TECHNISCHE UNIVERSITÄT MÜNCHEN
II. Medizinische Klinik Klinikum rechts der Isar
(Direktor: Univ.-Prof. Dr. R.M. Schmid)
Pankreas-spezifische Deletion von IκBα -
Implikationen für Pankreasentwicklung und Ausprägung des Schweregrades einer akuten experimentellen
Pankreatitis
Christiane Schwerdtfeger
Vollständiger Abdruck der von der Fakultät für Medizin der Technischen Universität München zur Erlangung des akademischen Grades eines
Doktors der Medizin genehmigten Dissertation. Vorsitzender: Univ.-Prof. Dr. E. J. Rummeny Prüfer der Dissertation: 1. Priv.-Doz. Dr. H. Algül 2. Univ.-Prof. Dr. R. M. Schmid 3. Univ.-Prof. Dr. J. H. Kleeff Die Dissertation wurde am 13.08.2012 bei der Technischen Universität München eingereicht und durch die Fakultät für Medizin am 18.09.2013 angenommen

Inhaltsverzeichnis
II
Pankreas-spezifische Deletion von IκBα Implikationen für Pankreasentwicklung und
Ausprägung des Schweregrades einer akuten experimentellen Pankreatitis
Inhaltsverzeichnis
I. Einleitung ............................................................................................ 1
1 Das Pankreas ......................................................................................................... 1 2 Die akute Pankreatitis im Menschen ...................................................................... 3 3 Modelle der akuten experimentellen Pankreatitis .................................................. 5 4 Pathophysiologie der akuten experimentellen Pankreatitis .................................... 5
4.1 Sekretionsblockade ................................................................................... 6
4.2 Intrazelluläre Trypsinaktivierung ................................................................ 6
4.3 Entzündungsmediatoren ............................................................................ 6
4.4 Nekrose und Apoptose .............................................................................. 7
5 Der Ikk/NF-κB/RelA-Signalweg .............................................................................. 8
5.1 NF-κB ........................................................................................................ 8
5.2 IκB ............................................................................................................. 8
5.3 IKK ........................................................................................................... 10
5.4 Der IKK/NF-kB/RelA–Signalweg .............................................................. 10
6 NF-κB und die akute Pankreatitis ........................................................................ 12 7 Ziel dieser Arbeit .................................................................................................. 13
II. Methoden .......................................................................................... 14
1 Tierhaltung ........................................................................................................... 14 2 Induktion einer akuten Caerulein-Pankreatitis ..................................................... 14 3 Induktion einer L-Arginin Pankreatitis .................................................................. 14 4 Präparation und Materialgewinnung .................................................................... 15 5 Zellkultur ............................................................................................................... 15
5.1 Azini-Isolierung ........................................................................................ 15
6 Genotypisierung ................................................................................................... 16 6.1 DNA-Extraktion ........................................................................................ 16
6.2 Poymerase-Chain-Reaktion (PCR) .......................................................... 17
6.3 DNA Detektion ......................................................................................... 17

Inhaltsverzeichnis
III
7 Genexpressionsanalysen ..................................................................................... 18 7.1 RNA-Isolation ........................................................................................... 18
7.2 Affymetrix Mikroarry ................................................................................. 18
7.3 GSEA ....................................................................................................... 19
8 Western Blot ......................................................................................................... 19 8.1 Proteinlyse und Proteinbestimmung ........................................................ 19
8.2 Gelelektrophoretische Auftrennung der Proteine ..................................... 20
8.3 Membrantransfer ..................................................................................... 20
8.4 Proteindetektion ....................................................................................... 20
8.5 Membranreaktivierung ............................................................................. 22
9 Morphologie ......................................................................................................... 22 9.1 Anfertigung von Paraffinschnitten ............................................................ 22
9.2 Anfertigung von Cryoschnitten ................................................................. 22
9.3 Hämatoxylin-Eosin-Färbung .................................................................... 22
9.4 Histoscore ................................................................................................ 23
9.5 Immunhistochemie. .................................................................................. 23
9.6 Immunfluoreszenz ................................................................................... 24
10 Enzymbiochemie ................................................................................................ 26 10.1 MPO-Aktivitätsmessung in Lungengewebe ............................................. 26
10.2 Trypsin–Aktivitätsmessung im Pankreas ................................................. 26
11 Serologie ............................................................................................................ 27 11.1 Amylase- und Lipase-Messung ............................................................... 27
11.2 IL6-Elisa ................................................................................................... 27
12 Statistische Auswertung ..................................................................................... 27
III. Material ............................................................................................. 28
1 Geräte .................................................................................................................. 28 2 Puffer und Medien ................................................................................................ 29
2.1 Reagenzien .............................................................................................. 29
2.2 Puffer ....................................................................................................... 30
2.3 Kits ........................................................................................................... 31
IV. Ergebnisse ........................................................................................ 32

Inhaltsverzeichnis
IV
1 Die Pankreas-spezifische Inaktivierung von ikba führt zu einer nukleären Translokation von NF-κB ..................................................................................... 32
1.1 Generierung der ikbaΔpanc
-Maus mittels des Cre/loxP-Systems ............. 32
1.2 Die pankreas-spezifische Deletion von IκBα führt zur nukleären
Translokation und transkriptionellen Aktivität von NF-κB ................................... 35
2 IκBα-defiziente Mäuse entwickeln ein unauffälliges Pankreas ............................ 37 3 Die Inhibition von IκBα mildert den Verlauf der akuten experimentellen
Pankreatitis .......................................................................................................... 38 3.1 Die Caerulein Pankreatitis ....................................................................... 39
3.2 Die L-Arginin-Pankreatitis ........................................................................ 41
4 Die Inhibition von IκBα reduziert die intrapankreatische Trypsinaktivierung während der Pankreatitis ..................................................................................... 42
5 Die Pankreatitis-assoziierten Komplikationen werden durch die Inhibition von IκBα attenuiert .............................................................................................................. 43
5.1 IL-6 – ein serologischer Marker für die schwere AEP .............................. 43
5.2 Systemische Komplikation der AEP am Beispiel der Lunge .................... 44
6 Der protektive Effekt in ikba-defizienten Mäusen ist abhängig von nukleärem RelA 46
V. Diskussion ......................................................................................... 50
VI. Zusammenfassung ............................................................................ 57
VII. Danksagung .................................................................................. 59
VIII. Referenzen ................................................................................ 60
Fehler! Textmarke nicht definiert.

Abkürzungsverzeichnis
V
Abkürzungsverzeichnis
ACh Acetylcholin
AEP Akute experimentelle Pankreatitis
APS Ammonium Peroxodisulfat
ARDS acute respiratory distress syndrome
Bak BCL-2 homologous antagonist
Bax BCL-2 associated Protein
BCL-2 B-cell lymphoma 2
BCL-xl B-cell lymphoma xl
BSA Bovines Serum Albumin
CCK Cholezystokinin
CFTR Cystic Fibrosis Transmembran Conductance Regulator
CK 8 Cytokeratin 8
Cre Causes rekombination
DAB 3,3` Diaminobenzidin
DNA Deoxyribonucleic acid
ELISA Enzyme Linked Immunosorbet Essay
ERCP Endoskopische retrograde Cholangiopancreaticographie
GSEA Gen Set Enrichment Analysis
H&E Hemtoxylin & Eosin Färbung
H2O2 Wasserstoffperoxid
HCL Chlorwasserstoff
HTAB Hexadecyl Trimethyl Ammonium Bromid
ICAM Intercellular adhesion molecule 1
IHC Immunhistochemie
ikbaF/F ikba homozygot gefloxt
ikbaΔpanc ikba-Knockout
IKK IκB Kinase Komplex
IL-6/-10 Interleukin-6/-10
IP-Puffer Immunoprecipitation Puffer
IκBα Inhibitor of NF-κB
kDA kilo Dalton
KG Körpergewicht

Abkürzungsverzeichnis
VI
loxP locus X-over P 1
MCP-1 Monocyte chemotactic Protein-1
MPO Myeloperoxidase
mRNA messanger Ribonucleic acid
NaCL Natriumchlorid
NF-κB Nuclear factor κB
PAP-1 Pankreatitis assoziertes Protein-1
PBS Phosphat buffered saline (phosphat gepufferte Salzlösung)
PCR Polymerase chain reaction
PFA Paraformaldehyd
PKA /C Proteinkinase A /C
PTF1a Pankreas specific transcription factor 1a
RDH Rel homology domain
rela x ikbaΔpanc rela x ikba Doppelknockout
relaΔpanc rela Knockout
RNA Ribonucleic acid
SDS Sodium dodecyl sulfate
SPINK 1 serine protease inhibitor kazal 1
TAE Tris acetat EDTA
Taq Thermus aquaticus
TBS Tris buffered saline (Tris gepufferte Salzlösung)
TE Tris EDTA
TEMED Tetramethylethylendiamin
TNF α Tumor Nekrose Faktor α
Tris Tris(hydroxymethyl)-aminom

Einleitung I
1
I. Einleitung
Die akute Entzündung der Bauchspeicheldrüse ist eine ernst zunehmende
Erkrankung. Zwar läuft die Erkrankung in ca. 2/3 der Fälle als milde interstitielle
ödematöse Entzündung ab, für einen Teil der Patienten aber endet die akute
Pankreatitis tödlich. Auch berühmte Persönlichkeiten der Geschichte blieben von der
Erkrankung nicht verschont. Neuesten Erkenntnissen zufolge verstarb Alexander der
Große 323 v. Chr. wenige Tage nach einem Trinkgelage an einer akuten Pankreatitis
(Sbarounis, 1997). Es sollte aber noch über 2000 Jahre dauern bis die Erkrankung
erstmals klinisch beschrieben wurde und man die Symptomatik mit der
Bauchspeicheldrüse in Verbindung brachte. Nach der Erstbeschreibung 1889
entwickelte Hans Chiari 1896 das pathophysiologische Konzept der Autodigestion
mit der Azinuszelle als Ausgangspunkt der akuten Pankreatitis (Chiari, 1896.). Ein
Modell, das sich in zahlreichen Experimenten als richtig erwies. Wenige Jahre später
fand Eugene Opie 1901 heraus, dass Gallengangssteine eine der Hauptursachen
der akuten Pankreatitis sind (Lerch et al., 1994). Dennoch stehen nach über 100
Jahren Forschung für die Therapie der akuten Pankreatitis nur symptomatische
Maßnahmen zur Verfügung. Eine zielgerichtete Therapie mit signifikanten Erfolg
konnte bislang noch nicht entwickelt werden. Dies ist unteranderem dem Umstand
geschuldet, dass die Pathogenese der akuten Pankreatitis noch nicht ausreichend
verstanden ist. Aus diesem Grund beschäftigt sich diese Arbeit mit der
Pathophysiologie der akuten Pankreatitis und geht im Speziellen der Frage nach,
welche Bedeutung der IKK/NF-κB/RelA-Signalweg für den Ablauf der akuten
Pankreatitis hat.
1 Das Pankreas
Die Aufgabe des Pankreas besteht in der Regulation des Blutzuckers und
Bereitstellung von Verdauungsenzymen.
Makroskopisch betrachtet lässt sich das Pankreas in Caput, Corpus und Cauda
unterteilen. Es lokalisiert sich retroperitoneal auf Höhe des 1. Lendenwirbels. Längs

I Einleitung
2
durch die Drüse zieht der Hauptausführungsgang. Dieser mündet zusammen mit
dem Ductus choledochus in der Papilla duodeni major im Duodenum (Fritsch, 2003).
Die Langerhans-Inseln bilden das endokrine Kompartiment. Die B-Zellen machen
80% einer Insel aus und sind für die Insulinproduktion verantwortlich. Die A-Zellen
produzieren Glukagon und die D-Zellen Somatostatin.
Der Großteil der Drüse wird vom exokrinen Kompartiment gebildet. Die Grundeinheit
der exokrinen Drüse bildet der Azinus, das Drüsenendstück. Mehrere Azinuszellen
bilden einen Azinus. Diese produzieren die Verdauungsenzyme. Die Azini werden
zusammen mit ihren Ausführungsgängen zu Lobuli oder Läppchen zusammen
gefasst. Neben den Azini bilden die duktalen Zellen die Drüsengänge (Welsch,
2003).
Graphik 1: Histologischer Schnitt Pankreas (H&E Vergrößerung 100x) und schematische Darstellung eines Azinus zusammen mit einem intralobulären Ausführungsgang
Die Mehrheit der in den Azini synthetisierten Verdauungsenzyme gelangt als inaktive
Vorstufen (Zymogene) in das Duodenum. Die Enteropeptidase (Enterokinase), eine
membranständige Serinprotease der duodenalen Mucosa, aktiviert die Peptidase
Trypsin durch katalytische Abspaltung eines N-terminalen Restes TAP ((Asp)4-Lys-
Ile) aus dem Proenzym Trypsinogen (Bricteux-Gregoire et al., 1972). Die
Endopeptidase Trypsin hat als einzige Peptidase die Fähigkeit sich selbst zu
aktivieren und ist für die Aktivierung der anderen Peptidasen im Darmlumen
verantwortlich (Rinderknecht, 1986, Gorelick et al., 1992). Eine vorzeitige
enzymatische Spaltung von Trypsinogen zu aktiven Trypsin im Pankreas wird unter
physiologischen Bedingungen durch verschiedene Mechanismen verhindert. Unter
anderem besitzt Trypsin eine calcium-abhängige autolytische Aktivität. Unter
physiologischen Bedingungen liegt in der Azinuszelle ein niedriger Calciumgehalt vor
Intralobulärer GangAzinuszelle
Zentroazinäre Zelle
Insel
Azini
Gang

Einleitung I
3
und aufgrund dessen inaktiviert Trypsin sich kontinuierlich selbst (Whitcomb et al.,
1996). Des weiteren wird aktives Trypsin durch den intrazellulären Trypsin-Inhibitor
SPINK 1 temporär gehemmt (Bartelt et al., 1977).
Die Verdauungsenzyme werden in der Azinuszelle in den Zymogengranula,
gespeichert und an der apikalen Membran in den Pankreasgang sezerniert. Die
Exozytose der Zymogengranula wird in den Azinuszellen der Säugetiere über das
Hormon Cholezystokinin (CCK) aus dem Dünndarm vermittelt (Wasle and
Edwardson, 2002). An dieser Stelle muss auf die Besonderheit des humanen
Pankreas hingewiesen werden. Humane Azinuszellen besitzen kaum CCK-
Rezeptoren (Reubi et al., 2003). CCK wirkt in diesem Fall nicht direkt auf die
Azinuszellen, sondern CCK stimuliert cholinerge Neurone, die über ACh und
muskarinische Rezeptoren wirken (Owyang and Logsdon, 2004).
2 Die akute Pankreatitis im Menschen
In den westlichen Industrieländern stellt die akute Pankreatitis ein zunehmendes
Problem dar. So stieg die Inzidenz in Kalifornien zwischen 1994 und 2001 von 33 auf
44 pro 100000 Einwohner (Frey et al., 2006). Studien Westeuropäischer Länder
kamen zu ähnlichen Ergebnissen (O'Farrell et al., 2007, Lindkvist et al., 2004, Yadav
and Lowenfels, 2006). Als Grund für diesen Inzidenzanstieg wird in den meisten
Fällen, neben verbesserten diagnostischen Methoden, vermehrter Alkoholkonsum
verantwortlich gemacht (Yadav and Lowenfels, 2006).
Die Hauptursachen der akuten Pankreatitis bilden Gallengangssteine (akute biliäre
Pankreatitis) mit 38% und exzessiver Alkoholabusus mit 35% (Whitcomb, 2006). Die
Genese der akuten biliären Pankreatitis wird dadurch erklärt, dass es durch eine
Verlegung des Gallengangs zu einer Druckerhöhung im Pankreasgang kommt,
woraufhin Azinuszellen geschädigt werden (Diehl et al., 1997). Der Zusammenhang
zwischen Alkohol und Pankreatitis ist komplex und zum Großteil noch nicht geklärt.
Obwohl in der Rangfolge der Verursacher einer Pankreatitis von untergeordneter
Bedeutung, doch für das Verständnis der Pathogenese von großer Wichtigkeit sind
die hereditären chronischen Pankreatitis Formen. Eine Mutation im humanen
kationischen Trypsinogen (PRSS1-Gen) liegt in ca. 50% der Fälle vor und ist somit
der häufigste nachgewiesene Gendefekt. Der Erbgang ist autosomal dominant und
führt dazu das Trypsin im Pankreas spontan aktiviert wird und resistent gegen

I Einleitung
4
Autolyse ist (Whitcomb et al., 1996). Daneben konnten noch Mutationen im SPINK1-
Gen, das für den intra-azinärer Trypsininhibitor kodiert, sowie Mutationen im CFTR-
Gen, das für einen Chloridkanal der Zellmembran kodiert, als Verursacher einer
chronischen hereditären Pankreatitis nachgewiesen werden (Cohn et al., 1998). In
der klinischen Praxis ist weiterhin die post ERCP–Pankreatitis von Bedeutung.
Darüber hinaus gibt es noch eine Vielzahl an Ursachen wie z.B Traumata,
Hypercalciämie, Hypertriglyceridämie, primärer Hyperparathyreoidismus und
verschiedenste Medikamente (Lankisch et al., 1995).
Klinisch manifestiert sich die Erkrankung meist mit plötzlich einsetzenden
Schmerzen, die in den Rücken, den Thorax und gürtelförmig um das Abdomen
ausstrahlen können.
Als laborchemischer diagnostischer Parameter dient die Serum-Lipase, ein Anstieg
ist spezifisch für die akute Pankreatitis (Johnson, 2005). Die Höhe des Anstiegs
korreliert aber nicht mit der Schwere der Erkrankung. Serum-Amylase steigt zu
Beginn der Erkrankung rasch an (Frossard et al., 2008), da sie aber nicht pankreas-
spezifisch ist, kommt ihr in der klinischen Praxis keine Bedeutung zu.
In 80% der Fälle nimmt die Erkrankung einen milden harmlosen Verlauf, 20% der
Patienten aber erleben eine schwere nekrotisierende Pankreatitis, wobei 10 - 30%
dieser Patienten sterben (Whitcomb, 2006). Eine schwere akute Pankreatitis ist
charakterisiert durch hämorrhagische Nekrosen und extrapankreatische
Komplikationen. Gefürchtet ist vor allem eine bakterielle Infektion der Nekrosen, da
diese meist einen septischen Verlauf nehmen. Das extrapankreatische Organ das
am häufigsten betroffen ist, ist die Lunge. In bis zu 75% der Fälle werden bei
Patienten mit akuter Pankreatitis pulmonale Komplikationen festgestellt (Browne and
Pitchumoni, 2006, Raghu et al., 2007). Diese variieren von einer leichten Hypoxämie
über Atelektasen bis zum Vollbild des ARDS (Acute Respiratory Distress Syndrome),
wobei die Häufigkeit der Lungenschäden mit dem Ausmaß der Nekrose im Pankreas
positiv korrelieren (Raghu et al., 2007). Das ARDS entsteht meist zwischen dem 3.
und 7. Tag der Erkrankung und 50% der Patienten, die ein ARDS entwickeln,
versterben (Pastor et al., 2003).

