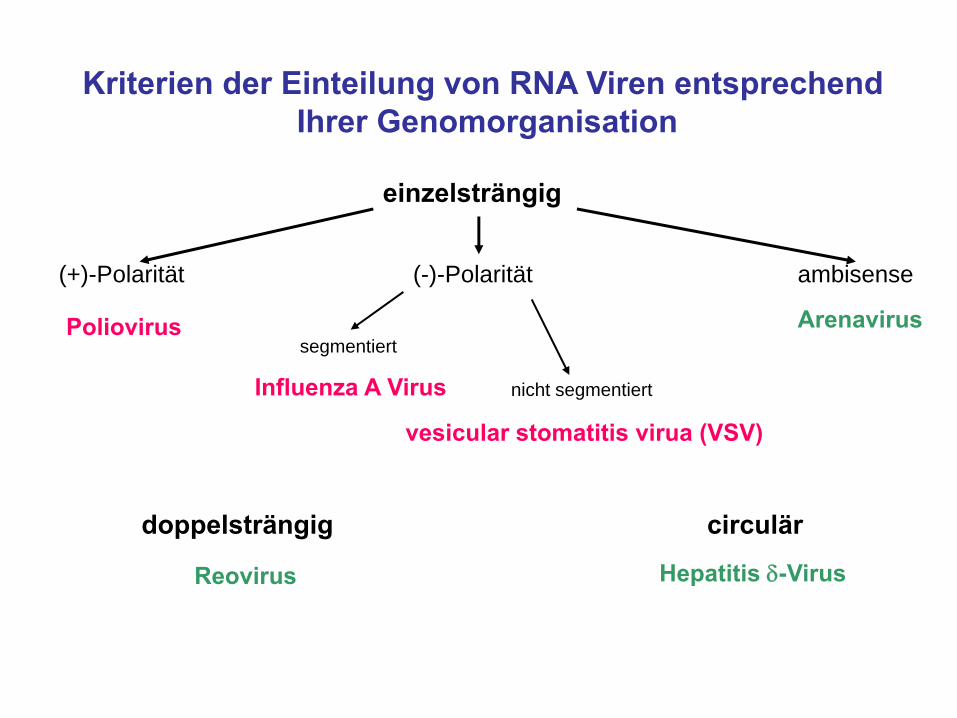
Kriterien der Einteilung von RNA Viren entsprechendIhrer Genomorganisation
einzelsträngig
doppelsträngig circulär
(+)-Polarität (-)-Polarität ambisense
Poliovirus
Influenza A Virus
Hepatitis δ-Virus
segmentiert
vesicular stomatitis virua (VSV)
Reovirus
nicht segmentiert
Arenavirus

RNA Genom(+),(-), ds Kopie des RNA Genoms
(+),(-), dsSynthese von mRNA
(+)
Translation in virale Proteine(Strukturproteine, regulatorische Proteine)
Verpackung in neue Virionen(Verpackungssequenzen, Strukturproteine)
RNA RNARNA-abhängige RNA Synthese
Virale RNA-abhängige RNA Polymerasen (RdRPs) !!
Replikation„Transkription“mRNA-Synthese
Ausnahme: Retroviren (RdDP)
Prinzipien der Replikation und mRNA-Synthesenicht-retroviraler RNA-Viren

RNA-abhängige RNA-Polymerasen
Die Aktivität konnte dem cytoplasmatischen viralen 3Dpol-Protein zugeschrieben werden
Zellextrakte Poliovirus-infizierter Zellen enthalten eine enzymatische Aktivität die die„primer“ und „template“ abhängige Inkorporation von Ribonukleotiden katalysiert (1962)
Die Aktivität ist insensitiv gegenüber Actinomycin D (im Gegensatz zu Pol II Aktivität)
Vergleichbare Aktivitäten sind später in Partikeln von (-) Strang und dsRNA Viren entdeckt worden
Die meisten RNA-abhängigen RNA Polymerasen (RdRPs) sind primerunabhängig(vergleichbar den zellulären DNA-abhängigen RNA Polymerasen) Ausnahme 3DPol
Aufgrund der präferentiellen Membranlokalisation ist die Reinigung der meisten RdRPs schwierig
Primärsequenzvergleiche verschiedener zellulärer und viraler Polymerasen weisen aufeinen gemeinsamen „Vorfahren“ hin
Initiation und Termination der RNA Synthese erfolgt an spezifischen RNA Stellen, RNA Synthese erfolgt von 5´- 3´.

