
MARYSE LESSARD
ÉTUDE DE L'EXPRESSION DES GÈNES ENCODANT· LES SOUS-UNITÉS D'INTÉGRINES a4 ET aS DANS LE CONTEXTE DES 'PROPRIÉTÉS TUMORIGÈNES
ET MÉTASTATIQUES DU MÉLANOME UVÉAL
Mémoire présenté à la Faculté des études supérieures de l'Université Laval
dans le cadre du programme de maîtrise en biologie cellulaire et moléculaire pour l'obtention du grade de Maître ès sciences (M.Sc.)
© Maryse Lessard, 2009
FACULTÉ DE MÉDECINE UNIVERSITÉ LAVAL
QUÉBEC
2009

II
Résumé
Les intégrines, des récepteurs trans-membranaires exerçant une fonction importante
dans l'adhésion et la migration cellulaire, pourraient contribuer aux propriétés,
tumorigènes et métastatiques du mélanome uvéa!. Nous avons démontré que le patron
d'expression de certaines intégrines différait entre les lignées tumorales: la sous-unité
d'intégrine a4 est anormalement élevée dans la lignée métastatique H79 alors que
l'expression de la sous-unité a5 est presque entièrement abolie dans des lignées à plus
fort potentiel tumorigène mais non dans les lignées peu agressives. La transcription du
gène a4 étant sous l' influence régulatrice du facteur de transcription (FT) Pax-6, nous
avons démontré que celui-ci n 'est cependant pas l'élément déterminant dans l'expression
aberrante du gène a4. Sachant que le promoteur du gène a5 est sous l'influence des FTs
Spl/Sp3, AP-l et NFI, nous avons démontré que l'agressivité des lignées de mélanomes
pouvait être étroitement liée' à des changements dans l'expression ou les propriétés de ces
FTs.

III
Avant-propos
Les résultàts obtenus pendant mon programme de maîtrise sont divisés en deux
chapitres (chapitre 2 et chapitre ·3) rédigés sous forme traditionnelle. Le troisième
chapitre de ce mémoire fera l'objet d'un prochain article, toujours en préparation au
moment de l'écriture de ce mémoire. Le Dr Sylvain L. Guérin, chercheur au centre de
recherche du CHUL, a agi à titre de directeur de recherche. Steeve Leclerc, assistant de
recherche dans l'équipe du Dr Sylvain L. Guérin, a agi à titre d'assistant de recherche
lors de la réalisation des travaux présentés dans ce mémoire.
Le premier chapitre de ce mémoire contient une introduction ainsi que les objectifs des
travaux présentés dans les chapitres suivants. Le second chapitre s'intitule: « L'influence
de Pax -6 sur la transcription du promoteur a4 dans le mélanome uvéal » et présente les
résultats d'une première étude concernant plus particulièrement la sous-unité d'intégrine
a4 dans le contexte de la progression métastatique du mélanome uvéa!. J'ai réalisé la
majeure partie des expériences de cette étude et quelques-unes avec l'aide de Solange
Landreville, PhD, étudiante dans le laboratoire du Dr Christian Salesse (Unité de
recherche en Ophtalmologie du CHUL) et de Steeve Leclerc, assistant de recherche,
MSc, dans le laboratoire du Dr Guérin. Le troisième chapitre est intitulé: « Influence de
la sous-unité a5 dans les propriétés tumorigènes du inélanome uvéal» et présente les
résultats d'une seconde étude portant sur la sous-unité d'intégrine a5 au niveau du
potentiel tumorigène du mélanome uvéa!. J'ai réalisé toutes les expériences de cette étude
dont certaines avec l'aide de Solange Landreville, PhD, étudiante dans le laboratoire du
Dr Salesse, et de Manon Gaudreault, assistante de recherche, PhD, dans le laboratoire du
Dr Guérin. Finalement, le dernier chapitre remet en contexte les conclusions tirées des
chapitres précédents ainsi que les perspectives qui sont ressorties suite à ce projet.

IV
Remerciements
Dans un premIer temps, j'aimerais remerCIer mon directeur de recherche, le Dr
Sylvain Guérin, pour ses conseils, son aide précieuse, mais surtout, sa présence tout au
long de la réalisation de cette maîtrise. Sa motivation et son soutien m'ont non seulement
permis ~e me dépasser tout au long de ces années, mais de le faire avec grande
satisfaction et fierté.
Un gros merci pour l'équipe Guérin et tout particulièrement Steeve Leclerc pour son
aide technique et sa façon extraordinaire ' de répondre à nos milliards de questions.
J'aimerais aussi remercier Karine Zaniolo qui m'a prise sous son aile dès mon
commencement. Un gros merci à Manon Gaudreault et Marie-Ève Gingras, pour leur
support moral, leur aide tout au long de ces années et avec qui j'ai pu partager de
nombreuses confidences. Merci les filles d'avoir su me poser les bonnes questions au bon
moment!
Je tiens également à remerCIer le Dr Christian Saless'e et son équipe, Solange
Landreville, Jennifer Gagné et Marj orie Bergeron pour leur précieuse collaboration dans
la réalisation de ce projet.
Je tiens à remercier le réseau vision, la banque d'Yeux du CHUL, la fondation de la
recherche en santé du Québec (FRSQ) et la Fondation Georges Phénix pour leur soutien
financier.
Finalement, merci à mes amis et à ma famille qui ont cru en moi du début à la fin. Un
merci tout spécial à mes parents et mon frère qui ont toujours été là pour moi et qui m'ont
toujours supportée. Un dernier merci, mais non le moindre, à mon conjoint, Pierre-Michel
Vallée qui m'a soutenue pendant les moments les plus difficiles et qui a toujours su dire
les mots pour me rassurer et m'encourager.
À tous, merci énormément!

v
Table des matières
Résumé ............................................................................................................................. 11
Avant-propos ................................................................................................................ 111
Remerciements .................................................................... : ........................................ IV
Table des matières ....................................................................................................... V
Liste des figures et tableaux .................................. .................................................. IX
Liste des abréviations ............................................................................................... XI
Chapitre 1
1 Introduction ................................................................................................................. 1
1.1 Généralités sur le cancer
1.2 Le mélanome uvéal 1.2.1 Généralités 1.2.2 Anatomie générale de l' œil 1.2.3 Type de mélanomes du tractus uvéal
1.2.3.1 Mélanomes de l'iris 1.2.3.2 Mélanomes du corps ciliaire 1.2.3.3 Mélanomes choroïdiens 1.2.3.4 Type cellulaire
1.2.3.4.1 Profil fusiforme 1.2.3.4.2 Profil épithélioïde 1.2.3.4.3 Profil mixte
1.2.4 Causes 1.2.5 Diagnostic et traitement
1.2.5.1 Diagnostic de la tumeur primaire 1.2.5.2 Choix du traitement
1.2.5.2.1 Notion de taille 1.2.5.2.2 Autres traitements possibles
1.2.6 Facteurs pronostics 1.2.6.1 La taille de la tumeur 1.2.6.2 Le type cellulaire et l'agressivité
1.2.7 Métastases 1.2.7.1 Processus métastatique 1.2.7.2 Pronostic et cause
1
3 3 3 5 5 6 6 6 7 7 7 8 8 8 9 9
10 10 10 Il Il Il 12

VI
1.3 Les molécules d'adhésion 14 1.3.1 Les sélectines 14 1.3.2 La superfamille des immunoglobulines 16 1.3.3 Les cadhérines 16 1.3.4 Les intégrines 17
1.3.4.1 Structure des intégrines 17 1.3.4.1.1 Sous-unité a 19 1.3.4.1.2 Sous-unité B 20
1.3.4.2 Ligands des intégrines 21 1.3.4.2.1 Matrice extracellulaire 21
1.3.4.3 Rôles des intégrines 22 1.3.4.4 Signalisation par les intégrines 23
1.3.5 La progression tumorale 25 1.3.6 L'intégrine a4B1 26
1.3.6.1 Structure de l'intégrirte a4~1 27 1.3.6.2 Régions régulatrices du gène a4 27 1.3.6.3 Le facteur de transcription Pax-6 29
1.3.6.3.1 La famille des gènes PAX 29 1.3.6.3.2 Pax-6 29
1.3.7 L'intégrine a5B1 31 1.3.7.1 Régions régulatrices du gène a5 32
1.4 Objectifs des travaux de recherche 34 1.4.1 Influence de Pax-6 sur l'activité du promoteur de la sous-unité a4 36 1.4.2 Influence .de la sous-unité a5 dans les propriétés tumorigènes du 36
mélanome uvéal
Chapitre 2
2 Influence de Pax-6 sur la transcription du promoteur ............................. 37
04 dans le mélanome uvéal
2.1 Introduction
2.2 Matériel et méthodes 2.2.1 Culture cellulaire 2.2.2 Extraits nucléaires et immunobuvardage de type western 2.2.3 Retard sur gel (EMSA) 2.2.4 Transfections transitoires
2.3 Résultats 2.3.1 Expression du facteur de transcription Pax-6 dans le
mélanome uvéal 2.3.2 Le facteur de transcription Pax-6 peut lier la séquence
de la région promotrice a4.1
37
38 38 38 39 40
41 41
41

1
1·
VII
2.3.3 Pax-6 est un répresseur de l'activité du promoteur a4 43
2.4 Discussion 46
Chapitre 3
3 Influence de la sous-unité aS dans les propriétés ......................................... 50 tumorigènes du mélanome uvéa)
3.1 Introduction
3.2 Matériel et méthodes 3.2.1 Culture cellulaire 3.2.2 Culture cellulaire en agar mou 3.2.3 Transfections transitoires 3.2.4 Extraction d'ARN et analyse en RT-PCR 3.2.5 Extraits nucléaires et immunobuvardage de type western 3.2.6 Retard sur gel (EMSA)
50
52 52 52 53 53 55 55
3.3 Résultats 57 3.3.1 Potentiel tumorigène et capacité des cellules à générer 57
des colonies en agar mou 3.3.2 Activité transcriptionnelle du promoteur du gène a5 59 3.3.3 Niveau d'expression des transcrits encodant les facteurs de 60
transcription SPI, SP3, AP-1 et NFI dans les lignées de mélanomes 3.3.4 Niveau d'expression des facteurs de transcription SPI, SP3, 62
AP-1 et NFI dans les lignées T97, T98, T108 et Tl15 3.3.5 Capacité de liaison des facteurs de transcription SP 1, AP-1 64
et NFI dans les lignées T97, T98, T108 et Tl15 3.3.6 Influence des facteurs SPI, AP-1 et NFI dans les lignées T97 69
et T108
3.4 Discussion 73
Chapitre 4
4 Conclusion .................................................................................................................. 80
4.1 Résumé 4.1.1 Influence de Pax -6 sur le promoteur du gène a4
80 80

4.1.2 Influence de la sous-unité a5 dans les propriétés tumorigènes du mélanome uvéal
4.2 Perspectives de recherche
VIII
81
83
Références bibliographiques ................................................................................... 85

Liste des figures et tableaux
Chapitre 1
Figure 1.1 Figure 1.2 Figure 1.3 Figure 1.4 Figure 1.5 Figure 1.6 Figure 1.7 Figure 1.8 Figure 1.9 Figure 1.10 Figure 1.11 Figure 1.12 Figure 1.13 Figure 1.14 Figure 1.15 Figure 1.16
Chapitre 2
Figure 2.1
Figure 2.2 Figure 2.3 Figure 2.4
Chapitre 3
Tableau 3.1 Tableau 3.2
Figure 3.1 Figure 3.2 Figure 3.3 Figure 3.4
Progression tumorale La structure générale de l'œil humain Différents types de mélanomes Différentes morphologies des cellules de mélanome uvéal Processus métastatique Molécules d'adhésion Structure générale des intégrines La famille des intégrines Signalisation par les intégrines Région régulatrice du gène a4 humain Éléments régulateurs Famille des gènes Pax PAX6 et PAX6 (Sa) Région régulatrice du gène aS humain Analyse en cytométrie de flu'x Expression des gènes encodant les sous-unités d'intégrines a4, aS et a6
Niveau d'expression du facteur de transcription Pax-6 dans les lignées de mélanomes Capacité de liaison de Pax-6 à la séquence régulatrice a4.1 Activité transcriptionnelle du promoteur du gène a4 Influence de Pax-6 sur l'activité du promoteur du gène a4
Résumé des cas d'où proviennent les lignées cancéreuses Séquences des amorces utilisées pour le RT -PCR . semi-quantitatif Morphologie cellulaire Indépendance à l'àncrage sur substrat d'agar mou Activité transcriptionnelle du promoteur du gène aS Niveaux des transcrits endogènes encodant la sous-~nité aS et des facteurs de transcription SpI, Ap-l et NFI
IX
Page
1 4 6 7
12 15 17 18 23 28 28 29 30 33 35 35
41
42 43 45
50 54
57 58 60 61

Figure 3.5.
Figure 3.6
Figure 3.7
Figure 3.8
Figure 3.9 Figure 3.10
Niveau d'expression des facteurs de transcription SpI, NFI et AP-I dans les lignées de mélanomes Capacité de liaison des facteurs de transcription SpI et Sp3 dans les lignées de mélanomes Capacité de liaison du facteur de transcription AP-I dans les lignées de mélanomes Capacité de liaison du facteur de transcription NFI dans les lignées de mélanomes Influence des sites SpI proximal et distal sur le promoteur aS Influence des facteurs de transcription SpI, AP-l et NFI sur le promoteur basal aS
x
63
65
66
68
70
71

Liste des abréviations
A ADN
AP-l
ARN
Asp
ATP
Bp
C
CAT
Cd49d1CD29
CIEBP
CIITA
Crk
DMEM
EDTA
EGF
EMSA
EPR
FACS
FAK
Fra
FT
FRE
G
GAPD
GFP
Glu
Gly
GRB
hGH HMBS
iARN
ICAM
Adénine
Acide désoxyribonucléique
Activator protein (protéine activatrice)
Acide ribonucléique
Aspartate
Adénosine triphosphate
Binding protein (protéine de liaison)
Cytosine
Chloramphénicol acétyl-transférase
Intégrine a4B 1
CCAAT/Enhancer Binding Prote in (protéine qui lie la séquence CCAAT)
class II, major histocompatibility complex, transactivator (transactivateur du complexe majeur d'histocompatibilité classe II) cdc-2 related kinase (kinase s'apparentant à cdc-2)
Dulbecco's modified Eagle's medium
Acide éthylène-diamine-tétraacétique
Epidermal growth factor (facteur de croissance épidermique)
Electrophoretic mobility shift assays (Rétention en gel)
Épithélium pigmentaire rétinien
Fluorescence-activated cell-sorting (Cytométtie de flux)
Focal adhesion kinase (kinase d'adhésion focale)
Fos-related antigen (antigène qui s'apparente à Fos)
F acteur de transcription
Fibronectin responsive element (élément de réponse à la fibronectine)
Guanine G lycéraldéhyde-3 -phosphate déhydrogénase
Green fluorescent protein (protéine fluorescente verte)
Glutamine
Glycine
Growth factor receptor -bound protein (protéine de liaison des récepteurs de facteurs de croissance)
Human growth hormone (hormone de croissance humaine)
hydroxyméthylbilane synthase
ARN interférent
Intercellular adhesion molecule (molécule d'adhésion intercellulaire)
XI

Ile
ILK
lm
Kb
KDa
Leu LM-peR MAPK
MeN
MEC
NCAM
NFI
NK NS
Pax
PCR
PKB
PLC
PNK PSI
PST Ras
RGD
rpm
RT-PCR SDSPAGE Ser
Shc
shRNA
SOS
Sp
Src
SSC
T
TTT Val
Isoleucine
Integrin linked kinase (kinase liée aux intégrines)
Initiateur
Kilobase
Kilodalton
Leucine ligation mediated PCR (peR médié par ligation)
Mitogen-activated prote in kinase (protéine kinase activée par des agents mitogènes)
Mélanocytes choroïdiens normaux
Matrice extracellulaire
Neural cell adhesion molecule (molécule d'adhésion des cellules neurales)
Nuclear factor 1 (facteur nucléaire 1)
N atural killer (tueuse naturelle) Non-spécifique
Paired box
Polymerase chain reaction (Réaction en chaîne de la polymérase)
Protéine kinase B
Phospholipase C Polynucléotide kinase Plexine/Semaphorine/Integrine
Pro line/ sérine/tyrosine
Retrovirus-associated DNA sequence (rétrovirus associé à l'ADN)
Arginine/ glycine/acide aspartique
Révolution par minute Reverse-transcriptase PCR (PCR par transcriptase inverse) Sodium dodecyl sulfate polyacrylamide gel electrophoresis
Sérine
Src homology 2 domain-containing (domaine contenant une région d'homologie 2 avec Src)
Short hairpin RNA (ARN en épingle à cheveux)
Son of sevenless
Specificity protein (protéine spécifique)
Sarcoma Rous cancer (cancer du sarcome de Rous)
Super-complexe
Tyrosine
Thermothérapie à travers la pupille
Valine
XII

