
Pécsi Tudományegyetem
Kémia Doktori Iskola
Mannich-ketonok antifungális hatásának vizsgálata és
antimikotikum-érzékenység meghatározása
chip-alapú módszerekkel
Phd értekezés
Bouquet Orsolya
Témavezetők:
Dr. Lóránd Tamás, Dr. Kocsis Béla
egyetemi docensek
PÉCS, 2014

1
KIVONAT
Munkám során Candida törzsek analitikai, elválasztás-technikai módszerekkel való
vizsgálata volt a célom. Két fő vizsgálati irányt céloztam meg mikrofluidikai áramlási
citometriás rendszer és chip-alapú kapilláris gélelektroforézis technika használatával.
Az áramlási citometriát régóta használják Candida sejtek vizsgálatára, antimikotikum-
érzékenység meghatározására, chip-alapú rendszer ilyen célú felhasználását eddig nem írták
le. Irodalmi adatok alapján áramlási citometriás vizsgálatokban élő/elölt sejtek elkülönítésére
alapozva antimikotikum-érzékenység meghatározását végeztem el. A módszert két terápiában
használt antifungális szerre (amphotericin B, fluonazol), valamint Candida albicans mellett
különböző non-albicans törzsekre is teszteltem.
Ezen kísérleteim során nyert adatok alapján, elsőként dolgoztam ki Candida sejtek
antimikotikum-érzékenységének mikrofluidikai módszerrel történő meghatározását. A
mikrofludikai áramlási citometria egy egyszerű, gyors, automatizálható alternatívát kínál a
hagyományos készülékek mellett. Ezzel a technikával megvalósítottam különböző
antimikotikumok különböző törzsekkel szembeni minimális gátló koncentráció (MIC)
értékeinek meghatározását.
Ezt követően egy bizonyítottan citotoxikus és antimikrobiális hatással rendelkező
vegyületcsalád, a Mannich-ketonok Candida albicans-ra gyakorolt hatásának vizsgálatára
tértem rá. Először azt vizsgáltam, hogy a fent leírt mikrofluidikai rendszer alkalmas-e egyes
telítetlen Mannich ketonok MIC értékének meghatározására. Ezeknél a vizsgálatoknál
továbbfejlesztettem a vizsgálati rendszert, több fluoreszcens jelölőt használtam
párhuzamosan. A MIC értékek meghatározása ebben az esetben is sikerült, habár a két
festékes módszer előnyei mellett sem váltotta be előzetes várakozásaimat.
A Mannich-ketonok gombaellenes hatása valószínűleg több összetevő eredménye, a
tiol-depléciós képesség mellett a gombák fehérje-összetételét is befolyásolhatják. A
kondenzált vázas Mannich-ketonok egy csoportjának (15 vegyület) vizsgálatával a Candidák
fehérje-összetételére gyakorolt hatását elemeztem, a vegyületek ezen csoportját részletesen,
ilyen céllal még nem vizsgálták. A mérésekhez egy mikrochip-alapú kapilláris
gélelektroforézis rendszert használtam. Bár általános szerkezet-hatás összefüggéseket nem
vonhattam le eredményeim alapján, de megvalósítottam a Mannich-ketonokkal kezelt
Candida minták fehérjekivonatainak vizsgálatát, ahol néhány Mannich-ketonnal történt
kezelésnél változást tapasztaltam a fehérjeprofilban.

2
TARTALOMJEGYZÉK
1. Rövidítések jegyzéke…………………………………………………………………. 4
2. Bevezetés……………………………………………………………………………... 5
3. Irodalmi összefoglaló………………………………………………………………… 6
3.1. Candida gombák és az antifungális terápia, antimikotikumok áttekintése,
diagnosztikai módszerek
3.1.1. A Candidákról általában……………………………………………………… 6
3.1.2. Az antifungális terápia általános megfontolásai, diagnosztikai módszerek….. 8
3.1.3. Antifungális szerek…………………………………………………………… 9
3.1.3.1. Amphotericin B………………………………………………………... 11
3.1.3.2. Azolok…………………………………………………………………. 12
3.1.3.3. Echinocandinok………………………………………………………... 16
3.1.3.4. Kitinszintézist gátló szerek…………………………………………..... 18
3.1.3.5. Sordarinok……………………………………………………………... 19
3.1.3.6. Egyéb antifungális hatású vegyületek…………………………………. 20
3.2. Mannich-reakció és Mannich-vegyületek………………………………………. 22
3.2.1. A Mannich-reakció jelentősége…………………………………………..… 22
3.2.2. Mannich-bázisok szintézise, enantioszelektív szintézis…………………….. 23
3.2.3. Mannich-reakció felhasználása bioaktív vegyületek szintézisében………… 26
3.2.3.1. α-metilén ketonok szintézise Mannich-bázis intermediereken
keresztül………………………………………………………………. 26
3.2.3.2. Mannich-reakció használata gyógyszervegyületek előállításában….... 26
3.2.3.3. Mannich-reakció természetes vegyületek szintézisében……………… 29
3.2.4. Mannich-bázisok biológiai hatásai…………………………...……………. 30
3.3. Módszerek……………………………………………………………………… 34
3.3.1. Áramlási citometria………………………………………………………..... 34
3.3.2. Chip-alapú áramlási citometria………………………………………….…… 36
3.3.3. Chip-alapú kapilláris gélelektroforézis………………………………………. 38
4. Célkitűzések………………………………………………………………………… 40

3
5. Kísérleti rész………..……………………………………………………………..... 41
5.1. Candida törzsek és tenyésztési körülmények………………………….………. 41
5.2. Fluoreszcens festékek …………………………………………………………. 41
5.3. Antimikotikumok…..………………………………………………………….. 42
5.4. Áramlási citometria és mikrofluidikai áramlási citometria (sejt-chip)………... 45
5.4.1. Candida minták előkészítése áramlási citometriás és sejt-chip
vizsgálatokhoz ……………………………………………………………. 45
5.4.2. Áramlási citometria ……………………………………………………….. 45
5.4.3. Sejt-chip – chip-alapú citometriás módszer………………………………... 46
5.5. Fehérjeösszetétel vizsgálatok…………………………………………………... 47
5.5.1. Mintaelőkészítés…………………………………………………………….. 47
5.5.2. Chip-alapú elektroforézis………………………………………………… 49
6. Eredmények………………………………………………………………………... 51
6.1. Áramlási citometria és sejt-chip……………………………………………….. 51
6.1.1. Candida sejtek detektálása, élő és elölt sejtek megkülönböztetése……….. 51
6.1.2. Candida törzsek megkülönböztetése………………………………………. 55
6.1.3. Antimikotikum-érzékenység vizsgálatok…………………………………… 57
6.1.3.1. Klasszikus antifungális szerek………………………………………. 57
6.1.3.2. Mannich-ketonok…………………………………………………… 65
6.2. Mannich-ketonok hatásának vizsgálata Candida albicans fehérjeprofiljára….. 67
7. Eredmények megvitatása………………………………………………………….. 70
7.1. Áramlási citometriás és sejt-chipes életképesség-vizsgálatok……………….. 70
7.2. Áramlási citometriás és sejt-chipes antimikotikum-érzékenység
meghatározás…………………………………………………………….......... 70
7.2.1. Egy fluoreszcens festékes vizsgálatok……………………………………. 70
7.2.2. Két fluoreszcens festékes vizsgálatok…………………………………….. 72
7.3 Fehérje-vizsgálatok…………………………………………………………… 73
8. Összefoglalás………………………………………………………………………. 74
9. Irodalomjegyzék…………………………………………………………….…….. 75
10. Köszönetnyilvánítás……………………………………………………………… 88

4
1. RÖVIDÍTÉSEK JEGYZÉKE
ADME = adsorption (felszívódás)-distribuiton (megoszlás)-metabolism (metabolizmus)-excretion (kiválasztás)
ATCC = American Type Culture Collection
CFU = colony forming unit (telep képző egység)
CLSI = Clinical and Laboratory Standards Institute
COST = color-space-time (adatgyűjtési mód)
CYP = citokróm P (enzimcsalád)
CZE = kapilláris zóna elektroforézis
DTT = ditiotreit
EDTA = etilén-diamin-tetraecetsav
EOF = electroosmotic flow (elektroozmotikus áramlás)
EUCAST = European Committee on Antimicrobial Susceptibility Testing
FSC = forward light scatter (előre irányuló fényszórás)
GDP = guanozin-difoszfát
LED = light-emitting diode
MALDI = matrix-assisted laser desorption/ionization
MIC = minimum inhibitory concentration (minimális gátló koncentráció)
MOPS = 3-[N-morfolin]-propánszulfonsav
MS = tömegspektrométer
NCCLS = National Committee for Clinical Laboratory Standards
PAS = perjódsav-Schiff (reakció)
PCR = polymerase chain reaction (polimeráz láncreakció)
PI = propidium-jodid
PMSF = fenil-metil-szulfonil-fluorid
QSAR = quantitative structure–activity relationship (kvantitaív szerkezet-hatás összefüggés)
rpm = rotation per minute
RPMI = Roswell Park Memorial Institute
SDS = nátrium-dodecil-szulfát
SSC = side light scatter (oldal irányú fényszórás)
TAMA = N-metilanilin-trifluoracetát

5
2. BEVEZETÉS
Az életet veszélyeztető, invazív gombás fertőzések számának növekedése világszerte
probléma. A leggyakoribb kórokozó továbbra is a Candida albicans, de egyre több a non-
albicans Candida fertőzés, valamint növekszik az Aspergillus sp. okozta megbetegedések
száma is. A rendelkezésre álló antifungális szerek között évtizedek óta használt, klasszikus
szerek mellett a legutóbbi évtizedben bevezetésre került hatóanyagcsalád is található. Mégis a
fertőzések növekvő száma, a rezisztencia terjedése, a kedvezőbb farmakokinetikai és
mellékhatás-profil kialakítása miatt a gomba-ellenes szerek kutatása továbbra is aktuális
terület. Ugyanezen okok miatt a hagyományos kimutatási módszerekhez képest gyorsabb,
kisebb idő – és munkaigényű diagnosztikai és anitmikotikum-érzékenységi vizsgálatok
fejlesztése is időszerű kérdés.
Az áramlási citometriát már régebb óta használják mikrobiológiai vizsgálatokra,
baktériumok, gombák, antimikrobiális szerekkel szembeni érzékenység vizsgálatára, de ilyen
jellegű chip vizsgálatokra kevesebb adat állt rendelkezésre. Napjainkban egyre jellemzőbb a
lab-on-a-chip rendszerek terjedése. Ezeknél a rendszereknél chip-alapon, mikroméretben, kis
mintaigénnyel, gyorsan valósíthatók meg különböző analitikai feladatok.
Munkám során Candida sp. antimikotikum-érzékenységének vizsgálatát valósítottam meg
hagyományos és mikrofluidikai áramlási rendszerben. A klasszikus antimikotikumok mellett
potenciális citotoxikus, antibakteriális és antifungális hatással rendelkező Mannich-
vegyületek Candida-ellenes hatását vizsgáltam chip-alapú áramlási citometriás és chip-alapú
kapilláris gélelektroforézis módszerek felhasználásával. Az eredményeimet a munkához
tartozó, általános irodalmi háttér rövid ismertetését követően mutatom be.

6
3. IRODALMI ÖSSZEFOGLALÓ
3.1. Candida gombák és az antifungális terápia, antimikotikumok áttekintése,
diagnosztikai módszerek
3.1.1. A Candidákról általában
A Candida fajok a mikroszkopikus gombák közé tartozó, eukarióta sejtfelépítésű
élőlények. Sarjadzó gombák, azaz egysejtűek, a gombasejtek mérete 3-6 µm, gömb vagy
ovális alakúak. A Candida sejtek is képesek un. pszeudohifákat képezni, itt az utódsejtek
nem válnak szét teljesen, hanem kolbászfüzér-szerű láncokat alkotva együtt maradnak
[Ádám É. 2001]. A pszeudohifa képzés a biofilmekben való megtelepedés és a patogenitás
szempontjából fontos, a környezeti tényezők, például az oxigén-ellátottság is befolyásolják
[Watanabe T. 2006].
A gombasejtek vázlatos felépítése: sejtfal, sejtmembrán, sejtmag, mitokondriumok,
Golgi-szerű részecskék. Az 1. ábrán mutatom be, kiemelve a gombák legjellegzetesebb
sejtalkotójának, a sejtfalnak szerkezetét. A sejtfalat főképp β-1,3-glukán- és β-1,6-glukán-
molekulák, valamint N-acetil-glükózamint tartalmazó hexóz-polimer, azaz kitin, alkotják.
Az ezek alkotta szilárdabb hálózathoz kapcsolódik egy fehérjékkel asszociált mannóz-
polimer, a gliko(manno)-proteinek rétege. Emellett egyéb fehérjék és kisebb mennyiségben
lipidek alkotják a sejtfalat. A sejtfal glikoproteinjeinek nagy része a β-1,6-glukán
molekulákhoz kapcsolódik a gliko-foszfatidil-inozitol részen keresztül, a fehérjék másik
része direkt módon kötődik a β-1,3-glukán molekulákhoz [Chaffin 2008.]. A
mannoprotein-réteg jelentős szerepet játszik a sejt adhéziójában, a vas megkötésében, a
biofilm-képződésben, a gazdaszervezet immunrendszerével való küzdelemben [Heilmann
és mtsai. 2012]. Fontos látnunk, hogy a sejtfal proteomja egy dinamikusan változó
rendszer, aminek összetételét nagyban befolyásolják a környezeti hatások.
A Candidák sejtmembránja koleszterin helyett ergoszterint tartalmaz, ezért több gomba-
ellenes szer támadáspontja az ergoszterin-bioszintézis és maga az ergoszterin.

7
1. ábra – A Candida sejtfal vázlatos felépítése [1]
A Candida genusba közel 100 faj tartozik, ezek közül a legjelentősebb humán kórokozó
a Candida albicans. A C. albicans mellett egyre gyakoribbak az un. non-albicans törzsek
okozta fertőzések, ezek közül a legfontosabbak a Candida krusei, C. glabrata, C.
tropicalis, C. parapsilosis [Dóczi és mtsai. 2012, Sandven és mtsai. 2006, Tragiannidis és
mtsai. 2012].
A Candida albicans a normál flóra tagja, a légutak, a gastrointesztinális traktus és a női
nemi szervek nyálkahártyáján fordul elő. A Candidák opportunista kórokozók, azaz
egészséges egyéneknél csak enyhébb lefolyású megbetegedéseket okoznak, általában
akkor, ha valamilyen átmeneti, lokális tényező hatására felborul az egyensúly a C. albicans
és a normál flóra többi tagja közt. A leggyakoribb ilyen megbetegedések a soor
(„szájpenész”), az interdigitális candidiasis (kéz- és lábujjközök megbetegedése), az
intertriginális candidiasis (a hajlatok érintettek), a paronychia (fájdalmas körömfertőzés) és
a vulvovaginitisszel járó fluor. Ezen kórképek prognózisa elég jó, bár gyakran
hosszadalmas, ismétlődő kezelést igényelnek. Legyengült immunrendszerű betegeknél a
Candidák gyakran súlyos – életveszélyes, szisztémás fertőzések okozói, mint a generalizált
orális candidiasis, oesophagitis, nephritis, pneumonia, endocarditis és a Candida – szepszis
[Gergely L. 2003]. Mivel a különböző okok miatt, pl. HIV-fertőzés, daganat-ellenes
terápia, szervtranszplantációk, immunszupresszált állapotban levő betegek száma
világszerte növekszik, ezen betegségek diagnosztikája és kezelése egyre jelentősebb.

8
3.1.2. Az antifungális terápia általános megfontolásai, diagnosztikai módszerek
A fertőzést okozó gombák gyors, pontos diagnosztikája és gombaellenes szerekkel
szembeni érzékenységének meghatározása a fentebb vázolt okok miatt fontos feladat.
A diagnosztika első lépése a direkt mikroszkópos vizsgálat. A gombák többsége a
baktérium-kimutatáshoz használt Gram-festéssel is festődik, így történik a Candida fajok
direkt mikroszkópos vizsgálata különböző vizsgálati anyagokból. A mintákban 3-4 µm
méretű sarjadzó gombasejtek láthatók szeptált hifákkal (5-20 µm), vagy nélkülük, végükön
esetleg chlamydospórákkal. A C. albicansra igen jellemző még a csíratömlőképzés, azaz
néhány órás, szérumtartalmú táptalajon történő tenyésztés után a gombasejtek
mikroszkópban jól látható jellegzetes nyúlványokat képeznek. Gombakimutatásra
használhatók más festési eljárások is, mint calcofluor-, PAS- és meténamin-ezüst-festés
(szövettani metszetekben).
A gombák tenyésztésére sokféle táptalajt dolgoztak ki, legelterjedtebb a Sabouraud-féle
glükózagar, amely szénforrásként 5% glükózt, nitrogénforrásként peptont tartalmaz és
enyhén savas kémhatású (pH=5,6), vagy az RPMI 1640 tápfolyadék. Candida speciesek
elkülönítésére szolgál az ún. chrom-agaron történő tenyésztés. Ennek lényege, hogy
különböző Candida fajok szelektív (baktériumokat kizáró) táptalajon tenyésztve a
kromogén szubsztrátokat különböző színnel inkorporálják.
A tenyésztés mellett biokémiai reakciókat is használnak a gombák identifikálására,
ilyen például a szénhidrát –asszimiláció vagy a szénhidrát-fermentáció vizsgálata. A
Candida-bőrpróba gyakorlatilag minden egészséges felnőttnél pozitív, negatív eredmény
inkább az immunválasz elégtelenségét jelzi. A szerológiai vizsgálatok jelentősek, főleg, ha
a tenyésztés negatív (fennálló klinikai tünetek ellenére), illetve candidiasisok esetén, de
kiértékelésük nehezebb, mint a baktérium- vagy vírusdiagnosztikában. A PCR alapú
gyorsdiagnosztikai eljárások is egyre elterjedtebbek, ezek időigénye a klasszikus
módszerekéinél jóval rövidebbek [Avni és mtsai. 2011].
A gombák identifikálása mellett a másik fontos feladat az antifungális szerekkel
szemben tanúsított rezisztencia/érzékenység megállapítása. Ez azért is jelentős, mert az
antifungális szerek egyre gyakoribb használata miatt növekszik a rezisztens törzsek
előfordulása. A rezisztencia főbb mechanizmusai: az antifungális szer csökkent felvétele a
sejtbe, illetve fokozott kiürülése efflux-pumpákon keresztül; a célmolekula módosulása
vagy a célenzim mennyiségének növekedése [Loeffler és Stevens, 2003]. Az
antimikotikum-érzékenységi vizsgálatok eredményét a vizsgálati körülmények nagyban
befolyásolják, így a módszerek standardizálására több ajánlás is létezik, legelterjedtebbek a

9
Clinical and Laboratory Standards Institute (CLSI, korábban NCCLS – National
Committee for Clinical Laboratory Standards) szabványok. Az európai standardizálási
törekvéseket az European Committee on Antimicrobial Susceptibility Testing (EUCAST)
ajánlások jelzik, a két rendszer átfedést mutat, de például a CLSI rendszer határértékei,
azaz azon koncentrációk, amik nem csak a laboratóriumi tesztben, de a terápiában is
hatékonyak, nem használhatók az EUCAST méréseknél [Espinell-Ingroff és mtsai. 2005].
Az antimikotikum-érzékenységet vizsgáló módszerek közé tartozik a hagyományos
csőhígítási-sor [M27-A3 CLSI, 2008], vagy másnéven makrodilúciós eljárás, illetve ennek
plateken kivitelezett mikrodiluciós változata. Az előző esetben az antimikotikumból a
megfelelő médium 1 – 1 ml-ével, a mikrodilució esetében 100 – 100 µl-ével hígítási sort
készítünk. Beoltás után a minimális gátló koncentráció (MIC) meghatározása vizuálisan
történik, eredmény minimum 24, jellemzően 48 óra múlva jelenthető ki. A rutin
labordiagnosztikában a fenti módszernél egyszerűbben alkalmazhatók az agar-diffúziós
tesztek, mint a korong-diffúziós módszer vagy az E-teszt, illetve használják még a
VITEK ® félautomata rendszereket [Arendrup 2013].
Ezek mellett számos alternatív, gyorsdiagnosztikai módszert is fejlesztettek
érzékenység-vizsgálatokra. Ilyen például a MIC meghatározása fluoreszcens próbákkal
[Peter és mtsai. 2005] vagy az a kolorimetriás próba, ahol az antimikotikumok fonalas- és
élesztőgombákra gyakorolt, sejtek lízisét okozó hatását határozták meg [Jewell és mtsai.
2002]. Az áramlási citometriát specifikus fluoreszcens jelölők használatával szintén
alkalmasnak találták antimikotikum-érzékenység vizsgálatára [Ramani és mtsai. 1997].
Az antifungális terápia megválasztásában fontos az időfaktor, gyakran a pontos
diagnosztikai és érzékenység-vizsgálat eredménye előtt szükséges az empirikus terápia
megkezdése. A nem-szisztémás, dermatológiai kórképek esetén is általában empirikus
módon, valamilyen azol-származékkal mielőbb megkezdik a terápiát. Az
anitmikotikumokat többnyire monoterápiában alkalmazzák, a különböző antifungális
hatóanyagok kombinált használata még nem kiforrott terület [Johnson és Perfect 2010]. A
következőkben a fontosabb antifungális gyógyszervegyületeket részletesebben tárgyaljuk.
3.1.3. Antifungális szerek
Az antifungális szereket több szempont alapján is csoportosíthatjuk. Először
beszélhetünk természetes antifungális hatású és szintetikusan előállított vegyületekről.
Előbbiek közé például a griseofulvin, a polién makrolidok, illetve részben az

