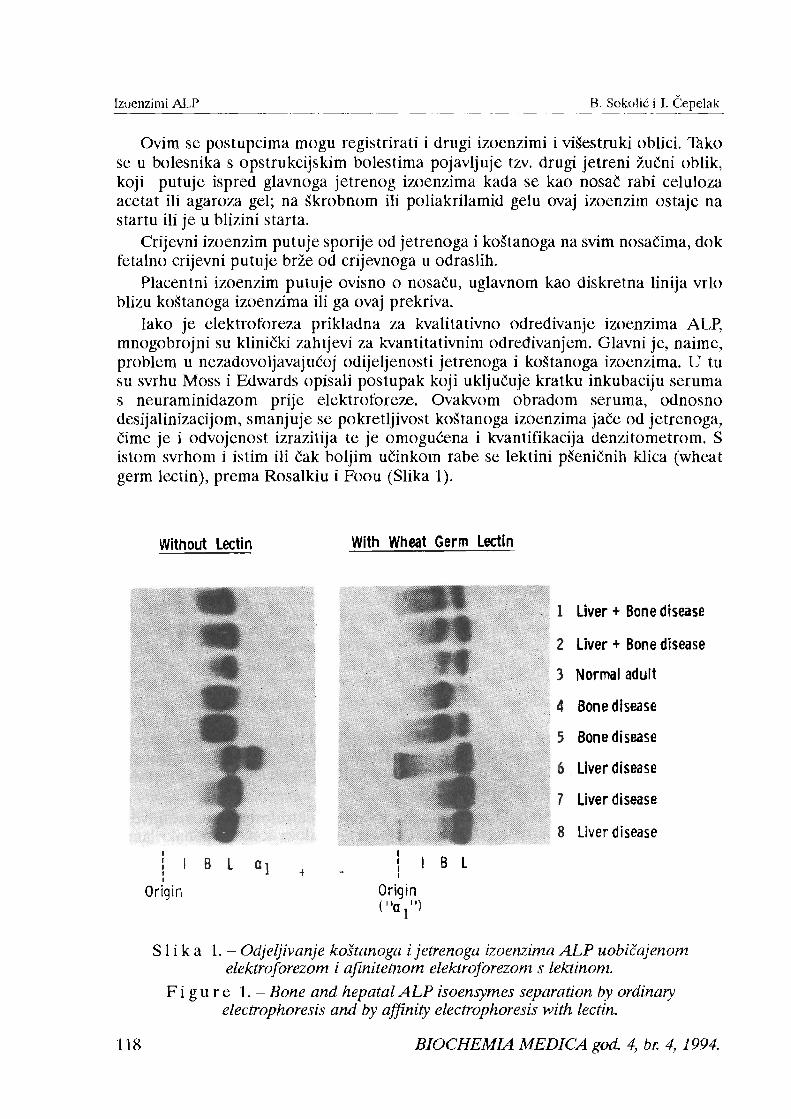
Iako je elektroforeza prikladna za kvalitativno odredivanje izoenzima ALP, mnogobrojni su klinicki zahtjevi z,a kvantitativnim odredivanjem. Glavni je, naime, problem u nezadovoljavaju6oj odijeljenosti jetrenoga i kogtanoga izoenzima. U tu su svrhu Moss i Edwards opisali postupak koji ukljutuje kratku inkubaciju seruma s neuraminidazom prije elektroforeze. Ovakvom obradom seruma, odnosno desijalinizacijom, smanjuje se pokretljivost koStanoga izoenzima jaCe od jetrenoga, Cime je i odvojenost izrazitija te je omogukna i kvantifikacija denzitometrom. S istom svrhom i istim ili h k boljim uCinkom rabe se lektini pSenitnih klica (wheat germ lectin), prema Rosalkiu i Foou (Slika 1).
Without Lectin With Wheat Germ Lectin
Liver + Bone disease
Liver + Bone disease
Normal adult
Bone disease
Bone disease
Liver disease
Liver disease
Liver disease
Origin
I I 1 I B L I
Origin i "a
S 1 i k a 1. - Odjeljivanje koitanoga i jetrenoga lzoenzimcl ALP uobiCcljenorn elektroforezom i afinitetnom elektrof-brezorn s lektinonz.
F i g u r e 1. - Bone and hepatal A L P isoenhymes separation by ordinary electrophoresis and by aflnity electrophoresis with kctin.