VCAM
VLA-4
Vascular cell adhesion molecule (molécule d'adhésion vasculaire)
Very-late antigen 4
XIII

1
1. INTRODUCTION
1.1 Généralités sur le cancer
À tout moment de notre vie, des mutations surviennent à l'intérieur du génome des
cellules constituant notre corps. Ce phénomène amène un dérèglement au niveau du cycle
cellulaire, de la croissance, de la différenciation en plus de la mort cellulaire programmée
appelée apoptose (Park, Bissell et al. 2000). Notre corps élimine généralement ces cellules
grâce à nos cellules immunitaires telles que les macrophages et les cellules NK (<< natural
killer »), mais il arrive parfois qu'elles deviennent résistantes, les rendant aptes à .se diviser
indéfiniment. Les principales caractéristiques des cellules cancéreuses sont d 'abord leur
caractère d'immortalité et leur pouvoir absolu de division cellulaire, mais aussi, leur
capacité à envahir d'autres tissus du corps humain (Mareel and Leroy 2003) (Figure 1.1).
Figure 1.1 - Progression tumorale. (a) Cellule acquérant une mutation; (b) Prolifération; (c) Cellule acquérant une seconde mutation; (d) Capacité invasive du cancer; (e) Infiltration d'une cellule dans un vaisseau sanguin; (f) Accumulation de cellules cancéreuses formant un petit embole dans le vaisseau sanguin (image tirée de Louvard et Thiery, 2003) . .
Les cellules cancéreuses possèdent également la capacité de modifier ' leur
environnement (Hanahan and Weinberg 2000), leur permettant ainsi de proliférer et de
progresser pour mener à la fonnation d'une tumeur maligne. Des cancers peuvent survenir
dans toutes les cellules du corps et leur degré d'agressivité peut varier selon le type
cellulaire atteint. Ils peuvent, par exemple, atteindre des endroits essentiels à la survie, tels

2
que le foie, le cerveau ou le pancréas. Il arrive tout de même parfois qu'ils atteignent des
endroits plus subtils et d'apparence moins dangereux. En effet, certains cancers peuvent
parfois mettre en péril la survie des patients en dépit du fait qu'ils semblent inoffensifs.
L'œil, cet organe des sens d'une incroyable complexité, en est un bon exemple, car bien
qu'il ne soit pas nécessaire à la survie, il revêt tout de même son importance notamment
dans la qualité de vie des patients.
Les cancers de l'oeil ne sont certainement pas parmi les cancers les plus fréquents dans
la population adulte. À titre indicatif, ces cancers représentent 0,01% de tous les cancers
répertoriés aux États-Unis et en Angleterre (Hakulinen, Teppo et al. 1978). Cependant,
l'étude des mécanismes sous-j acents à ses propriétés tumorigènes et métastatiques peut
aider à comprendre la progression d'autres formes de cancers et permettre d' en élucider
certaines particularités partagées.

3
1.2 Le mélanome uvéal
1.2.1 Généralités
Le mélanome uvéal correspond au type de tumeur intraoculaire le plus fréquent dans la
population adulte. Malgré sa rareté, environ 50% des patients atteints de ce type de cancer
trouveront la mort dans, au plus, les quinze années suivant le diagnostic initial de
mélanome uvéal (Kincaid 1998). Le mélanome uvéal n'est pas le cancer le plus commun,
son incidence annuelle n'étant que d'environ 7,3 nouveaux cas par million d'habitants
(Vidal, Bacin et al. 1995). Bien qu'une personne de tout âge et de toute race peut en être
atteinte, il semble y avoir une prépondérance pour le caucasien adulte âgé de 50 à 70 ans
(Hungerford 1995; Kincaid 1998). Ce cancer a déjà été diagnostiqué chez des patients de
moins de 30 ans, ces cas ne représentant cependant que 4% de l'ensemble des individus
atteints (McCartney 1995). De plus, ce type de cancer est très rare chez les noirs et les
asiatiques (McCartney 1995). En effet, il est retrouvé environ 150 fois plus fréquemment
chez .les individus blancs que chez les noirs (Hakulinen, Teppo et al. 1978; Singh and
Topham 2003). Finalement, il semble plus fréquent chez les personnes présentant un iris
pâle et des cheveux blonds que chez ceux ayant des yeux et des cheveux bruns ou plus
foncés (Shields, Annesley et al. 1977; Guenel, Laforest et al. 2001).
1.2.2 Anatomie générale de l'œil
L' œil est le seul organe des sens qui permet à l'animal de transformer la lumière en
images pour lui permettre de voir. Comme c'est le cas chez la plupart des mammifères,
l' œil humain comporte trois tuniques : la sclère, l'uvée et la rétine (Figure 1.2). La sclère,
ou sclérotique, est la partie blanche externe de l'œil qui est prolongée par la cornée et qui
protège l' œil des agressions mécaniques en plus de maintenir la pression intraoculaire.
L'uvée, ou tractus uvéal, est une tunique vasculaire pigmentée qui comprend l'iris~ le corps
ciliaire et la choroïde (Kincaid 1998). Finalement, la rétine comprend des cellules
ganglionnaires, des cellules bipolaires, des intemeurones (cellules amacrines et
horizontales), des photorécepteurs (cônes et bâtonnets), des . cellules de soutien (cellules

4
gliales de Müller) ainsi qu'un épithélium pigmentaire rétinien (EPR) pour permettre la
transformation de la lumière en images par notre cerveau (Young 1976).
Corps dliaire
Ugament
suspenseur
Cornée
Iris
Pupille
Humewaqueuse
Cristallin
Humeur vitrée
Uvée
r sdère
J. Choroide
Fovéa
Poin~
aveugle
Figure 1.2 - La structure générale de l'œil humain. (adaptée de Addison, Wesley et Longman, 1999).
De la partie antérieure vers la partie postérieure, l' œil se compose premièrement de la
cornée, une lentille transparente qui le protège de l'environnement extérieur et est capable
de concentrer la lumière sur le cristallin. On retrouve ensuite l'humeur aqueuse, un liquide
à faible viscosité responsable en partie du maintien de la pression intraoculaire et de la
forme globulaire de l'œil. L'iris, partie intégrante de l'uvée, est une membrane contractile,
tel un diaphragme, qui permet de modifier la quantité de lumière . pénétrant l'œil en
modifiant la taille de son orifice central, la pupille (Kincaid 1998). Le cristallin est une
lentille biologique biconvexe qui permet la mise au point et la formation d'une image sur la
rétine. Le corps ciliaire et la choroïde sont des structures musculaires qui se regroupent
avec l'iris pour former le tractus uvéal (Kincaid 1998). Le corps ciliaire permet

5
l'attachement du cristallin à la choroïde et cette dernière, richement vascularisée, est le tissu
nourricier de l' œil apportant l'oxygène et les nutriments aux cellules (Kincaid 1998). Elle
se compose principalement de quatre couches : la lamina fusca (suprachoroïde), le stroma,
la choriocapillaris et la membrane de Bruch (Kincaid 1998). La première couche est
constituée d 'un tissu ,?onjonctif élastique, de vaisseaux sanguins ainsi que de cellules
pigmentées appelées mélanocytes. Ces cellules présentent une morphologie dendritique et
contiennent le mélanosome qui est responsable de la synthèse de la mélanine et qui donne
la couleur brunâtre caractéristique à la choroïde (Boissy 1988). Le stroma contient
également plusieurs vaisseaux sanguins, mais de plus petit diamètre. La troisième couche
(choriocapillaris) contient un réseau fin de capillaires qui assure l'alimentation de la couche
externe de la rétine. La quatrième et dernière couche est la membrane de Bruch qui
représente la couche interne de la choroïde directement accolée à la couche pigmentaire de
la rétine et qui forme le lien entre cette dernière et la choroïde. Finalement, le point aveugle
ou tache aveugle correspond à l'endroit sur la rétine où s'insère le nerf optique et qui
permet de transmettre l'information visuelle jusqu'au cortex visuel, nous permettant ainsi
de voir des images (Schraermeyer, Addicks et al. 1998).
1.2.3 Types de mélanomes du tractus uvéal
Le mélanome uvéal peut se développer près de l'iris, près du corps ciliaire ou sur la
choroïde (Hungerford 1995).
1.2.3.1 Mélanomes de l'iris
Les mélanomes situés près de l'iris (Figure 1.3 A) comptent pour environ 3% de tous les
mélanomes du tractus uvéal malins (Damato 2006). Ce type de mélanome peut adopter un
patron nodulaire ou étendu avec un pourtour circonscrit ou diffus (Hungerford 1995). Étant
donné sa localisation, ce type de mélanome est le plus évident à repérer et les traitements
qui y sont associés varient beaucoup en fonction de son patron cellulaire (Kersten, Tse et al.
1985).

6
1.2.3.2 Mélanomes du corps ciliaire
Les mélanomes situés près du corps ciliaire (Figure 1.3 B) représentent
approximativement 7% de tous les cas de mélanome uvéal (Damato 2006). Les tumeurs
localisées près du corps ciliaire peuvent également être nodulaires ou diffuses. Il s 'agit du
type le plus difficile à diagnostiquer et la tumeur est souvent découverte à un stade plus
avancé, c'est-à-dire lorsqu'elle cause des troubles visuels ou lorsqu'elle produit un
détachement de la rétine suite à l'envahissement de la choroïde (Hungerford 1995).
1.2.3.3 Mélanomes choroïdiens
Le mélanome choroïdien (Figure 1.3 C) est celui le plus fréquemment observé (Scotto,
Fraumeni et al. 1976), correspondant à près de 90% de tous les cas répertoriés (Damato
2006). La plupart de ces tumeurs adoptent un patron nodulaire et une forte corrélation
existe entre la taille de la tumeur et le faible taux de survie des patients.
Figure 1.3 - Différents types de mélanomes. (À) Mélanome situé au niveau de l'iris; (B) Mélanome situé près du corps ciliaire; (C) Mélanome situé dans la choroïde (images tirées de www.etrf.org; "looc.org.uk; www.kimme1cancercenter.org).
1.2.3.4 Type cellulaire
Il est également possible de diviser les différents types de mélanomes du tractus uvéal
selon le ou les types cellulaires qui les constituent. Il existe trois grands profils qui peuvent
caractériser l ' apparence d 'une tumeur, soient les profils épithélioïde, fusiforme et mixte.

7
1.2.3.4.1 Profil fusiforme
Les cellules dites fusiformes sont plus longues et présentent des noyaux rayés (Figure
1.4 A). Cette classe se subdivise en deux types distincts, soit le type fusiforme A et
fusiforme B (Kincaid 1998). Le premier type (A) est le plus près du type bénin et se
présente avec un noyau plus fin. Le second type possède un noyau à forme plus arrondie
(Kincaid 1998).
1.2.3.4.2 Profil épithélioïde
Le profil dit épithélioïde présente des cellules plus volumineuses, ayant une forme
d'apparence cubique avec un plus grand volume de cytoplasme ainsi qu'un plus grand
noyau (McCartney 1995) (Figure 1.4 B). Ce profil n'est toutefois pas associé à un très bon
pronostic (Kincaid 1998)· et signifie souvent une plus grande agressivité des cellules
cancéreuses.
1.2.3.4.3 ProfIl mixte
Le profil mixte contient à la fois des cellules fusiformes et des ~ellules épithélioïdes. On
peut même associer l'agressivité de la tumeur avec le rapport entre les deux types de
cellules, car il semblerait que plus la tumeur contient de cellules épithélioïdes, plus elle est
susceptible d'être agressive (McLean, F oster et al. 1977).
Figure 1.4 - Différentes morphologies des cellules de mélanome uvéal. (A) Cellules fusiformes; (B) Cellules épithélioïdes (images tirées de Kincaid, 1998).

8
1.2.4 Causes
Le cancer apparaît suite · à la transformation des mélanocytes, cellules pigmentaires
situées dans le tractus uvéal. Ces dernières sont reconnues pour sécréter deux types de
mélanine soit l'eumélanine, d'une coloration qui va du beige au noir, et la phéomélanine,
. plus pâle et davantage jaune et rougeâtre. (Boissy 1988). Contrairement à ce qu 'on pourrait
penser, les facteurs environnementaux ne semblent pas Jouer un rôle dans la formation de
ce cancer. Plus particulièrement, les rayons ultraviolets connus pour endommager et
augmenter les risques de cancer de la peau, ne semblent pas prédisposer au développement
de la tumeur uvéale (Dolin, Foss et al. 1994; Moan, Porojnicu et al. 2008) et ne semblent
pas y être associés directement (Lutz, Cree et al. 2005). Toutefois, le mélanome uvéal
semble fortement associé aux mélanocytoses oculaires, notamment le Naevus d 'Ota, une
prolifération mélanocytaire ocul~ire et dermique touchant la conjonctive et les paupières en
plus de la peau adjacente, des sclérotiques et des muscles oculaires (Gonder, Shields et al.
1982).
Malgré la présence de quelques cas familiaux de mélanome uvéal répertoriés
(Oosterhuis, Went et al. 1982), ce type de cancer ne semble pas nécessiter une
prédisposition génétique et apparaît comme étant une tumeur sporadique (Kincaid 1998).
De plus, aucune mutation aléatoire sur plusieu~s chromosomes n'a été répertoriée, mais une
monosomie du chromoso~e 3 et ~ne duplication du chromosome 8q semblent davantage
associées au mélanome uvéal du corps ciliaire (Sisley, Cottam et al. 1992).
1.2.5 Diagnostic et traitements
1.2.5.1 Diagnostic de la tumeur primaire
La plupart des tumeurs du tractus uvéal sont généralement asymptomatiques et peuvent
souvent être dépistées pendant un simple examen de routine chez l' optométriste ou
l'ophtalmologiste (Kincaid 1998). Bien que les patients atteints ne présentent
habituellement aucun symptôme, les mélanomes situés près de l'iris entraînent parfois un
glaucome secondaire tandis que ceux situés près du corps ciliaire causent également de

9
l'astigmatisme ou des cataractes (Damato 2006). Qui plus est, les patients ayant un
mélanome choroïdal souffrent parfois aussi d'une perte de l'acuité visuelle ou d'une vision
embrouillée due au bris de l' épithélium pigmentaire rétinien ou du décollement de la rétine
(Damato 2006).
1.2.5.2 Choix du traitement
Traditionnellement, les patients atteints d'un mélanome uvéal subissaient l' énucléation
complète de l'œil touché. Aujourd'hui, certains centres spécialisés pour ce type de cancer
privilégient des techniques moins radicales permettant de conserver l'œil si les conditions
sont favorables. Il arrive même que le patient réussisse à conserver une certaine capacité
visuelle de l'œil touché. Néanmoins, et en dépit des avancées technologiques, il arrive que
le moyen choisi soit encore l' énucléation, mais combinée à d 'autres avenues afin de
s'assurer d'éradiquer complètement la tumeur. Le choix parmi les divers traitements
possibles est fait en fonction de plusieurs critères précis pouvant influencer les diverses
alternatives qui s'offrent au patient. De ces critères, on note plus particulièrement la taille et
l'origine de la tumeur, le patron de croissance, l'état du second œil, l'âge ainsi que la santé
générale du patient (Hungerford 1995).
1.2.5.2.1 La notion de taille
La notion de taille est un critère très important et précis qui peut donner beaucoup
d'indices sur la conduite à prendre pour détruire la tumeur. Un mélanome uvéal est
considéré comme petit si son diamètre est inférieur à 10 mm et possède une épaisseur
maximale de 3 mm; moyen si son diamètre se situe entre 10 et 15 mm et son épaisseur
entre 3 et 5 mm et, finalement, il est considéré comme gros s'il excède 15 mm de diamètre
et 5 mm d'épaisseur (McCartney 1995). Si la tumeur est de petite taille (moins de 5 mm),
les médecins préféreront une période de surveillance à tout autre traitement afin de voir
l'évolution de la tumeur (McCartney 1995). Ce choix peut s'expliquer en partie par la
controverse existant sur le fait que l'énucléation pourrait favoriser la migration de cellules
canc~reuses à travers les vaisseaux sanguins et la formation de métastases au niveau d'un
site secondaire (Zimmerman, McLean et al. 1978).