10
echinocandinok, míg az utóbbi csoportba az allilaminok, azolok, a flucitozin és a
fenilmorfolin-vázas vegyületek tartoznak.
Hatás szempontjából beszélhetünk fungicid, azaz gombaölő, illetve fungisztatikus, azaz
a gomba szaporodását gátló szerekről, bár ez a tulajdonság igen gyakran
koncentrációfüggő.
Kémiai szerkezet alapján is feloszthatjuk az antifungális szereket, mint:
- polién makrolidok (amphotericin B, nystatin)
- azolok: ezen belül is di – és triazolok
- echinocandinok
- egyéb: sordarinok, nikkomycinek, allilaminok (pl.:terbinafin), stb.
Leggyakrabban azonban hatásmechanizmus alapján csoportosítják az antifungális szereket
(ld. 2. ábra). Így beszélhetünk:
- a sejtfal-dezintegráló szerekről, mint az amphotericin B
- a gomba szterin-szintézisét gátló szerekről, mint az azolok, allilaminok
- a sejtfal-szintézis egyéb gátlóiról, mint az echinocandinok (glukán-szintáz gátlása)
vagy a nikkomycinek (kitin-szintáz gátlása)
- fehérje és/vagy nukleinsav szintézis gátló szerekről, pl. sordarinok, flucytosin
2. ábra – Leggyakoribb antimikotikumok hatásmechanizmusának vázlata [2]

11
A klinikai gyakorlatban régóta használt, illetve az újabb típusú antifungális szerek közül
néhányról részletesebben is érdemes beszélnünk.
3.1.3.1. Amphotericin B
Az amphotericin B (3. ábra) a Streptomyces nodosus által termelt heptaén makrolid
antibiotikum. Amfoter, amfipatikus bot-alakú molekula, amely hidrofób és hidrofil oldallal
is rendelkezik. Fungicid, alacsony koncentrációnál fungisztatikus hatású. Vízben
oldhatatlan, 37 oC-on labilis, 4 oC-on hetekig stabil. A gastrointestinalis traktusból alig
szívódik fel, orálisan adva csak a béllumenben levő gombákra hat. Szisztémás fertőzések
esetén intravénásan adagolandó, farmakokinetikája azonban kérdéses, a szérumszint nem
emelkedik párhuzamosan a dózissal. Plazmafehérjékhez erősen (>90%) kötődik, a
szövetekben felhalmozódik, kis mennyisége bejut a liquorba, átjut a placentán.
Eliminációjában a biliáris excretio jelentős, a vizeletben kis koncentrációban sokáig
kimutatható.
3. ábra – Az amphotericin B szerkezeti képlete
Az amphotericin B a membrán ergoszterin komponenséhez kötődik, pórusokat
képez, így metabolikus zavarokat, K+ és más ionok, kis molekulák elvesztését okozza,
ezeken keresztül pedig a sejt pusztulását eredményezi. Egyes vizsgálatok szerint
antifungális hatásában más folyamatok is, mint például az oxidatív stressz indukálása,
jelentős szerepet játszanak. Az amphotericin B erős immunmoduláló hatását kimutatták a
gazdaszervezet sejtjein is, ennek szerepe azonban egyelőre nem tisztázott [Mesa-Arango és
mtsai. 2012]. Emellett az amphotericin B kisebb mértékben az emlős sejtek
koleszterinjéhez is kötődik, ezért erős mellékhatásai vannak, úgy mint a nephrotoxicitás,
láz, fejfájás, máj- és csontvelő-károsodás, helyileg vénagyulladás.

12
Egyes tanulmányok vizsgálták, hogy az amphotericin B különböző funkciós
csoportjait érintő kovalens kémiai módosítások hogyan befolyásolják a hatását, de nem
találtak egyértelmű összefüggéseket, bár úgy a protonált amino-csoport pozitív töltésű N-
atomja szükséges az ergoszterinhez való kötődéshez. Érdekes lehetőség a szén
nanocsövekkel, vagy különböző poliszacharidokkal való konjugált komplexképzés is, bár
ezek klinikai felhasználása kétséges [Sedlák 2009].
A lipidekkel kapcsolt, illetve a liposzomális amphotericin B klinikai gyakorlatban
használt, nálunk is elérhető a liposzomális amphotericin B (Ambisome), az amphotericin B
lipid-komplex (Abelcet), és az amphotericin B kolloidális diszperzió (Amphocil). A
liposzomális amphotericin B liofilizált formában tartalmazza a hatóanyagot, amelyet
kisméretű (<100nm) egyrétegű lipid-kettősréteg (ezt főleg foszfatidilkolin, koleszterin és
diszteroil-foszfatidilglicerin alkotja) burkol. Ezek a formák hatékonyak, és csökkentik a
mellékhatások, elsősorban a vesekárosodás mértékét [Moen 2009]. Az amphotericinnel
szembeni rezisztencia ritka, non-albicans törzseknél írtak le ilyet néhány esetben.
3.1.3.2. Azolok
Az azolok antifungális terápiában leggyakrabban használt szerek, köszönhetően széles
hatás-spektrumuknak, kémiai stabilitásuknak és orális alkalmazhatóságuknak.
Mellékhatásaik a poliénekhez képest jóval enyhébbek, pl. hányinger, hányás, bőrkiütések.
Azonban mivel potenciális inhibitorai a citkróm P450 (CYP) 3A4 enzimnek, mely számos
gyógyszer metabolizmusában részt vesz, a gyógyszer-kölcsönhatásokra figyelni kell. Két
nagy csoportra bonthatók: az imidazolokra és a triazolokra.
Hatásukat az ergoszterin-szintézis gátlásával fejtik ki, megakadályozzák a
lanoszterin 14-α-demetileződését ergoszterinné, és a poszt-szkvalén szintézis további
elemeit, az oxidoszkvalén-ciklázt és a C-24-metiltranszferázt is gátolhatják. Ezen kívül
néhány gomba törzsnél a ∆22-deszaturáz enzimet, a szintézis utolsó lépését is gátolhatják.
A ketokonazol az egyik legrégebb használt diazol vegyület. A ketokonazol, ahogy
a 4. ábrán is látható, királis vegyület, 2 kiralitáscentrummal rendelkezik, gyógyszerként a
racém elegyet használják, mindkét enantiomer cis konfigurációjú. A cisz konfiguráció itt
azt jelenti, hogy a gyűrű 4-es számú szénatomjához fűződő hidrogén és a 2,4-diklórfenil-
csoport az öttagú dioxolán-gyűrű azonos oldalán található. Az enantiomerek reszolválása
HPLC-vel és szuperkritikus folyadékkromatográfiával is lehetséges [Thienpont és mtsai.
1999].

13
(+)-(2R, 4S) (+)-(2S, 4R)
4. ábra – Ketokonazol-enantiomerek
Imidazol-vázas antifungális hatású vegyületek még a klotrimazol, azaz az 1-[(2-
klórfenil)(difenil)metil]-1H-imidazol és az ekonazol, vagyis az (RS)-1-{2-[(4-
klórfenil)metoxi]-2-(2,4-diklórfenil)etil}-1H-imidazol. Ezeket főleg külsőleg, lokális
gombaellenes készítményekben használják. Az imidazol-vázas vegyületek antifungális
hatását vizsgáló tanulmányok szerint két vagy három aromás gyűrű jelenléte szükséges az
antifungális hatáshoz, az aromás gyűrűhöz kapcsolt halogénatom vagy apoláris funkciós
csoport, valamint 2- vagy 2,4- helyzetű szubsztitúció szintén növeli a hatásosságot [Rani és
mtsai. 2013].
A flukonazol (5. ábra) már a triazolok közé tartozik, molekulájában diklórfenil
csoport helyett difluorofenil-csoport található. Ez az egyik leggyakrabban használt azol,
hatását szintén az ergoszterin-szintézis gátlásával fejti ki. C. glabrata és C. krusei fajok
primer rezisztenciával rendelkeznek flukonazolra. Szerzett rezisztencia kialakulása jelentős
probléma, a rendszeres, profilaktikus flukonazol-szedéshez kapcsolódva írták le, C.
albicans és non-albicans törzsek esetében is. Flukonazol-rezisztens törzsek előretörésében
a korábbi antibiotikum-kezelés szerepét is kimutatták [Ben-Ami és Giladi 2012]. A
rezisztencia mechanizmusai: a C14-lanoszterin-demetiláz enzimet kódoló Erg11p gén
mutációja, az enzim fokozott expressziója, valamint a flukonazol sejtből való eltávolítását
segítő efflux-pumpák mennyiségének növekedése [Espinel-Ingroff 2008, White és mtsai.
1998].

14
5. ábra - Flukonazol
Az itrakonazol (6. ábra) is az orálisan alkalmazható azolok közé tartozik, bőr- és
körömfertőzések kezelésében a flukonazol mellett használják. Érdekesség, hogy az azolok
gyógyszerformulázása során ciklodextrin-származékokat is kipróbáltak, így készülhet
itrakonazolból orális szuszpenzió és intravénás oldat is [Mallié és Bastide, 2001].
6. ábra - Itrakonazol
Bőrfertőzések kezelésére külsőlegesen használt diazol-származék a bifonazol is (7. ábra),
antifungális hatása mellett Gram-pozitív coccusok ellen is hatékony. Bifonazol-származékok
enantioszelektív szintézisével Botta és mtsai foglalkoztak [2000]. A szintézisek a megfelelő
(S)- vagy (R)-1-fenil-2-propinilaminból indultak ki. A szerzők a tiszta enantiomerek
antifungális hatását is megvizsgálták. Várakozásaikkal ellentétben, egyetlen esetben sem volt
különbség a vizsgált enantiomer pár tagjainak antifungális aktivitásában. Ezt nem az anyagok
kísérlet közbeni racemizációja okozza, és nem is azt jelenti, hogy a célmolekula, a
lanoszterin-14α-demetiláz érzéketlen lenne a gátló anyag sztereokémiai felépítésére. A
jelenség oka valószínűleg a hatásmechanizmus más pontján van, valahol a sejtbe való felvétel
és a célenzim gátlása közt.

15
7. ábra – Bifonazol
Az újabb típusú antifungális vegyületek mind triazolok. A vorikonazol (8. ábra)
szerkezete a flukonazollal rokon, ahol az egyik triazol-gyűrűt fluorpirimidin-gyűrűre
cserélték és a propanol-vázhoz egy metilcsoportot kapcsoltak. Nagyon jó az orális
biohasznosulása, fungisztatikus a legtöbb Candida esetében, fungicid hatású egyes
Aspergillus és Fusarium törzseknél. Mivel penetrál a vér-agy gáton, központi idegrendszeri
aspergillozis esetén is alkalmazható. A májban CYP enzimeken át metabolizálódik, így
számos gyógyszer, mint például a ciklosporinok, benzodiazepinek, omeprazol, digoxin
plazmaszintjét emelheti, ezt fontos figyelembe venni alkalmazásakor [Odds és mtsai.
2003]. A metabolizáló-képesség eltérő erősséget mutat a humán populációban, így
esetenként nagyobb dózisok alkalmazása szükséges, ami a mellékhatásokat, például a látási
zavarokat, a májfunkció csökkenését, erősítheti.
8. ábra - Vorikonazol
9. ábra - Ravukonazol
A ravukonazol (9. ábra) is hatékony patogén Candida és Aspergillus törzsekre, előnye
hosszabb plazmafelezési ideje és enyhébb mellékhatásprofilja.
A posakonazol (10. ábra) az itrakonazollal rokon szerkezetű vegyület, fungicid hatású C.
albicans, C. krusei, Aspergillus, Fusarium törzsek esetén, valamint zygomycosisok esetén
is használható.

16
10. ábra - Posakonazol
Az azolok részletesebb bemutatásával kívántam szemléltetni azt az antifungális
szereknél is igaz tényt, hogy a gyógyszermolekulákon végrehajtott viszonylag nem nagy
módosítások jelentősen befolyásolhatják ezek farmakokinetikai tulajdonságait és
felhasználhatóságukat.
3.1.3.3. Echinocandinok
Az echinocandinok a klinikai gyakorlatban használt antifungális szerek újabb
képviselői. A 2000-es évektől kezdve kerültek forgalomba; a caspofungin mellett már a
micafungin és az anidulafungin is elérhető a világ nagy részén. Az echinocandinok nagy
molekulatömegű (~1200kDa) lipopeptidek, kémiailag ciklusos hexapeptidek, amelyek
nagy szénatomszámú zsírsavakkal vannak acilezve. Félszintetikus lipopeptidek, azaz
például a caspofungint a Glarea lozoyensis által termelt pneumoncandin B, míg az
anidulafungint az Aspergillus nidulans által termelt echinocandin B módosításával
állították elő. Az egyes vegyületek főleg farmakokinetikai tulajdonságaikban, illetve egyéb
gyógyszerekkel való interakciók területén különböznek. Részletes vizsgálatok
megmutatták, hogy például egy aminoprolin-csoport beépítése azon túl, hogy a
vízoldékonyságot javítja, növeli az in vivo hatékonyságot is [Klein és mtsai. 2000]. Az
echinocandin B, mint alapvegyület, valamint a caspofungin és a micafungin képletét a 11.
ábra mutatja be. Ez utóbbi molekula egy hosszú optimalizáció eredménye, a félszintetikus
micafungin acilcsoportja a természetes zsírsavaktól meglehetősen eltérő acil-oldallánc. Ez
a csoport a molekulának megfelelő logP értéket ad, amely optimális az antifungális hatás
szempontjából. A micafungin igen hatékony antifungális szer, mely alacsony hemolitikus
aktivitással rendelkezik. [Fujie 2007].

17
megj.: DMM=dimetilmerisztoil
11. ábra – Néhány echinocandin vegyület felépítése
Hatásmechanizmus: az echinocandinok a β-1,3-glukán-szintáz nem kompetitív
inhibitorai, mivel a β-1,3-D-glukán a gomba sejtfalának alapvető komponense, hiánya a
sejtek lízisét okozza. Az érzékenységi vizsgálatok alapján a hatáshoz valószínűleg más,
egyelőre nem tisztázott folyamatok is hozzájárulnak [Chen és mtsai. 2011]. Szelektivitásuk
alapja, hogy az emberi sejtek nem tartalmaznak β-1,3-D-glukánt. Előnyük még, hogy nem
szubsztrátjai és nem is befolyásolják CYP 450 enzimeket, így a gyógyszerkölcsönhatások
is ritkábbak.
Az echinocandinok hatékonyak a legtöbb Candida és Aspergillus törzzsel szemben,
különösen C. albicans és A. fumigatus ellen. Csak parenterálisan alkalmazhatók, infúzió
formájában, terápiás felhasználásuk a candidaemia, invazív és oesophagalis candidiasis,
különösen, ha egyéb szerek mellékhatás vagy rezisztencia miatt nem jöhetnek szóba.
Vizsgálták hatékonyságukat szervtranszplantációt és kemoterápiát követő gomba-
profilaxisban is [Scott 2012]. Az echinocandinokkal szemben rezisztencia nem jellemző,
mellékhatásaik a többi antifungális szerhez képest enyhék (bőrvörösség, kiütések),
felhasználásukat korlátozza még, hogy elég drágák [Kaufmann 2006].

18
3.1.3.4. Kitinszintézist gátló szerek
A kitin, bár kisebb mennyiségben van jelen, a sejtfal esszenciális alkotóeleme, a
glukán rétegtől elkülönül, de ahhoz kovalensen kapcsolódik, így adva meg a sejtfal
szilárdságát. A kitinszintézis gátlói közé tartoznak a nikkomycinek és a polyoxinek,
melyeket különböző Streptomyces fajokból izolálták.
A nikkomycinek családjából a leghatékonyabb vegyületek a nikkomycin X és a
nikkomycin Z, a 12. ábrán ez utóbbi képlete látható.
12.ábra – Nikkomycin Z 13.ábra – UDP-GlcNAc
A hatásmechanizmus azon alapul, hogy szerkezete hasonló a kitin-szintáz szubsztrátjához,
az uridin-difoszfát N-acetil-glükózaminéhoz (UDP-GlcNAc, 13. ábra), így ennek
kompetitív inhibitora. Elsősorban a kitin-szintáz I. és III. típusát képes gátolni. Általában
nem hatékony C. albicans, C. tropicalis és Aspergillus törzsek elllen, míg Blastomyces
dermatitidis vagy Histoplasma capsulatum esetén hatásosnak bizonyult [Lóránd és Kocsis
2007]. Az új típusú nikkomycinek fejlesztésénél cél a kitin-szintáz inhibitor hatás
erősítése, illetve a sejtbe való bejutás hatékonyságának növelése. Így vizsgálták például a
terminális aminosav-rész módosításait [Obi és mtsai. 2000]. Ezeknek a vegyületeknek
klinikai felhasználása még nem valószínű, de alapot adhatnak a további fejlesztésekhez
[Behr és mtasi. 2003].
A polyoxinek szerkezete a nikkomycinekhez hasonló, néhány polyoxin felépítését a 14.
ábra mutatja.

19
14. ábra – Néhány polyoxin felépítése
A polyoxinek in vitro hatékonyak a kitin-szintáz gátlásában, de Candida ellenes hatásuk
erősen kérdéses. Hatékonyságuk függ a gombasejt növekedési állapotától és a tenyésztési
körülményektől is, a sejtekbe való bejutásukhoz a peptid-transzport utakat veszik igénybe
[Mehta és mtsai. 1984].
3.1.3.5. Sordarinok
A sordarint (15. ábra) a Sordaria araneosa egy metabolitjaként izolálták az 1970-es
években. Kémiailag a sordarinok tetraciklusos diterpén glikozidok.
15. ábra – Sordarin
Az alapvegyület a sordaricin, itt R=H (lásd 15. ábra). A sordarinok a fehérje-szintézis
befolyásolásával fejtik ki hatásukat. Az elongációs fázisban az eEF2 elongációs faktorhoz
kötődnek, valamint stabilizálják az eEF-2 –GDP-riboszóma komplexet. A sordarin hatásos
többek közt C. albicans, C. tropicalis törzseken, ugyanakkor számos non-albicans törzs
primer rezisztenciát mutat [Shastry és mtsai. 2001].
A természetes sordarinok mellett, mint a zofimarin vagy az újabb moriniafungin (amely
non-albicans törzsek ellen is hatékony), számos szintetikus sordarin származékot is

20
előállítottak [Liang 2008]. Például sordaricin analógokat szintetizáltak ciklohexil gyűrű
beépítésével, itt a tioéter- és az oxim-származék bizonyultak hatékonynak (ld. 16. ábra)
[Kaneko és mtsai. 2002].
16. ábra - Antifungális hatású sordarin vegyületek: tioéter- és oxim-származék
Néhány módosított vegyület candidiasis, aspergillosis és pneumocystosis ellen
állatkísérletekben is hatékonynak bizonyult [Martinez és mtsai. 2000]. Nagy előrelépést
jelentett a (-) –sordarin totál szintézisének megvalósítása is [Chiba és mtsai. 2006].
Ezek az eredmények bíztatóak a jövőbeni klinikai felhasználás szempontjából.
3.1.3.6. Egyéb antifungális hatású vegyületek
A flucytosin, az uracil antimetabolitja, egy fluor tartalmú pirimidin-vázas vegyület,
a nukleinsav-szintézis gátlásán keresztül fejti ki hatását. Antifungális spektruma
korlátozott, vizsgálják kombinációban való alkalmazását egyéb szerekkel.
Az amorolfin fenilmorfolin-vázas vegyület, az ergoszterin-bioszintézisre gyakorol
hatást [Carillo-Munozés mtsai. 2006]. A terbinafin az allilaminok közé tartozik, az
ergoszterin-bioszintézisbe a szkvalén-epoxidáz szintjén avatkozik bele. Magyarországon is
forgalomban van, helyileg és orálisan alkalmazzák bőr- és körömfertőzések, esetenként
vulvovaginális candidiasis kezelésére [Rodriguez és Patrick, 2001].
A nystatin egy tetraén-dién tíusú makrolid, szerkezete az amphotericin B -hez
nagyon hasonló, külsőleg, valamint a száj-, garat- és a gasztrointestinális-traktus gombás
fertőzéseinél használják. A Nyotran® a nystatin liposzómális formája, melyet az
amphotericin B példájára intravénás alkalmazásra fejlesztettek [Johnson és mtsai. 1998].
A griseofulvin egy benzfuranon-származék, a spirociklusos vegyületek közé tartozik, a
Penicillium griseofulvum másodlagos metabolitja. Régóta használják, ez volt az első

21
gomba által termelt gombaellenes szer. Specifikusan a gombák microtubulus-
szerveződését befolyásolja, csak az un. dermatophytonokra (Tinea spp.) hat, melyek
bőrfertőzéseket okoznak. Mivel szájon át is szedhető és olcsó, ezért egyes országokban
napjainkban is használják [Odds 2003].
A felsoroltak mellett sok más anyag, például gyantasav-származékok [Savluchinske-
Feio és mtsai. 2006], polikationos vegyületekkel módosított felületek is potenciális
antifungális hatásúak. Ez utóbbi esetben különböző felületek (papír, pamut, selyem,
gyapjú) szabad primer hidroxil-csoportjait funkcionalizálták lipofil-láncot is hordozó
dikationos vegyületekkel. Az így módosított felületeken élesztő- és penészgombák
megtelepedése és szaporodása is akadályozott volt [Cohen és mtsai. 2004].
Antifungális hatású vegyületek szerkezet-hatás összefüggéseinek vizsgálatával számos,
egyéb célra használt gyógyszermolekula gombaellenes hatását is feltételezik. Ilyen szerek
például az antidepresszáns szertralin, amely feltételezett támadáspontja a gomba fehérje-
szintézise vagy az ösztrogén-receptor antagonista tamoxifen [Butts és Krysan 2012].
Figyelmet érdemelnek azok a vizsgálatok, melyek kisméretű fehérjék szerepét
vizsgálják különböző fertőzések elleni védekezésben. Hatásuk lehet a kórokozók direkt
elpusztítása, immunmodulátor hatás, vagy a patogének virulencia faktorainak gátlása.
Néhány ilyen antifungális hatású peptid, illetve proteináz-inhibitor az RNáz-7, lizozim,
lactoferrin, calprotectin, defensinek, dermcidin [Mehra és mtsai. 2012].
Bizonyos vaskelátoroknak ugyancsak lehet terápiás szerepe a gombás fertőzések
kezelésében, például a mucormycosis terápiájában a desferoxamin-származékok szerepet
kaphatnak [Ibrahim és mtsai. 2008]
A fenti felsorolás korántsem teljes, csupán egy vázlatos áttekintése az antifungális
hatású vegyületeknek.

