
UNIVERSIDAD DE BUENOS AIRES
FACULTAD DE FARMACIA Y BIOQUÍMICA
Área de Bacteriología – Instituto de Medicina Regional
Universidad Nacional del Nordeste
Rol del agua en la transmisión de cepas
diarreogénicas de Escherichia coli en la
Provincia del Chaco.
Tesis para acceder al Grado de Doctor en Bioquímica
Tesista: Liliana Silvina Lösch
Director: Dr. Luis Antonio Merino
Codirectora: Mgter. Marta Rivas
Consejera de Estudio: Dra. Sonia Korol
2017

ii
Copyright © 2017 por Liliana Silvina Lösch
Todos los derechos reservados
Todas las fotografías son propiedad del autor y podrán ser
utilizadas sin autorización previa mención de la fuente.

iii
Dedicada a:
A mis padres, promotores de mis sueños, por
creer y acompañarme en cada emprendimiento.
A mi esposo, por elegirnos.

iv
Mi sincero agradecimiento.....
A mi Director Luis Merino por acompañarme en cada etapa
de este proyecto con su experiencia y amistad.
A mi Codirectora, Marta Rivas, por sus consejos, orientación
y compromiso.
A los Chaqueños por abrir sus puertas para conocer un
poquito más de nuestra provincia.
A todos y cada uno de mis compañeros del Instituto de
Medicina Regional y con un cariño especial a los integrantes del
Área de Bacteriología: Luis Merino, Lidia Acevedo, Fernanda
Tracogna, Lucrecia Gariboglio, Verónica Gómez y Marcelo
Medina.
A la Administración Provincial del Agua del Chaco por
brindarme su apoyo para concretar este proyecto.
A mi familia y amigos por la comprensión, paciencia y el
ánimo recibidos.
A mi Consejera de Estudios por el apoyo.
Y a todos aquellos quienes de una u otra forma
contribuyeron a concretar este proyecto.

1
ÍNDICE
Página
Índice de Fotografías, Tablas y Gráficos.......................................................................2
Abreviaturas.....................................................................................................................3
Resumen...........................................................................................................................4
Introducción.....................................................................................................................6
-La Región Chaqueña..............................................................................................6
-La Provincia del Chaco...........................................................................................8
-Contaminación de las fuentes de agua................................................................10
-Microbiología de aguas.........................................................................................11
-Escherichia coli diarreogénico.............................................................................13
Estado actual del conocimiento....................................................................................23
Problemática..................................................................................................................28
Objetivos.........................................................................................................................29
Hipótesis.........................................................................................................................30
Materiales y métodos.....................................................................................................31
Resultados.......................................................................................................................40
Discusión.........................................................................................................................51
Conclusiones...................................................................................................................68
Bibliografía.....................................................................................................................69
Anexo I............................................................................................................................78
Anexo II..........................................................................................................................79
Anexo III.........................................................................................................................80
Anexo IV.........................................................................................................................81
Anexo V..........................................................................................................................82

2
ÍNDICE DE FIGURAS, TABLAS Y GRÁFICOS
FIGURAS
Página
Figura 1. Esquema cepas diarreogénicas de E. coli....................................................................14
Figura 2. Reservorios y vías de trasmisión cepas diarreogénicas de E. coli...............................24
Figura 3. Puntos de muestreo. ....................................................................................................32
Figura 4. Secuencia metodológica empleada..............................................................................33
Figura 5. Productos de amplificación patotipos diarreogénicos de E. coli.................................36
Figura 6. Productos de amplificación subtipificación E. coli enteroagregativo........................37
Figura 7. Productos de amplificación subtipificación E. coli enteropatogénico.......................39
Figura 8: Localización lagunas positivas para los diferentes patotipos......................................44
Figura 9: Localización fuentes superficiales positivas para los diferentes patotipos..................45
Figura 10: Localización fuentes subterráneas positivas para los diferentes patotipos................48
Figura 11: Productos de amplificación cepas diarreogénicas de referencia y ambientales.........50
Figura 12: Productos de amplificación subtipificación E. coli enteroagregativo........................50
TABLAS
Tabla 1: Características de cebadores en la identificación de cepas diarreogénicas...................35
Tabla 2: Cepas de referencia empleadas como controles positivos.............................................35
Tabla 3: Cebadores para subtipificación de E. coli enteroagregativo.........................................37
Tabla 4: Cebadores para subtipificación de E. coli enteropatogénico.........................................38
Tabla 5: Origen de muestras estudiadas distribuidas según la fuente de agua............................40
Tabla 6: Prevalencia cepas diarreogénicas en fuentes superficiales .........................................42
Tabla 7: Origen y frecuencia de muestras superficiales positivas ..............................................44
Tabla 8: Prevalencia cepas diarreogénicas en fuentes subterráneas............................................46
Tabla 9: Comparación resultados prevalencia del patotipo enteropatogénico ...........................53
Tabla 10: Comparación resultados prevalencia del patotipo enterotoxigénico ..........................54
Tabla 11: Comparación resultados prevalencia del patotipo enteroagregativo...........................56
GRÁFICOS
Gráfico 1: Distribución de patotipos en fuentes de agua superficiales.......................................43

3
ABREVIATURAS
ETEC: Enterotoxigenic Escherichia coli- Escherichia coli enterotoxigénico
EPEC: Enteropathogenic Escherichia coli- Escherichia coli enteropatogénico
STEC: Shiga toxin Producing Escherichia coli- Escherichia coli productor de toxina Shiga.
EIEC: Enteroinvasive Escherichia coli- Escherichia coli enteroinvasivo
EAEC: Enteroaggregative Escherichia coli- Escherichia coli enteroagregativo
EDTA: ácido etilendiaminotetracético
PCR : Polymerase Chain Reaction- reacción en cadena de la polimerasa.
pb: pares de bases
rpm: revoluciones por minuto
TAE: TRIS-Acetato- EDTA
TE: buffer Tris-EDTA
TRIS: 2-Amino-2-hydroxymethyl-propane-1,3-diol
UV: luz ultravioleta

4
1- RESUMEN
Las cepas diarreogénicas de Escherichia coli se consideran patógenos
emergentes y los diferentes patotipos participaron de brotes de origen hídrico en
distintas partes del mundo. Estos patotipos son los siguientes: E. coli enterotoxigénico
(ETEC), E. coli enteropatogénico (EPEC), E. coli enteroinvasivo (EIEC), E. coli
enteroagregativo (EAEC) y E. coli productor de toxina Shiga (STEC).
El objetivo del trabajo fue detectar la presencia de E. coli diarreogénico en
ambientes acuáticos de la Provincia del Chaco, mediante el reconocimiento de sus
factores de virulencia, a través de la técnica de reacción en cadena de la polimerasa
(PCR).
Se analizaron 362 muestras de agua obtenidas de fuentes superficiales,
subterráneas, aljibes, de red y bajadas de tanque. E. coli se recuperó en 184 muestras
(50,8%).
La caracterización de los aislamientos se realizó mediante la detección de genes
de virulencia característicos de cada patotipo: ETEC (elt y est), EPEC (eae/bfp), EIEC
(ipaH), EAEC (aggR) y STEC (stx).
Sobre 150 muestras de fuentes superficiales estudiadas, en 111 (74%) se
recuperaron cepas de E. coli. De ellas, 32 (29%) resultaron positivas para alguno de los
genes estudiados. La prevalencia de cepas diarreogénicas sobre el total de muestras
estudiadas fue del 21,3%. Los aislamientos clasificados como EAEC resultaron los más
prevalentes (15,3%). La prevalencia de los caracterizados como ETEC fue del 4%, y de
EPEC atípico del 2%.
Se analizaron 187 muestras de agua de fuentes subterráneas y de 61 (32,6%) se
recuperaron cepas de E. coli. En 19 (31%) de ellas se identificaron aislamientos
portadores de genes de virulencia. La prevalencia de cepas diarreogénicas sobre el total
de muestras estudiadas fue del 10,1%. Los aislamientos caracterizados como EAEC
resultaron los más prevalentes (8,6%). También se recuperaron cepas caracterizadas
como ETEC, EIEC y EPEC típico. La prevalencia de cada uno fue del 0,5%.
No se encontró diferencia estadísticamente significativa en la presencia de cepas
diarreogénicas de E. coli entre las fuentes superficiales y subterráneas.
En 5/8 (62,5%) muestras de agua de aljibes, se detectó la presencia de E. coli. En
3 (60%) de estas muestras se detectó al patotipo EAEC.

5
En el análisis de muestras de agua de red, 3/9 (33,3%) resultaron positivas para
E. coli sin detectarse genes de virulencia. De las 8 muestras de bajadas de tanque, 4
resultaron positivas para E. coli y 2 se clasificaron como EAEC.
La detección de los patotipos EPEC, ETEC y EAEC en fuentes de agua
superficiales evidencia su vulnerabilidad a la contaminación, principalmente de origen
antrópico, lo cual estaría relacionado con la falta de tratamiento de los efluentes
cloacales, una debilidad en toda la Cuenca del Plata.
Igual consideración puede hacerse con respecto a la detección de E. coli en
fuentes subterráneas que pueden actuar como reservorio y vehículo de estas cepas, con
el agravante de su no aptitud para el consumo humano. Cuando las cepas recuperadas
poseen genes de virulencia, como en este caso, se enfatiza aún más el riesgo para salud
de las personas que las consumen.
El presenta trabajo constituye el primer aporte, en la provincia del Chaco, sobre
la participación de las fuentes de agua en la cadena epidemiológica de estos patógenos.
La detección de cuatro de los cinco patotipos estudiados, con EAEC como el prevalente
y de carácter emergente en todo el mundo, evidencian el riesgo para la salud de las
personas expuestas a las mismas ante los diferentes usos que se les puede dar: consumo,
recreación, riego o fuente para potabilización.

6
2- INTRODUCCIÓN
La voz Chaco es de origen quechua y designaba originalmente a una forma de
caza que se practicaba en el occidente del actual Chaco boreal y central.
2.1 La región chaqueña
La vasta región del Chaco es compartida por Argentina, Paraguay y Bolivia. El
río Pilcomayo la divide en dos regiones: el Chaco boreal al Norte y el Chaco austral al
Sur.
La región del gran Chaco argentino ocupa una superficie de unos 400.000 Km2.
Constituye un área de gran diversidad biológica, ambiental y cultural. Se encuentra
limitada al Norte por el río Pilcomayo, al Este por los ríos Paraná y Paraguay, al Oeste
por las estribaciones de las sierras Subandinas y al Sur por la región comprendida entre
los ríos Salado y Dulce. La región se caracteriza por ser una vasta cuenca sedimentaria,
de relieve horizontal y con una suave pendiente Noroeste – Sureste que permite el lento
desplazamiento de los cursos de agua y de las inundaciones por exceso de lluvias, hacia
los ríos Paraná – Paraguay o hacia el río Salado. La llanura del gran Chaco se levanta
paulatinamente en plano inclinado desde los 50 m sobre el nivel del mar en el Este a
partir del surco Paraguay – Paraná hasta los 350 m en el occidente, en una distancia de
600 Km. Sobre estos soportes se agrega una masa boscosa constituida por bosque
xerófilo, sabanas y bosque hidrófilo.
El clima chaqueño es netamente subtropical (con estación seca). La diferencia
climática fundamental, entre el oriente y occidente de esta región, resulta de la desigual
frecuencia con que actúan las masas de aire marítimas y continentales. La ausencia de
relieves de jerarquía hace que las masas de aire cálido y húmedo del anticiclón
semipermanente del Atlántico se degrade paulatinamente desde el E hacia el O. En la
banda oriental las masas atlánticas son más frecuentes, lo que se traduce en mayor

7
humedad relativa y una mayor capacidad pluvial con montos anuales que superan los
1200 mm y se degradan hacia el Oeste donde los registros no llegan a los 500 mm. El
Chaco occidental presenta un marcado dominio de masas secas, preferentemente en
invierno y actividad pluvial durante el estío. El oriental presenta un régimen
pluviométrico de doble máximo, uno en primavera (noviembre) y el otro en otoño
(marzo) (1–3).
Hidrografía. La franja oriental aloja el sistema autóctono de ríos, alimentados
por las lluvias locales, cuyos colectores son los ríos Paraná y Paraguay. Los dos
períodos de lluvias máximas de la región y la escasa pendiente de la cuenca oriental
explican los meandros y la formación de depósitos aluvionales de margen (albardones).
Los albardones impiden el drenaje y contribuyen a la formación de cañadas, esteros y
lagunas que interconectan los conjuntos de cuencas.
El sistema alóctono, alimentado por lluvias orográficas, está constituido por
cuatro ríos que atraviesan la región: Pilcomayo, Bermejo, Salado y Dulce. Las lluvias
concentradas en la estación estival, con picos preferentemente en febrero y un período
de bajantes que se agudiza en invierno, la marcada pendiente de los cursos superiores y
la variación de caudal, explican la actividad erosiva de los cursos superiores de modo
que las aguas incorporan y llevan en suspensión un extraordinario volumen de material
sólido, especialmente en época de crecientes cuando la erosión y el trasporte son
dominantes. Al bajar a la planicie pierden los afluentes, decrecen los caudales por efecto
de la infiltración y la intensa evaporación y se deposita parte de su carga de sedimentos
produciendo el relleno de sus cauces y el futuro cambio del mismo (3).

8
2.2 Provincia del Chaco
La provincia del Chaco se encuentra ubicada entre los 28° y 25°39’21’’de latitud
S y 61º42´40´´ y 62°20´30´´de longitud O, en el nordeste de la Argentina.
Considerando las características climáticas y ecológicas, en líneas generales, el
Chaco puede dividirse en tres grandes ambientes o regiones naturales: el Chaco Oriental
o Húmedo, el Chaco Central o de Transición y el Chaco Occidental o Seco. Esto
clasificación está vinculada con la disminución hacia el Oeste de las precipitaciones y
por ende de la humedad ambiente, factor que incide en la distribución, acumulación y
escurrimientos de las aguas, asociado a la topografía de escasa pendiente.
El Chaco Occidental o Chaco Seco abarca el Noroeste de la provincia, es una
región semiárida con una estación seca marcada y una creciente disminución de las
lluvias hacia el Oeste. Se caracteriza por tener un Bosque Xerófilo Subtropical, con
dominancia de formas arbóreas combinadas con arbustos.
El Chaco Central es el área de transición entre el oriente húmedo y el occidente
seco. En esta región se alternan bosques con áreas no inundables cubiertas de pastizales,
a su vez es la más modificada por el hombre a través de sus obras y actividades
productivas; corresponde a la región agrícola por excelencia de la provincia. La
vegetación natural está compuesta por las mismas especies del oriente pero con un
predominio de árboles de maderas duras, ricas en tanino, y adaptados a una estación
seca más notoria y de mayor duración como quebrachos colorados y blancos,
algarrobos, guayacanes.
El Chaco Oriental está marcado por una pluviosidad con registros que oscilan
entre 1.000 mm y 1.200 mm al año, lo que determina una riqueza de ambientes
acuáticos que se alternan con otros más secos. Estos aspectos, sumados a los
microrelieves y la dinámica del agua, permiten diferenciar los siguientes paisajes

9
naturales: selva en galería, monte alto, bosques bajos abiertos, sabanas con palmeras y
esteros, cañadas y lagunas. A partir del 2 de febrero de 2004 un área de 508.000 ha del
Chaco Oriental, localizada en la franja oriental de los Departamentos San Fernando, 1º
de Mayo y Bermejo sobre el eje fluvial Paraguay-Paraná, se ha designado e inscripto
como sitio Humedales del Chaco en el listado de la Convención sobre los Humedales o
Convención de Ramsar.
Los principales ríos de la provincia son: al Norte (oficiando de límite con la
provincia de Formosa) los ríos Teuco y Bermejo y al Este los ríos Paraguay y Paraná,
todos ellos forman parte de la extensa Cuenca del Plata. A su vez la provincia posee en
su lado oriental ríos interiores que vierten sus aguas a los ríos Paraguay (Guaycurú, Oro,
Quiá) y Paraná (Negro, Tragadero, Palometa, Tapenagá) (1,3,4).
Indicadores básicos. La provincia del Chaco tiene una población estimada de
1.130.608 habitantes, de los cuales el 29,7% son menores de 15 años y el 6,7% de 65
años o más. El 23,3% de la población con necesidades básicas insatisfechas y el 5,5%
de 10 o más años en condición de analfabetismo. La tasa bruta de mortalidad para el año
2014 fue de 6,6 (por 1000 habitantes). El porcentaje de población urbana es del 84,6%.
En la Provincia del Chaco, 169.153 hogares (58,6%) tienen acceso al sistema de
distribución del agua potable dentro de la vivienda, mientras que en 51.595 (17,9%), el
acceso a la red pública se realiza fuera de la misma. El resto de los 67.675 hogares
(23,5%) se abastecen de agua a partir de fuentes subterráneas, superficiales, de lluvia o
bien por trasporte por cisternas. A su vez, sólo 76.107 (26,4%) de los hogares tienen
cloaca conectada a la red pública. En el resto de los hogares la descarga de los efluentes
cloacales se realiza a través de cámara séptica, pozo ciego o a excavaciones directas en
la tierra (5,6).