Einleitung I
5
3 Modelle der akuten experimentellen Pankreatitis
Da die wissenschaftliche Untersuchung der akuten Pankreatitis im Menschen in der
klinischen Praxis aufgrund der anatomischen Lage und des raschen Verlaufs sich
äußerst schwierig gestaltet, ist es notwendig, auf Tiermodelle zurückzugreifen. Ein
etabliertes Model der akuten Pankreatitis ist die Stimulation des Pankreas mit
supramaximalen Konzentrationen CCK bzw. seinem Analogon Caerulein in
Nagetieren.
Caerulein ist ein synthetisches Decapeptid, das mit dem Enzym CCK in seinem C-
Terminus identisch ist und somit an den CCK-Rezeptor der Azinuszellen des
Pankreas binden kann. Stimulation mit supramaximalen Konzentration CCK, die
intraperitoneal appliziert werden, führen zur Entwicklung einer akuten
experimentellen Pankreatitis in Nagetieren (Lampel and Kern, 1977). Die Caerulein-
induzierte Pankreatitis verläuft im Gegensatz zu anderen Modellen relativ mild, in der
Regel versterben die Tiere nicht. Sie ist gekennzeichnet durch Ödembildung,
Infiltration von Entzündungszellen und erhöhten Serum-Lipase und -Amylase
Parametern. Die zellulären Veränderungen sind gekennzeichnet durch
Vakuolenbildung, Trypsinaktivierung und intrazelluläre Zymogenaktivierung (Leach et
al., 1991).
Ein weiteres Model der akuten experimentellen Pankreatitis beinhaltet die einmalige
intraperitoneale Injektion einer hohen Dosis L-Arginin, die eine akute nekrotisierende
Pankreatitis induziert (Mizunuma et al., 1984). Der genaue Pathomechanismus ist
bislang noch nicht bekannt, aber wahrscheinlich sind freie Sauerstoffradikale und
Stickstoffoxid involviert (Hegyi et al., 2004). Dieses Modell kann, je nach applizierter
Dosis für akute oder chronische Pankreatitis-Studien verwendet werden.
4 Pathophysiologie der akuten experimentellen Pankreatitis Schon früh zeigte sich, dass der Ausgangspunkt der akuten Pankreatitis in der
Azinuszelle selbst liegt. In einer Vielzahl von Forschungsarbeiten hat sich die Ko-
lokalisationstheorie als pathophysiologisches Modell bestätigt. Durch eine
Überstimulation kommt es zu einer Akkumulation der Zymogengranula in der Zelle,
Kolokalisation der Zymogene mit lysosomalen Hydrolasen und dadurch zur

I Einleitung
6
intrazellulären Trypsinaktivierung. Dies hat eine Selbstandauung des Pankreas zur
Folge und die Induktion von Entzündung und Nekrose. Im Folgenden soll auf die
einzelnen Phasen eingegangen werden.
4.1 Sekretionsblockade Die Beziehung zwischen der Stimulation der Azinuszelle mit CCK und der Sekretion
von Zymogenen folgt nicht einem linearen Verlauf, sondern verhält sich biphasisch
(Grady et al., 1998). Bis zu einer bestimmten maximalen Konzentration von CCK
kommt es zu einer kontinuierlichen Ausschüttung der Verdauungsenzyme. Wird
diese Konzentration jedoch überschritten, ist eine Sekretion nicht mehr möglich und
die Zymogengranula akkumulieren in der Zelle (Watanabe et al., 1984). Diesem
Phänomen liegt eine Schädigung des Aktinzytoskeletts im apikalen Bereich der Zelle
zugrunde, aufgrund dessen eine Exozytose nicht mehr möglich ist (Jungermann et
al., 1995).
4.2 Intrazelluläre Trypsinaktivierung Eines der frühesten Ereignisse im Verlauf der akuten experimentellen Pankreatitis ist
die intrazelluläre Trypsinaktivierung (Whitcomb, 1999). Voraussetzung hierfür ist die
pathologische Fusion der physiologisch getrennten Zellkompartimente der
Lysosomen und der Zymogengranula (Willemer et al., 1990, Saluja et al., 1987,
Watanabe et al., 1984). Dies ist noch vor der Trypsinogenaktivierung zu beobachten
(van Acker et al., 2006). Unter den lysosomalen Enzymen ist Cathepsin B von
Bedeutung, welches hauptsächlich zur Trypsinaktivierung beiträgt. Ohne das
Vorhandensein von Cathepsin B findet die intrazelluläre Trypsinaktivierung nach
Induktion einer akuten Pankreatitis nur im geringen Maße statt und die Erkrankung
nimmt einen wesentlich milderen Verlauf (Halangk et al., 2000).
Als Folge der Schädigung der Azinuszelle durch die aktiven Verdauungsenzyme
kommt es nun zur Induktion von Entzündungsmediatoren.
4.3 Entzündungsmediatoren Unterschiedliche Entzündungsmediatoren sind verantwortlich für die Infiltration von
Leukozyten und das Ausmaß der lokalen und systemischen Komplikationen.
TNF−α wird vor allem von Makrophagen und Monozyten ausgeschüttet, aber auch
Azinuszellen können TNF-α produzieren und besitzen selbst TNF-α –Rezeptoren.

Einleitung I
7
(Gukovskaya et al., 1997). Die Wirkung von TNF-α ist dosisabhängig. Niedrige
Konzentrationen induzieren Apoptose und wirken protektiv (Yasuda et al., 1999).
Höhere Konzentrationen führen zur unkontrollierten Nekrosebildung und
konsekutiven Organschädigung (Malka et al., 2000).
In enger Beziehung zu TNF-α steht IL-1β. Es wird ebenfalls hauptsächlich von
Makrophagen und Monozyten ausgeschüttet, bewirkt die Aktivierung von
neutrophilen Granulozyten und wirkt synergistisch mit TNF-α (Bhatia et al., 2000).
Von prognostischer Bedeutung ist IL-6. In klinischen Studien konnte gezeigt werden,
dass erhöhte IL-6 Level im Serum mit einer schweren nekrotisierenden Pankreatitis
korrelieren (Aoun et al., 2009) und auch experimentell lies sich dieser
Zusammenhang nachweisen (Leindler et al., 2004).
IL-10 wirkt anti-inflammatorisch. In zahlreichen experimentellen Studien konnte
gezeigt werden, dass IL-10 in der Frühphase erhöht ist und schwere Verläufe
verhindert (Rongione et al., 1997), klinisch lies sich diese Erkenntnis aber noch nicht
nutzen (Bang et al., 2008).
4.4 Nekrose und Apoptose Der Schweregrad einer akuten Pankreatitis charakterisiert sich vor allem über das
Ausmaß der Nekrose-Areale. Durch den ungeordneten Zelltod, der Nekrose, kommt
es zur massenhaften Freisetzung von intrazellulären Zellbestandteilen in das
Interstitium, die eine unkontrollierte Entzündungsreaktion induzieren und zu weiteren
Organschäden führen (Gukovskaya and Pandol, 2004).
Im Gegensatz zur Nekrose, kommt es bei der Apoptose zur kontrollierten Aktivierung
bestimmter Proteasen, den Caspasen, sowie zur Freisetzung von Cytochrom C.
Dadurch kommt es zum kontrollierten Zelltod.
Caspase 8 und 9 sind die Hauptintiatoren, Caspase 3, 6 und 7 die Haupteffektoren
der Apoptose (Gukovskaya and Pandol, 2004). Während der akuten Pankreatitis
kann, stimuliert durch CCK, die Expression der Caspasen -8, -9 und -3 beobachtet
werden und damit einhergehend eine erhöhte Apoptoserate und milder verlaufende
Pankreatitis (Gukovskaya et al., 2002).
In diese Regelkreise greifen nun eine Vielzahl von übergeordneten
Regulatorproteinen ein. Neutrophile Granulozyten, rekrutiert durch verschiedene
Cytokine, schädigen die Zellen und verursachen Nekrose während der Pankreatitis

I Einleitung
8
(Fujimoto et al., 1997) Neben TNF-a spielt der NF-κB Signalweg eine entscheidende
Rolle bei der akuten Pankreatitis (Kucharczak et al., 2003).
5 Der Ikk/NF-κB/RelA-Signalweg
1986 entdeckten R. Sen und D. Baltimore einen Transkriptionsfaktor, der an ein
Enhancer-Element des Immunglobin κ light-chain Gens in B-Lymphozyten bindet
(Ghosh et al., 1998, Sen and Baltimore, 1986). Sie bezeichneten den
Transkriptionsfaktor als Nuklear Faktor κB (NF-κB) Bald darauf stellte sich heraus,
dass es sich hierbei um eine ubiquitär exprimierte Gruppe von heterogenen Dimeren
handelt, die essentiell in Prozesse wie zellulären Stress, Apoptose und Entzündung
involviert und für die Expression einer Vielzahl von Proteinen verantwortlich sind.
5.1 NF-κB
Die NF-κB/Rel Familie besteht aus 5 Mitgliedern. Diese sind NF-κB1 (p50) NF-κB2
(p52), c-Rel, RelA (p65 oder NF-κB3) und RelB. p50 und p52 entstehen erst durch
die Abspaltung eines C-terminalen Rests aus den Vorläuferproteinen p105 und p100
(Karin and Ben-Neriah, 2000, Perkins, 2000). Allen NF-κB/Rel Proteinen gemeinsam
ist eine ca. 300 Aminosäuren große N-terminale Region, die sogenannte Rel
homology Domain (RHD). Sie ist verantwortlich für die DNA-Bindung, nukleäre
Translokation und Dimerisation. Außerdem ist sie die Bindestelle für die
Inhibitorproteine der IκB-Familie. Zwar können alle NF-κBs an DNA binden, aber nur
RelA, RelB und c-Rel besitzen transkriptionelle Aktivität (Hayden and Ghosh, 2008).
Das bei weiten am häufigsten vorkommende Dimer ist p50:p65, so dass dieses oft
als Synonym für NF-κB steht. p50 und p52 Homodimere binden zwar ebenfalls an
die NF-κB-Bindestelle der DNA, können aber keine Transkription initiieren.
5.2 IκB
Die Aktivität von NF-κB wird durch eine bestimmte Gruppe von Inhibitorproteinen
kontrolliert den IκBs (Inhibitiores of NF-κB) Zu diesen gehören u.a. IκBα, IκBβ, IκBε
und BCL-3 (Karin and Ben-Neriah, 2000) Allen gemeinsam ist ein bestimmtes Motiv
bestehend aus 6-7 Ankyrin-Elementen (Blank et al., 1991, Mercurio et al., 1993, Rice
et al., 1992). Mit diesem Motiv binden die IκBs an die RHD-Sequenz von NF-κB und
maskieren so das Kernlokalisationssignal. Die Vorläuferproteine p105 und p100

Einleitung I
9
besitzen an ihrem C-Terminus ebenfalls dieses Ankyrinmotiv (Malek et al., 2001,
Ghosh and Karin, 2002). Während die Umwandlung von p105 zu p50 post-
translationell erfolgt, ist die Abspaltung des C-Terminus von p100 von zentraler
Bedeutung im alternativen NF-κB–Aktivierungsweg, der aber bei der Pathogenese
der akuten Pankreatitis nur eine untergeordnete Rolle spielt.
Am besten charakterisiert ist IκBα. An ihm ist der schnelle reguläre Aktivierungsweg
für NF-κB genau charakterisiert (s.u). IκBα ist ein 37 kDA großes Protein und im
Cytosol vor allem an p50:p65 Dimere gebunden. IκBα nur an die RHD von p65
während die von p50 frei bleibt. So kommt es im inaktiven Zustand von NF-κB zu
einem kontinuierlichen Eintritt der p50:p65:IκBα-Komplexe in den Nukleus, diese
Komplexe werden aber aufgrund der Eigenschaften von IκBα sofort wieder aus dem
Nukleus entfernt (Hayden and Ghosh, 2008)
IκBβ, das neben IκBα am häufigsten vorkommt, unterscheidet sich von Iκbα durch
einen langsameren Abbau und durch eine NF-κB unabhängige Expression. (Hayden
and Ghosh, 2008). IκBε besitzt ähnliche Eigenschaften wie IκBα wird aber nur in
hämatopoetischen Zellen exprimiert (Perkins, 2006, Kuwata et al., 2003). BCl-3
nimmt unter den IκBs eine Sonderrolle ein, da es nicht im Cytosol sondern im
Nukleus lokalisiert ist. Seine Rolle ist noch nicht genau geklärt.
Graphik 2: Die Proteine der NF-κB/Rel und IκB-Familie, schematische Darstellung modifiziert nach Karin 2000 (Karin and Ben-Neriah, 2000) und Ghosh 1998 (Ghosh et al., 1998). Die Zahlen an der rechten Seite der Proteine entspricht der Anzahl der Aminosäuren. RHD: Rel Homology Domain; TAD: Transaktivation Domain P: Aminosäure an der Phosphorylierung erfolgt U Ubiquitinierung; LZ Leuzin Zipper Region
U PU P
317
Rela/p65 RHD TAD551
RelB RHDLZ TAD579
c-Rel RHD TAD619
p100/p52
p105/p50
IκBα
IκBβ
IκBγ
IκBε
Bcl-3PP
446
PPU361
P
607
PPU
356
RHD969
RHD
PPU899
NF-κB-Familie
IκB-Familie
Ankyrin

I Einleitung
10
5.3 IKK
Die Phosphorylierung von IκBα und damit die Initiation seines Abbaus erfolgt durch
eine spezielle Kinase, die IκB-Kinase (IKK) (Hayden and Ghosh, 2008). Diese
besteht aus 3 Untereinheiten wobei IKKα (o. IKK1) und IKKβ (o. IKK2) katalytische
Aktivität besitzen, hingegen IKKγ (o. Nemo) regulatorisch wirkt. IKKα wird für beide
Signalwege benötigt, IKKβ nur für den regulären (s.u.). IKKα spielt außerdem bei der
Regulation der Transkription durch NF-κB eine Rolle (Hayden and Ghosh, 2004).
5.4 Der IKK/NF-kB/RelA–Signalweg
Der reguläre IKK/NF−κB/RelA–Signalweg führt zur schnellen Freisetzung von
p50:p65 Dimeren durch die Degradation von IκBα. Klassische Induktoren dieses
Signalwegs sind TNF-α und IL-1β. Bislang konnten die genauen Signalwege, die zur
Aktivierung der IκB-Kinase führen, noch nicht vollständig identifiziert werden (Karin
and Ben-Neriah, 2000, Perkins, 2000). Der Aktivierungsmechanismus des IKK-
Komplexes ist sehr umfangreich und involviert Phosphorylierung,
Transautophosphorylierung und Ubiquitinierung der einzelnen Untereinheiten (Brown
et al., 1995, Li et al., 1999).
IKKβ phosphoryliert IκBα an zwei spezifischen Serinresten (Serin 32 und 36) (Baldi
et al., 1996) dadurch kann IκBα an zwei Lysinresten (Lysin 21und 22) ubiquitiniert
werden (Karin and Ben-Neriah, 2000, Ghosh et al., 1998). Dies ist das Signal für den
Abbau durch das 26S Proteasom. Erst durch den Abbau von IκBα ist NF-κB
vollständig frei und kann dauerhaft in den Nukleus eintreten (Hoffmann et al., 2006).
Im Nukleus binden NF-κB-Dimere an sogenannte DNA κB-Sequenzen, die in einer
Vielzahl von Promotern und Enhancern enthalten sind (Ghosh, 1999). Die einzelnen
Untereinheiten sind für die Expression ganz unterschiedlicher Gene verantwortlich,
können unterschiedliche Coaktivatoren rekrutieren und sind verschiedensten
Modifikationen unterworfen. Ein bekannter Modulator von p65 ist zum Beispiel die
Proteinkinase A (PKA). Durch die Phosphorylierung durch PKA ist p65 in der Lage im
Nukleus mit weiteren Coaktivatoren zu interagieren (Darieva et al., 2004).
Zu den Proteinen die durch p50:p65 exprimiert werden, gehören Cytokine z.B.TNF-α
und IL-1β, Akut-Phase-Proteine und Adhesionsmoleküle (Tando et al., 1999).
Weiterhin besteht zwischen NF-κB und IκBα ein negativer Feedback-Mechanismus.
Um nur eine vorübergehende Aktivierung von NF-κB zu gewährleisten, induziert NF-

Einleitung I
11
κB die Expression von Iκbα. Das neu synthetisierte IκBα tritt in den Nukleus ein, löst
NF-κB von der DNA und transportiert es zurück in das Cytosol (Perkins, 2000). So
terminiert NF-κB selbst seine Aktivität. Graphik 3 zeigt zusammengefasst die
wichtigsten Schritte des IKK/NF−κB/RelA–Signalwegs.
Graphik 3: Der IKK/NF-κB/RelA- Signalweg: Wird die IκBα-Kinase IKK durch verschiedene Induktoren wie z.B. TNF-α oder IL-1 aktiviert, phosphoryliert sie IκBα an spezifischen Serinresten. Dies ist das Signal für die Ubiqitinierung und damit für die Proteasomen abhängige Degradation von IκBα. Das p65:p50 Dimer ist nun frei, kann in den Nukleus eintreten und durch Bindung an κB-Sites in der DNA die Expression verschiedener Proteine wie z.B IκBα, TNF-α, IL-1, IL-6 und PAP-1 induzieren. Durch Modifikationen der p50:p65 Dimere kann das Genexpressionsmuster verändert werden. IKK Untereinheiten: Nemo:IKKα:IKKβ; NF-κB: p50:p65; Proteinkinase A: PKA ;
P
p65
α β Nemo
p50 p65
IκBα
P
IκBα P
PKA
TNF α IL-1
Ubiquitinierung
p50 p65
IκBα P P
p50 p65
IκBα P P
p50 p65
p50
Proteasom
u.a Iκbα , TNF-α , IL-1, IL-6, PAP1
Modifikation
Phosphorylierung
Nukleus
Cytosol IKK

I Einleitung
12
6 NF-κB und die akute Pankreatitis
In der Caerulein-induzierten Pankreatitis verhält sich die NF-κB Aktivierung
biphasisch. Nach Stimulierung des Pankreas mit supramaximalen Dosen Caerulein
kann bereits innerhalb von 10 min freies NF-κB im Nukleus der Azinuszelle detektiert
werden (Brockhaus et al.) und nach 30 min hat die DNA-Bindungsaktivität von NF-κB
sein Maximum erreicht. Nach 90 min ist NF-κB nicht mehr aktiv, gleichzeitig mit der
Resynthese von IκBα. Nach ca. 3 h kommt es zu einer zweiten Welle der NF-κB
Aktivierung (Rakonczay et al., 2008). Dies rührt vermutlich von der langsam
verlaufenden Degradation von IκBβ her (Gukovskaya et al., 1997). Aber auch TNF-α
freigesetzt aus den Azinuszellen und infiltrierenden Leukozyten kann für diese zweite
NF-κB Welle verantwortlich gemacht werden (Steinle et al., 1999). Von den NF-κB
Untereinheiten können p50, p52 und RelA/p65 in der Azinuszelle nachgewiesen
werden, c-Rel dagegen nicht (Tando et al., 1999). Die Induktion von NF-κB verläuft
über den CCKA-Rezeptor, Calcium/Calcineurin und PKC sind vermutlich als
Messenger involviert (Yuan et al., 2008). Gleichzeitig mit der Aktivierung von NF-κB
kann die Degradation von IκBα detektiert werden, nach 3-6h aber ist die
Konzentration von IκBα, induziert durch NF-κB wieder auf dem Ausgangsniveau
(Algul et al., 2002). NF-κB terminiert so selbst seine Aktivität.
Die Aktivierung von NF-κB führt zur Induktion von Zytokinen und
Entzündungsmediatoren u.a TNF-α und IL-1β, MCP-1, ICAM und das Pankreatitis
assoziierte Protein I (Gukovsky et al., 2003, Gukovsky et al., 1998, Dunn et al., 1997,
Satoh et al., 1999, Ethridge et al., 2002, Steinle et al., 1999). Aber nicht nur in
Azinuszellen, auch in eingewanderten neutrophilen Granulozyten, periacinären
Myofibroblasten, in den Kupfferzellen der Leber und in der Lunge ist die NF-κB
Aktivität während der Pankreatitis hochreguliert (Algul et al., 2002). In die Apoptose-
und Nekrose-Signalwege während der akuten Pankreatitis ist NF-κB ebenfalls
involviert (Gukovskaya and Pandol, 2004). Bisher konnte nachgewiesen werden,
dass NF-κB u.a. die Expression der anti-apoptotisch Proteine BCL-2 und BCL-XL
induziert und über die Expression von TNF-a die Apoptose mit beeinflusst (Zhang et
al., 2007)

Einleitung I
13
In anderen Pankreatitis-Modellen, wie z.B in der L-Arginin induzierten Pankreatitis
oder der Taurocholate induzierten Pankreatitis konnte ebenfalls eine frühzeitige
Induktion von NF-κB beobachtet werden (Algul et al., 2002)
Aus den genannten Gründen ist der NF-κB-Signalweg in der akuten experimentellen
Pankreatitis von zentraler Bedeutung.
7 Ziel dieser Arbeit
Zahlreiche Forschungsarbeiten beschäftigten sich mit der Rolle des
Transkriptionsfaktors NF-κB in der akuten Pankreatitis. Bislang wurde u.a. durch
pharmakologische Inhibierung, konstitutive Aktivierung oder durch Verwendung
konditioneller knockout Mäuse die Bedeutung von NF-κB untersucht. Diese Arbeiten
kamen zum Teil zu widersprüchlichen Ergebnissen. Ob NF-κB, aktiviert in
Azinuszellen, die akute Pankreatitis aggraviert oder abgemildert, konnte noch nicht
hinreichend geklärt werden.
Deshalb beschäftigt sich diese Arbeit mit der Bedeutung des IKK/NF-κB/RelA-
Signalwegs in der Pathogenese der akuten Pankreatitis. Die Verwendung
konditioneller gewebespezifischer Knockout-Mäuse bietet den Vorteil, spezifische
Aussagen über Funktion und Bedeutung eines einzelnen Proteins in einem Organ
treffen zu können. Alguel et al. konnten bereits zeigen, dass eine pankreas-
spezifische Inaktivierung von NF-κB zu einer schwerer verlaufenden Pankreatitis
führt (Algul et al., 2007). In der nun folgenden Arbeit wurde untersucht, ob die
pankreas–spezifische Deletion des NF-κB Inhibtorproteins IκBa zu einer nukleären
Translokation und transkriptionellen Aktivität von NF-kB führt und welche Bedeutung
dies für die Entwicklung des Pankreas und den Verlauf einer akuten Pankreatitis hat.