22
3.2. Mannich-reakció és Mannich-vegyületek
3.2.1. A Mannich-reakció jelentősége.
Mannich-reakciónak nevezzük az α-helyzetű hidrogénatomot tartalmazó
karbonilvegyületek aminometilezését formaldehiddel és primer vagy szekunder aminokkal,
vagy ammóniával. Oxovegyületként aktív hidrogént tartalmazó vegyületeket, elsősorban
aldehideket vagy ketonokat, valamint észtereket vagy egyéb CH-savas vegyületeket
(nitroalkánokat és nitrileket) használhatunk, aminként primer vagy szekunder aminok sóit,
esetleg amidokat alkalmazhatunk [Faigl és mtsai 2011].
Klasszikus értelemben vett Mannich-reakcióban egy enolizálható aldehidet vagy
ketont hevítünk formaldehiddel és amin-hidrokloriddal savas közegben. A reakció általános
egyenletét a 17. ábra mutatja.
I
17. ábra – Klasszikus sav-katalizált Mannich-reakció egyenlete
A Mannich-reakcióval szintetizálható β-amino-ketonok és -aldehidek (Mannich-
bázisok), számos jól használható és értékes vegyület építőkövei, legfontosabb felhasználási
területük különböző gyógyszervegyületek szintézise. Emellett növényvédőszerek, festékek
előállításánál és a polimerkémiában is fontosak, mint szilárdságot fokozó, keresztkötő és
reakció-gyorsító anyagok [Tramontini és mtsai. 1988; Rotaru és mtsai. 2007].

23
3.2.2. Mannich-bázisok szintézise, enantioszelektív szintézis
A klasszikus Mannich-reakciónak (17. ábra) számos korlátja van. A drasztikus
reakciókörülmények és a hosszú reakcióidő miatt nem kívánatos termékek is képződnek,
probléma az alacsony szelektivitás és a kompetitív aldol-reakciók fellépése. Így primer
aminnál vagy ammóniánál a reakció addig folytatódhat, míg a nitrogén valamennyi H-atomja
helyettesítődik. Ezt megelőzendő a két reaktív α-helyzetű hidrogénatomot tartalmazó
ketonokat nagy feleslegben használják. A hagyományos reakciónál általában csak
formaldehid, illetve ketonok vagy aldehidek használhatók. Ez a típusú reakció általában nem
megfelelő a Mannich-bázisok enantioszelektív szintézisére, a termék racém elegy [Arend és
mtsai. 1998].
Az enantioszelektív, megfelelő termelést biztosító Mannich-típusú reakciók a
gyógyszervegyületek szintézise szempontjából kiemelkedően fontosak, ezért számos
módszert kidolgoztak ezen problémák megoldására.
A reagensek megfelelő megválasztásával biztosítható az elektrofil részecskék
magasabb koncentrációja, így a reakcióidő és a reakció hőmérséklete csökkenthető. A
hagyományosan használt karbonil-származékok (aldehidek, ketonok) helyett reaktívabb
megfelelőik, pl. enolátok vagy szilil-enol-éterek is használhatók. A reakcióval nem csak
aminometilezés, de aminoalkilezés, illetve a sztereoszelektív szintézis is megvalósítható.
Ilyen reagensek lehetnek még az enolizálható iminek, amelyeket felhasználnak karbonsav-
származékok sztereoszelektív aminoalkilezésére.
Az eddigi reakciók intermolekuláris Mannich-reakciók voltak. Az intramolekuláris
Mannich-reakció az azaheterociklusok szintézisében használható fel legsokoldalúbban.
Ezekben a reakciókban aciklusos kiindulási anyagokból képződnek gyűrűs termékek;
jellemző rájuk a nagyfokú kemoszelektivitás. Az intramolekuláris Mannich-reakciók
felhasználhatók természetes vegyületek - pl. alkaloidok szintézisére [Davis és mtsai. 2009]. A
3.2.3 fejezetben több konkrét példát is mutatok intramolekuláris Mannich-reakcióra. Egy
további fejlesztése ennek a reakciónak az ún. vinilóg típusú Mannich-reakció. Ennél egy α,
β-telítetlen ketont γ-helyzetben aminoalkileznek [Tanino és mtsai. 1992].
Enantioszelektív szintézisre ad lehetőséget a direkt, prolin-katalizált
háromkomponensű Mannich-reakció. Aceton, p-nitrobenzaldehid és p-anizidin kiindulási
anyagokból DMSO oldószerrel prolin jelenlétében 90% feletti enantiomer-felesleggel
képződött a Mannich-bázis megfelelő enantiomere (II - 18. ábra). A reakció előnyei az
enantioszelektivitás mellett, az olcsó, a reakcióelegyből visszanyerhető katalizátor (prolin), a

24
p-metoxifenil amin védőcsoport, amely oxidatív körülmények közt könnyen eltávolítható
[List 2000].
II
(S)-4-((4-metoxifenil)amino)-4-(4-nitrofenil)bután-2-on 94 ee%
18. ábra - Direkt, prolin-katalizált Mannich-reakció
A prolin-katalizált Mannich-reakcióval nukleofil donorként nem módosított
aldehideket használva is leírtak enantioszelektív szintéziseket. A reakció speciális
aminosavak, β-laktám-antibiotikumok és szerin proteáz inhibitorok szintézisében játszhat
szerepet. A katalizátort (S)-2-metoxi-metilpirrolidinre cserélve a megfelelő anti-
diasztereomerek direkt szintézisét valósították meg [Notz és mtsai. 2003]. A prolin-mediált,
aszimmetrikus, három-komponensű Mannich-reakciót nikkomycin B totálszintézisében is
felhasználták [Hayashi és mtsai. 2005]. Jó hatásfokú enantioszelektív szintéziseket
valósítottak meg Zn- , Cu-, vagy Pd-katalizátorokkal is.
Katalizátorként pirrolidin-2-karbonsavat (prolint) és származékait használva ketonok
direkt aszimmetrikus Mannich-reakcióját írták le, ahol termékként Mannich-bázisok
képződtek igen jó enantio- és diasztereoszelektivitással, itt a főtermékek a syn-izomerek.
Pirrolidin-3-karbonsavnak és származékainak katalizátorként való alkalmazásával az anti-
izomerek sztereoszelektív szintézisét valósították meg. A sztereokontrolban a pirrolidin 2-, ill.
3-helyzetű CH-savas csoportja játszik elsőleges szerepet, ezt a 19. ábrán szemléltetjük [Zhang
és mtsai. 2008].

25
syn-III – anti-III
19. ábra – A pirrolidin-karbonsav CH-savas csoportjának szerepe a Mannich-reakció
sztereokontroljában.
Érdekes lehetőség Mannich-bázisok diasztereoszelektív „zöld” szintézise vizes
közegben. A direkt szintézis adenin katalizátor használatával, 30 %-os hidrogén-peroxid
jelenlétében, vizes-etanolos közegben, szobahőmérsékleten zajlik. A reakció egyenletét a 20.
ábra mutatja.
O
R
NH2
R
CHO+
adenin (20 mol%)/H2O2
20 oC
O HN
R
R
O HN
R
R
+
R= H, Me, OMe, Br…
syn-IV anti-IV
20. ábra – Adenin-katalizált Mannich-reakció egyenlete, a keletkező syn/anti termékekkel (IV )
A kiindulási vegyületek szubsztituensei alapvető szerepet játszanak a keletkező β-
aminoketonok diasztereoszelektivitásában. Az amin elektronvonzó szubsztituensei a syn, míg
az elektronküldő szubsztituensek az anti formának kedveznek, de maximum 20:80 arányú
szelektivitás volt tapasztalható [Goswami és Das 2009].

26
3.2.3. Mannich-reakció felhasználása bioaktív vegyületek szintézisében
3.2.3.1. α-Metilén ketonok szintézise Mannich-bázis intermediereken keresztül
Az α-metilén ketonok szintézisét Mannich-bázis intermediereken keresztül a megfelelő
ketonból, s-trioxánból és N-metilanilin-trifluoracetátból (TAMA) kiindulva írták le. A 21.
ábra egy keton reakcióját mutatja egy s-trioxánnal és TAMA reagenssel száraz aprotikus
oldószerben, ami ekvivalens egy formaldehiddel végzett aldol-kondenzációval. A reakció
során egy Mannich-bázis képződik, amiből spontán β-eliminációval keletkezik egy
irreverzibilis lépésben az α-metilén keton (V) [Gras 1978]. A módszer jelentőségét az adja,
hogy az igen reaktív α-metilén ketonok így viszonylag könnyen előállíthatók.
R
CH3
O
+O O
O
+ CF3COO H2NCH3
C6H5
TAMA
THF
vagy dioxanR
O
H
H
H
V.
21. ábra – α-Metilén ketonok szintézise
3.2.3.2. Mannich-reakció használata gyógyszervegyületek előállításában
A Mannich-reakció használata gyógyszervegyületek előállításá során is nagyon gyakori. Az
alábbiakban erre mutatok néhány konkrét példát, a teljesség igénye nélkül [Subramaniapillai
2013]. A Parkinson-kór kezelésében használt osnervan vagy a fájdalomcsillapító hatású
tramadol (22. ábra) szintézise Mannich-intermedieren keresztül valósítható meg.
22. ábra – Tramadol
A nemszteroid fájdalomcsillapító-gyulladáscsökkentő hatású ibuprofen triazol-származékának
Mannich-bázisát is előállították formaldehid és szekunder amin reagensekkel (lásd 23. ábra),

27
mely szintén erős fájdalomcsillapító-gyulladáscsökkentő hatást mutatott [Sujith és mtsai.
2009].
ibuprofen triazol-számazéka ibuprofen triazol-származékának Mannich-analóg vegyülete
23. ábra – Ibuprofen triazol-származéka és annak Mannich-származéka
Az altató hatású zolpidem szintézisét Mannich-reakcióval Sumalatha és mtsai. írták le
[2009], a reakciót a 24. ábrán szemléltetjük (a köztes lépések kihagyásával).
R1 = 4-MeC6H4 zolpidem
24. ábra – Zolpidem szintézise Mannich-reakcióval
A Mannich-származék képzés gyakran hasznos a vízoldékonyság növelésére. Az
echinocandinok közé tartozó mulundocandin in vitro nagyon jól hat különböző Candida
törzsekre, de gyenge vízoldékonysága korlátozza alkalmazhatóságát [Lal és mtasi. 2004]. A
mulundocandin Mannich-származéka (25. ábra) a kiinduális vegyülethez hasonlóan jó
antifungális hatást mutat, míg vízoldékonysága lényegesen jobb annál.

28
HO
N O
NH
OHO
H3CH
OHO
NH
OCH3
H
HN MM
H
CH3
OH
H
N
O
HN
OHO
NH
OH
OH
H
HOOH
H
H
N
N
MM = 12-metilmirisztoil 25. ábra – Mulundocandin Mannich-származéka
Egy másik példa a vízoldékonyság növelésére a limeciklin, azaz az N-lizinometil-tetraciklin
(26. ábra), ez a tetraciklinek családjába tartozó antibiotikum, mely az alapvegyületnél sokkal
jobban oldódik vízben. Súlyos acne kezelésében használják belsőleg is [Bossuyt és mtsai.
2003].
26. ábra - Limeciklin (N-lizinometil-tetraciklin)
A kedvezőbb ADME (felszívódás-megoszlás-metabolizmus-kiválasztás)
tulajdonságok kialakítását szem előtt tartva fejlesztettek tovább egy létező maláriellenes
hatású vegyületet Guantai és mtsai. [2011]. A 27. ábra mutatja a reakció egyenletét és
körülményeit. A kiindulási vegyület triazol-gyűrűjét piperazinra cserélték és egy
kalkonszármazékkal vitték Mannich-reakcióba, formaldehides-etanolos közegben. A szintén
maláriaellenes hatású termék (VI ) farmakokinetikai paraméterei kedvezőbbek, mint a
kiindulási anyagé.
HN
HN
N(CH3)2
COOH
H
OH
HCH3HOOH
OOOHOOH
NH2

29
R=H, OMe
VI
27. ábra – Optimalizált karakterű maláriaellenes hatású vegyület előállítása Mannich-
reakcióval
3.2.3.3. Mannich-reakció felhasználása természetes vegyületek szintézisében
A Mannich-reakciót természes vegyületek, pl. alkaloidok szintézisére is használják. A
reakciót a 28. ábrán szemléltetjük egy nyílméregbékából izolált alkaloid, az epibatidin
szintézisével [Hernandez és mtsai. 1995]. Az intramolekuláris Mannich-reakcióban a
karbonsav-származéknak ketocsoportot kell tartalmaznia.
epibatidin
28. ábra – Az epibatidin szintézise
Egy alkaloidszerű vegyület, pontosabban egy furazocin-származék szintézisét Chernov
és mtsai. írták le [2003]. Kiindulási anyagnak lambertiánsavat, egy, a szibériai cédrusból
(Pinus sibirica J. Mayr.) kinyerhető diterpénsavat használtak. A szintézis egyik intermedierje
az intramolekuláris ciklizációban kapott azaciklooktán-származék (29. ábra).

30
Azaciklooktán-származék Furazocin-származék
29. ábra – Intramolekuláris Mannich-reakcióval szintetizált furazocin-származék, és a köztes
Mannich-bázis azaciklooktán-származék képlete
3.2.4. Mannich-bázisok biológiai hatásai
Hosszan sorolhatnám a Mannich-vegyületek biológiai hatásait, de a számos lehetőség
közül csak két területet szeretnék jobban kiemelni, a citotoxikus és az antimikrobiális hatást.
Egyes Mannich-bázisok citotoxikus hatása évtizedek óta ismert. Dimmock és mtsai.
[1980] sztiril-ketonok és ezek Mannich-származékainak tumorellenes hatását vizsgálták
egereken, ahol is a Mannich-bázisok számottevő tumorellenes aktivitást mutattak, míg a
megfelelő sztiril-ketonok nem vagy alig. Az antitumor-hatás jól korrelált a vegyületek tiol-
depléciós képességével. Ez azt jelenti, hogy a szer a tumorsejt létfontosságú tiol-vegyületeit,
mint például a glutationt vagy a ciszteint, képes alkilálni. Az elmélet szerint a tumorsejtek
érzékenyebbek a normál sejteknél erre a hatásra, különösen a szekvenciális citotoxicitást
mutató vegyületek esetén [Pati és mtsai. 2007].
H3CN
H2C
H2C C
H2C
H3CO
H2C N
CH3
CH3
x 2HClH3C
NH2C
H2C C C
HH3CO
CH2H N
CH3
CH3
x HCl+
HSR1
H3CN
H2C
H2C C
H2C
H3CO
H2C SR1H2C C
HC
H2C
O
H2C SR1H N
CH3
CH3
x HCl +
HSR2
R2SH2C
H2C C
H2C
O
H2C SR1
HSR1, HSR2 = celluláris tiol vegyület
30. ábra - Szekvenciális citotoxicitás menete

31
A szekvenciális citotoxicitás két vagy több citotoxikus anyag egymást követő
felszabadulását jelenti. Ahogy a 30. ábrán láthatjuk, a látens tioalkilező szerek először
dezaminálódnak, azaz 1,2-eliminációban vesznek részt, metilén-ketonokká, melyek reagálnak
egy celluláris tiol-vegyülettel. Ezt követően egy addíciós és dezaminációs lépést követően
újabb tioalkilezőszer szabadul fel.
Cikloalkanonok Mannich-származékainak vizsgálata során erős citotoxikus hatású
vegyületeket (VII , VIII ) szintetizáltak, ezek közül néhányat a 31. ábra mutat [Dimmock és
mtsai. 1993]. Bisz- és mono-származékokat is előállítottak, így vizsgálva a szekvenciális
citotoxicitást. A bisz-származékok két helyen is dezaminálódhatnak, így kétszeres alkilálásra
képesek. Ezen kísérletek során a bisz-származékok bizonyultak hatékonyabbnak, de a mono-
származékok közt is találtak hatásos vegyületeket.
VII. VIII.
31. ábra – Citotoxikus hatású cikloalkanon Mannich-származékok.
Kimutatták, hogy 1-aril-3-dialkilamino-1-propanon hidrogénhalogenid-származékai
számos egér ill. humán tumoros-sejtvonalon citotoxikus hatásúak. Ebből kiindulva
Vashishtha és mtsai. [2004] számos további vegyületet előállítottak. Az 1-(4-arilfenoxi)-3-
dietilamino-1-propanon-hidroklorid és származékai (IX -X, lásd 32. ábra) mind citotoxikus
hatásúnak bizonyultak.
R= H, CH3, F,Cl, Br X=N(CH3)2 X=O, S IX. X.
32. ábra - Citotoxikus hatású Mannich-bázisok: 1-(4-arilfenoxi)-3-dietilamino-1-propanon-
származékok

32
Citotoxikus hatású a gatifloxacin Mannich-származéka (33. ábra) is. Ez erősebb
antitumor-aktivitást mutatott, mint a terápiában használt etoposid.
N
O
NBr
SO2NH
N N
N
F
H3C
O
O
OHH3CO
N
N
H3CO OCH3
33. ábra - Gatifloxacin Mannich-származéka
A fenti, potenciális tumorellenes szerek mellett klinikai használatban levő citotoxikus
hatású Mannich-származékok közé tartozik például a topotecan, ami a camptotechin
vízoldékony származéka [Subramaniapillai, 2013].
Egyes Mannich-bázisokkal konjugált sztiril-ketonok antifungális aktivitása szintén
régóta ismert. A 34. ábrán egy Candida albicans ellen hatékony vegyület látható [Dimmock
és mtsai. 1994].
34. - ábra C. albicans ellen hatékony Mannich-keton vegyület
Kalkon-származékok antifungális hatását és annak mechanizmusát vizsgálva
megállapították, hogy ebben a gomba sejtfalára gyakorolt hatás is szerepet játszhat, a kitin-
szintáz és a β-1,3-D-glukán szintáz befolyásolásán keresztül [Lopez és mtsai. 2001].
Lóránd és mtsai. [2001] telítetlen Mannich-ketonok sorozatát állították elő. A szintézis
első lépésében 2-arilidéncikloalkanon köztes termékek keletkeztek bázis-katalizált aldol
kondenzációban. A Mannich-ketonokokat a megfelelő 2-arilidéncikloalkanon, szekunder

33
amin és paraformaldehid reakciójából sósavas sóként izolálták. A klasszikus Mannich-reakció
során etanolos közegben sósav katalizátort használtak, a szekunder-amint a kompetitív aldol-
kondenzáció visszaszorítására feleslegben alkalmazva. A vegyületek citotoxicitását, Gram-
pozitív és Gram-negatív baktériumokkal szemben mutatott hatását is vizsgálták. Néhány
vegyület számottevő antibakteriális aktivitást mutatott. A hatást feltételezhetőleg a tiol-
alkilálás mellett egyéb tényezők is befolyásolják. Kondenzált vázas Mannich-ketonok
szintézisét is megvalósították. Ez a vegyületcsalád is mutatott antibakteriális hatást [Lóránd és
mtsai. 2002].
A telítetlen Mannich-ketonok antifungális hatását Candida, Saccharomyces és
Aspergillus törzsekre is vizsgálták [Kocsis és mtsai. 2009]. A penészgomba törzsek ellen nem
voltak hatásosak ezek a vegyületek, míg a Candidákkal szemben a legtöbb anyag antifungális
hatást mutatott. A Mannich-ketonok ergoszterin-, kitin- és fehérje-szintézisre, valamint a
pszeudohifa-képzésre kifejtett hatását is tesztelték. Egyes vizsgált vegyületek befolyásolhatják
a C. albicans pszeudohifa-képzését, változásokat okoztak a fehérje-összetételében, illetve
befolyásolhatják a kitin-szintáz enzimet is.
Munkám során ezekkel a telítetlen és kondenzált vázas Mannich-ketonokkal foglalkoztunk,
így ezekről később még szólunk.

