
Extrapolatie van de functie van lipoproteïne lipase
in chronische lymfatische leukemie
Nore Robberecht
Verhandeling ingediend tot
het verkrijgen van de graad van
Master in de Biomedische Wetenschappen
Promotor: Prof. Dr. Jan Philippé Begeleidster: Drs Ans Rombout
Vakgroep Klinische Biologie
Academiejaar 2011-2012


Extrapolatie van de functie van lipoproteïne lipase
in chronische lymfatische leukemie
Nore Robberecht
Verhandeling ingediend tot
het verkrijgen van de graad van
Master in de Biomedische Wetenschappen
Promotor: Prof. Dr. Jan Philippé Begeleidster: Drs Ans Rombout
Vakgroep Klinische Biologie
Academiejaar 2011-2012

“De auteur en de promotor geven de toelating deze masterproef voor
consultatie beschikbaar te stellen en delen ervan te kopiëren voor persoonlijk
gebruik. Elk ander gebruik valt onder de beperkingen van het auteursrecht,
in het bijzonder met betrekking tot de verplichting uitdrukkelijk de bron te
vermelden bij het aanhalen van resultaten uit deze masterproef.”
Datum: 20 mei 2012
(handtekening student) (handtekening promotor)

Voorwoord
Deze thesis werd verwezenlijkt met vele uren door te brengen in het lab gecombineerd met
een zoektocht in een overvloed aan literatuur. Zonder de hulp van andere mensen had ik dit
werk niet kunnen realiseren. Elk wetenschappelijk onderzoek kent zijn hoogtes en zijn laagtes
en deze thesis ging dan ook gepaard met euforie en ontgoocheling.
Ik wil graag enkele mensen bedanken die me gesteund hebben om deze thesis te schrijven.
In de eerste plaats, wil ik mijn promotor, Professor Philippé, bedanken voor zijn snelle
verbeteringen, zijn enthousiasme en zijn goede raad tijdens de wekelijkse vergaderingen. Ik
ben hem dankbaar om me te mogen vervoegen bij het CLL-team. Dankzij hem, heb ik me
kunnen nuttig maken in het CLL-onderzoek.
In het bijzonder wil ik graag mijn begeleidster, Drs Ans Rombout, vermelden die me
uitstekend begeleid heeft. Ze heeft me gemotiveerd gedurende de hele stage.
‘Ans, dank je wel voor de leuke momenten, de enorme steun en de hulp gedurende het hele
jaar. Je bent een pracht van een begeleidster. Ik vond het aangenaam samenwerken met jou’
Professor Verhasselt mag zeker niet vergeten worden. Ik wil hem bedanken voor de vele
verduidelijkingen tijdens de vergaderingen en het aanreiken van mogelijke oplossingen.
Door haar onuitputtelijk doorzettingsvermogen, heeft Drs Valerie Pede me laten zien dat je
nooit mag opgeven in het CLL-onderzoek. Haar vele praktische tips in het lab waren voor mij
een enorme hulp.
Mijn ouders zou ik graag ophemelen omdat ze me gedurende de hele opleiding hebben
gesteund. Ze hebben me telkens gemotiveerd om in de zware examenperiodes door te zetten.
Ze hebben altijd geluisterd en interesse getoond in mijn onderzoek.
Tenslotte, wil ik Jonas Goossens bedanken om me op te vangen als ik de moed even liet
zakken. Het was motiverend om met iemand samen te leven die eveneens een
wetenschappelijke thesis schreef. Bedankt om mijn thesis na te lezen en te verbeteren. Jouw
ongelooflijke morele steun heeft me telkens opnieuw opgeladen.

Inhoudstafel
Samenvatting .............................................................................................................................. 1
I. Inleiding .................................................................................................................................. 2
1. Chronische lymfatische leukemie ...................................................................................... 2
1.1 Ontstaansmechanisme .................................................................................................. 2
1.2 Diagnose en behandeling ............................................................................................. 3
2. B-cel ontwikkeling ............................................................................................................. 5
3. Prognostische merkers bij CLL .......................................................................................... 7
3.1. Algemeen .................................................................................................................... 7
3.2 Genetische afwijkingen ................................................................................................ 8
3.3 IgVH mutatiestatus ........................................................................................................ 9
3.4 Surrogaatmerkers voorde IgVH mutatiestatus .............................................................. 9
3.5 Hypothetisch werkingsmechanisme van LPL in CLL ............................................... 12
4. Doelstelling ...................................................................................................................... 13
II. Materialen en methoden ...................................................................................................... 15
1. Stimulatie experiment ...................................................................................................... 15
1.1 Selectie van de patiënten ............................................................................................ 15
1.2 Isolatie en stimulatie ................................................................................................... 15
1.3 Oogsten van de cellen en flowcytometrie .................................................................. 16
1.4 Isolatie RNA ............................................................................................................... 17
1.5 cDNA synthese ........................................................................................................... 18
1.6 Kwantitatieve qPCR ................................................................................................... 18
2.pCMV-Sport6-LPL ........................................................................................................... 22
2.1 Vermenigvuldigen plasmide ...................................................................................... 22
2.2 Isolatie plasmide DNA ............................................................................................... 23
2.3 Digest en opzuivering ................................................................................................. 23
2.4 In vitro transcriptie ..................................................................................................... 23

2.6 mRNA Elektroporatie ................................................................................................ 24
3. pGEM4Z-LPL-A64bis ..................................................................................................... 25
3.1 Isolatie plasmide DNA pGEM4Z-EGFP-A64bis ....................................................... 25
3.2 Klonering .................................................................................................................... 25
3.3 mRNA elektroporatie ................................................................................................. 28
4. Transfectie HEK 293T-cellen .......................................................................................... 28
4.1 Uitplaten cellen .......................................................................................................... 28
4.2 Transfectie .................................................................................................................. 28
4.3 Analyse transfectie efficiëntie .................................................................................... 29
5.ELISA ................................................................................................................................ 29
III. Resultaten ........................................................................................................................... 31
1. Detectie van LPL in LPL-geëlektroporeerde CLL cellen ................................................ 31
1.1 Elektroporatie van LPL mRNA bekomen via IVT van pCMV-Sport6-LPL ............. 31
1.2 Elektroporatie van LPL mRNA bekomen via IVT van pGEM4Z-LPL-A64bis ........ 33
2. Detectie van LPL in HL-60 cellen en gestimuleerde CLL cellen .................................... 36
2.1 HL-60 cellen ............................................................................................................... 36
2.2 Gestimuleerde CLL cellen.......................................................................................... 37
3. Detectie van LPL in pCMV-Sport6-LPL getransfecteerde HEK 293T-cellen ................ 39
4. LPL eiwit detectie via ELISA .......................................................................................... 42
IV. Discussie ............................................................................................................................ 43
V. Referenties ........................................................................................................................... 47

Lijst met afkortingen
ADP Adenosine diphosphate
AP-1 Activator protein-1
ATM Ataxia telangiectasia mutated
Bag-1 Bcl-2-associated athanogene 1
Bax Bcl-2-associated X protein
Bcl-2 B-cell lymphoma-2
Bcl-xS Bcl-extra small
Bcl-xL Bcl-extra large
BCR B-cell receptor
BLNK B-cell linker
BSA Bovine serum albumine
CD Cluster of differentiation
CHOP Cyclofosfamide, doxorubicine, vincristine en prednison
CLL Chronische lymfatische leukemie
CMV Cytomegalovirus
CVP Cyclofosfamide, vincristine en prednison
CXCR4 C-X-C chemokine receptor type 4
DCs Dendritische cellen
DLEU2 Deleted in lymphocytic leukemia2
DMSO Dimethylsulfoxide
ECM Extracellulaire matrix
EDTA Ethylenediaminetetraacetic acid
EGFP Enhanced green fluorescent proteïne
ELISA Enzyme-linked immunosorbent assay
FACS Fluorescence-activated cell sorting
FCR Fludarabine, cyclofosfamide en rituximab
FCS Fetal calf serum
FITC Fluorescein isothiocyanate
FOS FBJ osteosarcoma
FSC Forward scatter
HBS Hanks’ balanced salts
HEK 293T-cel Human embryonic kidney 293T-cel
HL-60 Humane promyelocytaire leukemie-60
HRP Horseradish peroxidasde
HSPG Heparan sulfate proteoglycan
IFN Interferon
Ig Immunoglobuline
IL Interleukin
IMDM Iscove’s modified dulbecco’s medium
IRES Internal ribosome entry site
ITAM Immunoreceptor tyrosine-based activation motif
IVT in vitro transcriptie
LB Lysogeny broth
Lck Lymphocyte-specific protein tyrosine kinase
LDH Lactaat dehydrogenase
LiCl Lithiumchloride
LPL Lipoproteïne lipase
LRP LDL Receptor-related protein
MAPK Mitogen-activated protein kinase
MCL-1 Myeloid cell leukemia-1 gene
MDR Minimal deleted region
NAD Nicotinamide adenine dinucleotide
NFAT Nuclear factor of activated T-cells
NF-H2O Nuclease-free water
NFκB Nuclear factor κB
NK Natural killer cellen
NLC Nurse-like cellen
PBMCs Peripheral blood mononuclear cells
PE Phycoerythrin
PI Propidium iodide
PKC Protein kinase C
PLCγ Phospholipase C
qPCR Quantitative PCR
RISC RNA-induced silencing complex
SSC Side scatter

Syk Spleen tyrosine kinase
TAG Triacylglycerol
TCL-1 T-cell leukemia-1 gene
TCR T-cell receptor
VCAM-1 Vascular cell adhesion molecule-1
VLDL Very low density lipoprotein
ZAP-70 Zeta-chain-associated protein 70 kDa

1
Samenvatting
B-cel chronische lymfatische leukemie (CLL) is een neoplasie gekenmerkt door een
progressieve accumulatie van monoklonale mature B-cellen in bloed, beenmerg en lymfoïde
weefsels. Het ziekteverloop van CLL patiënten is zeer heterogeen. Sommige patiënten hebben
een indolente vorm en een lange overlevingstijd (10 jaar) terwijl anderen snel evolueren naar
een agressieve vorm met een korte overlevingstijd (2-3 jaar). Er zijn verschillende
prognostische merkers aangetoond voor CLL. Daarbij is de mutatiestatus van het
immunoglobine zware keten (IgVH) gen de gouden standaard om de prognose te voorspellen.
Een ongemuteerde IgVH status correleert met een ongunstige prognose. Omdat de analyse van
het gen tijdrovend en arbeidsintensief is, wordt er onderzoek gedaan naar surrogaatmerkers
voor deze status. Zo werd aangetoond dat lipoproteïne lipase (LPL) een goede en betrouwbare
prognostische surrogaatmerker is. Dit suggereert dat LPL een functie vervult in de
pathogenese van CLL cellen. Het werkingsmechanisme van LPL op CLL cellen is echter nog
niet gekend. In deze thesis werd een efficiënte methode gezocht om het LPL gen tot expressie
te brengen in LPL- CLL cellen. Door CLL cellen te vergelijken die enkel verschillen in het
eiwit van interesse, kan specifiek onderzoek verricht worden naar de functie van LPL in de
CLL cel. In een recent onderzoek werd reeds aangetoond dat elektroporatie van in vitro
getranscripteerd (IVT) zeta-chain-associated proteïne (ZAP-70) mRNA, aangemaakt van de
vector pGEM4Z-ZAP70-A64bis, een efficiënte methode is om CLL cellen te transfecteren. In
deze thesis werd eveneens elektroporatie uitgevoerd van IVT LPL mRNA, aangemaakt van
twee vectoren, een aangekochte vector pCMV-Sport6-LPL en een gekloneerde vector
pGEM4Z-LPL-A64bis. Aan de hand van qPCR werd het LPL mRNA aangetoond in de
geëlektroporeerde cellen. Maar het LPL eiwit werd niet gedetecteerd via flowcytometrische
kleuring. Vertrekkend van dit resultaat werden verschillende mogelijke verklaringen
onderzocht. De effectiviteit van de flowcytometrische kleuring werd bevestigd door een
kleuring uit te voeren op HL-60 cellen, gestimuleerde CLL cellen en HEK 293T-cellen
getransfecteerd met de pCMV-sport6-LPL vector. Ook mutaties in de sequentie van LPL
werden uitgesloten door de sequentiebepaling van het gen. Uit voorgaande resultaten wordt
gesuggereerd dat de aanwezigheid van LPL mRNA niet noodzakelijk LPL eiwit-expressie
impliceert. Onderzoek om een efficiënte ‘gene transfer’ methode te zoeken, moet nog verder
op punt gesteld worden.

2
I. Inleiding
1. Chronische lymfatische leukemie
B-cel chronische lymfatische leukemie (CLL) is een neoplasie gekenmerkt door een
progressieve accumulatie van monoklonale mature B-cellen in bloed, beenmerg en lymfoïde
weefsels [1]. Het is de meest voorkomende vorm van leukemie bij volwassenen in de westerse
wereld [2]. De ziekte komt vooral voor bij oudere mensen en treft voornamelijk mannen. De
diagnose wordt gesteld op een gemiddelde leeftijd van 65 jaar. In de Verenigde Staten worden
ongeveer 4,2 personen per jaar per 100.000 inwoners getroffen [3]. CLL cellen worden
gekenmerkt door een abnormaal immunofenotype, namelijk co-expressie van de oppervlakte
antigenen CD5, CD19 en CD23 [4]. Bovendien hebben ze een verminderde expressie van
CD20, CD22, CD79b en van het oppervlakte antilichaam IgM, in vergelijking met normale B-
cellen [5]. Het ziekteverloop van CLL is zeer heterogeen. Sommige patiënten evolueren snel
naar een agressieve vorm met korte overlevingstijd, in tegenstelling tot patiënten met een
indolente vorm die gepaard gaat met een langere overlevingstijd. Bij patiënten met de
agressieve vorm, treden klinische symptomen sneller op. De meest voorkomende symptomen
zijn vermoeidheid en lymfadenopathie. De helft van de symptomatische patiënten vertoont
een infiltratie van CLL cellen in de milt met een pijnlijke zwelling, splenomegalie, tot gevolg.
Ook hepatomegalie kan optreden maar dit komt minder frequent voor. In een verder
gevorderd stadium kunnen gewichtsverlies, frequente infecties, anemie en trombocytopenie
voorkomen. Dit laatste kan leiden tot bloedingen [6].
1.1 Ontstaansmechanisme
Het ontstaansmechanisme is tot op heden niet gekend, maar CLL is hoogstwaarschijnlijk te
wijten aan een misregulatie in apoptose die leidt tot een accumulatie van de monoklonale B-
cellen die zich in de G0/G1 fase van de celcyclus bevinden. Enerzijds wordt deze hypothese
bekrachtigd door een stijgende expressie van het anti-apoptotisch eiwit Bcl-2 bij 95% van de
CLL patiënten [7]. In normale weefsels wordt de homeostase tussen proliferatie en apoptose
onder andere in stand gehouden door de Bcl-2 eiwit familie. Deze familie bestaat uit de pro-
apoptotische eiwitten, Bax en Bcl-xS, en de anti-apoptotische eiwitten Bcl-xL, Bag-1 en Mcl-
1 [8]. Een verstoorde expressie van beide subgroepen kan leiden tot een daling van apoptose
in CLL cellen. Anderzijds worden in 80% van de CLL patiënten genetische afwijkingen
waargenomen met een toegenomen prevalentie van del13q14, trisomie 12, del11q22 en
3
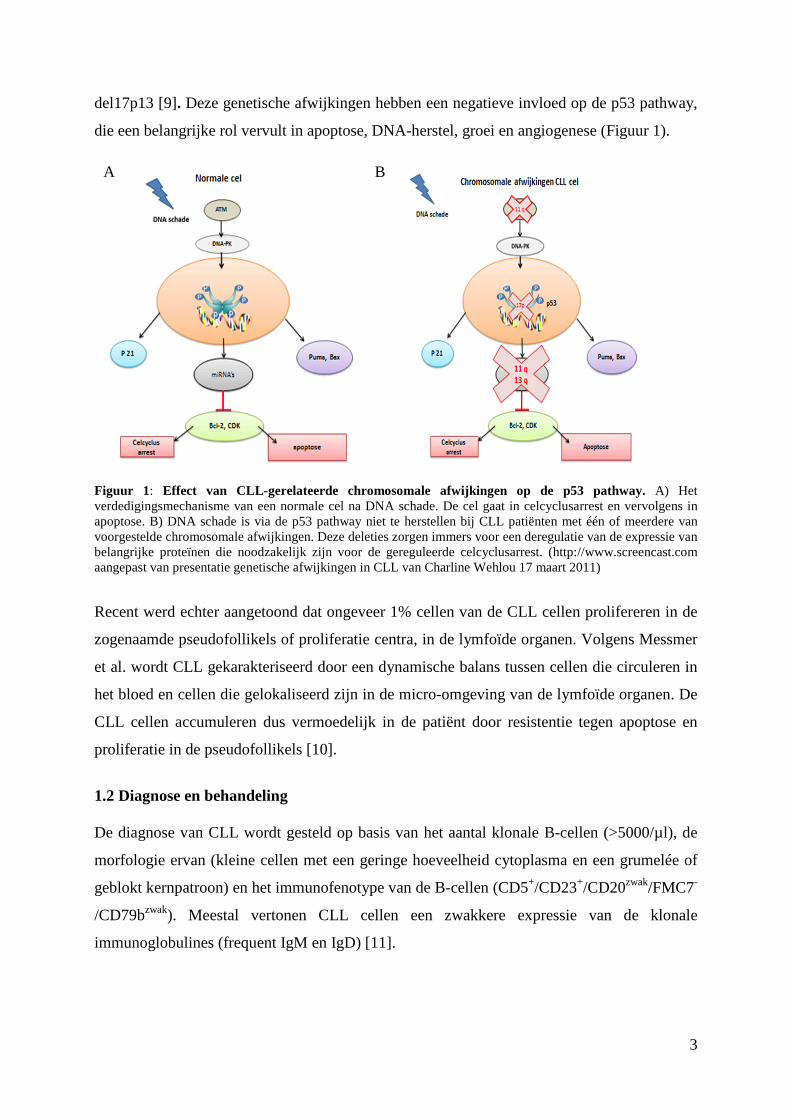
del17p13 [9]. Deze genetische afwijkingen hebben een negatieve invloed op de p53 pathway,
die een belangrijke rol vervult in apoptose, DNA-herstel, groei en angiogenese (Figuur 1).
Figuur 1: Effect van CLL-gerelateerde chromosomale afwijkingen op de p53 pathway. A) Het verdedigingsmechanisme van een normale cel na DNA schade. De cel gaat in celcyclusarrest en vervolgens in apoptose. B) DNA schade is via de p53 pathway niet te herstellen bij CLL patiënten met één of meerdere van voorgestelde chromosomale afwijkingen. Deze deleties zorgen immers voor een deregulatie van de expressie van belangrijke proteïnen die noodzakelijk zijn voor de gereguleerde celcyclusarrest. (http://www.screencast.com aangepast van presentatie genetische afwijkingen in CLL van Charline Wehlou 17 maart 2011)
Recent werd echter aangetoond dat ongeveer 1% cellen van de CLL cellen prolifereren in de
zogenaamde pseudofollikels of proliferatie centra, in de lymfoïde organen. Volgens Messmer
et al. wordt CLL gekarakteriseerd door een dynamische balans tussen cellen die circuleren in
het bloed en cellen die gelokaliseerd zijn in de micro-omgeving van de lymfoïde organen. De
CLL cellen accumuleren dus vermoedelijk in de patiënt door resistentie tegen apoptose en
proliferatie in de pseudofollikels [10].
1.2 Diagnose en behandeling
De diagnose van CLL wordt gesteld op basis van het aantal klonale B-cellen (>5000/µl), de
morfologie ervan (kleine cellen met een geringe hoeveelheid cytoplasma en een grumelée of
geblokt kernpatroon) en het immunofenotype van de B-cellen (CD5+/CD23+/CD20zwak/FMC7-
/CD79bzwak). Meestal vertonen CLL cellen een zwakkere expressie van de klonale
immunoglobulines (frequent IgM en IgD) [11].
A B

