Embed Size (px)
Citation preview

Struktur-Funktionsanalyse der QB-Bindenische im Photosystem II
von Chlamydomonas reinhardtii
Dissertation
zur Erlangung des Grades
eines Doktors der Naturwissenschaften
der Fakultät für Biologie
der Ruhr-Universität Bochum
angefertigt am
Lehrstuhl für Biochemie der Pflanzen
vorgelegt von
Grazyna Orawski
aus Nikolai
Bochum 2001

Tag der mündlichen Prüfung: 03.07.2001
Referent: Prof. Dr. W. Oettmeier
Korreferent: Prof. Dr. W. Hengstenberg

Herrn Prof. Dr. W. Oettmeier danke ich für die Vergabe des Themas und die gute
Betreuung der Arbeit.
Für die freundliche Übernahme des Korreferates bedanke ich mich bei Herrn Prof.
Dr. W. Hengstenberg.
Dem Sprecher Herrn Prof. Dr. E. Weiler, allen Dozenten, Kollegiatinnen und Kol-
legiaten des Graduiertenkollegs „Biogenese und Mechanismen komplexer Zell-
funktionen“ danke ich für die Unterstützung und das Interesse an meiner Arbeit.
Allen Mitarbeitern des Lehrstuhls für Biochemie der Pflanzen danke ich für ihre
ständige Hilfsbereitschaft und das angenehme Arbeitsklima.
Michael danke ich für die Unterstützung während des ganzen Studiums.

Inhaltsverzeichnis 1
Inhaltsverzeichnis
Inhaltsverzeichnis ...............................................................................................................1 Abkürzungsverzeichnis......................................................................................................3 1. Einleitung .........................................................................................................................5 2. Material und Methoden................................................................................................12
2.1. Chemikalien, Enzyme und Verbrauchsmaterialien........................................12 2.2. Algen- und Bakterienstämme ..............................................................................13
2.2.1. Algenstämme.....................................................................................................13 2.2.2. Bakterienstamm...............................................................................................13
2.3. Algenanzucht...........................................................................................................14 2.3.1. Anzuchtbedingungen ......................................................................................14 2.3.2. Medien ................................................................................................................14
2.4. Bakterienanzucht ...................................................................................................15 2.5. Oligonukleotide........................................................................................................17
2.5.1. Oligonukleotide für die ortsgerichtete Mutagenese..................................17 2.5.2. Oligonukleotide für die DNA-Sequenzierung ............................................18
2.6. Plasmid......................................................................................................................19 2.7. Molekularbiologische Methoden..........................................................................19
2.7.1. Gerichtete Mutagenese mit Anwendung der Polymerase- Kettenreaktion.............................................................................................................19 2.7.2. Polymerase-Kettenreaktionen mit Algen-DNA.........................................24 2.7.3. Agarose-Gelelektrophorese zur Auftrennung von DNA..........................25 2.7.4. DNA-Sequenzierung........................................................................................26 2.7.5. Isolierung von Gesamt-DNA aus Chlamydomonas reinhardtii .............26 2.7.6. Herstellung kompetenter Bakterienzellen .................................................27 2.7.7. Transformation kompetenter Bakterienzellen ..........................................27 2.7.8. Präparation von Plasmid-DNA .....................................................................28 2.7.9. Transformation des Chloroplasten von Chlamydomonas reinhardtii ..28
2.8. Präparation von Thylakoidmembranen aus Chlamydomonas reinhardtii30 2.9. Isolierung von Protein aus Chlamydomonas reinhardtii für die SDS-Polyacrylamid-Gelelektrophorese ...............................................................................31 2.10. SDS-Polyacrylamid-Gelelektrophorese von Protein aus Chlamydomonas reinhardtii ........................................................................................................................31 2.11. Immunologische Methoden.................................................................................33
2.11.1. Transfer von Proteinen auf Nitrozellulosemembranen.........................33 2.11.2. Immunologischer Nachweis des D1-Proteins aus Chlamydomonas reinhardtii.....................................................................................................................34
2.12. Photometrische Meßmethoden...........................................................................35 2.12.1. Bestimmung des Chlorophyllgehaltes.......................................................35 2.12.2. Bestimmung des Algenwachstums.............................................................35 2.12.3. Messung der Elektronentransportraten von PS II.................................36
3. Ergebnisse.......................................................................................................................37 3.1. Konstruktion von Chlamydomonas reinhardtii-Mutanten............................38
3.1.1. In vitro-Mutagenese des psbA-Gens.............................................................38 3.1.2. Sequenzierung der PCR-Produkte der Mutagenese.................................42 3.1.3. Transformation der Mutante Del1................................................................42

Inhaltsverzeichnis 2
3.2. Analyse der Transformanten mit Hilfe der PCR..............................................45 3.3. Sequenzierung der Transformanten-DNA........................................................52
3.3.1. Sequenzierung der Phe211/Met214-Transformanten..............................52 3.3.2. Sequenzierung der Phe206-Transformanten.............................................53 3.3.3. Sequenzierung der His215-Transformanten..............................................54 3.3.4. Sequenzierung der Val249-Transformanten .............................................55
3.4. Aufnahme von Wachstumskurven der Mutantenstämme.............................56 3.4.1. Aufnahme von Wachstumskurven der Doppelmutanten mit Aminosäureaustauschen an den Positionen Phe 211 und Met 214.................56 3.4.2. Aufnahme von Wachstumskurven der Mutanten mit Aminosäureaustauschen an der Position Phe 206 ..............................................59 3.4.3. Aufnahme von Wachstumskurven der Mutanten mit Aminosäureaustauschen an der Position Val 249 ...............................................61
3.5. Western Blot-Analyse der Mutanten..................................................................64 3.6. Messung der Herbizidresistenz von Chlamydomonas reinhardtii- Mutanten..........................................................................................................................68
3.6.1. Messung der Herbizidresistenz der Mutanten mit Aminosäureaustauschen an den Positionen Phe 211 und Met 214.................68 3.6.2. Messung der Herbizidresistenz der Mutanten mit Aminosäureaustauschen an der Position Val 249 ...............................................71 3.6.3. Messung der Herbizidresistenz der Mutanten mit Aminosäureaustauschen an der Position Phe 206 ..............................................77
4. Diskussion .......................................................................................................................79 4.1. Charakterisierung der Mutanten mit Aminosäureaustauschen an der Position Val 249 im D1-Protein ...................................................................................87
4.1.1. Hemmstoffexperimente ...................................................................................87 4.2. Charakterisierung der Doppelmutanten mit Aminosäureaustauschen an den Positionen Phe 211 und Met 214 im D1-Protein..............................................94
4.2.1. Hemmstoffexperimente ...................................................................................95 4.3. Mutationen der Aminosäure His 215 im D1-Protein.......................................97 4.4. Charakterisierung der Mutanten mit Aminosäureaustauschen an der Position Phe 206 im D1-Protein ..................................................................................99
5. Zusammenfassung.......................................................................................................101 6. Anhang ..........................................................................................................................104
6.1. Strukturformeln der Hemmstoffe ......................................................................104 6.1.1. Phenole .............................................................................................................104 6.1.2. Triazine ............................................................................................................105 6.1.3. Harnstoffe.........................................................................................................106 6.1.4. Triazinone........................................................................................................108 6.1.5. Biscarbamate...................................................................................................108 6.1.6. Sonstige Hemmstoffe .....................................................................................109
6.2. Sequenz des psbA-Gens aus Chlamydomonas reinhardtii und flankierende Bereiche ..........................................................................................................................110 6.3. Nukleotidsequenz des psbA-Gens mit flankierenden Bereichen und Aminosäuresequenz des D1-Proteins aus Chlamydomonas reinhardtii ..........112
7. Literaturverzeichnis ...................................................................................................114

Abkürzungsverzeichnis 3
Abkürzungsverzeichnis
A Adenin
A. dest. destilliertes Wasser
Abb. Abbildung
AK Antikörper
APS Ammoniumperoxodisulfat
ATP Adenosintriphosphat
bp Basenpaar
C Cytosin
Cyt. Cytochrom
DCPIP 2,6-Dichlorphenol-indophenol
Del Deletionsmutante
DMSO Dimethylsulfoxid
DNA Desoxyribonukleinsäure
DNP-INT 2-Jod-6-isopropyl-3-methyl-2’,4,4’-trinitrodiphenylether
dNTP Desoxyribonukleotid
EDTA Ethylendiaminotetraessigsäure
G Guanin
HEPES 2-(4-(2-Hydroxyethyl-)1-piperalzinyl)ethansulfonsäure
IL intronlose Mutante
kbp Kilobasenpaar
kDa Kilodalton
Lsg. Lösung
M molar
Mut. Mutante
N A, oder C, oder G, oder T
NADP Nicotinamidadenindinukleotidphosphat
OD optische Dichte
PAGE Polyacrylamid-Gelelektrophorese
PCR Polymerase-Ketten-Reaktion
PS Photosystem

Abkürzungsverzeichnis 4 rpm Umdrehungen pro Minute
SDS Natriumdodecylsulfat
T Thymin
TEMED Tetramethylendiamin
Tricin N-(Tris-(hydroxymethyl-)methyl-)glycin
Tris Tris-(hydroxymethyl-)aminomethan
WT Wildtyp

1. Einleitung 5
1. Einleitung
Die Sonne stellt die nahezu universelle Energiequelle der Biosphäre dar. Die
Lichtenergie wird von den photosynthetischen Organismen in chemisch gebunde-
ne Energie umgewandelt. Die Lichtreaktion der Photosynthese ist somit eine pho-
tochemische Reaktion, in der energiereiche Verbindungen in Form von ATP und
Reduktionsäquivalenten bereitgestellt werden. Diese Verbindungen werden dann
in der nachfolgenden Dunkelreaktion für den Aufbau von Kohlenhydraten ver-
wendet.
Die Lichtreaktion findet in den photosynthetischen Membranen verschiedener
Bakterien und in den Chloroplasten der Algen und höheren Pflanzen statt. Das
Licht wird zuerst von Antennenkomplexen absorbiert und dann zu den photosyn-
thetischen Reaktionszentren weitergeleitet. Dort kommt es zu einer photoindu-
zierten Ladungstrennung und dem anschließenden Elektronentransport über die
Membran.
An dem Elektronentransport sind bei Cyanobakterien, Algen und höheren Pflan-
zen drei integrale Membrankomplexe beteiligt: Photosystem I (PS I), Photosystem
II (PS II) und der Cytochrom b6/f-Komplex. Die Komplexe sind durch mobile Kom-
ponenten (Plastochinon und Plastocyanin) verknüpft. Die ATP-Synthase stellt
den vierten Membrankomplex dar (Abb. 1.1.).

1. Einleitung 6
Abb. 1.1.: Schematische Darstellung der Anordnung der Reaktionskomplexe des photosynthetischen
Elektronentransportes in Thylakoidmembranen von höheren Pflanzen und Algen. PS I, PS II und
Cyt. b6/f-Komplex enthalten mehrere Elektronencarrier, die an Polypeptiduntereinheiten gebunden
sind. Bewegliche Carriermoleküle wie Plastochinon (PQ) und Plastocyanin (PC) stellen die funktio-
nelle Verknüpfung der Komplexe her (nach Draber et al., 1991).
Im Licht erfolgt ein vektorieller Elektronentransport über die Membran, wobei
Elektronen von PS II auf den Plastochinonpool in der Membran übertragen wer-
den. Dabei findet an der Donorseite von PS II die Oxidation von Wasser zu Sau-
erstoff statt. Das an der Akzeptorseite reduzierte Plastochinon gibt die Elektronen
an den Cytochrom b6/f-Komplex ab. Von dort gelangen sie über das Plastocyanin
zum Photosystem I. PS I reduziert Ferredoxin. Das Ferredoxin überträgt die E-
lektronen auf NADP+. Der Elektronentransport ist mit einem Protonentransport
gekoppelt. Auf diese Weise wird ein Protonengradient über der Membran aufge-
baut, der durch die ATP-Synthase zur Bildung von ATP benutzt wird.
Die photosynthetischen Komplexe bestehen aus sehr hydrophoben transmembra-
nen Proteinuntereinheiten, mehr hydrophilen peripheren Proteinen und einer
bestimmten Anzahl von Cofaktoren.
Das PS II setzt sich aus mehr als 25 unterschiedlichen Proteinen (Barber et al.,
1997) zusammen. Aufgrund der Homologie mit dem Photosystem der Purpurbak-
terien (Trebst, 1986, 1987; Michel und Deisenhofer, 1988) und der kristallographi-

1. Einleitung 7 schen Daten (Holzenburg et al., 1993; Rhee et al., 1997; Rhee et al., 1998; Hanka-
mer et al., 1999) konnten Modelle für das PS II aufgestellt werden (Abb. 1.2.).
Abb. 1.2.: Schematische Darstellung des Photosystems II. Das PS II ist aus folgenden Komponenten
aufgebaut: dem D1-D2-Heterodimer, dem Cyt. b559, den Chlorophyll-bindenden Proteinen CP 43 und
CP 47, den Genprodukten der Gene psbH-N, den peripheren Proteinen des Wasserspaltungskomple-
xes (33, 23 und 16 kDa) und dem Lichtsammelkomplex II (LHC II).
Danach bilden die Proteine D1 und D2 den Kern des Reaktionszentrums von PS
II. Dieser Heterodimer bindet die Cofaktoren: P680 - ob es sich bei P680 um ein
Chlorophyll a-Monomer oder Dimer handelt, ist zur Zeit noch umstritten (Teten-
kin et al., 1989; Shuvalov et al., 1989; Braun et al., 1990; Durrant et al., 1995; No-
guchi et al., 1998)-, vier weitere monomere Chlorophylle, zwei Phaeophytine, ein
durch vier Histidinreste gebundenes Eisen-Ion, ein primäres Plastochinon QA am
D2-Protein und ein sekundäres Plastochinon QB am D1-Protein. Außerdem trägt
es die Bindestelle für den wasserspaltenden Mangankomplex.
PS II fungiert als Wasser-Plastochinon-Oxidoreduktase. Ein Komplex aus den
Proteinen D1 und D2 zusammen mit dem Cytochrom b559 (Nanba und Satoh,

1. Einleitung 8 1987; Barber et al., 1987) sowie einem kleinen psbI-Genprodukt (Tang et al., 1990)
ist bereits in der Lage, die primäre Ladungstrennung durchzuführen.
Die Primärreaktion im PS II-Komplex besteht in der Anregung des P680 durch
Lichtenergie, die von den äußeren Antennen über die inneren Antennen (CP47
und CP43) auf das P680 übertragen wird. Durch die absorbierte Energie tritt in
dem angeregten P680* die primäre Ladungstrennung ein. Das Elektron wird
dann auf ein Phaeophytin übertragen, das seinerseits das primäre Plastochinon
QA zum Semichinon reduziert. Dieses überträgt das Elektron auf den sekundären
Akzeptor QB, der in einer vom D1-Protein gebildeten Nische gebunden ist (Trebst,
1986). QB verläßt nach Aufnahme eines zweiten Elektrons von QA und zweier Pro-
tonen aus dem Stroma als Plastohydrochinon die Bindenische und setzt sich mit
dem freien Plastochinon/Plastohydrochinon-Pool im lipophilen Raum der Memb-
ran ins Gleichgewicht. Das Plastohydrochinon wird dann von dem Cytochrom b6/f-
Komplex oxidiert. Gleichzeitig besetzt ein anderes Plastochinonmolekül die Bin-
denische und wird zu einem neuen QB. Das oxidierte P680+ aus der primären La-
dungstrennung wird von dem Tyrosinrest 161 (TyrZ) im D1 Protein reduziert (De-
bus et al., 1988 a, b, 1992). Das Tyrosinradikal nimmt ein Elektron aus dem Man-
gankomplex auf. Der Mangankomplex oxidiert nach Abgabe von vier Elektronen
zwei Wassermoleküle, wobei ein Sauerstoffmolekül und vier Protonen im Thyla-
koidlumen freigesetzt werden.
Die QB-Bindestelle im D1-Protein ist der spezifische Angriffsort vieler Hemmstoffe
der Photosynthese. Nach einem Modell von Velthuys (1981) konkurrieren diese
Verbindungen mit dem mobilen Plastochinon QB um seine Bindenische und un-
terbinden damit seine Reduktion, was zur Hemmung des Elektronenflusses führt.
Die Hemmstoffe, die teilweise als Herbizide in der Landwirtschaft eingesetzt wer-
den, können aufgrund ihrer Struktur und Wirkung sowie Orientierung in der QB-
Bindenische in zwei Gruppen eingeteilt werden (Trebst, 1987). Zu der ersten
Gruppe, dem sog. DCMU-Typ, gehören Triazine, Harnstoffe, Triazinone, Biscar-
bamate, Cyanoacrylate, Pyrone, Chromone, Cyclohexandione, Hydroxychinole-N-
oxide, Benzochinone und Naphtochinone. Diese Inhibitoren werden auch als „Se-

1. Einleitung 9 rinfamilie“ bezeichnet, da sie sich zum Serin 264 in der Bindenische orientieren.
Zu der zweiten Gruppe, dem sog. Phenol-Typ, gehören Nitrophenole, Azaphe-
nanthrene, Thiazolyliden-Ketonitrile und Hydroxypyridine. Diese Hemmstoffe
bilden die „Histidinfamilie“. Sie sind in der Bindenische zum Histidin 215 hin ori-
entiert. Die Einteilung in diese zwei Gruppen darf aber nicht als absolut angese-
hen werden. So gehören z.B. die Cyanophenole wie Ioxynil keiner der beiden Fa-
milien an. Sie binden wie die Phenole, aber nicht unbedingt in der Nähe von
Histidin 215. Auch die Herbizide innerhalb einer Familie können unterschiedliche
Bereiche der QB-Bindenische besetzen (Oettmeier, 1992). So sind z.B. Triazinone
in zwei Gruppen zu unterteilen (Oettmeier und Hilp, 1991). Diese beiden Gruppen
unterscheiden sich durch ihre Orientierung in der Bindenische, die von den ver-
schiedenen Substituenten des Triazinonringes abhängig ist.
Das D1-Protein wird vom psbA-Gen des plastidären Genoms codiert (Zurawski et
al., 1982) und besitzt ein apparentes Molekulargewicht von 32 kDa. Vergleiche der
Gene von mehr als 40 Organismen (Jansson und Mäenpää, 1997) zeigen eine
starke Konservierung der Aminosäuresequenz und in fast allen Genen die Abwe-
senheit von Introns. Einzige bisher bekannte Ausnahmen stellen die psbA-Gene
in den Grünalgen Chlamydomonas reinhardtii (Erickson et al., 1984) und Euglena
gracilis (Karabin et al., 1984; Keller und Stutz, 1984) dar. Bei Chlamydomonas
reinhardtii ist das psbA-Gen in der inverted repeat-Region des Plastoms zu finden
und besteht aus 5 Exons, die von 4 Introns getrennt sind. Die Aminosäuresequenz
des D1-Proteins zeigt auffällige Homologien zu der Sequenz der L-Untereinheit
des Reaktionszentrums der Purpurbakterien, dessen Kristallstruktur bestimmt
wurde (Deisenhofer et al., 1985). Dies bildete die Basis für die Identifizierung des
D1-Proteins als Kernprotein des PS II-Reaktionszentrums (Michel und Deisenho-
fer, 1988).
Das D1-Protein konnte mit Hilfe von radioaktiv-markiertem Azidoatrazin (Pfister
et al., 1981) bzw. einer Triazin-resistenten Mutante von Amaranthus hybridus
(Hirschberg und McIntosh, 1983) als Herbizidbindeprotein identifiziert werden.
Sowohl der Einsatz radioaktiv-markierter Herbizide als auch die Analyse herbi-
zidtoleranter Mutanten ermöglicht nach wie vor eine strukturelle und funktionel-

1. Einleitung 10 le Charakterisierung des D1-Proteins. Die radioaktiv-markierten Herbizide wer-
den für Bindetests angewendet. Dabei wird ein radioaktiver Hemmstoff vorgege-
ben. Anschließend wird die Radioaktivität durch eine weitere Verbindung ver-
drängt. Dadurch lassen sich die Bindungsaffinitäten der Herbizide zum D1-
Protein bestimmen (Tischer und Strotmann, 1977). Ein anderes Verfahren ist die
Photoaffinitätsmarkierung. Diese Technik führt eine kovalente Bindung der Her-
bizide an das D1-Protein herbei. Bei Belichtung eines radioaktiven Azidoderivates
spaltet das Azid in ein sehr reaktives Nitren, das sich sofort an ein Atom im Prote-
in anbindet. Nach Proteaseverdauung des D1-Proteins und anschließender Prote-
insequenzierung kann festgestellt werden, an welche Aminosäure das Azidoderi-
vat kovalent gebunden ist. Photoaffinitätsmarkierungen und Verdrängungsver-
suche erlauben Aussagen über Strukturmerkmale der QB-Bindenische und über
die Bindungsstellen der Herbizide. Eine weitere Methode, mit deren Hilfe die
Struktur und Funktion des Herbizidbindeproteins untersucht werden kann, ist die
Mutantenanalyse. Die bisher einzige in freier Natur gefundene Mutation betrifft
Serin 264. Sie trat in höheren Pflanzen auf regelmäßig mit Atrazin behandelten
Flächen auf und führte zur Resistenz gegenüber dieser Substanz. Durch gezielte
Mutagenese wurden weitere zahlreiche Mutationen in Cyanobakterien, einzelli-
gen Algen und in Zellkulturen erzeugt. Die herbeigeführten Aminosäureaustau-
sche verändern die Bindungsaffinitäten der Herbizide in den Mutanten. Sie füh-
ren zu Resistenzen, Kreuzresistenzen bzw. negativen Kreuzresistenzen. Eine Mu-
tation kann die Bindungsaffinität eines Hemmstoffs verringern und damit der
Mutante eine Resistenz gegenüber dem Hemmstoff verleihen. Eine Mutation
kann auch Kreuzresistenzen verursachen. In diesem Fall ist die Mutante resistent
gegenüber verschiedenen Hemmstoffen, die zu verschiedenen Klassen gehören
können. Schließlich kann eine Mutation die Bindungsaffinität eines Inhibitors
erhöhen, was zur Supersensitivität führt. Wenn eine Mutation Resistenz gegen-
über einer Klasse von Hemmstoffen hervorruft und gleichzeitig Supersensitivität
gegenüber einer anderen Klasse von Hemmstoffen verursacht, wird das Phäno-
men als negative Kreuzresistenz bezeichnet. Das veränderte Verhalten der Mut-
anten gegenüber den Herbiziden liefert Informationen über die an der Herbizid-
bindung beteiligten Aminosäuren, über die Architektur der QB-Bindenische und

1. Einleitung 11 über die Orientierung der Herbizide in der Bindenische. Die bisher charakterisier-
ten zu Herbizidtoleranz bzw. Supersensitivität führenden Mutationen konzentrie-
ren sich auf dem Bereich zwischen den Aminosäuren Phe 211 und Leu 275 im D1-
Protein. Dieser Bereich umfaßt einen Teil der transmembranen Helix D, die paral-
lele Helix DE, einen Teil der transmembranen Helix E und sie verbindende
Schleifen.
Das Ziel der vorliegenden Dissertation ist die Charakterisierung der QB-
Bindenische im PS II von Chlamydomonas reinhardtii. Hierzu sollten durch Oli-
gonukleotid-gerichtete Mutagenese des psbA-Gens Mutanten mit Aminosäuresub-
stitutionen im D1-Protein erzeugt werden und hinsichtlich der Bindungsaffinitä-
ten verschiedener Inhibitoren untersucht werden. Folgende Aminosäuren im Be-
reich der QB-Bindenische sollten ausgetauscht werden: Val 249, His 215 und Phe
206. Val 249 befindet sich nach den Modellen für die Faltung des D1-Proteins am
Anfang der parallel zur Thylakoidmembran verlaufenden Helix DE und wurde als
Bindungsort von Azidoioxynil, einem Cyanophenol, erkannt (Oettmeier, 1989).
His 215 und Phe 206 liegen in der transmembranen Helix D. His 215 ist an der
Bindung des Eisen-Ions beteiligt. Der Austausch von Phe 206 ist bereits bei einer
Dreifachmutante bekannt (Orawski, 1998).
Ein weiteres Ziel dieser Arbeit war Doppelmutanten mit Aminosäureaustauschen
an Positionen Phe 211 und Met 214 zu charakterisieren. Diese Aminosäuren be-
finden sich ebenfalls in der transmembranen Helix D. Der Austausch von Phe 211
ist bereits bei Cyanobakterien bekannt (Gingrich et al., 1988; Kirilovsky et al.,
1989; Govindjee et al., 1990). Met 214 wurde durch Photoaffinitätsmarkierung als
Bindungsort von Azidoatrazin erkannt (Wolber et al., 1986; Whitelegge et al.,
1992).
Die Mutantenanalyse sollte zum Verständnis der Bedeutung der substituierten
Aminosäuren für die QB- und Herbizid-Bindung im PS II von Chlamydomonas
reinhardtii beitragen. Darüber hinaus sollte sie Aussagen über den Mechanismus
der Hemmwirkung untersuchten Substanzen erlauben.

2. Material und Methoden 12
2. Material und Methoden
2.1. Chemikalien, Enzyme und Verbrauchsmaterialien
Alle verwendeten Chemikalien stammten von den Firmen Baker, Biomol, Biorad,
Biozym, Difco, Fluka, Merck, Riedel de Haen, Serva bzw. Sigma und lagen in Ana-
lysequalität vor.
Die Enzyme wurden von den Firmen Gene Craft, MBI Fermentas, Peqlab und Ro-
che bezogen.
Der Molekulargewichtsmarker für Proteine (10 kDa-Leiter) stammt von der Firma
Gibco BRL.
Der DNA-Größenstandart (λ-DNA; HindIII/EcoRI geschnitten) stammt von der
Firma MBI Fermentas.
Antibiotika wurden von den Firmen Sigma und ICN Biomedicals Inc. bezogen.
Der Transfer von Proteinen erfolgte auf Nitrozellulosefolie BA-S 85 der Firma
Schleicher & Schuell in einer Naßblotkammer (Trans Blot Cell) der Firma Biorad.
Für die Transformation wurden die Algenzellen auf Membranfilter NL 17 (0,45
µm) der Firma Schleicher & Schuell gesaugt.
Die bei der Transformation der Algenzellen verwendete Treibladung (Spit T. I. A.)
stammt von der Firma Spit.
Für die Extraktion der PCR-Produkte aus Agarosegelen wurde der QIAquick Gel
Extraction Kit der Firma Qiagen, JETQUICK Gel Extraction Spin Kit der Firma
Genomed bzw. NucleoSpin Extract der Firma Macherey-Nagel verwendet.
Alle hitzebeständigen Lösungen, Glas- und Plastikmaterialien wurden durch Au-
toklavieren (121°C, 1 bar Überdruck, 25 Minuten) sterilisiert.

2. Material und Methoden 13 2.2. Algen- und Bakterienstämme
2.2.1. Algenstämme
Im Rahmen dieser Arbeit wurden folgende Algenstämme von Chlamydomonas
reinhardtii verwendet:
WT11/32-b:
Wildtyp (Sammlung von Algenkulturen, Göttingen).
IL:
Eine im psbA-Gen intronlose Mutante, die aus einer Komplementation der Deleti-
onsmutante FuD7 (Bennoun et al., 1986) mit der cDNA des psbA-Gens hervorge-
gangen ist (Heiß, 1992; Johanningmeier und Heiß, 1993). Die IL-Mutante synthe-
tisiert ein funktionsfähiges D1 Protein und wächst photoautotroph.
Del1:
Eine Deletionsmutante, die aus einer Cotransformation der intronlosen Mutante
IL hervorgegangen ist (Johanningmeier et al., 2000). Diesem Stamm fehlt die co-
dierende Sequenz für die Aminosäuren Alanin 152 bis Alanin 294 im D1 Protein.
Durch die Deletion wird der Leserahmen zerstört. Diese Mutante ist nicht zu pho-
toautotrophen Wachstum fähig, so daß sie auf Acetat als Kohlenstoffquelle ange-
wiesen ist. Sie ist resistent gegen Spectinomycin.
2.2.2. Bakterienstamm
Zur Vermehrung von Plasmid-DNA wurde der Escherichia coli-Stamm XL-1-Blue
(Stratagene) verwendet:
MRF’: ∆(mcrA)183, ∆(mcrCB-hsdSMR-mrr)173, endA1, supE44,
thi-1, recA1, relA1, gyrA96, lac-,
F’[proAB, lacIq, lacZ∆M15, Tn10(tetr)]

2. Material und Methoden 14 2.3. Algenanzucht
2.3.1. Anzuchtbedingungen
Die Anzucht der verschiedenen Algenstämme erfolgte in 100 ml Erlenmeyerkol-
ben auf einem Schütteltisch (60 rpm) unter permanenter Beleuchtung von 7000
Lux bei 18°C oder in Algenanzuchtröhren mit Begasung von 95% Luft und 5%
CO2 bei 7000 Lux Dauerlicht und 18°C. Die Zellen wuchsen entweder photoau-
totroph in HS (high-salt)-Medium (Sueoka, 1960) oder heterotroph in TAP (Tris-
Acetat-Phosphat)-Medium (Gorman und Levine, 1965). Stammkulturen wurden
auf HS- und TAP-Platten gehalten, bei denen die Medien mit 1,5%-igem Agar ver-
setzt sind. Die Anzucht der Deletionsmutante Del1 erfolgte in Spectinomycin-
haltigem TAP-Medium (100 mg Spectinomycin/1l Medium). Die Stammkultur der
Deletionsmutante Del1 wurde auf TAP-Medium gehalten, das mit 1,5%-igem Agar
versetzt war und Spectinomycin enthielt (100 mg Spectinomycin/1l Medium).
2.3.2. Medien
HS-Medium: Stocklsg. 5 ml
Phosphatlsg. 5 ml
Spurenelementelsg. (nach Hutner) 1 ml
A. dest. ad 1 l
TAP -Medium: Tris 2,42 g
Spurenelementelsg. (nach Hutner) 1 ml
Salzlsg. 50 ml
1 M Kaliumphosphat-Puffer; pH 7,2 1 ml
mit Essigsäure auf pH 7,2 eingestellt
A. dest. ad 1 l
Salzlsg.: MgSO4∗7H2O 2 g

2. Material und Methoden 15 NH4Cl 8 g
CaCl2∗2H2O 1 g
A. dest. ad 1 l
Stocklsg.: NH4Cl 100 g
MgSO4∗7H2O 4 g
CaCl2∗2H2O 2 g
A. dest. ad 1 l
Phosphatlsg.: K2HPO4 288 g
KH2PO4 144 g
A. dest. ad 1 l
Spurenelementelsg. nach Hutner : EDTA 50 g
(Harris, 1988) ZnSO4∗7H2O 22 g
H3BO3 11,4 g
MnCl2∗4H2O 5,06 g
FeSO4∗7H2O 4,98 g
CaCl2∗6H2O 1,61 g
CuSO4∗5H2O 1,57 g
(NH4)6Mo7O24 1,1 g
A. dest. ad 1 l
2.4. Bakterienanzucht
Die Anzucht von Bakterienzellen in kleinem Maßstab erfolgte in Flüssigkulturen
in sterilen Grainer-Röhren mit ca. 2 ml LB (Luria-Bertani)-Medium (Sambrook et

2. Material und Methoden 16 al., 1989) in einem Inkubationsschüttler (New Brunswick Scientific, Edison, New
Jersey) bei 37°C und 200 - 250 rpm über Nacht. Die Kulturen wurden mit Hilfe
einer sterilen Pipettenspitze von einer Einzelkolonie auf Agarplatten oder mit 50
µl einer Stammkultur angeimpft.
Bei größeren Kulturvolumina von ca. 10 - 100 ml wurden zur Anzucht sterile Er-
lenmeyerkolben verwendet, die mit einer über Nacht gewachsenen 2 ml Vorkultur
1%-ig angeimpft wurden. Die Anzucht des Bakterienstamms, der über das Plasmid
pSHc5 mit Ampicillinresistenz verfügte, erfolgte in Flüssigmedium, dem das Anti-
biotikum zugesetzt wurde.
Um Einzelkolonien zu erhalten, wurden die Bakterien mit einer Impföse oder ei-
ner gebogenen Pasteurpipette auf einer Festagarplatte ausgestrichen und 16 - 18
h bei 37°C bebrütet.
Zur Herstellung von Stammkulturen wurden 0,5 ml einer Übernachtkultur mit
0,5 ml 60% (v/v) Glycerin versetzt und bei -80°C gelagert.
LB-Medium: NaCl 10 g
Bacto-Pepton 10 g
Hefe-Extrakt 5 g
A. dest ad 1 l
mit 2 N NaOH auf pH 7,5 eingestellt
LB-Agar: 1,5% (w/v) Bacto-Agar in LB-Medium
Selektivmedium: Ampicillin 100 mg
Tetracyclin 10 mg
LB-Medium ad 1 l

