Famiglia dei Luteoviridae
Classificazione Gruppo IV (+ssRNA)
Ordine non assegnato
Famiglia Luteoviridae
Generi appartenenti alla famiglia Luteoviridae
1. Enamovirus, specie tipo Pea enation mosaic virus 1 (PEMV)
2. Luteovirus, specie tipo Barley yellow dwarf virus (BYDV)
3. Polerovirus, specie tipo Potato leafroll virus (PLRV)
4. Generi non assegnati.
a. Genere Enamovirus
1. Pea enation mosaic virus 1
b. Genere Luteovirus
1. Barley yellow dwarf virus-kerII
2. Barley yellow dwarf virus-kerIII
3. Barley yellow dwarf virus-MAV
4. Barley yellow dwarf virus-PAS
5. Barley yellow dwarf virus-PAV
6. Bean leafroll virus
7. Rose spring dwarf-associated virus
8. Soybean dwarf virus
c. Genere Polerovirus
1. Beet chlorosis virus
2. Beet mild yellowing virus
3. Beet western yellows virus
4. Carrot red leaf virus
5. Cereal yellow dwarf virus-RPS
6. Cereal yellow dwarf virus-RPV
7. Chickpea chlorotic stunt virus
8. Cotton leafroll dwarf virus
9. Cucurbit aphid-borne yellows virus
10. Maize yellow dwarf virus-RMV
11. Melon aphid-borne yellows virus
12. Pepper vein yellows virus
13. Potato leafroll virus
14. Suakwa aphid-borne yellows virus
15. Sugarcane yellow leaf virus
16. Tobacco vein distorting virus
17. Turnip yellows virus
d. Generi non assegnati
1. Barley yellow dwarf virus-GPV
2. Barley yellow dwarf virus-SGV

2
3. Chickpea stunt disease associated virus
4. Groundnut rosette assistor virus
5. Indonesian soybean dwarf virus
6. Sweet potato leaf speckling virus
7. Tobacco necrotic dwarf virus
Nella famiglia Luteoviridae vi sono 33 specie, divise in 3 generi, più un quarto gruppo costituito da
generi non ancora assegnati.
Luteoviridae è una famiglia di virus RNA a singolo filamento positivo.
Le piante servono come ospiti naturali. Il nome della famiglia deriva dalla parola latina virga (asta),
in quanto tutti i virus di questa famiglia sono a forma di asta, bacilliformi.
I virus della famiglia Virgaviridae sono caratterizzati da unica proteina di replicazione simile alle
proteine alfa.
I virus dei Luteoviridae non sono avvolti dal pericapside e presentano una geometria sferica con
virioni di circa 25-30 nm di diametro, con una simmetria icosaedrica formata da 180 proteine della
coat protein (figura 1).
Figura 1 – Virione di forma icosaedrica (a destra) di Luteoviridae. A sinistra si osserva
lo spessore della proteina di rivestimento CP (coat protein) e un (+)ssRNA.
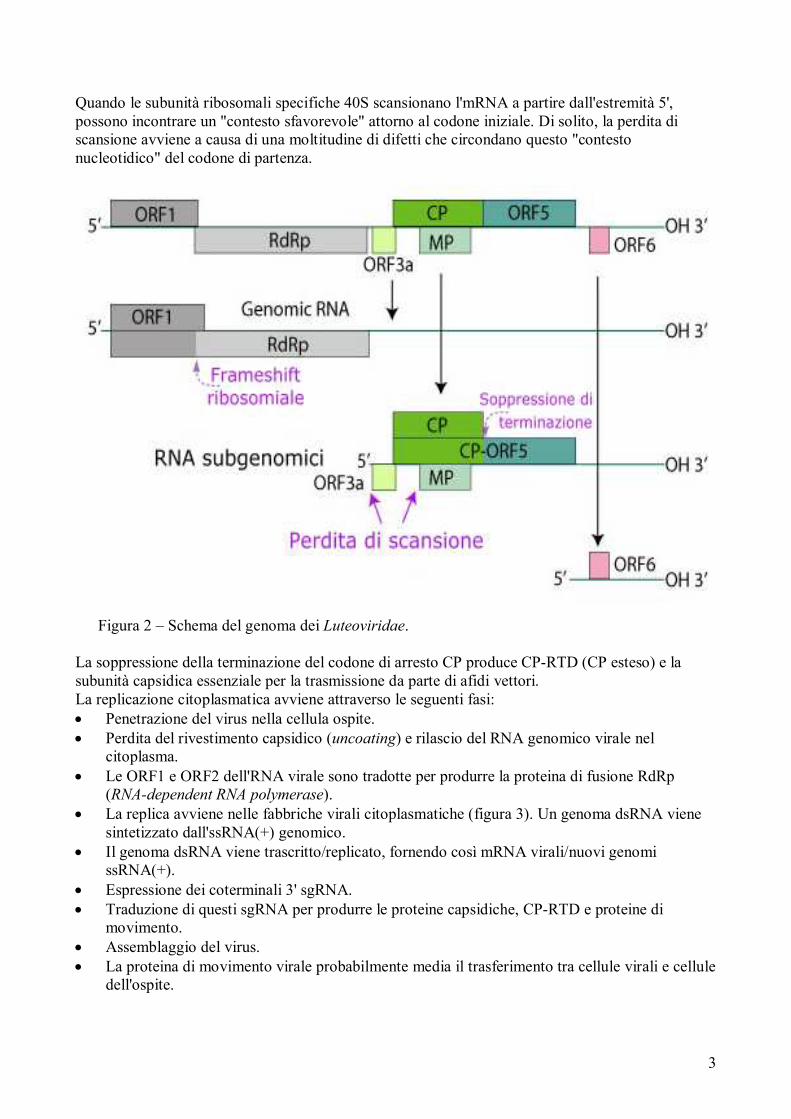
ssRNA (+) del genoma è monopartito, lineare, di 5,3-5,7 kb. I polerovirus ed enamovirus hanno un
VPg (viral protein genome) legato al terminale 5 '. Non esiste una struttura di poli (A) o simile a
tRNA al terminale 3' (figura 2).
L'RNA del virione è infettivo e funziona come genoma e come RNA messaggero. La polimerasi
virale è espressa da un frameshifting ribosomale alla fine di ORF1.
A seconda del genere, diverse proteine possono essere prodotte mediante la perdita di scansione.
Quest’ultima è un meccanismo utilizzato nella traduzione di cellule eucariotiche che consente per
40s subunità ribosomiali di bypassare il codone di avvio iniziale di AUG (adenina-uracile-guanina)
e di iniziare la traduzione in ulteriori codoni di inizio AUG a valle.
La traduzione in cellule eucariotiche secondo la maggior parte dei meccanismi di scansione si
verifica al codone di inizio AUG prossimale all'estremità 5 'di mRNA.
3
Quando le subunità ribosomali specifiche 40S scansionano l'mRNA a partire dall'estremità 5',
possono incontrare un "contesto sfavorevole" attorno al codone iniziale. Di solito, la perdita di
scansione avviene a causa di una moltitudine di difetti che circondano questo "contesto
nucleotidico" del codone di partenza.
Figura 2 – Schema del genoma dei Luteoviridae.
La soppressione della terminazione del codone di arresto CP produce CP-RTD (CP esteso) e la
subunità capsidica essenziale per la trasmissione da parte di afidi vettori.
La replicazione citoplasmatica avviene attraverso le seguenti fasi:
• Penetrazione del virus nella cellula ospite.
• Perdita del rivestimento capsidico (uncoating) e rilascio del RNA genomico virale nel
citoplasma.
• Le ORF1 e ORF2 dell'RNA virale sono tradotte per produrre la proteina di fusione RdRp
(RNA-dependent RNA polymerase).
• La replica avviene nelle fabbriche virali citoplasmatiche (figura 3). Un genoma dsRNA viene
sintetizzato dall'ssRNA(+) genomico.
• Il genoma dsRNA viene trascritto/replicato, fornendo così mRNA virali/nuovi genomi
ssRNA(+).
• Espressione dei coterminali 3' sgRNA.
• Traduzione di questi sgRNA per produrre le proteine capsidiche, CP-RTD e proteine di
movimento.
• Assemblaggio del virus.
• La proteina di movimento virale probabilmente media il trasferimento tra cellule virali e cellule
dell'ospite.