4
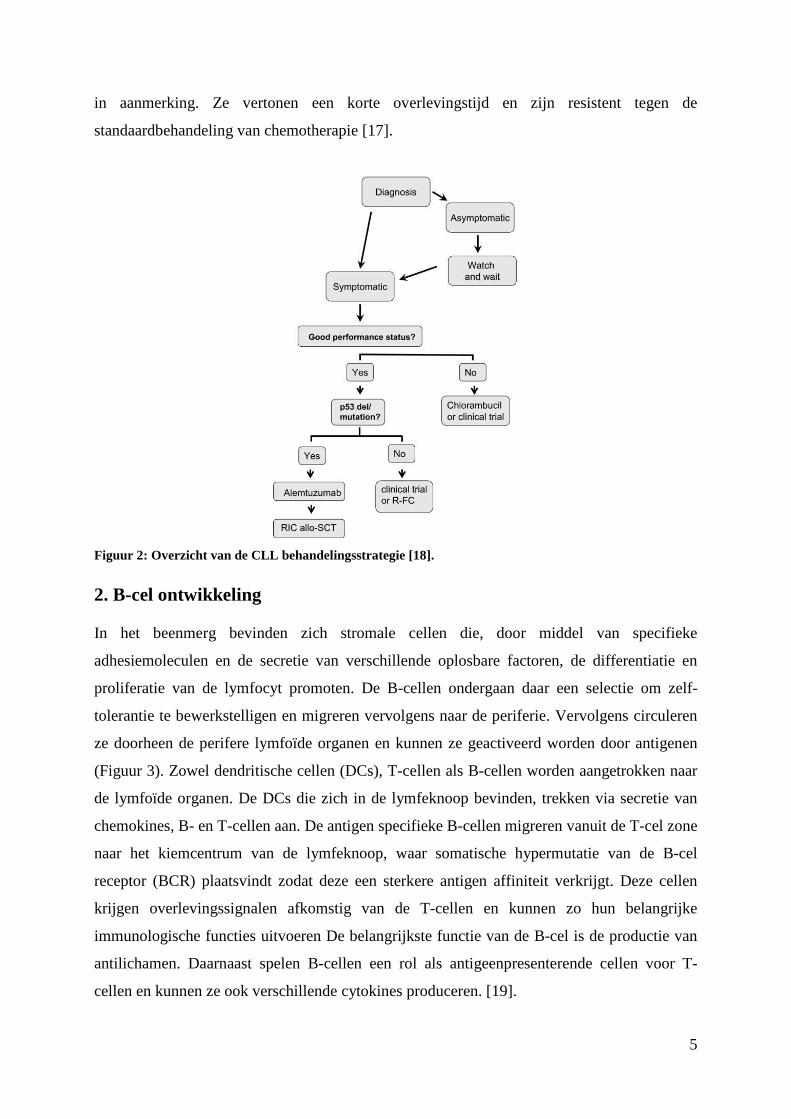
CLL is nog steeds niet te genezen maar de evolutie kan wel vertraagd worden. De
behandelingsstrategie wordt aangepast aan het type CLL en het stadium waarin de patiënt zich
bevindt (Figuur 2). De effecten van een eventuele behandeling moeten worden afgewogen
tegen de nadelen ervan [12]. In het vroege stadium van de ziekte wordt slechts therapie
toegepast van zodra de patiënt symptomen vertoont. Dit wordt de ‘watchfullwaiting approach’
genoemd. Na de beslissing of de patiënt al dan niet een behandeling krijgt, zijn er
verschillende mogelijkheden.
Chemotherapeutica en antilichaamtherapie
Celdelingsremmende chemotherapeutica worden frequent toegepast. Chloorambucil is een
oude en welgekende behandeling met een partiële remissie van de ziekte tot gevolg.
Alkylator/anthracycline combinaties zoals cyclofosfamide, vincristine en prednison (CVP) en
cyclofosfamide, doxorubicine, vincristine en prednison (ChOP) worden frequent gebruikt met
gelijkaardige of betere effecten dan chloorambucil, maar leiden niet tot een betere overleving.
Fludarabine is een purine analoog dat inhibitie van DNA polymerase en ribonucleotide
reductase veroorzaakt en bijgevolg apoptose promoot/stimuleert. Dit wordt frequent gebruikt
maar heeft verschillende toxische neveneffecten.
In sommige behandelingsschema’s worden de chemotherapeutica gecombineerd met
antilichaamtherapie. De nieuwe standaard eerstelijnsbehandeling voor CLL is de FCR, een
combinatie bestaande uit fludarabine, cyclofosfamide en het antilichaam rituximab. Er werd
aangetoond dat mensen met een p53 deletie (del17p13), vaak refractair zijn voor behandeling
met FC of FCR in tegenstelling tot patiënten met een 11q deletie, die een betere overleving
vertonen [13]. Bij aanwezigheid van een p53deletie zal gekozen worden voor alternatieve
therapieën zoals alemtuzumab (anti-CD52). Hoewel een gerandomiseerde studie heeft
aangetoond dat patiënten boven een leeftijd van 65 jaar een betere overleving hebben met
fludarabine monotherapie ten opzichte van chloorambucil, is chloorambucil nog steeds de
meest toegepaste eerstelijnstherapie in deze populatiegroep [14].
Bestraling
Radiotherapie wordt vooral gebruikt als patiënten last hebben van pijnlijke lymfeklier- of
miltvergroting.
Stamceltherapie
Patiënten met een voldoende goede basisconditie, die nog steeds een progressief verloop
vertonen na de eerstelijnsbehandeling komen in aanmerking voor een allogene
stamceltransplantatie [15, 16]. Ook patiënten met een p53 deletie (del17p13) komen hiervoor
5
in aanmerking. Ze vertonen een korte overlevingstijd en zijn resistent tegen de
standaardbehandeling van chemotherapie [17].
Figuur 2: Overzicht van de CLL behandelingsstrategie [18].
2. B-cel ontwikkeling
In het beenmerg bevinden zich stromale cellen die, door middel van specifieke
adhesiemoleculen en de secretie van verschillende oplosbare factoren, de differentiatie en
proliferatie van de lymfocyt promoten. De B-cellen ondergaan daar een selectie om zelf-
tolerantie te bewerkstelligen en migreren vervolgens naar de periferie. Vervolgens circuleren
ze doorheen de perifere lymfoïde organen en kunnen ze geactiveerd worden door antigenen
(Figuur 3). Zowel dendritische cellen (DCs), T-cellen als B-cellen worden aangetrokken naar
de lymfoïde organen. De DCs die zich in de lymfeknoop bevinden, trekken via secretie van
chemokines, B- en T-cellen aan. De antigen specifieke B-cellen migreren vanuit de T-cel zone
naar het kiemcentrum van de lymfeknoop, waar somatische hypermutatie van de B-cel
receptor (BCR) plaatsvindt zodat deze een sterkere antigen affiniteit verkrijgt. Deze cellen
krijgen overlevingssignalen afkomstig van de T-cellen en kunnen zo hun belangrijke
immunologische functies uitvoeren De belangrijkste functie van de B-cel is de productie van
antilichamen. Daarnaast spelen B-cellen een rol als antigeenpresenterende cellen voor T-
cellen en kunnen ze ook verschillende cytokines produceren. [19].

6
Figuur 3: Algemeen overzicht van de stappen in de ontwikkeling van B-cellen. B-cellen worden eerst gedifferentieerd mede onder invloed van de stromale beenmergcellen via herschikking van de V, (D) en J segmenten van de zware en lichte keten van de B-cel receptor. Vervolgens worden ze geactiveerd door antigenen die ze tegenkomen tijdens circulatie door de perifere lymfoïde organen. De mature B-cel ondergaat ten slotte affiniteit maturatie via somatische hypermutatie van de variabele regio van de zware keten en zal zo transformeren tot antilichaam secreterende plasmacellen [19].
De B-cel zal na deze hypermutatie oplosbare immunoglobulines uitscheiden. Zowel de
transmembranaire BCR als de gesecreteerde immunoglobulines bestaan uit twee zware en
twee lichte aminozuurketens die samengehouden worden door disulfidebindingen. Elke keten
is samengesteld uit een variabele regio en een constante regio. De genen van het variabele
deel van de immunoglobulineketen worden onderverdeeld in de lichte keten segmenten (V en
J segmenten) en de zware keten segmenten (V, D en J segmenten). De grote diversiteit aan
immunoglobulines is te wijten aan de grote variabiliteit aan combinaties die mogelijk zijn
door herschikking van de zware en lichte ketens en hun segmenten (Figuur 4). Deze
variabiliteit wordt nog verrijkt door introductie van P- en N- nucleotiden tijdens deze VDJ
recombinatie en de somatische hypermutatie in de variabele regio van de zware keten.

7
Figuur 4: B-cellen worden gedifferentieerd via VDJ recombinatie van de zware en lichte ketens van de B-cel receptor. Dit fenomeen is mede verantwoordelijk voor de grote diversiteit aan immunoglobulines. (http://ittakes30.wordpress.com)
3. Prognostische merkers bij CLL
3.1. Algemeen
CLL heeft een sterk heterogeen ziekteverloop. Sommige patiënten hebben een korte
overlevingstijd, terwijl andere meer dan 15 jaar kunnen overleven. Een vroege start van de
behandeling kan van cruciaal belang zijn voor patiënten met een slechte prognose.
De klinische prognose werd voornamelijk bepaald met behulp van de classificatie volgens Rai
en Binet [20, 21]. Deze classificatie geeft een betrouwbare indeling van patiënten in
risicogroepen gebaseerd op klinische observaties en standaard laboratoriumtesten. Zo wordt
een vroeg stadium (Rai 0, Binet A), een intermediair stadium (Rai I/II, Binet B) en een
gevorderd stadium (Rai III/IV, Binet C) gedefinieerd met een gemiddelde overlevingstijd van
respectievelijk >10, 5-7 en 1-3 jaar [12]. De classificatie volgens Rai en Binet is eenvoudig en
gemakkelijk toepasbaar. Het grote nadeel echter is dat de progressie van de ziekte niet
voorspeld kan worden in het vroege stadium van de ziekte. Aangezien meer dan 80% van de
CLL patiënten gediagnosticeerd worden in het vroege stadium van de ziekte, wordt er veel
onderzoek verricht naar klinische en biologische merkers die de klassieke classificatie kunnen
vervangen of er additioneel aan zijn. Klinische risicofactoren van de patiënt zoals de leeftijd
en het geslacht worden in rekening gebracht [12]. Ook de tumoractiviteit is een goede
indicator voor de progressie van de ziekte en kan bepaald worden via bepaling van het aantal
lymfocyten, de lymfocytverdubbelingstijd en de beenmerginfiltratie.

8
Ook worden verschillende gekende serummerkers toegepast, zoals lactaat dehydrogenase
(LDH), β2 microglobuline en oplosbaar CD23. Ook onderzoek naar angiogene merkers,
proliferatiemerkers en cytogenetische merkers werd verricht [22]. Op genetisch gebied blijken
de genetische afwijkingen in de genen voor p53 (del17p13) en ATM (del11q22–23) een
goede indicator om de prognose te bepalen. De mutatiestatus van de variabele regio van het
IgVH gen, blijkt tot nu toe de gouden standaard om prognose te bepalen.
3.2 Genetische afwijkingen
De meest frequente chromosomale afwijking is de 13q deletie die optreedt bij ongeveer 55%
van de CLL patiënten. De minimal deleted regio (MDR) regio wordt hierbij verwijderd. Deze
regio bevat het DLEU2 gen en het eerste exon van DLEU1. Intron 4 van DLEU2 bevat twee
tumor suppressorgenen die miRNA 15-a en miRNA 16-1 tot expressie brengen [23] (
Figuur 5). Deze veroorzaken post-transcriptionele gensilencing van anti-apoptitische genen
zoals Bcl-2 [24].
Figuur 5: Schematische representatie van de 13q14 locus waarvan de MDR regio frequent verwijderd is bij CLL patiënten. Deze regio bevat onder andere 2 tumor suppressorgenen, miRNA 15a en miRNA 16-1 [25].
Downregulatie van deze miRNAs werd eveneens waargenomen in cellen zonder 13q14
deletie waaruit men suggereerde dat deze genen eventueel epigenetisch onderdrukt worden
[26]. Bovendien zijn er vermoedelijk additionele genetische elementen aanwezig in deze
MDR regio die leiden tot een meer agressief ziekteverloop [27].
In tegenstelling tot dragers van deze 13q14 deletie, hebben patiënten met een 17p13 deletie
een uitgesproken slechte prognose. Hierbij treedt er een deletie op in het p53 gen. Ongeveer
20% van de CLL patiënten heeft een deletie in 11q22-q23 waarin het ATM gen gelokaliseerd
is. Deze mutatie leidt tot een matige prognose. De aanwezigheid van een trisomie 12 in CLL

9
cellen correleert eveneens met een intermediaire prognose. Deze trisomie wordt in ongeveer
10-25% van de CLL patiënten waargenomen [28].
3.3 IgVH mutatiestatus
Op basis van aanwezigheid van mutaties in de variabele regio van het IgVH gen, kan CLL
ingedeeld worden in twee klinische subgroepen [29]. Deze indeling is tot nu toe de gouden
standaard om de prognose te bepalen. Patiënten met CLL cellen die de somatische
hypermutatie niet hebben ondergaan, vertonen een slechtere prognose. Bij deze patiënten is
het belangrijk om de therapie zo vroeg mogelijk te starten [30]. Sequentieanalyse van het
IgVH gen is echter zeer tijdrovend en duur. Door middel van een klonaliteitsanalyse worden
de monoklonale B-cellen geselecteerd. Polyklonale B-cellen hebben elk een specifieke IgVH
lengte door een verschillende VDJ recombinatie en toevoeging van N- en P- nucleotiden.
Monoklonale B-cellen hebben één identieke IgVH sequentie met dezelfde lengte. Na
opzuivering van het geselecteerde IgVH gen wordt een sequenering en een alignment
uitgevoerd ten opzichte van het best gerelateerde kiemlijn IgVH gen. Aan de hand van de
homologie wordt de patiënt geclassificeerd als gemuteerde CLL met goede prognose of
ongemuteerde CLL met slechte prognose [31].
3.4 Surrogaatmerkers voorde IgVH mutatiestatus
Omdat bepaling van de mutatiestatus van het IgVH duur en tijdrovend is, wordt er veel
onderzoek verricht naar surrogaatmerkers voor de mutatiestatus. Onderzoek toonde aan dat bij
ongemuteerde CLL cellen een verhoogde expressie optreedt van CD38 [29], ZAP-70 [32-34]
en lipoproteïne lipase LPL [35].
3.4.1. CD38
CD38 is een multifunctioneel proteïne dat de hydrolyse van NAD+ tot cyclisch ADP-ribose
katalyseert. Daarnaast heeft dit transmembranair glycoproteïne een functie als receptor en is
het belangrijk bij celadhesie via binding van het CD31 ligand dat onder andere tot expressie
gebracht wordt door nurse-like cellen (NLC) en stromale cellen [36]. CD38+ CLL cellen
vertonen een hogere proliferatiesnelheid en migratiepotentieel. Recent onderzoek toonde
tevens aan dat deze CD38 ligatie, tyrosinefosforylatie van ZAP-70 veroorzaakt. Dit impliceert
een mogelijke functionele link tussen CD38 en ZAP-70 [37].

10
3.4.2. ZAP-70
ZAP-70 is een intracellulair tyrosine kinase dat tot expressie komt in T-cellen en natural killer
(NK) cellen [38]. Na T-cel receptor (TCR) activatie door onder andere antigen presenterende
cellen, worden Immunoreceptor Tyrosine-based Activation Motifs (ITAMs) gefosforyleerd
Deze ITAMs zijn geconserveerde sequenties in het intracellulaire deel van de TCR. Na
binding van ZAP-70 op deze ITAMs wordt een signaalcascade in de cel geïnduceerd.
Verschillende transcriptiefactoren worden geactiveerd zoals Nuclear Factor κB (NFκB),
Nuclear Factor of Activated T-cells (NFAT) en Activator Protein 1 (AP-1). Dit leidt tot
proliferatie en differentiatie van de T-cellen. B-cellen vertonen normaal geen ZAP-70
expressie maar wel expressie van Syk tyrosine kinase, een ZAP-70 homoloog. In CLL cellen
van patiënten met een slechte prognose, wordt wel een constante expressie van ZAP-70
waargenomen met een BCR-geïnduceerde signaalcascade tot gevolg [32, 33] (Figuur 6).
Figuur 6: Potentiële functie van ZAP-70 na BCR activatie in de CLL cel. CLL cellen die ZAP-70 tot expressie brengen, versterken de signalisatie na BCR activatie. Syk, BLNK, PLCγ, MAPK, PKC en NFκB zijn potentiële targets die gestimuleerd zouden kunnen worden door ZAP-70. Er wordt gesuggereerd dat ZAP-70 een verhoogde fosforylatie van Syk of een verhoogde Ca2+ influx zou kunnen veroorzaken [39].
3.4.3. MicroRNAs
MicroRNAs (miRNAs) zijn kleine (20-25 neocleotiden lang) niet coderende RNA moleculen
die post-transcriptioneel interfereren met de genexpressie van bepaalde genen. Ze worden
geïncorporeerd in het RNA-induced silencing complex (RISC) en binden vervolgens
complementaire sequenties van bepaalde mRNAs. Enerzijds kan dit mRNA geknipt worden
door argonaut, de katalytische component van RISC. Anderzijds toonden Mathonnet et al. aan
dat micro-RNAs eveneens de translatie kan inhiberen [40].
Dit proces maakt deel uit van de RNA interferentie pathway. De expressie van miRNAs werd
geanalyseerd en vergeleken tussen CLL patiënten met goede en slechte prognose. Hieruit

11
werd geconcludeerd dat een expressie “signature” van 13 miRNAs gerelateerd is met de
prognose van CLL. Dat expressieprofiel bevat onder andere de miRNAs, miR-15a en miR-16-
1 die, zoals reeds vermeld, het anti-apoptotisch eiwit Bcl-2 als target hebben [41]. Bij
verlaagde expressie van deze miRNAs zal Bcl-2 dus een hogere expressie vertonen,
geassocieerd met een slechtere prognose. Ook miR29c wordt neergereguleerd bij agressieve
vormen van CLL. De proto-oncogenen TCL-1 en MCL-1 zijn targets van dit miRNA en
vertonen bijgevolg hogere expressie [42].
3.4.4. Lipoproteïne lipase
De validiteit van bovenstaande prognostische factoren, CD38 en ZAP-70, wordt in vraag
gesteld door controversiële resultaten. Onderzoek in verschillende laboratoria gaven
verschillende resultaten voor deze twee merkers [48]. Daarnaast heeft analyse van ZAP-70
expressie met behulp van flowcytometrie enkele nadelen. Ten eerste moet bij gebruik van
deze techniek de integriteit van de celmembraan verstoord worden om een kleuring mogelijk
te maken. Een tweede nadeel is dat voorafgaande purificatie van de mononucleaire cellen van
cruciaal belang is aangezien ZAP-70 in grotere hoeveelheden aanwezig is in T- en NK cellen.
LPL expressie kan daartegen gemeten worden in ongezuiverde perifere mononucleaire cellen
en blijkt een meer betrouwbare merker te zijn om een onderscheid te maken tussen
gemuteerde en ongemuteerde CLL cellen in een vergevorderd stadium [43].
LPL is een eiwit bestaande uit 448 aminozuren met een ongekende kristalstructuur. De
modelstructuur werd wel gedemonstreerd aan de hand van het “molecular modeling system
INSIGHT II version 2000” (Accelrys Inc., Burlington, MA, USA). Dit model toont twee
domeinen, namelijk een N-terminaal domein en een C-terminaal domein. Het N-terminaal
domein is essentieel voor de katalytische activiteit van het enzyme en kan zowel in een open
en gesloten vorm voorkomen. LPL katalyseert de hydrolyse van de triacylglycerol (TAG)
component van circulerende chylomicronen en very low densitylipoproteins (VLDLs) tot
niet-veresterde vetzuren en 2-monoacylglycerol (Figuur 7). LPL is verankerd aan de luminale
zijde van de capillaire endotheelcellen via binding aan heparan sulfaat-proteoglycaan (HSPG)
ketens en bindt de circulerende lipoproteïnen via interactie met lipoproteïne receptoren zoals
LDL receptor related protein (LRP), LDL receptor, GP330/LRP2 en VLDL receptor. Het
apolipoproteïne C-II brengt de koppeling tussen LPL en de chylomicronen tot stand en heeft
een positieve invloed op de activiteit van LPL. Lipoproteïne partikels zijn te groot om door de
capillairen te diffunderen waardoor, via deze interactie, de gehydrolyseerde TAG producten