10
2.3 Contaminación de las fuentes de agua
El crecimiento de la población y la industrialización y también el cambio
climático, ejercen una continua presión sobre la calidad y cantidad de los recursos
hídricos. La escasez de fuentes de agua para consumo libres de contaminantes, es uno
de los mayores problemas que enfrenta la población mundial. Se estima que para el año
2025, el 60% de la población mundial sufrirá problemas de escasez de agua. Entre los
diferentes contaminantes de agua (compuestos orgánicos, inorgánicos y microbios), la
Organización Mundial de la Salud (OMS) considera a los microbios como la principal
amenaza, tanto en países desarrollados como en vías de desarrollo (7,8).
El agua es esencial para la vida y una fuente adecuada, segura y accesible debe
estar disponible para todos. Sin embargo millones de personas en todo el mundo no
tienen acceso a la misma. De acuerdo a la OMS la mortalidad por enfermedades
asociadas al agua excede los 5 millones de personas al año, de ellas más del 50% se
atribuyen a patógenos intestinales. Este organismo estimó que en el año 2008 2.5
millones de personas murieron a causa de diarrea y que el número de casos de cólera se
incrementó en un 85% en el 2011 con respecto al 2010. Una de las principales causas de
estas enfermedades es la ingestión de agua contaminada con material fecal humana o de
animales (9–11).
La descarga de efluentes cloacales, sin tratamiento previo, y de los efluentes de
la actividad ganadera a cursos de agua dulce o salada es la principal fuente de
microorganismos de origen fecal incluidos los patógenos. Como en otras partes del
mundo, esta problemática también se presenta en los cursos de agua que integran la
Cuenca del Plata (7,10,12,13).
La calidad bacteriológica del agua procedente de fuentes subterráneas, al estar
naturalmente protegidas, es generalmente mejor que aquella procedente de fuentes

11
superficiales no protegidas; condición que también se favorece por el propio proceso de
remoción de microorganismos en la infiltración del agua hacia los estratos subterráneos
durante su recarga. No obstante existen reportes de brotes por patógenos fecales
originados a partir de estas fuentes. La contaminación microbiológica del agua
subterránea se atribuye principalmente a la infiltración de agua contaminada con
materia fecal humana o de animales (14–16).
2.4 Microbiología de aguas
Garantizar la seguridad del agua es un cambio en curso para proteger la salud
pública. Para minimizar el riesgo de las infecciones transmitidas por el agua de bebida
se requiere aplicar el principio de barreras múltiples, dado que ningún tratamiento
individual puede garantizar la seguridad microbiológica del agua. Este principio se basa
en la protección de las fuentes de agua, la adecuada selección y operación de las
distintas etapas del tratamiento y finalmente la gestión y control del sistema de
distribución para mantener y proteger el agua ya tratada (17,18).
Entre los objetivos de la microbiología de aguas se encuentran i) evaluar la
eficiencia de la combinación de procesos físicos para la reducción del número de
microorganismos del agua destinada al consumo, ii) de los procesos químicos para
lograr la inactivación de los mismos y iii) evaluar la calidad del agua destinada a fines
recreativos, de riego y de captación, entre otros (10,11). El monitoreo de los patógenos
en los cuerpos de agua es dificultosa, costosa y compleja, ante la diversidad de
microorganismos que se reconocen que pueden estar presentes en el ambiente para lo
cual surge la utilización de indicadores bacterianos de contaminación (19,20).
El indicador bacteriano de contaminación fecal debe cumplir una serie de
requisitos: encontrarse en elevado número en el intestino y en las heces, no ser patógeno

12
para el ser humano y su detección en el agua debe ser fácil, de bajo costo y fiable.
Adicionalmente se pretende que el indicador no se multiplique fuera del intestino, que
su concentración en las fuentes de agua sea mayor que la de los gérmenes patógenos
pero que su sobrevida sea similar a la de éstos, que permita discriminar entre
contaminación fecal humana y la de origen animal y finalmente que su respuesta al
sistema de tratamiento de agua empleado sea similar a la de los patógenos (10,21).
Entre los indicadores de contaminación fecal de agua más utilizados se
encuentran los coliformes termotolerantes, enterococos y Escherichia coli. Las
técnicas para su detección deben ser relativamente sencillas y al alcance de la mayoría
de los laboratorios.
Escherichia coli es el indicador de contaminación fecal más fiable para evaluar
el riesgo para la salud de las personas que consuman o estén en contacto con una
determinada fuente de agua. Si bien estudios recientes demuestran que esta bacteria
pude sobrevivir y replicarse fuera del huésped, condición favorecida por la
concentración de nutrientes y las temperaturas del clima tropical y subtropical, continúa
siendo el microorganismo que mejor satisface los criterios de indicador (22,23). La
Comisión Europea, la Agencia de Protección Ambiental de Estados Unidos y la OMS
recomiendan el uso de E. coli como el mejor indicador de contaminación fecal para
predecir la presencia de patógenos en el agua de bebida y de recreación, criterio que fue
adoptado por numerosos países (9,24).
En Argentina el Código Alimentario Argentino (C.A.A.) en su capítulo XII
establece como organismos indicadores de las características microbiológicas de agua
potable de suministro público y agua potable de uso domiciliario, a los coliformes
totales, Escherichia coli y Pseudomonas aeruginosa. Por otro lado, nuestro país carece
de legislación propia referida a límites de calidad para aguas de uso recreativo, por lo

13
que se han tomado como valores guías niveles de indicadores bacterianos, Enterococos
y a Escherichia coli, de acuerdo con estándares internacionales los que se encuentran en
revisión (25,26).
2.5 Escherichia coli diarreogénico
Escherichia coli es un bacilo gramnegativo que pertenece a la familia
Enterobacteriaceae, coloniza el tracto intestinal del recién nacido a las pocas horas de
vida, y a partir de ese momento, huésped y microorganismo obtienen mutuo beneficio.
Sin embargo, existen clones de este microorganismo que han adquirido diferentes
factores de virulencia que les permiten adaptarse a nuevos nichos y causar un amplio
espectro de enfermedades. Se encuentran codificados en elementos genéticos que
pueden movilizarse entre diferentes cepas y crear nuevos factores de virulencia o bien
han quedado atrapados en el cromosoma bacteriano. Solo las combinaciones exitosas de
los factores de virulencia transforman a una cepa comensal de E. coli en un patotipo
específico capaz de causar enfermedades. Los cuadros clínicos más frecuentes
relacionados a estas cepas patogénicas son: enteritis, infecciones del tracto urinario y
sepsis/meningitis.
Su rol como patógeno entérico aumentó con la emergencia de E. coli O157:H7.
Las cepas causantes de enteritis se han clasificado en cinco grupos principales, a los
cuales se les agrega E. coli de adherencia difusa (DAEC). Cada uno de ellos engloba
diferentes serotipos, factores de virulencia y mecanismos de patogenia. Así, según los
genes de virulencia presentes en cada cepa, se pueden clasificar en: E. coli
enteropatógeno (EPEC), E. coli enteroinvasivo (EIEC), E. coli enterotoxigénico
(ETEC), E. coli productor de toxina Shiga (STEC) y E. coli enteroagregativo (EAEC)
(27–29).

14
Figura 1: Esquema cepas diarreogénicas de Escherichia coli. Fuente: Kaper y col (2004). (28)
Escherichia coli enteropatógeno (EPEC). Fue el primer patotipo descripto por
Bray en 1945. Produce una lesión característica en las células intestinales de adherencia
y borrado de las microvellosidades conocida como A/E (attaching/effacing) y la
formación de una estructura como pedestal rica en actina sobre la cual se adhieren las
células de este patotipo. Esta lesión es el resultado de la acción cooperativa de proteínas
que se encuentran codificadas en una isla de patogenicidad denominada locus LEE
(locus of enterocyte effacement). Este locus codifica a través del gen eae para una
proteína de membrana externa, denominada intimina, responsable de la adherencia
íntima de la bacteria a las células epiteliales y de la reorganización del citoesqueleto.
Esta región también se encuentra en el patotipo Escherichia coli enterohemorrágico
(EHEC). Por consenso, las cepas EPEC se identifican por la presencia de la región LEE
y la ausencia de los genes stx que codifican para la toxina Shiga.

15
A su vez este patotipo se divide en EPEC típico (tEPEC) y EPEC atípico
(aEPEC), clasificación que se basa en la presencia del plásmido pEAF, asociado a la
adherencia, presente en el primer grupo y ausente en las cepas atípicas de EPEC. Las
cepas típicas son más homogéneas en sus características de virulencia, en tanto que las
atípicas pueden portar genes de virulencia de otros patotipos (27,30).
Actualmente se estima que este patotipo es responsable del 5-10% de las
diarreas pediátricas en países como Brasil, Chile y Perú (31). Las cepas típicas de EPEC
son la principal causa de diarrea infantil en países en desarrollo y su único reservorio es
el ser humano. En tanto que las cepas aEPEC son importantes agentes causales de
diarrea en los países desarrollados y los seres humanos y los animales sus reservorios.
No obstante estudios epidemiológicos recientes sugieren un incremento en la
identificación de cepas atípicas en cuadros de diarrea infantil y de diarrea persistente,
tanto en países desarrollados como en vías de desarrollo (28,32–34).
La identificación molecular de este patotipo se basa en la detección del gen eae
en ausencia del stx y la presencia del gen bfpA o del plásmido EAF para la
subtipificación en cepas típicas o atípicas (35).
Escherichia coli enteroinvasivo (EIEC) es agente causal de cuadros de diarrea
acuosa y disentería en el ser humano, el cual también representa su principal reservorio.
Fue identificado como patógeno 50 años después de la primera descripción de Shigella
en 1897, con quien comparte algunas propiedades bioquímicas, genéticas y patogénicas.
Diversos estudios filogenéticos demuestran que Shigella y EIEC surgieron a partir de
cepas comensales de E. coli que adquirieron horizontalmente genes de virulencia, entre
ellos el plásmido Ipa. Ambos patógenos invaden la mucosa del intestino grueso y
provocan su destrucción acompañada de reacción inflamatoria (27,28).

16
A pesar de las semejanzas en los mecanismos de invasión, la dosis infectiva de
EIEC es superior (106-10
10 microorganismos) a la de Shigella (10
1-10
4
microorganismos), aunque en el primer caso el cuadro de diarrea puede ser
autolimitado. Ambos se pueden trasmitir por contacto entre personas o a través de
alimentos y agua contaminada. Los últimos brotes por este patotipo fueron reportados
en Italia en 2012 y en Inglaterra en 2014, a causa del consumo de vegetales
contaminados. En países limítrofes, Brasil reporta una prevalencia que oscila entre el
0,5-15% según la población estudiada y Bolivia del 2%. EIEC fue el responsable del
0,9% de los casos de diarrea en pacientes pediátricos en la ciudad de La Plata,
Argentina (35,36).
La dificultad en identificar bioquímicamente estos patógenos hace que los
estudios epidemiológicos subestimen a EIEC, por este motivo se requiere la
combinación de pruebas bioquímicas y técnicas moleculares para investigar genes de
virulencia del plásmido preferentemente el gen ipaH (37,38).
Escherichia coli enterotoxigénico (ETEC). Este patotipo fue asociado por
primera vez a casos de diarrea en 1971. Produce un cuadro de diarrea acuosa y
generalmente autolimitada.
En los países en vías de desarrollo existe una elevada tasa de morbilidad y
mortalidad asociada a los cuadros de diarreas por ETEC en niños menores de 5 años. De
acuerdo a la OMS cada año tienen lugar alrededor de 21 millones de casos de diarrea
por este patotipo en los países en desarrollo. En estos países las cepas de este patotipo
pueden ser recuperadas de portadores sintomáticos o asintomáticos. Es la principal
causa de la diarrea del viajero y agente causal de diarrea en animales.
ETEC coloniza la mucosa del intestino delgado a través de adhesinas proteicas
de superficie conocidas como factores de colonización y que fueron denominados:

17
como factor antigénico de colonización (CFA), antígeno de superficie de coli (CS) y
probable factor de colonización (PCF). ETEC puede producir una o dos toxinas. La
enterotoxina LT (termolábil) codificada por el gen lt y la toxina ST (termoestable)
codificada por el gen st. Ambas enterotoxinas producen diarrea. Sin embargo en
estudios de cohorte realizados en población pediátrica en diferentes países, entre ellos
Argentina, se demostró que en las cepas productoras de toxina ST era más frecuente el
cuadro de diarrea como así también de un incremento en el riesgo de muerte de los
niños. La definición de este patotipo se basa en la detección de las toxinas (27,39).
La estructura de la enterotoxina LT está estrechamente relacionada con la toxina
de Vibrio cholerae. Se la puede clasificar en LT-I expresada por cepas patógenas para
humanos y animales y LT-II recuperada principalmente de cepas patógenas de animales.
La toxina ST también se clasifica en dos: STa asociada a enfermedad en el ser humano
y STb asociada a diarrea en animales.
La principal vía de diseminación de este patotipo son los alimentos y el agua; la
transmisión entre personas no es frecuente. En la literatura se describen diversos brotes
hídricos causados por ECET (28,40–42).
Escherichia coli productor de toxina Shiga (STEC): fue reconocido por
primera vez como causal de enfermedad humana en 1982 y actualmente es considerado
uno de los patógenos emergentes más importantes, trasmitido por alimentos y de
distribución mundial. E. coli O157:H7 es el prototipo de esta categoría de cepas
diarreogénicas, implicado como agente etiológico de la mayoría de los grandes brotes.
No obstante las cepas de E. coli no O157 también pueden causar enfermedad humana.
El principal factor de virulencia de este patotipo es la toxina Shiga la que está
codificada en bacteriófagos. Estas citotoxinas se clasifican en dos tipos Stx1 y Stx2,

18
encontrándose con esta última mayor asociación con enfermedad severa. La familia de
las Stx1 es más homogénea e incluye a Stx1a, Stx1c y Stx1d. En tanto que el grupo de
la Stx2 es más heterogéneo y se han descrito numerosas variantes: Stx2a, Stx2b, Stx2c,
Stx2d, Stx2e, Stx2f y Stx2g. Diversos estudios epidemiológicos demostraron que más
de 200 serotipos de E. coli pueden producir la toxina Shiga, pero sólo algunos
serogrupos son responsables de enfermedad humana entre ellos O26, O45, O103, O111,
O121 y O145 (27,28).
La proteína de membrana externa intimina es otro factor de virulencia,
codificada por el gen eae que se encuentra en la isla de patogenicidad LEE del
cromosoma, también compartido con el patotipo EPEC. Esta proteína permite la
adherencia de la bacteria al enterocito y la formación de la lesión A/E
(attaching/effacing). La presencia de LEE le confiere a las cepas STEC una mayor
virulencia, sin embargo no es esencial para la patogénesis. Esto ha llevado a utilizar el
término STEC para designar a las cepas productoras de toxina Shiga y el de E. coli
enterohemorrágico (EHEC) a aquellas cepas que además de producir la toxina contienen
al LEE asociados a los cuadros de colitis hemorrágica. Finalmente algunas cepas de
STEC producen una enterohemolisina, codificada en el gen exhA del plásmido pO157
involucrada en la patogénesis (27,28,35,43).
El ganado vacuno es el principal reservorio de este patotipo, pero también fue
identificado en la materia fecal de otros animales como ovejas, cerdos, aves y peces.
Igualmente requiere atención su presencia en el ambiente dado que estos
microorganismos pueden sobrevivir en el estiércol, suelo y agua. En Argentina,
estudios realizados en terneros reportan una frecuencia de detección del 38,5% de no-
O157 y del 0,5% de O157:H7 en tanto que en Entre Ríos Tanaro y col. (2010) reportan
una frecuencia del 3,8% de E. coli O157:H7 (43,44).

19
La principal vía de transmisión de STEC O157 y no-O157 es la ruta fecal –oral,
por los alimentos contaminados y el agua contaminada, pero también el contacto directo
del hombre con los animales y la contaminación cruzada durante la preparación de
alimentos. La dosis infectiva capaz de ocasionar enfermedad por parte de este grupo
bacteriano es de 10 a 100 bacterias por gramo de alimento (27,43).
Este patotipo produce desde una diarrea leve, colitis hemorrágica y síndrome
urémico hemolítico (SUH). Este último se caracteriza por ser una entidad clínica de
presentación aguda, con daño renal, anemia hemolítica microangiopática y
trombocitopenia. La forma típica de SUH es de origen infeccioso y está precedida por
un período prodrómico con diarrea, generalmente sanguinolenta, fiebre, vómitos y dolor
abdominal. En Argentina el SUH es endémico y exhibe una de las incidencias más altas
del mundo en niños menores a 5 años. STEC productor de stx2 es el agente etiológico
prevalente y el serotipo O157:H7 el predominante. En el período 2010-2015 la tasa de
notificación de SUH fue de 8,5 casos cada 100.000 menores/año (43,45,46). En
contraposición Brasil reporta una baja incidencia de SUH como así también de
aislamientos de E. coli O157:H7. En Uruguay STEC representa el tercer patógeno en
orden de frecuencia después de Shigella y Campylobacter en las diarreas sanguinolentas
en niños menores de 5 años (35,47).
Escherichia coli enteroagregativo (EAEC): fue descripto por primera vez en el
año 1987 por Nataro y col. (29) Desde ese momento este patotipo fue identificado como
agente causal de diarreas endémicas, epidémicas, agudas y persistentes en el mundo
entero. Sin embargo su mayor impacto es como patógeno implicado en cuadros de
diarrea persistente tanto en pacientes portadores del VIH como en niños menores de
cinco años. Estudios epidemiológicos en Brasil relacionan casos de diarrea persistente

20
por EAEC en población pediátrica con malnutrición y retardo en el desarrollo físico e
intelectual (27,35,48).
Su mecanismo de patogénesis no está completamente dilucidado. Se postula que
coloniza la mucosa intestinal del duodeno, íleon o colon. Allí la bacteria exacerba la
producción de mucus que la atrapa en un biofilm, produce daños a la mucosa y secreta
enterotoxinas y citotoxinas. La formación del biofilm se ha relacionado con los cuadros
de diarrea persistente (49–51).
EAEC produce adhesinas fimbriales y afimbriales o proteínas de membrana
externa, enterotoxinas, citotoxinas y proteínas responsables de su patogénesis pero los
genes que los codifican no fueron detectados en todas las cepas de este patotipo. Una de
las principales características de EAEC es el carácter heterogéneo de los aislamientos
cuando se analiza su serotipo, marcadores genéticos de virulencia y relación
filogenética. A través de estudios por MLST (Multilocus sequence typing) se concluyó
que no existe una cepa que se pueda considerar representativa de este patotipo (49,51–
53). La presencia de un plásmido (pAA), donde se encuentran codificados el mayor
número de genes de virulencia, permite diferenciar entre cepas típicas y atípicas de este
patotipo. Las cepas portadoras de este plásmido representan un importante subgrupo
considerado como cepas típicas de EAEC (51,54).
El rasgo que define a las cepas de EAEC es su capacidad de adherirse en
cultivos a células epiteliales HEp-2 con una apariencia de ladrillos apilados. Si bien el
cultivo celular constituye la técnica de referencia para la identificación de estos
microorganismos, resulta engorrosa para poder ser desarrollada por laboratorios de
mediana complejidad. La clasificación en serogrupos O puede resultar problemática por
la capacidad de autoaglutinación de algunas cepas. Las técnicas moleculares permiten la

21
detección rápida del grupo heterogéneo de genes de virulencia presentes en el
aislamiento, constituyendo además una herramienta epidemiológica (27,50,55).
Entre los genes más estudiados de este patotipo se encuentran el aggR (activador
transcripcional) y AA probe (codifica una proteína de membrana externa la cual forma
parte de un sistema de proteínas transportadoras), ambos sólo fueron detectados en este
patotipo. En el caso del gen aap, que codifica para la dispersina también se detectó en
cepas no patogénicas de E. coli, y el gen astA (enterotoxina) fue detectado en los
patotipos ETEC, EHEC y EPEC. Todos estos genes están presentes en el plásmido de
las cepas típicas de EAEC, las que representan el grupo mayoritario de este patotipo
(54–56). Considerando el carácter emergente y heterogéneo de este patotipo, en la
actualidad se busca la estandarización de técnicas moleculares para la detección
simultánea de genes del cromosoma y del plásmido para la identificación de cepas
típicas y atípicas de EAEC. Mientras que el hombre es el principal reservorio de las
cepas típicas, las atípicas también fueron detectados en animales (27,57).
EAEC se trasmite por la ruta fecal-oral, a través de agua y alimentos
contaminados. Es el segundo agente causal de diarrea del viajero en Latinoamérica y
responsable de brotes de enfermedades trasmitidas por alimentos (35,58,59).
Fue reportada como agente causal de cuadros de diarrea tanto aguda como
persistente en diferentes países latinoamericanos como Chile, Brasil, Venezuela y Perú.
En nuestro país, Rüttler y col (2006) detectaron la presencia de EAEC en un brote de
diarrea aguda en niños menores a 2 años en la ciudad de Mendoza (53,60). Datos de la
región del noreste de Argentina (NEA) indican a EAEC como el patotipo más frecuente,
junto a ETEC, recuperados de muestras de pacientes pediátricos y adultos con diarrea
aguda en la ciudad de Corrientes. El híbrido EAEC/STEC, O104:H4, fue la cepa
responsable del brote en Alemania que se inició en el mes de mayo del 2011 (61,62).

