Embed Size (px)
Citation preview

1
Neuropeptidek retinoprotektív hatásának vizsgálata
Doktori (Ph.D.) értekezés
Dr. Werling Dóra
Témavezetők: Dr. Reglődi Dóra, egyetemi tanár
Dr. Biró Zsolt, egyetemi tanár
Programvezető: Dr. Csernus Valér, egyetemi tanár
Doktori iskola vezetője: Dr. Szekeres Júlia, egyetemi tanár
Pécsi Tudományegyetem Általános Orvostudományi Kar
Anatómiai Intézet
Pécs, 2016

2
Tartalomjegyzék
1. BEVEZETÉS ..................................................................................................................... 7
1.1. A hypophysis adenilát cikláz aktiváló polipeptidről (PACAP) általában ......................... 7
1.1.1. A PACAP előfordulása a szervezetben ............................................................................ 8
1.1.2. A PACAP receptorai ....................................................................................................... 8
1.1.3. A PACAP élettani hatásai................................................................................................ 9
1.1.4. A PACAP fragmensei, peptidcsalád tagjai .................................................................... 11
1.2. A retina szerkezete ............................................................................................................. 14
1.3. Bilateralis carotis communis okklúzió (BCCAO)............................................................. 16
1.4. Szemcseppes terápia ........................................................................................................... 18
1.5. A PACAP jellemzői a szemben .......................................................................................... 19
1.5.1. A PACAP előfordulása a szemben ................................................................................ 19
1.5.2. A PACAP receptorai a retinában ................................................................................... 20
1.5.3. A PACAP hatásai a retinában ........................................................................................ 21
1.5.4. A PACAP hatása ischaemiás retinopathiában................................................................ 22
2. CÉLKITŰZÉSEK ÖSSZEFOGLALÁSA .................................................................... 24
3. SZEKRETIN, GLÜKAGON ÉS KÜLÖNBÖZŐ PACAP FRAGMENSEK
VIZSGÁLATA ISCHAEMIÁS RETINOPATHIÁBAN ............................................. 25
3.1. Anyagok és módszerek ....................................................................................................... 25
3.1.1. Az arteria carotis communis permanens, bilaterális lekötésével létrehozott
retinadegenerációs modell ............................................................................................. 25
3.1.2. Rutin morfológiai és morfometriai analízis ................................................................... 26
3.2. Eredményeink ..................................................................................................................... 27
4. SZEMCSEPPKÉNT ALKALMAZOTT PACAP1-27 ÉS 1-38 VIZSGÁLATA
ISCHAEMIÁS RETINOPATHIÁBAN ........................................................................ 32
4.1. Anyag és módszer ............................................................................................................... 32
4.1.1. Bilaterális carotis communis műtétet követő szemcseppes terápia ................................ 32
4.1.2. A PACAP1-27 és 1-38 szemcseppel kezelt retinák hisztológiai analízise ..................... 33
4.1.3. PACAP1-27 és 1-38 radioaktív jelölése ........................................................................ 34
4.1.4. Gliális fibrilláris savas protein expressziójának mérése PACAP1-27 és 1-38
szemcseppes kezelést követően ..................................................................................... 34
4.1.5. Western blot analízis, BCCAO-t követő PACAP1-27 szemcseppes kezelés után ......... 35
4.1.6. PACAP1-27 szemcseppes kezelést követő citokin array vizsgálat ................................ 36
4.1.7. Elektroretinográfiás (ERG) vizsgálat PACAP1-27 szemcseppes terápiát követően ...... 37

3
4.2. Eredményeink ..................................................................................................................... 38
4.2.1. Rutin morfologiai és morfometriai analízis ................................................................... 38
4.2.2. 125I izotóppal jelölt PACAP1-27 és 1-38 koncentráció változása cseppentést követően 44
4.2.3. Müller glia sejtek immunhisztokémiai analízise ............................................................ 45
4.2.4. Akt és ERK1/2 foszforilációs változása PACAP1-27 kezelés hatására ......................... 47
4.2.5. PACAP1-27 szemcseppes kezelés hatása az ischaemia okozta citokin expresszió
változásra....................................................................................................................... 49
4.2.6. Funkcionális változások vizsgálata PACAP1-27 szemcseppes terápiát követően ......... 51
5. MEGBESZÉLÉS ............................................................................................................. 53
5.1. PACAP családjába tartózó neuropeptidek valamint fragmenseinek vizsgálatáról szóló
korábbi tanulmányok, illetve a mostani vizsgálatok eredményének összefoglalása .................. 53
5.2. Szemcseppként alkalmazott PACAP1-27 és 1-38 kutatási eredményeinek összegzése .. 56
6. ÖSSZEFOGLALÁS ........................................................................................................ 62
7. IRODALOMJEGYZÉK ................................................................................................. 63
8. PUBLIKÁCIÓS LISTA .................................................................................................. 88
8.1. Az értekezés alapjául szolgáló publikációk ....................................................................... 88
8.2. Az értekezés témájába közvetlenül nem illeszkedő egyéb közlemények ......................... 88
8.3. Konferencia szereplések ..................................................................................................... 89

4
Rövidítések jegyzéke
125I: jód izotóp
6-OHDA: 6-hidroxidopamin
AC: adenilát-cikláz
Akt/PKB: protein kináz B
Bad: „Bcl-2-associated death promoter” / Bcl-2 függő sejthalál promoter
Bax: „Bcl-2-associated X protein” / Bcl-2-hoz asszociált X protein
BCCAO: „bilateral common carotid artery occlusion” / kétoldali arteria carotis communis
okklúzió
Bcl-2: „B-cell lymphoma 2” / B-sejt lymphoma 2
cAMP: ciklikus adenozin monofoszfát
cGMP: ciklikus guanozin monofoszfát
CINC-1: „cytokine-induced neutrophil chemoattractant” / citokin indukálta neutrofil
kemoattraktáns
CNTF: „ciliary neurotrophic factor”/ ciliáris neurotrófikus faktor
CREB: „cAMP response element binding protein” / cAMP reszponzív elem-kötő protein
CXCL-7: thymus kemokin
DPPIV: dipeptidyl peptidáz-IV
ERG: elektroretinográfia
ERK: extracelluláris szignál regulációs kináz
GCL: „ganglion cell layer” / ganglion sejtek rétege
GFAP: „glial fibrillary acidic protein” / gliális fibrilláris savas protein
GHRH: „growth-hormone-releasing hormone” / növekedési hormon szabályozó hormon
GIP: „glucose-dependent insulinotropic polypeptide” / glükóz függő inzulinotróp polipeptid
GLP: „glucagon-like peptide” / glükagonhoz hasonló peptid
H2O2: hydrogén peroxid
HIF-1α: „hypoxia-inducible factor 1-alpha” / hypoxia-indukált faktor 1-alfa
HPLC: „High-performance liquid chromatography” / nagyhatékonyságú folyadékkromatográfia

5
IL: interleukin
ILM: „inner limiting membrane” / belső határhártya
IMP: inozitol monofoszfát
INL: „inner nuclear layer” / belső magvas réteg
IP3: inositol trifoszfát
I-PACAP: pituitary adenylate cyclase activating polypeptide jód izotóppal jelölve
IPL: „inner plexiform layer” / belső szinaptikus réteg
i.p.: intraperitoneális
i.v.: intravitravénás
JNK: c-Jun N-terminális kinázok
KIR: központi idegrendszer
KO: „knock out”
LED: „light-emitting diode” / fény-emissziós dióda
LIX: „lipopolysaccharide-induced cystein-X-cystein chemokine”/ lipopoliszacharid-indukált
cisztein-X-cisztein kemokin
MIP-3α: „macrophage inflammatory protein” / makrofág gyulladásos fehérje
mRNS: „messenger” ribonukleinsav
MSG: mononátrium-glutamát
NFL: „nerve fiber layer” / idegrostok rétege
NGF: „nerve growth factor” / ideg növekedési faktor
NMDA: N-metil-D-aszparaginát
OLM: „outer limiting membrane” / külső határhártya
ONL: „outer nuclear layer” / külső magvas réteg
OPL: „outer plexiform layer” / külső szinaptikus réteg
p38 MAPK: p38 mitogén-aktivált protein kináz
PACAP: „pituitary adenylate cyclase-activating polypeptide” / hypophysis adenilát cikláz
aktiváló polipeptid
PB: „phosphate buffer” / foszfát puffer
PBS: „phosphate buffered saline” / foszfát puffer fiziológiás sóoldatban
PE: pigment epithelium
PHM: peptid hisztidin methionin

6
PI: propidium jodid
PKA: protein kináz A
PKC: protein kináz C
PLC: foszfolipáz C
RANTES: „regulated on activation normal T cell expressed and secreted”/ normál T-sejt által
regulált, expresszált és szekretált citokin
SEM: „standard error of mean”
Sham: „sham-operated group” / áloperált csoport
sICAM: szolubilis intercelluláris adhéziós molekula
SOCB: solutio ophthalmica cum benzalkonio
TBS-T: „tris-buffered saline-Tween 20” / Tween-t tartalmazó Tris-pufferelt sóoldat
TIMP: „tissue inhibitor of metalloproteinase” / mátrix metalloproteináz szöveti gátlója
TNF: tumor nekrózis factor
VEGF: „vascular endothelial growth factor” / vaszkuláris endoteliális növekedési faktor
VIP: „vasoactive intestinal peptid” / vazoaktív intesztinális peptid

7
1. Bevezetés
1.1. A hypophysis adenilát cikláz aktiváló polipeptidről (PACAP)
általában
A PACAP egy mozaikszó, mely az angol „Pituitary adenylate cyclase activating
polypeptide” kezdőbetűiből áll össze. Egy 38 aminosavból álló C-terminálisán α-amidált
peptid, melyet 1989-ben Arimura és munkacsoportja izoláltak birka hypothalamusból. Nevét
az adenohypophysisben megtalálható sejtek adenilát cikláz aktiválása révén kapta, mely
következményes ciklikus adenozin-monofoszfát (cAMP) emelkedést idéz elő (Arimura 2007,
Miyata és mtsai 1989). A PACAP, a glükagon, a glükagonhoz hasonló peptid (glucagon-like
peptide - GLP) - 1 és 2, a növekedési hormont szabályozó hormon (growth-hormone releasing
hormone - GHRH), a vazoaktív intesztinális peptid (VIP), a hisztidin – metionin peptid (peptide
histidine methionine - PHM), a szekretin és a glükóz-függő inzulinotróp polipeptid (glucose-
dependent insulinotropic polypeptide - GIP) mellett a szekretin/glükagon/GHRH peptidcsalád
tagja (Arimura and Shioda 1995; Lee és mtsai 2007). Biológiailag aktív, N-terminális
struktúrája a filogenesis során nagymértékben konzervált szerkezetű maradt (a halaktól egészen
az emlősökig), bizonyítva a peptid esszenciális biológiai szerepét (Bourgault és mtsai 2011;
Sherwood és mtsai 2000). Egy évvel a PACAP1-38 felfedezése után azonosították rövidebb
fragmensét, a PACAP1-27-et, mely N terminálisa megegyezik a PACAP1-38-al (Miyata és
mtsai 1990). A PACAP1-27 szerkezetileg leginkább a VIP-hez hasonlít, mellyel N-terminális
szakasza 68%-ban megegyezik, azonban adenilát cikláz aktiváló hatását 1.000-10.000-szer
nagyobb mértékben fejti ki (Vaudry és mtsai 2009). Feltételezések szerint a PACAP1-27 és a
PACAP1-38 egymástól független módon hasítódik le a PACAP prekurzor proteinről, azaz nem
alternatív splice variánsokról van szó (Hosoya és mtsai 1992; Miyata és mtsai 1990). A PACAP
humán génjének lókusza a 18p11 kromoszóma, mely 5 exont tartalmaz, amelyről a 176
aminosav hosszúságú prepro-PACAP transzlálódik. A PACAP1-38 a 27 aminosavas formához
hasonlóan ebből a prekurzorból képződik, proteolítikus és amidációs lépéseket követően
(Hosoya és mtsai 1992). Gerincesekben 90 %-ban a 38 aminosavból álló forma dominál
(Vaudry és mtsai 2000). A PACAP gyors lebomlásáért a dipeptidyl peptidáz (DPP)-IV felelős,
felezési ideje a szisztémás keringésben mindössze néhány perc (Bourgault és mtsai 2008; Zhu
és mtsai 2003). Lebomlás során rövidebb peptidekre fragmentálódnak, melyek főként

8
antagonista hatásúak. Közülük a vizsgálatokban leggyakrabban használt forma a PACAP6-38
(Bourgault és mtsai 2008), melyet kezdetben a PAC1 receptor szelektív, kompetitív
antagonistájaként írtak le (Robberecht és mtsai 1992).
Számos kísérlet alátámasztotta, hogy a PACAP6-38 nemcsak a PAC1, hanem a VPAC2
receptor jelátvitelét is gátolja. Érdekes módon a tracheában, chondrocytákon, valamint
retinoblastoma és cytotrophoblast sejteken agonistaként viselkedik (Juhász és mtsai 2014; Liu
és mtsai 2000; Reglődi és mtsai 2008; Wojcieszak és Zawilska 2014).
1.1.1. A PACAP előfordulása a szervezetben
A PACAP központi idegrendszeri jelenlétét igazolták emlősök, madarak, halak valamint a
kétéltűek esetében is. Legnagyobb mennyiségben a hypothalamusban fordul elő, de
megtalálható a kéregállomány, az agytörzs, az amygdala, a hippocampus, a tobozmirigy, és a
kisagy területén is (Arimura és mtsai 1991; Hannibal 2002; Reglődi 2009). A perifériás
idegrendszerben, a spinális ganglionok kis érzőidegsejtjeiben, valamint a vegetatív pre- és
postganglionáris neuronokban is jelen van (Köves és mtsai 1990). A PACAP az idegrendszeren
kívül számos perifériás szervben is kimutatható. Megtalálható az endokrin mirigyekben, a
gasztrointesztinális rendszerben, a kardiovaszkuláris és a légzőrendszerben, továbbá az
urogenitális traktus területén is (Vaudry és mtsai 2009).
1.1.2. A PACAP receptorai
A PACAP hatásait G-proteinhez kötött receptorokon keresztül fejti ki. A receptorok közül
először a VPAC1-et, majd a VPAC2-őt fedezték fel (Ishihara és mtai 1992; Lutz és mtsai 1993).
Már a receptorok izolálásakor világossá vált, hogy a VPAC receptorokat a VIP és a PACAP is
egyaránt aktiválja. A PACAP specifikus receptorát, a PAC1 receptort csak ezt követően
azonosították (Pisegna és Wank 1993). Ehhez a PACAP mintegy 1000-szer nagyobb
affinitással kötődik, mint a VIP, ezzel szemben a VPAC1 és VPAC2 receptorokhoz azonos
erősséggel kötődnek. A receptor hét transzmembrán karból és egy intracellulárisan G-proteint
kötő doménből áll. A PAC1 receptor, agonista peptid bekötődés hatására adenilát cikláz (AC),
foszfolipáz C (PLC) és protein kináz C (PKC) által közvetített útvonalakat aktivál, de akár L-

9
típusú feszültségfüggő kálcium csatornákat is megnyithat. A PAC1 receptor nyolc splice-
variánssal rendelkezik, melyek a 3. intracelluláris karon található „hip” és „hop” szekvenciák
jelenléte vagy hiánya következtében különböző szekunder messengerek aktivációjához
vezetnek (Dickson és Finlayson 2009). A VPAC1 és VPAC2 receptorok aktiválódása cAMP,
vagy ciklikus guanozin-monofoszfát (cGMP) szint emelkedéséhez vezet (Murthy és mtsai
1997; Shioda és mtsai 2003). A PACAP receptorok szinte minden szervben expresszálódnak,
azonban az altípusok eloszlása szervrendszerenként változó. A PAC1 receptor magas
koncentrációban mutatható ki többek között az agyban, adenohypophysisben, gerincvelőben,
hasnyálmirigyben, mellékvesevelőben, vesében, herében és a petefészekben is. Megtalálható
még például a nagy nyálmirigyekben: fültőmirigyben, submandibularis és sublingualis
mirigyekben (Matoba és mtsai 2016). A VPAC1 receptor a központi idegrendszer mellett
előfordul többek között a tüdőben, májban, lépben, thymusban, mellékvesevelőben és a
gasztrointesztinális traktusban. A VPAC2 receptor a pancreas β-sejtjein, a mellékvese kortikális
részében és a gonádokban expresszálódik (Filipsson és mtsai 1998; Joo és mtsai 2004; Usdin
és mtsai 1994; Vaudry és mtsai 2009).
1.1.3. A PACAP élettani hatásai
A PACAP pleiotróp és multifunkcionális neuropeptid, melynek neuroprotektív és
neurotrófikus hatását széles körben tanulmányozzák. A PACAP rövid felezési ideje ellenére
számos biológiai hatással bír (Vaudry és mtsai 2009). A PACAP szerepet játszik élettani
folyamatok szabályozásában, így például a táplálkozás, alvás, termoreguláció, és a cirkadián
ritmus szabályozásában (Gray és mtsai 2002; Hannibal 2006; Matsuda és Maruyama 2007;
Matsuda és mtsai 2013; Murck és mtsai 2007; Rudecki and Gray 2016). Ezen felül ismert
anorexigén, befolyásolja a folyadékháztartást a folyadékfelvétel fokozásával, valamint szerepet
játszik az alvás, lokomotoros aktivitás, magatartási és nocicepciós folyamatokban, illetve
mindemellett serkenti a memóriát is (Agarwal és mtsai 2005; Ahnaou és mtsai 2006; Borbély
és mtsai 2013; Gray és mtsai 2002; Matsuda és mtsai 2013; Matsuyama és mtsai 2003; Pirger
és mtsai 2010; Reglődi és mtsai 2003; Sándor és mtsai 2009). Szorongásos betegségekben a
PACAP és a PAC1 receptor szintje megemelkedik az amygdala centrális magjában. Iemolo és
munkatársai is megerősítették a PACAP szerepét stressz hatást követő magatartási és endokrin
válaszok esetében, valamint a hiányából fakadó patológiás folyamatok - mint például a

10
szorongás - kialakulásában (Iemolo és mtsai 2016). Pszihés stresszt (pl.: depresszió) követően
fokozza a táplálékfelvételt (Jiang és mtsai 2016). Stresszreakcióban való szerepét több
tanulmányban is kimutatták (Gaszner és mtsai 2012, 2016). Endokrin hatásait is bizonyították
már: szabályozza az adeno- és neurohypophysis hormontermelését, a pancreas inzulin
termelését, stimulálja a pajzsmirigy tiroxin-, a mellékvese katekolamin-szintézisét (Chen és
mtsai 1993; Isobe és mtsai 2003; Okada és mtsai 2007; Winzell és Ahrén 2007).
A nyálmirigyek kivezetőcsöveiben az abszorbciós folyamatok befolyásolásán keresztül a
nyálszekréció szabályozásában is szerepet játszik (Matoba és mtsai 2016). Mindamellett hogy
vazodilatátor és bronchodilatátor, fontos szerepe van a pulmonális vaszkuláris tónus
szabályozásában is (Lindén és mtsai 1999; Otto és mtsai 2004; Warren és mtsai 1991). A
felsoroltakon túl még számos további élettani folyamatban leírták már hatásait. A továbbiakban
a neuroprotektív hatásait fogom részletesen bemutatni.
A PACAP neuronális védő szerepe először in vitro kísérletekben, toxikus hatásokkal
(glutamát, 6-OHDA, oxidatív stressz) szemben igazolódott (Somogyvári-Vígh és mtsai 2004).
A PACAP neuroprotektív hatását számos in vivo állatkísérletben is igazolták, mint például agyi
ischaemia, agyi trauma, gerincvelő sérülés, és Parkinson kór állatkísérletes modelljeiben
(Ohtaki és mtsai 2008; Reglődi és mtsai 2011, 2015; Shioda és mtsai 2006; Shioda és
Nakamachi 2015; Somogyvári-Vigh és Reglődi 2004; Vaudry és mtsai 2009; Waschek 2002).
Az endogén és az exogén PACAP általános citoprotektív hatását számos szövetben és sejten
kimutatták. Ezen hatását apoptotikus és inflammatorikus folyamatok gátlása révén fejti ki: B-
sejt lymphoma 2 (Bcl)-2, extracelluláris szignál regulációs kináz (ERK1/2), protein kináz B
(PKB/Akt) expressziójának növelése, valamint a proapoptotikus faktorok, mint például a
kaszpáz-3, a p38 mitogén-aktiváló protein kináz (MAPK), a c-Jun N-terminális kináz (JNK),
a Bcl-2- függő sejthalál promóter (Bad), a Bcl-2-függő X-protein (Bax), a hypoxia indukált
faktor 1-alfa (HIF-1α) aktivitásának csökkentése (Moody és mtsai 2011; Reglődi és mtsai
2012b, Vaudry és mtsai 2009; D’Amico és mtsai 2015).
Erős anti-apoptotikus hatását kezdetben kisagyi szemcsesejteken és az agyalapi mirigy
adenómasejtjein is vizsgálták (Cavallaro és mtsai 1996; Chang és mtsai 1996; Oka és mtsai
1999). A PACAP apoptózist gátló hatása más sejteken is megfigyelhető, mint például a T-
limfocitákon, preovulációs tüszőkön és tüdő alveoláris sejtjein (Delgado és Ganea 2001; Lee
és mtsai 1999; Masmoudi-Kouki és mtsai 2011; Onoue és mtsai 2004). A PACAP protektív
hatását a halló- és látószervekben is igazolták (Atlasz és mtsai 2010; Németh és mtsai 2014). A
retina egy speciális szövet, mint egy „kitelepülése” a központi idegrendszernek (KIR). Emiatt
kitűnő modellként szolgál a neuronális degeneráció mérésére. A PACAP retinális

11
neuroprotektív hatását számos in vitro és in vivo kísérletben igazolták (Atlasz és mtsai 2010;
Babai és mtsai 2006; Cheng és mtsai 2014; Fábian és mtsai 2012; Mester és mtsai 2011; Rabl
és mtsai 2002; Seki és mtsai 2011; Silveira és mtsai 2002). A könny termelődésében is szerepet
játszik a könnymirigy stimulációján keresztül. PACAP génhiányos (knock out – KO) egerekben
a csökkent könnytermelődésből fakadóan, száraz szem szindrómához hasonló tüneteket
tapasztaltak (Nakamachi és mtsai 2016).
1.1.4. A PACAP fragmensei, peptidcsalád tagjai
Az alacsony biológiai hasznosíthatósága és a gyors degradációja miatt a PACAP terápiás
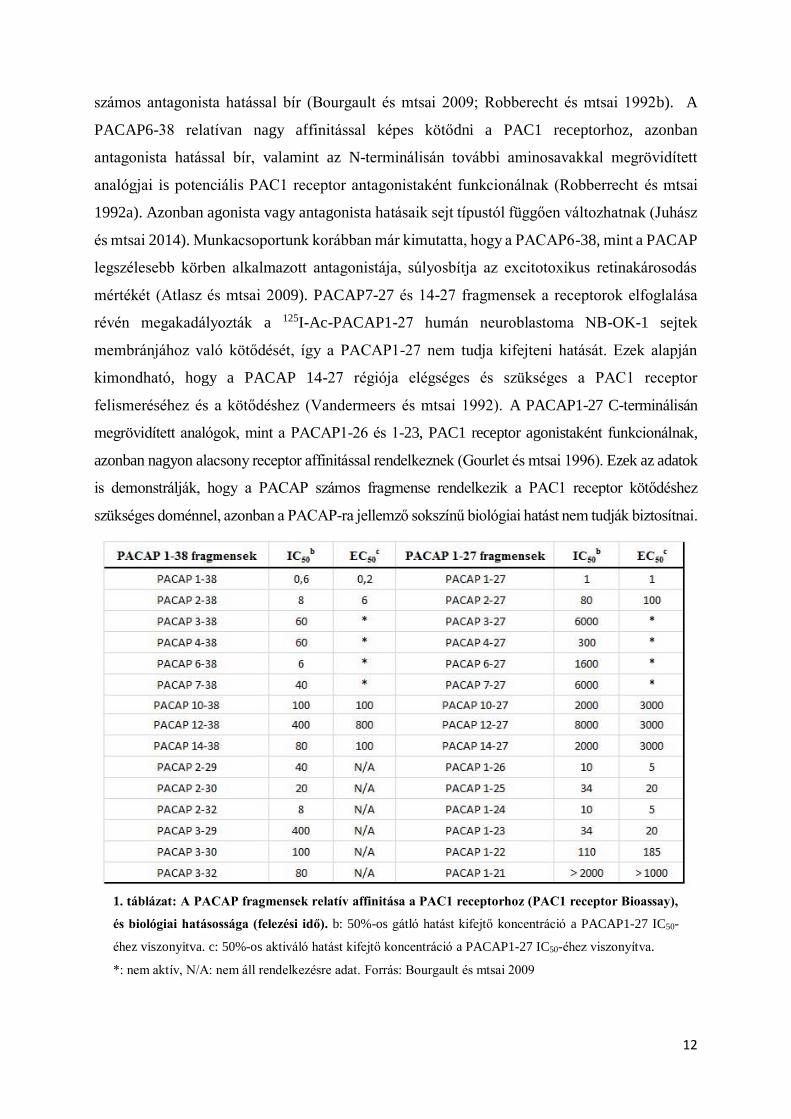
felhasználása nehezített. Emiatt rövidebb aminosavszámú fragmenseinek (1. táblázat) és a
peptidcsalád többi tagjának vizsgálata szükséges, ami a problémának egyik lehetséges
megoldása lehet. A fehérje N-terminális végéből lehasított aminosavakkal létrehozott
fragmensek általában antagonista hatással bírnak, bár néhány tanulmány agonista
tulajdonságról számol be sejt illetve szövet típustól függően (Bourgault és mtsai 2009; 2011;
Dejda és mtsai 2011a; Reglődi és mtsai 2008; Yu és mtsai 2012). A C-terminálisnál
megrövidített peptid fragmensei különböző erősséggel kötődnek a receptorokhoz (Bourgault és
mtsai 2009, 2011; Dejda és mtsai 2011b). A PACAP1-38 N-terminálisán 1-27-ig terjedő
aminosavak (PACAP1-27) konzervált szekvenciájúak (Chartel és mtsai 1991), az ezidáig
vizsgált gerincesekben megőrizték szerkezetüket: például a csirkékben (McRory és mtsai
1997), és a tokhalban (Adams és mtsai 2002). A legtöbb kísérlet szerint - azonban nem minden
esetben - a PACAP1-27 ugyanolyan biológiai hatásokkal bír, mint a PACAP1-38 (Bourgault
és mtsai 2008). PACAP1-38 intravénás (i.v.) adását követően a fehérjét a DPPIV rövid idő alatt
lebontja. Ez az enzim az N-terminális részét ismeri fel a fehérjének, azonban ez a régió
szükséges a PAC1 receptor kötődéshez is (Bourgault és mtsai 2008). A DPPIV hasítását
követően létrejött PACAP3-38 és 5-38 PAC1 receptor antagonistaként viselkednek
(Robberecht és mtsai 1992a). Egerekben vizsgálták, hogy az i.v.-an adott PACAP1-27 nem
bomlott le DPPIV hatására (Kastin és mtsai 2002). Ez arra utal, hogy a C-terminális 28-38
szekvenciája fontos szerepet játszik a PACAP-ot lebontó peptidázok interakciójában. Emiatt a
C-terminális felől megrövidített PACAP fragmensek ellenállóbbak lehetnek az enzimatikus
lebontási folyamatoknak (Bourgault és mtsai 2009). Azonban ezek a peptidek a PACAP1-38-
nál 50-szer kisebb affinitással kötődnek a PAC1 receptorhoz (Bourgault és mtsai 2009). A
PACAP1-27 és 1-38 N-terminálisát érintő deléciókat követően létrejött fragmensek közül
12
számos antagonista hatással bír (Bourgault és mtsai 2009; Robberecht és mtsai 1992b). A
PACAP6-38 relatívan nagy affinitással képes kötődni a PAC1 receptorhoz, azonban
antagonista hatással bír, valamint az N-terminálisán további aminosavakkal megrövidített
analógjai is potenciális PAC1 receptor antagonistaként funkcionálnak (Robberrecht és mtsai
1992a). Azonban agonista vagy antagonista hatásaik sejt típustól függően változhatnak (Juhász
és mtsai 2014). Munkacsoportunk korábban már kimutatta, hogy a PACAP6-38, mint a PACAP
legszélesebb körben alkalmazott antagonistája, súlyosbítja az excitotoxikus retinakárosodás
mértékét (Atlasz és mtsai 2009). PACAP7-27 és 14-27 fragmensek a receptorok elfoglalása
révén megakadályozták a 125I-Ac-PACAP1-27 humán neuroblastoma NB-OK-1 sejtek
membránjához való kötődését, így a PACAP1-27 nem tudja kifejteni hatását. Ezek alapján
kimondható, hogy a PACAP 14-27 régiója elégséges és szükséges a PAC1 receptor
felismeréséhez és a kötődéshez (Vandermeers és mtsai 1992). A PACAP1-27 C-terminálisán
megrövidített analógok, mint a PACAP1-26 és 1-23, PAC1 receptor agonistaként funkcionálnak,
azonban nagyon alacsony receptor affinitással rendelkeznek (Gourlet és mtsai 1996). Ezek az adatok
is demonstrálják, hogy a PACAP számos fragmense rendelkezik a PAC1 receptor kötődéshez
szükséges doménnel, azonban a PACAP-ra jellemző sokszínű biológiai hatást nem tudják biztosítnai.
1. táblázat: A PACAP fragmensek relatív affinitása a PAC1 receptorhoz (PAC1 receptor Bioassay),
és biológiai hatásossága (felezési idő). b: 50%-os gátló hatást kifejtő koncentráció a PACAP1-27 IC50-
éhez viszonyitva. c: 50%-os aktiváló hatást kifejtő koncentráció a PACAP1-27 IC50-éhez viszonyítva.
*: nem aktív, N/A: nem áll rendelkezésre adat. Forrás: Bourgault és mtsai 2009

13
A szekretin és glükagon peptidcsalád tagjai: glükagon, GLP1/2, szekretin, VIP, GIP,
GHRH, PHM, GIP, PACAP (1. ábra) (Sherwood és mtsai 2000). Korábban már a prekurzor
glükagon és a VIP teljes DNS-ét dekódolták. A preproglükagon és prepro-VIP mérete és
szekvenciája különböző, azonban a strukturális szerveződésük megegyezik (Bell és mtsai
1986). A peptidcsalád N-terminális doménje nagyfokú homológiát mutat (Bourgault és mtsai
2009). Munkacsoportunk korábban a VIP és a PACAP vizsgálatával foglalkozott, mostani
tanulmányomban a szekretin és glükagon retinoprotektív hatását részletezem, emiatt a
peptidcsalád ezen tagjait mutatom be részletesen az alábbiakban.
A VIP a PACAP-hoz hasonlóan megtalálható az agy számos területén: cortexben,
hippocampusban, amygdalában, suprachiasmaticus magban és a hypothalamusban. Szerepet
játszik a tanulásban, a memóriában és a kognitív folyamatokban (Acsády és mtsai 1996; Gozes
és mtsai 2008; Harmar és mtsai 2012; Loren és mtsai 1979). A VIP mindhárom receptorhoz
való kötődése révén tudja kifejteni retinoprotektív hatását, azonban a PAC1 receptorhoz
gyengébb affinitással kötődik (mintegy 1000-szer gyengébben) mint a PACAP, viszont a
VPAC1 és VPAC2 receptorokhoz ugyanolyan erősséggel képesek kötődni (Arimura és mtsai
1998; Harmar és mtsai 2012). A VIP a PACAP-hoz hasonlóan retinoprotektív hatású retinális
ischaemiát követően, azonban a PACAP hatásához hasonló mértékű protekcióhoz magasabb
dózisra van szükség (Szabadfi és mtsai 2012). A VIP fontos szerepet játszik a hippocampalis
szinaptikus transzmisszióban a VPAC1 receptoron keresztül kifejtett hatása révén (Cunha-Reis
és mtsai 2014). A PACAP1-38-at és különböző aminosavszámú fragmenseit, valamint a VIP-t
tumoros sejtvonalakon is vizsgálták, mint például retinoblastoma esetében (Wojcieszak és
Zawilska 2014).
Korábban közölt tanulmányban a VIP C-terminálisához kötött TAT peptid penetrációs
képességét tanulmányozták. Az így kapott VIP-TAT peptidet fluorescens jelölést követően
intraperitonealisan (i.p.) injektálták az egerekbe, majd a vér-agy gáton való átjutását vizsgálták.
Azt tapasztalták, hogy a VIP-TAT fehérje nagyobb mértékű protekcióra képes scopolamin
indukálta amnéziában, könnyebben penetrál a vér-agy gáton, valamint a PAC1 receptorhoz
nagyobb affinitással kötődik, mint a VIP önmagban (Yu és mtsai 2014).
A glükagon a proglükagonból alakul ki, szövetspecifikusan a pancreas α-sejtjeiben.
Hipovolémiás és hipoglikémiás stressz hatására szekretálódik. Hatására a májban elindul a
glükogenolízis és a glükoneogenezis, ami gyors vércukorszint emelkedést eredményez (Young
2005). A glükagon a szemmel is kapcsolatba hozható: Koh és Chader kimutatták, hogy számos
hormon valamint neurotranszmitter, többek között a VIP, glükagon, L-izoproterenol és a
hisztamin megemelik az intracelluláris cAMP koncentrációt retinális pigment epithelium

