Embed Size (px)
Citation preview

WISCONSIN STATE:
UNIVERSITY
STEVENS FOINT, TYISCONSIN
THE MUSEUM OF NIATURALHISTORY
Reports on the Fquns ond Floraof Wisconsin
NO. 18

"'. t.;
WORKS ON BREEDING BIRDS AND GIGANTIC TOADS
Charles A. Long
1. The Evolution of Nest·Slte Dls~ractlon Displays . . . Birds (Paridae and Sittidae). . . . . . . . • . . . . . . . • . . . . • . . .. .. .. . . . . . . • . . . .. .1
2. Rare Gigantic Toads ..• from Lake Michigan Isles ..••...•...........•....... 16
10 May 1982
THE MUSEUM OF NATURAL HISTORY Stevens Point, Wisconsin
Reports on the Fauna and Flora of Wisconsin No. 18

THE EVOLUTION OF NEST·SITE DISTRACTION DISPLAYS IN TWO HOLE-NESTING F AMILlES
OF. BIRDS (PARIDAE AND SITIIDAE)
Charles A. Long
The nature of nest-site distraction displays-Skutch (1976) suggests that various nest-site displays against nest·robbing predators be called "distraction displays". These vary from "feigning injury" (or more accurately injury simulation), to the "rodent run" (mbnicking a substitute prey), to threats of intimidation, to passive displays made by boldly exhibiting white spots, to death feigning, and even in one instance to the act of nest building in another bird's abandoned nest. Not all birds make nest·site distraction displays; many birds vigorously defend their nests as best they dare by overt attacks. Several authors have noted that the nest-site distraction display is often complex, comprised of several components of basic bird behavior such as the simulation of the gaping beak of fledglings, and so on (see Armstrong, 1949). It Is the aim of this paper to analyze distraction displays in nuthatches (Sittidae) and the Paddae to study their origin and differentiation.
Skutch (1976) has contrasted the conspicuous nest1ite distraction displays with the secretive behavior so often performed by parent birds, quietly slipping away from their nest. In communications the former is positive, an advertisement, and the latter is negative. Nest·site distraction displays include a variety of posi· tive communications, raising feathers, spreading the wings, flaring the tail, making noises, displaying bold patterns (usually of black and white), perfomiing unusu,1 and surprising movements (which often simulate injury) and so forth. Usually a distraction display involves several of these advertisements.
Skutch (1976). Hinde (1952) and others have cons~dered nest-site distraction displays as reproductive behavior, primarily because they seemgeneraUy confined from the season of incubation to that of fledging. Slmtch discussed how these displays are highly variable in relation to other kinds of reproductive behavior. such as nest buDding. He concludes that the bird's Intelligence in thwarting nest-robbing has influenced the evolution Qf the displays, because the danger during the display requires the bird to be funy alert and reactive whUe appearing in some cases just the opposite. .
An early theory, now disparaged, suggests that a conflict between the Urge to flee and the devotion to young or eggs results in muscular inhibitions. The opposing tendencies to fight and flee are important in aggressive displays (Hinde, 1952) and will be shown to be important in the expression of certain distraction displays. Nevertheless, the muscles are functional, and the bird displaying is alert and in fun command of its actions (Skutch, 1976). .
Terms such as "paralysis", "death feigning", "comatose", and so forth have been for the most part quite misleading, and "injury feigning" has been often misapplied to describe very aggressive displays. Some terms ouaht to be carefully qualified in discussing nest·,ite distractions, and many displays ought to be redescribed..
That nest·site displays are generally simUar within species and even within some families is evidence for their genetical basis. No one has ever clabned that the widespread but diverse displays observed in many
. families of birds are homologous. Skutch (1976) remarked that some wood warblers make distraction displays whereas their supposed ancestors, the vireos, do not.' ..
Nat-site distraction displays In Pariclae and Sittidae-Some nest-site distraction displays in the Great Tits, Chickadees, etc. and in the Nuthatches are very sironar. and all of them 10 far studied (Table I) are somewhat alike and amons birds, unusual. .Usually cavity nesting birds make no distraction displays. To my knowledge, only in antbirds is there any behavior reany comparable to that of parids and slttids. and in antbirds there is no strong awession apparent.
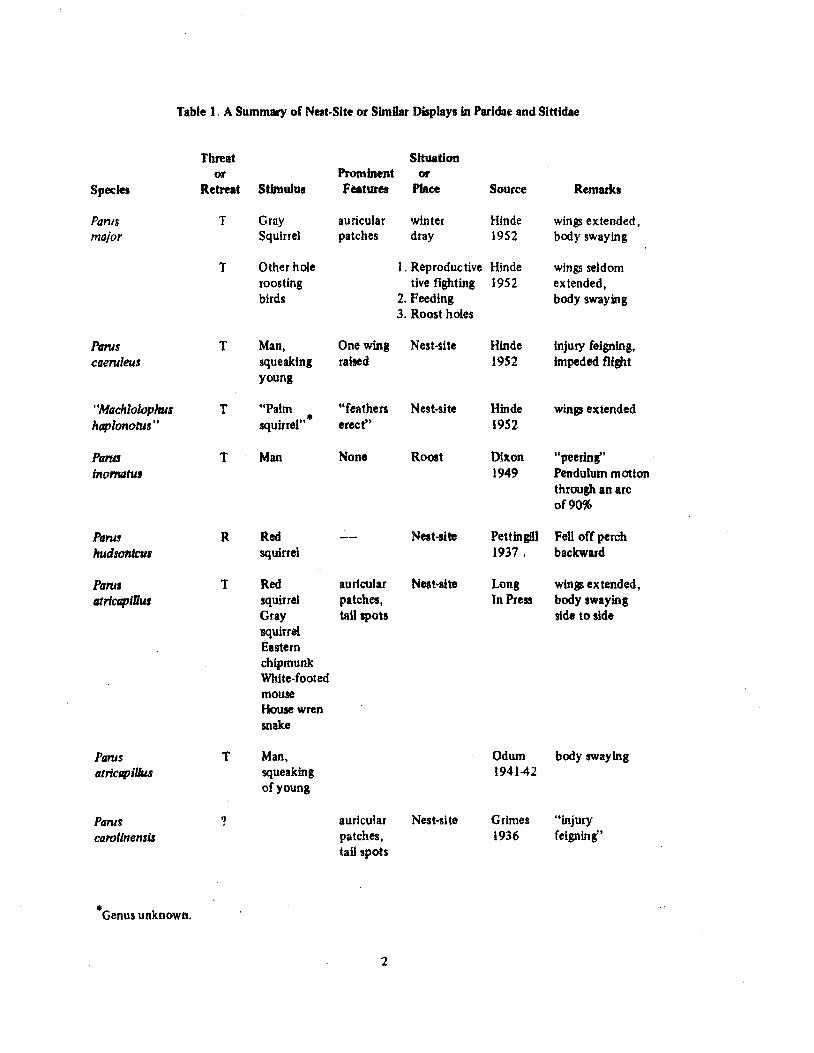
Table 1. A Summary of Nest-Site or Similar Displays in Paridae and Sittidae
Threat Stt~tion
or Prominent or Species Retreat Stimulus Features Place Source Rematks
Pams T Gray auricular winter Hinde wings extended. major Squirrel patches dray 1952 body swaying
T Other hole 1. Reproductive Hinde wings seldom roosting tive fighting 1952 extended, birds 2. Feeding body swaying
3. Roost holes
Parus T Man, One wing Nest-site Hinde injury feigning, caeruleus squeaking raised 1952 impeded flight
young
"Machlo/l)phus T "Palm "feathers Nest-site Hinde wings extended hapionolUs .. squirreJ" • erect" 1952
PatUI T Man None Roost Dixon "peerinr' inomahts 1949 Pendulum motion
through an arc of 90%
Parus R Red Nest-site Pettingill Fell off perch hudsontcus squirrel 1937. backward
Paru. T Red auricular Nest-site Long wings extended, fltricapiUus squirrel patches, In Press body swaying
Gray tail spots side to side 'Squirrel Eastern chipmunk White-footed mouse House wren snake
Parns T Man, Odum body swaying atrlcapillus squeaking 194142
of young
Pantg ? auricular Nest-site Grimes "injury Cflroiinensi.r patches, 1936 feigning"
tail spots
•Genus unknown.
2