4
Figura 3 - Le fabbriche virali sono compartimenti intracellulari (inclusioni) che aumentano l'efficienza
della replicazione virale e lo proteggono dalle difese ospitanti. Le fabbriche virali possono
essere citoplasmatiche o nucleari e spesso derivano da una riconversione estesa di
compartimenti citoscheletrici e/o cellulari delle cellule ospiti.
Il viroplasma (figura 3) costituisce delle dense inclusioni elettroniche citoplasmatiche in cui
avviene la replicazione virale e l'assemblaggio. Queste sono prodote dai virus nucleo-cytoplasmic
large DNA (NCLDV) come i Poxviridae, Asfarviridae e Iridoviridae, virus dsRNA come i
Reoviridae ssRNA (-) e come i Filoviridae.
Le sferule (tutte quelle di colore marrone che vediamo nella figura 3) sono invaginazioni della
membrana di diametro di 50-400 nm, in cui avviene la replicazione virale. Possono apparire in
diversi componenti cellulari, avvolte a seconda del virus: Tymoviruses nei cloroplasti, Tombusvirus
nei perossisomi, Nodaviridae nei mitocondri, Togaviridae negli endosomi e lisosomi e Flaviviridae
nel reticolo endoplasmatico. Tutti questi virus hanno un intermediario replicativo dsRNA e possono
replicarsi in compartimenti isolati per essere nascosti dall'immunità cellulare innata di dsRNA.
Le vescicole della doppia membrana (DMV) sono strutture membranose derivate dal reticolo
endoplasmatico o Golgi aventi un diametro di circa 200-300 nm (figura 3). Sono prodotte dalle
famiglie dei Picornaviridae e Nidovirales, Arteriviridae e Coronaviridae. Questi virus producono
dsRNA intermedio durante la replicazione e possono replicarsi in compartimenti isolati per essere
nascosti ai sensori dsRNA dell’immunità cellulare. Le strutture DMV dei Nidovirales sono
collegate da una sola rete di membrane derivate da ER.
I tubi virali (figura 3 e figura 4) costituiscono una struttura a membrana derivata dall’apparato del
Golgi, in cui avviene la replicazione nel Bunyaviridae. I tubi sono chiusi all’assemblaggio e la loro
funzione può essere quella di collegare la replicazione virale e la morfogenesi all'interno delle
fabbriche virali.

5
Figura 4 - Modello di struttura funzionale dei tubi virali integrati nella
fabbrica virale. I tubi virali si assemblano in associazione
con le membrane del Golgi, usando actina endogena e NSm
polimerizzata come impalcatura. I tubi si collegano alle
cisterne RER e ai mitocondri che forniscono i fattori
cellulari necessari per la replicazione virale e
l'assemblaggio. La polimerasi virale (L) si concentra nel
dominio globulare. L'RNA virale replicato (fase 1), viene
protetto dall'interazione con le molecole N, abbondanti
intorno alle membrane Golgi e nei tubi interni. Le molecole
N entrano nei tubi attraverso le loro aperture al citoplasma.e
con l'aiuto dell’actimiosina assemblano gli RNP e quindi si
spostano verso il citoplasma (fase 2), dove si legano ai
domini citoplasmatici delle glicoproteine virali (Gc/Gn),
concentrate nelle vicine membrane del Golgi. Nuovi eventi
(fase 3) portano a formare particelle virali che incorporano,
all'interno, alcune quantità di proteine delle impalcature di
NSm e di actina. Particelle virali immature assemblate
sono, quindi, pronte per la maturazione e per la loro
escrezione (fase 4).
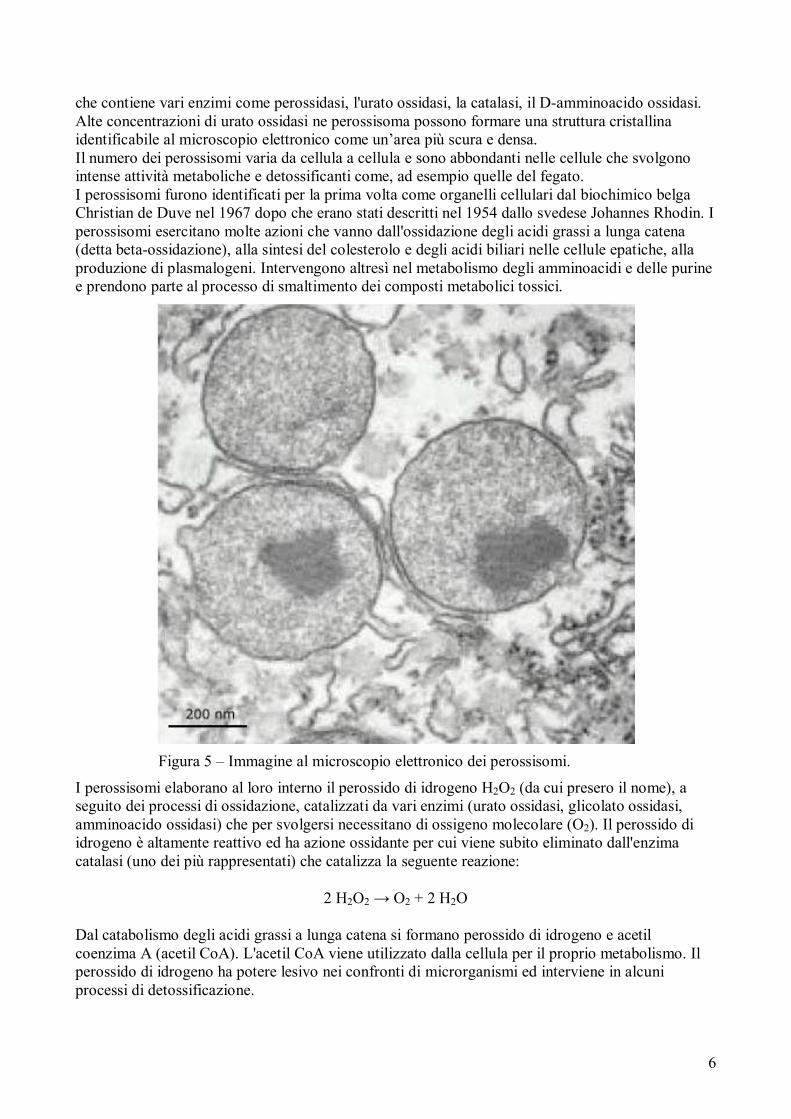
Il perossisoma è un organello cellulare vescicolare di circa 0,5-1 µm di diametro, ubiquitario negli
eucarioti (ad eccezione degli eritrociti), separato dal citoplasma da una membrana (figura 5) che
contiene almeno 50 enzimi ossidativi. In generale i perossisomi sono considerati comparti
metabolici specializzati, contenenti enzimi in grado di trasferire idrogeno da diverse sostanze e
legarlo all'ossigeno per la formazione di perossido di idrogeno (H2O2). In una cellula epatica vi
possono essere fino a 600 perossisomi all'interno dei quali è a volte rintracciabile un nucleo denso
6
che contiene vari enzimi come perossidasi, l'urato ossidasi, la catalasi, il D-amminoacido ossidasi.
Alte concentrazioni di urato ossidasi ne perossisoma possono formare una struttura cristallina
identificabile al microscopio elettronico come un’area più scura e densa.
Il numero dei perossisomi varia da cellula a cellula e sono abbondanti nelle cellule che svolgono
intense attività metaboliche e detossificanti come, ad esempio quelle del fegato.
I perossisomi furono identificati per la prima volta come organelli cellulari dal biochimico belga
Christian de Duve nel 1967 dopo che erano stati descritti nel 1954 dallo svedese Johannes Rhodin. I
perossisomi esercitano molte azioni che vanno dall'ossidazione degli acidi grassi a lunga catena
(detta beta-ossidazione), alla sintesi del colesterolo e degli acidi biliari nelle cellule epatiche, alla
produzione di plasmalogeni. Intervengono altresì nel metabolismo degli amminoacidi e delle purine
e prendono parte al processo di smaltimento dei composti metabolici tossici.
Figura 5 – Immagine al microscopio elettronico dei perossisomi.
I perossisomi elaborano al loro interno il perossido di idrogeno H2O2 (da cui presero il nome), a
seguito dei processi di ossidazione, catalizzati da vari enzimi (urato ossidasi, glicolato ossidasi,
amminoacido ossidasi) che per svolgersi necessitano di ossigeno molecolare (O2). Il perossido di
idrogeno è altamente reattivo ed ha azione ossidante per cui viene subito eliminato dall'enzima
catalasi (uno dei più rappresentati) che catalizza la seguente reazione:
2 H2O2 → O2 + 2 H2O
Dal catabolismo degli acidi grassi a lunga catena si formano perossido di idrogeno e acetil
coenzima A (acetil CoA). L'acetil CoA viene utilizzato dalla cellula per il proprio metabolismo. Il
perossido di idrogeno ha potere lesivo nei confronti di microrganismi ed interviene in alcuni
processi di detossificazione.

