
Enzime cu nichel
Pentru mult timp nichelul a fost singurul metal 3d pentru care rolul biologic nu a putut fi stabilit definitiv. Motivele acestei scăpări au fost variate: ionii de nichel nu au absorbţii caracteristice în prezenţa liganzilor fiziologici importanţi, spectrele Mossbauer nu sunt accesibile izotopilor nichelului. Ni(I) cu configiiratia d9 şi Ni(III) cu configuraţia d7 paramagnetici, nu pot fi detectaţi prin spectroscopia RPE datorită lipsei scindării hiperfine (abundenta naturală a 61Ni cu I = 3/2 este de numai 1,25%. S-a arătat ca nichelul este adesea singurul metal component al unor enzime complexe care mai deţin drept coenzime alte substanţe anorganice. Centrii cu nichel au rămas nedetectabili mult timp datorită mascării de catreclusterii Fe-S. Aplicând metode de detective mai sensibile cum ar fi spectroscopia de absorbţie şi de emisie atomică (AAS şi AES), măsurători magnetice (SQUID) sau spectroscopia RPE asupra izotopului 61Ni îmbogăţit s-au putut stabili caracteristicile câtorva enzime ce confin Ni din plante şi microorganisme.Nichelul, în mod particular, nu este un element rar în litosfera sau hidrosfera unde se găseşte dizolvat că Ni2+ iar în organisniele vii se găseşte în cantităţi mici, greu detectabile şi după părerea specialiştilor(Wolfgang Kaim şi Brigitte Schwederski) neesenţiale,nefiind raportat niciun simptom al carentei nichelului(Ni2+ este un toxic,responsabil al “alergiei la nichel”).Totuşi chiar nichelul din otelurile inoxidabile a fost mobilizt de anumite microorganisme pentru a şi satisfice necesarul corespunzător enzimelor esenţiale.După părerea altor specialist,una din ipotezele disparitfei dinozauriJor şi a altor creaturi de la sfârşitul cretacicului ar fi "otrăvirea cu niche” care provenea din materiale rneteorice.
Târziu, în 1960, nichelul este considerat component necesar pentru creşterea unor bacterii anaerobe; în 1975 metalul a fost evidenţiat drept component al ureazelor din plante. În concordanţă cu nivelul de cunoaştere actual, nichelul este prezent În patru tipuri de enzime esenţiale în stări de oxidare diferite cu o coordinare caracteristică mediului.Ureazele conţin Ni(II) penta sau hexacoordinat de liganzi cu atomi de N sau O donori, hidrogenazele mai multor bacterii anaearobe conţin nichel în principal coordinat de liganzi cu sulf( bacteria Knallgeas , CO-dehidrogenaza utilizată în sinteza bacterială a acetil-coenzimei A). Metil coenzima M reductaza bacterid metanogene, prezintă un complex al nichelului cu inelul tetrapirolic ca grupare prostetica denumit coenzima F430. Un alt complex tetrapirolic cu o funcfie nespecificata, ar fi tuniclorinul care conţine nichel, izolat din tunicate.
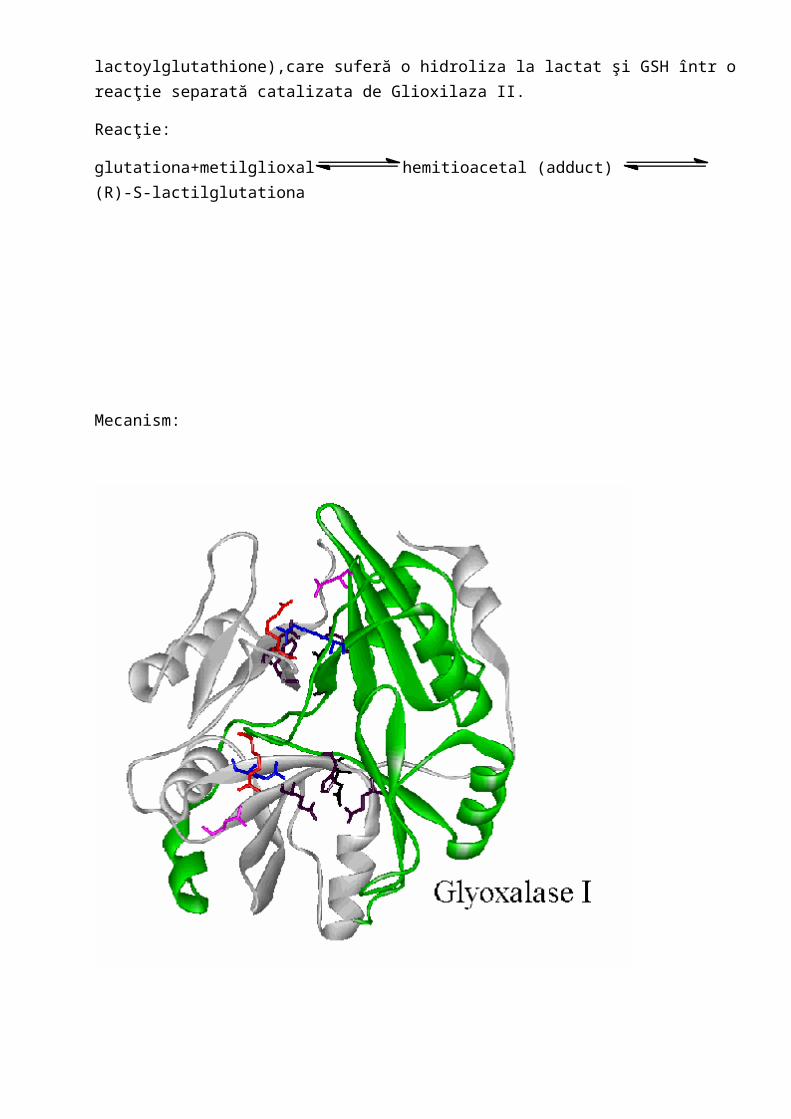
1. Glioxilaza I((EC4.4.1.5)(lactilglutationa liaza)) lactoylglutathione lyase
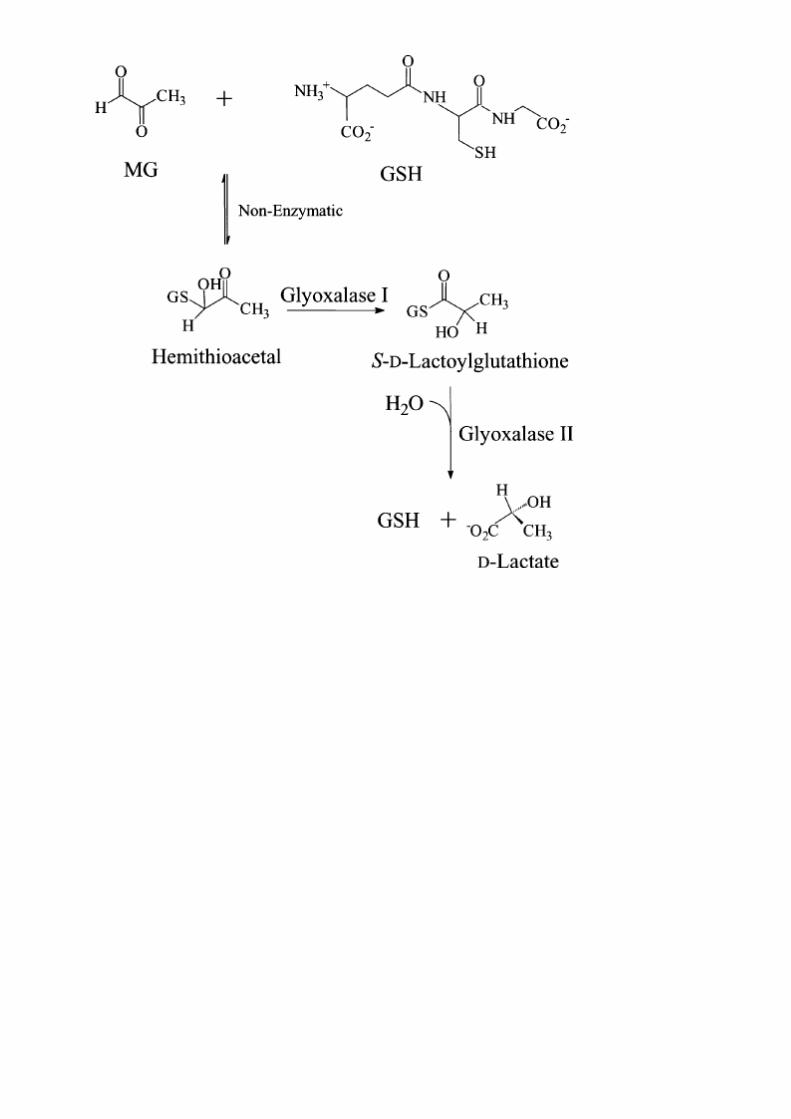
Glioxalaza I şi Glioxalaza ÎI catalizează conversia metilglioxalului,o specie toxică care formează legături covalente cu DNĂ,la lactat(lactate).Singurul ion de nichel se afla într o coordinatie octaedrica şi se comportă ca un acid Lewis.Acesta rămâne în stare de oxidare 2+ în timpul reacţiei de izomerizare,ceea ce include şi utilizarea Zn la glioxilaza I în unele
organisme,incluziv al omului.Ionul de Ni2+ se leagă hemoacetotiolic( binds the hemithioacetal adduct) între metilglioxal şi GSH,înlocuind un ligand de apă.Baza generală a catalizei duce la separarea unui proton din substrat(deprotonare),formând un intermediar coordinat enantiolat.Protonarea la C-2 iniţiază generarea de produşi S-d-lactilglutationati( promotes generation of the product S-d-lactoylglutathione),care suferă o hidroliza la lactat şi GSH într o reacţie separată catalizata de Glioxilaza II.
Reacţie:
glutationa+metilglioxal hemitioacetal (adduct) (R)-S-lactilglutationa
Mecanism:

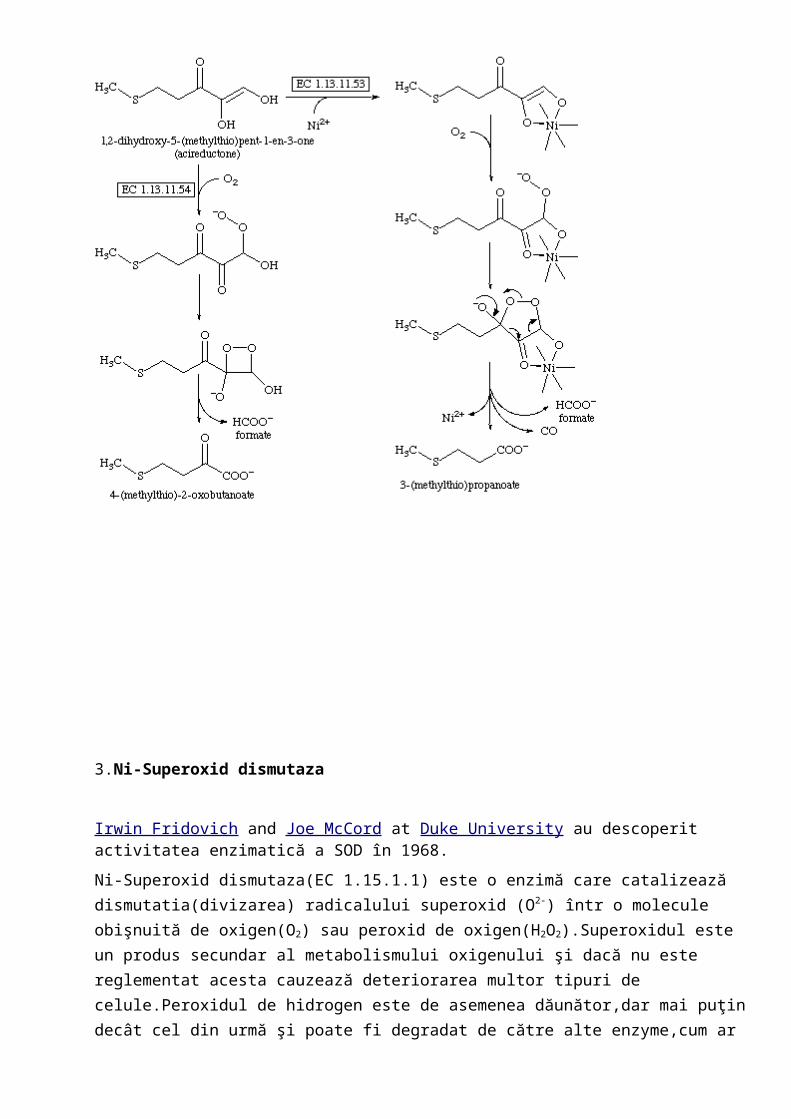
2.Acireductona deoxigenaza(EC1.13.11.53) este o metaloenzima care catalizeza reacţia chimică:
1,2-Dihidroxi-3-oxo-5-(metiltio)pent-1-en+ O2 HCOOH + metiltiopropionat + CO
Structura cristalografica cu centrul active de Ni(II):
ARD este un membru al clasei structurale a proteinelor cunoscute sub numele cupins, sau "jellyrolls." Nucleul structurii cupin este un (5-helix format din două antiparalele (i-fire). Dacă s-ar răsuci un ac de păr într-o spirală, s-ar obține topologia-pliere cupin.
Structura metaloenzimei dezvăluie o coordinatie octaedrica cu centrul active Ni(II),hexacoordinat de 3 liganzi de histidina,unul de acid aspartic şi 2 liganzi de apă.Nichelul nu suferă mudificari redox dar nici nu leagă O2,în schimb substratul acireductonei(1,2-dihidroxi-3-oxo-5-(metiltio)pent-1-en) reacţionează direct cu O2 formând speciile peroxidice (B).Ni rămâne pe tot parcurul reacţiei în starea de oxidare +2.Ni acţionează ca un acid Lewis promovând atacul intermediarului peroxola gruparea ligandului carbonil pentru a genera un intermediary cyclic(D),care se descompune ulterior în CO,acid formic şi acid carboxilic.
Mecanism:
3.Ni-Superoxid dismutaza
Irwin Fridovich and Joe McCord at Duke University au descoperit activitatea enzimatică a SOD în 1968.
Ni-Superoxid dismutaza(EC 1.15.1.1) este o enzimă care catalizează dismutatia(divizarea) radicalului superoxid (O2-) într o molecule obişnuită de oxigen(O2) sau peroxid de oxigen(H2O2).Superoxidul este un produs secundar al metabolismului oxigenului şi dacă nu este reglementat acesta cauzează deteriorarea multor tipuri de celule.Peroxidul de hidrogen este de asemenea dăunător,dar mai puţin decât cel din urmă şi poate fi degradat de către alte enzyme,cum ar fi catalaza.Astfel,SOD este un antioxidant de apărare important în aproape toate celulele vii expuse la oxigen.
Descriere:
Superoxid dismutaza (SOD) este o enzimă ce revitalizează celulele şi reduce rata distrugerii lor de către radicalii liberi. Neutralizează cei mai obişnuiţi şi poate cei mai periculoşi radicali