Table I (Cont). ~ summary of Nest-Site or Similar Displays in Partdae and Sittldae
Threat or Prominent
Species Retreat Stimulus F«'Altures
Sitra T Red and auricular carolinensis Gray patches,
squirrels tailspots Eastern underwing chipmunk spots
Sitta T Red, Gray tarolmensis and flying
squirrels Eastern chipmunks, White-footed mouse, snake
SUta T chipmunk carolinensll somebirds
Sitta T Birds TaU spots europaea
Sitta T Birds whitehead;
•This is probably a "head forward" feeding threat.
Situation or
Place
Nest-site
Nest-site
Bird feeder
Bird feeder
Nest-site
Soutee Remarks
Kilham wings extended 1957 body swaying Stoner 1942
Long wings extended, see Tables body swaying 2-3
Teale wings extended'" 1974
lohrl wings extended· 1967
lohrl feathers fluffed 1967
3

Hissing of females on the nest expressed toward predators has been discussed by Brewer (J 961) and others in chickadees and nuthat~hes. It is not studied here. The distraction displays discussed here involve !laring the tail and spreading the wings, not on the nest but near it.
Anyone who observes the full wingspread distraction displays of the White-breasted Nuthatch (SUra caro/inensis) and the Black-capped Chickadee (Parus atricapillus) to small rodents will be impressed by their similarity and may hypothesize that the behavior Is homologous in the two families. The similarities in size, color of plumage, pattern of spots, nesting habits, foraging habits. rearing the young in family groups, winter activities and so forth aU suggest the families are closely related. H()wever, a survey of other members of the two families reveals variation in color, pattern, habits, and even in size. Furthermore. there is variation in the observed nest-site distraction displays. Therefore, one may hypothesize that the similarities in the Whitebreasted Nuthatch and the Blackcapped Chickadee have led to analogous behavior. Even mimicry may be to some degree suspected; one of the birds possibly evolved a superior display adaptation while in association with the other, which consequently evolved a similar display.
A survey of observations on nest-site distraction displays in Paridae and Sittidae shows that most displays were threats, often accompanied by 'gressive attacks_ Usually. of course, the display was made near the nest, and in some of the exceptions there might be confusion between nest-site distractions and "head-forward" threats (FiS. 1). These are used in fighting over food and in supplanting encounters while foraging (Hinde, 1952).
Observations on the displays of the White-breasted Nuthatch are sumrnariZed in Tables 2-3, and observations on the Black~apped Chickadee are summarized by Long (1982). Displays were induced by hoisting a stuffed mammal or live snake to the nesHite by means of a screw eye and nylon fishing line.
The White-breasted Nuthatch has on the underside pf each wing a black-margined white spot (Fig. 2). Long (1982) has suggested these white spots have evolved in mimicry of e(!gS. and wing spots have evolved in a few other birds having nest-slte distraction displays. These birds do not attack the predator but instead they threaten menacingly.' displaying the spots prominently_However, in the nuthatch the display sometimes is le'\s threatening and seem. then to be only a pas$ive display of spots on head. wings and tail, much as moths display "eye-like" spots. The two displays are referred to as "Eg" or "By", respectively (fables 2,3).
Both in the Btack-capped Chickadee and the White-breasted Nuthatch, the displaying bird usually faces the predator, leans forward, spreads the tail and wing full, turns the head slowly from side to side (to display the auricular pattern), and continues this serp~ntine swaying up to 30 seconds (Fig. 3). The Chickadee may then attack. It does not possess underwing spots. The nuthatch may peck the predator's face as a last resort (Long, 1982).
The effects of nest predation-Kluijver (1951) reported that nests of Great Tits were destroyed by squirrels, rats (Rattus norvegicus), weasels (Mrntela nivali8), and martens <Manes manes). Of 2,022 clutches, 362 nests were destroyed (17.9%). In general, the proportion disturbed is not higher than 20%.
Hole-nesting birds gain security against ground foraging predators, but the nests are usually both obvious and accessible to scansorial mammals ranging into the nest area. The most common qtarrunals that endanger nest-hole sites are squirrels, and probably that is the chief reason most distraction nest-site displays reported for padds and sittids have been induced by the presence of various squirrels (fables }-3). Other predators studied include man, snakes, and birds of prey. Displays and reactions to them are often different from those made to sqUirrels, but are not always different when performed at the nest-site.