22
Escherichia coli de adherencia difusa (DAEC): implicado como agente
etiológico de varios casos de diarrea particularmente en niños mayores de 12 meses. Se
define por su característico patrón de adherencia difusa en cultivos de células HEp-2.
Los métodos de detección de este patotipo se encuentran bajo desarrollo, con lo cual su
epidemiología es poco clara (27).

23
ESTADO ACTUAL DEL CONOCIMIENTO
Los recursos hídricos superficiales se están transformado en reservorios de
bacterias que exhiben resistencia a múltiples agentes antimicrobianos o son portadores
de genes de virulencia, como resultado del vuelco de efluentes municipales,
hospitalarios y de las actividades agrícolas y ganaderas. Los genes de resistencia
antibiótica y los de virulencia se consideran como contaminantes ambientales
emergentes (63–65).
De igual manera, la calidad del agua de las fuentes subterráneas se puede alterar
como consecuencia de su localización, construcción y proximidad a fuentes de
contaminación como los pozos sépticos (66). El consumo de agua procedente de estas
fuentes, donde se detectó E. coli, es un riesgo para la salud de las personas que se
abastecen de la misma. El agua contaminada de origen subterráneo estuvo implicada en
más de la mitad de los brotes de origen hídrico en los Estados Unidos durante 1971-
2006 (15).
Los brotes de enfermedades relacionados al agua de bebida o de contacto
primario son una importante preocupación para la salud pública, a pesar de los avances
en materia de abastecimiento y saneamiento. La vigilancia integral realizada en los
países nórdicos entre los años 1998-2012 permitió identificar 175 brotes de origen
hídrico de los cuales 8 se debieron a E. coli patogénicas (67). Sin embargo, en esta
región del país a la fecha no se logró implementar una vigilancia coordinada de los
brotes de enfermedades trasmitidas por el agua. En la Figura 2 se esquematiza la
participación del agua y alimentos como vehículos y reservorios de los patotipos
diarreogénicos de E. coli.

24
Figura 2: potenciales reservorios y vías de trasmisión de patotipos diarreogénicos de E. coli.
Fuente: Croxen y col (2013) (27).
La Agencia de Protección Ambiental de Estados Unidos (EPA) identificó
alrededor de 500 patógenos potenciales relacionados con el agua de bebida, entre los
que se incluyen bacterias, virus, parásitos y protozoos. Entre los contaminantes
microbianos se encuentra E. coli O157. Las cepas diarreogénicas de E. coli se
consideran patógenos emergentes y los diferentes patotipos participaron de brotes de
origen hídrico en varias partes del mundo (24,68).
La prevalencia de las infecciones por EPEC difiere entre los estudios
epidemiológicos según el área geográfica y población estudiada. Datos recientes
evidencian que las infecciones por cepas aEPEC exceden a las causadas por cepas
típicas tanto en países en vías de desarrollo como desarrollados. La trasmisión de este
patotipo sigue la ruta fecal-oral a través de alimentos y agua contaminada. Yatsuyanagi

25
y col.(2003) reportan a E. coli enteropatógeno atípico como agente causal de un brote de
origen hídrico en Japón (31,69).
En los países en desarrollo ETEC es el agente etiológico del 13% de los cuadros
de diarrea en niños y causal de 325.000 defunciones en menores de 5 años. Además es
responsable del 33,6% de los casos de diarrea del viajero en América Latina (59,70). La
trasmisión de este patotipo sigue la ruta fecal-oral usualmente a través de alimentos y
agua contaminada. Se reportaron tres brotes por este patotipo relacionados al agua de
bebida en Estados Unidos entre los años 1996-2003. En Japón se informaron 131 brotes
relacionados al agua y la comida contaminada entre 1966 y 2009 y en Israel por el agua
contaminada. También se lo relacionó con brotes de diarrea en población pediátrica
ecuatoriana por el agua de bebida y fue el agente causal de un brote de gastroenteritis en
Bangladesh durante períodos de inundación (27,71).
La principal vía de diseminación de EAEC es el agua y los alimentos
contaminados. Este patotipo estuvo implicado en un brote asociado al agua en India, en
un brote a través de alimentos que afectó a 2697 niños en Japón. También Falcão y col.
2004) lo informan en muestras de hielo procedentes de diferentes fábricas (72–74). El
híbrido de EAEC y STEC fue responsable del brote en Alemania en el 2011, sin
embargo la presencia de este híbrido fue previamente reportado en Japón, Francia e
Irlanda (27,61).
Los alimentos y el agua contaminada son la principal vía de exposición a E. coli
productor de toxina Shiga (STEC). En la contaminación de las fuentes de agua están
implicados los escurrimientos procedentes de zonas de cría de ganado como lo
demuestran en nuestro país los trabajos de Tanaro y col. (2014) y Marucci y col. (2011)
(75,76). Estudios realizados por Avery y col. (2008) demostraron que este patotipo

26
puede sobrevivir hasta 2 meses en diferentes fuentes de agua, dependiendo de la
presencia de nutrientes, depredadores, radiación ultravioleta y temperatura (77).
La presencia de este patotipo en fuentes de agua constituye un riesgo para las
personas expuestas a las mismas, ya sea con fines recreativos o por ingesta cuando el
agua es consumida sin tratamiento o se produjo una falla en el mismo. Numerosos
trabajos evidencian el rol de STEC como patógeno de trasmisión hídrica. E. coli
O157:H7 fue el agente causal de un brote de 1000 casos de gastroenteritis en Nueva
York en el año 1999 por la contaminación del agua subterránea con efluentes
contaminados con estiércol. En el año 2000 este mismo agente y Campylobacter spp
contaminaron las fuentes municipales de agua de Ontario, Canadá, causando más de
2000 casos de gastroenteritis y en Finlandia se reportaron 1000 casos a través del agua
de bebida (27,78,79). En nuestro país Rivas y col. (2006) informaron dos brotes de
Síndrome Urémico Hemolítico en población pediátrica por contacto con agua de
recreación contaminada por este patógeno, como así también se lo identificó en aguas
del rio de La Plata próximo a una toma de agua. E. coli no O157 es responsable de
numerosos brotes y en algunos países es tan o más frecuente que E. coli O157H7 (43).
La aplicación de los métodos moleculares, principalmente aquellos basados en la
reacción en cadena de la polimerasa (PCR), surgen como herramientas muy específicas
y sensibles que permiten una rápida detección de los microorganismos de interés para la
salud pública, sean estos bacterias, virus o protozoos. Como elementos a favor de la
aplicación de estos métodos, en la determinación de la calidad microbiológica del agua,
cabe mencionar que para muchos patógenos y nuevos indicadores de calidad estas
técnicas son la única alternativa de detección y cuantificación. Su empleo no sólo
permite la identificación microbiana sino también su genotipificación y confirmación de
resultados. Las técnicas de PCR cuantitativas permiten la evaluación de la eficiencia en

27
la remoción de patógenos de los sistemas de tratamiento de efluentes. Finalmente
permiten la detección de patógenos por debajo de los límites de detección de otros
métodos, transformándose es una poderosa herramienta en la evaluación de riesgos.
Como factor en contra se pueden mencionar la presencia de inhibidores de los ensayos
moleculares, que dificultan su aplicación en el estudio de las muestras ambientales y
finalmente la revisión de las técnicas vigentes para lograr su estandarización y
aplicación a diferentes matrices (13,17,80) .

28
PROBLEMÁTICA
En la Provincia del Chaco el 76,5 % de los hogares tienen acceso al sistema de
distribución del agua potable. El 23,5% de los hogares restantes se abastecen de agua a
partir de fuentes subterráneas, superficiales, de lluvia o bien por trasporte por cisternas.
A su vez, sólo el 26,4%de los hogares tienen cloaca conectada a la red pública. La falta
de tratamiento de efluentes cloacales que se vierten a los recursos hídricos representa
una problemática común en los países integrantes de la Cuenca del Plata (6,12).
Las enfermedades diarreicas son una de las principales causas de mortalidad en
el mundo y las cepas patogénicas de E. coli uno de los principales agentes etiológicos.
A pesar que se reconoce el impacto de estos patotipos en la salud y la implicancia del
agua contaminada como vehículo de las mismas su distribución en las fuentes de agua
de la provincia del Chaco no está caracterizada.
El presente trabajo se planteó con el propósito de aportar datos acerca de la
posible participación de los ambientes acuáticos en la cadena epidemiológica de las
diarreas producidas por E. coli, ya que hasta este momento no se han realizado
actividades de vigilancia y control ambiental de estos patógenos en nuestra provincia.

29
OBJETIVOS
Objetivo general
Detectar la presencia de E. coli diarreigénicos, mediante el reconocimiento de
sus factores de virulencia a través de la técnica de PCR, en ambientes acuáticos de la
Provincia del Chaco.
Objetivos específicos
Poner a punto la técnica de recuperación e identificación de cepas diarreogénicas
de E. coli a partir de ambientes acuáticos.
Evaluar el rol de las aguas recreacionales y de bebida como reservorio
ambiental de los diferentes patotipos de E. coli.
Determinar la distribución de los patotipos diarreogénicos de E. coli en
ambientes acuáticos (superficiales y subterráneos) de la provincia del Chaco.

30
HIPÓTESIS DE TRABAJO
“Los ambientes acuáticos, superficiales y subterráneos, y el agua de bebida
de la provincia del Chaco pueden actuar como reservorio de alguno de los diferentes
patotipos de E. coli.”
.

31
3- MATERIALES Y MÉTODOS
3.1 Muestras
Se recogieron 362 muestras de agua procedentes de: fuentes subterráneas (n=
187), superficiales (n= 150), de aljibes (n=8) y de red y bajadas de tanque (n=17), de la
Provincia del Chaco. Los sitios en los cuales se realizó el muestreo se presentan en los
Anexos I, II, III y IV.
En el caso de las muestras de agua subterránea, se accedió a través de pozos o
perforaciones existentes en los hogares. Sólo se tomó muestras en aquellos hogares
donde el agua subterránea es la única fuente disponible para el consumo, por lo que su
contaminación representa un riesgo para la salud.
El muestreo de las perforaciones se realizó en el grifo de salida luego de realizar
la limpieza y bombear el agua durante dos minutos. En el caso de los pozos y de los
aljibes, la muestra se tomó a partir del recipiente con el cual la familia realizaba la
extracción del agua para su consumo.
Las muestras superficiales fueron tomadas de ríos y lagunas. En el caso de los
ríos, la recolección se realizó desde la costa enfrentando la corriente. En las lagunas, las
muestras también se tomaron de la costa, generando una corriente con el recipiente al
momento del muestreo. En la Figura 3 se muestran los distintos puntos de muestro.
Las muestras del agua de red se tomaron en grifos conectados a la misma dentro
de las viviendas y las de bajada de tanque en el grifo de la cocina. En ambos casos se
realizó una limpieza previa del grifo con un paño embebido en alcohol y luego se lo
flameó. Finalmente se dejó correr el agua durante dos minutos antes de realizar la toma
de la muestra.
Todas las muestras fueron recolectadas en botellas estériles de 150-250 cm3 de
capacidad dejando un espacio aéreo para facilitar la homogeneización antes del estudio,

32
y se mantuvieron refrigeradas hasta su llegada al laboratorio donde se realizó su
procesamiento (21,81).
Figura 3. Puntos de muestreo. A- pozo calzado. B- aljibe. C- Fuente superficial. Río Bermejo
3.2 Procesamiento de las muestras
3.2.1 Enriquecimiento
Las muestras se procesaron por la técnica de enriquecimiento directo de 100 ml
de agua en el mismo volumen de medio EC (Acumedia) doble concentración, lo cual
brindó información cualitativa sobre la presencia o ausencia del grupo de
microorganismos indicadores correspondientes a coliformes. Las botellas inoculadas
con las muestras previamente agitadas, se incubaron a temperaturas de 35±0,5 ºC y se
observaron a las 24 y 48 h. En la Figura 4 A se observan las botellas de enriquecimiento
(9,81).
3.2.2 Cultivo e identificación bioquímica
A partir de las botellas donde se evidenció desarrollo bacteriano, se realizaron
repiques en placas de Agar Eosina Azul de Metileno (EMB - Britania), las que se
incubaron a 35±0,5 ºC durante 24 h para la recuperación de E. coli. Las colonias
compatibles con este microorganismo fueron subcultivadas en agar tripticasa de soja
(ATS - Britania). A partir de este medio se realizó la identificación bioquímica y la
caracterización genotípica. En la Figura 4 B y C se observa el aislamiento en medio
EMB y la identificación bioquímica (81).

33
Todos los aislamientos fueron identificados mediante las siguientes pruebas
bioquímicas clásicas:
Prueba de oxidasa.
Utilización de citrato en medio de Simmons.
Fermentación de glucosa, lactosa y sacarosa con producción de ácido y gas en agar
hierro tres azúcares (TSI).
Producción de sulfuro, indol y determinación de movilidad en medio sulfuro, indol
movilidad (SIM).
Decarboxilación de lisina y ornitina en medio Moeller suplementado al 1% con el
aminoácido.
Producción de ureasa en medio de Christensen.
Producción de fenilalaninadesaminasa (FA)
Muestra positiva: se definió como muestra positiva aquella en la cual se
recuperaron cepas bioquímicamente identificadas como E. coli. Por cada muestra
positiva, se analizaron por PCR dos cepas de manera individual.
Figura 4. Secuencia metodológica empleada. A. Enriquecimiento directo. B. Cultivo en agar Eosina
Azul de Metileno (EMB). C. Identificación con pruebas bioquímicas, de izquierda a derecha: Citrato, TSI,
SIM, FA, Ureasa, Lisina y Ornitina decarboxilsas.

34
3.3 Detección genotípica de los factores de virulencia
La secuencia de pasos para la detección de los factores de virulencia en los
aislamientos de E. coli fueron los siguientes.
3.3.1 Preparación de los templados.
A partir de las estrías de ATS de las cepas bioquímicamente identificadas como
E. coli se tomó una ansada de cultivo. Esta ansada se resuspendió en 150 µl de buffer
Tritón X-100 al 1% en buffer TE 1X en tubos Eppendorf. Los tubos se sometieron a
100ºC en baño seco durante 10 minutos y posteriormente a centrifugación a 10000 rpm
durante 1 minuto. El sobrenadante obtenido se utilizó como templado de la reacción de
PCR múltiple. Este procedimiento es una modificación del propuesto por Toma y col.
(2003) en el cual se parte de bacterias provenientes de un cultivo de 18 h en caldo
Luria-Bertani y como agente lisante utilizan un buffer conteniendo Tween 20 y
proteinasa K (62,82).
3.3.2 Protocolo de amplificación.
El protocolo de amplificación utilizado fue el estandarizado por Toma y
col.(2003). La mezcla de reacción utilizada contenía 10 mM de TRIS-HCl (pH 8,3), 50
mM de KCl, 0,1% de gelatina, 1,5 mM de MgCl2, 2,5 U de Taq ADN polimerasa, 0,2
mM de desoxinucleótidostrifosfato, 5 µl del templado y los cebadores en las
concentraciones que se muestran en la Tabla 1. Volumen final de reacción 50 µl. (82)
Se utilizaron 6 pares de cebadores que corresponden a 6 genes codificantes de
factores de virulencia pertenecientes a 5 tipos patotipos de E. coli, a saber: ETEC (elt y
est), EPEC (eae), STEC (stx), EIEC (ipaH) y EAEC (aggR).
El programa de amplificación fue de 30 ciclos de 1 minuto de desnaturalización
a 95 ºC, 1 minuto de hibridación a 52 ºC y 1 minuto de extensión a 72 ºC más una
extensión final de 10 minutos a 72 ºC.

35
Tabla1: Características de los cebadores utilizados en la determinación genotípica de
los aislamientos de E. coli mediante PCR múltiple.
Cebador Secuencia del cebador (5´-3´)
Tamaño
de la
banda
(pb)
Gen Concentración
en la mezcla
SK1 CCCGAATTCGGCACAAGCATAAGC 881 eae
0,125 μM
SK2 CCCGGATCCGTCTCGCCAGTATTCG 0,125 μM
VTcom-u GAGCGAAATAATTTATATGTG 518 stx
0,25 μM
VTcom-d TGATGATGGCAATTCAGTAT 0,25 μM
AL65 TTAATAGCACCCGGTACAAGCAGG 147 est
0,5 μM
AL125 CCTGACTCTTCAAAAGAGAAAATTAC 0,5 μM
LTL TCTCTATGTGCATACGGAG 322 elt
0, 25 μM
LTR CCATACTGATTGCCGCAAT 0, 25 μM
ipa III GTTCCTTGACCGCCTTTCCGATACCGTC 619 ipaH
0,125 μM
ipa IV GCCGGTCAGCCACCCTCTGAGAGTAC 0,125 μM
aggRKs1 GTATACACAAAAGAAGGAAGC 254 aggR
0,25 μM
aggRkas2 ACAGAATCGTCAGCATCAGC 0,25 μM
Como controles positivos se incluyeron las cepas que se detallan en la Tabla 2.
Las mismas fueron gentilmente provistas por el Servicio Fisiopatogenia del INEI–
ANLIS “Dr. Carlos G. Malbrán”. En la Figura 5 se muestran los productos de
amplificación de controles y muestras.
Tabla 2: Controles positivos utilizados para la identificación de los diferentes patotipos
de E. coli. Cepa Descripción Amplicón (pb) Gen Referencias
E. coli 481 E. coli enteroinvasivo 619 ipaH Cerna y col.
(2003). Toma y
col. (2003).
Botkin y col.
(2012) (55,82,83)
E. coli 17-2 E. coli enteroagregativo 254 aggR
E. coli KNH-172 E. coli enterotoxigénico 147-322 est - elt
E. coli EDL933 E. coli O157:H7 881+518 eae / stx
Los productos de amplificación se separaron por electroforesis en gel de agarosa
al 2% (Biodynamics, Argentina) en buffer TRIS acetato EDTA (TAE) 1 X. Los geles se
tiñeron con GelRedTM
(Biotium) y se tomó registro fotográfico bajo luz UV (62,82).