10
1.2.5.2.2 Autres traitements possibles
Les diverses options qui ont évolué parallèlement à l'énucléation incluent la
radiothérapie par plaque, la radiothérapie avec l'aide de particules chargées, la
photocoagulation par laser, la thermothérapie à travers la pupille (TTT) , l'ablation locale
(Journee-de Korver and Keunen 2002), la chimiothérapie (Char 1989), la cryothérapie
(Fraunfelder, Boozman et al. 1977) et l ' immunothérapie (McLean, Berd et al. 1990). Ces
nouvelles techniques ont donc permis de restreindre l'énucléation aux plus grosses tuineurs,
à celles ne répondant pas aux traitements ou aux tumeurs récurrentes. Cette pratique,
autrefois employée dans 95% des cas, a vu son utilisation diminuer drastiquement jusqu'à
moins d'un cas sur cinq aujourd'hui (21 % des cas en 1991) (Shields, Shields et al. 1991 ;
Seregard 1996) afin de non seulement préserver la qualité de vie des patients, mais
également, de prévenir d'éventuelles complications qui pourraient survenir suite à
l'énucléation. Depuis la dernière décennie du 20e siècle, la combinaison de ces différentes
options demeure encor~ et toujours le meilleur traitement possible pour les patients atteints
d'un mélanome uvéal (De Potter, Shields et al. 1996; Kincaid 1998; Shields, Shields et al.
1998).
1.2.6 Facteurs pronostics
L'espérance de vie des patients atteints d'un mélanome uvéal varie beaucoup d'un
individu à l'autre. En effet, environ la moitié des persoruies atteintes trouveront la mort
dans les quinze années suivant le diagnostic (Kincaid 1998). L'évolution de la maladie et
l'avenir du patient dépendent beaucoup de la taille et de la localisation de la tumeur et si
cette dernière a été détectée de façon précoce.
1.2.6.1 La taille de la tumeur
La taille de la tumeur informe généralement sur l'espérance de vie du patient: une
masse plus importante indique habituellement un mauvais pronostic (Flocks, Gerende et al.
1955) et une tumeur ayant un diamètre inférieur à 7 mm est associée à une diminution dans
le risque de décès. Le taux de survie moyen après 10 ans est d'environ 85,1% pour les
petites tumeurs tandis qu'il diminue à 73% chez les patients ayant une tumeur moyenne

Il
(Seregard 1999). Le taux de mortalité moyen après 5 ans pour les patients avec une
moyenne tumeur est d'environ 6-10% et grimpe à 22-27% pour ceux qui ont des tumeurs
de plus grande surface (Journee-de Korver et Keunen 2002). Un mélanome uvéal situé sur
l'iris semble être associé à un taux de mortalité inférieur à ceux situés sur la choroïde ou sur
le corps ciliaire (Hungerford 1995). Toutefois, ceci pourrait être lié au fait qu'étant situé sur
l'iris, il serait plus facilement et plus précocement détectable. La taille imposante de la
tumeur au diagnostic est certainement associée à un mauvais pronostic, mais sa position
antérieure semble également jouer contre la survie (Shammas et Blodi 1977). De plus, une
tumeur ayant un volume inférieur à 1 cm3 situé près du corps ciliaire laisse présager
habituellement de bonnes chances de survie pour le patient alors qu'une tumeur de même
envergure située sur la choroïde annonce un mauvais pronostic (Sorensen, Gamel et al.
1993).
1.2.6.2 Le type cellulaire et l'agressivité
Le type cellulaire et l'agressivité sont également de bons indicatifs pour prédire les
chances de survie du patient (McCartney 1995). Une tumeur démontrant des cellules
épithélioïdes semble davantage agressive qu'une tumeur à cellules fusiformes (McLean,
Foster et al. 1977; Shammas et Blodi 1977). Qui plus est, comme les tumeurs épithélioïdes
sont souvent associées aux tumeurs de plùs grands gabarits, il ne fait aucun doute que ces
deux facteurs sont indicateurs d'un' mauvais pronostic pour le patient (Davidorf et Lang
1975).
1.2.7 Métastases
1.2.7.1 Processus métastatique
Les métastases résultent de la migration des cellules cancéreuses, qui se sont détachées
de la tumeur primaire, à travers la paroi des vaisseaux sanguins où elles peuvent alors
gagner accès à d'autres organes ou tissus de l'organisme (Figure 1.5). Le diagnostic de
métastases secondaires est souvent accompagné d'un mauvais pronostic de survie.
Grossièrement, la coopération temporelle et anatomique entre les . molécules d'adhésion

12
cellulaire, les protéases qui dégradent la matrice extracellulaire et certains peptides générant
la vascularisation tumorale est nécessaire pour permettre le développement de métastases
(Ahmad et Hart 1997).
A
Figure 1.5 - Processus métastatique. (A) Des cellules cancéreuses se détachent de la tumeur primaire et ga"gnent accès à la circulation sanguine; (B) Les cellules cancéreuses envahissent ensuite l'organe secondaire (image tirée de Sanofi-aventis, 2008).
De façon générale, le processus métastatique semble suivre un patron séquentiel qui
demeure sensiblement le même d'une tumeur à une autre. Celui-ci "débute par la croissance
de cellules néoplasiques et la néovascularisation de la tumeur. Par la suite, il. y a
détachement d'une cellule cancéreuse de la tumeur primaire, suivi de l'intravasation de
celle-ci dans un vaisseau sanguin (Figure 1.5 A). La cellule cancéreuse procède à
l'extravasation du vaisseau pour pénétrer à l'intérieur de l'organe secondaire et, finalement,
s 'y établir et y proliférer (Fidler 1991; Ahmad et Hart 1997) (Figure 1.5 B).
1.2.7.2 Pronostic et causes
Dans le cas du mélanome uvéal, un patient sur deux sera atteint de métastases
secondaires et dans 90% des cas, elles nidifieront au foie (Diener-West, Hawkins et al.

13
1992). Lorsque des métastases sont détectées, l'espérance de vie du patient se trouve
réduite à quelques mois et dépasse très rarement deux ans (Leyvraz, Spataro et al. 1997). Le
mélanome uvéal situé près de l' iris semble moins prédisposé à générer des métastases, mais
l~ résection de la tumeur pourrait permettre à certaines cellules cancéreuses de proliférer et
de traverser la barrière des vaisseaux sanguins pou~ ensuite envahir d'autres organes
(Bechrakis et Lee 1991). Toutefois, certaines évidences semblent démontrer que des
métastases invisibles à l'œil nu seraient présentes jusqu'à 5 ans avant le diagnostic de la
tumeur primaire (Eskelin, Pyrhonen et al. 2000). Ceci éliminerait donc la possibilité que
dans certains cas, les métastases ne surviennent qu'à la suite du traitement administré à la
tumeur primaire (Singh 200 l) ~ Le processus métastatique pourrait s'expliquer notamment
par la présence de molécules d'adhésion à la surface des cellules, comme les intégrines, qui
pourraient permettre à la cellule de migrer jusqu'à l'organe cible (Marshall, Rutherford et
al. 1995).

14
1.3 Les molécules d'adhésion
Les molécules d'adhésion sont responsables de la liaison entre les cellules et la matrice
extracellulaire (MEC) ainsi que des interactions cellules-cellules. La capacité des cellules à
interagir par le biais de leurs molécules d'adhésion joue un rôle clé dans plusieurs
processus biologiques impliquant l 'homéostasie, le développement, l 'embryogenèse,
l'inflammation et la réponse immunitaire (Frenette et Wagner 1996; Petruzzelli, Takami et
al. 1999). Considérant l'importance de ces derniers, il est raisonnable de croire qu 'un
dérèglement dans un de ces processus peut jouer un rôle dans la progression du cancer et
des métastases. En effet, plusieurs études indiquent que certaines familles de molécules
d'adhésion sont impliquées dans la tumorigenèse, dans la progression tumorale et dans le
processus métastatique (Ohene-Abuakwa et Pignatelli 2000). De ces familles, on compte
les cadhérines, les immunoglobulines, les sélectines et, finalement, les intégrines (Figure
1.6).
1.3.1 Les sélectines
La famille des sélectines se compose de glycoprotéines transmembranaires contenant un
domaine N-tenninal calcium-dépendant unique (Figure 1.6 A). Elles possèdent également
un domaine similaire au facteur de croissance EGF qui exerce une fonction importante dans
la régulation de son activation (Bock 1991).
Leur fonction principale se traduit par des interactions hétérotypiques entre cellules. En
effet, elles pennettent l'adhésion des cellules sanguines et des leucocytes avec les cellules
endothéliales des vaisseaux sanguins (Ohene-Abuakwa et Pignatelli 2000). Il existe
seulement trois membres dans cette famille qui sont la E-sélectine (endothéliale), la P
sélectine (plaquette) ainsi que la L-sélectine (leucocyte).
Il a été démontré que l'inhibition des fonctions de la E-sélectine hépatique dans un
modèle de souris entraîne une diminution dans le nombre de métastases au foie résultant de
l'implantation d'un carcinome du côlon (Brodt, Fallavollita et al. 1997; Khatib, Fallavollita
et al. 2002). L'adhésion des cellules cancéreuses à la paroi des vaisseaux sanguins leur

15
permettraient donc de traverser la parOI endothéliale afin d'envahir un site potentiel
secondaire.
A Domaine
NH :1-T erm inal
R é~on similaire au facteur de
croissanc e EG F
Répétitions des séquences deSpfotéinesré~aUcices
du C om plém ent
Dom aine intra cellulaire
c
cadhérine~
spécifiques
Domaines transm em brana1res
Dom ainesintracellu1aireS~~iI.
C atérunes
B
Dom aines uansm em brana.ire s
D
Dom aine calf-l
Dom aine calf-2
Pont disulfure
Domaines « 1 m m uno glubuline-lik e »
~Dolmaines ~ Fibronectine III
Dom aine intracellulaire
Dom aine Hybride Dom aine PSI
Figure 1.6 - Molécules d'adhésion. Représentation schématique de la structure des principales molécules d'adhésion responsables de l'interaction cellule-cellule et cellule-matrice; (A) Sélectine; (B) Immunoglobuline de type N-CAM; (C) Cadhérine; (D) Intégrine (image adaptée de Juliana, 2002).

16
1.3.2 La superfamille des immunoglobulines
La grande famille des immunoglobulines s'appelle ainsi en raison de leur domaine
extracellulaire « immunoglobuline-like » (Figure 1.6 B). Ce domaine se compose de
multiples chaînes . en boucles reliées par des ponts disulfures (Joseph-Silverstein et
Silverstein 1-998). Certaines de ces molécules contiennent également un domaine contenant
des répétitions de la séquence de la fibronectine III qui est reconnu pour avoir un rôle à
jouer dans la reconnaissance et la liaison de la cellule (Ohene-Abuakwa et Pignatelli 2000).
Ces molécules transmembranaires sont responsables non seulement de l' adhésion, mais
également de la migration des cellules (Beckner, Krutzsch et al. 1995) et ce, par des
interactions cellule-cellule. Parmi les membres de cette superfamille, on compte notamment
ICAM~1 (intercellulaire), VCAM-l (vasculaire) et NCAM (neural).
Certains membres de la famille des immunoglobulines ont vu leur expression augmentée
de façon aberrante à la surface de cellules malignes chez différents types de cancers (Maio,
Pinto et al. 1990; Passlick, Izbicki et al. 1994; Wolff, Stephenson et al. 1995). D ' autres
études semblent également démontrer que la surexpression de ICAM -1, par exemple, serait
associée à la progression de mélanomes et augmenterait le risque de métastases (Johnson,
Stade et al. 1989).
1.3.3 Les cadhérines
Les cadhérines constituent des glycoprotéines transmembranaires (Figure 1.6 C) qui
participent à l'adhésion cellule-cellule par une interaction homotypique calcium
dépendante (Yap, Niessen et al. 1998).
Ces récepteurs sont des déterminants fondamentaùx de l'organisation tissulaire des
organismes adultes ou en développement (Ohene-Abuakwa et Pignatelli 2000). Il existe
plusieurs membres dans la famille des cadhérines qui sont divisés en deux sous-groupes
soient les classiques ou celles associées aux desmosomes. On retrouve, entre autre, la E-

17
(épithéliale), la P- (placentaire) et la N- (neuronale) cadhérine qui font partie du type
classique (Moles et Watt '1997; Hatta, Takagi et al. 1987).
Les fonctions des cadhérines sont généralement régulées grâce à leur domaine
cytoplasmique via les caténines (a, ~ et y), des protéines intégrées au cytosquelette
(Gumbiner et McCrea 1993). Les cadhérines sont considérées comme des gènes
suppresseurs de tumeur et l ' inhibition de leurs fonctions ou de leur expression semble être
reliée à la progression tumorale (Ohene-Abuakwa et Pignatelli 2000).
1.3.4 Les intégrines
1.3.4.1 Structure des intégrines
Les intégrines sont des récepteurs transmembranaires localisés à la surface des cellules
(Figure 1.7). Ces dernières sont constituées d'une sous-unité a et d'une sous-unité ~ qui
s'hétérodimérisent de façon non-covalente afin d'acquérir leurs propriétés fonctionnelles
(Vigneault, Zaniolo et al. 2007).
Domaine H}imde
Domaine PSI
I-BGF (l a 4)
Domsiœ~
Figure 1.7 - Structure générale des intégrines. Représentation schématique de la structure de l'intégrine. On peut distinguer la sous-unité a et la sous-unité P ainsi que leurs principaux éléments structurants (image adaptée de Juliana, 2002).

18
Les sous-unités des intégrines sont des protéines transmembranaires de type l , c ' est-à
dire que leur extrémité N-terminale est extracellulaire tandis que leur extrémité C-terminale
est intracellulaire (Kuphal, Bauer et al. 2005). De plus, les sous-unités sont formées
intégralement de trois domaines distincts : un domaine extracellulaire ou exodomaine, qui
contient entre 700 et 1100' résidus, un court domaine transmembranaire de 20 à 30 résidus
et finalement, un domaine intracellulaire contenant environ 30 à 50 résidus (exception faite
de la sous-unité ~4 qui possède plus de 1 000 résidus) aussi appelé domaine cytoplasmique
(Kuphal, Bauer et al. 2005) .
. À ce jour, 24 sous-unités a différentes ainsi que 9 sous-unités ~ ont été identifiées dans
le génome humain. Toutefois, seulement 18 sous-unités a et 8 sous-unités ~ sont
actuellement répertoriées, celles-ci pouvant s'associer pour former 24 intégrines différentes
(Figure 1.8) (Hynes 1992; Meenan, Spaans et al. 1997; Plow, Haas et al. 2000).
Figure 1.8 - La famille des intégrines. Chacune des 24 intégrines représentées en hétérodimère composé d'une sous-unité a et une sous-unité ~. Les intégrines exprimées dans l'œil sont illustrées à l'intérieur du schéma oculaire. Orange: sous-unité ~; Vert: ligand ~ fibronectine; Rouge: ligand ~ collagène; Jaune: ligand ~ laminine; Violet: ligand ~ ténascine; Bleu: récepteur spécifique aux leucocytes (image tirée de Vigneault, 2007).