12
getransfereerd kunnen worden over het capillaire endothelium om verschillende weefsels
ervan te voorzien. Naast zijn katalytische functie heeft LPL ook een brugfunctie die niet
alleen een belangrijke rol speelt bij de opname van lipiden door weefsels [44]. Zo werd
aangetoond dat LPL de adhesie van cellen, zoals monocyten [45], aan endotheelcellen
promoot. LPL is vooral aanwezig in het hart-, spier- en vetweefsel en de melk producerende
borstklier. Lagere hoeveelheden worden geproduceerd door macrofagen, hormoon
producerende cellen van de ovaria en de bijnieren, thoracale aorta, milt, nier, testis en de long.
Figuur 7: Lipoproteïne lipase als brugvormend proteïne en als katalysator. LPL is verbonden met capillaire endotheelcellen via heparan sulfaat-proteoglycaan en bindt chylomicronen of VLDL partikels via specifieke receptoren en apolipoproteïne C-II. Via deze interactie wordt hydrolyse van triacylglycerol tot vrije vetzuren en 2-monoacylglycerol gekatalyseerd waarna deze reactieproducten migreren naar de verschillende weefsels [46].
3.5 Hypothetisch werkingsmechanisme van LPL in CLL
LPL expressie is positief gecorreleerd met verschillende andere prognostische factoren: de
IgVH mutatiestatus, CD38 expressie en ZAP-70 expressie [47]. Recent werd aangetoond dat
inductie van LPL optreedt door stimulatie van de BCR in CLL cellen in tegenstelling tot
normale B-cellen. CLL cellen van patiënten met de ongemuteerde IgVH status vertonen zelfs
verhoogde BCR autoreactiviteit en dus LPL expressie zonder stimulatie van de BCR [48].
De gemiddelde enzymatische activiteit van LPL is dan weer lager bij CLL patiënten met de
ongemuteerde status, in vergelijking met CLL patiënten waarbij het IgVH gen gemuteerd is.
Hieruit kan men suggereren dat LPL eerder een brugfunctie vervult in CLL cellen dan een
enzymatische functie [49].
Hypothetisch gezien zou LPL de interactie van de CLL cel met de micro-omgeving, die een
belangrijke functie in CLL vervult, kunnen versterken. De micro-omgeving wordt
gedefinieerd als het geheel van de extracellulaire matrix (ECM), stromale cellen en het

13
conventioneel milieu met cytokines en groeifactoren. De ECM bevat onder andere collageen,
laminine, fibronectine, vitronectine, tenascine en proteoglycanen. CLL cellen interageren met
verschillende celtypes in hun directe omgeving in de lymfoïde weefsels waardoor ze de
nodige overlevings- en proliferatiesignalen krijgen [50]. Veel onderzoek werd verricht om
deze interacties, die niet optreden bij normale B-cellen, te bestuderen. In het beenmerg komen
de monocyten in contact met de stromale beenmergcellen [51] en de folliculaire DCs [52]. In
de lymfeknopen zijn de CD4+ T-helper cellen dominant aanwezig en deze secreteren
cytokines (o.a. IL-4, INF-α en -γ) die CLL cel apoptose inhiberen. Tenslotte werd aangetoond
dat een subpopulatie van de CLL cellen, afkomstig uit het perifere bloed, in vitro
differentieert tot adherente NLCs. Deze beschermen eveneens de CLL cellen tegen spontane
apoptose in cultuur.
CLL cellen brengen verschillende adhesiemoleculen op hun oppervlak tot expressie die de
overleving van de CLL cel promoten. CLL cellen adhereren aan stromale cellen in de micro-
omgeving doordat de integrines CD49d/CD11a of CD11b/CD18 interageren met CD54 en
CD106, aanwezig op stromale cellen. Deze adhesie induceert bescherming tegen spontane
apoptose. De adhesiemolecule CD44 heeft eveneens een rol in de CLL overleving. CLL
cellen van patiënten met een agressief subtype vertonen een sterke CD44 expressie, wat
suggereert dat CD44 mogelijks een rol zou kunnen spelen in de CLL pathogenese en
prognose. Ten slotte kunnen CLL cellen ook via binding aan fibronectine en VCAM-1,
liganden gelokaliseerd in het endothelium, van de nodige overlevingssignalen voorzien
worden.
4. Doelstelling
Het verloop van de biosynthese en de functie van LPL bij CLL patiënten is nog steeds
ongekend. Dit onderzoek heeft als doel om inzicht te verwerven in de functie van deze
veelbelovende prognostische merker in de pathogenese van CLL. Dit kan eventuele nieuwe
therapeutische mogelijkheden aantonen en kan leiden tot een optimalisatie van de
hedendaagse behandelingen die te duur zijn en nog geen volledige genezing kunnen bieden.
LPL blijkt immers een goede target te zijn voor therapie. Onderzoek toonde aan dat
blootstelling van CLL cellen aan stijgende concentraties orlistat, apoptose veroorzaakt [48].
Orlistat is een effectieve inhibitor van LPL [53] en wordt gebruikt als geneesmiddel bij
obesitas.

14
In dit onderzoek werd een efficiënte manier gezocht om LPL binnen te brengen in LPL
negatieve CLL cellen om op deze manier de functie na te gaan. De opname van vreemd DNA
door eukaryote cellen, kan gebeuren aan de hand van verschillende technieken.
Zo bestaan er chemische transfectiemethoden waarbij DNA en een agens in de cel worden
gebracht, fysische methoden en transductiemethoden via virussen. In eerdere studies werd
mRNA van de prognostische merker, ZAP-70, succesvol binnengebracht in CLL cellen via
elektroporatie [54]. Bij deze methode wordt de celmembraan tijdelijk gepermeabiliseerd door
het toedienen van een kortstondig, high-voltage puls zodat het mRNA de cel kan
binnentreden. In deze thesis werd nagegaan of mRNA elektroporatie eveneens een efficiënte
methode is om het LPL eiwit in de CLL cel binnen te brengen. Het finaal doel blijft om aan
de hand van deze LPL+ en LPL- CLL cellen te vergelijken en zo de precieze functie van LPL
in de CLL cel na te gaan.

15
II. Materialen en methoden
1. Stimulatie experiment
1.1 Selectie van de patiënten
De patiënten worden geselecteerd op basis van de laboratoriumresultaten. De CLL patiënt
heeft per definitie minstens 5000 klonale B-cellen/µL in het perifeer bloed die een rijpcellige
cytomorfologie en expressie van antigenen, CD5 en CD23 op hun oppervlak vertonen. Er
worden stalen van patiënten geselecteerd met een gekende mutatiestatus en aan- of
afwezigheid van LPL. Dit onderzoek werd goedgekeurd door het ethisch comité van het UZ
Gent en de patiënten ondertekenden het informed consent.
1.2 Isolatie en stimulatie
Er werd gewerkt met ingevroren perifere bloed mononucleaire cellen (PBMCs) van twee CLL
patiënten die op een eerder tijdstip werden geïsoleerd uit verse bloedstalen. Patiënt 1 (P1) is
gemuteerd en negatief voor LPL. Patiënt 2 (P2) is ongemuteerd en positief voor LPL.
Bloedstolling van de verse stalen wordt bij afname verhinderd door EDTA, een
metaalchelator die onder andere Ca2+ verwijdert uit het bloed, een belangrijke cofactor voor
het stollingsproces. Isolatie van de PBMCs uit deze verse bloedstalen gebeurt als volgt: het
bloed (8ml) wordt 2X verdund in Iscove’s Modified Dulbecco’s Medium (IMDM,
Invitrogen, Merelbeke, België). Hierbij wordt 3 ml “lymphoprep” (Nycomed, Oslo,
Noorwegen) toegevoegd dat ervoor zorgt dat de suspensie, na centrifugatie (15’, 2350 rpm,
22°C), gescheiden wordt in vier fasen. De mononucleaire cellen worden afgezonderd in een
fase die voorzichtig wordt opgezogen en overgebracht naar een falconbuis. De cellen worden
2X gewassen met 8 ml IMDM medium. Vervolgens worden de cellen geteld via de “dye
exclusion test” met de Bürkertelkamer. Levende en dode cellen worden van elkaar
onderscheiden via trypaanblauw (Invitrogen, Merelbeke, België). Deze kleurstof wordt enkel
opgenomen door dode cellen die bijgevolg blauw kleuren, in tegenstelling tot de levende
cellen. De geïsoleerde cellen worden uiteindelijk ingevroren in DMSO + 10% FCS (Hyclone
Perbio, Erembodegem, België) aan een concentratie van 100 x106 cellen/ml.
Ontdooien gebeurt snel in een warm waterbad. De celsuspensie wordt gradueel verdund in
koud IMDM complete (IMDM met 10% FCS, 1% penicilline en strepomycine; Invitrogen,
Merelbeke, België), waarna de cellen worden afgedraaid en geresuspendeerd in warm IMDM

16
complete. De cellen worden tenslotte geteld en in cultuur gebracht aan 10 x 106 cellen/ml in
een 24-well plaat. Per patiënt worden 40 x 106 cellen in 4 ml verdeeld over 4 reactievaatjes.
IgM stimulatie van de cellen wordt uitgevoerd door toevoeging van 10 µl anti-humaan IgM
Immunobeads (Irvine Scientific, California, VS) per 10 x 106 cellen. Als negatieve controle
worden anti-humaan IgA Immunobeads gebruikt. Aangezien CLL cellen geen IgA op hun
oppervlak dragen, zullen de beads bijgevolg niet binden op deze cellen en geen stimulatie
veroorzaken. De 24-well plaat wordt vervolgens in de incubator geplaatst bij 37°C en 7%
CO2. Cellen worden geoogst en geanalyseerd 3 uur en 24 uur na stimulatie.
1.3 Oogsten van de cellen en flowcytometrie
Na 3 uur of 24 uur stimulatie wordt een deel van de cellen gekleurd voor flowcytometrische
analyse. De rest van de cellen wordt geoogst . De cellen worden gecentrifugeerd (10’, 13000
rpm) en gewassen met 500 µl PBS. Na centrifugatie (5’, 13000 rpm) worden de cellen in 1 ml
Trizol gebracht.
Flowcytometrie wordt gebruikt om verschillende oppervlakte- en/of intracellulaire merkers
van CLL cellen in suspensie te detecteren en te kwantificeren. De techniek is gebaseerd op het
principe van de hydrodynamische focussering van deeltjes in suspensie. Er worden specifieke
moleculen, meestal monoklonale antilichamen, toegevoegd. Deze zijn al dan niet gelabeld
met fluorochromen, die specifiek binden aan de oppervlaktemerkers of na permeabilisatie van
de membraan, aan specifieke intracellulaire merkers. Fluorochromen absorberen
hoogenergetische fotonen en emitteren energie onder de vorm van licht bij het terugvallen
naar de energetische grondtoestand. Deze fluorescentie wordt bij elke cel afzonderlijk
geanalyseerd doordat de celsuspensie door een capillaire naald gefocusseerd wordt. De
fluorescentie en de verstrooiing van het licht worden gedetecteerd en vervolgens
computationeel verwerkt. De forward scatter (FSC) meet de voorwaartse lichtverstrooiing en
bepaalt de relatieve verschillen in grootte tussen de geanalyseerde cellen. De side scatter
(SSC) meet de zijwaartse lichtverstrooiing die wordt bepaald door aanwezige granules in het
cytoplasma en door de complexiteit van de kernvorm.
In dit onderzoek wordt de aandacht gericht op de expressie van CD19 (B-cel merker), CD3
(T-celmerker) en LPL. Er wordt zowel gekeken naar de intracellulaire LPL expressie als naar
de LPL expressie op het oppervlak van de cellen.
Er worden 100.000 cellen overgebracht in een FACS-tube en vervolgens gewassen met
humane wasbuffer ((PBS)-BSA-Na3N). Bij een intracellulaire kleuring worden de cellen eerst

17
gefixeerd en vervolgens gepermeabiliseerd met behulp van de Fix&Perm Cell
Permeabilization kit (ImTec diagnostic, Antwerpen, België). Vervolgens worden de cellen
geïncubeerd met 1,5 µl ongelabeld muis anti-LPL antilichaam [LPL.A4] (Abcam, Cambridge,
VK). Hierna wordt het secundaire phycoerythrin (PE) gelabelde geit anti-muis antilichaam
(Caltage Laboratories, Buckingham, VK) toegevoegd. Ten slotte wordt een
oppervlaktekleuring uitgevoerd voor de B-cel of T-cel specifieke oppervlaktemerkers met
anti-CD19-APC en anti-CD3-FITC ( BD Biosciences, San josé, California, USA).
De cellen worden geanalyseerd op de LSRII of FACSCanto II flowcytometer (BD
Biosciences, België). Om na te gaan of stimulatie gelukt is, kan er eventueel een kleuring met
anti-CXCR4-PE (BD pharmingen, San Diego, California, USA) uitgevoerd worden.
Indien enkel de expressie van bovenstaande moleculen op het oppervlak geanalyseerd wordt,
wordt er geen fixatie en permeabilisatiestap uitgevoerd. Bovendien wordt bij elk staal
propidiumiodide toegevoegd om de viabiliteit van de cellen na te gaan.
1.4 Isolatie RNA
RNA isolatie wordt uitgevoerd met de RNAeasy mini kit (Qiagen, Hilden, Duitsland). De
isolatie gebeurt via een trizol gebaseerde methode. Aan de cellen in trizol (1 ml) wordt 140 µl
chloroform (Sigma-Aldrich, Bornem, België) toegevoegd. Na centrifugatie (15’, 13000 rpm,
4°C) ontstaan er drie fasen: een bovenste kleurloze fase die het RNA bevat, een tussenfase die
het DNA bevat en een onderste lichtroze organische fase die trizol, chloroform en eiwitten
bevat. De doorzichtige fase, die het RNA bevat, wordt geïsoleerd om hier vervolgens een 1,5
keer zo groot volume 100% EtOH (Chemical VWR Prolabo, Leuven, België) aan toe te
voegen (+/- 650 µl).
Het RNA-mengsel wordt op de RNeasy mini spin kolom gebracht en na een shortspin wordt
de flow-through verwijderd. Vervolgens wordt de kolommembraan, waaraan het RNA
gebonden is, gewassen met RWT buffer en RPE buffer. Tenslotte wordt het RNA geëlueerd
van de kolom met 30 µl RNase-vrij water. De concentratie van het eluaat wordt bepaald via
de Nanodrop 1000 Spectrofotometer (Isogene Life Science, St-Pieters-Leeuw, België).

18
1.5 cDNA synthese
Er wordt gebruik gemaakt van SuperScript III First-Strand Synthesis System kit (Invitrogen,
Merelbeke, België), waarbij het RNA wordt omgezet tot cDNA met behulp van reverse
transcriptase. Bij 1 µg RNA worden 1 µl random primers (250 µg/ml) en 1 µl dNTP mix (10
mM) gevoegd. Deze mix wordt aangelengd met nuclease vrij water (NF-H2O; Ambion,
Lennik, België) tot een totaalvolume van 13 µl en 5’ bij 65°C en 3’ bij 4°C gebracht in de
Thermalcycler AB 2720 (Applied Biosystems). Daarna wordt er bij elk staal 4 µl 5 x first
strand buffer, 1 µl RNase OUT inhibitor, 1 µl 0.1M DTT en 1 µl superscript III RT
toegevoegd. Dit mengsel (20µl) wordt vervolgens opnieuw in de Thermalcycler AB2720
gebracht (programma: 5’ bij 25°C, 50’bij 50°C, 15’ bij 70°C) (Applied Biosystems).
Tenslotte wordt het cDNA verdund met 30 µl NF-H2O (Ambion, Lennik, België).
1.6 Kwantitatieve qPCR
1.6.1 FOS en LPL
Om de LPL en FOS expressie te kwantificeren wordt een qPCR uitgevoerd gebruik makende
van de TaqMan-technologie. Hierbij wordt specifiek het target cDNA uit de verschillende
stalen geamplificeerd en gekwantificeerd met behulp van specifieke primers en taqmanprobes.
Een specifieke taqmanprobe bindt de DNA moleculen en is gelabeld met een fluorescente
reportermolecule aan het 5’ einde en een quencher aan het 3’ einde. Als de oligonucleotide
probe intact is en vrij in de oplossing voorkomt, zal de quencher het fluorescentiesignaal van
de reporter onderdrukken. Eenmaal de probe specifiek bindt op het DNA, hydrolyseert het taq
polymerase de probe en is de quencher niet meer in staat de reportermolecule te onderdrukken
met fluorescentie tot gevolg (Figuur 8B).

19
Figuur 8: Principe qPCR via (A) SYBR green detectie en (B) detectie via de taqmanprobe. (A) Stap 1: Na elke amplificatiecyclus, intercaleert het fluorescente SYBR green aspecifiek het geamplificeerde dsDNA. Stap 2: Bij aanvang van de volgende amplificatiecyclus wordt dsDNA gedenatureerd waardoor de fluorescentie drastisch daalt. Stap 3 en 4: Na polymerisatie stijgt opnieuw het fluorescentiesignaal door dsDNA binding van SYBR green. (B) Stap 1: De taqmanprobe bindt specifiek op de DNA sequentie en heeft dus een hogere betrouwbaarheid. De probe is gelabeld met enerzijds een fluorofoor reportermolecule aan het 5’ eind (groene R) en anderzijds met een quencher (grijze Q) aan het 3’ eind. Stap 2: De probe is intact waardoor de quencher, fluorescentie via de reportermolecule onderdrukt. Stap3: Na cleavage van de probe door polymerisatie van DNA, kan de quencher de reportermolecule niet meer onderdrukken met fluorescentie tot gevolg. (Aangepast van http://www.appliedbiosystems.com)
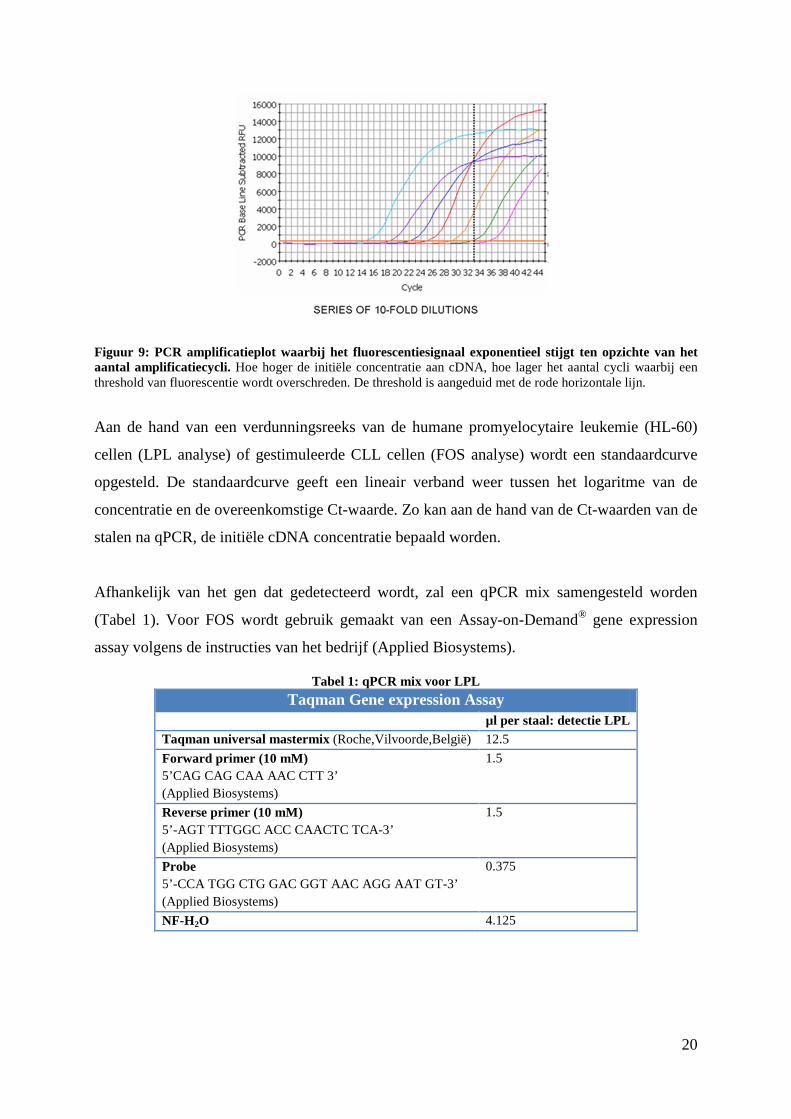
Bijgevolg stijgt het fluorescentiesignaal exponentieel met het aantal cycli die doorlopen zijn.
Het aantal cycli dat nodig is om een bepaalde threshold van fluorescentie te overschrijden (Ct-
waarde), wordt gebruikt om te analyseren hoeveel LPL cDNA er oorspronkelijk in het staal
aanwezig was. Hoe meer cycli nodig zijn om deze threshold te overschrijden, hoe minder
cDNA er oorspronkelijk aanwezig was (Figuur 9).
20
Figuur 9: PCR amplificatieplot waarbij het fluorescentiesignaal exponentieel stijgt ten opzichte van het aantal amplificatiecycli. Hoe hoger de initiële concentratie aan cDNA, hoe lager het aantal cycli waarbij een threshold van fluorescentie wordt overschreden. De threshold is aangeduid met de rode horizontale lijn.
Aan de hand van een verdunningsreeks van de humane promyelocytaire leukemie (HL-60)
cellen (LPL analyse) of gestimuleerde CLL cellen (FOS analyse) wordt een standaardcurve
opgesteld. De standaardcurve geeft een lineair verband weer tussen het logaritme van de
concentratie en de overeenkomstige Ct-waarde. Zo kan aan de hand van de Ct-waarden van de
stalen na qPCR, de initiële cDNA concentratie bepaald worden.
Afhankelijk van het gen dat gedetecteerd wordt, zal een qPCR mix samengesteld worden
(Tabel 1). Voor FOS wordt gebruik gemaakt van een Assay-on-Demand® gene expression
assay volgens de instructies van het bedrijf (Applied Biosystems).
Tabel 1: qPCR mix voor LPL
Taqman Gene expression Assay µl per staal: detectie LPL Taqman universal mastermix (Roche,Vilvoorde,België) 12.5
Forward primer (10 mM) 5’CAG CAG CAA AAC CTT 3’ (Applied Biosystems)
1.5
Reverse primer (10 mM) 5’-AGT TTTGGC ACC CAACTC TCA-3’ (Applied Biosystems)
1.5
Probe 5’-CCA TGG CTG GAC GGT AAC AGG AAT GT-3’ (Applied Biosystems)
0.375
NF-H2O 4.125