7
Il perossisoma contiene una membrana che separa il citoplasma cellulare dalla matrice, la parte
interna dell'organello. La membrana non ha solo funzione di barriera ma contiene molte proteine,
così come la matrice, necessarie per il funzionamento complessivo del perossisoma.
L'assemblaggio del perossisoma sembra avvenire in tre fasi:
1. all'inizio viene formata la membrana (pare che vi siano coinvolte almeno tre proteine PEX3,
PEX16, PEX19.),
2. successivamente vengono inserite le proteine transmembrana,
3. vengono, infine, inserite le proteine della matrice.
I perossisomi vengono assemblati a partire da vescicole che si distaccano dal reticolo
endoplasmatico (RE) e sono in grado di replicarsi per fissione binaria, pur non contenendo
materiale genetico.
Le proteine dei perossisomi, chiamate peroxine, vengono prodotte nel citoplasma ed importate nello
stesso perossisoma tramite il riconoscimento di una sequenza consenso della proteina da parte di un
recettore presente sulla membrana del perossisoma. In seguito all’importazione delle proteine, il
perossisoma aumenta di dimensioni e, raggiunto un punto critico, si divide per fissione binaria.
Si è detto che la membrana del perossisoma deriva dalla membrana dal RE. Difatti proteine
perossisomali di membrana sono state ritrovate in particolari regioni del RE. Si è pensato
inizialmente ad un problema di contaminazione, ma con l'avvento di nuove tecniche si è verificato
che queste proteine si trovano su particolari strutture del RE, dette estensioni lamellari. Queste
strutture sono visibili al microscopio e da queste prendono origine delle strutture tubulo-sacculari
che daranno luogo a un compartimento detto reticolo perossisomale. Da questo gemmano i
perossisomi che importano proteine della matrice.
Tutte le proteine del perossisoma vengono precedentemente sintetizzate nel citoplasma e
successivamente vengono condotte alla loro destinazione finale. Al momento si conoscono due
sequenze segnale (PTS1 e PTS2 da peroxisomal targeting signal) che vengono utilizzate per
indirizzare le proteine al perossisoma. Tali sequenze sono riconosciute da recettori specifici, PEX5
per PTS1 e PEX7 per PTS2, che trasportano la proteina interessata al perossisoma dove altre
proteine vengono utilizzate per effettuare il trasporto transmembrana fino alla matrice.
Il cloroplasto è un tipo di organulo presente nelle cellule dei vegetali (figura 6). All'interno di
questi organuli si svolge il processo della fotosintesi clorofilliana: l'energia luminosa viene catturata
dai pigmenti di clorofilla (e non solo) e viene convertita in energia chimica (ATP e NADPH).
Figura 6 – Schema di un cloroplasto.

8
Nonostante la loro importanza come agenti di malattie emergenti, la scala temporale ed i processi
evolutivi che formano l'aspetto di nuove specie virali sono in gran parte sconosciute.
Per affrontare queste problematiche, sono stati analizzati i processi evolutivi intra-e interspecifici
nella famiglia dei Luteoviridae.
Utilizzando il gene della proteina del capside di 12 membri della famiglia Luteoviridae , sono state
determinate le loro relazioni filogenetiche, i tassi di sostituzione dei nucleotidi, i tempi di origine
comune e modelli di speciazione.
Un’analisi multigenica associata ha permesso di dedurre la natura delle pressioni di selezione e la
distribuzione genomica degli eventi di ricombinazione.
Anche se i tassi di evoluzione, le variazioni e le pressioni di selezione variano tra i geni e le specie e
sono inferiori in alcune regioni sovrapposte del gene, tutte rientrano nell'ambito di quelle osservate
nei virus a RNA degli animali.
I punti di interruzione della ricombinazione sono comunemente osservati ai confini dei generi, ma
meno all'interno dei geni.
L’analisi dell'orologio molecolare ha suggerito che l’origine delle specie della famiglia dei
Luteoviridae, attualmente presenti in natura, si è verificata negli ultimi 4 millenni, con diversità
genetica intraspecifica che nasce negli ultimi cento anni.
La speciazione all’interno dei Luteoviridae può, quindi, essere associata all'espansione dei sistemi
agricoli.
Infine, l'analisi filogenetica ha suggerito che gli eventi di speciazione virale tendono a verificarsi
all'interno delle stesse specie di origine vegetale e del paese di origine, come è previsto se la
speciazione in natura è in gran parte simpatica, piuttosto che allopatrica.
L'endosoma è un corpo vescicolare, presente nella cellula, il cui compito è quello di partecipare
all'endocitosi, ovvero al meccanismo cellulare che permette il transito attraverso la membrana di
macromolecole e corpuscoli, le cui dimensioni non consentono l'ingresso attraverso i meccanismi
del trasporto di membrana.
L'interno del comparto endosomico si mantiene acido (pH 5-6) grazie alla pompa protonica
alimentata da ATP, presente nella membrana endosomica, la quale trasferisce protoni dal citosol al
lume. Questo compartimento funge da centrale di smistamento della via endocitica diretta verso
l'interno, proprio come il complesso di Golgi trans per la via secretoria diretta verso l'esterno.
L'ambiente acido ha un ruolo chiave nello smistamento, inducendo molti recettori a svincolarsi dal
loro bagaglio molecolare. Alcuni di questi usciranno per gemmazione come nel caso dei recettori
delle LDL (low density lipoprotein); altri finiranno nei lisosomi per essere degradati; altri ancora
subiranno un processo di transcitosi. Le proteine seguiranno il destino dei recettori rimanendoci
legate, altre finiranno degradate nei lisosomi, che poi è il destino della maggior parte del contenuto
endosomico. Resta ancora da capire in che modo le proteine dagli endosomi passino ai lisosomi. Si
ipotizza che il passaggio avvenga grazie a vescicole di trasporto o forse che gli endosomi stessi si
trasformino gradualmente in lisosomi.
L'endosoma corticale (o precoce) è una vescicola di 0,4-0,7 µm, situata nel citoplasma sottostante la
membrana plasmatica, con un pH interno pari a 6,0-6,2.
L'endosoma, infine, si divide in tardivo e precoce.
Precoce per via della grande vicinanza alla membrana plasmatica, mentre tardivo per la lontananza
dalla membrana plasmatica.
L’endosoma precoce si forma per trasformazione della vescicola rivestita in vescicola non rivestita
per perdita del rivestimento di clatrina. La clatrina è riciclata sotto la membrana plasmatica, mentre
la vescicola non rivestita si fonde con un endosoma a formare l’endosoma precoce (figura 7). Si
ricorda che la clatrina è una proteina esamerica, coinvolta nell’endocitosi mediata da recettori,
costituita da tre catene pesanti e da tre catene leggere, organizzate in una struttura a tre punte, detta
triskelion. I triskelion polimerizzano tra loro in modo da costituire un reticolo poliedrico formato da
unità esagonali e pentagonali; tale reticolo è presente, all’inizio dell’endocitosi, al di sotto delle
fossette rivestite e successivamente forma una gabbia che circonda le vescicole citoplasmatiche.
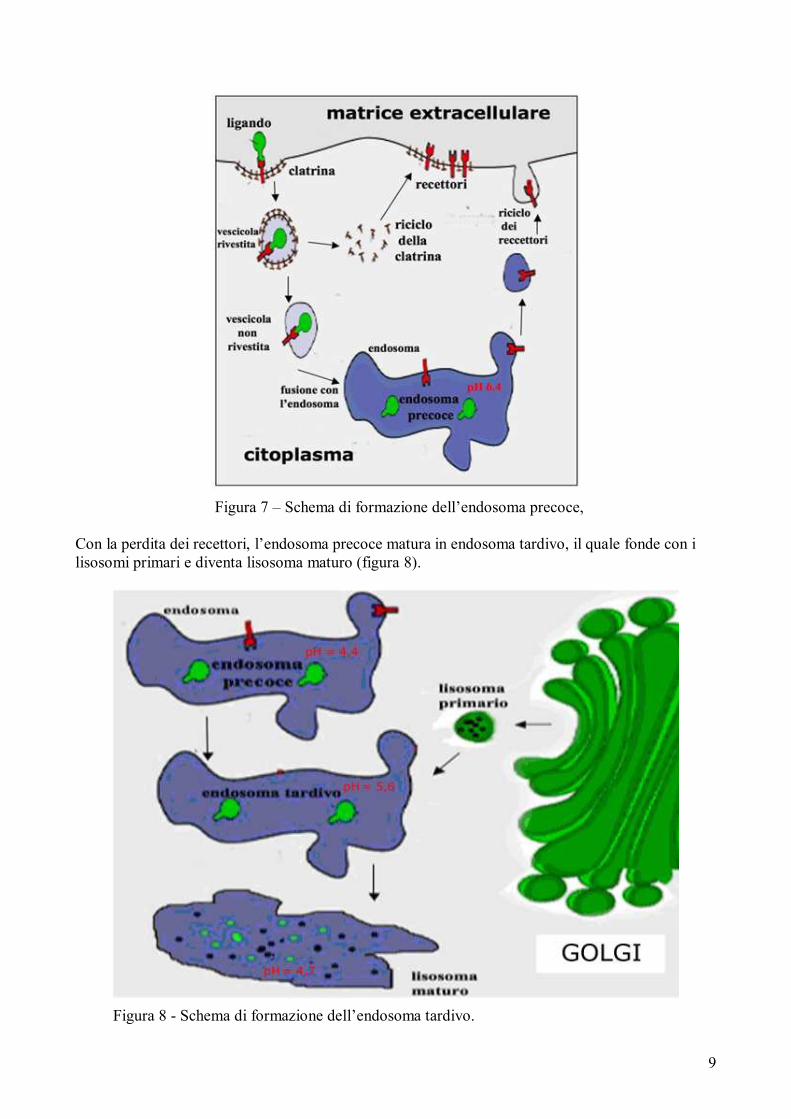
9
Figura 7 – Schema di formazione dell’endosoma precoce,
Con la perdita dei recettori, l’endosoma precoce matura in endosoma tardivo, il quale fonde con i
lisosomi primari e diventa lisosoma maturo (figura 8).
Figura 8 - Schema di formazione dell’endosoma tardivo.