Primärstrukturvergleich und Sequenzhomologie viralerund zellulärer Polymerasen
Poliovirus 3Dpol
HIV-I RT
Pol I KlenowRNA Pol T7
MetallbindungTeil des aktiven Zentrums
© Flint et al. Principles of Virology
dNTP – NTP Unterscheidung
Motive A-D in allen Polymerasen

Tertiärstruktur der Poliovirus 3Dpol-Polymerase und generelle Strukturhomologien von Polymerasen
YGDD
Klenow T7 RNAP HIV-I RT 3Dpol
Nur in RdXPs
© Flint et al. Principles of Virology

Die Beschaffenheit der RNA Matrizen von (-) und (+) Strang RNA Viren
RNA-Polymerasen kopieren die viralen (-)-Stränge nur im Proteinkomplex (nackte RNA wird nicht kopiert Bsp. VSV – N-Protein)
(-) Strang RNA Viren verpacken ihr Genom in Form von Nukleoprotein-haltigen Nukleokapsiden (diese enthalten die RdRP sowie aksessorische Proteine)
In dieser Form sind sie stabilisiert und resistent gegen RNAse
Die Genome von (+)-Strang RNA Viren sind nicht in gleicher Weise verpackt (Ausnahme: Retroviren)
Der Grund für diesen Unterschied liegt möglicherweise in der initialen Aufgabe der Matrize:Bei (+) Strang RNA Viren erfolgt zunächst TranslationBei (-) Strang RNA Viren mRNA-Synthese
Nukleoproteine sind kooperative Einzelstrang-RNA Bindeproteine die für die RNA-Synthese benötigt werden (analog den ssDNA Bindeproteinen bei der DNA-abhängigen DNA/RNA-Synthese).
Ihre Funktion besteht in der Aufrechterhaltung der Einzelsträngigkeit zum Verhindern der Basenpaarung zwischen Template und Produkt (ermöglicht mehrere Syntheserunden)
Bei vielen (+) Strang RNA Viren sind Helikasen an der Replikation beteiligt (Im Falle von Poliovirus hat 3DPol selbst (ATP-unabhängige) Helikaseaktivität).

RNA Sekundärstrukturen „stem-loops“ und „Pseudoknoten“
„stem-loop“ Strukturen Pseudoknoten
Basenpaarungen im Stamm (stem),Keine Basenpaarungen innerhalbder Schleife (loop)
Basenpaarungen im Stamm,Basenpaarungen der Schleife mitNukleotiden außerhalb der Schleife
© Flint et al. Principles of Virology
Vorhersage von RNA-Sekundärstrukturen mit ComputerprogrammenVerifikation der Strukturen durch spezifischen RNAase Verdaus und 2D-NMRVergleichbare RNA-Sekundärstrukturen müssen nicht notwendigerweise homolog sein

Einige repräsentative Viren mit RNA-Genomen
© Flint et al. Principles of Virology

Funktionen akzessorischer Proteine bei derRNA-abhängigen RNA Synthese
1. Direktion der RNA-Polymerase zum korrekten zellulären Kompartiment
Die Matrizenspezifität viraler RdRP wird durch spezifische Interaktionen derPolymerase mit akzessorischen viralen oder zellullären Proteinen, die anspezifische Stellen in der Matrize binden (RNA-Sekundärstrukturen), vermittelt.Beispiel: Poliovirus 3AB-3CD bindet an eine in Picornaviren konservierte PseudoknotenRNA Sekundärstruktur am 3´-Ende und initiiert so die Polymerase zur (-) Strangsynthese.
2. Direktion der RNA-Polymerase zur korrekten Stelle der viralen Matrize
RNA-Synthese findet typischerweise nicht frei im Cytoplasma, sondern in bzw. an definierten zellulären Strukturen statt:(-) Strang Segmente von Influenza A Virus werden vermittels eines NLS im NP Proteinsin den Kern dirigiert.(+) Strang des Poliovirusgenoms wird vermittels der Interaktion von 2C mit RNA und 3AB mit 3CD (enthält die 3DPol) an ein vesikuläres Kompartment dirigiert.
3. Stimulation der Polymeraseaktivität
4. Erleichterung der RNA-Synthese durch viruscodierte HelicasenBasengepaarte Bereiche innerhalb der RNA werden durch Helicasen aufgeschmolzen.Dies stellt einen möglichen Angriffspunkt für therapeutische Intervention dar (HCV-Helicase)