19
Les intégrines peuvent être divisées en famille selon leurs similarités fonctionnelles. On
retrouve donc les récepteurs RGD, les récepteurs des collagènes, les récepteurs des
laminines et les récepteurs des leucocytes (Hynes 2002). On peut regrouper les intégrines
selon la nature de leur sous-unité B. La famille BI est la plus importante et les intégrines
qui appartiennent à celle-ci ont principalement pour ligand des protéines de la MEC et sont
impliquées dans les interactions cellule-cellule (Alb~lda 1991).
1.3.4.1.1 Sous-unité a
Les sous-unités u représentent, le plus souvent, la partie la plus volumineuse de
l'hétérodimère. Elles ont une taille se situant entre 120 et 180 kDa et contiennent plus de
940 résidus (Petruzzelli, Takami et al. 1999; Berman et Kozlova 2000) (Figure 1.7). Il
existe non seulement beaucoup de variations inter-espèces, mais également intra-espèces du
point de vue de la séquence des ces sous-unités (Berman et Kozlova 2000).
Ces sous-~nités possèdent un domaine en forme d'hélice à sept lames, un domaine
« thigh », deux domaines «calf» et certaines possèdent également un domaine uA (ou
domaine 1) (Figure 1.7) (Arnaout, Mahalingam et al. 2005). Le premier de ces domaines
semble avoir un rôle à jouer dans la liaison de l'intégrine à son ligand, mais aussi dans
1 'hétérodimérisation du récepteur (Shimaoka, Takagi et al. 2002; Humphries, McEwan et
al. 2003). De plus, il contient des sites potentiels de liaison pour des cations divalents
comme le calcium, permettant ainsi son activation. (Hum_phries, McEwan et al. 2003). Le
domaine l, lorsque présent sur la sous-unité, semble lui aussi être impliqué dans la liaison
du ligand (Shimaoka et Springer 2003). Pour les sous-unités ne contenant pas ce domaine,
la sous-unité B ainsi que le domaine à sept lames semblent compenser pour cette absence
lors de la liaison du ligand (Kuphal, Bauer et al. 2005). La queue des sous-unités u contient
un domaine « thigh» homologue à l'immunoglobuline suivi de deux domaines, calf-l et
calf-2, disposés en feuillets-B qui correspondent au site de clivage protéolytique (Amaout,
Mahalingam et al. 2005). Finalement, près du domaine transmembranaire, on retrouve chez
toutes les sous-unités u la séquence particulière KXGF-FKR/X où X sont des résidus non
polaires (Berman et Kozlova 2000). Cette séquence, essentielle à l 'hétérodimérisation,

20
maintient les intégrines dans un état inactif et est très importante pour l'activation de ces
dernières (van der Flier et Sonnenberg 2001).
1.3.4.1.2 Sous-unité Il Les sous-unités ~ sont généralement plus petites que les sous-unités u et possèdent une
taille variant entre 90 et 115 kDa avec environ 640 résidus (Petruzzelli, Takami et al. 1999;
Shimaoka et Springer 2003) (Figure 1.7). Contrairement à sa sous-unité complémentaire,
cette sous-unité possède une séquence plus conservée entre les espèces (Albelda 1991 ;
Berman et Kozlova 2000).
La portion extracellulaire de la sous-unité ~ peut être subdivisée en 8 domaines
différents qui, bien que toutes leurs fonctions ne soient pas connues encore aujourd'hui,
semblent néanmoins importants compte tenu de leur grande conservation entre les espèces
(Figure 1.7) (Berman et Kozlova 2000). Le premier domaine consiste en un domaine
globulaire .dénommé ~A et inséré en partie dans le second domaine, le domaine hybride. Le
domaine ~A sert à compenser l'absence du domaine uA de la sous-unité u pour la liaison
du ligand (Shimaoka, Takagi et al. 2002; Shimaoka et Springer 2003). Le domaine hybride,
homologue à l'immunoglobuline, serait impliqué dans l'activation de l'intégrine (Arnaout,
Mahalingam et al. 2005). Le troisième domaine, le domaine PSI, riche en résidus cystéines,
semble être responsable du maintien de la position inactive de l'intégrine, contrairement au
second domaine (Shimaoka, Takagi et al. 2002). Les quatre domaines suivants surnommés
1 -EGF 1 à 4, sont également riches en résidus cystéines et semblent être importants au
niveau de la signalisation (Shimaoka, Takagi et al. 2002). Le dernier domaine de la portion
extracellulaire est le domaine ~TD dont la fonction est encore mal définie à ce jour
(Arnaout, Mahalingam et al. 2005). Finalement, deux séquences très conservées semblent
aussi impliquées dans l'activation de l'intégrine. Il y a tout d'abord la séquence de
localisation des plaques d'adhésion focales NPXY et une séquence située près de la queue
cytoplasmique de la sous-unité ~ (Dedhar and Hannigan 1996).
La queue cytoplasmique de la sous-unité ~ possède la capacité de lier plusieurs protéines
cytoplasmiques dont la taline, la filamine et l' a-actinine, qui pennettent un lien direct entre

21
les intégrines et le cytosquelette d' actine, les protéines F AK et ILK liées à la signalisation
et finalement, la paxilline et la GRB2 qui sont des protéines adaptatrices également
importantes dans la signalisation (Juliano et Haskill 1993; Schaller et Parsons 1994;
Giancotti et Ruoslahti 1999).
1.3.4.2 Ligands des intégrines
Les différentes combinaisons des sous-unités d'intégrines démontrent la capacité
d'induire diverses voies de signalisation, et ce, selon le ligand auquel elles se lient. Une
intégrine peut non seulement être reconnue par plusieurs ligands, mais plusieurs intégrines
peuvent également lier ce même ligand (Elner et Elner 1996). La plupart de leurs ligands
sont des protéines de la MEC impliquées dans l'adhésiori cellule-substrat (Vigneault,
Zaniolo et al. 2007). Toutefois, les intégrines peuvent également lier leurs contre-récepteurs
connus, des membres de la grande famille des immunoglobulines (ICAM et VCAM),
permettant ainsi l'adhésion cellule-cellule (Hynes 1992). La liaison du ligand à une
intégrine permet le passage de la forme inactive à la forme activée de celle-ci.
1.3.4.2.1 Matrice extracellulaire
La MEC fournit un support mécanique aux ·cellules et représente un réseau
tridimensionnel entrecroisé constitué de plusieurs macromolécules (Bosman et
Stamenkovic 2003). Le contact entre les cellules et la matrice permet la création d'une
barrière perméable en plus de maintenir l'intégrité tissulaire (Albelda 1991). Ce~e matrice
interagit avec les cellules afin d'activer et de réguler certaines fonctions cellulaires
importantes telles que la migration, l'adhésion, la prolifération, la différenciation ainsi que
la morphogenèse (Adams et Watt 1993; DeSimone 1994; Roskelley, Srebrow et al. 1995).
Plus précisément, la MEC est constituée de glycoprotéines, de collagènes, de
glycosaminoglycanes et de protéoglycan~s qui forment un enchevêtrement de fibres de
collagènes et de glycoprotéines (Zagris 2001). La MEC est une entité dynamique dont la
composition change . constamment . dans le but de permettre la modification du
comportement cellulaire via la signalisation par les récepteurs situés à la surface cellulaire
(Streuli 1999).

22
La MEC contient une variété de glycoprotéines connues sous le nom de fibronectine,
tenascine, entactine et laminine. Ces demièrés auront un rôle à jouer dans l'adhésion entre
la cellule et ' le tissu à un moment ou l'autre du développement (Zagris 2001). Les )
collagènes contenus dans la MEC (types l à XIX) en constituent quant à eux la composante
la plus abondante (Olsen 1995). Les glycosaminoglycanes comme l'acide hyaluronique, le
sulfate de chondroïtine, le sulfate d 'héparane et le kératane sulfate sont impliqués dans le
contrôle de la prolifération, de la migration, de la différenciation ainsi que dans le maintien
des structures morphqgéniques (Zagris 2001). Pour leur part, les protéoglycanes présents
dans la MEC peuvent être divisés en trois groupes distincts: les protéoglycanes de la
membrane basilaire (perlécan, bamacan, agrine), les hyalectanes (aggrécan, versican) et les
lectines (décorine, biglycan). Ces derniers peuvent notamment avoir un rôle à jouer dans la
prolifération cellulaire, mais aussi dans la maturation des tissus spécialisé~ (Iozzo 1998).
1.3.4.3 Rôles des intégrines
Les fonctions des intégrines traduisent par elles-mêmes l'importance qu'elles occupent
chez l'humain. Elles jouent non seulement un rôle dans la migration et dans l'adhésion des
cellules, mais elles affectent également, par signalisation intracellulaire, l'expression de
plusieurs gènes qui sont importants pour la différenciation; la prolifération et dans
l'organisation du cytosquelette (Hotchin et Hall 1995). Par des interactions cellule-cellule 1
ou cellule-MEC (Hynes 1992), les intégrines peuvent permettre la transduction de signaux
de l'extérieur vers l'intérieur de la ,cellule (<< outside-În signaling ») ou de l'intérieur vers
l'extérieur (<< inside-out signaling ») (Vigneault, Zaniolo et al. 2007). L'activation et la
signalisation des intégrines surviennent suite à un changement de conformation de ces
dernières (Giancotti 2003). En d'autres mots, le lien étroit entre l'extérieur et l'intérieur de
la cellule permet de répondre adéquatement aux changements par les intégrines qui
participent activement à des processus clés du cycle cellulaire tels que la survie et la mort
cellulaire. L' anoïkose, par exemple, est un phénomène qui traduit bien leur importance
dans la mort cellulaire. Il survient lorsque les interactions des cellules avec la matrice sont
instables ou absentes provoquant en retour la transmission d'un signal qui enclenc~e
l'apoptose (Frisch et Ruoslahti 1997). Les intégrines empêchent cette mort des cellules par

23
apoptose (anoïkose) lorsque celles-ci sont liées à la MEC dans l'environnement approprié
(Stupack et Cheresh 2002).
1.3.4.4 Signalisation par les intégrines
Les intégrines ont la capacité d'effectuer une signalisation bi-directionnelle (Danen et
Sonnenberg 2003). La liaison de la MEC provoque l'association des intégrines avec le
cytosquelette et active une cascade de signalisation à l'intérieur de la cellule (Figure 1.9)
(Giancotti et Ruoslahti 1999). Inversement, les signaux intracellulaires peuvent induire la
liaison entre les intégrines et la MEC (Liddington et Ginsberg 2002).
F Paxil line FSAK
J / rc ~ CAS !
CrK / PI31K ~ .~
DOC K 180 AKT! P ~ , KB
R,ac
~ PAK
~ JNK
'-.. Soc
/ Grb2-Sos
~ Ras ~
Raf-1
~ EK
~ ErK
Figure 1.9 - Signalisation par les intégrines. Lorsqu'elles sont activées par les protéines de la MEC, les intégrines activent diverses voies de signalisation dans la cellule (image adaptée de Hynes, 2002).
Les queues cytoplasmiques des intégrines jouent un rôle fondamental dans la
signalisation par les intégrines. Toutefois, étant donné qu'elles sont dépourvues d'activité
catalytique, elles doivent se lier à plusieurs protéines adaptatrices afin d'effectuer une
cascade de signalisation. Ces queues correspondent donc aux sites d'interaction de

24
plusieurs protéines cytoplasmiques et du cytosquelette d'actine (Hynes 2002). Seules, c ' est
à-dire lorsqu'elles ne sont pas liées à leur ligand, les intégrines diffusent librement dans la
membrane plasmique. Lorsqu'une intégrine se lie à la MEC, il survient une réorganisation
des filaments d'actine suivie d'une boucle de rétroaction positive qui amène d'autres
intégrines à s'assembler en liant plus fortement la MEC (Jockusch, Bubeck et al. 1995;
Aplin, Howe et al. 1998). Cette aggrégation de plusieurs intégrines au niveau de la
membrane cellulaire se nomme « complexe d'adhésion focal». La propagation du signal
implique plus de 50 protéines différentes qui provoquent une cascade de signalisation
spécifique (Romer, Birukov et al. 2006). Certains facteurs influencent le taux
d'assemblage, la taille et la signalisation qui résultent de l' assemblage des complexes
d'adhésion. De ces facteurs, on compte notamment les propriétés de la MEC, l'activation et
l'avidité des intégrines, l'état de contraction du cytosquelette et la spécificité tissulaire et
cellulaire du milieu où se déroulent les évènements (Romer, Birukov et al. 2006).
La signalisation par les intégrines survient via un grand nombre de seconds messagers
intracellulaires (Figure 1.9). Les protéines adaptatrices comme la taline, la paxilline, la
vinculine, l' a-actinine, la filamine, l 'ILK ainsi que la « focal adhesion kinase» ou F AK
(N obes et Hall 1995), peuvent se lier à la queue cytoplasmique de la sous-unité B (Lee et
Juliano 2004; Wiesner, Legate et al. 2005). La F AK, en s' autophosphorylant, s' associe
fréquemment à une kinase membre de la famille Src qui enclenche diverses voies de
signalisation comme la voie des PI3K (AKT/PKB), la voie de Crk/CAS et la voie des
MAPK (Giancotti 2003; Schatzmann, Marlow et al. 2003). Il existe également d'autres
protéines capables de se lier à la protéine F AK dont la phospholipase C-y (PLC-y), la
protéine Grb7 et Shc qui activeront à leur tour d'autres voies de signalisation (Schatzmann,
Marlow et al. 2003; Wiesner, Legate et al. 2005). Il arrive également que des intégrines
activent les membres de la famille Src sans la liaison préalable de la F AK. Ces intégrines
couplées à des membres de la famille Src tels que Fyn et Yes vont recruter Shc pour
s'associer au complexe Grb2/S0S et provoqueront l'activation de Ras et de la voie des
MAPK (Giancotti 2003).

25
-1.3.5 La progression tumorale
Comme d'autres molécules d'adhésion cellulaire, les intégrines sont également
reconnues pour avoir un rôle à jouer dans le cancer. Étant des récepteurs d'adhésion
cellulaire, on peut facilement les associer à la progression tumorale. La transformation
cellulaire, qui mène à la formation d'une cellule cancéreuse, est justement caractérisée par
une rupture dans l'intégrité du cytosquelette et une diminution de la capacité d ' adhésion
cellulaire (Mizejewski 1999; van der Flier et Sonnenberg 2001). De plus, le bris des liens
entre les intrégrines et la MEC serait impliqué dans le processus métastatique, ce qui
permettrait aux cellules cancéreuses d'envahir un ou des orgaries secondaires (Wehrle
Haller et Irnhof 2003). Par ailleurs, plusieurs évidences suggèrent également que les
cellules cancéreuses auraient la capacité de modifier le patron d'expression de certaines
intégrines à leur surface (Giancotti et Mainiero 1994). Le niveau d'expression de certaines
intégrines serait, de ce fait, augmenté alors que celui d'autres intégrines serait diminué afin
de permettre l'invasion, la progression et la migration des cellules cancéreuses (Mizejewski
1999; Kuphal, Bauer et al. 2005). Notons plus particulièrement l'intégrine a4B1 qui semble
jouer un rôle notamment dans la croissance tumorale et dans les processus métastatiques
lorsque sur-exprimée (Bao, Pigott et al. 1993) et l'intégrine a5B1 connue pour être associée
à un phénotype plus agressif lorsqu'elle est soit sur-exprimée (Gong, Wang et al. 1997) ou
encore réprimée (Sanders, Mainiero et al. 1998) chez certains types de cancer.
De plus, plusieurs études ont rapporté l'expression de certaines intégrines associées au
mélanome uvéal dont l'intégrine avB3 (Seftor, Seftor et al. 1999) qui est fortement
exprimée chez les cellules au profil fusiforme provenant d'un mélanome uvéal. Certaines
intégrines pourraient également être fonctionnellement impliquées dans la progression de
ce cancer et dans le processus métastatique (Przybylo, Pochec et al. 2008) bien qu'il soit
encore difficile de déterminer précisément par quels mécanismes elles y parviendraient.
Néanmoins, des travaux récemment publiés par Woodmard et collaborateurs (Woodward,
Rennie et al. 2005) associent l'expression des intégrines al, a4 et a6 à un phénotype plus
agressif du mélanome uvéal. Il devient donc intéressant d'étudier le patron d'expression des
intégrines chez le mélanome uvéal compte tenu de leur fonction probable dans le
développement de cette forme de cancer.