The distraction displays probably lessen predation (,'In eggs and young. Such predation is uncommon, but it doubtless is and always was important. Both Sittaand Pams display to Red Squirrels, which are considered notorious nest robbers. But according to Hatt (1929) the reputation is not fully deserved except in sorne local situations. In my observations Red Squirrels avoided or were driven away from nest sites of Pams and Sitta, but one did rob the nest of a flicker in the nuthatch nest tree. In spring 1980, a mother Gray Squirrel moved from a nest cavity to a leaf nest in an adjacent tree making the hole available to nuthatches. In the numerous distraction displays to tree squirrels that have been described, not once did a squirrel succeed in destroying the eggs or young. Hinde (1952) reported that Gray Squirrels "occasIonally" rob nests. Hvwever. one walked past a nest hole when fledgling Great Tits were departing the nest, and did not evoke
4

Table 2, Displays at Nest Site of White-breasted Nuthatch.
Possibly observations from the years 1975·1977 are of the same pair of nu thatches. Sometimes the displaying bird faces the predator with wing spots evident and possibly mimks a boldly patterned parent bird defending its "white eggs," Eg, but sometimes the 'displaying bird merely dings to a tree and flashes white auricular patches and eye·like spots on wings and taU like a huge moth, Ey, as it sways back and forth. Either display seldom lasts as long as 30 seconds, and either was successful against squirrels in Long's observations.
Observer Date Place Pose Remarks
Long, Wisconsin 15 April 1975 Nest Site ?Ey Three separate displays on this date.
Long, Wisconsin 11 June 1976 Nest site ? Display to squirrel as fledglings left nest.
Long, Wisconsin 12 April 1977 Nest site Eg Probably by male. Long, Wisconsin 13 April 1977 Nest site Eg Probably by male. Long, Wisconsin 15 April 1977 Nest site Eg Probably by male. long, Wisconsin 6 May 1980· Nest site Bg Both male and female
displayed. Long, Wisconsin 30 May 1980 Nest hole By Female alone, to red squirrel,
entrance young In nest, post vertical but head upright.
Stoner, New York 16 April 1942 Nest hole ?Eg Both male and female entrance displayed.
Kilham, Maryland 7 May 1957 Nest hole Ey Display by male. entrance
Kilham, Maryland 17 May 1957 Nest hole ? ? entrance
Kilham, Maryland 7 June 1957 Nest hole ? ? entrance
IGlliam, Maryland 11 June 1957 Nest hole ? ? entrance
Kilham, N. Hampshire 5 May 1966 Nest hole ? Display by female. entrance
Kilham, N. Hampshire 20 June 1966 Nest hole By Display by female. entrance
Kilham, N. Hampshire 7 Aprill~67 Nest hole ?ES Display by captive male to a entrance chipmunk.
Teale Bird feeder, ? Displayed to chipmunk; foraging on sometimes to birds. ground for sunflower seeds.
5

Table 3. Induced Displays at Nuthatch Nest Site
Eg. aposematic display, overt threat, wing spots evident. Ey, passive display of conspicuous spotted pattern. Nest site includes area within 2 m of entrance, on tree trunk. Pose is vertical. suspended unless otherwise noted. A futl display is characterized by fully extended wings, wing spots evident, and the body weaves back and forth.
Stimulus
Red Squirrel
Flying squirrel
Gray squirrel
leas! chipmunk
Date, 1980
21 May. 5:4S p.m.
21 May, 9:30 a.m.
22 May. 6:00p.m.
23 May, 9:30 p.m.
Place
Nest site
Nest site
3 m above nest site
1.Sm below hole
3 m above hole
Nest hole entrance
Nest entrance
6
Pose
Full 3 Eg 1 Ey
Partial 8 ES
Full 1 Ey t Eg
Full 2 Eg
Partial 3 Eg
Partial 2 ?Eg
Full Ey
FullEg
Full By
Partial Ey
Full Eg
Eg Eg
Remarks
Male, actively moved about tree trunk displaying intermittently. After 10 minutes, displays ceased, calling began.
Male advanced toward flying squirrel about 1 m below hole.
Female remained at entrance.
Male, turning right.
Female moved from entrance to better view squirrel. In part, the female's display seemed to stimulate the male to display fully _ Finally male began to caU.
Male displayed high above hole.
Female made no full display. briefly made a partial display and entered hole.
Male ran behind tree to display again. then returned to original place. Advanced to 0.3 m. from chipmunk. flew to branches. began to call.