118 BIOCHEMLA MEDICA god. 4, br. 4, 1994.

R. SokoliC i I. tepelak Izoenzimi ALP
Neuraminidaza ne utjeze na pokretljivost crijevnoga izoenzima, dok se placentni izoenzim preklapa s koStanim te su potrebni i dodatni postupci razlikovanja.
~ u ~ n i oblik ALP, ukoliko je prisutan, veZe sse s lektinom pa ili ostaje na startu ili blizu starta, za razliku od njegove alfa I-globulinske pokretljivosti tijekom uobihjene elektroforeze.
Afinitetni precipitacijski postupak
Rosalki i Foo su prvi izvijestili o precipitacijskom postupku i pokazali kako se otprilike 80% koStanoga izoenzima precipitira s lektinom pSeniCnih klica, samo manje od 10% jetrenoga, dok placentni i crijevni ne precipitiraju. UkoIiko je prisutan hCni oblik ALP, serum treba prethodno obraditi s Jkiton-X-100 detergentcm, kako bi sc sprijetila njegova precipitacija.
Behr i Barnert sw modificirali ovaj postupak te precipitirali koStani izoenzim 100% i dobili vrlo dobru korelaciju s toplinskim inaktiviranjem.
I z o e 1 e k t r i C n o f o k u s i r a n j e u poliakrilamidu, celuloza acetatu ili agarozi daje veki broj linija nego uobihjena elektroforeza na celuloza acetatnim vrpcama. Neki su autori tako identificirali i do 12 linija ALP u serumu zdravih osoba. Kako velik broj linija testo i zbunjuje, tehnizka prednost izoelektriCnog fokusiranja za sad nijc znabjno poboljSala kliniCku korisnost.
Veliko zanimanje zadnjih godina izazvala je uporaba visokotlaCne k r o m a t o- g r a f i j e za odjeljivanje i kvantificiranje izoenzima ALP u serumu. Ovim su postupkom odredene dvije linije kogtanoga izoenzima u serumu djece, dvije linije jetrenoga izoenzirna i 2ulini oblik u serumu bolesnika s kolestazom.
Kromatografijom s ionskom izmjenom u serumu zdrave osobe su odredena dva jctrena, jedan koStani i jedan crijevni izoenzim. Uporaba Iektina pSeniCnih kbca prigodom kromatografije nije pokazala zadovoljavajuCi uEinak. ~ u t n i oblik ALP se Jasno odjeljuje vekinorn krornatografskih postupaka, pa se kromatografija s ionskom izmjcnom i gel filtracijska kromatografija rabe posebice za identificiranje i b a n - tificiranje ovoga oblika.
1 m u n o k e m i j s k i p o s t u p c i. Prvi pisani dokumenti o protutijelirna protiv ALP opisani su joS 1954. godine. Kasnije se pokazalo kako su jetreni, koStani i bubre2ni izoenzimi antigeno vrlo sliCni, a crijevni i placentni razliCiti. Monoklonska protutijela protiv jetrenoga, koStanoga i bubrehoga izoenzima daju potpunu kriinu reakciju s ostala dva, dok su nedjelatna u odnosu na placentni i crijevni izoenzim.
Toplinska stabilnost Izoenzirni ALP u serumu denaturiraju se toplinom, i to specifiCnom brzinom
denaturacije. %ko je npr. zagrijavanjem na 65 "C kroz 10 minuta moguCe inaktivirati sve lzoenzime osim placentnoga, koji je stabilan. KoStani izoenzim, koji je vrlo termolabilan, potpuno se inaktivira vet zagrijavanjem kroz 10 minuta ha 56 "C. Jctreni i crijevni izoenzim pri ovoj temperaturi pokazuju srednju osjetljivost.
Za razlikovanje jetrenoga i koStanoga izoenzima stoga se moZe rabiti jednostavan postupak inkubacije seruma na 56 "C tijekom 10 minuta. Metoda zahtijeva preciznu kontrolu temperature, toCno odrecteni mali volumen uzorka, epruvetu tanke stijenke
BIOCHEML4 MEDICA god. 4, br 4,1994. 119

Izoenzirni ALP B. SokoliC i I. t q e l a k
i ledenu kupelj za prekidanje inkubacije. Ostatna aktivnost od manje od 10% ukazuje na postojanje kogtanoga izoenzima.