14
sejtkultúrában, ezzel befolyásolva a transzport folyamatokat és a fagocitózist is (Koh és Chader
1984). Egy másik tanulmányban arról számoltak be, hogy az exogén glükagon gátolja
csirkékben a myopia kialakulását, az üvegtesti tér elongációjának megakadályozása révén
(Vessey és mtsai 2005).
A szekretint kezdetben a duodenumból származó hormonnak tekintették, ami a pancreas
szekrécióját befolyásolja (Bayliss és Starling 1902). Később különböző tanulmányokban leírták
jelenlétét az agyban is (Charlton és mtsai 1981; Mutt és mtsai 1979). A szekretin és a receptora
számos helyen expresszálódik az agy különböző területén, ahol a neurotranszmisszióban,
génexpresszió szabályozásában, neurogenezisben, és neuroprotekcióban vesznek részt (Zhang
és Chow 2014). A szekretin és glükagon számos hatását és tulajdonságát korábban már
vizsgálták, azonban a retinában kifejtett esetleges protektív hatásáiról nem áll rendelkezésünkre
irodalmi adat.
1. ábra: A PACAP/VIP/secretin/glükagon peptidcsalád különböző tagjainak aminosav szekvenciája
(humán adat). Forrás: Vaudry és mtsai 2009
1.2. A retina szerkezete
A retina az agyhólyag kitüremkedő, kettős falú szemserleg származéka, emiatt különleges
differenciáltságú agyszövetnek tekinthető. A szemserleg külső fala végig egyrétegű (pars caeca
retinae), míg a belső fal hátsó 4/5-e fényérzékeny ideghártyává differenciálódott (pars optica
retinae). Kétféle neuronrendszer figyelhető meg a retinában. Az egyik a vertikális
neuronrendszer, amit a fotoreceptorsejtek, a bipoláris sejtek és a ganglionsejtek képeznek,

15
utóbbi axonjaiból szedődik össze a látóideg. A másik a horizontális neuronrendszer, melyet a
horizontális sejtek (fotoreceptor sejteket kötnek össze egymással) és az amakrin sejtek
(bipoláris sejtek között létesítenek kapcsolatot) alkotnak.
A retina külső határhártyáját (outer limiting membrane - OLM) csapok és pálcikák
beltagjainak töve, valamint a Müller glia sejtek scleralis nyúlványa alkotja. Az OLM a
fotoreceptor rétegét elhatárolja a retina mélyebb rétegeitől. Ezen kívül a vérellátásban is
egyfajta határt képez: csapok és pálcikák a ciliaris erekből a choroidea felől a pigmenthám
közvetítésével táplálkoznak, a retina mélyebb rétegeit viszont az arteria centralis retinae látja
el. A következő réteg a külső magvas réteg (outer nuclear layer - ONL), amit fotoreceptorok
perikaryonjai hoznak létre. A külső szinaptikus (rostos) rétegben (outer plexiform layer - OPL)
a csap- valamint a pálcikasejtek bipoláris sejtekkel képzett szinapszisai találhatóak. A belső
magvas rétegben (inner nuclear layer - INL) négy sejttípus perikaryonja található: bipoláris
sejtek, horizontális sejtek, amakrin sejtek, Müller sejtek. A belső szinaptikus (rostos) rétegben
(inner plexiform layer – IPL) kétféle szinapszis található, az egyik a ribbonszinapszis ahol
vertikális irányú -, a másik közönséges morfológiájú szinapszis, ahol lateralis irányú információ
áramlás történik. A ganglion sejtek rétegét (ganglion cell layer – GCL) multipoláris ganglion
sejtek képezik, dentritjeik bipoláris és amakrin sejtektől veszik át az információt, és axonjuk
elsősorban a corpus geniculatum lateraleban végződik. Kisebb részben végződnek a
nyúlványok a hypothalamus nucleus suprachiasmaticusában, a colliculus superiorban és az area
pretectalisban. A retina legnagyobb területén a ganglion sejtek egy sorban helyezkednek el. A
belső határhártya (inner limiting membrane – ILM) nem egy egységes lemez, hanem a Müller
sejtek vitralis végtalpaiból és a körülöttük lévő lamina basalisból áll. A Müller glia sejtek
hosszan elnyúlt sejtek, a két határhártyát (OLM – ILM) hidalják át és a sejtmagjuk a belső
magvas rétegben helyezkedik el. Vitrealis irányba haladó nyúlványuk talpszerű
megvastagodással hozza létre a belső határhártyát, a scleralis nyúlványuk pedig a
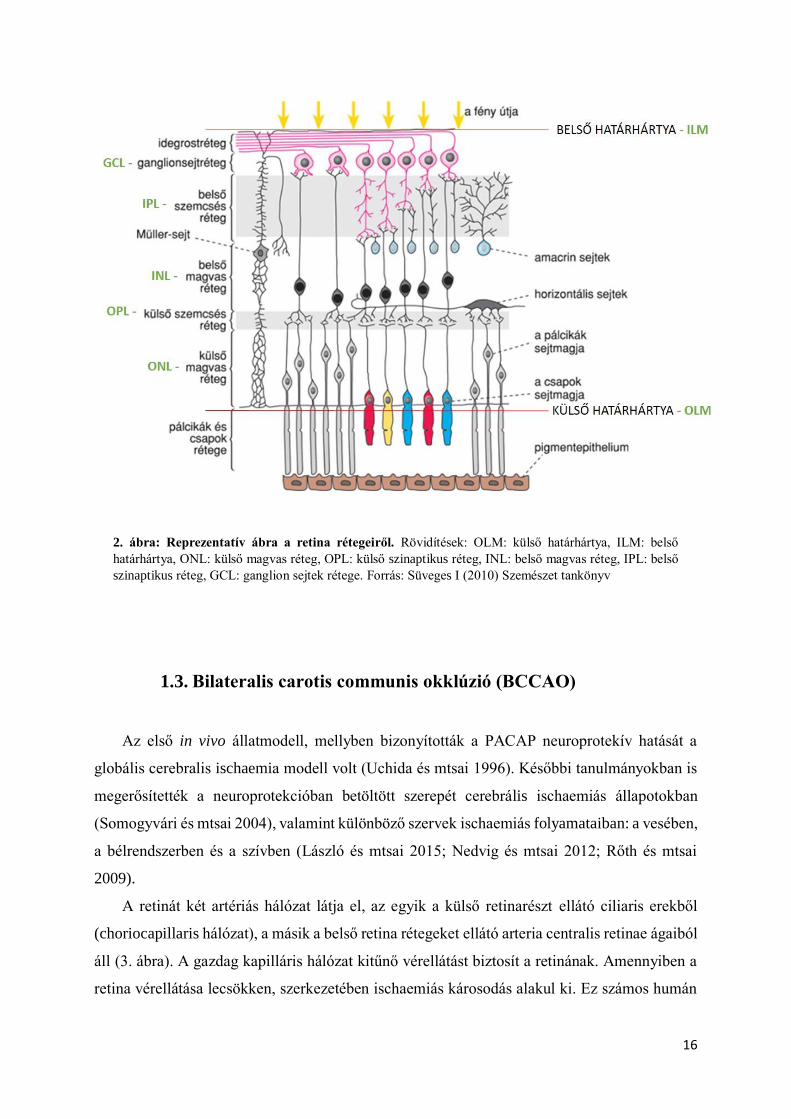
fotoreceptorok közé illeszkedik (2. ábra) (Röhlich 2006).
Nemzetközileg elfogadott a retina rétegeinek megnevezésénél az angol nómenklatúra
illetve annak rövidítése, ennek okán a továbbiakban én is ezt alkalmazom.
16
2. ábra: Reprezentatív ábra a retina rétegeiről. Rövidítések: OLM: külső határhártya, ILM: belső
határhártya, ONL: külső magvas réteg, OPL: külső szinaptikus réteg, INL: belső magvas réteg, IPL: belső
szinaptikus réteg, GCL: ganglion sejtek rétege. Forrás: Süveges I (2010) Szemészet tankönyv
1.3. Bilateralis carotis communis okklúzió (BCCAO)
Az első in vivo állatmodell, mellyben bizonyították a PACAP neuroprotekív hatását a
globális cerebralis ischaemia modell volt (Uchida és mtsai 1996). Későbbi tanulmányokban is
megerősítették a neuroprotekcióban betöltött szerepét cerebrális ischaemiás állapotokban
(Somogyvári és mtsai 2004), valamint különböző szervek ischaemiás folyamataiban: a vesében,
a bélrendszerben és a szívben (László és mtsai 2015; Nedvig és mtsai 2012; Rőth és mtsai
2009).
A retinát két artériás hálózat látja el, az egyik a külső retinarészt ellátó ciliaris erekből
(choriocapillaris hálózat), a másik a belső retina rétegeket ellátó arteria centralis retinae ágaiból
áll (3. ábra). A gazdag kapilláris hálózat kitűnő vérellátást biztosít a retinának. Amennyiben a
retina vérellátása lecsökken, szerkezetében ischaemiás károsodás alakul ki. Ez számos humán

17
retinát érintő betegségben megfigyelhető folyamat (artériás okklúzió, dibéteszes retinopathia,
stb), mely különböző mértékű látásromláshoz vezet (Chen és mtsai 2007; Harris és mtsai 2003;
Feigl és mtsai 2009; Kaur és mtsai 2008; Osborne és mtsai 2004; Pournaras és mtsai 2008).
3. ábra: A szemgolyó valamint a retina vérellátása. Forrás: Süveges I (2010) Szemészet Tankönyv
Számos állatmodell ismert retinális ischaemia előidézésére: magas intraocularis nyomás,
arteria ophthalmica lekötése, kétoldali arteria carotis communis lekötése arteria vertebralis
ligatioval, vagy anélkül (Osborne és mtsai 2004). A kétoldali arteria carotis communis lekötése
lehet időszakos illetve végleges, ami ischaemiás/reperfúziós változásokhoz vagy krónikus
retinális hipoperfúzióhoz vezet, mely a krónikus retinadegenerációk állatkísérletes
modelljeként szolgál (Szabadfi és mtsai 2010).
Kétoldali arteria carotis communis elzáródása patkányoknál csökkent agyi vérátáramlást
idéz elő (túléléshez szükséges vérmennyiséget az arteria vertebralis kollaterális keringés révén
biztosítja), az így kialakult biokémiai és magatartásbeli változások jól mérhetők. A BCCAO
következményes neuronális degenerációhoz, mikroglia aktivációhoz, asztrocitózishoz,
magatartásbeli változáshoz, valamint biokémiai paraméterek változásához vezet (Farkas és
mtsai 2005; Liu és mtsai 2005; Ni és mtsai 1994; Sarti és mtsai 2002; Schmidt-Kasner és mtsai

18
2005). A retinában karakterisztikus patológiás megjelenést okoz, hasonlóan a humán
gyógyászatban ismert arteria carotis elzáródás miatti retinopathiához (Slakter és mtsai 1984;
Spertus és mtsai 1984). Tanulmányokban elektroretinográfiás és morfológiai vizsgálati
módszerekkel is bizonyították a BCCAO okozta retinakárosodást (Block és mtsai 1992;
Osborne és mtsai 1999; Stevens és mtsai 2002; Yamamoto és mtsai 2006).
1.4. Szemcseppes terápia
Korábbi eredményeink alátámasztották a PACAP potenciális terápiás felhasználásának
lehetőségét ischaemiás károsodásokban. Az eddigi vizsgálatokban a PACAP-ot intravitreálisan
alkalmaztuk, ami az esetleges későbbi klinikai felhasználás során korlátozó tényező lehet.
Klinikai terápiás lehetőségként, az intravitreálisan bejuttatott hatóanyag elfogadott néhány
súlyos betegség esetében, ilyen például az időskori macula degeneráció bizonyos formája, és a
súlyos diabeteses retinopathia (Bandello és mtsai 2016). Mint minden invazív beavatkozásnak,
az intravitreális injektálásnak is előfordulhatnak szövődményei: megemelkedett intraokuláris
nyomás, infekció, vérzés, gyulladás és ritkán a szúrásból fakadóan bizonyos képletek (retina,
idegek) károsodása. Számos előnyét élvezhetjük a szemcseppes formának a klinikai
hétköznapok során, az intravitreális alkalmazással szemben. Erre példa az antibiotikumok
alkalmazása (általában 5 napon keresztül történik a cseppentés naponta több alkalommal), vagy
a glaukoma terápiáját képező szemcseppek adása, mely évekig tarthat.
Vizsgálatainkban a PACAP1-27 és 1-38 szemcseppes formájával kísérleteztünk, és
tanulmányoztuk, hogy a PACAP ebben a formában eljut-e a retináig, illetve a bejutott
koncentráció elégséges-e az eddig ismert retinoprotektív hatás kifejtéséhez.
Korábbi tanulmányokban hasonló módon vizsgálták az ideg növekedési faktort (nerve
growth factor – NGF). Vizsgálták, hogy az NGF intravitreális adást követően miként
befolyásolta a retinakárosodás mértékét retinitis pigmentosa-ban, illetve milyen más
növekedési faktorokat aktivált. Kimutatták, hogy az NGF befolyásolta a patkány retina
működését, valószínűleg számos növekedési faktor aktiválása révén (Lenzi és mtsai 2004). Ezt
követően az NGF lokális, szemcseppes alkalmazásáról közöltek összefoglaló tanulmányt,
melyben a corneális ulcusban, glaukómában, és a choroideában valamint a retinában kifejtett
hatásíról számoltak be (Steinle 2010).

19
Fukiage és munkatársai a PACAP ideg-regenerációban játszott szerepét vizsgálták
nyulakon. Ganglion trigeminale sejteket izoláltak nyulakból és vizsgálták a PACAP1-27 rájuk
kifejtett hatását. A PACAP1-27 hatására az ideg növekedését, regenerációját tapasztalták. A
nyulak szaruhártyáján egy speciális műtétet hajtottak végre (laser-assisted in situ keratomileusis
– LASIK), majd PACAP1-27 szemcseppes terápiát alkalmaztak. Azt tapasztalták, hogy műtétet
követően a PACAP1-27 segíti a cornea regenerálódását valamint csökkenti a szemszárazság
mértékét (Fukiage és mtsai 2007).
A PACAP topikális alkalmazásának vizsgálatára nemrég szintén sor került. A száraz szem
szindróma tüneteiként szolgáló csökkent könnytermelésre és corneális keratinizációra kifejtett
hatását vizsgálták. Kimutatták, hogy PACAP-KO egerekben a PACAP szemcseppes formában
képes fokozni a könny termelődését, valamint csökkenteni a keratinizáció mértékét (Nakamachi
és mtsai 2016).
1.5. A PACAP jellemzői a szemben
1.5.1. A PACAP előfordulása a szemben
Értekezésemben a PACAP retinoprotektív hatásai kerülnek tárgyalásra, emiatt a témával
kapcsolatos eddigi kutatási eredményeket részletesen mutatom be. A PACAP a szem számos
szövetében kimutatható, megtalálható az amakrin- és horizontális sejtekben a belső magvas
rétegben (INL), a ganglion sejtek rétegében (GCL), valamint a Müller-féle glia sejtekben is.
PACAP immunpozitív rostok detektálhatók az idegrostok rétegében (NFL) és a belső
szinaptikus rétegben (IPL), azonban nem lehet kimutatni őket a külső magvas rétegben,
valamint a pigment epitheliumban (PE) sem (D’Agata és Cavallaro 1998; Hannibal és
Fahrenkrug 2004; Izumi és mtsai 2000; Kubrusly és mtsai 2005; Seki és mtsai 2000a). A
PACAP a glutamát mellett a retinohypothalamikus pálya egyik fő transzmittere. A pálya a
ganglion sejtektől húzódik a suprachiasmaticus magba, feladata a fény diurnális ritmust
befolyásoló hatásainak közvetítése (Hannibal és Fahrenkrug 2004). A PACAP jelenlétét
kimutatták már patkány, egér (Mathis és Schaeffel 2007), hal (Grone és mtsai 2007), teknős
(Reglődi és mtsai 2001) valamint csirke (Józsa és mtsai 2001) retinákban. Emlősökben a
retinohypothalamikus pálya a retina ganglionsejtjeiből ered és a hypothalamus

20
suprachiasmatikus magjában végződik, ez tekinthető az emlősök biológiai órájának. Ezen
ganglionsejtek melanopszin tartalmuk miatt direkt fényérzékenyek. A fotoszenzitív ganglion
sejtekben kimutatható a PACAP, ahol a melanopszin expressziójának szabályozásában játszik
szerepet. Ezekben a sejtekben a PACAP expressziója dopamin kontroll alatt áll (Sakamoto és
mtsai 2005; Wan és mtsai 2006). PACAP-ot és a melanopszint tartalmazó retinális ganglion
sejtek fontos szerepet játszanak számos fény által aktiválható folyamatban. Ez azt mutatja, hogy
a glutamáterg szignáltranszdukcióban a PACAP fontos szerepet játszik (Delwig és mtsai 2013;
Purrier és mtsai 2014). A kiscsirkék fejlődése során a PACAP expressziója az INL rétegben
már az embrionális 8. napon detektálható (Borba és mtsai 2005). A PACAP és receptorai az
emlősök fejlődése során is kimutathatóak (Bagnoli és mtsai 2003).
A PACAP nemcsak a retinában, hanem a szem többi szövetében is megtalálható, mint
például az irisben, corpus ciliareban és a conjunctivában is. Jelen van a trigeminális,
sphenopalatinalis és ciliaris ganglionokban is, melyeken keresztül befolyásolja a mirigyek
szekrécióját. Munkacsoportunk bizonyította a PACAP jelenlétét a könnyben is (Gaál és mtsai
2008).
1.5.2. A PACAP receptorai a retinában
Korai tanulmányokban leírták a PACAP receptorok eloszlását, és lokalizációját a retinában.
Nilsson és munkatársai (1994) albino patkányokban vizsgálták a receptorok jellemzőit,
valamint a PACAP kötődést követő adenilát cikláz aktivitást. A PACAP/VIP receptor
expresszió varianciáját, valamint a foszfolipáz C útvonal aktivációját D’Agata és Cavallaro
vizsgálták (1998). Csirkéknél már az embrionális 6. napon kimutatták a PAC1 receptor mRNS-
ének jelenlétét (Borba és mtsai 2005). Humán foetalis retinában immunhisztokémiai
módszerekkel igazolták a PACAP és receptorainak jelenlétét. Ezt az eredményt a PACAP
receptorok mRNS-ének RT-PCR-ral történő kimutatásával is alátámasztották (Olianas és mtsai
1997).
A szelektív PACAP receptor felelős a PACAP retinális kötődésének 80%-áért (Nilsson és
mtsai 1994). Újszülött patkány retinájának minden rétegéből izolálták a PAC1 receptor mRNS-
ét (Silveira és mtsai 2002). Pontos lokalizációra irányuló vizsgálatok szerint patkány retinákban
különösen erős PAC1 receptor sűrűség észlelhető a NFL, INL és GCL rétegben, míg kevesebb
receptor mutatható ki az IPL, OPL, ONL rétegekben valamint a fotoreceptorok külső
szegmenseiben (Seki és mtsai 1997, 2000b). Különböző PAC1 receptor mRNS variánsokat

21
találtak, a ganglion és amakrin sejtek testében valamint a nyúlványaikban (Seki és mtsai 1997).
Müller gliasejt sejttenyészetben is igazolták PAC1 receptorok jelenlétét (Kubrusly és mtsai
2005; Seki és mtsai 2006).
A retina pigmentepithel sejtjeiben mindegyik PACAP receptor mRNS-ét detektálták
(Zhang és mtsai 2005). Más vizsgálatok is megerősítették a VPAC receptorok jelenlétét a
retinában (D’Agata és Cavallaro 1998). Humán retinoblastoma sejtek is tartalmaznak PACAP
receptorokat (Olianas és mtsai 1996).
1.5.3. A PACAP hatásai a retinában
Patkányban, nyúlban, sertésben valamint borjúban is megfigyelték a PACAP1-27 és 1-38
adenilát cikláz aktiváló hatását, és a 38 aminosavból álló forma hatékonyabbnak bizonyult, mint
a 27-ből álló (Onali és Olianas 1994). A retinában a PACAP1-27 és 38 a cAMP expresszióját
hozzávetőleg ötszörösére fokozza (Nilsson és mtsai 1994). Fejlődő patkány, csirke és humán
retinában a PACAP1-27 és 1-38 nemcsak a cAMP képződést fokozza, hanem az inozitol-
monofoszfát (inositol-monophosphate - IMP) szintet is megemeli (Borba és mtsai 2005;
D’Agata és Cavallaro 1998; Olianas és mtsai 1997; Silveira és mtsai 2002). A cAMP válasz
függő transzkripciós fehérjét (cAMP response element binding protein - CREB) az
intracelluláris cAMP aktiválja. PACAP-al kezelt újszülött patkánynál a CREB foszforilációját
már a kezelést követő 5. percben detektálni lehet (Silveira és mtsai 2002). Két-három nappal a
PACAP adását követően IL-6 expresszió figyelhető meg a retina egész területén (Nakatani és
mtsai 2006). A Müller glia sejteken is megtalálható a PAC1 receptor, itt a PACAP már kis
koncentrációban (pmol) is aktivációt okoz (Kubrusly és mtsai 2005; Nakatani és mtsai 2006;
Seki és mtsai 2006).
A PACAP szerepet játszik a retina fejlődésében is. Fejlődő csirke retinájában PACAP
kezelés hatására fokozódik a tyrosin hidroxiláz (tyrosin hydroxilase - TH) aktivitása, mely a
dopaminerg idegsejtek fejlődésére utal (Borba és mtsai 2005). Lang és munkatársai (2010)
kimutatták PAC1 transzgenikus egereken, hogy a PACAP által elindított jelátviteli útvonalak
szükségesek a retina és a vizuális funkció fejlődéséhez.
Glutamáttal kölcsönhatásba lépve fontos szerepet játszik a PACAP a fényinger
elvezetésében és a cirkadián ritmus szabályozásában (Hannibal és Fahrenkrug 2004). A corpus
pinealéban található pinealocytákban is kimutatták, hogy a PACAP fokozza a melatonin

22
termelést (Csernus és mtsai 2004; Faluhelyi és mtsai 2004; Nagy és Csernus 2007; Rácz és
mtsai 2008; Rékási és Czömpöly 2002).
A retina vérellátását a kapillárisok körüli pericyták kontraktilitása befolyásolja. A PACAP
a protein kináz A, valamint a foszfolipáz C mediálta útvonalakon a pericyták relaxációjához
vezet (Markhotina 2007). A PACAP1-27 és 1-38 hatással van a choroidea vérellátására is,
csökkentve az uveális vaszkuláris rezisztenciát, és ezáltal fokozva a vérátáramlást (Nilsson
1994).
1.5.4. A PACAP hatása ischaemiás retinopathiában
Számos humán gyógyászatban előforduló retinadegeneráció állatkísérletes modelljéül
szolgálhatnak az ischaemiás/reperfúziós károsodást, illetve a krónikus hipoperfúziót előidéző
modellek. A kétoldali arteria carotis communis lekötése ischaemiás károsodást okoz az agyban,
ami a neurológiai cerebrovaszkuláris betegségek (Fehér és mtsai 2011) háttérfolyamatainak
vizsgálatára is alkalmas lehet, emellett a modell a retinális ischaemiás folyamatokat is jól
reprezentálja. További lehetőség a mesterségesen megemelt intraoculáris nyomás okozta
ichemiás ideghártya károsodás, ami a klinikai gyakorlatban előforduló glaukóma modelljéül
szolgálhat (Szabadfi és mtsai 2010). A PACAP retinoprotektív hatása mindegyik ischaemiás
modellben igazolódott.
Az emberi test szövetei közül a retina az egyik legérzékenyebb hipoxiára. Számos kóros
elváltozás előidézheti a retina oxigénhiányos állapotát: artériás elzáródás, diabeteses
retinopathia, magas intraocularis nyomás. Permanens kétoldali carotis communis elzáródás is
csökkent agyi vérátáramláshoz vezet, mely következményesen retinadegenerációt idéz elő
(Atlasz és mtsai 2007; Mester és mtsai 2009). Kutatócsoportunk korábbi tanulmányokban
igazolta az intravitreálisan beadott PACAP retinoprotektív hatását. A BCCAO jelentős
károsodást idézett elő a retina összes rétegében. Kontroll retinákhoz hasonlítva, két héttel a
carotis lekötést követően, a szinaptikus rétegvastagságok csökkenése volt tapasztalható, ami a
külső- és belső határhártya (OLM-ILM) közötti távolság csökkenését vonta maga után (Atlasz
és mtsai 2007). Műtétet követően, intravitreálisan beadott PACAP1-38 jelentős protektív
hatással bírt. A retina rétegei a kontroll állatokéhoz hasonlóak voltak, ami morfometriai
vizsgálatokkal is igazolódott (Atlasz és mtsai 2007, 2010). Újabb tanulmányok is
megerősítették korábbi eredményeinket, Seki és munkatársai (2011) magas intraocularis

23
nyomás okozta ischaemiás retinopathiában vizsgálták a PACAP szerepét. Intravitreális beadást
követően a PACAP citoprotektív hatását tapasztalták a ganglion sejtek rétegében.
A molekuláris mechanizmusok feltárására irányuló korábbi kísérleteinkben a MAP kináz,
és Akt, valamint a különböző citokinek aktivitásbeli változásait vizsgáltuk (Szabó és mtsai
2012). A PACAP kezelés önmagában nem növelte az Akt és MAP kináz foszforilációját. A
PACAP hipoperfúziós károsodást követően a p38MAP kináz és JNK aktivációját csökkentette,
a protektív Akt valamint ERK1/2 foszforilációt viszont megnövelte (Szabó és mtsai 2012).
BCCAO-t követően a citokin profilban számos változás következett be. Néhány citokin és
kemokin expressziója megnőtt, mint például: CINC, CNTF, fractalkine, sICAM, IL-1, LIX,
MIP-1, RANTES és a TIMP-1. PACAP kezelés hatására az említett fehérjék aktivitása
csökkent. A PACAP megemelte a vaszkuláris endoteliális növekedési faktor valamint a thymus
kemokin szintjét (Szabó és mtsai 2012).
Munkacsoportunk az eddig tapasztalt morfológiai változásokat funkcionális vizsgálatokkal
(elektroretinográfia - ERG) is alátámasztotta (Dányádi és mtsai 2014). Műtétet követő második
illetve tizennegyedik napon elvégzett ERG vizsgálat eredményeit értékelték. BCCAO-t
követően, intravitreálisan PACAP-al kezelt szemek ERG görbéi a kontrollhoz hasonlóak
voltak, szemben a szintén érlekötésen átesett, fiziológiás sóoldatot kapott szemekkel. A
posztoperatív 14. napon a PACAP kezelés szignifikáns mértékben csökkentette az ischaemiás
károsodásból fakadó elváltozásokat. Az a- és b-hullám amplitúdója - az ischaemiás károsodást
követően - lecsökkent, ennek ütemét a PACAP szignifikáns mértékben csökkentette. Az öt
egymást követő oszcillációs potenciál közben eltelt idő a fiziológiás sóoldatot kapott
szemekben szignifikáns mértékben lecsökkent, míg PACAP kezelés hatására nem csökkent
ilyen ütemben (Dányádi és mtsai 2014).
D’Alessandro és munkatársai nátrium-aziddal előidézett, ex vivo ischaemiás károsodást,
valamint a károsodást követő PACAP kezelés metabolikus változásait vizsgálták. Többek
között azt tapasztalták, hogy a PACAP megakadályozta a pro-inflammatorikus faktorok és
peroxidálódott lipidek akkumulációját. Például ischaemiás károsodást követően 3 nitrát-oxid
metabolit szintjének emelkedését is leírták: arginin, ornithin és citrullin. A PACAP kezelés
visszafordította ezeket a változásokat. A PACAP ezen kívül csökkentette az α-ketoglutarát
szintjét is, ami az ischaemiás károsodást követő glutamát bioszintézisében játszik szerepet
(D’Alessandro és mtsai 2014).

24
2. Célkitűzések összefoglalása
Korábbi tanulmányokban bebizonyították, hogy a PACAP1-38 és 1-27 azonos mértékű
protekciót idéz elő. Azonban a kisebb aminosavszámú fragmenseknél és a peptidcsalád többi
tagjánál (szekretin és glükagon) eddig nem vizsgálták az esetleges retinoprotektív hatást. Az
eddigi tanulmányokban a retinális folyamatok tekintetében a PACAP6-38 és 6-27 antagonista
szerepe nyert bizonyítást (Atlasz és mtsai 2009).
1. Vizsgálataink egyik felében célul tűztük ki a PACAP kisebb aminosavszámú
fragmenseinek (PACAP 4-13, 4-22, 6-10, 6-15, 11-15, 20-31), valamint a PACAP
peptid családjába tartozó szekretin és glükagon retinoprotektív hatásának vizsgálatát
ischaemiás retinopathiában.
2. Arra kerestük a választ, hogy a permanens bilaterális carotis communis lekötést
követően a szemcseppes forma (mely könnyebb klinikai felhasználást tenne lehetővé)
képes-e az eddig ismert retinoprotektív hatás kifejtésére, mind a PACAP1-27 mind az
1-38 alkalmazása során.

25
3. Szekretin, glükagon és különböző PACAP fragmensek
vizsgálata ischaemiás retinopathiában
3.1. Anyagok és módszerek
3.1.1. Az arteria carotis communis permanens, bilaterális lekötésével
létrehozott retinadegenerációs modell
Kísérleteink során 3-4 hónapos Wistar patkányokat (sham n=7, BCCAO n=32)
vizsgáltunk, melyeket standard laboratóriumi körülmények között, igény szerinti táplálék- és
folyadékellátás mellett tartottunk 12 órás világos-sötét ciklusban. Az állatok elhelyezését,
gondozását és a kísérletek kivitelezését a jelenleg érvényes etikai szabályoknak és az egyetemi
protokollnak megfelelően végeztük (BA02/2000-31/2011, Pécsi Tudományegyetem).
Izoflurános altatás alatt, centrális nyaki metszést követően, mindkét oldali arteria carotis
communist 3-0-s fonállal permanensen lekötöttük (BCCAO) (4. ábra). Ezáltal agyi hipoxiát
idéztünk elő, ami ischaemiás retinakárosodáshoz vezetett. A műtét végén a sebet zártuk, és ezt
követően a szemeket operációs mikroszkóp alatt intravitreálisan kezeltük. A corneán ejtett
metszést követően a jobb szembe 30 G Hamilton tűvel intravitreálisan – operációs
mikroszkóppal kontrollálva - PACAP1-38-at, illetve különböző PACAP fragmenseket
(PACAP4-13, 4-22, 6-10, 6-15, 11-15, 20-31), szekretint valamint glükagont juttattunk,
mindegyiket 100pM/5µl koncentrációban. Az ischaemiás kontrollt képező bal szemekbe
intravitreálisan azonos mennyiségű fiziológiás sóoldatot juttattunk. Az áloperált állatok (sham)
képezték az intakt kontrollt: altatást követően nyaki metszést végeztünk, majd sebüket bezártuk.
A sham csoportba tartozó patkányokat is az ischaemiás állatokkal megegyező intravitreális
terápiában részesítettük.