Table 3 (Cont.). Induced Displays at Nuthatch Nest Site
Stimulus Date, 1980
Least chipmunk 23 May. 9:30p.m.
Long·tailed weasel :24 May, (hoisted often) 7:30 p.m.
White-footed mouse 24 May, 10:00 p.m.
Leng-tailed weasel 2S May, 9:30 a.m.
White-footed mouse 25 May, 10:00 a.m.
Fox snake (hoisted 25 May, repeatedly) 10:45 a.m.
8-inch stick 25 May, ll:lSa.m.
Place
Branches
Branches
Trunk, near ground
HOrizontal branch
Nest site
Trunk near mouse
Nest hole entrance
Nest site
Tree trunk 1 m above ground
Nest site
Branch
Tree trunk
7
Pose
Horizontal, FuJI Eg
Partial?
Full Eg
Full Eg
Full Eg
Full Eg
Full Ey
Full Eg
Partial?
Full Eg
Full Eg
Partial? FuU Eg
No displays
Remarks
Female did not display,
Male flicked one wing, did not fully display. Calle d for 10 minutes. Female did not display, went into hole.
Male ran down tree 1 S nun near the ground.
Male ran up tree, onto horizontal branch. No sound.
Male displayed, ran down tree 0.3 m from meuse.
Displayed 10 minutes. No sound.
Female bill-sweeping, entered hole without displaying. Re·emerged, after 10 minutes, one brief display. No. sound.
Male called intermittently.
Male ran down tree to 1 m above ground, then ran alongside in partial display up the tree.
No calls.
Male fmally displayed.
Male ran down to stick, examined It intently, rapid weaving back and forth. No calls.

a display from the parents or attempt to prey upon the young birds. Chipmunks have preyed on some nesting birds. such as Bluebirds and Tree Swallows (Miller, 1968; Genevan, 1911), but such observations were reported as "unusuar'. Both nuthatches and chickadees successfully defended their nests against chipmunks in my study.
Another factor not previously considered for nest-site distraction displays is the struggle for occupancy of the tree cavity. Such encounters might be expected between hole·nesting parids, sittids, and squirrels. In my study both chickadees and nuthatches displayed regularly and with full intensity (Fig. 3) to the small White-footed Mouse (Table 3; Long. 1981). A mouse conceivably might prey on nestlings on rare occasions. These birds and the mouse likely would compete for free cavities as homes.
In the fan of 1979, and again in 1980, the nuthatch's nest was utilized by a family of Red Squirrels. After their departure, prior to both winters, the cavity was used by Gray Squirrels. Once it was investigated in winter by a pair of nuthatches (January, 1981). This same cavity has been used by nuthatches to rear young on a regular basis in spring. Therefore, occupancy is temporal in this case, and by following this schedule both kinds of squirrels and the nuthatch as well utilized the cavity regularly. In 1981, Gray Squirrels maintained possession of the dray past Easter when it was visited by a pair of nuthatches. No nest· ing was observed and the dray was abandoned; the nuthatches nested elsewhere. Nest-site displays probably aid the nuthatch to maintain possession of the nest hole.
Hinde (1952) reported aggressive displays ofseveral sorts made in winterin struggles fOJ hr,.e occupancy between Great Tits and other hole-nesting birds, especially Great Spotted Woodpeckers. These displays resembled some kln~s of nest-site distraction displays seen in pands and sittids. Especially remarkable was a single display of a Great Tit in January to a Gray Squirrel in a dray repeated several times. Such a display was never seen again, even when parent tits and young at the n~st-site were approached closely by a squirrel. This display was remarkably similar to observed distraction ~isplays in both Black·capped Chickadees and nuthatches, and "all the ingredients" were there (squirrel, cavity, bird) except that the breeding season had hardly commenced. (However, some pairing does take place In Great Tits in January., Probably Hinde ob· served a tit ..squirrel occupancy conflict and not an out-of·seasQn nest·site display.
The origin ofnest-sUe dlsplays-From a variety of aggressive threat displays described by Hinde (1952) and others, one may compare the nest-site displays to determine, if pOSSible, which primitive display might have been nest-site distraction.
One might first hypothesize that the reproductive distrac~ion displays are derived from win ter struggles for hole occupancy. However, the tatter have been reported only in the Great Tit (Hinde, 1952). One display observed by Hinde (on two successive nights, to a woodpecker) was a "head up" display which was usually an intraspecific display. It is used in reproductive fish-ting in ki:nds of tits having conspicuous black marking, present on their throats and breasts.
Other winter occupancy displays by Great Tits resemble~ the head-forward htterspeciflc threat. This interesting interspecific aggressive display of the Great Tit could be an ancestral pattern readny
modified in the Paridae to serve as either intraspecific or nest-site distraction displays, and serving iri Great Tits to threaten competitors for roosting holes in winter. Th~ head-forward display (Fig_ 1) is usually expressed when another bird approaches too closely. Common in feeding encoun ters and used against different ~pedes of birds, the display Is simUar enough to nest·slte displays to be considered as possibly their forerunnel'. Both in Great Titsand Chickadees, birds bringing food to young have eaten the Insects while making distraction displays (Hinde, 1952; Long. 1982). A defense agflinst supplanting attacks has been reported in several parids. Nothing much like it has been observed in chicl\adees. Brewer (1961) mentioned that the variety of aggressive displays seen in Great Tits "are absent or difficult to detect" in chickadees and never are very specific. A threatening nuthatch I observed in summer (July 11) was foraging on a tree trunk and was the object of a supplanting attack. It moved its head left and right three to four times and flared its tail (showing the spots), displaying not to a squirrel but to another nuthatch.
Lohrl (1967) described a somewhat simBar threat display at feeders by the Kleiber (Sittll europaea). It differed somewhat from its "tail up" intraspecific territorial display, but in both kinds of display the wings spread. the tail flared, and black and white spots in the tail exposed. The Cor~ican Nuthatch (Sitta whiteheadl) fluffed its feathers out and spread its wings to threaten various parids near its nest hole, and on one occasion actually attacked, pushing with its head, a Spotted Woodpecker that perched near its nest-site.
8