Zelluläre Orte viraler RNA Synthese
Die meisten RNA-Viren replizieren ihr Genom und synthetisieren ihre mRNAs im Cytoplasma
Cytoplasma:
Synthese erfolgt nicht frei sondern ist an zelluläre Strukturen gebunden (Vesikel, Membranen, Cytoskelett)
Die hohe lokale Konzentration der für die Replikation notwendigen Komponenten erhöhtdie Effizienz der Replikation
Membranen stellen häufig den Ort der Verpackung neuer Virionen dar.
Problem: Alle normalerweise nukleären Aktivitäten des Wirtes müssen von viralen Proteinenbereitgestellt werden (z.B. capping von mRNAs).
Nukleus:Von den RNA Viren replizieren nur Influenza und Borna Viren im Nukleus
Vorteil besteht in der Möglichkeit der Ausnutzung des zellulären „splicing“-Apparates
Es müssen Mechanismen des Kerntransportes entwickelt werden. NP-Protein enthält Kernlokalisationssequenz.

Replikation und RNA-Synthese von (+) Strang RNA Viren
RNA von (+) Strang RNA Viren ist infektiös- (+) Strang genomische RNA kann als direkte Matrize für die Synthese von RdRP dienen
(+) Strang RNA Viren bedürfen keiner aktiven RdRP im Viruspartikel
Genomreplikation erfolgt über ein komplettes (-) Strang Intermediat
Polivirus hat ein 5´VpG statt CAP

Struktur und Genomorganisation des (+) StrangPicornavirus: Poliovirus
© Flint et al. Principles of Virology

Replikationszyklus des (+) StrangPicornavirus: Poliovirus
Bindung an PVR(Kapsidöffnung, RNA-Freisetzung)
Polyproteinsynthese(VPg-Entfernung, Ribosomenbindungan IRES Element, Initiation an nt 741)
Polyproteinprozessierung(P1:Struktur; P2, P3: Proteasen,
akzessorische Proteine und RdRP)
Proteintransport in Vesikel(vesikuläre RNA-Synthese, teilweiser
Rücktrasport von (+) RNA ans Ribosom)
VPg (-) Strangsynthese
VPg (+) Strangsynthese
Zellkern
Membranvesikel
© Flint et al. Principles of Virology

Struktur des 5´-Endes des Poliovirus (+) Strang RNA Genoms
22 AminosäurenVPg Peptid
Phosphosäureester mit Tyr 3 von VPg
5´-Uridylrest
Hydrolyse durch zelluläre Esterase(Generierung eines 5´-Up-Endes)
Kleeblattstruktur des 5´-Endes desPoliovirus RNA-Genoms
© Flint et al. Principles of Virology
Kovalente VPg-Bindung am 5´Ende des (+) und (-) Strangs der PV-RNA führten zur Hypothese, dass VPg als primer fungiert.Die Präsenz von VPg-pUpU in Zellextrakten infizierter Zellen bestätigte dies. VPg-pUpU wirkt als Proteinprimer für die Polio RdRP.
(Viele RdRPs können ähnlich den DdRPs primerunabhängig initiieren!!