34
3.3. Módszerek
3.3.1. Áramlási citometria
Az áramlási citometria (= flow citometria) egy eljárás vagy mérési módszer, amellyel
folyadékáramban önálló részecskék tulajdonságait vizsgálhatjuk. Előnye, hogy 1 µm feletti
részecskékről egyedi információkat nyerhetünk. Az áramlási citometria segítségével nemcsak
a sejtek méretéről tájékozódhatunk, hanem különböző fluoreszcens festékeket használva
speciális tulajdonságokat is vizsgálhatunk. A flow sorting technikával sejtek vagy részecskék
elkülönítése, elválasztása is lehetséges mért paraméterek alapján.
A klasszikus áramlási citométerek vázlatos technikai felépítése: fényforrás, áramlási
rendszer, detektáló rendszer, adatgyűjtés – analízis, opcionális sorting-rendszer. Az áramlási
citometriás méréseknél a minta szuszpenzió formájában tartalmazza a sejteket, melyeket
túlnyomás segítségével juttatnak el egy fejhez vagy áramlási cellához.
Az áramlási cellához kerülve a sejtek hidrodinamikai fókuszálással rendeződnek, azaz
a nyomásviszonyok megfelelő beállításával lineáris áramlás jön létre, a köpenyfolyadék és a
minta nem keverednek, hanem a segédanyag körülöleli a mintát. A fejet a sejteket tartalmazó
mintacseppek sorban egymás után hagyják el, megfelelő beállítások mellett egy
folyadékcseppbe egy sejt kerül.
Ezután a sejteket megvilágítják egy fókuszált fénysugárral, amelyet lézerrel,
lámpákkal vagy diódával generálnak, majd detektálják az egyes sejtekről érkező optikai
jeleket, úgy mint:
- előre irányuló szórás = forward scatter (FSC): a sejt méretét jellemzi
- oldalirányú szórás = side scatter (SSC): a sejt belső strukturáltságáról informál
- fluoreszcencia: a sejtek speciális jellemzőiről adhat tájékoztatást, a mért emissziós
intenzitás arányos a sejtekhez kötött fluorofór mennyiségével.
A detektorok elhelyezése függ a készülék optikai elrendezésétől, lézeres fényforrás esetén a
detektor merőlegesen helyezkedik el a megvilágító fényhez és a folyadékoszlophoz képest is.
Az oldalirányú szórás és a fluoreszcencia detektálására fotoelektron-sokszorozókat (PMT), az
előre irányuló szóráshoz fotodiódákat használnak, ezek érzékenysége elmarad a PMT-től, de
kisebbek, olcsóbbak, stabilak. A fluoreszcens jel specifikus régióinak detektálásához az
emittált fényt hullámhossza alapján szét kell osztani, erre a célra az interferencia elvén
alapuló optikai szűrőket és dikroikus tükröket használnak. A jeleket analóg-digitális átalakítók

segítségével konvertálják, az adattárolás és analízis természetesen számítógépes pr
segítségével lehetséges [Lustyik 2008].
35. ábra – Áramlási citométer vázlatrajza: lézer, optikai rendszer (gy
dikroikus tükrök), detektorok [3].
Az áramlási citométerek történetében nagy el
(szortolásra) alkalmas készülékek megjelenése
piezoelektromos kristály segítségével a cseppekre bontott folyadékáramban lev
alapján szétválaszthatók. A modern készülékek tervezésénél egyik irány a minél összetettebb,
több lézert, így több, akár 12 uta
megjelentek egészen egyszerű
készülékek is. A chip-alapú mikrofluidikai rendszerekr
Napjainkban az áramlási citomet
immunológiai, immundiagnosztikai,
van, ezeket jelen munkánknak nem célja részletezni.
Az egyedi sejtek vizsgálata azonban a mikrobiológiában is font
módszer az áramlási citometria [
eukarióta sejtek vizsgálatára használták, ma már
Baktériumok, köztük Staphylococcus aureus, Esch
kimutatását, antibiotikum-érzékenység
Vives-Rego 1998, Walberg és mtsai. 1997
35
segítségével konvertálják, az adattárolás és analízis természetesen számítógépes pr
[Lustyik 2008]. A rendszer felépítését az 35. ábra mutatja.
Áramlási citométer vázlatrajza: lézer, optikai rendszer (gyűjtő
dikroikus tükrök), detektorok [3].
Az áramlási citométerek történetében nagy előrelépés volt a sejtszeparálásra
(szortolásra) alkalmas készülékek megjelenése [Mattanovich és Borth
piezoelektromos kristály segítségével a cseppekre bontott folyadékáramban lev
alapján szétválaszthatók. A modern készülékek tervezésénél egyik irány a minél összetettebb,
több lézert, így több, akár 12 utas detektálást használó rendszerek fejlesztése
egészen egyszerű, rutindiagnosztikai feladatokra terepen is használható
mikrofluidikai rendszerekről lentebb részletesen szólunk.
ainkban az áramlási citometria egy széles körben elterjedt módszer, számtalan
immunológiai, immundiagnosztikai, biotechnológiai, rutin labor- és kísérleti felhasználása
van, ezeket jelen munkánknak nem célja részletezni.
Az egyedi sejtek vizsgálata azonban a mikrobiológiában is fontos, erre egy lehetséges
ometria [Brehm-Stecher és Johnson, 2004]. Míg kezdetben csak
eukarióta sejtek vizsgálatára használták, ma már prokarióták elemzésére is
Staphylococcus aureus, Escherichia coli, Salmonella
érzékenység meghatározását írták le a módszerrel
lberg és mtsai. 1997].
segítségével konvertálják, az adattárolás és analízis természetesen számítógépes programok
mutatja.
űjtő lencsék, filterek,
relépés volt a sejtszeparálásra
anovich és Borth, 2006]. Beépített
piezoelektromos kristály segítségével a cseppekre bontott folyadékáramban levő sejtek töltés
alapján szétválaszthatók. A modern készülékek tervezésénél egyik irány a minél összetettebb,
s detektálást használó rendszerek fejlesztése, illetve
iagnosztikai feladatokra terepen is használható
lentebb részletesen szólunk.
ria egy széles körben elterjedt módszer, számtalan
és kísérleti felhasználása
os, erre egy lehetséges
Míg kezdetben csak
elemzésére is elterjedt.
Salmonella spp., stb.
meghatározását írták le a módszerrel [Comas és

36
Változatos fluoreszcens jelölőket alkalmazva használják életképesség-vizsgálatokra,
mely tulajdonság meghatározása nem is mindig olyan egyértelmű [Davey 2011].
Alkalmazható sejtciklus-vizsgálatokban, például élesztőgombák növekedési fázisainak
követésében és modellezésében [Porro és mtsai. 2009]. Page és mtsai. [2006] polisztirol
gyöngy alapú olgionukleotid próbákkal klinikai mintákból Candida törzsek kimutatását
valósították meg. Aspergillus és Candida törzsek antimikotikumokkal, úgymint, amphotericin
B, azolok, caspofungin szembeni érzékenységének vizsgálatára sikerrel használtak áramlási
citometriát [Kirk és mtsai. 1997, Pina-Vaz és mtsai. 2005]. Az áramlási citometriás
eredmények jól korreláltak a nemzetközi standard és rutin diagnosztikai érzékenység-
vizsgálatokkal [Chaturvedi és mtsai. 2004, Favel és mtsai. 1999].
3.3.2. Chip-alapú áramlási citometria
A mikronizálási törekvés napjaink jellemző trendje. A hagyományos áramlási
citométerek összetett, ezért drágább rendszereivel szemben jelentkezett igény a kisebb
méretű, kisebb minta – és anyagigényű, olcsóbb rendszerek fejlesztésére.
A mikrofluidikai áramlási citométer fejlesztések esetében felmerülő alapvető
problémák a folyadék-áramlási rendszerek miniatürizálása, az optikai rendszerek
lekicsinyítése illetve optimalizálása a vizsgálható mintákra. További lehetőség rejlik a mikro-
sorting rendszerek kidolgozásában. Megoldandó feladatként merült fel különböző feladatok
intergálásának megoldása egy felületen. Ezért a különböző, un. lab-on-a-chip rendszerek
fejlesztése ma is zajlik, sőt ilyen megoldások már kereskedelmi forgalomban is elérhetők
[Lapsley és mtsai. 2013].
A megfelelő folyadékáramlás biztosításánál fontos, hogy például a perisztaltikus
pumpáknál gyakran tapasztalható pulzálást elkerüljük, így a csatornák méretét is figyelembe
véve a megfelelő áramlási profil alakuljon ki. A mikroflow citométerek többségénél külső,
nyomás-vezérelt pumpákkal oldották meg a folyadékáramlást, ide értve a fecskendő-alapú,
sűrített gázos vagy vákuum-pumpákat, de gravitáció-hajtott áramlást is leírtak. A nyomás-
vezérelt áramlás helyett elektrokinetikus fókuszálás is használható, ezek a módszerek az
elektroozmotikus áramlás (EOF) és/vagy az elektroforézis elvén alapulnak. Egydimenziós
fókuszálás a gyakoribb, de két-dimenziós fókuszálást is alkalmaznak már mikroflows
rendszereknél [Ateya és mtsai. 2008].
Az optikai rendszer kivitelezésére szintén számos lehetőség létezik. Mivel mind a
gerjesztést, mind a detektálást meg kell valósítania, így a pontszerű (20-60 µm) gerjesztési
sugár mellett a fényszórás mérésénél is lokalizált, pontos adatgyűjtés szükséges.

37
Fényforrásnak többnyire LED-et vagy lézert, a detektáláshoz optikai filtereket, szűrőket,
mikrolencséket használnak. Alternatív lehetőségként leírták a kizáráson-alapuló optikai
rendszerek, illetve a párhuzamosan sok szín detektálását lehetővé tevő COST (color-space-
time) adatgyűjtés használatát is. [Cho és mtsai. 2010].
A mikrosorting, azaz a sejtek szeparálása is elvégezhető már chip-méretű
rendszerekben is. Ehhez használhatnak mechanikus csapdázást, ilyen lehetőség a
mikrofilterek, pl. vér alakos elemeinek szeparálására [Chen és mtsai. 2008], vagy a sejt-
filterek (állati és baktérium sejt elválasztására) alkalmazása, de a sejtek különböző adhéziós
tulajdonságait is kiaknázhatják e célra [Andresson és Berg, 2003]. Az elektromos mező által
vezérelt szeparálás általában véve jól illeszthető a mikrofluidikai rendszerekbe, előnyei a
gyorsaság, kontrollálhatóság, változatos felhasználás. Az elválasztandó részecskék típusa
határozza meg az elektromos mező típusát. Cho és mtsai. [2009] fluoreszcencia-aktivált mikro
sejt-szorter hatékonyságát bizonyították.
A gyakorlatban számos, chip-alapú citometriás rendszert hoztak létre kísérleti céllal,
ezek közül több kereskedelmi forgalomban is elérhető, lab-on-a-chip rendszerek részeként is.
A 36. ábrán példaképp egy ilyen készülék, az Agilent 2100 Bioanalyzer áramlási citometriás
chipjének rajzát mutatom be [Preckel és mtsai. 2002]. Az ábrán kinagyítva látható a chip
csatorna-hálózata, a mintafelviteli helyek kék, míg a segédanyagként használt puffer felviteli
helye piros színű karikával van jelölve. Az elválasztás nyomásvezérelt rendszerben zajlik,
fluoreszcens detektálást alkalmaznak. Közvetlen a detektálás előtt a csatorna adott
szegmensébe fókuszálják a sejteket hidrodinamikai fókuszálással. Így a detektálási zónába
egymást követően érnek a sejtek, azaz itt is egyedi sejtekről nyerhetők információk.
36. ábra - Az Agilent áramlási citometriás chipének vázlatos rajza, kinagyítva a detektális
régió előtti terület, ahol a hidrodinamikai fókuszálás történik. [4]

38
A fentiekből is látszik, hogy a mikroflow rendszerek felhasználása egy dinamikusan
fejlődő terület. Csak néhány példa a felhasználásra: Palkova és mtsai. [2004] élesztőgombák,
emlőssejtek és gombaspórák kimutatását írták le. Alkalmasnak találták a rendszert
baktériumok vizsgálatára [Gerdts és Luedke, 2006], vagy humán sejtek fenotípus-elemzésére
[Chien és mtsai. 2003]. Alshareef és mtsai. [2013] tumorsejtek szétválasztását valósították
meg elektroforézis - alapú mikrofluidikai chipen.
3.3.3. Chip-alapú kapilláris gélelektroforézis
Napjaink leghatékonyabb elválasztási technikái az elektroforézis elvén alapulnak,
amely szerint az oldat töltéssel rendelkező molekulái elektromos erőtér hatására elmozdulnak.
A poliakrilamid és agaróz gélt alkalmazó elektroforézis technikák alapvető fontosságúak a
genomikában és proteomikában. A hagyományos gélelektroforézis egy-, illetve kétdimenziós
elválasztási rendszerek alkalmazását is lehetővé teszi.
A kapilláris elektroforézis technikákat szintén több évtizede használják, főleg
biomolekulák analízisére. A kapilláris zónaelektroforézis (CZE) a leggyakrabban használt
kapilláris elektroforetikus módszer, az oldott részecskék különböző elektroforetikus
mozgékonyságán alapszik. Ennek megfelelően az elválasztás a tömeg/töltés arány alapján
zajlik az elektrolittal töltött kapillárisokban. Fehérjék elválasztására a CZE jól alkalmazható,
de figyelembe kell venni a fehérjék és a kapilláris fala közt fellépő, nem kívánatos
adszorpciós kölcsönhatásokat. Ezek kiküszöbölésére bevont vagy módosított felületű
kapillárisokat használnak, vagy segédanyagként megfelelő puffer-adalékokat [Gáspár 2000].
Kapilláris gélelektroforézis alatt azt értjük, mikor az elektroforézis kivitelezésére
géllel vagy polimer-oldattal töltött kapillárisokat használunk. Ennél a módszernél a fehérjéket
felületaktív anyaggal kezelik a mérés előtt, a vándorlás a töltésüknek köszönhető, de
molekulaméret alapján választódnak el. A technika fehérjék mellett, DNS, illetve komplex
szénhidrátok elválasztására is alkalmas [Guttman 2003].
A mikrochip-elektroforézis a kapilláris elektroforézisből fejlődött ki. Napjaink
legmodernebb elválasztástechnikai módszerei közé tartozik, alkalmas biológiailag fontos
minták, DNS, RNS, fehérjék elválasztására. A molekulák elválasztása kisméretű üveglapba
maratott, 10-40 µm mély és 50-200 µm hosszú csatornákban történik, üvegchipek mellett ma
már műanyagchipeket is használnak. A csatornák a mikrochip felszínén levő nyílásokban
végződnek, melyek a minták és segédanyagok bejuttatására szolgálnak. A tényleges

39
mintaigény néhány nanoliternyi, a mérések időtartama néhány perc nagyságrendű. A
detektálás leggyakrabban lézer-indukált fluoreszcencia méréssel történik, de léteznek direkt
MS-kapcsolt mikrochip rendszerek is. Egyéb detektális módok, mint az elektrokémiai
detektálás vagy a kemilumineszcencia mérése kevésbé elterjedtek [Kustos és mtsai. 2007].
A rengeteg, adott kísérlethez tervezett chip-alapú elektroforézis-rendszer mellett
számos, kereskedelmi forgalomban is elérhető változata létezik. Ilyen például a Bio-Rad
ExperionTM nevű mikrofluidika-alapú elektroforézis-készüléke vagy az Agilent Bioanalyzer
rendszere [Kuschel és mtsai. 2005].
A chip alapú kapilláris gélelektroforézis rendszereket önállóan vagy lab-on-a-chip
rendszer formájában használják.

40
4. CÉLKIT ŰZÉSEK
Munkám során Candida törzsek analitikai, elválasztás-technikai módszerekkel való
vizsgálata volt a célom. Ezen igen tág témán belül az alábbi vizsgálati irányokat terveztem
megvalósítani:
• Candida sejtek vizsgálata áramlási citometriás módszerrel, elsősorban az
antimikotikum-érzékenység meghatározása (irodalmi adatok alapján).
• Különböző antimikotikumok vizsgálata, Candida albicans és non-albicans törzseken.
• Ezen eredményekre alapozva hasonló elven működő, de chip-alapú antimikotikum-
érzékenység meghatározás tervezése, mivel ilyen célú felhasználását nem találtam
ezen rendszernek.
• A hagyományosan használt antimikotikumok mellett más típusú, gombaellenes
hatással is rendelkező vegyületek, a Mannich-ketonok vizsgálata.
• Candida albicans fehérjeprofiljában a Mannich-ketonok hatására bekövetkező
minőségi és mennyiségi változások vizsgálata chip-alapú elektroforézises módszer
felhasználásával.
• Különböző típusú Mannich-ketonok vizsgálata, a fehérje összetételre gyakorolt hatás
és a vegyületek szerkezete közti összefüggések elemzése.

41
5. KÍSÉRLETI RÉSZ
5.1. Candida törzsek és tenyésztési körülmények
Kísérleteim során nemzetközi standard Candida törzseket használtam: Candida
albicans ATCC 90028, Candida tropicalis ATCC 90874, Candida glabrata 90030,
Candida krusei 30068. A gombákat Sabouraud-agar (Oxoid Ltd., Basingstoke, Hants,
England) táptalajon tenyésztve tartottam fent, 30 °C-on, hetente átoltva.
5.2. Fluoreszcens festékek
Az áramlási citometriás és sejt-chip vizsgálatoknál a gomba sejteket különböző
fluoreszcens festékekkel jelöltem. A különböző kísérletek során az alábbi festékeket
használtam:
• Propidium-jodid (Sigma, USA). 10 µg/ml-es koncentrációjú oldatát használtam, ez a
vegyület fénytől védve, 4 °C-on tárolható. Nukleinsav festék, az elölt sejtek jelölésére
alkalmas. Gerjesztési/ emissziós maximuma 535/617 nm.
• Sytox Green (Molecular Probes, Eugene, OR, USA) 5 mmol/l oldat DMSO-ban, nem
permeábilis nukleinsav festék, az elölt sejtek jelölésére alkalmas. A Sytox Green
hozzávetőleges gerjesztési/emissziós maximuma 504/523 nm.
• Syto 60 (Molecular Probes, Eugene, OR, USA) permeábilis nukleinsav festék,
egyaránt festi az élő és az elhalt sejteket, így az összsejtszám jelölésre alkalmas,
gerjesztési/emissziós maximuma 652/678 nm.
A propidium-jodid és a Sytox Green festékek szerkezeti képletét a 37. ábrán mutatom be
(a Syto 60 vegyület szerkezeti képletét/ racionális nevét a gyártó nem tette elérhetővé).
Propidium-jodid Sytox Green
37. ábra - Fluoreszcens festékek szerkezeti képlete

42
5.3. Antimikotikumok
Az áramlási citometriás és sejtchip kísérleteimhez amphotericin B-t (ld. 3.ábra) és
flukonazolt (ld. 5. ábra) használtam.
Az amphotericin B-t Fungizone® injekcióhoz való por (Bristol-Myers Squibb,
Epemon, France) formában használtam, ez porampullánként 50 mg amphotericin B mellett
41 mg nátrium-dezoxikolátot tartalmaz és 20,2 mg nátrium-foszfátot pufferként. A
kristályos amphotericin B vízben nem oldódik, a hozzáadott nátrium-dezoxikolát
segítségével intravénás használatra alkalmas kolloidális diszperziót tud képezni. A 1
mg/ml-s amphotericin B törzsoldatot desztillált vízzel mindig a kísérletek előtt közvetlenül
készítettem el.
A flukonazolt Mycosyst® 2 mg/ml infúzióhoz való oldat (Richter-Gedeon Nyrt.,
Budapest, Magyarország) formájában használtam.
Mannich-ketonok
A kísérleteim során használt Mannich-ketonokat témavezetőm, Dr. Lóránd Tamás
szintetizálta a PTE ÁOK Biokémiai és Orvosi Kémiai Intézetében. A mérésekhez
szükséges mennyiségű, por formájú szereket a felhasználásig -20 °C-on tároltuk, ezeket
közvetlen az alkalmazásuk előtt oldottam adott mennyiségű steril desztillált vízben.
Az antimikotikum-érzékenységi sejt-chip vizsgálatok során néhány telítetlen Mannich-
ketont is teszteltem, ezeket az anyagokat az 1. táblázat mutatja.
Vegyület n Ar R C. albicans MIC (µg/ml)
1 1 4’ -OCH3-C6H4 4-morfolil 3,125
2 1 2’-OCH3-C6H4 1-piperidil 6,25
3 3 fenil 1-piperidil 50
1. táblázat – Chip alapú áramlási citometriás kísérletek során tesztelt telítetlen vázas
Mannich-ketonok

43
Egy rokon vegyületcsoport, a kondenzált vázas Mannich-ketonok Candida
albicans fehérjeprofiljára gyakorolt hatását vizsgáltam. A kondenzált vázas Mannich-
ketonok szintézisének vázlatát a 38. ábrán mutatom be. Kiindulási anyagként a
megfelelő biciklusos ketont, szekunder amint és paraformaldehidet használva
etanolos-vizes közegben klasszikus sav-katalizált Mannich-reakcióval kaptuk a kívánt
termékeket. A Mannich-ketonok hidroklorid formában izolálhatók, így is használtam
ezeket.
OO
CH2
H2+
N
(CH2)nNH+
(CH2)n
Cl-
+ + CH2O
Cl-
n = 1, 2
38. ábra - Kondenzált Mannich-ketonok szintézisének általános egyenlete
A 2. táblázat sorolja fel a fehérje-vizsgálatok során általam tesztelt, kondenzált
vázas Mannich-ketonokat, néhány főbb jellemzőjükkel. Ezekhez a vizsgálatokhoz az
előkészített gomba-szuszpenziókat a kísérleti anyag 0,5xMIC illetve 2xMIC
koncentrációival kezeltük 1 órán át (30 °C, 200 rpm rázótermosztátban).