21
Aan 20 µl qPCR mix wordt 5 µl cDNA toegevoegd. Vervolgens ondergaat deze mix
verschillende cycli van amplificatie. Dit gebeurt bij volgende condities: 10’ bij 95°C en 40
cycli van: 15” bij 95°C en 1’ bij 60°C (7300 Real Time PCR system, Applied Biosystems,
Foster City, CA, USA).
1.6.2 Actine
Om de LPL en FOS expressie te normaliseren wordt ook de expressie van een huishoudgen,
in dit geval actine, bepaald. Huishoudgenen komen in de cellen constitutief tot expressie. Als
probe wordt gebruik gemaakt van Sybr Green (Eurogentec, Luik, België). Sybr Green bindt
op het dubbelstrengig DNA gevormd na de elongatiestap van de primers en dit
fluorescentiesignaal wordt gemeten (Figuur 8A)
Bovendien is het bij Sybr Green mogelijk om de smelttemperatuur van de gevormde
dubbelstrengige DNA moleculen te bepalen. Via verhitting denatureren dsDNA moleculen en
daalt bijgevolg het fluorescentiesignaal. Twee verschillende DNA sequenties hebben
verschillende lengtes waardoor de daling in fluorescentiesignaal bij verhitting een
verschillend verloop kent. Indien er dus meerdere verschillende smeltcurves aangetoond
worden na qPCR, heeft er een aspecifieke amplificatie plaatsgevonden zoals bijvoorbeeld
gebeurt door mispriming, primer-dimeer artefacten en contaminatie.
De qPCR voor actine wordt als volgt samengesteld (Tabel 2):.
Tabel 2 qPCR mix voor actine
qPCR Core Kit voor SYBR green (Eurogentec,Luik,België) µl per staal: detectie actine 2xmastermix 12.5
Forward primer (10µM) [55] 5’-TGA CCC AGA TCA TGT TTG AGA-3’
0.75
Reverse primer (10 µM) [55] 5’AGA GGC GTA CAG GGA TAG CA-3’
0.75
SYBR green I 0.75
NF-H2O 5.25
Aan 15 µl qPCR mix wordt 5 µl cDNA toegevoegd. Vervolgens ondergaat deze mix
verschillende cycli van amplificatie. Dit gebeurt bij volgende condities: 10’ bij 95°C en 40
cycli van 15” bij 95°C en 1’ bij 60°C. De smeltcurve wordt eveneens toegevoegd (7300 Real
Time PCR system, Applied Biosystems, Foster City, CA, USA).
22
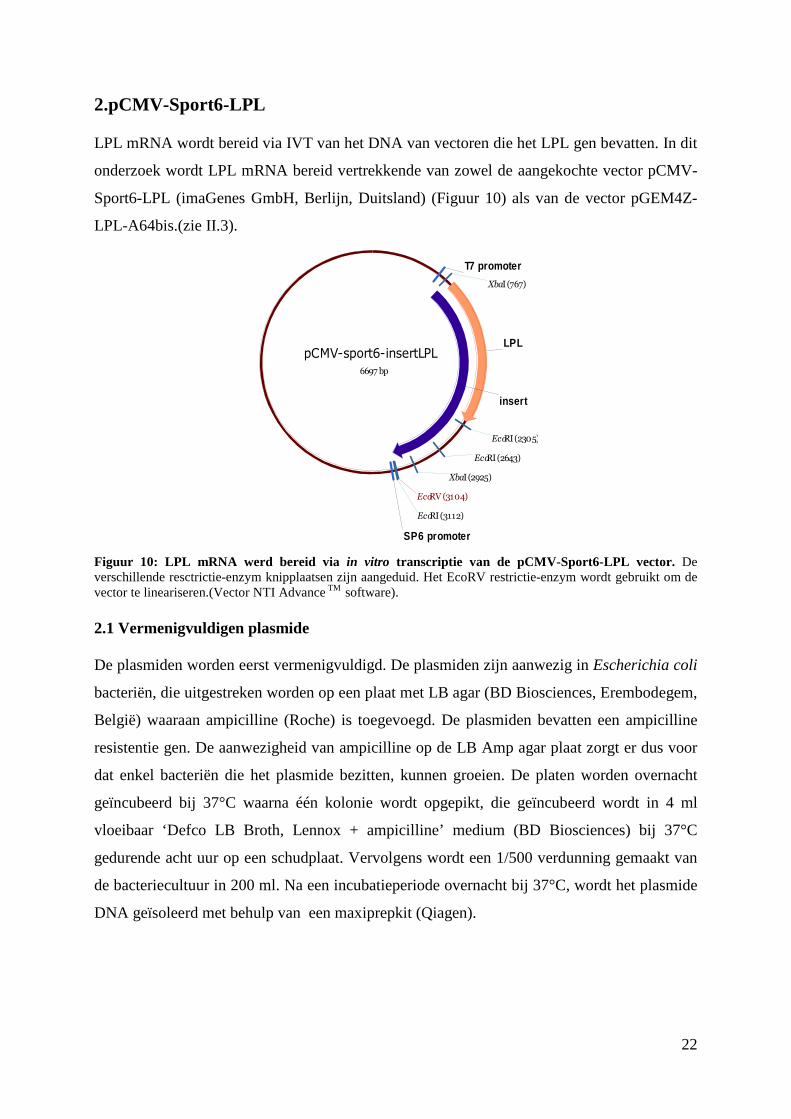
2.pCMV-Sport6-LPL
LPL mRNA wordt bereid via IVT van het DNA van vectoren die het LPL gen bevatten. In dit
onderzoek wordt LPL mRNA bereid vertrekkende van zowel de aangekochte vector pCMV-
Sport6-LPL (imaGenes GmbH, Berlijn, Duitsland) (Figuur 10) als van de vector pGEM4Z-
LPL-A64bis.(zie II.3).
Figuur 10: LPL mRNA werd bereid via in vitro transcriptie van de pCMV-Sport6-LPL vector. De verschillende resctrictie-enzym knipplaatsen zijn aangeduid. Het EcoRV restrictie-enzym wordt gebruikt om de vector te lineariseren.(Vector NTI Advance TM software).
2.1 Vermenigvuldigen plasmide
De plasmiden worden eerst vermenigvuldigd. De plasmiden zijn aanwezig in Escherichia coli
bacteriën, die uitgestreken worden op een plaat met LB agar (BD Biosciences, Erembodegem,
België) waaraan ampicilline (Roche) is toegevoegd. De plasmiden bevatten een ampicilline
resistentie gen. De aanwezigheid van ampicilline op de LB Amp agar plaat zorgt er dus voor
dat enkel bacteriën die het plasmide bezitten, kunnen groeien. De platen worden overnacht
geïncubeerd bij 37°C waarna één kolonie wordt opgepikt, die geïncubeerd wordt in 4 ml
vloeibaar ‘Defco LB Broth, Lennox + ampicilline’ medium (BD Biosciences) bij 37°C
gedurende acht uur op een schudplaat. Vervolgens wordt een 1/500 verdunning gemaakt van
de bacteriecultuur in 200 ml. Na een incubatieperiode overnacht bij 37°C, wordt het plasmide
DNA geïsoleerd met behulp van een maxiprepkit (Qiagen).
pCMV-sport6-insertLPL
6697 bp
LPL
insert
T7 promoter
SP6 promoter
EcoRV (3104)
XbaI (767)
XbaI (2925)
EcoRI (2305)
EcoRI (2643)
EcoRI (3112)

23
2.2 Isolatie plasmide DNA
Om het plasmide DNA te isoleren, wordt geopteerd voor een maxiprep. Dit geeft een grotere
opbrengst en dus een voldoende hoge concentratie om IVT mogelijk te maken. Hiervoor
wordt gebruik gemaakt van de maxiprepkit volgens de instructies van het bedrijf (Qiagen,
Venlo, Nederland, zie ‘QIAGEN® Plasmid Purification Handbook’ voor protocol)
De concentratie wordt bepaald via de Nanodrop1000 Spectrofotometer (Isogen Life Science,
St-Pieters-Leeuw, België).
2.3 Digest en opzuivering
De pCMV-Sport6-LPL vector wordt gelineariseerd met behulp van het restrictie-enzyme
EcoRV (New England Biolabs), dat uniek knipt buiten het LPL gen. Er wordt 20 µg plasmide
toegevoegd aan 10X NEbuffer 3, 3 µl EcoRV, 1 ul 100X Bovine Serum Albumine (BSA) en
dit wordt aangelengd met NF-H2O tot een totaalvolume van 100 µl. Dit mengsel wordt
geïncubeerd bij 37°C voor een periode van 3 uur.
Het digest wordt vervolgens opgezuiverd via de QIAquick PCR Purification kit (Qiagen,
Venlo, Nederland). Aan het digest wordt 500 µl PB buffer toegevoegd. Dit wordt op de
QiaQuick kolom gebracht en gewassen met 750 µl PE buffer. De gelineariseerde vector wordt
geëlueerd met 30 µl NF-H2O. Hiervan wordt de concentratie bepaald via de Nanodrop1000
Spectrofotometer (Isogen Life Science, St-Pieters-Leeuw, België).
2.4 In vitro transcriptie
Vervolgens wordt het construct in vitro overgeschreven via de mMessage mMachine T7 Ultra
kit (Invitrogen, Merelbeke, België). Deze kit bevat een T7 polymerase dat LPL overschrijft
vanaf de T7 promotor. Hiervoor worden 1 µg lineair DNA, 10 µl 2X T7 NTP/ARCA, 2 µl
10X T7 reactiebuffer en 2 µl enzymmix samengevoegd en aangelengd met NF-H2O tot een
totaal volume van 20 µl. Dit wordt gedurende 2u30 geïncubeerd bij 37°C. Daarna wordt het
resterende DNA verwijderd door toevoeging van 1 µl TURBO DNase en dit wordt
15’geincubeerd bij 37 °C.
Het LPL mRNA, afgeschreven van de pCMV-Sport6-LPL vector, bevat nog geen polyA-
staart. Met behulp van het polyA-polymerase worden een reeks adenosine nucleotiden aan het
3’ einde van het LPL mRNA geligeerd. De polyA-staart is noodzakelijk voor de stabiliteit van
het mRNA en voor een goede terminatie van de translatie in eukaryote cellen. Verschillende

24
reagentia worden samengevoegd in de volgende volgorde: 20 µl IVT-eindproduct, 36 µl NF-
H2O, 20µl 5X E-PAP buffer, 10 µl 25mM MnCl2, 10 µl ATP oplossing en 4 µl E-PAP
enzyme. Na 45’ incubatie bij 37°C wordt dit mengsel op ijs geplaatst.
Om de overgebleven nucleotiden en eiwitten te verwijderen wordt het mRNA geprecipiteerd
via lithiumchloride (LiCl). LiCl zorgt voor een goede precipitatie van mRNA, bestaande uit
meer dan 300 nucleotiden. Er wordt 50µl LiCl toegevoegd aan het mRNA en dit wordt
overnacht geïncubeerd bij -20°C. Na centrifugatie (30’, 13000 rpm, 4°C) wordt de pellet
gewassen in 70% ethanol. Tenslotte wordt het mRNA opgelost in NF-H2O en wordt de
concentratie bepaald met de Nanodrop1000 Spectrofotometer (Isogen Life Science, St-
Pieters-Leeuw, België).
2.6 mRNA Elektroporatie
Voor de elekroporatie experimenten met LPL mRNA worden de cellen van een LPL- CLL
patiënt gebruikt (P1). De cellen worden ontdooid zoals hierboven beschreven (zie II.1.3) Per
IVT mRNA elektroporatie zijn er 10x106 cellen nodig. De cellen worden drie keer gewassen,
waarvan twee keer met koud IMDM blanco medium en één keer met koude Washing solution
A (CellProjects, Kent, VK). De celpellet wordt geresuspendeerd in 200 µl Electroporation
buffer per elektroporatie (CellProjects, Kent, VK). Aan deze celsuspensie wordt 40 µg IVT
mRNA toegevoegd waarna het geheel van cellen en mRNA wordt overgebracht in een 0,4 cm
elektroporatiecuvette. Vervolgens wordt de elektroporatie uitgevoerd met de Put Gene pulser
II (Biorad, Nazareth, Belgïe) met volgende condities: capaciteit 150 µF, voltage 0,500 kV. De
geëlektroporeerde cellen worden vervolgens in 1 ml warm IMDM complete medium gebracht
in een 24-well plaat. Deze plaat wordt geïncubeerd bij 37 °C, 7% CO2. Naast een
elektroporatie met LPL mRNA worden drie controlestalen meegenomen: een cultuur (Cult) en
een mock controle (Mock) als negatieve controles en een EGFP controle (EGFP) als positieve
controle. In de cultuurcontrole (Cult) blijven de cellen onbehandeld. Voor de mockcontrole
(Mock) worden de cellen geëlektroporeerd zonder IVT mRNA. Deze controle laat toe het
effect van de elektroporatie zelf op de cellen na te gaan. Ten slotte worden voor de EGFP
controle de cellen geëlektroporeerd met EGFP IVT mRNA. Dit EGFP IVT mRNA werd
gesynthetiseerd zoals reeds beschreven voor LPL mRNA maar zonder de polyA-tailing stap.
(zie II.3.2).
Na 3 uur of 24 uur stimulatie wordt een klein deel van de cellen (100.000) gekleurd voor
flowcytometrische analyse. De helft van de overblijvende cellen wordt geoogst en in trizol

25
gebracht voor RNA isolatie (zie II.1.3) om vervolgens een qPCR (LPL) uit te voeren. De
andere helft van de cellen wordt in een lysebuffer gebracht. Hiervoor worden de cellen (+/-
500 µl) twee keer gewassen met 1 ml PBS (5’, 13000 rpm) (Lonza, Verviers, België).
Vervolgens wordt 100 µl 2X lysebuffer (0.25% Triton X-100, 50 mM KCl, 100 mM TrisHCl,
40% glycerol (pH 7.4)) toegevoegd.
3. pGEM4Z-LPL-A64bis
Figuur 11: De pGEM4Z vector gekloneerd met LPL pGEM4Z-LPL-A64bis (B) wordt verkregen door LPL te kloneren in PGEM4Z-EGFP-A64bis (A) (Vector NTI Advance TM software).
Om het plasmide pGEM4Z-LPL-A64bis (Figuur 11A) te verkrijgen wordt LPL DNA
geïsoleerd uit de pCMV-Sport6-LPL vector (Figuur 10) en vervolgens gekloneerd in de
vector pGEM4Z-EGFP-A64bis (Figuur 11B). Deze klonering heeft tot doel om de polyA-
tailing stap na IVT te vermijden. Door zijn 64bp-lange polyA-staart is de pGEM4Z-A64bis
vector immers optimaal voor IVT.
3.1 Isolatie plasmide DNA pGEM4Z-EGFP-A64bis
Het DNA van de pGEM4Z-EGFPA64bis vector werd geïsoleerd op dezelfde manier als
eerder beschreven (zie II.2.2)
3.2 Klonering
3.2.1 Restrictie pGEM4Z-EGFP-A64bis en pCMV-Sport6-LPL
LPL (1428bp) wordt uit de geïsoleerde vector pCMV-Sport6-LPL geknipt door gebruik te
maken van de restrictie-enzymen XbaI en EcoRI (New England Biolabs, Leusden, Nederland)
(Figuur 10). Het reactiemengsel bestaat uit 2 µg vector, 10X NEbuffer 2, 1 µLXbaI, 1
A B

26
µLEcoRI, 100X BSA en dit wordt aangelengd tot 50 µL met NF-H2O. Dit mengsel wordt
gedurende 3 uur bij 37 °C geplaatst.
De pGEM4Z-EGFP-A64bis vector wordt eveneens geknipt met de restrictie-enzymen XbaI
en EcoRI om EGFP te verwijderen.
3.2.2 Opzuivering
De gewenste restrictiefragmenten (insert: LPL en backbone: pGEM4Z-A64bis zonder EGFP)
worden opgezuiverd via gelelektroforese. Gelelektroforese maakt het mogelijk om DNA
moleculen te scheiden op basis van grootte. Een 1,5% agarosegel wordt geladen met een
100bp ladder, een 1 kb ladder en de restrictieproducten (50 µl restrictieproduct + 10 µl 6X
loading dye (Fermentas, St. Leon-Rot, Duitsland)). Aan de hand van de ladders kunnen de
gewenste DNA fragmenten uit de gel gesneden worden en opgezuiverd via het Wizard SV gel
and PCR Clean-up system (Promega).
3.2.3 Ligatie
Ligatie van het LPL gen in het geknipte pGEM4Z-A64bis plasmide wordt uitgevoerd aan een
3:1 of 1:1 vector:insert ratio. De starthoeveelheid van het acceptor plasmide is 50 ng. De
hoeveelheid insert die moet worden toegevoegd, wordt berekend aan de hand van de bp ratio.
In dit geval is de insert (1538 bp) 1,68 keer kleiner dan de geknipte vector (2595bp).
Rekening houdend met deze ratio, kan berekend worden dat er 29 ng (1:1 ligatie) of 88 ng
(1:3 ligatie) insert moet worden toegevoegd. Hieraan wordt nog 100x T4 DNA ligase en 10x
T4 DNA buffer (Invitrogen, Merelbeke, België) toegevoegd. De stalen worden overnacht
geïncubeerd bij 4°C. Er wordt eveneens een controle meegenomen waarbij het insert
vervangen wordt door water, dit dient om de achtergrond autosluiting te detecteren.
3.2.4 Transformatie
Om de gekloneerde pGEM4Z-LPL-A64bis vector te vermenigvuldigen wordt een
transformatie in DH5α E.coli competente cellen uitgevoerd. Deze competente cellen bezitten
een endA1 mutatie, die expressie van een endonculease inhibeert en bijgevolg plasmide
degradatie vermijdt. De competente cellen (100 µl) worden samengebracht met 10 µl van het
ligatiemengsel en 45’ op ijs geplaatst. Daarna worden de cellen gedurende 2’ blootgesteld aan
een hitteschok van 42°C. De getransformeerde bacteriële cellen worden onmiddellijk terug op
ijs geplaatst waarna 700 µl SOC medium wordt toegevoegd. Na een incubatieperiode van één