liberi- radicalii superoxid. Radicalul superoxid produce deficitul de fluid sinovial, important lubrifiant pentru încheieturile corpului, ceea ce duce la frecare şi în cele din urmă la inflamaţii.SOD (Superoxid dismutaza) acţionează sinergetic cu enzima catalaza, care se găseşte din abundenţă în organism. Catalaza transformă hidrogenul peroxid din subprodusele sale create de reacţii SOD.SOD ajuta la asimilarea zincului, cuprului şi manganului. Nivelul tinde să scadă odată cu vârsta, ceea ce creşte formarea de radicali liberi. Potenţialul ca tratament antiimbatranire se afla încă în studii de cercetare.Din punct de vedere chimic, există două forme din această enzimă. Forma cupru/zinc (cunoscută Cu/Zn SOD) îşi exercita calităţile de antioxidant în citoplasmă celulelor. Acesta este un fluid apos care înconjoară toate celalalte component celulare.Forma mangan (Mn SOD)este activ în mitocondrii, stucturi din celule în care se produce energia. Producerea energiei celulare duce la crearea radicalilor liberi şi forma nichel(Ni SOD).
Utilizări medicale
SOD (Superoxid dismutaza) este utilizat pentru a trata artrită, probleme de prostată, ulcere, arsuri, boli inflamatorii, dar şi intoxicaţii de la expunerea la fum şi radiaţii, şi pentru a preveni efectele secundare ale medicamentelor împotriva cancerului. Mai poate ajuta şi la reducerea ridurilor faciale, la cicatrizarea şi vindecarea rănilor şi arsurilor.SOD face ca fiecare celulă a corpului să fie mai rezistentă împotriva atacurilor radicalilor liberi şi a altor substanţe nocive. Puterea antioxidantă a SOD o depăşeşte pe cea a celor mai cunoscuţi antioxidanţi: flavonoide, carotenoide, vitamina A, C şi E, mai mult, nici chiar coenzima Q10, glutation peroxideaza şi catalaza nu ating puterea acestuia. Câteva studii legată de procesul de îmbătrânire au remarcat faptul că animalele care produc cel mai mare nivel de SOD au şi cea mai mare durată de viaţă.
Surse
SOD exista, în mod natural în: orz, broccoli, varza de Bruxelles, varza, germeni de grâu şi multe alte plante verzi.
Administrarea suplimentelor de SOD
SOD este o moleculă fragilă ce este distrusă uşor în stomac, astfel tabletele trebuie să fie comprimate filmate - adică acoperite cu o substanţă protectoare care ajuta tableta să treacă intactă prin acidul din stomac până în intestinal subţire unde poate fi absorbită la întreaga potenta.
Reacţii:
Ni3+-SOD + O2- → Ni2+-SOD + O2
Ni2+-SOD + O2- + 2H+ → Ni3+-SOD + H2O2.
Structura:
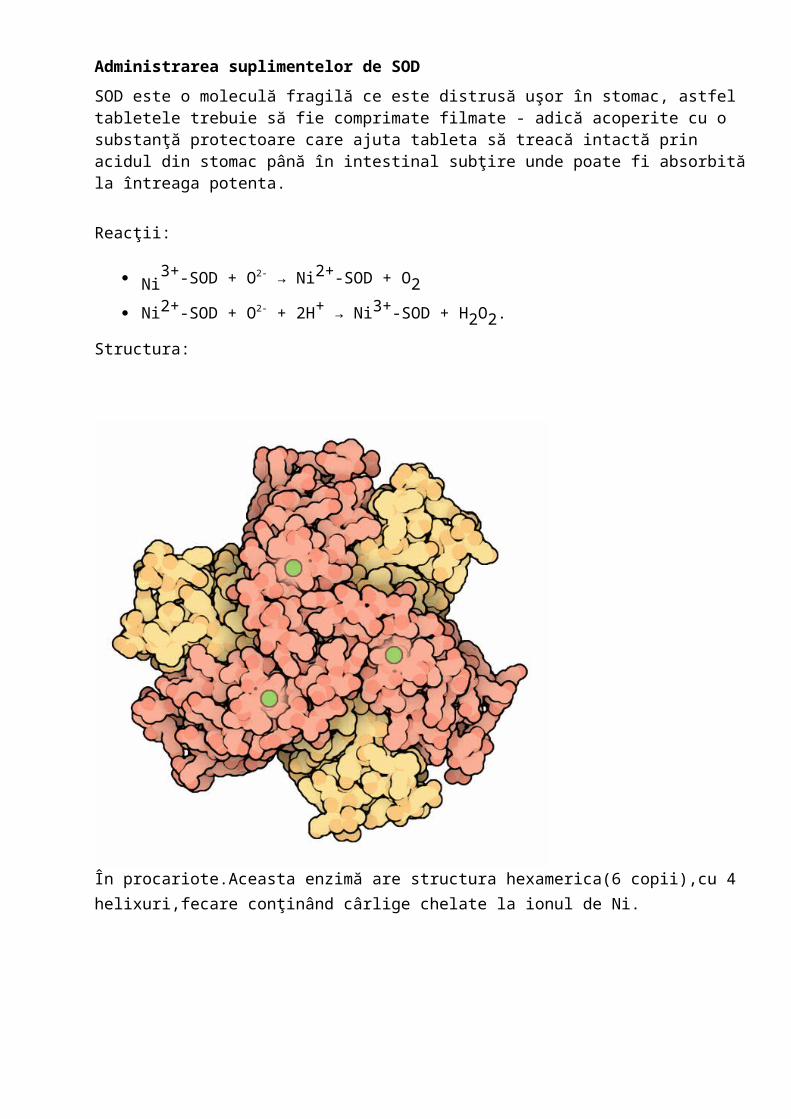
În procariote.Aceasta enzimă are structura hexamerica(6 copii),cu 4 helixuri,fecare conţinând cârlige chelate la ionul de Ni.