2. Material und Methoden 17 2.5. Oligonukleotide
Die als Primer verwendeten Oligonukleotide wurden von der Firma MWG-Biotech
AG synthetisiert.
2.5.1. Oligonukleotide für die ortsgerichtete Mutagenese
Folgende Oligonukleotide wurden als Primer für die ortsgerichtete Mutagenese
verwendet:
opsbA -28
5’-CGGAGAAATTAAAACTTTAAAAAAATTAACATATGACAGC-3’
(40 Nukleotide)
Komplementäre Sequenz zum codogenen Strang des psbA-Gens im
5’flankierenden Bereich. Das Startcodon ist fett und kursiv gedruckt.
opsbA stop
5’-CCAGATTTATTTTAGTTTAAAAAAAAATTAGTTG-3’
(34 Nukleotide)
Komplementäre Sequenz zum nicht-codogenen Strang des psbA-Gens im
3’flankierenden Bereich. Das Stopcodon ist fett und kursiv gedruckt.
opsbA 206
5’-AGAATAATGAACCACCNNCTACACCAGCAACACCT-3’ G T
(35 Nukleotide)
Komplementäre Sequenz zum nicht-codogenen Strang des psbA-Gens mit 3 ge-
wobbelten Nukleotiden. Fett gedruckter Bereiche kennzeichnet die gewobbelten
Nukleotide.
opsbA 215
5’-AAGTAACTAAAGAACCNNACATAGCTGAGAATAAT-3’ C T
(35 Nukleotide)

2. Material und Methoden 18 Komplementäre Sequenz zum nicht-codogenen Strang des psbA-Gens mit 3 ge-
wobbelten Nukleotiden. Die gewobbelten Nukleotide sind fett gedruckt.
opsbA 249
5’-AGTAACCATGAGCAGCNNNAATGTTGTAAGTTTCT-3’
(35 Nukleotide)
Komplementäre Sequenz zum nicht-codogenen Strang des psbA-Gens mit 3 ge-
wobbelten Nukleotiden. Die gewobbelten Nukleotide sind fett gedruckt.
opsbA 215Gln
5’-AAGTAACTAAAGAACCTTGCATAGCTGAGAATAAT-3’
(35 Nukleotide)
Komplementäre Sequenz zum nicht-codogenen Strang des psbA-Gens mit 2 Ba-
senaustauschen. Die Basenaustausche sind fett gedruckt.
opsbA 215Ile
5’-AAGTAACTAAAGAACCGATCATAGCTGAGAATAAT-3’
(35 Nukleotide)
Komplementäre Sequenz zum nicht-codogenen Strang des psbA-Gens mit 2 Ba-
senaustauschen. Die Basenaustausche sind fett gedruckt.
opsbA 215Phe
5’-AAGTAACTAAAGAACCGAACATAGCTGAGAATAAT-3’
(35 Nukleotide)
Komplementäre Sequenz zum nicht-codogenen Strang des psbA-Gens mit 2 Ba-
senaustauschen. Die Basenaustausche sind fett gedruckt.
2.5.2. Oligonukleotide für die DNA-Sequenzierung
Folgende Oligonukleotide wurden als Primer bei den Sequenzierreaktionen einge-
setzt:

2. Material und Methoden 19 opsbA 412
5’-GGTATGCGTCCATGGATCGC-3’
(20 Nukleotide)
Komplementäre Sequenz zum codogenen Strang des psbA-Gens im Bereich von
Exon III.
opsbA 928
5’-GTGAGTCTACTACTGATTGG-3’
(20 Nukleotide)
Komplementäre Sequenz zum nicht-codogenen Strang des psbA-Gens im Bereich
von Exon V.
2.6. Plasmid
Als Matrize in den Polymerase-Kettenreaktionen der ortsgerichteten Mutagenese
diente der Vektor pSHc5 (Heiß, 1992; Johanningmeier und Heiß, 1993). Dieses
Plasmid enthält das intronlose psbA-Gen und ca. 1,5 kb 3’flankierende Sequenz im
Bluescript Vektor pBLUESKRIPT-II-SK(+). Der Vektor pSHc5 ist ein Derivat des
Plasmids pSHc1 (Heiß, 1992). pSHc1 entstand durch Klonierung der cDNA des
psbA-Gens in einen EcoRV-geschnittenen pBLUESKRIPT-II-SK(+)-Vektor. In das
Plasmid pSHc1 wurde anschließend ein 1,5 kb BsmI/BamHI-Fragment kloniert,
das einen Teil des Exons V des psbA-Gens und 3’flankierenden Bereich umfaßt.
2.7. Molekularbiologische Methoden
2.7.1. Gerichtete Mutagenese mit Anwendung der Polymerase-Kettenreaktion
Die ortsspezifische Mutagenese mit Hilfe der PCR wurde nach der Methode von
Landt et al. (1990) durchgeführt. Ausgehend von doppelsträngiger DNA als Mat-
rize erfordert diese Methode ein spezifisches mutagenes Oligonukleotid als Primer
sowie 2 Primer, die die zu mutierende Region in 3’- und 5’-Richtung flankieren.
Die Mutagenese erfolgte in 2 Schritten (Abb.2.1.).

2. Material und Methoden 20
Abb. 2.1.: Schematische Darstellung der ortsgerichteten Mutagenese.
Als Matrize diente der Vektor pSHc5 (Heiß, 1992; Johanningmeier und Heiß,
1993). Dieses Plasmid enthält das intronlose psbA-Gen und ca. 1,5 kb
3’flankierende Sequenz im Bluescript Vektor pBSSK(+) (vgl. Kap. 2.6.). Im 1.
Schritt wurde mit dem mutierten Primer A (opsbA 206, opsbA 215, opsbA 249,
opsbA 215Gln, opsbA 215Ile, bzw. opsbA 215Phe; vgl. Kap. 2.5.1.) und dem Primer

2. Material und Methoden 21 B (opsbA -28) ein doppelsträngiges Fragment erzeugt, das die Basenaustausche
enthält. Dieses 1. PCR-Produkt wurde durch Elektrophorese in einem 0,8%-igen
(w/v) Agarosegel aufgetrennt und durch daran anschließende Elution aus dem Gel
gereinigt (vgl. Kap. 2.7.3.). Im 2. PCR-Schritt diente der Einzelstrang des gerei-
nigten Fragments, der die Basenaustausche an seinem 3’-Ende trug, als Primer.
Als 2. Primer bei dieser Reaktion diente Primer C (opsbA stop). Das Produkt der 2.
PCR wurde nach der Reinigung über ein 0,8%-iges Agarosegel als Matrize für ei-
ne 3. PCR eingesetzt. In der 3. PCR wurde das Produkt der Mutagenese amplifi-
ziert.
Als Kontrolle wurde eine PCR mit dem Vektor pSHc5 als Vorlage und den flankie-
renden Oligonukleotiden B (opsbA -28) und C (opsbA stop) als Primer durchge-
führt.
Alle PCR-Reaktionen der Mutagenese wurden mit Hilfe eines von Fischer-
Technik konstruierten Roboters, der von einem PC mit entsprechender Software
gesteuert wurde, in drei unterschiedlich temperierten Wasserbädern durchge-
führt.
1. PCR:
Die Durchführung der 1. PCR erfolgte nach folgendem Ansatz:
10 µl Reaktionspuffer (10fach)
10 µl dNTPs (2 mM)
10 µl Primer B (5 µM)
10 µl Primer A (5 µM)
1 µl pSHc5 (100 ng/ µl)
6 µl MgSO4 (25 mM)
2,5 U Pwo-DNA-Polymerase
ad 100 µl mit A. dest.
Die gewählten Bedingungen sind in der Tabelle 2.1. aufgeführt.

2. Material und Methoden 22
Zyklus-Nummer 1 2-34 35
Denaturierung 92°C 3 1 1
Primer-Anbindung 40°C 1 1 1
Elongation 72°C 1,5 1,5 3
Tab. 2.1.: Bedingungen der 1. PCR. Die Zykluszeiten sind in Minuten angegeben.
2. PCR:
Die Durchführung der 2. PCR erfolgte nach folgendem Ansatz:
10 µl Reaktionspuffer (10fach)
10 µl dNTPs (2 mM)
10 µl Primer C (5 µM)
variable Mengen des Produktes der ersten PCR (1/10 bis 1/2 der gereinigten DNA)
2 µl pSHc5 (10 ng/ µl)
10 µl MgSO4 (25 mM)
5 µl DMSO
2,5 U Pwo-DNA-Polymerase
ad 100 µl mit A. dest.
Die gewählten Bedingungen sind in der Tabelle 2.1. aufgeführt.
Zyklus-Nummer 1 2-29 30
Denaturierung 92°C 3 2 2
Primer-Anbindung 40°C 1,5 1,5 1,5
Elongation 72°C 1 1 3
Tab. 2.2.: Bedingungen der 2. PCR. Die Zykluszeiten sind in Minuten angegeben.

2. Material und Methoden 23 3. PCR:
Die Durchführung der 3. PCR erfolgte nach folgendem Ansatz:
10 µl Taq-DNA-Polymerase-Puffer (10fach)
10 µl dNTPs (2 mM)
10 µl opsbA -28 (5 µM)
10 µl opsbA stop (5 µM)
100 ng Produkt der zweiten PCR
2 U Taq-Polymerase
ad 100 µl mit A. dest.
Die gewählten Bedingungen sind in der Tabelle 2.3. aufgeführt.
Zyklus-Nummer 1 2-34 35
Denaturierung 92°C 3 1 1
Primer-Anbindung 55°C 1 1 1
Elongation 72°C 2 2 3
Tab. 2.3.: Bedingungen der 3. PCR. Die Zykluszeiten sind in Minuten angegeben.
Kontroll-PCR:
Die Durchführung der Kontroll-PCR erfolgte nach folgendem Ansatz:
10 µl Taq-DNA-Polymerase-Puffer (10fach)
10 µl dNTPs (2 mM)
10 µl opsbA -28 (5 µM)
10 µl opsbA stop (5 µM)
1 µl pSHc5 (50 ng/µl)
2 U Taq-Polymerase
ad 100 µl mit A. dest.

2. Material und Methoden 24 Die gewählten Bedingungen sind in der Tabelle 2.4. aufgeführt.
Zyklus-Nummer 1 2-34 35
Denaturierung 92°C 3 1 1
Primer-Anbindung 55°C 1 1 1
Elongation 72°C 2 2 3
Tab. 2.4.: Bedingungen der Kontroll-PCR. Die Zykluszeiten sind in Minuten angegeben.
2.7.2. Polymerase-Kettenreaktionen mit Algen-DNA
Die aus den Algen isolierte Gesamt-DNA (vgl. Kap. 2.7.5.) wurde als Matrize bei
den PCR-Reaktionen eingesetzt.
Die Durchführung der PCR erfolgte nach folgendem Ansatz:
10 µl Taq-DNA-Polymerase-Puffer (10fach)
10 µl dNTPs (2 mM)
10 µl opsbA -28 (5 µM)
10 µl opsbA stop (5 µM)
100 ng Gesamt-DNA aus Chlamydomonas reinhardtii
2 U Taq-Polymerase
ad 100 µl mit A. dest.
Die gewählten Bedingungen sind in der Tabelle 2.5. aufgeführt.

2. Material und Methoden 25
Zyklus-Nummer 1 2-34 35
Denaturierung 92°C 3 1 1
Primer-Anbindung 55°C 1 1 1
Elongation 72°C 2 2 3
Tab. 2.5.: Die Bedingungen der PCR mit Algen-DNA als Matrize. Die Zykluszeiten sind in Minuten
angegeben.
2.7.3. Agarose-Gelelektrophorese zur Auftrennung von DNA
Die Auftrennung von PCR-Produkten erfolgte in einem Spannungsfeld mittels
Agarose-Gelelektrophorese (Sambrook et al., 1989).
Für analytische und präparative Gele wurden 0,8% (w/v) Agarose in 1 x TAE-
Puffer (s. u.) durch Erhitzen gelöst und mit 2,5 µl Ethidiumbromidlösung (10
mg/ml in H2O) pro 100 ml flüssigem Gel versetzt. Die Auftrennung der DNA-
Fragmente erfolgte für ca. 1 h bei 60 - 80 V mit dem entsprechenden Laufpuffer (1
x TAE) und 6 x DNA-Probenpuffer. Der Größenvergleich erfolgte mit dem DNA-
Größenstandart (λ-DNA; HindIII/EcoRI geschnitten).
Die im Gel mit Ethidiumbromid assoziierte DNA konnte mit Hilfe von UV-Licht
(312 nm) sichtbar gemacht werden.
Die Elution der PCR-Produkte aus Agarosegelen erfolgte mit Gel-Extraction-Kit
(vgl. Kap. 2.1.) nach den Angaben des Herstellers.
TAE-Puffer: 0,04 M Tris-Acetat
0,001 M EDTA; pH 8,0
DNA-Probenpuffer (6x): 0,25% (w/v) Bromphenolblau
0,25% (w/v) Xylencyanol
30% (w/v) Glycerin

2. Material und Methoden 26 2.7.4. DNA-Sequenzierung
Die Sequenzierungen wurden von der Firma Sequence Laboratories in Göttingen
durchgeführt. Sequenziert wurden PCR-Fragmente (Produkte der Mutagenese
bzw. Produkte der PCR mit Algen-DNA als Matrize ). Folgende Oligonukleotide
wurden als Primer bei den Sequenzierreaktionen eingesetzt: opsbA 412, opsbA
928 (vgl. Kap. 2.5.2.).
2.7.5. Isolierung von Gesamt-DNA aus Chlamydomonas reinhardtii
Zur Extraktion der Gesamt-DNA aus Chlamydomonas reinhardtii wurde die Me-
thode von Newman et al. (1990) in geringfügig modifizierter Form angewandt.
Die Zellen einer 50 ml Kultur wurden durch 5-minütige Zentrifugation bei 4°C
und 5000 rpm (Sorvall-Zentrifuge RC-5B, Du Pont Instruments; SS 34-Rotor,
Kontron) sedimentiert. Das Algenpellet wurde in 1 ml TEN-Puffer (s. u.) aufge-
nommen. Anschließend wurden die Zellen 30 s bei 13000 rpm in einer Eppendorf-
zentrifuge (5415 C, Sigma) sedimentiert und in 400 µl TEN resuspendiert. Die Ly-
se der Zellen erfolgte nach Zugabe von 40 µl 20% (w/v) Sarkosyl, 40 µl 20% (w/v)
SDS und 30 µl Pronase-Lösung (s. u.) während einer 10-minütigen Inkubation un-
ter leichtem Schütteln bei Raumtemperatur. Die anschließende Zugabe von 0,65
ml PCI (s. u.) diente der Trennung der Nukleinsäuren von anderen Zellbestand-
teilen. Der Ansatz wurde 5 min bei Raumtemperatur auf einem Vortexgerät (Reax
2000, Omnilab) mild geschüttelt, dann 5 min bei 13000 rpm in der Eppendorfzen-
trifuge zentrifugiert. Die obere wäßrige Phase wurde abgenommen. Die Extrakti-
on der Nukleinsäuren mit PCI wurde wiederholt. Die Zugabe von 2 Volumina
100% Ethanol führte bei – 80°C nach 1-2 h zur Fällung der DNA. Durch eine 10-
minütige Zentrifugation bei 13000 rpm und 4°C konnte die DNA sedimentiert
werden. Das DNA-Pellet wurde zweimal mit 70%-igem Ethanol gewaschen, kurz
angetrocknet und in 0,3 ml TE-Puffer (s. u.) gelöst. Nach Zugabe von 30 µl 3 M
Ammonium-Acetat und 30 µl 0,5 M MgCl2 folgte eine 2. Ethanolfällung. Nach 2
Waschschritten mit 70%-igem Ethanol wurde die DNA in der Speed Vac (Speed
Vac DNA 110, Savant) getrocknet und anschließend in 50 µl TE-Puffer aufge-
nommen. Zur Bestimmung der DNA-Konzentration wurde die Extinktion bei 260
nm gemessen.

2. Material und Methoden 27 TEN-Puffer: 150 mM NaCl
10 mM EDTA
10 mM Tris-HCl; pH 8,0
TE-Puffer: 10 mM Tris-HCl; pH 8,0
1 mM EDTA
Pronase-Lsg.: 1 g Pronase
11 mg CaCl2
1 ml Tris-HCl; pH 7,5
ad 5 ml mit A. dest.
PCI: Phenol/Chloroform/Isoamylalkohol im Verhältnis 25:24:1
2.7.6. Herstellung kompetenter Bakterienzellen
Die Präparation kompetenter Zellen des Escherichia coli-Stamms XL-1-Blue MRF’
(vgl. Kap. 2.2.2.) erfolgte nach der RbCl2-Methode von Hanahan (Protokoll 3; Ha-
nahan, 1985). Hierzu wurden Bakterienzellen der logarithmischen Wachstums-
phase (OD595 = 0,5 - 0,8) verwendet. Die kompetenten Zellen konnten entweder di-
rekt zur Transformation eingesetzt oder bei -80°C bis zu ihrem Gebrauch gelagert
werden1.
2.7.7. Transformation kompetenter Bakterienzellen
Zur Vermehrung des pSHc5-Plasmids (ein Derivat des Bluescript Vektors
pBSSK(+), das das intronlose psbA-Gen und ca. 1,5 kb 3’flankierende Sequenz
1 Die kompetenten Zellen wurden mir freundlicherweise von A. Wiesmann (Lehrstuhl für Biochemie der Pflanzen, Prof. Dr. W. Oettmeier, Ruhr-Universität Bochum) für meine Arbeit zur Verfügung gestellt.

2. Material und Methoden 28 enthält; vgl. Kap. 2.6.) wurde der Escherichia coli-Stamm XL-1-Blue (vgl. Kap.
2.2.2.) verwendet. Zur Transformation kompetenter Bakterienzellen wurde die
Methode nach Sambrook (Sambrook et al., 1989) eingesetzt. 200 µl kompetente
Zellen wurden mit ca. 100 ng Plasmid-DNA vorsichtig gemischt und anschließend
für 30 min auf Eis inkubiert. Nach einem 90-sekündigen Hitzeschock bei 42°C in
einem Wasserbad wurden die Zellen sofort auf 0°C abgekühlt. Nach Zugabe von
800 µl LB-Medium (Sambrook et al., 1989; vgl. Kap. 2.4.) und 1 h Inkubation bei
37°C wurden die Bakterien durch Zentrifugation (Eppendorfzentrifuge, 4 min,
4000 rpm, Raumtemperatur) sedimentiert. Der erhaltene Überstand wurde grob
abgegossen, so daß mit dem restlichen Medium die Zellen resuspendiert werden
konnten. Im Anschluß wurden die Zellen auf Selektivmedium ausplattiert.
2.7.8. Präparation von Plasmid-DNA
Die Präparation von Plasmid-DNA wurde mit Hilfe des Plasmid Purification Kit
der Firma Qiagen nach Angaben des Herstellers durchgeführt.
2.7.9. Transformation des Chloroplasten von Chlamydomonas reinhard-tii
Die Transformation des Chloroplasten von Chlamydomonas reinhardtii erfolgte
mit Hilfe der Partikelkanone, die von Klein und Sanford (1987) entwickelt wurde.
Bei dieser Methode wird die in den Chloroplasten einzubringende DNA auf Wolf-
rampartikel präzipitiert und durch eine Treibladung beschleunigt.
Der zu transformierende Stamm Del1 (vgl. Kap. 2.2.1.) wurde in TAP-Medium
(vgl. Kap. 2.3.2.) angezogen. Jeweils 1 bis 2×107 Zellen wurden über eine Glasfritte
mit leichtem Unterdruck auf einen Nylonfilter gesaugt. Die Filter wurden auf
TAP-Agarplatten (vgl. Kap. 2.3.2.) übertragen und über Nacht bei Raumtempera-
tur im Dunkeln gehalten.
Zur Transformation wurden Wolframpartikel mit einem Durchmesser von 1,3 µm
eingesetzt. 60 mg von diesen Partikeln wurden zuvor in 1 ml 100% Ethanol auf-
genommen und durch eine 3-minütige Ultraschallbehandlung (Lehfeldt) fein

2. Material und Methoden 29 dispergiert. Nach dreimaligem Waschen mit sterilem A. dest. wurden die Partikel
schließlich in 1 ml A. dest. resuspendiert.
Folgende Fällungsansätze wurden für jeweils 5 Schüsse bei der Transformation
mit den PCR-Produkten der Mutagenese mit den mutagenen Primern opsbA 206,
opsbA 215 bzw. opsbA 249 angesetzt:
25 µl Wolframpartikel-Suspension (s. o.)
5 µl eluiertes Produkt der 3. PCR; dies ent-
spricht 1500 ng DNA
25 µl CaCl2 (2,5 M)
10 µl Spermidin (0,1 M)
Folgende Fällungsansätze wurden für jeweils 1 Schuß bei der Transformation mit
den PCR-Produkten der Mutagenese mit den mutagenen Primern opsbA 215Gln,
opsbA 215Ile bzw. opsbA 215Phe angesetzt:
7 µl Wolframpartikel-Suspension
2 µl eluiertes Produkt der 3. PCR; dies ent-
spricht 2 µg DNA
7,4 µl CaCl2 (2,5 M)
2,8 µl Spermidin (0,1 M)
Folgende Fällungsansätze wurden für jeweils 1 Schuß bei der Transformation mit
dem Produkt der Kontroll-PCR angesetzt:
5 µl Wolframpartikel-Suspension
1 µl eluiertes Produkt der Kontroll-PCR; dies ent-
spricht 300 ng DNA
5 µl CaCl2 (2,5 M)
2 µl Spermidin (0,1 M)

2. Material und Methoden 30 Die Ansätze wurden jeweils für 10 min bei Raumtemperatur inkubiert. Nach dem
Sedimentieren der Partikel wurde der Überstand (52,5 µl, 16,7 µl bzw. 10,5 µl)
verworfen, der restliche Ansatz resuspendiert. Pro Schuß wurden 2,5 µl in die
Bohrung des dafür vorgesehenen Trägers eingefüllt. Durch Beschleunigung mit-
tels einer Treibladung und Verteilung der einzelnen Partikel mit Hilfe einer
Blende wurde die Suspension auf die vorbereiteten Platten mit Chlamydomonas
reinhardtii-Zellen (s. o.) geschossen. Nach Inkubation der transformierten Zellen
über Nacht im Dunkeln bei Raumtemperatur wurden die Filter auf HS-
Agarplatten (vgl. Kap. 2.3.2.) übertragen und bei 7000 Lux im Dauerlicht bei
Raumtemperatur gehalten. Transformierte Kolonien wurden nach etwa 4 Wochen
mit bloßem Auge sichtbar.
2.8. Präparation von Thylakoidmembranen aus Chlamydomonas rein-hardtii
Die Thylakoidmembranen wurden nach der leicht modifizierten Methode von
Chua und Bennoun (1975) aus Chlamydomonas reinhardtii isoliert.
200 ml Algenkultur der spätlogarithmischen Phase wurden durch 5-minütige
Zentrifugation bei 4°C und 5000 rpm (Sorvall-Zentrifuge RC-5B, Du Pont In-
struments; GSA-Rotor, Kontron) sedimentiert. Das Pellet wurde in Lösung A (s.
u.) resuspendiert, so daß 1/10 des ursprünglichen Volumens erreicht wurde. Die
Zellen wurden erneut 5 min bei 4°C und 5000 rpm (Sorvall-Zentrifuge RC-5B; SS
34-Rotor, Kontron) sedimentiert. Das Pellet wurde in Lösung B (s. u.) aufgenom-
men und auf ca. 0,3 mg Chlorophyll/ml eingestellt. Anschließend wurden die Zel-
len mit Hilfe der Yeda-Presse bei einem Druck von ca. 40 atm und einer Durch-
flußgeschwindigkeit von ca. 1 ml/s aufgebrochen. Durch eine 5-minütige Zentrifu-
gation bei 4°C und 5000 rpm (Sorvall-Zentrifuge RC-5B; SS 34-Rotor) wurden die
Thylakoide sedimentiert. Das Pellet wurde in Lösung B resuspendiert und auf et-
wa 1 mg Chlorophyll/ml eingestellt.
Lösung A: 20 mM HEPES-NaOH; pH 7,0
2,5 mM MgCl2
50 mM KCl

2. Material und Methoden 31 Lösung B: 20 mM HEPES-NaOH; pH 7,5
5 mM MgCl2
1 mM MnCl2
2 mM KCl
0,3 M Sorbit
2.9. Isolierung von Protein aus Chlamydomonas reinhardtii für die SDS-Polyacrylamid-Gelelektrophorese
Zur Proteinisolierung wurden Thylakoide nach der unter Punkt 2.8. beschriebe-
nen Methode präpariert. Die Thylakoide wurden nach der letzten Zentrifugation
statt in Lösung B in 0,1 M Na2CO3 Lösung aufgenommen, so daß eine Chloro-
phyllendkonzentration von 0,5 mg Chlorophyll in 1 ml vorlag. Zur Solubilisierung
der Proteine wurde Solubilisierungspuffer (Chua, 1980; s. u.) und β-
Mercaptoethanol hinzugefügt, so daß der Solubilisierungspuffer 40% (v/v) und β-
Mercaptoethanol 5% (v/v) des Gesamtvolumens ausmacht (Endkonzentrationen:
12% (w/v) Saccharose, 2% (w/v) SDS, 5% (v/v) β-Mercaptoethanol). Der Ansatz
wurde 25 min bei Raumtemperatur auf einem Vortexgerät geschüttelt. Ungelöste
Partikel wurden durch eine anschließende Zentrifugation von 3 min bei 13000
rpm in einer Eppendorfzentrifuge sedimentiert. Der Überstand wurde bei der
nachfolgenden SDS-PAGE (vgl. Kap. 2.10.) eingesetzt.
Solubilisierungspuffer: 30% (w/v) Saccharose
5% (w/v) SDS
2.10. SDS-Polyacrylamid-Gelelektrophorese von Protein aus Chla-mydomonas reinhardtii
Die elektrophoretische Auftrennung von Proteinen wurde nach Jagow (Schägger
et al., 1985) durchgeführt. Die Elektrophorese erfolgte über Nacht bei einer Span-
nung von 65 V in einem 15%-igen SDS-Polyacrylamidgel (1×140×180 mm). Die
Chlorophyllmenge der aufgetragenen Proteinproben betrug jeweils 6 µg. Nach
erfolgter elektrophoretischer Auftrennung wurden die Proteine entweder mit

2. Material und Methoden 32 Coomassie-Brilliant-Blue ca. 1 h unter Schütteln angefärbt oder auf Nitrozellulo-
se (vgl. Kap. 2.11.) überführt. Die Gelentfärbung erfolgte zunächst für 10 min in
der Entfärbelösung I (s. u.) und anschließend für mehrere Stunden in der Entfär-
belösung II (s. u.).
Acrylamid/ Bisacrylamid -Lsg.: 45% (w/v) Acrylamid
1,5% (w/v) Bisacrylamid
Gelpuffer (3fach): 3 M Tris
1 M HCl
0,3% (w/v) SDS
Gelkomponenten (Angaben für 6 Gele; 1×140×180 mm):
Trenngel: 40 ml Acrylamid/Bisacrylamid-Lsg.
40 ml Gelpuffer
24 g Glycerin
300 µl APS (10%)
40 µl TEMED
ad 120 ml A. dest.
Sammelgel: 3,3 ml Acrylamid/Bisacrylamid-Lsg
10 ml Gelpuffer
200 µl APS (10%)
30 µl TEMED
ad 30 ml A. dest.
Anodenpuffer: 0,2 M Tris-HCl; pH 8,9

2. Material und Methoden 33 Kathodenpuffer: 0,1 M Tris
0,1 M Tricin
0,1% (w/v) SDS
Färbelösung: 2,5 g Coomassie Brilliant Blue R 250
500 ml Methanol
70 ml Eisessig
ad 1 l A. dest.
Entfärbelsg. I: 50% Methanol
7,5% Eisessig
Entfärbelsg. II: 5% Methanol
7,5% Eisessig
Zur Bestimmung des Molekulargewichtes der aufgetrennten Proteine wurde ein
Molekulargewichtsmarker (10 kDa-Leiter mit 10-200 kDa-Referenzproteinen) auf-
getragen (vgl. Kap. 2.1.).
2.11. Immunologische Methoden
2.11.1. Transfer von Proteinen auf Nitrozellulosemembranen
Der Transfer von Proteinen aus SDS-Polyacrylamidgelen (vgl. Kap. 2.10.) auf Nit-
rozellulosemembranen (vgl. Kap. 2.1.) erfolgte nach der Methode von Towbin et al.
(1979) in einer Naßblotkammer (vgl. Kap. 2.1.) für 2 h bei 400 mA und bei 4°C.
Zur Überprüfung des Transfererfolges wurden die Proteine sofort nach Beendi-
gung des Blotvorganges auf der Nitrozellulose mit Ponceaureagenz (0,2% (w/v)

2. Material und Methoden 34 Ponceau S, 3% (v/v) Trichloressigsäure) sichtbar gemacht und anschließend mit A.
dest. wieder entfärbt.
Transferpuffer: 25 mM Tris-HCl; pH 8,3
129 mM Glycin
20% (v/v) Methanol
2.11.2. Immunologischer Nachweis des D1-Proteins aus Chlamydomonas reinhardtii
Zum Nachweis der elektrophoretisch transferierten Proteine auf einer Nitrozellu-
losemembran mittels spezifischen, polyklonalen Antikörpers wurde die Membran
zunächst zur Absättigung freier Proteinbindestellen für 1 h in Blocklösung (s. u.)
inkubiert. Die Bindung des 1. Antikörpers erfolgte für 14-16 h in Blocklösung. An-
schließend wurde die Nitrozellulosemembran 4-mal für 15 min mit TBS (s. u.) ge-
waschen. Die Inkubation mit dem 2. Antikörper erfolgte für ca. 1 h in Blocklösung.
Anschließend wurde die Membran, wie oben beschrieben, mit TBS gewaschen.
Sämtliche Schritte des Immunnachweises wurden bei RT unter ständigem Schüt-
teln durchgeführt. Der Nachweis der enzymatischen Aktivität der Peroxidase er-
folgte mit der Entwicklerlösung (s. u.). Nach Erscheinen violetter Banden wurde
die Enzymreaktion mit H2O abgestoppt.
TBS: 20 mM Tris-HCl; pH 7,5
500 mM NaCl
Blocklösung: 3% (w/v) Magermilchpulver in TBS
Entwickler: 50 ml TBS
10 ml Methanol
30 mg HPR Color Development Reagent (Biorad)
30 µl H2O2 (37%)

2. Material und Methoden 35 1. Antikörper: der polyklonale Kaninchenantikörper AK 271-t; gerichtet gegen den
Bereich des D1 Proteins zwischen Aminosäure 167 und Aminosäure 353 (Johan-
ningmeier, 1987a); der Antikörper wurde 1:450 in Blocklösung verdünnt.
2. Antikörper: Ziege-Antikaninchen-Antikörper mit Meerrettich-Peroxidase ge-
koppelt (Biorad); Verdünnung 1:3000 in Blocklösung.
2.12. Photometrische Meßmethoden
2.12.1. Bestimmung des Chlorophyllgehaltes
Die Chlorophyllbestimmung wurde nach Arnon (1949) durchgeführt. Sie ist zur
Messung des Chlorophyllgehaltes von ganzen Zellen sowie von isolierten Thyla-
koiden einsetzbar. Es wird die optische Dichte einer chlorophyllhaltigen, 80%-igen
(v/v) Acetonlösung bei 652 nm bestimmt und daraus die Chlorophyllmenge pro
Volumen mit Hilfe des Lambert-Beerschen-Gesetzes nach folgender Formel be-
rechnet:
OD652 × Verdünnungsfaktor
———————————————— = Chlorophyll a+b [mg/ml]
34,5
OD652: optische Dichte bei 652 nm
34,5: molarer Extinktionskoeffizient
2.12.2. Bestimmung des Algenwachstums
Je 250 ml HS-Medium (vgl. Kap. 2.3.2.) wurden mit den zu untersuchenden Chla-
mydomonas reinhardtii-Stämmen angeimpft, so daß die Ausgangswerte der opti-
schen Dichte bei 750 nm ca. 0,01 betrugen. Die Anzucht erfolgte in Algenröhren,
wie im Kap 2.3.1. beschrieben. In definierten Zeitintervallen wurden Aliquots zur
Bestimmung der optischen Dichte der Algensuspension bei 750 nm entnommen
(Sager und Granick, 1953).