26
1.3.6 L'intégrine a4pl
L'intégrine a4Bl est également connue sous le nom de « very-Iate antigen-4 » (VLA-4)
ou encore CD49d/CD29 et constitue l'une des deux seules intégrines portant la sous-unité
a4, l'autre étant l'intégrine a4~7 (Zhang et al. 1991; Vigneault, Zaniolo et al. 2007).
Contrairement à d'autres intégrines, l'intégrine a4Bl ne présente pas un patron
d'expression ubiquitaire (exprimé par presque tous les tissus). Elle est, entre autre,
exprimée à la surface des fibroblastes du derme (Gailit, Pierschbacher et al. 1993), des
cellules des muscles squelettiques en développement (Rosen, Birkenmeier et al. 1991), des
neurones (Dufour, Duband et al. 1988), de l'épithélium cornéen (Lauweryns, van den Oord
et al. 1991) ainsi qu'à la surface des cellules provenant de lignées hématopoïétiques
(Remler, Elices et al. 1990; Ager et Rumphries 1991; Rosen, Barks et al. 1994). En effet,
l'intégrine VLA-4 semble jouer un rôle dans l'adhési<?,n des lymphocytes, des monocytes,
des éosinophiles, des basophiles, des cellules NK et quelques cellules de mélanome
(Kassner, Teixido et al. 1992) via notamment son contre-récepteur VCAM-l présent à la
surface des cellules endothéliales (Remler, Elices et al. 1990; Allavena, Paganin et al.
1991). Cette intégrine possède aussi la capacité de lier un second ligand: la fibronectine, un
constituant important de la MEC, au niveau de sa région IIICS contenant le motif RGDS
(Arg-Gly-Asp-Ser) et EILDV (Glu-Ile-Leu-Asp-Val). L'interaction entre cette dernière et
l'intégrine a4Bl est essentielle à la localisation des lymphocytes antigène-spécifiques au
site de l'antigène (Ferguson, Mizutani et al. 1991). De plus, cette interaction permet de
faciliter la co-stimulation des lymphocytes T (Davis, Oppenheimer-Marks et al. 1990;
Nojima, Humphries et al. 1990; Shimizu, van Seventer et al. 1990), semble avoir une
importante fonction dans la maturation des cellules ' progénitrices de la moelle osseuse
(Williams, Rios et al. 1991) et finalement, semble être importante dans , la migration des
cellules de la crête neurale durant l'embryogenèse (Dufour, Duband et al. 1988).
L'intégrine a4B 1 est également impliquée dans de nombreuses pathophysiologies comme
l'inflammation, l'asthme, les allergies, l'arthrite, l'athérosclérose et la cicatrisation (Laffon,
Garcia-Vicuna et al. 1991; Striz et Costabel 1992; Walsh et Murphy 1992; Jaspers, Wu et
al. 1995; Lobb, Abraham et al. 1996). Finalement, cette intégrine semble exercer un rôle

27
dans la progression du cancer compte tenu de son implication dans des processus tels que la
croissance des cellules tumorales et la propagation métastatique (Bao, Pigott et al. 1993).
1.3.6.1 Structure de l'intégrine a4pl
L' intéwine u4~ 1 possède certaines caractéristiques structurales spécifiques. Par
exemple, sa sous-unité u4 constituante est la seule sous-unité u connue pour subir un
clivage protéolytique variable près du milieu de la molécule; produisant ainsi des fragments
de 70 et 80 kDa (Hemler, Elices et al. 1990). Il semble que les différentes formes de la
molécule générées par ce phénomène possèderaient également différentes capacités
fonctionnelles (Wayner, Garcia-Pardo et al. 1989). La queue cytoplasmique de cette même
sous-unité semble cependant très conservée d'une espèce à l'autre suggérant un rôle
important exercé par ce segment (Kassner, Teixido et al. 1992). Elle ·est toutefois très
différente des autres sous-unités u, conférant ainsi une ' fonction unique à cette sous-unité
(Kassner, Teixido et al. 1992). La queue cytoplasmique de la sous-unité ~ joue, quant à
elle, un rôle critique dans la signalisation de l'intégrine (O'Toole, Katagiri et al. 1994).
1.3.6.2 Régions régulatrices du .gène a4
La présence de nombreux éléments agissant en cis a été identifiée sur le promoteur et la
région 5' du gène a4 (Figure 1.10). Ceux-ci incluent, entre autre, des sites pour les facteurs
de transcription (FT) AP-1 et AP-2 (<< Activator Protein ») à -377 et +590 respectivement,
un site de liaison pour MyoD à -274 et un site probable pour Mitf dans la région s'étendant
de -89 à -84 (Rosen, Birkenmeier et al. 1991; Planque, Leconte et al. 2001; Vigneault,
Zaniolo et al. 2007). De plus, on remarque également la présence de sites de liaison pour le
facteur de transcription ZEB dans les régions -400 et -130 (Vigneault, Zaniolo et al. 2007).
La région promotrice contient aussi des sites de liaison (boîtes PU) pour des FTs membres
de la grande famille Ets localisés à -67, -54 et -50 et pour le FT Pax-6 dans les régions
situées entre -953 et -936 et aux environs de -96 et de -45 (Rosen, Barks et al. 1994;
Zaniolo, Leclerc et al. 2004).

28
· ~~ B Oites < PU ) a4.1 a4.2
Figure 1.10 - Région régulatrice du gène (14 humain. Représentation schématique de la . région promotrice du gène de l'intégrine a4. On peut y observer les principaux sites de liaison pour des facteurs de transcription (image adaptée de Vigneault, 2007).
Des travaux réalisés dans notre laboratoire ont permis de mettre en évidence deux
régions homologues agissant en cis requises à différents niveaux afin de moduler l'activité
de base du promoteur a4 (Audet, Masson et al. 1994). Ces éléments, désignés a4.1 et a4.2
(Figure 1.11), sont compris entre les régions -51 à -30 et -27 à -6, respectivement (Audet,
Masson et al. 1994). Des étu~es antérieures nous ont permis d'identifier des protéines
nucléaires pro,:,enant de cellules épithéliales de cornée de lapin capables de lier les régions
régulatrices du promoteur a4 (Larouche, Leclerc et al. 1996). Ces protéines, désignées Bp 1
à Bp5 (<< Binding Protein» 1 à 5), possèdent des poids moléculaires allant de 91 kDa pour
BpI à 39 kDa pour Bp5 (74 kDa pour Bp2, 59 kDa pour Bp3 et 45 kDa pour Bp4)
(Larouche, Leclerc et al. 1996). Elles pourraient avoir un rôle à jouer dans la modulation de
l'expression du gène de l'intégrine a4 (Larouche, Leclerc et al. 1996). Plus récemment,
nous avons identifié un de ces facteurs, Bp5, comme étant le FT Pax -6 (Zaniolo, Leclerc et
al. 2004). L'identité des autres protéines du complexe demeure toutefois encore inconnue à
ce jour (Larouche, Leclerc et al. 1996).
a4 .. 1 .1-67 ... GGAAGTGTGGGGAGGAAGGAAGTGGGTATAGAAGGGTG ... - 3 0 . 1 1 Il . 1 1 r r Il Il Il 1 I l 1 t l' ,. Il' Il r
«4.2 1-43 ... GGTATAGAAGGGTGC TGAGATGTGGGTCTTGAAGAGAA ... - 6
Figure 1.11 - Éléments régulateurs. Représentation schématique des éléments régulateurs a4.1 et a4.2 identifiés dans le promoteur du gène a4. Les caractères en gras indiquent les résidus conservés entre les deux réglons (image tirée de Larouche, 1996).

29
1.3.6.3 Le facteur de transcription Pax-6
1.3.6.3.1 La famille des gènes PAX
Les gènes PAX font partie d'une famille de gènes (Figure 1.12) contenant neuf membres
chez l'humain (Pax-l à Pax-9) qui codent pour des facteurs de transcription nucléaires
impliqués dans le développement précoce ainsi que dans le maintien de l'intégrité tissulaire
de certains organes (Strachan et Read 1994; Macdonald et Wilson 1996). Ces gènes sont
classés selon leur similitude de séquence: tous les membres de la famille PAX ont la
particularité d'avoir un domaine «Paired », certains possèdent un motif octapeptide et
certains ont un homéodomaine (Strachan et Read 1994).
Gène Local isation Groupe Structure
PAX 1 20pll -PAX 9 14q12 -PAX 2 2q35 -PAX 5 9p13 Il -PAX 8 2q12 -PAX 3 2q35
III -PAX 7 lp36 -PAX 4 7q32 IV
PAX 6 I1p13
Figure 1.12 - Famille des gènes Pax. Représentation schématique de la structur~ des gènes Pax selon leurs domaines structuraux (image adaptée de Ewan, 2006).
1.3.6.3.2 Pax-6
Un des membres de la famille PAX, Pax-6, est notamment exprimé dans l'oeil pendant
le développement, mais également au niveau de,s voies nasales, du pancréas, de
l'hypophyse ainsi que du système nerveux central (Zaniolo, Leclerc et al. 2004). Pax-6
possède également des rôles chez l'adulte et continue d'être exprimé, par exemple, dans la
rétine, l'épithélium cornéen, le cerveau et le pancréas (Mathers et lamrich 2000; Zaniolo,
Leclerc et al. 2004).
Le gène Pax-6, localisé sur le chromosome I1p13 (Callaerts, Halder et al. 1997),
comporte 12 exons et encode un ARN messager de 2,7 kb qui produira une protéine avec

30
un poids moléculaire de 46 kDa (P AX-6) (Callaerts, Halder et al. 1997). La protéine
contient un domaine «Paired», un homéodomaine ainsi qu'un domaine PST riche en
prolines, sérines et thréonines (Callaerts, Halder et al. 1997) (Figure 1.13). Le domaine
« Paired» est un domaine de liaison à l'ADN constitué de 128 acides aminés (Bopp, Jamet
et al. 1989; Xu, Rould et al. 1999). Il se subdivise lui-même en deux parties distinctes, soit
le domaine « PAl », en position amino-terminale, qui permet la stabilité du contact entre la
protéine et l'ADN et le domaine « RED » en position carboxy-terminale, qui augmente la
spécificité de la protéine en contribuant à la reconnaissance du site de liaison à l'ADN
(Callaerts, Halder et al. 1997; Xu, Rould et al. 1999). L'homéodomaine situé au centre de
la protéine est séparé du domaine «Paired» par une région liante comportant une forte
proportion de résidus glycines et glutamines (Callaerts, Halder et al. 1997). Il est constitué
de 60 acides aminés assemblés en trois hélices a qui sont suivies d'un petit domaine
globulaire. Il s'agit également d'un domaine de liaison à l'ADN, qui, par coopération avec
le domaine «Paired», permet la reconnaissance par la protéine d'une séquence d'ADN
pouvant aller jusqu'à 40 pb, lui conférant ainsi une forte affinité et une grande spécificité de
liaison à l'ADN (Callaerts, Halder et al. 1997; S trachan et Read 1994; Duncan, Cvekl et al.
2000). Finalement, le domaine PST est le plus imposant domaine de la protéine avec 152
acides aminés. Étant un domaine de transactivation, il peut être phosphorylé par des
protéines kinases comme les MAP kinases et exercerait un rôle clé dans plusieurs processus
cellulaires (Mikkola, Bruun et al. 1999).
422 .......... ------w
Sa
Figure 1.13 - PAX6 et PAX6 (Sa). Représentation schématique des différents domaines de PAX6 et PAX6 (5a) (image adaptée de Bharesh, 2004).
Il existe cependant certains isoformes de cette protéine issues d'un épissage alternatif.
La plus connue est PAX-6(5a), une protéine de 48 kDa qui possède un exon supplémentaire

31
( 5 a) de 14 acides aminés dans le domaine « Paired » de la protéine (Epstein, Glaser et al.
1994) (Figure 1.13). Il existe également des variants de 43, 32 ou 33 kDa (Carriere, Plaza et
al. 1993) résultant de l' épissage alternatif du transcrit Pax -6 et bien qu'elles demeurent
encore aujourd'hui beaucoup moins caractérisées que PAX-6 et PAX-6(5a), on soupçonne
néanmoins qu'elles possèdent des fonctions particulières et contrôlent des gènes distincts
(Carriere, Plaza et al. 1993). L'isoforme de 32/33 kDa ne possède pas de domaine
« Paired » tandis que l'isoforme de 43 kDa ne possède pas le cinquième exon, créant ainsi
un nouveau domaine de liaison à l'ADN (Carriere, Plaza et al., 1993).
Les mutations dans le gène Pax -6 évoquent elles-mêmes l'importance des rôles que ce
FT détient. Ainsi, la majorité des mutations identifiées dans ce gène provoquent une perte
de fonction de la protéine (van Heyningen et Williamson 2002). Chez la souris, ces
mutations provoquent un phénotype présentant des yeux plus petits ou l'absence totale des
yeux (Tsonis et Fuentes 2006). Plusieurs anomalies ou désordres oculaires sont liés à un
défaut d'expression de Pax-6 chez l'humain: l'aniridie (hypoplasie de l'iris ou l'absence de
cette dernière) survient par transmission autosomique dominante par haplo-insuffisance du
gène Pax-6 muté; l'hypoplasie fovéale (des cataractes préséniles) due à une mutation
hétérozygote du gène Pax-6; le syndrome de Peter's (défaut de la chambre antérieure de
l' œil et opacité centrale de la cornée) qui se transmet de façon autosomique récessive (van
Heyningen et Williamson 2002; Tsonis et Fuentes 2006).
1.3.7 L'intégrine «SpI
. L'iiltégrine a5~1 représente le seul récepteur qui contient la sous-unité a5. Néanmoins,
cette intégrine est exprimée dans beaucoup de tissus du corps humain notamment la peau,
les reins, le foie, le pancréas, la rate, les intestins, les amygdales, le cœur, les nerfs
périphériques, le cortex cérébral, les vaisseaux sanguins ainsi que dans plusieurs parties du
globe oculaire (rétine, cornée, choroïde, cristallin, conjonctive) (Sincock, Mayrhofer et al.
1997; Brem et al. 1994; Sherry, Proske 2001).

1
1
32
Via son interaction avec son principal ligand, la fibronectine, l'intégrine a5Bl semble
jouer un rôle au niveau de plusieurs processus cellulaires importants. En effet, la liaison
entre la fibronectine et cette intégrine semble être essentielle à la cicatrisation cornéenne,
l' intégrité tissulaire ainsi que l' assemblage de la matrice de fibronectine et la survie '
cellulaire (Stepp, Spurr-Michaud et al. 1993; Pulai, Del Carlo et al. 2002; Danen et
Sonnenberg 2003; Robinson, Zazzali et al. 2003). De plus, cette même intégrine semble
impliquée au niveau de la prolifération, de la migration et de l'adhésion cellulaire
(Akiyama, Yamada et al. 19?9; Kim, Zhang et al. 1992; Loeser 1994; Robbins, Brem et al.
1994; Symington 1995; Urbich, Dernbach et ,al. 2002). L'intégrine a5Bl semble également
jouer un rôle au niveau du cancer. En effet, plusieurs évidences indiquent que la
surexpression ou la répression de cette intégrine serait associée à un phénotype plus
agressif des cellules cancéreuses (Vigneault, Zaniolo et al. 2007; Gong, Wang et al. 1997).
Une étude portant sur des cellules de carcinomes du .côlon semble démontrer que plus
l'expression de l'intégrine a5Bl est élevée, plus les cellules cancéreuses démontrent un
haut potentiel invasif (Gong, Wang et al. 1997). Cependant, d'autres études démontrent que
la diminution de l'expression de cette intégrine est également associée à certains types de
ca~cers comme l'adénocarcinonie ou le carcinome du pancréas (Stallmach, von Lampe et
al. 1992; Sanders, Mainiero et al. 1998).
1.3.7.1 Régions régulatrices du gène aS
Le promoteur du gène a5, très riche en Guanine-Cytosine (G-C), ne possède ni boîte
TATA, ni de boîte CCAAT (Birkenmeier, McQuillan et al. 1991; Vigneault, Zaniolo et al.
2007). De plus, on remarque la présence de .deux sites majeurs d'initiation de la
transcription soit à -45 et + 1, ces derniers étant situés dans une séquence initiatrice (lnr)
(Birkenmeier, McQuillan et al. ,1991). On signale également l'existence de plusieurs sites
de liaison pour les facteurs de transcription suivants sur .la région promotrice (Figure .1.14) :
SpI, entre les positions -117 et -101 (site distal) et entre -60 et -52 (site proximal); AP-l,
entre les positions -51 et -45; AP-2, entre les positions +77 et +84; Ets, entre les positions -
302 et -297; C/EBPa et C/EBPB (<< CCAAT/enhancer binding prote in »), entre les positions
-73 et -66; Pax -6, entre les positions -701 et -672 et finalement, NFI (<< Nuclear Factor» l),
entre les positions -71 et -67 (Corbi, Jensen et al. 2000; Gingras, Larauche et al. 2003;

33
Vigneault, Zaniolo et al. 2007). Les facteurs de transcription AP-1, Sp1/Sp3 et Pax-6
semblent tous contribuer positivement à l'activité du promoteur a5 (Corbi, Jensen ~t al.
2000; Duncan, Kozmik et al. 2000; Larouche, Leclerc et al. 2000; Gingras, Larouche et al.
2003). Les membres de la famille Ets semblent également exercer un effet positif sur
l'activité du promoteur a5, bien que l'endroit exact où se lient ces protéines sur le
promoteur n'ait pas encore été identifié (Kita, Takino et al. 2001). Les facteurs de
transcription C/EBP semblent exercer une influence différentielle selon le type cellulaire
étudié. En effet, ils activent le promoteur a5 dans des cellules d'hépatome alors qu'ils le
répriment dans des kératinocytes humains (Corbi, Jensen et al. 2000). Finalement, NFI
semble exercer une influence négative sur l'activité du promoteur (Gingras, Larouche et al.
2003; Gingras et al. 2008).
-702 __ -672
~ lm
Figure 1.14 - Région régulatrice du gène aS humain. Représentation schématique de la région promotrice du gène de l' intégrine a5. On peut y observer les principaux sites de liaison pour des facteurs de transcription (image adaptée de Vigneault, 2007).