II Methoden
14
II. Methoden
1 Tierhaltung
Alle Versuchstiere wurden in den Tierställen des Klinikums rechts der Isar gemäß
den Protokollen des Zentrums für präklinischen Forschung der Technischen
Universität München gehalten. Die Räume speziell konzipiert für die
Versuchstierhaltung sind klimatisiert und verfügen über einen Tag/Nacht – Rhythmus
(12h hell/12h dunkel). Die Mäuse wurden in einzelbelüfteten Mäusekäfigen gehalten
und erhielten autoklaviertes Mäusefutter und Wasser zur freien Verfügung. Die
Pflege erfolgte durch speziell ausgebildete Tierpfleger.
2 Induktion einer akuten Caerulein-Pankreatitis
Wie bereits beschrieben verursacht die Stimulation des Pankreas mit
supramaximalen Konzentrationen Caerulein eine akute Pankreatitis. Um dies zu
erreichen, wird der Maus in stündlichen Abständen 50 µg/kg/KG Caerulein gelöst in
NaCl intraperitoneal appliziert.
Für die Induktion der Pankreatitis wurden jeweils ikbaΔpanc
- und ikbaF/F-Mäuse
verwendet, das Geschlechtsverhältnis war in beiden Gruppen gleich. Die Mäuse
waren bei Durchführung des Experiments jeweils zw. 2,5 – 3,5 Monate alt.
Im Rahmen dieser Arbeit wurde eine 8h- und eine 24h-Pankreatitis induziert. Vor der
ersten Injektion mussten die Mäuse eine mindestens 12-stündige Nahrungskarenz
einhalten, wobei Wasser ad libidum verfügbar war. Sowohl für die 8h-, wie auch 24h-
Pankreatitis wurde Caerulein 8mal injiziert und 8h bzw. 24h nach Beginn wurden die
Tiere mit Isofluran euthanasiert.
3 Induktion einer L-Arginin Pankreatitis Für die L-Arginin Pankreatitis wurde eine 8% sterile L-Arginin-HCL Lösung (pH 7)
verwendet. Den Mäusen wurden jeweils 4g/kg/KG Arginin intraperitoneal injiziert. Es
erfolgten 2 Injektionen im Abstand von einer Stunde. Eine Nahrungskarenz wurde

Methoden II
15
nicht eingehalten. 72h nach der Erstinjektion wurden die Mäuse getötet und
Pankreas und Lunge zu weiteren Analysen entnommen.
4 Präparation und Materialgewinnung
Zu den Zeitpunkten 0h, 4h, 8h und 24h wurde den Mäusen Blut aus der
Schwanzspitze entnommen. Die Blutproben wurden bei 4°C abzentrifugiert und das
gewonnene Serum für die Bestimmung von IL-6 und die Amylase und Lipase-
Messung verwendet.
Nach 8h bzw 24h wurden die Mäuse mit Isofluran euthanasiert. Das Körpergewicht
wurde gewogen. Nach Öffnung des Abdomens und des Thorax wurden die Mäuse
durch Punktion der V. cava inf. entblutet und Pankreas, Duodenum, Milz, Leber und
die Lunge herauspräpariert. Das Gewicht des Pankreas wurde gewogen.
Gewebeproben von Pankreas und Lunge für die Proteingewinnung und die
Serumproben wurden in flüssigem Stickstoff schockgefroren und bei -80°C
aufbewahrt.
Für die RNA Gewinnung wurde Pankreasgewebe in 1200 µl RLT-Puffer, (RNeasy-
Kit, Quiagen) versetzt mit 1% β-Mercaptoethanol, gegeben, homogenisiert und in
flüssigen Stickstoff schockgefroren.
Für die Histologie wurden die Organe über Nacht in 4% Paraformaldehyd fixiert, die
Entwässerung erfolgte durch das Institut für Pathologie des Klinikums rechts der Isar,
München, anschließend wurden die Organe in Paraffin eingebettet. Zur Herstellung
von Gefrierschnitten wurde ein Teil des Pankreas direkt in Tissue Tec
schockgefroren.
5 Zellkultur
5.1 Azini-Isolierung
Die Auftrennung des Pankreasgewebes in einzelne Azini ist eine Methode, die auf
dem Prinzip der enzymatischen Andauung durch Kollagenase beruht .
Für die Gewinnung des Pankreas wurden die Mäuse mit Isufloran eingeschläfert und
nach Eröffnung des Bauchraums mittels Punktion der V. cava inferior entblutet.

II Methoden
16
Nach der Explantation des Mäusepankreas, muss dieser in einer Gewebeschale mit
PBS gewaschen und auf Eis gelagert werden. Nun wurde das Pankreas mit 5 ml
Kollagenase–Lösung infundiert. Für eine effektive Andauung des Pankreas muss die
Kollagenase–Lösung fächerförmig in den Pankreas injiziert werden, damit das
Pankreas in einzelne Läppchen aufgetrieben wird. Anschließend wurde das
Pankreasgewebe mittels einer Schere zerkleinert, so dass die einzelnen Läppchen
freigelegt werden. Die Pankreas-Kollagenase-Suspension wurden für 10 min bei
37°C inkubiert. Als nächster Schritt wurde die Suspension mit 10 ml Waschlösung
verdünnt und bei 300 rpm für 5 min bei Raumtemperatur zentrifugiert. Der Überstand
wurde mit einer Glaspipette abgesaugt und das Pellet mit 5 ml Kollagenase–Lösung
resuspendiert. In einer Gewebeschale folgte eine weitere Inkubation bei 37°C für 10
min. Nach dem zweiten Andauungsschritt müssten nun die Azini von den nicht
aufgetrennten Gewebeanteilen mittels eines sterilen Zellsiebs separiert werden. Um
den Erhalt einer möglichst großen Menge an Azini zu gewährleisten, wurde das
Zellsieb mit 15 ml Waschlösung gespült. Die 20 ml Azini–Suspension wurde nun ein
weiteres mal bei 300 rpm für 3min zentrifugiert, der Überstand verworfen und das
Pellet mit 20 ml Waschlösung resuspendiert. Dieser Zentrifugationschritt wurde noch
einmal wiederholt. Das Zellpellet wurde anschließend McCoys 5a Medium
resuspendiert. Vor der weiteren Bearbeitung wurden die Azini 30 min bei 37°C
inkubiert.
6 Genotypisierung
6.1 DNA-Extraktion
Als Ausgangsmaterial wurden die Schwanzspitzen der Mäuse verwendet. Die
Schwanzspitze wurde zuerst in 750 µl Lysepuffer, der die Proteinase K enthält,
mindestens 5 h bei 50°C verdaut.
Nachdem die Verdauung abgeschlossen war, wurden 250 µl einer 6M NaCl–Lösung
zugegeben. Anschließend wurde die Suspension bei 13000 rpm für 30 min bei 4°C
zentrifugiert. Es wurden 200 µl aus der Mitte der Lösung entnommen, der Rest
verworfen. Nach Zugabe von 175 µl Isopropanol, fällt die DNA aus. Das
überschüssige Isopropanol wurde bei 13000 rpm 10 min lang abzentrifugiert und
verworfen. Es folgten 2 Waschschritte mit -20°C kalten 75%igem Ethanol, der

Methoden II
17
ebenfalls jeweils 10 min lang abzentrifugiert wurde. Die als Pellet sichtbar gewordene
DNA wird anschließend ca 15 min bei RT getrocknet und in 50 µl TE–Puffer
aufgenommen. Bevor die DNA weiter verwendet werden konnte, musste sie noch 2 h
bei RT inkubieren.
6.2 Poymerase-Chain-Reaktion (PCR) Die PCR wurde mit Hilfe des RedTaq ReadyMix PCR Reaction Mix (#2523, Sigma)
nach Angaben des Herstellers durchgeführt. Hierfür wurden jeweils 12,5 µl Red-Taq-
Ready-Mix©, 10,5 µl H20, 1 µl Primer in einer Konzentration von 10 pmol/µl und 1 µl
der Probe gemischt. Anschließend wurden die DNA-Abschnitte im Thermocycler®
vervielfältigt.
In Tabelle 1 sind die PCR-Bedingungen aufgeführt, in Tabelle 2 die verwendeten
Primer-Sequenzen.
Temperatur Dauer Wiederholungen Funktion
95°C 1 min 1x Denaturierung
der DNA
94°C 30 sec
40x
“ “ “
58°C 30 sec Anlagerung der
Primer
72°C 1 min 30 sec Elongation
72°C 10 min 1x Extension Tabelle 1: PCR Temperaturzyklen
Primer Sequenzen
ptf1acreex1 FW 5’- GTCCAATTTACTGACCGTACACCAA-3’
Re 5’- CCTCGAAGGCGTCGTTGATGGACTGCA-3’
ikba FW 5’ – CCA AGC AGA GAC GTG TAT TTC T – 3’
RE 5’ – TCC AGA CAG TAA GGG CCA GGT – 3’ Tabelle 2: Primer und Sequenzen
6.3 DNA Detektion

II Methoden
18
Um die amplifizierten DNA-Abschnitte sichtbar zu machen, wurden die Proben
gelelektrophoretisch aufgetrennt. Für die Agarosegele wurden 2 g Agarose auf 100
ml TAE-Puffer (entspricht einem 25%igem Gel) durch Aufkochen in der Mikrowelle
gelöst und anschließend mit 0,01% (v/v) Ethidiumbromid nach kurzem abkühlen
versetzt. Die ausgehärteten Gele wurden mit den PCR-Produkten beladen. Die
Auftrennung erfolgte in einer Elektrophoresekammer bei 100 V für ca 2 h. Die
Detektion der DNA-Abschnitte erfolgte mit dem Imager Gel Doc XR System unter
UV-Licht.
7 Genexpressionsanalysen 7.1 RNA-Isolation
Die RNA-Isolation aus Pankreasgewebe erfolgte mit dem RNeasy® Mini Kit von
Quiagen nach Angaben des Herstellers.
Nachdem die aufgereinigte RNA mit 50 µl H2O aus der Membran gefällt wurde,
bestimmte man die Konzentration mit Hilfe des Nanodrop® photometrisch und
analysierte die Qualität der isolierten RNA mittels Gelelektrophorese. Hierfür wurden
1% Agarosegele verwendet. Zu 2 µl RNA werden 8 µl H2O und 1,7 µl Orange G
gegeben und das Gel damit beladen. Die Auftrennung der Proben erfolgte bei 60V
für 1,5h.
7.2 Affymetrix Mikroarry Die Mikroarry-Technologie erlaubt die qualitative und quantitative Analyse eines
umfassenden Genexpressionsprofils.
Auf einem Array, der in unserem Fall das gesamte Maus-Genom enthält, wird die
biotin markierte cRNA des Pankreas eines Versuchstieres hybridisiert. Nach der
Hybridisierung wird der Array fluoreszenz-gefärbt und gescannt. Anhand des Orts
und der Stärke des Fluoreszenzsignals kann dann ein exprimiertes Gen detektiert
und das Expressionsniveau quantifiziert werden.
In dieser Arbeit wurde der Mouse Genom 430 A 2.0 Array verwendet. Die Analyse
und das Scannen des Genchips erfolgte mit der Affymetrix Microarray Suite 5.0
software (MAS 5.0). Die Durchführung des Experiments sowie die Datenauswertung

Methoden II
19
erfolgte durch Dr. med. Henrik Einwächter, II.Medizinische Klinik, Klinikum rechts der
Isar mit Hilfe des Affymetrix one cycle target labelling kit und GeneChip Hybridization
Wash and Stain Kit nach Angaben des Herstellers.
7.3 GSEA Werden die Affymetrixdaten zweier Phänotypen miteinander verglichen, kann man
anhand der unterschiedlichen Expressionsstärken Ranglisten der einzelnen Gene
erstellen. Gene am unteren bzw am oberen Ende dieser Liste unterscheiden sich am
stärksten, Gene in der Mitte werden ungefähr gleich stark in beiden Phänotypen
exprimiert.
Die Gen Set Enrichment Analysis Methode (kurz GSEA) ist ein statistisches
Verfahren, die es erlaubt, diese Ranglisten auf bestimmte Gen-Gruppen hin zu
untersuchen. Es kann festgestellt werden, ob sich die Expression einer bestimmten
Gruppe von Genen (Gen-Set) in zwei verschiedenen Phänotypen signifikant
unterscheidet oder diese willkürlich innerhalb der ermittelten Rangliste verteilt sind
(Subramanian et al., 2005). Je mehr Gene eines Gen Sets am oberen oder unteren
Ende der Rangliste zu finden sind, um so wahrscheinlicher ist dass dieses Gen-set
signifikant stärker im jeweiligen Phänotyp exprimiert wird.
In den durchgeführten Analysen wurden jeweils die Array-Daten der ikbaΔpanc
bzw
relaΔpanc
mit denen der ikbaF/F Maus verglichen. Die GSEA Software und das Gen-
Set wurden vom “Broad Institute of MIT and Harvard“ zur Verfügung gestellt, die
voreingestellten Standardparameter der Software wurden übernommen.
8 Western Blot 8.1 Proteinlyse und Proteinbestimmung
Um das Pankreasgewebe für die Analyse von Proteinen aufzubereiten, wurde das
Gewebe in 500 µl IP-Puffer homogenisiert und die Suspension bei 13000 rpm und
4°C für 15 min zentrifugiert. Der Überstand, der aus Proteinlysat besteht, konnte nun
für die weiteren Analysen verwendet werden.
Für die Western Blot Analyse wurde der Proteingehalt der Gewebelysate mit Hilfe
des BCA Protein-assay-kits (Thermo scientific) mittels photometrischer Analyse nach

II Methoden
20
den Angaben des Herstellers bestimmt. Die Proben wurden jeweils im Verhältnis 1:5
mit IP-Puffer verdünnt. Als Standardkontrolle diente eine Albumin-Verdünnungsreihe
mit Konzentrationen von 2000 bis 0 µg/ml. Die zumessende Probenlösung, bzw. die
Standardkontrolle wurden im Verhältnis 1:20 mit der Kit-Lösung gemischt und bei
37°C für 30 min inkubiert. Die photometrische Messung erfolgte bei 562 nm.
Daraufhin wurden die Proben auf eine Proteinkonzentration zwischen 1 und 2 µg/µl
eingestellt und mit 5x Lämmli- Puffer versetzt.
8.2 Gelelektrophoretische Auftrennung der Proteine Die Auftrennung der Proteine erfolgte mit diskontinuierlichen SDS-Page-Gelen
bestehend aus einer Phase Sammelgel und einer Phase Trenngel. Es wurden 10%
Trenngele verwendet. Bevor das Gel mit den Proben beladen werden konnte,
wurden die Lämmli-Proteinlysate bei 95°C für 5 min denaturiert. Pro Probe wurden
zwischen 5 und 10 µl Proteinsuspension verwendet.
In der Gelelektrophoresezelle erfolgt die molekulargewichts-abhängige Auftrennung
in Laufpuffer.
Trenngel 10% Sammelgel
dH20 2,0 ml 1,5 ml
Puffer 1,3 ml (Trenngel-Puffer) 750µl (Sammelgelpuffer)
30% Acrylamid 1,65 ml 375µl
10 % SDS 50 µl 25 µl
10% APS 25 µl 12,5 µl
TEMED 7,5 µl 5 µl Tabelle 3: Zusammensetzung Trenn- und Sammelgel
8.3 Membrantransfer Nach Aktivierung der PVDF-Membran in Methanol, wurden sowohl die Membran, als
auch das mit Protein beladene Gel in Nasstransferpuffer getränkt und zwischen
Whatman Papier eingebettet. Es folgte die elektrophoretische Übertragung der
Proteine auf die Membran in Nasstransferpuffer in der Blottingzelle. Für diesen
Vorgang wurde eine Stromstärke von 350 mA für 120 min angelegt.
8.4 Proteindetektion

Methoden II
21
Für die Detektion von Proteinen mittels spezifischer Antikörper war es notwendig,
unspezifische Bindungsstellen der Membran mit dem Medium des Antikörpers, für 1h
bei RT abzublocken Zum Blocken der Membran wurde je nach verwendetem
Antikörper 5% Magermilchpulver oder 5% BSA in TBST verwendet. Anschließend
erfolgt die Inkubation der Membran mit dem jeweiligen Antikörper über Nacht bei
4°C. Zur Detektion des gebundenen Antikörpers erfolgte die Inkubation mit dem
entsprechenden Zweitantikörper für 1h bei RT. Die Sekundärantikörper sind mit dem
Enzym Merrettich-Peroxidase gekoppelt. Zwischen all diesen Schritten wurde die
Membran mit TBST gewaschen, um überschüssige Medien und nichtgebundene
Antikörper zu entfernen. Um die nun gebundenen Antiköprperkomplexe mittels
Chemilumineszenz visualisieren zu können wurden Amersham ECL Western Blotting
Detection Reagants auf die Membran gegeben. In der Dunkelkammer wurden die
Antikörpersignale auf Film übertragen und mit Hilfe des Amersham Hyperprocessors
entwickelt.
Tabelle 4 und 5 geben Aufschluss über die verwendeten Erst- und Zweitantikörper.
Tabelle 4: Erstantikörper Western Blot
Zweitantikörper Seriennr. Hersteller
Blocklösung Verdünnung
ECL Anti - Rabbit #NA934 5% Trockenmilch 1: 500
Erstantikörper (Spezies)
Seriennr. Hersteller
Molekular- gewicht
Block-Lösung Verdünnung
Anti - IκBα (C21)
(Rabbit)
# sc-371
Santa cruz 37 kDA
5% Trockenmilch
in TBST 1: 1000
Anti - Iκbβ C20)
(Mouse)
#sc
945/Santa
cruz
45 kDA 5% Trockenmilch
in TBST 1: 1000
Anti - p65 (C20)
(Rabbit)
#sc-372
Santa cruz 65 kDA
5% Trockenmilch
in TBST 1: 1000
Anti-pp65 (Ser 276)
(Rabbit)
#3037 Cell
Signaling 80 kDa 5% BSA in TBST 1: 1000
Anti - p50
(Rabbit)
#3035 Cell
Signaling 50 kDA 5%BSA in TBST 1: 1000

II Methoden
22
IgG
(HRP-conjugated)
GE Healthcare in TBST
ECL Anti - Mouse
IgG
(HRP-conjugated)
# NA931
GE Healthcare
5% Trockenmilch
in TBST 1: 500
Tabelle 5: Zweitantikörper Western Blot
8.5 Membranreaktivierung Um die Membran erneut mit Antikörper beladen zu können, muss diese zunächst mit
Methanol für 1 min reaktiviert werden. Daraufhin werden die gebundenen
Antikörperkomplexe mit Stripping-Puffer für 20 min gelöst. Nun kann die Membran
erneut mit Antikörper beladen werden.
9 Morphologie 9.1 Anfertigung von Paraffinschnitten
Nachdem die Organe der Maus entnommen wurden, wurden sie für 24h bei 4°C in
4% PFA inkubiert, im Institut für Pathologie, Klinikum Rechts der Isar nach einem
Standardprotokoll dehydriert und in Paraffin gegossen. Die Paraffinblöcke werden
auf -20°C gekühlt, mit dem Mikrotrom zwischen 3,5 und 4 µm dick geschnitten und
auf Objektträger fixiert.
9.2 Anfertigung von Cryoschnitten Für die Anfertigung von Cryoschnitten ist es notwendig das Gewebe sofort nach der
Präparation in Tissue Tec® mit Stickstoff schockzugefrieren. Anschließend wurden
die Blöcke mit dem Cryotrom geschnitten und auf Objektträger fixiert.
9.3 Hämatoxylin-Eosin-Färbung Für die Hämatoxylin-Eosin-Färbung (kurz H&E-Färbung) wurden die Paraffinschnitte
zunächst in einer absteigenden Alkoholreihe deparaffinisiert und rehydriert (5 min
Histoclear, je 3 min in 100%, 96% und 80% Ethanol, 5 min in dH2O). Anschließend
für 10 min mit Hämalaun gefärbt. Daraufhin wurden die Schnitte für 10 min mit
Leitungswasser gewaschen werden. Die Gegenfärbung erfolgte für 5 min mit Eosin.
Abschließend erfolgte die Fixierung der Schnitte der Reihe nach für je 5 min mit 96%

Methoden II
23
Ethanol, Isopropanol und Histoclear und die Schnitte wurden in Mounting-Medium
eingebettet.
9.4 Histoscore
Die Quantifizierung der Nekrose und Inflammation im Pankreasgewebe erfolgte an
H&E gefärbten Paraffin-Schnitten. Es wurden die Pankreata von ikbaΔpanc
und ikbaF/F-
Mäusen (s.u.) nach Induktion einer Caerulein-Pankreatitis verglichen. Die
Auswertung erfolgte verblindet durch 2 unabhängige Untersucher. Als Kriterien für
Nekrose galten Verlust der Zellmembran–Integrität, Zellödem, Austritt von
Zellorganellen in das Interstitium. Die Entzündung des Pankreas wurde anhand des
Ausmaß der Infiltration von inflammatorischen Zellen bewertet. Die Evaluation
erfolgte zum Zeitpunkt 24h. Die histologischen Schnitte des Pankreas wurden jeweils
für Nekrose und Inflammation mit einem Punktwert zwischen 0 (normal) und 3
(schwer) bewertet.
9.5 Immunhistochemie. Die Paraffin-Schnitte mussten als erstes deparafinisiert und rehydriert werden (5 min
Histoclear, je 3 min in 100%, 96% und 80% Ethanol, 5 min in dH2O). Für die
Demaskierung der Antigene wurden die Schnitte für ca. 10 min in Antigen
Unmasking Solution (10 mM Natrium-Citrat Puffer) in der Mikrowelle gekocht.
Für die nächsten Schritte wurden die Objektträger in feuchte Kammern umgestellt.
Nachdem die Objektträger ca 15 min auskühlten, wurden sie 3x für je 5 min mit PBS
gewaschen. Anschließend wurden die Schnitte für 15 min mit 3% v/v H2O2
abgesättigt und erneut 3x für je 5 min mit PBS gewaschen.
Für die Reduktion unspezifischer Hintergrundreaktionen wurden 4 Tropfen Avidin
und 5% Serum (v/v) in PBS gelöst und für 30 min auf die Gewebeschnitte gegeben.
Dabei wurden ca. 200 µl angesetzte Lösung pro Schnitt verwendet. Es erfolgt ein
weitere Waschschritt.
Die Primärantikörperlösung setzt sich aus 85% PBS, 2,5% Serum des
Sekundärantikörpers, 4 Trpf Biotin und unkonjugierten Primärantikörper. Diese wird
über Nacht bei 4°C auf den Objektträgern belassen.
Nach einem weiteren Waschschritt werden die Objektträger für 30 min bei RT mit
dem Sekundärantikörper inkubiert. Diese Lösung setzt sich aus 97% PBS, 3%
Serum und biotinyliertem Sekundärantikörper zusammen.