Such displays would be inappropriate against raptors, snakes, and even man, but it might have proven effective when tried out against squirrels and other small mammals. Since both head-forward and nest-site distraction displays of either parids or sittids are threats often accompanied by pecking the opponent's face or body, then other positlve advertisemen ts of the displ ay (weaving movemen ts, ex tension of the wing, evolution of spots) would be selected for. In comparing the Paridae and Sittidae there Is more reason to choose the supplanting defense while foraging as ancestral to nest-site displays than any intraspecific territorial display at this thne. However, territorial aggression is a breeding phenomenon appropriate for the breeding season. Neither type of aggression seems too dissimilar to be adapted to distraction displays, and both kinds have been seen in strife for winter holes.
Overt behavior functional in other kinds of fighting, when accompanied by a bold threat, might gradu· ally evolve to become a ritualized, conspicuous nest· site display, and might encourage the evolution of bold patterns of markings and colors. Overall agressive postures used in several sitUations probably undergo con· Unuous and variable selection. A genome favoring aggressive protection of a food site, Ii winter roost, the nest-site, or the mate, or all of these together may reasonably account for the evolution of an unusual but effective response at the nest-site in time of predation. The genome, long established in the families, will incorporate mutations for the positive advertisement of the aggressive behavior, the manifestation of which may be somewhat variable at the nest-site.
The heads-up display (Fig. 4) described by Hinde (1952) in the Great and Coal Tits (Parus ater) may be disregarded as the ancestral pattern giving rise to nest-site distraction displays. The behavior is correlated with the presence on the breast of bold black markings which are in this display conspicuously advertised. Furthermore, the display is usually intraspecific.
The Acadian Chickadee on Grand Manon Island (Parus hudsonicUlI) lures the predator (Red Squirrel) with a somewhat similar display. The bird raised upright on its perch and slowly fell over backwards, clinging upside down, "wings fluttering helplessly". Alighting on another perch, the bird fell over sideways (unfortunately, only one report, Pettingill, 1937). Fleeing and threat seem to be in close balance.
Another distraction display mentioned by Hinde (1952) in Blue Tits, and mentioned as occurring in "glimpses" in Great Tits, is an Impeded "moth-like" flight near the disturbed nest, the parents "often gliding a foot or two with slightly raised, not fully spread wings". One bird "perched on a branch about three feet away, rolled over on one side and held one wing up" for two or three seconds. There is the resemblance to "feigning injury" in this display and in that of P. hudsonicul mentioned above. Squeaking by the young evoked this display, in Ute presence of man (a much larger predator than any squirrel).
The evolution of advertisement color patterns, accompanied by threats arid overt aggression all incor· porated in a ritualized display is evident in most nest-~ite distraction displays as well as in the heads-up posture. For example, the weaving of the head right and left is related to white auricular patches (Whitebreasted Nuthatch, Black-capped Chickadee, head-forward display of Great Tit). The flaring of the taU is enhanced by white feathers (nuthatch, Black-capped Chickadee). The active presentation of the wings is reo lated to the presence of white egg-like underwing spots in the nuthatch. Such advanced patterns of coloration and elaborate displays are probably not important in the genesis of parid and sittld distraction displays. More likely the best source of distraction display in this family is a simple and pervasive form of aggressive behavior such as the head· forward display and defense during supplan ting attacks. .
In discussing the 'differences between and the simlarities of reproductive and other fonns of fighting in Paddae, Htnde (1952: 157) stated, .. Jt is conceivable that all the motor patterns of the different forms of figh ting were primitively the same ... different types offlghting were thus evolved to suit the differen t situations in which they were evolved." Long (in press) discussed how the general "scolding" of the mother Red Squirrel (a hole.nestingmammal) is in.tensifled when her nest is endangered. and becomes an intense bi ...·ocal, ventriloquistic call (apparently mimicking in part the squeaking of young squirrels). This call is used in a display involving threat and fleeing performed to lure the predator away from the nest. In other situations, in other season, the caU is merely "scolding". .
As Hinde (1952) mentioned, there are some fundamental differences between reproductive fighting and fighting for food or roosting holes. Reproductive fighting is "evoked by differen t situations", dependent on a different "internal state", and, one might add, often is a characteristic of breeding males. He pointed out some relations between these forms of aggression, namely resemblance in the basic patterns involving
9