34
1.4 Objectifs des travaux de recherche
Le mélanome uvéal n'est pas le cancer ayant la plus forte incidence dans la population.
Toutefois, il demeure le cancer intraoculaire le plus fréquent chez l'individu adulte et son
agressivité est telle qu'un patient sur deux décèdera des suites de cette maladie (Diener
West, Hawkins et al. 1992). Certaines tumeurs provenant de patients différents semblent
démontrer des niveaux d'agressivité variables et ne semblent pas toutes posséder la capacité
de produire des métastases secondaires~ Nous savons déjà que les intégrines pourraient
avoir un rôle prépondérant à jouer, notamment dans les processus métastatiques, mais
également dans l'agressivité de la tumeur (potentiel tumorigène). Il devient donc important
d'étudier ces récepteurs cellulaires dans le contexte du mélanome uvéal afin d'identifier les
différences entre les tumeurs uvéales de patients qui décèdent rapidement et celles de
patients toujours en vie.
De récentes études réalisées dans notre laboratoire ont permis de démontrer que le
patron d'expression de certaines intégrines différait d'une lignée tumorale (provenant d'une
tumeur uvéale primaire ou de métastases hépatiques secondaires) à une autre (Figures 1.15
et 1.16). En effet, dans une lignée cellulaire dérivée des métastases hépatiques d'un patient
ayant développé un mélanome uvéal primaire (H79), nous avons remarqué une hausse de
l'expression de l'intégrine a4~1 en comparaison à des lignées tumorales dérivées de
mélanomes uvéaux primaires n'ayant pas encore produit de métastases au foie (SP6.5,
TP31, SP8.0, T97, T98, T108, Tl15). De plus, nous avons également remarqué un patron
d'expression particulier pour l'intégrine a5~1 entre des lignées non métastatiques (Figures
1.15 et 1.16). Celui-ci pourrait donc être lié à l'agressivité tumorale variable de ces
différentes lignées.

35
Lignee Sous-~nités des intégrines Cellulaire
a l a2 a3 a.4 a5 a6 av ~ 1 ~3 ~4 ~5
MeN ++ + - T +
SP6.5 ++ - ++
SP8 .0 ++ - + .. +++ - -
TP31 - - - -H79 ++++ +++ +++ ~ +++ +++ + ' ++ -
T97 -
T98 - +0+
TiOS - +++
Tl 15 -
Figure 1. 15 - Analyse en cytométrie de flux. Niveau d 'expression de certaines sous-unités d ' intégrines à la surface de cellules dérivées de mélanome uvéal. MeN: mélanocytes normaux; H79: lignée dérivée de métastases hépatiques provenant d 'un patient ayant développé un mélanome uvéal; SP6.5, SP8. 0, TP31, T97, T98, Tl 08, T 115: lignées cellulaires dérivées de mélanomes uvéaux primaires. (-): aucune expression; (+): faible expression; (++): expression modérée; (+++): forte expression; (++++): très forte expression (d' après S. Landreville, 1. Gagné et M. Bergeron).
500
200
500
200
~·'''~ - 11 8S
- a4
-188 - (X4
....... 0> 1-
ex) lO CO 0 ~
0> ...... ...... ..... 1-' t-
- 18S
-0.5
...... CO 0> œ 1-1-
co 0 ~
l-
Ln ,...-~
I-
- 18S
- a6
1:88 aS
Figure 1.16 - Expression des gènes encodant les sous-unités d'intégrines a4, a5, et a6. Analyse par RT-PCR semi-quantitatif de l 'expression des gènes des sous-unités a4, a5 et a6 dans des lignées de mélanome uvéal. UVM: mélanocytes normaux; H79: lignée dérivée de métastases hépatiques provenant d'un patient ayant développé un mélanome uvéal; SP6.5, T97, T98, Tl 08, Tl15: lignées cellulaires dérivées de mélanomes u'véaux primaires; Jurkat: lignée immortalisée de lymphocytes . T (d'après S. Landreville et J. Gagné).

36
Ces différentes observations ont permis d'élaborer plusieurs objectifs quant à l'étude de la
régulation de l'expression des gènes des intégrines a4~ I et a5 ~ I en relation avec les
propriétés métastatiques et tumorigènes du mélanome uvéa!.
1.4.1 Influence de Pax-6 sur l'activité du promoteur de la sous-unité a4 .
Dans un premier temps, et puisque nous avons déjà démontré que la transcription du
gène a4 (Zaniolo, Leclerc et al. 2004) est sous l'influence régulatrice du FT Pax-6 (Zaniolo,
Leclerc et al. 2004), nous étudierons le rôle de ce facteur par l'entremise de trois sections
présentées au chapitre 2 de cet ouvrage. Premièrement, nous allons tenter d'évaluer les
niveaux d'expression du facteur Pax -6 dans un éventail de lignées de mélanome uvéal. En
second lieu, il sera question d'évaluer la capacité de liaison du même FT à la séquence du
promoteur a4. Finalement, nous tenterons de déterminer quelle influence possède Pax-6 sur
l'activité transcriptionnelle du promoteur du gène a4.
1.4.2 Influence de la sous-unité aS dans les propriétés tumorigènes du
mélanome uvéal
Le chapitre ' 3 de ce mémoire sera consacré à l' intégrine a5 ~ I en relation avec
l'agressivité tumorale et se divise lui-même en trois parties. Tout d'àbord, nous allons
tenter de déterminer qualitativement le potentiel de ces lignées à générer des tumeurs.
Ensuite, nous allons évaluer les niveaux d'activité du promoteur du gène de l'intégrine a5
dans des lignées ciblées du mélanome uvéal. Finalement, nous évaluerons les niveaux
d'expression des principaux FTs qui modulent l'activité du promoteur a5 (SpI, AP-I et
NFI) afin de déterminer les facteurs qui pourraient être impliqués dans l'expression
différenti~lle de cette intégrine dans les lignées de mélartome uvéal.

37
2. Influence du facteur de transcription Pax-6 sur l'activité du promoteur du gène a4 dans le mélanome uvéal
2.1 Introduction
Les métastases issues d'un mélanome uvéal primaire réduisent l'espérance de vie des
patients à seulement quelques mois (Leyvraz, Spataro et al. 1997). Les intégrines pourraient
contribuer à ce processus en modifiant les propriétés adhésives et migratrices des cellules
cancéreuses. De récentes études réalisées par notre groupe ont démontré que le niveau
d'expression de certaines intégrines différait d'une lignée tumorale à une autre. En effet,
chez une lignée cellulaire dérivée des métastases hépatiques d'un patient ayant développé
un mélanome uvéal primaire (H79), nous avons remarqué une hausse de l'expression de
l'intégrine a4p 1 en comparaison à des lignées tumorales dérivées de mélanomes uvéaux
primaires n'ayant pas entraîné la formation de métastases au foie (SP6.5, TF31, SP8.0, T97
et T98). Plus récemment, on a démontré ' que la transcription du promoteur a4 était sous
l'influence régulatrice du FT Pax-6 (Zaniolo, Leclerc et al. 2004). Pax-6 est notamment
exprimé pendant le développement des structures oculaires et il est responsable de
l'intégrité de l'œil chez les vertébrés (Mathers et larnrich 2000; Zaniolo, Leclerc et al.
2004). Ceci nous a donc amené à 1) comparer les niveaux d'expression de Pax-6 chez
différents types cellulaires provenant de tumeurs métastatiques et non-métastatiques du
mélanome uvéal, 2) vérifier la capacité de liaison de ce FT à la ·séquence du promoteur a4
et finalement, 3) évaluer l'activité transcriptionnelle du promoteur du gène a4 dans ces
mêmes lignées afin de mettre en évidence des différences susceptibles d'expliquer la
progression métastatique de certaines tumeurs.

38
2.2 Matériel et méthodes
2.2.1 Culture cellulaire
Les lignées cellulaires SP6.5, SP8.0, TP31, T97 et T98 sont dérivées de tumeurs
primaires différentes isolées de patients ayant développé un mélanome uvéal (Beliveau,
Bérubé et al. 2000) alors que la lignée H79 est dérivée de métastases secondaires obtenues
du foie d'un patient atteint d 'un mélanome uvéal (Bérubé, Talbot et al. 2005). Nous avons
également mis en culture des mélanocytes (MCN) provenant de la choroïde d'yeux 1
humains sains. Les cellules ont été cultivées dans le milieu de culture DMEM (Dulbecco ' s
modified Eagle's medium) additionné de 10% de sérum de veau fœtal, de 1 mM de
glutamine et de 15 Jlg/JlI de gentamicine à 37°C sous une atmosphère de 5% de CO2. Le
milieu a été changé environ deux fois par semaine afin de permettre aux cellules de
proliférer jusqu'à la densité souhaitée (recouvrement d'environ 80% de la surface du
flacon) à des fins d'extraction des protéines nucléaires.
2.2.2 Extraits nucléaires et immunobuvardage de type western
Les extraits nucléaires ont été préparés à partir des lignées de mélanomes (H79, SP6.5,
SP8.0, TP31, T97 et T98) cultivées à 80% de confluence cellulaire selon la ,procédure
développée antérieurement dans le laboratoire du Dr Guérin. (Roy, Gosselin et al. 1991).
Les extraits nucléaires des différentes lignées cancéreuses provenant de mélanomes
uvéaux (25 Jlg) ont été fractionnés sur gel SDS-PAGE (75 Volts) et ~ransférés sur une
membrane de nitrocellulose (50 Volts). Ces membranes ont été lavées avec du tampon TS
(150 mM NaCI, 10 mM Tris-HCI pH 7,4) et TSM (tampon TS auquel on ajoute 5% de lait
sans gras et 0,1% de Tween 20) pour ensuite être incubées avec un anticorps primaire dirigé
contre le FT Pax -6 (Santa Cruz Biotechnology ; dilution 1 : 1000) ou un anticorps dirigé
contre l'actine (Jackson Immuno Research laboratories inc. ; diluti9n 1 : 35 000) pendant
Ih30. Les membranes ont ensuite été lavées avec du TSM et -incubées avec un anticorps
secondaire dirigé contre les IgG de lapin couplé à la péroxydase pendant 1 heure (Jackson).

39
Les complexes immunoréactifs ont été révélés par chimiluminescence (<<western blot
detection kit»; Amersham Biosciences).
2.2.3 Retard sur gel (EMSA)
Un oligonucléotide synthétique double-brin (50 ng) correspondant à la séquence de la
région régulatrice a4.1 située entre les positions -62 et -28 par référence au site d ' initiation
de la transcription du gène a4 (5'-GATCTGTGGGAGGAAGGAAGTGGGTATAG
AAGGGTGCT -3 ' ; Audet, Masson et al. 1994) a été radiomarqué au 32p et utilisé en guise
de sonde dans les essais de retard sur gel de la façon suivante: 7,0 ,.d de tampon kinase
10X, 3,0 }.lI d'ATP radiomarqué (y32p ATP; PerkinElmer precise/y, Boston) et 1,0 }.lI
d'enzyme T4 PNK (polynucléotide kinase; 10,000 U/ml) ont été ajoutés à l 'ADN. Le
volume total a été complété à 50 ,.1I avec de l'eau. Le mélange réactionnel a été incubé à
37°C pendant 90 minutes. Un volume de 90 }.lI de Tris 10 mM a été ajouté et la sonde a
ensuite été purifiée sur Elutip-D (BioAmerica Inc.), tel que recommandé par le fournisseur.
L'ADN marqué a été précipité par l'ajout de 1/10 du volume de NaOAC 3 M et 2,5
volumes d'éthanol 95%, à -20°C pendant 2 heures. La radioactivité totale de l'échantillon a
été mesurée à l'aide d'un compteur Bêta (Wallac, « WinSpectral : 1414 Liquid Scintillation
Counter ») et le culot re-suspendu dans du Tris 1 0 mM à raison de 40 000 cpml}.lI. Les
sondes ont été conservées à -20°C pour une période maximale de deux semaines.
Les essais de retard sur gel ont été réalisés selon une technique déjà éprouvée dans le
laboratoire du Dr Guérin (Schneider, Gander et al. 1986). Des gels de polyacrylamide natifs
6% et 8% ont été utilisés (acrylamide 40%; 39: 1) afin de séparer les complexes ADN
protéine. Le gel a préalablement été soumis à une tension de 120 volts à 4°C pendant 1
heure avant d'y déposer les échantillons. Chaque échantillon contenait 16 000 cpm de
sonde Pax 6, 200 à 500 ng de poly dIdC, ,10 }.lg d'extrait nucléaire, 12 }.lI de Tampon D2X
(20 mM Hepes pH 7,9 + 20% glycérol + 0,2 mM EDTA +O,5mM PMSF), 1,2 mM de KCI
et le volume est complété avec de l'ea jusqu'à 24 }.lI. La migration du gel a été réalisée à
120 volts à 4°C pendant environ 4-5 heures pour un gel à 6% ou 6-8 heures pour un gel à
8% contre une solution de Tris-Glycine IX (0,25 M Tris + 2 M Glycine). Le gel a ensuite

40
été autoradiographié pendant 16 heures à -80°C. Des super-rétentions ont également été
réalisées par l'ajout d'un anticorps spécifique (2 JlI et 4 JlI) dirigé contre Pax-6.
2.2.4 Transfections transitoires
Les plasmides a4-7 6CA T et a4-1 OOOCA T couplés au gène rapporteur CA T
(chloramphénicol acétyl transférase) (Birkenmeier, McQuillan et al. 1991) dans le vecteur
pskCAT ont été utilisés pour les transfections. Les différentes lignées cancéreuses (H79,
SP6.5, SP8.0 et TP31) ont été transfectées de façon transitoire avec les différents vecteurs
(1 Jlg par puits) dans des plaques à 6 puits contenant 200 000 cellules, à l'aide d'un lipide
polycationique (Lipofectamine; Invitrogen). Le plasmide pXGH5 (0,5 Jlg), qui code pour
un variant sécrété de l'hormone de croissance humaine (hGH), a été ajouté à chaque
échantillon. L'activité CAT a été évaluée par dosage enzymatique (Pothier, Ouellet et al.
1992) et la transféction a été normalisée par rapport à la quantité de GH sécrétée dans le
milieu de culture (ImmuChem Double antibody hGH 1251 RIA Kit; ICN Phatmaceuticals
Inc.) ainsi qu'à la quantité de protéine utilisée (dosée par la technique de Bradford;
Fermentas, Life Science). Les activités CAT devaient être au moins trois fois supérieures
au bruit de fond causé par le mélange réactionnel seul pour être considérées significatives.
Les valeurs représentent la moyenne d'au moins trois transfections effectuées en triplicata.
Des analyses statistiques (Test Student) ont été réalisées et les différences ont été
considérées statistiquement significatives lorsque p < 0,05.