44
Vegyület n R1 R2 C. albicans MIC (µg/ml)
4 1 H 1-piperidil 3,125
5 1 H 1-pirrolidil 0,8
6 1 5-OCH3 1-piperidil 3,125
7 2 H 1-piperidil 12,5
8 2 H 1-pirrolidil 12,5
9 2 H 4-morfolil 3,125
10 2 H 2-(1,2,3,4-tetrahidro)-izokinolil
6,25
11 2 5-OCH3 1-piperidil 12,5
12 2 6-OCH3 1-pirrolidil 100
13 2 6-OCH3 4-morfolil 25
14 2 6-OC3H7 4-morfolil 25
15 2 7-OCH3 1-piperidil 6,25
16 2 7-OCH3 4-morfolil 12,5
17 3 H 1-piperidil 6,25
18
3 H 4-morfolil 12,5
2. táblázat – Fehérje-vizsgálatok során tesztelt, kondenzált vázas Mannich-vegyületek

45
5.4. Áramlási citometria és mikrofludikai áramlási citometria (sejt-chip)
5.4.1. Candida minták előkészítése áramlási citometriás és sejt-chip vizsgálatokhoz
A szükséges gombatörzseket RPMI 1640 tápfolyadékban (Sigma-Aldrich Chemie
GmbH, Steinheim, Germany) tenyésztettem. Ehhez az ionmentes vízben feloldott RPMI
1640 pH-ját MOPS-val (Sigma-Aldrich Chemie GmbH, Steinheim, Germany) 7,1-re
állítottuk be és membrán szűrőn (Millipore Millex-GP Filter Unit, Millipore Corporation,
Bedford, MA USA, 0.22 µm pórus méret) sterilre szűrtük. A gombatörzseket ebben a
tápfolyadékban szuszpendáltam, majd 2 órán át 30 °C-on inkubáltam, míg a
sejtszuszpenziók optikai denzitása 600 nm-en A=0.4-t (UV-VIS fotometer, Perkin-Elmer
GmbH, Überlingen, Germany) elérte.
Az antimikotikumokból felező hígítási sort készítettem, ez egy olyan hígítási sor, ahol
az antimikotikum koncentráció a csősorozat minden tagjánál az előző koncentráció fele.
Amphotericin B esetén az első csőben az antimikotikum koncentrációja 10 µg/ml volt,
flukonazol esetén a 64 µg/ml-es koncentrációtól indult a hígítási sor. A kiindulási
koncentrációkat a Candida törzsek MIC értékei alapján tervezett előzetes vizsgálatokkal
határoztem meg. Az előkészített gomba-szuszpenzióból 10 µl-t adtam mindegyik csőhöz.
A hígítási sor mellett kontrol csöveket is készítettem, a pozitív kontrol cső nem
tartalmazott antimikotikumot, míg a negatív kontrol cső nem tartalmazott sem
antimikotikumot, sem gombát. A csöveket 30 °C-on 24 órán át inkubáltam, a Candida
sejtek növekedését vizuálisan ellenőriztem. Minden csőből 10 µl-t Sabouraud agar
lemezekre oltottam ki, majd megfelelő inkubáció (30 °C, 48 h) után a telepképző egységek
számát (CFU) leolvastam.
A minimális gátló koncentráció (MIC) értékét úgy határoztam meg, mint az
antimikotikum azon legkisebb koncentrációja, mely képes meggátolni a gomba sejtek
növekedését. A vizsgálatokat mindig legalább háromszor ismételtem.
5.4.2. Áramlási citometria
Az áramlási citometriás mérésekhez a fentebb leírt módon előkészített, inkubált
hígítási sorokat használtam. Minden egyes csőből 350 µl-t gőzsterilizálóba tettem, 100 °C-
ra, 1 órán keresztül. Ezeket a hőkezelt mintákat és az eredeti szuszpenziókból 350 µl-ket
megfelelő fluoreszcens festékkel festettem. Propidium-jodid (PI) festéshez a mintákat
Eppendorf-centrifugában centrifugáltam, majd az eredeti térfogattal egyező mennyiségű PI
oldatba vettem fel. Sytox Green, illetve Syto 60 festékből 0,5 µl-t adtam mindegyik

46
mintához, majd ezeket 30 percig 30 °C-on inkubáltam. Az így előkészített mintákból 100
µl-t használtam fel az áramlási citometriás vizsgálatokhoz. A festékek tesztelése során
különböző koncentrációban próbáltam ki ezeket. Úgy találtam, hogy 0,1 - 0,2 % -os
töménységben alkalmazva is hatékonyan jelölnek, ezért használtam a fenti mennyiségeket.
Kísérleteimhez a Becton Dickinson FACS Calibur áramlási citométerét használtam
(Becton Dickinson, Fraklin Lakes, NJ, USA). A készülék egy 15 mW (488 nm) argonion
lézerrel és egy vörös dióda lézerrel (~635 nm) is fel van szerelve. Az adatgyűjtés log/log
módban történt, a fluoreszcencia intenzitás mellett az előre, ill. oldalra irányuló fényszórást
detektáltam (FSC ill. SSC). Küszöb paraméternek (trigger signal) az FSC -t használtam, a
detektálási küszöb érték 52 volt, E = 650 – nél.
A Sytox Green-nel jelölt sejteket FL1 tartományban (530/30 nm), a propidium-
jodiddal jelölt sejteket FL2 (585/30 nm), míg a Syto 60-val jelölteket FL3 tartományban
(661/30 nm) detektáltam. A mérésekhez és a kiértékeléshez a készülékhez elérhető Cell
QuestPro szoftvert használtam.
5.4.3. Sejt-chip – chip-alapú citometriás módszer
Az Agilent 2100 Bioanalyzer lab-on-a-chip rendszerét használtam chip-alapú áramlási
citometriás (továbbiakban sejt-chip). Ebben a rendszerben a chipen 25 x 75 µm-es
csatornákban zajlik a mérés, a sejteket vákuum segítségével áramoltatva. A folyadékáramlást
perisztaltikus pumpa vezérli, a vákuum 0-140 mbar közti, nyomás szenzorral pontosan
szabályozható. Ez biztosítja a rendszerben a stabil folyadékáramlást, a chip és a nyomást
kontrolálló egység egy speciális feltéten keresztül kapcsolódik. A készülékbe egy kék LED és
egy vörös lézer van beépítve, így fluoreszcens detektálásra képes két tartományban. A
mérésekhez a sejteket pufferben szuszpendáljuk, a mérés előtt a chip kapillárisait egy vizes
oldattal, míg az egyik csatornát egy fluoreszcens festékkel töltjük fel, a mérés során ez szolgál
referenciaként az optikai rendszer számára. A rendszer 2x106 sejt/ml-es koncentrációig
ideális.
Tehát a sejt-chipes mérések az áramlási citometria elvén alapulnak, de mivel itt csak
fluoreszcens jeleket detektálhatunk, így csak festett sejtek vizsgálhatók. A vörös lézer
gerjesztési maximuma 630 nm körül van, a detektálási maximum 680 nm, míg a kék LED
gerjesztési/detektálási maximuma ~470/525 nm. Ezek alapján chipes vizsgálatokhoz a
propidium-jodid nem használható, így itt a másik két festékkel dolgoztam. Az áramlási
citometriás méréshez előkészített, festett minták másik részét mértem sejt-chipen.

47
A chip előkészítését a felhasználási kézikönyv alapján végeztem, azaz: a chip
felszínén található megfelelő helyre (39. ábra) 10 µl feltöltő oldatot vittem fel, amivel
feltöltöttem folyadékkal a chip kapillárisait. 60 másodperc elteltével 10 µl fókuszáló
festéket, majd 30-30 µl sejtpuffert pipettáztam a megfelelő oldatfelviteli helyekre.
A sejtszuszpenziókból 200-200 µl-t 10 percig 10.000 rpm sebességgel centrifugáltam. A
felülúszót leszívtam, majd az üledéket, azaz a gombasejteket, azonos térfogatú (200 µl)
sejtpufferbe felvettem és óvatos vortexeléssel reszuszpendáltam. A mintákból 10-10 µl-t
késedelem nélkül betöltöttem a már előkészített chip megfelelő mintahelyeire. Az így
betöltött chipet az Agilent 2100 Bioanalyzer készülékbe helyeztem, és azonnal elindítottam
a mérést.
Egy chipen hat mintát mérhetünk meg, a mérés ideje körülbelül 20 perc. A méréshez a
készülékkel elérhető Agilent 2100 Expert szoftver Antibody Staining® ill. Generic®
programját használtam. A kiértékelést szintén a 2100 Expert program segítségével
végeztem.
PS – Feltöltő oldat (Priming Solution)
FD – Fókuszáló festék (Focusing Dye)
CB – Sejt puffer (Cell Buffer)
1-6 – minta helyek
39. ábra - Sejt-chip [4]
5.5. Fehérjeösszetétel-vizsgálatok
5.5.1. Mintaelőkészítés
Ezekhez a vizsgálatokhoz Candida albicans ATCC 90028 törzset használtam. Mivel a
fehérje-összetétel változásának vizsgálatához nagyobb mennyiségű gombára volt
szükségem, így itt az alábbi módszerrel készítettem elő a mintákat:
2 l Mueller-Hinton Broth (Oxoid Ltd., Basingstoke, Hants, England) húsleves táptalajt 5
egyenlő részre osztottam steril 1l-es infúziós üvegekbe. A táptalajokat ezt követően

48
legalább 1 órára (100C°) gőzsterilizálóba tettem. Kihűlés után a táptalajokhoz 20-20 ml 50
% -os frissen készített, mikrohullámú készülékben 800W-on 5 percig kezelt, steril glukóz
oldatot adtam.
Az 5-5 ml steril tápfolyadékban szuszpendált gombasejteket 30 °C-on 2 órán át
inkubáltam, míg azok optikai sűrűsége az A = 0,4 (600 nm) értéket elérte, majd
hozzáadtam a glukózos bouillon tápfolyadékhoz. A tenyészeteket 30 °C-ra
rázótermosztátba tettem. 24 óra elteltével a gombaszuszpenziók koncentrációját
ellenőriztem, szükség esetén steril tápoldattal beállítottam A= 0,4 (600 nm) értékre. A
szuszpenziókból 10 µl mintát kioltottam Sabouraud agar táptalajra, ezzel ellenőrizve a
CFU számot, és hogy nem fertőződött-e be a tenyészet.
Steril infúziós üvegekbe osztottam szét a Candida albicans tenyészeteket, majd
kezeltem a kiválasztott antifungális szerrel. Az antimikotikumokat 0,5xMIC és 2xMIC
koncentrációban használtam, 1 órás kezelési idővel, 30 °C-on, rázótermosztátban.
Antimikotikumot nem tartalmazó kontrol mintát minden esetben készítettem. Az
inkubációt követően a szuszpenziókat lecentrifugáltam.
A felülúszót eldobva a gomba sejteket fiziológiás só oldattal reszuszpendáltam, majd
újra lecentrifugáltam. Ezt a mosási lépést még egyszer ismételtem. Az így nyert, kezelt és
kezeletlen, mosott C. albicans sejteket steril üvegekben lefagyasztottam (minimum 1
éjszakán át) és további felhasználásig -20 °C-on tároltam.
A Candida sejtek fehérje összetételének vizsgálatához első lépésben ki kellett nyerni a
fehérjéket az előzetesen kezelt, fagyasztva tárolt gombasejtekből. Erre a szakirodalomban
több módszer található, mely ultrahanggal, üveggyöngyök segítségével vagy extrakciós
pufferrel oldja meg a gombasejtek feltörését [Al-Rawi és Kavanagh 1998, Lim és mtsai.
2008]. Mivel az ultrahangos módszer előzetes kísérletek során nekünk nem működött
megfelelő hatásfokkal, más módszert választottam.
A fagyasztott gombasejtekből 2 grammnyit dörzsmozsárba mértem, kevés folyékony
nitrogénnel összetörtem, 2 ml extrakciós puffert mértem minden mintára, majd jégen 45
percig inkubáltam ezeket. Az extrakciós puffer összetétele: 0,37 M tris-HCl (pH=7,5),
1mM EDTA, 5mM 2-merkapto-etanol, 1% Triton X-100, melyhez közvetlen felhasználás
előtt adtam még 1mM fenil-metil-szulfonil-fluoridot (PMSF; M=174,19 g/mol). Inkubálás
után a fel nem tört sejteket centrifugálással eltávolítottam, a kinyert fehérjéket tartalmazó
felülúszót vizsgáltam tovább.

49
5.5.2. Chip-alapú elektroforézis
Méréseimhez itt is az Agilent 2100 Bioanalyzer rendszerét használtam, a Protein 80
ill. Protein 230 reagens készletekkel. A rendszer működése a kapilláris gélelektroforézis
elvén alapul. A 10 µm mély és 50 µm széles kapillárisok egy üveg chipbe vannak maratva.
Az előkészítés első lépéseként ezeket a kapillárisokat nyomás alkalmazásával egy
polimer oldattal töltjük fel. A polimer oldat egy nem keresztkötött poliakrilamid
származék, mely lecsökkenti az elektroozmootikus áramlást (EOF-t), ezért használhatunk
nem bevont kapillárisokat. A polimer oldathoz előzetesen hozzáadunk egy fluoreszcens
festéket, ez fogja majd jelölni a mintában található fehérjéket, tehát a jelölés a chipen
történik.
A mérés során használunk még egy un. festékmentesítő (destaining) oldatot is, ez
valójában festék-mentes polimer oldat, mely a detektálás előtt kihigítja a mintát. A mérés
előtt a fehérje-mintákhoz denaturáló oldatot adunk, ez egy nátrium-dodecil-szulfátot (SDS)
is tartalmazó gyári puffer oldat, melyhez még merkaptoetanolt (én ezt használtam) vagy
ditiotreitet (DTT) adunk redukálószerként. A denaturáló oldat ezeken kívül tartalmaz egy
standard fehérjét, ezt belső standardként használjuk a kiértékelés során, ez lesz a felső
marker, alsó markernek az SDS-festék-komplex jelét használja a program.
Külső standardként egy fehérjesorozatot használunk (ez is a reagenskészlet része),
mely az alsó és felső markeren kívül 6, ill. 7 fehérjét tartalmaz. Ezek mérete a Protein 80-
as készletnél: 3,5; 6,5; 15; 28; 46; 63 kDa, a Protein 230-as változatnál: 7; 15; 28; 46; 63;
95; 150 kDa. A felső marker 90 ill. 240 kDa. Ezeket a sorozatokat hívják gélképük alapján
„létrának” is.
Közvetlen mérés előtt a vizsgált minták 4 µl-éhez 2 µl denaturáló oldatot adtam, és 5
percig 100 °C-os termosztátba tettem az Eppendorf-csöveket. Kihűlés és néhány
másodperces centrifugálás után a létrát és a mintákat hígítottam 84 µl ill. híg minták esetén
14 µl ionmentes desztillált vízzel.
A chipet a gyári leírásnak megfelelően feltöltöttem a polimer oldattal, majd a
megfelelő minta-felvivő helyekre felvittem a segédoldatokat és a mintákat (lásd 40. ábra),
és haladéktalanul elindítottam a mérést. Minden wellbe egy-egy elektród merül, a minták
injektálása feszültség hatására történik, az elválasztás a szeparáló csatornában megy végbe.
A detektálási pont előtt történik az SDS kihígítása, ennek következtében csökken a
háttérzaj és egy nagyságrenddel megnő a minta csúcsmagassága. A detektálás lézer
indukált fluoreszcens detektorral történik. Egy chipen 10 minta futtatható, a mérés

50
időigénye 25 perc, az alkalmazott programtól függően 5-80, illetve 15-230 kDa méretű
fehérjéket választhatunk el.
G – polimer oldat
DS – festékmentesítő oldat
1-10 - minták
létra – külső standard-sor
40. ábra - Protein chip vázlatos felépítése [4].
A fehérjemintákat hagyományos akrilamid gélelektroforézissel is elválasztottuk. A
gélekből a PTE ÁOK Biokémiai és Orvosi Kémiai Intézetében tömegspektrometriás
vizsgálatokat végeztünk, azaz:
a gélelektroforézist követően a fehérjéket tartalmazó gél-darabkákat kivágtuk és
NH4HCO3 puffer jelenlétében 5 ng/µl koncentrációjú tripszin-oldattal megemésztettük. A
fehérjék triptikus emésztményének 1–1 µl-ét Bruker rozsdamentes acél mintatartó tálcára
cseppentettük. A vizsgálatok során mátrixként α–ciano–4–hidroxi–fahéjsav (CHCA)
telített 0,1%-os trifluor-ecetsav (TFA) – acetonitril (2/1 V/V) oldatát alkalmaztuk. A
minták beszáradása után az elemzéseket Bruker Autoflex II típusú MALDI TOF/TOF
tömegspektrométerrel reflektordetektálási módban végeztük el. Az ionizáláshoz 337 nm-es
nitrogén lézert alkalmaztunk, a vizsgálatok során 1000 lövés tömegspektrumát
összesítettük, a lézer frekvenciája 50 Hz volt. A tömegspektrumokat pozitív ionizációs
módban 800 és 5000 m/z tartományban regisztráltuk a gyorsító feszültség 20,0 kV, a
késleltetési idő 120 ns volt. A fehérjék azonosítása PMF és MS/MS adatbázis kereséssel
történt, amely során Mascot 2.2 kereső szoftvert és NCBInr, Swissprot és MSDB
adatbázisokat használtunk.

51
6. EREDMÉNYEK
6.1. Áramlási citometria és sejt chip
Az áramlási citometriát régóta használják élesztő gombák, Candida törzsek vizsgálatára.
A sejt-chip rendszerrel azonban kevés gomba-vizsgálatot végeztek eddig. Kísérleteim során
áramlási citometriás eredményekből kiindulva terveztem meg a chipes vizsgálatokat, ezért az
eredményeket is így mutatom be.
6.1.1. Candida sejtek detektálása, élő és elölt sejtek megkülönböztetése
Az élő és elölt gombasejtek megkülönböztetésére propidum-jodid (PI), Sytox Green és
Syto 60 nukleinsav-festékeket használtam. A PI, illetve a Sytox Green festék az elölt sejteket
jelölik, mivel az élő sejtek membránján nem jutnak át. A Syto 60 az élő és elölt sejteket is
megfesti.
A 41. ábra antimikotikummal nem kezelt, élő és hővel elölt Candida albicans minták
áramlási citometriás elemzéssel nyert képét mutatja. Ezen a különböző tartományban
detektált fluoreszcenciát az oldalirányú szórás (SSC) függvényében ábrázolom, az ábrán egy
pont egy sejtet jelöl. Az egyes ábrákon látható régiókat a megfelelő kontrol-minták
eredményei alapján jelöltem ki.
A 41. a, c élő, míg a b,d ábra hővel elölt minták eredményeit mutatja, Sytox Green
(a,b), illetve Syto 60 (c,d) festékkel jelölve. Sytox Green festés mellett az élő sejtek a jobb
alsó kvadrátban (41. ábra: a), míg az elpuszult sejtek a jobbfelső kvadrátba találhatók (41.
ábra: b). Az esetleges sejttörmelékek a bal alsó tartományban jelennek meg. Sytox Green és
PI (az eredményeket nem ábrázoltam) festéssel az élő és elölt sejtek egyértelműen
elkülöníthetők. A Syto 60 permeábilis nukleinsav festék, így az élő (41. c) és elölt (41. d)
sejteket egyformán jelöli.
Az élő és elölt C. albicans mintákat sejt-chipen is megmértem, a készülékkel
kompatibilis Sytox Green és Syto 60 fluoreszcens festékeket használva. Az eredményeket a
42. és 43. ábrán jelenítem meg, „dot plot”, illetve hisztogram-ábrázolásban. A dot plot (42.
ábra) a két fluoreszcens tartományt jeleníti meg, az ábrán egy pont egy detektált eseményt,
azaz egy sejtet jelöl. Ezeket az eseményeket mutatja a hisztogram (43.ábra) is, a
fluoreszcencia (kék vagy vörös tartományú) függvényében

52
41. ábra – Candida albicans áramlási citometriás képe. Az a, c élő, míg a b,d ábra hővel
elölt sejteket mutat, Sytox Green (a,b) és Syto 60 (c,d) festékkel jelölve, a különböző
tartományban detektált fluoreszcenciát az SSC függvényében ábrázolva.