26
4. ábra: Bilateralis carotis communis okklúzió műtét során készült képek. Az első képen a nyakizmok
láthatóak, a másodikon a csipesz felett az arteria carotis communis, a harmadik kép pedig a lekötést követő
pillanatot ábrázolja.
3.1.2. Rutin morfológiai és morfometriai analízis
Két héttel a műtétet követően az állatokat túlaltattuk, majd a szemeken rutin szövettani
vizsgálatot, valamint morfometriai analízist végeztünk. Az enukleálást követően a szemeket
fixáló oldatba 4%-os paraformaldehid (PFA) 0,1M-os foszfát pufferbe oldva (PB) (Sigma-
Aldrich, Budapest) helyeztük. A szövettani protokollt a korábbi tanulmányaink alapján
alkalmaztuk: 3 napos gyantás beágyazást követően Durcupan ACM (Sigma-Aldrich,
Budapest), az elkészült gyanta blokkokból ultramikrotómmal 2µm-es retinametszeteket
készítettünk, melyeket tárgylemezre téve toluidin kékkel (Sigma-Aldrich, Budapest) festettünk,
majd lefedtünk. A blokkok metszése során a viszonyítási pontot a nervus opticus jelentette
(Atlasz és mtsai 2007). Ezt követően a tárgylemezen lévő metszeteket lefotóztuk, és a retina
rétegeinek vastagságát lemértük, majd táblázatban összesítettük (Microsoft Excel szoftver
segítségével). A retina rétegei közül a következőket mértük: külső határhártya és a belső
határhártya közötti távolságot, a külső magvas réteget, a külső szinaptikus réteget, a belső
magvas réteget, a belső szinaptikus réteget, valamint 100µm-es retinahosszon a ganglionsejtek
rétegében a sejtek számát. Morfometriai vizsgálataink során kapott eredményeink értékeléséhez
egy-utas ANOVA tesztet használtunk, amit Tukey-B féle post hoc analízis követett.

27
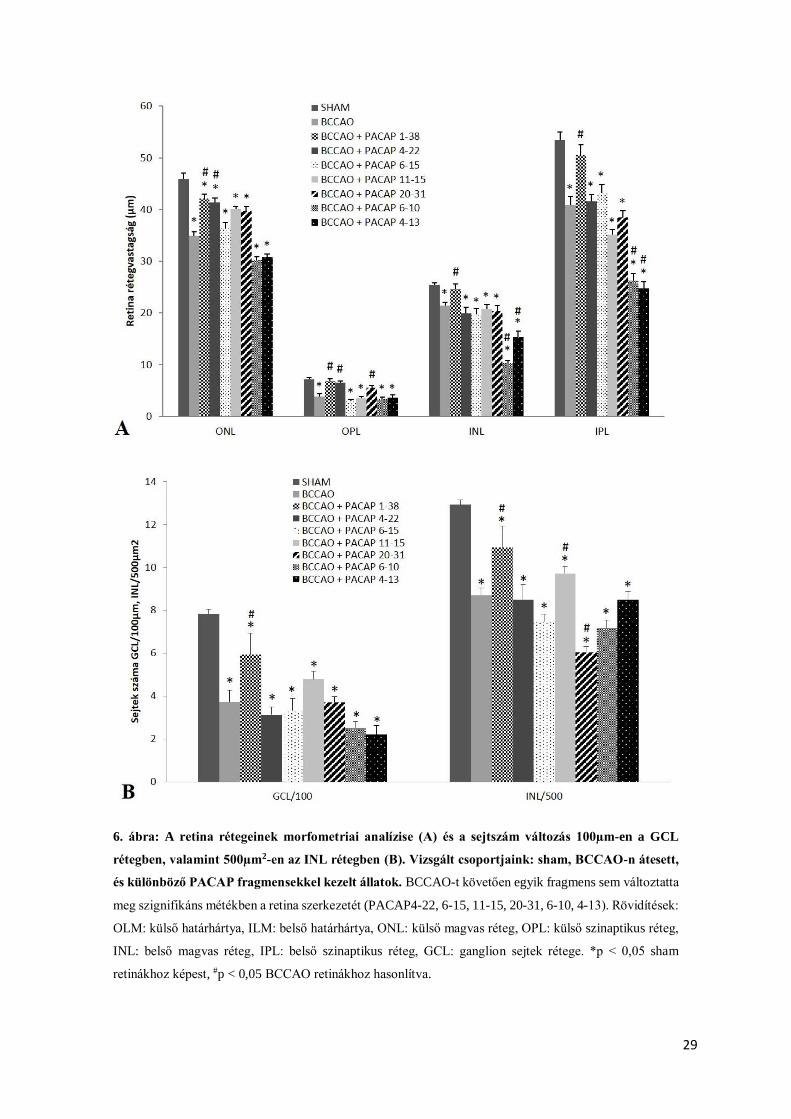
3.2. Eredményeink
A BCCAO által előidézet retinakárosodás mértéke a retinát alkotó különböző rétegek
vastagságának csökkenésében nyilvánult meg. A mért értékeket a sham operált állat retinájához
viszonyítottuk (5. ábra). A károsodás mértéke rétegenként eltérő volt (6/A. ábra). A legnagyobb
fokú rétegvastagság csökkenést az OPL rétegben tapasztaltuk (6/A. ábra). BCCAO-t követően
a fotoreceptorok rétegének vastagságában is csökkenést figyeltünk meg (5/B. ábra). Ez a réteg
szignifikánsan vékonyabb volt a sham retinákhoz képest (6/A. ábra).
A műtétet követően a ganglion sejtek rétegében nagymértékű sejtveszteséget
tapasztaltunk (5. ábra). Ezt a károsodást jól tükrözte a sejtszám csökkenés 100µm
retinahosszon a GCL rétegben, valamint 500µm2-en az INL rétegben (6/B. ábra). A műtét
hatására kialakult károsodás mértékét az intravitreálisan beadott PACAP1-38 jelentősen
csökkentette (5/C. ábra), melyet a morfometriai analízis eredménye is alátámasztott (6/A.
ábra). A PACAP1-38 kezelést követően a retinarétegek vastagsága hasonló volt a sham
állatoknál mért értékekhez (5.,6/A. ábra.). Szignifikánsan nagyobb volt a retina különböző
rétegeinek vastagsága az intravitreálisan PACAP1-38-al kezelt szemek esetében, a műtéten
átesett kontroll retinákhoz képest (intravitreálisan fiziológiás sóoldattal kezelt bal szemek) (6/A.
ábra). Legnagyobb mértékű különbséget az OPL rétegben tapasztaltunk, azonban az ONL és
INL rétegvastagságban is számottevő különbség mutatkozott a fentebb említett két csoport
között (6/A. ábra). A carotis lekötésen átesett állatok retinái esetében a sejtszám csökkenés a
GCL, valamint INL rétegekben szignifikáns mértékű volt a sham állatokhoz képest (6/B. ábra).
A metszeteken számos károsodott sejt volt látható, amely a sejtszám csökkenésének
magyarázatául szolgálhat.
Intravitreálisan beadott PACAP fragmensek (PACAP4-13, 4-22, 6-10, 6-15, 11-15, 20-
31) nem változtatták meg szignifikáns mértékben a retina szerkezetét (5/D-I. ábra). Ezeknél a
retináknál a szövettani képeken jól látható volt a sejtes (INL, ONL), valamint szinaptikus (IPL,
OPL) rétegek vastagságának csökkenése (5/D-I. ábra), hasonlóan az ischaemiás kontroll
retinákhoz (5/B. ábra). A sham állathoz viszonyítva a PACAP fragmens terápiában részesült
retinák esetében a sejtszám is szignifikánsan csökkent a GCL és INL rétegekben (6/A. ábra).
Az intravitreálisan beadott PACAP6-10 és 4-13 esetében nagyobb mértékű károsodás volt
megfigyelhető az INL és IPL rétegekben az ischaemiás kontrollhoz képest (5/I., H. ábra).

28
5. ábra: Reprezentatív fénymikroszkópos felvételek a retina rétegeiről (toluidine kék festés). A sham
(A) illetve a BCCAO-n (B) átesett állatok retinametszetei láthatók. Reprezentatív retina keresztmetszetek
láthatók a műtéten átesett, intravitreálisan PACAP1-38-al (C), valamint különböző fragmenseivel:
PACAP4-22 (D), 6-15 (E), 11-15 (F), 20-31 (G), 6-10 (H), 4-13 (I) kezelt szemeknek. A PACAP
különböző fragmensei (C-F) nem befolyásolták szignifikánsan az ischaemia okozta retinakárosodás
mértékét. Némelyik fragmens esetében (PACAP6-10, 4-13) azt a tendenciát tapasztaltuk, hogy fokozták a
károsodás mértékét (antagonista hatás), ami a rétegvastagság, valamint az ONL és INL rétegben taláható
sejtek számának csökkenésén mutatkozott meg. (lépték: 20µm)
29
6. ábra: A retina rétegeinek morfometriai analízise (A) és a sejtszám változás 100µm-en a GCL
rétegben, valamint 500µm2-en az INL rétegben (B). Vizsgált csoportjaink: sham, BCCAO-n átesett,
és különböző PACAP fragmensekkel kezelt állatok. BCCAO-t követően egyik fragmens sem változtatta
meg szignifikáns métékben a retina szerkezetét (PACAP4-22, 6-15, 11-15, 20-31, 6-10, 4-13). Rövidítések:
OLM: külső határhártya, ILM: belső határhártya, ONL: külső magvas réteg, OPL: külső szinaptikus réteg,
INL: belső magvas réteg, IPL: belső szinaptikus réteg, GCL: ganglion sejtek rétege. *p < 0,05 sham
retinákhoz képest, #p < 0,05 BCCAO retinákhoz hasonlítva.

30
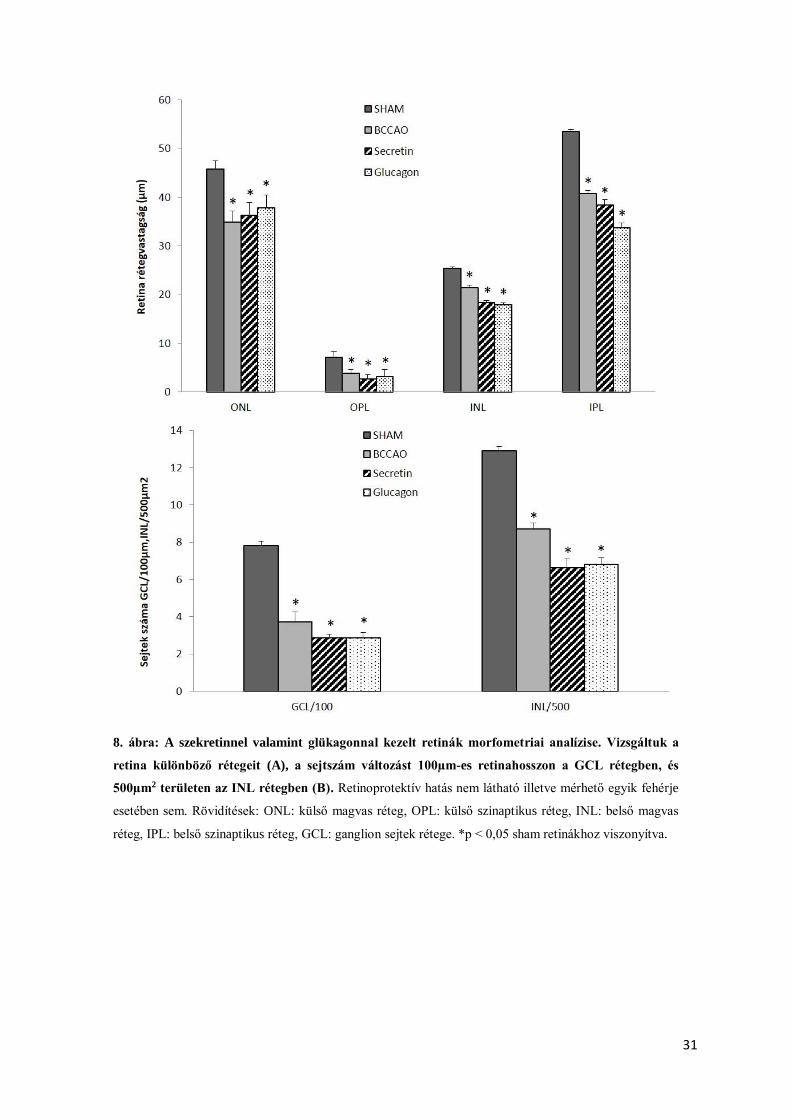
A PACAP családjába tartozó szekretin és glükagon sem bizonyult retinoprotektívnek
ischaemiás retinopathiában. A BCCAO okozta retina rétegvastagság csökkenése hasonló
mértékű volt az intravitreálisan szekretinnel vagy glükagonnal kezelt retinákéhoz (7. ábra). Ezt
az eredményt a morfometriai analízis is alátámasztotta (8. ábra). A szekretinnel és glükagonnal
kezelt retinák esetében a sejtszám csökkenés is szignifikáns mértékű volt a GCL és INL
rétegekben a sham állatokhoz viszonyítva (8. ábra).
7. ábra: Fénymikroszkópos felvétel a retina rétegeiről (toluidin kék festés), BCCAO + szekretinnel
(A) és BCCAO + glükagonnal (B) kezelt retinákból. Egyik esetben sem bizonyosodott be retinoprotektív
hatás. (lépték: 20µm)
31
8. ábra: A szekretinnel valamint glükagonnal kezelt retinák morfometriai analízise. Vizsgáltuk a
retina különböző rétegeit (A), a sejtszám változást 100µm-es retinahosszon a GCL rétegben, és
500µm2 területen az INL rétegben (B). Retinoprotektív hatás nem látható illetve mérhető egyik fehérje
esetében sem. Rövidítések: ONL: külső magvas réteg, OPL: külső szinaptikus réteg, INL: belső magvas
réteg, IPL: belső szinaptikus réteg, GCL: ganglion sejtek rétege. *p < 0,05 sham retinákhoz viszonyítva.

32
4. Szemcseppként alkalmazott PACAP1-27 és 1-38 vizsgálata
ischaemiás retinopathiában
4.1. Anyag és módszer
4.1.1. Bilaterális carotis communis műtétet követő szemcseppes terápia
Kísérleteinkben 3-4 hónapos hím Wistar patkányokat (Ʃn=130: n=43 szövettani
analízis, n=11 immunhisztokémiai analízis, n=20 immuno-blot analízis, n=28 citokin array,
n=28 elektroretinográfiás vizsgálat) vizsgáltunk, melyeket standard laboratóriumi körülmények
között, igény szerinti táplálék- és folyadékellátás mellett tartottunk 12 órás világos-sötét
ciklusban. Az állatok elhelyezését, gondozását és a kísérletek kivitelezését a jelenleg érvényes
etikai szabályoknak és az egyetemi protokollnak megfelelően végeztük (BA02/2000-31/2011,
Pécsi Tudományegyetem). A fentebb már részletezett BCCAO-s műtéti eljárást alkalmaztuk
ebben a kísérletben is. A műtétet követően az állatok az első szemcseppes terápiában
részesültek. Az áloperált állatok (sham) képezték a kontroll csoportot, ők is ugyanúgy
részesültek szemcseppes kezelésben. A szemcsepp 400µl vivőanyagban feloldott 10µg
PACAP1-27-et tartalmazott (1µg/csepp). Vivőanyagként fiziológiás sóoldatot, aqua ad
iniectabilat, solvens viscosa pro oculoguttis cum thiomersalot és solutio ophthalmica cum
benzalkoniot (0,005%-os oldat) alkalmaztunk. Az állatok jobb szemét az éppen aktuális
vivőanyagban feloldott PACAP1-27-el kezeltük, míg a bal szemükbe az aktuálisan alkalmazott
vivőanyagot cseppentettük. Kezelésünk öt napig tartott, naponta kétszer cseppentettünk,
délelőtt és délután ugyanabban az időintervallumban (9. ábra). A cseppentések alatt az állatokat
izofluránnal bebódítottuk.
Kapott eredményeink birtokában ugyanezt a műtéti eljárást alkalmazva, a kísérletet
solutio ophthalmica cum benzalkonioban oldott PACAP1-38 szemcseppel (400µl
vivőanyagban oldott 10µg PACAP1-38, 1µg/csepp) is megismételtük. Az állatokat a fent leírt
módon kezeltük.

33
9. ábra: A szemcseppes kezelés sémás ábrája. A kék cseppek a napi kétszeri szemcseppes kezelést
jelképezik, a nyilak az adott vizsgálatok időpontjait mutatják. Számokkal a műtétet követő napokat jeleztük.
4.1.2. A PACAP1-27 és 1-38 szemcseppel kezelt retinák hisztológiai analízise
A szövettani vizsgálatokhoz 43 állatot használtunk, melyből 10 képezte a sham
csoportot, míg 33 permanens kétoldali carotis lekötésen esett át (PACAP1-27: n=7 sham, n=24
BCCAO; PACAP1-38: n=3 sham, n=9 BCCAO). Két héttel a műtét után az állatokat túlaltattuk,
és szemük eltávolítását követően a szemserleget kipreparáltuk, majd 4%-os
paraformaldehidben (PFA) fixáltuk. A szövettani protokollt a fent már részletesen leírt módszer
szerint végeztük. A retina rétegeinek vastagságát lemértük, majd táblázatban összesítettük
(Microsoft Excel szoftver segítségével), és diagramokon ábrázoltuk. A retina rétegei közül a
következőket mértük: a külső és a belső határhártya távolságát, a külső magvas réteget, a külső
szinaptikus réteget, a belső magvas réteget, a belső szinaptikus réteget, valamint 100µm-es
retinahosszon, a ganglionsejtek rétegében a sejtek számát. Eredményeink értékelésénél, a
különbségek szignifikanciájának meghatározásához a megfelelő vivőanyag megtalálásáig egy-
utas ANOVA tesztet használtunk. Később, a leghatásosabb vivőanyaggal végzett további
vizsgálatokhoz kétutas ANOVA tesztet alkalmaztunk, amit Fischer féle post hoc analízis
követett.

34
4.1.3. PACAP1-27 és 1-38 radioaktív jelölése
Ezt a vizsgálatot amerikai kollaborációban végeztük William A. Banks, és Therese S.
Salameh segítségével (Education, and Clinical Center, Veterans Affairs Puget Sound Health
Care System, Seattle, WA, USA; Division of Gerontology and Geriatric Medicine, Department
of Medicine, University of Washington School of Medicine, Seattle, WA, USA). A PACAP1-27-et
valamint 1-38-at 125-ös jódizotóppal (125I) jelöltünk, laktoperoxidáz módszert alkalmazva
(Nonaka és mtsai 1983). Először 10µg PACAP1-27-et és 1-38-at (0,25M PB-ben oldva), 30µl
0,4M-os Na-acetáttal (pH 5,6) és 10µl laktoperoxidázzal (10µg/ml oldva 0,1M Na-acetátban,
pH 5,6), valamint 1mCi 125I-al elegyítettük, és további vizsgálatainkhoz ezt alkalmaztuk. A
reakciót 10µl 30%-os hidrogén-peroxid (H2O2) hozzáadásával indítottuk el. Tíz perc elteltével
újabb 10µl 30%-os H2O2 –ot adtunk, és újabb 10 percet vártunk, majd az így létrejött oldatot
high-performance liquid chromatography (HPLC, Shimadzu USA Manufacturing Inc.,
Columbia, MD) segítségével megtisztítottuk, melyhez C18 oszlopot (P.J. Cobert Associates,
Inc., St. Louis, MO) használtunk.
4.1.4. Gliális fibrilláris savas protein expressziójának mérése PACAP1-27 és
1-38 szemcseppes kezelést követően
Két héttel a műtétet követően 4 sham - és 7 BCCAO-n átesett állat (PACAP1-27: n=2
sham, n=5 BCCAO; PACAP1-38: n=2 sham, n=2 BCCAO) szemeit eltávolítottuk, majd a
szemserlegeiket kipreparáltuk, és felszálló cukoroldatba helyeztük őket. Óránként egyre
töményebb koncentrációjú szacharóz oldatba (szacharóz + PBS) tettük a szemserlegeket: 10%-
os, 20%-os, végül éjszakára 30%-os cukros oldatba kerültek. A preparátumokat másnap a
kriosztáthoz szükséges médiumba (Tissue-Tek OCT Compound, Sakura Finetek) tettük, majd
10 perc alatt lefagyasztottuk. Ezt követően a mintát lemetszettük, és a 10-12µm-es metszeteket
zselatinozott tárgylemezre helyeztük. Felhasználásig -20 ◦C-on tároltuk. Ezt követően kezdtük
el a 2 napos immunhisztokémiai protokollt. Első nap a tárgylemezekre fiziológiás sóoldatban
oldott foszfát puffert (PBS, Sigma-Aldrich, Budapest) cseppentve mostuk őket, 3 x 10 percen
keresztül. Egy órára nedves kamrába tettük a blokkolóval átmosott metszeteket. A blokkoló
oldat 3%-os szamár szérumot tartalmazott. Ezt követően 3 x 10 percig mostuk a metszeteket
PBS-sel, majd a 3%-os szamár szérumot tartalmazó antitest hígító oldatba tettük a GFAP-ra

35
specifikus primer antitestünket (nyúl anti-GFAP, 1:1000, Sigma-Aldrich, Budapest), és
éjszakára nedves kamrában inkubáltuk a metszeteket. A protokoll szerint, másnap 3 x 10 perc
PBS-es mosást követően, 2 órára a primer antitestre specifikus szekunder fluorescens antitestet
(1:200, szamár, anti-nyúl antitest, Alexa Fluor „488” (Life Technologies, Budapest) tartalmazó
oldattal, sötét helyen inkubáltuk a retinametszeteket. Kétszer 5 perc PBS mosást követően
propidium jodidot (PI, 1:500, Sigma-Aldrich, Budapest) tartalmazó oldatot cseppentettünk
rájuk 1 percig, majd ismét 2 x 5 percig mostuk őket PBS-el, végül Fluoromount-G géllel
(Sigma-Aldrich, Budapest) lefedtük. A fentebb leírt antitestek alkalmazásával
retinametszeteinket kétlépcsős immunhisztokémiai eljárásnak vetettük alá. Metszeteinket
fluorescens mikroszkóp (Nikon Eclipse Ci) segítségével tettük láthatóvá, illetve fotózhatóvá.
4.1.5. Western blot analízis, BCCAO-t követő PACAP1-27 szemcseppes
kezelés után
Két héttel a műtétet követően az állatok retináin western blot vizsgálatot végeztünk (3
sham-es és 9 BCCAO-n átesett állat). Eredményeink alapján 2 héttel a műtétet követően, nem
volt mérhető különbség a kezelt és nem kezelt csoportok között. Emiatt a kísérletet
újraterveztük, és a retinákat nem két héttel a műtétet követően, hanem 24 óra elteltével
preparáltuk ki (4 sham operált és 4 műtéten átesett állat). A rövidebb kezelési időintervallum
miatt a cseppentések számát arányaiban megnöveltük: 4 x 1 csepp az első nap, másnap a szemek
eltávolítása előtt 2 x 1 csepp. Mindkét esetben (2 hét illetve 24 órával a műtétet követően) az
állatokat túlaltattuk, majd szemeiket eltávolítottuk, és retináikat kipreparáltuk, majd folyékony
nitrogénben azonnal lefagyasztottuk, végül -80 ◦C –on tároltuk. A minták előkészítése a western
blot analízishez munkacsoportunk által már korábban leírtak szerint történt (Szabó és mtsai
2012). Ezt követően a mintákat nitrocellulóz membránon 4 ◦C-on inkubáltuk a primer
antitestekkel: anti-totál Akt (anti-tAkt, 1:1000; R&D Systems, Budapest), foszfospecifikus
anti-Akt (1:500; Cell Signaling technology, Danvers, USA), foszfospecifikus anti-
extracelluláris szignál regulációs kináz 1/2 (ERK1/2, 1:1000; Cell Signaling Technology,
Danvers, Massachusetts, USA), anti-aktin (1:10000; Sigma-Aldrich, Budapest) antitestek. A
foszforilált Akt (pAkt) belső kontrollja a tAkt volt, míg az ERK1/2-é az aktin. Másnap 6 x 5
percen át 0,2 % Tweent tartalmazó Tris-pufferelt sóoldatban (pH 7,5) mostuk a membránokat.
Ezt követően kecske anti-nyúl (1:3000) vagy anti-egér (1:1500) tormaperoxidázzal konjugált

36
másodlagos antitesteket (1:3000, BioRad, Budapest) használtunk. Az antigén-antitest komplex
vizualizálása erősített kemilumineszcencia segítségével történt, a reakciót ECL Plus western
blotting detektáló rendszerrel rögzítettük (GE Healthcare). Scanningelést követően
eredményeinket NIH ImageJ program segítségével kvantifikáltuk, szemikvantitatív pixel
denzitás mérést követően. A normalizációt a pAkt esetében tAkt-re, a pERK1/2 esetében aktinra
végeztük. Statisztikai elemzésünkhöz kétutas ANOVA tesztet alkalmaztunk, Fischer féle post
hoc analízissel.
4.1.6. A PACAP1-27 szemcseppes kezelést követő citokin array vizsgálat
A western blot analízis során nyert tapasztalatokból kiindulva a vizsgálathoz szükséges
retinákat a műtétet követő 24 óra után távolítottuk el. Kísérletünkhöz 11 sham és 17 ischaemiás
károsodást szenvedett állatot használtunk. A cseppentések számát ennél az eljárásnál is
megnöveltük, a western blotnál leírtakhoz hasonlóan. A műtétet követő 24 óra elteltével az
állatokat túlaltattuk, majd szemeik eltávolítás után a retinákat kipreparáltuk, és az azonos
csoportba tartozó szemeket ugyanabban az Eppendorf csőben tároltuk (pooloztuk), és
folyékony nitrogénben azonnal lefagyasztottuk. A citokin array vizsgálatig -80◦C-on tároltuk a
retinákat. A szemikvantitatív módszer a nitrocellulóz membránokra előre duplikátumban kötött
antitestek, és a mintában lévő proteinek kötődésén alapul. Az array-ek szenzitivitása az egyes
proteinek esetében eltérő, alacsony pg/ml-es tartományban van, detekciós maximuma 2-10
ng/ml, azonban ez a detekciós eljárástól és az expozíciós időtől függő. Vizsgálatunkhoz Rat
Cytokine Array Panel A Array kitet használtunk (R&D System, Biomedica, Budapest). A kit
tartalmazza a kísérlethez szükséges puffert, mosóoldatokat, detektáló antitest koktélt és a négy
membránt (10. ábra). Az array-ket a gyártó cég előírása szerint alkalmaztuk. Egy órás
membránblokkolást követően 1,5ml médiumot adtunk a membránokhoz, majd azokat detektáló
koktél jelenlétében rázólemezen inkubáltuk 2-8◦C-on, egy éjszakán keresztül. Másnap
háromszori mosást követően adtuk a rendszerhez a tormaperoxidázhoz konjugált streptavidint,
melyet kemilumineszcens előhívás követett (reagens: Amersham Biosciences, Budapest), végül
röntgenfilmen rögzítettük az eredményeket (Szabó és mtsai 2012). A filmeket scanneltük, végül
ImageJ Protein Array Analiser program segítségével a denzitás változásokat lemértük. Az így
kapott adatokat Microsoft Excel programmal tovább árnyaltuk és pontosítottuk: a referencia

37
spotok segítségével a hátteret egységesítettük, és ezt követően a kontroll spotok átlagához
normalizáltuk adatainkat. Így a relatív változás mértékét tudtuk meghatározni, illetve
összehasonlítani. Eredményeinket a változások százalékos értékében határoztuk meg, és a
kontroll (sham) retinákhoz képest 20%-nál nagyobb fokú expressziós eltérést mutató
citokineket ábrázoltuk diagramokon. A citokin array méréseinket négyszer ismételtük meg.
10. ábra: Citokin array
4.1.7. Elektroretinográfiás (ERG) vizsgálat PACAP1-27 szemcseppes terápiát
követően
Két héttel a műtétet követően, állatainkat (11 sham operált és 17 BCCAO műtéten
átesett) az ERG vizsgálat előtt 24 órán keresztül sötét adaptációnak tettük ki. Korábbi
vizsgálatainkban leírtak szerint, intraperitoneálisan alkalmazott ketamin – xylazil koktéllal
altattuk a patkányokat (Dányádi és mtsai 2014). A vizsgálathoz Homatropin szemcseppet
(gyógyszertári magisztrális készítmény) használtunk pupillatágítóként, illetve oxybuprocaine
(Humacain, TEVA) cseppet érzéstelenítőként. Az elektroretinogram rögzítéséhez minden
mérésnél öt elektródot használtunk: 2 db mérő, 2 db referencia, és 1 általános föld elektródot.
Miután a pupilla teljesen kitágult, az állatok mindkét szaruhártyájának felszínére mérő
elektródot helyeztünk. A két referencia elektródot az pofa bőre alá helyeztük el, míg az általános
föld elektródot az állat középvonalában, a glabella területébe szúrtuk. Mindezt sötét
helyiségben, fejen rögzített, piros LED fényben kiviteleztük, így a kialakult sötét adaptációt
nem befolyásoltuk. A felvillanó fényre adott retinális elektromos jeleket (5,0cd/m2, 0,25Hz,
503nm zöld LED fény) az elektródokkal vezettük el, majd a kapott válaszokat felerősítettük