41
2.3 Résultats
2.3.1 Expression du facteur de transcription Pax-6 dans le mélanome
uvéal
Les essaIS d'immunobuvardage de type western ont permIS d'évaluer les nIveaux
d'expression de Pax-6 dans les lignées métastatiques et non-métastatiques provenant de
patients atteints d'un mélanome uvéa!. La présence de l'isoforme Pax-6 (5a) de 48 kDa est
évidente dans toutes les lignées tumorales (Figure 2.1). On remarque également que la
protéine Pax-6 de 46 kDa est exprimée seulement dans les mélanocytes normaux (MeN) et
semble absente chez les lignées cancéreuses (Figure 2.1).
H79 8P6_5
48 kDa ::p,..rr .... ' 46 kDa
A.ctine
TP3 l T97 T98
Psx~ Ca) - Psx~
Figure 2.1 - Niveau d'expression du facteur de transcription Pax-6 dans les lignées de mélanomes. Analyse d' immunobuvardage de type western réalisée avec 25 Jlg de protéines nucléaires de chacune des lignées. Les extraits ont été fractionnés sur gel, transférés sur membrane de nitrocellulose et incûbés avec des anticorps dirigés contre le facteur de transcription Pax-6" et contre l'actine. La position des marqueurs de poids moléculaire (kDa) est indiquée sur la figure.
2.3.2 Le facteur de transcription Pax-6 peut lier la séquence de la région
promotrice «4.1
Afin d'évaluer l'activité de lîaison de Pax-6 à la séquence de la région promotrice a4.1,
nous avons réalisé des essais de retard sur gel (EMSA). Dans un premier temps, ces essais
ont été réalisés avec les lignées cancéreuses H79, SP6.5, SP8.0 ainsi que TP31 dans le but
de comparer la lignée métastatique et les lignées non-métastatiques. La sonde utilisée
correspond à la région promotrice a4.1 marquée au 32p. Il est possible d'observer un patron

42
de liaison identique pour les quatre lignées indépendamment de leur pouvoir métastatique
(Figure 2.2 A).
B. A. 1·
~
u
Figure 2.2 - Capacité de liaison de Pax-6 à la séquence régulatrice a4.1. A) Essais de retard sur gel réalisés avec 10 Ilg de protéines nucléaires de chacune des lignées. Ces extraits ont été incubés avec une sonde radiomarquée correspondant à la séquence régulatrice a4.1. B) Afin de vérifier la spécificité des complexes obtenus, des anticorps dirigés spécifiquement contre Pax-6, Ets-l (2 III et 4 Ill) et Sp 1 (4 Ill) ont été ajoutés. Les complexes ADN-protéines ont été séparés sur gel de polyacrylamide (8%) et révélés par autoradiographie. U : sonde libre.
Dans un deuxième temps, des essais de super-rétention en gel ont été réalisés afin de
s'assurer de la spécificité du signal avec la lignée métastatique H79 et une lignée non
métastatique, SP6.5, en utilisant la même sonde radiomarquée ainsi qu'un anticorps dirigé
contre Pax-6. L'ajout de l'anticorps Pax-6 entraîne la disparition du signal spécifique
produit par ce facteur (Figure 2.2 B). Ce phénomène s'observe autant dans la lignée
métastatique (H79) que dans la lignée non-métastatique (SP6.5). La disparition du signal,
plutôt que sa super-rétention, est due au fait que l'anticorps dirigé contre Pax-6 se lie au
domaine de liaison à l'ADN de la protéine l'empêchant ainsi de lier la sonde. Cependant,
lorsqu'un anticorps dirigé contre les FTs Ets-l ou Sp 1 est ajouté au mélange réactionnel, on

43
ne remarque aucune disparition du signal ni aucune formation d'un super-complexe quel
qu'il soit avec les extraits H79 ou SP6.5 (Figure 2.2 B). Ces résultats démontrent ainsi la
spécificité du signal correspondant à Pax-6. La présence de la bande correspondant au
signal spécifique de Pax-6 dans les quatre lignées cancéreuses (Figure 2.2 A) suggère que
Pax-6 possède la capacité de lier la séquence de la région promotrice a4.1 autant dans la
lignée métastatique H79 que dans les lignées non-métastatiques SP6.5, SP8.0 et TP31.
2.3.3 Pax-6 est un répresseur de l'activité du promoteur a4
Des essais en transfection transitoire ont été réalisés afin de définir l'influence
régulatrice de Pax-6 sur le promoteur du gène a4. Les résultats obtenus permettent
d'affirmer que l'activité dirigée par le plasmide portant le plus long segment du promoteur
a4 (-1 000a4CAT) est plus iinportante dans la lignée métastatique H79 que dans les lignées
non-métastatiques SP6.5, SP8.0 et TP31 (Figure 2.3). Bien qu'une activité moindre que
celle observée dans les H79 ait été mesurée dans la lignée SP6.5, cette dernière demeure
tout de même facilement mesurable comparativement à SP8.0 et TP31, toutes deux étant
incapables d'assurer une transcription dirigée par le plasmide recombinant -1000 a4CAT.
H79
TP3l
....... o o o cr
1
~ 16 =211
A'ctivité CAT (%CAI/hGH) t-J o o o
~
o o o
9
~ o o o 9
----t 3 659:: 14
18 841 = 334
!
Figure 2.3 - Activité transcriptionnelle du promoteur du gène a4. Le plasmide -IOOOa4CAT a été transfecté dans les lignées de mélanome uvéal H79, SP6.5, SP8.0 et TP31. Le graphique représente la moyenne des activités CA T (normalisées par rapport à la GH) obtenues suite à trois transfections réalisées en triplicata.

44
De plus, des essais ont été réalisés en transfectant de petits ARN interférents (iARN) de
21 à 25 pb dirigés contre le transcrit Pax-6 conjo~ntement avec les vecteurs contenant soit le
plus court (-7 6a4CA T) ou le plus long (-1 000a4CA T) segment du promoteur a4 dans la
lignée métastatique H79 et dans la lignée non-métastatique SP6.5. Par l'ajout d'iARN, il est
possible de diminuer considérablement l'expression de Pax-6 en stimulant l'élimination de
son transcrit dans le but de définir son influence sur l'activité du promoteur a4. On
remarque que les iARN Pax-6 dans la lignée H79 (Figure 2.4 A) provoquent une
augmentation de l'activité du promoteur a4 d'environ 2,4 fois lorsque transfectés avec lé
plasmide -76a4CAT et d'environ 3,6 fois avec le plasmide -1000a4CAT en comparaison
avec l'activité initiale mesurée en absence d'iARN (correspondant à 100%). Une tendance
similaire, mais d'amplitude accrue, est également observée dans la lignée non-métastatique
SP6.5 (Figure 2.4 B). En effet, en présence d'iARN, on remarque une augmentation plus
prononcée de l'activité du promoteur a4 dans la lignée SP6.5 pour le plasmide -76a4CAT
(environ 4 fois) et pour le plasmide -1000a4 CAT (près de 9 fois) par rapport à l ' activité
obtenue sans iARN (100%).
Afin de confirmer le rôle du FT Pax-6 dans les lignées de mélanome, nous avons
également transfecté transitoirement un vecteur sur-exprimant la protéine Pax -6 dans les
lignées H79 et SP6.5 simultanément avec les mêmes vecteurs utilisés pour l'interférence à
l' ARN. Ainsi, la transfection du vecteur Pax -6 dans la lignée .H79, entraîne une diminution
de 3 fois de l'activité du promoteur contenu sur le plasmide -76a4CAT et de 2 fois avec
celui porté par ~ 1000a4CAT (Figure 2.4 A) en référence à l'activité initiale du promoteur
ayant été transfecté avec le vecteur vide (pUCI9). Ce même phénomène s'observe
également dans la lignée non-métastatique SP6.5 dans laquelle une diminution de près de 2
fois l'activité initiale du promoteur est observée, tant avec le plasmide -7 6a4CAT, qu'avec
le plasmide -1000a4CAT (Figure 2.4 B).
Selon les résultats obtenus, dans chacun· des cas, Pax-6 semble jouer un rôle de
répresseur de l'activité du promoteur a4, tant dans la lignée métastatique H79 que dans la
lignée non-métastatique SP6.5.

45
A. ACÜ'l'i é CAT rela ti\ i'e
0.4-6
a. - 6+ iARN
0.4-1 000
a -1000 + iARN
04-1 000 + V~ctellr
B.
a4- 6
,a4-16 + iARN
,0.4-1000
'M-l 000 + iARN 1---+---...-...----4 989 = 11].4-1000 + v ecteur
Figure 2.4 - Influence de Pax-6 sur l'activité du promoteur du gène a4. Les plasmides -76a4CAT (a4-76) et -lOOOa4CAT (a4-1000) ontété transfectés eJans la lignée métastatique R79 (A) ou dans la lignée non-métastatique SP6.5 (B) cultivées à confluence. Les transfections ont également été réalisées en présence d'ARN interférent ciblant le facteur de transcription Pax-6 (iARN) ou en présence d'un vecteur sur-exprimant Pax-6 (Vecteur). Le graphique représente la moyenne relative des activités CAT (normalisées par rapport à la GR) obtenues suite à trois transfections réalisées en triplicat.

46
2.4 Discussion
Il est maintenant admis que la fonnation de métastases résulte de la migration des
cellules cancéreuses d'un mélanome uvéal primaire vers d'autres organes cibles du corps,
principalement le foie (dans près de 90% des cas; Diener-West, Hawkins et al. 1992). Ce
phénomène pourrait être dicté par l' intégrine a4 puisqu'elle est responsable, entre autre, de
la migration cellulaire (Aplin, Howe et al. 1998) et qu'elle est surexprimée dans la lignée
H79. Nous savons également, par de récentes études, que le gène encodant cette intégrine
voit sa transcription modulée par le FT Pax-6 (Zaniolo, Leclerc et al. 2004) et l'expression
aberrante de ce dernier pourrait donc être la cause du développeinent d'une tumeur à un
foyer secondaire.
Nos résultats d'immunobuvardage de type western démontrent cependant que Pax-6
(46kDa) est exprimé seulement dans les mélanocytes nonnaux (Figure 2.1) alors que
l'isoforme Pax-6 (5a) (48 kDa) est exprimé dans toutes les lignées cancéreuses
. indépendamment de leur pouvoir métastatique (Figure 2.1). De plus, on remarque que Pax-
6 (5a) semble autant exprimé dans toutes les lignées cancéreuses et aucune différence
significative n'a été observée entre les lignées. Selon la littérature sur ce sujet, les isoformes
de Pax-6 semblent, à ce jour, exercer la même fonction que Pax-6 de 46kDa (Epstein,
Glaser et al. 1994; Kozmik, Czerny et al. 1997; Chauhan, Reed et al. 2002), ce qui pennet
de croire que Pax-6 (5a) pourrait agir de manière similaire à Pax-6 sur le promoteur du gène
de l'intégrine a4.
Même si le niveau de Pax-6 (5a) semble identique dans toutes les lignées cancéreuses, la
présence de la protéine ne nous renseigne aucunement sur la capacité de ce facteur à se lier
au promoteur a4 et d'agir ainsi en tant que FT. Cependant, les résultats obtenus suite aux
analyses de retard sur gel permettent de confinner que l'isofonne Pax-6 (5a) est capable de
lier la séquence régulatrice a4.1 dans toutes les lignées cancéreuses, métastatiques ou non
(Figure 2.2 A). De plus, nos résultats confinnent non seulement l'identité de Pax-6, mais
également la spécificité de sa liaison à la sonde et ce, grâce au signal obtenu in vitro pour
Pax-6 (Pax-6 (5a) correspondant au même signal) dans les lignées H79, SP6.5, SP8.0 et

47
TP31 (Figure 2.2 B-C). Nous savons maintenant que non seulement Pax-6 (5a) est présent
dans toutes les lignées cancéreuses, mais qu'il est également abilité à lier la séquence a4.1
autant dans la lignée métastatique que dans les lignées non-métastatiques.
Les résultats des transfections transitoires démontrent qu'effectivement, l'activité du
promoteur a4 est plus élevée dans la lignée métastatique H79 que dans les lignées SP6.5,
SP8.0 et TP31 (Figure 2.3). L'inhibition de l'expression de Pax-6 par iARN ou sa sur
expression avec le vecteur sur-exprimant Pax-6 (Figure 2.4 A-B), ont permis de confirmer
le rôle de Pax-6 sur l'activité transcriptionnelle du promoteur du gène a4. Considérant que
les isoformes de Pax-6 possèdent sensiblement les mêmes fonctions et que, par analyse
d'immunobuvardage de type western, seul Pax-6 (5a) semble exprimé dans nos lignées, ce
dernier pourrait donc vraisemblable~ent exercer une influence négative sur l '·activité du
promoteur a4. En effet, en inhibant l'expression de Pax-6 par l'ajout de petits ARNs
. interférents, nous obtenons une augmentation de l'activité du promoteur (Figure 2.4 A)
comparativement à une diminution lorsque ce FT est sur-exprimé dans ces cellules (Figure
2.4 B). Ainsi, Pax-6 ne semble pas être l'élément déterminant qui permettrait d'expliquer
l'expression aberrante de la sous-unité a4 puisqu'il est présent dans toutes les lignées,
qu'elles soient métastatiques ou non. De plus, cet isoforme (Pax-6 (5a)) semble être un
puissant répresseur de l'activité dirigée par le promoteur a4 dans les lignées cancéreuses
dérivées d'un mélanome uvéal, ce qui corrobore une étude qui a rapporté Pax-6 comme
étant un répresseur de la transcription (Duncan, Haynes et al. 1998).
Ces résultats suggèrent donc qu'un autre facteur pourrait également se lier à la séquence
du promoteur a4 et moduler l'expression aberrante de ce gène contribuant ainsi à la
migration des cellules cancéreuses des vaisseaux sanguins vers le foie. Nous croyons qu'il
pourrait se produire une compétition entre les protéines se liant aux régions régulatrices
a4.1 et a4.2, empêchant ainsi Pax-6 de se lier à sa séquence cible dans le promoteur a4
dans la lignée métastatique H79, empêchant de surcroît Pax-6 de réprimer l'activité de ce
promoteur. De plus, puisque le FT. Pax-6 fait partie du complexe protéique Bp5 (Zaniolo,
Leclerc et al. 2004), il se pourrait donc qu'un autre facteur présent dans l'un ou l'autre des
complexes Bp puisse compétitionner directement avec la liaison de Pax-6. En effet, certains

48
travaux visant à caractériser les protéines Bp 1 à Bp5 appuient nos résultats (Larouche,
Leclerc et al. 1996). Ainsi, lorsque le complexe protéique Bp2 est mis en présence du
complexe Bp5, on observe la compétition pour le site de" liaison commun qui entraîne une
baisse du signal de Bp5. De plus, certains travaux ont permis de démontrer la présence du
complexe Bp2 dans la lignée non-métastatique SP6.5 (Larouche, Larouche et al. 1998).
Ceci suggère que malgré la présence de Pax-6 (5a) dans cette lignée, une compétition entre
" les complexes Bp2 et Bp5 pour la liais?n sur le promoteur du gène a4 pourrait survenir.
Ainsi, si le même phénomène survient également dans les autres lignées, Pax -6 ne pourrait
interagir avec l'élément a4.1 afin de réprimer le promoteur a4 dans la lignée H79.
Alternativement, un activateur fort pourrait présenter plus d'affinité pour la séquence du
promoteur a4 dans cette lignée et ainsi activer plus fortement ce promoteur éclipsant, par le
fait même, l'influence négative de Pax-6.
Il importe également de souligner" la présence de sites de liaison pour d'autres facteurs
de transcription dans la région promotrice du gène a4. En effet, la présence de sites pour
des FTs appartenant à la famille Ets près de la région a4.1 a été rapportée par Rosen et ses
collaborateurs (Figure 1.9) (Rosen, Birkenmeier et al. 1991; Rosen, Barks et al. 1994).
Cette famille de facteurs de transcription contient plus de 30 membres dont la plupart sont
de puissants activateurs. Sachant qu'il existe des sites de liaison près des séquences
régulatrices a4.1 et a4.2, la protéine constituant le complexe Bp2 pourrait fort bien
appartenir à la famille Ets.
Il est également possible que Pax-6 puisse subir des modifications post-traductionnelles,
telles que la phosphorylation ou la glycosylation, l'empêchant ainsi d'agir en tant que
répresseur dans la lignée métastatique H79. En effet, plusieurs études démontrent que ces
modifications peuvent changer et moduler les fonctions de Pax -6 (Lefebvre, Planque et al.
2002; Yan, Liu et al. 2007) ce qui pourrait expliquer la diffé~ence observée entre les lignées
métastatiques ou non.
En résumé, les résultats obtenus nous indiquent qu'un évènement additionnel survient
afin d'activer le promoteur a4 dans la lignée métastatique H79 et que le FT Pax-6 ne peut

49
être le responsable de cette expreSSIon aberrante puisqu'il semble réprimer l 'activité
transcriptionnelle du promoteur a4 dans les lignée~ cancéreuses dérivées de mélanomes
~véaux primaires ou de métastases hépatiques secondaires. Suivant les hypothèses
mentionnées précédemment afin d'expliquer ce phénomène, il serait intéressant
premièrement de déterminer si Pax-6 subit"des modifications post-traductionnelles dans la
lignée métastatique H79, car celles-ci pourraient notamment changer la nature de son
influence sur l'activité transcriptionnelle du promoteur a4 ou, tout simplement, l ' empêcher
de lier la séquence du promoteur et d'agir en tant que répresseur. Toutefois, la sur
expression de Pax-6 dans les lignées H79 et SP6.5 entraînant un effet (répression)
d'amplitude similaire, il est donc improbable que des différences dans d'éventuelles
modifications post-traductionnelles soient en cause dans l'expression différentielle du gène
a4. De plus, il serait important d'approfondir l'étude des facteurs de transcription membres
de la famille Ets afin de définir si un de ceux-ci pourrait activer le promoteur du gène a4,
permettant ainsi aux cellules cancéreuses d'acquérir un pouvoir métastatique leur
permettant d'établir de nouveaux foyers cancéreux au foie.