10
I mitocondri, si ricorda, sono organelli cellulari presenti negli organismi eucarioti e sono
considerati la centrale energetica della cellula. Al loro interno avviene la respirazione cellulare, con
cui sono in grado di produrre grandi quantità di energia sotto forma di molecole di adenosina
trifosfato (ATP).
I mitocondri (figura 9), di forma allungata, con un diametro di 0,5-1 µm ed una lunghezza di 1-4
µm, assieme ai cloroplasti, sono gli unici organelli cellulari che contengono un proprio genoma (di
tipo procariotico). I mitocondri si riproducono all’interno della cellula in maniera indipendente.
I mitocondri si sono originati circa 1,5 miliardi di anni fa, in seguito ad una endosimbiosi avvenuta
tra i progenitori delle cellule eucariote con alcune cellule procariote in grado di utilizzare ossigeno
.
Figura 9 – Schema di un mitocondrio. Le creste mitocondriali hanno lo scopo di
aumentare la superficie di contatto, ai fini della respirazione cellulare.
Trasmissione dei Luteoviridae
Le specie dei Luteoviridae sono trasmesse dagli afidi in un modo circolativo e non propagativo.
A tal proposito si ricorda che i numerosi agenti patogeni (virus, batteri, micoplasmi, funghi,
protozoi e nematodi) possono essere trasmessi, alle piante ed animali, dagli insetti. Con la
trasmissione meccanica il patogeno è trasmesso attraverso il semplice contatto delle appendici
boccali, delle zampe, delle feci o del rigurgito dell’insetto (veicolo). La trasmissione è immediata,
ma di scarsa durata.
Con la trasmissione biologica: esiste un adattamento anche molto specifico fra il patogeno e
l’insetto vettore. Questi, dopo una fase di acquisizione e di latenza (o incubazione) risulta capace di
trasmettere la malattia nella fase di inoculazione, anche per un lungo periodo di tempo (persistenza).
Durante l‘incubazione il patogeno entra spesso in circolo nel vettore, penetrando nelle
sue ghiandole salivari, da dove può essere inoculato (trasmissione circolativa). L’interazione
biologica è così stretta che il patogeno può addirittura replicarsi (virus) o moltiplicarsi (batteri,
fitoplasmi) all’interno del vettore (trasmissione propagativa), potendo addirittura penetrare nelle
sue gonadi ed essere trasmesso ai suoi discendenti.
Ciò richiede che il virus venga acquisito attraverso il tessuto intestinale nell’emocele degli afidi e
quindi uscire attraverso i tessuti delle ghiandole salivari.
Questo processo è specifico per ogni specie di afidi e comporta un riconoscimento specifico del
virus da parte di componenti non identificati sulle membrane del tessuto intestinale e salivare.

11
Il trasporto attraverso i tessuti è un processo di endocitosi/esocitosi. Le proteine strutturali del virus
sono coinvolte nel processo di trasmissione, con più domini che regolano il movimento e la
sopravvivenza del virus negli afidi e nel sistema virus-ospite.
Una modalità specifica di interazione tra virus e artropodi vettori è stata ampiamente descritta nei
virus delle piante nelle tre famiglie Luteoviridae, Geminiviridae e Nanoviridae, ma mai negli
arbovirus degli animali.
Si ricorda che gli arbovirus, secondo una classificazione ormai datata, sono un vasto gruppo di virus
appartenenti a famiglie diverse e che possono accrescersi negli artropodi e nei vertebrati. Sono i
responsabili di zoonosi, trasmesse occasionalmente agli uomini da artropodi vettori tramite una
puntura. Il termine deriva dall'acronimo di arthropod-borne viruses. Gli arbovirus sono virus a
RNA, appartenenti a diverse famiglie (Flaviviridae, Togaviridae, Bunyaviridae, Reoviridae,
Rhabdoviridae).
Dopo questo inciso, il precedente discorso riprende con l’affermazione che la cosiddetta
trasmissione non propagativa circolatoria contrasta con la trasmissione biologica classica degli
arbovirus degli animali, in quanto i virus corrispondenti attraversano le barriere cellulare vettoriali,
dal lume intestinale all’emolimfa e alle ghiandole salivari, senza esprimere nessuno dei loro geni e
senza replicare.
Monitorando la composizione genetica delle popolazioni virali durante il ciclo di vitale di Faba
bean necrotic stunt virus (FBNSV), genere Nanovirus, si dimostrano i cambiamenti genetici
riproducibili durante il transito del virus all'interno del corpo dell’afide vettore.
Questi cambiamenti non si adattano alla concezione che i virus attraversino semplicemente i corpi
dei loro artropodi vettori e suggeriscono interazioni più intime, mettendo in discussione l’attuale
comprensione della trasmissione circolativa, non propagativa.
La trasmissione dei virus può avvenire in modo:
1. non persistente,
2. semipersistente,
3. persistente.
Trasmissione non persistente. Sono le virosi stilettari, dette anche stylet borne, che contaminano
solo temporaneamente l’apparato boccale dell’afide e vengono eliminati con la muta o con ripetute
punture (figura 10).
Figura 10 – I virus a localizzazione epidermica si legano
alle parti distali degli stiletti con l’ingestione,
mediante una proteina-colla (helper) indotta
dal patogeno (A). Il virus è rilasciato in una
pianta sana con l’immissione di saliva (B).

12
Sono i più comuni, in quanto sono facilmente prelevati dalle cellule epidermiche della pianta,
attraverso le punture di assaggio, che l’afide esegue per identificare il substrato di alimentazione
idoneo, e ospitati temporaneamente in microscopiche anfrattuosità presenti sulla superficie dello
stiletto.
Tali virosi sono tipiche degli afidi, che sono sprovvisti di sensilli gustativi sugli apici delle
appendici boccali ed effettuano punture di assaggio sulle potenziali piante ospiti.
Il contenuto delle cellule epidermiche e parenchimatiche viene inviato in prossimità della pompetta
salivare dove è analizzato dai sensilli.
Se il succo non contiene i fagostimolanti adatti, viene travasato nelle cellule punte e l’afide riprende
la ricerca di una pianta idonea.
Nel caso in cui la pianta sia infettata da un virus a localizzazione epidermica, l’afide lo assume
immediatamente in quanto i virioni aderiscono nella porzione distale degli stiletti. In questa fase i
Potyvirus (ma non i Cucumovirus) aderiscono agli stiletti a mezzo della proteina-colla (helper) che
la pianta è indotta a produrre dall’RNA virale.
Figura 11 - Virosi non persistenti della patata. PVY, Potato virus Y, in pieno campo (A) e
sintomi su foglia (B) e su tuberi (C). Virus A della patata (D).
Le particelle virali si distaccano grazie probabilmente all’azione di una proteasi. Il virus non ha
periodo di incubazione, ma l’infettività viene presto perduta per disattivazione od in seguito alla
muta.
Nella figura 11 sono rappresentate alcune virosi non persistenti con i relativi sintomi che
interessano la patata.
Trasmissione semi-persistente. Questo tipo di trasmissione è anche detta stomodeale (figura 12),
poiché i virioni aderiscono a mezzo della proteina helper all’intima dell’intestino anteriore
(stomodeo).
L’afide, dopo aver accettato la pianta come ospite, raggiunge con gli stiletti i fasci floematici, dove
il virus è localizzato e successivamente acquisito.
Anche in questo caso non c’è periodo di latenza, ma il periodo infettivo è superiore rispetto ai virus
non persistenti.