II Methoden
24
30 min bevor man sie auf die Schnitte gibt, muss Avidin Biotin Complex-Lösung nach
Angaben des Herstellers (ABC Elite Kit, Vektor Laboratories) vorbereitet werden.
Nachdem die Schnitte gewaschen wurden werden sie mit der Lösung für 30 min bei
RT inkubiert. Es erfolgt eine weiterer Waschschritt. Um die gebunden Antigen-
Antikörperkomplexe anzufärben wird DAB-Lösung für 2 min auf die Schnitte
gegeben. Für die DAB-Lösung (Vektor Laboratories) werden je ein Tropfen Buffer
Stock Lösung, Wasserstoffperoxid Lösung und zwei Tropfen DAB in 2,5 ml dH2O
gelöst. Nun erfolgt kein Waschschritt mit PBS mehr, sondern die Reaktion der DAB
Lösung wird mit H2O abgestoppt. Nachdem man die Schnitte kurz mit Hämatoxilin
gegen gefärbt hat werden sie wieder dehydriert (2 min 80 % Ethanol, 2 min 96%
Ethanol, 2 min 100% Ethanol, 5 min Histoclear) und in Mounting Medium eingebettet.
Tabelle 6 gibt Aufschluss über die verwendeten Antiköper.
Tabelle 6: Antikörper Immunhistochemie
9.6 Immunfluoreszenz Die Cryoschnitte mussten zunächst für 10 min in 4% PFA bei 4°C fixiert werden.
Anschließend wurden sie für 5 min mit H2O gewaschen bevor man für 10 min 3%
Erstantikörper (Spezies) (Hersteller)
Verdünnung
Zweitantikörper (Spezies) (Hersteller)
Verdünnung
Serum
Anti - CK 8
(Mouse) (#sc101459,santa cruz)
1:250 Biotinylated Anti-Mouse (Vector Laboratories)
1:500 5% Goat-
Serum
Anti - Insulin
(Guinea-Pig) (#MAB 1417, R&D
System)
1:500 Biotinylated Anti-Guinea
Pig (Dianova)
1:500 5% Goat-
Serum Anti - Glukagon
(Guinea Pig) (# 4031-01F, Linco)
1:500

Methoden II
25
H2O2–Lösung auf die Schnitte gibt. Nachdem die Schnitte noch einmal mit H2O
gewaschen wurden, erfolgen alle weiteren Waschschritte mit PBS.
Danach wurde eine Blocklösung bestehend aus 50 µl Serum des Primärantikörpers
und 2 Trpf. Avidin gelöst in PBS für 1h auf die Schnitte gegeben. Es schlossen sich 2
Waschschritte mit PBS an. Die Inkubation des Primärantikörpers erfolgte bei 4°C
über Nacht. Diese Lösung setzt sich aus 50 µl Serum, 2 Trpf Biotin und
Erstantikörper gelöst in PBS zusammen. War dieser Inkubationsschritt vollzogen,
wurden die Schnitte erneut mit PBS gewaschen und die Sekundärantikörperlösung,
bestehend aus 50 µl Serum und biotinylierten Zweitantikörper (Verdünnung 1:1000)
gelöst in 1ml PBS, für 1h inkubiert. Nachdem die Schnitte mit PBS gewaschen
wurden, erfolgt die Detektion und Anfärbung der gebundenen Immunkomplexe mit
Hilfe des “TSA–Kit with HRP-conjugated Streptavidin and Alexa Fluor® 568
Tyramidin“ (Invitrogen) nach Angaben des Herstellers. Hierfür wurden die Schnitte
mit HRP-conjugated Streptavidin (in PBS, 1:100) für 30 min inkubiert. Die nächsten
Schritte mussten in Dunkelheit erfolgen. Die Lösung bestehend aus 20 µl einer
0,15% H2O2 Lösung, 10 µl Alexa Fluor 568 und 2 ml Puffer (Tyramidin stock solution
gelöst in Amplifikationspuffer, 1: 100) wurde für 5 min auf die Schnitte gegeben. Das
Abstoppen der Reaktion erfolgt mit 3 Waschschritten H20 für je 10 min. Anschließend
wurden die Zellkerne mit Sytox green (in TBS, 1:10000) gefärbt und die Schnitte in
Mounting Medium eingebettet.
Die Auswärtung der Immunfluoreszenzgefärbten Schnitte erfolgte mit dem Axiovert
200 Mikroscop von Zeiss. Das Fluoreszenz-Spektrum für Alexa Fluor 568 liegt bei
579/604 nm und für Sytox Green bei 504/523 nm. Die mit Sytox gefärbten Zellkerne
erscheinen grün, die an das Zielprotein gebundenen Antikörperkomplexe erscheinen
rot. Im Falle von nuklear lokalisierten p65 ergibt sich als Fusionsfarbe orange.
Tabelle 7 gibt eine Übersicht über die verwendeten Antikörper.
Erstantikörper (Spezies) (Hersteller)
Verdünnung Zweitantikörper (Spezies) (Hersteller)
Verdünnung Serum
Anti - IκBα
(Rabbit) (#sc-371 Santa cruz)
1:100
Biotinylated Anti
Rabbit (Goat) (#BA-1000 Vektor
Laboratories)
1:1000 5% Goat
Serum
Anti - p65 (C20)

II Methoden
26
Tabelle 7: Antikörper Immunfluoreszenz
10 Enzymbiochemie
10.1 MPO-Aktivitätsmessung in Lungengewebe
Um die systemischen Komplikationen einer akuten Pankreatitis quantifizieren zu
können, wurde die Myeloperoxidase-Aktivität in der Lunge, als Ausdruck für die
granulozytäre Infiltration, bestimmt.
Für die Messung wurde pro Maus ein Lungenflügel verwendet. Dieser wurde
zunächst mit 600 µl Homogenisierungspuffer homogenisiert. Daraufhin erfolgte eine
Ultraschalldesintegration der Suspension. Um die Zellwände aufzubrechen wurde die
Suspension mehrfach in flüssigem Stickstoff schockgefroren und wieder aufgetaut.
Anschließend wurde der Zelldetritus 15000 rpm für 10 min abzentrifugiert und der
Überstand für die Messung verwendet.
Die Messung erfolgte photometrisch in einem Spektrophotometer mit Kinetik bei 470
nm. Die Reaktionslösung setzt sich aus 630 µl Phosphatpuffer (10mM pH6), 50 µl
Probe und den Substraten 0,22% Guiacol und 0,1% H2O2 zusammen, wobei H2O2
erst unmittelbar vor der Messung hinzugefügt wurde. Die gemessene
Extinktionsänderung verläuft zu Beginn der Reaktion linear ansteigend, wobei die
Steigung der Geraden proportional zur Emzymaktivität ist. Die so gemessene
Aktivität wird um das Gewicht des verwendeten Lungenflügels korrigiert und die
Daten mit Hilfe von Exel 2003® (Microsoft) ausgewertet und grafisch dargestellt
10.2 Trypsin–Aktivitätsmessung im Pankreas
Die Messung der Trypsinaktivität erfolgte fluorimetrisch nach der Methode von
Halangk et al. (Halangk et al., 2000). Hierfür wurde der Pankreas homogenisiert und
25 ml Homogenisat bei 37°C mit 2ml Tris/HCL Puffer (pH 8) , der 15 mM NaCL, 1
mM CaCl2 und 0,1 mg/ml BSA enthält, versetzt. Als Substrat dient ein fluorogenes
Peptid-Substrat (Boc-Gln-Ala-Arg-7-amino-4-methyl-coumarin).
(Rabbit) (#sc-372 Santa cruz)

Methoden II
27
Die Fluoreszenzemission wurde bei 380 nm gemessen nachdem die Substrat-
Enzym-Lösung bei 440 nm angeregt wurde. Die Trypsin-Aktivität wurde anhand einer
Standardkurve aus gereinigten Trypsin errechnet. Die Auswertung der Daten erfolgte
mit Hilfe von Exel 2003® (Microsoft)
11 Serologie
11.1 Amylase- und Lipase-Messung Die Bestimmung der Enzyme Amylase und Lipase im Serum erfolgte durch das
Institut für Klinische Chemie des Klinikums Rechts der Isar.
11.2 IL6-Elisa Die quantitative Bestimmung von IL-6 im Serum erfolgte mittels des “Mouse IL-6
ELISA Ready-SET-GO“-Kits [eBioscience] nach Angaben des Herstellers.
12 Statistische Auswertung Die statistische Auswertung und graphische Darstellung erfolgte mittels dem
Programm Microsoft Windows Exel 2003®. Die erhoben Daten werden als Mittelwerte
± Standardabweichung dargestellt. Zur Gewährleistung eines statistisch eindeutigen
Ergebnisses wurden die Gruppengrößen jeweils entsprechend festgelegt. Die
Signifikanzanalyse der erhobenen Unterschiede zweier Vergleichsgruppen erfolgte
mittels T-Test. P-Werte ≤ 0,05 werden als signifikant angesehen.

III Material
28
III. Material
1 Geräte Bezeichung Name/Seriennummer Firma 96-Well Plate ELISA Corning Costar 9018 eBioscience
Chromatographie Papier Whatman
Cryotom HM 569 Microm
Digitalkamera AxioCam HRc Zeiss
Feinwaage A200S Sartorius Analytic
Filmentwickler Hyperprocessor GE Healthcare
Fluoreszenzmikroskop Axivert 200M Zeiss
Gen chip array Mouse Genom 430A 2.0 Array Affymetrix
Gen-Chip Scanner GenChip scanner 3000 Affymetrix
Homogenisator DIAX 900 Heidolph
Hybridisierungsofen GeneChip Hybridization Oven Affymetrix
Lichtmikroskop Axioimager A1 Zeiss
Magnetrührer MR 3001 Heidolph
Microarray Waschstation GeneChips Fluidics Station 450 Affymetrix
Mikrotom HM 3555 Microm
Objektträger Superfrost Plus Menzel-GläserWh
PCR-Elektrophoresekammer Sub-cell ® Bio RAD
PCR-Gelentwickler GelDoc XR Bio RAD
PCR-Maschine Primus 16+ MWG AG Biotec
pH-Meter MP220 Mettler-Toledo
Photometer (Protein) Smartspec plus Bio RAD
Photometer (RNA) Nanotrop ND 1000 PEQLab
Pipetten research/reference/easypet Eppendorf
Pipetten (PCR) safe seal professional Biozym
Thermoblock Thermomixer compact Eppendorf
Tischzentrifuge Zentrifuge#5415R Eppendorf
Ultraschalldesintegrator UW2070 Bandelin sanoplus
Western Blot-Transfermembran Immobilion-P Transfer Membran Millipore
Western Blot Film Hyperfilm,#RPN3103K GE Healthcare

Material III
29
Western Blot-Transferkammer MINI(TransBlot) Cell Bio-RAD
2 Puffer und Medien
2.1 Reagenzien Bezeichung Name/Seriennummer Firma Acetylsäure 100% #100063 Merck
Acrylamid Rotiphorese gel, #3029.2 Roth
Agarose #840.004 Biozym
Ammoniumpersulfat #9592 Roth
Antigen Unmasking Solution #H-3300 Vektor Lab.
BSA #8076 Roth
Caerulein #C9026 Sigma
Eosin #2C-140 Croma
Glycin #H50753 Promega
Goat Serum #G9023 Sigma
Guiacol #sc205337 santa cruz
Haematoxilin #105174 Merck
HCL #109057 Merck
HEPES #H7523 Sigma#
Histoclear Roti-Histol #6640.1 Roth
HTAB #H9151 Sigma
Isofluran #B506 Forene Abbott
Methanol #4627.5 Roth
Mounting Medium #PER 20000 Pertex, Medite
PBS #L182-50 Biochrom
Rabbit Serum #R9133 Sigma
SDS #811030 ICN
Sytox green #S7020 Invitrogen
TEMED #87689 Fluka
Tissue freezing Medium Tissue-Tec #4583 Sakura
Tris-Base #5429.2 Roth
Trockenmilchpulver #70166 Fluka
Tween-20 #9127.1 Roth

III Material
30
Wasserstoffperoxid 30% #108597 Merck
β-Mercaptoethanol # M7522 Sigma
2.2 Puffer Bezeichnung Zusammensetzung
PBS 1M PBS
PBS-T 1xPBS, 0,1% v/v Tween 20
TBS 10mM Tris, 150 mM NaCl
TBS-T 1xTBS +0,05% Tween-20
Azini-Isolation
Solution I McCoys 5a Medium, 0,1% w/v BSA, 0,12% w/v
Collagenase
Solution II McCoys 5a Medium, 0,1% w/v BSA
Western Blot:
Blockpuffer 5% Trockenmilchpulver, in TBS-T
IP-Puffer 50 mM HEPES pH 7,9, 150 mM NaCl, 1 mM EDTA, 0,5%
NP-40, 10% Glycerol
Lämmli: 10% SDS, 300 mM Tris-HCL pH 6,8, 0,05%
Bromphenolblau 5% β-Mercaptoethanol
Laufpuffer 25 mM Tris, 192 mM Glycin, 0,1% w/v SDS
Sammelgelpuffer 0,5 M Tris/HCl pH 6,8
Stripping-Puffer 25 mM Glycin-HCl pH 2, 1% w/v SDS
Transferpuffer 25 mM Tris-HCl pH 8,3, 150 mM Glycerin
Trenngelpuffer 1,5 MTris-HCl pH 8,8
PCR:
Lyse-Puffer: 50mM Tris-HCL pH 8, 100mM EDTA pH 8, 100mM
NaCl, 1% w/v SDS, 0,5% w/v Proteinase K
MPO-Aktivität
Homogenisierungspuffer 0,5% w/v HTAB 50mM Phosphatpuffer pH 6

Material III
31
2.3 Kits Name Seriennummer Firma
ABC Elite Kit #PK-6100 Vektor Lab.
Affymetrix one cycle target labelling kit #900493 Affymetrix
Amersham ECL WB Detection Reag. #RPN2106 GE Healthcare
Avidin/Biotin Blocking Kit #SP-2001 Vektor Lab.
BCA-Protein-Assay Kit #23227 Thermo scientific
DAB Lösung #SK-4100 Vektor Lab.
GeneChip Hybridization Wash and Stain Kit #900720 Affymetrix
Mouse IL-6 ELISA Ready-Set-Go! #88-7064 eBioscience
RedTaq ReadyMix PCR Reaction Mix #R2523 Sigma
RNeasy Mini Kir #74104 Quiagen
TSA – Kit #T20934 Invitrogen

IV Ergebnisse
32
IV. Ergebnisse
1 Die Pankreas-spezifische Inaktivierung von ikba führt zu einer nukleären
Translokation von NF-κB
Um die Rolle von NF-κB speziell im Pankreas untersuchen zu können, ist es
notwendig pankreas-spezifische ikba-knockout Mäuse zu generieren. Zudem sterben
Mäuse, die nicht in der Lage sind IκBα systemisch zu exprimieren, ca. 9-10 Tage
post partum (Beg et al., 1995, Klement et al., 1996).
1.1 Generierung der ikbaΔpanc-Maus mittels des Cre/loxP-Systems
Die Inhibition von IκBα im exokrinen Pankreas wurde mit Hilfe des Cre/loxP-Systems
erreicht. Der Knockout des ikba-Gens hat zur Folge, dass zu keinem Zeitpunkt ein
funktionsfähiges Protein in den Azinuszellen der Maus exprimiert werden kann.
Die Cre-Rekombinase wurde aus dem Bakteriophagen P1 isoliert und kann auch in
Säugetieren zu exprimiert werden. Sie hat die Eigenschaft in der DNA
Rekombination zwischen spezifischen DNA-Erkennungssequenzen, den
sogenannten loxP-Sites (locus of X-over P1) zu vollziehen. Dies geschieht autonom
unabhängig von der Art der DNA und ohne dabei auf weitere Kofaktoren angewiesen
zu sein (Lukowski R., 2005). Bei den loxP-sites handelt es sich um 34 Basenpaar
große DNA-Abschnitte, bestehend aus einer zentralen asymmetrischen Sequenz, die
von zwei Palindromen flankiert wird. Diese loxP-sites sind wie die Cre-Rekombinase
im Genom der Maus nicht enthalten. Für die intramolekulare Deletion eines Gens
müssen nun zwei gleichgerichtete loxP-sites in Intron-Sequenzen, die funktionell
wichtige Bereiche des Gens umgeben, eingebracht werden. Nach Deletion des
DNA–Abschnitts bleibt eine loxP-site zurück.
Die Pankreas-spezifische Expression von Cre wird erreicht, indem man eine knock-in
Maus generiert, in der die Cre-Rekombinase anstelle des Exon 1 in das PTF1a (p48)
Gen eingebracht wird. PTF1 ist basisches Helix-loop-helix Protein bestehend aus
den Untereinheiten p48 (PTF1a), p64 und p75, wobei p48 essentiell für die
Entwicklung des exokrinen Pankreas ist (Krapp et al., 1998). Somit wird die

Ergebnisse IV
33
Rekombinase nur zusammen mit der, für den exokrinen Pankreas spezifischen, p48-
Untereinheit des Transkriptionsfaktors PTF1 exprimiert. Da homozygote Ptf1acreex1–
Mäuse nicht lebensfähig sind, werden nur heterozygote Ptf1acreex1–Mäuse
verwendet (Nakhai et al., 2007). Kreuzt man nun transgene Mäuse, in deren Genom
ein bestimmtes Gen homozygot mit zwei loxP-sites markiert ist, mit transgenen
Ptf1acreex1–Mäusen, so wird das Gen im exokrinen Pankreas nicht exprimiert (Krapp
et al., 1996). In allen anderen Zellen aber ist das Protein intakt, da die loxP-sites
alleine die Expression des Gens nicht beeinflussen. Die Ptf1acreex1–Maus wurde von
Nakhai et al. 2007 im gastroenterologischen Labor der II. Medizinischen Klinik des
Klinikums rechts der Isar generiert (Nakhai et al., 2007).
Die homozygot gefloxte ikba-Maus (ikbaF/F –Maus) wurde großzügiger Weise von R.
Rupec von der Dermatologischen Klinik der Ludwigs Maximilians Universität zur
Verfügung gestellt (Rupec et al., 2005). Bei den ikbaF/F-Mäusen handelt sich um
129/svJ/C57BL/6 Hybriden die über 7 Generationen mit C57BL/6 Mäusen
zurückgekreuzt wurden. In der ikbaF/F
–Maus sind in der DNA zwei loxP-sites
integriert, die die Promoterregion mit der NF-κB-Bindestelle, den Core-Promoter,
Exon 1 und 2 umfassen. Eine homozygote ikbaF/F-Mauslinie wurde mit heterozygoten
Ptf1acreex1–Mäusen gekreuzt, um eine Mauslinie zu erhalten in denen IκBα in den
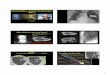
Azinuszellen des Pankreas nicht exprimiert werden kann. Abbildung 1 A zeigt das
Genkonstrukt der ikbaF/F-Maus.
Zum Nachweis der konstitutiven Inaktivierung von IκBα auf DNA-Ebene führten wir
PCR-Analysen für ikba und Ptf1acreex1 durch. Bei homozygot gefloxten ikba-Mäusen
erhält man in der PCR ein 365 bp großes Produkt, während das ikba-Gen der
Wildtyp Maus nur 275 bp groß ist. Wie in Abbildung 1B zu sehen ist, ergeben sich für
das Gen ikba 3 verschiedene Maus-Linien. Homozygot gefloxte ikba Mäuse (ikbaF/F
),
die wt-Linie (ikbawt/wt) und eine heterozygote Linie (ikbaF/wt). Kreuzt man homozygote
loxP-Mäuse (ikbaF/F
) mit Mäusen die, wie in Abbildung 1C dargestellt, heterozygot für
Ptf1acreex1 sind, so erhält man ikbaΔpanc
–Mäuse. Diese Mäuse expremieren IκBα im
Pankreas nicht.
Für die weiteren Nachweise verwendeten wir als Kontrolle die ikbaF/F
-Mauslinie, um
mögliche Interferenzen der loxP-sites auszuschließen.
Zur Analyse der Expression von IκBα im Pankreas wurden Western Blots aus
Pankreasgewebslysaten von 3 Monate alten ikbaF/F
– und ikbaΔpanc
-Mäusen durch