38
(2,000 x, Bioamp SbA4-V6, Supertech, Hungary), és A/D konverterrel rögzítettük. Minden
mérés alkalmával, a válaszértékek átlagát A/D konverter rajzolta ki görbék formájában. Az így
kapott görbéket Origin 8 szoftver segítségével értékeltük, ahol lehetőség volt a különböző
hullámok amplitúdójának lemérésére, és az implicit idő, valamint az oszcillációs potenciál
meghatározására. Kapott adatainkat Microsoft Excel programban összesítettük.
Kiértékelésükhöz kétutas ANOVA tesztet használtunk Fischer féle post hoc analízissel.
4.2. Eredményeink
4.2.1. Rutin morfologiai és morfometriai analízis
Két héttel a műtétet követően retina metszeteinken az ischaemiás károsodás jelei jól
láthatóak illetve mérhetőek (11.,12.,13.,14. ábra) voltak. A sham operált állatok jobb és bal
szemei között (PACAP1-27-el kezelt – nem kezelt szemek) nem detektálható szignifikáns
különbség, így megállapítható, a PACAP1-27 kezelés önmagában nem okozott változást a
retina szerkezetében (14. ábra). Kísérleteink kezdeti szakaszán a megfelelő vivőanyag
megtalálását tűztük ki célul. Kipróbált vivőanyagaink a következők voltak: fiziológiás sóoldat,
aqua ad iniectabilia, solvens viscosa pro oculoguttis cum thiomersalo, solutio ophthalmica cum
benzalkonio.
Szövettani metszeteinken a retina szerkezetében súlyos ischaemiás károsodásra utaló
jeleket láttunk BCCAO-t követően, valamint a retina rétegeinek vastagságában szignifikáns
mértékű különbségeket mértünk az operált, illetve sham csoportba tartozó állatok között. A
PACAP1-27 eddig már ismert retinoprotektív hatása azonban nem igazolódott sem a fiziológiás
sóoldat, sem az aqua ad iniectabila, és a solvens viscosa pro oculoguttis cum thiomersalo
vivőanyag esetében sem (11.,12.,13. ábra).
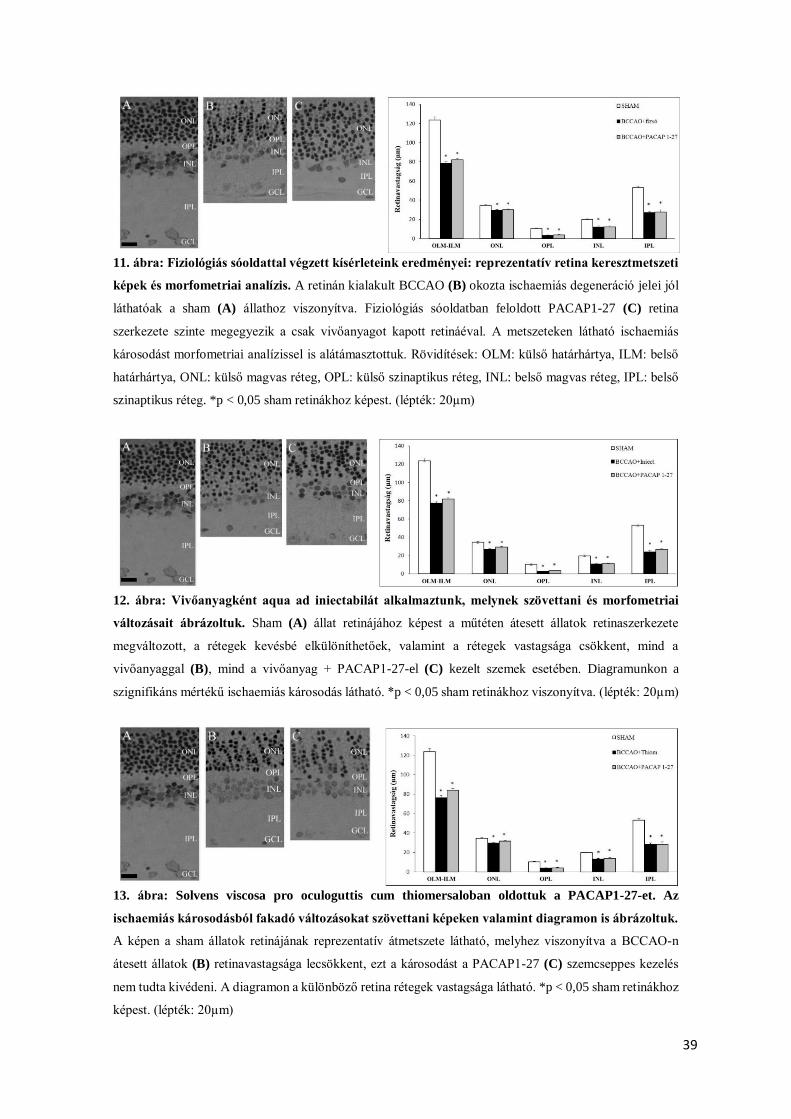
39
11. ábra: Fiziológiás sóoldattal végzett kísérleteink eredményei: reprezentatív retina keresztmetszeti
képek és morfometriai analízis. A retinán kialakult BCCAO (B) okozta ischaemiás degeneráció jelei jól
láthatóak a sham (A) állathoz viszonyítva. Fiziológiás sóoldatban feloldott PACAP1-27 (C) retina
szerkezete szinte megegyezik a csak vivőanyagot kapott retináéval. A metszeteken látható ischaemiás
károsodást morfometriai analízissel is alátámasztottuk. Rövidítések: OLM: külső határhártya, ILM: belső
határhártya, ONL: külső magvas réteg, OPL: külső szinaptikus réteg, INL: belső magvas réteg, IPL: belső
szinaptikus réteg. *p < 0,05 sham retinákhoz képest. (lépték: 20µm)
12. ábra: Vivőanyagként aqua ad iniectabilát alkalmaztunk, melynek szövettani és morfometriai
változásait ábrázoltuk. Sham (A) állat retinájához képest a műtéten átesett állatok retinaszerkezete
megváltozott, a rétegek kevésbé elkülöníthetőek, valamint a rétegek vastagsága csökkent, mind a
vivőanyaggal (B), mind a vivőanyag + PACAP1-27-el (C) kezelt szemek esetében. Diagramunkon a
szignifikáns mértékű ischaemiás károsodás látható. *p < 0,05 sham retinákhoz viszonyítva. (lépték: 20µm)
13. ábra: Solvens viscosa pro oculoguttis cum thiomersaloban oldottuk a PACAP1-27-et. Az
ischaemiás károsodásból fakadó változásokat szövettani képeken valamint diagramon is ábrázoltuk.
A képen a sham állatok retinájának reprezentatív átmetszete látható, melyhez viszonyítva a BCCAO-n
átesett állatok (B) retinavastagsága lecsökkent, ezt a károsodást a PACAP1-27 (C) szemcseppes kezelés
nem tudta kivédeni. A diagramon a különböző retina rétegek vastagsága látható. *p < 0,05 sham retinákhoz
képest. (lépték: 20µm)

40
A solutio ophthalmica cum benzalkonio vivőanyagban feloldott PACAP1-27-nél
szignifikáns különbséget mértünk a BCCAO és BCCAO + PACAP1-27 csoport retináinak
rétegvastagságai között, a PACAP1-27 ismert retinoprotektív hatása igazolódott, a vivőanyag
hatékonynak bizonyult (14. ábra). A hatásosnak vélt solutio ophthalmica cum benzalkonio
vivőanyaggal vizsgálatainkat többször megismételtük. Kísérleteinkben az állatok szemeit 4
csoportba soroltuk be: sham, sham + PACAP1-27, BCCAO, BCCAO + PACAP1-27.
Többszöri méréseink alátámasztották korábbi eredményeinket, a PACAP1-27 szignifikánsan
csökkentette a károsodás mértékét a teljes retina vastagságában (OLM-ILM: 50,7%-al csökkent
BCCAO, 36,1%-al BCCAO + PACAP1-27 csoportba), a külső szinaptikus rétegben (OPL:
54,2%-al csökkent BCCAO, míg 47,9%-al BCCAO + PACAP1-27 kezelés hatására) és a belső
magvas rétegben is (INL: 46,5%-al csökkent BCCAO-t követően, 30,7%-al BCCAO +
PACAP1-27 kezelés után), valamint a belső szinaptikus rétegben (IPL: BCCAO: 61,4%,
BCCAO + PACAP1-27: 38,3%). A két csoport között a külső magvas rétegvastagságnál
szignifikáns mértékű különbség nem volt mérhető (ONL: BCCAO: 35,3%-al, BCCAO +
PACAP1-27: 36,3%-al csökkent) (15. ábra).
14. ábra: Fénymikroszkópos felvétel a retina rétegeiről. A BCCAO (C) okozta ischaemiás károsodás
jelei láthatóak a retina szerkezetében a sham (A) illetve a sham + PACAP1-27 (B) állat retinájához képest.
A műtétet követően solutio ophthalmica cum benzalkonioban oldott PACAP1-27 szemcseppes kezelés
hatására a retina rétegeiben enyhébb mértékű degeneráció figyelhető meg (D). (lépték: 50µm)
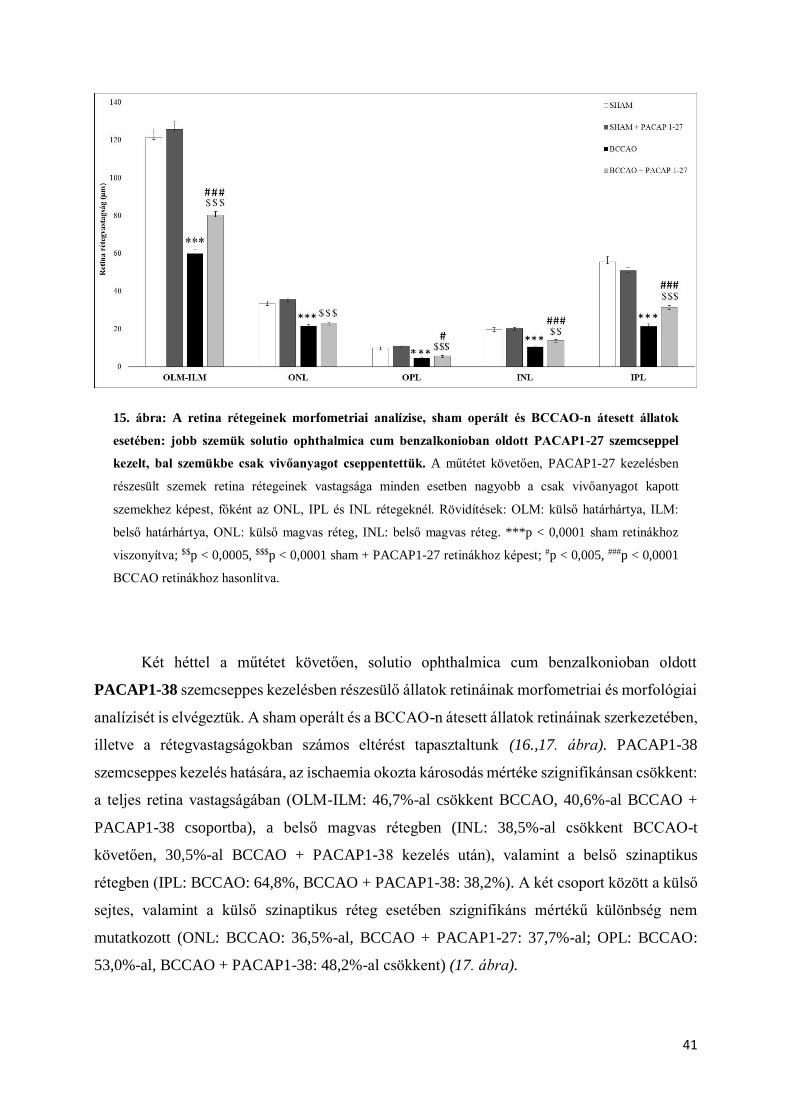
41
15. ábra: A retina rétegeinek morfometriai analízise, sham operált és BCCAO-n átesett állatok
esetében: jobb szemük solutio ophthalmica cum benzalkonioban oldott PACAP1-27 szemcseppel
kezelt, bal szemükbe csak vivőanyagot cseppentettük. A műtétet követően, PACAP1-27 kezelésben
részesült szemek retina rétegeinek vastagsága minden esetben nagyobb a csak vivőanyagot kapott
szemekhez képest, főként az ONL, IPL és INL rétegeknél. Rövidítések: OLM: külső határhártya, ILM:
belső határhártya, ONL: külső magvas réteg, INL: belső magvas réteg. ***p < 0,0001 sham retinákhoz
viszonyítva; $$p < 0,0005, $$$p < 0,0001 sham + PACAP1-27 retinákhoz képest; #p < 0,005, ###p < 0,0001
BCCAO retinákhoz hasonlítva.
Két héttel a műtétet követően, solutio ophthalmica cum benzalkonioban oldott
PACAP1-38 szemcseppes kezelésben részesülő állatok retináinak morfometriai és morfológiai
analízisét is elvégeztük. A sham operált és a BCCAO-n átesett állatok retináinak szerkezetében,
illetve a rétegvastagságokban számos eltérést tapasztaltunk (16.,17. ábra). PACAP1-38
szemcseppes kezelés hatására, az ischaemia okozta károsodás mértéke szignifikánsan csökkent:
a teljes retina vastagságában (OLM-ILM: 46,7%-al csökkent BCCAO, 40,6%-al BCCAO +
PACAP1-38 csoportba), a belső magvas rétegben (INL: 38,5%-al csökkent BCCAO-t
követően, 30,5%-al BCCAO + PACAP1-38 kezelés után), valamint a belső szinaptikus
rétegben (IPL: BCCAO: 64,8%, BCCAO + PACAP1-38: 38,2%). A két csoport között a külső
sejtes, valamint a külső szinaptikus réteg esetében szignifikáns mértékű különbség nem
mutatkozott (ONL: BCCAO: 36,5%-al, BCCAO + PACAP1-27: 37,7%-al; OPL: BCCAO:
53,0%-al, BCCAO + PACAP1-38: 48,2%-al csökkent) (17. ábra).
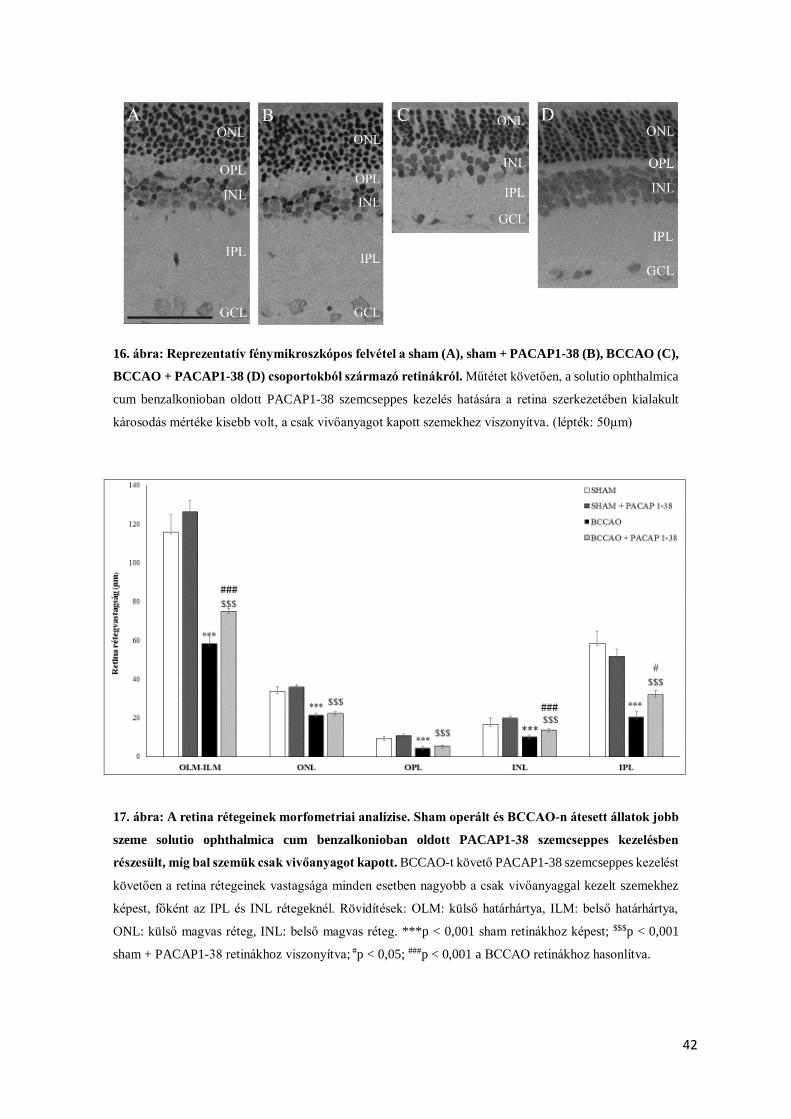
42
16. ábra: Reprezentatív fénymikroszkópos felvétel a sham (A), sham + PACAP1-38 (B), BCCAO (C),
BCCAO + PACAP1-38 (D) csoportokból származó retinákról. Műtétet követően, a solutio ophthalmica
cum benzalkonioban oldott PACAP1-38 szemcseppes kezelés hatására a retina szerkezetében kialakult
károsodás mértéke kisebb volt, a csak vivőanyagot kapott szemekhez viszonyítva. (lépték: 50µm)
17. ábra: A retina rétegeinek morfometriai analízise. Sham operált és BCCAO-n átesett állatok jobb
szeme solutio ophthalmica cum benzalkonioban oldott PACAP1-38 szemcseppes kezelésben
részesült, míg bal szemük csak vivőanyagot kapott. BCCAO-t követő PACAP1-38 szemcseppes kezelést
követően a retina rétegeinek vastagsága minden esetben nagyobb a csak vivőanyaggal kezelt szemekhez
képest, főként az IPL és INL rétegeknél. Rövidítések: OLM: külső határhártya, ILM: belső határhártya,
ONL: külső magvas réteg, INL: belső magvas réteg. ***p < 0,001 sham retinákhoz képest; $$$p < 0,001
sham + PACAP1-38 retinákhoz viszonyítva; #p < 0,05; ###p < 0,001 a BCCAO retinákhoz hasonlítva.
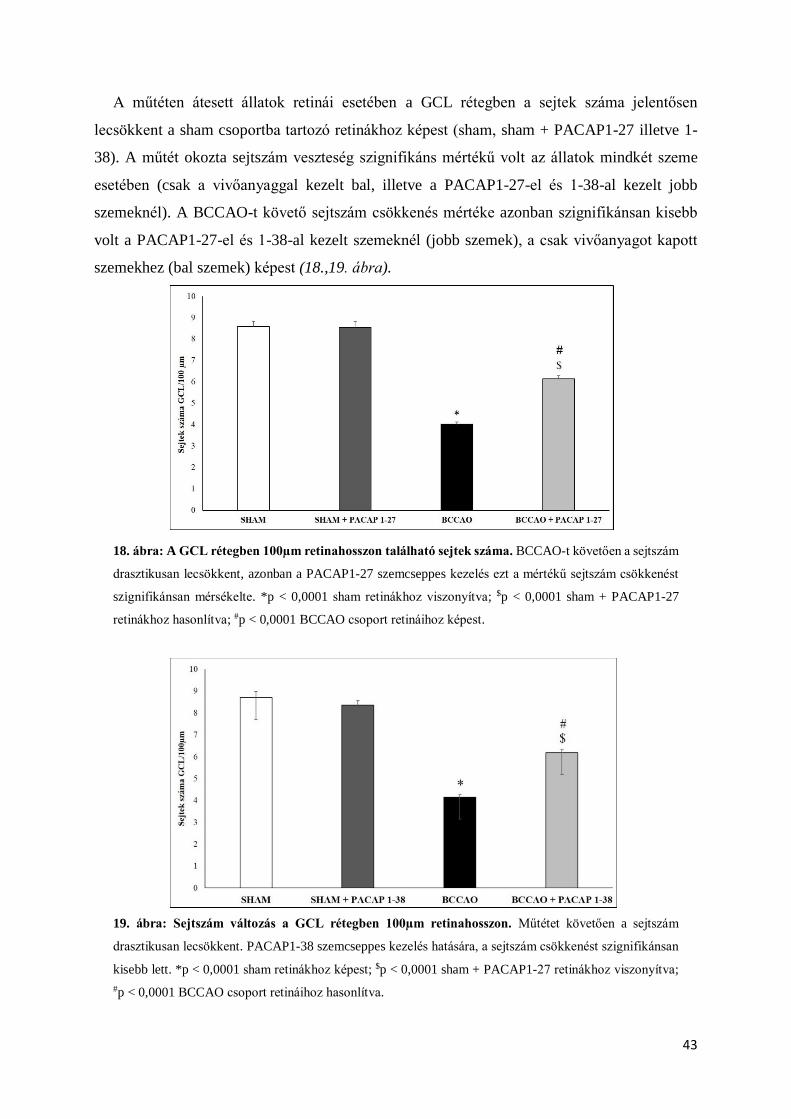
43
A műtéten átesett állatok retinái esetében a GCL rétegben a sejtek száma jelentősen
lecsökkent a sham csoportba tartozó retinákhoz képest (sham, sham + PACAP1-27 illetve 1-
38). A műtét okozta sejtszám veszteség szignifikáns mértékű volt az állatok mindkét szeme
esetében (csak a vivőanyaggal kezelt bal, illetve a PACAP1-27-el és 1-38-al kezelt jobb
szemeknél). A BCCAO-t követő sejtszám csökkenés mértéke azonban szignifikánsan kisebb
volt a PACAP1-27-el és 1-38-al kezelt szemeknél (jobb szemek), a csak vivőanyagot kapott
szemekhez (bal szemek) képest (18.,19. ábra).
18. ábra: A GCL rétegben 100µm retinahosszon található sejtek száma. BCCAO-t követően a sejtszám
drasztikusan lecsökkent, azonban a PACAP1-27 szemcseppes kezelés ezt a mértékű sejtszám csökkenést
szignifikánsan mérsékelte. *p < 0,0001 sham retinákhoz viszonyítva; $p < 0,0001 sham + PACAP1-27
retinákhoz hasonlítva; #p < 0,0001 BCCAO csoport retináihoz képest.
19. ábra: Sejtszám változás a GCL rétegben 100µm retinahosszon. Műtétet követően a sejtszám
drasztikusan lecsökkent. PACAP1-38 szemcseppes kezelés hatására, a sejtszám csökkenést szignifikánsan
kisebb lett. *p < 0,0001 sham retinákhoz képest; $p < 0,0001 sham + PACAP1-27 retinákhoz viszonyítva;
#p < 0,0001 BCCAO csoport retináihoz hasonlítva.

44
Összefoglalva, a szövettani méréseink bizonyították, hogy a PACAP1-27 solutio
ophthalmica cum benzalkonioban oldva - szemcseppként alkalmazva - kifejti eddig ismert
retinoprotektív hatását ischaemiás károsodás esetén. Emiatt további vizsgálatainkat ezzel a
vivőanyaggal folytattuk. Az említett vivőanyagban oldott PACAP1-38 szintén
retinoprotektívnek bizonyult.
4.2.2. 125I izotóppal jelölt PACAP1-27 és 1-38 koncentráció változása
szemcseppes kezelés esetén
A radioaktív jódizotóppal jelölt PACAP1-27 és 1-38 radioaktivitását 5 – 120 perces
periódus között többször mértük hím CD-1 egereken. A jelölt PACAP1-27 és 1-38 már 5 perc
elteltével kimutatható volt a szemben, az agyban és a szérumban egyaránt. Elsőként a corneán
való átjutásukat ábrázoltuk, csúcskoncentrációjukat a PACAP1-27 esetében a 60. percnél, míg
PACAP1-38 esetében a 120. percnél érték el (20/A. ábra). Az üvegtestben detektált PACAP1-
27 koncentrációja a szaruhártyához hasonlóan, 60 perc után érte el a csúcsértéket. A PACAP1-
38 szintén a corneánál mért csúcsértékhez hasonlóan, a 120. percben (20/C. ábra). A retinánál
a PACAP1-27 csúcskoncentrációját szintén a 60. percben detektáltuk, azonban ebben az
esetben később sem csökkent a koncentrációja, hanem plato szerűen megtartott maradt. Az
ideghártyánál a PACAP1-38 koncentrációja már a 30. percben hirtelen megnőtt, és a szintje
alig változott a 120. percig (20/B. ábra). A PACAP1-27 gyorsan bejutott az agy területére is
(30 perc), azonban ezt követően koncentrációja rohamosan csökkent, 120 perc után szinte alig
volt kimutatható. A PACAP1-38 kisebb mértékben jutott be az agyba, mint a 27 aminosavból
álló forma, csúcskoncentrációját ebben az esetben a 60. percnél érte el (20/D. ábra). PACAP1-
27 nagyon alacsony koncentrációban jutott a szérumba, és a csúcskoncentráció elérése is
hosszabb időt vett igénybe, 120 percet. A 38 aminosavból álló forma kis koncentrációban jutott
csak be a szérumba (20/E. ábra). A retinát kivéve, a PACAP1-27 és 1-38 csúcskoncentrációinak
értéke a corneától a szérumig folyamatosan csökkent (20. ábra). Az agyban, valamint a
szérumban mért csúcskoncentrációk mindkét peptid esetében töredékei a corneában mértnek
(PACAP1-38: corneában 420%Inj/g, agyban 0,05%Inj/g, szérumban 0,02%Inj/ml; PACAP1-
27: corneában 300%Inj/g, agyban 0,25%Inj/g, szérumban 1,2%Inj/ml) (20. ábra).

45
20. ábra: Radioaktív izotóppal jelölt PACAP1-27 és 1-38 eloszlása szemcseppes terápia során. 125I
izotóppal jelölt PACAP1-27 (teli karikás, folytonos vonal) valamint 1-38 (üres karikás, szaggatott vonal)
megjelenése a corneán (A), az üvegtestben (C), a retinán (B), az agy területén (D) illetve a szérumban (E),
5 – 30 – 60 – 120 perc elteltével.
4.2.3. Müller glia sejtek immunhisztokémiai analízise
A GFAP filamentumok intermedier filamentumok, melyek főként az astrocytákban és
az ependimális sejtekben expresszálódnak a központi idegrendszerben. A retinában normális
esetben a Müller glia sejtek végtalpaiban vannak jelen, az ILM rétegben (21/A.,B.; 22/A.,B.
ábra) (Bjorklund és Dahl 1985). Carotis lekötést követően - az ischaemiás károsodás miatt
- a GFAP szintje megemelkedett, és a Müller glia sejtek teljes hosszában kimutathatóvá vált
(az OLM-től az ILM rétegekig) (21/C.; 22/C. ábra). PACAP1-27 illetve 1-38 kezelés
hatására, a GFAP kisebb mértékben expresszálódott, szinte csak a végtalpakra korlátozódott,
ami a kisebb mértékű károsodásra utal (21/D.; 22/D. ábra).
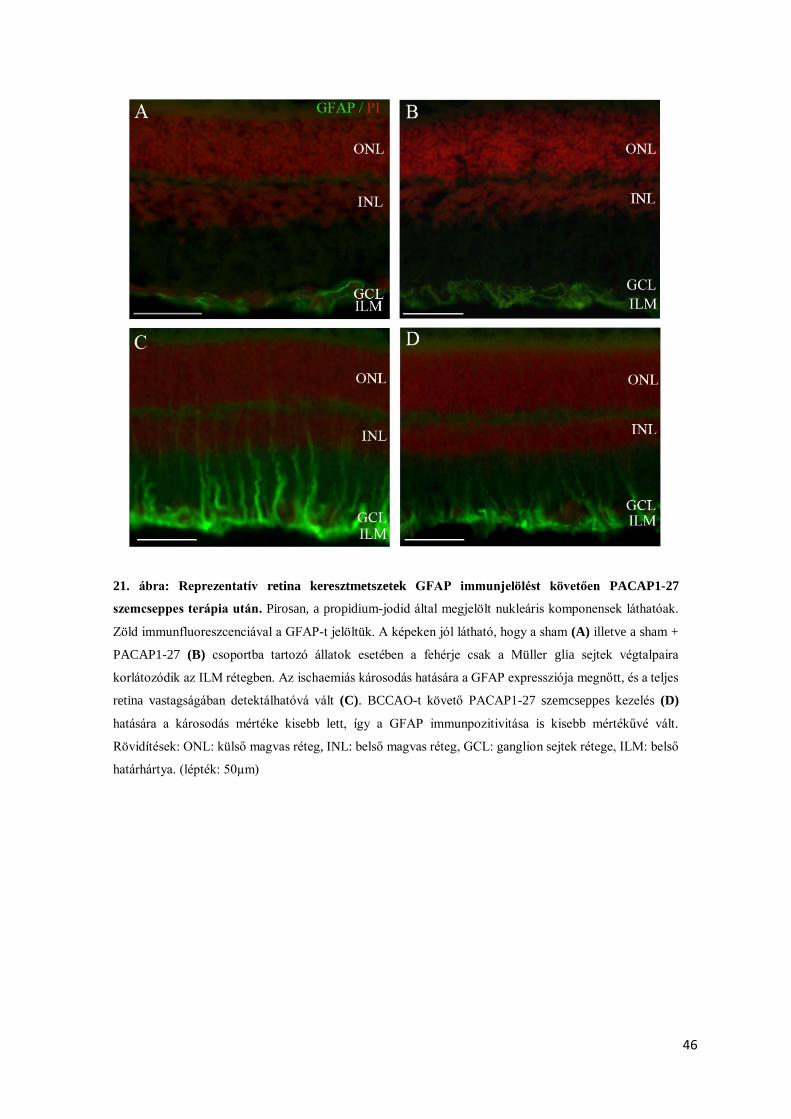
46
21. ábra: Reprezentatív retina keresztmetszetek GFAP immunjelölést követően PACAP1-27
szemcseppes terápia után. Pirosan, a propidium-jodid által megjelölt nukleáris komponensek láthatóak.
Zöld immunfluoreszcenciával a GFAP-t jelöltük. A képeken jól látható, hogy a sham (A) illetve a sham +
PACAP1-27 (B) csoportba tartozó állatok esetében a fehérje csak a Müller glia sejtek végtalpaira
korlátozódik az ILM rétegben. Az ischaemiás károsodás hatására a GFAP expressziója megnőtt, és a teljes
retina vastagságában detektálhatóvá vált (C). BCCAO-t követő PACAP1-27 szemcseppes kezelés (D)
hatására a károsodás mértéke kisebb lett, így a GFAP immunpozitivitása is kisebb mértékűvé vált.
Rövidítések: ONL: külső magvas réteg, INL: belső magvas réteg, GCL: ganglion sejtek rétege, ILM: belső
határhártya. (lépték: 50µm)
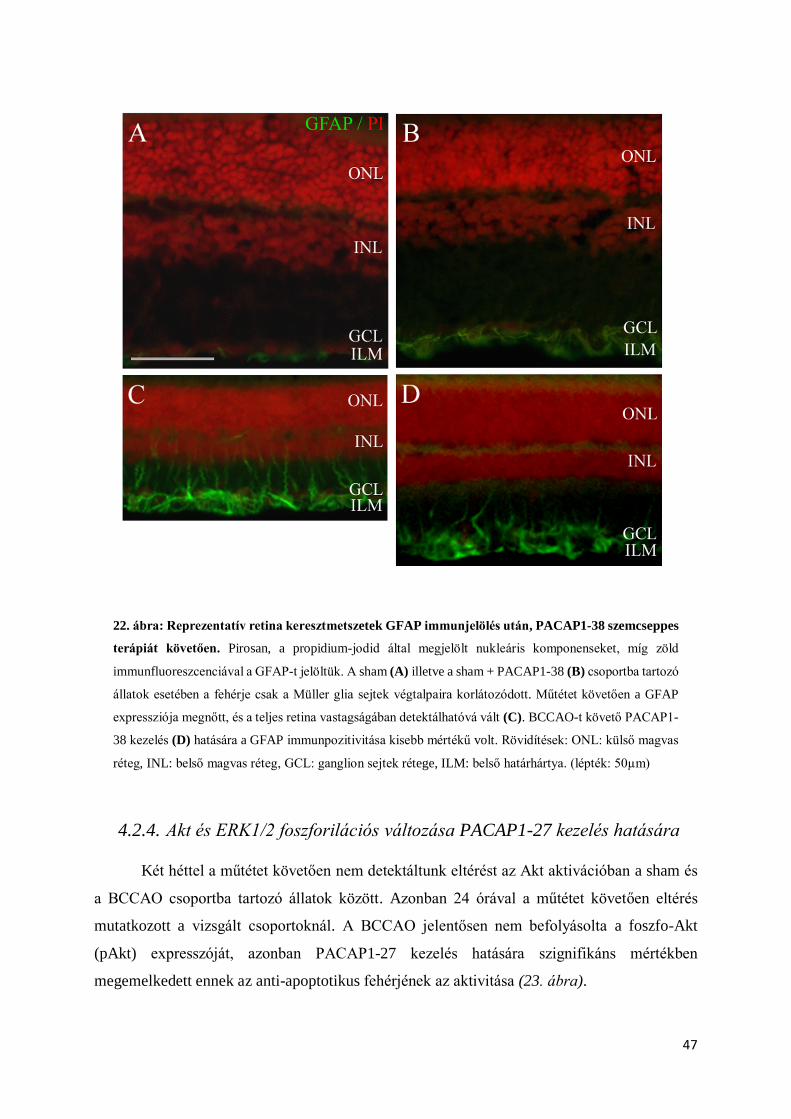
47
22. ábra: Reprezentatív retina keresztmetszetek GFAP immunjelölés után, PACAP1-38 szemcseppes
terápiát követően. Pirosan, a propidium-jodid által megjelölt nukleáris komponenseket, míg zöld
immunfluoreszcenciával a GFAP-t jelöltük. A sham (A) illetve a sham + PACAP1-38 (B) csoportba tartozó
állatok esetében a fehérje csak a Müller glia sejtek végtalpaira korlátozódott. Műtétet követően a GFAP
expressziója megnőtt, és a teljes retina vastagságában detektálhatóvá vált (C). BCCAO-t követő PACAP1-
38 kezelés (D) hatására a GFAP immunpozitivitása kisebb mértékű volt. Rövidítések: ONL: külső magvas
réteg, INL: belső magvas réteg, GCL: ganglion sejtek rétege, ILM: belső határhártya. (lépték: 50µm)
4.2.4. Akt és ERK1/2 foszforilációs változása PACAP1-27 kezelés hatására
Két héttel a műtétet követően nem detektáltunk eltérést az Akt aktivációban a sham és
a BCCAO csoportba tartozó állatok között. Azonban 24 órával a műtétet követően eltérés
mutatkozott a vizsgált csoportoknál. A BCCAO jelentősen nem befolyásolta a foszfo-Akt
(pAkt) expresszóját, azonban PACAP1-27 kezelés hatására szignifikáns mértékben
megemelkedett ennek az anti-apoptotikus fehérjének az aktivitása (23. ábra).
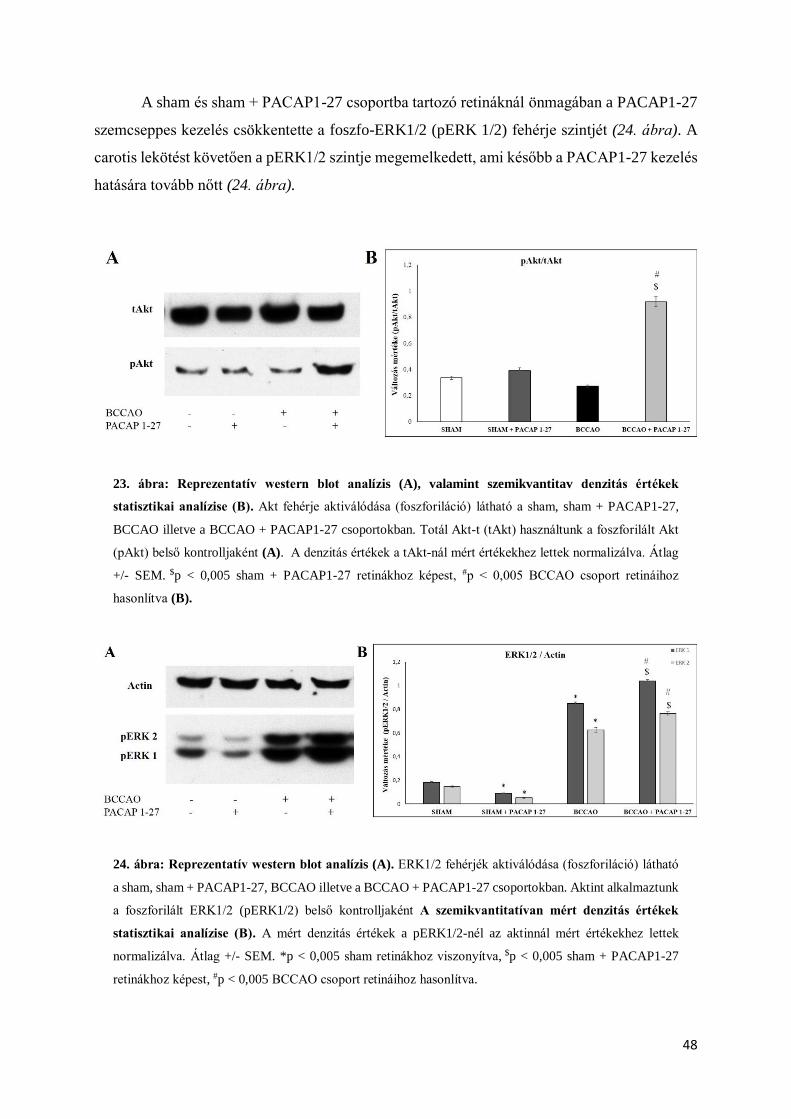
48
A sham és sham + PACAP1-27 csoportba tartozó retináknál önmagában a PACAP1-27
szemcseppes kezelés csökkentette a foszfo-ERK1/2 (pERK 1/2) fehérje szintjét (24. ábra). A
carotis lekötést követően a pERK1/2 szintje megemelkedett, ami később a PACAP1-27 kezelés
hatására tovább nőtt (24. ábra).
23. ábra: Reprezentatív western blot analízis (A), valamint szemikvantitav denzitás értékek
statisztikai analízise (B). Akt fehérje aktiválódása (foszforiláció) látható a sham, sham + PACAP1-27,
BCCAO illetve a BCCAO + PACAP1-27 csoportokban. Totál Akt-t (tAkt) használtunk a foszforilált Akt
(pAkt) belső kontrolljaként (A). A denzitás értékek a tAkt-nál mért értékekhez lettek normalizálva. Átlag
+/- SEM. $p < 0,005 sham + PACAP1-27 retinákhoz képest, #p < 0,005 BCCAO csoport retináihoz
hasonlítva (B).
24. ábra: Reprezentatív western blot analízis (A). ERK1/2 fehérjék aktiválódása (foszforiláció) látható
a sham, sham + PACAP1-27, BCCAO illetve a BCCAO + PACAP1-27 csoportokban. Aktint alkalmaztunk
a foszforilált ERK1/2 (pERK1/2) belső kontrolljaként A szemikvantitatívan mért denzitás értékek
statisztikai analízise (B). A mért denzitás értékek a pERK1/2-nél az aktinnál mért értékekhez lettek
normalizálva. Átlag +/- SEM. *p < 0,005 sham retinákhoz viszonyítva, $p < 0,005 sham + PACAP1-27
retinákhoz képest, #p < 0,005 BCCAO csoport retináihoz hasonlítva.