36
3.3.3 Subtipificación de aislamientos de E. coli enteroagregativo.
En los aislamientos identificados como EAEC se realizó la búsqueda de los
siguientes factores de virulencia: aap (dispersina), AA probe (también conocido como
pCVD432, codifica una proteína de membrana externa la cual forma parte de un sistema
de proteínas transportadoras). La secuencia de los oligonucleótidos utilizados y el
protocolo de PCR fue el estandarizado por Cerna y col.(2003). Se incluyó como control
positivo a la cepa E. coli 17-2 (55). En la Figura 6 se muestran los productos de
amplificación de controles y muestras.
La mezcla de reacción utilizada contenía 10 mM de TRIS-HCl (pH 8,3), 50 mM
de KCl, 2 mM de MgCl2, 1U de Taq ADN polimerasa, 2 mM de
desoxinucleótidostrifosfato, 2µl del templado y los cebadores en las concentraciones
que se muestran en la Tabla 3. Volumen final de reacción 25 µl.
Figura 5. Productos de
amplificación por PCR para
la detección de los genes eae,
stx, lt, st, aggR e ipaH en
cepas de referencia y aisladas
de agua. Línea 1: marcador
de tamaño molecular cien pb.
Línea 2: control negativo E.
coli ATCC 25922. Líneas 3:-
E. coli 933. Línea 4: E. coli
KNH 172. Línea 5: E. coli
17-2 Línea 6: E. coli 481.
Línea 7: cepa de agua aggR +
Líneas 8-10: cepas de agua
negativas

37
El programa de amplificación fue de 1 ciclo a 50 ºC durante 2 minutos, 1 ciclo a
95 ºC durante 5 minutos, 40 ciclos de 45 segundos cada uno de desnaturalización a
95ºC, de hibridación a 55 ºC y de extensión a 72 ºC más una extensión final de 10
minutos a 72 ºC.
Tabla 3: Características de los cebadores utilizados en la determinación genotípica de
los factores de virulencia de EAEC mediante PCR múltiple.
Cebador Secuencia del cebador (5´-3´) Tamaño de la
banda (pb) Gen
Concentración
en la mezcla
aap-F CTTGGGTATCAGCCTGAATG
310 aap 10 pMol
aap-R AACCCATTCGGTTAGAGCAC
aggR-F CTAATTGTACAATCGATGTA 457 aggR 15 pMol
aggR-R AGAGTCCATCTCTTTGATAAG
AAprobe-F CTGGCGAAAGACTGTATCAT
629 AA
probe
20 pMol
AAprobe-R CAATGTATAGAAATCCGCTGTT
La electroforesis se realizó en gel de agarosa al 2,5% (Biodynamics, Argentina)
en buffer TAE 1X. Los geles se tiñeron con GelRedTM
(Biotium) y se tomó registro
fotográfico bajo luz UV (55).
Figura 6. Productos de
amplificación por PCR
para la detección de los
genes aap, aggR, AA
probe en cepas de
referencia y aisladas de
agua. Líneas 2 y 6: E.
coli 17-2. Línea 3:
control negativo E. coli
ATCC 25922. Línea 5:
cepa de agua negativa.
Línea 4: marcador de
tamaño molecular cien
pb.

38
3.3.4 Subtipificación de aislamientos de E. coli enteropatógeno.
En los aislamientos identificados como EPEC se realizó la detección del gen
bfpA, marcador que permite discriminar entre cepas típicas (bpf+) y atípicas (bpf-) de
este patotipo. Las secuencias de los cebadores utilizados fue la descripta por Aranda y
col. (2007) y el protocolo de PCR el descripto por Gunzburg y col. (1995). Se incluyó
como control positivo a la cepa E. coli 2348/69 (eae+ y bfp+) gentilmente provista por
el Servicio Fisiopatogenia del INEI–ANLIS “Dr. Carlos G. Malbrán”.
La mezcla de reacción utilizada contenía 10 mM de TRIS-HCl (pH 8,3), 50 mM
de KCl, 2 mM de MgCl2, 2 U de Taq ADN polimerasa, 2 mM de
desoxinucleótidostrifosfato, 5 µl del templado y los cebadores en las concentraciones
que se muestran en la Tabla 4. Volumen final de reacción 50 µl (84,85).
El programa de amplificación fue de 29 ciclos de 30 segundos de
desnaturalización a 94 ºC, 1 minuto de hibridación a 56 ºC y 2 minutos de extensión a
72 ºC.
Tabla 4: Características de los cebadores utilizados en la determinación genotípica del
factor de virulencia bfp de EPEC mediante PCR.
Cebador Secuencia del cebador (5´-3´) Tamaño de la
banda (Pb) Gen
Concentración
en la mezcla
bfpA-F AATGGTGCTTGCGCTTGCTGC 326 bfp
0,5 µM
bfpR-R GCCGCTTTATCCAACCTGGTA
La electroforesis se realizó en gel de agarosa al 2% (Biodynamics, Argentina) en
buffer TAE 1X. Los geles se tiñeron con GelRedTM
(Biotium) y se tomó registro
fotográfico bajo luz UV. En la Figura 7 se muestran los productos de amplificación de
controles y muestras.
En todas las reacciones se utilizó como control negativo a E. coli ATCC 25922
sin factores de virulencia y en todas las electroforesis marcador de peso molecular de
cien pares de bases (pb).

39
El análisis de resultados se llevó a cabo mediante el programa EpiInfo (CDC).
Figura 7. Productos de
amplificación por PCR
para la detección del gen
bfp en cepas de referencia
y aisladas de agua. Líneas
1, 2,3 y 9: E. coli 2348/69.
Línea 4: control negativo
E. coli ATCC 25922.
Línea 5: cepa de agua
positiva. Línea 6:
marcador de tamaño
molecular cien pb. Líneas
7 y 10: cepas de agua
negativas. Línea 8: sin
producto de amplificación

40
4- RESULTADOS
4.1 Origen de las muestras
El muestro se realizó entre los años 2009 y 2014. Se analizaron 362 muestras de
agua obtenidas de fuentes superficiales, subterráneas, aljibes, de red y bajadas de tanque
procedentes de distintos puntos de la Provincia, cuya frecuencia de distribución se
presenta en la Tabla 5. Durante la realización del muestreo no se consideró la
variabilidad estacional o fenómenos meteorológicos previos.
Tabla 5: Origen de las muestras estudiadas distribuidas según la fuente de agua.
Fuente Nº Porcentaje
Superficiales 150 41,4
Subterráneas 187 51,7
Aljibe 8 2,2
Bajada de tanque 8 2,2
Red 9 2,5
Total 362 100
E. coli se recuperó en 184 muestras (50,8%), en tanto que las 178 (49,2%)
restantes resultaron negativas para este microorganismo.
4.2 Fuentes superficiales
4.2.1 Prevalencia de E. coli en fuentes superficiales.
Se analizaron 150 muestras de agua procedentes de diferentes cuerpos y cursos
superficiales de la Provincia. De ellas, 62 provenían de lagunas, 82 de ríos y 6 de
represas. Se constató la ausencia de descargas de efluentes industriales, cloacales, de
frigoríficos o de feedlots 300 metros aguas arriba del punto de toma de muestra.

41
En el Anexo I se detalla el origen geográfico de las muestras de fuentes
superficiales analizadas y la frecuencia de muestreo.
Sobre las 150 muestras estudiadas, en 111 (74%) se recuperaron cepas de E.
coli. De éstas, 50 (33,3%) provenían de lagunas, 58 (38,7%) de ríos y 3 (2%) de
represas.
4.2.2 Distribución de factores de virulencia en los aislamientos de E. coli de
fuentes superficiales
Por cada muestra positiva se estudiaron dos aislamientos de E. coli. Cuando se
realizó la caracterización de los mismos mediante la detección de los genes de
virulencia de ETEC (elt y est), EPEC (eae), STEC (stx), EIEC (ipaH) y EAEC (aggR),
de las 111 muestras en las cuales se aisló E. coli, 32 (29%) resultaron positivas para
alguno de los genes estudiados. La prevalencia de aislamientos pertenecientes a alguno
de los patotipos en estas fuentes fue del 21,3%. En cada muestra sólo se recuperó un
patotipo.
En la Tabla 6 se muestran las prevalencias de los tres patotipos detectados en
relación al número total de fuentes de agua superficiales estudiadas. Los aislamientos
portadores del gen aggR, clasificados como EAEC, resultaron los prevalentes (15,3%).
Su frecuencia entre los aislamientos de E. coli fue del 20,7%.
En estos aislamientos se realizó la investigación de otros genes de virulencia de
EAEC: el aap y AA probe (pCDV432). En 3 (13%) de ellos se determinó la presencia
del gen app que codifica para la dispersina, todos provenientes de la misma fuente
superficial, laguna Argüello, en muestreos realizados en diferentes momentos (años
2009-2010). En 2 (8,7%) se determinó la presencia del AA probe, procedentes del R.
Negro y laguna Los Teros.

42
Tabla 6: Prevalencia de cepas diarreogénicas de E. coli en las fuentes de agua
superficiales evaluadas del Chaco.
Patotipos Nº (%)
EAEC 23 (15,3)
ETEC 6 (4,0)
EPEC 3 (2,0)
E. coli sin factores de virulencia 79 (52,7)
Muestras sin E. coli 39 (26)
Total 150 (100)
Abrev. EAEC: Escherichia coli enteroagregativo. ETEC: E. coli enterotoxigénico. EPEC: E. coli
enteropatogénico.
Entre las muestras positivas en 6 (5,4%) se identificaron aislamientos del
patotipo ETEC, de los cuales 2 de ellos poseían solamente el gen lt, 2 el gen st, y los 2
restantes la combinación lt-st.
En 3 (2,7%) muestras positivas se determinó la presencia de aislamientos
portadores del gen eae los que fueron identificados como EPEC. Por consenso, la
presencia de este gen en ausencia de otro marcador de virulencia es suficiente para
definir este patotipo (32,82).
La discriminación de los aislamientos caracterizados como EPEC en cepas
típicas ó atípicas se realizó por la posterior detección del gen bfpA por PCR. No se
detectó la presencia del citado gen en los aislamientos por lo que fueron clasificados
como cepas aEPEC.
En las fuentes superficiales evaluadas no se recuperaron cepas portadoras de
genes de los patotipos de EIEC y STEC, ni se detectó la presencia de dos patotipos
simultáneamente.

43
4.2.3 Prevalencia de cepas diarreogénicas en las diferentes fuentes de agua
superficiales
En 17 (11,3%) de las 62 muestras de agua de lagunas se determinó la presencia
de patotipos diarreogénicos de E. coli, y sobre las 82 muestras de ríos, 15 (10%)
resultaron positivas para estos tipos patogénicos. En ambas fuentes superficiales EAEC
resultó el patotipo más prevalente (15,3%).
En ninguna de las 6 muestras de represa se constató la presencia de cepas
diarreogénicas de E. coli.
No se encontró diferencia estadísticamente significativa entre la presencia de
cepas diarreogénicas de E. coli en las lagunas y los ríos estudiados (P=0,35).
En el Gráfico 1 se compara la distribución de los patotipos detectados en lagunas
y ríos, y en la Tabla 7 se detalla el origen geográfico de cada aislamiento.
Gráfico 1: Distribución de los patotipos EAEC, EPEC y ETEC según el origen de los
aislamientos.
13
2 2
10
1
4
0
2
4
6
8
10
12
14
EAEC EPEC ETEC
N°
de
mu
est
ras
po
siti
va
s
Patotipo
Lagunas
Ríos

44
Tabla 7: Origen y frecuencia de las muestras superficiales positivas para los patotipos
EAEC, ETEC y EPEC detectados.
PATOTIPOS ORIGEN Y FRECUENCIA
EAEC
Lagunas N°=13 ( 11,7%): Argüello (7), Francia (1), El Sauzal (2),
La Verde (1), Los Lirios (1) y Los Teros (1).
Ríos N°=10 (9% ): Negro (8) y Paraná (2)
ETEC
Lagunas N°=2 (1,8%): Argüello (1) y Moncholo (1)
Ríos N°=4 (3,6%): Bermejito (1), Guaycurú (1), Negro (1) y
Paraguay (1)
EPEC
Lagunas N°=2 (1,8%): Ávalos (1) y Moncholo (1)
Ríos N°=1 (0,9%): Negro
Abrev. EAEC: Escherichia coli enteroagregativo. ETEC: E. coli enterotoxigénico. EPEC: E. coli
enteropatogénico
En las Figuras 8 y 9 se indican los puntos geográficos de toma de muestra a
partir de los cuales se recuperaron cepas de E. coli de los distintos los patotipos.
Figura 8: Localización geográfica de las lagunas de la ciudad de Resistencia positivas para los
diferentes patotipos. EAEC: (1) Lag. Argüello, (2) Lag. Francia, (3) Lag. Los Lirios (4) Lag. Los
Teros. ETEC: (1) Lag. Argüello. EPEC: (5) Lag. Ávalos. Fuente: Google maps.

45
4.3 Fuentes subterráneas
4.3.1 Prevalencia de E. coli en fuentes subterráneas
Las muestras de agua subterránea se obtuvieron a través de los pozos o
perforaciones existentes en los hogares. Las mismas fueron tomadas en 22 de los 25
Departamentos de la provincia del Chaco. Solo se tomaron muestras en aquellos
hogares donde el agua subterránea es la única fuente disponible para el consumo, por lo
que su contaminación representa un riesgo para la salud. En el Anexo II se detalla las
1
2
5
3
6
7
1-Lag. Sauzal
2-Lag. La Verde
4-R. Negro
3-R. Paraná
7-R. Guaycurú
5-Lag. Moncholo 6-R. Paraguay
4
8-R. Bermejito 8
Figura 9: Localización geográfica de las fuentes superficiales de agua positivas para los
diferentes patotipos. EAEC: (1) Lag. El Sauzal, (2) Lag. La Verde, (3) R. Paraná (4) R. Negro.
ETEC:(4) R. Negro, (5) Laguna Moncholo, (6) R. Paraguay, (7) R. Guaycurú, (8) Río Bermejito.
EPEC:(4) R. Negro, (5) Laguna Moncholo. Fuente: adaptación de mapa de mapoteca.edu.ar

46
localidades y frecuencia de muestreo por Departamento y en el Anexo III la localización
geográfica.
Se analizaron 187 muestras de agua, en 61 (32,6%) de ellas se recuperaron cepas
de E. coli. La presencia de E. coli en estas muestras determina que deban ser
consideradas como no aptas para el consumo humano, según lo establecido en el
Capítulo XII del Código Alimentario Argentino (26).
5.3.2 Distribución de factores de virulencia en los aislamientos de E. coli
provenientes de fuentes subterráneas.
En 19 de las 61 muestras positivas para E. coli, se identificaron aislamientos
portadores de alguno de los factores de virulencia estudiados, por lo que la prevalencia
de E. coli diarreogénicos en estas fuentes fue del 10,1%.
En la Tabla 8 se muestran las prevalencias para los cuatro patotipos detectados
en relación al número total de muestras de agua subterránea estudiadas.
Tabla 8: Prevalencia de cepas diarreogénicas de E. coli en las muestras de fuentes
subterráneas evaluadas del Chaco.
.Patotipos Nº (%)
EAEC 16 (8,6)
ETEC 1 (0,5)
EPEC 1 (0,5)
EIEC 1 (0,5)
E. coli sin factores de virulencia 42 (22,5)
Fuentes sin E. coli 126 (67,4)
Total 187 (100)
Abrev. EAEC: Escherichia coli enteroagregativo. ETEC: E. coli enterotoxigénico. EPEC: E. coli
enteropatogénico. EIEC: E. coli enteroinvasivo

47
Entre las muestras positivas, en 16 (26,2%) se determinó la presencia de
aislamientos portadores del gen aggR los que fueron clasificados como EAEC,
resultando el patotipo más prevalente, al igual que en las fuentes superficiales.
Las muestras que resultaron positivas para EAEC y su frecuencia fueron
tomadas en las siguientes localidades: Charata (2), Sauzalito (1), Gral. Pinedo (2), Las
Breñas (1), Las Palmas (1), Machagai (1), Pampa del Infierno (3), Paraje La Matanza
(1), Resistencia (2), Sáenz Peña (1) y Tres Isletas (1).
De la misma manera que en fuentes superficiales, en los aislamientos de estas
muestras se realizó la investigación de otros genes de virulencia de EAEC; en 4 (25%)
de ellos se detectó la presencia del gen app y en 1 (6,2%) se determinó la presencia de
la combinación AA probe-aap. Los aislamientos portadores de estos genes procedían de
las localidades de Charata, Gral. Pinedo, Pampa del Infierno, Resistencia y Sáenz Peña.
En 1 (1,6%) muestra de la localidad de Tres Isletas se determinó la presencia de
aislamientos portadores del gen st, los que fueron clasificados como ETEC.
En la localidad de Basail se detectó 1 (1,6%) muestra donde los aislamientos
fueron positivos para el gen eae y se clasificaron como EPEC. En la posterior detección
del gen bfpA, en cada uno de ellos por PCR, permitió su caracterización como cepas
típicas de EPEC (bfp+).
El patotipo EIEC se detectó en a través del gen ipaH en los aislamientos de 1
(1,6%) muestra procedente de Puerto Tirol.
Al igual que en las fuentes superficiales, no se recuperaron cepas portadoras de
genes del patotipo STEC, ni la presencia simultánea de más de un patotipo en la misma
muestra.
En las 42 (68,8%) muestras restantes no se detectaron aislamientos de E. coli
portadores de los genes de virulencia estudiados.