27
uur bij 37°C worden de cellen uitgestreken op LB Amp agar platen. Als positieve controle
wordt een ongeknipte vector getransformeerd (Qiagen).
3.2.5 Miniprep
Na een incubatieperiode overnacht bij 37°C worden per plaat meerdere kolonies opgepikt en
elk in 4 ml LB brothAmp medium gebracht. De bacterieculturen worden overnacht
geïncubeerd, waarna het plasmide DNA wordt geïsoleerd door middel van een miniprep kit
(Qiagen, Venlo, Nederland, zie ‘QIAprep® Miniprep Handbook’ voor protocol)).
3.2.6 Screening na transformatie
De vector wordt geknipt om na te gaan of de LPL insert inderdaad geligeerd is in de
pGEM4Z-A64bis backbone. Er worden twee restricties uitgevoerd op de pGEM4Z-LPL-
A64bis clones. Voor de eerste restrictie wordt geknipt met Nde1 (New England Biolabs,
Leusden, Nederland) dat in de backbone knipt en met PflMI (New England Biolabs, Leusden,
Nederland) dat in de insert knipt. Voor de tweede restrictie wordt er geknipt met dezelfde
enzymen als diegene waarmee de insert in de backbone werd gebracht.
3.2.7 Cyclesequencing
Er wordt een PCR uitgevoerd om de LPL-sequentie te amplificeren. Volgende primers
werden ontworpen op basis van de sequentie van de pGEM4Z-EGFP-A64 bis vector en de
pCMV-Sport6-LPL vector; T7 Forward primer (5’-TAA TAC GAC TCA CTA TAG GG-3’), LPL
Forward primer 1 (5’- AAA CTG GTG GGA CAG GAT G-3’), LPL Forward primer 2 (5’-GTT GTA
GAA AGA ACC GCT GC-3’), LPL Foward primer 3 (5’-TAC TTT AGC TGG TCA GAC TGG-3’), LPL
Reverse primer 1 (5’-GCA TTG CAG GAA GTC TGA C-3’). De sequencing PCR wordt uitgevoerd
met de AB BigDye Terminator kit (Applied Biosystems). Per staal wordt een mix bereid
bestaande uit 3,5 µl 5X sequencing buffer, 1 µl sequencing RR 100 mix (Ready Reaction
mix) en 11 µl NF-H2O. Hieraan wordt 2 µl DNA en 2,5 µl 2µM primer toegevoegd. De PCR
wordt gelopen in de Veriti 96 well Thermal Cycler (Applied Biosystems) bij volgend
programma: (6’ bij 96°C, 25 cycli 10” bij 96°c, 5” bij 50°C, 4’ bij 60°C). Vervolgens worden
de PCR producten opgezuiverd aan de hand van de BigDye X-Terminator Purifaction Kit
(Applied Biosystems). Bij elk staal wordt 20 µl X terminator Solution en 90 µl SAM Solution
gevoegd. De stalen worden 30’ gevortexd en vervolgens gecentrifugeerd gedurende 2’ aan
1000 rpm. 30 µl van de bovenstaande oplossing wordt overgebracht in een 96-well plaat en de
sequentie wordt bepaald via de ABI PRISM® 310 Genetic Analyzer (Applied Biosystems)

28
door het personeel van het ARL (Aids Referentie Labo). Via DNAman wordt gekeken of de
pGEM4Z-LPL-A64bis clones het LPL gen bevatten en of het geen puntmutaties bevat.
3.3 mRNA elektroporatie
Van het plasmide pGEM4Z-LPL-A64bis wordt eveneens een IVT uitgevoerd. Deze vector
heeft als voordeel dat er geen polyA-staart moet aangemaakt worden (zie II.2.4). De CLL
cellen worden opnieuw geëlektroporeerd met het mRNA. Na 3 uur of 24 uur stimulatie wordt
een deel van de cellen gekleurd voor flowcytometrische analyse. Een deel van de cellen wordt
geoogst en in trizol gebracht voor RNA isolatie om vervolgens een qPCR uit te voeren. De
rest van de cellen wordt in een lysebuffer gebracht voor ELISA (II.2.6).
4. Transfectie HEK 293T-cellen
4.1 Uitplaten cellen
Voor DNA transfectie wordt gebruik gemaakt van HEK 293T-cellen die 70% confluent zijn.
Hiervoor worden de adherente HEK 293T-cellen één dag voor transfectie uitgeplaat aan
750.000 cellen in 6 ml IMDM complete medium. Er worden 6 plaatjes gemaakt die in de
incubator worden geplaatst bij 37°C en 7% CO2.
4.2 Transfectie
DNA transfectie in HEK 293T-cellen gebeurt met behulp van de calcium fosfaat transfectiekit
(Invitrogen, Merelbeke, België). Deze transfectiemethode is gebaseerd op de vorming van een
calcium fosfaat-DNA precipitaat. Calcium fosfaat bevordert de binding van het DNA op het
cel oppervlak waarna het DNA de cel binnentreedt via endocytose. Er worden vier
transfecties uitgevoerd: een transfectie met het DNA van de pCMV-Sport6-LPL vector en
twee transfecties met twee verschillende hoeveelheden (10 µg en 40 µg) LPL mRNA in vitro
overgeschreven van de pGEM4Z-LPL-A64bis vector. Daarnaast wordt ook een transfectie
met het DNA van de pLKO.1- EGFP controle vector gedaan als positieve controle. De cellen
getransfecteerd met deze vector zouden na transfectie groen moeten kleuren onder de
fluorescentiemicroscoop of een sterk FITC signaal geven op flowcytometrie. Als negatieve
controle worden niet getransfecteerde 293T-cellen gebruikt. Volgende mixen van DNA,
Transfection Qualified Water (TQW) en calcium chloride worden bereid in 1,6 ml eppendorf
buisjes:

29
Tabel 3: Samenstelling mixen.
DNA TQW CaCl2 pCMV-Sport6-LPL [1190 ng/µl] 10 µg (8,4µl) 255,6 µl 36 µl
IVT mRNA LPL[1660 ng/µl] 10 µg (6,0µl) 258 µl 36 µl
IVT mRNA LPL[1660 ng/µl] 40 µg (24,1µl) 239,9 µl 36 µl
pLKO.1 shRNA EGFP scrambled [202 ng/µl]
10 µg (49,5µl) 214,5 µl 36 µl
Negatieve controle - - -
TQW : Transfection Qualified Water, CaCl2: calciumchloride
Bovenstaande DNA mixen worden druppelsgewijs toegevoegd aan 300 µl 2X HBS oplossing
terwijl er lucht doorheen de oplossing wordt ‘gebubbeld’. Dit heeft tot doel te vermijden dat
er DNA klonters worden gevormd aangezien de binding aan de cel dan niet efficiënt verloopt.
Dit mengsel wordt geïncubeerd op kamertemperatuur gedurende 30’ en 5’ voor het einde van
deze incubatieperiode wordt chloroquine toegevoegd aan de te transfecteren HEK 293T-
cellen. Het medium (6ml) wordt verwijderd van de cellen, waarna 6 ml IMDM complete
medium met chloroquine aan een 1/8000 concentratie op de cellen wordt gebracht.
Vervolgens wordt het DNA mengsel verspreid over de cellen in het 6 cm plaatje.
Eén dag na transfectie wordt het medium van de cellen ververst. De volgende dag worden de
cellen geanalyseerd.
4.3 Analyse transfectie efficiëntie
Om te zien of de transfectie succesvol is, wordt de EGFP expressie gecontroleerd onder de
fluorescentiemicroscoop. Vervolgens worden de cellen losgemaakt met behulp van trypsine
waarna een kleine fractie cellen overgebracht wordt in een FACS buisje voor
flowcytometrische kleuring. Er wordt zowel een intracellulaire als een oppervlakte kleuring
voor LPL uitgevoerd (zie II.1.3). Er wordt niet gekleurd voor andere oppervlaktemerkers. De
stalen worden geanalyseerd op de FACS-Calibur. Analyse gebeurt met het CellQuest Pro
programme (BD Biosciences). Zoals hoger beschreven wordt de helft van de resterende cellen
in 700 µl trizol gebracht, waarna RNA isolatie, cDNA synthese en een qPCR voor LPL
plaatsvindt. De andere helft van de cellen wordt gelyseerd voor ELISA.
5.ELISA
Naast flowcytometrie wordt ook via een ELISA kit (AlpcoDiagnostics, Salem, VS) (Figuur
12), de LPL eiwit-expressie gemeten. Het LPL eiwit wordt via ELISA nagegaan in het
cellysaat van drie stalen: CLL cellen geëlektroporeerd met mRNA verkregen via IVT van
PGEM4Z-LPL-A64bis, getransfecteerde HEK 293T-cellen met pCMV-Sport6-LPLvector,

30
getransfecteerde HEK 293T-cellen met pLKO.1-EGFP vector als negatieve controle. Een 96-
well plaat is gecoat met anti-LPL monoclonaal antilichaam. In deze plaat wordt per
reactievaatje 50 µl van staal toegevoegd. De plaat wordt 1 uur geïncubeerd bij 15-25°C,
zodanig dat het LPL eiwit kan binden op het antilichaam waarmee de plaat gecoat is.
Vervolgens wordt de ongebonden fractie verwijderd door de plaat drie keer te wassen met 350
µl wasbuffer. Daarna wordt er 50 µl anti-LPL polyclonaal antilichaam (anti-rund LPL
kippenserum) toegevoegd dat bindt op het LPL aanwezig in de welletjes. Dit wordt 30’
geïncubeerd bij kamertemperatuur. Na drie keer wassen, wordt 50 µl van het enzyme gelabeld
antlichaam (horseradish peroxidase (HRP) - gelabeld anti-kip IgG goat serum) toegevoegd.
Dit antilichaam zal binden op het anti-LPL polyclonaal antilichaam. Er volgt opnieuw een
incubatieperiode van 30’ bij kamertemperatuur. Na het wassen, wordt het substraat, o-
phenylenediamine, toegevoegd. Indien er LPL aanwezig was in de well, zal het substraat door
inwerking van HRP resulteren in een gele kleur. De absorbantie wordt tenslotte gemeten bij
492 nm met behulp van een microplaat reader Versamax™ (Molecular Devices Inc,
Sunnyvale, VS).
Figuur 12 Principe van de LPL ELISA (aangepast van http://www.cellsignal.com). l