IV Ergebnisse
34
geführt. Wie in Abbildung 1D zusehen ist, konnte in ikbaΔpanc
-Mäusen, im Gegensatz
zu den ikbaF/F
–Mäusen nur eine schwache Bande bei 37 kDa detektiert werden.
Dieses schwache Signal ist auf den IκBα–Gehalt des nicht–azinären
Pankreasgewebes zurückzuführen. Das Expressionslevel von IκBβ in den
verschiedenen Mäusen ist annähernd gleich. Auch in der morphologischen Analyse
des Pankreasgewebes mittels Immunfluoreszenzfärbung gegen IκBα (Abbildung 1E)
zeigt sich, dass IκBα (rot) in ikbaΔpanc-Mäusen ebenfalls nur in nicht-azinärem
Gewebe vorhanden ist. In diesem Fall handelt es sich dabei um Blutgefäße und
duktalen Zellen. Im Zytosol der Azinuszellen ist IκBα nicht nachweisbar. In ikbaF/F
–
Mäusen hingegen färbt sich das Zytosol der Azinuszellen homogen rot an, da hier
IκBα regulär exprimiert wird.
Diese Ergebnisse weisen nach, dass es gelungen ist mittels des Cre/loxP Systems
eine konditionale pankreas-spezifische ikba Knockout Maus zu generieren

Ergebnisse IV
35
Abbildung 1 Die Pankreas-spezifische Inaktivierung von IκBα mittels des Cre/loxP Systems (A) Genkonstrukt: zwei gleichgerichtete loxP Sequenzen (hellblauer Pfleil) umfassen den Promoter (blauer Kreis), Exon 1 und 2 (rotes Rechteck). Im deletierten Gen (ikba
Δpanc) bleibt eine loxP-site
zurück. Pfeil: PCR Primer Ansatzstelle (B) PCR von ikba: Darstellung eines homozygot gefloxten (F/F), eines heterozygot gefloxten (F/wt) und eines wt (wt/wt) Genotyps (C) PCR von Ptf1acreex1
Darstellung einer positiv (+) und einer wt (-) Kontrolle; (D) Western Blot Analyse vom IκBα und IκBβ aus Proteinlysaten von isolierten Azinuszellen (E) Imumfluoreszenz zur Darstellung von IκBα im Pankreas; rot (Alexa Fluor 568) anti-IκBα, grün (Sytox green) Zellkerne;
1.2 Die pankreas-spezifische Deletion von IκBα führt zur nukleären
Translokation und transkriptionellen Aktivität von NF-κB
Intaktes IκBα verhindert die nukleäre Translokation von RelA/p65. Im weiteren
Verlauf wurden die Auswirkungen der IκBα-Blockade auf das Verhalten von
RelA/p65 untersucht
Hierfür wurden Azini aus dem Pankreas von unbehandelten ikbaF/F- und ikbaΔpanc
-
Mäusen separiert und anhand von Western Blot Analysen p50 und RelA/p65
untersucht. Hierbei zeigt sich für p50, das mit p65 ein Heterodimer bildet, sowohl in
ikbaΔpanc
-Mäusen als auch in ikbaF/F–Mäusen eine gleich starke Proteinexpression
ikbaF/F ikbaΔpancIκBα
E
IκBα
IκBβ
ikbaF/F ikbaΔpanc
D
ikba
F/F wt/wt
F/wt
B
Ptf1acreex1
+ -
C
5'
5'
3'
3'
ikba flox Allel
ikba Δ Allel
1 2 3 4 5A

IV Ergebnisse
36
bei 50 kDa. Für RelA/p65 hingegen stellt sich in ikba-defizienten Mäusen bei 65 kDa
eine schwächere Bande dar. Dies deckt sich mit der Western Blot von
phosphorylierten, d.h. aktiviertem RelA/p65 (pp65). Der verwendete Antikörper bindet
spezifisch an p65, das von der Proteinkinase A an Serin 276 phosphoryliert wurde. In
den unstimulierten Azinilysaten der ikbaΔpanc
-Maus kann pp65 als schwache Bande
detektiert werden, in den Azinilysaten der ikbaF/F-Maus hingegen ist keine Bande
nachweisbar. Dies ist ein Hinweis, dass durch die Abwesenheit von IκBα die NF-κB-
Dimere im Ruhezustand der Zelle zum Teil aktiv sind und in den Nukleus der
Azinuszelle eintreten können.
Abbildung 2B zeigt Pankreasgewebe von ikbaΔpanc
-Mäusen, das gegen RelA/p65 und
gegen Sytox Green, einen Nukleus-spezifischen Antikörper, gefärbt wurde. In der
Immunfluoreszenz ist RelA/p65 in einigen azinären Zellen sowohl im Nukleus als
auch im Zytosol des Pankreas lokalisierbar, während im endokrinen Kompartment,
dass heißt in den Zellen in denen IκBα funktionsfähig ist, RelA/p65 nur im Zytosol
nachzuweisen ist.
Zur weiteren Klärung, ob das nukleäre RelA/p65 nun auch transkriptionell aktiv ist,
wurden Genexpressionsanalysen durchgeführt. Hierfür wurde das Genom der
ikbaΔpanc
-Maus und der relaΔpanc
-Maus jeweils mit dem Wildtyp verglichen. Bei den
relaΔpanc
-Mäusen wurde analog wie bei den ikbaΔpanc
-Mäusen unter zu Hilfenahme
des Cre/loxP-Systems RelA/p65 im Pankreas deletiert (Algul et al., 2007). Hierfür
wurden aus den unstimulierten Azini von ikbaΔpanc
-, ikbaF/F
-, relaΔpanc
- und relaF/F
-
Mäusen die RNA extrahiert, mittels des Affymetrix Genchip aufgeschlüsselt und
durch die GSEA Software unter Verwendung eines Gensets, dass NF-κB abhängige
Gene enthält, analysiert. Wie in Abbildung 2C zu sehen zeigt sich im Vergleich zum
Wildtyp für die ikbaΔpanc
-Maus eine signifikante Hochregulation RelA/p65 abhängiger
Gene, während in der relaΔpanc
-Maus die Expression NF-κB abhängiger Gene im
Vergleich zum Wildtyp signifikant reduziert ist. Somit konnte gezeigt werden, dass
bereits in Ruhe durch die Deletion von IκBα RelA/p65 in den Nukleus eintreten kann
und transkriptionell aktiv ist.

Ergebnisse IV
37
Abbildung 2: Nukleäre Lokalisation von RelA/p65 im Pankreas von IκBα defizienten Mäusen. (A) Western Blot von nukleären und Cytosol-Extrakten isolierter Azini mit Antikörpern gegen IκBα, IκBb, p65 und phospho-p65. (*) Nukleus. (B) Fluoreszenzfärbung von Pankreasgewebe ikba
Δpanc -
Maus wurde gegen p65 floureszenz gefärbt. Sytox (Grün) Zellkerne, Rot /Alexa 568) anti- p65, Orange nukleäres p65. Die gestrichelte weiße Linie : Langerhansche Insel (C) GSEA: Die RNA des Pankreas von 3 Monate alten ikba
Δpancund ikbaF/F-Mäusen wurde extrahiert. Die Analyse der
Affymetrix Expression Array Daten mittels der GSEA-Software zeigt eine signifikante Hochregulation einer RelA Signatur in ikba
Δpanc und eine signifikante Unterexpression in relaΔpanc Proben (jeweils im Vergleich zum Wildtyp) 2 IκBα-defiziente Mäuse entwickeln ein unauffälliges Pankreas
Um herauszufinden, ob die Deletion von IκBα Auswirkung auf die Entwicklung des
Pankreas und die nicht azinären Anteile des Pankreas hat, wurde die Morphologie
der Pankreata von 3 und 13 Monate alten Mäusen untersucht.
Wie in Abbildung 3A zu sehen ergeben sich weder in den jüngeren noch in der
älteren Pankreata morphologische Unterschiede zwischen den Vergleichsgruppen.
Vor allem ist darauf hinzuweisen dass sich in den älteren IκBα-defizienten Mäusen
kein Hinweis auf einen chronisch entzündlichen Prozess, Degeneration der Azini und
Fibrose des Pankreas ergibt.
Des Weiteren wurde untersucht, ob sich Auswirkungen der Deletion von IκBα auf die
nicht azinären Anteile des Pankreas, d.h. die Langerhansschen Inseln und duktale
ikba
Δpanc
p65 mergeSytox
B
IκBαp50
p-p65
Acini
ikbaF/F ikbaΔpanc
Cytoplasma
*
A
p650
1
2
-2
-1
relaΔpanc
GSEA-NES
score
*
*C ikbaΔpanc

IV Ergebnisse
38
Anteile, ergeben. Hierfür wurden immunhistochemische Färbungen mit Antikörpern
gegen Insulin, Glukagon und CK 8, einem Cytokeratin charakteristisch für duktale
Zellen, durchgeführt (Abbildung 3B). Auch hier ergeben sich keine Unterschiede in
den ikbaΔpanc
- und ikbaF/F
–Mäusen. So kann man zusammenfassend sagen, dass die
Abwesenheit von IκBα und damit vermehrte Aktivität von RelA/p65 keinen Einfluss
auf Pankreasentwicklung und auf die extra-azinären Anteile des Pankreas
Abbildung 3 ikbaΔpanc –Mäuse entwickeln ein phänotypisch unauffälliges Pankreas
(A) Morphologischer Vergleich des Pankreas H&E Färbung des Pankreas 3 und 13 Monate alter Mäuse, Vergrößerung 1:100; (B) Darstellung der endokrinen und duktalen Anteile des Pankreas, IHC Färbung gegen Insulin, Glukagon und CK 8 (duktale Zellen), Vergrößerung 1:100;
3 Die Inhibition von IκBα mildert den Verlauf der akuten experimentellen Pankreatitis
In ikbaΔpanc
- und ikbaF/F
-Mäusen erfolgte die Induktion einer akuten experimentellen
Pankreatitis sowohl mit Caerulein als auch mit L-Arginin. Zur Beurteilung und
Vergleich der beiden Modelle wurden morphologische Analysen anhand von
Gewebeschnitten (Ödem-Entwicklung, Nekroserate, Inflamation) sowie
laborchemische Verfahren (Serumwerte für Amylase und Lipase) angewendet.
ikbaF/F ikbaΔpanc
H&E
3Monate
13Monate
A B
Insulin
Glukagon
CK8
ikbaF/F ikbaΔpanc

Ergebnisse IV
39
3.1 Die Caerulein Pankreatitis
Abbildung 4A zeigt H&E Färbungen des Pankreas zu den Zeitpunkten 0h, 8h und
24h nach Induktion einer Caerulein Pankreatitis. Während die Pankreata im
Ruhezustand mikroskopisch (H&E gefärbte Schnitte) keine signifikanten
Unterschiede aufwiesen, zeigten sich nach 8 bzw. 24h in den Pankreata der
ikbaΔpanc
-Mäusen weniger Ödem, Nekrosen und eine geringere Anzahl an
eingewanderten Entzündungszellen als in ikbaF/F
-Mäusen.
Ein messbarer Parameter für das Pankreasödem ist das relative Pankreasgewicht,
der Quotient aus dem Pankreasgewicht und dem Körpergewicht der entsprechenden
Maus. In unstimulierten Mäusen unterscheidet sich das relative Gewicht der beiden
Gruppen nicht voneinander (Abb 4B). Nach Induktion einer akuten Pankreatitis steigt
nach 8h das Pankreasgewicht beider Gruppen im Vergleich zum Ausgangsgewicht
an. Das relative Pankreasgewicht der Kontroll–Mäuse ist nach 8h aber signifikant
höher als das der ikbaΔpanc
-Mäusen (p<0,05). Somit entwickeln die ikbaF/F
–Mäuse in
dem verwendeten Model signifikant mehr Ödem im Pankreas.
Mit Hilfe eines Histoscores (siehe Methodenteil) lassen sich das Ausmaß der
Nekrose und der Infiltration von inflammatorischen Zellen semiquantitiv darstellen.
Abb. 4C zeigt eine signifikant höhere Nekroserate der Pankreata der ikbaF/F
-Mäuse
zum 8h Zeitpunkt wie auch einen signifikant höheren Anteil an inflammatorischen
Zellen im Vergleich zu den Pankreata der ikbaΔpanc
-Mäuse.
Auch im Serum spiegeln sich diese Unterschiede wieder. Den Mäusen wurden zu
den Zeitpunkten 0h, 4h und 8h Blut aus der Schwanzspitze entnommen und Amylase
und Lipase im Serum als Parameter für zelluläre Schäden bestimmt. Wie in den
Abbildungen 4D und E zu sehen ist, lässt sich im Ruhezustand kein Unterschied
feststellen, sowohl Lipase als auch Amylase sind in geringen Mengen im Serum
nachweisbar. Bereits sehr früh, 4h nach Induktion einer AEP steigen die Enzyme im
Serum beider Mausgruppen an. In ikbaF/F
-Mäusen sind nach 4h und 8h die Werte für
Amylase signifikant höher als bei den ikbaΔpanc
-Mäuse.
In Zusammenschau der Ergebnisse ließ sich feststellen, dass durch die Inhibition von
IκBα die lokalen Organschäden und entzündlichen Veränderungen im Pankreas
induziert durch Caerulein geringer ausfallen. Die akute experimentelle Pankreatitis
verläuft in ikbaΔpanc
-Mäusen milder.

IV Ergebnisse
40
Abbildung 4: Die Caerulein–Pankreatitis verläuft in ikbaΔpanc – Mäusen milder
(A) Histologische Schnitte des Pankreas von ikbaF/F und ikbaΔpanc
–Mäusen zu den Zeitpunkten 0h, 8h und 24h nach Induktion von Caerulein mit 50 µg/kg/KG, H&E Vergrößerung 1:200; (B) Vergleich des relativen Pankreasgewichts (Pankreasgewicht/Körpergewicht) als Maß für Ödem-Entwicklung im Pankreas zu den Zeitpunkten 0h und 8h, (n=4) (C) Semiquantitative Darstellung von Inflamamtion (=Ausmaß der Infiltration von entzündlichen Zellen) und Nekrose zum Zeitpunkt 24h, (n=4 pro Gruppe); (D und E) Serologischer Nachweis von Amylase und Lipase im Blut zu den Zeitpunkten 0h, 4h und 8h nach Induktion von Caerulein, als Parameter für den Azinuszellschaden * p<0,05, ** p<0,005;
24 h
H&E
0 h 8 h
ikba
F/F
ikba
Δpanc
0
0.005
0.01
0.015
0.02
0.025 *
0h 8h
*
RelativesPankreasgewicht
ikbaF/FikbaΔpanc
D
5000
10000
15000
20000
25000
30000
0
Serum-AmylaseU/ml
****
ikbaF/FikbaΔpanc
0h 4h 8h
B
500
1000
1500
2000
2500
3000
Serum-LipaseU/ml **
**
ikbaF/FikbaΔpanc
0
0h 4h 8h
CInflammation
0
0.5
1
1.5
2
2.5
3
24h
Nekrose
0
0.5
1
1.5
2
2.5 *
24h
**
ikbaF/FikbaΔpanc
E
A

Ergebnisse IV
41
3.2 Die L-Arginin-Pankreatitis
Auch in einem anderen Model der akuten Pankreatitis, der L-Arginin-Pankreatitis,
ließen sich die beschriebenen Unterschiede beobachten, jedoch zu einem späteren
Zeitpunkt als im Vergleich zur Caerulein-Pankreatitis. Die L-Arginin-Pankreatitis
wurde durch die zweimalige Injektion von L-Arginin induziert. Zu den Zeitpunkten 0h,
24h, 48h und 72h wurden den Mäusen Blutproben aus der Schwanzspitze
entnommen, nach Ablauf von 72h wurden die Mäuse getötet und die Pankreata nach
ihrer Morphologie analysiert. Wie in Abbildung 5A zu sehen, zeigten die Pankreata
der ikbaΔpanc
-Maus weniger entzündliche Veränderungen und weniger
Gewebsschäden. In den Serumproben wurden Amylase und Lipase bestimmt. Nach
72h sind die Serum-Level für Amylase und Lipase in den ikbaF/F
-Mäusen signifikant
höher als in der ikbaΔpanc
-Maus. Bereits nach 24h und 48h zeigen sich erst
Unterschiede, jedoch sind diese nicht signifikant. Daraus ergibt sich, dass die in der
Caerulein-Pankreatitis beobachteten Veränderungen auch auf andere Pankreatitis-
Modelle übertragbar sind.
Abbildung 5 Die L-Arginin Pankreatitis in ikbaΔpanc –Mäusen zeigt einen attenuierten Verlauf
(A) Histologische Schnitte des Pankreas 72h nach Gabe von insgesamt 8 mg L-Arginin, H&E Vergößerung 1:100; (B und C) Serologischer Nachweis von Amylase und Lipase im Blut zu den Zeitpunkten 0h, 24h, 48h und 72h nach Induktion (n=5).
ikbaF/F ikbaΔpanc
L-Arginin-Pankreatitis 72hA
****
**
ikbaF/FikbaΔpanc
0
100
200
300
400
500
600
Serum-LipaseU/ml
0h 24h 48h 72h
B
**
0h 24h 48h 72h0
2000
4000
6000
8000
10000
12000
14000
Serum-AmylaseU/ml
ikbaF/FikbaΔpanc
C

IV Ergebnisse
42
4 Die Inhibition von IκBα reduziert die intrapankreatische Trypsinaktivierung während der Pankreatitis
Ein Element in der Pathophysiologie der akuten Pankreatitis ist die intra-azinäre
Trypsinaktivierung. Es wurde untersucht, ob in der ikbaΔpanc
-Maus die Aktvierung von
Trypsin im Pankreas während der AEP beeinflusst wird. Die Bestimmung der
Enzymaktivität erfolgte fluorometrisch im Pankreashomogenisat von ikbaΔpanc
- und
ikbaF/F
-Mäusen im Ruhezustand und 1h bzw 8h nach Induktion einer Caerulein-
Pankreatitis. Im unstimulierten Pankreas ergaben sich keine Unterschiede. Es ist in
beiden Fällen eine geringe Aktivität nachweisbar. Da die Trypsinaktivierung zu den
frühesten Ereignissen der AEP gehört, ist schon nach 1h Stunde ein starker Anstieg
erreicht. In den Kontrollmäusen hat sich nach 1h die Aktivität im Vergleich zum
Ausgangswert im Durchschnitt verdreifacht. Im Pankreas der ikbaΔpanc
-Maus ist nach
1h ebenfalls eine Aktivierung von Trypsin nachweisbar, diese ist aber signifikant
niedriger als in der ikbaF/F
-Mäusen. Nach 8h ist es in beiden Fällen zu einem
Rückgang der Trypsinaktivität gekommen, im Pankreas der ikbaF/F
-Mäuse ist aber
weiterhin eine signifikant höhere Aktivität nachweisbar.
Damit konnte bewiesen werden, dass durch eine azinäre Inhibition von IκBα die intra-
azinäre Trypsinaktivierung während der akuten Caerulein-Pankreatitis reduziert wird.
Abbildung 6 Die Inhibition von IκBα reduziert die intrapankreatische Trypsinaktivierung während der Caerulein-Pankreatitis Fluorometrische Aktivitätsbestimmung von Trypsin aus Pankreas-Homogenisaten zu den Zeitpunkten 0h, 1h und 8h nach Induktion einer Caeruleinpankreatitis; aktives Trypsin kann im Pankreas IκBα defizienter Mäuse signifikatifikant weniger nachgewiesen werden. * p<0,05, ** p<0,005;
TrypsinAktivität(fmol/mg)
0
500
1000
1500
2000ikbaF/Fikba!panc
****
0h 1h 8h

Ergebnisse IV
43
5 Die Pankreatitis-assoziierten Komplikationen werden durch die Inhibition von IκBα attenuiert
Ein serologischer Parameter und frühzeitiger Indikator für die schwere akute
Pankreatitis und extrapankreatische Komplikationen ist IL-6 (Dambrauskas et al.,
2010). Während einer akuten experimentellen Pankreatitis kommt es nicht nur zu
lokalen Veränderungen im Pankreas, sondern der gesamte Organismus wird
involviert. Der Hauptmanifestationsort der extrapankreatischen Komplikationen ist die
Lunge. Zur Beurteilung des Schweregrads der extrapankreatischen Komplikation
wurde IL-6 im Serum gemessen und die inflammatorischen Veränderungen in der
Lunge analysiert.
5.1 IL-6 – ein serologischer Marker für die schwere AEP
Mittels eines Enzym Linked Absorbtion Essays (ELISA) wurde IL-6 im Serum
quantitativ gemessen. Die Serumkonzentrationen von IL-6 in 3 Monate alten Mäusen
zu den Zeitpunkten 0h, 8h und 24h nach Ablauf einer Caerulein induzierten
Pankreatitis (Abbildung 7) wurden miteinander verglichen. Zu Beginn unterscheidet
sich die Level von IL-6 in ikbaΔpanc
und ikbaF/F
-Mäusen nicht voneinander und liegt
auf einem niedrigen Niveau. Nach 8h jedoch ist in beiden Mauslinien die
Konzentration von IL-6 angestiegen. Dabei zeigten sich signifikant höhere Il-6
Spiegel in den ikbaF/F
-Mäuse als in den ikbaΔpanc
-Mäusen. Nach Ablauf von 24h ist
die Konzentration von IL-6 in beiden Gruppen wieder abgesunken.
Somit wird die Induktion von IL-6 durch die Inhibition von IκBα abgeschwächt..