Acţiunea superoxid dismutazei (SOD) în organism:1. împiedica producerea agenţilor liberi nocivi 2. antioxidant natural 3. eficient în cazul bolilor cronice 4. apară organismul împotriva impactului radicalilor liberi 5. încetineşte îmbătrânirea şi apăra împotriva radiaţiilor UV 6. ajută în stările de convalescenţă şi împotriva dependenţei de droguri 7. conferă protecţie împotriva radiaţiilor solare, poluare şi pentru utilizatorii de
calculatoare 8.combate radicalii liberi şi substanţele cancerigene.
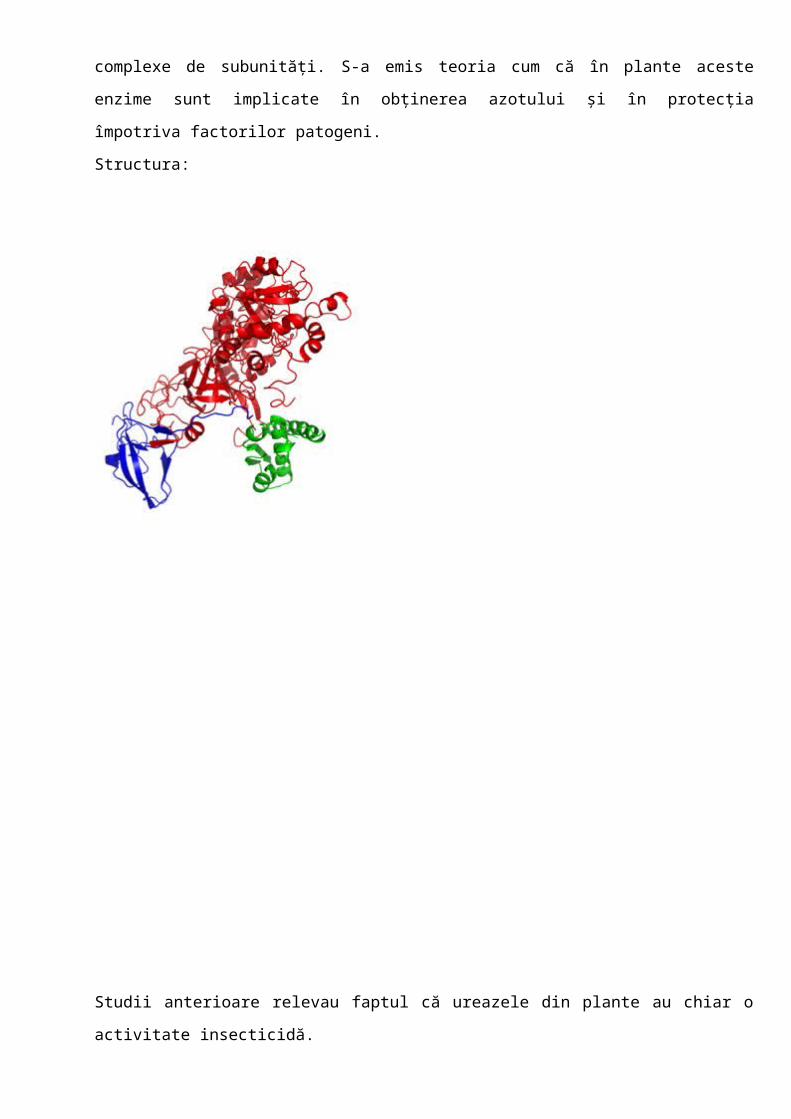
4.Ureaze
Urează (E.C.3.5.1.5.) este o metaloenzimă dependentă de Ni care catalizează hidroliza
ureei la amoniac și dioxid de carbon. Este produsă de plante, fungi și bacterii, dar nu și de
animale; ureazele prezintă omologii semnificative și mecanisme similare de catalizare, dar
diferă în structurile cuaternare. În timp ce ureazele produse de plante și fungi sunt proteine
homo-oligomerice de 90 kDa, cele produse de bacterii sunt multimere de două (ex.
Helicobacter pylori) sau trei complexe de subunități. S-a emis teoria cum că în plante aceste
enzime sunt implicate în obținerea azotului și în protecția împotriva factorilor patogeni.
Structura:
Studii anterioare relevau faptul că ureazele din plante au chiar o activitate insecticidă.
Ureazele pot fi izolate din bacterii sau plante ,cum ar fi o specie de fasole şi au o
istorie interesantă.Urează din Canavalia ensiformis, contrar opiniei lui Richard Willstatter a fost
prima enzimă obţinută în stare cristalină pură. Totuşi dear 50 de ani mai târziu a fost
determinat conţinutul de nichel al acestei enzime.Ureazele catalizează degradarea ureei la
dioxid de carbon şi amoniac. Ureea este o moleculă foarte stabilă care în mod normal
hidrolizează foarte încet obţinându-se acid izocianic şi amoniac. Durata reacţiei necatalizate
este de 3.6 ani la 38°C.
H2N-CO-NH2 + H20 [H2N-COO" + NH4+]] 2 NH3 + CO2
H2N-CO-NH2 + H20 -NH3 +H20 + H-N=C=0
Activitatea catalitică a enzimei determina o creştere a vitezei de reacţie de aproape 1014 ori asemenea accelerare poate fi explicate doar prin schimbarea mecanismului de reacţie . În timp ce reacţia necatalizata implica o eliminare directă a moleculei de amoniac, enzima probabil catalizează o reacţie hidrolitică cu ionul carbamat ca prim intermediar. Realizarea unei legături Ni-substrat facilitează ultima reacţie amecanismului. O dovadă în plus pentru legarea metalului de substrat este faptul că activitatea enzimei este inhibata prin Iegarea nichelului de un derivat fosfat.
Formarea legăturii Ni-substrat:
Secvenţele de aminoacizi ale diferitelor ureaze sunt cunoscute dar informaţiile privind structuraşi activitatea centrilor de nichel sunt puţine. Haloenzima ureazei din fasole este formată din şase unităţi identice, fiecare subunitate (91kDa) conţinând doi ioni de nichel în poziţii apropiate şi aparent diferiţi.

Măsurătorile EXAFS au evidenţiat prezenta azotului şi oxigenului în prima sferă de coordinare şi au sugerat numărul de coordinare cinci sau/şi şase, în concordanţă cu masurato rile magnetice spectroscopia de absorbfie. Măsurătorile de succeptibilitate magnetică au evdentiat existenţa unui echilibru Între stările spin-inalt (S=1) şi spin- scăzut (S=0), ceea ce ar fi tipic pentru coordinarea 5 sau 6 de octaedru deformat a Ni(ll) datorită diferenţelor mici şi variabile de energie dintre orbitalii dz2 şi dx2-y2. Mecanismul propus implica un atac electrofil al unuiadintre centrii nichelului la atomul de oxigen al grupării carbonil, şi un atac nucleofil al specie nichel hidroxo asupra atomului de carbon din gruparea carbonil (mecanism push-pull). Ipoteze similare există şi pentru mecanismul de funcţionare a enzimelor hidrolitice ce conţin zinc, unde protonii funcţionează adesea ca specii electrofile,. Enzimele hidrolitice cu zinc necesita astfel numai un centru metallic care să furnizeze hidroxidul nucleofil prin activarea intr- una din etape a unei molecule de apă.
Nu este evident de ce centrii activi hidrolitici de Ni sunt prezenţi doar în ureaze. Prin substituţia Zn cu Ni în carboxipeptidaza A (CPA) nu sc modifica semnificativ activitatea enzimei.
Schema mecanismului de degradare a ureei sub acţiunea catalitică a ureazelor ce conţin nichel(mecanism push-pull)
Poate că ceriţele mai puţin stringente privind selectivitatea numărul mare de heteroatomi coordinaţi din ureaze favorizează Ni(II) faţă de Zn(II) care prezintă compuşi cu numere de coordinare mici. În concordanţă cu acestea avem legarea ureei de gruparea guanidin din arginina favorizată de prezenţa catalitica a hidrolazelor dinucleare cu Mn(II).
5.Hidrogenaze
Hidrogenazele sunt enzime care catalizează oxidarea reversibilă a hidrogenului molecular cu implicarea în proces a doi electroni:
H2 2H+ (aq) + 2e-
Această reacţie joacă un rol important în cursul fixării dinitrogenului, în fosforilarea microbiala sau fermentaţia substanţelor biologice la

metan. Microorganismele anaerobe şi câteva aerobe conţin hidrogenaze; hidrogenul poate servi ca sursa de energie în locul NADH-ului sau poate fi un produs final al proceselor de reducere. Majoritatea dar nu toate hidrogenazele conţin clusteri Fe-S. Enzimele "numai-Fe" ce conţin unităţi [4Fe-4S] în special "clusterii-H" catalitici (6Fe) nu cer alte metale sau coenzime. În funcţie de prezenţa altor grupări prostetice, de exemplu flavinele sau elemente "anorganice" neuzuale, hidrogenazele sunt împărţite în: Ni/Fe-hidrogenaze, cea mai cunoscută forma şi Ni/Fe/Se-hidrogenaze. Ni/Fe-hidrogenazele conţin un atom de Ni în plus înafara clusterului Fe-S;
Ni/Fe/Se-enzimele prezintaNi şi Se (Ni este coordinat de selenocisteina) în raport echimolecular cu cel al clusterilor Fe-S. Nitrogenazele au şi o activitate diferită de a hidrogenazelor mai ales că în substratul primar de N2 lipseşte. Catalizaiea reacţiei este necesară din cauza transferului simultan al celor doi electroni.
Reducerea unui proton, cu transferul unui singur electronsi obţinerea unui atom de hidrogen ar necesita mai multe potenţiale negative, mai mici ca valoare decât -2V. Aceste condiţii nu se întâlnesc în mediul fiziologic.Componentele anorganice ale hidrogenazelor îndeplinesc rolul de rezervor de eiectroni şi de centri catalitici.
Dispunerea centrului proteic ce conţine hidrogenaze în ciano bacteria Anacystis Nidularis.
La un număr mare de bacterii fotosintetic active şi alge, s-a observant că activitatea unor hidrogenaze depinde de un sens preferat al reacţiei,astfel putem distinge hidrogenaza unidirecţională (H2 -»2H+)
dintre enzimele bidirecţionale reversibile. Majoritatea hidrogenazelor sunt de mărime medie (40-100 kDa) şi de regulă au funcţiuni revesibile. Cu toate acestea, potenţialele de transfer electronic dintre clusterii Fe/S şi componentele flavinice sunt adesea apropiate ca valoare de potenţialele proceselor câtă izate preferenţial într-o singură direcţie, valoare inferioară celei ce caracterizează mediile fiziologice. Obţinerea hdrogenului molecular are loc doar în condiţii strict anaerobe în timp ce oxidarea hidrogenului are loc atât În condiţii aerobe cât şi anaerobe.
Câteva organisme printre care şi Desulfovibrio gigas, pe lângă hidrogenazele dizolvate în citoplasmă, mai conţin altele la marginea membranei care sunt implicate în reducerea CO2, NO3 ,O2 sau ŞO4
2- În condiţii anaerobe.În acest caz, forma solubila catalizează reducera NAD+ cu H2 iar electronii rezultaţi din
procesul de oxidare la nivelulul membranei a hidrogenului molecular sunt captaţi de catenă respiratorie şi contribuie la formarea fosfaţilor cu legături macroergice.