31
III. Resultaten
1. Detectie van LPL in LPL-geëlektroporeerde CLL cellen
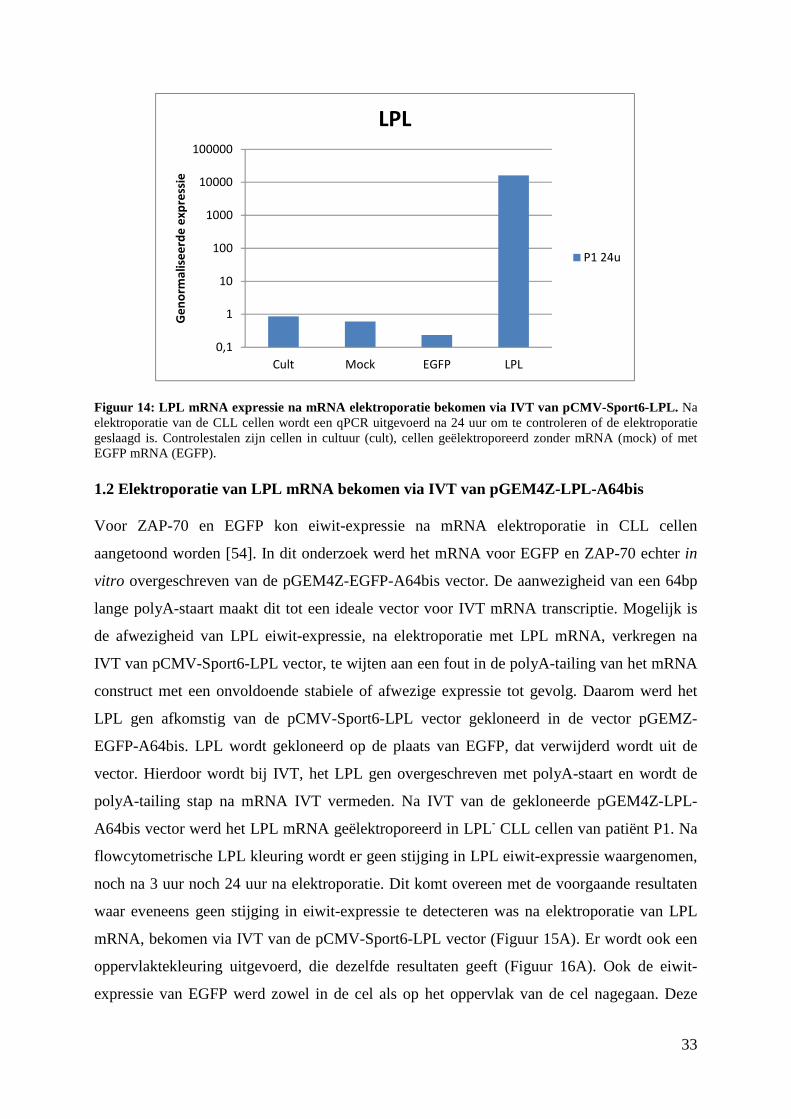
1.1 Elektroporatie van LPL mRNA bekomen via IVT van pCMV-Sport6-LPL
Om LPL tot expressie te brengen in LPL- CLL cellen van patiënt P1, worden de cellen
geëlektroporeerd met LPL mRNA. Dit mRNA is afgeschreven van de pCMV-Sport6-LPL
vector via IVT.
Via flowcytometrie wordt translatie van het geëlektroporeerde mRNA nagegaan. Volgende
gatingstrategie werd toegepast om specifiek de eiwit-expressie van de B-cellen te analyseren.
Eerst worden de lymfocyten geselecteerd op basis van de grootte en de granulositeit via
analyse van respectievelijk de FSC en de SSC Vervolgens worden ze afgezonderd van de
dode cellen op basis van PI kleuring. Onderscheid tussen B- en T-cellen gebeurt op basis van
expressie van respectievelijk CD19 en CD3. Vervolgens kan de expressie van LPL in de B-
cellen worden geanalyseerd. Negatieve controles zijn onbehandelde cellen (cult) en cellen
geëlektroporeerd zonder mRNA (mock). Cellen geëlektroporeerd met EGFP mRNA (EGFP)
worden gebruikt als positieve controle. De niet-behandelde CLL cellen vertonen nauwelijks
LPL expressie (Figuur 13). Bij de LPL-geëlektroporeerde cellen wordt een stijging van het
aantal LPL+ CLL cellen waargenomen. Deze stijging wordt echter ook waargenomen bij
mock-behandelde cellen en er wordt zelfs een grotere stijging waargenomen bij EGFP-
geëlektroporeerde CLL cellen.
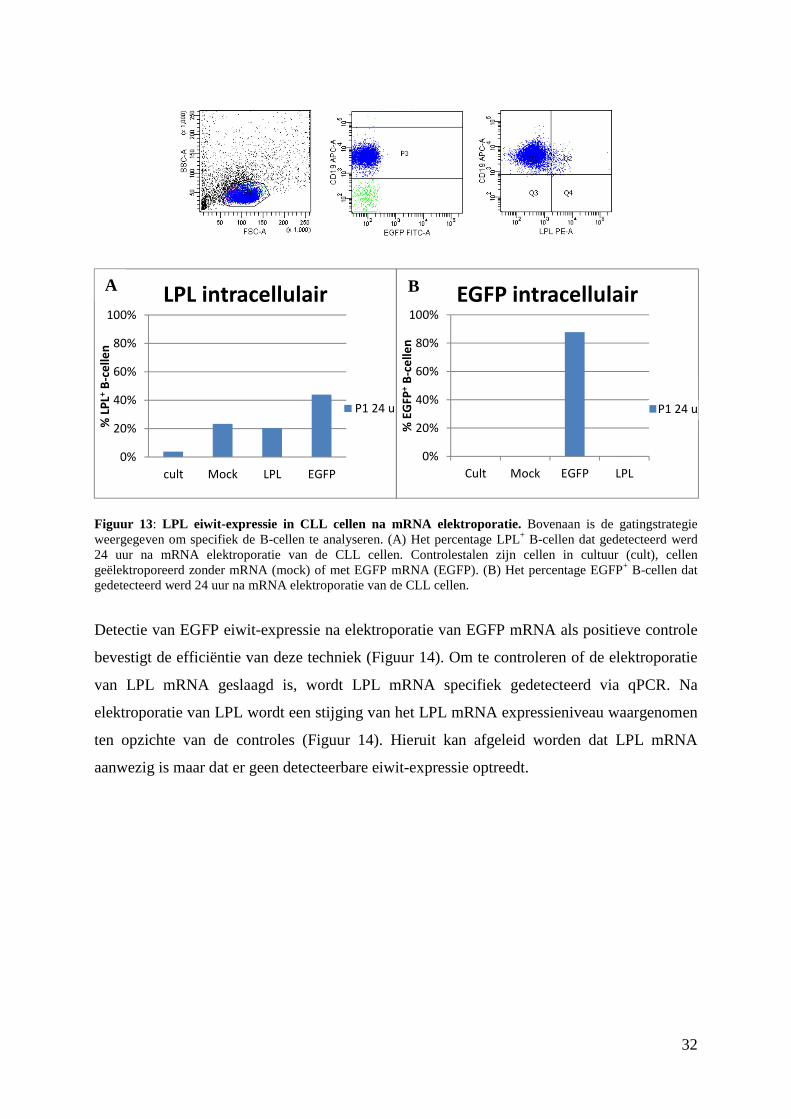
32
Figuur 13: LPL eiwit-expressie in CLL cellen na mRNA elektroporatie. Bovenaan is de gatingstrategie weergegeven om specifiek de B-cellen te analyseren. (A) Het percentage LPL+ B-cellen dat gedetecteerd werd 24 uur na mRNA elektroporatie van de CLL cellen. Controlestalen zijn cellen in cultuur (cult), cellen geëlektroporeerd zonder mRNA (mock) of met EGFP mRNA (EGFP). (B) Het percentage EGFP+ B-cellen dat gedetecteerd werd 24 uur na mRNA elektroporatie van de CLL cellen.
Detectie van EGFP eiwit-expressie na elektroporatie van EGFP mRNA als positieve controle
bevestigt de efficiëntie van deze techniek (Figuur 14). Om te controleren of de elektroporatie
van LPL mRNA geslaagd is, wordt LPL mRNA specifiek gedetecteerd via qPCR. Na
elektroporatie van LPL wordt een stijging van het LPL mRNA expressieniveau waargenomen
ten opzichte van de controles (Figuur 14). Hieruit kan afgeleid worden dat LPL mRNA
aanwezig is maar dat er geen detecteerbare eiwit-expressie optreedt.
0%
20%
40%
60%
80%
100%
Cult Mock EGFP LPL
% E
GF
P+
B-c
ell
en
EGFP intracellulair
0%
20%
40%
60%
80%
100%
cult Mock LPL EGFP
% L
PL+
B-c
ell
en
LPL intracellulair
P1 24 u P1 24 u
A B
33
Figuur 14: LPL mRNA expressie na mRNA elektroporatie bekomen via IVT van pCMV-Sport6-LPL. Na elektroporatie van de CLL cellen wordt een qPCR uitgevoerd na 24 uur om te controleren of de elektroporatie geslaagd is. Controlestalen zijn cellen in cultuur (cult), cellen geëlektroporeerd zonder mRNA (mock) of met EGFP mRNA (EGFP).
1.2 Elektroporatie van LPL mRNA bekomen via IVT van pGEM4Z-LPL-A64bis
Voor ZAP-70 en EGFP kon eiwit-expressie na mRNA elektroporatie in CLL cellen
aangetoond worden [54]. In dit onderzoek werd het mRNA voor EGFP en ZAP-70 echter in
vitro overgeschreven van de pGEM4Z-EGFP-A64bis vector. De aanwezigheid van een 64bp
lange polyA-staart maakt dit tot een ideale vector voor IVT mRNA transcriptie. Mogelijk is
de afwezigheid van LPL eiwit-expressie, na elektroporatie met LPL mRNA, verkregen na
IVT van pCMV-Sport6-LPL vector, te wijten aan een fout in de polyA-tailing van het mRNA
construct met een onvoldoende stabiele of afwezige expressie tot gevolg. Daarom werd het
LPL gen afkomstig van de pCMV-Sport6-LPL vector gekloneerd in de vector pGEMZ-
EGFP-A64bis. LPL wordt gekloneerd op de plaats van EGFP, dat verwijderd wordt uit de
vector. Hierdoor wordt bij IVT, het LPL gen overgeschreven met polyA-staart en wordt de
polyA-tailing stap na mRNA IVT vermeden. Na IVT van de gekloneerde pGEM4Z-LPL-
A64bis vector werd het LPL mRNA geëlektroporeerd in LPL- CLL cellen van patiënt P1. Na
flowcytometrische LPL kleuring wordt er geen stijging in LPL eiwit-expressie waargenomen,
noch na 3 uur noch 24 uur na elektroporatie. Dit komt overeen met de voorgaande resultaten
waar eveneens geen stijging in eiwit-expressie te detecteren was na elektroporatie van LPL
mRNA, bekomen via IVT van de pCMV-Sport6-LPL vector (Figuur 15A). Er wordt ook een
oppervlaktekleuring uitgevoerd, die dezelfde resultaten geeft (Figuur 16A). Ook de eiwit-
expressie van EGFP werd zowel in de cel als op het oppervlak van de cel nagegaan. Deze
0,1
1
10
100
1000
10000
100000
Cult Mock EGFP LPL
Ge
no
rma
lise
erd
e e
xp
ress
ie
LPL
P1 24u
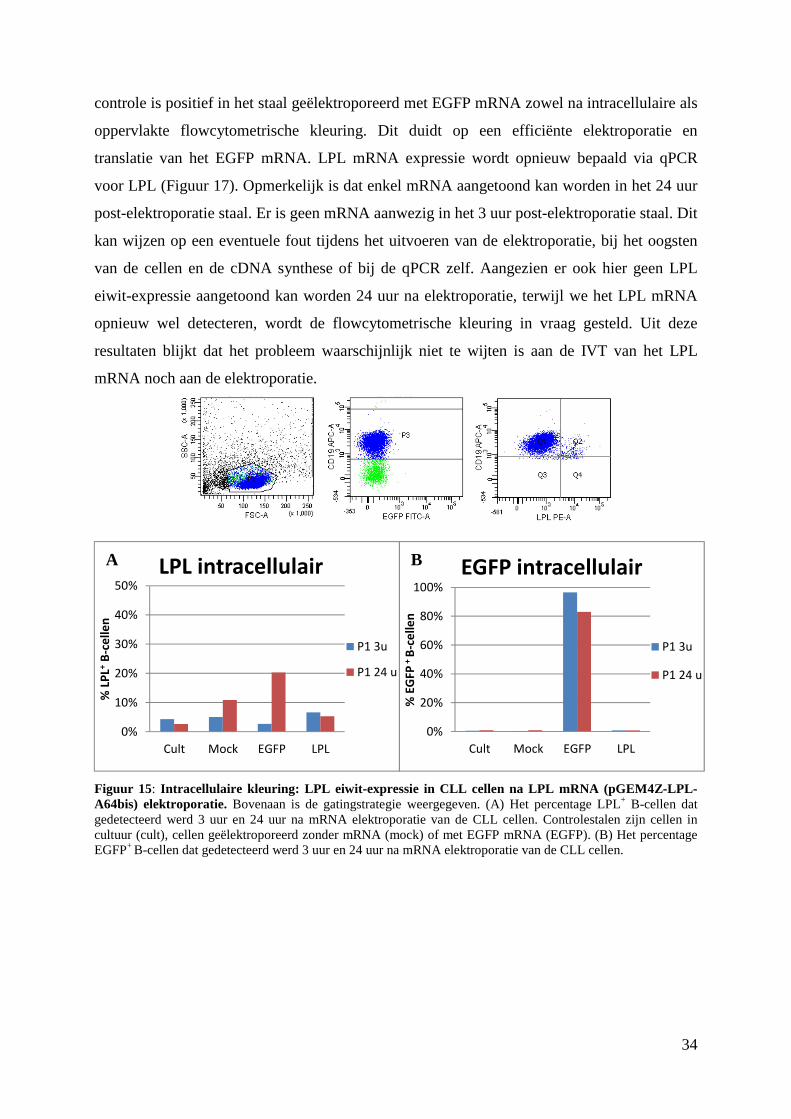
34
controle is positief in het staal geëlektroporeerd met EGFP mRNA zowel na intracellulaire als
oppervlakte flowcytometrische kleuring. Dit duidt op een efficiënte elektroporatie en
translatie van het EGFP mRNA. LPL mRNA expressie wordt opnieuw bepaald via qPCR
voor LPL (Figuur 17). Opmerkelijk is dat enkel mRNA aangetoond kan worden in het 24 uur
post-elektroporatie staal. Er is geen mRNA aanwezig in het 3 uur post-elektroporatie staal. Dit
kan wijzen op een eventuele fout tijdens het uitvoeren van de elektroporatie, bij het oogsten
van de cellen en de cDNA synthese of bij de qPCR zelf. Aangezien er ook hier geen LPL
eiwit-expressie aangetoond kan worden 24 uur na elektroporatie, terwijl we het LPL mRNA
opnieuw wel detecteren, wordt de flowcytometrische kleuring in vraag gesteld. Uit deze
resultaten blijkt dat het probleem waarschijnlijk niet te wijten is aan de IVT van het LPL
mRNA noch aan de elektroporatie.
Figuur 15: Intracellulaire kleuring: LPL eiwit-expressie in CL L cellen na LPL mRNA (pGEM4Z-LPL-A64bis) elektroporatie. Bovenaan is de gatingstrategie weergegeven. (A) Het percentage LPL+ B-cellen dat gedetecteerd werd 3 uur en 24 uur na mRNA elektroporatie van de CLL cellen. Controlestalen zijn cellen in cultuur (cult), cellen geëlektroporeerd zonder mRNA (mock) of met EGFP mRNA (EGFP). (B) Het percentage EGFP+ B-cellen dat gedetecteerd werd 3 uur en 24 uur na mRNA elektroporatie van de CLL cellen.
0%
20%
40%
60%
80%
100%
Cult Mock EGFP LPL
% E
GF
P+
B-c
ell
en
EGFP intracellulair
P1 3u
P1 24 u
0%
10%
20%
30%
40%
50%
Cult Mock EGFP LPL
% L
PL+
B-c
ell
en
LPL intracellulair
P1 3u
P1 24 u
A B
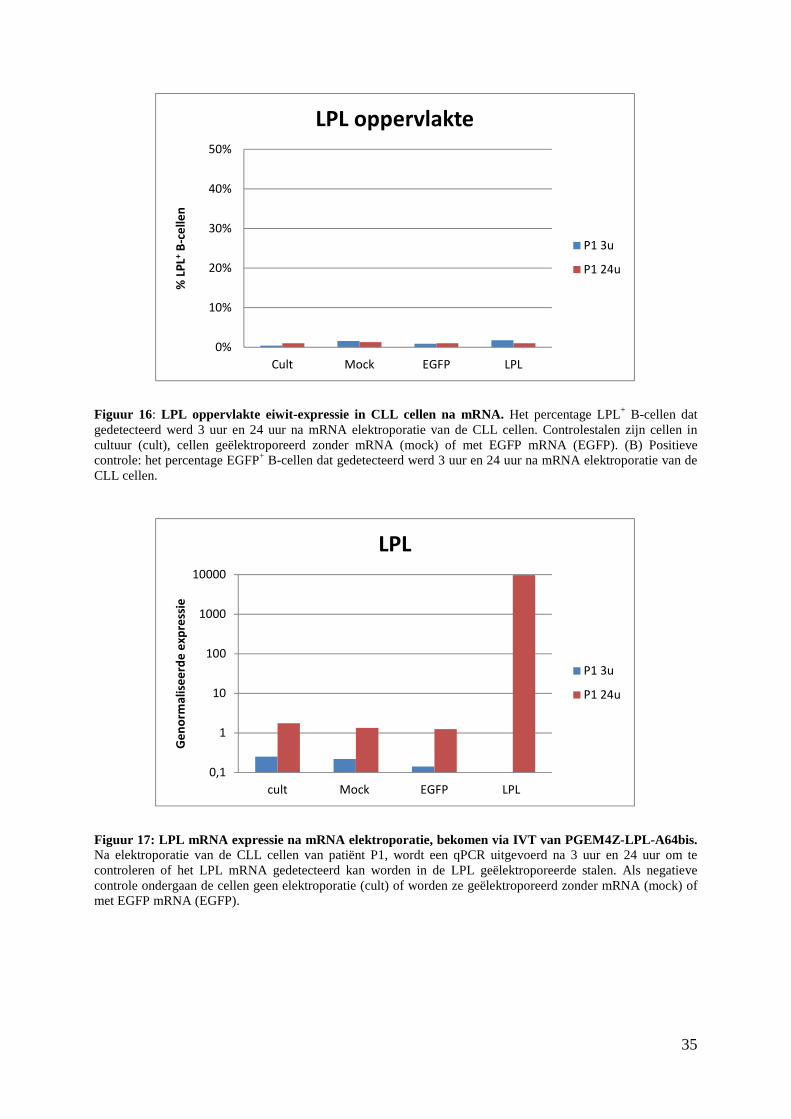
35
Figuur 16: LPL oppervlakte eiwit-expressie in CLL cellen na mRNA. Het percentage LPL+ B-cellen dat gedetecteerd werd 3 uur en 24 uur na mRNA elektroporatie van de CLL cellen. Controlestalen zijn cellen in cultuur (cult), cellen geëlektroporeerd zonder mRNA (mock) of met EGFP mRNA (EGFP). (B) Positieve controle: het percentage EGFP+ B-cellen dat gedetecteerd werd 3 uur en 24 uur na mRNA elektroporatie van de CLL cellen.
Figuur 17: LPL mRNA expressie na mRNA elektroporatie, bekomen via IVT van PGEM4Z-LPL-A64bis. Na elektroporatie van de CLL cellen van patiënt P1, wordt een qPCR uitgevoerd na 3 uur en 24 uur om te controleren of het LPL mRNA gedetecteerd kan worden in de LPL geëlektroporeerde stalen. Als negatieve controle ondergaan de cellen geen elektroporatie (cult) of worden ze geëlektroporeerd zonder mRNA (mock) of met EGFP mRNA (EGFP).
0%
10%
20%
30%
40%
50%
Cult Mock EGFP LPL
% L
PL+
B-c
ell
en
LPL oppervlakte
P1 3u
P1 24u
0,1
1
10
100
1000
10000
cult Mock EGFP LPL
Ge
no
rma
lise
erd
e e
xp
ress
ie
LPL
P1 3u
P1 24u
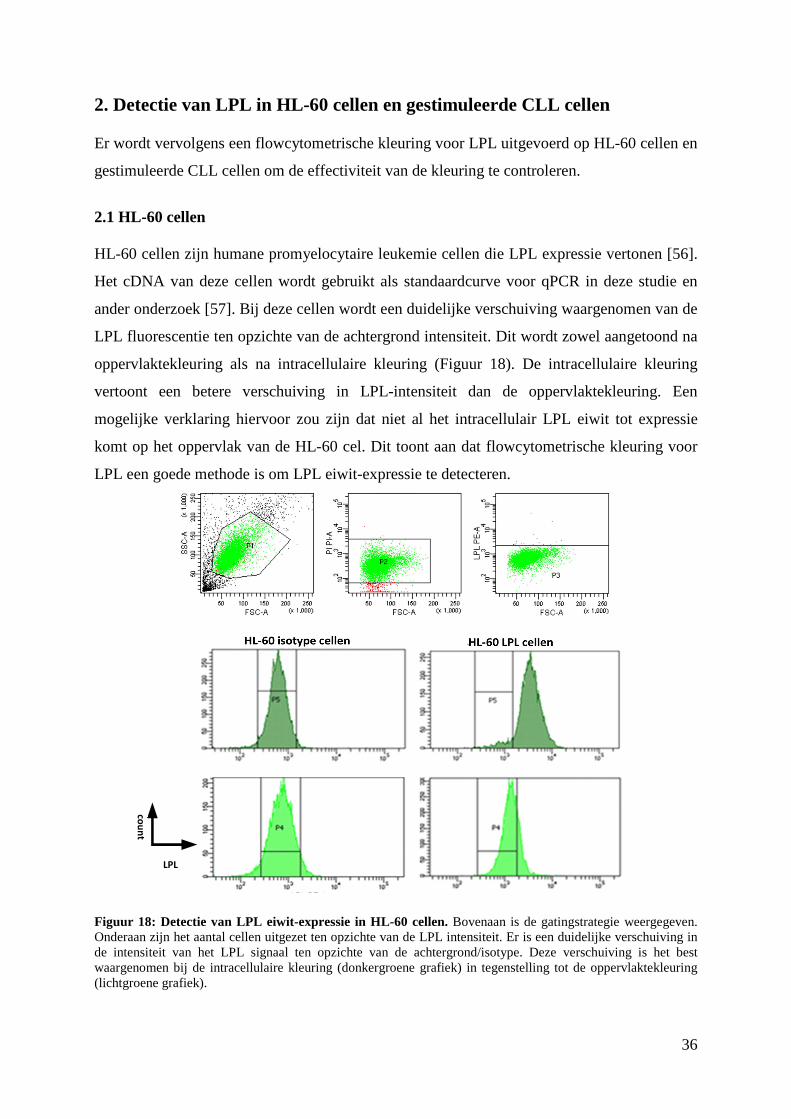
36
2. Detectie van LPL in HL-60 cellen en gestimuleerde CLL cellen
Er wordt vervolgens een flowcytometrische kleuring voor LPL uitgevoerd op HL-60 cellen en
gestimuleerde CLL cellen om de effectiviteit van de kleuring te controleren.
2.1 HL-60 cellen
HL-60 cellen zijn humane promyelocytaire leukemie cellen die LPL expressie vertonen [56].
Het cDNA van deze cellen wordt gebruikt als standaardcurve voor qPCR in deze studie en
ander onderzoek [57]. Bij deze cellen wordt een duidelijke verschuiving waargenomen van de
LPL fluorescentie ten opzichte van de achtergrond intensiteit. Dit wordt zowel aangetoond na
oppervlaktekleuring als na intracellulaire kleuring (Figuur 18). De intracellulaire kleuring
vertoont een betere verschuiving in LPL-intensiteit dan de oppervlaktekleuring. Een
mogelijke verklaring hiervoor zou zijn dat niet al het intracellulair LPL eiwit tot expressie
komt op het oppervlak van de HL-60 cel. Dit toont aan dat flowcytometrische kleuring voor
LPL een goede methode is om LPL eiwit-expressie te detecteren.
Figuur 18: Detectie van LPL eiwit-expressie in HL-60 cellen. Bovenaan is de gatingstrategie weergegeven. Onderaan zijn het aantal cellen uitgezet ten opzichte van de LPL intensiteit. Er is een duidelijke verschuiving in de intensiteit van het LPL signaal ten opzichte van de achtergrond/isotype. Deze verschuiving is het best waargenomen bij de intracellulaire kleuring (donkergroene grafiek) in tegenstelling tot de oppervlaktekleuring (lichtgroene grafiek).
LPL
cou
nt
37
2.2 Gestimuleerde CLL cellen
2.2.1. Bepaling LPL-genexpressie in gestimuleerde CLL cellen
Na stimulatie van CLL cellen komt LPL tot expressie. LPL expressie treedt eveneens op
zonder B-cel stimulatie bij CLL patiënten met de niet-gemuteerde status. LPL komt niet tot
expressie in normale B-cellen of niet-gestimuleerde gemuteerde CLL cellen [48]. Dit wordt
nagegaan aan de hand van een qPCR analyse van stalen van twee verschillende CLL patiënten
P1 en P2. De CLL cellen van P1 bezitten een gemuteerde IgVH status (M), terwijl de cellen
van P2 een ongemuteerde IgVH status (UM) hebben. De stalen worden gestimuleerd met anti-
IgM beads en na 3 uur en 24 uur wordt een analyse uitgevoerd van deze gestimuleerde cellen.
Om te controleren of de cellen reageren op de stimulatie wordt de FOS expressie
geanalyseerd die na IgM stimulatie van B-cellen stijgt [58]. Aangezien er geen IgA op het
oppervlak van de CLL cel aanwezig is, worden de cellen gestimuleerd met anti-IgA beads als
negatieve controle. Zoals in Figuur 19 wordt weergegeven, stijgt de expressie van FOS drie
uur na stimulatie. Na 24 uur is deze reeds afgenomen, maar nog steeds hoger dan de expressie
in de controlestalen. Na stimulatie met de IgM beads, is de LPL expressie zowel gestegen in
de CLL cellen met de gemuteerde IgVH status als de CLL cellen met ongemuteerde IgVH
status (Figuur 20). In het staal van P2 met ongemuteerde CLL cellen, treedt een grotere
stijging van LPL expressie in tegenstelling tot het staal van P1 met de gemuteerde CLL
cellen. Dit resultaat sluit aan bij eerder onderzoek [48].
Figuur 19: Stijging van de FOS expressie na IgM stimulatie van CLL cellen. De genormaliseerde c-fos expressie wordt vergeleken tussen IgM gestimuleerde en IgA gestimuleerde CLL cellen. mRNA expressie wordt gedetecteerd 3 uur en 24 uur na stimulatie voor patiënten P1 (M) en P2 (UM).
0
50
100
150
200
250
300
P1 3u P2 3u P1 24u P2 24u
Ge
no
rma
lise
erd
e e
xp
ress
ie
FOS
IgA
IgM
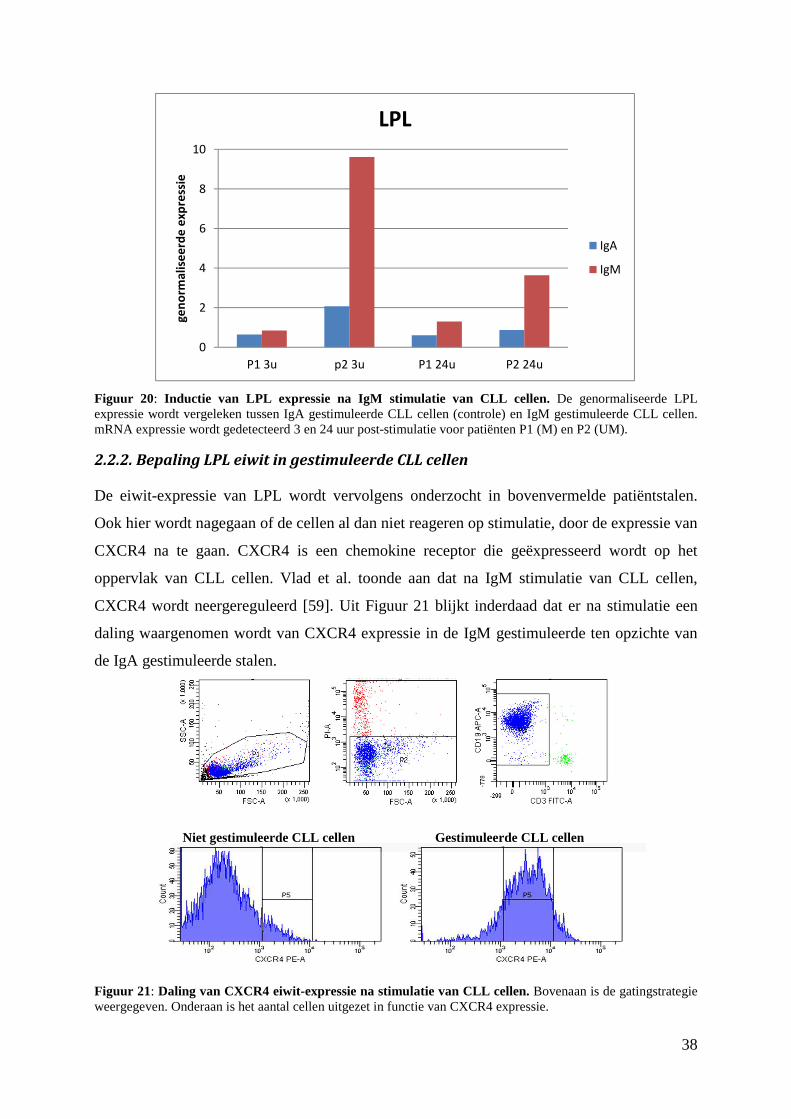
38
Figuur 20: Inductie van LPL expressie na IgM stimulatie van CLL cellen. De genormaliseerde LPL expressie wordt vergeleken tussen IgA gestimuleerde CLL cellen (controle) en IgM gestimuleerde CLL cellen. mRNA expressie wordt gedetecteerd 3 en 24 uur post-stimulatie voor patiënten P1 (M) en P2 (UM).
2.2.2. Bepaling LPL eiwit in gestimuleerde CLL cellen
De eiwit-expressie van LPL wordt vervolgens onderzocht in bovenvermelde patiëntstalen.
Ook hier wordt nagegaan of de cellen al dan niet reageren op stimulatie, door de expressie van
CXCR4 na te gaan. CXCR4 is een chemokine receptor die geëxpresseerd wordt op het
oppervlak van CLL cellen. Vlad et al. toonde aan dat na IgM stimulatie van CLL cellen,
CXCR4 wordt neergereguleerd [59]. Uit Figuur 21 blijkt inderdaad dat er na stimulatie een
daling waargenomen wordt van CXCR4 expressie in de IgM gestimuleerde ten opzichte van
de IgA gestimuleerde stalen.
Figuur 21: Daling van CXCR4 eiwit-expressie na stimulatie van CLL cellen. Bovenaan is de gatingstrategie weergegeven. Onderaan is het aantal cellen uitgezet in functie van CXCR4 expressie.
0
2
4
6
8
10
P1 3u p2 3u P1 24u P2 24u
ge
no
rma
lise
erd
e e
xp
ress
ie
LPL
IgA
IgM
Niet gestimuleerde CLL cellen Gestimuleerde CLL cellen
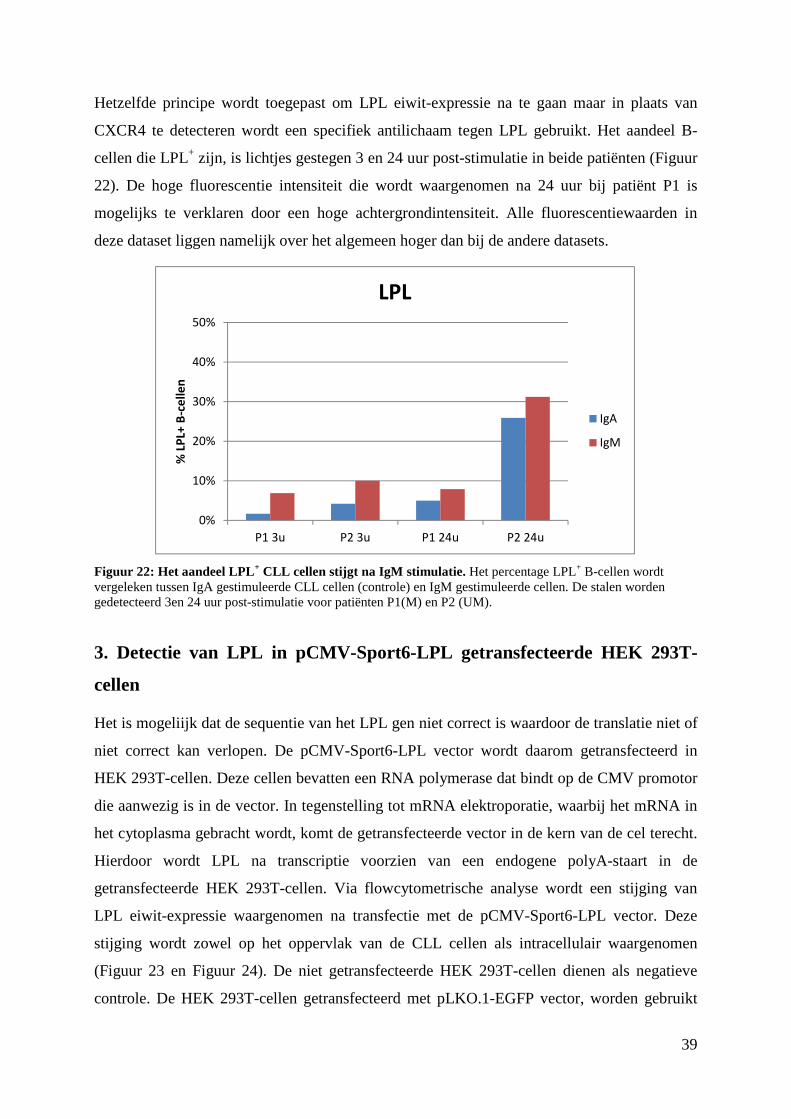
39
Hetzelfde principe wordt toegepast om LPL eiwit-expressie na te gaan maar in plaats van
CXCR4 te detecteren wordt een specifiek antilichaam tegen LPL gebruikt. Het aandeel B-
cellen die LPL+ zijn, is lichtjes gestegen 3 en 24 uur post-stimulatie in beide patiënten (Figuur
22). De hoge fluorescentie intensiteit die wordt waargenomen na 24 uur bij patiënt P1 is
mogelijks te verklaren door een hoge achtergrondintensiteit. Alle fluorescentiewaarden in
deze dataset liggen namelijk over het algemeen hoger dan bij de andere datasets.
Figuur 22: Het aandeel LPL+ CLL cellen stijgt na IgM stimulatie. Het percentage LPL+ B-cellen wordt vergeleken tussen IgA gestimuleerde CLL cellen (controle) en IgM gestimuleerde cellen. De stalen worden gedetecteerd 3en 24 uur post-stimulatie voor patiënten P1(M) en P2 (UM).
3. Detectie van LPL in pCMV-Sport6-LPL getransfecteerde HEK 293T-
cellen
Het is mogeliijk dat de sequentie van het LPL gen niet correct is waardoor de translatie niet of
niet correct kan verlopen. De pCMV-Sport6-LPL vector wordt daarom getransfecteerd in
HEK 293T-cellen. Deze cellen bevatten een RNA polymerase dat bindt op de CMV promotor
die aanwezig is in de vector. In tegenstelling tot mRNA elektroporatie, waarbij het mRNA in
het cytoplasma gebracht wordt, komt de getransfecteerde vector in de kern van de cel terecht.
Hierdoor wordt LPL na transcriptie voorzien van een endogene polyA-staart in de
getransfecteerde HEK 293T-cellen. Via flowcytometrische analyse wordt een stijging van
LPL eiwit-expressie waargenomen na transfectie met de pCMV-Sport6-LPL vector. Deze
stijging wordt zowel op het oppervlak van de CLL cellen als intracellulair waargenomen
(Figuur 23 en Figuur 24). De niet getransfecteerde HEK 293T-cellen dienen als negatieve
controle. De HEK 293T-cellen getransfecteerd met pLKO.1-EGFP vector, worden gebruikt
0%
10%
20%
30%
40%
50%
P1 3u P2 3u P1 24u P2 24u
% L
PL+
B-c
ell
en
LPL
IgA
IgM
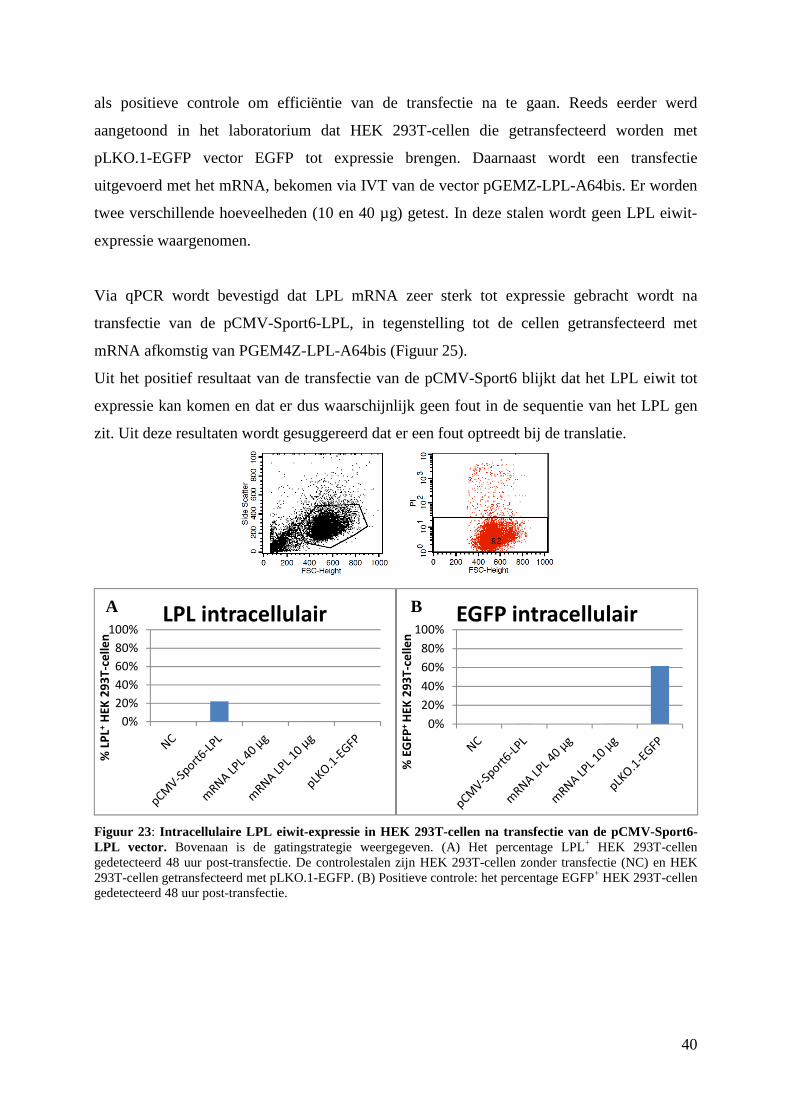
40
als positieve controle om efficiëntie van de transfectie na te gaan. Reeds eerder werd
aangetoond in het laboratorium dat HEK 293T-cellen die getransfecteerd worden met
pLKO.1-EGFP vector EGFP tot expressie brengen. Daarnaast wordt een transfectie
uitgevoerd met het mRNA, bekomen via IVT van de vector pGEMZ-LPL-A64bis. Er worden
twee verschillende hoeveelheden (10 en 40 µg) getest. In deze stalen wordt geen LPL eiwit-
expressie waargenomen.
Via qPCR wordt bevestigd dat LPL mRNA zeer sterk tot expressie gebracht wordt na
transfectie van de pCMV-Sport6-LPL, in tegenstelling tot de cellen getransfecteerd met
mRNA afkomstig van PGEM4Z-LPL-A64bis (Figuur 25).
Uit het positief resultaat van de transfectie van de pCMV-Sport6 blijkt dat het LPL eiwit tot
expressie kan komen en dat er dus waarschijnlijk geen fout in de sequentie van het LPL gen
zit. Uit deze resultaten wordt gesuggereerd dat er een fout optreedt bij de translatie.
Figuur 23: Intracellulaire LPL eiwit-expressie in HEK 293T-cellen na transfectie van de pCMV-Sport6-LPL vector. Bovenaan is de gatingstrategie weergegeven. (A) Het percentage LPL+ HEK 293T-cellen gedetecteerd 48 uur post-transfectie. De controlestalen zijn HEK 293T-cellen zonder transfectie (NC) en HEK 293T-cellen getransfecteerd met pLKO.1-EGFP. (B) Positieve controle: het percentage EGFP+ HEK 293T-cellen gedetecteerd 48 uur post-transfectie.
0%
20%
40%
60%
80%
100%
% E
GF
P+
HE
K 2
93
T-c
ell
en
EGFP intracellulair
0%
20%
40%
60%
80%
100%
% L
PL+
HE
K 2
93
T-c
ell
en
LPL intracellulairA B
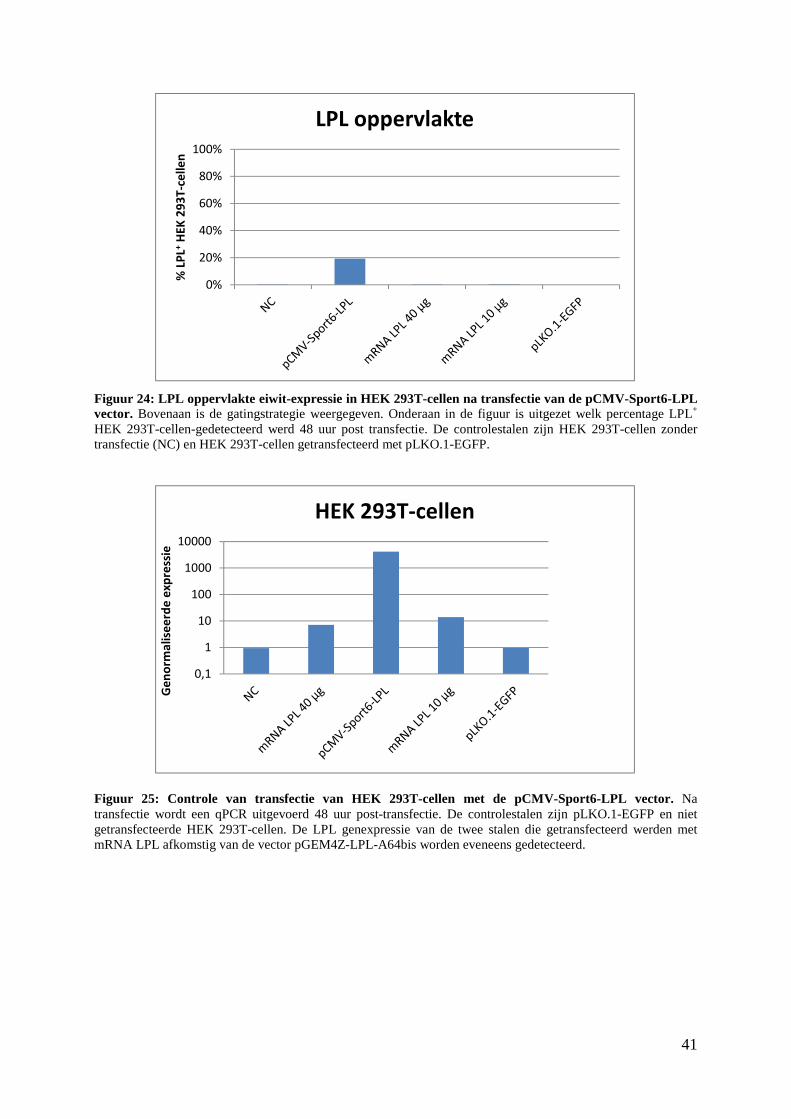
41
Figuur 24: LPL oppervlakte eiwit-expressie in HEK 293T-cellen na transfectie van de pCMV-Sport6-LPL vector. Bovenaan is de gatingstrategie weergegeven. Onderaan in de figuur is uitgezet welk percentage LPL+ HEK 293T-cellen-gedetecteerd werd 48 uur post transfectie. De controlestalen zijn HEK 293T-cellen zonder transfectie (NC) en HEK 293T-cellen getransfecteerd met pLKO.1-EGFP.
Figuur 25: Controle van transfectie van HEK 293T-cellen met de pCMV-Sport6-LPL vector. Na transfectie wordt een qPCR uitgevoerd 48 uur post-transfectie. De controlestalen zijn pLKO.1-EGFP en niet getransfecteerde HEK 293T-cellen. De LPL genexpressie van de twee stalen die getransfecteerd werden met mRNA LPL afkomstig van de vector pGEM4Z-LPL-A64bis worden eveneens gedetecteerd.
0%
20%
40%
60%
80%
100%
% L
PL+
HE
K 2
93
T-c
ell
en
LPL oppervlakte
0,1
1
10
100
1000
10000
Ge
no
rma
lise
erd
e e
xp
ress
ie
HEK 293T-cellen
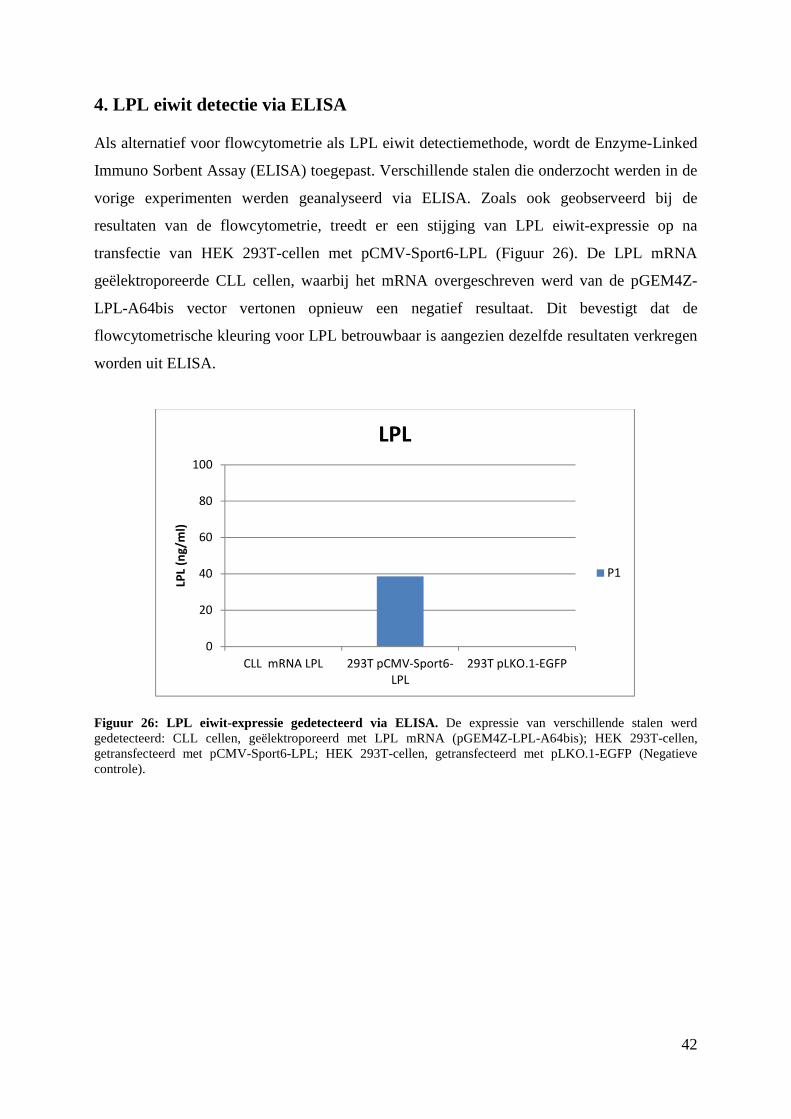
42
4. LPL eiwit detectie via ELISA
Als alternatief voor flowcytometrie als LPL eiwit detectiemethode, wordt de Enzyme-Linked
Immuno Sorbent Assay (ELISA) toegepast. Verschillende stalen die onderzocht werden in de
vorige experimenten werden geanalyseerd via ELISA. Zoals ook geobserveerd bij de
resultaten van de flowcytometrie, treedt er een stijging van LPL eiwit-expressie op na
transfectie van HEK 293T-cellen met pCMV-Sport6-LPL (Figuur 26). De LPL mRNA
geëlektroporeerde CLL cellen, waarbij het mRNA overgeschreven werd van de pGEM4Z-
LPL-A64bis vector vertonen opnieuw een negatief resultaat. Dit bevestigt dat de
flowcytometrische kleuring voor LPL betrouwbaar is aangezien dezelfde resultaten verkregen
worden uit ELISA.
Figuur 26: LPL eiwit-expressie gedetecteerd via ELISA. De expressie van verschillende stalen werd gedetecteerd: CLL cellen, geëlektroporeerd met LPL mRNA (pGEM4Z-LPL-A64bis); HEK 293T-cellen, getransfecteerd met pCMV-Sport6-LPL; HEK 293T-cellen, getransfecteerd met pLKO.1-EGFP (Negatieve controle).
0
20
40
60
80
100
CLL mRNA LPL 293T pCMV-Sport6-
LPL
293T pLKO.1-EGFP
LPL
(ng
/ml)
LPL
P1