Bei der Poliovius-RNA Synthese wird der WirtsfaktorPoly (rC) Bindeprotein 2 benötigt. Ein weiterer Faktor ist das
Poly (A) Bindprotein
Abhängigkeit der Poliovirus RNA Synthese von Wirtsfaktoren
I. Inkubation von Poliovirus (+) Strang RNA mit zytoplasmatischen Extrakten aus nichtInfizierten aber permissiven Zellen:⇒ Translation der genomische RNA (u.a. Synthese der viralen Polymerase 3DPol.⇒ Weiterhin: Synthese des (-) Stranges (in Abwesenheit von Guanidinhydrochlorid)In Anwsesenheit von letzterem findet nur assembly mit Pol statt, aber keine Initiation. Die kann erst durch erneute Zugabe von cytoplasmatischem Extrakt erreicht werden.
II. Verwendet man nicht permissive Zellen (z.B. Xenopus laevis Oocyten) und injiziertPoliovirus RNA, kann diese nicht repliziert werden. Nach zusätzlicher Injektion vonCytoplasmatischen Extrakten human Zellen erfolgt RNA Synthese.
⇒ Die Poliovirus RNA Synthese benötigt einen zellulären Faktor für die RNA Synthese

Die Spezifität der Poliovirus RNA Synthese wird durch spezifische Interaktionenviraler und zellulärer Proteine an RNA-Sekundärstrukturelemente gewährleistet
Bindung von Poly (rC) Bindeprotein 2 an stem loop B ermöglicht die Bindung von 3CD an
stem loop D.
Bindung von UTP und Anlagerung des Komplexesan Membrangebundenes 3AB Protein
(Dies ist in Wirklichkeit sehr viel komplizierter)
„Priming“ und proteolytische Abspaltung von VPg aus 3AB durch die Protease 3C;Bindung von
3Dpol/VPg-primer an 3´-Ende; Erkennung des3´“pseudoknot“ durch 3Dpol bedingt Spezifität
Elongation und (-) Strang Synthese
© Flint et al. Principles of Virology

„Priming“ Reaktion am 3´-Ende der Poliovirus RNA
Bindung von 3AB an Membranen(membranous web)
Uridylylierung von Tyr 3 des VPg-Teils in 3AB durch 3Dpol
Rekrutierung des polyadenylierten3´-Endes der Poliovirus RNA
Ausbildung eines „geprimten“Ribonukleoproteinkomplexesmit 2 Uriditylresten (VPgpUpU)
Proteolytische Abspaltung vonVpG durch virale 3C Protease
Elongation des (-) Stranges
Synthese eines vollständigenVPg gebundenen (-) Stranges
© Flint et al. Principles of Virology

Poliovirus (+) Strang Synthese benötigt die Helikase 2C
1. Trennung der beiden Stränge der replikativen Form (RF) durch die Helikase 2C. Diese bindet den (-) Strang
2. Bindung des Poly (rC) Bindeproteins 2 an den B-loop der Kleeblatsttruktur des (+) Strangs
3. Bindung der 3CD-Protease an den D-loop der gleichen Struktur und Assoziation mit 3AB an der Membrane
4. Proteolytische Freisetung von VPg und 3DPol durch 3CDPro (Hetero-und Autoproteolyse).
5. Synthese von VPg-pUpU durch 3DPol am 3´-Ende des (-)Strangs
6. Elongation des VPgpUpU-primers durch 3Dpol bis zur vollständigen Synthese des (+) Strangs.
7. Währenddessen können multiple Reinitiationen erfolgen.
Sinn dieser komplexen Interaktionen viraler und zellulärer Proteine sowie struktur- und
sequenzspezifischer RNA-Elemente ist die Gewährleistung der Spezifität der Poliovirus RNA
Synthese.

Translation viraler RNA in Proteine und RNA-Synthese sind koordinierte Prozesse
© Flint et al. Principles of Virology
Um Kollisionen von Ribosomen bei der Translation mit Polymerasekomplexen bei der (-) Strang Synthese zu verhindern, müssen beide Prozesse koordiniert, d.h voneinander abhängig ablaufen.Bei Poliovirus spielt, neben anderen Prozessen, die Rekrutierung der RNA durch neu synthetisiertes 3CD zu Orten der Replikation eine Rolle. In Abwesenheit von 3CD erfolgt präferentiell die Translation durch Ribosomen.