Schema generală de degradare a hidrogenului molecular sub acţiunea catalitică a hidrogenazelor
Hidrogenazele catalizează degradarea hidrogenului molecular liber obţinut în urma reacţiilor de reducere care poate fi periculos pentru organisme, iar reacţia catalizată de hidrogenaza din Knallgas apara formele de viaţă anaerobe procesând resturile de oxigen prin dezactivare, de exemplu în timpul fixării N2.
Hidrogenazele au fost studiate atent datorită posibilei lor implicări Indegradarea microbiala controlată a materiei organice cu obţinere de CH4 sau H2 pentru energie. Caracterizarea lor structuralaestedificila, hidrogenazele fiind sensibile şi labile. Din analiza câtorva Ni/Fe-hidrogenaze a rezultat o analogie a aminoacizilor, în special în ceea ce priveşte gruparea cisteinica. Enzimele ce conţin nichel sunt mai bine caracterizate, ele fiind mai puţin sensibile decât hidrogenazele numai-Fe care au o activitate mai mare.
Informaţii privind structura şi activitatea catalitică a centrului activ de nichel din Ni/Fe hidrogenaze.
Spectrele RPE obţinute pentru oxidarea hidrogenazelor de la marginea membranei pot fi interpnetate doar prin presupunerea prezenţei Ni(III) paramagnetic (spin scăzut d7 ,S = 1/2). Prezenţa nichelului paramagnetic a fost confirmată şi de experimentele cu hidrogenaze ce conţin 61Ni marcat, când în spectroscopia RPE s-au obţinut scindări ale spinului nuclear la I = 3/2.Geometria centrilor activi de nichel a fost investigate prin XANES §i EXAFS. Conform datelor obţinute nichelul este înconjurat de 2 ± 1 liganzi cu sulf iar lungimea medie a legăturilor Ni-S este de 223 pm 3 ± 1 din atomii de coordinare pot fi oxigen sau azot Distanţa dintre nichel şi cel mai apropiat ion de fier a fost estimată de unii specialişti la 430 pm. Coordinarea şi geometria pentra stările de oxidare invocate: Ni(I), Ni(II), Ni(III) cu configuraţiile d9, d8 respectiv d7 nu sunt determinate fără ambiguităţi. Pentru Ni/Fe hidrogenaze se presupune existenta în sfera de coordinare a cinci sau şase liganzi dispuşi după o geometric de dipiramida trigonala sau de octaedru deformat.Mecanismul catalizei Ni/Fe-hidrogenazei precum şi natura speciilor participante şi stările lor de oxidare sunt în curs de investigare.Apariţia sau dispariţia semnalelor RPE indica modificarca stării de oxidare a nichelului în timpul activităţii reductive a catalizatorului. Studiul spectroscopic ENDOR a unei forme active a enzimei arata interacţiunea 1H/ spin - electron ce implică probabil şi un proton din gruparea (beta - CH2 a cisteinei şi unul din substratul H2. Rezultatul nu este cert întrucât nu s-a dovedit că nichelul poate coordina H2 după descoperirea hidrogenazelor care nu conţin Ni. S-a verificat accesibilitatea gazelor inerte şi de reacţie la centrul activ al hidrogenazei din Desulfovihrio gigas efectuad analize cu raze X asupra cristalelor unor compuşi model în atmosfeara de Xe la presume ridicată. Astfel de date obţinute pentru cristale ce conţin nichel în diferite stări de oxidare (|), (II),(III) permit interpretări ale mecanismelor de reacţie.
Specroscopia RPE pentru Ni(II) din formă oxidata (inactivă) a enzimei prezintă alte semnale atribuite Ni(I) sau N(III). Absenţa semnalelor ar putea fi atribuită Ni(II) cu număr par de electroni to subnivelul "d" sau cuplării puternic antifearomagnetice dintre un atom de nichel cu S=l/2 şi forma redusă a clusteului [4Fe-4S).
0 compoziţie tipică în componente anorganice a fost găsită În hidrogenaza cu masa 88 kDa din bacteria Desulfovibrio gigas. Aceasta este o proteină periplasmatica hetrodimerica

formats din două subunităţi de 28 kDa şi 60 kDa. Pe lângă ionul de nichel, hidrogenaza mai conţine un cluster stabil [3Fe-4S] şi două cluster [4Fe-4S] mai puţin întâlnite, care participă la transferul celor doi electroni.
Structura şi geometria clusterlui [3Fe-4S] din bacteria Desulfovibrio gigas Pentru forma activată (redusă) a ezimei se indica prezenţa Ni(Îl) a unui cluster [3Fe-4S]
redus. Este unanim acceptată de specialişti existenta a 12 ± 1 atomi de fier intro moleculă de enzimă, deci apare posibilitatea de a avea un atom de Fe suplimentar, care nu aparţine nici unui cluster Fe-S şi care poate fi al doilea centru activ metalic al enzimei. Existenţa lui creşte posibilitatea legării hidrogenului molecular prin realizarea unui complex metalic cu hidrogen în punte, în care Fe este înconjurat octaedric sau plan pătratic iar nichelul piramidal-patratic.
Prezenţa H2 face ca semnalul RPE să se piardă (poate datorită formării Ni(II)-H2), urmată de apariţia altei specii RPE active cu formula Ni(l))-H-1 sau formă de rezonanţă Ni(I)-H+1. Pe baza acestor studii, fără a cunoaşte exact mediul de reacţie în care are loc coordinarea, nu puten face distincţia netă între Ni(I) şi Ni(III). Studiile asupra fermelor inactive de Ni(III) au demonstrat că potenţialele de reducere Ni(III)/Ni(II) sunt surprinzător de mici. Potenţiale de reducere mici s-au obţinut pentrucomplecşii model NiS4, NiS4N2 sau NiN6 în mediul de coordinare. Mecanismul trebuie să ţină cont şi de faptul că aceasta enzimă accelerează schimbul H/D În apă conform reacţiei:
H2 + D2O HD + HDO
Viteza reacţiei de descompunere fotochimică este mult micşorata în apă deuterata (D20) ceea ce ne îndreptăţeşte să presupunem că speciile fotolitice sunt sub formă de hidruri ale Ni(I) sau Ni(II). Mecanismul implica ruperea heterolitica a H2, ionul hidrura (H-) rămânând pe ionul metalic, În timp ce componenta protica (H+) se leagă de un atom de sulf coordinat la metal, reacţia producând încă un centru bazic în interiorul proteinei. Natura presupusei legături dintre hidrogen şi nichelul catalitic prezintă un interes deosebit, hidrurile de nichel fiind binecunoscute în chimia organo-metalica. A fost discutată şi posibilitatea unei coordinări "side-on".
Fe/Ni hidrogenazele prezintă o competiţie în realizarea legăturii cu H2 şi CO2 ; reacţiile de scindare fotochimică ale complexului carbonilic generează intermediari catalitici care au fost studiatf prin cinetică cu schimb izotopic. Efectul izotopic de schimb H/D este mai pronunţat în cazul complexului format de enzime cu hidrogenul, ceea ce confirmă ipotezele în legătură cu mecanismul de reacţie.