43
IV. Discussie
Onderzoek toonde aan dat LPL een goede prognostische merker is voor CLL [35, 57, 60].
Hieruit kan afgeleid worden dat LPL een belangrijke functie heeft in het ziekteverloop van
CLL. Toch is er nauwelijks informatie gekend over de rol van LPL in CLL. LPL expressie is
bij CLL patiënten geassocieerd met een slechte prognose van de ziekte. Omdat er geen
stijging is in de enzymatische LPL activiteit in deze cellen, kan verondersteld worden dat LPL
eerder een brugfunctie zou vervullen in CLL cellen [49]. Hypothetisch gesteld zouden de
CLL cellen meer anti-apoptotische signalen kunnen ontvangen, door een LPL gemedieerde
versterkte interactie met de micro-omgeving. Op die manier kunnen de cellen sneller
accumuleren in de patiënt en bijdragen tot een agressiever ziekteverloop. In deze studie werd
een efficiënte manier gezocht om LPL tot expressie te brengen in LPL- CLL cellen. Dit zou
toelaten LPL- en LPL+ celpopulaties te vergelijken in een aantal biologische assays en zo de
rol van LPL in de CLL pathogenese te achterhalen.
Om efficiënt de functie en de gevolgen van LPL expressie te onderzoeken, werd er gebruik
gemaakt van een methode om LPL mRNA te elektroporeren in LPL- CLL cellen. De
effectiviteit van elektroporatie is reeds beschreven voor ZAP-70, een andere prognostische
factor in CLL [54]. De constructie van de pGEM4Z-ZAP70-A64bis vector werd in hun studie
bekomen vertrekkend van de pGEM4Z-EGFP-A64bis en pcDNA3-ZAP70. Via IVT kon
ZAP-70 mRNA afgeschreven worden van deze vector om vervolgens dit mRNA in cellen te
transfecteren via elektroporatie. Van Bockstaele et al. toonden zowel aan de hand van
flowcytometrie als een western blot analyse een overexpressie van ZAP-70 eiwit in CLL
cellen aan [54].
In dit onderzoek werd LPL mRNA bereid via IVT van de aangekochte pCMV-Sport6-LPL
vector. Via flowcytometrie werd echter geen intracellulaire LPL eiwit-expressie in de LPL
geëlektroporeerde cellen aangetoond. De CLL cellen geëlektroporeerd met EGFP (positieve
controle) gaven wel een duidelijk expressie van het EGFP eiwit. Om na te gaan of de
elektroporatie gelukt was, werd een qPCR uitgevoerd die aantoonde dat het LPL mRNA
duidelijk aanwezig is in de CLL cellen na elektroporatie. Deze bevindingen wezen eerder op
fouten bij de translatie van het IVT mRNA tot het LPL eiwit. De afwezigheid van LPL eiwit-
expressie kon te wijten zijn aan de in vitro polyA-tailing van het mRNA construct. De
pCMV-SPORT6-LPL vector bevat namelijk het LPL gen zonder polyA-staart. Daarom wordt
na IVT, in vitro polyA tailing uitgevoerd op het LPL mRNA. Indien er fouten gebeuren bij de