48
En la Figura 10 se presenta la localización geográfica de las localidades cuyas
muestras de agua resultaron positivas para los diferentes patotipos.
No se encontró diferencia estadísticamente significativa entre la presencia de
cepas diarreogénicas de E. coli en las fuentes superficiales y subterráneas estudiadas
(P=0,75)
5.4 Muestras de aljibe
Se estudiaron 8 muestras procedentes de agua de aljibes, en 5 (62,5%) de ellas se
detectó la presencia de E. coli.
Figura 10: Localización geográfica de las fuentes de agua subterráneas positivas para los diferentes
patotipos. EAEC: (1) Charata, (2) Sauzalito, (3) Gral. Pinedo, (4) Las Breñas, (5) Las Palmas, (6)
Machagai, (7) Pampa del Infierno, (8) Paraje La Matanza, (9) Resistencia, (10) Sáenz Peña, y (11)
Tres Isletas. ETEC: (11) Tres Isletas. EPEC (12) Basail. EIEC (13) Puerto Tirol

49
La aplicación de la técnica de PCR múltiple permitió caracterizar a las cepas
como pertenecientes al patotipo EAEC en 3 (60%) de las muestras positivas para E.
coli. Las mismas procedían de las localidades de Campo Largo, General Pinedo y
Resistencia.
En estos aislamientos no se identificaron cepas portadoras de los genes de
virulencia aap y AA probe. La prevalencia de EAEC con respecto a la totalidad
demuestras de agua de aljibes fue del 37,5%.
5.5 Muestras de red y de bajada de tanque
Muestras de red. Se analizaron 9 muestras de agua procedentes de red
colectadas a la salida de las plantas de tratamiento. Las mismas fueron tomadas en
Barrio Pedro Pescador (1), General Vedia (2) Miraflores (1), Tartagal (1) y Villa Río
Bermejito (4).
Tres (33,3%) muestras resultaron positivas para E. coli. Cuando se realizó el
estudio molecular de las mismas, no se detectó ninguno de los genes de virulencia de los
patotipos estudiados.
Bajada de tanque. Se estudiaron 8 muestras de bajadas de tanque en domicilios
particulares en General Vedia (2) y Villa Río Bermejito (6). De ellas, 4 (50%) resultaron
positivas para E. coli y en el análisis molecular de los genes de virulencia se determinó
la presencia del gen aggR correspondiente a EAEC en 2 de las muestras tomadas en
Villa Río Bermejito. En estos aislamientos no se detectaron los genes aap y AA probe.
En el Anexo IV se detalla las localidades y frecuencia de muestreo por
Departamento de las muestras de aljibes, red y bajadas de tanque.
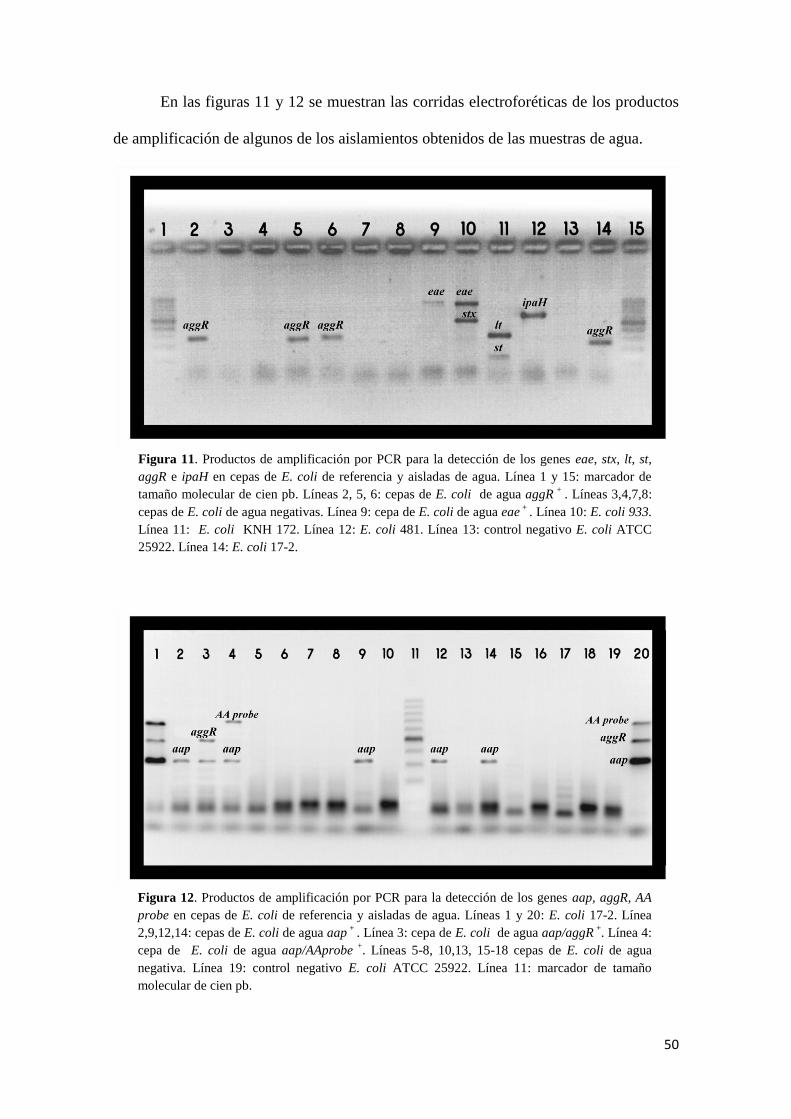
50
En las figuras 11 y 12 se muestran las corridas electroforéticas de los productos
de amplificación de algunos de los aislamientos obtenidos de las muestras de agua.
Figura 11. Productos de amplificación por PCR para la detección de los genes eae, stx, lt, st,
aggR e ipaH en cepas de E. coli de referencia y aisladas de agua. Línea 1 y 15: marcador de
tamaño molecular de cien pb. Líneas 2, 5, 6: cepas de E. coli de agua aggR +
. Líneas 3,4,7,8:
cepas de E. coli de agua negativas. Línea 9: cepa de E. coli de agua eae +
. Línea 10: E. coli 933.
Línea 11: E. coli KNH 172. Línea 12: E. coli 481. Línea 13: control negativo E. coli ATCC
25922. Línea 14: E. coli 17-2.
Figura 12. Productos de amplificación por PCR para la detección de los genes aap, aggR, AA
probe en cepas de E. coli de referencia y aisladas de agua. Líneas 1 y 20: E. coli 17-2. Línea
2,9,12,14: cepas de E. coli de agua aap +
. Línea 3: cepa de E. coli de agua aap/aggR +
. Línea 4:
cepa de E. coli de agua aap/AAprobe +
. Líneas 5-8, 10,13, 15-18 cepas de E. coli de agua
negativa. Línea 19: control negativo E. coli ATCC 25922. Línea 11: marcador de tamaño
molecular de cien pb.

51
5- DISCUSIÓN
Este trabajo buscó aportar datos sobre la presencia y distribución de E. coli
diarreogénicos en ambientes acuáticos de la Provincia del Chaco. Al momento de la
realización del mismo no se contaba con información de la participación de las fuentes
de agua de esta región del país en la cadena epidemiológica de estos patógenos.
5.1- Hallazgos en fuentes de agua superficiales
Se analizaron 150 muestras de agua procedentes de fuentes superficiales de la
provincia. A partir de las mismas en 111 (74%) se aislaron cepas de Escherichia coli.
De éstas, 50 (33,3%) provenían de lagunas, 58 (38,7%) de ríos y 3 (2%) de represas, en
tanto que en las 39 (26%) muestras restantes no se aisló la bacteria.
Treinta y dos de las 60 lagunas estudiadas se encuentran en el área del Gran
Resistencia al igual que la zona elegida del Río Negro (26 muestras). El Departamento
San Fernando (ANEXO I), donde se encuentra la capital provincial, presenta la mayor
densidad demográfica de la provincia, con el 86,7% de los hogares conectados a la red
de agua potable y el 46,8% conectado al sistema público de cloacas (6,86). Sin
embargo, un número importante de personas conformaron asentamientos en terrenos
públicos ribereños de las lagunas, los cuales no cuentan con la infraestructura adecuada.
En trabajos previos se demostró la contaminación bacteriológica de estos cuerpos de
agua, situación atribuible a lo descripto previamente, y a las conexiones clandestinas a
los sistemas de drenaje pluvial de Resistencia. donde las lagunas y el río Negro actúan
como cuerpo receptor de los mismos (86–88).

52
La prevalencia de aislamientos portadoras de genes de virulencia en las fuentes
superficiales fue del 21,3%. Este resultado difiere con los hallazgos de otros grupos de
investigación en ecosistemas acuáticos de distintos países.
El porcentaje establecido en el presente trabajo es superior al 11,2% reportado
por Rebello y col. (2014) en Río de Janeiro, Brasil (89), pero inferior al 85% informado
por Sidhu y col. (2013) en Brisbane, Australia (90), y al 61,1% reportado por Ram y
col. (2008) en el río Gomti en India (65).
En Taiwán, Huang y col. (2016) establecieron una prevalencia de cepas
diarreogénicas del 3,3% en reservorios de agua potable y del 36,1% en ríos (91).
Esta prevalencia también es superior al rango del 0,9-10% publicados por Chern
y col.(2004) (92) y Masters y col.(2011) para cepas diarreogénicas en fuentes
superficiales (93).
En estas fuentes no se detectaron, en un mismo aislamiento, genes de virulencia
correspondientes a diferentes patotipos discrepando de los resultados obtenidos por
Ranjbar y col.(2016) (94), Stange y col. (2016) (64), Sidhu y col (2013) (90) y Titilawo
y col. (2015) (95).
La prevalencia de Escherichia coli enteropatógeno (EPEC) en fuentes
superficiales del Chaco fue del 2% (n=3). La clasificación de las mismas se realizó en
función de la presencia del gen eae que codifica la intimina en ausencia de otro
marcador genético (32). Posteriormente, en estos aislamientos se realizó la
identificación del gen bfp. Todos los aislamientos de las fuentes superficiales fueron
clasificados como cepas atípicas de EPEC. Este hallazgo podría estar relacionado con el
carácter emergente de estas cepas como productoras de diarrea en países en desarrollo o
vincularse a reservorios animales (27,34).

53
En la tabla 9 se comparan los resultados encontrados en el presente estudio con
los hallazgos sobre este patotipo de otros grupos de investigación, se detalla en cada
caso los genes estudiados.
Huang y col.(2016) estudiaron muestras de agua en Taiwán y determinaron que
el 17% de muestras de ríos y el 1,7% de reservorios eran positivas para EPEC, por la
detección directa del gen de virulencia eae (91). Por la misma metodología Masters y
col.(2011) también detectaron al gen eae en el 100% de muestras de aguas superficiales
en Brisbane, Australia, durante la estación húmeda (93). Akter y col. (2013)
determinaron que EPEC era el segundo patotipo más prevalente en muestras de fuentes
superficiales de Bangladesh a partir de la detección directa del gen eae, con porcentajes
que oscilaron entre el 65% en invierno y el 19% en verano (96).
Tabla 9: Prevalencia de los genes de virulencia del patotipo EPEC en este estudio en
comparación con otras publicaciones
Lugar Nº de
aislamientos
Genes
estudiados Resultados Referencias
Estados Unidos, Ohio
(Lago Erie)
472 eae 0,01%, sin discriminar
entre típicas (tEPEC) y
atípicas (aEPEC)
Lauber y col.(2003)
(97)
Brasil, San Pablo 133 eae probe/
bfpAprobe
0,01% (2) aEPEC Orsi y col (2007) (98)
Japón, Río Yamato. 549 eae 1% sin discriminar entre
típicas y atípicas
Gomi y col. (2015)
(99)
Argentina, Chaco
111 eae, bfp 2% de aEPEC
Brasil, San Pablo. 99 eae/bfpA 2% de aEPEC Rodrigues y col.
(2016) (100)
Brasil, Río de Janeiro. 178 escV/bfpB 2,2% de aEPEC Rebello y col.(2014)
(89)
Estados Unidos,
California. Bahía de
Avalon
24493 eae y
plásmido EAF
3,6%, con 0,9% tEPEC Hamilton y col.
(2010) (101)
Alemania, Düsseldorf,
Río Rin
47 eae/bfp 4,3%, con 2,1% tEPEC Stange y col. (2016)
(64)
Nigeria, estado de
Osun
300 eae/bfp 6% con 4% tEPEC Titilawo y col. (2015)
(95)
Australia, Brisbane. 300 eae/bfp 24% de tEPEC Sidhu y col. (2013)
(90)
Irán, Provincia de
Alborz
100 eae/bfpA 31% con 3% de tEPEC Ranjbar y col. (2016)
(94)
Bolivia, Río La Paz 48 eae/bfp 35,4% de aEPEC Poma y col. (2016)
(102)

54
La prevalencia de Escherichia coli enterotoxigénico (ETEC) fue del 4% (n=6).
De estos aislamientos, dos eran portadores del gen st, dos del lt y los restantes de la
combinación st-lt.
A diferencia de los resultados aquí presentados, estudios realizados en fuentes
superficiales de California (101), en el río Yamato de Japón (99), en las aguas del lago
Erie (97) y en las del río Rin (64) no detectaron cepas portadoras de los genes de
virulencia de este patotipo. En este último trabajo realizado en Alemania Stange y col.
(2016) postulan que la ausencia de ETEC podría estar relacionada con la baja
temperatura del agua.
En la tabla 10 se comparan los resultados encontrados en el presente estudio con
los hallazgos sobre ETEC de otros grupos de investigación y se detallan en cada caso
los genes estudiados.
Tabla 10: Prevalencia de los genes de virulencia del patotipo ETEC en este estudio en
comparación con otras publicaciones
Lugar Nº de
aislamientos Genes estudiados Prevalencia Referencia
Brasil, San Pablo 133 LT-1 0,01% (2) Orsi y col (2007) (98)
Brasil, Río de
Janeiro
178 elt, estla/esltb 0,6% Rebello y col.(2014)
(89)
Brasil, San Pablo 99 elt/ esth/estp 2% Rodrigues y col.
(2016) (100)
Australia, Brisbane 300 lt, st 2% y 4% de cepas
portadoras de los
genes lt y st
respectivamente.
Sidhu y col. (2013)
(90)
Argentina, Chaco 111 lt, st 4%
China, Río
Minjiang
2788 elt, estA, estB 19,3% Chen y col. (2011)
(103)
Nigeria, Estado de
Osun
300 lt 45% Titilawo y col. (2015)
(95)
Bolivia, Río La
Paz.
48 muestras lt/sth/stp 71% Poma y col. (2016)
(102)
Otros grupos de trabajo también detectaron este patotipo por medio de la
detección directa de los genes de virulencia en muestras de agua. Akter y col.(2013)
demostraron, por medio de la detección de los genes estA y eltB, que ETEC es el
patotipo prevalente en diferentes fuentes de agua en Bangladesh, con valores de

55
prevalencia que oscilaron entre el 63% y el 76% en invierno y verano, respectivamente
(96). Masters y col.(2011) detectaron al gen est1 en el 5% de muestras analizadas
durante la estación seca, y a los genes est1 y eltA durante la estación húmeda, en aguas
superficiales en Brisbane, Australia (93). En Taiwán, Huang y col. (2016) determinaron
un 17% de muestras positivas en ríos y 1,7% en reservorios, mediante la detección del
gen lt (91).
Tal como se expuso anteriormente, ETEC es la principal causa de diarreas en
niños menores de 5 años y el agente etiológico de la diarrea del viajero. Los trabajos
realizados por Lothigius y col. (2010) demostraron la sobrevida de este patotipo en
aguas dulces y salobres durante tres meses sin que se altere la expresión de los genes de
virulencia, por lo cual los ecosistemas acuáticos actúan de reservorios de estas cepas
(40,104). La capacidad de sobrevida de ETEC y el brote de gastroenteritis ocasionado
por este patotipo en Bangladesh durante las inundaciones (71), alertan sobre el posible
riesgo para la salud derivado de su presencia en fuentes superficiales, teniendo en
cuenta las inundaciones que a lo largo de la historia sufrió la provincia del Chaco.
Los aislamientos portadores del gen aggR, correspondiente al patotipo
enteroagregativo (EAEC) resultaron los más prevalentes en las fuentes superficiales de
la provincia del Chaco. En 23 (15,3%) muestras de agua se determinó la presencia de
aislamientos portadores del gen aggR. Por la heterogeneidad de este patotipo, en estos
aislamientos se investigaron otros genes de virulencia de EAEC: el aap y AA probe
(pCDV432). En 3/23 (13%) se determinó la presencia del gen app y en 2/23 (8,7%) se
determinó la presencia del AA probe. Todos los genes estudiados permitieron
determinar la presencia de cepas típicas de este patotipo (27,55,57).

56
En la tabla 11 se comparan los resultados encontrados en el presente estudio con
los hallazgos de otros grupos de investigación y se detallan en cada caso los genes de
EAEC estudiados.
Tabla 11: Comparación de la prevalencia de los genes de virulencia del patotipo EAEC
hallados en este estudio con otras publicaciones
Lugar Nº de
aislamientos
Genes
estudiados Prevalencia Referencia
Brasil, San Pablo 133 AA probe
(hibridación)
0,04% (6) Orsi y col (2007) (98)
Río Yamato, Japón 549 aggR 0,2% Gomi y col. (2015)
(99)
Nigeria, Estado de Osun 300 eagg 2% Titilawo y col. (2015)
(95)
Alemania, Düsseldorf,
Río Rin
47 aggR, astA 4,3% para el gen
aggR, y 4,3% para del
gen astA.
Stange y col. (2016)
(64)
Argentina, Chaco
111 aggR 15,3%
Australia, Brisbane 300 aggR, astA 29%
36% en estación seca y
26% en la húmeda.
Sidhu y col. (2013)
(90)
Bolivia, Río La Paz. 48 muestras paa 46% Poma y col. (2016)
(102)
A diferencia de los resultados aquí obtenidos, Rodrigues y col. (2016) en San
Pablo Brasil no detectaron cepas portadoras del gen aggR, pero sí 35% portadoras del
gen astA (100). También en el estado de San Pablo, en 15 aislamientos recuperados del
río Tiete, Carlos y col. (2011) no detectaron cepas de EAEC utilizando el método de
hibridación (105). Coincidentemente, Rebello y col. (2014) tampoco detectaron
aislamientos portadores de los genes aggR, astA y pic en fuentes superficiales de la
región de Río de Janeiro (89).
En diferentes fuentes de agua dulce en Bangladesh, Akter y col. (2013)
demostraron por medio de la detección del gen pCVD432, la presencia y fluctuación
estacional de EAEC, con valores de prevalencia que oscilaron entre el 18% y el 4% en
invierno y verano, respectivamente (96).