IV Ergebnisse
44
Abbildung 7 IL-6 – ein serologischer Marker für die schwere AEP Nachweis von IL-6 im Serum mittels ELISA; Das Ausmaß der IL-6 Expression korreliert positiv mit dem Schweregrad der akuten Pankreatitis IL-6 wird in ikba
Δpanc–Mäusen in der Frühphase der akuten
Caerulein-Pankreatitis signifikant weniger exprimiert. * p<0,05, ** p<0,005
5.2 Systemische Komplikation der AEP am Beispiel der Lunge
Zur Beurteilung des Schweregrads der Lungenveränderungen erfolgten
morphologische Analysen an H&E gefärbten Lungengewebsschnitten und die
Bestimmung der Myeloperoxidase – Aktivität (MPO) im Lungengewebe als
Parameter für das Ausmaß der Leukozyteninfiltration.
Zum Nachweis, dass das Lungengewebe nicht von der ikba-Deletion betroffen ist,
wurden Western Blot Analysen aus Proteinlysaten des Lungengewebes der beiden
Mauslinien durchgeführt. Wie in Abb. 8A zu sehen ist, wird das Protein IκBα in der
Lunge von ikbaF/F
- und ikbaΔpanc
-Mäusen regulär exprimiert,
Abb 8B zeigt H&E Färbungen der Lungen unstimulierter Mäuse, sowie nach Ablauf
von 24h nach Induktion der AEP in der Übersicht bei 100x und detaillierter in 200x
Vergrößerung. Der 24h Zeitpunkt wurde gewählt, da systemische Veränderungen
einer AEP sich erst zu einem späteren Zeitpunkt manifestieren.
In unstimulierten Tieren zeigten sich normale Lungenmorphologien. 24h nach
Induktion einer Pankreatitis sind die Lungen der Kontrollmäuse deutlich entzündlich
verändert, die Alveolarsepten verdickt, das Gewebe ödematös und hämorrhagisch
imbibiert. Auch im Lungengewebe der ikbaΔpanc
-Linie zeigten entzündliche
Veränderungen im Vergleich zum Ausgangsbefund zu erkennen, diese waren aber
deutlich schwächer als bei den ikbaF/F
-Mäusen. Zur Quantifizierung der
0
10
20
30
40
50
60
70
80
90
1
Serum-IL-6pg/ml
**
0 h 8 h 24 h
ikbaΔpancikbaF/F

Ergebnisse IV
45
mikroskopischen Befunde diente die spektrometrische Bestimmung der MPO-
Aktivität im Lungengewebe, ein Enzym spezifisch für neutrophile Granulozyten. Die
Höhe der gemessenen Aktivität korreliert mit dem Ausmaß der Leukozyteninfiltration.
(Abb 8C) Nach 8h ergaben sich noch keine signifikanten Unterschiede der MPO-
Aktivität, auch ist die Aktivität noch auf einem relativ niedrigen Niveau. 24h nach
Ablauf der akuten Pankreatitis kommt es in der Lunge ikbaF/F
–Mäuse zu einem 4-
fachen Anstieg der Aktivität im Vergleich zum 8h Wert. Die Lungen der ikbaΔpanc
-
Mäusen wurden im Vergleich weit weniger stark von Neutrophilen infiltriert. 24 nach
Induktion der AEP ist die MPO – Aktivität ebenfalls angestiegen, aber signifikant
geringer als in der Kontrollgruppe.
Diese Befunde ergeben, dass durch die Deletion von IκBα im Pankreas, der Ablauf
der akuten Pankreatitis nicht nur lokal beeinflusst, sondern auch die
extrapankreatischen Komplikationen reduziert wurden.
Abbildung 8 Die pankreas-spezifische Inhibition von IκBα mildert die Pankreatitis assoziierten Komplikationen in der Lunge (A) Nachweis der regulären IκBα Expression in der Lunge; Western Blot Analyse von Lungenzell-Lysaten (20 µg) von ikba
Δpanc und ikbaF/F-Mäusen; (B) Paraffinschnitte von Lungengewebe zu den
Zeitpunkten 0h und 24h nach Caerulein-Injektion, Es zeigt sich ein geringeres Ausmaß an entzündlichen Veränderungen in der ikba
Δpanc-Maus, H&E 1:200; (C) MPO-Aktivität: Zu den
Zeitpunkten 8h und 24h wurde die Myeloperoxidase-Aktivität (ΔOD/min/µg Protein) im Lungengewebe photometrisch bestimmt. (n=4);* p<0,05, ** p<0,005; (n=4 pro Mauslinie)
24h
0h
ikbaΔpancikbaF/F
B
ikbaF/F ikbaΔpanc
LungeIκbα
n.n.
A
0
20
60
80
100
8h 24h
**
MPO-AktivitätΔA/min/pg ikbaF/F
ikbaΔpanc
40
120C

IV Ergebnisse
46
6 Der protektive Effekt in ikba-defizienten Mäusen ist abhängig von nukleärem RelA
Die bisherigen Untersuchung zeigten eine mild verlaufende Pankreatitis in ikba
knockout Mäusen. Im nächsten Schritt gilt es festzustellen, ob die beobachteten
Veränderungen auf die konstitutive nukleäre Aktivität von RelA/p65 zurückzuführen
sind. Hierfür induzierten wir eine Pankreatitis in ikba p65 Doppel-Knockout Mäusen.
Um Doppel-Knockout Mäuse (rela x ikbaΔpanc
) zu erhalten, wurden die ikbaΔpanc
-
Mäuse mit relaF/F
–Mäusen verpaart (Algul et al., 2007). Im Genom der relaF/F-Maus
sind die Exons 7-10 von loxp Sites flankiert. Diese Exons codieren Teile der RHD.
Das deletierte RelA/p65 ist nicht in der Lage in den Nukleus einzutreten und
transkriptionell aktiv zu werden. Abbildung 10 zeigt das Genkonstrukt der relaF/F-
Maus.
Der Pankreas-spezifische Knockout von IκBα und RelA/p65 wurde durch Western
Blot Analysen an isolierten unstimulierten Azinuszellen der verschiedenen Mauslinien
verifizert. In den relaΔpanc
-Mäusen zeigt sich die deletierte Form von p65 auf Höhe
von 42 kDA, Wildtyp p65 kann nicht nachgewiesen werden. ß-Actin diente als
Ladungs kontrolle.
Es erfolgten in den 3 Mausgruppen analoge akute Caerulein-Pankreatitis-Studien
über 24 h wie vorbeschrieben, Analysen zur Morphologie, der Grad der Inflammation
und Nekroserate und die Konzentration von Amylase im Serum wurden bestimmt.
Abbildung 9B zeigt H&E Färbungen des Pankreas der ikbaF/F
-, der ikbaΔpanc
- und der
rela x ikbaΔpanc
-Maus nach Ablauf von 24h nach Induktion der AEP. Der Pankreas
der ikbaΔpanc
-Maus weist wie vorbeschrieben nur geringe morphologische
Veränderungen auf, das Parenchym ist kompakt, das Gewebe der Kontrollmaus und
der rela x ikbaΔpanc
-Maus hingegen sind deutlich ödematös verändert. Das Ausmaß
der Nekrose und der entzündlichen Veränderungen wurde wieder mit Hilfe eines
Histoscores beurteilt. 24h nach Beginn der AEP ist die Nekrose Rate im Pankreas
der rela x ikbaΔpanc
-Maus signifikant höher als in der ikbaΔpanc
-Maus (Abbildung 9D)
und auch im Vergleich zur Kontrollmaus ist die Nekroserate leicht erhöht. Analog
verhält sich die Entzündungsrate. Analog verhielten sich die Messergebnisse im
Serum. Die Konzentrationen für Amylase in allen 3 Mauslinien sind im Ruhezustand

Ergebnisse IV
47
annähernd gleich. Nach 8h jedoch kommt es in jedem Fall zu einem Anstieg der
Amylasekonzentration, in der rela x ikbaΔpanc
-Maus aber ist dieser am höchsten und
signifikant höher als in der ikbaΔpanc
-Mauslinie (Abbildung 9C).
Zusammengefasst verläuft die AEP in rela x ikbaΔpanc
-Mäusen signifikant schwerer
als in der ikbaΔpanc
-Maus und tendenziell schwerer als in den Kontrollmäusen, deren
NF-κB-Signalweg in den Azinuszellen regulär funktionsfähig ist. Diese
Beobachtungen lassen den Schluss zu, dass die protektiven Effekte während der
AEP in ikba-defizienten Mäusen auf die nukleäre Aktivität von RelA zurückzuführen
ist.
Abbildung 10: Das relaΔpanc Genkonstrukt 2 gleichgerichtete loxP-Sites (hellblaus Dreieck) flankieren die Exons 7-10 (rote Rechtecke). In diesem Genabschnitt ist u.a. die RHD enthalten Durch die Cre-Rekombinase wird dieser Bereich deletiert eine loxP-Site bleibt zurück (rela Δ Allel). BglII: Ansatzstelle für das Restriktionsenzym BglII
rela flox Allel
rela Δ Allel
5'
5'
BglII BglIIBglII
BglII BglII
6 7 8 9 10

IV Ergebnisse
48
Abbildung 9: Die Coinhibiton von IκBα und RelA/p65 führt zu einer schwer verlaufenden Pankreatitis (A) Western Blot Analyse von isolierten Azini der 3 Mausllnien, Verwendung von Antikörpern gegen IκBα und p65, β-Actin als Kontrolle, Bei rela x ikba
Δpanc-Mäusen ist anstelle von p65 nur eine
H&E
ikba!panc rela!panc x ikba!pancikbaF/F
24 h
B
I"B#
p65
!p65$-actin
ikbaF/F
ikba!panc
0
5000
10000
Serum-AmylaseU/ml
15000
20000
**
0h 8h
0
Inflammation
1
2
3 *
*
24h0
1
2
3
Nekrose
*
*
24h
ikba!pancikbaF/F
rela x ikba!panc
A
C
D
rela
x ikba!panc

Ergebnisse IV
49
trunkierte Form (Δp65) nachweisbar; (B) Paraffinschnitte des Pankreas der 3 Mauslinien zum Zeitpunkt 24h nach Induktion einer Caerulein-Pankreatitis (50 µg/kg/Kg), H&E Vergrößerung 1:100;(C) Messung der Serum-Amylase Konzentration (U/ml) im Blut zu den Zeitpunkten 0h und 8h nach Induktion von Caerulein; (D) Semiquantitative Darstellung von Inflammation und Nekrose mittels Histoscore (siehe Material und Methoden);* p<0,05, ** p<0,005; (n=4 pro Mauslinie);

V Diskussion
50
V. Diskussion
Die Pathophysiologie der akuten Pankreatitis wird bereits seit vielen Jahren erforscht
und relativ bald kristallisierte sich heraus, dass der IKK/NF-κB/Rel Signalweg den
Verlauf der akuten Pankreatitis von Beginn an beeinflusst. NF-κB gehört zu den
ersten Signalkaskaden, die während der akuten experimentellen Pankreatitis in den
Azinuszellen induziert werden (Gukovsky et al., 1998). Seine Schlüsselrolle als
allgemeiner proinflammatorischer Transkriptionsfaktor, Regulator von Apoptose und
Kanzerogenese ist hinreichend bekannt (Perkins, 2000).
Da durch die Blockade NF-κB abhängiger Proteine u.a. von TNF-α und IL-1 eine
milder verlaufende Pankreatitis beobachtet werden konnte, verwundert es nicht, dass
man NF-κB a priori eine proinflammatorische Funktion im Pankreas zuschrieb (Algul
et al., 2002). Dies lies sich initial auch in einer Vielzahl von Arbeiten bestätigen.
Durch pharmakologische Inhibierung oder auch transgene Aktivierung von NF-κB
konnte ein schwer verlaufende Pankreatitis beobachtet werden.
In Gegensatz dazu stehen die Ergebnisse der Arbeit von Algül et al, die gleichzeitig
auch den direkten Vorläufer zu der vorgestellten Arbeit darstellt. Algül et al konnten
durch Generierung einer pankreas-spezifischen RelA/p65 Knockout Maus zeigen,
dass die Caerulein induzierte Pankreatitis in diesen Mäusen schwerwiegender
verläuft und sowohl die systemischen als auch die lokalen Komplikationen
ausgeprägter sind (Algul et al., 2007).
Die in dieser Arbeit vorgestellten Ergebnisse weisen nach, dass durch die Inhibition
von IκBα und der damit verbundenen endogenen Aktivität von NF-κB die
Azinuszellen während der akuten Pankreatitis vor entzündlichen Schäden geschützt
sind.
Dieses Resultat steht nun im Widerspruch zu den bisherigen Forschungsarbeiten,
die besagen, dass NF-κB entscheidend für die Induktion von Entzündung und
Gewebsschäden während der AEP ist. Wodurch ergeben sich diese Unterschiede?
Zunächst muss daraufhin gewiesen werden, dass die inflammatorischen Vorgänge
während der Pankreatitis einem komplexen Zusammenspiel zwischen Immunzellen
und Azinuszellen unterliegen (Vonlaufen et al., 2007). Bereits an anderer Stelle im
Bereich der Epidermis zeigte sich, dass durch die organ-spezifische Deletion von
IκBα keine lokale Entzündung induziert werden kann. Erst die gleichzeitige Inhibition

Diskussion V
51
von IκBα in epidermalen und inflammatorischen Zellen führte zu entzündlichen
Veränderungen (Rebholz et al., 2007).
Im Weiteren muss man die vorangegangenen Arbeiten genauer analysieren. In den
ersten experimentellen Studien, die sich mit der Bedeutung von NF-κB in der
Pathogenese der akuten Pankreatitis beschäftigten, arbeitete man mit
pharmakologischer Inhibierung von NF-κB. In den meisten Fällen zeigte sich durch
die so erreichte Reduktion der Aktivierung von NF-κB eine milder verlaufende
Pankreatitis. Aber gleichzeitig brachte die pharmakologische Inhibition von NF-κB
einige Limitierungen mit sich. So erwiesen sich die häufig verwendeten Inhibitoren
NAC und PDTC als nicht NF-κB-spezifisch (Rakonczay et al., 2008), des weiteren
konnte eine organ- und zellspezifische Inhibition nicht gewährleisten werden. So
ergibt es sich, dass durch Verwendung der gleichen Inhibitoren ganz
unterschiedliche Ergebnisse erzielt wurden. So sahen z.B. Steinle et al im Gegensatz
zu Gukovsky et al und Satoh et al eine schwerer verlaufende Pankreatitis durch
Verwendung von NAC und PDTC (Steinle et al., 1999, Satoh et al., 1999, Gukovsky
et al., 1998). Deshalb sind die meisten Forschungsarbeiten, aufgrund der
Verwendung relativ unspezifischer Inhibitoren, den unterschiedlichen
Applikationsformen und der geringen Zellspezifität nur eingeschränkt aussagekräftig.
Daraufhin ging man dazu über, die Funktion von NF-κΒ auf genetischer Ebene direkt
zu untersuchen. Nur so kann gewährleistet werden, dass die beobachteten
Veränderungen während der akuten Pankreatitis tatsächlich mit der Aktivität von NF-
κB zusammenhängen. 2003 wiesen Altavilla et al eine milder verlaufende
Pankreatitis in p105/p50 defizienten Mäusen nach (Altavilla et al., 2003). Es handelte
sich aber hierbei nicht um Pankreas-spezifische knockout Mäuse, sondern es wurde
ein systemischer Knockout verwendet. Somit ist nicht unterscheidbar, ob die
beobachteten Ergebnisse auf die Aktivität von NF-κB in Azinuszellen oder aber in
inflammatorischen Zellen zurückzuführen sind.
Aleksic und Baumann zeigten durch Verwendung transgener Mäuse, die eine durch
Doxycyclin induzierbare Pankreas-spezifische konstitutive Aktivierung von IKKβ
aufwiesen, dass die so induzierte NF-κB/RelA–Überexpression zu vermehrten
Gewebeschäden während der akuten Pankreatitis führt. Auch konnten sie
beobachten, dass in ihrem Modell die Aktivierung von IKKβ allein ausreicht, um eine
Entzündung im Pankreas zu induzieren. (Baumann et al., 2007, Aleksic et al., 2007)
Zu einem ähnlichen Ergebniss gelangte die Forschergruppe um Chen et al, die durch

V Diskussion
52
einen adenoviralen Transfer von aktiven RelA/p65 über den Pankreashauptgang in
die Azinuszellen eine Pankreatitis auslösen konnten (Chen et al., 2002).
In dieser Arbeit nun wurden die Auswirkungen einer Inhibition von IκBα für die
Entwicklung des Pankreas und den Verlauf einer akuten Pankreatitis untersucht.
Dies wurde durch die Generierung einer konditionalen Pankreas-spezifischen ikba-
knockout Maus erreicht. Mit Hilfe des Cre/loxP Systems gelang die Deletion von Iκbα
in den Azinuszellen des Pankreas und somit eine konstitutive Aktivität von RelA
erzielt.
In den Western Blot Analysen und auch in der Immunfluoreszenzfärbung konnte die
Deletion von IκBα verifiziert werden. IκBβ dagegen blieb von der IκBα-Deletion
unbeeinflusst. Ein Hinweis, dass IκBβ nicht die Funktion von Iκbα ersetzt.
Dieser “loss of function“ Ansatz erlaubt die Imitation einer regulären NF-κB
Aktivierung im Gegensatz zu z.B einer transgenen Überexpression von NF-κB.
Mittels Immunfluoreszenz und Western Blot wurde nachgewiesen, dass RelA/p65
bereits im Ruhezustand in den Nukleus eintritt. Durch die GSEA Analyse der
pankreatischen mRNA bestätigte sich, dass NF-κB transkriptionell aktiv ist, da in der
ikbaΔpanc
-Maus NF-κB abhängige Gene im Vergleich zum Wildtyp signifikant
hochreguliert sind.
Bemerkenswert ist, dass sich dadurch keine Auswirkungen auf den pankreatischen
Phänotyp ergeben. Zu keinem Zeitpunkt wurden vermehrt entzündliche oder auch
degenerative Veränderungen im Vergleich zum Wildtyp gesehen. Auch die nicht
azinären Anteile des Pankreas blieben von der azinären IκBα Inaktivierung
unbeeinflusst. Es konnte also bewiesen werden, dass RelA/p65 nicht in der Lage ist
von sich aus eine akute Pankreatitis zu induzieren.
Im Gegensatz zu Chen et al., die durch eine viral induzierte Überexpression von
RelA/p65 eine Pankreatitis induzieren konnten, ließ sich deren Beobachtung in
unserem Modell nicht reproduzieren. Eine Erklärung für diese unterschiedlichen
Ergebnisse könnte sein, dass durch den adenoviralen Transfer von RelA/p65 über
den Pankreasgang nicht nur azinäre Zellen sondern auch inflammatorische Zellen
involviert wurden. Desweiteren könnten durch Verwendung von Viren allein
inflammatorische Prozesse aktiviert worden sein
Aleksic und Baumann wiederum gelang durch die konstitutive Aktivierung von
IKKβ eine Überexpression von NF-κB/RelA und sahen dadurch vermehrte zelluläre