44
in vitro polyA-tailing zal de stabiliteit van het mRNA verloren gaan. Deze polyA-staart is
eveneens cruciaal voor de initiatie van de translatie van LPL mRNA.
Om de in vitro polyA-tailing te vermijden, wordt de pGEM4Z-LPL-A64bis vector, die reeds
een polyA-staart bevat, gebruikt. Van bockstaele et al. hebben de efficiëntie van deze vector
voor IVT mRNA elektroporatie reeds aangetoond [54]. Het LPL gen werd gekloneerd van de
pCMV-Sport6-LPL vector in de pGEM4Z-EGFP-A64bis vector. Na intracellulaire en
oppervlaktekleuring werd ook hier echter geen eiwit-expressie aangetoond via
flowcytometrie. De efficiëntie van de elektroporatie werd opnieuw bevestigd aan de hand van
de positieve controle (EGFP). Na IVT en LPL mRNA elektroporatie werd echter enkel LPL
mRNA gedetecteerd via qPCR 24 uur na elektroporatie. Dit wijst op een fout bij het uitvoeren
van het protocol. De resultaten bekomen 24 uur na elektroporatie, tonen aan dat LPL mRNA
elektroporatie van de CLL cel niet noodzakelijk impliceert dat er eiwit-expressie plaatsvindt.
Op basis van dit resultaat werd ook de flowcytometrische kleuring voor LPL in vraag gesteld.
Daarom werd de eiwit-expressie nagegaan op gestimuleerde CLL-cellen. Pallasch et al. [48]
toonden reeds aan dat LPL expressie geïnduceerd wordt na stimulatie van CLL cellen in
vergelijking met gezonde CD5+ B-cellen. Daarbij werd ook een hogere expressie van LPL in
ongemuteerde CLL-cellen waargenomen in vergelijking met gemuteerde CLL cellen. Deze
bevinding werd bevestigd in ons experiment. Bij IgM gestimuleerde CLL cellen van de
gemuteerde en ongemuteerde patiënt werd een gestegen expressie van LPL mRNA ten
opzichte van de IgA gestimuleerde CLL cellen
Om de eiwit-expressie na te gaan van LPL, werd een flowcytometrische analyse toegepast.
Het aandeel LPL+ B-cellen is licht, doch significant, gestegen bij IgM gestimuleerde CLL
cellen ten opzichte van de IgA gestimuleerde CLL cellen (controle). Deze flowcytometrische
resultaten zijn minder duidelijk dan de qPCR. Hieruit kan verondersteld worden dat de
oppervlakte eiwit-expressie niet overeenkomt met de expressie van mRNA LPL. Heintel et al
hadden dit al eerder gesuggereerd in hun onderzoek [35]. Zij maakten eveneens gebruik van
flowcytometrie om LPL eiwit-expressie op het oppervlak van CLL cellen aan te tonen. Ze
toonden via qPCR aan dat LPL expressie significant hoger is bij CLL patiënten met een
ongunstige prognose en dus bij CLL cellen met een ongemuteerde IgVH status. Toch was na
flowcytometrische oppervlakte kleuring de gemiddelde fluorescentie intensiteit voor LPL
slechts lichtjes lager in de gemuteerde patiënten in vergelijking met de ongemuteerde
patiënten.

45
Aangezien de stalen afkomstig waren van twee patiënten, is de steekproef te klein. Om een
meer betrouwbaar resultaat te bekomen, zouden er in de toekomst meerdere CLL stalen
onderzocht moeten worden op LPL eiwit-expressie via flowcytometrie. Dit resultaat geeft
onvoldoende aan of flowcytometrische detectie van LPL een betrouwbare methode is.
Daarom werd de flowcytometrische analyse ook toegepast op LPL expresserende HL-60
cellen. Bij deze cellen werd een duidelijke verschuiving van LPL fluorescentie intensiteit
waargenomen ten opzichte van de achtergrond intensiteit. Dit bevestigt het vermoeden dat de
snelle en eenvoudige analyse met flowcytometrie een goede detectiemethode voor LPL eiwit-
expressie is.
Een ander mogelijke verklaring voor de negatieve resultaten inzake eiwit-expressie na mRNA
elektroporatie is een fout in de sequentie van LPL in de vector pCMV-Sport6-LPL. Daarom
werd de sequentie van het LPL gen op de pCMV-Sport6-LPL vector geanalyseerd. Er werden
geen mutaties of andere fouten gevonden in deze sequentie. Er wordt nagegaan of LPL eiwit-
expressie vertrekkende van de pCMV-Sport6-LPL vector mogelijk is in HEK 293T-cellen.
Via transfectie werd de vector in de kern gebracht. Vervolgens werd door middel van
flowcytometrie een stijging van LPL eiwit-expressie geobserveerd na transfectie. Dit
bevestigt dat de sequentie van het LPL gen correct is en translatie van het eiwit goed verloopt.
Dit resultaat bevestigt nog eens dat fouten in de flowcytometrische analyse niet waarschijnlijk
zijn.
Om na te gaan of er in de toekomst een alternatief bestaat voor flowcytometrie als
detectiemethode van LPL eiwit-expressie, werd een ELISA toegepast op verschillende stalen.
Zoals ook geobserveerd via flowcytometrische analyse, wordt er geen stijging van LPL eiwit-
expressie waargenomen na elektroporatie van CLL cellen met LPL mRNA (pGEM4Z-LPL-
A64bis). Er werd wel een stijging in eiwit-expressie waargenomen na transfectie van HEK
293T-cellen met de pCMV-Sport6-LPL vector. Dit bevestigt de resultaten van de
flowcytometrische analyse. Bovendien kunnen in de toekomst verschillende stalen
geanalyseerd worden via beide detectiemethoden om vals negatieve of vals positieve
resultaten zoveel mogelijk uit te sluiten.
Uit voorgaande resultaten kan verondersteld worden dat er problemen zijn opgetreden tijdens
de IVT. Daarom zal in de toekomst het IVT protocol opnieuw uitgevoerd vertrekkende van de

46
pGEM4Z-LPL-A64bis. Indien dit opnieuw een negatief resultaat geeft, zal de volledige
klonering van de pGEM4Z-LPL-A64bis herstart worden.
Als alternatief zal er eveneens geprobeerd worden om CLL cellen te transduceren. Frecha et
al. [61] toonden aan dat lentivirale vectoren geïncorporeerd met hemagglutinin- en fusion-
glycoproteinen, van het mazelenvirus ongeveer 50% van de B-cellen in rusttoestand kunnen
transduceren. Ook CLL cellen, geblokkeerd in de G0/G1 fase van de celcyclus werden
efficiënt getransduceerd met deze lentivirale vectoren. Dit protocol werd reeds getest in het
lab en bleek inderdaad een succes. Klonering van het LPL gen in de gebruikte transfervector,
laat mogelijks toe om op deze manier LPL in de CLL cellen te brengen en de effecten van dit
eiwit te analyseren.
Uit deze resultaten kan geconcludeerd worden dat er nog veel onderzoek verricht moet
worden om LPL op een efficiënte manier in de CLL cellen te transfecteren. De synthese van
IVT mRNA vergt weinig tijd in tegenstelling tot de labo-intensieve lentivirale transductie
methode. Daarbij is het een veilige en efficiënte methode om CLL cellen te transfecteren [54].
Aangezien deze gen transfer methode in de toekomst functionele studies naar LPL in CLL
kan vergemakkelijken, is verder onderzoek naar de eerder beschreven fouten in de eiwit-
expressie na LPL mRNA elektroporatie noodzakelijk.

47
V. Referenties
1. Dighiero, G., CLL biology and prognosis. Hematology Am Soc Hematol Educ Program, 2005: p. 278-84.
2. Weiss, N.S., Geographical variation in the incidence of the leukemias and lymphomas. Natl Cancer Inst Monogr, 1979(53): p. 139-42.
3. Altekruse, S.F., et al. SEER Cancer Statistics Review. 2010 2009; Available from: http://seer.cancer.gov/csr/1975_2007/
4. Ginaldi, L., et al., Levels of expression of CD19 and CD20 in chronic B cell leukaemias. Journal of clinical pathology, 1998. 51(5): p. 364-9.
5. Moreau, E.J., et al., Improvement of the chronic lymphocytic leukemia scoring system with the monoclonal antibody SN8 (CD79b). American journal of clinical pathology, 1997. 108(4): p. 378-82.
6. Kipps, T.J., Chronic Lymphocytic Leukemia and Related Diseases, in Williams Hematology. p. 2460. 7. Hanada, M., et al., bcl-2 gene hypomethylation and high-level expression in B-cell chronic lymphocytic
leukemia. Blood, 1993. 82(6): p. 1820-8. 8. Gottardi, D., et al., In leukaemic CD5+ B cells the expression of BCL-2 gene family is shifted toward
protection from apoptosis. British journal of haematology, 1996. 94(4): p. 612-8. 9. Ripolles, L., et al., Genetic abnormalities and clinical outcome in chronic lymphocytic leukemia.
Cancer Genet Cytogenet, 2006. 171(1): p. 57-64. 10. Messmer, B.T., et al., In vivo measurements document the dynamic cellular kinetics of chronic
lymphocytic leukemia B cells. The Journal of clinical investigation, 2005. 115(3): p. 755-64. 11. Kater, A.P., et al., Dutch guidelines for diagnosis and treatment of chronic lymphocytic leukaemia
2011. Neth J Med, 2011. 69(10): p. 422-9. 12. Byrd, J.C., S. Stilgenbauer, and I.W. Flinn, Chronic lymphocytic leukemia. Hematology Am Soc
Hematol Educ Program, 2004: p. 163-83. 13. Hallek, M., et al., Addition of rituximab to fludarabine and cyclophosphamide in patients with chronic
lymphocytic leukaemia: a randomised, open-label, phase 3 trial. Lancet, 2010. 376(9747): p. 1164-74. 14. Eichhorst, B.F., et al., First-line therapy with fludarabine compared with chlorambucil does not result
in a major benefit for elderly patients with advanced chronic lymphocytic leukemia. Blood, 2009. 114(16): p. 3382-91.
15. Moreno, C., et al., Allogeneic stem-cell transplantation may overcome the adverse prognosis of unmutated VH gene in patients with chronic lymphocytic leukemia. Journal of Clinical Oncology, 2005. 23(15): p. 3433-3438.
16. Dreger, P., et al., Indications for allogeneic stem cell transplantation in chronic lymphocytic leukemia: the EBMT transplant consensus. Leukemia : official journal of the Leukemia Society of America, Leukemia Research Fund, U.K, 2007. 21(1): p. 12-7.
17. Hallek, M., et al., Guidelines for the diagnosis and treatment of chronic lymphocytic leukemia: a report from the International Workshop on Chronic Lymphocytic Leukemia updating the National Cancer Institute-Working Group 1996 guidelines. Blood, 2008. 111(12): p. 5446-56.
18. Gribben, J.G., How I treat CLL up front. Blood, 2010. 115(2): p. 187-97. 19. Murphy, K.P., P. Travers, and M. Walport, Immunobiology. 7th ed2007. 20. Rai, K.R., et al., Clinical staging of chronic lymphocytic leukemia. Blood, 1975. 46(2): p. 219-34. 21. Binet, J.L., et al., A clinical staging system for chronic lymphocytic leukemia: prognostic significance.
Cancer, 1977. 40(2): p. 855-64. 22. Van Bockstaele, F., B. Verhasselt, and J. Philippe, Prognostic markers in chronic lymphocytic
leukemia: a comprehensive review. Blood Rev, 2009. 23(1): p. 25-47. 23. Calin, G.A., et al., Frequent deletions and down-regulation of micro- RNA genes miR15 and miR16 at
13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci U S A, 2002. 99(24): p. 15524-9. 24. Calin, G.A., et al., MiR-15a and miR-16-1 cluster functions in human leukemia. Proc Natl Acad Sci U S
A, 2008. 105(13): p. 5166-71. 25. Klein, U. and R. Dalla-Favera, New insights into the pathogenesis of chronic lymphocytic leukemia.
Semin Cancer Biol, 2010. 20(6): p. 377-83. 26. Mertens, D., et al., Down-regulation of candidate tumor suppressor genes within chromosome band
13q14.3 is independent of the DNA methylation pattern in B-cell chronic lymphocytic leukemia. Blood, 2002. 99(11): p. 4116-21.
27. Klein, U., et al., The DLEU2/miR-15a/16-1 cluster controls B cell proliferation and its deletion leads to chronic lymphocytic leukemia. Cancer Cell, 2010. 17(1): p. 28-40.
28. Stilgenbauer, S., et al., Molecular cytogenetic analysis of B-cell chronic lymphocytic leukemia. Ann Hematol, 1998. 76(3-4): p. 101-10.

48
29. Damle, R.N., et al., Ig V gene mutation status and CD38 expression as novel prognostic indicators in chronic lymphocytic leukemia. Blood, 1999. 94(6): p. 1840-7.
30. Hamblin, T.J., et al., Unmutated Ig V(H) genes are associated with a more aggressive form of chronic lymphocytic leukemia. Blood, 1999. 94(6): p. 1848-54.
31. Ghia, P., et al., ERIC recommendations on IGHV gene mutational status analysis in chronic lymphocytic leukemia. Leukemia, 2007. 21(1): p. 1-3.
32. Crespo, M., et al., ZAP-70 expression as a surrogate for immunoglobulin-variable-region mutations in chronic lymphocytic leukemia. N Engl J Med, 2003. 348(18): p. 1764-75.
33. Chen, L., et al., Expression of ZAP-70 is associated with increased B-cell receptor signaling in chronic lymphocytic leukemia. Blood, 2002. 100(13): p. 4609-14.
34. Wiestner, A., et al., ZAP-70 expression identifies a chronic lymphocytic leukemia subtype with unmutated immunoglobulin genes, inferior clinical outcome, and distinct gene expression profile. Blood, 2003. 101(12): p. 4944-51.
35. Heintel, D., et al., High expression of lipoprotein lipase in poor risk B-cell chronic lymphocytic leukemia. Leukemia : official journal of the Leukemia Society of America, Leukemia Research Fund, U.K, 2005. 19(7): p. 1216-23.
36. Deaglio, S., et al., Human CD38 (ADP-ribosyl cyclase) is a counter-receptor of CD31, an Ig superfamily member. J Immunol, 1998. 160(1): p. 395-402.
37. Deaglio, S., et al., CD38 and ZAP-70 are functionally linked and mark CLL cells with high migratory potential. Blood, 2007. 110(12): p. 4012-21.
38. Fischer, A., et al., ZAP70: a master regulator of adaptive immunity. Semin Immunopathol, 2010. 32(2): p. 107-16.
39. Pede, V., et al., The biological Relevance of ZAP-70 in CLL, in Chronic Lymphocytic Leukemia, P. Oppezzo, Editor 2012, InTech. p. 448.
40. Mathonnet, G., et al., MicroRNA inhibition of translation initiation in vitro by targeting the cap-binding complex eIF4F. Science, 2007. 317(5845): p. 1764-7.
41. Calin, G.A., et al., A MicroRNA signature associated with prognosis and progression in chronic lymphocytic leukemia. N Engl J Med, 2005. 353(17): p. 1793-801.
42. Mraz, M., et al., MicroRNAs in chronic lymphocytic leukemia pathogenesis and disease subtypes. Leuk Lymphoma, 2009. 50(3): p. 506-9.
43. Oppezzo, P., et al., The LPL/ADAM29 expression ratio is a novel prognosis indicator in chronic lymphocytic leukemia. Blood, 2005. 106(2): p. 650-7.
44. Mead, J.R., S.A. Irvine, and D.P. Ramji, Lipoprotein lipase: structure, function, regulation, and role in disease. J Mol Med (Berl), 2002. 80(12): p. 753-69.
45. Obunike, J.C., et al., Lipoprotein lipase can function as a monocyte adhesion protein. Arterioscler Thromb Vasc Biol, 1997. 17(7): p. 1414-20.
46. Schaefer, J.R., Potential Polymorphisms of the VLDL Receptor and other Obesity Candidate Genes in PAtients with and without Coronary Artery Disease, in Obesity and Related DisordersUnknown, Philipps University.
47. Xu, W., et al., Expression level of lipoprotein lipase in Chinese patients with chronic lymphocytic leukemia and its correlation with other prognostic factors. Int J Lab Hematol, 2009. 31(5): p. 552-9.
48. Pallasch, C.P., et al., Targeting lipid metabolism by the lipoprotein lipase inhibitor orlistat results in apoptosis of B-cell chronic lymphocytic leukemia cells. Leukemia, 2008. 22(3): p. 585-92.
49. Mansouri, M., et al., Lipoprotein lipase is differentially expressed in prognostic subsets of chronic lymphocytic leukemia but displays invariably low catalytical activity. Leuk Res, 2010. 34(3): p. 301-6.
50. Munk Pedersen, I. and J. Reed, Microenvironmental interactions and survival of CLL B-cells. Leuk Lymphoma, 2004. 45(12): p. 2365-72.
51. Panayiotidis, P., et al., Human bone marrow stromal cells prevent apoptosis and support the survival of chronic lymphocytic leukaemia cells in vitro. Br J Haematol, 1996. 92(1): p. 97-103.
52. Chilosi, M., et al., Immunohistochemical demonstration of follicular dendritic cells in bone marrow involvement of B-cell chronic lymphocytic leukemia. Cancer, 1985. 56(2): p. 328-32.
53. Lookene, A., N. Skottova, and G. Olivecrona, Interactions of lipoprotein lipase with the active-site inhibitor tetrahydrolipstatin (Orlistat). European journal of biochemistry / FEBS, 1994. 222(2): p. 395-403.
54. Van Bockstaele, F., et al., Efficient gene transfer in CLL by mRNA electroporation. Leukemia : official journal of the Leukemia Society of America, Leukemia Research Fund, U.K, 2008. 22(2): p. 323-9.
55. Cheung, V.G., et al., Polymorphic cis- and trans-regulation of human gene expression. PLoS Biol, 2010. 8(9).

49
56. Auwerx, J.H., et al., Transcriptional activation of the lipoprotein lipase and apolipoprotein E genes accompanies differentiation in some human macrophage-like cell lines. Biochemistry, 1988. 27(8): p. 2651-5.
57. Van Bockstaele, F., et al., Lipoprotein lipase mRNA expression in whole blood is a prognostic marker in B cell chronic lymphocytic leukemia. Clinical chemistry, 2007. 53(2): p. 204-12.
58. Shu, J., S.H. Benedict, and M.A. Chan, Anti-IgM and anti-IgG modulate c-fos RNA levels differently in human B lymphocytes. Cellular immunology, 1994. 159(2): p. 170-83.
59. Vlad, A., et al., Down-regulation of CXCR4 and CD62L in chronic lymphocytic leukemia cells is triggered by B-cell receptor ligation and associated with progressive disease. Cancer research, 2009. 69(16): p. 6387-95.
60. Kaderi, M.A., et al., LPL is the strongest prognostic factor in a comparative analysis of RNA-based markers in early chronic lymphocytic leukemia. Haematologica, 2011. 96(8): p. 1153-60.
61. Frecha, C., et al., Efficient and stable transduction of resting B lymphocytes and primary chronic lymphocyte leukemia cells using measles virus gp displaying lentiviral vectors. Blood, 2009. 114(15): p. 3173-80.
Recommended

