attack and fleeing. stating that "the basic patterns of attacking and fleeing in 'predator fighting' are similar to those in intraspecific fighting". It is a hypothesis presented here that distraction displays in padds and sittids are aggressive in origin, and that advertisement distraction displays were derived from the threat displays expressed usually while fighting with other birds for food. The distraction displays became generally synchroriized with breeding behavior so as to be expressed most fully in the proximity 9f the nest (eggs or young), and in the presence of a potential nest robber. Luring or injury simulation seen rarely in Blue Tits and probably in the Boreal Chickadee (Paros hudsonicus) probably results from a ritualized fleeing combined with a more primitive aggressive display, formerly an interspecific or intraspecific threat.
In fact, the conflict of threat and flight seems to evoke in some species highly variable responses, displacement activities, and seemingly irrelevant actions (see Armstrong, 1949).
In the evolution of luring, an advertisement display initially creates a successful distraction near the nest: eventually the ritualized behavior involves fleeing as wen as threat. These closely balanced factors are important also in the evolution of the head· forward and other displays of several parids (Hinde, 1952). How the balance can readily be adapted to luring (injury simulation) is apparent in my pursuit of a displaying parent (Parus atricapilhls) while carrying a snake held aloft (Long. 1982). The bird retreated several times, displaying at each new perch. However, when retreat of the parent was not occasioned by my intervention, in displays toward smaller predators. there was only threat and attack, and never any fleeing. The evolution ofluring requires that if retreat is necessary. then the distraction or advertisement display mus: ;'e performed often or continuously along th~ way. and the stimulus for the display is, if the display is effec~ive. following close behind. The distance from the nest·site wUllower the threshold for making the display', as the proximity to the nest seems to raise it (Skutch, 1976; see Tables 2. 3).
Movements that shnulate or mimic injureti animals eventually would probably be strongly seleCted for encounters with fairly large or dangerous predators, the evolution of "injury feigning" (i.e., injury simula· tion) evolving gradually and perhaps following an obligatory tendency (with survival increasingly enhanced) as a natural consequence. The ritualized threats observed within Parids and Sittids are probably more likely hornologQuS than are various elaborate display patterns ofluring which function analogously and may have evolved independently in birds.
10

LITERAiURE CITED
Armstrong, E. A. 1949. Diversionary display. Ibis, 91: 88·97, 179·188.
Brewer, R. 1961. Comparative notes on the life history of the Carolina Chickadee. Wilson Bull., 73: 348·373.
Dixon, K. t. 1949. Behavior of the plain titmouse. Condor, 51: 110-136.
Genevan, M. E. 1971. Chipmunk predation on bank swallows. Wilson Bull., 83: 102.
Grimes, S. A. 1936. "Injury feigning" by birds. Auk, 53: 478480.
Hatt, R. T. 1929. The red sqUirrel ... Roosevelt Wildlife Annals, 2(1): 57-59.
Hinde, R. A. 1952. The behavior of the great tit (Paros major) and some other related species. Behaviour, Suppl. II, Leiden, 201 p.
IGlham, L. 1957. Reproductive behavior of the white-breasted nuthatches. Auk, 85: 477492.
Kluijver, H. N. 1951. The population ecology of the great tit, Parus m. major L. Ardea, 39: 1·135.
Lohrl. H. 1967. Die Kleiber Europas ... A. Ziemsen Verlag, Wittenberg Lutherstadt, 120 p.
Long, C. A. 1982. Comparison of the nest·site distraction displays of hlack-capped chickadee and whitebreasted nuthatch. Wilson Bul1., in press.
Long, C. A. 1982. Nest-site distraction displays by birds with egg.llke spots in the wings. Jack·pine Warbler.
Miller, W. 1968. Predation of bluebirds by an Eastern chipmunk. Blue Jay 26(3): 145.
Odum, E. P. 1941·1942. Annual cycle of the black-capped chickadee. Auk, 58: 314-33, 518·35, also 59: 499-531.
Pettingill, E. R. 1937. Grant Manon's Acadian Chickadee. Bird·lore, 39: 277-282.
Skutch, A. F. 1976. Parent birds and their young. Unlv. of Texas Press, Austin and London, 503 p.
Stoner, D. 1943. Defensive behavior ofwhlte breasted nuthatch. Auk, 60: 95·96.
Teale, E. W. 1974. Windows on wildlife. Audubon, 76(6): 50-55.
11

.......--- .,~------......, ....._--,- ._~"~6jJ
Fig. 1 Head·forwarddisplay of Great Tit used against birds that approach too closely. Tail mayor may not be flared. After Hinde, 1952.
10mm
Fig. 2 White underwing egg·like spot of White-breasted Nuthatch. After Long, 1982.
12

fig. 3 Nest·slte distraction display of Black~apped Chickadee to White·footed Mouse. After Long, 1982.
Fig. 4 Head-up threat display of Great Tit. used in reproductive fighting. conspicuously revealing black markings on throat and breast. After Hinde, 1952.
13