Replikation und RNA-Synthese von (-) Strang RNA Viren
RNA von (-) Strang RNA Viren ist nicht infektiös- (-) Strang kann durch zelluläre Enzyme weder kopiert noch translatiert werden.
(-) Strang RNA Viren enthalten aktive RdRP im Viruspartikel
Genomreplikation erfolgt über ein komplettes (+) Strang Intermediat
© Flint et al. Principles of Virology
Segmentiert (z.B. Influenza) Unimolekular (VSV)

Struktur und Genomorganisation des nicht segmentierten (-) StrangRhabdovirus: vesicular stomatitis virus (VSV)
leader© Flint et al. Principles of Virology

Replikationszyklus des nicht segmentierten (-) StrangRhabdovirus: vesicular stomatitis virus (VSV)
Bindung und Fusion(Freisetzung des helicalen
viralen Nukleokapsids)
Synthese von 5 mRNAs(ausgehend von leader-Sequenz)
Translation viraler mRNAs(an freien und ER-gebundenen Ribosomen)
Prozessierung und Lokalisierung des G-Proteins
(+) Strang Synthese(unter Beteiligung von N-, P- und L-Protein)
(-) Strang Synthese(unter Beteiligung von N-, P- und L-Protein)
Verpackung (unter Beteiligung von M-Protein)
© Flint et al. Principles of Virology
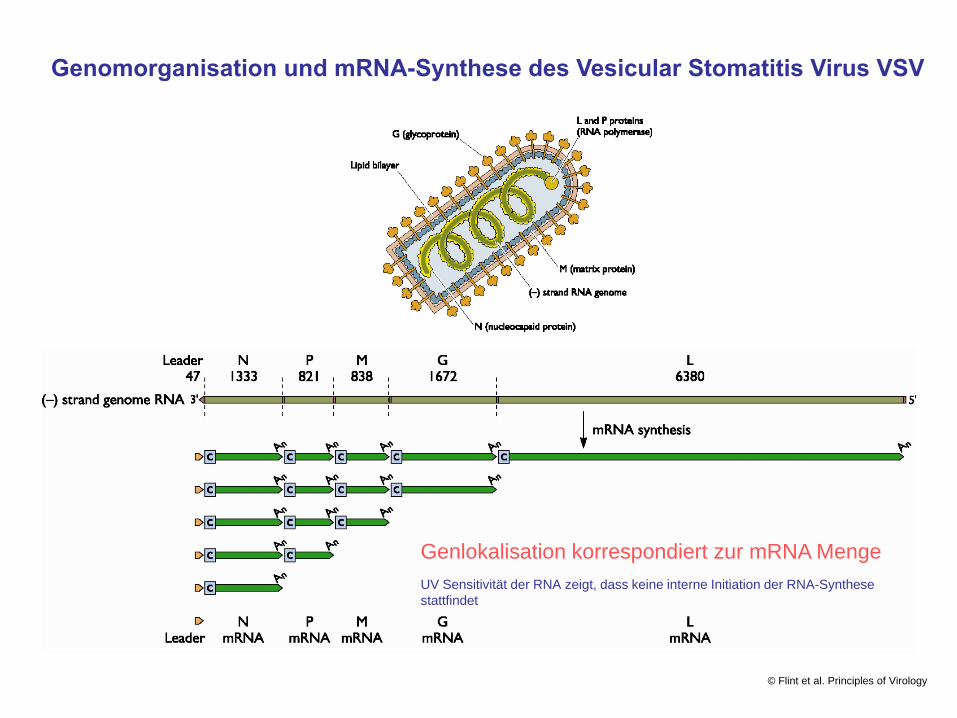
Genomorganisation und mRNA-Synthese des Vesicular Stomatitis Virus VSV
Genlokalisation korrespondiert zur mRNA Menge
© Flint et al. Principles of Virology
UV Sensitivität der RNA zeigt, dass keine interne Initiation der RNA-Synthese stattfindet