2. Material und Methoden 36 2.12.3. Messung der Elektronentransportraten von PS II
Die photosynthetische Aktivität von PS II wurde im DCPIP-System gemessen.
Dabei wird der entkoppelte Elektronenfluß vom Wasser zu dem künstlichen E-
lektronenakzeptor DCPIP erfaßt. Die Messungen wurden an einer Querbelich-
tungsapparatur (Zeiss PMQ II, bei 0,1 W/cm2 Rotlicht) bei 600 nm durchgeführt.
Meßpuffer: 30 mM HEPES-NaOH; pH 7,0
10 mM MgCl2
60 µM DCPIP
2,5 µM Gramicidin
1 µM DNP-INT
Der Meßansatz bestand aus 2 ml Meßpuffer und 10-20 µl Thylakoidsuspension
(vgl. Kap. 2.8.). Das Volumen der Thylakoidsuspension entsprach jeweils 10 µg
Chlorophyll. Die Messungen der Elektronentransportraten erfolgten in Ab- und
Anwesenheit von Hemmstoffen. Die Hemmwirkung der Hemmstoffe wurde bei ver-
schiedenen Konzentrationen des jeweiligen Hemmstoffs bestimmt.

3. Ergebnisse 37
3. Ergebnisse
Das Reaktionszentrum von Photosystem II besteht aus den Proteinen D1 und D2.
Sie tragen die Komponenten der Elektronentransportkette. Das D1-Protein ent-
hält zudem die Bindestelle für das im lipophilen Raum der Membran lösliche
Plastochinon QB.
Die Struktur des Photosystems II ist noch nicht endgültig aufgeklärt. Dank der
Kristallstrukturanalyse ist der Verlauf der α-Helices im Reaktionszentrum be-
kannt. Detaillierte Angaben zu den Aminosäuren, der Ausrichtung ihrer Seiten-
ketten und der Fixierung der prosthetischen Gruppen fehlen (Zouni et al., 2001).
Die Modelle für das Photosystem II basieren auf der Röntgenstruktur des Reakti-
onszentrums der Purpurbakterien. Diese Modelle werden durch Photoaffinitäts-
markierungen und durch Mutantenanalyse unterstützt.
Im Rahmen dieser Arbeit sollte die Mutantenanalyse für die Charakterisierung
der QB-Bindenische im Photosystem II von Chlamydomonas reinhardtii angewen-
det werden. Es sollten Mutanten mit Aminosäuresubstitutionen im D1-Protein
erzeugt werden. Dazu wurde die Methode der Oligonukleotid-gerichteten Mutage-
nese gewählt. Sie ermöglicht gezielte Punktmutationen in das für das D1-Protein
codierende psbA-Gen einzuführen. Folgende Aminosäuren des D1-Proteins sollten
substituiert werden: Val 249, His 215 und Phe 206. Val 249 wurde durch Photoaf-
finitätsmarkierung von Spinatthylakoiden als Bindungsort von Azido-
[125I]ioxynil, einem Cyanophenol, erkannt (Oettmeier et al., 1989). His 215 ist an
der Bindung des Fe-Ions beteiligt und stabilisiert folglich die dreidimensionale
Faltung des D1-Proteins. Der Austausch von Phe 206 ist bereits in einer Drei-
fachmutante von Chlamydomonas reinhardtii bekannt, die ein verändertes Ver-
halten gegenüber Hemmstoffen zeigt (Mutante 12: Tyr206/Ala211/Thr214 aus der
Diplomarbeit; Orawski, 1998). Im Hinblick auf diese Erkenntnisse sind die Amino-
säureaustausche an Positionen Val 249, His 215 und Phe 206 von großem Interes-
se.

3. Ergebnisse 38 Die Charakterisierung der Mutanten mit Hilfe von Hemmstoffen sollte nähere
Aussagen über Struktur und Funktion der QB- und Herbizid-Bindenische liefern.
Für die Analyse der Mutanten mit Hemmstoffen wurden Verbindungen verschie-
dener Klassen gewählt: Harnstoffe, Triazine, Triazinone, Biscarbamate, Cyanoac-
rylate, Nitrophenole und Cyanophenole.
Bereits in der vorhergehenden Arbeit (Orawski, 1998) wurde ortsgerichtete Muta-
genese durchgeführt, bei der Aminosäureaustausche an den Positionen Phe 211
und Met 214 im D1-Protein herbeigeführt wurden. Die Mutation von Phe 211 zu
Ser wurde bereits bei Cyanobakterien charakterisiert (Gingrich et al., 1988; Kiri-
lovsky et al., 1989; Govindjee et al., 1990). Diese Mutation veränderte die Hemm-
eigenschaften von Atrazin, Diuron und Metribuzin. Met 214 wurde durch Photoaf-
finitätsmarkierung von Spinatthylakoiden als Bindungsort von Azido-[14C]atrazin
erkannt (Wolber et al., 1986; Whitelegge et al., 1992). Die mutagenisierte DNA
wurde erfolgreich bei der Transformation in die Zellen von Chlamydomonas rein-
hardtii eingeschleust. Von den 35 erzeugten Klonen wurden 5 charakterisiert (O-
rawski, 1998). Im Rahmen dieser Dissertation sollten die weiteren 30 Transfor-
manten analysiert werden. Diese Analyse sollte die Erkenntnisse über die Bedeu-
tung der Aminosäuren an den Positionen 211 und 214 für die Struktur der QB-
Bindenische und für die Bindung verschiedener Hemmstoffe vertiefen.
3.1. Konstruktion von Chlamydomonas reinhardtii-Mutanten
3.1.1. In vitro-Mutagenese des psbA-Gens
Es sollten Mutanten von Chlamydomonas reinhardtii mit Aminosäureaustau-
schen im D1-Protein erzeugt werden. Vom Interesse waren die Aminosäuren: Phe
206, His 215 und Val 249 (Abb. 3.1.). Gezielte Punktmutationen lassen sich mit
Hilfe der Oligonukleotid-gerichteten Mutagenese einfügen.
Im Rahmen dieser Arbeit wurde die ortsspezifische Mutagenese mit Hilfe der PCR
nach einer Methode von Landt et al. (1990) durchgeführt (vgl. Kap. 2.7.1.).

3. Ergebnisse 39
Abb. 3.1.: Modell der QB- und
Herbizid-Bindenische des D1-
Proteins von PS II; die invers
dargestellten Aminosäuren
sollten mutiert werden.
Als Vorlage für die PCR diente das Plasmid pSHc5 (Johanningmeier und Heiß,
1993). Dieses Plasmid enthält das intronlose psbA-Gen und ca. 1,5 kb
3’flankierende Sequenz im Bluescript Vektor pBSSK(+) (vgl. Kap. 2.6.). Der muta-
gene Primer, der bei der Mutagenese von Phe 206 eingesetzt wurde (opsbA 206),
war an 3 Stellen gewobbelt (Abb. 3.2.). Durch die verschiedenen Kombinations-
möglichkeiten der Nukleotide bei der Oligonukleotidsynthese waren folgende 16
Aminosäuren anstelle von Phe 206 möglich: Ala, Arg, Asn, Asp, Gln, Glu, Gly, His,
Ile, Leu, Lys, Met, Pro, Thr, Trp oder Val.

3. Ergebnisse 40
206 Gly Val Ala Gly Val Phe Gly Gly Ser Leu Phe (a): 5’-A GGT GTT GCT GGT GTA TTC GGT GGT TCA TTA TTC T-3’ (b): 3’-T CCA CAA CGA CCA CAT CNN CCA CCA AGT AAT AAG A-5’ G T
Abb. 3.2. a: Abschnitt des psbA-Gens zwischen den Codons 201 und 211 mit der entsprechenden
Aminosäuresequenz; die zu mutierende Aminosäure ist fett gedruckt; b: mutagener Primer opsbA
206; die ausgetauschten Nukleotide sind kursiv gedruckt.
Die Mutagenese von His 215 wurde mit 4 verschiedenen Primern (opsbA 215,
opsbA 215Gln, opsbA 215Ile, opsbA 215Phe) durchgeführt. Der mutagene Primer
opsbA 215 war an 3 Stellen gewobbelt (Abb. 3.3.). Durch die verschiedenen Kom-
binationen waren an der Position 215 im D1-Protein alle Aminosäuren außer His,
Gln und Pro möglich. Die anderen 3 Oligonukleotide (opsbA 215Gln, opsbA 215Ile,
opsbA 215Phe) waren so gewählt, daß His 215 entweder durch Gln, Ile oder Phe
substituiert werden konnte (Abb. 3.3.)
Das mutagene Oligonukleotid für den Austausch von Val 249 war an 3 Stellen ge-
wobbelt, so daß alle 64 Tripletts möglich waren und folglich die Wildtypsequenz
nicht ausgeschlossen war (Abb. 3.4.).
215 Leu Phe Ser Ala Met His Gly Ser Leu Val Thr (a): 5’-A TTA TTC TCA GCT ATG CAC GGT TCT TTA GTT ACT T-3’ (b): 3’-T AAT AAG AGT CGA TAC ANN CCA AGA AAT CAA TGA A-5’ C T (c): 3’-T AAT AAG AGT CGA TAC GTT CCA AGA AAT CAA TGA A-5’ (d): 3’-T AAT AAG AGT CGA TAC TAG CCA AGA AAT CAA TGA A-5’ (e): 3’-T AAT AAG AGT CGA TAC AAG CCA AGA AAT CAA TGA A-5’
Abb. 3.3. a: Abschnitt des psbA-Gens zwischen den Codons 210 und 220 mit der entsprechenden
Aminosäuresequenz; die zu mutierende Aminosäure ist fett gedruckt; b: mutagener Primer opsbA
215; c: mutagener Primer opsbA 215Gln; d: mutagener Primer opsbA 215Ile; e.: mutagener Primer
opsbA 215Phe; die ausgetauschten Nukleotide sind kursiv gedruckt.

3. Ergebnisse 41
249 Glu Thr Tyr Asn Ile Val Ala Ala His Gly Tyr (a): 5’-A GAA ACT TAC AAC ATT GTA GCT GCT CAT GGT TAC T-3’ (b): 3’-T CTT TGA ATG TTG TAA NNN CGA CGA GTA CCA ATG A-5’
Abb. 3.4. a: Abschnitt des psbA-Gens zwischen den Codons 244 und 254 mit der entsprechenden
Aminosäuresequenz; die zu mutierende Aminosäure ist fett gedruckt; b: mutagener Primer opsbA
249; die ausgetauschten Nukleotide sind kursiv gedruckt.
Die Mutagenese erfolgte jeweils in 2 Schritten (vgl. Kap. 2.7.1.). Das Produkt der 1.
PCR diente als Primer in der 2. PCR. Da die Ausbeute bei der 2. PCR nicht immer
zufriedenstellend war, wurde das Produkt dieser PCR als Matrize bei einer weite-
ren PCR eingesetzt (Abb. 3.5.). Das Produkt der 3. PCR wurde in die Mutante Del1
transformiert.

3. Ergebnisse 42 Abb. 3.5. (vorherige Seite): Analyse der PCR-Produkte der ortsgerichteten Mutagenese. Gele-
lektrophoretische Auftrennung (Agarosegel, 0,8% (w/v), Ethidiumbromid-Färbung) der PCR-
Produkte. a: M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1: Produkt der 1. PCR (666 bp)
mit dem Primer opsbA 206; 2: Produkt der 1. PCR (693 bp) mit dem Primer opsbA 215; 3: Produkt
der 1. PCR (795 bp) mit dem Primer opsbA 249; 4: Produkt der Kontroll-PCR (1118 bp); b: M: Grö-
ßenstandard (λ-DNA HindIII/EcoRI geschnitten); 1, 2 bzw. 3: Produkt der 2. PCR (1118 bp) der Mu-
tagenese von Phe 206, von His 215 bzw. von Val 249; 4, 5, 6: Produkte der entsprechenden 3. PCR
(1118 bp); c: M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1, 2 bzw. 3: Produkt der 1.
PCR (693 bp) mit dem Primer opsbA 215Gln, opsbA 215Ile bzw. opsbA 215Phe; 4, 5, 6: Produkte der
entsprechenden 2. PCR (1118 bp); d: M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1, 2
bzw. 3: Produkt der 3. PCR (1118 bp) für die Mutagenese von His 215 zu Gln, Ile bzw. Phe.
3.1.2. Sequenzierung der PCR-Produkte der Mutagenese
Um zu überprüfen, ob die Mutagenese gelungen war und ob die PCR-Produkte
keine zusätzlichen unbeabsichtigten Mutationen aufwiesen, wurden die Mutage-
neseprodukte sequenziert (vgl. Kap. 2.7.4.). Die Sequenzierungen zeigten, daß die
Mutagenese mit allen sechs mutagenen Oligonukleotiden gelungen war. Es waren
keine zusätzlichen Mutationen aufgetreten. Die mutagenisierten PCR-Produkte
konnten zur Transformation eingesetzt werden.
3.1.3. Transformation der Mutante Del1
Die einzellige Grünalge Chlamydomonas reinhardtii besitzt zwei Kopien des psbA-
Gens in der inverted repeat-Region des Genoms (Erickson et al., 1984). Etwa 80
Kopien des Chloroplastengenoms sind pro Zelle vorhanden. Mit Hilfe der Parti-
kelkanone (Klein et al., 1987; Boynton et al., 1988) können Mutationen in das
Chloroplastengenom eingeführt werden. Diese Technik sollte im Rahmen dieser
Arbeit angewandt werden, um Punktmutationen in das psbA-Gen einzuführen.
Die Modifikation des psbA-Gens muß beide Kopien des Gens auf dem zirkulären
Genom betreffen, und alle der ca. 80 Genomkopien müssen sie enthalten, d. h. es
muß Homoplasmizität vorliegen. Heteroplasmische Mutanten sind oft instabil und
liefern heterogene Genprodukte.
Zur Transformation (vgl. Kap. 2.7.9.) wurde der Chlamydomonas reinhardtii
Stamm Del1 eingesetzt (Johanningmeier et al., 2000). Del1 entstand durch die

3. Ergebnisse 43 Transformation der Mutante IL. Diese besitzt ein intronloses psbA-Gen, welches
für ein D1-Protein codiert, das dem des Wildtyps entspricht (Heiß, 1992; Johan-
ningmeier und Heiß, 1993). Del1 wurde auf der Grundlage dieses intronlosen
psbA-Gens erstellt. Sie besitzt eine Deletion von 425 bp und codiert für ein ver-
kürztes D1-Protein, das die Helices I und II sowie etwa die Hälfte von Helix III
aufweist. Zudem zerstört die Deletion den Leserahmen, so daß das D1-Protein der
Mutante Del1 am C-Terminus einen Schwanz von 30 überwiegend hydrophilen
Aminosäuren trägt (Abb. 3.6.). Entscheidend für die hier durchgeführten Experi-
mente ist, daß die Helices IV, IV/V und V in der Mutante Del1 deletiert sind und
somit der Bereich der QB-Bindenische fehlt. In dieser Region des Proteins liegen
die zu mutierenden Aminosäuren. Außerdem ist die Mutante Del1 nicht in der
Lage photosynthetisch zu wachsen, wodurch nach der Transformation, die zum
Wiedererlangen der Fähigkeit zum photoautotrophen Wachstum führt, photosyn-
thetisch kompetente von photosynthetisch nicht kompetenten Zellen selektiert
werden können.
Abb. 3.6.: Schematische Darstellung des intakten (D1) und des deletierten (tD1) D1- Proteins des PS
II; angegeben sind die Kofaktoren P680 und Fe; der deletierte Bereich ist markiert.

3. Ergebnisse 44 Zur Transformation der Mutante Del1 wurden die Produkte der Mutagenesen
sowie das Produkt der Kontroll-PCR eingesetzt. Die Selektion der transformierten
Zellen erfolgte auf HS-Medium (vgl. Kap. 2.3.2.), das nur das Wachstum von pho-
tosynthetisch kompetenten Zellen ermöglicht. Die Ausbeuten der durchgeführten
Transformationen sind in der Tabelle 3.1. aufgeführt.
Donor-DNA beschossene Zellen (×1-2×107)
positive Transformanten
Transformations-frequenz (×1-2×10-7)
PCR 206 9 18 2
PCR 215 20 11 0,6
PCR 249 9 42 4,7
PCR 215Gln 20 12 0,6
PCR 215Ile 7 3 0,4
PCR 215Phe 20 5 0,3
Kontroll-PCR 6 184 31
ohne DNA 2 0 0
Tab. 3.1.: Ausbeuten der Transformationen. 1 bis 2×107 Zellen der Mutante Del1 wurden mit Muta-
geneseprodukten sowie mit dem Produkt der Kontroll-PCR transformiert. Die Einschleusung der
DNA in den Chloroplasten erfolgte mit einer Partikelkanone. Nach der Transformation wurden die
Zellen 4 Wochen im Dauerlicht inkubiert. Zur Negativkontrolle wurden die Zellen mit Wolframpar-
tikeln ohne DNA beschossen. PCR 206: Produkt der Mutagenese mit dem mutagenen Primer opsbA
206; PCR 215: entsprechend mit dem Primer opsbA 215; PCR 249: mit dem Primer opsbA 249; PCR
215Gln: mit dem Primer opsbA 215Gln; PCR 215Ile: mit dem Primer opsbA 215Ile; PCR 215Phe: mit
dem Primer opsbA 215Phe.
Das PCR-Produkt, mit dem die Zellen transformiert wurden, enthielt das vollstän-
dige psbA-Gen. Die Transformation führt nur dann zu photosynthetisch wachsen-
den Kolonien, wenn die im psbA-Gen erzeugte Punktmutation nicht letal ist oder
wenn die eingeschleuste DNA zum Wildtyp führt. Die Wiederherstellung von
Wildtyp war bei der Transformation mit der DNA für die Mutagenese von Val 249
möglich. Bei dem mutagenen Primer für diese Mutagenese waren alle 64 Tripletts
möglich. Dies erklärt die Tatsache, daß die Transformationsfrequenz in diesem
Fall am höchsten war.

3. Ergebnisse 45 Es wurden sowohl Positiv- als auch Negativkontrollen durchgeführt. Zur Negativ-
kontrolle wurden die Zellen mit Wolframpartikeln ohne DNA beschossen. In die-
sem Fall gab es keine positiven Transformanten. Zur Positivkontrolle wurden die
Algenzellen mit der DNA der Kontroll-PCR transformiert. Die Transformations-
frequenz der Kontrolle betrug 31×1 bis 2×10-7 (Tab. 3.1.).
Die Transformation mit der DNA der Kontroll-PCR führt in jedem Fall zum Wild-
typ. Die Transformationsbedingungen bei den Kontrollen waren vergleichbar mit
den Bedingungen der mutagenen Transformationen. Die Transformationsfre-
quenz der Positivkontrollen ist deutlich höher als die der mutagenen Transforma-
tionen. Die Frequenz bei der Transformation mit den PCR-Produkten für die Sub-
stitution von His 215 ist am niedrigsten.
3.2. Analyse der Transformanten mit Hilfe der PCR
Es ist erforderlich, daß nach der Transformation alle der etwa 80 Kopien des
Plastoms wieder das vollständige psbA-Gen enthalten. Um dies festzustellen, wur-
den die Transformanten mit Hilfe der PCR analysiert. Die Gesamt-DNA wurde
aus den Transformanten sowie aus den Mutanten Del1 und IL als Kontrolle iso-
liert (vgl. Kap. 2.7.5.) und als Matrize in einer PCR-Reaktion eingesetzt (vgl. Kap.
2.7.2.). Als Primer dienten die Oligonukleotide opsbA -28 und opsbA stop. Diese
Oligonukleotide flankieren das psbA-Gen. Die resultierenden PCR-Fragmente
wurden über 0,8%-igen Agarosegele aufgetrennt (Abb. 3.7. bis Abb. 3.18.). Die
Größe der PCR-Produkte der Transformanten wurde mit der Größe der PCR-
Produkte der Mutanten Del1 und IL verglichen.
Die PCR mit der DNA der Mutante IL ergab ein Fragment von 1,1 kbp. Dies ent-
spricht dem intronlosen psbA-Gen. Das PCR-Produkt der Mutante Del1 war 0,7
kbp groß und entsprach dem deletierten psbA-Gen. Es wurden insgesamt 115 Ko-
lonien analysiert. 30 davon (Klon 1, 2, 3, 5, 7, 9, 10, 11, 13, 14 bis 23 und 25 bis 35)
stammen aus der vorhergehenden Arbeit (Orawski, 1998). 18 entstanden durch
die Transformation mit dem PCR-Produkt, bei dem Phe 206 substituiert wurde
(Klon 206/1 bis 206/18). 31 weitere stammen aus der Transformation mit den PCR-
Produkten, bei denen His 215 substituiert wurde (Klon 215/1 bis 215/11, 215Gln/1

3. Ergebnisse 46 bis 215Gln/12, 215Ile/1 bis 215Ile/3, 215Phe/1 bis 215Phe/5) und 36 aus der Trans-
formation mit dem PCR-Produkt, bei dem Val 249 ausgetauscht wurde (Klon 249/1
bis 249/8, 249/13 bis 249/35 und 249/38 bis 249/42). Wie die Abbildungen zeigen,
besitzen die transformierten Algen wieder ein vollständiges intronloses psbA-Gen.
Ihre PCR-Produkte laufen bei der Agarosegelelektrophorese auf der gleichen Hö-
he wie das Produkt der PCR mit der DNA der Mutante IL als Matrize. Die analy-
sierten Transformanten zeigen keine Bande bei 0,7 kbp. Dies bedeutet, daß alle
Kopien des Chloroplastengenoms das vollständige psbA-Gen tragen und damit
homoplasmisch sind.
Abb. 3.7.: PCR-Analyse der Klone aus der Transformation mit dem Mutageneseprodukt bei dem
Phe211 und Met214 substituiert wurden. Gelelektrophoretische Auftrennung (Agarosegel, 0,8%
(w/v), Ethidumbromid-Färbung) von PCR-Produkten aus Reaktionen mit dem Oligonukleotidpaar
opsbA -28 und opsbA stop. M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1: Del1; 2 IL; 3
bis 12: Klon 1, 2, 3, 5, 7, 9, 10, 11, 13, 14.

3. Ergebnisse 47
Abb. 3.8.: PCR-Analyse der Klone aus der Transformation mit dem Mutageneseprodukt, bei dem
Phe211 und Met214 substituiert wurden. Gelelektrophoretische Auftrennung (Agarosegel, 0,8%
(w/v), Ethidumbromid-Färbung) von PCR-Produkten aus Reaktionen mit dem Oligonukleotidpaar
opsbA -28 und opsbA stop. M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1: Del1; 2: IL; 3
bis 12: Klon 15, 16, 17, 18, 19, 20, 21, 22, 23, 25.
Abb. 3.9.: PCR-Analyse der Klone aus der Transformation mit dem Mutageneseprodukt, bei dem
Phe211 und Met214 substituiert wurden. Gelelektrophoretische Auftrennung (Agarosegel, 0,8%
(w/v), Ethidumbromid-Färbung) von PCR-Produkten aus Reaktionen mit dem Oligonukleotidpaar
opsbA -28 und opsbA stop. M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1: Del1; 2: IL; 3
bis 12: Klon 26 bis 35.

3. Ergebnisse 48
Abb. 3.10.: PCR-Analyse der Klone aus der Transformation mit dem Mutageneseprodukt, bei dem
Phe206 substituiert wurde. Gelelektrophoretische Auftrennung (Agarosegel, 0,8% (w/v), Ethidum-
bromid-Färbung) von PCR-Produkten aus Reaktionen mit dem Oligonukleotidpaar opsbA -28 und
opsbA stop. M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1: Del1; 2: IL; 3 bis 11: Klon
206/1 bis 206/9.
Abb. 3.11.: PCR-Analyse der Klone aus der Transformation mit dem Mutageneseprodukt, bei dem
Phe206 substituiert wurde. Gelelektrophoretische Auftrennung (Agarosegel, 0,8% (w/v), Ethidum-
bromid-Färbung) von PCR-Produkten aus Reaktionen mit dem Oligonukleotidpaar opsbA -28 und
opsbA stop. M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1: Del1; 2: IL; 3 bis 11: Klon
206/10 bis 206/18.

3. Ergebnisse 49
Abb. 3.12.: PCR-Analyse der Klone aus der Transformation mit dem Mutageneseprodukt, bei dem
His215 substituiert wurde. Gelelektrophoretische Auftrennung (Agarosegel, 0,8% (w/v), Ethi-
dumbromid-Färbung) von PCR-Produkten aus Reaktionen mit dem Oligonukleotidpaar opsbA -28
und opsbA stop. M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1: Del1; 2: IL; 3 bis 13:
Klon 215/1 bis 215/11.
Abb. 3.13.: PCR-Analyse der Klone aus der Transformation mit dem Mutageneseprodukt, bei dem
His215 substituiert wurde. Gelelektrophoretische Auftrennung (Agarosegel, 0,8% (w/v), Ethi-
dumbromid-Färbung) von PCR-Produkten aus Reaktionen mit dem Oligonukleotidpaar opsbA -28
und opsbA stop. M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1: Del1; 2: IL; 3 bis 14:
Klon 215Gln/1 bis 215Gln/12.

3. Ergebnisse 50
Abb. 3.14.: PCR-Analyse der Klone aus der Transformation mit dem Mutageneseprodukt, bei
dem His215 substituiert wurde. Gelelektrophoretische Auftrennung (Agarosegel, 0,8% (w/v), Ethi-
dumbromid-Färbung) von PCR-Produkten aus Reaktionen mit dem Oligonukleotidpaar opsbA -28
und opsbA stop. M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1: Del1; 2: IL; 3: Klon
215Ile/1; 4: Klon 215Ile/2; 5: Klon 215Ile/3; 6: Klon 215Phe/1; 7: Klon 215Phe/2; 8: Klon 215Phe/3; 9:
Klon 215Phe/4; 10: Klon 215Phe/5.
Abb. 3.15.: PCR-Analyse der Klone aus der Transformation mit dem Mutageneseprodukt, bei
dem Val249 substituiert wurde. Gelelektrophoretische Auftrennung (Agarosegel, 0,8% (w/v), Ethi-
dumbromid-Färbung) von PCR-Produkten aus Reaktionen mit dem Oligonukleotidpaar opsbA -28
und opsbA stop. M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1: Del1; 2: IL; 3 bis 13:
Klon 249/1 bis 249/8, 249/13 bis 249/15.
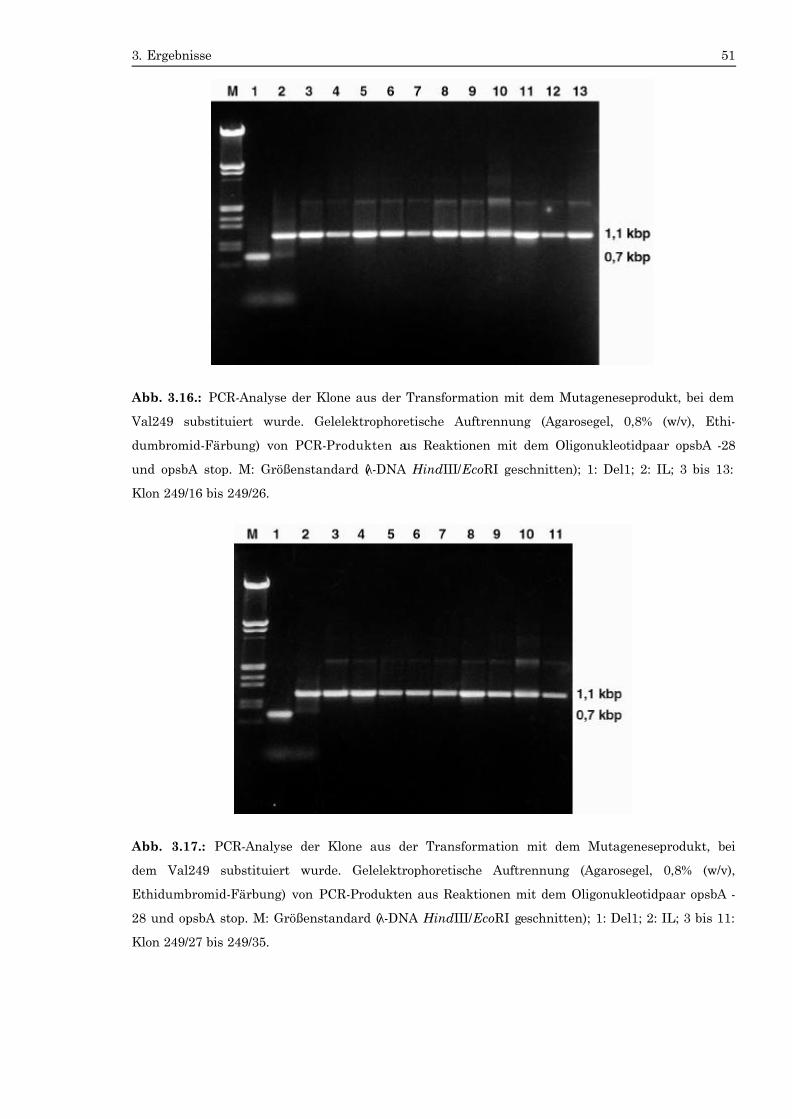
3. Ergebnisse 51
Abb. 3.16.: PCR-Analyse der Klone aus der Transformation mit dem Mutageneseprodukt, bei dem
Val249 substituiert wurde. Gelelektrophoretische Auftrennung (Agarosegel, 0,8% (w/v), Ethi-
dumbromid-Färbung) von PCR-Produkten aus Reaktionen mit dem Oligonukleotidpaar opsbA -28
und opsbA stop. M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1: Del1; 2: IL; 3 bis 13:
Klon 249/16 bis 249/26.
Abb. 3.17.: PCR-Analyse der Klone aus der Transformation mit dem Mutageneseprodukt, bei
dem Val249 substituiert wurde. Gelelektrophoretische Auftrennung (Agarosegel, 0,8% (w/v),
Ethidumbromid-Färbung) von PCR-Produkten aus Reaktionen mit dem Oligonukleotidpaar opsbA -
28 und opsbA stop. M: Größenstandard (λ-DNA HindIII/EcoRI geschnitten); 1: Del1; 2: IL; 3 bis 11:
Klon 249/27 bis 249/35.

3. Ergebnisse 52
Abb. 3.18.: PCR-Analyse der Klone aus der
Transformation mit dem Mutageneseprodukt, bei
dem Val249 substituiert wurde. Gelelektro-
phoretische Auftrennung (Agarosegel, 0,8% (w/v),
Ethidumbromid-Färbung) von PCR-Produkten aus
Reaktionen mit dem Oligonukleotidpaar opsbA -28
und opsbA stop. M: Größenstandard (λ-DNA
HindIII/EcoRI geschnitten); 1: Del1; 2: IL; 3 bis 8:
Klon 249/38 bis 249/42.
3.3. Sequenzierung der Transformanten-DNA
Die in die Mutante Del1 transformierten PCR-Produkte waren Populationen ver-
schiedener DNA-Moleküle, weil die mutagenen Primer so gewählt waren, daß an
der zu mutierenden Position möglichst viele verschiedene Aminosäureaustausche
zu erwarten waren. Um die erwarteten Mutationen zu identifizieren, sollte die
Transformanten-DNA sequenziert werden (vgl. Kap. 2.7.4.). Für die Sequenzie-
rung wurden PCR-Produkte eingesetzt, die bei der Analyse der Transformanten
mit Hilfe der PCR synthetisiert wurden.
3.3.1. Sequenzierung der Phe211/Met214-Transformanten
30 der im Rahmen dieser Dissertation analysierten Klone stammen aus der Dip-
lomarbeit (Orawski, 1998). Das Ziel der Mutagenese waren Aminosäureaustausche
an den Positionen Phe 211 und Met 214. Der mutagene Primer war so gewählt,
daß sowohl anstelle des Codons für Phe 211 als auch des Codons für Met 214 alle
64 Tripletts möglich waren. Von den insgesamt 35 Transformanten haben sich 18
als Mutanten herausgestellt, darunter sind 16 verschiedene Mutanten vorhanden,
2 kommen zweimal vor. Die restlichen 17 Kolonien waren Wildtyp. Die Sequen-
zierergebnisse sind in der Tabelle 3.3. aufgeführt.