53
42. ábra – Candida albicans sejt-chip vizsgálatokkal nyert eredményei, a mért
fluoreszcenciát dot plot ábrán bemutatva. Az a, c ábra élő, a b, d ábra hővel elölt sejteket mutat, Sytox Green (a,b) illetve Syto 60 (c,d) festéssel jelölve.

54
A 42. a, c ábra élő, a b, d ábra hővel elölt sejteket mutat, Sytox Green (a,b) illetve
Syto 60 (c,d) festéssel jelölve. Az elölt sejtek Sytox Grenn-nel festve a készülék „kék”
fluoreszcens detektálási régiójában (525+/-30 nm) észlelhetők. A Syto 60-nal jelölt, élő és
elölt sejtek a vörös régióban láthatók (680+/-30 nm).
A 43. a hisztogram élő, míg a b elölt, Sytox Green-nel jelölt sejteket ábrázol, az észlelt
események (sejtek) számát a fluoreszcencia függvényében mutatva. Az élő és elölt sejtek
egyértelműen elkülöníthetők a mért fluoreszcencia-értékek alapján.
43. ábra – Candida albicans sejt-chip vizsgálati eredményei, Sytox Green festéssel. Az a hisztogram élő, míg a b elölt sejteket ábrázol, az észlelt események (sejtek) számát a fluoreszcencia függvényében mutatva.

55
6.1.2. Candida törzsek megkülönböztetése
Az általam vizsgált Candida albicans, C. tropicalis, C.krusei és C. glabrata törzsek
sejtjeinek a mérete, felépítése meglehetősen hasonló, így áramlási citometriásan is hasonló
képet mutatnak. A 44. ábra ezen törzsek hagyományos áramlási citometriával nyert képét
mutatja, az előre és oldalra irányuló szórást (FSC/SSC) ábrázolva. Az ábrán a gombasejtek a
körülkerített régióban jelennek meg.
44. ábra - C. albicans (a), C. tropicalis (b), C. krusei (c) és C. glabrata (d) törzsek áramlási citometriás képe, az előre ill. oldalra mutató fényszórás értékeket (FSC, ill. SSC) ábrázolva.
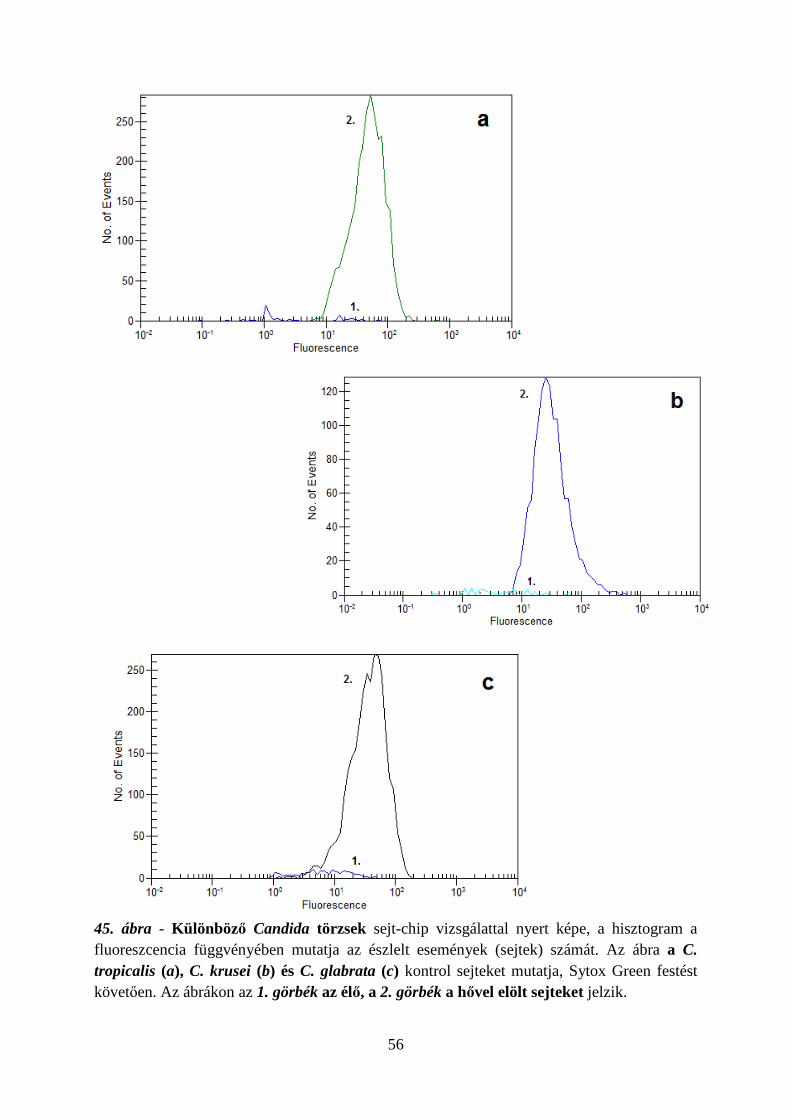
56
45. ábra - Különböző Candida törzsek sejt-chip vizsgálattal nyert képe, a hisztogram a fluoreszcencia függvényében mutatja az észlelt események (sejtek) számát. Az ábra a C. tropicalis (a), C. krusei (b) és C. glabrata (c) kontrol sejteket mutatja, Sytox Green festést követően. Az ábrákon az 1. görbék az élő, a 2. görbék a hővel elölt sejteket jelzik.

57
A különböző Candida törzseket a sejt-chip rendszerben is megmértem, a Sytox Green
festést követő a eredményeket a 45. ábrán prezentálom. A 45. a ábra a C. tropicalis, a 45. b a
C. krusei, míg a 45. c a C. glabrata élő (1.görbe) és elölt (2. görbe) kontrol sejteket mutatja.
Bár a különböző törzsek egymástól nem voltak elkülöníthetők, az élő és elölt sejtek
megkülönböztetése minden esetben egyértelmű volt.
6.1.3. Antimikotikum-érzékenység vizsgálatok
6.1.3.1. Klasszikus antifungális szerek
Az élő/elölt sejtek megkülönböztetésére alapozva terveztem meg a Candidák
antimikotikum-érzékenységének vizsgálatát. Elsőként a C. albicans törzs amphotericin B- és
flukonazol-érzékenységet vizsgáltam. Ezekhez a vizsgálatokhoz fluoreszcens jelölőként Sytox
Green festéket használtam, a továbbiakban, ha külön nem jelölöm, Sytox Green-nel festett
sejtekről beszélek.
A 46. és 47. ábrán C. albicans sejtek antimikotikum-hígítási sorainak áramlási
citometriás eredményeit jelenítem meg. Ahogy korábban említettem a hígítási sorok egyes
csöveiből vett minták egyik részét közvetlen festés után mértem. A minták másik részét a
festés előtt hőkezelésnek vetettem alá, majd így vizsgáltam tovább.
A 46. ábrán az amphotericin B hígítási sor hőkezelt mintáinak eredményeit mutatom
be a fluoreszcenciát az oldalirányú szórás függvényében megjelenítve. Az amphotericin B
koncentráció a 46. a ábrán 0.625 µg/ml, a b ábrán 0.312 µg/ml, míg c ábránál 0.156 µg/ml.
A 47. ábra a flukonazol kezelt minták áramlási citometriás eredményeit mutatja, itt
szintén a hőkezelt minták diagramjait láthatjuk. A flukonazol koncentrációja a 47. a ábrán 1
µg/ml, a b ábrán 0.5 µg/ml, míg a c ábránál 0.25 µg/ml.
A 48. és 49. ábrákon a párhuzamos sejt-chip vizsgálatok eredményeit mutatjuk be. Itt is
a hőkezelt minták eredményeit látjuk, Sytox Green festést követően, de az Agilent
Bioanalyzer chipes áramlási rendszerén mérve.
A 48. ábra az amphotericin B-vel kezelt minták sejt-chipes eredményeit demonstrálja,
az amphotericin B koncentrációja: a - 0.625 µg/ml, b - 0.312 µg/ml, c - 0.156 µg/ml.
A 49. ábra a flukonazollal kezelt minták eredményeit mutatja, a flukonazol
koncentrációja: a - 1µg/ml, b - 0.5 µg/ml, c - 0.25 µg/ml.

58
46. ábra – C. albicans amphotericin B hígítási sor áramlási citometriás képe, Sytox Green festéssel, hőkezelt minták, a fluoreszcenciát az SSC függvényében megjelenítve. Az amphotericin B koncentráció az a ábrán 0.625 µg/ml, b ábrán 0.312 µg/ml, míg c ábránál 0.156 µg/ml.

59
47. ábra – C. albicans flukonazol-hígítási sor áramlási citometriás képe, Sytox Green festéssel, hőkezelt minták, a fluoreszcenciát SSC függvényében megjelenítve. A flukonazol koncentráció az a ábrán 1 µg/ml, b ábrán 0,5 µg/ml, míg c ábránál 0,25 µg/ml.

60
48. ábra – C. albicans amphotericin B-hígítási sor sejtchip eredményei, hőkezelt minták,
Sytox Green jelöléssel. Az amphotericin B koncentrációja: a 0.625 µg/ml, b 0.312 µg/ml, c
0.156 µg/ml.
49. ábra – C. albicans flukonazol-hígítási sor sejtchip eredményei, hőkezelt minták, Sytox
Green jelöléssel. A flukonazol koncentrációja a 1µg/ml, b 0.5 µg/ml, c 0.25 µg/ml.

61
Non-albicans Candida törzsek amphotericin B- és flukonazol-érzékenységi vizsgálatát
is elvégeztem. Az áramlási citometriás és sejt-chip vizsgálatokat mindenben a C. albicans
törzsnél alkalmazott módon végeztem, a hígítási sorok kiindulási koncentrációját előzetes
MIC meghatározások alapján állapítottam meg. Az áramlási citometriával sikerült mindhárom
törzsnél meghatározni a MIC értéket (ld. 3. táblázat) amphotericin B esetében. Flukonazolra,
az előzetes várakozásainknak megfelelően, gyakorlatilag nem voltak érzékenyek ezek a
törzsek.
A non-albicans Candida törzsek áramlási citometriás eredményeit az 50. ábrán
demonstrálom, a C. glabrata amphotericin B-hígítási sorának különböző koncentrációjú,
hőkezelt mintáinak bemutatásával. Az amphotericin B koncentrációja az 50. a ábrán 1, 25
µg/ml, a b ábrán 0,625 µg/ml, míg a c ábrán 0,312 µg/ml.
Ezt követően a mikrofluidikai módszert teszteltem ugyan erre a célra. Az 51. ábrán a 3
különböző Candida törzs amphotericin B érzékenységi vizsgálatának sejt-chip
eredményét mutatom be. Az 51. a ábra C. tropicalis, a b ábra C. krusei, míg a c ábra C.
glabrata eredményeit demonstrálja. Az 51. ábra is hőkezelt, Sytox Green-nel festett minták
eredményeit mutatja be. Az amphotericin B koncentrációja az ábrákon szereplő számozott
görbéknél részletesen:
- C. tropicalis (a): 1. 0,312 µg/ml; 2. 0,156 µg/ml; 3. 0,078 µg/ml
- C. krusei (b): 1. 0,312 µg/ml; 2. 0,156 µg/ml; 3. 0,078 µg/ml
- C. glabrata (c): 1. 1,25 µg/ml; 2. 0,625 µg/ml; 3. 0, 312 µg/ml.
A flukonazol-érzékenység vizsgálatok során itt is egyértelműen kiderült, hogy ez a szer
nem hatásos ezen törzsek ellen. A gombasejtek a kontrolhoz hasonló eredményeket adtak,
mindegyik mintában sok, erősen fluoreszkáló eseményt mértem (az adatokat nem mutatom be
külön).

62
50. ábra – C. glabrata amphotericin B-hígítási sor áramlási citometriás képe, Sytox Green festéssel, hőkezelt minták, a fluoreszcenciát SSC függvényében megjelenítve. Az amphotericin B koncentráció az a ábrán 1,25 µg/ml, b ábrán 0,625 µg/ml, míg c ábránál 0,312 µg/ml.

63
51. ábra – C. tropicalis (a), C. krusei (b), C. glabrata (c) amphotericin B-hígítási sorának sejt-chip vizsgálati eredményei (hőkezelt minták, Sytox Green jelöléssel). Az amphotericin B koncentrációja: a és b ábra: 1. 0,312 µg/ml; 2. 0,156 µg/ml; 3. 0,078 µg/ml. c ábra: 1. 1,25 µg/ml; 2. 0,625 µg/ml; 3. 0,312 µg/ml.

64
A különböző Candida törzsek 3 módszerrel meghatározott MIC értékeit (µg/ml) a
3. táblázatban foglalom össze.
Amphotericin B Flukonazol Csőhígítás FC Sejt-chip Csőhígítás FC Sejt-chip
C. albicans ATCC 90028 0,312
0,312-0,625
0,312-0,625
1 1 0.5
C. glabrata ATCC 90030 0,625 0,625 0,625 >64 >64 >32
C. krusei ATCC 30068 0,312 0,312 0,156 >64 >64 >64
C. tropicalis ATCC 90874 0,156 0,156 0,156 >64 >64 >64
3. táblázat - Különböző Candida törzsek csőhígítással, áramlási citometriával és sejt-chip módszerrel meghatározott MIC értékei (µg/ml).

65
6.1.3.2. Mannich-ketonok
Néhány, előzetesen gombaellenes hatást mutató telítetlen Mannich-keton sejtchipes
antimikotikum-érzékenységi vizsgálatát is megkíséreltem, a vizsgált anyagokat a 1. táblázat
mutatja. Ezeknél a vizsgálatoknál párhuzamosan Sytox Green és Syto festést is használtam a
C. albicans sejtek jelölésére, a többi vizsgálati körülmény megegyezett az eddigiekkel.
A két festékes vizsgálat, ezen belül is az 1 vegyület eredményeit az 52. ábrán
demonstrálom. Az 52. a ábra Syto 60, míg a b ábra Sytox Green festést követően ábrázolja a
mintákat. Az 1 vegyület koncentrációja az egyes görbéknél: 1. 3,125 µg/ml, 2. 1,56 µg/ml, 3.
0,78 µg/ml.
A vizsgált telítetlen Mannich-ketonok sejt-chip vizsgálattal, és csőhígítással
meghatározott MIC értékeit, az irodalmi adatokkal [*Kocsis és mtsai. 2009] összevetve a 4.
táblázatban mutatjuk be.
4. táblázat – Telítetlen Mannich-vegyületek különböző módszerekkel meghatározott MIC
értékei C. albicans ATCC 90028 törzsre. *[Kocsis és mtsai. 2009]
Vegyület Irodalmi adat (µg/ml)*
Csőhígítás (µg/ml)
Sejt-chip (µg/ml)
1 3,125 3,125 3,125
2 6,25 6,25 6,25
3 50 100 100

66
52. ábra – 1 Mannich-keton MIC meghatározása C. albicansra sejt-chippel, Syto
60 (a ábra) és Sytox Green (b) festéssel. Az 1 vegyület koncentrációja: 1. 3,125 µg/ml, 2.
1,56 µg/ml, 3. 0,78 µg/ml.
1

67
6.2. Mannich-ketonok hatásának vizsgálata Candida albicans fehérje-profiljára
Méréseim során az 2. táblázatban felsorolt 15 kondenzált vázas Mannich-keton hatását
vizsgáltam C. albicans fehérje-profiljára. A Mannich-ketonokat két különböző
koncentrációban alkalmaztam, 1 órás kezelési idővel. Párhuzamosan minden anyaghoz
készítettem antimikotikum-mentes kontrolt. A tenyésztést, kezelést és a méréseket többször
ismételtem.
A fehérje mintákat, a szokványos poliakrilamid-gélelektroforézis mellett, a kapilláris
gélelektroforézis elvén működő chip-alapú rendszerrel teszteltem. Az elválasztás itt méret
alapján történik, köszönhetően a felhasznált redukálószernek és az SDS-nek. A rendszerben
fluoreszcens detektálást alkalmazunk, a kapott elektroferogramokon a fluoreszcenciát a
migrációs idő függvényében ábrázoltam. Az eredmények kiértékelését a készülékhez elérhető
2100 Expert programmal végeztem. Az alkalmazott program segítségével a migrációs időből
számítva a fehérjék méretét (kDa) közvetlen meghatározhatjuk. A méret meghatározást és a
mérések összehasonlítását külső és belső standardok használata is segíti. A kvalitatív
meghatározás mellett szemikvantitatív analízisre is alkalmas a mérés, ehhez a felső marker
koncentrációját használja standardnak a program.
A fehérje mintákat a Protein 230 és Protein 80 elválasztási protokollokkal vizsgáltam.
Ezek olyan futtatási programok, melyekkel 15-230 kD, illetve 5-80 kDa mérettartományba
eső fehérjéket választhatunk el.
A vizsgált fehérjemintáknál jelentősebb változásokat a 30 - 90 kDa közötti régióban
detektáltam, ezért a Protein 80 protokol szerint folytattam a további méréseket. Példaképp az
53. és 54. ábrán a 11 vegyülettel kezelt minta elektroferogramját mutatom be. A kontrol
fehérje minta elektroferogramja zöld, a 0,5xMIC koncentrációval kezelt mintát kék, a
2xMIC mintát piros szín jelöli. A két marker belső, gyári standard (1,6 kDa és 95 kDa),
meghatározzák a méret-tartományt, melyet vizsgálhatunk (53. ábra). A 54. ábrán ezen
elektroferogram kinagyított részlete látható. Az ábrán jelöltem azon csúcsok számított
molekulatömegét, melyekre valószínűleg hatást gyakorolt a vizsgált vegyület.
Az 55. ábrán az 5, 13 és 15 Mannich-ketonok fehérje-chip méréssel nyert
eredményeit mutatom be, kiemelve a régiót, ahol a fehérjemennyiség-változás jellemző. Az
ábra a kontrol mintákat (kék színnel) és az adott Mannich-keton 2xMIC koncentrációjával
kezelt minták eredményeit (piros szín) demonstrálja.

68
53. ábra – C. albicans 11 Mannich-ketonnal kezelt minta elektroferogramja chip-alapú
gélelektroforézissel mérve (a keretes rész nagyítva a 49.ábrán)
54. ábra - 11 Mannich-ketonnal kezelt minta elektroferogramjának részlete. Zöld szín:
kontrol, kék: 0,5xMIC, piros: 2xMIC.
O N
OCH3
*

69
55. ábra – C.albicans fehérjevizsgálati eredményei. A kék elektroferogram a kontrol, míg a piros az 5, 13 ill. 15 Mannich-keton 2xMIC koncentrációjával kezelt minták eredményeit mutatja.
5
13
15

70
7. EREDMÉNYEK MEGVITATÁSA
7.1. Áramlási citometriás és sejt-chipes életképesség-vizsgálatok
Áramlási citometriás kísérleteim során Candida törzsek antimikoitkum-érzékenységének
meghatározásával foglalkoztam. Elsőként hagyományos áramlási citometriás vizsgálatokat
végeztem, irodalmi adatokra alapozva. Majd ezeket az eredményeket felhasználva a Candida
törzsek vizsgálatára az áramlási citometria elvén alapuló chip-alapú rendszert (sejt-chip)
próbáltam ki.
Először egy standard C. albicans törzs élő, és hőkezeléssel elölt tenyészeteit vizsgáltam.
Áramlási citometriával nem csak fluoreszcensen jelölt, hanem festetlen sejtek is vizsgálhatók.
Fluoreszcens jelölők közül PI, Sytox Green és Syto 60 festéket használtam. A Syto 60 mind
az élő, mind az elölt sejteket jelöli, így ezek elválasztására nem alkalmas, összsejtszám
vizsgálatra használható. A PI és Sytox Green csak az elölt sejteket festi meg, mivel az élő sejt
membránján nem jut át. Mindkét festékkel megkülönböztethetők voltak az élő, ill. elölt
minták.
A sejt-chipen csak fluoreszcens jeleket detektálunk, így itt csak festett sejtek
vizsgálhatók. Az általam tesztelt festékek közül a PI nem kompatibilis a rendszerrel, ezért
Syto 60 és Sytox Green festéket használtam a mérésekhez. A Syto 60 itt is az összes sejt
jelölésére volt jó, Sytox Green festékkel sikerült elkülöníteni az élő, illetve az elölt
gombasejteket.
Ezt követően különböző Candida törzseket vizsgáltam azonos módon. Mindkét módszer
esetében, az összes törzsnél egyértelműen elkülöníthetőek voltak az élő és elölt sejteket
tartalmazó tenyészetek. Ugyanakkor a fentebb felsorolt festékekkel jelölve a különböző
törzsek egyik módszerrel sem voltak megkülönböztethetők.
7.2. Áramlási citometriás és sejt-chipes antimikotikum-érzékenység meghatározás
7.2.1. Egy fluoreszcens festékes vizsgálatok
Az antimikotikum-érzékenységi vizsgálatokat az élő/elölt sejtek megkülönböztetésére
alapozva terveztem meg és végeztem el. Ezeknél a vizsgálatoknál a célom annak
meghatározása volt, hogy egy adott törzs mennyire érzékeny az adott antimikotikumra. Ennek
jellemzésére a MIC értéket használtam, mely az a legkisebb koncentrációja az antifungális
szernek, mely képes gátolni a gomba növekedését.