Modelul legării şi scindării heterolitice a H2 la compuşii de coordinatie ai nichelului cu liganzi de sulf.
Măsurătorile RPE şi XAS efectuate pe hidrogenaza din Desulfovibrio baculatus au indicat o coordinare a selenocistcinei la atomul de nichel (distanţa Se-Ni este 244 pm); astfel liganzi care conţin sulf se afla la o distanţă de 217 pm, iar trei sau patru atomi de oxigen (sau azot) la o distanţă de 206 pm de metal. Ni/Fe/Se hidrogenazele întâlnite într-o serie de microorganisme au o mare afinitale pentru reţinerea hidrogenului (H2) dar prezintă o scădere a posibilităţii de schimb H/D, şu gerând rolul de bază al anionului de seleniu în scindarea heterolitica a H2.
6.Metil coenzima m reductaza
Metil-coenzima M reductază (MCR),este enzima responsabilă pt formarea microbiană a metanului, este o proteină de 300 kilodaltoni organizată ca un hexamer într-un aranjament α2β2γ2.În enzimologie, coenzima-B sulfoetiltiotransferaza, de asemenea, cunoscută sub numele de metil-coenzima M reductază (MCR) sau mai sistematic ca 2- (metiltio) etansulfonat: N-(7-tioheptanoil) -3-O-fosfotreonină S- (2-sulfoetil) tiotransferaza este o enzimă care catalizează etapa finală în formarea de metan. Ea face acest lucru prin combinarea hidrogenului donor al coenzimei B și metilul din coenzima M. Prin aceasta enzimă,se produce cea mai mare partea gazelor naturale de pe pământ. Rumegătoarele (de exemplu, vaci) produc metan, deoarece stomacul lor conține procariote metanogene (Archaea) ,care codifică și exprimă un set de gene de acest complex enzimatic. Enzima are două zone active, ocupate de fiecare conținând nichel F430 drept cofactor.
2- (metiltio) etansulfonat (metil-CM) + N- (7-mercaptoheptanoyl) treonină 3-O-fosfat (coenzima B) (reversibil)CoM-SS-DA + metan. Astfel, cele două substraturi ale acestei enzime sunt 2- (metiltio) etansulfonat și N- (7-mercaptoheptanoil) treonină 3-O-fosfat, în timp ce cele două produsele sale sunt CoM-SS-DA și metan. La unele specii, enzima reacționează în sens invers (-un proces numit metanogeneza inversă), catalizând oxidarea anaerobă a metanului, prin urmare, eliminarea din mediu. Aceste organisme sunt metanotrope. Această enzimă aparține familiei de transferaze,mai exactl cele care transfera grupuri tio.
Această enzimă participă la biosinteza de acid folic .
Ruperea din bacteria metanogenă a metil-ceoenzimei M (2- metiletansulfonat), cu eventuala formare şi eliberare a metariului, este catalizata de metil coenzima M reductaza. În

figură este reprezentată ultima etapă în producerea energiei sintetizând CH4 din CO2 de către arheobacterii autotrofe, cum ar fi Metanobactenum termoautotroficum.
Bacteriiie metanogene participa la degradarea microbiaia anaeroba a componentelor organice din plante, proces care prin volum este considerat ca fiind unul dintre cele mai importante procese biotehnologice. Producerea metanului prin degradare microbiană are loc în sedimente (se obţin gaze naturale), în timpul cultivării orezului şi în sistemul digestiv al rumegătoarelor. În ultimul timp s-a acordat multă atenţie urmelor de metan din atmosfera datorită contribuţiei lor la “efectul de seră”. Deşi cea mai mare parte a metanului biogenic rezulta prin degradarea radicalului acetat, formarea sa din C02 este mult mai interesantă din punct de vedere chimic.
Ultima etapă în procesul de obţinere a metanului
Procesul. global necesită 8 electroni şi prezenţa mai multor enzime esenţiale. "Metanofuranul” permite fixarea CO2- ului şi transformarea funcţiunii -COOH în timpul primei reduceri, cu implicarea a doi electroni. Gruparea formil rezultată este transferată tetrahidrometanopterinei care funcţional se aseamănă cu acidul tetrahidrofolic din eucariote. După alte două etape de transfer a câte doi electroni (formil -> hidroximetil -> metil), gruparea metil rezultată este transferată coenzimei M care pune în libertate metanul printr-o reacţie catalizată de metil-coenzima M reductaza. Forţa motrice a acestui proces este formarea legăturii S-S între coenzima M şi HS-HTP. Electronii necesari diferitelor etape ale reducerii sunt obţinuţi direct prin oxidarea hidrogenului molecular, proces catalizat de diferite hidrogenaze.Metil-coenziina M reductaza este o enzimă complexă şi foarte sensibilă. Dimerul proteic are o masă moleculară de aproximativ 300 kDa şi este format din trei subunităţi cu diferite de 68,47 şi 38 kDa.Din enzima a fost izolată o substanţă ce conţine Ni, de culoare galbenă, cu masa molecular mică, care prezintă absorbţie maximă la 430 nm şi care a fost denumită F430.Structura acestei prime coenzime biogene a fost stabilită în urma unor biosinteze de laborator şi a aplicării metodelor spectroscopice de studiu (RMN). Ea prezintă un ciclu porfirinic înalt hidrogenat condensat cu gama-lactama şi ciclohexanona. Structura ciclica a fost denumită “hidrocorfin” pentru a arăta asemfinarea ei structurală, cu porfirina şi nu cu ciclul conjugal corinic (F430 are o legătură lipsă în inelul tetrapirolic).

Schema de obţinere a metanului biogenic din CO2
Caracterul parţial săturat al macrociclului precumn şi condensarea suplimentară cu cicluri săturate, determină apariţia unei flexibilităţi a stucturii, în special în sensul plierii ciclului tetrapirolic după direcţia axei S4 .
Devieri de la geometria tipică a compuşilor macrociclici cu inel tetrapirolic

O consednta importantă a acestui fapt este modificarea stării de spin. Astfel Ni(II) S = 0 spin scăzut suferă o tranziţie d9, facilitând o altă orientare neaxiala a Ni(II) spin înalt cu S = 1. O configuraţie d8 spin scăzut (spinii sunt împerecheaţi) rezultă doar în urma unei distorsiuni a câmpului octaedric al liganzilor, degenerarea orbitalului eg, care defavorizează împerecherea spinilor, joacă în acest caz un rol important. Cum este de aşteptat, pentru speciile Ni(I) = d9, spectrul RPE al F430 reduse indică un orbital dx2-y2 semiocupat, delocalizat peste atomii de azot din planul ecuatorial şi un centru nucleofil putenic, dat de perechea de electroni din orbitalul dz2.Pentru o reacţie de adiţie oxidativă, ambele coordinări ale Ni(II) şi Ni(I) spin scăzut cu liganzii nesaturaţi, sunt importante pentru activarea metil-derivatilor E - CH3. Totuşi, din cauza flexibilităţii geometriei ligandului macrociclic, parametrii legăturilor sunt diferiţi: spin înalt Ni(II) - N = 210 pm, spin scăzut Ni(II) - N =190 pm şi Ni(I) - N aproximativ 200 pm. La fel ca alţi liganzi tetrapirolici, sistemul conjugat corfinic poate accepta un electron mai ales atunci când prezintă o grupare carbonil conjugată cu inelul ciclohexanonei. Pentru reducerea complexului trebuie creată o ambivalentă, în sensul reducerii alternative a metalului sau ligandului; o ambivalentă similară întâlnim şi în Fe- porflirine. O altă particularitate este prezenţa unei singure sarcini negative În interiorul macrociclului hidrocorfinic care fixează metalul, contrar ligandului porfirinic care coordinează două specii deprotonate.
Schema ipotetică a mecanismului de funcţionare a Ni ca centru active în enzima ce leagă cofactorul F430
Detaliile despre mecanismul de funcţionare a nichelului ca centru activ în enzima ce leagă cofactorul F430 nu sunt complete. În schema ipotetică din figura nichelului i se atribuie un rol în transferul electronului şi un rol în coordinarea radicalului CH3. în scurta sa viaţă (intermediar organo-metalic). Legarea directă de substrat a tioeterului sau a centrilor ce conţin S, este una diritre posibilele funcţii ale ionului de nichel. Scindarea legăturii alchil-tiol ( C-S) prin intermediarul speciilor active cu nichel este o tehnică cunoscută de la Ni Raney şi catalizarea similară a reacţiei de desulfatare.
7. CO Dehidrogenaza = CO Oxidoreductaza = Acetil-CoA Sintetaza
Multe bacterii metanogene şi acetogene (care produc metan şi acid acetic) conţin “CO dehidrogenaza”, enzima ce catalizează oxidarea CO/C02. În biochimie oxidarea este prezentată ca fiind o dehidrogenare, totuşi CO nu conţine hidrogen, iar termenul de CO oxidoreductaza pare mai potrivit.CO + H20 CO2 + 2H+ +2eReacţia enzimatică este reversibilă şi de aceea poate servi ca modalitate altenativa de fixare (asimilare) a C02 de către bacteria activa în procesul de fotosinteză. Cealaltă funcţie biologică a enzimei este să catalizeze formarea reversibilă a acetil-coenzimei A prin combinarea coenzimei Acuo sursa de metil. Proteinele corinoide cu funcţionalitate CH3-[Co] ale metil transferazei şi disulfit reductazele ce conţin Fe/S, contribuie de asemenea la această reacţie.CH3-[Co] + CO + HS-CoA CH3-C(O)S-CoA + H+ + [Co]- co2