Sbbilnost na kemijska sredstva
UCinak ureje na aktivnost izoenzima ovisi o vremenu inaktiviranja, temperaturi i koncentraciji ureje. Pri niskim koncentracijama, ureja izaziva reverzibilno inak- tiviranje, dok pri viSim koncentracijama izaziva ireverzibilno inaktiviranje. Mnoga izvje3t.a pokazuju kako je koStani izoenzim osjetljiviji u odnosu na ostale izoenzime. Raznolikost reakcijskih uvjeta, odnosno mnogobrojne modifikacije postupka, razlog su rijetke praktiCne primjene. Kao alternativa ureji, kao bolji inaktivator prema Shepardu i Peakeu, rabi se gvanidin hidroklorid, kojemu je potrebno k r a k vrijeme inkubiranja. Kod toCno odredenih uvjeta koStani se izoenzim inaktivira 86%, a jetreni 53%. Ovi su autori pokazali kako je kombinacijom inaktiviranja s gvanidin hidrokloridom, toplinskog inaktiviranja na 60 "C i L-fenilalanin inhibicije moguCe odrediti 4 izoenzima (jetreni, koStani, crijevni i placentni), ali samo ako su njihove aktivnosti povebne.
L-fenilalanin (5-10 mmol/L) inhibira oko 80% crijevnoga i placentnoga izoen- zima ALP. Radi se o nekompetitivnoj inhibiciji koja je u svezi sa sprijehvanjem defosforilacije enzima. I druge aminokiseline, kao histidin, Iizin, L-homoarginin, glutaminska kiselina, L-triptofan te neki lijekovi (levamisol, bormtetramisol), takoder u razliCitom stupnju inhibiraju neke izoenzime ALP.
Referentne vrijednosti
Prema dosadahjim saznanjima, ne postoji znaCajniji dnevni ritam aktivnosti ALP, a niti umjerena tjelesna aktivnost ili mirovanje ne mijenjaju aktivnost enzima. Nakon masnoga obroka, ukupna se aktivnost lagano poveh, Sto se prete5ito odnosi na povehnje crijevnoga izoenzima. Osim vitamina D, nema znatajnijega utjecaja os- talih prehrambenih tvari na aktivnost ALP (zabiljezene su u sporadihim sluCajevima smanjene aktivnosti uslijed manjka unosa proteina, cinka i magnezija).
Utjecaj dobi na referentne vrijednosti ALP dobro je poznat. Giavni je razlog rast kostiju, dakle osteoblastna aktivnost, i pojava koStanoga izoenzima i cirkulaciji. Ovaj izoenzim ima maksimalnu aktivnost u p n i h 6 mjeseci Zivota, smanjuje se na odredenu razinu u razdoblju izmedu 2. i 9. godine Zvota, ponovno se povehva u pubertetu te se smanjuje do granim referentnih vrijednosti oko 18-20. godine.
U pwim mjesecima Zivota jetreni izoenzim ima vrlo malu ili nikakvu aktivnost, koja se povehva do odredene razine do 6. mjeseca iivota i na toj se razini zadrgava i daIje.
Manje aktivnosti crijevnoga izoenzima nalaze se u osoba s krvnom skupinom A ili AB te u osoba koje prije oduzimanja krvi nisu uzele masni obrok.
Ovisno o primijenjenu postupku razdvajanja izoenzima ALP, u literaturi Cemo na6i i razlieite referentne raspone za pojedine izoenzime. Sve dok se ne standar- diziraju postupci i nomenklatura za izoenzime, preporuka je da svaki laboratorij naliini svoje referentne raspone.
BIOCHEMU MEDICA god 4, bl: 4,1994.

ISOENZYMES AND MU1,TIPLE FORMS OF ALP: THE SIGNIFICANCE AND THE METHODS OF ANALYSIS
SUMMARY - The characterisalion of isoenzymes and multiple forms of the alkaline phosphatase (ALfl EC.3.1.3.1) have been described in detail. The methods of analysis as. well a,s. diagnostic significance have been discused The recommandations o n reference vcrlues were given on the end-
LITERATURA
1. Tietz NV Rinker AD: The measurement of alkalinc phosphatase. U: Clinical and Analyti- cal Concepts in Enzymology. Homburger HA College of American Pathologists, Skokic, 1982. 195.