Stop-Start Modell der VSV mRNA Synthese
Initiation der RNA Synthese am 3´-Ende des (-) StrangGenoms von VSV (Primerunabhängig)
Synthese einer 47 nt langen „leader“-Sequenz; Termination durch Ablösen der RNA von Polymerase
Reinitiation der RNA-Synthese am 3´-Ende des 1. Gens(N) (nur ein Teil der Polymerasemoleküle reinitiieren)
Synthese der N mRNA und Termination in der folgenden intergenischen Region.
Reinitiation der RNA-Synthese am 3´-Ende des 2. Gens (P)(nur ein Teil der Polymerasemoleküle reinitiieren)
© Flint et al. Principles of Virology
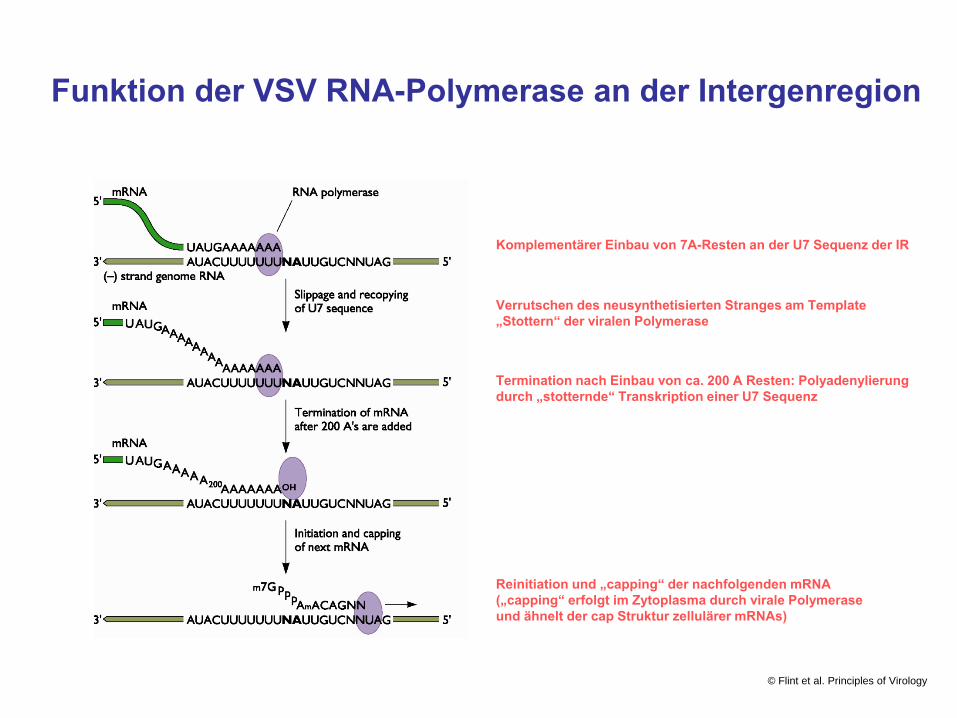
Funktion der VSV RNA-Polymerase an der Intergenregion
Komplementärer Einbau von 7A-Resten an der U7 Sequenz der IR
Verrutschen des neusynthetisierten Stranges am Template„Stottern“ der viralen Polymerase
Termination nach Einbau von ca. 200 A Resten: Polyadenylierungdurch „stotternde“ Transkription einer U7 Sequenz
Reinitiation und „capping“ der nachfolgenden mRNA(„capping“ erfolgt im Zytoplasma durch virale Polymeraseund ähnelt der cap Struktur zellulärer mRNAs)
© Flint et al. Principles of Virology
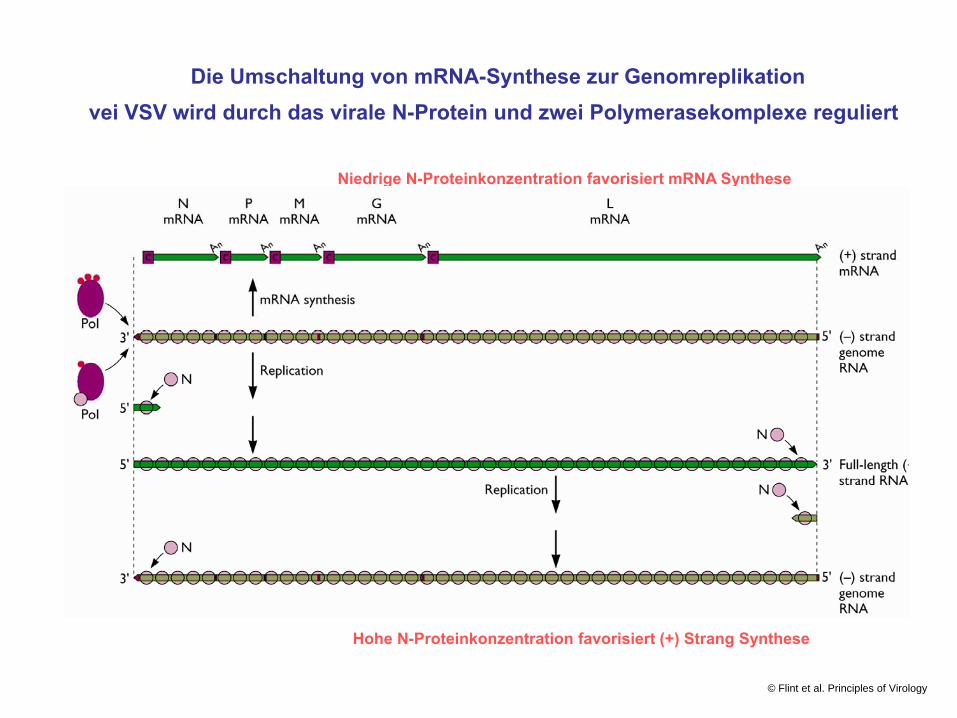
Die Umschaltung von mRNA-Synthese zur Genomreplikationvei VSV wird durch das virale N-Protein und zwei Polymerasekomplexe reguliert
Niedrige N-Proteinkonzentration favorisiert mRNA Synthese
Hohe N-Proteinkonzentration favorisiert (+) Strang Synthese
© Flint et al. Principles of Virology