71
A hagyományos áramlási citometriával különböző antibiotikumokra és antifungális
szerekre is leírtak érzékenységi vizsgálatokat [Mason és mtsai. 1994, Wenisch és mtsai. 1997,
2001].
Ezeknél a MIC meghatározására többféle módszert ajánlanak, például:
- definiálják úgy, mint az antimikotikumnak az a legalacsonyabb koncentrációja, mely 50%-
os emelkedést produkál az átlag fluoreszcencia értékben a kontrolhoz képest [Ramani és
Chaturvedi 2000].
- más megközelítésben a MIC az antifungális szer azon koncentrációja, mely a sejtek
szignifikáns (50%) elmozdulását okozza az élő sejtek régiójából vagy szignifikáns
csökkenést okoz az 1 másodperc alatt mért sejtszámban [Mitchell és mtsai. 2005].
Jelen munkámban a MIC értéket az élő és elhalt sejtek arányából határoztam meg.
Az antimikotikum-érzékenységi vizsgálatokhoz Sytox Green festéket használtam
fluoreszcens jelölőnek. Az antimikotikum-hígítási sor minden egyes koncentrációjánál a
minta felét megfestettem, a minták másik felét festés előtt hőkezelésnek vetettem alá, majd
mindegyik mintát azonos módon megmértem. Így meg tudtam határozni az eredeti mintákban
levő elpusztult sejtek mennyiségét, a hővel kezelt mintákból pedig (mivel itt az összes
gombasejtet elöltem) megkaptam az össz-sejtszámot az adott antimikotikum-koncentrációhoz.
Az élő sejtek mennyisége ezek különbségéből számítható.
Ez alapján MIC értéknek azt a koncentrációt határoztam meg, ahol az élő sejtek aránya
szignifikánsan (min.50%) csökkent az antimikotikummal nem kezelt, pozitív kontrolhoz
képest. Ezeket az eredményeket a 46. és 47. ábrákon szemléltettem. Az antimikotikum
nagyobb koncentrációjánál a gombák növekedése akadályozott, a mintában viszonylag kevés
sejt látható, azok nagy része nem élő. MIC alatti koncentrációnál a gombák növekedése nem
akadályozott, a mintákban sok sejt található, ezek jó része élő sejt, Sytox Green festésnél ezért
láthatók hőkezelés után. A MIC érték az a koncentráció, amelynél a váltás történik.
Az áramlási citometriás eredményekre alapozva a módszert a mikrofluidikai, chipes
rendszerre is kiterjesztettem. Ezt korábban, ilyen célra nem használták. A sejt-chipes mérés
előnye a minimális mintaigény, egyszerűség, automatizált mérés, mely egy diagnosztikai
módszernél hasznos lehet. A MIC meghatározás az áramlási citometriás kiértékelésnek
megfelelően történt.

72
A hagyományos és a mikrofludikai áramlási citometriával 4 különböző, nemzetközi
standard Candida törzs amphotericin B-vel és flukonazollal szembeni érzékenységét
vizsgáltam meg. A MIC értékeket minden esetben sikerült meghatározni, az így nyert
eredmények jól korreláltak egymással és a standard csőhígításos módszer eredményeivel (ld.
3. táblázat).
Az amphotericin B-, illetve a flukonazol-érzékenység vizsgálata során (48., ill. 49. ábra)
a sejt-chipes vizsgálatok egy érdekes eredményt is szolgáltattak. Az amphotericin B
magasabb koncentrációinál csupán néhány sejtet észleltem, alacsony fluoreszcenciával. A
flukonazol esetében még a szer magas koncentrációi mellett is detektálhattam néhány
fluoreszcens sejtet. Itt a MIC-et jelző váltás sem volt olyan éles, mint az amphotericin B
esetében, bár a flukonazol-érzékenység is egyértelműen meghatározható volt. A különbség
egyik lehetséges oka, hogy az amphotericin B fungicid szer, azaz elpusztítja a sejteket, míg a
flukonazol fungisztatikus hatású, a gombasejtek növekedését gátolja.
7.2.2. Két fluoreszcens festékes vizsgálatok
A két festékes atimikotikum-érzékenységi vizsgálatoknál Sytox Green mellett Syto 60
festést is használtam. Néhány, korábban antifungális hatást mutató telítetlen Mannich-keton
érzékenységi vizsgálatát végeztem el.
Ez a módszer mutatott némi előnyt az egy festék használatával szemben, de nem felelt
meg teljesen a várakozásaimnak. A két festék használata leegyszerűsítette az előkészítés
folyamatát, azaz a hőkezeléses lépést, az elölt kontrol készítés kivételével, el tudtuk hagyni.
Sikerült észlelni a MIC értékeket, ezek az eredmények megfeleltek az irodalmi adat alapján
vártnak, de a MIC érték pontos meghatározása ebben az esetben jóval nehezebb.
Nem sikerült megvalósítani, hogy a sejteket egyszerre jelöljem a két festékkel, habár
számos ilyen előzetes kísérletet végeztem. Ennek oka valószínűleg az, hogy mindkét festék a
nukleinsavakhoz kapcsolódik, és a Sytox festéket részben leszorítja a hozzáadott Syto. Így
minden mintát kettéosztottam és külön-külön mértem. A Syto festék használata miatt egy
desztillált vizes mosási lépést is be kellett iktatni, hogy a háttérzajt csökkentsem. Ez növelte a
módszer anyag és időigényét, ezzel együtt a költségeit is, ezért ezeket a vizsgálatokat nem
folytattam tovább.

73
7.3. Fehérje-vizsgálatok
A fehérje vizsgálatok során azt vizsgáltam milyen változást okoz 1 órás Mannich-keton
kezelés a C. albicans fehérjeprofiljában. A Mannich-ketonokat 0,5xMIC és 2xMIC
koncentrációkban alkalmaztam. Korábbi hasonló vizsgálatok kimutatták, hogy a telítetlen
vázas Mannich-ketonok közt van, amely, hasonlóan a flukonazolhoz, jelentős változásokat
képes indukálni [Kustos és mtsi. 2006]. Ezekre a vizsgálatokra alapozva én a kondenzált
vázas Mannich-ketonok vizsgálatára fókuszáltam.
A kondenzált Mannich-ketonok vizsgálata alapján kijelenthető, hogy egyes vegyületek
hatással vannak a C. albicans fehérjéire. Változásnak értékeltem új fehérje-csúcs
megjelenését az elektroferogramon, meglevő fehérje mennyiségének növekedését vagy
csökkenését (a kontrolhoz képest min. 20 %).
Az általam vizsgált 15 anyag közül 4: az 5, 11, 13, 15 anyagok okoztak értékelhető
változást. Új fehérje megjelenését nem sikerült kimutatnom egyik mintánál se, a hatás adott
fehérjék mennyiségének növekedésében jelentkezett, a 30-70 kDa közti tartományban. A
számított fehérjetömegek azért hozzávetőlegesek, mivel összetett fehérje-mintákat
vizsgáltunk és az fehérjék elválasztása nem volt teljes, az elektroferogramon egy csúcsként
jelentkező fehérje nem feltétlen jelent valóban egyetlen fehérjét. A fehérjék identifikálására
MS vizsgálatokat végeztünk, melynek során egy- és kétdimenziós elektroforézis
eredményeképp nyert gélekből is vettünk mintát. Egyelőre nem sikerült azonosítani a
fehérjéket, sem új-fehérje megjelenését igazolni a kezelés hatására, további vizsgálatok
szükségesek. Itt kell megemlítenem, hogy a chip-alapú elektroforézis-eredmények
reprodukálhatósága sem volt minden esetben tökéletes. Ennek számos oka lehet, például,
hogy az extrakció során kis mennyiségű mintát sikerült kinyerni, illetve az esetleges puffer-
maradványok is befolyásolhatják a mérést.
A 15 vizsgált kondenzált Mannich-keton fenti eredményei alapján általános
összefüggéseket a vegyületek szerkezete és a C. albicans fehérjéire gyakorolt hatás között
nem tudtam levonni. Az eredményeim és a vegyületek logP értéke között professzor Prókai
László (Department of Pharmacology & Neuroscience, University of North Texas, Health
Science Center at Fort Worth, USA) által végzett QSAR számítások (atom-típuson alapuló
elemzés [Ghose és mtsai.1988]) alapján úgy tűnik, hogy a fehérjeprofilra gyakorolt hatás
szempontjából ideális logP érték 1,8-2,5 közt van, míg a túl lipofil (logP > 2,5) vegyületeink
nincsenek hatással a fehérjeprofilra [Prókai L. nem publikált eredmények].

74
8. ÖSSZEFOGLALÁS
Kísérleteim során Candida sejtek áramlási citometriás antimikotikum-érzékenységi
meghatározását valósítottam meg. A vizsgálatban két klinikai gyakorlatban használt
antimikotikumot, egy fungicid és egy fungisztatikus hatású szert is bevontam. Az
érzékenység-vizsgálatot Candida albicans és non-albicans törzseken teszteltem.
Ezen eredményeket adaptáltam egy hasonló célra eddig nem használt, mikrofluidikai
áramlási rendszerre. Így elsőként sikerült kifejlesztenem egy gyors, könnyen kivitelezhető
chip-alapú áramlási citometriás diagnosztikai módszert antimikotikum-érzékenység
meghatározására.
Ezt követően a klasszikus antifungális szerek mellett egy érdekes, gombaellenes hatással
is rendelkező vegyületcsalád, a Mannich-ketonok vizsgálatára koncentráltam. A kifejlesztett
mikrofludikai módszer egy változatával meghatároztam néhány telítetlen Mannich-keton
standard C. albicans törzs elleni hatékonyságát (MIC). A Mannich-ketonok citotoxikus és
antimikrobiális hatása több tényezőre vezethető vissza. Elsődleges a célsejt tiol-csoportjainak
alkilálása, emellett azonban más tényezők, mint a fehérje-szintézis gátlása és gombáknál az
ergoszterin- és kitin-bioszintézis gátlása is fontos lehet.
Ezek közül a mechanizmusok közül a fehérje-összetétel megváltoztatását választottam ki.
Ennek monitorozására egy korszerű, chip-alapú gélelektroforézis technikát használtam. A
mikrochip elektroforézis alkalmasnak bizonyult Candida albicans fehérjéinek elválasztására.
Célom volt a C. albicans fehérjeprofiljában Mannich-keton kezelés hatására fellépő minőségi
és mennyiségi változások megfigyelése; és ilyen értelmű szerkezet-hatás összefüggések
felállítása. A Mannich-ketonok közül a kondenzált vázas vegyületek egy csoportját (15
vegyület) teszteltem. Megállapítottam, hogy a vegyületcsalád egyes tagjai az alkalmazott egy
órás kezelési idő mellett is befolyásolták a fehérjeprofilt, a hatás egyes fehérjék
mennyiségének növekedésében jelentkezett. Szerkezet-hatás összefüggések levonásához
egyelőre nem sikerült elég eredményet gyűjtenem.
Vizsgálataim alapján megállapítható, hogy a mikrofluidikai citometriás és elektroforézis
rendszerek alkalmasak egyes élesztőgombák vizsgálatára. Kiterjedtebb mikrobiológiai
felhasználásuk a jövőben várható.

75
9. IRODALOMJEGYZÉK
MEGJELENT KÖZLEMÉNYEK
Az értekezés alapjául szolgáló publikációk
Bouquet O, Kocsis B, Kilár F, Lóránd T, Kustos I.: Amphotericin B and fluconazole
suscepitibility of Candida species determined by cell-chip technology.
Mycoses 2012, 55 (3): e90-96. IF: 1,278
Bouquet O, Kocsis B, Kilár F, Kustos I.: Application of chip-based flow cytometry for
amphotericin B and fluconazole susceptibility testing on Candida strains.
Methods in Molecular Biology 2013, 968: 149-154.
Rossignol T, Kocsis B, Bouquet O, Kustos I, Kilár F, Nyul A, Jakus PB, Rajbhandari K,
Prókai L, d’Enfert C, Lóránd T.: Antifungal activity of fused Mannich ketones triggers an
oxidative stress response and is Cap1-dependent in Candida albicans.
PLoS One 2013, 8(4): e62142. IF: 3,73
Az értekezés témájában készült nem referált konferencia absztraktok
Bouquet O, Kustos I, Kocsis B, Lóránd T, Kilár F.: Rapid susceptibility testing of Candida
albicans by flow cytometry and cell-chip method. 7th International Symposium and Summer
School on Bioanalysis, CEEPUS project. 10-15 June, 2007, Pécs, Hungary. Abstract Book,
p.16.
Bouquet O, Kustos I, Lórand T, Kilár F, Kocsis B.: Determination of amphotericin B
susceptibility of Candida albicans by cell-chip method. ISAAC XXIV International
Congress. 17-21 May, 2008. Budapest, Hungary. Abstract Book p.253.

76
Bouquet O, Kustos I, Lórand T, Kilár F, Kocsis B: Application of cell-chip method to
rapid susceptibility testing. 9th International Symposium on Instrumental Analysis. 29 June –
2 July, 2008, Pécs, Hungary. Abstract Book p.55.
Bouquet O, Kustos I, Kocsis B, Kilár F, Lórand T.: Effect of fused Mannich ketones on the
protein profile of Candida albicans. The Fungal Cell, Annual Scientific Meeting of the British
Mycological Society, 1-4 September, 2009, Dundee, UK. Abstract Book p.25.
Bouquet O, Kustos I, Kocsis B, Kilár F, Lóránd T.: Examination of the protein profile of
Candida albicans - effect of fused Mannich ketons. 6th International Interdisciplinary Meeting
on Bioanalysis. 5-8 November, 2009, Pécs, Hungary. Abstract Book p.41.
Bouquet O.: Különböző antifungális vegyületek hatása Candida albicans fehérjeprofiljára.
Biológus Doktoranduszok Konferenciája, Pécsi Akadémiai Bizottság Biológiai Tudományok
Szakbizottságának rendezvénye, Pécs, 2009. november 12-13. előadás

77
FELHASZNÁLT IRODALMAK:
Ádám É. Gyógyszerészi mikrobiológia; egyetemi tankönyv. Semmelweis kiadó, Budapest
2001.
Al-Rawi N, Kavanagh K. A rapid method for the extraction of whole cell proteins from
Candida species. J. Microbiol. Meth. 1998; 34: 107-112.
Alshareef M, Metrakos N, Juarez-Perez E, Azer F, Yang F, Yang X Wang G. Separation of
tumor cells with dielectrophoresis-based microfluidic chip. Biomicrofluidics. 2013; 7:
011803.
Andresson H, van den Berg A. Microfluidic devices for cellomics: a review. Sensor. Actuat.
B-Chem. 2003; 92: 315-325.
Arend M, Westermann B, Risch N. Modern variants of the Mannich reaction. Angew. Chem.
Int. Ed. 1998; 37: 1044-1070.
Arendrup MC. Candida and candidaemia. Susceptibility and epidemiology. Dan. Med. J.
2013; 60: B4698.
Ateya AD, Erickson JS, Howell PB, Hilliard LR, Golden JP, Ligler FS. The good, the bad,
and the tiny: a review of microflow cytometry. Anal. Bioanal. Chem. 2008; 391: 1485-1498.
Avni T, Leibovici L, Paul M. PCR diagnosis of invasive candidiasis: systematic review and
meta-analysis. J. Clin. Microbiol. 2011; 49: 665-670.
Behr JB, Gourlain T, Helimi A and Guillerm G. Design, synthesis and biological evaluation
of hetaryl-nucleoside derivatives as inhibitors of chitin synthase. Bioorg. Med. Chem. Lett.
2003; 13: 1713-1716.
Ben-Ami R, Giladi M. Fluconazole-resistant Candida: collateral damage associated with prior
antibacterial exposure? Future Microbiol. 2012; 7: 1029-31.
Bossuyt L, Bosschaert J, Richert B, Cromphaut P, Mitchell T, Al Abadie M, Henry I, Bewlwy
A, Polyner T, Mann N, Czernielewski J. Lymcycline in the treatment of acne: an efficacious,
safe and cost-effective alternative to minocycline. Eur. J. Dermatol. 20013; 13: 130-135.

78
Botta M, Corelli F, Gasparrini F, Messina F, Mugnaini C. Chiral azole derivatives. 4.
Enantiomers of bifonazole and related antifungal agents: synthesis, configuration assignment
and biological evaluation. J. Org. Chem. 2000; 65: 4736-4739.
Brehm-Stecher BF, Johnson EA. Singel-cell microbiology: tools, technologies, and
applications. Microbiol. Mol. Biol. R. 2004; 68: 538-559.
Butts A, Krysan DJ. Antifungal drug discovery: something old and something new. PLOS
Pathogens 2012; 8: e1002870.
Carillo-Munoz AJ, Giusiano G, Ezkurra PA, Quindós G. Antifungal agents: mode of action in
yeast cells. Rev. Esp. Quim. 2006; 19: 130-139.
Chaffin W Lajean. Candida albicans cell wall proteins. Microbiol. Mol. Biol. R. 2008; 72:
495-544.
Chaturvedi V, Ramani R, Pfaller MA. Collaborative study of the NCCLS and flow cytometry
methods for antifungal susceptibility testing of Candida albicans. J. Clin. Microbiol. 2004;
42: 224-2251.
Chen SCA, Slavin MA, Sorrell TC. Echinocandin antifungal drugs in fungal infections – a
comparison. Drugs 2011; 71: 11-41
Chen X, Cui DF, Liu CC, Li H. Microfluidic chip for blood cell separation and collection
based on crossflow filtration. Sensor. Actuat. B-Chem. 2008; 130: 216-221.
Chernov SV, Shults EE, Makhmut MS, Bagrjanskaja IY, Gatilov YV, Tolstikov GA.
Synthetic transformations of natural diterpenes. Synthesis of alkaloid-like compounds from
lambertianic acid. Arkivoc 2003; 13: 172-183.
Chiba S, Kitamura M, Narasaka K. Synthesis of (-)-sordarin. J. Am. Chem. Soc. 2006; 128:
6931-7.
Chien CM, Cheng JL, Chang WT, Tien MH, Wu WY, Chang YH, Chang HY, Chen ST. Cell
phenotype analysis using a cell fluid-based microchip with high sensitivity and accurate
quantitation. J. Chromatogr. B 2003; 795: 1-8.

79
Cho SH, Chen CH, Tsai FS, Lo YH. Micro-fabricated fluorescence-activated cell sorter. Conf.
Proc. IEEE Eng. Med. Biol. Soc. 2009; 1075-1078.
Cho SH, Godin JM, Chen CH, Qiao W, Lee H, Lo YH. Review article: Recent advancements
in optofluidic flow cytometer. Biomicrofluidics. 2010; 4: 43001.
Cohen JI, Abel T, Burkett D, Engel R, Escalera J, Filshtinskaya M, Hatchett T, Leto M,
Melgar Y, Melkonian K. Polycations. 15. Polyammonium surfaces – a new approach to
antifungal activity. Lett. Drug. Des. Discov. 2004; 1: 88-90.
Comas J, Vives-Rego J. Enumeration, viability and heterogenity in Staphylococcus aureus
cultures by flow cytometry. J. Microbiol. Meth. 1998; 32: 45-53.
Davey HM. Life, death and in-between: meanings and methods in microbiology. Appl.
Environ. Microbiol. 2011; 77: 5571-5576.
Davis FA, Theddu N, Gaspari PM. Assymetric synthesis of substituted tropinones using the
intramolecular Mannich cyclization reaction and acyclic N-sulfinyl β-amino ketone ketals.
Org. Lett. 2009; 11: 1647-1650.
Dimmock JR, Kumar P, Manavathu EK, Obedeanu N, Grewal J. Activity of some Mannich
bases of conjugated styryl ketones against Candida albicans. Pharmazie. 1994; 49: 909-912.
Dimmock JR, Sidhu KK, Chen M, Reid RS, Allen TM, Kao GY, Truitt GA. Evaluation of
Mannich bases of cycloalkanones and related compunds for cytotoxic activity. Eur. J. Med.
Chem. 1993; 28: 313-322.
Dimmock JR, Smith LM, Smith PJ. The reaction of some nuclear substituted acyclic
conjugated styryl ketones and related Mannich bases with ethanethiol. Can. J. Chemistry
1980; 58: 984-991.
Dóczi I, Pető Z, Fodor E, Bereczki L, Nagy E, Hajdú E. Evaluation of fungaemia infections in
a Hungarian university hospital between 1996 and 2009. Acta Microbiol. Immunol. Hung.
2012; 59: 29-41.
Espinel-Ingroff A. Mechanisms of resistance to antifungal agents: yeasts and filamentous
fungi. Rev. Iberoam. Micol. 2008; 25: 101-106.