3. Ergebnisse 53
2 Phe211→Ala Met214→Asn
3 Phe211→Thr Met214→Thr Val219→Ala
4* Phe211→Ile Met214→Gly
6* Phe211→Gly Met214→Ser
8* Phe211→Thr Met214→Gly
10 Phe211→Thr Met214→Ala
12* Phe211→Ala Met214→Thr Phe206→Tyr
13 Phe211→Met Met214→Thr
14 Phe211→Gln
19=24 Phe211→Gly Met214→Gln
21 Phe211→Leu Met214→Gly
24*=19 Phe211→Gly Met214→Gln
27 Phe211→Gly Met214→Ala
28=30 Phe211→Gln Met214→Gly Val219→Ala
30=28 Phe211→Gln Met214→Gly Val219→Ala
32 Phe211→Cys Met214→Thr Val219→Ala
34 Phe211→Gly Met214→Ser Val219→Ala
35 Phe211→Gly Met214→Thr
Tab. 3.3.: Ergebnisse der Sequenzierung des psbA-Gens im Bereich der QB-Bindenische der Klone 1
bis 35. Die nicht aufgeführten Klone waren Wildtyp. Die mit Stern * markierten Mutanten wurden
bereits in der Diplomarbeit charakterisiert (Orawski, 1998).
Bei vier Mutanten trat an der Stelle Val 219 eine nicht geplante Mutation zu Ala
auf, die auf einen Fehler der Taq-Polymerase zurückzuführen ist. Das Gleiche gilt
für die Mutation von Phe 206 zu Tyr bei der Mutante 12. 5 von diesen Mutanten
(Mutante 4, 6, 8, 12 und 24) wurden bereits in der vorhergehenden Arbeit charak-
terisiert (Orawski, 1998).
3.3.2. Sequenzierung der Phe206-Transformanten
Die Transformation mit dem Mutageneseprodukt, bei dem Phe 206 substituiert
wurde, ergab 18 Kolonien, bezeichnet als Klone 206/1 bis 206/18. Die Sequenzie-
rergebnisse sind in der Tabelle 3.4. aufgelistet.

3. Ergebnisse 54
206/1 WT
206/2 Phe 206→Ile
206/3 Phe 206→Glu
206/4 Phe 206→Glu
206/5 Phe 206→Glu
206/6 Phe 206→Ser
206/7 Phe 206→Ala
206/8 Phe 206→Ile Ala 154→Thr
206/9 Phe 206→His
206/10 Phe 206→Asn
206/11 Phe 206→Val
206/12 Phe 206→Ala
206/13 Phe 206→Ile
206/14 Phe 206→Ala
206/15 Phe 206→Asn
206/16 WT
206/17 Phe 206→Val
206/18 Phe 206→Lys
Tab. 3.3.: Ergebnisse der Sequenzierung des psbA-Gens im Bereich der QB-Bindenische der Klone
206/1 bis 206/18.
Die Klone 206/1 und 206/16 erwiesen sich als Wildtyp. Nur 2 Codons codieren für
Phenylalanin: TTT und TTC. In dem mutagenen Primer trat an der 1. Position
des Tripletts kein T auf, so daß die Wildtypsequenz ausgeschlossen war. An der 2.
und 3. Position waren alle vier Basen möglich. Das Entstehen vom Wildtyp ist auf
einen Fehler der Taq-Polymerase zurückzuführen, die keine Korrekturlese-
Funktion besitzt. Die Taq-Polymerase wurde zum Amplifizieren der Mutagenese-
Produkte verwendet (vgl. Kap. 2.7.1. und 3.1.1.). Bei den Kombinationen A(G,C)TT
und A(G,C)TC würde ein Fehler in der ersten Position, nämlich ein Austausch ge-
gen T, zum Codon für Phenylalanin führen. Die Mutante 206/8 weist eine zusätz-
liche nicht geplante Mutation von Ala 154 zu Thr auf, die ebenfalls auf einen Feh-
ler während der PCR zurückzuführen ist. Insgesamt wurden 8 verschiedene Mut-
anten in Bezug auf Phe 206 erzeugt.
3.3.3. Sequenzierung der His215-Transformanten
Die Transformation mit dem gewobbelten Produkt für die Mutagenese von His 215
ergab 11 Kolonien, bezeichnet als Klone 215/1 bis 215/11. Alle 11 Transformanten
stellten sich als Wildtyp heraus, obwohl das mutagene Oligonukleotid so syntheti-
siert wurde, daß das Wildtyp-Codon nicht auftreten durfte (vgl. Kap. 3.1.1.). Histi-
din wird nur durch 2 Basentripletts codiert: CAT und CAC. Der Primer für die
Mutagenese wurde so gewählt, daß an der 1. Position kein C auftrat. An der 2. und
3. Position waren alle 4 Basen möglich, darunter auch die Kombination AT und

3. Ergebnisse 55 AC. Das Entstehen vom Wildtyp ist mit einem Fehler der Taq-Polymerase zu er-
klären, die keine Korrekturlese-Funktion besitzt. Bei den Kombinationen
G(A,T)AT und G(A,T)AC, die möglich waren, würde ein Fehler in der 1. Position,
nämlich ein Austausch gegen C, zum Wildtyp führen. Um diese Möglichkeit aus-
zuschließen, wurde die Mutagenese von His 215 mit 3 anderen Primern wieder-
holt. Das Codon für His 215 wurde durch folgende Tripletts ersetzt: CAA (Gln),
ATC (Ile), TTC (Phe). Die Tripletts ATC und TTC stimmen nur in einer Base mit
dem Wildtyp-Codon überein. Es ist fast unmöglich, daß während einer PCR zwei
Fehler nebeneinander vorkommen, was in diesem Fall zur Wildtypsequenz führen
könnte. Das Triplett CAA unterscheidet sich von dem Wildtyp-Codon nur durch
eine Base. Hier war also die Wahrscheinlichkeit des Auftretens vom Wildtyp
durch einen Fehler der Taq-Polymerase mindestens so groß wie bei der ersten Mu-
tagenese. Es wurden 20 Transformanten erzeugt, die nach der Sequenzierung sich
als Wildtyp herausstellten. Die Transformationsfrequenzen waren bei den 4 ver-
schiedenen Mutagenesen vergleichbar, so daß das Entstehen von Wildtyp nicht
allein mit einem Fehler der Taq-Polymerase während der PCR erklärt werden
kann.
3.3.4. Sequenzierung der Val249-Transformanten
In dem mutagenen Primer für den Austausch von Val 249 waren an der Position
249 alle 64 Tripletts möglich. Folglich konnte auch Wildtyp entstehen. Bei der
Transformation wurden 42 Klone erzeugt, darunter sind 19 Mutanten vorhanden.
Die restlichen 23 Kolonien sind Wildtyp. Es wurden 10 verschiedene Substitutio-
nen von Val 249 erreicht. Die Sequenzierergebnisse sind in der Tabelle 3.4. aufge-
führt.

3. Ergebnisse 56
249/3 Val 249→Arg
249/5 Val 249→His
249/8 Val 249→Ile
249/13 Val 249→Ala
249/16 Val 249→Leu
249/17 Val 249→Ser
249/19 Val 249→Asn
249/21 Val 249→Arg
249/24 Val 249→Gln
249/26 Val 249→Ser
249/28 Val 249→Cys
249/29 Val 249→Asn
249/30 Val 249→Ser
249/31 Val 249→Ala
249/32 Val 249→Gly
249/34 Val 249→His
249/38 Val 249→Leu
249/40 Val 249→Leu
249/42 Val 249→Ala
Tab. 3.4.: Ergebnisse der Sequenzierung des psbA-Gens im Bereich der QB-Bindenische der Klone
249/1 bis 249/42. Die nicht aufgeführten Klone waren Wildtyp.
3.4. Aufnahme von Wachstumskurven der Mutantenstämme
Die Wachstumskurven der unter photoautotrophen Bedingungen angezogenen
Chlamydomonas-Stämme sollten erste Aufschlüsse darüber geben, ob die Photo-
syntheseleistung der transformierten Algen durch die eingeführten Punktmutati-
onen beeinträchtigt ist. Grundlage für eine Vergleichbarkeit der Messungen wa-
ren identische Anzuchtbedingungen bezüglich Belichtung, Temperatur und Be-
gasung während des Wachstums sowie die Animpfung mit gleichen Zellzahlen in
identischen Volumina des HS-Mediums (vgl. Kap. 2.12.2.). Die obligat hete-
rotrophe Del1-Mutante kann aufgrund ihrer Unfähigkeit zur Photosynthese nicht
in HS-Medium wachsen und konnte daher nicht mit den anderen Stämmen vergli-
chen werden.
3.4.1. Aufnahme von Wachstumskurven der Doppelmutanten mit Amino-säureaustauschen an den Positionen Phe 211 und Met 214
Die Wachstumsraten der Mutanten 2 (Ala211/Asn214), 10 (Thr211/Ala214), 13
(Met211/Thr214), 14 (Gln211), 21 (Leu211/Gly214), und 35 (Gly211/Thr214) un-
terscheiden sich in der logarithmischen Phase nicht signifikant von denen des
Wildtyps und der intronlosen Mutante (Abb. 3.19. bis Abb. 3.21.). Die Mutanten 3
(Thr211/Thr214/Ala219), 27 (Gly211/Ala214), 28 (Gln211/Gly214/Ala219), 32
(Cys211/Thr214/Ala219) und 34 (Gly211/Ser214/Ala219) zeigen gegenüber dem

3. Ergebnisse 57 Wildtyp und der intronlosen Mutante ein verändertes Wachstum (Abb. 3.19., Abb.
3.21., Abb. 3.22.). Die Dauer der Anlaufphase ist bei diesen Mutanten verlängert
und sie erreichen das Wachstum des Wildtyps und der intronlosen Mutante in der
logarithmischen Phase nicht. Dieses Ergebnis läßt darauf schließen, daß die her-
beigeführten Aminosäureaustausche einen negativen Einfluß auf den photosyn-
thetischen Elektronentransport der Algen haben und damit die Wachstumsrate
der Mutantenstämme verringern. Die geringere Photosyntheseleistung der Mut-
anten kann verschiedene Ursachen haben. Einerseits wäre es möglich, daß in den
Mutanten 3, 27, 28, 32 und 34 weniger D1-Protein synthetisiert wird, oder daß
durch die Mutationen das D1-Protein schneller abgebaut wird. In beiden Fällen
müßte die D1-Konzentration in den Mutanten niedriger sein als im Wildtyp. An-
dererseits wäre es denkbar, daß die Menge an D1-Protein gleichbleibt, aber der
Elektronentransport im PS II durch die Mutationen eingeschränkt wird, z.B.
durch eine verminderte Bindung von Plastochinon in der QB-Bindenische.
Abb. 3.19.: Photosynthetisches Wachstum der
Chlamydomonas reinhardtii-Stämme: WT, IL,
Mut. 2, 3 und 10. Die Anzucht der Algen
erfolgte photoautotroph in HS-Medium in An-
zuchtröhren mit Begasung von 95% Luft und
5% CO2 bei 7000 Lux Dauerlicht und 18°C.

3. Ergebnisse 58
Abb. 3.20.: Photosynthetisches Wachstum der
Chlamydomonas reinhardtii-Stämme: WT, IL,
Mut. 13, 21 und 35. Die Anzucht der Algen
erfolgte photoautotroph in HS-Medium in
Anzuchtröhren mit Begasung von 95% Luft
und 5% CO2 bei 7000 Lux Dauerlicht und 18°C.
Abb. 3.21.: Photosynthetisches Wachstum der
Chlamydomonas reinhardtii-Stämme: WT, IL,
Mut. 14, 27 und 28. Die Anzucht der Algen
erfolgte photoautotroph in HS-Medium in An-
zuchtröhren mit Begasung von 95% Luft und
5% CO2 bei 7000 Lux Dauerlicht und 18°C.

3. Ergebnisse 59
Abb. 3.22.: Photosynthetisches Wachstum der
Chlamydomonas reinhardtii-Stämme: WT, IL,
Mut. 32 und 34. Die Anzucht der Algen erfolgte
photoautotroph in HS-Medium in
Anzuchtröhren mit Begasung von 95% Luft
und 5% CO2 bei 7000 Lux Dauerlicht und 18°C.
3.4.2. Aufnahme von Wachstumskurven der Mutanten mit Aminosäure-austauschen an der Position Phe 206
Unter den Stämmen mit Aminosäuresubstitutionen an der Position Phe 206 zei-
gen die Mutanten 206/6 (Ser206) und 206/7 (Ala206) eine verlängerte Anlaufpha-
se und eine verminderte Wachstumsrate der logarithmischen Phase. Die Ursa-
chen dafür können wie in dem Fall der Mutanten 3, 27, 28, 32 und 34 unterschied-
lich sein (vgl. Kap. 3.4.1.). Das Wachstum anderer Mutanten dieser Gruppe ist
vergleichbar mit dem Wachstum des Wildtyps und der intronlosen Mutante (Abb.
3.23. bis Abb. 3.25.).

3. Ergebnisse 60
Abb. 3.23.: Photosynthetisches Wachstum
der Chlamydomonas reinhardtii-Stämme:
WT, IL, 206/2 (Ile206) und 206/3 (Glu206). Die
Anzucht der Algen erfolgte photo- autotroph in
HS-Medium in Anzuchtröhren mit Begasung
von 95% Luft und 5% CO2 bei 7000 Lux
Dauerlicht und 18°C.
Abb. 3.24.: Photosynthetisches Wachstum der
Chlamydomonas reinhardtii-Stämme: WT, IL,
206/6 (Ser206), 206/7 (Ala206) und 206/9
(His206). Die Anzucht der Algen erfolgte
photoautotroph in HS-Medium in
Anzuchtröhren mit Begasung von 95% Luft
und 5% CO2 bei 7000 Lux Dauerlicht und
18°C.

3. Ergebnisse 61
Abb. 3.25.: Photosynthetisches Wachstum der
Chlamydomonas reinhardtii-Stämme: WT, IL,
206/10 (Asn206), 206/11 (Val206) und 206/18
(Lys206). Die Anzucht der Algen erfolgte
photoautotroph in HS-Medium in
Anzuchtröhren mit Begasung von 95% Luft
und 5% CO2 bei 7000 Lux Dauerlicht und 18°C.
3.4.3. Aufnahme von Wachstumskurven der Mutanten mit Aminosäure-austauschen an der Position Val 249
Die Mutanten 249/13 (Ala249) und 249/16 (Leu249) zeigen gegenüber dem Wild-
typ und der intronlosen Mutante eine leicht verlängerte Anlaufphase. Die ande-
ren Mutanten dieser Gruppe unterscheiden sich nicht in ihrem Wachstum vom
Wildtyp und der intronlosen Mutante (Abb. 3.26. bis Abb. 3.29.).

3. Ergebnisse 62
Abb. 3.26.: Photosynthetisches Wachstum der
Chlamydomonas reinhardtii-Stämme: WT, IL,
249/3 (Arg249), 249/5 (His249) und 249/8
(Ile249). Die Anzucht der Algen erfolgte
photoautotroph in HS-Medium in
Anzuchtröhren mit Begasung von 95% Luft
und 5% CO2 bei 7000 Lux Dauerlicht und
18°C.
Abb. 3.27.: Photosynthetisches Wachstum der
Chlamydomonas reinhardtii-Stämme: WT, IL,
249/13 (Ala249) und 249/16 (Leu249). Die
Anzucht der Algen erfolgte photoautotroph in
HS-Medium in Anzuchtröhren mit Begasung
von 95% Luft und 5% CO2 bei 7000 Lux
Dauerlicht und 18°C.

3. Ergebnisse 63
Abb. 3.28.: Photosynthetisches Wachstum der
Chlamydomonas reinhardtii-Stämme: WT,
IL, 249/17 (Ser249), 249/19 (Asn249) und
249/24 (Gln249). Die Anzucht der Algen
erfolgte photoautotroph in HS-Medium in
Anzuchtröhren mit Begasung von 95% Luft
und 5% CO2 bei 7000 Lux Dauerlicht und
18°C.
Abb. 3.29.: Photosynthetisches Wachstum
der Chlamydomonas reinhardtii-Stämme:
WT, IL, 249/28 (Cys249) und 249/32
(Gly249). Die Anzucht der Algen erfolgte
photoautotroph in HS-Medium in An-
zuchtröhren mit Begasung von 95% Luft
und 5% CO2 bei 7000 Lux Dauerlicht und
18°C.

3. Ergebnisse 64 3.5. Western Blot-Analyse der Mutanten
Um zu überprüfen, ob die Menge an D1-Protein der einzelnen Mutanten und des
Wildtyps vergleichbar ist, wurde Gesamt-Protein isoliert (vgl. Kap. 2.9.). Eine je-
weils 6 µg Chlorophyll enthaltende Proteinprobe wurde auf einem 15%-igen
SDS/Acrylamidgel aufgetrennt und auf eine Nitrozellulosemembran elektrotrans-
feriert (vgl. Kap. 2.10. und 2.11.). Die Detektion des D1-Proteins erfolgte durch den
Antikörper 271-t (Abb. 3.30. bis Abb. 3.33.). Dieser Antikörper ist gegen den Be-
reich des D1 Proteins zwischen der Aminosäure 167 und der Aminosäure 353 ge-
richtet. Dieses Bruchstück wurde in Escherichia coli überexprimiert und anschlie-
ßend als Antigen zur Synthese des Antikörpers eingesetzt (Johanningmeier,
1987a). Alle Mutanten zeigen die Anwesenheit des D1-Proteins im Bereich von 30
kDa. Das deletierte D1-Protein der Mutante Del1 ist mit dem verwendeten Anti-
körper nicht detektierbar. Die Mengen an D1-Protein, auf der Basis gleicher Chlo-
rophyllmengen, sind im Wildtyp und in den Mutanten vergleichbar.
Abb. 3.30. a: SDS-PAGE (15%, Coomassie-Färbung) von Gesamt-Protein. Die Chlorophyllmenge der
aufgetrennten Proteine betrug jeweils 6 µg. M: Molekulargewichtsmarker (10 kDa-Leiter mit 10-200
kDa Referenzproteinen); 1: intronlose Mutante; 2: Deletionsmutante; 3: Wildtyp. b: Immunblot zu
Proben aus a. 1. AK: 271-t ist gegen den Bereich des D1-Proteins zwischen Aminosäuren 167 und
353 gerichtet. 2. AK: Ziege-Antikaninchen-AK mit Meerrettich-Peroxidase gekoppelt.

3. Ergebnisse 65
Abb. 3.31. a: SDS-PAGE (15%, Coomassie-Färbung) von Gesamt-Protein. Die Chlorophyllmenge der
aufgetrennten Proteine betrug jeweils 6 µg. M: Molekulargewichtsmarker (10 kDa-Leiter mit 10-200
kDa Referenzproteinen); 1: Mut. 2 (Ala211/Asn214); 2: Mut. 3 (Thr211/Thr214/Ala219); 3: Mut. 10
(Thr211/Ala214); 4: Mut. 13 (Met211/Thr214); 5: Mut. 14 (Gln211); 6: Mut. 21 (Leu211/Gly214); 7:
Mut. 27 (Gly211/Ala214); 8: Mut. 28 (Gln211/Gly214/Ala219); 9: Mut. 32 (Cys211/Thr214/Ala219);
10: Mut. 34 (Gly211/Ser214/Ala219); 11: Mut. 35 (Gly211/Thr214).
Abb. 3.31. b: Immunblot zu Proben aus a. 1. AK: 271-t ist gegen den Bereich des D1-Proteins zwi-
schen Aminosäuren 167 und 353 gerichtet. 2. AK: Ziege-Antikaninchen-AK mit Meerrettich-
Peroxidase gekoppelt.

3. Ergebnisse 66
Abb. 3.32. a: SDS-PAGE (15%, Coomassie-Färbung) von Gesamt-Protein. Die Chlorophyllmenge der
aufgetrennten Proteine betrug jeweils 6 µg. M: Molekulargewichtsmarker (10 kDa-Leiter mit 10-200
kDa-Referenzproteinen); 1: 206/2 (Ile206); 2: 206/3 (Glu206); 3: 206/6 (Ser206); 4: 206/7 (Ala206); 5:
206/9 (His206); 6: 206/10 (Asn206); 7: 206/11 (Val206); 8: 206/18 (Lys206).
Abb. 3.32. b: Immunblot zu Proben aus a. 1. AK: 271-t ist gegen den Bereich des D1-Proteins zwi-
schen Aminosäuren 167 und 353 gerichtet. 2. AK: Ziege-Antikaninchen-AK mit Meerrettich-
Peroxidase gekoppelt.

3. Ergebnisse 67
Abb. 3.33. a: SDS-PAGE (15%, Coomassie-Färbung) von Gesamt-Protein. Die Chlorophyllmenge der
aufgetrennten Proteine betrug jeweils 6 µg. M: Molekulargewichtsmarker (10 kDa-Leiter mit 10-200
kDa-Referenzproteinen); 1: 249/3 (Arg249); 2: 249/5 (His249); 3: 249/8 (Ile249); 4: 249/13 (Ala249); 5:
249/16 (Leu249); 6: 249/17 (Ser249); 7: 249/19 (Asn249); 8: 249/24 (Gln249); 9: 249/28 (Cys249); 10:
249/32 (Gly249).
Abb. 3.33. b: Immunblot zu Proben aus a. 1. AK: 271-t ist gegen den Bereich des D1-Proteins zwi-
schen Aminosäuren 167 und 353 gerichtet. 2. AK: Ziege-Antikaninchen-AK mit Meerrettich-
Peroxidase gekoppelt.

3. Ergebnisse 68 3.6. Messung der Herbizidresistenz von Chlamydomonas reinhardtii-Mutanten
Um den Einfluß der Punktmutationen auf die Bindung von Herbizidmolekülen zu
untersuchen, wurden die pI50-Werte einer Reihe von charakteristischen Hemm-
stoffen verschiedener Typen in den einzelnen Mutanten bestimmt. Der pI50-Wert
ist als negativer dekadischer Logarithmus des I50-Wertes definiert. Als I50-Wert
wird die Konzentration an Herbizid bezeichnet, bei der der Elektronentransfer um
50% reduziert ist. Die pI50-Werte der Mutanten wurden mit den entsprechenden
pI50-Werten beim Wildtyp verglichen und die R/S-Werte (Verhältnis Resistenz zu
Sensitivität) wurden berechnet. Der R/S-Wert ergibt sich aus dem Quotienten der
I50-Werte von Mutante zu Wildtyp. Ein R/S-Wert>1 bedeutet Resistenz, ein R/S-
Wert<1 Supersensitivität. Die Messungen erfolgten an isolierten Thylakoiden. Es
wurde der entkoppelte Elektronentransport von Wasser zu dem artifiziellen Elek-
tronenakzeptor DCPIP in Ab- und Anwesenheit von Hemmstoffen gemessen (vgl.
Kap. 2.12.3.). Die Hemmwirkung wurde bei 7 bis 10 verschiedenen Konzentratio-
nen des Hemmstoffs bestimmt. Die pI50-Werte wurden graphisch ermittelt. Der
Korrelationskoeffizient für die lineare Regression war größer als 0,95. Der in dem
Meßansatz enthaltene Hemmstoff des Cytochrom b6/f-Komplexes DNP-INT
(Trebst et al., 1978) verhindert die Reoxidation von Plastohydrochinon. Dadurch
wird eine Reduktion des DCPIP durch PS I verhindert und nur die Photosynthe-
seaktivität des PS II gemessen.
3.6.1. Messung der Herbizidresistenz der Mutanten mit Aminosäureaus-tauschen an den Positionen Phe 211 und Met 214
Bei den Mutanten mit Aminosäuresubstitutionen an den Positionen 211 und 214
sollten die Hemmeigenschaften einer Reihe von charakteristischen Inhibitoren
untersucht werden. Bereits in der vorhergehenden Arbeit (Orawski, 1998) wurden
5 Mutanten dieser Gruppe hinsichtlich der Bindungsaffinitäten verschiedener
Herbizide analysiert. Es wurde gezeigt, daß die Substitution der beiden Amino-
säuren die Hemmwirkung der Herbizide verändert. Diese Erkenntnisse sollten
durch die Charakterisierung weiterer Mutanten mit Austauschen von Phe 211
und Met 214 vertieft werden. Die ermittelten pI50-Werte der Mutanten sind im
Vergleich zum Wildtyp in den Tabellen 3.5., 3.6. und 3.7. aufgeführt.

3. Ergebnisse 69
WT Mut. 2 Ala211/ Asn214
Mut. 3 Thr211/ Thr214/ Ala219
Mut. 10 Thr211/ Ala214
Mut. 13 Met211/ Thr214
Hemmstoff pI50 pI50 R/S pI50 R/S pI50 R/S pI50 R/S Atrazin 6,4 5,9 3,2 5,5 7,9 6,4 1,0 6,9 0,3 DCMU 7,1 6,7 2,5 7,1 1,0 6,9 1,6 7,6 0,3
Metribuzin 6,7 6,4 2,0 5,9 6,3 6,9 0,6 5,6 12,6 Metamitron 5,5 5,1 2,5 4,8 5,0 4,8 5,0 5,3 1,6
Ioxynil 5,9 5,3 4,0 6,0 0,8 5,8 0,8 5,3 4,0 Phen-
medipham 7,1 6,9 1,6 6,0 12,6 5,9 16,0 6,5 4,0
Bromnitro- thymol
7,1 7,4 0,5 6,7 2,5 7,2 0,8 7,0 1,3
Cyanoacrylat 7,4 6,9 3,2 6,5 7,9 7,0 2,5 7,0 2,5 Hydroxy- chinolin
7,1 8,0 0,1 7,7 0,3 7,5 0,4 6,7 2,5
Ketonitril 7,7 7,6 1,3 7,7 1,0 7,4 2,0 7,1 4,0
Tab. 3.5.: Ergebnisse der Messungen der Herbizidresistenz von Wildtyp und den Mutanten. Die
Messungen erfolgten an Thylakoiden. Meßansatz: 60 µM DCPIP, 2,5 µM Gramicidin, 1 µM DNP-
INT, 10 µg Chlorophyll. Die Strukturformeln der einzelnen Hemmstoffe befinden sich im Anhang.
WT Mut. 14 Gln211
Mut. 21 Leu211/ Gly214
Mut. 27 Gly211/ Ala214
Mut. 28 Gln211/ Gly214/ Ala219
Hemmstoff pI50 pI50 R/S pI50 R/S pI50 R/S pI50 R/S Atrazin 6,4 5,0 25,0 6,7 0,5 6,3 1,3 6,1 2,0 DCMU 7,1 7,2 0,8 6,5 4,0 7,6 0,3 7,6 0,3
Metribuzin 6,7 6,4 2,0 6,5 1,6 6,5 1,6 6,7 1,0 Metamitron 5,5 5,1 2,5 5,2 2,0 5,1 2,5 4,9 4,0
Ioxynil 5,9 6,2 0,5 6,3 0,4 5,5 2,5 5,6 2,0 Phen-
medipham 7,1 6,5 4,0 6,7 2,5 6,1 10,0 5,7 25,0
Bromnitro- thymol
7,1 6,9 1,6 7,8 0,2 7,2 1,3 7,4 0,5
Cyanoacrylat 7,4 6,7 5,0 6,9 3,2 6,6 6,3 6,6 6,3 Hydroxy- chinolin
7,1 7,4 0,5 6,8 2,0 6,5 4,0 7,5 0,4
Ketonitril 7,7 7,4 2,0 7,5 1,6 7,3 2,5 8,3 0,3
Tab. 3.6.: Ergebnisse der Messungen der Herbizidresistenz von Wildtyp und den Mutanten. Die
Messungen erfolgten an Thylakoiden. Meßansatz: 60 µM DCPIP, 2,5 µM Gramicidin, 1 µM DNP-
INT, 10 µg Chlorophyll. Die Strukturformeln der einzelnen Hemmstoffe befinden sich im Anhang.

3. Ergebnisse 70
WT Mut. 32 Cys211/ Thr214/ Ala219
Mut. 34 Gly211/ Ser214/ Ala219
Mut. 35 Gly211/ Thr214
Hemmstoff pI50 pI50 R/S pI50 R/S pI50 R/S Atrazin 6,4 5,7 5,0 5,5 7,9 5,9 3,2 DCMU 7,1 7,4 0,5 6,7 2,5 7,1 1,0
Metribuzin 6,7 6,6 1,3 6,6 1,3 6,6 1,3 Metamitron 5,5 5,4 1,3 5,0 3,2 5,5 1,0
Ioxynil 5,9 5,2 5,0 6,4 0,3 5,8 1,3 Phen-
medipham 7,1 6,0 12,6 5,6 31,6 6,1 10,0
Bromnitro- thymol
7,1 6,6 3,2 7,6 0,3 6,9 1,6
Cyanoacrylat 7,4 6,8 4,0 6,7 5,0 6,6 6,3 Hydroxy- chinolin
7,1 7,3 0,6 7,5 0,4 7,1 1,0
Ketonitril 7,7 7,4 2,0 7,6 1,3 7,2 3,2
Tab. 3.7.: Ergebnisse der Messungen der Herbizidresistenz von Wildtyp und den Mutanten. Die
Messungen erfolgten an Thylakoiden. Meßansatz: 60 µM DCPIP, 2,5 µM Gramicidin, 1 µM DNP-
INT, 10 µg Chlorophyll. Die Strukturformeln der einzelnen Hemmstoffe befinden sich im Anhang.
Die Mutanten 14, 3 und 34 zeigen eine erhöhte Resistenz gegenüber dem Triazin-
Herbizid Atrazin. Bei anderen Mutanten sind die Hemmeigenschaften dieses Tri-
azins nur leicht verändert im Vergleich zum Wildtyp.
Das Harnstoffderivat DCMU zeigt nur leicht veränderte Hemmeigenschaften. Ei-
ne geringe Supersensitivität wurde bei Mutante 13, 27, 28 und 32 beobachtet.
Das Triazinon Metribuzin zeigt eine gesteigerte Resistenz in den Mutanten 13
und 3. Bei dem Triazinon Metamitron sind dagegen keine wesentlichen Unter-
schiede gegenüber dem Wildtyp erkennbar.
Die Wirkung des Cyanophenols Ioxynil ist nur geringfügig beeinflußt.
Am stärksten verändert wurde das Verhalten gegenüber dem Biscarbamat Phen-
medipham. Alle Mutanten zeigen eine gesteigerte Resistenz gegenüber dieser
Substanz.
Für das phenolische Herbizid Bromnitrothymol wurden etwa die gleichen pI50-
Werte in den Mutanten und im Wildtyp gemessen.

3. Ergebnisse 71 Beim Cyanoacrylat wurde eine leicht erhöhte Resistenz aller Mutanten festge-
stellt.
Hydroxychinolin zeigt bei den meisten Mutanten eine verminderte
Bindungsaffinität.
Ketonitril zeigt nur geringe Änderungen der Hemmwirkung in den Mutanten.
3.6.2. Messung der Herbizidresistenz der Mutanten mit Aminosäureaus-tauschen an der Position Val 249
Bei den Mutanten mit Aminosäuresubstitutionen an der Position 249 sollten die
Hemmeigenschaften verschiedener Inhibitoren untersucht werden, darunter einer
Reihe von Harnstoffderivaten, Triazinen, Triazinonen, Nitrophenolen und Cya-
nophenolen. Die ermittelten pI50-Werte der Mutanten sind im Vergleich zum
Wildtyp in den Tabellen 3.8. bis 3.15. aufgeführt.
Hemmstoff
WT 249/3
Arg249 249/5
His249 249/8
Ile249 249/13 Ala249
249/16 Leu249
Phenole pI50 pI50 R/S pI50 R/S pI50 R/S pI50 R/S pI50 R/S
Cyanophenole
Ioxynil 5,9 6,9 0,1 6,9 0,1 6,8 0,13 6,5 0,25 6,3 0,4 Bromoxynil 4,7 5,5 0,16 5,2 0,32 5,0 0,5 5,5 0,16 5,0 0,5
Nitrophenole
Bromnitro- thymol
7,1 7,2 0,8 7,1 1 6,9 1,6 6,9 1,6 7,0 1,26
Dinoterb 6,2 6,2 1 6,5 0,5 5,9 2,0 5,6 4,0 6,3 0,8 Dinoseb 6,0 6,1 0,8 5,8 1,6 5,4 4,0 5,5 3,16 5,6 2,5 DNOC 5,0 5,0 1 4,8 1,6 5,1 0,8 4,5 3,16 4,9 1,26
Tab. 3.8.: Ergebnisse der Messungen der Herbizidresistenz von Wildtyp und den Mutanten gegen-
über Phenolen. Die Messungen erfolgten an Thylakoiden. Meßansatz: 60 µM DCPIP, 2,5 µM Grami-
cidin, 1 µM DNP-INT, 10 µg Chlorophyll. Die Strukturformeln der einzelnen Hemmstoffe befinden
sich im Anhang.