Diskussion V
53
Schäden. Nun ist diese Art der transgenen Überexpression aber unphysiologisch und
die Stärke der IKKβ Aktivierung nur schwer zu regulieren (Rakonczay et al., 2008)
Auch die intra-azinäre Trypsinaktivierung wird durch die Inhibition von IκBα reduziert.
Die Trypsin-Aktivierung ist eines der frühesten und wichtigsten Ereignisse in der
Pathophysiologie der akuten Pankreatitis (Saluja et al., 2007). Die Tatsache, dass in
ikbaΔpanc
-Mäusen eine Reduktion der Trypsinaktivierung nach Induktion der akuten
Pankreatitis nachzuweisen ist, unterstreicht zum einen, dass NF-κB möglichweise
auf direkte Weise den Verlauf der AEP beeinflusst, zum anderen dass der IKK/NF-
κB/RelA Signalweg sehr früh im Verlauf der Pankreatitis aktiviert wird.
Ob ein Zusammenhang zwischen der frühzeitigen Aktivierung von Trypsin und NF-κB
besteht, war lange umstritten. Zudem herrschte Uneinigkeit darüber in welcher Form
sich beide beeinflussen könnten. Tando et al stellten fest, dass nach Blockade von
Trypsinogen mittels eines Serin-Protease Inhibitors der IKK/NF-κB/Rel Signalweg
durch CCK nicht aktiviert wurde und postulierten deshalb, dass Trypsinogen NF-κB
während der AEP aktiviert (Tando et al., 2002). Andere Forschergruppen sahen die
in beiden Ereignisse als unabhängig von einander (Han et al., 2001, Hietaranta et al.,
2001, Bi and Williams, 2004). Han et al zum Beispiel sahen in verschiedenen
Zellkulturexperimenten mit modifizierten Ovarialzellen und adenoviralen Transfer von
p65 in Azinuszellen keinen Zusammenhang zwischen der CCK abhängigen
Aktivierung von NF-κB und Trypsinogen. Nun handelte sich in keiner Arbeit um ein
endogenes Modell und die so gewonnene Ergebnisse unterliegen somit einigen
Limitierungen.
Dawra et al verwendeten erstmals ein konditionales trypsinogen-Knockout
Mausmodel und fanden so heraus, dass die Aktivierung von NF-κB während der
AEP im Pankreas von Trypsinogen nicht beeinflusst wird und dass Trypsinogen
verantwortlich für die ersten Schäden im Verlauf der AEP ist (Dawra et al., 2011).
Dies deckt sich mit den Ergebnissen unserer Arbeit. Man kann also den Schluss
ziehen, dass NF-κB unabhängig von Trypsinogen aktiviert wird und NF-κB die
Trypsinogen-Aktivierung abschwächt und somit protektiv in der Frühphase wirkt. Ob
NF-κB die Aktivierung von Trypsinogen direkt beeinflusst und wie der genaue
zeitlichen Zusammenhang abläuft, muss noch heraus gefunden werden.

V Diskussion
54
Auch auf systemischer Ebene bestätigte sich die protektiven Eigenschaften der
Inhibition von IκBα während der AEP. Die IL-6 Konzentrationen im Serum waren
signifikant reduziert und auch die Veränderungen in der Lunge vielen in den ikba-
knockout Mäusen milder aus. IκBα wurde in der Lunge der ikbaΔpanc
-Mäuse regulär
exprimiert. Dies zeigt zum einen, dass die Expression von IκBα in anderen Organen
durch den Knockout im Pankreas unbeeinflusst bleibt, d.h. dass es sich um einen
gewebsspezifischen Knockout handelt und zum anderen, dass die in der Lunge
beobachteten Veränderung auf die Inhibition von IκBα im Pankreas zurückzuführen
sind.
Um letztendlich sicherzustellen, ob die Versuchsergebnisse tatsächlich auf eine
vermehrte endogene Aktivität von RelA/p65 zurückzuführen sind, wurde der Verlauf
einer AEP in rela x ikbaΔpanc
-Mäusen untersucht. Durch Kreuzung der ikbaΔpanc
-Maus
mit einer relaF/F
-Maus wurden Mäuse generiert in denen sowohl IκBα als auch
RelA/p65 im Pankreas deletiert sind, das heißt die Azinuszellen verfügen nicht mehr
über den regulären IKK/NF-κB/Rel Signalweg.
Folgt man der Annahme, dass azinäres NF-κB ein entscheidender
proinflammatorischer Faktor während der AEP ist, müsste durch Deletion des
Signalwegs eine deutlich mildere Pankreatitis ablaufen. Das Gegenteil war der Fall.
Das Pankreas der Doppel-Knockout-Mäuse war nach Induktion einer akuten
Pankreatitis deutlich entzündlich verändert und dies signifikant stärker sowohl im
Vergleich zum Wildtyp wie auch zu den ikbaΔpanc
-Mäusen. Dies ließ sich sowohl
histologisch, als auch serologisch nachweisen. Die Inhibition von IκBα und RelA
führte zu einer schwer verlaufenden Pankreatitis. Dies beweist, dass die in der ikba-
knockout Maus beobachteten Veränderungen, der milde Verlauf einer akuten
experimentellen Pankreatitis, auf die nukleäre Aktivität von NF-κB/RelA
zurückzuführen sind.
Die große Widersprüchlichkeit dieses Ergebnisses im Vergleich zu den bisherigen
Forschungsarbeiten zeigt aber auch wie komplex und divers die Rolle des
ubiquitären Transkriptionsfaktors NF-κB ist. Die akute Pankreatitis spielt sich nicht
nur isoliert im Pankreas ab, sondern ist eine systemische Erkrankung. Involviert sind
Entzündungszellen, das Endothel, Leber und Lunge. Für ein umfassendes

Diskussion V
55
Verständnis ist es sicherlich notwendig, nicht nur die Funktion von NF-κB in der
Azinuszelle zu untersuchen, sondern unter anderem auch seine Aufgaben in
neutrophilen Granulozyten, die das Pankreas infiltrieren.
Für ein besseres Verständnis der Pathogenese der akuten Pankreatitis und der
Bedeutung von NF-kB gilt es im weiteren zu untersuchen über welche Mechanismen
der IKK/RelA/NF-kB Signalweg die akute Pankreatitis moduliert. Hierfür ist unter
anderem die Identifikation protektiver NF-κB abhängiger Gene notwendig. Mit der
Affymetrix Analyse der mRNA der relaΔpanc
und ikbaΔpanc
-Mäuse die Grundlage
geschaffen. Durch die weitere Auswertung dieser Daten wird die Identifikation von
Kandidaten-Genen möglich sein.
Zusammenfassend konnte gezeigt werden, dass die Inhibition von IκBα den Verlauf
der akuten experimentellen Pankreatitis abgemildert. Ursächlich hierfür ist die
vermehrte endogene Aktivität von NF-κB in der Azinuszelle. Die Beobachtung von
Chen et al, dass RelA/p65 allein in der Lage ist eine akute Pankreatitis zu induzieren,
konnte in unserem Modell nicht nachgewiesen werden. Zwar konnte gezeigt werden,
dass NF-κB bereits im Ruhezustand in der Azinuszelle aktiv ist, entzündliche
Veränderungen im Phänotyp ließen sich aber nicht nachweisen. Während der AEP
erwies sich die vermehrte Aktivität von NF-κB als protektiver Faktor, die
Ausschaltung des IKK/NF-κB/RelA Signalwegs führte zu schweren Schäden im
Pankreas während der AEP.
Die bislang gültige Annahme, dass NF-κB ein proinflammatorischer Faktor der
akuten Pankreatitis ist und eine vermehrte Aktivität in der Azinuszelle mit einer
schwer verlaufenden Pankreatitis verbunden ist, muss überdacht werden. Im
Gegenteil, es konnte gezeigt werden, dass NF-κB protektive und
antiinflammatorische Bedeutung hat. Eine wichtige Erkenntnis zum weiteren
Verständnis der Pathogenese der akuten Pankreatitis konnte somit entschlüsselt
werden.
Bislang gibt es keine gezielten therapeutischen Strategien zur Therapie einer akuten
Pankreatitis. Da die akute Pankreatitis nicht nur eine milde selbstlimitierende
Erkrankungen ist, sondern in einigen Fällen ein komplikationsreichen schweren
Verlauf nimmt, ist die Entwicklung therapeutischer Ansatzpunkte von besonderer
Bedeutung. Dafür ist das Verständnis der Pathogenese von entscheidender
Bedeutung. Mit der Erkenntnis, dass der IKK/NF-κB/RelA Signalweg protektive

V Diskussion
56
Eigenschaften während der AEP hat, können im weiteren vielleicht therapeutische
Strategien entwickelt werden.

Zusammenfassung VI
57
VI. Zusammenfassung
In dieser Arbeit wurden die Auswirkungen der Inhibition von IκBα auf den Verlauf
einer akuten experimentellen Pankreatitis untersucht. IκBα ist der Inhibitor von NF.kB
einem ubiquitär vorkommenden Transkriptionsfaktor. Seine Rolle in der akuten
Pankreatitis ist bislang stark umstritten und eine Vielzahl von Studien beschäftigen
sich mit diesem Thema.
Um die Rolle von aktivem NF-κB in der Frühphase der akuten Pankreatitis spezifisch
in den Azinuszellen untersuchen zu können, ist es notwendig auf Tiermodelle
zurückzugreifen. Durch Verwendung einer konditionellen pankreas-spezifischen ikba
Knockout Maus (ikbaΔpanc
) steht ein Tiermodell zur Verfügung, das eine endogene
Aktivierung von NF-κB in den Azinuszellen gewährleistet. In einem zweiten Ansatz
wurde der Verlauf einer akuten experimentellen Pankreatitis in ikba/rela
Doppelknock-Mäusen (rela x ikbaΔpanc
) untersucht, d.h. in Mäusen in denen sowohl
IκBα als auch RelA/p65 deletiert wird.
Zunächst verifizierten die Inhibition von IκBα auf Proteinebene durch Western Blot
Analysen und Immunfluoreszenz-Darstellung. Die Inhibiton von IκBα hat zur Folge,
dass NF-κB bereits im Ruhezustand in den Nukleus eintreten kann und dort aktiv ist.
Dies lies im Western Blot durch Nachweis von phosphorylierten p65, in der
Immunfluoreszenz durch Detektion von nukleär lokalisierten p65 und v.a in der
Genexpressionsanalyse darstellen, die zeigte, dass in den ikbaΔpanc
-Mäusen NF-κB
abhängige Gene signifikant hochreguliert sind. Dadurch ergaben sich aber keine
Auswirkungen auf den Phänotyp des Pankreas, auch 13 Monate alte Mäuse wiesen
keine entzündlichen Veränderung im Pankreas auf. Die duktalen und endokrinen
Kompartimente blieben ebenfalls unbeeinflusst.
Die akute experimentelle Pankreatitis wurde anhand zwei verschiedener Modelle, der
Caerulein und der L-Arginin-Pankreatitis studiert. Nach Induktion der Caerulein-
Pankreatitis wurden die Tiere nach 8h bzw. 24h euthanasiert und die entnommen
Gewebe und Serumproben weiter analysiert. Bereits morphologisch zeigten sich im
Wildtyp ausgeprägtere entzündliche Veränderungen als in den Knockout Mäusen.
Dies lies sich in allen untersuchten Parametern für die akute Pankreatitis weiter
bestätigen. Das relative Pankreasgewicht, die Quantifizierung von Nekrose und
Inflammation sowie die Konzentration von Amylase und Lipase im Serum waren
durch die Inhibition von IκBα signifikant erniedrigt. Analog lies sich dies auch in der L-

VI Zusammenfassung
58
Arginin Pankreatitis bestätigen. Auch konnte gezeigt werden, dass durch die
Inhibition von IκBα die Trypsin-Aktivierung in den Azinuszellen während der
Pankreatitis reduziert ist. Analog liesen sich diese Beobachtungen in der L-Arginin-
Pankreatitis bestätigen.
Die extrapankreatischen Auswirkungen der akuten experimentellen Pankreatitis
wurden durch die azinäre Inaktivierung von IκBα ebenfalls reduziert. Mittels ELISA
konnte die Ausschüttung vom IL-6 im Serum quantifizieren werden und das Ausmaß
der neutrophilen Granulozyten – Infiltration in der Lunge als Indiktator für den Grad
der Entzündung wurde bestimmt. Diese Parameter waren in den Knockout Mäusen
im Vergleich zum Wildtyp signifikant reduziert.
Um letztendlich zu beweisen, dass all diese Beobachtungen tatsächlich auf die
endogene Aktivität von NF-κB zurückzuführen sind, wurde in einem zweiten Schritt
der Verlauf einer akuten Pankreatitis in rela x ikbaΔpanc
–Mäusen untersucht. In
diesem Fall ist der Ikk/NF-κB/RelA-Signalweg nicht mehr intakt, zusätzlich zu IκBα ist
RelA ebenfalls inaktiv. In diesen Mäusen zeigte sich nun eine wesentlich schwerer
verlaufende Pankreatitis sowohl im Vergleich zu den Kontrollmäusen wie auch zu
den ikbaΔpanc
-Mäusen.
Insgesamt konnte also gezeigt werden, dass die pankreas-spezifische Inaktivierung
von IκBα protektive Bedeutung für den Verlauf einer akuten Pankreatitis hat. Diese
Beobachtung ist auf eine endogene Aktivität von NF-κB in den Azinuszellen
zurückzuführen. Im Weiteren gilt es nun zu analysieren durch welche Zielproteine
NF-κB seine protektive Wirkung vermittelt, zum einem um die Pathogenese der
akuten Pankreatits weiter zu entschlüsseln, vor allem aber um letztendlich
spezifische therapeutische Strategien entwickeln zu können.

Referenzen IX
59
VII. Danksagung
In besonderen Maße möchte ich mich bei Herrn PD Dr. H. Algül für die Vergabe des
Themas, Unterstützung, Vetrauen sowie die freundliche und lehrreiche Betreuung
bedanken.
Ebenfalls gilt mein Dank Herrn Prof Dr. R.M. Schmid für die Arbeitsmöglichkeit und
Bereitstellung der technischen Voraussetzungen.
Herrn PD Dr R. Rupec möchte ich für die Bereitstellung der ikbaF/F-Maus danken.
Für die Durchführung der Micro-Array-Analysen bedanke ich mich sehr herzlich bei
Herrn Dr H. Einwächter, des weiteren auch für seine Ratschläge und Tipps während
der Laborarbeit.
Dies gilt ebenso und in besonderem Maße für Frau Dr. M. Lesina und Herrn Dr. M.
Treiber ohne deren Unterstützung ich diese Arbeit sicherlich nicht so gut hätte
bewältigen können.
Des weiteren möchte ich mich bei Elisabeth Anetsberger bedanken, ohne die die
Arbeit im Labor nur halb so viel Spass gemacht hätte, sowie bei allen Kollegen und
Mitarbeitern der II. Medizinischen Klinik des Klinikums rechts der Isar, die mich bei
der Fertigstellung dieser Dissertation unterstützt haben.

VIII Danksagung
60
VIII. Referenzen
ALEKSIC, T., BAUMANN, B., WAGNER, M., ADLER, G., WIRTH, T. & WEBER, C. K. 2007. Cellular immune reaction in the pancreas is induced by constitutively active IkappaB kinase-2. Gut, 56, 227-36.
ALGUL, H., TANDO, Y., SCHNEIDER, G., WEIDENBACH, H., ADLER, G. & SCHMID, R. M. 2002. Acute experimental pancreatitis and NF-kappaB/Rel activation. Pancreatology, 2, 503-9.
ALGUL, H., TREIBER, M., LESINA, M., NAKHAI, H., SAUR, D., GEISLER, F., PFEIFER, A., PAXIAN, S. & SCHMID, R. M. 2007. Pancreas-specific RelA/p65 truncation increases susceptibility of acini to inflammation-associated cell death following cerulein pancreatitis. J Clin Invest, 117, 1490-501.
ALTAVILLA, D., FAMULARI, C., PASSANITI, M., GALEANO, M., MACRI, A., SEMINARA, P., MINUTOLI, L., MARINI, H., CALO, M., VENUTI, F. S., ESPOSITO, M. & SQUADRITO, F. 2003. Attenuated cerulein-induced pancreatitis in nuclear factor-kappaB-deficient mice. Lab Invest, 83, 1723-32.
AOUN, E., CHEN, J., REIGHARD, D., GLEESON, F. C., WHITCOMB, D. C. & PAPACHRISTOU, G. I. 2009. Diagnostic accuracy of interleukin-6 and interleukin-8 in predicting severe acute pancreatitis: a meta-analysis. Pancreatology, 9, 777-85.
BALDI, L., BROWN, K., FRANZOSO, G. & SIEBENLIST, U. 1996. Critical role for lysines 21 and 22 in signal-induced, ubiquitin-mediated proteolysis of I kappa B-alpha. J Biol Chem, 271, 376-9.
BANG, U. C., SEMB, S., NOJGAARD, C. & BENDTSEN, F. 2008. Pharmacological approach to acute pancreatitis. World J Gastroenterol, 14, 2968-76.
BARTELT, D. C., SHAPANKA, R. & GREENE, L. J. 1977. The primary structure of the human pancreatic secretory trypsin inhibitor. Amino acid sequence of the reduced S-aminoethylated protein. Arch Biochem Biophys, 179, 189-99.
BAUMANN, B., WAGNER, M., ALEKSIC, T., VON WICHERT, G., WEBER, C. K., ADLER, G. & WIRTH, T. 2007. Constitutive IKK2 activation in acinar cells is sufficient to induce pancreatitis in vivo. J Clin Invest, 117, 1502-13.
BEG, A. A., SHA, W. C., BRONSON, R. T. & BALTIMORE, D. 1995. Constitutive NF-kappa B activation, enhanced granulopoiesis, and neonatal lethality in I kappa B alpha-deficient mice. Genes Dev, 9, 2736-46.
BHATIA, M., BRADY, M., SHOKUHI, S., CHRISTMAS, S., NEOPTOLEMOS, J. P. & SLAVIN, J. 2000. Inflammatory mediators in acute pancreatitis. J Pathol, 190, 117-25.
BI, Y. & WILLIAMS, J. A. 2004. Receptor biology and signal transduction in pancreatic acinar cells. Current opinion in gastroenterology, 20, 427-34.
BLANK, V., KOURILSKY, P. & ISRAEL, A. 1991. Cytoplasmic retention, DNA binding and processing of the NF-kappa B p50 precursor are controlled by a small region in its C-terminus. Embo J, 10, 4159-67.
BRICTEUX-GREGOIRE, S., SCHYNS, R. & FLORKIN, M. 1972. Phylogeny of trypsinogen activation peptides. Comp Biochem Physiol B, 42, 23-39.
BROCKHAUS, M., SCHOENFELD, H. J., SCHLAEGER, E. J., HUNZIKER, W., LESSLAUER, W. & LOETSCHER, H. 1990. Identification of two types of tumor necrosis factor receptors on human cell lines by monoclonal antibodies. Proc Natl Acad Sci U S A, 87, 3127-31.
BROWN, K., GERSTBERGER, S., CARLSON, L., FRANZOSO, G. & SIEBENLIST, U. 1995. Control of I kappa B-alpha proteolysis by site-specific, signal-induced phosphorylation. Science, 267, 1485-8.
BROWNE, G. W. & PITCHUMONI, C. S. 2006. Pathophysiology of pulmonary complications of acute pancreatitis. World J Gastroenterol, 12, 7087-96.