49
4.2.5. PACAP1-27 szemcseppes kezelés hatása az ischaemia okozta citokin
expresszió változásra
Különböző citokinek expressziós változásainak méréséhez citokin arrayt használtunk (25.
ábra). Az ischaemiás károsodás számos citokin aktivációját befolyásolta (25. ábra). A lemért
denzitásértékek normalizálását követően a citokinek relatív változását hasonlítottuk össze. Azokat
a citokineket vizsgáltuk, illetve ábrázoltuk diagramon, amelyeknél a sham retinákhoz képest a
relatív változás mértéke a 20%-ot meghaladta. A különböző vizsgált csoportok között ilyen mértékű
változást a citokin-indukált neutrophil chemoattraktáns-1 (CINC-1), makrofág gyulladásos fehérje
(MIP-3α), thymus kemokin (CXCL-7), mátrix metalloproteináz szöveti gátlója (TIMP-1) valamint
a vaszkuláris endoteliális növekedési faktor esetében mértünk. Műtétet követően mennyiségük
megemelkedett a sham csoport állataihoz képest, azonban PACAP1-27 kezelés hatására szintjük
lecsökkent (26. ábra,2. táblázat). A sham csoportba tartozó PACAP1-27-el kezelt és nem kezelt
állatoknál is mértünk különbséget a citokinek expressziójában, azonban ez a 20%-ot nem haladta
meg egyik esetben sem.
25. ábra: Reprezentatív citokin array. A filmeken: Sham, sham + PACAP1-27, BCCAO és BCCAO +
PACAP1-27 csoportok láthatók. Karikával azoknak a citokineknek a helyét jelöltük, melyek expressziója
az ischaemiás károsodást követően nagymértékben megváltozott, és ezt a relatív változást a PACAP1-27
szemcseppes terápia 20%-nál nagyobb mértékben befolyásolta.
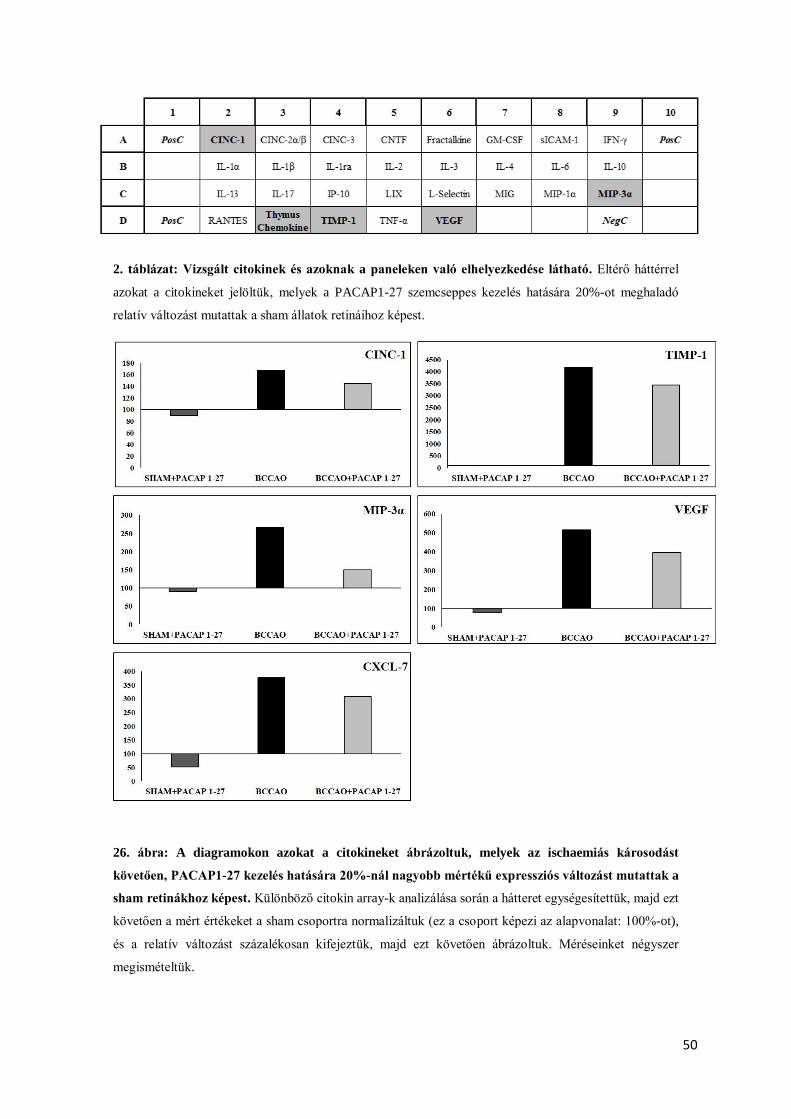
50
2. táblázat: Vizsgált citokinek és azoknak a paneleken való elhelyezkedése látható. Eltérő háttérrel
azokat a citokineket jelöltük, melyek a PACAP1-27 szemcseppes kezelés hatására 20%-ot meghaladó
relatív változást mutattak a sham állatok retináihoz képest.
26. ábra: A diagramokon azokat a citokineket ábrázoltuk, melyek az ischaemiás károsodást
követően, PACAP1-27 kezelés hatására 20%-nál nagyobb mértékű expressziós változást mutattak a
sham retinákhoz képest. Különböző citokin array-k analizálása során a hátteret egységesítettük, majd ezt
követően a mért értékeket a sham csoportra normalizáltuk (ez a csoport képezi az alapvonalat: 100%-ot),
és a relatív változást százalékosan kifejeztük, majd ezt követően ábrázoltuk. Méréseinket négyszer
megismételtük.
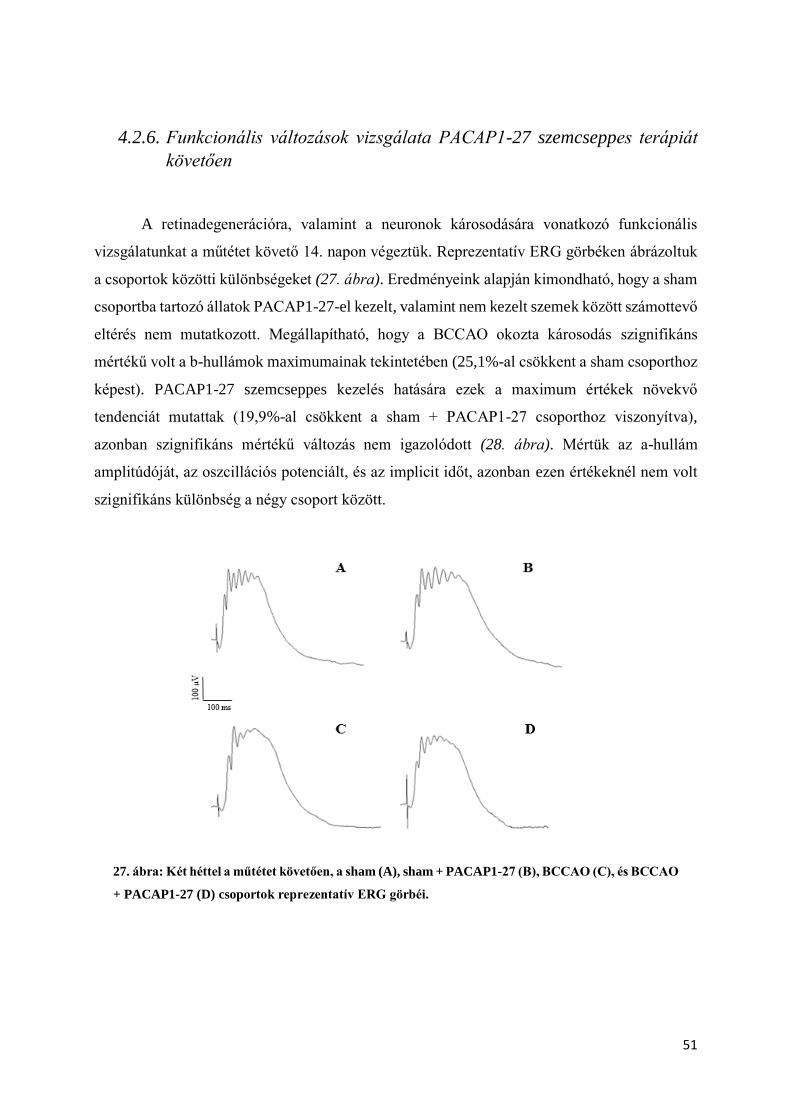
51
4.2.6. Funkcionális változások vizsgálata PACAP1-27 szemcseppes terápiát
követően
A retinadegenerációra, valamint a neuronok károsodására vonatkozó funkcionális
vizsgálatunkat a műtétet követő 14. napon végeztük. Reprezentatív ERG görbéken ábrázoltuk
a csoportok közötti különbségeket (27. ábra). Eredményeink alapján kimondható, hogy a sham
csoportba tartozó állatok PACAP1-27-el kezelt, valamint nem kezelt szemek között számottevő
eltérés nem mutatkozott. Megállapítható, hogy a BCCAO okozta károsodás szignifikáns
mértékű volt a b-hullámok maximumainak tekintetében (25,1%-al csökkent a sham csoporthoz
képest). PACAP1-27 szemcseppes kezelés hatására ezek a maximum értékek növekvő
tendenciát mutattak (19,9%-al csökkent a sham + PACAP1-27 csoporthoz viszonyítva),
azonban szignifikáns mértékű változás nem igazolódott (28. ábra). Mértük az a-hullám
amplitúdóját, az oszcillációs potenciált, és az implicit időt, azonban ezen értékeknél nem volt
szignifikáns különbség a négy csoport között.
27. ábra: Két héttel a műtétet követően, a sham (A), sham + PACAP1-27 (B), BCCAO (C), és BCCAO
+ PACAP1-27 (D) csoportok reprezentatív ERG görbéi.
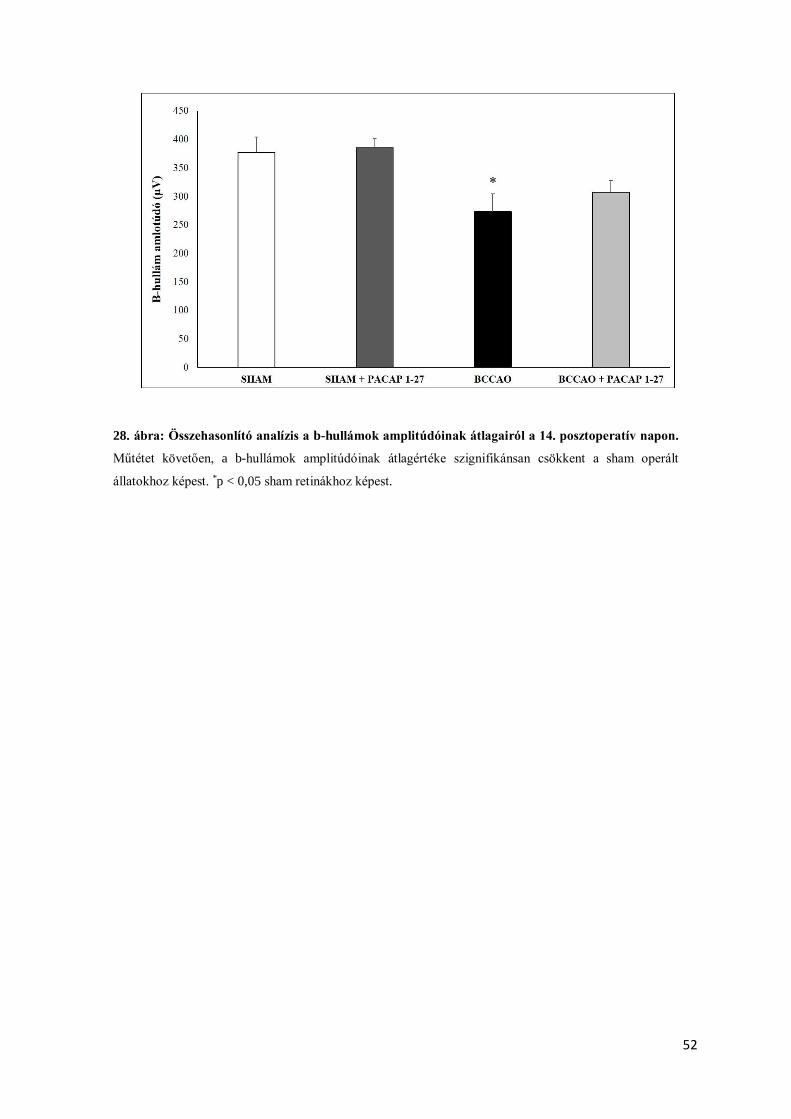
52
28. ábra: Összehasonlító analízis a b-hullámok amplitúdóinak átlagairól a 14. posztoperatív napon.
Műtétet követően, a b-hullámok amplitúdóinak átlagértéke szignifikánsan csökkent a sham operált
állatokhoz képest. *p < 0,05 sham retinákhoz képest.

53
5. Megbeszélés
5.1. PACAP családjába tartozó neuropeptidek valamint
fragmenseinek vizsgálatáról szóló korábbi tanulmányok, illetve
a mostani vizsgálatok eredményének összefoglalása
A PACAP a szekretin/glükagon/VIP peptidcsalád tagja, receptorai megtalálhatóak a
retinában is (Nakamachi és mtsai 2012). A PACAP fontos szerepet játszik a látás
mechanizmusában, a retinohypothalamicus pálya fontos kotranszmittere, ahonnan az
információk a suprachiasmaticus magba jutnak (Kawaguchi és mtsai 2010). Amellett, hogy a
PACAP feltehetően részt vesz különböző fiziológiás folyamatokban, bizonyítottan
retinoprotektív sajátosságokkal is rendelkezik. Endogén védő hatását bizonyították olyan
egereken, melyek nem termelnek endogén PACAP-ot. Az egereknél ischaemiás
retinakárosodást (Reglődi és mtsai 2012), valamint NMDA indukálta excitotoxikus károsodást
(Endo és mtsai 2011) idéztek elő, és azt tapasztalták, hogy a PACAP génhiányos egerek
retinakárosodásának mértéke fokozottabb volt a kontroll állatokhoz képest. Ezt a tulajdonságát
további vizsgálatok is megerősítették: PACAP antagonista intravitreális adását követően az
apoptózis mértéke fokozódott (Rácz és mtsai 2006). A PACAP egy elfogadott retinoprotektív
ágens különböző retinakárosodást előidéző állatkísérletes modellekben, emiatt számos
tanulmány próbálta feltárni a retinaprotekció molekuláris hátterét (Rácz és mtsai 2006).
Munkacsoportunk korábban kimutatta, hogy toxikus retinakárosodást követően a PACAP
hatására az anti-apoptotikus útvonalak aktiválódnak, gátlása során pedig a proapoptotikusak
(Rácz és mtsai 2006). Munkacsoportunk retinális ischaemia kialakulását követően azt
tapasztalta, hogy a PACAP retinoprotektív hatását részben az Akt és MAPK útvonalak
aktiválódásán, valamint az anti-inflammatorikus mechanizmusok beindításán keresztül éri el
(Szabó és mtsai 2012).
A PACAP retinális, illetve más szerveket/szöveteket érintő patológiás folyamatokban
nagyon hatásos molekulának bizonyult, ez indukálta új analógok valamint fragmensek
vizsgálatának igényét, melyek könyebb hozzáférhetőséget és nagyobb stabilitást jelentenének
az esetleges későbbi klinikai felhasználás során (Bourgault és mtsai 2009, 2011). PACAP
carrier-molekulákhoz való kötése feltehetően megnöveli a biológiai membránokon keresztüli
transzportálódását, így könnyebben ki tudja fejteni citoprotektív hatását (Yu és mtsai 2012).

54
Kimutatták, hogy a PACAP miután átjut a sejtmembránokon, receptorokhoz nem köthető
folyamatokban is részt vesz (Doan és mtsai 2012). A fentebb részletezett feltételezésből
fakadóan számos PACAP analóg fejlesztése megkezdődött (Bourgault és mtsai 2009, 2011;
Doan és mtsai 2011). Érdekeség, hogy stroke modellben a stabil analógok nem tudnak nagyobb
mértékű neuroprotekciót kifejteni az eredeti peptidnél (Dejda és mtsai 2011a). Sajnos a számos
próbálkozás ellenére, nem sikerült kifejleszteni az eredeti peptidnél hatásosabb molekulát, vagy
egy olyan stabilabb analógot létrehozni, ami ugyanolyan mértékű protektív hatással
rendelkezik, mint a PACAP1-38. A PACAP 38 aminosavból álló formájának fontosságát az is
mutatja, hogy az evolúció során csaknem változatlan szerkezetű maradt, mind a gerincesek,
mind a gerinctelen egyedekben is (Bourgault és mtsai 2009). Számos tanulmány vizsgálta a
PACAP fragmensek, valamint analógok farmakológiai és receptorokra vonatkozó
tulajdonságait, azonban kevesen nézték ezeknek a formáknak a biológiai hatásosságát (Dejda
és mtsai 2011a). Korábbi tanulmányok kimutatták, hogy a PACAP nem konvencionális
bekötődése szokatlan válaszreakciókat indít el néhány szövetben, amiből azt a következtetést
vonták le, hogy más receptor splice-variánsok is léteznek, melyekről még nincs információnk
(Muller és mtsai 2007; Reglődi és mtsai 2008; Walker és mtsai 2014). Emiatt fontos a
fragmensek biológiai tulajdonságainak kutatása, amihez számos korábbi tanulmány által már
standardizált módszer áll rendelkezésre.
Tanulmányunkban kimutattuk, hogy az általunk vizsgált rövidebb PACAP fragmensek
neuroprotektív hatással nem rendelkeznek, a vizsgált fragmensek többsége nem befolyásolta a
károsodás mértékét. A PACAP6-10 és 4-13-nál néhány morfometriai paraméter esetében azt a
tendenciát tapasztaltuk, hogy az ischaemia okozta károsodás fokozódott. Eredményeink
összhangban vannak korábbi tanulmányokkal is, melyekben leírták, hogy a fragmensek
biológiai viselkedését nem lehet teljesen kiszámítani, akár szövetenként változó lehet. Erre
példa, hogy a peptid N-terminális végéről leszakított aminosavak révén létrejött fragmensek
általában antagonista tulajdonsággal rendelkeznek, mint a leggyakrabban alkalmazott,
korábban a PAC1 receptor szelektív antagonistájaként számon tartott PACAP6-38 (Bourgault
és mtsai 2009, Robberecht és mtsai 1992a). Azóta számos kísérletben igazolták, hogy nemcsak
a PAC1, hanem a VPAC2 receptor jelátvitelét is gátolja, sőt érdekes módon a tracheában,
chondrocytákon, valamint retinoblastoma és cytotrophoblast sejteken agonistaként viselkedik
(Liu és mtsai 2000; Juhász és mtsai 2014; Reglődi és mtsai 2008; Wojcieszak és Zawilska
2014). A C-terminális felől megrövidített peptidek is viselkedhetnek agonistaként valamint
antagonistaként is. Bár a C-terminális felől megrövidített peptidtől agonista hatású
fragmenseket várnánk, hiszen az így létrejött peptidek a peptidázokkal szemben

55
ellenállóbbakká válnak (Borgault és mtsai 2009). Eredményeink összhangban vannak
kutatócsoportunk korábbi eredményeivel, melyekben leírtuk, hogy a rövidebb aminosavszámú
fragmensek antagonistaként is viselkedhetnek, mivel a PACAP6-38 és 6-27 adását követően a
retinadegeneráció mértéke súlyosabb volt MSG indukálta retinakárosodás esetében (Atlasz és
mtsai 2009).
A retinára vonatkozóan kevesebb ismeretünk van a peptidcsalád többi tagjáról. Számtalan
tagjuk megtalálható a gerincesek retinájában, ahol különböző funkciókkal bírnak (Mathis és
Shaeffel 2007; Swedlund és Rosenzweig 1990). Az amakrin sejtek egy kis subpopulációját
képezik a glükagonerg amakrin sejtek, melyek feltételezhetően a vizuális információ
feldolgozásában játszanak szerepet (Diedrich és Schaeffel 2009; Eldred és mtsai 1996). Korábbi
tanulmányokban leírták, hogy a glükagon befolyásolja a bulbus növekedését, ami felveti az
emmetropiában betöltött szerepét. Az exogén glükagon olyan receptorokhoz képes szelektíven
kötődni, amelyekhez csak a glükagon és az oxyntomodulin tud. A myopia endogén
szabályozásban vélhetően a GLP és receptorai játszanak szerepet (Vessey és mtsai 2005).
Korábbi vizsgálatokban a glükagon megnövelte a cAMP szintjét csirke retinális Müller glia
sejtekben úgy, mint a VIP is (Koh és mtsai 1984). A szekretin és glükagon agyi patológiás
folyamatokban betöltött protektív hatásáról kevés tanulmány számol be (Fanne és mtsai 2011a,
2011b; Lam és mtsai 2008). In vivo kísérletben, agyi ischaemiát követően a glükagon
neuroprotektív hatását írták le (Fanne és mtsai 2011b). Ezidáig nem vizsgálták a szekretin és
glükagon esetleges retinoprotektív hatását ischaemiás károsodást követően. Vizsgálatunkban
az alkalmazott koncentrációban sem a szekretin, sem a glükagon esetében nem tapasztaltunk
védő hatást ischaemiás retinopathiában. A korábbi és a jelen tanulmány eredményeit alapul
véve elmondhatjuk, hogy a PACAP erős neuroprotektív hatással bír. A peptidcsaládból még a
VIP-nél tapasztaltak hasonló protektivitást (azonban nagyobb dózisban alkalmazva), a
szekretinnél és glükagonnál nem számolhatunk be hasonló hatásról.
Tanulmányunk megerősítette, hogy a PACAP1-38 és 1-27 izoformákban a leghatásosabbak
retinális ischaemiaban. A 38 valamint 27 aminosavból álló peptideket nem lehet az általunk
vizsgált rövidebb fragmensekkel illetve a peptidcsalád többi tagjával helyettesíteni.
Kísérleteink újfent alátámasztották a PACAP retinoprotektív hatását, ez a további tanulmányok
alapjául is szolgálhat, ezáltal közelebb kerülve az esetleges klinikai felhasználáshoz.