13
Con la muta le particelle virali sono allontanate, insieme alla cuticola dello stomodeo. I
Caulimovirus, anche se hanno localizzazione stilettare, hanno caratteristiche di virus semi-
persistenti.
In questi virus un componente dell’helper (la proteina P2, una proteina estrinseca di membrana che
appartiene alla superfamiglia delle lipocaline e alla famiglia delle proteine che legano gli acidi
grassi, FABP, favorendone il trasporto attraverso la membrana cellulare) interagisce con un
recettore non glicosilato immerso nella chitina degli stiletti.
Figura 12 – Trasmissione semi-persistente stomodeale dei virus.
Trasmissione dei virus persistenti. I virus persistenti sono divisi in due categorie:
a. virus persistenti circolativi (che non si moltiplicano nel vettore) e
b. virus persistenti propagativi, che si moltiplicano nel vettore e possono essere trasmessi alla
discendenza (figura 13).
I virus si muovono all’interno del vettore dal lume intestinale all’emolinfa o ad altri tessuti (tessuto
nervoso), per giungere infine alle ghiandole salivari, da cui sono nuovamente inoculate nella pianta
durante l’alimentazione dell’insetto vettore.
I virus persistenti possono essere trasmessi alla pianta anche dopo essere stati inoculati
sperimentalmente nell’emocele dell’insetto vettore.
Nella trasmissione dei virus persistenti circolativi, il virus dopo essere stato assunto dal floema
raggiunge l’intestino e da quest’ultimo (in particolare l’intestino medio) passa nell’emocele,
raggiungendo, infine, le ghiandole salivari.
All’adesione e veicolazione attraverso i tessuti dell’ospite partecipa il capside virale. Sebbene
l’adattamento al vettore sia elevato, non vi è replicazione virale al suo interno. I tempi di latenza
vanno da qualche ora a 1-2 giorni, ma l’infettività, una volta manifestatasi, può durare molto a
lungo.
Luteoviridae, Geminiviridae e Nanoviridae sono le famiglie di virus trasmesse in maniera
circolativa. Questi virus hanno particelle icosaedriche in cui gli acidi nucleici sono contenuti in un
capside proteico senza involucro lipidico. Luteovirus e nanovirus sono trasmessi esclusivamente da
afidi, mentre i geminivirus sono trasmessi da aleirodidi e cicadellidi.

14
Il movimento e/o la replicazione dei virus persistenti e propagativi nei loro insetti ospiti
richiedono specifiche interazioni per superare quattro principali barriere:
1. barriera dell'infezione dell’intestino medio;
2. disseminazione attraverso la fuoriuscita dall'intestino medio ed infezione delle ghiandole
salivari;
3. fuoriuscita dalle ghiandole salivari;
4. superamento della barriera dell’infezione transovarica.
Figura 13 – Schema di trasmissione dei virus persistenti e propagativi, per cui i
virus replicano (si moltiplicano) anche all’interno del vettore.
Interazione Luteoviridae-vettore. Il capside di rivestimento dei Luteoviridae consiste di due
proteine di rivestimento: una proteina di 22 kDa ed una proteina (ReadThrough) di 76 kDa. La
proteina RT contiene un dominio readthrough (RTD) di 54-kDa al terminale C che protrude
all’esterno del virione.
I Luteoviridae interagiscono nelle diverse specie con differenti proteine vettori per muoversi
attraverso l’intestino e le ghiandole salivari attraverso meccanismi geneticamente distinti (luciferasi,
ciclofillina, omologo del rack-1).
I Luteoviridae entrano nelle cellule epiteliali dell’intestino del vettore per endocitosi e ne
fuoriescono invadendo l’emocele per esocitosi.
Entrambe le proteine capsidiche sono implicate nella trasmissione del virus, ma la capacità del virus
di invadere esclusivamente l’intestino medio è sotto il controllo di RTD. Entrambe queste proteine
sono glicosilate ed il nucleo glucidico gioca un ruolo centrale nell’invasione delle cellule epiteliali.
La glicosilazione riveste anche un ruolo indiretto, prevenendo la degradazione del virus, grazie
all’interazione della regione glicosilata con la chaperonina simbionina prodotta dal batterio
simbionte Buchnera aphidicola.
Evoluzione a lungo termine dei Luteoviridae: speciazione dei virus.

15
Anche se i virus a RNA sono gli agenti più comuni di malattie emergenti, gli aspetti chiave della
loro evoluzione sono ancora solo parzialmente comprensibili. Questo è di importanza accademica e
pratica, poiché l'evoluzione del virus può compromettere le strategie di controllo delle malattie,
compresa la rapida generazione di genotipi in grado di eludere risposte immunitarie o di quelli che
sono resistenti agli antivirali o alla resistenza genetica delle colture.
La maggior parte delle nostre conoscenze sulla rapidità dell'evoluzione dei virus a RNA deriva
dallo studio dei virus animali, per i quali le stime dei tassi di sostituzione dei nucleotidi
normalmente rientrano in un ordine di grandezza di 1 x 10-3
sostituzioni nucleotidi per sito all'anno
(sub/sito/anno) e in gran parte riflettono il programma del tasso di mutazione.
Gli studi equivalenti sui virus a RNA dei vegetali hanno riportato tassi più eterogenei. I primi studi
suggerivano che alcuni virus a RNA dei vegetali si sviluppassero più lentamente dei virus a RNA
che infettano gli animali. Ad esempio, le stime del tasso di sostituzione dei nucleotidi nell'intervallo
di 1 x 10-6
a 1 x 10-8
sub/sito/anno Sono stati ottenuti per Turnip yellow mosaic virus e per alcuni
tobamovirus.
Al contrario, stime più recenti attuate mediante l’uso del metodo della coalescenza bayesiana,
applicato a sequenze con dati noti di campionamento e che hanno permesso la variazione dei tassi
tra le linee, hanno riportato tassi di sostituzione nello stesso range dei virus a RNA riguardanti gli
animali, suggerendo tassi relativamente elevati di mutazione, come previsto, data la natura
intrinsecamente incline all'errore della replicazione di RNA. Così, come le differenze di questi tassi
stimati, hanno una ragionevole spiegazione biologica, per la presenza di polimorfismi mutazionali
inferiori in lungo periodo, perché qualsiasi mutazione deleteria sarebbe stata rimossa, purificando la
selezione. In particolare, molti ostacoli della popolazione alla trasmissione consentirebbero
mutazioni deleterie ad alta frequenza, a causa di una forte deriva genetica. Alcuni effetti rendono
pericoloso estrapolare i tassi di lungo termine di un cambiamento evolutivo dall'analisi dei dati di
una sequenza intraspecifica. Differenze nella forza dell'evoluzione adattativa potrebbero anche
causare eterogeneità dei tassi, inclusi alcuni processi come la competizione per individui suscettibili
e la colonizzazione di nuove specie ospiti.
Anche se c'è un crescente numero di dati sui processi intraspecifici evolutivi nei virus delle piante a
RNA, compresi i tassi di sostituzione nucleotidica, vi è stata una negligenza generale dei modelli
evolutivi a lungo termine, compresi i fattori determinanti della speciazione virale.
La speciazione è un processo evolutivo grazie al quale si formano nuove specie da quelle
preesistenti. Il fenomeno opposto è l’estinzione.
Il concetto di speciazione, essenzialmente sviluppato da Ernst Mayr, risulta dalla selezione naturale
e/o dalla deriva genetica, che sono i due motori dell'evoluzione. Questo fenomeno è una delle
colonne portanti del neodarwinismo.
Le diverse modalità di speciazione possono essere classificate in due ampie categorie:
• la divergenza adattiva, che consiste nello sviluppo graduale e in tempi lunghi dell’isolamento
riproduttivo, sotto la spinta della selezione naturale,
• la speciazione improvvisa, che avviene bruscamente e riguarda in massima parte la nascita di
individui (di solito vegetali) con corredo cromosomico poliploide (tabacco, cotone, patata).
Eccezioni sono le recenti analisi del Potyviridae e dei Sobemovirus, i quali associarono la
speciazione virale con lo sviluppo dell'agricoltura (Fargette et al., 2008; Gibbs et al., 2008).
La divergenza adattiva include 4 differenti modalità di speciazione, associate al livello in cui le
popolazioni in speciazione si sono geograficamente isolate le une dalle altre:
speciazione allopatrica, prevede l’esistenza di barriere geografiche che, separando due popolazioni
di individui della stessa specie in due territori differenti (definiti isole), interrompono il flusso
genico della popolazione iniziale, portando alla diversificazione di due specie differenti.
L’isolamento geografico può essere legato alla presenza di barriere naturali preesistenti quali
montagne, deserti, mari, oppure essere il risultato di una modificazione ambientale (innalzamento
del livello del mare, deviazione di un corso d’acqua, costruzione di barriere artificiali). Quando due
popolazioni di individui della stessa specie si trovano in condizioni di isolamento, le nuove