3. Ergebnisse 72
Hemmstoff
WT 249/17 Ser249
249/19 Asn249
249/24 Gln249
249/28 Cys249
249/32 Gly249
Phenole pI50 pI50 R/S pI50 R/S pI50 R/S pI50 R/S pI50 R/S
Cyanophenole
Ioxynil 5,9 6,7 0,16 6,5 0,25 6,9 0,1 6,4 0,32 6,8 0,13 Bromoxynil 4,7 5,4 0,2 5,2 0,32 5,3 0,25 5,7 0,1 5,3 0,25
Nitrophenole
Bromnitro- Thymol
7,1 6,9 1,6 6,7 2,5 6,7 2,5 6,9 1,6 7,2 0,8
Dinoterb 6,2 5,7 3,16 6,0 1,6 6,6 0,4 6,1 1,26 6,0 1,6 Dinoseb 6,0 6,1 0,8 6,2 0,63 6,4 0,4 6,2 0,63 6,1 0,8 DNOC 5,0 4,6 2,5 4,6 2,5 4,2 6,3 5,2 0,63 5,1 0,8
Tab. 3.9.: Ergebnisse der Messungen der Herbizidresistenz von Wildtyp und den Mutanten gegen-
über Phenolen. Die Messungen erfolgten an Thylakoiden. Meßansatz: 60 µM DCPIP, 2,5 µM Grami-
cidin, 1 µM DNP-INT, 10 µg Chlorophyll. Die Strukturformeln der einzelnen Hemmstoffe befinden
sich im Anhang.
Alle Mutanten sind supersensitiv gegenüber den Cyanophenolen Ioxynil und
Bromoxynil.
Das Nitrophenol Bromnitrothymol zeigt eine leicht verminderte Bindungsaffinität
in den Mutanten Asn249 und Gln249.
Die Hemmeigenschaften der Dinitrophenole sind in den einzelnen Mutanten un-
terschiedlich. Die Mutanten Ala249 und Ser249 sind resistent gegenüber Dino-
terb, Ile249, Ala249 und Leu249 gegenüber Dinoseb, Gln249, Ala249, Ser249 und
Asn249 gegenüber DNOC. Supersensitivität wurde nur bei Mutante Gln249 ge-
genüber Dinoterb und Dinoseb beobachtet. Gleichzeitig zeigt aber diese Mutante
eine verminderte Bindungsaffinität zu DNOC.

3. Ergebnisse 73
Hemmstoff
WT 249/3 Arg249
249/5 His249
249/8 Ile249
249/13 Ala249
249/16 Leu249
pI50 pI50 R/S pI50 R/S pI50 R/S pI50 R/S pI50 R/S
Triazine
Atrazin 6,4 6,7 0,5 6,5 0,8 6,3 1,26 6,4 1 6,5 0,8 Atraton 6,2 5,8 2,5 6,3 0,8 6,3 0,8 5,8 2,5 6,5 0,5 Simazin 6,0 6,1 0,8 6,1 0,8 6,4 0,4 6,3 0,5 6,4 0,4
Prometryn 7,1 7,1 1 7,1 1 7,6 0,32 7,2 0,8 7,2 0,8 Terbutryn 8,0 7,8 1,6 7,8 1,6 7,8 1,6 7,8 1,6 8,0 1 Ametryn 7,1 7,6 0,32 7,5 0,4 7,6 0,32 7,5 0,4 7,7 0,25
Prometon 5,7 6,1 0,4 5,7 1 6,2 0,32 5,8 0,8 6,3 0,25
Tab. 3.10.: Ergebnisse der Messungen der Herbizidresistenz von Wildtyp und den Mutanten gegen-
über Triazinen. Die Messungen erfolgten an Thylakoiden. Meßansatz: 60 µM DCPIP, 2,5 µM Grami-
cidin, 1 µM DNP-INT, 10 µg Chlorophyll. Die Strukturformeln der einzelnen Hemmstoffe befinden
sich im Anhang.
Hemmstoff
WT 249/17 Ser249
249/19 Asn249
249/24 Gln249
249/28 Cys249
249/32 Gly249
pI50 pI50 R/S pI50 R/S pI50 R/S pI50 R/S pI50 R/S
Triazine
Atrazin 6,4 6,7 0,5 6,5 0,8 6,4 1 6,5 0,8 6,4 1 Atraton 6,2 6,2 1 6,3 0,8 6,0 1,6 6,7 0,32 5,9 2,0 Simazin 6,0 6,4 0,4 6,5 0,32 6,2 0,63 6,1 0,8 6,3 0,5
Prometryn 7,1 6,9 1,6 7,2 0,8 7,1 1 7,5 0,4 7,2 0,8 Terbutryn 8,0 7,6 2,5 7,9 1,26 7,9 1,26 7,8 1,6 8,0 1 Ametryn 7,1 7,4 0,5 7,6 0,32 7,2 0,8 7,4 0,5 7,3 0,63
Prometon 5,7 6,0 0,5 6,1 0,4 6,0 0,5 6,0 0,5 6,1 0,4
Tab. 3.11.: Ergebnisse der Messungen der Herbizidresistenz von Wildtyp und den Mutanten gegen-
über Triazinen. Die Messungen erfolgten an Thylakoiden. Meßansatz: 60 µM DCPIP, 2,5 µM Grami-
cidin, 1 µM DNP-INT, 10 µg Chlorophyll. Die Strukturformeln der einzelnen Hemmstoffe befinden
sich im Anhang.
Die Bindung der Triazine ist in vielen Fällen positiv beeinflußt. So ist Mutante
Ile249 supersensitiv gegenüber Prometryn, Prometon, Ametryn und Simazin,
Mutante Leu249 gegenüber Prometon, Ametryn und Simazin und Mutante
Asn249 gegenüber Ametryn, Simazin und Prometon. Eine geringe Resistenz wur-
de nur gegen Terbutryn bei der Mutante Ser249 festgestellt.

3. Ergebnisse 74
Hemmstoff
WT 249/3 Arg249
249/5 His249
249/8 Ile249
249/13 Ala249
249/16 Leu249
pI50 pI50 R/S pI50 R/S pI50 R/S pI50 R/S pI50 R/S
Harnstoffe
DCMU 7,1 7,3 0,63 7,6 0,32 7,5 0,4 7,1 1 7,6 0,32 Cycluron 5,6 6,0 0,4 5,6 1 5,8 0,63 5,9 0,5 6,1 0,32 Linuron 7,0 7,7 0,2 7,3 0,5 7,2 0,63 7,1 0,8 7,6 0,25
Chloroxuron 7,7 7,4 2 7,3 2,5 7,5 1,6 7,6 1,26 7,6 1,26 Neburon 6,9 7,6 0,2 7,7 0,16 7,5 0,25 7,6 0,2 7,8 0,13
Chlortoluron 6,8 7,2 0,4 6,9 0,8 6,9 0,8 6,8 1 7,2 0,4 Benzthiazuron 6,5 6,4 1,3 6,3 1,6 6,7 0,63 6,5 1 6,8 0,5
Metabenz- thiazuron
6,5 6,4 1,3 6,2 2,0 6,5 1 6,7 0,63 6,5 1
Fluometuron 5,7 5,9 0,63 6,0 0,5 6,0 0,5 5,8 0,8 6,2 0,32 Thiazfluron 5,8 5,7 1,26 6,0 0,63 6,1 0,5 6,5 0,2 6,1 0,5
Buturon 5,7 6,0 0,5 6,2 0,32 6,2 0,32 6,7 0,1 6,2 0,32 Fenuron 5,6 5,5 1,26 5,3 2,0 5,4 1,6 5,3 2,0 5,8 0,63
Tab. 3.12.: Ergebnisse der Messungen der Herbizidresistenz von Wildtyp und den Mutanten gegen-
über Harnstoffen. Die Messungen erfolgten an Thylakoiden. Meßansatz: 60 µM DCPIP, 2,5 µM Gra-
micidin, 1 µM DNP-INT, 10 µg Chlorophyll. Die Strukturformeln der einzelnen Hemmstoffe befinden
sich im Anhang.
Hemmstoff
WT 249/17 Ser249
249/19 Asn249
249/24 Gln249
249/28 Cys249
249/32 Gly249
pI50 pI50 R/S pI50 R/S pI50 R/S pI50 R/S pI50 R/S
Harnstoffe
DCMU 7,1 7,2 0,8 7,7 0,25 7,0 1,26 7,0 1,26 7,4 0,5 Cycluron 5,6 6,0 0,4 5,8 0,63 5,8 0,63 5,8 0,63 6,0 0,4 Linuron 7,0 7,5 0,32 7,5 0,32 7,2 0,63 7,0 1 7,6 0,25
Chloroxuron 7,7 7,5 1,6 7,4 2,0 7,6 1,26 7,4 2,0 7,7 1 Neburon 6,9 7,7 0,16 7,6 0,2 7,7 0,16 7,5 0,25 7,7 0,16
Chlortoluron 6,8 7,3 0,32 7,0 0,63 6,9 0,8 6,9 0,8 7,1 0,5 Benzthiazuron 6,5 6,5 1 6,6 0,8 6,5 1 6,8 0,5 6,8 0,5
Metabenz- thiazuron
6,5 6,6 0,8 6,4 1,3 6,4 1,3 6,6 0,8 6,5 1
Fluometuron 5,7 5,8 0,8 6,0 0,5 6,1 0,4 6,0 0,5 6,4 0,2 Thiazfluron 5,8 6,1 0,5 6,1 0,5 5,6 1,6 5,9 0,8 6,3 0,32
Buturon 5,7 6,1 0,4 6,2 0,32 6,2 0,32 6,0 0,5 6,4 0,2 Fenuron 5,6 5,4 1,6 5,7 0,8 5,4 1,6 5,4 1,6 5,7 0,8
Tab. 3.13.: Ergebnisse der Messungen der Herbizidresistenz von Wildtyp und den Mutanten gegen-
über Harnstoffen. Die Messungen erfolgten an Thylakoiden. Meßansatz: 60 µM DCPIP, 2,5 µM Gra-
micidin, 1 µM DNP-INT, 10 µg Chlorophyll. Die Strukturformeln der einzelnen Hemmstoffe befinden
sich im Anhang.

3. Ergebnisse 75 Ein ähnliches Bild ergibt sich für die getesteten Harnstoffderivate. Bei keiner
Mutante wurde Resistenz beobachtet. Die Mutanten Leu249 und Gly249 sind su-
persensitiv gegenüber den meisten getesteten Harnstoffen. Auch Mutanten mit
polaren Aminosäuren an der Position 249 zeigen erhöhte Bindungsaffinitäten für
diese Herbizide. Mutante Ser249 ist supersensitiv gegenüber Neburon, Chlortolu-
ron, Linuron, Cycluron und Buturon, Mutante Asn249 gegenüber Neburon,
DCMU, Linuron und Buturon, Mutante Arg249 gegenüber Neburon, Linuron,
Cycluron und Chlortoluron. Neburon und Buturon zeigen in allen untersuchten
Mutanten gesteigerte Bindungsaffinitäten.
Hemmstoff
WT 249/3 Arg249
249/5 His249
249/8 Ile249
249/13 Ala249
249/16 Leu249
pI50 pI50 R/S pI50 R/S pI50 R/S pI50 R/S pI50 R/S
Triazinone
Metribuzin 6,7 7,0 0,5 7,1 0,4 6,7 1 6,9 0,63 7,2 0,32 Metamitron 5,5 5,6 0,8 5,5 1 5,5 1 5,5 1 5,6 0,8
Biscarba-mate
Phen- medipham
7,1 7,0 1,26 7,3 0,63 6,9 1,6 6,8 2,0 7,1 1
Phenisopham 6,9 7,1 0,63 6,6 2,0 6,9 1 6,9 1 6,8 1,26
Sonstige
Hydroxy- chinolin
7,1 7,8 0,2 7,5 0,4 7,3 0,63 7,1 1 7,5 0,4
Ketonitril 7,7 7,4 2 7,8 0,8 7,7 1 8,1 0,4 7,9 0,63 Cyanoacrylat 7,4 7,9 0,32 8,1 0,2 7,8 0,4 7,8 0,4 8,0 0,25
Tab. 3.14.: Ergebnisse der Messungen der Herbizidresistenz von Wildtyp und den Mutanten. Die
Messungen erfolgten an Thylakoiden. Meßansatz: 60 µM DCPIP, 2,5 µM Gramicidin, 1 µM DNP-
INT, 10 µg Chlorophyll. Die Strukturformeln der einzelnen Hemmstoffe befinden sich im Anhang.

3. Ergebnisse 76
Hemmstoff
WT 249/17 Ser249
249/19 Asn249
249/24 Gln249
249/28 Cys249
249/32 Gly249
pI50 pI50 R/S pI50 R/S pI50 R/S pI50 R/S pI50 R/S
Triazinone
Metribuzin 6,7 6,6 1,26 6,9 0,63 7,0 0,5 7,0 0,5 7,0 0,5 Metamitron 5,5 5,6 0,8 5,4 1,26 5,5 1 5,4 1,26 5,7 0,63
Biscarba-mate
Phen- medipham
7,1 6,9 1,6 7,0 1,26 6,8 2,0 6,9 1,6 7,1 1
Phenisopham 6,9 7,5 0,25 7,2 0,5 6,9 1 7,0 0,8 6,9 1
Sonstige
Hydroxy- chinolin
7,1 7,2 0,8 7,9 0,16 7,5 0,4 7,2 0,8 7,6 0,32
Ketonitril 7,7 8,1 0,4 7,6 1,26 7,8 0,8 7,7 1 7,7 1 Cyanoacrylat 7,4 7,6 0,63 7,9 0,32 7,8 0,4 7,9 0,32 7,9 0,32
Tab. 3.15.: Ergebnisse der Messungen der Herbizidresistenz von Wildtyp und den Mutanten. Die
Messungen erfolgten an Thylakoiden. Meßansatz: 60 µM DCPIP, 2,5 µM Gramicidin, 1 µM DNP-
INT, 10 µg Chlorophyll. Die Strukturformeln der einzelnen Hemmstoffe befinden sich im Anhang.
Bei den beiden getesteten Triazinonen sind im Fall von Metamitron keine Ände-
rungen in der Hemmwirkung zu verzeichnen. Metribuzin ist in den Mutanten
Leu249 und His249 leicht supersensitiv.
Die eingeführten Mutationen haben keine großen Auswirkungen auf die Bindung
der Biscarbamate Phenmedipham und Phenisopham.
Die Hemmeigenschaften des phenolischen Herbizids Ketonitril sind durch die
meisten Mutationen nicht beeinflußt. Das andere phenolische Herbizid Hydroxy-
chinolin zeigt in den Mutanten Asn249, Arg249, Gly249, His249, Leu249 und
Gln249 eine erhöhte Bindungsaffinität.
Für Cyanoacrylat ergibt sich ein einheitliches Bild. Seine Bindungsaffinität ist in
allen Mutanten leicht gestiegen.

3. Ergebnisse 77 3.6.3. Messung der Herbizidresistenz der Mutanten mit Aminosäureaus-tauschen an der Position Phe 206
Die bis jetzt bekannten Aminosäuresubstitutionen im D1-Protein, die das Bin-
dungsverhalten der Hemmstoffe verändern, konzentrieren sich auf dem Bereich
zwischen den Aminosäuren Phe 211 und Leu 275. Dieser Bereich wird als Herbi-
zid-Bindenische bezeichnet.
Eine Ausnahme stellt eine Mutante von dem Cyanobakterium Synechocystis sp.
PCC 6803 dar, bei der Phe 295 gegen Ser ausgetauscht ist (Narusaka et al., 1998).
Diese Mutation führt zu leicht erhöhter Resistenz gegenüber Atrazin (R/S-
Wert=2).
Ein anderer Aminosäureaustausch außerhalb des als Herbizid-Bindenische defi-
nierten Bereichs wurde bei einer Dreifachmutante von Chenopodium rubrum be-
schrieben (Schwenger-Erger et al., 1999). Es handelt sich um eine Metribuzin-
resistente Zellinie mit den Aminosäuresubstitutionen: Ile 184 gegen Asn, Val 219
gegen Ile und Ala 251 gegen Val.
Bereits in der vorhergehenden Arbeit (Orawski, 1998) wurde eine Dreifachmutan-
te von Chlamydomonas reinhardtii erzeugt, bei der folgende Aminosäuren ausge-
tauscht waren: Phe 206 gegen Tyr, Phe 211 gegen Ala und Met 214 gegen Thr.
Diese Mutante zeigte ein verändertes Verhalten gegenüber Hemmstoffen. Die
Charakterisierung der im Rahmen dieser Dissertation erzeugten Mutanten mit
Aminosäuresubstitutionen an der Position Phe 206 hinsichtlich der Bindung von
3 repräsentativen Hemmstoffen sollte zeigen, ob diese Mutation außerhalb der bis
jetzt bekannten Herbizid-Bindenische allein das Bindungsverhalten der Herbizi-
de beeinflußt. Die ermittelten pI50-Werte der Mutanten sind im Vergleich zum
Wildtyp in der Tabelle 3.16. aufgeführt.

3. Ergebnisse 78
WT 206/2 Ile206
206/3 Glu206
206/6 Ser206
206/7 Ala206
Hemmstoff pI50 pI50 R/S pI50 R/S pI50 R/S pI50 R/S Atrazin 6,4 6,7 0,5 6,7 0,5 6,9 0,32 6,7 0,5 DCMU 7,1 7,1 1 7,0 1,3 7,3 0,6 7,1 1 Phen-
medipham 7,1 7,8 0,2 7,4 0,5 7,7 0,25 7,6 0,32
WT 206/9 His206
206/10 Asn206
206/11 Val206
206/18 Lys206
Hemmstoff pI50 pI50 R/S pI50 R/S pI50 R/S pI50 R/S Atrazin 6,4 6,6 0,6 6,7 0,5 6,7 0,5 6,9 0,32 DCMU 7,1 7,0 1,3 7,0 1,3 7,0 1,3 7,2 0,8 Phen-
medipham 7,1 7,3 0,6 7,3 0,6 7,7 0,25 7,6 0,32
Tab. 3.16.: Ergebnisse der Messungen der Herbizidresistenz von Wildtyp und den Mutanten. Die
Messungen erfolgten an Thylakoiden. Meßansatz: 60 µM DCPIP, 2,5 µM Gramicidin, 1 µM DNP-
INT, 10 µg Chlorophyll. Die Strukturformeln der einzelnen Hemmstoffe befinden sich im Anhang.
Das getestete Triazin Atrazin zeigt in allen Mutanten eine leicht gesteigerte Bin-
dungsaffinität.
Die Hemmeigenschaften des Harnstoffs DCMU sind in den Mutanten und im
Wildtyp gleich.
Die Bindung des Biscarbamats Phenmedipham ist in vielen Fällen positiv
beeinflußt. Die Mutanten Ile206, Val206, Ser206, Lys206 und Ala206 sind super-
sensitiv gegenüber diesem Hemmstoff.

4. Diskussion 79
4. Diskussion
Das Photosystem II ist ein Multiproteinkomplex. Sein Kern wird von einem Hete-
rodimer gebildet, der aus den Proteinen D1 und D2 besteht. Diese Proteine tragen
die Cofaktoren der photosynthetischen Elektronentransportkette.
Die dreidimensionale Struktur des Photosystems II ist noch nicht endgültig auf-
geklärt. Die kristallographischen Daten liefern Informationen über die Anzahl
und räumliche Anordnung der Sekundärstrukturelemente der Proteinunterein-
heiten und über die räumliche Verteilung der Cofaktoren (Rhee et al., 1998; Han-
kamer et al., 1999; Zouni et al., 2001). Das D1- und D2-Protein besitzen jeweils 5
transmembrane α-Helices, bezeichnet als Helix A bis E (bzw. I bis V). Zwischen
den transmembranen Helices C und D sowie D und E befinden sich parallel zur
Thylakoidmembran verlaufende Helices, bezeichnet als Helix C-D (bzw. III/IV)
und D-E (bzw. IV/V). Helix C-D liegt an der Lumenseite der Membran, Helix D-E
an der Stromaseite (Abb. 4.1.). Genaue Angaben zu den Aminosäuren, der Aus-
richtung ihrer Seitenketten und der Fixierung der prosthetischen Gruppen fehlen.
Abb. 4.1.: Zweidimensionales Strukturmodell des D1-Proteins. Das D1-Protein besitzt 5 trans-
membrane und 2 parallel zur Thylakoidmembran verlaufende Helices (nach Trebst, 1987).
Für die detaillierte dreidimensionale Faltung des D1/D2-Heterodimers wurden
Modelle vorgeschlagen (Trebst, 1987; Tietjen et al., 1991; Ruffle et al., 1992; Xiong

4. Diskussion 80 et al., 1996; Xiong et al., 1998). Als Grundlage für diese Modelle dient die Rönt-
genstruktur des Reaktionszentrums der Purpurbakterien, dessen L- und M-
Untereinheit Homologien mit den Untereinheiten D1 und D2 des Photosystems II
aufweisen. Diese Modelle werden durch Hemmstoffexperimente unterstützt.
Der photosynthetische Elektronentransport kann an mehreren Stellen unterbun-
den werden. Einer der Angriffsorte der Photosynthesehemmstoffe ist die Akzep-
torseite von Photosystem II. Zahlreiche Substanzen verdrängen den sekundären
Elektronenakzeptor QB aus seiner Bindenische im D1-Protein, wodurch der Elekt-
ronentransport zum Erliegen kommt. Diese Substanzen sind von großem Interes-
se. Einerseits tragen sie zum Verständnis der Struktur und Funktion des Reakti-
onszentrums von PS II bei, insbesondere der QB-Bindenische, andererseits werden
viele von ihnen kommerziell als Herbizide eingesetzt. 20% aller eingesetzten Her-
bizide sind Photosystem II-Hemmstoffe.
Für die Charakterisierung des D1-Proteins und seiner QB-Bindenische mit Hilfe
von Hemmstoffen können zwei Methoden angewendet werden. Die eine ist die
Photoaffinitätsmarkierung, die andere die Analyse herbizidtoleranter Mutanten.
Bei Photoaffinitätsmarkierungen wird der Hemmstoff mit einer Azidogruppe ver-
sehen. Nach Inkubation der Thylakoiden mit dem Hemmstoffderivat wird aus der
Azidogruppe mit einer UV-Lichtquelle Stickstoff abgespalten. Dabei entsteht ein
extrem elektrophiles Nitren, das sofort kovalent an ein Protein bindet. Der Ein-
satz eines radioaktiv markierten Azido-Herbizids (Azido-[14C]atrazin) führte zur
Erkenntnis, daß eine der Untereinheiten des Photosystems II, das D1-Protein, die
Hemmstoffe bindet (Pfister et al., 1981). Photoaffinitätsmarkierungen ermöglichen
auch die Identifizierung von Aminosäuren, die direkt an der Bindung von Herbi-
ziden beteiligt sind. Mittels Proteinsequenzierung eines photoaffinitätsmarkierten
Proteins kann festgestellt werden, an welche Aminosäure das Nitrenderivat ge-
bunden ist. Auf diese Weise wurde Met 214 im D1-Protein als Bindungsort von
Azido-[14C]atrazin erkannt (Wolber et al., 1986; Whitelegge et al., 1992). Tyr 237
und Tyr 254 wurden durch Photoaffinitätsmarkierung mit Azido-[14C]monouron,

4. Diskussion 81 einem Analogon von DCMU, identifiziert (Dostatni et al., 1988). Val 249 wurde
mit Azido-[125I]ioxynil markiert (Oettmeier et al., 1989).
Eine andere Methode, die auf dem Einsatz von Hemmstoffen basiert, ist die Ana-
lyse herbizidtoleranter Mutanten. Die Sequenzierung des in freier Natur gefun-
denen Atrazin-resistenten Amaranthus hybridus bestätigte, daß das D1-Protein
des Reaktionszentrums von Photosystem II das Herbizid-Bindeprotein ist
(Hirschberg und McIntosh., 1983). Es wurde gezeigt, daß die Herbizidresistenz
dieser Pflanze auf einer Aminosäuresubstitution von Ser 264 zu Gly im D1-
Protein beruht. Die gleiche Mutation trat in weiteren herbizidtoleranten Pflanzen
auf. Sie wurde bei Solanum nigrum (Hirschberg et al., 1984; Goloubinoff et al.,
1984), Senecio vulgaris (Bleyden und Gray, 1986), Chenopodium album (Bettini et
al., 1987), Phalaris paradoxa (Schönfeld et al., 1987; Ohad und Hirschberg, 1990),
Amaranthus retroflexus und Amaranthus bouchonii (Solymosi und Lehoczki,
1989) beschrieben.
Weitere zahlreiche Mutationen des D1-Proteins in Cyanobakterien, einzelligen
Algen und in Zellkulturen höherer Pflanzen wurden entweder durch gezielte Mu-
tagenese oder durch Selektion gegen Herbizide erzeugt. Die dabei herbeigeführ-
ten Aminosäureaustausche verändern die Bindungsaffinitäten der Herbizide.
Diese Aminosäureaustausche konzentrieren sich auf dem Bereich zwischen den
Aminosäuren Phe 211 und Leu 275. Es wurden Einfach-, Doppel- und Dreifach-
mutanten charakterisiert (Abb. 4.2.).

4. Diskussion 82 F N F M I V F Q A E H N I L M H P F H M L G V A G 180 190 200
V F G G S L F S A M H G S L V T S S L I R E T T E 210 Helix D 220
N E S A N E G Y R F G Q E E E T Y N I V A A H G Y 230 240 250 Helix
F G R L I F Q Y A S F N N S R S L H F F L A A W P D-E 260 270
V I G I W F T A L G L S T M A F N L N G F N F N Q 280 Helix E 290 300 Abb. 4.2.: Aminosäuresequenz des D1-Proteins im Bereich zwischen Phe 180 und Glu 304. Die Ami-
nosäuren, deren Austausche zu einer veränderten Hemmwirkung von Herbiziden führen, sind fett
geschrieben. Kursiv erscheinen Aminosäuren, die durch Photoaffinitätsmarkierung identifiziert
wurden.
Die Mutation von Phe 211 wurde bei Cyanobakterien identifiziert. Bei Synecho-
coccus sp. PCC 7002 wurde die Mutation von Phe 211 zu Ser beschrieben (Gin-
grich et al., 1988). Dieser Aminosäureaustausch verleiht dem Bakterium Kreuzre-
sistenz gegenüber Atrazin und Diuron. Die gleiche Aminosäuresubstitution führt
bei einer Synechocystis sp. PCC 6714 Mutante zu Kreuzresistenz gegenüber Atra-
zin und Metribuzin (Kirilovsky et al., 1989; Govindjee et al., 1990). Die Mutation
von Val 219 zu Ile wurde bei Chlamydomonas reinhardtii beschrieben. Sie führt
zur Resistenz gegenüber Metribuzin (R/S-Wert=200) und Diuron (R/S-Wert=15-
32) (Erickson et al., 1985; Erickson et al., 1989; Wildner et al., 1990). Die gleiche
Mutation ist bei dem Cyanobakterium Synechococcus sp. PCC 7002 bekannt, bei
dem sie ebenfalls Diuronresistenz hervorruft (Gingrich et al., 1988). Die Substitu-
tion von Ile 248 gegen Thr führt bei einer Mutante von Synechocystis sp. PCC
6714 zur Metribuzinresistenz (Perewoska et al., 1994). 7 verschiedene Aminosäu-
reaustausche (Arg, Asn, Asp, His, Ile, Phe, Tyr) wurden an der Position Ala 250
bei Chlamydomonas reinhardtii eingeführt. Diese Mutanten sind resistent gegen-
über Phenmedipham und supersensitiv gegenüber phenolischen Herbiziden (Jo-
hanningmeier et al., 2000). An der Position Ala 251 sind bei Chlamydomonas
reinhardtii 5 Mutationen beschrieben, die das Bindungsverhalten von Herbiziden
beeinflussen. Der Austausch gegen Val verleiht der Alge eine hohe Metribuzinre-

4. Diskussion 83 sistenz (R/S-Wert=1000) (Pucheu et al., 1984; Johanningmeier et al., 1987b). Die
Austausche gegen Ile und Leu vermindern ebenfalls die Bindungsaffinität von
Metribuzin (Förster et al., 1997). Die Austausche gegen Cys und Gly führen zur
Resistenz gegenüber phenolischen Herbiziden (Johanningmeier et al., 2000). Der
in Chlamydomonas reinhardtii zur Metribuzinresistenz führende Aminosäure-
austausch von Ala 251 gegen Val wurde auch in dem Cyanobakterium Synecho-
cystis sp. PCC 6714 erzeugt und führt bei dem Bakterium ebenfalls zur Metribu-
zinresistenz (R/S-Wert=200) (Perewoska et al., 1994). Weitere Mutationen an der
Position Ala 251 wurden in Synechocystis sp. PCC 6803 durchgeführt. Die Amino-
säuresubstitutionen von Ala 251 gegen Asp, Ser, Leu und Lys verleihen den Mut-
anten Diuronresistenz (Mäenpää et al., 1998). Die Mutation von Phe 255 zu Tyr
führt zur Atrazinresistenz. Dies wurde sowohl bei Chlamydomonas reinhardtii
(Galloway und Mets, 1984; Erickson et al., 1985; Erickson et al., 1989) als auch bei
Synechococcus sp. PCC 7942 beobachtet (Ohad et al., 1990). Kreuzresistenz ge-
genüber Atrazin und Diuron wurde bei Mutanten beobachtet, bei denen die Ami-
nosäuren Gly 256 und Arg 257 substituiert waren. Eine Mutation von Gly 256 zu
Asp mit dieser Wirkung wurde in Chlamydomonas reinhardtii erzeugt (Erickson
et al., 1989), eine Mutation von Arg 257 zu Val in Synechocystis sp. PCC 6803
(Kless et al., 1994). Der Austausch von Ala 263 gegen Pro wurde in Synechocystis
sp. PCC 6803 durchgeführt und verursacht eine hohe Resistenz gegenüber Atra-
zin (R/S-Wert=2000), Metribuzin (R/S-Wert=1600), Terbutryn (R/S-Wert=160)
und Diuron (R/S-Wert=60) (Dalla Chiesa et al., 1997). Der Austausch von Ser 264
beeinflußt am stärksten die Bindungsaffinitäten von Hemmstoffen. Die Mutation
von Ser 264 zu Gly ist, wie bereits erwähnt, die einzige Mutation, die bei höheren
Pflanzen in vivo vorkommt. Sie verleiht den Pflanzen Atrazinresistenz mit R/S-
Werten bis zu 3000. Der Austausch von Ser 264 gegen Gly wurde durch ortsge-
richtete Mutagenese in das Cyanobakterium Synechococcus sp. PCC 7942 einge-
führt. Diese Mutante zeigt Resistenz gegenüber Metribuzin (R/S-Wert=1500), ver-
schiedenen Triazinen und Harnstoffen (Ohad und Hirschberg, 1990; Ohad et al.,
1990). Der Austausch von Ser 264 gegen andere Aminosäuren verändert ebenfalls
die Hemmwirkung von Herbiziden. Bei Chlamydomonas reinhardtii wurde eine
Mutation zu Ala erzeugt, die Resistenz gegenüber Metribuzin (R/S-Wert=5000-