Acetil-CoA rezultată (“acid acetic activat”) poate fi carboxilata de către bacterii anaerobe la piruvat CH3C(O)COO. În bacteriile metanogene degradarea acidului acetic la C02 şi CH4 decurge probabil cu obţinerea ca intermediar a CO.
Este cunoscut că, CO dehidrogenazele din bacteriile a aerobe conţin nichel În timp ce speciile aerobe necesita molibdenopterine. A fost studiată CO dehidrogenaza bacteriei acetogene Clostridium thermoaceticum care
este sensibilă la prezenţa oxigenului. Ea este alcătuită din 3x2 subunităţi ( )3 de mărime 82 şi 73 kDa. Nu este încă stabilită identitatea metalului conţinut.S-au găsit cantităţi mici de nichel (un atom de Ni pentru fiecare unitate hexamera de proteină) şi cantităţi mari de fier şi
sulf (mai mult de 10 atomi / ( ) dimer) precum şi diferite cantităţi de zinc. Datele privind structura centrului activ care conţine Ni au fost obţinute în cea mai mare măsură prin metoda RPE, spectroscopie Mossbauer şi EXAFS. Datele RPE, S = 1/2 indica interacţia electronului cu trei sau mai mulţi atomi de fier, cu un atom de nichel şi un atom de carbon (pentru studiul scindării hiperfine se folosesc izotopi îmbogăţiţi: 61Ni, 57Fe şi13CO). Datele EXAFS sugerează pentru nichel coordinarea a patru liganzi cu S aflaţi la o distanţă de aproximativ 216 pm Conform spectroscopiei Mossbauer distant Ni-Fe este estimate la 325 pm şi indică interacrtia nichelului cu un cluster Fe/S. Studii, recente structurează enzima în patru subunităţi: un cluster A ce cuprinde atomi de Ni,şi unităţi Fe-S active în sinteza acetil coenzimei A ; un cluster C care reprezintă partea activa în oxidarea CO; un cluster [Fe4S4] 2+/1+ şi o “componentă feroasă”.
Enzima cu structura cuaternara de tipul 2 2 la tratare cu cantităţi mici de detergent SDS
suferă o descompunere într-o unitate metalică alfa şi una 1 2 care prezintă activitate oxidantă asupra CO. Subunitatea metalică alfa conţine un atom de nichel şi patru atomi de fier. Ni este coordinat la liganzi ce conţin doi atomi de sulf şi doi atomi de oxigen sau azot, geometria nichelului fiind de plan-patrat deformat, iar Fe este dispus într-un cluster [Fe4S4]2+/1+ care în stare redusă are S=3/2. În experimentele de laborator carbonilarea CO dehidrogenazei s-a dovedit a fi reversibila, schema mecanismului de reactfe putând explica rezultatul coordinării reversibile a CO la Ni.
Modelul mecanismului de coordinare reversibilă a CO la nichel
După demonstrarea existenţei a doi centri activi metalici unul de Ni şi unul de Fe, M. Kumar propune un mecanism bimetalic pentru sinteza acetil coenzimei A în care atât Fe cât şi Ni au roluri catalitice. Pentru transferul grupării metil de la metil-Co3+ (Co= coenzima) din C/Fe-SP la CO dehidrogenaza (CODH) s-a propus următoarea reacţie:
CH3 - Co3+ -C/Fe-SP + Ni-CODH Co1+-C/Fe-SP + CH3-Ni-CODH.
De vreme ce produsul este Co1+,reacţia trebuie să implice o deplasare nucleofilă între gruparea CH3-Co 3+ şi Ni1+ pentru a forma CH3-Ni 3+ care apoi poate accepta un electron pentru a forma o specie mai stabilă CH3- Ni 2+ .

Compus model pentru funcţiunile Ni(II)-Me,Ni(II)-COMe,Ni(II)-H şi Ni(I)-CO
Compuşi model
Atunci când concepem un compus de modielare a centrului activ metalic, nu este suficientă caracterizarea Ni-proteinei deoarece apar şi alte probieme, cum ar fi aparenta stabilitate a unor stări de oxidare “exotice” sau surprinzătoare. În compuşii organo-metalici nichelul se afla frecvent în stări joase de oxidare +1 sau zero, care sunt importante în tehnica catalitică a hidrogenărilor (hidrogenaze), desulfurizări (metil-coenzima M reductaza) şi în reacţii de carbonilare (CO dehidrogenaza sau CO oxidoreductaza).
Pe de altă parte Ni(III) a fost considerat mult timp ca fiind într-o stare de oxidare improprie lumii vii. Totuşi, pot fi sintetizaţi un număr mare de compuşi de coordinatie ai Ni(III) cu liganzi ca : amidele chelatizante, peptidele deprotonate, oxime şi tioli, sau poate fi stimulată cel puţin detecţia Ni(III) ,dacă nu este necesară enzima activă.Un aspect important în modelaiea Ni(III) este geometria coordinatiei care s-a dovedit a fi restrictiva. De exemplu, pentru aceeaşi liganzi chelatizanti configuraţiile electronice d8 (Ni(II)) şi d7 (Ni(III)) manifestă preferinţe pentru geometrii diferite. Aplicând aceasta strategic şi folosind liganzi donori bogaţi în electroni, de tipul RS , S2- sau amide R2N- , a fost posibilă micşorarea potenţialului de oxidare a Ni(II)/Ni(III) astfel că speciile Ni(III) devin stabile chiar şi în condiţii fiziologice.
Compus model al Ni(III) cu Gly-Gly-Gly3-
Printre compuşii de coordinate cu Ni(III) stabili sunt şi compuşii cu peptidele deprotonate de dimensiuni mici. Centrul Ni(III) în complexul din figura este localizat în planul format de tripeptida în timp ce pozitfile axiale sunt ocupate de molecule de apă slab coordinate. Asemenea compuşi pot fi uşor preparaţi prin oxidarea electrochimică a compuşilor cu Ni(II) corespunzători. Ca şi tiolatii, peptidele deprotonate la azot (carboxiamidele) sunt putenic
donoare atât cât şi şi sunt capabile sastabilizeze stările înalte de oxidare ale metalului. Pentru trecerea NiII/III complexul prezentat are un potenţial redox relativ mare, scăderea semnificativă a acestei valori întâlnindu-se în hidrogenaze şi alte enzime cu nichel,fiind datorată probabil multitudinii de liganzi cisteinici.
Modelarea Ni-hidrogenazelor
După cum s-a arătat, hidrogenazele catalizează oxidarea reversibilă a moleculei de hidrogen permiţând celulelor să genereze sau să elibereze în timpul respiraţiei echivalenţi de reducitori:H2 2H++2e-
Publicarea recentă a unui studiu de analiza cu raze X asupra structurii cristaline a centrului activ din enzimă, deschide noi perspective în elucidarea şi modelarea hidrogenazei din Desulfovibrio gigas.