56
5.2. Szemcseppként alkalmazott PACAP1-27 és 1-38 kutatási
eredményeinek összegzése
Tanulmányunkban a PACAP szemcseppes formában való hatásosságát vizsgáltuk solutio
ophthalmica cum benzalkonioban oldva ischaemiás retinopathiában. Eredményeinkkel a
PACAP ezidáig ismert retinoprotektív hatását támasztottuk alá szemcseppes formában is,
valamint sikerült egy olyan vivőanyagot találnunk, ami megfelelő koncentrációban átjuttatja a
PACAP-ot a szem különböző barrierein keresztül egészen a retináig, ahol ki tudja fejteni
korábban már bizonyított protektív hatását.
Az in vivo PACAP kísérleteknél a fő hátráltató tényezőt az jelentette, hogy a peptid eredeti
szerkezeti formáját rövid ideig őrzi meg - mivel a szervezetben megtalálható DPPIV enzim
hasítja, és az így létrejött fragmensek antagonista hatásúak - ezáltal a biológiai hasznosulása
kismértékű (Vaudry és mtsai 2009). Szérumban a felezési ideje csupán néhány perc. A fehérje
ezen tulajdonságai magyarázatul szolgálhatnak korábbi eredményeinkre, miszerint újszülött
patkányoknál excitotoxikus glutamát adását követően a szisztémásan beadott PACAP
kismértékben vagy egyáltalán nem fejtette ki retinoprotektív hatását (Kiss és mtsai 2006). A
rövid metabolizmusa ellenére fontos szerepet játszik bizonyos biológiai folyamatokban,
valamint széles spektrumú hatással rendelkezik. A peptid magas konformációs stabilitása,
illetve a lipid membránokhoz való kötődése, ami megakadályozza in situ enzimatikus
degradációját (Komi és mtsai 2007; Krishnadas és mtsai 2007). A rövid felezési ideje ellenére,
a receptoraihoz való kötődést követően hosszabb távú hatásokat is képes kifejteni (Vaudry és
mtsai 1998).
A PACAP-nak szisztémás hatásai is ismertek intravénás, illetve intraperitoneális adást
követően: csökkenti a vérnyomást, arckipirulást, valamint migrén szerű rohamot idéz elő
migrénre hajlamos betegekben (Bhatt és mtsai 2014; Lamine és mtsai 2015; Schytz és mtsai
2009; Steinhoff és mtsai 2013). Az említett szisztémás kezelést követő mellékhatások
ismeretében, és számos szemészeti megbetegedés tudatában, vizsgálatainkhoz lokálisan
alkalmazható formák (szemcsepp vagy kenőcs) közül választottunk. Krónikus retinális
hipoperfúziós kísérleteinkhez végül a szemcseppes formában feloldott PACAP-ot választottuk.
Morfometriai és morfológiai vizsgálataink során azt tapasztaltuk, hogy a PACAP 27 és 38
aminosavas formában is, solutio ophthalmica cum benzalkonioban oldva, szignifikánsan
csökkentette a BCCAO indukálta retinadegeneráció mértékét. A protektív hatása mind a rétegek
vastagságának, mind a ganglion sejtek rétegében megtalálható sejtek számának kisebb mértékű
csökkenésén is megmutatkozott. A retina rétegeinek szerkezete megtartottabb volt a BCCAO-

57
t követően csak vivőanyag terápiában részesült szemekhez képest. Korábban már kimutatták,
hogy a PACAP1-38 és 1-27 átjut különböző barriereken, mint például a vér-agy gáton (Banks
és mtsai 1993a), valamint a vér-here gáton is (Banks és mtsai 1993b). Vizsgálataink azt
mutatták, hogy a PACAP megfelelő vivőanyagban oldva a szem különböző barrierein keresztül
képes eljutni a retináig. Eredményeinket radioaktívan 125-ös jód izotóppal jelölt PACAP1-27-
el és 1-38-al is alátámasztottuk: a jelölt molekulákat a corneától az üvegtesten keresztül, a
retinában is ki lehetett mutatni. A PACAP solutio ophthalmica cum oldva szemcseppként
alkalmazva kis koncentrációban volt kimutatható az agyban illetve a szérumban, ami a fent
említett mellékhatások elkerülése szempontjából is fontos adatnak bizonyult (PACAP1-27
csúcskoncentrációja a retinában: 220%Inj/g, agyban: 0,25%Inj/g, szérum: 1,25%Inj/ml;
PACAP1-38 csúcskoncentrációja a retinában: 180%Inj/g, agyban: 0,2%Inj/g, szérum:
0,5%Inj/ml).
Összegezve, a korábbi vizsgálatokban főként intravénásan, illetve intraperitoneálisan
alkalmazott PACAP terápiával szemben, az újabb tanulmányokban leírtak alapján lehetőség
nyílik lokális alkalmazásra is, ami csökkenti a szisztémás mellékhatások előfordulásának
gyakoriságát. Intranasálisan alkalmazott PACAP a neurodegeneratív betegségek terápiás
lehetőségeként szolgálhat (Meredith és mtsai 2015; Nonaka és mtsai 2012). Lokálisan a bőrre
kenve dermális vazodilatációt okoz (Warren és mtsai 1992), szájon át történő alkalmazását
pedig a II típusú diabetes terápiájában javasolta egy tanulmány (Langoth és mtsai 2005). A
PACAP lokális szemészeti alkalmazásmódja további corneális hatások vizsgálatára is alkalmas
lehet. Korábbi tanulmányokban a PACAP szemcseppes formában fokozta a corneális
sebgyógyulást (Farkas és mtsai 2010; Ma és mtsai 2015; Nakamachi és mtsai 2011), stimulálta
a könny szekrécióját (Ma és mtsai 2015; Nakamachi és mtsai 2011), és felgyorsította az
idegsejtek regeneratióját (Fukiage és mtsai 2007) (29. ábra). Nemrég közölt tanulmányban a
száraz szem szindróma terápiájában betöltött szerepét vizsgálták (Nakamchi és mtsi 2016). A
szindróma tüneteiként megjelenő corneális keratinizációt valamint a csökkent könnytermelést
vizsgálták. Kimutatták, hogy PACAP KO egerekben PACAP1-38 és1-27 szemcsepp fokozza a
könnytermelést. Ennek hátterében a könnymirigyben található aquaporin csatornáknak is
szerepe van, az aquaporin 5 csatorna megemeli a membrán permeábilitását, ezáltal befolyásolja
a könnytermelést (Ishikawa és mtsai 2005). A csatorna nagymértékben csökkent
immunreaktivitást mutat a könnymirigyben található acináris sejtekben számos humán betegség
esetében: Sjögren szindrómában, néhány krónikus autóimmun betegségben (Tsubota és mtsai
2001). A PACAP hatására megsokszorozódik az aquaporin 5 csatornák száma, ezáltal
fokozódik a könny termelődése is, valamint csökken a corneális keratinizáció mértéke

58
(Nakamachi és mtsai 2016). A lokálisan, szemcseppként alkalmazott PACAP1-27 és 1-38
retinális hatásait azonban ezidáig még nem tanulmányozták. Néhány lokálisan alkalmazott
növekedési faktort már több tanulmányban is vizsgáltak, ilyen például az idegi növekedési
faktor (nerve growth factor - NGF). A lokálisan alkalmazott NGF corneális betegségekben
hatásosnak bizonyult, valamint protektív hatást fejtett ki a retinára is (Lambiase és mtsai 2005;
Steinle és mtsai 2010). Kutyákban vizsgálták az NGF hatását könnymirigy műtétet követően
kialakult száraz szem szindrómában, és azt tapasztalták, hogy kenőcsként alkalmazva enyhített
a betegség tünetein (Coassin és mtsai 2005). Farmakokinetikáját tekintve az NGF eltér a
PACAP-tól: míg az NGF a retinális csúcskoncentrációját a szemcseppentést követően 6 óra
elteltével érte el, addig a PACAP már fél - 1 órát követően (PACAP1-27: 1 óra elteltével,
PACAP1-38: fél óra után). A különbség a fehérjék molekulasúlyából adódhat, a PACAP egy
kis neuropeptid (PACAP1-38: 4534,26Da, PACAP1-27: 3147,61Da), míg az NGF egy nagy
molekulasúlyú fehérje (16,5kDa) (Steinle 2010).
Tanulmányunkban újfent bizonyítottuk, hogy a PACAP retinoprotektív hatását komplex
protekciós mechanizmusokon keresztül fejti ki úgy, mint korábban intravitreálisan alkalmazott
terápia esetén. Anti-apoptotikus, anti-inflammatorikus, valamint anti-oxidáns hatásai korábban
is már igazolódtak (D’Alessandro és mtsai 2014; Rácz és mtsai 2006a, 2006b; Wada és mtsai
2013). Kutatócsoportunk már kimutatta glutamát indukálta excitotoxikus (Rácz és mtsai 2006a)
valamint hipoperfúzió okozta (Szabó és mtsai 2012) retinadegeneratió után, hogy a PACAP
retinán található receptorkötődést követően anti-apoptotikus útvonalakat aktivál (ERK1/2
valamint Akt mediálta szignáltranszdukciós útvonalak), melyek egyúttal gátolják a pro-
apoptotikus jelátviteli útvonalakat (Rácz és mtsai 2006a, 2006b, 2007; Szabó és mtsai 2012).
Az Akt neuroprotektivitásért felelős fehérje, mely foszforilációt követően az Akt kinázon
keresztül az apoptózishoz vezető jelátviteli utakat inaktiválja. Az Akt foszforilációjának gátlása
a fotoreceptorok apoptózisához vezethet, amit már korábban egér modellekben is
tanulmányoztak (Jomary és mtsai 2006). Korábban leírták, hogy az Akt hatására elindult
jelátviteli utak általában összefüggésben vannak a sejtvédelemmel, például retinális
ischaemiában (Dreixler és mtsai 2009). Kísérleteinkben a korábbi eredményeknek megfelelően,
az Akt foszforilációja nagymértékben megemelkedett, ezáltal elindított citoprotektív
útvonalakat. Számos modell alátámasztotta, hogy a MAPK család fontos szerepet játszik a
PACAP indukálta sejtvédelemben (May és mtsai 2010; Vaudry és mtsai 2002). Kisagyi
szemcsesejtekben az ERK és a JNK MAPK közötti egyensúly lényeges tényezőnek bizonyul
egy esetleges károsodás során. A PACAP indukálja az ERK foszforilációját, ami a JNK
foszforilációjának következményes gátlásához vezet, ezáltal megakadályozza a sejtpusztulást

59
(Vaudry és mtsai 2003). Eredményeink megerősítették ezen összefüggést, a PACAP
szignifikánsan megemelte a foszforilált ERK1 és 2 szintjét, mely anti-apoptotikus hatást
biztosított.
Már régóta ismert a PACAP gyulladásban betöltött szerepe, azonban hatását számos
folyamat befolyásolja, mint például a sejt és szövet típusa, a kor, valamint a különböző
patológiás állapotok. A PACAP-ot általánosságban anti-inflammatórikus peptidként tartják
számon (Delgado és mtsai 2003). Retinális ischaemia hatására számos citokin aktiválódik
(Gustavsson és mtsai 2008; Szabó és mtsai 2012; Yoneda és mtsai 2001). Kutatócsoportunk
korábban vizsgálta, hogy a retinában a PACAP képes megváltoztatni a károsodásból fakadó
gyulladásos citokin/kemokin profilt, egy anti-inflammatórikus profil irányába (Szabó és mtsai
2012). A CINC (1-3) család tagjai pro-inflammatórikus kemokinek közé tartoznak, szerepük
van számos gyulladásos folyamatban, főként az akut gyulladásos válaszban játszanak szerepet
(Ghaly és mtsai 2010). Károsodás hatására a szövetekben, így a retinában is a szintjük
megemelkedik, és a szöveti károsodás csökkenésekor kezd a szintjük újból lecsökkeni (Asaga
és mtsai 2008; Junnarkar és mtsai 2010). A PACAP hatására a szintjük szintén lecsökkent
(Nedvig és mtsai 2012). Az interleukinok expressziós változása PACAP kezelést követően
különböző lehet (Szabó és mtsai 2012). Adhéziós molekulák mennyisége számos patológiás,
szemet érintő folyamatban változik (Jonas és mtsai 2010). A mátrix metalloproteinázoknak is
szerepe van a retinális ischaemiás folyamatokban (Mathalone és mtsai 2007). Például, a MIP1-
α szintje megemelkedik ischaemiás retinakárosodást követően (Yoshida és mtsai 2003).
Munkacsoportunk emellett az ICAM, szelektin, TIMP-1 expressziójának fokozódását is
igazolta (Szabó és mtsai 2012). Korábbi kutatásokban leírták, hogy stressz hatására a VEGF
befolyásolja a sejt túlélést a retinában, valamint diabeteses retinopathiában az anti-VEGF
tartalmú készítmények neuroprotektívnek bizonyultak. Intraocularisan adott PACAP hatására
a VEGF szintje lecsökkent (Amato és mtsai 2016). Kísérleteinkben a korábban közölt
eredményekhez hasonlót tapasztaltunk. BCCAO-t követően kialakult ischaemiás károsodás
következtében néhány citokin expressziója nagymértékben fokozódott: CINC-1, MIP-3α,
thymus kemokin, TIMP-1 valamint a VEGF, azonban szemcseppes PACAP1-27 kezelés
hatására szintjük lecsökkent, ezáltal kifejtve az anti-inflammatorikus hatást.
A Müller glia sejtek speciális sejtek, amik egészen az OLM-től az ILM-ig terjednek,
ezáltal a retina teljes vastagságában felelősek a szövet funkciójának és szerkezetének
megőrzéséért. Hatásukat számos mechanizmus aktivációjával fejtik ki, erre példa a kálium és a
glutamát metabolizmus szabályozása (Newman és Reichenbach 1996). A legtöbb ismeretünk a
Müller glia sejt aktivitásának jelét mutató fehérjék közül a GFAP-ről és a glutmain szintázról

60
van (Lam és mtsai 1995). A GFAP egy fehérje, egy intermedier filamentum, ami az
asztrocytákban és az ependimális sejtekben expresszálódik a központi idegrendszerben. A
retinában normális esetben a Müller glia sejtek végtalpaiban van jelen, az ILM rétegben
(Björklund és mtsai 1985). A GFAP expressziója megemelkedik glaukóma (Xue és mtsai
2006), valamint kainát indukálta neuronpusztulás esetén (Honjo és mtsai 2000). Retinális
ischaemia a gliális sejtek aktivációján keresztül patológiás útvonalakat indít el. Wada és
munkatársai kimutatták, hogy a PACAP glia sejtekre is hatással van a retinában: csökkenti az
NMDA indukálta retinadegeneráció mértékét a mikroglia/makrofág rendszer aktivációján
keresztül (Wada és mtsai 2013). Korábbi tanulmányainkban ischaemiás és diabetes okozta
retinadegeneráció esetében is a GFAP szintje megemelkedett, azonban a változás PACAP
kezelés hatására kismértékű volt (Atlasz és mtsai 2010; Szabadfi és mtsai 2012, 2014, 2016).
Vizsgálatunkban a korábbiakban leírtakhoz hasonlót tapasztaltunk: BCCAO-t követően a
GFAP expressziója fokozódott, a retina összes rétegében kimutatható volt, míg a solutio
ophthalmica cum benzalkonioban oldott PACAP1-27 és 1-38 szemcseppel kezelt retinákon
kisebb mennyiségben volt detektálható, és megjelenése a kontroll állatokéhoz hasonlóan inkább
az ILM rétegre korlátozódott.
Az ischaemiás károsodást követő funkcionális javulás mértékének a mérését ERG
vizsgálattal végeztük, ahogy kutatócsoportunk korábbi tanulmányaiban is ezt a módszert
választotta (Dányádi és mtsai 2014). Az ERG vizsgálat reprezentálja és mérhetővé teszi a
bonyolult retinális elektromos aktivitást, ami a pigment epithelium és a legbelső réteg között
megtalálható különböző sejtektől függ (Noell és mtsai 1963). Ez a módszer jól alkalmazható
neuronális degeneráció mérésére, mivel nem invazív, valamint felvilágosítást ad a retina
rétegeiről is. Kutatócsoportunk korábban kimutatta, hogy a BCCAO okozta ischaemiás
károsodás mérésére a b-hullám amplitúdó változása a legalkalmasabb (Dányádi és mtsai 2014).
Ezt alátámasztja az a korábbi tanulmány is, amiben leírták, hogy az oszcillációs potenciál
változása és a b-hullám amplitúdó változása az abnormális retinális keringésről ad
felvilágosítást (Kergoat és Lovasik 1990). Az oszcillációs potenciál változása egy kvantitatív
és objektív indexe az ischaemiás és diabeteses retina diszfunkciójának és manifesztációjának is
(Galloway és mtsai 1971; Yonemura és mtsai 1962). Kutatócsoportunk kimutatta, hogy a
retinában mind a BCCAO, mind a PACAP kezelés neuronális aktivácio változáshoz vezetett.
A legfőbb eltérés a b-hullám amplitúdójában mutatkozott, amit a BCCAO átlagosan
lecsökkentett, azonban PACAP kezelést követően a kontrollhoz közeli amplitúdójú maradt
(Dányádi és mtsai 2014). Kísérleteinkben a korábbiakhoz hasonlóan, BCCAO-t követően a b-

61
hullám amplitúdójának csökkenése bekövetkezett, azonban nem tapasztaltunk PACAP kezelés
hatására szignifikáns mértékű amplitúdó változást, csupán tendenciát.
29. ábra: összefoglaló ábra a PACAP szemcseppként kifejtett hatásairól.
Összegezve, eredményeink alapján kimondható, hogy a PACAP solutio ophthalmica
cum benzalkonioban oldva, szemcseppes formában is ki tudja fejteni eddig ismert
retinoprotektív hatását ischaemiás retinopathiában. A szemcsepp egy könnyen alkalmazható
forma, ezáltal egy lépést jelenthet a PACAP esetleges klinikai felhasználásához vezető úton.

62
6. Összefoglalás
Értekezésem első felében a PACAP peptidcsaládjába tartozó szekretint és glükagont,
valamint különböző aminosavszámú fragmenseit vizsgáltam, intravitreális beadást követően
ischaemiás retinopathiában. Tanulmányunk megerősítette, hogy a PACAP1-38 izoformában a
leghatásosabb retinalis ischaemiában. A 38 valamint 27 aminosavból álló peptideket nem lehet
más rövidebb fragmenssel illetve szekretinnel vagy glükagonnal helyettesíteni. Kísérleteink
újfent alátámasztották a PACAP - ezidáig már többször bizonyított - retinoprotektív hatását.
Eredményeink további tanulmányok alapjául is szolgálhat.
Értekezésem második felében a PACAP kezelés egy új formáját, a szemcseppes terápiát
vizsgáltuk ischaemiás retinopathiában. Vizsgálataink kezdetén a megfelelő vivőanyag
megtalálása volt a cél, amiben oldva a PACAP1-27 ki tudja fejteni a korábban már bizonyított
retinoprotektivitását. Eredményeink vizsgálatához morfológiai és morfometriai analízist
végeztünk. A hatásos vivőanyagnak a solutio ophthalmica cum benzalkonio (solutio
ophthalmica cum benzalkonio) bizonyult, további kísérleteinket emiatt ezzel a szerrel végeztük.
solutio ophthalmica cum benzalkonioban oldott PACAP1-27-el és 1-38-al megismételtük
szövettani vizsgálatainkat, valamint eredményeinket alátámasztottuk immunhisztokémiai és
radioaktív jelöléses vizsgálatokkal is. Vizsgálataink bizonyítják, hogy a PACAP mindkét
biológiailag aktív formája szemcseppként alkalmazva eljut a retináig, és a koncentrációja
megfelelő a retinoprotektív hatás kifejtéséhez. A PACAP1-27 szemcsepp hatásait továbbá
western blot és citokin array vizsgálatokkal is megvizsgáltuk. Western blot vizsgálattal
azonosítottunk a védő hatás hátterében álló folyamatok egy részét, miszerint a kezelés hatására
csökkent a proapoptotikus, viszont emelkedett az anti-apoptotikus fehérjék foszforilációja. A
retinoprotektív hatáshoz hozzájárult a PACAP anti-inflammatorikus, illetve anti-oxidatív
hatása is, melyet a kezelést követő csökkent pro-inflammatorikus citokin expresszió jelzett.
PACAP1-27 szemcseppes terápiát követően funkcionális ERG vizsgálatot is végeztünk.
Eredményeink a korábban már munkacsoportunk által kapott eredményekkel azonos tendenciát
mutatott, vizsgálatainkban a PACAP1-27 a funkcionális károsodás mértékét nem csökkentette
szignifikáns mértékben, azonban a tendencia megegyező volt a korábban leírtakkal. Kísérleteinkben
a PACAP szemészeti felhasználásának ezidáig retinális szinten nem tanulmányozott formáját
vizsgáltuk (szemcsepp), ezáltal eredményeinkkel közelebb kerülve az esetleges klinikai
felhasználáshoz.

63
7. Irodalomjegyzék
Acsády L, Arabadzisz D, Freund TF (1996) Correlated morphological and neurochemical
features identify different subsets of vasoactive intestinal polypeptide-immunoreactive
interneurons in rat hippocampus. Neuroscience 73:299-315.
Adams BA, Lescheid DW, Vickers ED, Crim LW, Sherwood NM (2002) Pituitary adenylate
cyclase activating polypeptide and growth hormone-releasing hormone-like peptide in
sturgeon, whitefish, grayling, flounder and halibut: cDNAsequence, exonskipping and
evolution. Regul Pept 109:27-37.
Agarwal A, Halvorson LM, Legradi G (2005) Pituitary adenylate cyclase-activating
polypeptide (PACAP) mimics neuroendocrine and behavioral manifestations of stress:
Evidence for PKA-mediated expression of the corticotropin-releasing hormone (CRH)
gene. Mol Brain Res 138:45-57.
Ahnaou A, Yon L, Arluison M, Vaudry H, Hannibal J, Hamon M, Adrien J, Bourgin P (2006)
Immunocytochemical distribution of VIP and PACAP in the rat brain stem:
implications for REM sleep physiology. Ann N Y Acad Sci 1070:135-142.
Amato R, Biagioni M, Cammalleri M, Dal Monte M, Casini G (2016) VEGF as
a Survival Factor in Ex Vivo Models of Early Diabetic Retinopathy. Invest Ophthalmol Vis
Sci 57:3066-3076.
Arimura A (1998) Perspectives on pituitary adenylate cyclase activating polypeptide (PACAP)
in the neuroendocrin, endocrine, and nervous system. Jpn J Physiol 48:301-331.
Arimura A (2007) PACAP: the road to discovery. Peptides 28:1617-1619.
Arimura A, Somogyvári-Vigh A, Miyata A, Mizuno K, Coy DH, Kitada C (1991) Tissue
distribution of PACAP as determined by RIA: highly abundant in the rat brain and
testes. Endocrinology 129:2787-2789.
Asaga T, Ueki M, Chujo K, Taie S (2008) JTE-607, an inflammatory cytokine synthesis
inhibitor, attenuates ischaemia/reperfusion-induced renal injury by reducing
neutrophil activation in rats. J Biosci Bioeng 106:22-26.

64
Atlasz T, Szabadfi K, Kiss P, Babai N, Kőszegi Z, Tamás A, Reglődi D, Gábriel R (2008)
PACAP-mediated neuroprotection of neurochemically identified cell types in MSG-
induced retinal degeneration. J Mol Neurosci 36:97-104.
Atlasz T, Babai N, Kiss P, Reglődi D, Tamás A, Szabadfi K, Tóth G, Hegyi O, Lubics A, Gábriel
R (2007) Pituitary adenylate cyclase activating polypeptide is protective in bilateral carotid
occlusion-induced retinal lesion in rats. Gen Comp Endocrinol 153: 108-114.
Atlasz T, Szabadfi K, Kiss P, Tamás A, Tóth G, Reglődi D, Gábriel R (2010) Evaluation of the
protective effects of PACAP with cell-specific markers in ischaemia-induced retinal
degeneration. Brain Res Bull 81: 497-504.
Atlasz T, Szabadfi K, Kiss P, Rácz B, Gallyas F, Tamás A, Gaál V, Marton Z, Gábriel
R, Reglődi D (2010) Review of pituitary adenylatecyclase activating polypeptide in
the retina: focus on the retinoprotective effects. Ann N Y Acad Sci 1200:128-139.
Atlasz T, Szabadfi K, Reglődi D, Kiss P, Tamás A, Tóth G, Molnár A, Szabó K, Gábriel R
(2009) Effects of pituitary adenylate cyclase activating polypeptide and its fragments
on retinal degeneration induced by neonatal monosodium glutamate treatment. Ann N
Y Acad Sci 1163: 348-352.
Atlasz T, Szabadfi K, Kiss P, Marton Z, Griecs M, Hamza L, Gaál V, Biró Z, Tamás A, Hild
G, Nyitrai M, Tóth G, Reglődi D, Gábriel R (2011) Effects of PACAP in UV-A
radiation-induced retinal degeneration models in rats. J Mol Neurosci 43:51-57.
Atlasz T, Váczy A, Werling D, Kiss P, Tamás A, Kovács K, Fábián E, Kvárik T, Mammel B,
Dányádi B, Lőkös E, Reglődi D (2016) Neuroprotective effects of PACAP in the
retina. in Pituitary Adenylate Cyclase Activating Polypeptide – PACAP, edited by
Dóra Reglődi and Andrea Tamás. New York: Springer Nature 501-527.
Babai N, Atlasz T, Tamás A, Reglődi D, Tóth G, Kiss P, Gábriel R (2006) Search for the
optimal monosodium glutamate treatment schedule to study the neuroprotective
effects of PACAP in the retina. Ann NY Acad Sci 1070: 149-155.
Bagnoli P, Dal Monte M, Casini G (2003) Expression of neuropeptides and their receptors in
the developing retina of mammals. Histol Histopathol 18:1219-1242.

65
Bandello F, Corvi F, La Spina C, Benatti L, Querques L, Capuano V, Naysan J, Chen X, Sarraf
D, Parodi MB, Souied E, Freund KB, Querques G (2016) Outcomes of intravitreal
anti-VEGF therapy in eyes with both neovascular age-related macular degeneration
and diabetic retinopathy. Br J Ophthalmol doi: 10.1136/bjophthalmol-2016-308400.
Banks WA, Kastin AJ, Komaki G, Arimura A (1993a) Passage of pituitary adenylate cyclase
activating polypeptide1-27 and pituitary adenylate cyclase activating polypeptide1-38
across the blood-brain barrier. J Pharmacol Exp Ther 267: 690-696.
Banks WA, Kastin AJ, Komaki G, Arimura A (1993b) Pituitary adenylate cyclase activating
polypeptide (PACAP) can cross the vascular component of the blood-testis barrier in
the mouse. J Androl 14:170-173.
Bayliss WM, Starling EH (1902) The mechanism of pancreatic secretion. J Physiol 28:325–
353.
Bell GI (1986) The glucagon superfamily: precursor structure and gene organization. Peptides
7:27-36.
Bhatt DK, Gupta S, Olesen J, Jansen-Olesen I (2014) PACAP-38 infusion causes sustained
vasodilation of the middle meningeal artery in the rat: possible involvement of mast
cells. Cephalalgia 34:877-886.
Björklund H, Bignami A, Dahl D (1985) Immunohistochemical demonstration of glial
fibrillary acidic protein in normal rat Müller glia and retinal astrocytes. Neurosci Lett
54:363-368.
Block F, Schwarz M, Sontag KH (1992) Retinal ischaemia induced by occlusion of both
common carotid arteries in rats as demonstrated by electroretinography. Neurosci Lett
144:124–126.
Borba JC, Henze IP, Silveira MS, Kubrusly RC, Gardino PF, de Mello MC, Hokoç JN, de
Mello FG (2005) Pituitary adenylate cyclase activating polypeptide (PACAP) can act
as a determinant of the tyrosine hydroxylase phenotype of dopaminergic cells during
retina development. Dev Brain Res 156:193-201.
Borbély É, Scheich B, Helyes Z (2013) Neuropeptides in learning and memory. Neuropeptides
47:439-450.

66
Bourgault S, Vaudry D, Botia B, Couvineau A, Laburthe M, Vaudry H, Fournier A (2008)
Novel stable PACAP analogs with potent activity towards the PAC1 receptor. Peptides
29:919-932.
Bourgault S, Chatenet D, Wurtz O, Doan ND, Leprince J, Vaudry H, Fournier A, Vaudry D
(2011) Strategies to convert PACAP from a hypophysiotropic neurohormone into a
neuroprotective drug. Curr Pharm Des 17:1002-1024.
Bourgault S, Vaudry D, Dejda A, Doan ND, Vaudry H, Fournier A (2009) Pituitary adenylate
cyclase-activating polypeptide: focus on structure-activity relationships of a
neuroprotective Peptide. Curr Med Chem 16:4462-4480.
Cavallaro S, Copani A, D'Agata V, Musco S, Petralia S, Ventra C, Stivala F, Travali S,
Canonico PL (1996) Pituitary adenylate cyclase activating polypeptide prevents
apoptosis in cultured cerebellar granule neurons. Mol Pharmacol 50:60-66.
Chang JY, Korolev VV, Wang JZ (1996) Cyclic AMP and pituitary adenylate
cyclaseactivating polypeptide (PACAP) prevent programmed cell death of cultured rat
cerebellar granule cells. Neurosci Lett 206:181-184.
Charlton CG, O’Donohue TL, Miller RL, Jacobowitz DM (1981) Secretin immunoreactivity
in rat and pig brain. Peptides 2:45–91.
Chartrel N, Tonon MC, Vaudry H, Conlon JM (1991) Primary structure of pituitary adenylate
cyclase-activating polypeptide (PACAP) and effects of PACAP on frog pituitary.
Endocrinology 129:3367-3371.
Chen CS, Miller NR (2007) Ocular ischemic syndrome: review of clinical presentations,
etiology, investigation, and management. Compor Ophthalmol 8:17–28.
Chen W, Inui T, Hachiya T, Ochi Y, Nakajima Y, Kajita Y (1993) Stimulatory action of
pituitary adenylate cyclase-activating polypeptide (PACAP) on thyroid gland.
Biochem Biophys Res Commun 194:923-929.
Cheng H, Ding Y, Yu R, Chen J, Wu C (2014) Neuroprotection of a novel cyclopeptide
C*HSDGIC* from the cyclization of PACAP (1-5) in cellular and rodent models of
retinal ganglion cell apoptosis. PLoS One 9:e108090.
Coassin M, Lambiase A, Costa N, De Gregorio A, Sgrulletta R, Sacchetti M, Aloe L, Bonini S
(2005) Efficacy of topical nerve growth factor treatment in dogs affected by dry eye.
Graefes Arch Clin Exp Ophthalmol 243:151-155.

67
Cunha-Reis D, Aidil-Carvalho Mde F, Ribeiro JA (2014) Endogenous inhibition of
hippocampal LTD and depotentiation by vasoactive intestinal peptide VPAC1
receptors Hippocampus 24:1353-1363.
Csanaky K, Doppler W, Tamás A, Kovács K, Tóth G, Reglődi D (2014) Influence of terminal
differentiation and PACAP on the cytokine, chemokine, and growth factor secretion
of mammary epithelial cells. J Mol Neurosci 52: 28-36.
Csernus V, Józsa R, Reglődi D, Hollósy T, Somogyvári-Vigh A, Arimura A (2004) The effect
of PACAP on rhythmic melatonin release of avian pineals. Gen Comp Endocrinol
135:62-69.
D`Agata V, Cavallaro S (1998) Functional and molecular expression of PACAP/VIP receptors
in the rat retina. Mol Brain Res 54:161-164.
D'Alessandro A, Cervia D, Catalani E, Gevi F, Zolla L, Casini G (2014) Protective effects of
the neuropeptides PACAP, substance P and the somatostatin analogue octreotide in
retinal ischemia: a metabolomic analysis. Mol Biosyst 10: 1290-1304.
D'Amico AG, Maugeri G, Reitano R, Bucolo C, Saccone S, Drago F, D’Agata V (2015)
PACAP modulates expression of hypoxia-inducible factors in streptozotocin-induced
diabetic rat retina. J Mol Neurosci 57: 501-509.
Dányadi B, Szabadfi K, Reglődi D, Mihalik A, Dányádi T, Kovács Zs, Batai I, Tamás A, Kiss
P, Tóth G, Gábriel R (2014) PACAP application improves functional outcome of
chronic retinal ischemic injury in rats-evidence from electroretinographic
measurements. J Mol Neurosci 54: 293–299.
Dejda A, Seaborn T, Bourgault S, Touzani O, Fournier A, Vaudry H, Vaudry D (2011a)
PACAP and a novel stable analog protect rat brain from ischemia: Insight into the
mechanisms of action. Peptides 32:1207-1216.
Dejda A, Bourgault S, Doan ND, Létourneau M, Couvineau A, Vaudry H, Vaudry D, Fournier
A (2011b) Identification by photoaffinity labeling of the extracellular N-terminal
domain of PAC1 receptor as the major binding site for PACAP. Biochimie 93: 669-
677.
Delgado M, Abad C, Martinez C, Juarranz MG, Leceta J, Ganea D, Gomariz RP (2003)
PACAP in immunity and inflammation. Ann N Y Acad Sci 992:141-157.

68
Delgado M, Ganea D (2001) Vasoactive intestinal peptide and pituitary adenylate cyclase-
activating polypeptide inhibit expression of Fas ligand in activated T lymphocytes by
regulating c-Myc, NF-kappa B, NF-AT, and early growth factors 2/3. J Immunol
166:1028-1040.
Delwig A, Majumdar S, Ahern K, LaVail MM, Edwards R, Hnasko TS, Copenhagen DR (2013)
Glutamatergic neurotransmission from melanopsin retinal ganglion cells is required
for neonatal photoaversion but not adult pupillary light reflex. PLoS One 8:e83974.
Dickson L, Finlayson K (2009) VPAC an PAC receptors: From ligands to function. Pharmacol
Ther 121:294-316.
Diedrich E, Schaeffel F (2009) Spatial resolution, contrast sensitivity, and sensitivity to
defocus of chicken retinal ganglion cells in vitro. Vis Neurosci 26:467-476.
Doan ND, Bourgault S, Dejda A, Létourneau M, Detheux M, Vaudry D, Vaudry H, Chatenet
D, Fournier A (2011) Design and in vitro characterization of PAC1/VPAC1-selective
agonists with potent neuroprotective effects. Biochem Pharmacol 81:552-561.
Doan ND, Chatenet D, Létourneau M, Vaudry H, Vaudry D, Fournier A (2012) Receptor-
independent cellular uptake of pituitary adenylate cyclase-activating polypeptide.
Biochim Biophys Acta 1823:940-949.
Dragich JM, Loh DH, Wang LM, Vosko AM, Kudo T, Nakamura TJ, Odom IH, Tateyama S,
Hagopian A, Waschek JA, Colwell CS (2010) The role of the neuropeptides PACAP
and VIP in the photic regulation of gene expression in the suprachiasmatic nucleus.
Eur J Neurosci 31:864-875.
Dreixler JC, Hemmert JW, Shenoy SK, Shen Y, Lee HT, Shaikh AR, Rosenbaum DM, Roth
S (2008) The role of Akt/protein kinase B subtypes in retinal ischemic preconditioning.
Exp Eye Res 88:512-521.
Eldred WD, Ammermüller J, Schechner J, Behrens UD, Weiler R (1996) Quantitative
anatomy, synaptic connectivity and physiology of amacrine cells with glucagon-like
immunoreactivity in the turtle retina. J Neurocytol 25:347-364.
Endo K, Nakamachi T, Seki T, Kagami N, Wada Y, Nakamura K, Kishimoto K, Hori
M, Tsuchikawa D, Shinntani N, Hashimoto H, Baba A, Koide R, Shioda S (2011)
Neuroprotective effect of PACAP against NMDA-induced retinal damage in the
mouse. J Mol Neurosci 43:22-29.