4. Diskussion 84 10000), Atrazin (R/S-Wert=125-500) und Diuron (R/S-Wert=200) hervorruft (Ja-
natkova und Wildner, 1982; Pucheu et al., 1984; Przibilla et al., 1991). Ähnliches
Hemmverhalten wird durch die gleiche Mutation bei Synechocystis sp. PCC 6714
bewirkt (Ajlani et al., 1989a; Govindjee et al., 1990). In Zellkulturen höherer
Pflanzen wurden weitere Aminosäuresubstitutionen von Ser 264 gefunden. Eine
Terbutryn-resistente Zellkultur von Nicotiana plumbaginifolia weist die Mutation
zu Asn auf (Pay et al., 1988). Eine Mutation zu Thr wurde in Zellkulturen von
Nicotiana tabacum (Sato et al., 1988) und Solanum tuberosum (Smeda et al., 1993)
identifiziert. Beide Zellinien zeigen Atrazinresistenz. Der Austausch von Ser 264
gegen Pro wurde bei dem Cyanobakterium Synechocystis sp. PCC 6803 erzeugt
und verursacht ebenfalls Atrazinresistenz (R/S-Wert=10000) (Dalla Chiesa et al.,
1997). An der Position Asn 266 wurden bei Synechocystis sp. PCC 6714 zwei
Austausche gefunden, gegen Thr (Ajlani et al., 1989b) und gegen Asp (Creuzet et
al., 1990). Beide Mutanten sind resistent gegenüber Ioxynil. Die Substitution von
Arg 269 gegen Gly führt bei einer Mutante von Chlamydomonas reinhardtii zur
Terbutrynresistenz (Xiong et al., 1997). Der Austausch von Leu 275 gegen Phe ist
bei Chlamydomonas reinhardtii bekannt und führt zur Resistenz gegenüber
Metribuzin und Diuron (Erickson et al., 1989; Wildner et al., 1990).
Veränderte Bindungsaffinitäten der Hemmstoffe wurden bei einer Reihe von Dop-
pelmutanten beobachtet. Bei einigen treten Mutationen auf, die bereits bei Ein-
zelmutanten identifiziert wurden, bei anderen ist eine der beiden Mutationen be-
reits in Einzelmutanten bekannt. Durch ortsgerichtete Mutagenese wurden 5 ver-
schiedene Mutanten von Chlamydomonas reinhardtii mit Aminosäure-
austauschen an den Positionen Phe 211 und Met 214 erzeugt. Alle Mutanten zei-
gen Resistenz gegenüber Atrazin und Phenmedipham (Orawski, 1998). Die Muta-
tion von Phe 211 kommt, wie bereits erwähnt, in Einzelmutanten von Cyanobak-
terien vor. Der Austausch von Met 214 ist in Einzelmutanten nicht bekannt. Eine
Mutation, die ebenfalls nicht in Einzelmutanten auftritt, ist der Austausch von
Leu 218 gegen Phe bei einer Doppelmutante von Euglena gracilis. Die zweite Mu-
tation betrifft Ser 264 (bei Euglena gracilis Leu 219 und Ser 265, wegen der zu-
sätzlichen Aminosäure am Aminoterminus). Diese Mutante ist resistent gegen-
über Diuron und Atrazin (Thuillier-Bruston et al., 1996). In Doppelmutanten von

4. Diskussion 85 Synechocystis sp. PCC 6803 wurden Austausche von Ser 221 gegen Leu, Ser 222
gegen Ala sowie Thr 227 gegen Ala, Thr 228 gegen Ala erzeugt. Beide Mutanten
zeigen Resistenz gegenüber DCMU und Atrazin (Kless et al., 1994).
Die Analyse herbizidtoleranter Mutanten erlaubt die Identifizierung von Amino-
säuren, die für die Herbizidwirkung essentiell sind, und liefert Informationen über
den Mechanismus der Hemmstoffbindung. Diese Erkenntnisse bilden die Grund-
lage für die zukünftige Entwicklung künstlicher Herbizidresistenzen in Nutz-
pflanzen einerseits, und für das Modellieren neuer wirksamer Herbizide anderer-
seits.
Im Rahmen dieser Arbeit wurde die Mutantenanalyse für die Charakterisierung
der QB-Bindenische im Photosystem II von Chlamydomonas reinhardtii ange-
wandt.
Die Punktmutationen in dem für das D1-Protein codierenden psbA-Gen wurden
mit Hilfe der Oligonukleotid-gerichteten Mutagenese erzeugt. Die Codons für fol-
gende Aminosäuren wurden mutagenisiert: Phe 206, His 215 und Val 249. Die
Einschleusung der in vitro-manipulierten DNA des psbA-Gens in den Organismus
erfolgte mit Hilfe der Partikelkanone, die von Klein und Sanford entwickelt wurde
(Klein et al., 1987; Boynton et al., 1988). Die Technik der particle gun ermöglicht
die Transformation von Zellorganellen. Das Prinzip dieser Methode beruht dar-
auf, daß sehr kleine, mit DNA beladene Wolframpartikel durch eine Treibladung
beschleunigt werden, die Zellwand durchdringen und in den Chloroplasten gelan-
gen können. Im Chloroplasten löst sich die DNA vom Partikel und steht zur Re-
kombination zur Verfügung
Im Rahmen dieser Dissertation wurde die Chloroplastentransformation direkt mit
PCR-Produkten durchgeführt. Plasmide als Träger der einzuschleusenden DNA
wurden nicht verwendet. Diese Abwandlung, deren Vorteil im Wegfall der Klonie-
rungsschritte liegt, wurde bereits in der vorhergehenden Diplomarbeit etabliert.
Die einzellige Grünalge Chlamydomonas reinhardtii bietet als Modellorganismus
im Vergleich mit höheren Pflanzen mehrere Vorteile. Der PS II-Komplex der Alge

4. Diskussion 86 entspricht strukturell und funktionell dem PS II-Komplex der höheren Pflanzen.
Die Alge ist im Labor leicht zu kultivieren und hat eine kurze Generationsdauer.
Der Chloroplast der Grünalge kann relativ leicht mit der Partikelkanone trans-
formiert werden, und das Chloroplastengenom ist zur homologen Rekombination
mit eingebrachter DNA fähig. Im Gegensatz zu höheren Pflanzen stehen Mutan-
ten zur Verfügung, die trotz Defekten im Photosyntheseapparat heterotroph mit
Acetat als Kohlenstoffquelle wachsen können. Der Nachteil bei der Verwendung
von Chlamydomonas reinhardtii liegt in der Struktur des psbA-Gens. Die Alge
besitzt zwei Kopien dieses Gens in der inverted repeat-Region des Chloroplasten-
genoms. Außerdem ist der codierende Bereich des Gens durch vier Introns unter-
brochen, was die Mutagenese wegen der Größe der einzuschleusenden DNA er-
schwert. Dieses Problem konnte umgangen werden, da für die Transformation die
Chlamydomonas reinhardtii-Mutante Del1 zur Verfügung stand (Johanningmeier
et al., 2000). Diese Mutante zeichnet sich durch Modifikationen ihres psbA-Gens
aus. Del1 besitzt ein intronloses psbA-Gen mit einer 425 bp großen Deletion, die
die Zerstörung des Leserahmens zur Konsequenz hat. Infolgedessen codiert das
Gen für ein deletiertes D1-Protein, das aus den ersten 152 Aminosäuren und einer
fremden Sequenz von 30 Aminosäuren am Carboxyterminus besteht. Der Mutan-
te fehlen somit die Bindestellen für die Cofaktoren P680 und Fe, sowie die Binde-
nische für den sekundären Chinonakzeptor QB. Sie wächst ausschließlich hete-
rotroph, weil sie keine PS II-Aktivität besitzt. Hierdurch war es möglich, nach der
Transformation photosynthetisch kompetente von photosynthetisch inaktiven Zel-
len zu selektieren. Auf diese Weise können Mutationen in das psbA-Gen einge-
führt werden, die die Plastochinonreduktion erlauben, aber unbekannte Auswir-
kungen auf die Bindung von Inhibitoren haben. Es ist jedoch nicht möglich,
Transformanten zu identifizieren, bei denen die eingebrachten Mutationen die
Inaktivierung der Photosynthese zur Folge haben und damit für den Organismus
letal sind.

4. Diskussion 87 4.1. Charakterisierung der Mutanten mit Aminosäureaustauschen an der Position Val 249 im D1-Protein
Die Aminosäure Val 249 befindet sich nach den Modellen für das D1-Protein am
Anfang der parallel zur Thylakoidmembran verlaufenden Helix D-E. Sie wurde
durch Photoaffinitätsmarkierung als Bindungsort von Azido-[125I]ioxynil, einem
Cyanophenol, identifiziert (Oettmeier et al., 1989).
Im Rahmen dieser Arbeit wurden Mutanten von Chlamydomonas reinhardtii mit
Aminosäuresubstitutionen an der Position Val 249 erzeugt. Das mutagene Oligo-
nukleotid für den Austausch von Val 249 war an 3 Stellen gewobbelt, so daß alle
64 Tripletts möglich waren und folglich die Wildtypsequenz nicht ausgeschlossen
war. Nach der Transformation wurde mit Hilfe der PCR-Analyse festgestellt, daß
alle Kopien des Plastoms in den Transformanten wieder das vollständige psbA-
Gen enthalten und damit alle Transformanten homoplasmisch sind. Mittels DNA-
Sequenzierung wurde die Identität der einzelnen Transformanten bestimmt. Von
42 Transformanten stellten sich 19 als Mutanten heraus, darunter sind 10 ver-
schiedene Mutanten vorhanden. Folgende Aminosäureaustausche wurden er-
reicht: Ala, Arg, Asn, Cys, Gln, Gly, His, Ile, Leu und Ser.
Das D1-Protein zeichnet sich durch eine hohe Konservierung seiner Sequenz aus
(Jansson und Mäenpää, 1997). Eine drastische Sequenzmodifikation kann daher
potentiell seine Funktion beeinträchtigen oder ganz ausschalten. Das photoau-
totrophe Wachstum der Mutanten schloß eine Funktionsunfähigkeit ihrer D1-
Proteine bereits aus. Ob die Mutationen die photosynthetische Leistung beein-
trächtigen, wurde mit physiologischen Experimenten überprüft. Dazu wurden die
Wachstumskurven der Mutanten im Vergleich zum Wildtyp und zu der intronlo-
sen Mutante IL aufgenommen. Es wurden keine signifikanten Unterschiede in
dem Wachstum festgestellt.
4.1.1. Hemmstoffexperimente
Der Einfluß der Punktmutationen an der Position Val 249 auf die Bindung von
Herbizidmolekülen wurde für verschiedene Klassen der Hemmstoffe untersucht,
nämlich Phenole, unter Phenolen waren Cyanophenole und Nitrophenole vertre-

4. Diskussion 88 ten, Triazine, Harnstoffderivate, Triazinone und Biscarbamate. Außerdem wurde
die Bindungsaffinität von Hydroxychinolin, Ketonitril und Cyanoacrylat be-
stimmt.
Gly 249
Bei der Mutante 249/32 wurde Valin, eine Aminosäure mit aliphatischer Seiten-
kette, gegen Glycin, die Aminosäure mit dem minimalen Raumbedarf, ausge-
tauscht. Diese Mutante zeigt Supersensitivität gegenüber den Cyanophenolen
Ioxynil und Bromoxynil (R/S-Werte<1). Die Bindung der Nitrophenole wurde da-
gegen nicht beeinflußt (R/S-Werte≅1). Die erhöhte Bindungsaffinität des Ioxynils
ist ein überraschendes Ergebnis. Oettmeier et al. (1989) bewies, daß Val 249 an
der Bindung von Ioxynil direkt beteiligt ist. Es wurde erwartet, daß durch den
Austausch von Val 249 gegen Glycin stabilisierende Wechselwirkungen zwischen
der Aminosäure und dem Herbizidmolekül wegfallen und folglich die Bindungsaf-
finität vermindert wird. Das vorliegende Ergebnis deutet darauf hin, daß der ste-
rische Effekt die Bindung von Ioxynil und Bromoxynil begünstigt. Triazine zei-
gen nur leicht veränderte Hemmeigenschaften, so daß der Einbau einer kleinen
Aminosäure an der Position 249 die Bindung von Hemmstoffen dieser Klasse nur
geringfügig beeinflußt. Lediglich die Bindung von Prometon und Simazin ist
leicht erhöht. Die Bindung von Harnstoffen ist in fast allen Fällen positiv
beeinflußt. 9 von 12 getesteten Substanzen zeigen Supersensitivität. Harnstoffe
gehören zu der „Serinfamilie“ (Trebst, 1987). In der Bindenische sind sie zum Ser
264 orientiert. Ihr Carbonylsauerstoffatom ist vermutlich an einer Wasserstoff-
brücke zum peptidischen Amidwasserstoffatom des dem Serin folgenden Phe 265
beteiligt (Trebst, 1987). Der Austausch von Val 249 gegen Glycin modifiziert an-
scheinend die Bindenische der Harnstoffe in der Weise, daß die Bindungsaffinität
steigt. Die Bindungsaffinität der beiden getesteten Triazinone Metribuzin und
Metamitron ist leicht gestiegen. So wie die Harnstoffe gehören Triazinone zu der
„Serinfamilie“. Damit läßt sich die gleiche Tendenz der beiden Substanzklassen
erklären. Die Hemmwirkung der Biscarbamate Phenmedipham und Phenisopham
zeigt keine Änderung (R/S-Wert=1). Erhöhte Bindungsaffinität wurde bei Hydro-

4. Diskussion 89 xychinolin und Cyanoacrylat beobachtet. Wie im Fall der Cyanophenole scheint
auch hier der kleine Raumbedarf des Glycins eine Rolle zu spielen.
Ala 249
Alanin gehört, so wie Valin, zu den unpolaren Aminosäuren mit aliphatischen
Kohlenwasserstoffresten. Alanin ist kleiner und weniger hydrophob als Valin.
Mutante 249/13, bei der Val 249 gegen Alanin ausgetauscht wurde, ist supersensi-
tiv gegenüber Cyanophenolen und zeigt damit das gleiche Verhalten, wie die
Mutante Gly 249. Sie ist gleichzeitig leicht resistent gegenüber Nitrophenolen
(R/S-Wert>1), eine Eigenschaft, die bei der Mutante Gly 249 nicht beobachtet
wurde. Das Hemmverhalten der Triazine ist nur geringfügig beeinflußt. Dies ist
eine weitere Gemeinsamkeit der Mutanten Gly 249 und Ala 249. Die Supersensi-
tivität gegenüber Harnstoffen ist weniger ausgeprägt als bei der Mutante Gly 249.
Nur 4 Substanzen dieser Klasse zeigen eine erhöhte Bindungsaffinität. Einerseits
sind die sterischen Effekte bei dieser Substitution nicht so stark, andererseits sind
hydrophobe Wechselwirkungen zwischen der Seitenkette des Alanins und dem
Hemmstoffmolekül, ähnlich wie beim Wildtyp, möglich. Dies erklärt die Tatsache,
daß die Bindung vieler Harnstoffe unbeeinflußt bleibt. Triazinone und Biscarba-
mate zeigen keine signifikanten Änderungen der Hemmwirkung.
Leu 249
Bei der Mutante 249/16 ist Val 249 gegen Leucin ausgetauscht. Die Seitenkette
von Leucin ist um ein Kohlenstoffatom länger als die von Valin. Die Veränderun-
gen der Bindungsaffinitäten der Hemmstoffe sind also in erster Linie auf einen
sterischen Effekt zurückzuführen. Die Supersensitivität gegenüber Ioxynil und
Bromoxynil ist auch bei dieser Mutante zu verzeichnen, aber sie ist weniger deut-
lich. Das Hemmverhalten der Nitrophenole ist nicht signifikant verändert. Die
Bindung der Triazine ist im Fall von Prometon, Ametryn, Simazin und Atraton
positiv beeinflußt. Die Verengung der Bindenische führt zur besseren Bindung
dieser Substanzen. Die Mutante Leu 249 ist supersensitiv gegenüber fast allen
getesteten Harnstoffen. Die Gegenüberstellung der Mutanten Gly 249, Ala 249
und Leu 249 läßt interessante Aussagen zu. Das Wirkungsmuster der Harnstoff-

4. Diskussion 90 derivate ist in den Mutanten Gly 249 und Leu 249 sehr ähnlich aber abweichend
von dem in der Mutante Ala 249. Die Supersensitivität in den Mutanten Gly 249
und Leu 249 ist deutlicher ausgeprägt als in der Mutante Ala 249. Einerseits
führt die Vergrößerung der Bindenische, wie im Fall von Gly 249, zu einer erhöh-
ten Bindungsaffinität, andererseits aber auch eine Verengung, wenn stärkere
hydrophobe Wechselwirkungen möglich sind, wie im Fall von Leu 249. Der Aus-
tausch von Val 249 gegen Ala bewirkt eine Vergrößerung der Bindenische, aber in
diesem Fall müssen auch die möglichen hydrophoben Wechselwirkungen berück-
sichtigt werden, die bei Glycin wegfallen und bei Leucin stärker sind. So unter-
scheidet sich die Mutante Ala 249 hinsichtlich der Bindung von Harnstoffen am
wenigsten vom Wildtyp. Die Bindungsaffinität von Metribuzin ist in der Mutante
Leu 249 gestiegen. Die hydrophoben Wechselwirkungen scheinen auch hier zum
Tragen zu kommen. Die Hemmwirkung von Hydroxychinolin und Cyanoacrylat
ist ebenfalls begünstigt.
Ile 249
Bei der Mutante 249/8 wurde Valin gegen Isoleucin ausgetauscht. Leucin und Iso-
leucin unterscheiden sich nur durch die Konstitution ihrer Seitenketten. Die Mut-
ante Ile 249 zeigt dementsprechend ein ähnliches Verhalten gegenüber den
Hemmstoffen, wie die Mutante Leu 249. Die Bindungsaffinität der Cyanophenole
ist erhöht. Die Mutante Ile 249 ist resistent gegenüber dem Nitrophenol Dinoseb
(R/S-Wert=4). Die Mutante Leu 249 zeigt ebenfalls Resistenz gegenüber dieser
Substanz (R/S-Wert=2,5). Die Bindung der Triazine ist im Fall von Prometon,
Ametryn, Prometryn und Simazin positiv beeinflußt, ähnliches Wirkungsmuster
wie bei Mutante Leu 249. Die Supersensitivität gegenüber den Harnstoffderivaten
ist bei der Mutante Ile 249 weniger ausgeprägt als bei Leu 249, wobei auch hier
beide Mutanten weitgehende Ähnlichkeiten zeigen. Die Verengung der Bindeni-
sche durch die Substitution von Val 249 durch Isoleucin führt zur besseren Bin-
dung von Cyanoacrylat.

4. Diskussion 91 Arg 249
Das Arginin besitzt eine große basische Seitenkette. Die Mutante 249/3 trägt diese
Aminosäure an der Stelle von Val 249. Diese Mutante zeigt Supersensitivität ge-
genüber den Cyanophenolen Ioxynil und Bromoxynil. Das Bindungsverhalten
von Nitrophenolen ist dagegen nicht beeinflußt. Val 249 liegt in direkter Nach-
barschaft von Ala 250. Eine Reihe von Chlamydomonas reinhardtii-Mutanten mit
Aminosäureaustauschen an der Position Ala 250 ist bekannt (Sopp, 1995; Johan-
ningmeier et al., 2000). Bei einer diesen Mutanten wurde Ala 250 gegen Arginin
ausgetauscht. Diese Mutante zeigt Supersensitivität gegenüber Ioxynil und Bro-
moxynil. Der Einbau einer positiv geladenen Aminosäure sowohl an der Position
Val 249 als auch Ala 250 verstärkt die Bindung der Cyanophenole. Unter den
Triazinen ist eine erhöhte Bindung im Fall von Ametryn, Prometon und Atrazin
festzustellen. Erhöhte Atrazinsensitivität wurde auch bei der oben erwähnten
Mutante Arg 250 beobachtet (Sopp, 1995; Johanningmeier et al., 2000). Die Mut-
ante Arg 249 ist supersensitiv gegenüber einigen der getesteten Harnstoffe. Linu-
ron, Neburon, Cycluron, Chlortoluron und Buturon zeigen R/S-Werte<1. Mögli-
cherweise tritt die große und polare Seitenkette des Arginins mit dem jeweiligen
Hemmstoffmolekül in Wechselwirkung, was einen positiven Einfluß auf die Bin-
dung ausübt. Die Substitution von Val 249 durch Arginin führt zur besseren Bin-
dung von Hydroxychinolin und Cyanoacrylat. Bei dieser Substitution können die
veränderten Hemmeigenschaften der Herbizide sowohl auf sterische Effekte als
auch auf polare Wechselwirkungen zurückzuführen sein.
His 249
Bei der Mutante 249/5 ist Val 249 zu Histidin mutiert, einer Aminosäure mit
schwach basischer Seitenkette. Durch den Einbau von Histidin wird die Bindeni-
sche eingeengt, da seine Seitenkette größer ist als die Seitenkette von Valin. Der
sterische Effekt ist aber nicht so stark wie bei der Mutante Arg 249. Die Cya-
nophenole Ioxynil und Bromoxynil zeigen höhere Bindungsaffinitäten, während
die Nitrophenole in ihrer Bindung nicht signifikant beeinflußt werden. Die Tria-
zine zeigen keine Änderung ihrer Bindungsaffinitäten. Lediglich für Ametryn
wurde eine leichte Supersensitivität verzeichnet (R/S-Wert=0,4). Die Hemmeigen-

4. Diskussion 92 schaften der Harnstoffderivate sind in der Mutante His 249 unterschiedlich. So
zeigen Neburon, DCMU, Buturon, Fluometuron und Linuron erhöhte Hemmwir-
kung (R/S-Werte zwischen 0,16 und 0,5), während Chloroxuron, Metabenzthiazu-
ron und Fenuron leicht verminderte Hemmwirkung aufweisen (R/S-Werte zwi-
schen 2,5 und 2). Die Mutante His 249 ist demnach ein Beispiel dafür, daß sogar
Herbizide der gleichen Substanzklasse etwas unterschiedliche Bindenischen ha-
ben können. Zu den Gemeinsamkeiten der Mutanten His 249 und Arg 249 gehört
die Supersensitivität gegenüber Metribuzin, Cyanoacrylat und Hydroxychinolin.
Ser 249
Bei Mutante 249/17 wurde Val 249 gegen Serin ausgetauscht. Die Seitenkette des
Serins besitzt eine Hydroxylgruppe, die mit einem Hemmstoffmolekül potentielle
Wasserstoffbrückenbindungen ausbilden kann. Die Cyanophenole Ioxynil und
Bromoxynil zeigen erhöhte Bindungsaffinität (R/S-Wert=0,16 bzw. 0,2). Eine
Doppelmutante von Chlamydomonas reinhardtii mit den Aminosäureaustauschen
Ala 250 gegen Serin und Phe 255 gegen Isoleucin zeigt die gleichen R/S-Werte für
die beiden Cyanophenole (Johanningmeier et al., 2000). Der Einbau einer polaren
Aminosäure sowohl an der Position Val 249 als auch Ala 250 verstärkt die Bin-
dung der Cyanophenole. Unter den Nitrophenolen ist bei Dinoterb und DNOC ei-
ne leichte Resistenz zu verzeichnen (R/S-Wert=3,2 bzw. 2,5), Dinoseb und Brom-
nitrothymol binden in der Mutante und im Wildtyp etwa gleich. Auch in der oben
erwähnten Doppelmutante Ser250/Ile255 ist die Dinosebsensitivität nicht
beeinflußt (Johanningmeier et al., 2000). Die Hemmeigenschaften der Triazine
sind im Fall von Simazin, Atrazin, Ametryn und Prometon geringfügig erhöht
(R/S-Wert≈0,5). Bei der Doppelmutante Ser250/Ile255 wurde der gleiche R/S-Wert
für Atrazin ermittelt (Johanningmeier et al., 2000). Das Wirkungsmuster der
Harnstoffderivate in der Mutante Ser 249 weist Ähnlichkeiten mit dem in der
Mutante Arg 249 auf. Es wurde Supersensitivität gegenüber Neburon, Linuron,
Chlortoluron, Cycluron, Buturon und Thiazfluron beobachtet. Die Aminosäuren
Arginin und Serin unterscheiden sich sehr deutlich in der Größe ihrer Seitenket-
ten. Der Einbau von Arginin führt zur Verengung der Bindenische, während der
von Serin keinen solchen Effekt hat. Das ähnliche Wirkungsmuster läßt vermu-

4. Diskussion 93 ten, daß vor allem die polaren Wechselwirkungen bei der Bindung der genannten
Harnstoffe eine Rolle spielen.
Cys 249
Cystein zeigt eine starke Polarität, die durch die Thiolgruppe seiner Seitenkette
bedingt ist. Cystein tritt bei der Mutante 249/28 an der Stelle von Val 249 auf.
Durch diesen Austausch kann eine zusätzliche Wasserstoffbrückenbindung zu
dem Hemmstoffmolekül gebildet werden. Die Bindungsaffinität der Cyanophenole
ist gestiegen, während die Bindung der Nitrophenole keine Änderungen zeigt. Der
Einbau von Cystein beeinflußt positiv die Bindung von Atraton, Simazin, A-
metryn und Prometon, die zu den Triazinen gehören. Harnstoffderivate zeigen
nur leicht verändertes Hemmverhalten. Nur 4 Vertreter dieser Gruppe zeigen ge-
ringfügig erhöhte Bindungsaffinität: Neburon, Benzthiazuron, Fluometuron und
Buturon (R/S-Werte zwischen 0,25 und 0,5). Die Mutante Cys 249 ist supersensitiv
gegenüber Cyanoacrylat.
Asn 249
Asparagin gehört zu den Aminosäuren mit ungeladenen polaren Resten. Mutante
249/19 weist den Austausch von Val 249 gegen Asparagin auf. Die veränderten
Hemmeigenschaften, die bei dieser Mutante beobachtet werden, hängen mit der
Fähigkeit des Asparagins zusammen, Wasserstoffbrückenbindungen mit dem
Hemmstoffmolekül einzugehen. Die getesteten Cyanophenole Ioxynil und Bromo-
xynil zeigen eine erhöhte Bindungsaffinität (R/S-Wert=0,25 bzw. 0,32). Unter den
Nitrophenolen zeigen Bromnitrothymol und DNOC leicht verminderte Bindungs-
affinität (R/S-Wert=2,5). Die Amidgruppe des Asparagins scheint die Bindung die-
ser beiden Nitrophenole zu stören. Die Mutante Asn 249 ist supersensitiv gegen-
über einigen der Triazine, nämlich gegenüber Simazin, Ametryn und Prometon.
Auch die Bindung der Harnstoffe ist in vielen Fällen positiv beeinflußt. So ist die
Sensitivität gegenüber DCMU, Neburon, Linuron, Buturon, Fluometuron und
Thiazfluron gestiegen. Eine erhöhte Bindungsaffinität ist auch bei Hydroxychino-
lin und Cyanoacrylat zu vermerken.
Gln 249

4. Diskussion 94 Bei der Mutante 249/24 ist Val 249 zu Glutamin mutiert. Glutamin unterscheidet
sich von Asparagin nur durch eine Verlängerung der Seitenkette um ein Kohlen-
stoffatom. Die Unterschiede zwischen den Mutanten Asn 249 und Gln 249 kom-
men also durch einen sterischen Effekt zustande. Glutamin ist in der Lage, so wie
Asparagin, mit dem Hemmstoffmolekül Wasserstoffbrückenbindungen zu bilden.
So wie bei allen anderen Mutanten wurde auch bei der Mutante Gln 249 eine er-
höhte Sensitivität gegenüber Ioxynil und Bromoxynil beobachtet. Als eine ge-
meinsame Eigenschaft der Mutanten Gln 249 und Asn 249 ist die Resistenz ge-
genüber den Nitrophenolen DNOC und Bromnitrothymol zu verzeichnen (R/S-
Wert=6,3 bzw. 2,5). Überraschenderweise ist die Bindungsaffinität der beiden an-
deren Nitrophenole Dinoterb und Dinoseb in der Mutante Gln 249 gestiegen (R/S-
Wert=0,4). In der Mutante Asn 249 zeigen diese Nitrophenole keine signifikante
Änderung in der Hemmwirkung. Dieser Unterschied ist auf die Verlängerung der
Seitenkette des Glutamins zurückzuführen. Die Bindung der Triazine ist nicht
beeinflußt. Lediglich Prometon zeigt eine leicht erhöhte Bindungsaffinität (R/S-
Wert=0,5). Auch die Bindung der Harnstoffe ist weniger beeinflußt als bei der
Mutante Asn 249. Supersensitivität wurde gegenüber Neburon, Buturon und Flu-
ometuron beobachtet, nur gegenüber 3 der 12 getesteten Substanzen. Die Mutan-
te Asn 249 ist supersensitiv gegenüber 6 der 12 Harnstoffe. Die Bindung des Tria-
zinons Metribuzin ist durch die längere Seitenkette des Glutamins positiv
beeinflußt (R/S-Wert=0,5). Hydroxychinolin und Cyanoacrylat zeigen ein ähnli-
ches Wirkungsmuster wie in der Mutante Asn 249, wobei die Supersensitivität in
der Mutante Gln 249 weniger ausgeprägt ist.
4.2. Charakterisierung der Doppelmutanten mit Aminosäureaustau-schen an den Positionen Phe 211 und Met 214 im D1-Protein
Die Aminosäuren Phe 211 und Met 214 befinden sich nach den Modellen des Re-
aktionszentrums von Photosystem II in der transmembranen Helix IV und sind
Bestandteil der QB-Bindenische. Sie liegen in der Nähe bzw. in der unmittelbaren
Nachbarschaft von His 215, das an der Bindung des Fe-Ions beteiligt ist und folg-
lich die dreidimensionale Faltung des D1-Proteins stabilisiert. Die Mutation des
Codons 211 war bereits in Cyanobakterien bekannt, bei denen die Substitution

4. Diskussion 95 von Phe 211 gegen Serin zur Resistenz gegenüber Atrazin, Diuron und Metribu-
zin führt (Gingrich et al., 1988; Kirilovsky et al., 1989; Govindjee et al., 1990). Met
214 wurde durch Photoaffinitätsmarkierung von Spinatthylakoiden als Bindung-
sort von Azido-[14C]atrazin erkannt (Wolber et al., 1986; Whitelegge et al., 1992).
Bereits in der vorhergehenden Arbeit (Orawski, 1998) wurde ortsgerichtete Muta-
genese durchgeführt, bei der Aminosäureaustausche an den Positionen Phe 211
und Met 214 im D1-Protein herbeigeführt wurden. Mit der in vitro manipulierten
DNA wurden 35 Transformanten erzeugt, von denen 30 im Rahmen dieser Disser-
tation charakterisiert wurden.
Die PCR-Analyse zeigte, daß alle Transformanten homoplasmisch sind. Mittels
DNA-Sequenzierung wurden 11 neue Mutanten identifiziert: Ala211/Asn214,
Thr211/Thr214/Ala219, Gly211/Ser214/Ala219, Thr211/Ala214, Met211/Thr214,
Gln211, Leu211/Gly214, Gly211/Thr214, Gly211/Ala214, Gln211/Gly214/Ala219
und Cys211/Thr214/Ala219. Die Mutanten Thr211/Thr214/Ala219 (Mut. 3),
Gly211/Ala214 (Mut. 27), Gln211/Gly214/Ala219 (Mut. 28), Cys211/Thr214/
Ala219 (Mut. 32) und Gly211/Ser214/Ala219 (Mut. 34) zeigen ein vermindertes
Wachstum im Vergleich zum Wildtyp und zu der intronlosen Mutante. Die einge-
führten Mutationen haben einen negativen Einfluß auf den photosynthetischen
Elektronentransport. Sie vermindern anscheinend die Bindung des Plastochinons
in der QB-Bindenische.
4.2.1. Hemmstoffexperimente
Für die Charakterisierung der erzeugten Doppelmutanten wurden folgende
Hemmstoffe verwendet: Atrazin, DCMU, Metribuzin, Metamitron, Ioxynil, Phen-
medipham, Bromnitrothymol, Cyanoacrylat, Hydroxychinolin und Ketonitril.
Atrazin
Ein verändertes Bindungsverhalten des Atrazins wurde erwartet. Einerseits ruft
die Mutation von Phe 211 zu Serin in Cyanobakterien Atrazinresistenz hervor.
Andererseits deuten Photoaffinitätsmarkierungsversuche darauf hin, daß Met 214
an der Bindung des Atrazins beteiligt ist. Die höchste Atrazinresistenz wurde bei