Struktur und Genomorganisation des segmentierten (-) StrangOrthomyxovirus: Influenza A
© Flint et al. Principles of Virology

Replikationszyklus des segmentierten (-) StrangOrthomyxovirus: Influenza A
Bindung über HA an Sialinsäure(Endozytose)
pH-induzierte Fusion(Freisetzung der 8 Nukleokapsidsegmente)
Kerntransport des Nukleokapsids(NP/P/RNA-Komplex über NLS)
mRNA-Synthese/Export(cap-snatching)
mRNA-splicing(NS2/M2)
Synthese von HA, NA(am rER, Transport)
Synthese von PA, PB1, PB2(Kernimport)
© Flint et al. Principles of Virology

Nukleäre RNA-Synthese bei dem Orthomyxovirus Influenza A
8 Segmente mit (-) Strang Polaritätals Nukleoproteinkomplex im Virion
Synthese gecappter und polyadenylierter mRNA aus (-) Strang Segmenten Inhibierbar
durch α-Amanitin und Actinomycin D !?
Genomisches RNA-Segment mit5´und 3´konservierten Regionen
(vRNA)
„Gecappte“ mRNA mit 10-13 zellulärenNukteotiden und 3´-Poly A (mRNA)
Vollständige Kopie des (+) Segmentes(cRNA)
?
© Flint et al. Principles of Virology

Capping eukaryontischer mRNAs
Capping ist ein Prozess des Editings eukaryontischer mRNAs
Es involviert mehrere Transferasen-katalysierte Reaktionen
Es bildet sich u.a. ein 7N-methylguanosyl cap am 5´Ende
Cap-Struktur stabilisiert mRNA
Gecappte mRNAs sind monocistronisch und polyadenyliert
Capping ist notwendige Voraussetzung für die Initiation derTranslation. Cap bindet an den eukariontischen Initiations-Faktor eIF4F der die Bindung der RNA ans Ribosom vermittelt.
© Flint et al. Principles of Virology

mRNA-Synthese bei Influenza A Virus: „cap snatching“„Cap“-Raub im Nucleus der Wirtszelle
Hydrolyse einer beliebigen nukleären, zellulären mRNA („capping“ erfolgte
durch zelluläre Transferase) durch virale Nuklease (Teil der Polymerase).
Initiation der (+) Strang Synthese durchEinbau eines komplementären GTP an
vorletztem C im (-) Strang Segment:Cap-primer-abhängige Initiation
Elongation durch sukkzessivem Einbaukomplementärer Nukleotide
CpG
G
© Flint et al. Principles of Virology