69
Fábián E, Reglődi D, Mester L, Szabó A, Szabadfi K, Tamás A, Tóth G, Kovács D (2012)
Effects of PACAP on intracellular signaling pathways in human retinal pigment
epithelial cells exposed to oxidative stress. J Mol Neurosci 48:493-500.
Falcone JA, Salameh TS, Yi X, Cordy BJ, Mortell WG, Kabanov AV, Banks WA (2014)
Intranasal administration as a route for drug delivery to the brain: Evidence for a
unique pathway for albumin. J Pharmacol Exp Ther 351: 54-60.
Faluhelyi N, Reglődi D, Lengvári I, Csernus V (2004) Development of the circadian melatonin
rhythm and the effect of PACAP on melatonin release in the embryonic chicken pineal
gland. An in vitro study. Regul Pept 123:23-28.
Fanne RA, Nassar T, Heyman SN, Hijazi N, Higazi AA (2011b) Insulin and glucagon share
the same mechanism of neuroprotection in diabetic rats: role of glutamate. Am J
Physiol Regul Integr Comp Physiol 301:668-673.
Fanne RA, Nassar T, Mazuz A, Waked O, Heyman SN, Hijazi N, Goelman G, Higazi AA
(2011a) Neuroprotection by glucagon: role of gluconeogenesis. J Neurosurg 114:85-91.
Farkas E, Timmer NM, Domoki F, Mihály A, Luiten PG, Bari F (2005) Post-ischemic
administration of diazoxide attenuates long-term microglial activation in the rat brain
after permanent carotid artery occlusion. Neurosci Lett 387:168–172.
Farkas J, Mester L, Kovács K, Reglődi D, Matkovits A, Gaál V, Biró Zs, Szabó A, Rácz B,
Szabadfi K, Lubics A, Tamás A, Atlasz T, Shioda S, Nakamachi T, Kiss P (2010)
Effects of pituitary adenylate cyclase activating polypeptide (PACAP) in corneal
epithelial regeneration and signal transduction in rats. Presented at 5th International
Peptide Symposium. Kyoto Japan.
Fehér A, Pusch G, Harang G, Gasztonyi B, Papp E, Werling D, Menyhárt M, Komáromy H,
Szapáry L, Fehér G (2011) Aspirin resistance in cerebrovascular patients. Int J Cardiol
152:111-112.
Feigl B (2009) Age-related maculopathy-linking aetiology and pathophysiologycal changes to
the ischaemia hypothesis. Prog Retin Eye Res 28:63–86.
Filipsson K, Sundler F, Hannibal J, Ahrén B (1998) PACAP and PACAP receptors in insulin
producing tissues: localization and effects. Regul Pept 74:167-175.

70
Fukiage C, Nakajima T, Takayama Y, Minagawa Y, Shearer TR, Azuma M (2007) PACAP
induces neurite outgrowth in cultured trigeminal ganglion cells and recovery of corneal
sensitivity after flap surgery in rabbits. Am J Ophthalmol 143: 255-262.
Gaál V, Márk L, Kiss P, Kustos I, Tamás A, Kocsis B, Lubics A, Németh V, Németh A, Lujber
L, Pytel J, Tóth G, Reglődi D (2008) Investigation of the effects of PACAP on the
composition of tear and endolymph proteins. J Mol Neurosci 36:321-329.
Galloway NR, Wells M, Barber C (1971) Changes in the oscillatory potential in relation to
different types diabetic retinography. Vision Res 11:1218.
Gaszner B, Kormos V, Kozicz T, Hashimoto H, Reglődi D, Helyes Zs (2012) The behavioral
phenotype of pituitary adenylate-cyclase activating polypeptide-deficient mice in
anxiety and depression tests is accompanied by blunted c-Fos expression in the bed
nucleus of the stria terminalis, central projecting Edinger-Westphal nucleus, ventral
lateral septum, and dorsal raphe nucleus. Neuroscience 202:283-299.
Gasz B, Rácz B, Rőth E, Borsiczky B, Ferencz A, Tamás A, Cserepes B, Lubics A, Gallyas Jr
F, Tóth G, Lengvári I, Reglődi D (2006) Pituitary adenylate cyclase activating
polypeptide protects cardiomyocytes against oxidative stress-induced apoptosis.
Peptides 27: 87–94.
Ghaly A, Marsh DR (2010) Ischaemia-reperfusion modulates inflammation and fibrosis of
skeletal muscle after contusion injury. Int J Exp Pathol 91:244-255.
Gourlet P, Vandermeers A, Vandermeers-Piret MC, Rathe J, de Neef P, Robberecht P (1996)
C-terminally shortened pituitary adenylate cyclase activating polypeptide (PACAP)
discriminate PACAP I, PACAP II-VIP1 and PACAP II-VIP2 recombinant receptors.
Regul Pept 62:125-130.
Gourlet P, Woussen-Colle MC, Robberecht P, de Neef P, Cauvin A, Vandermeers-Piret MC,
Vandermeers A, Christophe J (1991) Structural requirements for the binding of the
pituitary adenylate cyclase activating polypeptide to receptors and pituitary adenylate
cyclase activation in pancreatic and neuronal membranes. Eur J Biochem 195:534-541.
Gozes I (2008) VIP, from gene to behavior and back: summarizing my 25 years of research. J
Mol Neurosci 36:115-124.

71
Gray SL, Yamaguchi N, Vencová P, Sherwood NM (2002) Temperature-sensitive phenotype
in mice lacking pituitary adenylate cyclase-activating polypeptide. Endocrinology
143:3946-3954.
Grone BP, Zhao S, Chen CC, Fernald RD (2007) Localization and diurnal expression of
melanopsin, vertebrate ancient opsin, and pituitary adenylate cyclase activating
peptide mRNA in a teleost retina. J Biol Rhythms 22:558-561.
Gustavsson C, Agardh CD, Hagert P, Agardh E (2008) Inflammatory markers in nondiabetic
and diabetic rat retinas exposed to ischemia followed by reperfusion. Retina 28:645-652.
Hannibal J (2002) Pituitary adenylate cyclase-activating peptide in the rat central nervous
system: an immunohistochemical and in situ hybridization study. J Comp Neurol
453:389-417.
Hannibal J (2006) Roles of PACAP-containing retinal ganglion cells in circadion timing. Int
Rev Cytol 251:1-39.
Hannibal J, Fahrenkrug J (2004) Target areas innervated by PACAP-immunoreactive retinal
ganglion cells. Cell Tissue Res 316:99-113.
Harmar AJ, Fahrenkrug J, Gozes I, Laburthe M, May V, Pisegna JR, Vaudry D, Vaudry H,
Waschek JA, Said SI (2012) Pharmacology and functions of receptors for vasoactive
intestinal peptide and pituitary adenylate cyclase activating polypeptide: IUPHAR
review 1. Br J Pharmacol 166:4-17.
Harris A, Jonescu-Cuypers CP, Kagemann L, Krieglstein GK (2003) Atlas of Ocular Blood
Flow–Vascular Anatomy, Pathophysiology, and Metabolism; Imprint of Butterworth
Heinemann: Philadelphia, PA, USA.
Honjo M, Tanihara H, Kido N, Inatani M, Okazaki K, Honda Y (2000) Expression of ciliary
neurotrophic factor activated by retinal Müller cells in eyes with NMDA- and kainic
acid-induced neuronal death. Invest Ophthalmol Vis Sci 41:552-560.
Hosoya M, Kimura C, Ogi K, Ohkubo S, Miyamoto Y, Kugoh H, Shimizu M, Onda H,
Oshimura M, Arimura A, et al (1992) Structure of the human pituitary adenylate
cyclase activating polypeptide (PACAP) gene. Biochim Biophys Acta 1129:199-206.
Iemolo A, Seiglie M, Blasio A, Cottone P, Sabino V (2016) Pituitary adenylate cyclase-
activating polypeptide (PACAP) in the central nucleus of the amygdala induces
anxiety via melanocortin receptors. Psychopharmacology 233:3269-3277.

72
Ishihara T, Shigemoto R, Mori K, Takahashi K, Nagata S (1992) Functional expression and tissue
distribution of a novel receptor for vasoactive intestinal polypeptide. Neuron 8:811–819.
Ishikawa Y, Yuan Z, Inoue N, Skowronski MT, Nakae Y, Shono M, Cho G, Yasui M, Agre P,
Nielsen S (2005) Identification of AQP5 in lipid rafts and its translocation to apical
membranes by activation of M3 mAChRs in interlobular ducts of rat parotid gland.
Am J Physiol Cell Physiol 289:C1303-311.
Isobe K, Tatsuno I, Yashiro T, Nanmoku T, Takekoshi K, Kawakami Y, Nakai T (2003)
Expression of mRNA for PACAP and its receptors in intra- and extra-adrenal human
pheochromocytomas and their relationship to catecholamine synthesis. Regul Pept
110:213-217.
Izumi S, Seki T, Shioda S, Zhou CJ, Arimura A, Koide R (2000) Ultrastructural localization of
PACAP immunoreactivity in the rat retina. Ann NY Acad Sci 921:317-320.
Jiang SZ, Eiden LE (2016) Activation of the HPA axis and depression of feeding behavior
induced by restraint stress are separately regulated by PACAPergic neurotransmission
in the mouse. Stress 26:1-9.
Jomary C, Cullen J, Jones SE (2006) Inactivation of the Akt survival pathway during
photoreceptor apoptosis in the retinal degeneration mouse. Invest Ophthalmol Vis Sci
47:1620-1629.
Jonas JB, Tao Y, Neumaier M, Findeisen P (2010) Monocyte chemoattractant protein 1,
intercellular adhesion molecule 1, and vascular cell adhesion molecule 1 in exudative
age-related macular degeneration. Arch Ophthalmol 128:1281-1286.
Joo KM, Chung YH, Kim MK, Nam RH, Lee BL, Lee KH, Cha CI (2004) Distribution of
vasoactive intestinal peptide and pituitary adenylate cyclase-activating polypeptide
receptors (VPAC1, VPAC2, and PAC1 receptor) in the rat brain. J Comp Neurol
476:388-413.
Józsa R, Somogyvári-Vigh A, Reglődi D, Hollosy T, Arimura A (2001) Distribution and daily
variations of PACAP in the chicken brain. Peptides 22:1371-1377.
Juhász T, Matta C, Katona É, Somogyi C, Takács R, Gergely P, Csernoch L, Panyi G, Tóth
G, Reglődi D, Tamás A, Zákány R (2014) Pituitary adenylate cyclase activating
polypeptide (PACAP) signalling exerts chondrogenesis promoting and protecting
effects: Implication of calcineurin as a downstream target. PLoS One 9:e91541.

73
Juhász T, Matta C, Katona É, Somogyi C, Takács R, Hajdú T, Helgadottir SL, Fodor J,
Csernoch L, Tóth G, Bakó É, Reglődi D, Tamás A, Zákány R (2014) Pituitary
adenylate cyclase-activating polypeptide (PACAP) signalling enhances osteogenesis
in UMR-106 cell line. J Mol Neurosci 54:555-73.
Junnarkar SP, Tapuria N, Mani A, Dijk S, Fuller B, Seifalian AM, Davidson BR (2010)
Attenuation of warm ischemia-reperfusion injury in the liver by bucillamine through
decreased neutrophil activation and Bax/Bcl-2 modulation. J Gastroenterol Hepatol
25:1891-1899.
Kastin AJ, Akerstrom V, Pan W (2002) Interactions of glucagon-like peptide-1 (GLP-1) with
the blood-brain barrier. J Mol Neurosci 18:7-14.
Kaur C, Foulds WS, Ling EA (2008) Blood-retinal barrier in hypoxic ischaemic conditions:
basic concepts, clinical features and management. Prog Retin Eye Res 27:622–647.
Kawaguchi C, Isojima Y, Shintani N, Hatanaka M, Guo X, Okumura N, Nagai K, hashimoto
H, Baba A (2010) PACAP-deficient mice exhibit light parameter-dependent
abnormalities on nonvisual photoreception and early activity onset. PLoS One 5:9286.
Kergoat H, Lovasik JV (1990) The effects of altered retinal vascular perfusion pressure on the
white flash scotopic ERG and oscillatory potentials in man. Electroencephalogr Clin
Neurophysiol 75:306–322.
Kiss P, Tamás A, Lubics A, Lengvári I, Szalai M, Hauser D, Horváth Zs, Rácz B, Gábriel
R, Babai N, Tóth G, Reglődi D (2006) Effects of systemic PACAP treatment in
monosodium glutamate-induced behavioral changes and retinal degeneration. Ann N
Y Acad Sci 1070: 365-370.
Koh SW, Kyritsis A, Chader GJ (1984) Interaction of neuropeptides and cultured glial (Müller)
cells of the chick retina: elevation of intracellular cyclic AMP by vasoactive intestinal
peptide and glucagon. J Neurochem 43:199-203.
Komi N, Okawa K, Tateishi Y, Shirakawa M, Fujiwara T, Akutsu H (2007) Structural analysis
of pituitary adenylate cyclase-activating polypeptides bound to phospholipid
membranes by magic angle spinning solid-state NMR. Biochim Biophys Acta 1768:
3001-3011.

74
Kormos V, Gáspár L, Kovács L, Farkas J, Gaszner T, Csernus V, Balogh A, Hashimoto H,
Reglődi D, Helyes Zs, Gaszner B (2016) Reduced response to chronic mild stress in
PACAP mutant mice is associated with blunted FosB expression in limbic forebrain
and brainstem centers. Neuroscience 330:335-358.
Kovács K, Hanto K, Bognár Z, Tapodi A, Bognár E, Kiss NG, Szabó A, Rappai G, Kiss T,
Sümegi B, Gallyas F (2009) Prevalent role of Akt and ERK activation in
cardioprotective effect of Ca2+ channel- and beta-adrenergic receptor blockers. Mol
Cell Biochem 321: 155-164.
Köves K, Arimura A, Somogyvári-Vigh A, Vigh S, Miller J (1990) Immunohistochemical
demonstration of a novel hypothalamic peptide, pituitary adenylate cyclase-activating
polypeptide, in the ovine hypothalamus. Endocrinology 127:264-271.
Krishnadas A, Onyüksel H, Rubinstein I (2003) Interactions of VIP, secretin and PACAP (1-
38) with phospholipids: a biological paradox revisited. Curr Pharm Des 9: 1005-1012.
Kubrusly RC, da Cunha MC, Reis RA, Soares H, Ventura AL, Kurtenbach E, de Mello MC,
de Mello FG (2005) Expression of functional receptors and transmitter enzymes in
cultured Muller cells. Brain Res 1038:141-149.
Lam IP, Siu FK, Chu JY, Chow BK (2008) Multiple actions of secretin in the human body. Int
Rev Cytol 265:159-190.
Lam TK, Chan WY, Kuang GB, Wei H, Shum AS, Yew DT (1995) Differential expression of
glial fibrillary acidic protein (GFAP) in the retinae and visual cortices of rats with
experimental renal hypertension. Neurosci Lett 198:165-168.
Lambiase A, Tirassa P, Micera A, Aloe L, Bonini S (2005) Pharmacokinetics of conjunctivally
applied nerve growth factor in the retina and optic nerve of adult rats. Invest
Ophthalmol Vis Sci 46:3800-3806.
Lamine A, Létourneau M, Doan ND, Maucotel J, Couvineau A, Vaudry H, Chatenet D, Vaudry
D, Fournier A (2015) Characterizations of a synthetic pituitary adenylate cyclase-
activating polypeptide analog displaying potent neuroprotective activity and reduced
in vivo cardiovascular side effects in a Parkinson's disease model. Neuropharmacology
108:440-450.

75
Lang B, Zhao L, McKie L, Forrester JV, McCaig CD, Jackson IJ, Shen S (2010) GABAergic
amacrine cells and visual function are reduced in PAC1 transgenic mice.
Neuropharmacology 58:215-225.
Langoth N, Kalbe J, Bernkop-Schnürch A (2005) Development of a mucoadhesive and
permeation enhancing buccal delivery system for PACAP (pituitary adenylate cyclase-
activating polypeptide). Int J Pharm 296:103-111.
László E, Varga A, Kovács K, Jancsó G, Kiss P, Tamás A, Szakály P, Fülöp B, Reglődi D
(2015) Ischemia/reperfusion-induced kidney injury in heterozygous PACAP-deficient
mice. Transplant Proc 47:2210-2215.
Lee J, Park HJ, Choi HS, Kwon HB, Arimura A, Lee BJ, Choi WS, Chun SY (1999)
Gonadotropin stimulation of pituitary adenylate cyclase-activating polypeptide
(PACAP) messenger ribonucleic acid in the rat ovary and the role of PACAP as a
follicle survival factor. Endocrinology 140:818-826.
Lee LT, Siu FK, Tam JK, Lau IT, Wong AO, Lin MC, Vaudry H, Chow BK (2007) Discovery
of growth hormone-releasing hormones and receptors in nonmammalian vertebrates.
Proc Natl Acad Sci USA 104:2133-2138.
Lenzi L, Coassin M, Lambiase A, Bonini S, Amendola T, Aloe L (2005) Effect of exogenous
administration of nerve growth factor in the retina of rats with inherited retinitis
pigmentosa. Vision Res 45:1491-1500.
Lindén A, Cardell LO, Yoshihara S, Nadel JA (1999) Bronchodilation by pituitary adenylate
cyclase-activating peptide and related peptides. Eur Respir J 14:443-451.
Liu DM, Cuevas J, Adams DJ (2000) VIP and PACAP potentiation of nicotinic AChevoked
currents in rat parasympathetic neurons is mediated by G-protein activation. Eur J
Neurosci 12:2243-2251.
Liu HX, Zhang JJ, Zheng P, Zhang Y (2005) Altered expression of MAP-2, GAP-43, and
synaptophysin in the hippocampus of rats with chronic cerebral hypoperfusion
correlates with cognitive impairment. Mol Brain Res 139:169–177.
Loren I, Emson PC, Fahrenkrug J, Bjorklund A, Alumets J, Hakanson R, Sundler F (1979)
Distribution of vasoactive intestinal polypeptide in the rat and mouse brain.
Neuroscience 4:1953-1976.

76
Lutz EM, Sheward WJ, West KM, Morrow JA, Fink G, Harmar AJ (1993) The VIP2 receptor:
molecular characterisation of a cDNA encoding a novel receptor for vasoactive
intestinal peptide. FEBS Lett 334:3-8.
Ma Y, Zhao S, Wang X, Shen S, Ma M, Xu W, Hong A (2015) A new recombinant PACAP-
derived peptide efficiently promotes corneal wound repairing and lacrimal secretion.
Invest Ophthalmol Vis Sci 56:4336-4349.
Markhotina N, Liu GJ, Martin DK (2007) Contractility of retinal pericytes grown on silicone
elastomer substrates is through a protein kinase A-mediated intracellular pathway in
response to vasoactive peptides. IET Nanobiotechnol 1:44-51.
Masmoudi-Kouki O, Douiri S, Hamdi Y, Kaddour H, Bahdoudi S, Vaudry D, Basille M,
Leprince J, Fournier A, Vaudry H, Tonon MC, Amri M (2011) Pituitary adenylate
cyclase-activating polypeptide protects astroglial cells against oxidative stress-induced
apoptosis. J Neurochem 117:403-411.
Mathalone N, Lahat N, Rahat MA, Bahar-Shany K, Oron Y, Geyer O (2006) The involvement
of matrix metalloproteinases 2 and 9 in rat retinal ischemia. Graefes Arch Clin Exp
Ophthalmol 245:725-732.
Mathis U, Schaeffel F (2007) Glucagon-related peptides in the mouse retina and the effects of
deprivation of form vision. Graefes Arch Clin Exp Ophthalmol 245:267-275.
Matoba Y, Nonaka N, Takagi Y, Imamura E, Narukawa M, Nakamachi T, Shioda S, Banks
WA, Nakamura M (2016) Pituitary adenylate cyclase-activating polypeptide enhances
saliva secretion via direct binding to PACAP receptors of major salivary glands in
mice. Anat Rec 299:1293-1299.
Matsuda K, Azuma M, Maruyama K, Shioda S (2013) Neuroendocrine control of feeding
behavior and psychomotor activity by pituitary adenylate cyclase-activating
polypeptide (PACAP) in vertebrates. Obes Res Clin Pract 7:88.
Matsuda K, Maruyama K (2007) Regulation of feeding behavior by pituitary adenylate
cyclase-activating polypeptide (PACAP) and vasoactive intestinal polypeptide (VIP)
in vertebrates. Peptides 28:1761-1766.
Matsuyama S, Matsumoto A, Hashimoto H, Shintani N, Baba A (2003) Impaired long-term
potentiation in vivo in the dentate gyrus of pituitary adenylate cyclase activating polypeptide
(PACAP) or PACAP type I receptor-mutant mice. Neuroreport 14:2095-2098.

77
May V, Lutz E, MacKenzie C, Schutz KC, Dozark K, Braas KM (2010) Pituitary adenylate
cyclase-activating polypeptide (PACAP)/PAC1HOP1 receptor activation coordinates
multiple neurotrophic signaling pathways: Akt activation through phosphatidylinositol 3-
kinase gamma and vesicle endocytosis for neuronal survival. J Biol Chem 285:9749-9761.
McRory JE, Parker RL, Sherwood NM (1997) Expression and alternative processing of a
chicken gene encoding both growth hormone-releasing hormone and pituitary
adenylate cyclase-activating polypeptide. DNA Cell Biol 16:95-102.
Meredith ME, Salameh TS, Banks WA (2015) Intranasal delivery of proteins and peptides in
the treatment of neurodegenerative diseases. AAPS J 17:780-787.
Mester L, Kovács K, Rácz B, Solti I, Atlasz T, Szabadfi K, Tamás A, Reglődi D (2011)
Pituitary adenylate cyclase-activating polypeptide is protective against oxidative stress
in human retinal pigment epithelial cells. J Mol Neurosci 43:35-43.
Mester L, Szabó A, Atlasz T, Szabadfi K, Reglődi D, Kiss P, Rácz B, Tamás A, Gallyas F Jr,
Sümegi B, Hocsák E, Gábriel R, Kovács K (2009) Protection against chronic
hypoperfusion-induced retinal neurodegeneration by PARP inhibition via activation
of PI3-kinase Akt pathway and suppression of JNK and p38 MAP kinases. Neurotox
Res 16:68-76.
Miyata A, Arimura A, Dahl RR, Minamino N, Uehara A, Jiang L, Culler MD, Coy DH (1989)
Isolation of a novel 38 residue-hypothalamic polypeptide which stimulates
adenylate cyclase in pituitary cells. Biochem Biophys Res Commun 164:567-574.
Miyata A, Jiang L, Dahl RD, Kitada C, Kubo K, Fujino M, Minamino N, Arimura A (1990)
Isolation of a neuropeptide corresponding to the N-terminal 27 residues of the
pituitary adenylate cyclase activating polypeptide with 38 residues (PACAP38).
Biochem Biophys Res Commun 170:643-648.
Moody TW, Ito T, Osefo N, Jensen RT (2011) VIP and PACAP: recent insights into their
functions/roles in physiology and disease from molecular and genetic studies. Curr
Opin Endocrinol Diabetes Obes 18:61-67.
Muller JM, Debaigt C, Goursaud S, Montoni A, Pineau N, Meunier AC, Janet T (2007)
Unconventional binding sites and receptors for VIP and related peptides PACAP and
PHI/PHM: an update. Peptides 28:1655-1666.

78
Murck H, Steiger A, Frieboes RM, Antonijevic IA (2007) Pituitary adenylate cyclase
activating peptide affects homeostatic sleep regulation in healthy young men. Am J
Physiol Endocrinol Metab 292:E853-857.
Murthy KS, Jin JG, Grider JR, Makhlouf GM (1997) Characterization of PACAP receptors
and signaling pathways in rabbit gastric muscle cells. Am J Physiol 272:1391-1399.
Mutt V, Carlquist M, Tatemoto K (1979) Secretin-like bioactivity in extracts of porcine
brain. Life Sci 25:1703–1710.
Nagy AD, Csernus VJ (2007) The role of PACAP in the control of circadian expression of
clock genes in the chicken pineal gland. Peptides 28:1767-1774.
Nakamachi T, Aizawa Y, Ohtaki H, Seki T, Wada Y, Kagami N, Arata S, Hashimoto H,
Shintani N, Baba A, Farkas J, Reglodi D, Shioda S (2011) PACAP stimulates the
lacrimation via aquaporin 5 translocation in mouse. Presented at the 10th International
Symposium on VIP-PACAP and Related Peptides, Eilat, Izrael.
Nakamachi T, Matkovits A, Seki T, Shioda S (2012) Distribution and protective function of
pituitary adenylate cyclase-activating polypeptide in the retina. Front Endocrinol
(Lausanne) 3:145.
Nakamachi T, Ohtaki H, Seki T, Yofu S, Kagami N, Hashimoto H, Shintani N, Baba A, Mark
L, Lanekoff I, Kiss P, Farkas J, Reglodi D, Shioda S (2016) PACAP suppresses dry
eye signs by stimulating tear secretion. Nat Commun 7:12034.
Nakatani M, Seki T, Shinohara Y, Taki C, Nishimura S, Takaki A, Shioda S (2006) Pituitary
adenylate cyclase-activating peptide (PACAP) stimulates production of interleukin-6
in rat Müller cells. Peptides 27:1871-1876.
Nedvig K, Wéber G, Németh J, Kovács K, Reglődi D, Kemény A, Ferencz A (2012) Changes
of PACAP immunoreactivities and cytokine levels after PACAP-38 containing
intestinal preservation and autotransplantation. J Mol Neurosci 48:788-794.
Németh A, Szabadfi K, Fülöp B, Reglődi D, Kiss P, Farkas J, Szalontai B, Gábriel R,
Hashimoto H, Tamás A (2014) Examination of calcium-binding protein expression in
the inner ear of wild-type, heterozygous and homozygous pituitary adenylate cyclase-
activating polypeptide (PACAP)-knockout mice in kanamycin-induced ototoxicity.
Neurotox Res 25:57-67.

79
Newman E and Reichenbach A (1996) The Müller cell: a functional element of the retina.
Trends Neurosci 19:307-312.
Ni JW, Ohta H, Matsumoto K, Watanabe H (1994) Progressive cognitive impairment following
chronic cerebral hypoperfusion induced by permanent occlusion of bilateral carotid
arteries in rats. Brain Res 653:231–236.
Nilsson SF, De Neef P, Robberecht P, Christophe J (1994) Characterization of ocular receptors
for pituitary adenylate cyclase activating polypeptide (PACAP) and their coupling to
adenylate cyclase. Exp Eye Res 58:459-467.
Noell WK (1963) Cellular physiology of the retina. J Opt Sot Am 53:3643.
Nonaka N, Farr SA, Nakamachi T, Morley JE, Nakamura M, Shioda S, Banks WA (2012)
Intranasal administration of PACAP: uptake by brain and regional brain targeting with
cyclodextrins. Peptides 36:168-175.
Ohtaki H, Nakamachi T, Dohi K, Shioda S (2008) Role of PACAP in ischemic neural death.
J Mol Neurosci 36:16-25.
Oka H, Jin L, Kulig E, Scheithauer BW, Lloyd RV (1999) Pituitary adenylate cyclase-
activating polypeptide inhibits transforming growth factor-beta1-induced apoptosis
in a human pituitary adenoma cell line. Am J Pathol 155:1893-1900.
Okada R, Yamamoto K, Ito Y, Mochida H, Tonon MC, Fournier A, Leprince J, Vaudry H,
Kikuyama S (2007) VIP and PACAP stimulate TSH release from the bullfrog
pituitary. Peptides 28:1784-1789.
Olianas MC, Ennas MG, Lampis G, Onali P (1996) Presence of pituitary adenylate cyclase-
activating polypeptide receptors in Y-79 human retinoblastoma cells. J Neurochem
67:1803-1809.
Olianas MC, Ingianni A, Sogos V, Onali P (1997) Expression of pituitary adenylate cyclase-
activating polypeptide (PACAP) receptors and PACAP in human fetal retina. J
Neurochem 69:1213-1218.
Onali P, Olianas MC (1994) PACAP is a potent and highly effective stimulator of adenylyl
cyclase activity in the retinas of different mammalian species. Brain Res 641:132-134.
Osborne NN, Casson RJ, Wood JP, Chidlow G, Graham M, Melena J (2004) Retinal ischemia:
mechanisms of damage and potential therapeutic strategies. Prog Retin Eye Res 23:91–147.

80
Osborne NN, Safa R, Nash MS (1999) Photoreceptors are preferentially aVected in the rat
retina following permanent occlusion of the carotid arteries. Vis Res 39:3995–4002.
Otto C, Hein L, Brede M, Jahns R, Engelhardt S, Gröne HJ, Schütz G (2004) Pulmonary
hypertension and right heart failure in pituitary adenylate cyclase-activating
polypeptide type I receptor-deficient mice. Circulation 110:3245-3251.
Pirger Z, László Z, Kemenes I, Tóth G, Reglődi D, Kemenes G (2010) A homolog of the
vertebrate pituitary adenylate cyclase-activating polypeptide is both necessary and
instructive for the rapid formation of associative memory in an invertebrate. J
Neurosci 30:13766-13773.
Pisegna JR, Wank SA (1993) Molecular cloning and functional expression of the
pituitary adenylate cyclase-activating polypeptide type I receptor. Proc Natl Acad Sci
USA 90:6345–6349.
Pournaras CJ, Rungger-Brandle E, Riva CE, Hardarson SH, Stefansson E (2008) Regulation
of retinal blood flow in health and disease. Prog Retin Eye Res 27:284–330.
Purrier N, Engeland WC, Kofuji P (2014) Mice deficient of glutamatergic signaling from
intrinsically photosensitive retinal ganglion cells exhibit abnormal circadian
photoentrainment. PLoS One 9:e111449.
Rábl K, Reglődi D, Bánvölgyi T, Somogyvári-Vigh A, Lengvári I, Gábriel R, Arimura A
(2002) PACAP inhibits anoxia-induced changes in physiological responses in
horizontal cells in the turtle retina. Regul Pept 109:71-74.
Rácz B, Gallyas F Jr, Kiss P, Tamás A, Lubics A, Lengvári I, Rőth E, Tóth G, Hegyi O, Verzár
Zs, Fabricsek Cs, Reglődi D (2007) Effects of pituitary adenylate cyclase activating
polypeptide (PACAP) on the PKA-Bad-14-3-3 signaling pathway in glutamate-
induced retinal injury in neonatal rats. Neurotox Res 12:95-104.
Rácz B, Gallyas F Jr., Kiss P, Tóth G, Hegyi O, Gasz B, Borsiczky B, Ferencz A, Rőth E,
Tamás A, Lengvári I, Lubics A, Reglődi D (2006b) The neuroprotective effects of
PACAP in monosodium glutamate-induced retinal lesion involves inhibition of
proapoptotic signaling pathways. Regul Pept 137:20-26.
Rácz B, Horváth G, Faluhelyi N, Nagy AD, Tamás A, Kiss P, Gallyas F Jr, Tóth G, Gaszner
B, Csernus V, Reglődi D (2008) Effects of PACAP on the circadian changes of
signaling pathways in chicken pinealocytes. J Mol Neurosci 36:220-226.