-------

RARE GIGANTIC TOADS, BUFO AMERfCANUS. FROM LAKE MICHIGAN ISLES
Charles A. Long
One of the rarest kinds of amphibians in the State of Wisconsin, if not the rarest of aU. is the gigantic kind of toad found on Rock Island, Wisconsin (beyond Washington Island, off the Door Peninsula) and first mentioned by Long and Long, 1976. Gigantic toads Occur also at some other localities in Lake MichIgan. This /onn or variety of gigantic toad, named alan! here, is of great biological interest, but its rarity limits investigations on numerous problems and questions. A number of these problems will be discussed. State legal protection by the Wisconsin Department of Natural Resources for this form of toad is suggested.
History and Distributlon-Hatt et a1. .(1948) reported gigantic toads from South Fox Island in the Beaver Islands of eastern Lake Michigan. I have examined 13 gravid females preserved from there. In biological surveys made by my sons and myself (Long and Long, 1976; Long et al., 1978) gigantic toads were reported from Rock and tittle Summer islands and the Garden Peninsula.
The gigantic toads occur from a fairly wide area, but the actual areas of these isles and peninsula are extremely small. The range Is patchy (discontinuous) in that the populations from South Fox Island Is about 80 Ian east of the populations of Green Bay. Giapt toads were not found on some isles of Green Bay. Although some toads were regarded as large on Garde~ Island (phillip, Ologa and Drew. 1965) there seems no evidence that the a1ani toads are found there or On Beaver, Trout, Whiskey and Squaw islands (Hatt et aI., 1948; Ologs. and Phillips, 1964) or on North and South Manitou islands (Hatt et aI., 1948; Scharf, 1973). Hatt reported toads from both North and South Fox islands. but discussed giants only from South Fox Island. Thus, on the 14 Beaver Island archipelago the alani toads are known on only one. The giants, whether having either slight or significant genetic basta, may have evolved polyphyletically from toads of simUar genomes on scattered isles because the envirofllllents are so similar. If the alani toads are monophyletic, rafting might explain their disjunct and pat~hy insular distribution. However. the evolution or rafting of these toads and their establishment on Lake Michigan isles occurred In tess than 8,000 to 10,000 years because the area populated today was fonnedy covered by ice of the Wisconsin glaciation (Hough, 1958).
Size-The gigantic toads observed are all femall'lS which, of course, exceed males in size in Bulo americanus. These females seem to illustrate insular gigantism, and are Significantly larger (20-30 percent. larger in six measurements) than females from Centr~ Wisconsin. Large size is somewhat related to age and on islands very old age may be possible to attain because of few predators (Cadquist, 1974). However, there is some observed predation of toads on Rock Islllnd, and large captive toads from the Mainland fed intensively, over a period of years, have gained consititerable weight but have not approached the size of the insular giants.
Three of the largest giants measured in the field, exceeding S inches (125 mm) in snout-vent length, escaped and were not preserved. Tom Jessen conected the largest males and females of nearly 100 pairs in a shallow breeding pond on Rock Island, and five of the largest females varied from five to perhaps SYl inches. Five males averaged approximately three inches. The means and observed ranges in mm of 13 gravid females preserved in alcohol from South Fox Island (Michigan State University Vert. Museum) and of one female and male from Rock Island are, respectively as follows: snout-vent length, 98.S (89-104), 107, 78.5; length hind foot from heel, 67.4 (60·72),70,59; body width, 65.0 (57·73).68.5,34; length
Fig. 1. Gigantic toad from Rock Island, Wisconsin. Drawn from actual female specimen by K. VoUmer, Natural size.
16

parotoid gland, 19.06 (17.4-21.8). 19, 16; width mandible, 37.3(34.540.3), 39.2, 31.0; inter-narial \\oidth, 6.45 (5.9-7.7), 5.9. --,
Giant toads, even the smallest observed, had wide mouths and large parotoid protruberances. The males occasionally were greenish on Rock Island, the females dark. Giants from South Fox Island showed large, scattered and discrete wartS. There is little or usually no overlap in size of the gigantic females and the largest females from Central Wisconsin.
Size alone may be a poor criterion to base a taxonomic name upon, but often it has been done. I have been advised both, to name the toads, and to not name them. The gigantic size may have a genetical basis. It is found in several populations over a fairly wide range. The toads, therefore, seem to be a geographic race. A way to identify such a fonn of toad is to use a subspecies name. The advice to the contrary points out that toads show conservative evolution, ant\ that speciation In less than 10,000 years was very unJikely.
Guttman (1975) noted quite insignifcant genetical differentiation among toads from 11 localities in Northern Wisconsin. Nevertheless, toads from this area and across the lake in Michigan were differentiated in several isozymes from toads to the eastward, northward, and southward. The isozyme IDH·2C was found in Northern Wisconsin and Michigan, but not northward or eastward. PGM·3a and GOT-lb were found in Wisconsin populations but not elsewhere except the latter occurred as far eastward as some populations near Toronto. Unfortunately no studies yet on isozymes have been made on the giant toads, an obvious problem for future study.
Characterized only by large size, and in one sex at that, the giant toads are not described as a subspecies here. Some toads exceeded 130 mm tn a few giant specimens measured in the field (but possibly these measurements were inaccurate). A few of the largest toads throughout the ranse ofBufo tunerlcanus attain lengths of 1()()'110 mm.
Instead of using a subspecific name for the gigantic toads, they may go by the variety name alan!. A precedent for using an intraspec:ific name for an inSt\lar variety is given by Degerbol (1940). Buckner (1966:464) described a surprisingly rapid increase in size In shrews introduced onto Newfoundland. My son Alan carried out most of the work on reptiles and amphlblana in my surveys 011 the islands of Green Bay. Alani toads may represent only a deme, the phenotypes i;>f which result trom a locaUzed high frequency of genes favoring large size not seen elsewhere.
Ell Mass-Another character. consequential but obviously adaptive, is the larger size of the eg mass in giant females. Since the e8P appear normal in size they probably outnumber the ova of small females. More descendants, then, Ukely result from large egg masses. The largest toads being more abundant would be more likely to persist on small islands, and the genes for large size may be fixed when toads cycle low in abundance. Such weak natural selection and chance fixation of the combinations of such genes for large size might lead to the rapid appearance of extremely large toads even though new mutations for giant size mayor may not appear in the isolated populations. In my opinion, the large size of the egg mass seems exceptionally important in the rapid appearance of giant· toads, an origin which seems mysterious in light of the short span of evolutionary time. Such origin may be monophyletic only in the source of genes causing large size, which genes may be widespread not only on islands but on the mainland.
Impaired locomotion-If genes for large size result in the origin of large females with large numbers of progeny. then why are such females not predominant throughout the range of Hula american"s? There is one obviol.luy detrimental character of giant toads, so detrimental u to be in effect probably a lethal character of older and larger toads. ThiJ probable lethal character i. impaired mobility. That is one reason that such gigantic toads were never seen on the mainland. Hatt (1948) mentioned impaired locomotion in toads from South Pox Island. I observed that giant toads walk more than hop 00 Rock Island. Uttle Summer Island, and on the beach of the Garden Peninsula (Fairport,Michigan). On theae small islands and on the beach of this small peninsula predation may be insignificant. What we know of any comes from birds of prey, for which cryptic behavior Is more adaptive than hopping. Migration on small .is1~ to water for egg deposition is no great problem. Although incapable of leaping. these 'giant toads showed surprising strength. Apparently size and weight alone limit their mobility.
17