16
condizioni ambientali favoriscono, per selezione ulteriori, cambiamenti genetici, per cui, se
l’isolamento persiste per un periodo sufficiente, la neo-specie non sarà più in grado di incrociarsi
con la popolazione di origine. L’esempio classico, da cui prende origine questa teoria, è quello dei
“fringuelli di Darwin”, osservati da Charles Darwin sulle isole Galapagos;
speciazione parapatrica, quando la divergenza avviene all’interno di popolazioni che non sono
totalmente isolate geograficamente, ma possiedono una ristretta zona di contatto. Le migrazioni tra
popolazioni sono, tuttavia, limitate poiché queste ultime si perpetuano all’interno di condizioni
ambientali differenti (per esempio gradienti climatici). La selezione naturale ha dunque un ruolo
importante in questa modalità di speciazione.
Se le due specie hanno acquisito completo isolamento riproduttivo possono sovrapporsi, in base alle
preferenza di habitat.
Se si sono sviluppate barriere di isolamento riproduttivo, ma non compatibilità ecologica, gli areali
si mantengono parapatrici.
Se non sono state acquisite barriere anti-ibridazione, si forma una zona di contatto che porta alla
formazione di ibridi, nei quali possono evolvere barriere post-copula;
speciazione peripatrica o speciazione per "effetto del fondatore", avviene quando un piccolo
numero di individui costituisce una nuova popolazione ai margini dell’areale della specie di origine,
ad esempio, colonizzando una piccola isola vicina alla costa. La nuova popolazione può
rapidamente evolvere in una nuova specie. A questo modello sono riferibili i casi di semi-specie,
circoli di specie, super specie;
speciazione simpatrica avviene quando due popolazioni non isolate geograficamente si evolvono
in specie distinte, grazie alla presenza di polimorfismo nel tempo. In questo caso la selezione
naturale gioca un ruolo cruciale nella divergenza delle popolazioni. Il fenomeno presenta, tuttora,
degli aspetti controversi, ma è stato ben documentato in alcuni casi, per esempio nei pesci di acqua
dolce della famiglia dei Ciclidi.
I principali studi in questo campo riguardano gli esperimenti sulla Drosophila. La speciazione
simpatrica può avvenire per formazione di una criptospecie, ossia di una popolazione non più
interfeconda con la specie di origine, a causa di alterazioni del cariotipo. Questo meccanismo può
rientrare tra le modalità di speciazione improvvisa.
La speciazione improvvisa o speciazione istantanea, è un processo che riguarda prevalentemente
gli organismi vegetali e si verifica in tempi brevi. Tipica causa è il fenomeno della poliploidia, che
consiste, come è noto, in un aumento del numero di cromosomi rispetto al normale assetto diploide.
La poliploidia può avvenire per:
• una non-disgiunzione dei cromosomi durante il processo mitotico o meiotico;
• un processo meiotico o mitotico avvenuto senza errori, ma senza la successiva citodieresi.
Pertanto, prendendo in esame un qualunque individuo diploide, si ha una situazione del genere:
o l'individuo effettua la meiosi;
o durante la meiosi, avviene una non-disgiunzione dei cromosomi: i gameti risulteranno quindi
diploidi;
o i gameti si incrociano tra loro: lo zigote risultante sarà tetraploide, ossia avrà un assetto
cromosomico doppio rispetto al suo ascendente originario, cioè rispetto all’organismo iniziale
preso in considerazione.
Tuttavia, la formazione di criptospecie avviene per lo più a livello di ibridi, cioè di individui frutto
dell’incrocio di due specie differenti e quindi sterili. Nel caso degli organismi vegetali, gli ibridi
sterili sopperiscono attraverso la riproduzione. Nel 2011 è stato documentato il caso di speciazione
simpatrica in organismi termofili
Nel contesto di virus a RNA, la speciazione allopatrica può essere considerata come la
diversificazione genetica che si verifica quando i virus salgono verso nuove specie ospitanti e,
successivamente, si evolvono in modo indipendente, come comunemente associato al processo di
emergenza virale. Al contrario, la speciazione simpatrica avrebbe luogo quando i virus si
diversificheranno all'interno di una singola specie ospitante, forse sfruttando diversi tipi di cellule

17
(Holmes, 2009). Nonostante l'importanza di questi processi per la comprensione della
macroevoluzione dei virus a RNA, i loro rispettivi ruoli sono attualmente sconosciuti.
Per comprendere meglio la natura dei processi evolutivi a lungo termine nei virus a RNA dei
vegetali, è stata intrapresa un’ampia analisi molecolare evolutiva della famiglia Luteoviridae, una
famiglia eterogenea di virus vegetali suddivisi in tre generi, Luteovirus, Polerovirus ed Enamovirus,
come detto all’inizio, contenenti rispettivamente cinque, nove ed una specie classificate, nonché una
serie di specie non classificate (Fauquet et al., 2005). I luteoviridi possiedono un genoma RNA a
singolo filamento di 5.600-6.000 nucleotidi (nt). Questi genomi possono ospitare cinque o sei ORF.
Il 5'–prossimale parzialmente sovrapponibile a ORF1 e ORF2 codifica le proteine P1 e P2, che sono
coinvolte nella replicazione del virus.
La bassa frequenza di –1 ribosomiale nella regione sovrapposta provoca la fusione di P1-P2 RNA
dipendente dalla proteina RNA polimerasi (RdRp). ORF3 codifica la proteina del capside (CP) e
contiene completamente ORF4, che non è presente negli Enamovirus ed è necessaria per il
movimento del virus nella pianta (proteina di movimento, MP). L’ORF5, che è necessaria per la
trasmissione da parte degli afidi (Brault et al., 2005; Gray e Gildow, 2003; Miller et al., 2002) ed è
anche coinvolta nel movimento del virus e nella limitazione del floema dei Luteoviridae (Peter et
al., 2009), è tradotto attraverso il codone terminatore di ORF3, esistente come un dominio di lettura
(RTD) fuso a CP. I membri del genere Polerovirus hanno una extra ORF0 supplementare nel
terminale 5' del genoma che si sovrappone parzialmente a ORF1. Il suo prodotto di traduzione (P0)
funge da repressore della risposta alla difesa dell’impianto di silenziamento RNA (Mangwende et
al., 2009; Pfeffer et al., 2002). Infine, alcune specie di Luteovirus hanno un ulteriore ORF6 con una
funzione sconosciuta nel terminatore 3' del genoma (Fauquet et al., 2005; Miller e Rasochova,
1997; Smith e Barker, 1999). Come conseguenza di questa particolare organizzazione genomica,
circa un terzo del genoma dei Polerovirus ed una frazione minore nel genoma dei Luteovirus sono
composte da regioni sovrapposte.
A causa della loro importanza agronomica, i dati della sequenza genica, insieme ad informazioni
sulla gamma degli ospiti e sulla distribuzione geografica, sono disponibili per un numero
relativamente elevato di membri della famiglia Luteoviridae. Tuttavia, fino ad oggi l’unico
Luteovirus per il quale sono stati stimati i tassi di cambiamento evolutivo è il Barley yellow dwarf
virus (BYDV). In questo caso, un’analisi dei tassi di sostituzione basati su RNA virale estratto da
esemplari di erbario ha prodotto stime tra i 6,2 x 10-4
e 9,7 x 10-5
sub/sito/anno (Malmstrom et al.,
2007). Allo stesso modo, solo una stima del punto in cui la diversità genetica è nata nella famiglia
Luteoviridae è stata rilevata, cioè circa 9.000 anni fa, forse associata al’'aumento di sviluppo
dell’agricoltura (Fargette et al., 2008). Tuttavia, solo un numero limitato di specie e sequenze di
Luteoviridae sono stato incluso in questa analisi.
Nessun studio ha ancora considerato i meccanismi della speciazione nella famiglia Luteoviridae.
La famiglia Luteoviridae rappresenta anche un set di dati utile per studiare altri due fenomeni
evolutivi: il pattern e le determinanti della ricombinazione, che sembra essere comune all’interno
della famiglia Luteoviridae (Gibbs, 1995; Miller et al., 2002; Moonan et al., 2000; Smith e Barker,
1999; Stevens et al., 2005) e le diverse dinamiche evolutive geniche con ORF sovrapposte. Esistono
ipotesi contrastanti sul fatto che le ORF sono utilizzate in modo così comune nei virus a RNA.
Secondo una certa visione, la sovrapposizione genica massimizza l’informazione genetica in
genomi più piccoli (Barrell et al., 1976; Jordan et al., 2000). In alternativa, è stato suggerito che la
sovrapposizione genica genera robustezza mutazionale (cioè la capacità di preservare i fenotipi,
malgrado il carico genomico mutazionale) a livello di popolazione (Belshaw et al., 2007; Elena e
Sanjua, 2005; Krakauer e Plotkin. 2002). Secondo questa ipotesi, la sovrapposizione genica genera
un’ipersensibilità a mutazioni deleterie, in quanto queste influenzano più di un gene.
Anche se questa ipersensibilità riduce la capacità di ciascun individuo a tamponare gli effetti della
mutazione, essa rappresenta un vantaggio selettivo per i genotipi di tipo selvatico, che poi aumenta
la robustezza a livello di popolazione (Elena e Sanjua, 2005; Krakauer e Plotkin. 2002). Come
conseguenza di questo elevato carico di mutazione deleteria, i virus a RNA con proporzioni più