81
Rácz B, Tamás A, Kiss P, Tóth G, Gasz B, Borsiczky B, Ferencz A, Gallyas F Jr, Rőth E, and
Reglődi D (2006a) Involvement of ERK and CREB signalling pathways in the
protective effect of PACAP on monosodium glutamate-induced retinal lesion. Ann NY
Acad Sci 1070:507-511.
Reglődi D (2009) A PACAP neuroprotektív és általános citoprotektív hatásainak vizsgálata in
vitro és in vivo modellekben. MTA Doktori értekezés. Pécsi Tudományegyetem,
Általános Orvostudományi Kar, Anatómiai Intézet, Pécs.
Reglődi D, Börzsei R, Bagoly T, Boronkai A, Rácz B, Tamás A, Kiss P, Horváth G, Brubel
R, Németh J, Tóth G, Helyes Z (2008) Agonistic behavior of PACAP6-38 on sensory
nerve terminals and cytotrophoblast cells. J Mol Neurosci 36:270-278.
Reglődi D, Kiss P, Lubics A, Tamás A (2011) Review of the protective effects of PACAP in
models of neurodegenerative diseases in vitro an in vivo. Curr Pharm Des 17:962-972.
Reglődi D, Kiss P, Tamás A, Lengvári I (2003) The effects of PACAP and PACAP antagonist
on the neurobehavioral development of newborn rats. Behav Brain Res 140:131-139.
Reglődi D, Kiss P, Szabadfi K, Atlasz T, Gábriel R, Horváth G, Szakaly P, Sándor B, Lubics
A, László E, Farkas J, Matkovits A, Brubel R, Hashimoto H, Ferencz A, Vincze
A, Helyes Z, Welke L, Lakatos A, Tamás A (2012) PACAP is an endogenous
protective factor – insights from PACAP deficient mice. J Mol Neurosci 48:482-492.
Reglődi D, Renaud J, Tamás A, Tizabi Y, Socías B, Del-Bel E, Raisman-Vozari R (2015)
Novel tactics for neuroprotection in Parkinson's disease: Role of antibiotics,
polyphenols and neuropeptides. Prog Neurobiol S0301-0082 (15):00128-8.
Rékasi Z, Czömpöly T (2002) Accumulation of rat pineal serotonin N-acetyltransferase mRNA
induced by pituitary adenylate cyclase activating polypeptide and vasoactive intestinal
peptide in vitro. J Mol Endocrinol 28:19-31.
Robberecht P, Gourlet P, De Neef P, Woussen-Colle MC, Vandermeers-Piret MC,
Vandermeers A, Christophe J (1992a) Structural requirements for the occupancy of
pituitary adenylate-cyclase-activating-peptide (PACAP) receptors and adenylate
cyclase activation in human neuroblastoma NB-OK-1 cell membranes. Discovery of
PACAP (6-38) as a potent antagonist. Eur J Biochem 207:239-246.

82
Robberecht P, Gourlet P, De Neef P, Woussen-Colle MC, Vandermeers-Piret MC,
Vandermeers A, Christophe J (1992b) Receptor occupancy and adenylate cyclase
activation in AR 4-2J rat pancreatic acinar cell membranes by analogs of pituitary
adenylate-cyclase-activating-peptide amino-terminally shortened or modified at
position 1,2,3,20, or 21. Mol Pharmacol 42:347-355.
Röhlich P (2006) Szövettan Tankönyv: 465-481.
Rőth E, Wéber G, Kiss P, Horváth G, Tóth G, Gasz B, Ferencz A, Gallyas F Jr, Reglődi D,
Rácz B (2009) Effects of PACAP and preconditioning against ischemia/reperfusion-
induced cardiomyocyte apoptosis in vitro. Ann NY Acad Sci 1163:512-516.
Rudecki AP, Gray SL (2016) PACAP in the defense of energy homeostasis. Trends Endocrinol
Metab 9:620-632.
Sakamoto K, Liu C, Kasamatsu M, Pozdeyev NV, Iuvone PM, Tosini G (2005) Dopamine
regulates melanopsin mRNA expression in intrinsically photosensitive retinal
ganglion cells. Eur J Neurosci 22:3129-3136.
Salameh TS, Bullock KM, Hujoel IA, Niehoff ML, Wolden-Hanson T, Kim J, Morley JE, Farr
SA, Banks WA (2015) Central nervous system delivery of intranasal insulin:
mechanisms of uptake and effects on cognition. J Alzheimers Dis 47:715-728.
Sándor K, Bölcskei K, McDougall JJ, Schuelert N, Reglődi D, Elekes K, Pethő G, Pintér E,
Szolcsányi J, Helyes Z (2009) Divergent peripheral effects of pituitary adenylate
cyclase-activating polypeptide-38 on nociception in rats and mice. Pain 141:143-150.
Sarti C, Pantoni L, Bartolini L, Inzitari D (2002) Persistent impairment of gait performances
and working memory after bilateral common carotid artery occlusion in the adult
Wistar rat. Behav Brain Res 136:13–20.
Schmidt-Kastner R, Aguirre-Chen C, Saul I, Yick L, Hamasaki D, Busto R, Ginsberg MD
(2005) Astrocytes react to oligemia in the forebrain induced by chronic bilateral
common carotid artery occlusion in rats. Brain Res 1052:28–39.
Schytz HW, Birk S, Wienecke T, Kruuse C, Olesen J, Ashina M (2009) PACAP38 induces
migraine-like attacks in patients with migraine without aura. Brain 132:16-25.
Seki T, Itoh H, Nakamachi T, Endo K, Wada Y, Nakamura K, Shioda S (2011) Suppression of
rat retinal ganglion cell death by PACAP following transient ischemia induced by high
intraocular pressure. J Mol Neurosci 43:30-34.

83
Seki T, Itoh H, Nakamachi T, Shioda S (2008) Suppression of ganglion cell death by PACAP
following optic nerve transection in the rat. J Mol Neurosci 36:57-60.
Seki T, Shioda S, Izumi S, Arimura A, Koide R (2000a) Electron microscopic observation of
pituitary adenylate cyclase activating polypeptide (PACAP)-containing neurons in the
rat retina. Peptides 21:109-113.
Seki T, Izumi S, Shioda S, Zhou CJ, Arimura A, Koide R (2000b) Gene expression for PACAP
receptor mRNA in the rat retina by in situ hybridization and in situ RT-PCR. Ann NY
Acad Sci 921:366-369.
Seki T, Nakatani M, Taki C, Shinohara Y, Ozawa M, Nishimura S, to H, Shioda S (2006)
Neuroprotective effects of PACAP against kainic acid-induced neurotoxicity in rat
retina. Ann NY Acad Sci 1070:531-534.
Seki T, Shioda S, Ogino D, Nakai Y, Arimura A, Koide R (1997) Distribution and
ultrastructural localization of a receptor for pituitary adenylate cyclase activating
polypeptide and its mRNA in the rat retina. Neurosci Lett 238:127-130.
Sherwood NM, Krueckl SL, McRory JE (2000) The origin and function of hte pituitary
adenylate cyclase activating polypeptide (PACAP)/glucagon superfamily. Endocr Rev
21:619-670.
Shioda S, Nakamachi T (2015) PACAP as a neuroprotective factor in ischemic neuronal
injuries. Peptides 72:202-207.
Shioda S, Ohtaki H, Nakamachi T, Dohi K, Watanabe J, Nakajo S, Arata S, Kitamura S, Okuda
H, Takenoya F, Kitamura Y (2006) Pleiotropic functions of PACAP in the CNS:
neuroprotection and neurodevelopment. Ann NY Acad Sci 1070:550-60.
Shioda S, Zhou CJ, Ohtaki H, Yada T (2003) PACAP receptor signaling. Pituitary Adenylate
Cyclase-Activating Polypeptide 95-124. Springer 2003
Shoge K, Mishima HK, Saitoh T, Ishihara K, Tamura Y, Shiomi H, Sasa M (1999) Attenuation
by PACAP of glutamate-induced neurotoxicity in cultured retinal neurons. Brain Res
839:66-73.
Silveira MS, Costa MR, Bozza M, Linden R (2002) Pituitary adenylate cyclase activating
polypeptide prevents induced cell death in retinal tissue through activation of cyclic
AMP-dependent protein kinase. J Biol Chem 277:16075-80.

84
Slakter JS, Spertus AD, Weissman SS, Henkind P (1984) An experimental model of carotid
artery occlusive disease. Am J Ophtalmol 97:168–172.
Somogyvari-Vigh A, Reglődi D (2004) Pituitary adenylate cyclase activating polypeptide: a
potential neuroprotective peptide. Review. Curr Pharm Des 10:2861-2889.
Spertus AD, Slakter JS, Weissman SS, Henkind P (1984) Experimental carotid occlusion:
fundoscopic and fuorescein angiographic findings. Br J Ophtalmol 68:47–57.
Steinhoff M, Schauber J, Leyden JJ (2013) New insights into rosacea pathophysiology: a
review of recent findings. J Am Acad Dermatol 69:S15-26.
Steinle JJ (2010) Topical administration of adrenergic receptor pharmaceutics and nerve
growth factor. Clin Ophthalmol 4:605-610.
Stevens WD, Fortin T, Pappas BA (2002) Retinal and optic nerve degeneration after chronic
carotid ligation. Stroke 33:1107–1112.
Süveges I (2013) Szemészet Tankönyv, Budapest.
Swedlund AP, Rosenzweig SA (1990) Characterization of vasoactive intestinal peptide
receptors in retina. Exp Eye Res 51:317-23.
Szabadfi K, Atlasz T, Kiss P, Reglődi D, Szabó A, Kovács K, Szalontai B, Sétáló Gy, Bánki
E, Csanaky K, Tamás A, Gábriel R (2012) Protective effects of the neuropeptide
PACAP in diabetic retinopathy. Cell Tissue Res 348:37-46.
Szabadfi K, Mester L, Reglődi D, Kiss P, Babai N, Rácz B, Kovács K, Szabó A, Tamás A,
Gábriel R, Atlasz T (2010) Novel neuroprotective strategies in ischemic retinal lesions.
Int J Mol Sci 11:544-561.
Szabadfi K, Reglődi D, Szabó A, Szalontai B, Valasek A, Sétáló Gy Jr, Kiss P, Tamás
A, Wilhelm M, Gábriel R (2016) Pituitary adenylate cyclase activating polypeptide, a
potential therapeutic agent for diabetic retinopathy in rats: focus on the vertical
information processing pathway. Neurotox Res 29:432-446.
Szabadfi K, Szabó A, Kiss P, Reglődi D, Sétáló G Jr, Kovács K, Tamás A, Tóth G, Gábriel R
(2014) PACAP promotes neuron survival in early experimental diabetic retinopathy.
Neurochem Int 64:84-91.

85
Szabó A, Dányádi B, Bognár E, Szabadfi K, Fábián E, Kiss P, Mester L, Manavalan S, Atlasz
T, Gábriel R, Tóth G, Tamás A, Reglődi D, Kovács K (2012) Effect of PACAP on
MAP kinases, Akt and cytokine expressions in rat retinal hypoperfusion. Neurosci Lett
523:93-98.
Tajiri M, Hayata-Takano A, Seiriki K, Ogata K, Hazama K, Shintani N, Baba A, Hashimoto
H (2012) Serotonin 5-HT(7) receptor blockade reverses behavioral abnormalities in
PACAP-deficient mice and receptor activation promotes neurite extension in primary
embryonic hippocampal neurons: therapeutic implications for psychiatric disorders. J
Mol Neurosci 48:473-81.
Tsubota K, Hirai S, King LS, Agre P, Ishida N (2001) Defective cellular trafficking of lacrimal
gland aquaporin-5 in Sjögren's syndrome. Lancet 357:688-689.
Uchida D, Arimura A, Somogyvari-Vigh A, Shioda S, Banks WA (1996) Prevention of
ischemia-induced death of hippocampal neurons by pituitary adenylate cyclase
activating polypeptide. Brain Res 736:280-286.
Usdin TB, Bonner TI, Mezey E (1994) Two receptors for vasoactive intestinal polypeptide with
similar specificity and complementary distributions. Endocrinology 135:2662-2680.
Vandermeers A, Vanderborre S, Hou X, de Neef P, Robberecht P, Vandermeers-Piret MC,
Christophe J (1992) Antagonistic properties are shifted back to agonistic properties by
futher N-terminal shortening of pituitary adenylate-cyclase-activating-peptide in
human neuroblastoma NB-OK-1 cell membranes. Eur J Biochem 208:815-819.
Vaudry D, Falluel-Morel A, Basille M, Pamantung TF, Fontaine M, Fournier A, Vaudry H,
Gonzalez BJ (2003) Pituitary adenylate cyclase-activating polypeptide prevents C2-
ceramide-induced apoptosis of cerebellar granule cells. J Neurosci Res 72:303-316.
Vaudry D, Falluel-Morel A, Bourgault S, Basille M, Burel D, Wurtz O, Fournier A, Chow
BK, Hashimoto H, Galas L, Vaudry H (2009) Pituitary adenylate cyclase activating
polypeptide and its receptors: 20 years after the discovery. Pharmacol Rev 61: 283-357.
Vaudry D, Gonzalez BJ, Basille M, Anouar Y, Fournier A, Vaudry H (1998) Pituitary
adenylate cyclase-activating polypeptide stimulates both c-fos gene expression and
cell survival in rat cerebellar granule neurons through activation of the protein kinase
A pathway. Neuroscience 84:801-812.

86
Vaudry D, Gonzalez BJ, Basille M, Yon L, Fournier A, Vaudry H (2000) Pituitary adenylate
cyclase-activating polypeptide and its receptors: from structure to functions.
Pharmacol Rev 52:269-324.
Vaudry D, Pamantung TF, Basille M, Rousselle C, Fournier A, Vaudry H, Beauvillain JC,
Gonzalez BJ (2002) PACAP protects cerebellar granule neurons against oxidative
stress-induced apoptosis. Eur J Neurosci 15:1451-1460.
Vessey KA, Lencses KA, Rushforth DA, Hruby VJ, Stell WK (2005) Glucagon receptor
agonists and antagonists affect the growth of the chick eye: a role for glucagonergic
regulation of emmetropization? Invest Ophthalmol Vis Sci 46:3922-3931.
Vigneswara V, Esmaeili M, Deer L, Berry M, Logan A, Ahmed Z (2015) Eye drop delivery of
pigment epithelium-derived factor-34 promotes retinal ganglion cell neuroprotection
and axon regeneration. Mol Cell Neurosci 68:212-221.
Wada Y, Nakamachi T, Endo K, Seki T, Ohtaki H, Tsuchikawa D, Hori M, Tsuchida
M, Yoshikawa A, Matkovits A, Kagami N, Imai N, Fujisaka S, Usui I, Tobe K, Koide
R, Takahashi H, Shioda S (2013) PACAP attenuates NMDA-induced retinal damage
in association with modulation of the microglia/macrophage status into an acquired
deactivation subtype. J Mol Neurosci 51:493-502.
Walker CS, Sundrum T, Hay DL (2014) PACAP receptor pharmacology and agonist bias:
Analysis in primary neurons and glia from the trigeminal ganglia and transfected cells.
Br J Pharmacol 171:1521-1533.
Warren JB, Cockcroft JR, Larkin SW, Kajekar R, Macrae A, Ghatei MA, Bloom SR (1992)
Pituitary adenylate cyclase activating polypeptide is a potent vasodilator in humans. J
Cardiovasc Pharmacol 20:83-87.
Warren JB, Donnelly LE, Cullen S, Robertson BE, Ghatei MA, Bloom SR, MacDermot J
(1991) Pituitary adenylate cyclase-activating polypeptide: a novel, long-lasting,
endothelium independent vasorelaxant. Eur J Pharmacol 197:131-134.
Waschek JA (2002) Multiple actions of pituitary adenylyl cyclase activating peptide in nervous
system development and regeneration. Dev Neurosci 24:14-23.
Winzell MS, Ahrén B (2007) Role of VIP and PACAP in islet function. Peptides 28:1805-1813.
Wojcieszak J, Zawilska JB (2014) PACAP38 and PACAP6-38 exert cytotoxic activity against
human retinoblastoma Y79 cells. J Mol Neurosci 54:463-468.

87
Xue LP, Lu J, Cao Q, Hu S, Ding P, Ling EA (2006) Müller glial cells express nestin coupled
with glial fibrillary acidic protein in experimentally induced glaucomain the rat retina.
Neuroscience 139:723-732.
Yamamoto H, Schmidt-Kasner R, Hamasaki DI, Yamamoto H, Parel JM (2006) Complex
neurodegeneration in retina following moderate ischemia induced by bilateral
common carotid artery occlusion in Wistar rats. Exp Eye Res 82:767–779.
Yan Y, Zhou X, Pan Z, Ma J, Waschek JA, DiCicco-Bloom E (2013) Pro- and anti-mitogenic
actions of pituitary adenylate cyclase-activating polypeptide in developing cerebral
cortex: potential mediation by developmental switch of PAC1 receptor mRNA
isoforms. J Neurosci 33:3865-3878.
Yoneda S, Tanihara H, Kido N, Honda Y, Goto W, Hara H, Miyawaki N (2001) Interleukin-
1beta mediates ischemic injury in the rat retina. Exp Eye Res 73:661-667.
Yonemura D, Aoki T, Tzuzuki K (1962) Electroretinogram in diabetic retinopathy. Arch
Ophthalmol 68:19–24.
Yoshida S, Yoshida A, Ishibashi T, Elner SG, Elner VM (2003) Role of MCP-1 and MIP-
1alpha in retinal neovascularization during postischemic inflammation in a mouse
model of retinal neovascularization. J Leukoc Biol 73:137-144.
Young A (2005) Inhibition of glucagon secretion. Adv Pharmacol 52:151-171.
Yu R, Guo X, Huang L, Zeng Z, Zhang H (2012) The novel peptide PACAP-TAT with
enhanced traversing ability attenuates the severe lung injury induced by repeated
smoke inhalation. Peptides 38:142-149.
Yu R, Yang Y, Cui Z, Zheng L, Zeng Z, Zhang H (2014) Novel peptide VIP-TAT with higher
affinity for PAC1 inhibited scopolamine induced amnesia. Peptides 60:41-50.
Zhang L, Chow BK (2014) The central mechanisms of secretin in regulating multiple
behaviors. Front Endocrinol 5:77.
Zhang XY, Hayasaka S, Chi ZL, Cui HS, Hayasaka Y (2005) Effect of pituitary adenylate
cyclase-activating polypeptide (PACAP) on IL-6, IL-8, and MCP-1 expression in
human retinal pigment epithelial cell line. Curr Eye Res 30:1105-1111.
Zhu L, Tamvakopoulos C, Xie D, Dragovic J, Shen X, Fenyk-Melody JE, Schmidt K, Bagchi
A, Griffin PR, Thornberry NA, Sinha Roy R (2003) The role of dipeptidyl peptidase
IV in the cleavage of glucagon family peptides: in vivo metabolism of pituitary
adenylate cyclase activating polypeptide-(1-38). J Biol Chem 278:22418-22423.

88
8. Publikációs lista
8.1. Az értekezés alapjául szolgáló publikációk
Werling D, Reglődi D, William AB, Theresa SS, Kovács K, Kvárik T,
Váczy A, Kovács L, Lőkös E, Tamás A, Tóth G, Biró Zs, Atlasz T (2016)
Ocular delivery of PACAP1-27 protects the retina from ischemic damage in
rats. Invest Ophthalmol Vis Sci 57:6683-6691. IF: 3,427
Werling D, Reglődi D, Kiss P, Tóth G, Szabadfi K, Tamás A, Biró Z, Atlasz
T (2014) Investigation of PACAP fragments and related peptides in chronic
retinal hypoperfusio. J Ophthalmol 2014:563812. IF: 1,425
Atlasz T, Váczy A, Werling D, Kiss P, Tamás A, Kovács K, Fábián E,
Kvárik T, Mammel B, Dányádi B, Lőkös E, Reglődi D (2016)
Neuroprotective effects of PACAP in the retina. in Pituitary Adenylate
Cyclase Activating Polypeptide – PACAP, edited by Dóra Reglődi and
Andrea Tamás. New York: Springer Nature 501-527.
Témához kapcsolódó publikációk összesített impakt faktora: 4,852
8.2. Az értekezés témájába közvetlenül nem illeszkedő egyéb
közlemények
Fehér A, Pusch G, Harang G, Gasztonyi B, Papp E, Werling D, Menyhárt
M, Komáromy H, Szapáry L, Feher G (2011) Aspirin resistance in
cerebrovascular patients. Int J Cardiol 152:111-112. IF: 3,47

89
Fehér A, Pusch G, Harang G, Gasztonyi B, Papp E, Werling D, Menyhárt
M, Komáromy H, Szapáry L, Feher G (2011) Az acetilszalicilsav-
rezisztencia klinikai jelentősége a cerebrovascularis betegek esetében. LAM
21:719-724.
Kvárik T, Mammel B, Reglődi D, Kovács K, Werling D, Bede B, Váczy A,
Fábián E, Tóth G, Kiss P, Tamás A, Ertl T, Gyarmati J, Atlasz T (2016)
PACAP is protective in a rat model of retinopathy of prematurity. J Mol
Neurosci 60:179-85. IF: 2,352
Témához közvetlenül nem kapcsolódó publikációk összesített impakt faktora: 5,822
8.3. Konferencia szereplések
Fehér G, Werling D, Menyhárt M, Szapáry L (2011) Aspirin non-responsio
cerebrovascularis betegségekben. Magyar Stroke Társaság X. konferenciája,
Mickolc.
Fehér G, Werling D, Menyhárt M, Szapáry L (2011) Statinterápia szerepe
az ishemiás cerebrovascularis betegségek primer és secunder
prevenciójában. Magyar Stroke Társaság X. konferenciája, Mickolc.
Fehér G, Werling D, Menyhárt M, Szapáry L (2011) Infektív endocarditis a
neurológia szemszögéből. Magyar Stroke Társaság X. konferenciája,
Mickolc.
Horváth R, Werling D, Bosnyák E, Deli G, Csécsei P, Komáromi H, Imre
M, Bóné B, Gyimesi Cs, Fehér G, Komoly S, Janszky J, Szapáry L (2011)
Epilepszia mint az agyi vénás thrombosisok következménye. Magyar Stroke
Társaság X. konferenciája, Mickolc.
Werling D, Atlasz T, Kiss P, Tamás A, Szabadfi K, Gábriel R, Tóth G,
Reglődi D (2013) Investigating the retinoprotective effects of PACAP
fragments in ischemic retinopathy. The 11th International Symposium on
VIP, PACAP and Related Peptides, Pécs.
Werling D, Atlasz T, Kiss P, Tamás A, Szabadfi K, Gábriel R, Tóth G,
Reglődi D (2013) Investigating the retinoprotective effects of PACAP

90
fragments in ischemic retinopathy. Magyar Szemorvostársaság Retina
Szekció Kongresszusa, Magyarország, Velence.
Werling D, Atlasz T, Kiss P, Tamás A, Szabadfi K, Gábriel R, Tóth G,
Reglődi D (2014) Investigating the retinoprotective effects of PACAP
fragments in ischemic retinopathy. IBRO Workshop, Debrecen.
Werling D, Kvárik T, Varga R, Nagy N, Mayer F, Váczy A, Reglődi D, Kiss
P, Tamás A, Biró Zs, Tóth G, Atlasz T (2014) Szemcseppként alkalmazott
PACAP retinoprotektív hatásának vizsgálata ischaemiás retinopathiában.
Magyar Szemorvostársaság 2014. évi Kongresszusa, Pécs.
Kvárik T, Balogh M, Bloomfield S, Werling D, Zhang Yi, Völgyi B, Atlasz T,
Reglődi D (2014) Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP)
Prevents Functional Disturbances in the Mouse Retina Monosodium Glutamate
(MSG) Induced. 11th ISOPT Clinical – The International Symposium on Ocular
Pharmacology and Therapeutics, Iceland, Reyklavic.
Mammel B, Kvárik T, Atlasz T, Werling D, Reglődi D, Kiss P, Ertl T,
Dányádi B, Gyarmati J (2014) Effects of Early Hyperglycaemia on the
Retinal Structure of OIR Rats. 11th ISOPT Clinical – The International
Symposium on Ocular Pharmacology and Therapeutics, Iceland, Reyklavic.
Kvárik T, Mammel B, Werling D, Kiss P, Ertl T, Gyarmati J, Dányádi B,
Reglődi D, Atlasz T (2014) PACAP Administration can Ameliorate Vascular
Changes in Retinopathy of Prematurity. 11th ISOPT Clinical – The
International Symposium on Ocular Pharmacology and Therapeutics,
Iceland, Reyklavic.
Tamás A, Atlasz T, Szabadfi K, Werling D, Kiss P, Dányádi B, Reglődi D, Gábriel
R (2014) Protective Effects of Pituitary Adenylate Cyclase Activating Polypeptide
(PACAP) in Different Models of Retinal Injuries. 11th ISOPT Clinical – The
International Symposium on Ocular Pharmacology and Therapeutics, Iceland,
Reyklavic.
Reglődi D, Werling D, Váczy A, Kiss P, Tamás A, Dányádi B, Fülöp BD,
Gábriel R, Kovács K, Tamás A (2015) Protective effects of the
Secretin/Glucagon/VIP/PACAP peptide family in adult retinal lesions in rodents.
12th International Symposium on VIP, PACAP and Related Peptides,
Törökország, Cappadocia.

91
Werling D, Kvárik T, Varga R, Nagy N, Mayer F, Váczy A, Reglődi D, Kiss
P, Tamás A, Biró Zs, Tóth G, Atlasz T (2015) Investigating the
retinoprotective effects of PACAP eye-drop in ischemic retinopathy. 12th
International Symposium on VIP, PACAP and Related Peptides,
Törökország, Cappadocia.
Banks WA, Salameh TS, Atlasz T, Werling D, Váczy A, Reglődi D (2015)
Uptake and distribution to the vitreous humor and retina of PACAP27 after
its intraocular administration. 12th International Symposium on VIP,
PACAP and Related Peptides, Törökország, Cappadocia.
Lökős E, Atlasz T, Maász G, Schmidt J, Márk L, Kvárik T, Werling D,
Fülöp B, Tamás A, Kocsis B, Hashimoto H, Reglődi D (2015) Proteome
Analisys of tears from wild type and PACAP-deficient mice. 12th
International Symposium on VIP, PACAP and Related Peptides,
Törökország, Cappadocia.
Atlasz T, Váczy A, Kovács K, Lökős E, Kvárik T, Werling D, Kocsis B,
Fülöp B, Tamás A, Hashimoto H, Reglődi D (2015) The protective role of
the endogenous PACAP in LPS-induced inflammation in the retina. 12th
International Symposium on VIP, PACAP and Related Peptides,
Törökország, Cappadocia.
Kvárik T, Mammel B, Werling D, Váczy A, Tóth G, Tamás A, Reglődi D,
Ertl T, Gyarmati J, Atlasz T (2015) Does PACAP-treatement have any
influence on the outcome of retinopathy of prematurity? 12th International
Symposium on VIP, PACAP and Related Peptides, Törökország,
Cappadocia.
Werling D, Kvárik T, Varga R, Nagy N, Mayer F, Váczy A, Reglődi D, Kiss
P, Tamas A, Biró Zs, Tóth G, Atlasz T (2015) Szemcseppként alkalmazott
PACAP retinoprotektív hatásának vizsgálata ischaemiás retinopathiában.
Magyar Szemorvostársaság Retina Szekció Kongresszusa, Kecskemét.
Werling D, Kvárik T, Varga R, Nagy N, Mayer F, Váczy A, Reglődi D, Kiss
P, Tamas A, Biró Zs, Tóth G, Atlasz T (2016) Szemcseppként alkalmazott
PACAP retinoprotektív hatásának vizsgálata ischaemiás retinopathiában.
Magyar Farmakológiai, Anatómus, Mikrocirkulációs és Élettani Társaságok
közös Tudományos Konferenciája, Pécs.

92
Atlasz T, Werling D, Banks WA, Salameh TS, Kovács K, Kvárik T, Váczy
A, Kovács L, Dányádi B, Lökős E, Tamás A, Tóth G, Biró Zs, Mayer F,
Reglődi D (2016) A novel rout of neuroprotective action of PACAP1-27 in
ischemic retinopathy. International Regulatory Peptide Society (RegPep)
2016, Franciaország, Rouen.
Szabó E, Kvárik T, Mammel B, Werling D, Váczy A, Tóth G, Reglődi D,
Ertl T, Gyarmati J, Atlasz T (2016) Does endogen PACAP have any role in
the developement of retinopathy of prematurity? International Regulatory
Peptide Society (RegPep) 2016, Franciaország, Rouen.
Váczy A, Somoskőy T, Reglődi D, Kovács K, Szabó E, Lökős E, Werling D,
Tamás A, Atlasz T (2016) The protective role of PAC1-receptor agonist maxadilan
in BCCAO-induced retinal degeneration. International Regulatory Peptide Society
(RegPep) 2016, Franciaország, Rouen.

93
Köszönetnyilvánítás
Köszönettel tartozom elsősorban témavezetőimnek Dr. Reglődi Dóra egyetemi tanárnak, aki
kezdetektől irányította és mindenben támogatta kutatási munkámat, és akinek kitartása és
szakmai tudása mindig példaként áll majd előttem.
Köszönet illeti Dr. Biró Zsolt egyetemi tanárt, akinek a szakmai támogatása nélkülözhetelen
volt.
Köszönetemet fejezem ki Dr. Atlasz Tamás egyetemi adjunktusnak, aki a vizsgálati módszerek
és kísérleti protokollok elsajátításánál mindenkor önzetlenül segített, valamint az adatok
feldolgozásában, értékelésében, illetve a tudományos publikáció folyamatába bevezetett; időt
és energiát nem kímélve támogatta tudományos munkámat.
Hálásan köszönöm a sok segítséget és támogatást Dr. Kvárik Tímea rezidens orvosnak,
kutatótársamnak az időt, fáradságot nem kímélő közös munkát és mindenkori támogató
együttműködését.
Köszönet illeti William A. Banks-et, és Therese S. Salameh-et akikkel kollaborációban
dolgozva segítettük egymás munkáját.
Köszönöm a sok segítséget Dr. Kovács Krisztina egyetemi docensnek, aki a molekuláris
biológiai vizsgálatokban és módszertanuk elsajátításában nyújtott segítséget.
Köszönettel tartozom Váczy Alexandra egyetemi tanársegédnek, aki a várandóságom alatt
segített a magzatra káros kísérleti folyamatok elvégzésében, és támogatott a munkám során.
Hálásan köszönöm Dr. Kovács László egyetemi tanársegédnek, munkatársamnak a statisztikai
elemzésekben nyújtott fáradhatalan segítségét.
Köszönet illeti Dr. Dányádi Bese rezidens orvost, aki az ERG vizsgálatokban volt
segítségemre.

94
Köszönettel tartozom néhai Dr. Szabadfi Krisztina tudományos munkatársnak, aki az
értekezésben ismertetett szövettani vizsgálatok lépéseinek elsajátításában volt segítségemre.
Köszönet illeti Nagy Noémi és Varga Rita orvostanhallgatókat, akik a várandóságom alatt
sokat segítettek a kísérletek magzatra ártalmas szakaszainak kivitelezésében. Mayer Flóra
TDK hallgatómat külön köszönet illeti, a kísérletekben nyújtott segítsége, és eredményeink
konferenciákon való prezentálása miatt.
Köszönettel tartozom az Anatómiai Intézet minden dolgozójának, kiváltképpen Kiss
Anikónak, aki a szövettani metszetek készítésében rengeteget segített, és Dittrich
Erzsébetnek, aki a szövettani beágyazáshoz szükséges oldatok elkészítésével támogatta
munkámat.
Kitüntetett köszönettel fordulok Családom felé, akik mindenben és mindig támogattak; olykor
jó szóval, olykor határozottsággal, olykor azzal, hogy nélkülözték a társaságomat vagy átvették
az otthoni feladataimat, hogy én a kísérletek egész sorát folytathassam, és annak eredményét
feldolgozhassam. Hálás vagyok érte!
Támogatók:
AZ EMBERI ERŐFORRÁSOK MINISZTÉRIUMA ÚJ NEMZETI KIVÁLÓSÁG PROGRAMJÁNAK TÁMOGATÁSÁVAL KÉSZÜLT
OTKA K104984, K119759; Arimura Foundation, PTE AOK KA Research Grant, National
Brain Research Programme [KTIA_13_NAP-A-III/5]; PTE MTA “Lendület”; GINOP-2.3.2-
15- 2016-00050 “PEPSYS”.





