1
1. Bindung der konservierten (-) Strang Segmentsequenz durch PB1 induziert Bindung gecappter zellullärer mRNA an PB2
2
2. Zur mRNA Synthese aktivierter Polymerasekomplex (Komplex ist inaktiv im freien Zustand)
3
3. Bindung des 3´-Endes an PB1 (hier erfolgt Selektion) und Positionierung des hydrolysierten gecappten zellulärenmRNA-Fragmentes zum 3´Ende
4
4. Templatespezifische Elongation des gecappten primers bis zu einer U7 Sequenz vor dem gebundenen 5´-Ende.Die RNA wird dabei durch den Polymerasekomplex gefädelt (Polymerase ist Fixpunkt, RNA wandert).
5
5. RNA blockiert eigene Elongation und reiterativer Einbau von As (150) führt zur Polyadenylierung und Termination.
© Flint et al. Principles of Virology
0. Inaktiver (keine Cap-Bindung, keine Pol-aktivität) heterotrimerer Polymerasekomplex, bestehend aus PB1, PB2 und PA
Aktivierung des Influenza A Virus PolymerasekomplexesSynthese gecappter polyadenylierter nRNA aus genomischen (-) Strang Segmenten

Regulation von mRNA-Synthese und Genomreplikation beiInfluenza A Virus durch Polymerasemodifikation und NP
PB1 Polymerase wird durch die Bindungvon (-) Strang aktiviert.
Daraufhin bindet PB2 mRNAPA hat hier keine Funktion.
In Gegenwart des viralen NP Proteins erfolgt Bindung an PA. PB2 spielt hier keine Rolle. PA initiiert primerunabhängig die Synthese
vollständiger (+) und (-) Stränge. Die Polymerase PB1 wirkt primer-unabhängig und katalysiert eine komplette cRNA-Synthese
© Flint et al. Principles of Virology
Funktionell verschieden Formen der Influenza RNA-Polymerase führen mRNA Syntese und Genomreplikation aus
mRNA-Synthese Genomereplikation

Genetische Diversität bei Viren mit RNA-Genomen
Die fehlende Akkuratesse von RNA-Polymerasen bedingt den Fehleinbau von Nukleotiden
RNA-abhängige RNA Polymerasen haben keine „proof-reading“ Aktivität (keine Exonuklease).Dies führt zu einem Fehleinbau von jedem 1.000sten bis 10.000sten Nukleotid.Für die Translation hat dies gegebenenfalls Aminosäureaustausche und Funktionsverlustdes Proteins zur Folge.Bezüglich der Replikation bedeuted dies, das typischerweise jedes neue synthetisierte Virion 1-10Mutationen in seinem Genom aufweist.Eine Population von RNA-Viren ist immer heterogen und bildet somit eine „Quasispezies“
Die Segmentierung viraler Genome (Influenza A) ermöglicht Neukombinationen
Koinfektion einer Wirtszelle mit zwei verwandten segmentierten Viren erlaubt eine Neuver-Teilung der viralen Gensegmente.Bei Influenza A Viren führt dies regelmäßig zur neuen Virussubtypen mit antigener Diversität.
RNA-Rekombination ermöglicht den Austausch von viralen und zellulären Gensegmenten
Koinfektion einer Wirtszelle mit zwei verwandten Viren erlaubt Rekombination durch SprungDer Polymerase auf den anderen Strang (Template-abhängig).Rekombination viraler und zellulärer RNAs kann zur Integration neuer gene führen(z.B. Eibau zellulärer Gene der Zellzyklusregulation).
Recommended










![iochemiczne metody badania aktywności i funkcji enzymów ... · WT Dis3 lub Dis3 D551N w obecnościsubstratu RNA znakowanego [32P]-pUpU na końcu 3’ zy mutacja D551N znosi aktywność](https://img.pdfslide.tips/doc/110x75/5d0135cc88c993c93d8c26eb/iochemiczne-metody-badania-aktywnosci-i-funkcji-enzymow-wt-dis3-lub-dis3.jpg)