18
grandi dei loro genomi presenti come ORF sovrapponibili, dovrebbero presentare bassi tassi di
sostituzione dei nucleotidi (Krakauer, 2000; Miyata e Yasunaga, 1978). Tale riduzione di tasso è
stata osservata in molti virus a DNA e RNA (Hughes et al., 2001; Hughes e Hughes, 2005; Pavesi,
2006; Zaaijer et al., 2007; Zhao et al., 2007), anche se solo alcuni studi hanno considerato, in
questo contesto, i virus a RNA patogeni dei vegetali (Guyader e Ducray, 2002; Ranjith-Kumar,
1998).
Bibliografia
1. Balboni B, 2003. Evoluzione ed evoluzionismo Alpha Test, pp. 128.
2. Barrell BG, Air GM, Hutchison CA III. 1976. Overlapping genes in bacteriophage ФX174.
Nature 264:34–41.
3. Belshaw R, Pybus OG, Rambaut A, 2007. The evolution of genome compression and
genomic novelty in RNA viruses. Genome Res. 17:1496–1504.
4. Beuve MM, Stevens HY, Liu WM, Witermantel SH, Lemaire O, 2008. Biological and
molecular characterization of an American sugar beet-infecting Beet western yellows virus
isolate. Plant Dis. 92:51–60.
5. Blok J, Mackenzie A, Guy P, Gibbs A, 1987. Nucleotide sequence comparisons of Turnip
yellow mosaic virus isolates from Australia and Europe. Arch. Virol. 97:283–295.
6. Botstein D, 1980. A theory of modular evolution for bacteriophages. Ann. N. Y. Acad. Sci.
354:484–491.
7. Brault V, Perigon S, Reinbold C, Erdinger M, Scheidecker D, Herrbach E, Richards K,
Ziegler-Graff V, 2005. The polerovirus minor capsid protein determines vector specificity and
intestinal tropism in the aphid. J. Virol. 79:9685–9693.
8. Chay CA, Gunasinge UB, Dinesh-Kumar SP, Miller WA, Gray SM, 1996. Aphid
transmission and systemic plant infection determinants of Barley yellow dwarf Luteovirus-
PAV are contained in the coat protein readthrough domain and 17-kDa protein, respectively.
Virology 219:57–65.
9. Cornish PV, Hennig M, Giedroc DP, 2005. A loop 2 cytidine-stem 1 minor groove
interaction as a positive determinant for pseudoknot-stimulated -1 ribosomal frameshifting.
Proc. Natl. Acad. Sci. U. S. A. 102: 12694–12699.
10. Dolja VV, Carrington JC, 1992. Evolution of positive-strand RNA viruses. Semin. Virol.
3:315–326.
11. Drake JW, Charlesworth B, Charlesworth D, Crow JF, 1998. Rates of spontaneous mutation.
Genetics 148:1667–1686.
12. Drummond AJ, Rambaut A, 2007. BEAST: Bayesian evolutionary analysis by sampling trees.
BMC Evol. Biol. 7:214.
13. Drummond AJ, Ho SYW, Phillips MJ, Rambaut A, 2006. Relaxed phylogenetics and dating
with confidence. PLoS Biol. 4:e88.
14. Duffy S, Shackelton LA, Holmes EC, 2008. Rates of evolutionary change in viruses: patterns
and determinants. Nat. Rev. Genet. 9:267–276.
15. Edgar RC, 2004. MUSCLE: multiple sequence alignment with high accuracy and high
throughput. Nucleic Acids Res. 32:1792–1797.
16. Elena SF, Agudelo-Romero P, Carrasco P, Codoner FM, Martin S, Torres-Barcelo C, Sanjuán
R, 2008. Experimental evolution of plant RNA viruses. Heredity 100:478–483.
17. Elena SF, Sanjuán R, 2005. Adaptive value of high mutation rates of RNA viruses:
separating causes from consequences. J. Virol. 79:11555–11558.
18. Fargette D, Pinel-Galzi A, Sereme D, Lacombe S, Hebrard E, Traore O, Konate G, 2008.
Diversification of Rice yellow mottle virus and related viruses spans the history of agriculture
from the Neolithic to the present. PLoS Pathog. 4:e1000125.

19
19. Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball BA, 2005. Virus taxonomy:
classification and nomenclature of viruses. 8th report of the International Committee on the
Taxonomy of Viruses. Elsevier-Academic Press, San Diego, CA.
20. Fitch WN, 1971. Toward defining the course of evolution: minimal change for a specific tree
topology. Syst. Zool. 20:406–416.
21. García-Arenal F, McDonald BA, 2003. An analysis of the durability of resistance to plant
viruses. Phytopathology 93:941–952.
22. Gibbs AJ, 1980. How ancient are the tobamoviruses? Intervirology 14:101–108.
23. Gibbs AJ, Fargette D, Garcia-Arenal F, Gibbs MJ, 2010. Time—the emerging dimension of
plant virus studies. J. Gen. Virol. 91:13–22.
24. Gibbs AJ, Blok J, Coates DJ, Guy PL, Mackenzie A, Pigram N, 1986. Turnip yellow mosaic
virus in an endemic Australian alpine Cardamine, p. 289–300. In B. A. Barlow (ed.), Flora
and fauna of alpine Australasia; ages and origins. CSIRO, Collingwood, Victoria, Australia.
25. Gibbs AJ, Ohshima K, Phillips MJ, Gibbs MJ, 2008. The prehistory of potyviruses: their
initial radiation was during the dawn of agriculture. PLoS One 3:e2523.
26. Gibbs AJ, Gibbs MJ, Ohshima K, Garcia-Arenal F, 2008. More plant virus evolution; past
present and future, p. 229–250. In E. Domingo, C. R. Parrish and J. J. Holland (ed.), Origin
and evolution of viruses, 2nd ed. Academic Press, London, United Kingdom.
27. Gibbs AJ, 1995. The luteovirus supergroup: rampant recombination and persistent
partnerships, p. 351–368. In A. Gibbs, C. H. Calisher, and F. García-Arenal (ed.), Molecular
basis of virus evolution. Cambridge University Press, Cambridge, United Kingdom.
28. Gray S, Gildow FE, 2003. Luteovirus-aphid interactions. Annu. Rev. Phytopathol. 41:539–
566.
29. Guyader S, Ducray DG, 2002. Sequence analysis of Potato leafroll virus isolates reveals
genetic stability, major evolutionary events and differential selection pressure between
overlapping reading frame products. J. Gen. Virol. 83:1799–1807.
30. Hanada K, Suzuki Y, Gojobori T, 2004. A large variation in the rates of synonymous
substitution for RNA viruses and its relationship to a diversity of viral infection and
transmission modes. Mol. Biol. Evol. 21:1074–1080.
31. Hasegawa M, Kishino H, Yano T, 1985. Dating the human-ape splitting by a molecular clock
of mitochondrial DNA. J. Mol. Evol. 22:160–174.
32. Holmes EC, 2003. Patterns of intra- and inter-host nonsynonymous variation reveal strong
purifying selection in dengue virus. J. Virol. 77:11296–11298.
33. Holmes EC, 2003. Molecular clocks and the puzzle of RNA virus origins. J. Virol. 77:3893–
3897.
34. Holmes EC, 2003. Error thresholds and the constraints to RNA virus evolution. Trends
Microbiol. 11:543–546.
35. Holmes EC, 2009. The evolution and emergence of RNA viruses. Oxford University Press,
Oxford, United Kingdom.
36. Hughes AL, Westover K, Da Silva J, O’Connor DH, Watkins DI, 2001. Simultaneous
positive and purifying selection on overlapping reading frames of the tat and vpr genes of
simian immunodeficiency virus. J. Virol. 75:7966–7972.
37. Hughes AL, Hughes MA, 2005. Patterns of nucleotide difference in overlapping and non-
overlapping reading frames of papillomavirus genomes. Virus Res. 113:81–88.
38. Jenkins GM, Rambaut A, Pybus OG, Holmes EC, 2002. Rates of molecular evolution in RNA
viruses: a quantitative phylogenetic analysis. J. Mol. Evol. 54:156–165.
39. Jin Z, Wang X, Chang S, Zhou G, 2004. The complete nucleotide sequence and its
organization of the genome of Barley yellow dwarf virus-GAV. Sci. China C Life Sci.
47:175–182.
40. Jordan IK, Sutter BA IV, McClure MA, 2000. Molecular evolution of the Paramyxoviridae
and Rhabdoviridae multiple-protein-encoding P gene. Mol. Biol. Evol. 17:75–86.