57
La heterogeneidad de las cepas de EAEC representa el principal obstáculo para
encontrar un gen que permita su identificación. El gen aggR, es uno de los más
estudiados y se encuentra en el 80% de las cepas de este patotipo, en tanto que el gen
aap también se lo detecta en cepas no patogénicas de E. coli. AA probe sólo fue
detectado en cepas de EAEC. Los tres genes de virulencia están presentes en el
plásmido AA, es decir que en este trabajo se identificaron genéticamente a las cepas
típicas de EAEC, las que representan el grupo mayoritario de este patotipo (54–56).
En las muestras tomadas de estas fuentes no se detectaron aislamientos
portadores del gen stx, es decir no se detectaron cepas de Escherichia coli productor de
toxina Shiga (STEC). Estos resultados concuerdan con los publicados por Hamilton y
col. (2010) en la Bahía de Avalon en California (101), con los de Gomi y col. (2015) en
el Río Yamato en Japón (99), con los de Lauber y col. (2003) en el lago Erie (97) y con
los de los reservorios de agua en Taiwán publicados por Huang y col. (2016) (91).
También coinciden con los resultados de Carlos y col. (2011) en el río Tiete, San Pablo
Brasil, y con los de Poma y col.(2016) en el Río la Paz de Bolivia (102,105).
Con la finalidad de evaluar la presencia o ausencia de los diferentes patotipos en
las fuentes superficiales y no en efluentes, el muestreo no se realizó en los puntos de
descarga, o aguas debajo de efluentes cloacales, de frigoríficos o de feedlots,
considerando que el ganado vacuno es uno de los principales reservorios de este
patotipo. Este criterio en la etapa de muestreo podría condicionar los resultados
obtenidos para este patotipo. En una zona ganadera de la provincia de Entre Ríos,
Tanaro y col. (2014) determinaron la presencia de STEC O157:H7 en el 12,7% de las
251 muestras tomadas aguas abajo de la descarga de efluentes de feedlots, y también en
el 10% de las 60 muestras tomadas aguas arriba de las descargas (75).

58
En muestras superficiales de Brasil, Rodrigues y col. (2016) determinaron un 1%
de cepas portadoras del gen stx1 en el estado de San Pablo, en tanto que Rebello y col.
(2014) detectaron un 2,8% de cepas portadoras del gen stx1 en ecosistemas acuáticos de
Río de Janeiro (89,100). En balnearios de la región de Sierra de la Ventana, provincia de
Buenos Aires, Marucci y col (2011) identificaron 4/35 aislamientos de E. coli
portadores del gen stx2 (76).
En las fuentes superficiales no se detectaron aislamientos portadores del gen
ipaH correspondiente al patotipo EIEC. Este resultado concuerda con los hallazgos de
estudios realizados en Japón (99), Brasil (89), y Nigeria (95). Pero, a la vez difiere de
aquellos procedentes de otros ecosistemas superficiales. En el río Rin Stange y col
(2016) detectaron un 2,1% de aislamientos portadores de este gen, al tiempo que
Ranjbar y col. (2016) informaron un 97% de aislamientos portadores a partir de fuentes
superficiales en Irán (64,94).
5.1.1 Consideraciones finales sobre las fuentes de agua superficiales
La detección de los patotipos, EPEC, ETEC, EAEC en las fuentes de agua
superficiales estudiadas es evidencia de la vulnerabilidad de los mismos a la
contaminación principalmente de origen antrópico. Esta última apreciación se sustenta
en que EAEC fue el patotipo más prevalente, su reservorio es el ser humano, y en la
cobertura de red cloacal de la provincia.
La presencia de cepas portadoras de genes de virulencia representa una
oportunidad de intercambio genético en los microorganismos y un riesgo para la salud
ante los diferentes usos que se les puede dar a las mismas: recreación, riego o fuente
para potabilización.

59
El ejemplo de este intercambio genético más reciente es la emergencia del
híbrido EAEC/STEC causante de un brote de diarrea sanguinolenta y SUH en
Alemania, en el año 2011. La cepa causal fue caracterizada como E. coli
enteroagregativo O104:H4, que adquirió por transferencia horizontal el gen stx de E.
coli productor de toxina Shiga (6,27,61,89).
Los brotes de gastroenteritis relacionados a actividades recreativas en aguas
contaminadas con E. coli diarreogénicos fueron reportados en Estados Unidos (106) y
Europa, gracias al sistema de vigilancia existente en esos países, resultando ser los niños
los más susceptibles por la alta ingesta de agua durante dichas actividades (100).
La falta de tratamiento de los efluentes cloacales representa una debilidad en
toda la Cuenca del Plata, y un punto a tener en cuenta por las empresas proveedoras de
agua potable. Los resultados aquí obtenidos constituyen una herramienta al momento de
realizar una evaluación de riesgos de la cuenca de captación, particularmente en lo
referente al Área Metropolitana del Gran Resistencia, dado que actualmente la toma de
agua del principal acueducto del Chaco se encuentra aguas abajo de la descarga del Río
Negro, el cual es el receptor final del sistema lacustre de la zona Norte y receptor de
efluentes cloacales e industriales (12,18).
A diferencia de lo que se observó en el presente trabajo, cuando se protegen los
ecosistemas acuáticos se manifiesta una mejor calidad microbiológica. Esto se reflejó en
los trabajos de Huang y col. y Poma y col. (91,102).
5.2 Hallazgos en fuentes subterráneas
5.2.1-Contaminación de las fuentes de agua subterráneas
En la provincia del Chaco, el 14% de los hogares se abastecen de agua a partir
de fuentes subterráneas ubicadas tanto dentro como fuera del terreno, la cual es utilizada

60
directamente, sin tratamiento previo. A su vez, sólo el 26,4% de los hogares tienen
cloaca conectada a la red pública. En el resto, la descarga de los efluentes cloacales se
realiza a través de cámara séptica, pozo ciego o a excavaciones directas en la tierra (6).
La contaminación microbiológica del agua subterránea se atribuye
principalmente a la infiltración de agua contaminada con materia fecal humana o de
animales. Los factores que favorecen este proceso son el diseño inadecuado, ubicación,
construcción, operación y mantenimiento de los pozos y/o perforaciones utilizadas para
acceder al agua, como así también las características geológicas y condiciones
climáticas. Raramente la contaminación de estas fuentes se deba a un solo factor.
Complementariamente, las bombas manuales utilizadas para la extracción de agua
pueden actuar como reservorios microbianos y como fuente de contaminación del agua
subterránea. En las viviendas en las cuales se realizó el muestreo no poseían agua
potable ni conexión al sistema público de cloacas. La contaminación del agua de estos
pozos y perforaciones se debería principalmente a la infiltración de los efluentes
domésticos, siendo la ubicación y el mantenimiento los principales factores para que
ello ocurra (107–109).
5.2.2 Calidad microbiológica y distribución de cepas diarreogénicas en las
fuentes estudiadas
Las muestras de agua subterránea se tomaron en aquellos hogares donde el agua
subterránea es la única fuente disponible para el consumo. Se analizaron 187 muestras
de agua, en 61 (32,6%) de ellas se recuperaron cepas de E. coli. Las 126 (67,4%)
restantes fueron negativas para el indicador.
El Código Alimentario Argentino, en su capítulo XII, establece la ausencia de E.
coli en 100 ml como uno de los criterios microbiológicos del agua destinada al consumo

61
humano (26). La presencia de E. coli en el 32,6% de las muestras, determina que se
deban considerar como no aptas para el consumo humano. En el 67,4% restante, si bien
no se detectó la presencia de este microorganismo, al no realizarse la determinación de
la totalidad de los parámetros microbiológicos no se puede dictaminar si las mismas son
bacteriológicamente aptas para el consumo humano.
Los valores encontrados de E. coli (32,6%) en estas fuentes son similares a datos
publicados por otros grupos de investigación en diferentes países. Así Ferguson y col.
(2012) determinan la presencia de este indicador en el 40% de las 50 muestras
estudiadas en Bangladesh (16); Batabyal y col. (2013) en el 24,3% de las 74 muestras
en Bengala Occidental, India (110) y Brown y col. (2208) en el 21% de 12.194 muestras
en Camboya (111). En Inglaterra, Richardson y col. (2009) reportan un valor de 9,85%
hallado en 6588 muestras, inferior al valor del presente estudio (66).
En Argentina, Gambero y col. (2014) determinaron la presencia de E. coli en
19% de las 36 muestras tomadas en la zona de Río Cuarto, Córdoba (112). Por otra
parte, Juárez y col. (2015), en su trabajo de detección de patógenos virales y
parasitarios, informaron la presencia de este indicador en fuentes de agua subterráneas
de la ciudad de Salta, sin mostrar datos de su cuantificación (113).
De las 61 muestras positivas para E. coli, en 19 se identificaron aislamientos
portadores de alguno de los genes de virulencia estudiados. La prevalencia de E. coli
diarreogénicas en estas fuentes fue del 10,1%. Se determinó la presencia de los
patotipos EAEC, ETEC, EPEC y EIEC.
La prevalencia de EAEC fue del 8,6%. En tanto que para los patotipos restantes
la prevalencia de cada uno de ellos fue del 0,5%. No se detectó la presencia
aislamientos del patotipo STEC, portadores del gen stx, en las muestras de estas fuentes.

62
La distribución de cepas diarreogénicas en fuentes de agua subterráneas está
poco estudiada. Los resultados de prevalencia total obtenidos en el presente trabajo son
similares a los datos que informaron Ferguson y col. del 8% a través de la detección de
los genes ipaH, elt, eae y stx. en Bangladesh, aunque este grupo no estudió la presencia
del gen aggR (16).
En India se identificó un 9% (8/88) de aislamientos de E. coli de los patotipos
EPEC, ETEC y EIEC en todos las muestras estudiadas, entre las que se incluyen las de
fuentes subterráneas (110).
En los trabajos citados no se detectó al patotipo EAEC, el más prevalente en las
fuentes de la provincia del Chaco. Sin embargo, se lo reportó como agente causal de
diarrea en la India a partir del consumo de agua subterránea (74,79). La presencia de un
determinado patógeno en estas fuentes está en función de su circulación en la población;
para el caso particular de EAEC y EIEC los animales no constituyen un reservorio
importante. Además, en el caso de EAEC los portadores adultos sanos tendrían un
importante rol en la epidemiología de este patotipo (105,114).
5.2.3 Vulnerabilidad de las fuentes de agua subterráneas
La detección de E. coli en las fuentes subterráneas evidencia su vulnerabilidad a
la contaminación fecal y la no aptitud para el consumo humano. En estas fuentes, la
ausencia del indicador de contaminación fecal denota que es improbable la existencia de
otros patógenos bacterianos. Por el contrario, tan pronto como se detecten estos
indicadores, la probabilidad de aparición de los mismos se incrementa
significativamente. En las aguas subterráneas la sobrevida de las bacterias aumenta por
las bajas temperaturas, la humedad del suelo, el pH neutro o alcalino y la presencia de
carbón orgánico. La presencia y actividad de los microorganismos nativos también

63
juega un rol preponderante en el control de los microorganismos contaminantes
(10,114,115).
La población expuesta a las mismas, en especial los niños, poseen mayor riesgo
de desarrollo de enfermedad diarreica, condición agravada por la presencia de cepas
diarreogénicas como en este caso. En Estados Unidos, el 30,3% (248/818) de los brotes
relacionados al agua de consumo se debieron al agua subterránea no tratada. De los
casos en los cuales se confirmó el agente etiológico, el 11,7% se debió a E. coli. En la
provincia del Chaco, como en otras partes del mundo, los brotes relacionados a estas
fuentes que son de uso particular raramente son reportados a las autoridades de salud,
generando un subregistro de las mismas (15,109).
En el presente trabajo se determinó una prevalencia de cepas diarreogénicas del
10,1% con lo cual se puede concluir que estas fuentes de agua pueden actuar como
reservorio y vehículo de estas cepas. Ante los resultados obtenidos, no se debe
subestimar la presencia de un potencial riesgo para la salud en las personas que
consumen de las fuentes de agua estudiadas.
5.3 Hallazgos en otras muestras estudiadas
5.3.1 Resultados en aljibes
El aljibe representa una alternativa de almacenamiento de agua de origen pluvial
destinada principalmente al consumo humano. En 5/8 (62,5%) de las muestras se
detectó la presencia de E. coli, y de ellas 3 se correspondieron al patotipo EAEC.
Los resultados obtenidos en este trabajo concuerdan con los Dobrowsky y col.
(2016) en Sudáfrica, donde reportaron a EAEC como el patotipo más prevalente en
estas muestras pero también la presencia de EPEC y EHEC (116). En tanto que en
Australia, Ahmed y col. (2011) detectaron cepas portadoras de genes de virulencia de

64
los patotipos EPEC y ETEC (117). A diferencia de los hallazgos de ambos grupos, en
las muestras analizadas del Chaco sólo se encontró un patotipo, posiblemente por el
número de muestras estudiadas.
Los resultados obtenidos demuestran que esta alternativa de captación y
almacenamiento de agua representa un riesgo para la salud de no mediar una forma de
tratamiento para transformarla en bacteriológicamente apta para el consumo humano
(26,116,117).
Hasta tanto se amplíe la cobertura de agua potable en la provincia, se requieren
acciones conjuntas de los laboratorios de ambiente y salud pública a fin de realizar
controles periódicos a las fuentes de agua subterránea y de aljibes y la educación de la
población para el tratamiento del agua antes de destinarla al consumo humano,
principalmente la destinada a población pediátrica.
5.3.2 Resultados en muestras de agua de red y de bajadas de tanque
Los resultados obtenidos sobre estas muestras, además de no ser aptas para el
consumo humano según la normativa vigente, también evidencian la necesidad del
manejo integral del abastecimiento de agua a las poblaciones, desde su cuenca de
captación hasta la su almacenamiento en el domicilio. Esta acción se justifica por la
presencia de EAEC en las muestras de fuentes de agua y en las de los tanques
domiciliarios. (18,118).
6.4 Gastroenteritis en el Chaco
En el período en el que se llevó a cabo el presente estudio (2009-2014) se
reportaron 200.580 casos de diarrea y 4 casos de SUH en la provincia del Chaco. Para el
mismo período la tasa de SUH fue menor al 0,5/100.000 habitantes en la región NEA.

65
(46). Durante este período, en la provincia no se realizaron los estudios moleculares
para discriminar los diferentes patotipos de E. coli en los aislamientos clínicos, por lo
cual se desconoce la prevalencia de cepas diarreogénicas de E. coli en la población del
Chaco.
Datos de la ciudad de Corrientes, ubicada a 20 km de Resistencia, y con la cual
existe un intenso intercambio de trabajadores y estudiantes, indican a EAEC junto a
ETEC como los patotipos más frecuentes recuperados de muestras de pacientes
pediátricos y adultos con diarrea aguda (62,119).
Los resultados de los tipos patogénicos encontrados en las fuentes de agua
coinciden con estos datos reportados en pacientes de la región NEA. Representan
además una estimación indirecta de la circulación de cepas diarreogénicas en la
población de la provincia del Chaco, principalmente en lo referente a EAEC, al ser el
hombre su reservorio. Finalmente, resaltan la importancia de la vigilancia para el
conocimiento de la circulación de los diferentes patotipos.
EAEC se considera un patógeno emergente. Responsable de la hospitalización
de niños con diarrea aguda. Es el segundo agente más frecuente implicado en los casos
de diarrea del viajero en América Latina. El elevado inóculo requerido para el desarrollo
de infección por EAEC sugiere al agua y los alimentos como vehículo de este patotipo
(53,58,120).
6.5 Metodología empleada
La secuencia metodológica empleada que incluyó enriquecimiento, cultivo,
identificación bioquímica y posterior caracterización molecular permitió la detección de
cepas de E. coli viables, portadoras de genes de virulencia y su clasificación dentro de
los cinco patotipos estudiados. Al trabajar directamente sobre los aislamientos se evitó

66
las sustancias inhibidores de las técnicas moleculares que pueden encontrarse en
muestras ambientales. También es la secuencia sugerida por otros autores para
diferenciar Shigella spp. y EIEC dado que el gen ipaH es compartido por ambos
géneros, pero las dosis infectivas de ambos patógenos es diferente (9,13,38).
Otros grupos de investigación plantearon la detección directa de genes de
virulencia en los ecosistemas acuáticos. Sin embargo, esta última secuencia
metodológica no permite determinar la prevalencia de cepas patogénicas de E. coli y su
viabilidad, concepto fundamental para interpretar el riesgo para la salud que conlleva su
presencia (38,90,91,93).
Como factores negativos de esta secuencia metodológica se debe citar que no se
reduce el tiempo de análisis, condición crítica cuando el agua se destina al consumo
humano y tampoco se detectan cepas viables no cultivables, las que mantienen todo su
potencial patogénico. Diferentes investigaciones confirman la persistencia de estas
últimas cepas de E. coli en el ambiente (9,13,23).
La selección de dos cepas de E. coli por muestra positiva para su estudio
molecular se realizó por razones operativas. Las técnicas moleculares aplicadas al
control de aguas complementan las limitaciones de los métodos basados en cultivo pero
requieren estandarización y validación (17,80,108). Por el contrario las técnicas de
detección de indicadores bacterianos basadas en su cultivo y/o cuantificación se
encuentran estandarizadas. Esta estandarización permite asegurar la calidad higiénica
del agua destinada a diferentes usos (21,81).
Los resultados obtenidos en fuentes superficiales y subterráneas esbozan la
necesidad y el enfoque de próximas líneas de investigación atendiendo a los siguientes
puntos:
Variabilidad o no en la prevalencia de los diferentes patotipos luego de lluvias.

67
Variabilidad en la distribución estacional de los patotipos.
Presencia de genes de resistencia antibiótica junto con los de virulencia como
contaminantes emergentes.
Estudios en paralelo, en muestras clínicas y ambientales de la provincia, a fin de
evaluar la implicancia o no de las diversas fuentes en posibles brotes por cepas
diarreogénicas de E. coli.