80
Espinel-Ingroff A, Barchiesi F, Cuenca-Estrella M, Pfaller MA, Rinaldi M, Rodriguez-Tudela
JL, Verwij PE. International and multicenter comparison of EUCAST and CLSI M27-A2
broth microdilution methods for testing susceptibilities of Candida spp. to fluconazole,
intraconazole, posaconazole and voricaonazole. J. Clin. Microbiol. 2005; 43: 3884-3889.
Faigl F, Kovács E, Mátravölgyi B, Thurner A. Gyógyszerkémiai alapfolyamatok – egyetemi
jegyzet. 2011 Typotex kiadó.
Favel A, Peyron F, De Méo M, Michel-Nguyen A, Carriere J, Chastin C, Regli P.
Amphotericin B susceptibility testing of Candida lusitaniae isolates by flow cytofluorometry:
comparison with Etest and the NCCLS broth macrodilution method. J. Antimicrob.
Chemother. 1999; 43: 227-232.
Fujie A. Discovery of micafungin (FK463): a novel antifungal drug derived from a natural
product lead. Pure Appl. Chem. 2007; 79: 603–614.
Gáspár A. Kapilláris zónaelektroforézis. Kossuth egyetemi kiadó, Debrecen, 2000.
Gerdts G, Luedke G. FISH and chips: Marine bacteria communities analyzed by flow
cytometry based on microfluidics. J. Microbiol. Method. 2006; 64: 232-240.
Gergely L. Orvosi mikrobiológia; egyetemi tankönyv második, átdolgozott kiadás. Alliter
Kiadói és Oktatásfejlesztő Alapítvány, Budapest 2003.
Ghose AK, Pritchett A, Crippen GM. Atomic physicochemical parameters for three
dimensional structure directed quantitative structure-activity relationships III: Modeling
hydrophobic interactions. J. Comput. Chem. 1988, 9, 80-90.
Goswami P, Das B. Adenine as aminocatalyst for green sythesis of diastereoselective
Mannich products in aqueous medium. Tetrahedron Lett. 2009; 50: 2384-2388.
Gras JL. A direct synthesis of α – methylene ketones. Tetrahedron Lett. 1978 April; 19: 2111-
2114.
Gras JL. A facile entry to vinyl ketones. Tetrahedron Lett. 1978 June; 19: 2955-2958.
Guantai EM, Ncokazi K, Egan TJ, Gut J, Rosenthal PJ, Bhampidipati R, Kopinathan A, Smith
PJ, Chibale K. Enone- and chalcone-chloroquinoline hybrid analogs: in silico-guided design,

81
synthesis, antiplasmodial activity, in vitro metabolism and mechanistic studies. J. Med. Chem.
2011; 54: 3637-3649.
Guttman A. Gel and polymer-solution mediated separation of biopolymers by capillary
electrophoresis. J. Chromatogr. Sci. 2003; 41: 449-459.
Hayashi Y, Urushima T, Shin M, Shoji M. The stereoselective synthesis of α-substitued β-
amino secondary alcohols based on the proline-mediated, assymetric, three-component
Mannich reaction and its application to the formal total sythesis of nikkomycins B and Bx.
Tetrahedron 2005; 61: 11393-11404.
Heilmann CJ, Sorgo AG, Klis FM. News from the fungal front: wall proteome dynamics and
host-pathogen interplay. PLoS Pathog.s 2012; volume 8, issue 12
Hernandez A, Marcos M, Rapoport H. Synthesis of (+)- and (-)-N-BOC-7-
azabicyclo[2.2.1]heptan-2-ones, versatile intermediates for the enantiospecific synthesis of
(+)- and (-)-epibatidine and analogs. J. Org. Chem. 1995; 60: 2683-2691.
Ibrahim A, Spellberg B, Edwards J. Prospective on mucormycosis pathogenesis and
treatment. Curr. Opin. Infect. Dis. 2008; 21: 620-625.
Jewell SN, Waldo Rh, Cain CC, Falkinham JO. Rapid detection of lytic antimicrobial activity
against yeast and filamentous fungi. J. Microbiol. Method. 2002; 49: 1-9.
Johnson EM, Ojwang JO, Szekely A, Wallace TL, Warnock DW. Comparison of in vitro
antifungal activities of free and liposome-encapsulated nystatin with those of four
amphotericin B formulations. Antimicrob. Agents Ch. 1998; 42: 1412-1416.
Johnson MD, Perfect JR. Use of antifungal combination therapy: agents, order and timing.
Curr Fungal Infect Rep. 2010; 4: 87-95.
Kaneko S, Uchida T, Shibuya S, Honda T, Kawamoto I, Harasaki T et al. Synthesis of
sordaricin analogues as potent antifungal agents against Candida albicans. Bioorg. Med.
Chem. Lett. 2002; 12: 803-806.
Kauffmann CA. Clinical efficacy of new antifungal agents. Curr. Opin. Microbiol. 2006; 9:
483-488.

82
Kirk SM, Callister SM, Lim LCL, Schell RF. Rapid susceptibility testing of Candida albicans
by flow cytometry. J. Clin. Microbiol 1997; 35: 358-363.
Klein LL, Li L, Chen HJ, Curty CB, DeGoey DA, Grampovnik DJ, Leone CL, Thomas SA,
Yeung CM, Funk KW, Kishore V, Lundell EO, Wodka D, Meulbroek JA, Alder JD, Nilius
AM, Lartey PA, Plattner JJ. Total synthesis and antifungal evaluation of cyclic
aminohexapeptides. Bioorgan. Med. Chem. 2000; 8: 1677-1696.
Kocsis B, Kustos I, Kilár F, Nyul A, Jakus PB, Kerekes Sz, Villarreal V, Prókai L, Lóránd T.
Antifungal unsaturated cyclic Mannich ketones and amino alcohols: study of mechanism of
action. Eur. J. Med. Chem. 2009; 44: 1823-1829.
Kuschel M, Buhlmann C, Preckel T High-throughput protein and DNA analysis based on
microfluidic on-chip electrophoresis. JALA 2005; 10: 319-326.
Kustos I, Kocsis B, Kilar F. Bacterial outer membrane protein analysis by electrophoresis and
microchip technology. Expert. Rev. Proteomics. 2007; 4: 91-106.
Kustos I, Nyul A, Lóránd T, Kocsis B, Kilár F. Effect of antifungal agents on protein
composition of Candida albicans studied by capillary electrophoresis and chip technology.
J. Biochem. Biophys. Methods 2006; 69: 57-65.
Lal B, Gund VG, Bishe NB, Gangopadhyay AK. Mannich reaction: an approach for the
synthesis of water soluble mulundocandin analogues. Bioorg. Med. Chem. 2004; 12: 1751-
1768.
Lapsley MI, Wang L, Huang TJ. On-chip flow cytometry: where is it now and where is it
going ? Biomark Med. 2013; 7: 75-78.
Liang H. Sordarin, an antifungal agent with a unique mode of action. Beilstein J. Org. Chem.
2008; 4: 1-14.
Lim CSY, Tung CH, Rosli R, Chong PP. An alternative Candida spp. cell wall disruption
method using a basic sorbitol lysis buffer and glass beads. J. Microbiol. Meth. 2008; 75: 576-
578.
List B. The direct catalytic asymmetric three-component Mannich reaction. J. Am. Chem. Soc.
2000; 122: 9336-9337

83
Loeffler J, Stevens DA. Antifungal drug resistance. Clin. Infect. Dis. 2003; 36(suppl 1): S31-
41.
Lopez SN, Castelli MV, Zacchino SA, Domnguez JN, Lobo G, Charris-Charris J, Cortes JC,
Ribas JC, Devia C, Rodirguez AM, Enriz RD. In vitro antifungal evaluation and structure-
activity relationships of a new series of chalcone derivatives and synthetic analogues, with
inhibitory properties against polymers of the fungal cell wall. Bioorgan. Med. Chem. 2001; 9:
1999-2013.
Lóránd T, Kocsis B. Recent advances in antifungal agents. Mini-Rev. Med.Chem. 2007; 7:
900-911.
Lóránd T, Kocsis B, Sohár P, Nagy G, Kispál Gy, Krane HG, Schmitt H, Weckert E.
Synthesis and antibacterial study of unsaturated Mannich ketones. Eur. J. Med. Chem. 2001;
36: 705-717.
Lóránd T, Kocsis B, Sohár P, Nagy G, József P, Kispál Gy, László R, Prókai R. Synthesis and
antibacterial activity of fused Mannich ketones. Eur. J. Med. Chem. 2002; 37: 803-812.
Lustyik György: Az áramlási citometria és a konfokális mikroszkópia alapelvei és
alkalmazásuk az orvos-biológiai kutatásban. PTE ÁOK Doktori Iskola Ph.D. kurzusának
anyaga, 2004.november, 2008.január
Mattanovich D, Borth N. Applications of cell sorting in biotechnology. Microb. Cell. Fact.
2006; 5:12.
M27-A3 - Reference method for broth dilution antifungal susceptibility testing of yeast;
Approved Standard - Third Edition. 2008; Vol. 28. No.14.
Mallié M, Bastide JM. Les nouveaux antifongiques et leur stratégie de recherché. Revue
Francaise des Laboratoires 2001; N° 332: 63-70.
Martinez A, Aviles P, Jimenez E, Caballero J, Gargallo-Viola D. Activities of sordarins in
experimental models of candidiasis, aspergillosis and pneumocytosis. Antimicrob. Agents
Chemother. 2000; 44: 3389-3394.
Mason DJ, Allman R, Stark JM, Lloyd D. Rapid estimation of bacterial antibiotic
susceptibility with flow cytometry. J. Microsc.1994; 176: 8-16.

84
Mehra T, Köberle M, Braunsdorf C, Mailänder-Sanchez D, Borelli C, Schaller M. Alternative
approaches to antifungal therapies. Exp. Dermatol. 2012; 21: 778-782.
Mehta RJ, Kingsbury WD, Valenta J, Actor P. Anti-Candida activity of polyoxin: example of
peptide transport in yeasts. Antimicrob. Agents Chemother. 1984; 25: 373-374.
Mesa-Arango AC, Scorzoni L, Zaragoza O. It only takes one to do many jobs: Amphotericin
B as antifungal and immunomodulatory drug. Front. Microbiol. 2012; 3: 286
Mitchell M, Hudspeth M, Wright A. Flow cytometry susceptibility testing for the antifungal
caspofungin. J. Clin. Microbiol. 2005; 43: 2586–2589.
Moen DM, Lyseng-Williamson KA, Scott LJ. Liposomal Amphotericin B: a review of its use
as empirical therapy in febrile neutropenia and in the treatment of invasive fungal infections.
Drugs 2009; 69: 361-392.
Notz W, Tanaka F, Watanabe SI, Chowdari NS, Turner JM, Tahyumanavan R, Barbas CF.
The direct organocatalytic asymmetric Mannich reaction: unmodified aldehydes as
nucleophiles. J. Org. Chem. 2003; 68: 9624-9634.
Obi K, Uda JI, Iase K, Sugimoto O, Ebisu H, Matsuda A. Novel nikkomycin analogues:
inhibitors of the fungal cell wall biosynthesis enzyme chitin synthase. Bioorg. Med. Chem.
Lett. 2000; 10: 1451-1454.
Odds FC. Antifungal agents: their diversity and increasing sophistication. Mycologist 2003;
17: 51-55.
Odds FC, Brown AJP, Gow NAR. Antifungal agents: mechanisms of action. Trends
Microbiol. 2003; 11: 272-279.
Palkova Z, Vachova L, Valer M, Preckel T. Single-cell analysis of yeast, mammalian cells,
and fungal spores with a microfluidic pressure-driven chip-based system. Cytometry Part A.
2004; 59A: 246-253.
Page BT, Shields CE, Merz WG, Kurtzman CP. Rapid identification of ascomycetous yeasts
from clinical specimens by a molecular method based on flow cytometry and comparison with
identifications from phenotypic assays. J. Clin. Microbiol. 2006; 44: 3167-3171.

85
Pati HN, Das U, Sharma RK, Dimmock JR. Cytotoxic thiol alkylators. Mini. Rev. Med. Chem.
2007; 7: 131-139.
Peter J, Armstrong D, Lyman CA, Walsh TJ. Use of fluorescent probes to determine MICs of
amphotericin B and caspofungin against Candida spp. and Aspergillus spp. J. Clin. Microbiol.
2005; 43: 3788-3792.
Pina-Vaz C, Costa-de-Oliveira S, Rodrigues AG, Espinel-Ingroff A. Comparison of two
probes for testing susceptibilities of pathogenic yeasts to voriconazole, itraconazole, and
caspofungin by flow cytometry. J. Clin. Microbiol. 2005; 43: 4674-4679.
Porro D, Vai M, Vanoni M, Alberghina L, Hatzis C. Analysis and modeling of growing
budding yeast populations at the single cell level. Cytometry A 2009; 75: 114-120.
Preckel T, Luedke G, Chan SDH, Wang BN, Dubrow R, Buhlmann C. Detection of cellular
parameters using of a microfluidic chip-based system. JALA 2002; 7: 85-89.
Ramani R, Chaturvedi V. Flow cytometry antifungal susceptibility testing of pathogenic
yeasts other than Candida albicans and comparison with the NCCLS broth microdilution test.
Antimicrob. Agents Chemother. 2000; 44: 2752-2758.
Ramani R, Ramani A, Wong SJ. Rapid flow cytometric susceptibility testing of Candida
albicans. J. Clin. Microbiol. 1997; 35: 2320-2324.
Rani N, Sharma A, Gupta GK, Singh R. Imidazoles as potential antifungal agents: a review.
Mini. Rev. Med. Chem. 2013; 13: 1626-55.
Rodriguez CA, Patrick CC. New therapeutic options for invasive fungal diseases. Semin.
Pediat. Infect. Dis. 2001; 12: 301-308.
Rotaru I, Ionescu M, Donescu D, Vuluga M, Purcar V. Synthesis of new aromatic Mannich
polyols for rigid polyurethane foams. U.P.B. Sci. Bull., Series B. 2007; 69: 35-42.
Sandven P, Bevanger L, Digranes A, Haukland HH, Mannsaker T, Gaustad P, Norwegian
Yeast Study Group. Candidemia in Norway (1991 to 2003): results from a nationwide study.
J. Clin. Microbiol. 2006; 44: 1977-81.

86
Savluchinske-Feio S, Curti MJM, Gigante B, Roseiro JC. Antimicrobial activity of resin acid
derivatives. Appl. Microbiol. Biotechnol. 2006; 72: 430-436.
Scott LJ. Micafungin – a review of its use in the prophylaxis and treatment of invasive
Candida infections. Drugs 2012; 72: 2141-2165.
Sedlák M. Amphotericin B: from derivatives to covalent targeted conjugates. Mini-Rev.
Med.Chem. 2009; 9: 1306-16.
Shastry M, Nielsen J, Ku T, Hsu MJ, Liberator P, Anderson J, Schmatz D, Justice MC.
Species-specific inhibition of fungal protein synthesis by sordarin: identification of a sordarin-
specificity region in eukaryotic elongation factor 2. Microbiology 2001; 147: 383-390.
Subramaniapillai SG. Mannich reaction: a versatile and convenient approach to bioactive
skeletons. Chem. Sci. J. 2013; 125: 467-482.
Sujith KV, Rao JN, Shetty P, Kalluraya B. Regioselective reaction: synthesis and
pharmacological study of Mannich bases containing ibuprofen moiety. Eur. J. Med. Chem.
2009; 44: 3697-3702.
Sumalatha Y, Reddy PP, Reddy R, Satyanarayana B. Sythesis and spectral characterization of
process-realeted substances to the hypnotic agent zolpidem. Arkivoc 2009; vii: 143-149.
Tanino K, Takahashi M, Murayama K, Kuwajima I. Generation of a Lewis acid activated
formaldimine and its reaction with enol silyl ethers. N-unsubstituted aminomethylation. J.
Org. Chem. 1992; 57: 7009-7010.
Thienpont A, Gal J, Aeschlimann C, Félix G. Studies on stereoselective separations of the
„azole” antifungal drugs ketoconazole and itraconazole using HPLC and SFC on silica-based
polysaccharides. Analysis 1999; 27: 713-718.
Tragiannidis A, Fegeler W, Rellensmann G, Debus V, Muller V, Hoering –Franz I, Siam K,
Pana ZD, Jurgens H, Groll AH. Candidaemia in a European Paediatric University Hospital: a
10-year observational study. Clin. Microbiol. Infect. 2012; 18: E27-30.
Tramontini M, Angiolini L, Ghedini N. Mannich bases in polymer chemistry. Polym. 1988;
29: 771-788.

87
Vashishtha SC, Zello GA, Nienaber KH, Balzarini J, De Clercq E, Stables JP, Dimmock JR.
Cytotoxic and anticonvulsant aryloxyaryl Mannich bases and related compunds. Eur. J. Med.
Chem. 2004; 39: 27-35.
Walberg M, Gaustad P, Steen HB. Rapid assessment of ceftazidime, ciprofloxacin, and
gentamicin suscepitibility in exponentially-growing E. coli cells by means of flow cytometry.
Cytometry 1997; 27: 169-178.
Watanabe T, Ogasawara A, Mikami T, Matsumoto T. Hyphal formation of Candida albicans
is controlled by electron transfer system. Biochem. Bioph. Res. Co. 2006; 348: 206-211.
Wenisch C, Linnau KF, Parschalk B, Zedwitz-Liebenstein K, Georgopoulos A. Rapid
susceptibility testing of fungi by flow cytometry using vital staining. J. Clin. Microbiol.
1997; 35: 5-10.
Wenisch C, Moore CB, Krause R, Presterl E, Pichna P, Denning DW. Antifungal
susceptibility testing of fluconazole by flow cytometry correlates with clinical outcome.
J. Clin. Microbiol. 2001; 39: 2458-2462.
White TC, Marr KA, Bowden RA. Clinical, cellular and molecular factors that contribute to
antifungal drug resistance. Clin. Microbiol. Rev. 1998; 11: 382-402.
Zhang H, Mitsumori S, Utsumi N, ImaiM, Garcia-Delgado N, Mifsud M, Albertshofer K,
Cheong PH, Houk KN, Tanaka F, Barbas CF. Catalysis of 3-pyrrolidinecarboxylic acid and
related pyrrolidine derivatives in enantioselective anti-Mannich-type reactions: Importance of
the 3-acid group on pyrrolidine for stereocontrol. J. Am. Chem. Soc. 2008; 130: 875-886.
ÁBRÁK FORRÁSAI:
[1] http://www.medschool.lsuhsc.edu/pharmacology/docs/Fox_2009.pdf
[2] Lóránd T, Kocsis B. Recent advances in antifungal agents. Mini-Rev. Med.Chem.
2007; 7: 901.o.
[3] http://www.abdserotec.com/flow-cytometry-signal-processing.html
[4] Preckel T, Luedke G, Chan SDH, Wang BN, Dubrow R, Buhlmann C. Detection of
cellular parameters using of a microfluidic chip-based system. JALA 2002; 7: 86.o.
[5] http://www.chem.agilent.com/

88
10. KÖSZÖNETNYILVÁNÍTÁS
Köszönöm témavezetőimnek, Dr. Lóránd Tamásnak és Dr. Kocsis Bélának, hogy
hosszú évek óta, kitartóan segítették kutatási munkám; ötleteikkel, javaslataikkal
támogattak. Külön köszönöm Kocsis tanár úrnak, hogy a munkámhoz nélkülözhetelen
mikrobiológiai alapokkal megismertett, Lóránd tanár úrnak pedig, hogy fáradhatatlanul
terelt a kémiai gondolkodásmód és szemlélet felé.
Köszönöm Dr. Kilár Ferencnek, volt témavezetőmnek, hogy szakmai javaslataival és
gyakorlati tanácsaival mindvégig figyelemmel kísérte tevékenységem.
Köszönettel tartozom dr. Kustos Ildikónak, aki ezen a kutatási vonalon elindított,
később is mindig segített és készséggel javította munkáim.
Köszönöm Dr. Prókai László professzor úrnak a logP számításokat.
Köszönöm a PTE TTK Kémia Intézet Analitikai és Környezeti Kémia Tanszék, a
PTE ÁOK Orvosi Mikrobiológiai és Immunitástani Intézet, a PTE ÁOK Bioanalitikai
Intézet, a PTE ÁOK Biokémiai és Orvosi Kémiai Intézet, és a PTE ÁOK Biofizikai
Intézet valamennyi munkatársának, akik munkámban bármilyen formában segítségemre
voltak.
Külön köszönöm Sajtiné Pintér Krisztinának a képletek, ábrák szerkesztését, Kocsis
Bélánénak a mikrobiológiai előkészítést, Jámbor Évának az MS mérésekben nyújtott
segítségét.
Végül pedig köszönöm családomnak és barátaimnak, hogy mindig támogattak,
bátorítottak, bíztattak és elviseltek.
Recommended

