20
41. Kosakovsky Pond SL, Frost SDW, 2005. Datamonkey: rapid detection of selective pressure
on individual sites of codon alignments. Bioinformatics 21:2531–2533.
42. Krakauer DC, 2000. Stability and evolution of overlapping genes. Evolution 54:731–739.
43. Krakauer DC, Plotkin JB, 2002. Redundancy, antiredundancy, and the robustness of genomes.
Proc. Natl. Acad. Sci. U. S. A. 99:1405–1409.
44. Malmstrom C, Shu R, Linton EW, Newton LA, Cook M, 2007. Barley yellow dwarf viruses
(BYDVs) preserved in herbarium specimens illuminate historical disease ecology of invasive
and native grasses. J. Ecol. 95:1153–1166.
45. Mangwende T, Wang ML, Borth W, Hu J, Moore PH, Mirkov ET, Albert HH, 2009. The P0
gene of Sugarcane yellow leaf virus encodes an RNA silencing suppressor with unique
activities. Virology 384:38–50.
46. Martin DP, Williamson C, Posada D, 2005. RDP2: recombination detection and analysis from
sequence alignments. Bioinformatics 21:260–262.
47. Martin RR, Keese PK, Young MJ, Waterhouse PM, Gerlach WL, 1990. Evolution and
molecular biology of luteoviruses. Annu. Rev. Phytopathol. 28:341–363.
48. Mayr E, 1970. L'evoluzione delle specie animali. G. Einaudi ed., voll.I-II, pp. 833.
49. McDonald BA, Linde C, 2002. The population genetics of plant pathogens and breeding
strategies for durable resistance. Euphytica 124:163–180.
50. Miller WA, Rasochova L, 1997. Barley yellow dwarf viruses. Ann. Rev. Phytopathol.
35:167–190.
51. Miller WA, Liu S, Beckett R, 2002. Barley yellow dwarf virus: Luteoviridae or
Tombusviridae? Mol. Plant Pathol. 3:177–183.
52. Miyata T, Yasunaga T, 1978. Evolution of overlapping genes. Nature 272:532–535.
53. Moonan F, Molina J, Mirkov TE, 2000. Sugarcane yellow leaf virus: an emerging virus that
has evolved by recombination between luteoviral and poleroviral ancestors. Virology
269:156–171.
54. Moonan F, Mirkov TE, 2002. Analyses of genotypic diversity among North, South, and
Central American isolates of Sugarcane yellow leaf virus: evidence for Colombian origins and
for intraspecific spatial phylogenetic variation. J. Virol. 76:1339–1348.
55. Moya A, Holmes EC, González-Candelas F, 2004. The population genetics and evolutionary
epidemiology of RNA viruses. Nat. Rev. Microbiol. 2:279–288.
56. Nosil P, Sandoval CP, 2008. Ecological Niche Dimensionality and the Evolutionary
Diversification of Stick Insects, in PLoS ONE. 2008; 3(4): e1907.
57. Parker J, Rambaut A, Pybus OG, 2008. Correlating viral phenotypes with phylogeny:
accounting for phylogenetic uncertainty. Infect. Genet. Evol. 8:239–246.
58. Pavesi A, 2006. Origin and evolution of overlapping genes in the family Microviridae. J. Gen.
Virol. 87:1013–1017.
59. Pazhouhandeh M, Dieterle M, Marrocco K, Lechner E, Berry B, Brault V, Hemmer O,
Kretsch T, Richards KE, Genschik P, Zieger-Graff V, 2006. F-box-like domain in the
Polerovirus protein P0 is required for silencing suppressor function. Proc. Natl. Acad. Sci. U.
S. A. 103:1994–1999.
60. Peter KA, Liang D, Palukaitis P, Gray SM, 2008. Small deletions in the Potato leafroll virus
readthrough protein affect particle morphology, aphid transmission, virus movement and
accumulation. J. Gen. Virol. 89(Pt. 8):2037–2045.
61. Peter KA, Gildow F, Palukaitis P, Gray SM, 2009. The C terminus of the polerovirus P5
readthrough domain limits virus infection to the phloem. J. Virol. 83:5419–5429.
62. Pfeffer S, Dunoyer P, Heim F, Richards KE, Jonard G, Ziegler-Graff V. 2002. P0 of Beet
western yellow virus is a suppressor of posttranscriptional gene silencing. J. Virol. 76:6815–
6824.
63. Posada D, Crandall KA, 1998. Modeltest: testing the model of DNA substitution.
Bioinformatics 14:817–818.

21
64. Ranjith-Kumar CT, Gopinath K, Jacob ANK, Svrividhya V, Elango P, Savithri SH, 1998.
Genomic sequence of Physalis mottle virus and its evolutionary relationship with other
tymoviruses. Arch. Virol. 143:1489–1500.
65. Robertson NL, French R, 2007. Genetic structure in natural populations of barley/cereal
yellow dwarf virus isolates from Alaska. Arch. Virol. 152:891–902.
66. Roossinck M, Ali A, 2007. Mechanisms of plant virus evolution and identification of genetic
bottlenecks: impact on disease management, p. 109–124. In Z. K. Punja, S. H. De Boer, and
S. Sanfac¸on (ed.), Biotechnology and plant disease management. CABI Publishing,
Wallingford, United Kingdom.
67. Sadowy E, Juszczuk M, David C, Gronenborn B, Hulanicka MD, 2001. Mutational analysis
of the proteinase function of Potato leafroll virus. J. Gen. Virol. 82:1517–1527.
68. Sanjuán R, Agudelo-Romero P, Elena SF, 2009. Upper-limit mutation rate estimation for
plant RNA virus. Biol. Lett. 5:394–396.
69. Schierup MH, Hein J, 2000. Recombination and the molecular clock. Mol. Biol. Evol.
17:1578–1579.
70. Schluter D, 2001. Ecology and the origin of species. Trends Ecol. Evol. 16:372–380.
71. Schluter A, Real-Chicharro A, Gabaldon T, Sanchez-Jimenez F, Pujol A, 2010.
PeroxisomeDB 2.0: an integrative view of the global peroxisomal metabolome. Nucleic Acids
Res, 38, D800-5.
72. Simmonds P, Smith DB, 1999. Structural constraints on RNA virus evolution. J. Virol.
73:5787–5794.
73. Simmons HE, Holmes EC, Stephenson AG, 2008. Rapid evolutionary dynamics of Zucchini
yellow mosaic virus. J. Gen. Virol. 89:1081–1085.
74. Smith HG, Barker H, 1999. The Luteoviridae. CABI Publishing, Wallingford, United
Kingdom.
75. Stevens M, Freeman B, Liu H-Y, Herrbach E, Lemaire O, 2005. Beet polerovirus: close
friends or distant relatives? Mol. Plant Pathol. 6:1–9.
76. Terauchi HK, Honda N, Yamagishi S, Kanematsu K, Ishiguro Hidaka S, 2003. The N-
terminal region of the readthrough domain is closely related to aphid vector specificity of
Soybean dwarf virus. Phytopathology 93:1560–1564.
77. Van der Wilk F, Verbeek M, Dullemans AM, van den Heuvel JF, 1997. The genome-linked
protein of Potato leafroll virus is located downstream of the putative protease domain of the
ORF1 product. Virology 234:300–303.
78. Viswanathan R, Balamuralikrishnan M, Karuppaiah R, 2008. Identification of three genotypes
of Sugarcane yellow leaf virus causing yellow leaf disease from India and their molecular
characterization. Virus Genes 37:368–379.
79. Wang TH, Donaldson YK, Brettle RP, Bell JE, Simmonds P, 2001. Identification of shared
populations of human immunodeficiency virus type 1 infecting microglia and tissue
macrophages outside the central nervous system. J. Virol. 75:11686–11699.
80. Whelan S, Goldman N, 2001. A general empirical model of protein evolution derived from
multiple protein families using a maximum-likelihood approach. Mol. Biol. Evol. 18:691–
699.
81. Zaaijer HL, van Hemert FJ, Koppelman MH, Lukashov VV, 2007. Independent evolution of
overlapping polymerase and surface protein genes of hepatitis B virus. J. Gen. Virol.
88:2137–2143.
82. Zhao X, McGirr KM, Buehring GC, 2007. Potential evolutionary influences on overlapping
reading frames in the bovine leukemia virus pXBL region. Genomics 89:502–511.
Recommended