Structura prezintă un atom de Ni coordinat prin intermediul sulfului de 4 grupări cisteinice, două din ele formând o punte cu un al doilea metal, presupus a fi Fe. Modelul structural va permite apoi stabilirea mecanismului de reacţie şi a activităţii redox a proteinei. Pentru aceasta trebuiesc sintetizaţi compuşi mononucleari ai Ni cu grupe tiol, cum ar fi compusul de coordinatie ai Ni(II) cu N,N’-dimetil-N,N’-bis(P- mercaptoetil) etilendiamina,în solventfi aprotici, care să formeze prin punţi de sulf compuşi heterodinucleari, prin reacţia cu compuşi coordiriativi mononucleari ai fierului (este preferata sinteză “în situ”) în solvenţi protici. Similar sinteza decurge plecând de la ditioli prin condensare cu baze Schiff în prezenţa Ni(II) (reacţie templata).
Exemplu de reactive template pentru modelarea centrului active cu Ni din hidrogenaza
În ultimii ani s-au sintetizat liganzi N şi S donori cum ar fi:
Liganzi model pentru simularea structurii şi funcţionalităţii hidrogenazelor pe bază de Ni
Cu L1 au puitut fi izolaţi mai mulţi compuşi ai nichelului. Când anionul este [BF4]- sau [B(Ph)4]- complexul trinuclear are formula [Ni3(L1)4](anion)2 şi poate fi izolat. Există indicaţii biliografice privind existenţa unui compus mononuclear [Ni(L1)2] dar nu s-au putut separa cristale. Reducerea chimică a unei soluţii de compus trinuclear cu un echiyalent de [BF4]- da spedi detectabile RPE.Reducerea electrochimică este ireveisibila şi se realizează cu Ag/AgCl. În atmosfera de hidrogen se observe oxidarea catalitică a H2.Cu ligandul L2 s-au obţinut cristalele unui compus cu formula [Ni(L2)] [BF4]. Structurile acestor compuşi sunt cunoscute, reactivitatea lor este încă în studiu.Eforturi considerabile s-au făcut de către A.Berkessel şi col. în sensul imitării activităţii catalitice a [Ni,Fe]-hidrogenazelor de către chelati ai Ni cu mase moleculare mici. Liganzii implicaţi sunt: tiosemicarbazona plana , derivată din compuşi carbonilici aromatici orto- hidroxi-substituiti (1) şi sisteme N, O, S - “tripodal” (2) cum ar fi rac – 2.
Liganzi chelatizanti;modele pentru hidrogenaza.
R3Şi-H + H-O-R’ H2 + R3 Şi-O-R’Compusul Ni(II) cu tiosemicarbazona şi liganzi de tipul (1) activează legătura Şi-H din silani favorizând un atac nucleofil ca în reacţia de alcooliza a silanilor. Pentru a elucida mecanismul acestei reacţii, care este corelat cu activarea moleculei de hidrogen de către hidrogenaze, trebuie luate în considerare, mărimea liganzilor ai ionului metalic central şi condiţiile de reacţie.În cazul liganzilor de tip “tripodal” rac-2 noul cluster Ni3N3S4 rac- 3 rezultă din reacţia dintre tiol rac-2a cu acetatul de Ni(II). Benzilul şi trimetil- sililtioeterul rac-2b, permit amândoi pentru numărul de coordinare 4 geometria plan-patratica, iar pentru numărul de coordinare 6 geometria pseudooctaedrica impuse de condiţiile de reacţie.

Structura compusului rac-3= Ni3N3S4
Au fost sintetizaţi compuşi ai Ni(II) prin reacţia clorurii de nichel cu o serie de derivaţi ai bazelor Schiff obţinute prin condensarea P,P-bis- 3(4-amino-5-mercapto-1,2,4-triazolil) benzen cu salicil aldehide substituite. La un raport stoechiometric 1:1 compusul este de tipul NiL.2H20. Valorile molare ale conductibilităţii în DMF sunt prea mici pentru a putea fi luate În considerare în procesul de disociere a compusului. Aceşti compuşi funcţionează ca filtru cu activitate antibacteriană şi antifungica împotriva bacteriilor Escherichia coJi, B. cirrafigellosus şi citipercilor C. albican şi A. niger.U. Gauding şi coL au sintetizat un număr mare de compuşi model pentru Ni-hidrogenaze de tipul celor din figura, în care L poate fi imidazol, piridina sau fosfina; X este oxigen sau sulf, grupările R1, R2 şi R având diferite efecte electronice:
Tipuri de liganzi tridentati [ONS]2-
Aceşti compuşi sunt diferiţi în ceea ce priveşte structură, reactivitatea şi mecanismul lor de acţiune. În prezenfa solventului s-a pus în evident un dimer cu oxigen sau sulf, numărul de coordinare formal fiind 3. În solvenţi care nu coordinează la Ni s-au obţinut oligomeri superiori care sunt în echilibru cu dimerii. Pentru diferiţi liganzi L avem:monomeri plani tip aduct 1:1 pentru L = baze cu azot, fosfine; dimeri pentacoordinati aduct 1:1 pentru L = piridina; tetrameri octaedrici aduct 1:1 pentru L = MeOH iar X = O;monomeri octaedrici aduct 1:3 pentru L=piridina. În funcţie de natura atomilor donori şi substituenţii ligandului tridental chelatant.
Echilibrele de substitute a ligandului care pot fi examinate spectrofotometric, indica clar că compuşii, coordinativi ce conţin X = S şi/sau R = 1,2 fenilen sunt mai “moi” decât cei cu X = O şi/sau R = etilen. Activitatea catalitică a acestor compuşi a fost testată pe reacţia de alcooliza a trietilsilanilor:
Et3Şi-H + HO-R H2 + Et3Şi-OR
R. Crabtree a afirmat că această reacţie este un test potrivit pentru modelarea hidrogenazelor. Doar compuşii cu X = S §i R = 1,2 fenilen au activitate catalitică bună şi ea
creşte în seria: 1f < 1e < 1d < 1a 1b < 1c. Toţi compuşii cu R = etilen şi X = 0 sunt inactivi.

Pentru caracterizarea cât mai exactă a geometriei nichelului şi naturii legăturii lui cu substratul enzimatic Paul K.Ross a efectuat un studiu asupra a trei serii de compuşi model cu Ni(III), folosind o combinaţie a metodei dicroismului magnetic circular (DMC) la temperatura variabilă cu spectroscopia RPE. Prima serie este reprezentată de [Ni(III)(PS3*)(t-butilpiridina)] şi [Ni(III)(PS3*)(N-metilimidazol)] sintetizaţi de Millar pe baza împiedicărilor sterice din ligandul PS3*= tri(6-feni]-2-mercaptofenil)fosfina. Aceşti compuşi au o geometrie de bipiramidă trigonala, cu atomii de sulf plasaţi ecuatorial. Spectrele DMC prezintă tranziţii caracteristice S-Ni şi P-Ni dar nu s-a evidetiat nici o tranziţie d-d. Al doilea grup conţine [Ni(siclam)(SO4)](ClO4) şi [Ni(siclam)(Cl2)](ClO4) cu geometrie de plan-patrat, în planul ecuatorial fiind coordinaţi atomii de N. Spectrele DMC ale acestor compuşi dau o tranziţie d-d la 690 nm pentru compusul cu clor şi la 810 nm pentru cel cu sulfat. Ultima serie consta din [Ni(tacn)2](ClO4) şi [Ni(tacn)2](ClO4)3 compuşi în care Ni are o geometrie octaedrică şi un număr de coordinare 6, fiind coordinaţi tioeteri.
Recommended

