4. Diskussion 96 der Mutante Gln 211 ermittelt (R/S-Wert=25). Im Gegensatz zu den anderen ist
diese Mutante eine Einfachmutante. Der Austausch von Phenylalanin, einer un-
polaren Aminosäure mit aromatischem Ring, gegen Glutamin mit einer A-
midgruppe hat einen negativen Einfluß auf die Bindung des Atrazins. Die Mutan-
te Gln211/Gly214/Ala219 trägt an der Position 211 ebenfalls ein Glutamin, zeigt
aber nur geringe Atrazinresistenz (R/S-Wert=2). Anscheinend kompensieren die
zusätzlichen Mutationen teilweise den negativen Einfluß des Glutamins. Ent-
scheidend scheint dabei das Vorhandensein einer kleinen unpolaren Aminosäure
an der Position 214 zu sein. Die Bindungsaffinität des Atrazins ist bei Mutanten
mit Glycin oder Alanin an der Position 214 am wenigsten beeinflußt.
Triazinone
Wie bereits erwähnt führt der Austausch von Phe 211 gegen Serin in Synecho-
cystis sp. PCC 6714 zur Metribuzinresistenz (Kirilovsky et al., 1989; Govindjee et
al., 1990). Unter den neu erzeugten Chlamydomonas reinhardtii-Mutanten weisen
die Mutanten Met211/Thr214 und Thr211/Thr214/Ala219 eine verminderte
Metribuzinsensitivität auf (R/S-Wert=12,6 bzw. 6,3). Das zu der gleichen Hemm-
stoffklasse gehörende Metamitron zeigt leicht verminderte Bindungsaffinität in
den Mutanten Thr211/Ala214, Thr211/Thr214/Ala219, Gln211/Gly214/Ala219
und Gly211/Ser214/Ala219 (R/S-Werte zwischen 5 und 3). Erwartungsgemäß sind
die meisten Folgen der Mutationen in den Dreifachmutanten zu verzeichnen. A-
minosäuresubstitutionen, die 3 Positionen betreffen, führen offensichtlich zu weit-
gehenden Veränderungen in der Struktur der Bindenische.
Phenmedipham
Die Hemmeigenschaften des Biscarbamats Phenmedipham sind am stärksten
verändert. Alle Mutanten zeigen gesteigerte Resistenz gegenüber diesem Herbi-
zid. Die höchsten R/S-Werte zeigen die Dreifachmutanten Gly211/Ser214/Ala219
(R/S-Wert=32), Gln211/Gly214/Ala219 (R/S-Wert=25), Cys211/Thr214/Ala219
(R/S-Wert=13) und Thr211/Thr214/Ala219 (R/S-Wert=13). Unter den Doppelmut-
anten zeigt die Mutante Thr211/Ala214 (R/S-Wert=16), Gly211/Thr214 (R/S-
Wert=10) und Gly211/Ala214 (R/S-Wert=10) ebenfalls Resistenz. Sowohl der Ein-

4. Diskussion 97 bau einer polaren Aminosäure (Threonin, Cystein, Glutamin) als auch der Einbau
der kleinsten Aminosäure (Glycin) an der Position 211 führt zur Erhöhung der
Resistenz.
Cyanoacrylat
Cyanoacrylat zeigt in allen Mutanten verminderte Bindungsaffinität. Offensicht-
lich wird die Bindung des Cyanoacrylats im Wildtyp durch die Wechselwirkungen
des aromatischen Restes im Cyanoacrylatmolekül mit der aromatischen Seiten-
kette des Phenylalanins an der Position 211 stabilisiert. In den Mutanten fallen
diese Wechselwirkungen weg.
Hydroxychinolin
Die Hemmeigenschaften des Hydroxychinolins sind in den einzelnen Mutanten
unterschiedlich. Mutante Gly211/Ala214 zeigt eine leichte Resistenz (R/S-
Wert=4). Diese Mutante trägt sowohl an der Stelle von Phe 211 als auch an der
Stelle von Met 214 sehr kleine Aminosäuren. Die Vergrößerung der Bindenische
kann die Bindung des Herbizidmoleküls abschwächen. In einigen Fällen wurde
Supersensitivität gegenüber Hydroxychinolin beobachtet. Die Mutanten
Ala211/Asn214, Thr211/Thr214/Ala219, Thr211/Ala214, Gln211/Gly214/Ala219
und Gly211/Ser214/Ala219 zeigen eine erhöhte Bindungsaffinität. Offensichtlich
sind zusätzliche Interaktionen zwischen dem Hemmstoff und den eingeführten
Aminosäuren möglich.
Die Hemmeigenschaften von Diuron, Ioxynil, Bromnitrothymol und Ketonitril
sind nur geringfügig verändert. Die Eingeführten Aminosäuresubstitutionen ha-
ben keinen signifikanten Einfluß auf die Bindung dieser Hemmstoffe.
4.3. Mutationen der Aminosäure His 215 im D1-Protein
Im Rahmen dieser Arbeit wurde mit Hilfe der ortsgerichteten Mutagenese das Co-
don für His 215 im D1-Protein von Chlamydomonas reinhardtii mutiert. Es wur-
den 4 verschiedene mutagene Oligonukleotide eingesetzt, darunter ein gewobbel-
tes und 3 mit festgelegten Sequenzen. Durch den Einsatz des gewobbelten Oligo-

4. Diskussion 98 nukleotids waren 17 verschiedene Aminosäureaustausche an der Position His 215
denkbar. Durch den Einsatz der 3 anderen Oligonukleotide war der Austausch
von His 215 gegen Glutamin, Isoleucin und Phenylalanin möglich. Mittels DNA-
Sequenzierung wurde festgestellt, daß die Mutagenese mit allen 4 Primern erfolg-
reich war. Die Mutageneseprodukte wurden zur Transformation der Deletionsmu-
tante Del1 von Chlamydomonas reinhardtii eingesetzt. Nach Sequenzierung des
psbA-Gens der gewonnenen Transformanten stellte sich heraus, daß alle 31 Trans-
formanten Wildtypsequenz aufweisen. Es konnten keine photosynthetisch aktiven
Mutanten mit Aminosäuresubstitutionen an der Position His 215 erzeugt werden.
Die im Rahmen der vorliegenden Arbeit angewendete Methode ermöglicht nicht
die Identifizierung der Mutanten, bei denen die Aminosäureaustausche zur Inak-
tivierung der Photosynthese führen.
Aufgrund der Homologie des D1- und D2-Proteins mit der L- und M-Untereinheit
des bakteriellen Reaktionszentrums wurden detaillierte Modelle für das Photosys-
tem II aufgestellt (Trebst, 1987; Ruffle et al., 1992; Xiong et al., 1996; Xiong et al.,
1998). Nach diesen Modellen koordinieren 4 Histidinreste das Fe-Atom, nämlich
His 215 und His 272 im D1-Protein sowie His 215 und His 269 im D2-Protein (in
Cyanobakterien His 214 und His 268 im D2-Protein). Der Histidinrest an der Posi-
tion 215 im D1- bzw. im D2-Protein fungiert außerdem als Ligand für QB bzw. QA.
Der Mißerfolg bei der Konstruktion von photosynthetisch aktiven Chlamydomo-
nas reinhardtii-Mutanten mit Aminosäuresubstitutionen von His 215 im D1-
Protein läßt vermuten, daß das His 215 eine essentielle Rolle für die Struktur und
Funktion des Photosystems II spielt. In dem Cyanobakterium Synechocystis sp.
6803 wurde durch ortsgerichtete Mutagenese der Austausch von His 214 gegen
Asparagin im D2-Protein eingeführt (Vermaas et al., 1987). Diese Mutation führte
zur Inaktivierung des Photosystems II. Auch die Mutation von His 268 zu Gluta-
min im D2-Protein von Synechocystis sp. 6803 hat zur Folge den Verlust des pho-
toautotrophen Wachstums (Vermaas et al., 1994). Sie führt zu veränderten Eigen-
schaften sowohl der QA- als auch der QB- und Herbizid-Bindenische. Vermaas et al.
(1994) stellte eine Hypothese auf, wonach die Mutation von His 268 zu Glutamin
im D2-Protein zum funktionellen Verlust des Fe-Ions im PS II führt. Das Fe-Atom
scheint für den Elektronentransport im PS II wichtig zu sein, wenn auch seine

4. Diskussion 99 Rolle im Elektronentransport noch nicht bekannt ist. Es ist denkbar, daß der Aus-
tausch von His 215 im D1-Protein die gleichen oder vergleichbaren Effekte hat,
wie die Austausche von His 214 und His 268 im D2-Protein. Dies würde das Aus-
bleiben der photoautotrophen Mutanten mit Aminosäuresubstitutionen an der
Position His 215 im D1-Protein erklären.
Das Auftreten von Wildtypkolonien nach der Transformation der Deletionsmutan-
te von Chlamydomonas reinhardtii kann zwei Ursachen haben. Einerseits ist ein
Fehler der Taq-Polymerase denkbar. Andererseits kann eine Kontamination der
Proben mit Wildtyp-DNA während der Transformation nicht ausgeschlossen wer-
den. Die Taq-Polymerase wurde zum Amplifizieren der Mutageneseprodukte ver-
wendet. Bei der Sequenzierung der amplifizierten Mutageneseprodukte können
Fehler, die in fortgeschrittenen Zyklen der PCR auftreten, nicht erfaßt werden.
Erst die Sequenzierung der Transformanten-DNA ermöglicht die endgültige Veri-
fizierung der Sequenz.
4.4. Charakterisierung der Mutanten mit Aminosäureaustauschen an der Position Phe 206 im D1-Protein
Die Aminosäureaustausche im D1-Protein, die das Bindungsverhalten der Hemm-
stoffe beeinflussen, konzentrieren sich auf dem Bereich zwischen den Aminosäu-
ren Phe 211 und Leu 275. Eine Ausnahme stellt die Mutante von Synechocystis
sp. PCC 6803 dar, bei der Phe 295 im D1-Protein gegen Serin ausgetauscht ist
(Narusaka et al., 1998). Dieser Aminosäureaustausch verursacht eine leicht er-
höhte Atrazinresistenz (R/S-Wert=2). Phe 295 befindet sich nach den Modellen für
das D1-Protein am Ende der transmembranen Helix V an der Lumenseite und ist
nicht direkt an der Bildung der QB-Bindenische beteiligt. Möglicherweise führt der
Aminosäureaustausch an dieser Position zu einer Konformationsänderung des
D1-Proteins, so daß die Bindung eines Hemmstoffmoleküls beeinflußt wird (Naru-
saka et al., 1998).
Im Rahmen der vorliegenden Arbeit wurde die Aminosäure Phe 206 im D1-
Protein von Chlamydomonas reinhardtii substituiert. Es wurden 8 verschiedene
Aminosäureaustausche eingeführt, nämlich gegen Ala, Asn, Glu, His, Ile, Lys, Ser

4. Diskussion 100 und Val. Phe 206 liegt in der transmembranen Helix IV außerhalb des Bereiches,
der als Herbizid-Bindenische definiert wird. Der Austausch von Phe 206 ist bereits
in einer Dreifachmutante von Chlamydomonas reinhardtii bekannt
(Tyr206/Ala211/Thr214; Mutante 12 aus der Diplomarbeit; Orawski, 1998), die ein
verändertes Verhalten gegenüber Hemmstoffen zeigt. In den erzeugten Mutanten
wurde das Bindungsverhalten von einem Triazin, einem Harnstoffderivat und
einem Biscarbamat untersucht. Das getestete Triazin Atrazin zeigt in allen Mut-
anten außer der Mutante His 206 eine leicht erhöhte Bindungsaffinität. Die Bin-
dung des Biscarbamats Phenmedipham ist ebenfalls in den meisten Mutanten
positiv beeinflußt. Die Hemmeigenschaften des Harnstoffs DCMU sind nicht ver-
ändert. Diese Ergebnisse deuten darauf hin, daß Phe 206 einen Einfluß auf die
Struktur und Funktion der Herbizid-Bindenische haben kann. Unklar bleibt, ob
Phe 206 direkt mit einem Hemmstoffmolekül interagieren kann, oder ob es indi-
rekt die Eigenschaften der Bindenische beeinflussen kann. Um nähere Aussagen
über die Bedeutung dieser Aminosäure für die Struktur der QB- und Herbizid-
Bindenische machen zu können, sind weitere experimentelle Daten notwendig.

5. Zusammenfassung 101
5. Zusammenfassung
Die QB-Bindenische ist der spezifische Angriffsort vieler Hemmstoffe der Photo-
synthese. Die Wirkung dieser Substanzen beruht auf der kompetitiven Verdrän-
gung des sekundären Plastochinons QB aus seiner Bindenische. Hemmstoffe wer-
den einerseits kommerziell als Herbizide eingesetzt, andererseits tragen sie zum
Verständnis der Struktur und Funktion des Reaktionszentrums von PS II bei.
Im Rahmen dieser Dissertation wurde die Mutantenanalyse für die Charakterisie-
rung der QB-Bindenische im Photosystem II von Chlamydomonas reinhardtii an-
gewandt. Die Punktmutationen in dem für das D1-Protein codierenden psbA-Gen
wurden mit Hilfe der Oligonukleotid-gerichteten Mutagenese erzeugt. Die Codons
für folgende Aminosäuren wurden mutagenisiert: Phe 206, His 215 und Val 249.
Es wurden 10 verschiedene Aminosäureaustausche an der Position Val 249 er-
zielt: Ala, Arg, Asn, Cys, Gln, Gly, His, Ile, Leu und Ser. Der Einfluß der Punkt-
mutationen auf die Bindung von Herbizidmolekülen wurde für verschiedene
Hemmstoffe untersucht. Die Cyanophenole Ioxynil und Bromoxynil weisen in al-
len Mutanten erhöhte Bindungsaffinität auf. Im Gegensatz dazu zeigen Nitro-
phenole kein einheitliches Wirkungsmuster. Eine leichte Supersensitivität wurde
nur bei der Mutante Gln 249 gegenüber Dinoterb und Dinoseb sowie bei der
Mutante His 249 gegenüber Dinoterb beobachtet. Der Austausch von Val 249 ge-
gen Aminosäuren mit polaren ungeladenen Seitenketten (Asparagin, Glutamin,
Serin) führt zur leichten Resistenz gegenüber Bromnitrothymol und DNOC. Eine
verminderte Bindungsaffinität einiger Nitrophenole wurde auch bei Mutanten
mit hydrophoben Aminosäuren an der Position Val 249 beobachtet. So führt der
Austausch gegen das kleine Alanin zur Resistenz gegenüber Dinoterb, Dinoseb
und DNOC. Der Austausch gegen größere unpolare Aminosäuren (Leucin, Isoleu-
cin) führt zur Resistenz gegenüber Dinoseb. Triazine zeigen nur leicht veränderte
Hemmeigenschaften. Die Verengung der Bindenische durch den Einbau von gro-
ßen unpolaren Aminosäuren (Leucin, Isoleucin) führt zur besseren Bindung von
Prometon, Ametryn und Simazin. Eine ähnliche Auswirkung hat die Mutation
von Val 249 zu Arginin, einer Aminosäure mit einer großen basischen Seitenket-

5. Zusammenfassung 102 te. Die Supersensitivität gegenüber Prometon und Ametryn bei der Mutante Arg
249 ist anscheinend ebenfalls auf sterische Effekte zurückzuführen. Das Bin-
dungsverhalten der Harnstoffderivate ist in der Mutante Gly 249 am stärksten
beeinflußt. 9 von den 12 getesteten Harnstoffen zeigen erhöhte Bindungsaffinität.
Interessanterweise zeigen die Mutanten Leu 249 und Ile 249 ähnliches Verhalten
gegenüber dieser Klasse von Herbiziden, wie die Mutante Gly 249, obwohl der
Einbau von Leucin bzw. Isoleucin zur Verengung der Bindenische führt. Hier
kommen aber die hydrophoben Wechselwirkungen zum Tragen, die in der Mutan-
te Gly 249 wegfallen und im Wildtyp schwächer sind.
Im Rahmen dieser Arbeit wurden 11 Mutanten mit Aminosäureaustauschen an
den Positionen Phe 211 und Met 214 identifiziert und im Hinblick auf die Bin-
dung von Herbiziden untersucht. Am stärksten sind die Hemmeigenschaften des
Biscarbamats Phenmedipham verändert. Alle Mutanten zeigen gesteigerte Resis-
tenz. Die höchsten Resistenzwerte zeigen Mutanten, bei denen an der Position Val
219 eine zusätzliche Mutation zu Alanin auftritt: Gly211/Ser214/Ala219,
Gln211/Gly214/Ala219, Cys211/Thr214/Ala219 und Thr211/Thr214/Ala219. So-
wohl der Einbau einer polaren Aminosäure (Threonin, Cystein, Glutamin) als
auch der Einbau der kleinsten Aminosäure (Glycin) an der Position 211 führt zur
Erhöhung der Resistenz. Die Bindungsaffinität des Cyanoacrylats ist in allen
Mutanten vermindert. Offensichtlich wird die Bindung des Cyanoacrylats im
Wildtyp durch die Wechselwirkungen des aromatischen Restes im Cyanoacrylat-
molekül mit der aromatischen Seitenkette des Phenylalanins an der Position 211
stabilisiert.
Der Austausch von His 215 blieb erfolglos. Die Transformation des Del1-Stammes
mit der in vitro manipulierten DNA ergab keine photoautotrophen Mutanten. Dies
läßt vermuten, daß His 215 eine essentielle Rolle für die Struktur und Funktion
des D1-Proteins spielt.
Es wurden 8 Mutanten mit Aminosäuresubstitutionen an der Position Phe 206
erzeugt. Folgende Austausche wurden erzielt: Ala, Asn, Glu, His, Ile, Lys, Ser und
Val. Die Mutagenese von Phe 206 führt zu einem veränderten Bindungsverhalten

5. Zusammenfassung 103 von Atrazin und Phenmedipham, während das Bindungsverhalten von DCMU
keine Änderungen zeigt. Diese Ergebnisse deuten darauf hin, daß Phe 206, das
außerhalb des als Herbizid-Bindenische definierten Bereiches liegt, zumindest
einen indirekten Einfluß auf die Struktur und Funktion der Bindenische im D1-
Protein haben kann.

6. Anhang 104
6. Anhang
6.1. Strukturformeln der Hemmstoffe
6.1.1. Phenole
OH
JJ
CN
Ioxynil: 4-Hydroxy-3,5-dijodobenzonitril
OH
CN
BrBr
Bromoxynil: 4-Hydroxy-3,5-dibromobenzonitril
OH
Br
CH3
NO2
(CH3)2CH
Bromnitrothymol: 2-Bromo-4-nitrothymol
OH
NO2 C(CH )3 3
NO2 Dinoterb: 2-Tertiärbutyl-4,6 dinitrophenol

6. Anhang 105
2NO
2NO
OH
CHCH3
C H2 5
Dinoseb: 2,4-Dinitro-6-isobutylphenol
2NO
32NO
OH
CH
DNOC: 4,6-Dinitro-o-cresol
6.1.2. Triazine
N
N
N
Cl
NHCH(CH3)2C2H5NH
Atrazin: 2-Chlor-4-ethylamino-6-isopropylamino-1,3,5-triazin
N
N
N
OCH3
NH CH(CH )3 2NHC H2 5
Atraton: 2-Amino-4-ethylamino-6-methoxy-1,3,5-triazin
52C H NH NH
N
N
N
C H2 5
Cl
Simazin: 2,4-Bis(ethylamino)-6-chlor-1,3,5-triazin

6. Anhang 106
2 NH 23CH(CH )NH
3
N
N
N
SCH
CH(CH )3 Prometryn: 2,4-Bis(isopropylamino)-6-methylthio-1,3,5-triazin
52C H NH NH
3
N
N
N
SCH
C(CH )3 3 Terbutryn: 2-(1,1-Dimethylethylamino)-4-ethylamino-6-methylthio-1,3,5-triazin
52C H NH 23CH(CH )NH
3
N
N
N
SCH
Ametryn: 2-Ethylamino-4-isopropylamino-6-methylthio-1,3,5-triazin
3(CH ) CH
N
N
N
3
NH CH(CH )3 2NH2
OCH
Prometon: 2,4-Bis(isopropylamino)-6-methoxy-1,3,5-triazin
6.1.3. Harnstoffe
CH3NNCl
Cl
H CH3
O
DCMU=Diuron: 3-(3,4-Dichlorphenyl)-1,1-dimethylharnstoff
N
H
C
O
NCH
CH
3
3
Cycluron: 3-Cyclooctyl-1,1-dimethylharnstoff

6. Anhang 107
O
NNCl
Cl
H CH3
O CH3
Linuron: 3-(3,4-Dichlorphenyl)-1-methoxy-1-methylharnstoff
OCl NH CO N(CH )23
Chloroxuron: 3-(4-(4-chlorophenoxy)phenyl)-1,1-dimethylharnstoff
Cl
Cl NH CO NCH3
C H4 9 Neburon: 1-Butyl-3-(3,4-dichlorophenyl)-1-methylharnstoff
Cl
NH CO N(CH )23CH3
Chlortoluron: 3-(3-Chloro-4-methylphenyl)-1,1-dimethylharnstoff
N
S
3CHCONH NH
Benzthiazuron: 1-(Benzothiazol-2-yl)-3-methylharnstoff
N
S3CHCO NHN
CH 3 Metabenzthiazuron: 1-(Benzothiazol-2-yl)-1,3-dimethylharnstoff
NH CO N(CH )23
CF3 Fluometuron: 3-(3-(Trifluoromethyl)phenyl)-1,1-dimethylharnstoff
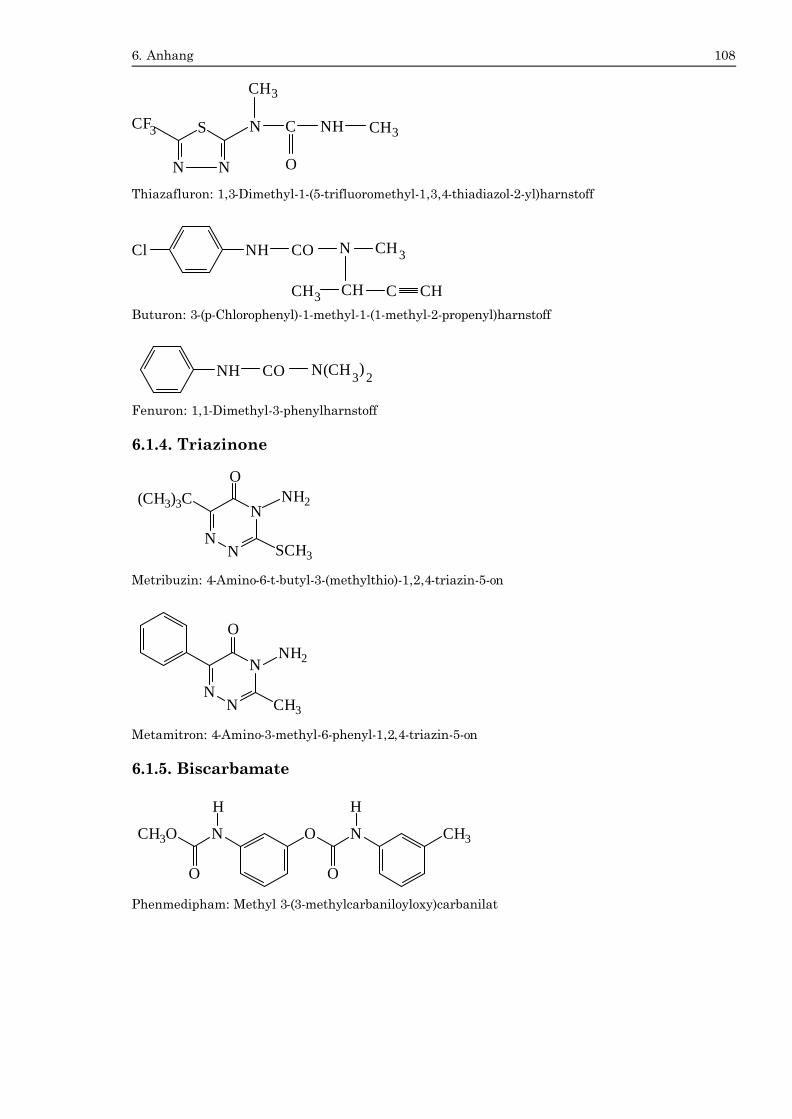
6. Anhang 108
N N
SCF3 N C
O
NH CH3
3CH
Thiazafluron: 1,3-Dimethyl-1-(5-trifluoromethyl-1,3,4-thiadiazol-2-yl)harnstoff
Cl NH CO N CH3
CHCH3 C CH
Buturon: 3-(p-Chlorophenyl)-1-methyl-1-(1-methyl-2-propenyl)harnstoff
NH CO N(CH )23
Fenuron: 1,1-Dimethyl-3-phenylharnstoff
6.1.4. Triazinone
NH2
NN
N
SCH3
(CH3)3CO
Metribuzin: 4-Amino-6-t-butyl-3-(methylthio)-1,2,4-triazin-5-on
NH2
O
N
N
CH3N
Metamitron: 4-Amino-3-methyl-6-phenyl-1,2,4-triazin-5-on
6.1.5. Biscarbamate
NONCH3O
OO
HH
CH3
Phenmedipham: Methyl 3-(3-methylcarbaniloyloxy)carbanilat

6. Anhang 109
NON
OO
H
OCH(CH 3)2
C H2 5
Phenisopham: Isopropyl 3-(ethyl(phenyl)carbamoyloxy)carbanilat
6.1.6. Sonstige Hemmstoffe
N
Br
CF3
CF3
OH
Hydroxychinolin: 3-Brom-2,6-di-trifluoromethyl-4-hydroxy-chinolin
N
S
CN
OH
Ketonitril: 2-Phenylthiazol-3-hydroxy-4-phenyl-butenonitril
COOCH2CH2OCH2CH3
CN
NH
CH3CH2
CH2Cl
Cyanoacrylat: 2-Cyano-3-ethyl-3-(4-chlor-benzyl)-amino-acrylat-ethoxy- Ethylester
O
NO
NO
2
2
I 2NO
CH3
CH 3CH( )2 DNP-INT: Dinitrophenylether von Nitro-iod-thymol

6. Anhang 110 6.2. Sequenz des psbA-Gens aus Chlamydomonas reinhardtii und flankierende Bereiche
Die in der Arbeit verwendeten Primer erscheinen in Fettdruck.
1424 bp; 381 A; 311 C; 268 G; 464 T. 10 20 30 40 50 5' CGTCCTATTT TAATACTCCG AAGGAGGCAG TTGGCAGGCA ACTGCCACTG 3' GCAGGATAAA ATTATGAGGC TTCCTCCGTC AACCGTCCGT TGACGGTGAC 60 70 80 90 100 ACGTCCCGTA AGGGTAAGGG GACGTCCACT GGCGTCCCGT AAGGGGAAGG TGCAGGGCAT TCCCATTCCC CTGCAGGTGA CCGCAGGGCA TTCCCCTTCC 110 120 130 140 150 GGACGTAGGT ACATAAATGT GCTAGGTAAC TAACGTTTGA TTTTTTGTGG CCTGCATCCA TGTATTTACA CGATCCATTG ATTGCAAACT AAAAAACACC 160 170 180 190 200 TATAATATAT GTACCATGCT TTTAATAGAA GCTTGAATTT ATAAATTAAA ATATTATATA CATGGTACGA AAATTATCTT CGAACTTAAA TATTTAATTT 210 220 230 opsbA-28→→ 250 ATATTTTTAC AATATTTTAC GGAGAAATTA AAACTTTAAA AAAATTAACA TATAAAAATG TTATAAAATG CCTCTTTAAT TTTGAAATTT TTTTAATTGT Start 260 270 280 290 300 TATGACAGCA ATTTTAGAAC GTCGTGAAAA TTCTAGCCTA TGGGCTCGTT ATACTGTCGT TAAAATCTTG CAGCACTTTT AAGATCGGAT ACCCGAGCAA 310 320 330 340 350 TTTGTGAGTG GATCACTTCA ACTGAAAACC GTTTATACAT CGGTTGGTTC AAACACTCAC CTAGTGAAGT TGACTTTTGG CAAATATGTA GCCAACCAAG 360 370 380 390 400 GGTGTAATCA TGATCCCATG TCTTCTTACT GCAACATCAG TATTCATCAT CCACATTAGT ACTAGGGTAC AGAAGAATGA CGTTGTAGTC ATAAGTAGTA 410 420 430 440 450 CGCTTTCATC GCTGCTCCGC CAGTAGACAT CGATGGTATC CGTGAACCAG GCGAAAGTAG CGACGAGGCG GTCATCTGTA GCTACCATAG GCACTTGGTC 460 470 480 490 500 TTTCAGGTTC TCTTCTTTAC GGTAACAACA TCATTACAGG TGCTGTAATC AAAGTCCAAG AGAAGAAATG CCATTGTTGT AGTAATGTCC ACGACATTAG 510 520 530 540 550 CCAACTTCTA ACGCAATCGG TCTTCACTTC TACCCAATTT GGGAAGCTGC GGTTGAAGAT TGCGTTAGCC AGAAGTGAAG ATGGGTTAAA CCCTTCGACG 560 570 580 590 600 TTCTCTAGAC GAGTGGTTAT ACAACGGTGG TCCTTACCAA CTTATCGTTT AAGAGATCTG CTCACCAATA TGTTGCCACC AGGAATGGTT GAATAGCAAA 610 620 630 640 650 GTCACTTCCT TCTAGGTGTA TACTGCTACA TGGGTCGTGA GTGGGAATTA CAGTGAAGGA AGATCCACAT ATGACGATGT ACCCAGCACT CACCCTTAAT 660 opsbA412→→ 680 690 700 TCTTTCCGTT TAGGTATGCG TCCATGGATC GCTGTAGCTT ACTCAGCTCC AGAAAGGCAA ATCCATACGC AGGTACCTAG CGACATCGAA TGAGTCGAGG

6. Anhang 111 710 720 730 740 750 AGTAGCTGCA GCTTCAGCTG TATTCTTAGT TTACCCTATC GGCCAAGGTT TCATCGACGT CGAAGTCGAC ATAAGAATCA AATGGGATAG CCGGTTCCAA 760 770 780 790 800 CATTCTCTGA CGGTATGCCT TTAGGTATCT CTGGTACTTT CAACTTCATG GTAAGAGACT GCCATACGGA AATCCATAGA GACCATGAAA GTTGAAGTAC 810 820 830 840 850 ATCGTATTCC AAGCAGAACA CAACATCCTT ATGCACCCAT TCCACATGTT TAGCATAAGG TTCGTCTTGT GTTGTAGGAA TACGTGGGTA AGGTGTACAA 860 870 880 890 900 AGGTGTTGCT GGTGTATTCG GTGGTTCATT ATTCTCAGCT ATGCACGGTT TCCACAACGA CCACATNNNC CACCAAGTAA TAAGAGTCGA TACNNNCCAA 910 ←←opsbA206 930 ←←opsbA215 CTTTAGTTAC TTCATCTTTA ATCCGTGAAA CAACTGAAAA CGAATCAGCT GAAATCAATG AAGTAGAAAT TAGGCACTTT GTTGACTTTT GCTTAGTCGA 960 970 980 990 1000 AACGAAGGTT ACCGTTTCGG TCAAGAAGAA GAAACTTACA ACATTGTAGC TTGCTTCCAA TGGCAAAGCC AGTTCTTCTT CTTTGAATGT TGTAANNNCG 1010 1020 1030 1040 ←←opsbA249 TGCTCATGGT TACTTTGGTC GTCTAATCTT CCAATACGCT TCTTTCAACA ACGAGTACCA ATGAAACCAG CAGATTAGAA GGTTATGCGA AGAAAGTTGT 1060 1070 1080 1090 1100 ACTCTCGTTC ATTACACTTC TTCTTAGCTG CTTGGCCGGT AATCGGTATT TGAGAGCAAG TAATGTGAAG AAGAATCGAC GAACCGGCCA TTAGCCATAA 1110 1120 1130 1140 1150 TGGTTCACTG CTTTAGGTTT ATCAACTATG GCATTCAACT TAAACGGTTT ACCAAGTGAC GAAATCCAAA TAGTTGATAC CGTAAGTTGA ATTTGCCAAA 1160 1170 1180 1190 1200 CAACTTCAAC CAATCAGTAG TAGACTCACA AGGTCGTGTA CTAAACACTT GTTGAAGTTG GTTAGTCATC ATCTGAGTGT TCCAGCACAT GATTTGTGAA 1210 ←←opsbA928 1230 1240 1250 GGGCAGACAT CATCAACCGT GCTAACTTAG GTATGGAAGT AATGCACGAG CCCGTCTGTA GTAGTTGGCA CGATTGAATC CATACCTTCA TTACGTGCTC 1260 1270 1280 1290 1300 CGTAACGCTC ACAACTTCCC TCTAGACTTA GCTTCAACTA ACTCTAGCTC GCATTGCGAG TGTTGAAGGG AGATCTGAAT CGAAGTTGAT TGAGATCGAG Stop 1320 1330 1340 1350 AAACAACTAA TTTTTTTTTA AACTAAAATA AATCTGGTTA ACCATACCTA TTTGTTGATT AAAAAAAAAT TTGATTTTAT TTAGACCAAT TGGTATGGAT 1360 ←←opsbAstop 1380 1390 1400 GTTTATTTTA GTTTATACAC ACTTTTCATA TATATATACT TAATAGCTAC CAAATAAAAT CAAATATGTG TGAAAAGTAT ATATATATGA ATTATCGATG 1410 1420 CATAGGCAGT TGGCAGGACG TCCC 3' GTATCCGTCA ACCGTCCTGC AGGG 5'