The giants Hatt observed were, so to speak. egg·bound, carrying eggs beyond the normal time of . mating. This might impair mobility so as to account for their discribedconditlon of helplessness. However,
toads mated in mid·May on Rock Island. which is not at aU an unusual season. Non·gravid giants observed there were impaired in their locomotion too. An assumption made here is that the large size impairs the leaping ability, not merely the egg-bound condition.
lug (1978) found no difference in leaping ability of male and female Bufo americanu$. Their linear and power regression curves were nearly identical. Furthermore, the largest toads leaped the farthest. However, lug's largest toad~, both male and female, were aU less than 80 mm in snout-vent length. None ofhls jumpers approached the size of gigantic toads of lake Michigan.
Zug listed two reasons why some frogs show a poorer jumping ability in adult females than in males. The weight of the egg masses is an obvious factor, but "the effect of landing forces on developing ova perhaps placed a selective premium on females with shorter jumps" was another hypothesis. The lake Michigan giant toads are excluded from the latter not only because the time for such selection to have become effective is short (less than 10,000 years). but their jumping when gravid seems handicllpped much beyond the point necessary to dimJnish shock to the ova. Large size in giant toads limits the ability to jump whether the toads are gravid Of not. Zug's curves for Bufo americal1us obviously do not apply to giant toads of Bufo americanus.
Toads as a group were poor leapers, In comparison to various frogs. and lug referred to toads as "earthbound." Low in mobility in general, then perhaps only on small islands where water is close at hand, and predation on toads less Significant, is development ofa gigantic size possible. In these habitats gigantic toads probably are even more fit than mainland toads. A giant female was observed to feed on larger prey items (huge ground beetles) than did the smaller males, and prey abundance is cyclic on islands. In such a situation differential niche separation of the sexes is encouraged evtl1 if not fully realized (tee Amodon, 1959; Selander, 1966; and Schoener, 1968). The number of progeny of giant toads is probably increased by their larger egg muse •. These progeny may possess ever increasing proportions of genes for large-stud females and large egg malS, and when the abundance of toads is scarce theJr frequenele. may be fixed.
ConlelV8.tion-The scarce alani toads are in peril of extinction, or at least alteration of the phenotypic character of size (which may be weakly differentiated anyhow). Found in small numbers on a few tiny islands the giant toads are vulnerable. CoHectors have selectively removed the largest specimens. In small populations such collecting tends to rapidly remove any genes from the small populations that cause large size.
The populations of gigantic toads are interestins and distinctive, occurring patchily over a range of 80 km, but in very small areas. They seem to show evolutionary processetl tending toward such phenomena as niche separation of sexes, insular speciation, impaired locomotion, and rapid post·Wisconsin evolutionary change. If these phenomena are not fully established and clearly evident, their indications, nevertheless, suggest very important evolutionary processes at work in the brief geological time theorized for them to be working. Certainly such unusual toads ought to be preserved and protected by law. Otherwise, this rare and interesting form described as Buro americanus alani may vanish forever.
18

LITERAlURE CITED
Amodon, D. 1959. The significance of sexual differences in size among birds. Prod. Amer. Phi. 1. Soc. 103;531·536.
Buckner, C. H. 1966. The role of vertebrate predators in the biological control offorest insects. Ann. Rev. Entomol., 11 :449470.
Carlquist, S. 1974. Island biology. Columbia Univ. Press, New York, 660 p.
Degerbol, M. 1940. Zoology of the Faeros, Mammalia. Copenhagen.
Guttman, S.1. 1975. Genetic variation in the genus Buto, I1.lsozymes in northern allopatrlc populations of the American toad, Buro americanus. In Isozymes, ed. by C. L. Markert, Vol. IV, Genetics and Evolution, Acad. Press.
lIatt, R. T., et al. 1948. Island life: A survey of the land vertebrates of the islands of eastern Lake Michigan. Cranbrook Inst. Sci., Bull. No. 27, 179 p.
Hough. J. L. 1958. Geology of the Great lakes, Univ. nUnois Press. Urbana.
Long. C. A. 1978. Mammals of tile islands of Green Bay. Lake Micltigan. Jack·Pine Warbler. 56:59-82.
Long. C. Alan, and C. A. tong. 1976. Some amphibians and reptiles collected on Wands In Green Bay, Lake Michigan. Jack-Pine Warbler, 54:54-58.
Long. C. Alan, J. E. Long and C. A. Long. 1978. Some amphibians and reptiles from ChambeR Island, Green Bay. Lake MJchigan. Jack·Pine Warbler, 56:46-47.
Ozoga. J. T. and C. 1. Phillips. 1964. Mammals of Beaver Island, Michigan. Michigan State University Public Museum, Bioi. Ser.• 2:307·347.
Phillips, C. J., J. T. Ozoga and L. C. Drew. 1965. The land vertebrates of Garden Island, Mighigan. Jack· Pine Warbler, 43:20-25.
Scharf, W. C. 1973. Birds and land vertebrates of South Manitou Island. Jack·Pine Warbler, 51 :3·19.
Schoener, T. W. 1968. The Anolil lizards of Bimini: Resource partitioning in a complex fauna. Ecology, 49:704-726.
Selander, R. K. 1966. Sexual dimorphism and differentia! niche utilization in birds. Condor, 68:113·151.
Zua. G. R. 1978. Anuran locomotion-Structure and Function, 2: Jumping performance of semiaquatic. terrestrial. and arboreal frop. Smithsonian Contr. Zool.. 216: 1·5I.
19