Referenzen IX
61
CHEN, X., JI, B., HAN, B., ERNST, S. A., SIMEONE, D. & LOGSDON, C. D. 2002. NF-kappaB activation in pancreas induces pancreatic and systemic inflammatory response. Gastroenterology, 122, 448-57.
CHIARI, H. 1896. Über die Selbstverdauung des menschlichen Pankreas. Z. Heilkunde, 17, 69-96. COHN, J. A., FRIEDMAN, K. J., NOONE, P. G., KNOWLES, M. R., SILVERMAN, L. M. & JOWELL, P.
S. 1998. Relation between mutations of the cystic fibrosis gene and idiopathic pancreatitis. N Engl J Med, 339, 653-8.
DAMBRAUSKAS, Z., GIESE, N., GULBINAS, A., GIESE, T., BERBERAT, P. O., PUNDZIUS, J., BARAUSKAS, G. & FRIESS, H. 2010. Different profiles of cytokine expression during mild and severe acute pancreatitis. World J Gastroenterol, 16, 1845-53.
DARIEVA, Z., LASUNSKAIA, E. B., CAMPOS, M. N., KIPNIS, T. L. & DA SILVA, W. D. 2004. Activation of phosphatidylinositol 3-kinase and c-Jun-N-terminal kinase cascades enhances NF-kappaB-dependent gene transcription in BCG-stimulated macrophages through promotion of p65/p300 binding. J Leukoc Biol, 75, 689-97.
DAWRA, R., SAH, R. P., DUDEJA, V., RISHI, L., TALUKDAR, R., GARG, P. & SALUJA, A. K. 2011. Intra-acinar trypsinogen activation mediates early stages of pancreatic injury but not inflammation in mice with acute pancreatitis. Gastroenterology, 141, 2210-2217 e2.
DIEHL, A. K., HOLLEMAN, D. R., JR., CHAPMAN, J. B., SCHWESINGER, W. H. & KURTIN, W. E. 1997. Gallstone size and risk of pancreatitis. Arch Intern Med, 157, 1674-8.
DUNN, J. A., LI, C., HA, T., KAO, R. L. & BROWDER, W. 1997. Therapeutic modification of nuclear factor kappa B binding activity and tumor necrosis factor-alpha gene expression during acute biliary pancreatitis. Am Surg, 63, 1036-43; discussion 1043-4.
ETHRIDGE, R. T., HASHIMOTO, K., CHUNG, D. H., EHLERS, R. A., RAJARAMAN, S. & EVERS, B. M. 2002. Selective inhibition of NF-kappaB attenuates the severity of cerulein-induced acute pancreatitis. J Am Coll Surg, 195, 497-505.
FREY, C. F., ZHOU, H., HARVEY, D. J. & WHITE, R. H. 2006. The incidence and case-fatality rates of acute biliary, alcoholic, and idiopathic pancreatitis in California, 1994-2001. Pancreas, 33, 336-44.
FRITSCH, H. 2003. Bauchspeicheldrüse. In: FRITSCH H., K. W. (ed.) Taschenatlas der Anatomie. 8 ed. Stuttgart, New York: Thieme.
FROSSARD, J. L., STEER, M. L. & PASTOR, C. M. 2008. Acute pancreatitis. Lancet, 371, 143-52. FUJIMOTO, K., HOSOTANI, R., DOI, R., WADA, M., LEE, J. U., KOSHIBA, T., MIYAMOTO, Y. &
IMAMURA, M. 1997. Role of neutrophils in cerulein-induced pancreatitis in rats: possible involvement of apoptosis. Digestion, 58, 421-30.
GHOSH, S. 1999. Regulation of inducible gene expression by the transcription factor NF-kappaB. Immunol Res, 19, 183-9.
GHOSH, S. & KARIN, M. 2002. Missing pieces in the NF-kappaB puzzle. Cell, 109 Suppl, S81-96. GHOSH, S., MAY, M. J. & KOPP, E. B. 1998. NF-kappa B and Rel proteins: evolutionarily conserved
mediators of immune responses. Annu Rev Immunol, 16, 225-60. GORELICK, F. S., MODLIN, I. M., LEACH, S. D., CARANGELO, R. & KATZ, M. 1992. Intracellular
proteolysis of pancreatic zymogens. Yale J Biol Med, 65, 407-20; discussion 437-40. GRADY, T., MAH'MOUD, M., OTANI, T., RHEE, S., LERCH, M. M. & GORELICK, F. S. 1998.
Zymogen proteolysis within the pancreatic acinar cell is associated with cellular injury. Am J Physiol, 275, G1010-7.
GUKOVSKAYA, A. S., GUKOVSKY, I., JUNG, Y., MOURIA, M. & PANDOL, S. J. 2002. Cholecystokinin induces caspase activation and mitochondrial dysfunction in pancreatic acinar cells. Roles in cell injury processes of pancreatitis. J Biol Chem, 277, 22595-604.
GUKOVSKAYA, A. S., GUKOVSKY, I., ZANINOVIC, V., SONG, M., SANDOVAL, D., GUKOVSKY, S. & PANDOL, S. J. 1997. Pancreatic acinar cells produce, release, and respond to tumor necrosis factor-alpha. Role in regulating cell death and pancreatitis. J Clin Invest, 100, 1853-62.
GUKOVSKAYA, A. S. & PANDOL, S. J. 2004. Cell death pathways in pancreatitis and pancreatic cancer. Pancreatology, 4, 567-86.
GUKOVSKY, I., GUKOVSKAYA, A. S., BLINMAN, T. A., ZANINOVIC, V. & PANDOL, S. J. 1998. Early NF-kappaB activation is associated with hormone-induced pancreatitis. Am J Physiol, 275, G1402-14.
GUKOVSKY, I., REYES, C. N., VAQUERO, E. C., GUKOVSKAYA, A. S. & PANDOL, S. J. 2003. Curcumin ameliorates ethanol and nonethanol experimental pancreatitis. Am J Physiol Gastrointest Liver Physiol, 284, G85-95.
HALANGK, W., LERCH, M. M., BRANDT-NEDELEV, B., ROTH, W., RUTHENBUERGER, M., REINHECKEL, T., DOMSCHKE, W., LIPPERT, H., PETERS, C. & DEUSSING, J. 2000. Role

IX Referenzen
62
of cathepsin B in intracellular trypsinogen activation and the onset of acute pancreatitis. J Clin Invest, 106, 773-81.
HAN, B., JI, B. & LOGSDON, C. D. 2001. CCK independently activates intracellular trypsinogen and NF-kappaB in rat pancreatic acinar cells. Am J Physiol Cell Physiol, 280, C465-72.
HAYDEN, M. S. & GHOSH, S. 2004. Signaling to NF-kappaB. Genes Dev, 18, 2195-224. HAYDEN, M. S. & GHOSH, S. 2008. Shared principles in NF-kappaB signaling. Cell, 132, 344-62. HEGYI, P., RAKONCZAY, Z., JR., SARI, R., GOG, C., LONOVICS, J., TAKACS, T. & CZAKO, L.
2004. L-arginine-induced experimental pancreatitis. World J Gastroenterol, 10, 2003-9. HIETARANTA, A. J., SALUJA, A. K., BHAGAT, L., SINGH, V. P., SONG, A. M. & STEER, M. L. 2001.
Relationship between NF-kappaB and trypsinogen activation in rat pancreas after supramaximal caerulein stimulation. Biochem Biophys Res Commun, 280, 388-95.
HOFFMANN, A., NATOLI, G. & GHOSH, G. 2006. Transcriptional regulation via the NF-kappaB signaling module. Oncogene, 25, 6706-16.
JOHNSON, C. D. 2005. UK guidelines for the management of acute pancreatitis. Gut, 54 Suppl 3, iii1-9.
JUNGERMANN, J., LERCH, M. M., WEIDENBACH, H., LUTZ, M. P., KRUGER, B. & ADLER, G. 1995. Disassembly of rat pancreatic acinar cell cytoskeleton during supramaximal secretagogue stimulation. Am J Physiol, 268, G328-38.
KARIN, M. & BEN-NERIAH, Y. 2000. Phosphorylation meets ubiquitination: the control of NF-[kappa]B activity. Annu Rev Immunol, 18, 621-63.
KLEMENT, J. F., RICE, N. R., CAR, B. D., ABBONDANZO, S. J., POWERS, G. D., BHATT, P. H., CHEN, C. H., ROSEN, C. A. & STEWART, C. L. 1996. IkappaBalpha deficiency results in a sustained NF-kappaB response and severe widespread dermatitis in mice. Mol Cell Biol, 16, 2341-9.
KRAPP, A., KNOFLER, M., FRUTIGER, S., HUGHES, G. J., HAGENBUCHLE, O. & WELLAUER, P. K. 1996. The p48 DNA-binding subunit of transcription factor PTF1 is a new exocrine pancreas-specific basic helix-loop-helix protein. Embo J, 15, 4317-29.
KRAPP, A., KNOFLER, M., LEDERMANN, B., BURKI, K., BERNEY, C., ZOERKLER, N., HAGENBUCHLE, O. & WELLAUER, P. K. 1998. The bHLH protein PTF1-p48 is essential for the formation of the exocrine and the correct spatial organization of the endocrine pancreas. Genes Dev, 12, 3752-63.
KUCHARCZAK, J., SIMMONS, M. J., FAN, Y. & GELINAS, C. 2003. To be, or not to be: NF-kappaB is the answer--role of Rel/NF-kappaB in the regulation of apoptosis. Oncogene, 22, 8961-82.
KUWATA, H., WATANABE, Y., MIYOSHI, H., YAMAMOTO, M., KAISHO, T., TAKEDA, K. & AKIRA, S. 2003. IL-10-inducible Bcl-3 negatively regulates LPS-induced TNF-alpha production in macrophages. Blood, 102, 4123-9.
LAMPEL, M. & KERN, H. F. 1977. Acute interstitial pancreatitis in the rat induced by excessive doses of a pancreatic secretagogue. Virchows Arch A Pathol Anat Histol, 373, 97-117.
LANKISCH, P. G., DROGE, M. & GOTTESLEBEN, F. 1995. Drug induced acute pancreatitis: incidence and severity. Gut, 37, 565-7.
LEACH, S. D., MODLIN, I. M., SCHEELE, G. A. & GORELICK, F. S. 1991. Intracellular activation of digestive zymogens in rat pancreatic acini. Stimulation by high doses of cholecystokinin. J Clin Invest, 87, 362-6.
LEINDLER, L., MORSCHL, E., LASZLO, F., MANDI, Y., TAKACS, T., JARMAI, K. & FARKAS, G. 2004. Importance of cytokines, nitric oxide, and apoptosis in the pathological process of necrotizing pancreatitis in rats. Pancreas, 29, 157-61.
LERCH, M. M., WEIDENBACH, H., HERNANDEZ, C. A., PRECLIK, G. & ADLER, G. 1994. Pancreatic outflow obstruction as the critical event for human gall stone induced pancreatitis. Gut, 35, 1501-3.
LI, Z. W., CHU, W., HU, Y., DELHASE, M., DEERINCK, T., ELLISMAN, M., JOHNSON, R. & KARIN, M. 1999. The IKKbeta subunit of IkappaB kinase (IKK) is essential for nuclear factor kappaB activation and prevention of apoptosis. J Exp Med, 189, 1839-45.
LINDKVIST, B., APPELROS, S., MANJER, J. & BORGSTROM, A. 2004. Trends in incidence of acute pancreatitis in a Swedish population: is there really an increase? Clin Gastroenterol Hepatol, 2, 831-7.
LUKOWSKI R., W. S., WEINMEISTER P., FEIL S., FEIL R., 2005. Cre/loxP-vermittelte konditionale Mutagenese des cGMP-Signalwegs in der Maus. BIOspektrum, 3, 287-90.
MALEK, S., CHEN, Y., HUXFORD, T. & GHOSH, G. 2001. IkappaBbeta, but not IkappaBalpha, functions as a classical cytoplasmic inhibitor of NF-kappaB dimers by masking both NF-kappaB nuclear localization sequences in resting cells. J Biol Chem, 276, 45225-35.
MALKA, D., VASSEUR, S., BODEKER, H., ORTIZ, E. M., DUSETTI, N. J., VERRANDO, P., DAGORN, J. C. & IOVANNA, J. L. 2000. Tumor necrosis factor alpha triggers antiapoptotic

Referenzen IX
63
mechanisms in rat pancreatic cells through pancreatitis-associated protein I activation. Gastroenterology, 119, 816-28.
MERCURIO, F., DIDONATO, J. A., ROSETTE, C. & KARIN, M. 1993. p105 and p98 precursor proteins play an active role in NF-kappa B-mediated signal transduction. Genes Dev, 7, 705-18.
MIZUNUMA, T., KAWAMURA, S. & KISHINO, Y. 1984. Effects of injecting excess arginine on rat pancreas. J Nutr, 114, 467-71.
NAKHAI, H., SEL, S., FAVOR, J., MENDOZA-TORRES, L., PAULSEN, F., DUNCKER, G. I. & SCHMID, R. M. 2007. Ptf1a is essential for the differentiation of GABAergic and glycinergic amacrine cells and horizontal cells in the mouse retina. Development, 134, 1151-60.
O'FARRELL, A., ALLWRIGHT, S., TOOMEY, D., BEDFORD, D. & CONLON, K. 2007. Hospital admission for acute pancreatitis in the Irish population, 1997 2004: could the increase be due to an increase in alcohol-related pancreatitis? J Public Health (Oxf), 29, 398-404.
OWYANG, C. & LOGSDON, C. D. 2004. New insights into neurohormonal regulation of pancreatic secretion. Gastroenterology, 127, 957-69.
PASTOR, C. M., MATTHAY, M. A. & FROSSARD, J. L. 2003. Pancreatitis-associated acute lung injury: new insights. Chest, 124, 2341-51.
PERKINS, N. D. 2000. The Rel/NF-kappa B family: friend and foe. Trends Biochem Sci, 25, 434-40. PERKINS, N. D. 2006. Post-translational modifications regulating the activity and function of the
nuclear factor kappa B pathway. Oncogene, 25, 6717-30. RAGHU, M. G., WIG, J. D., KOCHHAR, R., GUPTA, D., GUPTA, R., YADAV, T. D., AGARWAL, R.,
KUDARI, A. K., DOLEY, R. P. & JAVED, A. 2007. Lung complications in acute pancreatitis. Jop, 8, 177-85.
RAKONCZAY, Z., JR., HEGYI, P., TAKACS, T., MCCARROLL, J. & SALUJA, A. K. 2008. The role of NF-kappaB activation in the pathogenesis of acute pancreatitis. Gut, 57, 259-67.
REBHOLZ, B., HAASE, I., ECKELT, B., PAXIAN, S., FLAIG, M. J., GHORESCHI, K., NEDOSPASOV, S. A., MAILHAMMER, R., DEBEY-PASCHER, S., SCHULTZE, J. L., WEINDL, G., FORSTER, I., HUSS, R., STRATIS, A., RUZICKA, T., ROCKEN, M., PFEFFER, K., SCHMID, R. M. & RUPEC, R. A. 2007. Crosstalk between keratinocytes and adaptive immune cells in an IkappaBalpha protein-mediated inflammatory disease of the skin. Immunity, 27, 296-307.
REUBI, J. C., WASER, B., GUGGER, M., FRIESS, H., KLEEFF, J., KAYED, H., BUCHLER, M. W. & LAISSUE, J. A. 2003. Distribution of CCK1 and CCK2 receptors in normal and diseased human pancreatic tissue. Gastroenterology, 125, 98-106.
RICE, N. R., MACKICHAN, M. L. & ISRAEL, A. 1992. The precursor of NF-kappa B p50 has I kappa B-like functions. Cell, 71, 243-53.
RINDERKNECHT, H. 1986. Activation of Pancreatic Zymogens, Normal Activation, Premature Intrapancreatic Activation, Protective Mechanisms Against Inappropriate Activation. Digestive Diseases and Science, 31, pp. 314 - 321.
RONGIONE, A. J., KUSSKE, A. M., KWAN, K., ASHLEY, S. W., REBER, H. A. & MCFADDEN, D. W. 1997. Interleukin 10 reduces the severity of acute pancreatitis in rats. Gastroenterology, 112, 960-7.
RUPEC, R. A., JUNDT, F., REBHOLZ, B., ECKELT, B., WEINDL, G., HERZINGER, T., FLAIG, M. J., MOOSMANN, S., PLEWIG, G., DORKEN, B., FORSTER, I., HUSS, R. & PFEFFER, K. 2005. Stroma-mediated dysregulation of myelopoiesis in mice lacking I kappa B alpha. Immunity, 22, 479-91.
SALUJA, A., HASHIMOTO, S., SALUJA, M., POWERS, R. E., MELDOLESI, J. & STEER, M. L. 1987. Subcellular redistribution of lysosomal enzymes during caerulein-induced pancreatitis. Am J Physiol, 253, G508-16.
SALUJA, A. K., LERCH, M. M., PHILLIPS, P. A. & DUDEJA, V. 2007. Why Does Pancreatic Overstimulation Cause Pancreatitis?
doi:10.1146/annurev.physiol.69.031905.161253. Annual Review of Physiology, 69, 249-269. SATOH, A., SHIMOSEGAWA, T., FUJITA, M., KIMURA, K., MASAMUNE, A., KOIZUMI, M. &
TOYOTA, T. 1999. Inhibition of nuclear factor-kappaB activation improves the survival of rats with taurocholate pancreatitis. Gut, 44, 253-8.
SBAROUNIS, C. N. 1997. Did Alexander the Great die of acute pancreatitis? J Clin Gastroenterol, 24, 294-6.
SEN, R. & BALTIMORE, D. 1986. Multiple nuclear factors interact with the immunoglobulin enhancer sequences. Cell, 46, 705-16.
STEINLE, A. U., WEIDENBACH, H., WAGNER, M., ADLER, G. & SCHMID, R. M. 1999. NF-kappaB/Rel activation in cerulein pancreatitis. Gastroenterology, 116, 420-30.
SUBRAMANIAN, A., TAMAYO, P., MOOTHA, V. K., MUKHERJEE, S., EBERT, B. L., GILLETTE, M. A., PAULOVICH, A., POMEROY, S. L., GOLUB, T. R., LANDER, E. S. & MESIROV, J. P.

IX Referenzen
64
2005. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc Natl Acad Sci U S A, 102, 15545-50.
TANDO, Y., ALGUL, H., SCHNEIDER, G., WEBER, C. K., WEIDENBACH, H., ADLER, G. & SCHMID, R. M. 2002. Induction of IkappaB-kinase by cholecystokinin is mediated by trypsinogen activation in rat pancreatic lobules. Digestion, 66, 237-45.
TANDO, Y., ALGUL, H., WAGNER, M., WEIDENBACH, H., ADLER, G. & SCHMID, R. M. 1999. Caerulein-induced NF-kappaB/Rel activation requires both Ca2+ and protein kinase C as messengers. Am J Physiol, 277, G678-86.
VAN ACKER, G. J., PERIDES, G. & STEER, M. L. 2006. Co-localization hypothesis: a mechanism for the intrapancreatic activation of digestive enzymes during the early phases of acute pancreatitis. World J Gastroenterol, 12, 1985-90.
VONLAUFEN, A., APTE, M. V., IMHOF, B. A. & FROSSARD, J. L. 2007. The role of inflammatory and parenchymal cells in acute pancreatitis. J Pathol, 213, 239-48.
WASLE, B. & EDWARDSON, J. M. 2002. The regulation of exocytosis in the pancreatic acinar cell. Cell Signal, 14, 191-7.
WATANABE, O., BACCINO, F. M., STEER, M. L. & MELDOLESI, J. 1984. Supramaximal caerulein stimulation and ultrastructure of rat pancreatic acinar cell: early morphological changes during development of experimental pancreatitis. Am J Physiol, 246, G457-67.
WELSCH, U. 2003. Bauchspeicheldrüse. In: WELSCH, U. (ed.) Lehrbuch Histologie. 1 ed. München, Jena: Urban & Fischer Verlag.
WHITCOMB, D. C. 1999. Early trypsinogen activation in acute pancreatitis. Gastroenterology, 116, 770-2.
WHITCOMB, D. C. 2006. Clinical practice. Acute pancreatitis. N Engl J Med, 354, 2142-50. WHITCOMB, D. C., GORRY, M. C., PRESTON, R. A., FUREY, W., SOSSENHEIMER, M. J., ULRICH,
C. D., MARTIN, S. P., GATES, L. K., JR., AMANN, S. T., TOSKES, P. P., LIDDLE, R., MCGRATH, K., UOMO, G., POST, J. C. & EHRLICH, G. D. 1996. Hereditary pancreatitis is caused by a mutation in the cationic trypsinogen gene. Nat Genet, 14, 141-5.
WILLEMER, S., BIALEK, R. & ADLER, G. 1990. Localization of lysosomal and digestive enzymes in cytoplasmic vacuoles in caerulein-pancreatitis. Histochemistry, 94, 161-70.
YADAV, D. & LOWENFELS, A. B. 2006. Trends in the epidemiology of the first attack of acute pancreatitis: a systematic review. Pancreas, 33, 323-30.
YASUDA, H., KATAOKA, K., ICHIMURA, H., MITSUYOSHI, M., IIDA, T., KITA, M. & IMANISHI, J. 1999. Cytokine expression and induction of acinar cell apoptosis after pancreatic duct ligation in mice. J Interferon Cytokine Res, 19, 637-44.
YUAN, J., LUGEA, A., ZHENG, L., GUKOVSKY, I., EDDERKAOUI, M., ROZENGURT, E. & PANDOL, S. J. 2008. Protein kinase D1 mediates NF-kappaB activation induced by cholecystokinin and cholinergic signaling in pancreatic acinar cells. Am J Physiol Gastrointest Liver Physiol, 295, G1190-201.
ZHANG, X. P., LIN, Q. & ZHOU, Y. F. 2007. Progress of study on the relationship between mediators of inflammation and apoptosis in acute pancreatitis. Dig Dis Sci, 52, 1199-205.
Recommended