6. Anhang 112 6.3. Nukleotidsequenz des psbA-Gens mit flankierenden Bereichen und Aminosäuresequenz des D1-Proteins aus Chlamydomonas rein-hardtii 1 CG TCC TAT TTT AAT ACT CCG AAG GAG GCA GTT GGC AGG CAA CTG CCA 47 48 CTG ACG TCC CGT AAG GGT AAG GGG ACG TCC ACT GGC GTC CCG TAA GGG 95 96 GAA GGG GAC GTA GGT ACA TAA ATG TGC TAG GTA ACT AAC GTT TGA TTT 143 144 TTT GTG GTA TAA TAT ATG TAC CAT GCT TTT AAT AGA AGC TTG AAT TTA 191 192 TAA ATT AAA ATA TTT TTA CAA TAT TTT ACG GAG AAA TTA AAA CTT TAA 239 1 Met Thr Ala Ile Leu Glu Arg Arg Glu Asn Ser Ser 12 240 AAA AAT TAA CAT ATG ACA GCA ATT TTA GAA CGT CGT GAA AAT TCT AGC 287 13 Leu Trp Ala Arg Phe Cys Glu Trp Ile Thr Ser Thr Glu Asn Arg Leu 28 288 CTA TGG GCT CGT TTT TGT GAG TGG ATC ACT TCA ACT GAA AAC CGT TTA 335 29 Tyr Ile Gly Trp Phe Gly Val Ile Met Ile Pro Cys Leu Leu Thr Ala 44 336 TAC ATC GGT TGG TTC GGT GTA ATC ATG ATC CCA TGT CTT CTT ACT GCA 383 45 Thr Ser Val Phe Ile Ile Ala Phe Ile Ala Ala Pro Pro Val Asp Ile 60 384 ACA TCA GTA TTC ATC ATC GCT TTC ATC GCT GCT CCG CCA GTA GAC ATC 431 61 Asp Gly Ile Arg Glu Pro Val Ser Gly Ser Leu Leu Tyr Gly Asn Asn 76 432 GAT GGT ATC CGT GAA CCA GTT TCA GGT TCT CTT CTT TAC GGT AAC AAC 479 77 Ile Ile Thr Gly Ala Val Ile Pro Thr Ser Asn Ala Ile Gly Leu His 92 480 ATC ATT ACA GGT GCT GTA ATC CCA ACT TCT AAC GCA ATC GGT CTT CAC 527 93 Phe Tyr Pro Ile Trp Glu Ala Ala Ser Leu Asp Glu Trp Leu Tyr Asn 108 528 TTC TAC CCA ATT TGG GAA GCT GCT TCT CTA GAC GAG TGG TTA TAC AAC 575 109 Gly Gly Pro Tyr Gln Leu Ile Val Cys His Phe Leu Leu Gly Val Tyr 124 576 GGT GGT CCT TAC CAA CTT ATC GTT TGT CAC TTC CTT CTA GGT GTA TAC 623 125 Cys Tyr Met Gly Arg Glu Trp Glu Leu Ser Phe Arg Leu Gly Met Arg 140 624 TGC TAC ATG GGT CGT GAG TGG GAA TTA TCT TTC CGT TTA GGT ATG CGT 671 141 Pro Trp Ile Ala Val Ala Tyr Ser Ala Pro Val Ala Ala Ala Ser Ala 156 672 CCA TGG ATC GCT GTA GCT TAC TCA GCT CCA GTA GCT GCA GCT TCA GCT 719 157 Val Phe Leu Val Tyr Pro Ile Gly Gln Gly Ser Phe Ser Asp Gly Met 172 720 GTA TTC TTA GTT TAC CCT ATC GGC CAA GGT TCA TTC TCT GAC GGT ATG 767 173 Pro Leu Gly Ile Ser Gly Thr Phe Asn Phe Met Ile Val Phe Gln Ala 188 768 CCT TTA GGT ATC TCT GGT ACT TTC AAC TTC ATG ATC GTA TTC CAA GCA 815 189 Glu His Asn Ile Leu Met His Pro Phe His Met Leu Gly Val Ala Gly 204 816 GAA CAC AAC ATC CTT ATG CAC CCA TTC CAC ATG TTA GGT GTT GCT GGT 863 205 Val Phe Gly Gly Ser Leu Phe Ser Ala Met His Gly Ser Leu Val Thr 220 864 GTA TTC GGT GGT TCA TTA TTC TCA GCT ATG CAC GGT TCT TTA GTT ACT 911 221 Ser Ser Leu Ile Arg Glu Thr Thr Glu Asn Glu Ser Ala Asn Glu Gly 236 912 TCA TCT TTA ATC CGT GAA ACA ACT GAA AAC GAA TCA GCT AAC GAA GGT 959 237 Tyr Arg Phe Gly Gln Glu Glu Glu Thr Tyr Asn Ile Val Ala Ala His 252 960 TAC CGT TTC GGT CAA GAA GAA GAA ACT TAC AAC ATT GTA GCT GCT CAT 1007

6. Anhang 113 253 Gly Tyr Phe Gly Arg Leu Ile Phe Gln Tyr Ala Ser Phe Asn Asn Ser 268 1008 GGT TAC TTT GGT CGT CTA ATC TTC CAA TAC GCT TCT TTC AAC AAC TCT 1055 269 Arg Ser Leu His Phe Phe Leu Ala Ala Trp Pro Val Ile Gly Ile Trp 284 1056 CGT TCA TTA CAC TTC TTC TTA GCT GCT TGG CCG GTA ATC GGT ATT TGG 1103 285 Phe Thr Ala Leu Gly Leu Ser Thr Met Ala Phe Asn Leu Asn Gly Phe 300 1104 TTC ACT GCT TTA GGT TTA TCA ACT ATG GCA TTC AAC TTA AAC GGT TTC 1151 301 Asn Phe Asn Gln Ser Val Val Asp Ser Gln Gly Arg Val Leu Asn Thr 316 1152 AAC TTC AAC CAA TCA GTA GTA GAC TCA CAA GGT CGT GTA CTA AAC ACT 1199 317 Trp Ala Asp Ile Ile Asn Arg Ala Asn Leu Gly Met Glu Val Met His 332 1200 TGG GCA GAC ATC ATC AAC CGT GCT AAC TTA GGT ATG GAA GTA ATG CAC 1247 333 Glu Arg Asn Ala His Asn Phe Pro Leu Asp Leu Ala Ser Thr Asn Ser 348 1248 GAG CGT AAC GCT CAC AAC TTC CCT CTA GAC TTA GCT TCA ACT AAC TCT 1295 349 Ser Ser Asn Asn 1296 AGC TCA AAC AAC TAA TTT TTT TTT AAA CTA AAA TAA ATC TGG TTA ACC 1343 1344 ATA CCT AGT TTA TTT TAG TTT ATA CAC ACT TTT CAT ATA TAT ATA CTT 1391 1392 AAT AGC TAC CAT AGG CAG TTG GCA GGA CGT CCC 1424

7. Literaturverzeichnis 114
7. Literaturverzeichnis
Ajlani, G.; Kirilovsky, D. L.; Picaud, M.; Astier, C.; (1989a)
Molekular analysis of psbA mutations responsible for various herbicide resistance phenotypes in Synechocystis 6714
Plant Mol. Biol. 13, 469-479
Ajlani, G.; Meyer, I.; Vernotte, C.; Astier, C.; (1989b)
Mutation in phenol-type herbicide resistance maps within the psbA gene in Synechocystis 6714
FEBS Lett. 246, 207-210
Arnon, D. I.; (1949)
Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris.
Plant Physiol. 24, 1-15
Barber, J.; Chapmann, D. J.; Telfer, A.; (1987)
Characterisation of a PSII reaction centre isolated from the chloroplasts of Pisum sativum
FEBS Lett. 220, 67-73
Barber, J.; Nield, J.; Morris, E. P.; Zheleva, D.; Hankamer, B.; (1997)
The structure, function and dynamics of photosystem two
Physiol. Plant. 100, 817-827
Bennoun, P.; Spieler-Herz, M.; Erickson, J.; Girard-Bascou, J.; Pierre, Y.; Delosme, M.; Rochaix, J.-D.; (1986)
Characterisation of photosystem II mutans of Chlamydomonas reinhardtii lacking the psbA gene
Plant Mol. Biol. 6, 151-160
Bettini, P.; McNally, S.; Sevignac, M.; Darmency, H.; Gasquez, J.; Dron, M.; (1987)
Atrazine Resistance in Chenopodium album
Plant Physiol. 84, 1442-1446

7. Literaturverzeichnis 115 Blyden, E. R.; Gray, J. C.; (1986)
The molecular basis of atrazine herbicide resistance in Senecio vulgaris L.
Biochem. Soc. Trans. 14, 62
Boynton, J. E.; Gilham, N. W.; Harris, E. H.; Hosler, J. P.; Johnson, A. M.; Jones, A. R.; Randolph-Anderson, B. L.; Robertson, D.; Klein, T. M.; Sherk, K. B.; Sanford, J. C.; (1988)
Chloroplast Transformation in Chlamydomonas with High Velocity Microprojectiles
Science 240, 1534-1537
Braun, P.; Greenberg, B. M.; Scherz, A.; (1990)
D1-D2-cytochrom b559 Complex from the Aquatic Plant Spirodela oligorrhiza: Correlation between Complex Integrity, Spectroscopic Properties, Photochemical Activity, and Pigment Composition
Biochemistry 29, 10376-10387
Chua, N.-H.; Bennoun, P.; (1975)
Thylakoid Membrane Polypeptides of Chlamydomonas reinhardtii: Wilde-Type and Mutant Strains Deficient in Photosystem II Reaction Center
Proc. Natl. Acad. Sci. USA 72, 2175-2179
Chua, N.-H.; (1980)
Electrophoretic Analysis of Chloroplast Proteins
Meth. Enzymol. 69, 434-446
Creuzet, S.; Ajlani, G.; Vernotte, C.; Astier, C.; (1990)
A New Ioxynil-Resistant Mutant in Synechocystis PCC 6714; Hypothesis on the Interaction of Ioxynil with the D1 Protein
Z. Naturforsch. 45c, 436-440
Dalla Chiesa, M.; Friso, G.; Deak, Z.; Vass, I.; Barber, J.; Nixon, P. J.; (1997)
Reducet turnover of the D1 polypeptide and photoactivation of electron transfer in novel herbicide resistant mutants of Synechocstis sp. PCC 6803
Eur. J. Biochem. 248, 731-740

7. Literaturverzeichnis 116
Debus, R. J.; Barry, B. A.; Babcock, G. T.; McIntosh, L.; (1988a)
Site-directed mutagenesis identifies a tyrosins involved in the photosynthetic oxygen-evolving system
Proc. Natl. Acad. Sci. USA 58, 427-430
Debus, R. J.; Barry, B. A.; Sithole, L.; Babcock, G. T.; McIntosh, L.; (1988b)
Directed mutagenesis indicates that the donor to P680+ in photosystem II is tyrosine-161 of the D1-polypeptide
Biochemistry 27, 9071-9074
Debus, R. J.; (1992)
The manganese and calcium ions of photosynthetic oxygen evolution
Biochim. Biophys. Acta 1102, 269-352
Deisenhofer, J.; Epp, O.; Miki, K.; Huber, R.; Michel, H.; (1985)
Structure of protein subunits in the photosynthetic reaction centre of Rhodopseudomonas viridis at 3 Å resolution
Nature 318, 618-624
Dostatni, R.; Meyer, H. E.; Oettmeier, W.; (1988)
Mapping of two tyrosine residues involved in the quinone-(QB) binding site of the D-1 reaction center polypeptide of photosystem II
FEBS Lett. 239, 207-210
Draber, W.; Kluth, F. J.; Tietjen, K.; Trebst, A.; (1991)
Herbizide in der Photosyntheseforschung
Angew. Chem. 103, 1650-1663
Durrant, J. R.; Giorgi, L. B.; Barber, J.; Klug, D. R.; Porter, G.; (1990)
Charakterisation of triplet states in isolated Photosystem II reaction centres: oxygen quenching as a mechanism for photodamage
Biochim. Biophys. Acta 1017, 167-175

7. Literaturverzeichnis 117 Durrant, J. R.; Klug, D. R.; Kwa, S. L. S.; van Grondelle, R.; Porter, G.; (1995)
A multimer model for P680, the primary electron donor of photosystem II
Proc. Natl. Acad. Sci. USA 92, 4798-4802
Erickson, J. M.; Rahire, M.; Rochaix, J.-D.; (1984)
Chlamydomonas reinhardtii gene for the 32 000 mol. wt. protein of photosystem II contains four large introns and is located entirely within the chloroplast inverted repeat
EMBO J. 3, 2753-2762
Erickson, J. M.; Rahire, M.; Rochaix, J.-D.; (1985)
Herbicide Resistance and Cross-Resistance: Changes at Three Distinct Sites in the Herbicide-Binding Protein
Science 228, 204-207
Erickson, J. M.; Pfister, K.; Rahire, M.; Togasaki, R. K.; Mets, L.; Rochaix, J.-D.; (1989)
Molecular and Biophysical Analysis of Herbicide-Resistant Mutants of Chlamydomonas reinhardtii: Structure-Function Relationship of the Photosystem II D1 Polypeptide
Plant Cell 1, 361-371
Förster, B.; Heifetz, P. B.; Lardans, A.; Boynton, J. E.; Gillham, N. W.; (1997)
Herbicide Resistance and Growth of D1 Ala 251 Mutants in Chlamydomonas
Z. Naturforsch. 52c, 654-664
Galloway, R. E.; Mets, L. J.; (1984)
Atrazine, bromacil and diuron resistance in Chlamydomonas. A single non-mendelian genetic locus controls the structure of the thylakoid binding site
Plant Physiol. 74, 469-474
Gingrich, J. C.; Buzby, J. S.; Stirewalt, V. L.; Bryant, D. A.; (1988)
Genetic analysis of two new mutations resulting in herbicide resistance in the cyanobacterium Synechococcus sp. PCC 7002
Photosyn. Research 16, 83-90

7. Literaturverzeichnis 118 Goloubinoff, P.; Edelman, M.; Hallick, R. B.; (1984)
Chloroplast-coded atrazine resistance in Solanum nigrum: psbA loci from susceptible and resistant biotypes are isogenic except for a single codon change
Nucleic Acids Res. 12, 9489-9496
Gorman, D. S.; Levine, R. P.; (1965)
Cytochrome f and plastocyanin: their sequence in the photosynthetic electron transport chain of Chlamydomonas reinhardtii
Proc. Natl. Acad. Sci. USA 54, 1665-1669
Govindjee; Vernotte, C.; Peteri, B.; Astier, C.; Etienne, A. L.; (1990)
Differential sensitivity of bicarbonate-reversible formate effects on herbicide-resistant mutants of Synechocystis 6714
FEBS Lett. 267, 273-276
Hanahan, D.; (1985)
Techniques for transformation of Escherichia coli.
In: DNA-cloning 1, 109-135
Hrsg. Glover, D. M.
IRL Press, Oxford
Hankamer, B; Morris, E. P.; Barber, J.; (1999)
Revealing the structure of the oxygen-evolving core dimer of photosystem II by cryoelectron crystallography
Nat. Struct. Biol. 6, 560-564
Harris, E. H.; (1988)
The Chlamydomonas Sourcebook: a comprehensive guide to biology and laboratory use
Academic Press, San Diego
Heiß, S.; (1992)
Untersuchung zur Transformation des Chloroplasten der Grünalge Chlamydo-monas reinhardtii am Beispiel des psbA-Gens
Dissertation, Ruhr-Universität Bochum

7. Literaturverzeichnis 119 Hirschberg, J.; McIntosh, L.; (1983)
Molecular Basis of Herbicide Resistance in Amaranthus hybridus
Science 222, 1346-1348
Hirschberg, J.; Bleecker, A.; Kyle, D. J.; McIntosh, L.; Arntzen, C. J.; (1984)
The molecular basis of triazine-herbicide resistance in higher-plant chloroplasts
Z. Naturforsch. 39c, 412-420
Holzenburg, A.; Bowley, M. C.; Wilson, F. H.; Nicholson, W. V.; Ford, R. C.; (1993)
Three-dimensional structure of photosystem II
Nature 363, 470-472
Janatkova, H.; Wildner, G. F.; (1982)
Isolation and characterization of metribuzin-resistant Chlamydomonas reinhardtii cells
Biochim. Biophys. Acta 682, 227-233
Jansson, C.; Mäenpää, P.; (1997)
Site-Directed Mutagenesis for Structure-Function Analyses of the Photosystem II Reaction Center Protein D1
in: Progress in Botany, 352-367
Hrsg.: Behnke, H.-D.; Lüttge, U.; Esser, K.; Kadereit, J. W.; Runge, M.
Springer Verlag; Berlin, Heidelberg, New York
Johanningmeier, U.; (1987a)
Expression of the psbA Gen in E. coli
Z. Naturforsch. 42c, 755-757
Johanningmeier, U.; Bodner, U.; Wildner, G. F.; (1987b)
A new mutation in the gene coding for the herbicide-binding protein in Chlamydomonas
FEBS Lett. 211, 221-224

7. Literaturverzeichnis 120 Johanningmeier, U.; Heiss, S.; (1993)
Construction of a Chlamydomonas reinhardtii mutant with an intronless psbA gene
Plant. Mol. Biol. 22, 91-99
Johanningmeier, U., Sopp, G., Brauner, M., Altenfeld, U., Orawski, G., Oettmeier, W.; (2000)
Herbicide Resistance and Supersensitivity in Ala250 or Ala251 Mutants of the D1 Protein in Chlamydomonas reinhardtii
Pestic. Biochem. Physiol. 66, 9-19
Karabin, G. D.; Farley, M.; Hallick, R. B.; (1984)
Chloroplast gene for Mr 32.000 polypeptide of photosystem II in Euglena gracilis is interruptet by four introns with conserved boundary sequences
Nucl. Acids Res. 12, 5801-5812
Keller, M., Stutz, E.; (1984)
Structure of Euglena gracilis chloroplast gene (psbA) coding for the 32-kDa protein of photosystem II
FEBS Lett. 175, 173-177
Kirilovsky, D. L.; Ajlani, G.; Picaud, M.; Etienne, A.-L.; (1989)
Mutations responsible for high lihgt sensitivity in an atrazin-resistant mutant of Synechocystis 6714
Plant. Mol. Biol. 13, 355-363
Klein, T. M.; Wolf, E. D.; Sanford, L. C.; (1987)
High-velocity microprojectiles for delivering nucleic acids into living cells
Nature 327, 70-73
Kless, H.; Oren-Shamir, M.; Malkin, S.; McIntosh, L.; Edelman, M.; (1994)
The D-E Region of the D1 Protein Is Involved in Multiple Quinone and Herbicide Interactions in Photosystem II
Biochemistry 33, 10501-10507

7. Literaturverzeichnis 121 Landt, O.; Grunert, H.-P.; Hahn, U.; (1990)
A general method for rapid site-directed mutagenesis using the polymerase chain reaction
Gene 96, 125-128
Mäenpää, P.; Sippola, K.; Rokka, A.; Aro, E.-M.; (1998)
Substitution of Ala-251 of the D1 reaction centre polypeptide with a charged residue results in impaired function of photosystem II
Plant Mol. Biol. 38, 1191-1200
Michel, H.; Deisenhofer, J.; (1988)
Relevance of the Photosynthetic Reaction Center from Purple Bacteria to the Structure of Photosystem II
Biochemistry 27, 1-7
Nanba, O.; Satoh, K.; (1987)
Isolation of a photosystem II reaction center consisting of D-1 and D-2 polypeptides and cytochrome b-559
Proc. Nat. Acad. Sci. USA 84, 109-112
Narusaka, Y.; Narusaka, M.; Kobayashi, H.; Sato, K.; (1998)
The Herbicide-Resistant Species of the Cyanobacterial D1 Protein Obtained by Thorough and Random in vitro Mutagenesis
Plant Cell Physiol. 39, 620-626
Newman, S. M.; Boynton, J. E.; Gillham, N. W.; Randolph-Anderson, B. L.; Johnson, A. M.; Harris, E. H.; (1990)
Transformation of Chloroplast Ribosomal RNA Genes in Chlamydomonas: Molecular and Genetic Characterization of Integration Events
Genetics 126, 875-888
Noguchi, T.; Tomo, T.; Inoue, Y.; (1998)
Fourier transform infrared study of the cation radical of P680 in the photosystem II reaction center: evidence for charge delocalization on the chlorophyll dimer
Biochemistry 37, 13614-13625

7. Literaturverzeichnis 122 Oettmeier, W.; Hilp, U.; (1991)
Structure-Activity Relationships of Triazinone Herbicides on Resistent Weeds and Resistent Chlamydomonas reinhardtii
Pestic. Sci. 33, 399-409
Oettmeier, W.; (1992)
Herbicides of photosystem II
in: The photosystems: Structure, function and molecular biology, 349-408
Hrsg.: Barber, J.
Elsevier Science Publishers; Amsterdam
Ohad, N.; Hirschberg, J.; (1990)
A similar structure of the herbicide binding site in photosystem II of plants and cyanobacteria is demonstrated by site specific mutagenesis of the psbA gene
Photosynth. Res. 23, 73-79
Ohad, N.; Amir-Shapira, D.; Koike, H.; Inoue, Y.; Ohad, I.; Hirschberg, J.; (1990)
Amino acid substitutions in the D1 protein of photosystem II affect QB-stabilization and accelerate turnover of D1
Z. Naturforsch. 45c, 402-407
Orawski, G.; (1998)
Herbizidresistenz in Mutanten von Chlamydomonas reinhardtii
Diplomarbeit, Ruhr-Universität Bochum
Páy, A.; Smith, M. A.; Nagy, F.; Márton, L.; (1988)
Sequence of the psbA gene from wild type and triazine resistant Nicotiana plumbaginifolia
Nucleic Acids Res. 16, 8176
Perewoska, I.; Etienne, A. L.; Miranda, T.; Kirilovsky, D.; (1994)
S1 destabilization and higher sensitivity to light in metribuzin-resistant mutants
Plant. Physiol. 104, 235-245

7. Literaturverzeichnis 123 Pfister, K.; Steinback, K. E.; Gardner, G.; Arntzen, C. J.; (1981)
Photoaffinity labeling of an herbicide receptor protein in chloroplast membranes
Proc. Natl. Acad. Sci. USA 78, 981-985
Przibilla, E.; Heiss, S.; Johanningmeier, U.; Trebst, A.; (1991)
Site-specific mutagenesis of the D1 subunit of photosystem II in wild-type Chlamydomonas
Plant Cell 3, 169-174
Pucheu, N.; Oettmeier, W.; Heisterkamp, U.; Masson, K.; Wildner, G. F.; (1984)
Metribuzin-Resistant Mutants of Chlamydomonas reinhardtii
Z. Naturforsch. 39c, 437-439
Rhee, K. H.; Morris, E. P.; Zhelva, D.; Hankamer, B.; Kühlbrandt, W.; Barber, J.; (1997)
Two-dimensional structure of the plant photosystem II reaction centre at 8 Å resolution
Nature 389, 522-526
Rhee, K. H.; Morris, E. P.; Barber, J.; Kühlbrandt, W.; (1998)
Three-dimensional structure of the plant photosystem II reaction centre at 8 Å resolution
Science 396, 283-286
Ruffle, S. V.; Donnelly, D.; Blundell, T. L.; Nugent, J. H. A.; (1992)
A three-dimensional model of the Photosystem II reaction centre of Pisum sativum
Photosynth. Res. 34, 287-300
Sager, R.; Granick, S.; (1953)
Nutrtional studies with Chlamydomonas reinhardtii
Ann. N. Y. Acad. Sci. 56, 831-838
Sambrook, J.; Fritsch, E. F.; Maniatis, T.; (1989)
Molecular cloning - a laboratory manual, second edition
Cold Spring Harbor Laboratory Press, New York

7. Literaturverzeichnis 124 Sato, F.; Shigematsu, Y.; Yamada, Y.; (1988)
Selection of anatrazine-resistant tobacco cell line having a mutant psbA gene
Mol. Gen Genet. 214, 358-360
Schägger, H.; Borchart, U.; Aquila, H.; Link, T. A.; von Jagow, G.; (1985)
Isolation and amino acid sequence of the smallest subunit of beef heart bc1 complex
FEBS Lett. 190, 89-94
Schönfeld, M.; Yaacoby, T.; Ben-Yehuda, A.; Rubin, B.; Hirschberg, J.; (1987)
Triazine resistance in Phalaris paradoxa: physiological and molecular analyses
Z. Naturforsch. 42c, 779-782
Schwenger-Erger, C.; Böhnisch, N.; Barz, W.; (1999)
A New psbA Mutation Yielding an Amino-Acid Exchange at the Lumen-Exposed Site of the D1 Protein
Z. Naturforsch. 54c, 909-914
Shuvalov, V. A.; Heber, U.; Schreiber, U.; (1989)
Low temperature photochemistry and spectral properties of a photosystem 2 reaction center complex containing the proteins D1 and D2 and two hemes of Cyt b-559
FEBS Lett. 258, 27-31
Smeda, R. J.; Hasegawa, P. M.; Goldsbrough, P. B.; Singh, N. K.; Weller, S. C.; (1993)
A serine-to-threonine substitution in the triazine herbicide-binding protein in potato cells results in atrazine resistance without impairing productivity
Plant Physiol. 103, 911-917
Solymosi, P.; Lehoczki, E.; (1989)
Co-resistance of atrazine-resistant Chenopodium and Amaranthus biotypes to other photosystem II inhibiting herbicides
Z. Naturforsch. 44c, 119-127

7. Literaturverzeichnis 125 Sopp, G.; (1995)
Struktur- und Funktionanalyse der QB-Bindenische photosynthetischer Reakti-onszentren mit Hilfe von Hemmstoffen
Dissertation, Ruhr-Universität Bochum
Sueoka, N.; (1960)
Mitotic Replication of Deoxyribonucleic Acid in Chlamydomonas reinhardtii
Proc. Natl. Acad. Sci. USA 46, 83-91
Tang, X. S.; Fushimi, K.; Satoh, K.; (1990)
D1-D2 complex of the photosystem II reaction center from spinach
FEBS Lett. 273, 257-260
Tetenkin, V. L.; Gulyaev, B. A.; Seibert, M.; Rubin, A. B.; (1989)
Spectral properties of stabilized D1/D2/cytochrome b-559 photosystem II reaction center complex
FEBS Lett. 250, 459-463
Thuillier-Bruston, F.; Calvayrac, R.; Duval, E.; (1996)
Partial molecular analysis of the psbA gene in Euglena gracilis mutants exhibiting resistance to DCMU and atrazine
Z. Naturforsch. 51c, 711-720
Tietjen, K. G.; Kluth, J. F.; Andree, R.; Haug, M.; Linding, M.; Müller, K. H.; Wroblowsky, H. J.; Trebst, A.; (1990)
The Herbicide Binding Niche of Photosystem II - A Model
Pestic. Sci. 31, 65-72
Tischer, W.; Strotmann, H.; (1977)
Relationship between inhibitor binding by chloroplasts and inhibition of photosynthetic electron transport
Biochim. Biophys. Acta 460, 113-125

7. Literaturverzeichnis 126 Towbin, H.; Staehelin, T.; Gordon, J.; (1979)
Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications
Proc. Natl. Acad. Sci. USA 76, 4350-4354
Trebst, A.; Wietoska, H.; Draber, W.; Knops, H. J.; (1978)
The Inhibition of Photosynthetic Electron Flow in Chloroplasts by the Dinitrophenylether of Bromo- or Iodo-nitrothymol
Z. Naturforsch. 33c, 919-927
Trebst, A.; (1986)
The Topology of the Plastoquinone and Herbicide Binding Peptides of Photosystem II on the Thylakoid Membrane
Z. Naturforsch. 41c, 240-245
Trebst, A.; (1987)
The Three-Dimensional Structure of the Herbicide Binding Niche on the Reaction Center Polypeptides of Photosystem II
Z. Naturforsch. 42c, 742-750
Velthuys, B. R.; (1981)
Electron-dependent competition between plastoquinone and inhibitors for binding to photosystem II
FEBS Lett. 126, 277-281
Vermaas, W. F. J.; Williams, J. G. K.; Arntzen, C. J.; (1987)
Site-Directed Mutations of Two Histidine Residues in the D2 Protein Inactivate and Destabilize Photosystem II in the Cyanobacterium Synechocystis 6803
Z. Naturforsch. 42c, 762-768
Vermaas, W.; Vass, I.; Eggers, B.; Styring, S.; (1994)
Mutation of a putative ligand to the non-heme iron in Photosystem II: implications for QA reactivity, electron transfer, and herbicide binding
Biochim. Biophys. Acta 1184, 263-272

7. Literaturverzeichnis 127 Whitelegge, J. P.; Jewess, P.; Pickering, M. G.; Gerrish, C.; Camilleri, P.; Bowyer, J. R.; (1992)
Sequence analysis of photoaffinity-labelled peptides derived by proteolysis of photosystem-2 reaction centres from thylakoid membranes treated with [14C]azidoatrazine
Eur. J. Biochem. 207, 1077-1084
Wildner, F. G.; Heisterkamp, U.; Trebst, A.; (1990)
Herbicide Cross-Resistance and Mutations of psbA Gene in Chlamydomonas reinhardtii
Z. Naturforsch. 45c, 1142-1150
Wolber, P. K.; Eilmann, M.; Steinback, K. E.; (1986)
Mapping of the triazine binding site to a highly conserved region of the QB-Protein
Arch. of Biochem. and Biophys. 248, 224-233
Xiong, J.; Subramaniam, S.; Govindjee; (1996)
Modeling of the D1/D2 proteins and cofactors of the photosystem II reaction center: Implications for herbicide and bicarbonate binding
Protein Sci. 5, 2054-2073
Xiong, J.; Hutchinson, R. S.; Sayre, R., T.; Govindjee; (1997)
Modification of the photosystem II acceptor side function in a D1 mutant (arginine-269-glycine) of Chlamydomonas reinhardti
Biochim. Biophys. Acta 1322, 60-76
Xiong, J.; Subramaniam, S.; Govindjee (1998)
A knowledge-based three dimensional model of the photosystem II reaction center of Chlamydomonas reinhardtii
Photosynth. Res. 56, 229-254
Zouni, A.; Witt, H. T.; Kern ,J.; Fromme, P.; Krauß, N.; Saenger, W.; Orth, P.; (2001)
Crystal structure of photosystem II from Synechococcus elongatus at 3,8 Å resolution
Nature 409, 739-743

7. Literaturverzeichnis 128 Zurawski, G.; Bohnert, H. J.; Whitfeld, P. R.; Bottomley, W.; (1982)
Nucleotide sequence of the gene for Mr 32.000 thylakoid membrane protein from Spinacia oleracea and Nicotiana debneyi pridicts a totally conserved primary translation product of Mr 38.950
Proc. Natl. Acad. Sci. USA 79, 7699-7703

Lebenslauf
Persönliche Daten
Name: Orawski, geb. Maszka
Vorname: Grazyna
Geburtsdatum: 07.02.1967
Geburtsort: Nikolai / Polen
Familienstand: verheiratet
Schulbildung
IX. 1974 - VI. 1982 Grundschule in Nikolai
IX. 1982 - V. 1986 Allgemeinbildendes Lyzeum in Nikolai
6. VI. 1986 Abschluß: Abitur
Studium
X. 1986 - VI. 1988 Studium der Biologie an der Schlesischen Universität
in Kattowitz / Polen
V. - XII. 1989 Sprachkurs der GFBA e.V. in Geseke-Eringerfeld
IX. 1991 -V. 1998 Studium der Biologie an der Ruhr-Universität Bochum
20. V. 1998 Abschluß: Diplom
Thema der Diplomarbeit: “Herbizidresistenz in Mutanten
von Chlamydomonas reinhardtii”
Angefertigt am Lehrstuhl für Biochemie der Pflanzen
Betreuer der Arbeit: Prof. Dr. W. Oettmeier
Seit VII. 1998 Promotion am Lehrstuhl für Biochemie der Pflanzen mit
dem Thema “Struktur-Funktionsanalyse der QB-
Bindenische im Photosystem II von Chlamydomonas
reinhardtii”
Betreuer der Arbeit: Prof. Dr. W. Oettmeier
VII. 1998 - IX. 2000 Stipendiatin des Graduiertenkollegs
“Biogenese und Mechanismen komplexer Zellfunktionen”
Berufliche Tätigkeit
Seit X. 2000 Einstellung als wissenschaftliche Hilfskraft von
Prof. Dr. W. Oettmeier





