68
6- CONCLUSIONES
El presente trabajo constituye el primer aporte sobre la participación de las
fuentes de agua en la cadena epidemiológica de estos patógenos. en esta región del
país.
La combinación de la técnica de cultivo y los métodos moleculares representan
una alternativa válida en la detección de E. coli diarreogénicos en las fuentes de agua.
En los cuerpos y cursos de agua superficiales de la provincia del Chaco se
detectó la presencia de los patotipos diarreogénicos de Escherichia coli
enteroagregativo, enterotoxigénico y enteropatogénico.
En las fuentes de agua subterránea de la provincia se detectó la presencia de los
patotipos diarreogénicos de Escherichia coli enteroagregativo, enterotoxigénico,
enteropatogénico y enteroinvasivo
En ambas fuentes E. coli enteroagregativo resultó el patotipo prevalente.
Las diferentes fuentes de agua actúan como reservorio y vehículo de cepas
patogénicas de E. coli.
Ante los resultados obtenidos, no se debe subestimar la presencia de un potencial
riesgo para la salud en las personas que consumen de las fuentes de agua estudiadas.
En la provincia del Chaco se requiere seguir trabajando para ampliar la cobertura
de agua potable y de cloacas.
Autor Director Codirectora Consejera de Estudios

69
7- BIBLIOGRAFÍA
1. Maeder E. Historia del Chaco. 1°. Buenos Aires: Editorial Plus Ultra; 1996. 295 p.
2. Martinez G, Beccaglia A, Llinares A. Problemática hídrico-sanitaria, percepción local y
calidad de fuentes de agua en una comunidad toba (qom) del Impenetrable (Chaco,
Argentina). Salud Colect. 2014;10(2):225–42.
3. Bruniard E. El gran Chaco Argentino. Ensayo de interpretación geográfica. Geográfica.
1978;4:259.
4. Alberto J. El Chaco Oriental y sus fisonomías vegetales. Geográfica Digital. [Internet].
2006;3(5). Disponible en: http://hum.unne.edu.ar/revistas/geoweb/homeig0.htm
5. Ministerio de Salud de la Nación, Organización Panamericana de la Salud. Indicadores
básicos. Argentina 2016 [Internet]. CABA, Argentina; 2016. Disponible en:
http://www.deis.msal.gov.ar/index.php/indicadores-basicos/
6. Instituto Nacional de Estadística y Censos- INDEC. Censo Nacional de Población,
Hogares y Viviendas 2010 [Internet]. CABA, Argentina; 2010. Disponible en:
http://www.indec.gov.ar/nivel4_default.asp?id_tema_1=2&id_tema_2=41&id_tema_3=1
35
7. Páll E, Niculae M, Kiss T, Şandru C, Spînu M. Human impact on the microbiological
water quality of the rivers. J Med Microbiol. 2013;62(PART 11):1635–40.
8. Bridle H. Introduction. En: Bridle H, editor. Waterborne Pathogens Detection methods
and applications. 1°. Londres, UK: Academic Press of Elsevier; 2013. p. 416.
9. Mendes Silva D, Domingues L. On the track for an efficient detection of Escherichia
coli in water: A review on PCR-based methods. Ecotoxicol Environ Saf. 2015;113:400–
11.
10. Cabral J. Water microbiology. Bacterial pathogens and water. Int J Environ Res Public
Health. 2010 Oct;7(10):3657–703.
11. World Health Organization. Water quality and health strategy 2013-2020 [Internet].
World Health Organization. Ginebra, Suiza; 2012. Disponible en:
http://www.who.int/water_sanitation_health/publications/2013/water_quality_strategy/e
n/
12. Comité Intergubernamental Coordinador de los Países de la Cuenca del Plata. Programa
para la Gestión Sostenible de los Recursos Hídricos de la Cuenca del Plata en relación
con los efectos de la variabilidad climática y el Cambio Climático [Internet]. CABA,
Argentina; 2011. Disponible en: file:///Users/crogelis/Downloads/Programa Marco -
version marzo 2011.pdf
13. Girones R, Ferrús M, Alonso J, Rodriguez-Manzano J, Calgua B, Corrêa A, Hundesa A,
Carratala A, Boffill-Mas S. Molecular detection of pathogens in water--the pros and cons
of molecular techniques. Water Res. 2010 Aug;44(15):4325–39.
14. Macler B, Merkle J. Current knowledge on groundwater microbial pathogens and their
control. Hydrogeol J. 2000;8(1):29–40.
15. McKay L. Foreword: pathogens and fecal indicators in groundwater. Ground Water.

70
2011;49(1):1–3.
16. Ferguson A, Layton A, Mailloux B, Culligan P, Daniel E, Smartt A, Sayler G, Feighery
J, McKay L, Knappett P, Alexandrova E, Arbit T, Emch M, Escamilla V, Ahmed K,
Alam J, Streatfield P, Yunus M, van Geen A. Comparison of fecal indicators with
pathogenic bacteria and rotavirus in groundwater. Sci Total Environ. 2012;431:314–22.
17. Brettar I, Höfle M. Molecular assessment of bacterial pathogens - a contribution to
drinking water safety. Curr Opin Botechnology. 2008 Jun;19(3):274–80.
18. Bartram J, Corrales L, Davison A, Deere D, Drury D, Gordon B, Howard G, Rinehold A,
Stevens M. Manual para el desarrollo de planes de seguridad del agua: metodología
pormenorizada de gestión de riesgos para proveedores de agua de consumo. [Internet].
1°. Organización Mundial de la Salud, International Water Association, editors. Ginebra,
Suiza: Organización Mundial de la Salud; 2009. 116 p. Disponible en:
http://whqlibdoc.who.int/publications/2009/9789243562636_spa.pdf
19. Harwood V, Staley C, Badgley B, Borges K, Korajkic A. Microbial source tracking
markers for detection of fecal contamination in environmental waters: relationships
between pathogens and human health outcomes. FEMS Microbiol Rev. 2014;38(1):1–
40.
20. Hamelin K, Bruant G, El-Shaarawi A, Hill S, Edge T, Bekal S, Fairbrother J, Harel J,
Maynard C, Masson L, Brousseauet R. A virulence and antimicrobial resistance DNA
microarray detects a high frequency of virulence genes in Escherichia coli Isolates from
Great Lakes recreational waters. Appl Environ Microbiol. 2006;72(6):4200–6.
21. Organización Mundial de la Salud. Guías para la calidad del agua potable. 3°. Vol. 1.
Ginebra, Suiza; 2008. 393 p.
22. Dirk Van Elsas J, Semenov A, Costa R, Trevors J. Survival of Escherichia coli in the
environment: fundamental and public health aspects. ISME J. 2011;5(10):173–83.
23. Ishii S, Sadowsky M. Escherichia coli in the Environment: Implications for Water
Quality and Human Health. Microbes Environ. 2008;23(2):101–8.
24. Saxena G, Bharagava R, Kaithwas G, Raj A. Microbial indicators, pathogens and
methods for their monitoring in water environment. J Water Health. 2015;13(2):319–39.
25. Gutiérrez Cacciabue D, Juárez M, Poma H, Garcé B, Rajal V. Cuantificación y
evaluación de la estacionalidad de elementos parasitarios en ambientes acuáticos
recreativos de la provincia de Salta, Argentina. Rev Argent Microbiol. 2014;46(2):150–
60.
26. Administración Nacional de Medicamentos, Alimentos y Tecnología Médica. Código
Alimentario Argentino. Capítulo XII- Bebidas Hídricas, Agua y Agua Gasificada.
[Internet]. Buenos Aires; 2007. Disponible en:
http://www.anmat.gov.ar/alimentos/codigoa/CAPITULO_XII.pdf
27. Croxen M, Law R, Scholz R, Keeney K, Wlodarska M, Finlay B. Recent advances in
understanding enteric pathogenic Escherichia coli. Clin Microbiol Rev. 2013;26(4):822–
80.
28. Kaper J, Nataro J, Mobley H. Pathogenic Escherichia coli. Nat Rev Microbiol. 2004
Feb;2(2):123–40.
29. Nataro J, Steiner T, Guerrant R. Enteroaggregative Escherichia coli. Emerg Infect Dis.

71
1998;4(2):251–61.
30. Nguyen R, Taylor L, Tauschek M, Browne-Robins R. Atypical Enteropathogenic
Escherichia coli Infection and Prolonged Diarrhea in Children. Emerg Infect Dis.
2006;12(4):1–5.
31. Hu J, Torres A. Enteropathogenic Escherichia coli: Foe or innocent bystander? Clin
Microbiol Infect. 2015;21(8):729–34.
32. Chen H, Frankel G. Enteropathogenic Escherichia coli: Unravelling pathogenesis.
FEMS Microbiol Rev. 2005;29(1):83–98.
33. Trabulsi L, Keller R, Tardelli Gomes T. Typical and atypical enteropathogenic
Escherichia coli. Emerg Infect Dis. 2002;8(5):508–13.
34. Hernandes R, Elias W, Vieira M, Gomes T. An overview of atypical enteropathogenic
Escherichia coli. FEMS Microbiol Lett. 2009;297(2):137–49.
35. Gomes T, Elias W, Scaletsky I, Guth B, Rodrigues J, Piazza R, Ferreira L, Martinez M.
Diarrheagenic Escherichia coli. Brazilian J Microbiol. 2016;47S:3–30.
36. Giugno S, Oderiz S. Etiología bacteriana de la diarrea aguda en pacientes pediátricos.
Acta Bioquím Clín Latinoam. 2010;44(1):63–9.
37. Ud-Din A, Wahid S. Relationship among Shigella spp. and enteroinvasive Escherichia
coli (EIEC) and their differentiation. Brazilian J Microbiol. 2014;45(4):1131–8.
38. Hsu B, Wu S, Huang S, Tseng Y, Ji D, Chen J, Shih F. Differentiation and identification
of Shigella spp. and enteroinvasive Escherichia coli in environmental waters by a
molecular method and biochemical test. Water Res. 2010;44(3):949–55.
39. Viboud G, Jouve M, Binsztein N, Vergara M, Rivas M, Quiroga M, Svennerholm A.
Prospective cohort study of enterotoxigenic Escherichia coli infections in Argentinean
children. J Appl Microbiol. 1999;37(9):2829–33.
40. Vila Estapé J, Zboromyrska Y. Brotes epidémicos causados por Escherichia coli
diarreagénicas. Gastroenterol Hepatol. 2012;35(2):89–93.
41. Daniels N, Neimann J, Karpati A, Parashar U, Greene K, Wells J, Srivastava A, Tauxe
R, Mintz E, Quick R. Traveler’s diarrhea at sea: three outbreaks of waterborne
enterotoxigenic Escherichia coli on cruise ships. J Infect Dis. 2000;181(4):1491–5.
42. Huerta M, Grotto I, Gdalevich M, Mimouni D, Gavrieli D, Yavzori D, Cohen D,
Shpilberg O. A waterborne outbreak of gastroenteritis in the Golan Heights due to
enterotoxigenic Escherichia coli. Infection. 2000;28(5):267–71.
43. Rivas M, Miliwebsky E, Chinen I, Deza N, Leotta G. Epidemiología del sindrome
urémico hemolítico en Argentina. Diagnóstico del agente etiológico, reservorios y vías
de transmisión. Medicina (B Aires). 2006;66(SUPPL. 3):27–32.
44. Tanaro J, Leotta G, Lound L, Galli L, Piaggio M, Carbonari C, Araujo S, Rivas M.
Escherichia coli O157 in bovine faeces and surface water streams in a beef cattle farm of
Argentina. Foodborne Pathog Dis. 2010;7(4):475–7.
45. Leotta G, Miliwebsky E, Chinen I, Espinosa E, Azzopardi K, Tennant S, Robins-Browne
R, Rivas M. Characterisation of Shiga toxin-producing Escherichia coli O157 strains
isolated from humans in Argentina, Australia and New Zealand. BMC Microbiol.
2008;8:46.

72
46. Secretaría de Promoción y Programas Sanitarios. Boletín Integrado de Vigilancia
[Internet]. CABA; 2016. Disponible en:
http://www.msal.gob.ar/index.php/home/boletin-integrado-de-vigilancia
47. Varela G, Chinen I, Gadea P, Miliwebsky E, Mota M, González S, González G,
Gugliada M, Carbonari C, Algorta G, Bernadá M, Sabelli R, Pardo L, Rivas M,
Schelotto F. Detección y caracterización de Escherichia coli productor de toxina Shiga a
partir de casos clínicos y de alimentos en Uruguay. Rev Argent Microbiol. 2008;40:93–
100.
48. Okhuysen P, DuPont H. Enteroaggregative Escherichia coli (EAEC): a cause of acute
and persistent diarrhea of worldwide importance. J Infect Dis. 2010;202(4):503–5.
49. Weintraub A. Enteroaggregative Escherichia coli: epidemiology, virulence and
detection. J Med Microbiol. 2007 Jan 1;56(1):4–8.
50. Nataro J. Enteroaggregative Escherichia coli pathogenesis. Curr Opin Gastroenterol.
2005 Jan;21(1):4–8.
51. Harrington S, Dudley E, Nataro J. Pathogenesis of enteroaggregative Escherichia coli
infection. FEMS Microbiol Lett. 2006;254(1):12–8.
52. Okeke I, Wallace-Gadsden F, Simons H, Matthews N, Labar A, Hwang J, Wain J. Multi-
locus sequence typing of enteroaggregative Escherichia coli isolates from nigerian
children uncovers multiple lineages. PLoS One. 2010;5(11):1–12.
53. Villaseca J, Hernández U, Sainz-Espuñes T, Rosario C, Eslava C. Enteroaggregative
Escherichia coli an emergent pathogen with different virulence properties. Rev Latinoam
Microbiol. 2005;47(3–4):140–59.
54. Jenkins C, Chart H, Willshaw G, Cheasty T, Smith H. Genotyping of enteroaggregative
Escherichia coli and identification of target genes for the detection of both typical and
atypical strains. Diagn Microbiol Infect Dis. 2006 May;55(1):13–9.
55. Cerna J, Nataro J, Estrada-Garcia T. Multiplex PCR for detection of three plasmid-borne
genes of enteroaggregative Escherichia coli strains. J Clin Microbiol. 2003;41(5):2138–
40.
56. Monteiro B, Campos L, Sircili M, Franzolin M, Bevilacqua L, Nataro J, Elias W. The
dispersin-encoding gene (aap) is not restricted to enteroaggregative Escherichia coli.
Diagn Microbiol Infect Dis. 2009;65(1):81–4.
57. Andrade F, Gomes T, Elias W. A sensitive and specific molecular tool for detection of
both typical and atypical enteroaggregative Escherichia coli. J Microbiol Methods.
2014;106:16–8.
58. Estrada-Garcia T, Navarro-Garcia F. Enteroaggregative Escherichia coli pathotype: a
genetically heterogeneous emerging foodborne enteropathogen. FEMS Immunol Med
Microbiol. 2012 Dec;66(3):281–98.
59. Shah N, DuPont H, Ramsey D. Global etiology of travelers’ diarrhea: systematic review
from 1973 to the present. Am J Trop Med Hyg. 2009;80(4):609–14.
60. Rüttler M, Yanzón C, Cuitiño M, Renna N, Pizarro M, Ortiz A. Evaluation of a
multiplex PCR method to detect enteroaggregative Escherichia coli. Biocell. 2006
Aug;30(2):301–8.
61. Muniesa M, Hammerl J, Hertwig S, Appel B, Brüssow H. Shiga toxin-producing

73
Escherichia coli O104:H4: A new challenge for microbiology. Appl Environ Microbiol.
2012;78(12):4065–73.
62. Esquivel P, Lifschitz V, Lösch L, Medina M, Pato A, Cacciamani A, Merino L.
Caracterización molecular de aislamientos de Escherichia coli productores de diarrea en
niños y adultos de la ciudad de Corrientes, Argentina. Rev Panam Infectología.
2010;12(3):17–21.
63. Pruden A, Pei R, Storteboom H, Carlson K. Antibiotic resistance genes as emerging
contaminants: studies in Northern Colorado. Environ Sci Technol. 2006;40(23):7445–50.
64. Stange C, Sidhu J, Tiehm A, Toze S. Antibiotic resistance and virulence genes in
coliform water isolates. Int J Hyg Environ Health. 2016;219(8):823–31.
65. Ram S, Vajpayee P, Tripathi U, Singh R, Seth P, Shanker R. Determination of
antimicrobial resistance and virulence gene signatures in surface water isolates of
Escherichia coli. J Appl Microbiol. 2008;105(6):1899–908.
66. Richardson H, Nichols G, Lane C, Lake I, Hunter P. Microbiological surveillance of
private water supplies in England - The impact of environmental and climate factors on
water quality. Water Res. 2009;43(8):2159–68.
67. Guzman-Herrador B, Carlander A, Ethelberg S, De Blasio B, Kuusi M, Lund V, Löfdahl
M, MacDonald E, Nichols G, Schönning C, Sudre B, Trönnberg L, Vold L, Semenza J,
Nygård K. Waterborne outbreaks in the Nordic countries, 1998 to 2012.
Eurosurveillance. 2015;20(24):1–10.
68. Ashbolt N. Microbial contamination of drinking water and human health from
community water systems. Curr Environ Heal Reports. 2015;2(1):95–106.
69. Yatsuyanagi J, Saito S, Miyajima Y, Amano K, Enomoto K. Characterization of atypical
enteropathogenic Escherichia coli strains harboring the astA gene that were associated
with a waterborne outbreak of diarrhea in Japan. J Clin Microbiol. 2003;41(5):2033–9.
70. Gupta S, Keck J, Ram P, Crump J, Miller M, Mintz E. Analysis of data gaps pertaining
to enterotoxigenic Escherichia coli infections in low and medium human development
index countries, 1984–2005. Epidemiol Infect. 2008;136:721–38.
71. Qadri F, Khan A, Faruque A, Begum Y, Chowdhury F, Nair G, Salam M, Sack D,
Svennerholm A. Enterotoxigenic Escherichia coli and Vibrio cholerae diarrhea,
Bangladesh, 2004. Emerg Infect Dis. 2005;11(7):1104–7.
72. Itoh Y, Nagano I, Kunishima M, Ezaki T. Laboratory investigation of enteroaggregative
Escherichia coli O Untypeable: H10 associated with a massive outbreak of
gastrointestinal illness. J Clin Microbiol. 1997;35(10):2546–50.
73. Falcão J, Falcão D, Gomes T. Ice as a vehicle for diarrheagenic Escherichia coli. Int J
Food Microbiol. 2004;91(1):99–103.
74. Pai M, Kang G, Ramakrishna B, Venkataraman A, Muliyil J. An epidemic of diarrhoea
in south India caused by enteroaggregative Escherichia coli. Indian J Med Res.
1997;106:7–12.
75. Tanaro J, Piaggio M, Galli L, Gasparovic A, Procura F, Molina D, Vitón M, Zolezzi G,
Rivas M. Prevalence of Escherichia coli O157:H7 in surface water near cattle feedlots.
Foodborne Pathog Dis. 2014;11(12):960–5.
76. Marucci P, Olivera N, Brugnoni L, Sica M, Cubitto M. The occurrence of Shiga toxin-