2. Kim EE, Wyckofi HW Structure of alkaline phosphatases. Clin Chim Acta 1989;186:175-8.
3. Moss DW Multiple forms of acid and alkaline phosphatascs: Genetics, exprcssion and tlssue- specific modification. Clin Chim Acta 1986: 161: 123-35.
4. Moss DW Alkaline phosphatase isoenzyme. Clin Chem 1982;28:2007-16.
5. Wrpooten G 4 Nuyrs GD, Hoylaerts MF I' sul: Immunoassay in urine of a specific markcr for proximal tubular segment. Clin Cheni 1992; 38: 642-7.
6. Boyer SH. AIkaline phosphatase in human sera and placentae, Science 1961;134:1002-4.
7. Burlina A. Diagnostic significance of alkaline phosphatase isoenzymes. MediLab 1985; 6/7: 18-26.
8. Schoneau E. f f enog KH, Roehies HHJ: Liquid chromatographic determination of isoenzymes of alkaline phosphatase in serum and tissue homogenates. Clin Chem 1986;32:816-8.
9. Roberts W, Variations in the phosphatase ac- tivity in the blood in disease. Rr J Exp Path01 1930;11:90.
10. Mom DW Perspectives in alkaline phosphatase research. Clin Chem 1992;38:2486-92.
11. Schallaeger R, Haux I: Kattcrrnann R Studies on he mechanism of the increase in serum alkaline phosphatase activity in cholestasis: Significance of the hepatic biie acid concentra- tion for the leakage of alkaline phosphatase from rat liver. Enzyme 1982;28:3-13.
12. Korninami 7; Miki A, Ikehara Y Electro- phorctic characterisation of hepatic alkaline phosphatase released by phosphatidilinositol- specitic phmpholipase C. Biochem J 1985; 227: 183-6.
13. Joplin G c Slevenson JC. Paget's disease of bone. U: StevensonJC. New Techniques in Me- tabolic Bone Disease. London: Wright, 1990: 266-78.
14. Millian JL. Alkaline phosphatase as a reporter of cancerous transformation. Clin Chim Acta 1992;209:123-9.
15. Jeppsorl A. Wharen B, Brehmer-Andersson E, Silfversward C. Stigbrarld 7: Millan JL. Eutopic expression of placental-like alkaline phos- phatase in testicular tumours. J Cancer 1984; 34: 757-61.
16. Sturk A, Sanders GB7: Macro enzymes: Prevalence, composition, detection and clinical relevance. J Clin Chem Clin Biochem 1990; 28: 65-81.
17. Crofton PM. What is the cause of benign tran- sient hyperphosphatasemia? A study of 35 cases. Clin Chem 1988;34:335-40.
IS. Moss DW Edwards RK. Improvcd electro- p!loretic resolution of bone and liver alkaline phosphatases resulting from partial digestion with neuraminidase. Clin Chim Acta 1984; 143: 177-82.
19. Rosalki SB. Foo AY 3vo ncw methods for separating and quantifyng bone and liver alkaline phosphatase isoenzymes in plasma. Clin Chem 1984;30:1182-6.
20. Shephard MDS. Peake M,! Quantitative method for determining serum alkaline phos- phatase lsoenzyme activity. I Guanidine hydrochloride: New rcagent for selectively in- hibiting major serum isoenzymes of alkaline phosp hatase. J Cl~n Path01 1986;39:1025-30.
21. vatz Hoof VO. Hoylaerts M 4 G q f t-I, van Mnl- !ern M, Lepourte LG) De Broe ME. Agc and scx distribution of alkaline phosphatase isocn- zymes by agarose electrophoresis. CIin Chem 1990;36:575-8.
22. Panteghini M. Hepatlc alkaline phosphatase isoenzyme: I1 Clinical aspects. Giorn I t Chim Clin 1990;15:301-8.
Rad prirnljen: 25. 10. 1994.
Rad prihvaien: 22. 12. 1994,
BIOCHEMIA MEDICA gad. 4, 61: 4, 1994.
















![Ngày [ ] tháng [ ] năm 2010pentstudiotayho.org/wp-content/uploads/2017/07/HDMB-Mau-PentSt… · về việc nhà ở hình thành trong tương lai được đưa vào kinh doanh](https://img.pdfslide.tips/doc/110x75/5f0e83bb7e708231d43f9d44/ngy-thng-nfm-v-vic-nh-hnh-thnh-trong-tng-lai-c.jpg)