74
producing Escherichia coli in bathing water of the Sierra de la Ventana region, Buenos
Aires Province, Argentina. Environ Monit Assess. 2011;175:1–8.
77. Avery L, Williams A, Killham K, Jones D. Survival of Escherichia coli O157:H7 in
waters from lakes, rivers, puddles and animal-drinking troughs. Sci Total Environ.
2008;389(2–3):378–85.
78. Saxena T, Kaushik P, Krishna Mohan M. Prevalence of E. coli O157: H7 in water
sources: An overview on associated diseases, outbreaks and detection methods. Diagn
Microbiol Infect Dis. 2015;82(3):249–64.
79. Hunter P. Drinking water and diarrhoeal disease due to Escherichia coli. J Water Health.
2003;2(1):65–72.
80. Jofre J, Blanch A. Feasibility of methods based on nucleic acid amplification techniques
to fulfil the requirements for microbiological analysis of water quality. J Appl Microbiol.
2010;109:1853–67.
81. APHA AWWA WEF. Standart Methods for the Examination of Water and Wastewater.
20a. Eaton A, Clesceri L, Rice E, Greenberg A, editors. Washington D.C.: American
Public Health Association; 2005. 2671 p.
82. Toma C, Lu Y, Higa N, Nakasone N, Chinen I, Baschkier A, Rivas M, Iwanaga M.
Multiplex PCR assay for identification of human diarrheagenic Escherichia coli. J Clin
Microbiol. 2003;41(6):2669–71.
83. Botkin D, Galli L, Sankarapani V, Soler M, Rivas M, Torres A. Development of a
multiplex PCR assay for detection of Shiga toxin-producing Escherichia coli,
enterohemorrhagic E. coli, and enteropathogenic E. coli strains. Front Cell Infect
Microbiol. 2012;2(February):1–10.
84. Aranda K, Fabbricotti S, Fagundes-Neto U, Scaletsky I. Single multiplex assay to
identify simultaneously enteropathogenic, enteroaggregative, enterotoxigenic,
enteroinvasive and Shiga toxin-producing Escherichia coli strains in Brazilian children.
FEMS Microbiol Lett. 2007;267(2):145–50.
85. Gunzburg S, Tornieporth N, Riley L. Identification of enteropathogenic Escherichia coli
by PCR-based detection of the bundle-forming pilus gene. J Clin Microbiol.
1995;33(5):1375–7.
86. Bianucci S, Clothos S. Influencia de la contaminación de lagunas en la salud pública. En:
III Congreso De Ingeniería Civil, Territorio Y Medio Ambiente [Internet]. Zaragoza,
España: Colegio de Ingenieros de Caminos, Canales y Puertos; 2006. p. 1–16.
Disponible en: http://www.ciccp.es/biblio_digital/Icitema_III/congreso/pdf/010309.pdf
87. Hervot E, Prieto L, Farías A, Roshdestwensky S, Utgés E, Tenev M, väzquez F.
Comparación de los Indices de Calidad de Agua de las Lagunas Argüello y Los Lirios –
Resistencia – Chaco. En: Sosa C, Dalfaro N, editors. Investigación y Desarrollo en la
Facultad Regional Resistencia [Internet]. Resistencia, Argentina; 2011. p. 1–10.
Disponible en:
http://www.edutecne.utn.edu.ar/investigacion_fr_res/Farias_Ambiente.pdf
88. Bianucci S, Clemente M, Depettris C. Degradación ambiental de las lagunas ubicadas en
áreas urbanas. En: Universidad Nacional del Nordeste, editor. Comunicaciones
Científicas y Tecnológicas de la Universidad Nacional del Nordeste [Internet].
Resistencia, Argentina; 2004. p. 4. Disponible en:

75
http://www.unne.edu.ar/unnevieja/Web/cyt/com2004/7-Tecnologia/T-029.pdf
89. Rebello-Costa De Luca R, Regua-Mangia AH. Potential enterovirulence and
antimicrobial resistance in Escherichia coli isolates from aquatic environments in Rio de
Janeiro, Brazil. Sci Total Environ. 2014;490:19–27.
90. Sidhu J, Ahmed W, Hodgers L, Toze S. Occurrence of virulence genes associated with
diarrheagenic pathotypes in Escherichia coli isolates from surface water. Appl Environ
Microbiol. 2013 Jan;79(1):328–35.
91. Huang W, Hsu B, Kao P, Tao C, Ho Y, Kuo C, Huang Y. Seasonal distribution and
prevalence of diarrheagenic Escherichia coli in different aquatic environments in
Taiwan. Ecotoxicol Environ Saf. 2016;124:37–41.
92. Chern E, Tsai Y, Olson B. Occurrence of genes associated with enterotoxigenic and
enterohemorrhagic Escherichia coli in agricultural waste lagoons. Appl Environ
Microbiol. 2004;70(1):356–62.
93. Masters N, Wiegand A, Ahmed W, Katouli M. Escherichia coli virulence genes profile
of surface waters as an indicator of water quality. Water Res. 2011;45(19):6321–33.
94. Ranjbar R, Hosseini S, Zahraei-Salehi T, Kheiri R, Khamesipour F. Investigation on
prevalence of Escherichia coli strains carrying virulence genes ipaH, estA, eaeA and
bfpA isolated from different water sources. Asian Pacific J Trop Dis. 2016;6(4):278–83.
95. Titilawo Y, Obi L, Okoh A. Occurrence of virulence gene signatures associated with
diarrhoeagenic and non-diarrhoeagenic pathovars of Escherichia coli isolates from some
selected rivers in South-Western Nigeria. BMC Microbiol. 2015;15(204):1–14.
96. Akter S, Islam M, Afreen K, Azmuda N, Khan S, Birkeland N. Prevalence and
distribution of different diarrhoeagenic Escherichia coli virulotypes in major water
bodies in Bangladesh. Epidemiol Infect. 2013;141(12):2516–25.
97. Lauber C, Glatzer L, Sinsabaugh R. Prevalence of pathogenic Escherichia coli in
recreational waters. J Great Lakes Res. 2003;29(2):301–6.
98. Orsi R, Stoppe N, Sato M, Gomes T, Prado P, Manfio G, Ottoboni L. Genetic variability
and pathogenicity potential of Escherichia coli isolated from recreational water
reservoirs. Res Microbiol. 2007 Jun;158(5):420–7.
99. Gomi R, Matsuda T, Fujimori Y, Harada H, Matsui Y, Yoneda M. Characterization of
pathogenic Escherichia coli in river water by simultaneous detection and sequencing of
14 virulence genes. Environ Sci Technol. 2015;49(11):6800–17.
100. Rodrigues V, Rivera I, Lim K, Jiang S. Detection and risk assessment of diarrheagenic
E. coli in recreational beaches of Brazil. Mar Pollut Bull. 2016;109(1):163–70.
101. Hamilton M, Hadi A, Griffith J, Ishii S, Sadowsky M. Large scale analysis of virulence
genes in Escherichia coli strains isolated from Avalon Bay, CA. Water Res.
2010;44(18):5463–73.
102. Poma V, Mamani N, Iñiguez V. Impact of urban contamination of the La Paz River
basin on thermotolerant coliform density and occurrence of multiple antibiotic resistant
enteric pathogens in river water, irrigated soil and fresh vegetables. Springerplus.
2016;5(1):499.
103. Chen B, Zheng W, Yu Y, Huang W, Zheng S, Zhang Y, Guan X, Zhuang Y, Chen N,
Topp E. Class 1 integrons, selected virulence genes, and antibiotic resistance in

76
Escherichia coli isolates from the Minjiang River, Fujian Province, China. Appl Environ
Microbiol. 2011;77(1):148–55.
104. Lothigius A, Sjöling A, Svennerholm A, Bölin I. Survival and gene expression of
enterotoxigenic Escherichia coli during long-term incubation in sea water and
freshwater. J Appl Microbiol. 2010;108(4):1441–9.
105. Carlos C, Alexandrino F, Vieira M, Stoppe N, Sato M, Gomes T, Ottoboni L. Prevalence
of virulence factors in Escherichia coli isolated from healthy animals and water sources
in Brazil. J Water Health. 2011;9(1):138–42.
106. Rangel J, Sparling P, Crowe C, Griffin P, Swerdlow D. Epidemiology of Escherichia
coli O157:H7 outbreaks, United States, 1982-2002. Emerg Infect Dis. 2005;11(4):603–9.
107. Ferguson A, Mailloux B, Ahmed K, van Geen A, McKay L, Culligan P. Hand-pumps as
reservoirs for microbial contamination of well water. J Water Health. 2011;9(4):708–17.
108. Momba M, Malakate V, Theron J. Abundance of pathogenic Escherichia coli,
Salmonella typhimurium and Vibrio cholerae in Nkonkobe drinking water sources. J
Water Health. 2006;4(3):289–96.
109. Wallender E, Ailes E, Yoder J, Roberts V, Brunkard J. Contributing factors to disease
outbreaks associated with untreated groundwater. Groundwater. 2014;52(6):886–97.
110. Batabyal P, Mookerjee S, Sur D, Palit A. Diarrheogenic Escherechia coli in potable
water sources of West Bengal, India. Acta Trop. 2013;127(3):153–7.
111. Brown J, Proum S, Sobsey M. Escherichia coli in household drinking water and
diarrheal disease risk: evidence from Cambodia. Water Sci Technol. 2008;58(4):757–63.
112. Gambero M, Blarasin M, Bettera S, Albo G. Cuadernos de uso y manejo de aguas
subterráneas. Evaluación de la calidad del agua subterránea mediante la caracterización
fenotípica y genotípica de bacterias Escherichia coli aisladas. Primera. UniRío-
Universidad de Río Cuarto, editor. Río Cuarto, Córdoba; 2014. 15 p.
113. Juárez M, Poma H, Rajal V. ¿Cumplir con la legislación nos garantiza consumir agua
segura? RIBAGUA - Rev Iberoam del Agua. 2015;2(2):71–9.
114. Payment P, Locas A. Pathogens in water: value and limits of correlation with microbial
indicators. Ground Water. 2011;49(1):4–11.
115. John D, Rose J. Review of factors affecting microbial survival in groundwater. Environ
Sci Technol. 2005;39(19):7345–56.
116. Dobrowsky P, van Deventer A, De Kwaadsteniet M, Ndlovu T, Khan S, Cloete T, Khan
W. Prevalence of virulence genes associated with pathogenic Escherichia coli strains
isolated from domestically harvested rainwater during low- and high-rainfall periods.
Appl Environ Microbiol. 2014;80(5):1633–8.
117. Ahmed W, Hodgers L, Masters N, Sidhu J, Katouli M, Toze S. Occurrence of intestinal
and extraintestinal virulence genes in Escherichia coli isolates from rainwater tanks in
Southeast Queensland, Australia. Appl Environ Microbiol. 2011;77(20):7394–400.
118. Clasen T, Bastable A. Faecal contamination of drinking water during collection and
household storage: the need to extend protection to the point of use. J Water Health.
2003;1(3):109–15.
119. Medina M, Esquivel P, Lifschitz V, Medina M, Lösch L, Merino L. Detección de

77
Escherichia coli diarreogénicos en niños de barrios humildes de Corrientes, Argentina.
Rev Cubana Med Trop. 2010;2(1):42–7.
120. Huang D, Mohanty A, DuPont H, Okhuysen P, Chiang T. A review of an emerging
enteric pathogen: enteroaggregative Escherichia coli. J Med Microbiol.
2006;55(10):1303–11.

78
ANEXO I
Origen geográfico de las muestras superficiales y frecuencia de muestreo
Departamento Fuente superficial Frecuencia (N°)
San Fernando
Laguna Argüello 20
Laguna Ávalos 2
Laguna Colussi 3
Laguna Francia 2
Laguna Los Lirios 2
Laguna Los Teros 1
Laguna Prosperidad 2
Río Negro 26
Riacho Barranqueras 9
Río Paraná 10
Bermejo Laguna Moncholo 24
Río Paraguay 21
Libertad Laguna Beligoy 1
General Güemes Laguna La Verde 1
Laguna Aranda 1
Launa El Sauzal 2
Río Bermejito 3
Río Guaycurú 1
Río Teuco 2
Arroyo Malá 1
Represa –Miraflores 1
1º de Mayo Río Tragadero 2
General Güemes-
Libertador Gral. San
Martín- Bermejo
Río Bermejo* 7
Libertador Gral. San
Martín
Arroyo Correntoso 1
Represa –Presidencia Roca 2
9 de Julio Represa –Las Breñas 3
* Las muestras del Río Bermejo se tomaron en diferentes puntos de su curso, por tal
motivo se citan los tres Departamentos

79
ANEXO II
Origen geográfico de las muestras subterráneas y frecuencia de muestreo
Departamento Localidades Frecuencia
(Nº)
1 de Mayo Colonia Benítez, Margarita Belén, Tres Horquetas 8
12 de octubre General Pinedo, Mesón de Fierro, Pampa Landriel 5
25 de Mayo Machagai 4
9 de Julio Las Breñas 16
Almirante Brown Concepción del Bermejo, Los Frentones, Pampa del
Infierno, Río Muerto
11
Bermejo General Vedia, La Leonesa, Las Palmas 5
Chacabuco Charata, Tres Estacas 9
Comandante
Fernández
Pcia. R. Sáenz Peña 19
Donovan La Escondida, Makallé 6
Fontana Coronel Du Graty, Villa Ángela 6
Fray Justo Santa
Ma de Oro
Chorotis, Tres Mojones 2
General Güemes Comandancia Frías, El Espinillo, El Simbolar, El
Sauzalito, Fortín Belgrano, Fortín Lavalle, Fuerte
Esperanza, J.J.Castelli, Pje. El Asustado, Víbora
Blanca, Tartagal, Tres Pozos
29
Gral. Belgrano Corzuela 1
Independencia AviaTerai – Campo Largo 3
Libertad Puerto Tirol- Puerto Bastiani 13
Libertador San
Martin
General San Martín, Pampa Almirón 7
Maipú Tres Isletas, Villa Rural Carlos Palacios 10
O´Higgins San Bernardo 3
Pcia de la Plaza Presidencia de La Plaza 4
Quitilipi Pje. La Matanza- Quitilipi- Colonia Saavedra 3
San Fernando Basail, Colonia Baranda, Colonia Tacuarí, Fontana,
Puerto Vilelas, Resistencia
20
San Lorenzo Villa Berthet 3

80
ANEXO III
Ubicación geográfica de los puntos de muestreo de las muestras subterráneas. Fuente: mapoteca.edu.ar
81
ANEXO IV
Origen geográfico y frecuencia de muestreo de las muestras de aljibes, red y
bajadas de tanque
Muestras de red y bajadas de tanque
Departamento Localidades Frecuencia de muestreo Nº
Bermejo General Vedia 4
General Güemes Miraflores, Tartagal, Villa Río Bermejito 12
San Fernando Barrio Pedro Pescador 1
Muestras de aljibes
Departamento Localidades Frecuencia de muestreo Nº
12 de octubre General Pinedo 2
Almirante Brown Pampa del Infierno 1
Independencia Campo Largo 2
San Fernando Resistencia 1
San Lorenzo Villa Berthet 2

82
ANEXO V
Producción científica en el marco del Doctorado
Publicaciones científicas:
- Detección de genes de virulencia del patotipo enteroagregativo en cepas de Escherichia
coli aisladas en fuentes de agua subterránea de la Provincia del Chaco, Argentina. Liliana
S. Lösch, María L. Gariboglio Vázquez, Marta Rivas, Luis A. Merino. Revista Argentina de
Microbiología. 2015; 47(2)88-94.
- Detección de cepas diarreogénicas de Escherichia coli en fuentes de agua de la
Provincia del Chaco mediante la Reacción en Cadena de la Polimerasa”. Liliana Silvina
Lösch, Luis Antonio Merino. Anales de la Fundación Alberto J. Roemmers . Volumen XXII.
2012; 223-232
Publicaciones de divulgación:
- Microbiología de aguas: nuevos desafíos. Liliana Silvina Lösch, Luis Antonio Merino.
Boletín Electrónico de la Asociación Argentina de Microbiología. Boletín Nº198 Octubre-
Diciembre 2012
Capítulo de libro:
- Detección de genes de virulencia en aislamientos de Escherichia coli. Lösch, Liliana
Silvina. Merino, Luis Antonio. Capítulo 81. Manual de Métodos Moleculares para Estudios
Microbiológicos. Primera Edición. Asociación Argentina de Microbiología. 2011. ISBN 978-
987-26716-0-0.
Presentaciones a eventos científicos:
- Distribución de Escherichia coli diarreigénico en fuentes de aguas superficiales del
Chaco, Argentina. L Merino, L Gariboglio Vázquez, M Rivas, L Lösch. XXIII Congreso
Latinoamericano de Microbiología- XIV Congreso Argentino de Microbiología. 26-30 de
septiembre de 2016. Rosario, Santa Fe. Argentina.
- Detección de Escherichia coli enteroinvasivo en cuerpos de agua superficiales de la
provincia del Chaco, Argentina. Lösch, LS.; Gariboglio Vázquez, ML.; Rivas M.; Merino LA.
III Congreso Bioquímico del Litoral. XVI Jornadas Argentinas de Microbiología. 5-7 agosto de
2015. Santa Fe. Argentina.
- Detección de genes de virulencia de Escherichia coli enteroagregativa en fuentes de
agua subterráneas del oeste de la provincia del Chaco, Argentina. Lösch, LS.; Tracogna,
MF; Gariboglio Vázquez, ML; Merino, LA. XV Jornadas Argentinas de Microbiología. 14-16
agosto de 2014. Córdoba capital. Argentina.
- Detección de de genes de virulencia de Escherichia coli enteroagregativa en fuentes de
agua superficiales de la provincia del Chaco, Argentina. LS Lösch, MF Tracogna, ML
Gariboglio Vázquez, LA Merino. XIII Congreso Argentino de Microbiología 2013. II Congreso

83
de Microbiología Agrícola y Ambiental. 23 al 26 de septiembre de 2013. Buenos Aires,
Argentina
- Detección de Escherichia coli y de sus genes de virulencia en depósitos domiciliarios de
agua en la provincia del Chaco, Argentina. LS Lösch, MF Tracogna, ML Gariboglio
Vázquez, M Rivas, LA Merino. XI Congreso latinoamericano de Microbiología e Higiene de los
Alimentos - MICROAL 2012 Buenos Aires, Argentina.
- Detección de cepas diarreogénicas de Escherichia coli en fuentes de agua subterráneas
de la Provincia del Chaco, Argentina. Lösch LS, Tracogna MF, Gariboglio ML y Merino
LA. XIV Jornadas Argentinas de Microbiología. III Jornadas de Microbiología e Infectología
del NEA. 29 de septiembre al 1 de octubre de 2011. Resistencia, Chaco. Argentina.
Recommended

