50
3. Influence de la sous-unité aS dans les propriétés tumorigènes du mélanome uvéal
3.1 Introduction
La plupart des individus atteints d'un mélanome. uvéal ne développeront aucune
complication métastatique liée à cette forme de cancer. Toutefois, dans un certain nombre
de cas, la maladie évoluera plus rapidement et causera le décès du patient dans les dix
années suivant le diagnostic (Kincaid 1998) en raison de l'apparition de métastases au foie
des individus atteints. Des variations dans l'agressivité des cellules de ces tumeurs
pourraient expliquer la différence entre survie et mort chez les patients atteints d'un
mélanome uvéal. Des études réalisées dans notre laboratoire ont démontré que d'après le
suivi de plusieurs patients atteints d'un mélanome uvéal, ceux démontrant une plus forte
expression de la sous-unité d'intégrine a5 semblaient survivre plus longtemps que les
patients démontrant une plus faible expression (Tableau 3.1).
T bl a eau 3 1 R' 'd esume es cas d' ' ou provlennen tl r es 12nees cancereuses Lignée
Origine Âge, Date de Date de Suivi
Histologie cellulaire Sexe l'énuclation l'invasion au foie (mois)
SP6.5 TP 77,F 01/26/88 - 111 Mixte
SP8.0 TP 48,M 04/05/88 01101196 138* Mixte
TP31 TP 62,M 10/08/91 10/13/94 38* Mixte T97 TP 69,M 05/06/02 - 75 Mixte
T98 TP 32,M OS/27/02 - 69 Mixte T108 TP 44,M 04/20/04 - 44 Mixte
Tl15 TP 30,F 02/03/06 - 65 Mixte . Les lignées cellulaires sont dérivées de tumeurs uvéales primaires (TP). L'âge correspond à l'âge des patients au moment du diagnostic de la tumeur primaire. Le suivi débute suite au diagnostic du mélanome uvéa!. On a perdu la trace du patient duquel la lignée SP6.5 a été dérivée. Aucune métastase au foie n'avait été décelée lors de sa dernière visite. L'astérisque indique la fin du suivi après la mort du patient suite à des métastases secondaires.
Nous avons donc supposé que la sous-unité a5 pouvait jouer un rôle dans l'agressivité
tumorale chez les patients atteints d'un mélanome uvéal. Des lignées cellulaires dérivées de

51
différentes tumeurs uvéales primaires (T97, T98, T108 et T115) ont été utilisées en guise
de modèle pour l'étude de l'agressivité tumorale. Selon des analyses réalisées en cytométrie
de flux dans notre .laboratoire, les lignées T97 et T98 expriment des niveaux très faibles de
la sous-unité a5 à leur surface cellulaire tandis qu'elle est fortement exprimée dans les
lignées Tl 08 et T 115 (Figure 1.15). La différence entre les lignées se dénote également à
l'analyse du transcrit a5 par RT -PCR (Figure 1.16). Compte tenu de ces résultats, nous
avons évalué qualitativement le potentiel tumorigène . de ces lignées puisque celles-ci
n'avaient encore jamais été caractérisées. Par la suite, nous avons évalué les niveaux
d'activité transcriptionnelle du promoteur du gène a5 dans ces mêmes lignées. Finalement,.
sachant, par des études antérieures réalisées dans notre laboratoire, que le promoteur du
. gène a5 est principalement sous l'influence régulatrice des facteurs de transcription Sp 1,
AP-1 et NFI (Gingras, Larouche et al. 2003), nous avons déterminé les niveaux
d'expression et l'influence de chacun de ces facteurs dans les quatre lignées tumorales
ciblées (T97, T98, T108 et Tl15).

52
3.2 Matériel et méthodes
3.2.1 Culture cellulaire
Les lignées cellulaires T97, T98, TI08 et T115 sont dérivées directement de tumeurs
primaires isolées de patients ayant été diagnostiqués avec un mélanome uvéal. Les cellules
ont ~té cultivées dans le milieu de culture DMEM (Dulbecco's modified Eagle ' s medium)
additionné de 10% de sérum de veau fœtal, de 1 mM de glutamine et de 15 JlgIJlI de
gent~micine et incubées à 37°C sou~ une atmosphère de 5% de CO2. Le milieu a été changé
environ à tous les deux jours afin de permettre aux cellules de proliférer jusqu'à la densité
souhaitée (recouvrement d'environ 80% de la surface du flacon) à des fins d' extraction des
protéines nucléaires.
3.2.2 Culture cellulaire en agar mou
Les essais d'indépendance à l'ancrage ont ' été réalisés en mélangeant de l' agarose 2%
liquéfié à un volume égal de DMEM 2X sans sérum supplémenté de 15 Jlg/JlI de
gentamicine. Un millilitre de ce mélange a été versé çlans des pétris de 35 mm de diamètre
afin de créer une base d'agarose à 1 %. De l'agarose 0,6% liquéfié a ensuite été mélangé à
un volume égal de DMEM 2X et 1 ml de cette solution a été mélangé à 100 JlI de cellules
en suspension (correspondant à 104 cellules/ml) dans du milieu DMEM complet. Cette
suspension cellulaire de 0,27% d'agarose a ensuite été étendue sur la base d'agarose 1 %
gélifiée et 1 ml de DM~M complet contenant 10% de sérum de veau fœtal y a été ajouté
par-dessus cette préparation. Les pétris ont é,té incubés à 37°C sous une atmosphère de 5%
de CO2 pendant 7 jours et ont été photographiés en contraste de phase en utilisant un
microscope (Nikon TE 2000-S Eclipse) équipé d'une caméra numérique (Qimaging Retiga
1300) (Desprez, Lin et al. 1998).

53
3.2.3 Transfections transitoires
Les plasmides -92a5-CAT, -132a5-CAT et -954a5-CAT, qUI contiennent
respectivement la séquence du promoteur a5 comprise entre les positions -92 à +23 , - 132 à
+23 et -954 à +23, couplée au gène rapporteur de la chloramphénicol acétyle transférase
(CAT) ont été utilisés pour les transfection transitoires (Birkenmeier, McQuillan et al.
1991). Ces séquences contiennent ou non des mutations dans les sites de liaison pour les
facteurs de transcription SpI (dans les sites proximal et distal; -132a5-CAT/Mp, -132a5-
CAT/Md, -132a5-CAT/Mp+d), AP-1 et NFI (introduites par mutagenèse dirigée ; -92a5-
CAT/AP-1m+NFIm, -92a5-CAT/NFIm+SP1m, -92a5-CAT/SP1m+AP-1m, -92a5-
CAT/SP1m+AP-1m+NFIm (triple mutants)). Les différentes lignées de mélanome uvéal
(T97, T98, T108 et Tl15) ont été transfectées de façon transitoire avec les différents
vecteurs (1 flg par puits), dans des plaques à 6 puits contenant 200 000 cellules, à l'aide
d'un lipide polycationique (Lipofectamine; Invitrogen). Le plasmide pXGH5 (0,5 flg) , qui
code pour un variant sécrété de l'hormone de croissance humaine (hGH), a été ajouté à
. chaque puits. L'activité CAT a été évaluée par dosage enzymatique (Pothier, Ouellet et al.
1992) et la transfection a été normalisée par rapport à la quantité de GH sécrétée dans le
milieu de culture (ImmuChem Double antibody hGH 1251 RIA Kit; ICN Pharmaceuticals
Inc.) ainsi qu'à la quantité de protéine utilisée pour chaque mesure (dosage Bradford). Les
activités CAT devaient être au moins trois fois supérieures au bruit de fond, causé par le
mélange réactionnel seul, pour être considérées significatives. Les valeurs représentent la
moyenne d'au moins trois transfections effectuées en triplicata. Des tests statistiques (test
Student) ont été réalisés et les différences ont été considérées statistiquement significatives
lorsque p < 0,05.
3.2.4 Extraction d'ARN et analyse en RT-PCR
L'ARN total des lignées T97, T98, T108 et Tl15 a été isolé par l'ajout de la solution
d'extraction Tri Reagent (Molecular Researchcenter, Inc) à partir des cellules cultivées à
80%. La transcription inverse de l' ARN isolé a été réalisée en utilisant la trousse
«Superscript II Transcriptase» (Invitrogen Gibco) (Gingras, Larouche et al. 2003). L'ADN
complémentaire nouvellement transcrit a été purifié sur une colonne «QIAquick nucleotide

54
removal kit» (Qiagen) et utilisé pour l'amplification PCR du transcrit a5, des transcrits
encodant les facteurs de transcription SpI, AP-I et NFi et des enzymes HMBS et GAPD.
Les amorces utilisées pour l'amplification (Tableau 3.2) ont été dérivées à partir du logiciel
GenBankTM. Les amplifications PCR ont été réalisées avec la Taq polymérase PlatÏnum
(Invitrogen) en utilisant les paramètres suivants: 30 secondes à 95°C (dénaturation), 30
secondes à 58°C (hybridation) et 30 secondes à 72°C (élongation) pendant une période de
25 à 40 cycles selon le gène (a5 et GAPD : 25 cycles ; SpI, c-Fos, Fra-2 et NFI: 27 cycles ;
SP?: 35 cycles ; membres de la famille Jun, FosB, Fra-I et HMBS : 40 cycles). Les
produits PCR ont été révélés sur un gel d'agarose (1,6%) coloré au bromure d 'éthidium
(0,5 Jlg/ml).
Tableau 3.2 Séquences des amorces utilisées pour le RT-PCR semi-quantitatif Brin Sens Référence
Brin Anti-sens Produit PCR (bp) Genbank Gène
. _______________________________ i~~l'l _______________________________________ ~_______ Accession
a5
SpI
Sp3
NFI-A
NFI-B
NFI-C
NFI-X
c-Jun
JunB
JunD
c-Fos
FosB
Fra-l
Fra-2
GAPD
HMBS
CCAGGATGGCTACAATGATG CCCACAATCAGATCAGGATA
ATTGAGTCACCCAATGAGAACAG CAGCCACAACATACTGCCC
TCTCATACCACAGACTGGTCAA TTTATCAAATGTCCGTCGGCAT
ACGCTGAAAGAATTTGTCCAACT GGGGTCAGGTGGTCTGTCT
TATTCGCCAGGAGTATCGAGAG CTAGATCCAGACGCCAGACTT
TACCTGGCCTACTTCGTGC AGTTGGGTCCTGTTCCAGTCA
GAAGCCCGAGATCAAGCAGAA GAGGCGACTTGTAGAGCCG
TGGAAACGACCTTCTATGACGA GTTGCTGGACTGGATTATCAGAA
TACCACGACGACTCATACACA CGCTTTGAGACTCCGGTAGG
AAACACCCTTCTACGGCGATG GTCAGCGCGTCCTTCTTCAT
GTCTCCAGTGCCAACTTCATT CCTCCTGTCATGGTCTTCACA TTTCCCCGGAGACTACGACTC
CTGGTTGTGATCGCGGTGA TTCCGAGACTTCGGGGAAC
CCCCAGGAAATGAGGCTGTA AGAAATTCCGGGTAGATATGCCT
GGTATGGGTTGGACATGGAGG ATGCAACGGATTTGGTCGTAT TCTCGCTCCTGGAAGATGGTG
GGCAATGCGGCTGCAA GGGTACCCACGCGAATCAC
222 NM_002205
241 NM 138473
216 AF494280
300 NM 005595
149 NM_005587
205 NM_005597
275
242 NM 002228
129 NM_002229
148 .NM 005354
191 . NM_005252
180 NM_006723
177
129 NM 005253
220 NM 002046
64 NM 000190

55
3.2.5 Extraits nucléaires et immunobuvardage de type western
Les extraits nucléaires ont été préparés à partir des lignées de mélanomes (T97, T98,
T108 et Tl15) cultivées entre 80-90% de confluence cellulaire selon la procédure
développée antérieurement dans le laboratoire du Dr Guérin (Roy, Gosselin et al. 1991).
Les extraits nucléaires des différentes lignées cancéreuses provenant de mélanomes
uvéaux (25 Jlg) ont été fractionnés sur un gel SDS-PAGE (75 Volts) et transférés sur une
membrane de nitrocellulose (50 Volts). Ces membranes ont été lavées avec du tampon TS
(150 mM NaCl, 10 mM Tris-HCI pH7,4) et TSM (tampon TS auquel on ajoute 5% de lait
sans gras et 0,1% de Tween 20) pour ensuite être incubées avec un anticorps primaire dirigé
contre les FTs Sp 1, Sp3 et NFI (Santa Cruz ~iotechnology ; dilution 1 :2000) ainsi flue
contre les sous-unités JunB, c-Jun, JunD, FosB, c-Fos, Fra-1 et Fra-2 du facteur AP-1
(Santa Cruz Biotechnology ; dilution 1: 1000) ou avec un anticorps dirigé contre l' actine
(Jackson Immuno Research Laboratories Inc. ; dilution 1 :35 000) pendant 2 heures. Les
membranes ont ensuite été lavées avec du TSM et incubées avec un anticorps secondaire
dirigé contre les IgG de lapin couplé à la péroxydase pendant 1 heure. Les complexes
immunoréactifs ont été révélés par chemiluminescence (<<Western blot detection kit»;
Amersham Biosciences).
3.2.6 Retard sur gel (EMSA)
Les analyses de retard sur gel ont été réalisés selon le protocole décrit précédemment
(Chapitre 2; Section 2.2.3) en utilisant, en guise de sonde radiomarquée, des
oligonucléotides à haute affinité pour: SpI (5'GATCATATCTGCGGGGCGGGGCAG
ACACAG-3') (Dynan et Tjian 1983), NFI (5'-GATCTTATTTTGGATTGAAGCC
AATATGAG-3') (de Vries, van Driel et al. 1987) et AP-1 (5'-GATCCCCGCGTTGAGTC
ATTCGCCTC-3') (Coibi, Jensen et al. 2000). Les complexes ADN-protéines ont été
séparés sur un gel de polyacrylamide natif 8% en utilisant le tampon Tris-Glycine (50 mM
Tris, 2,5 mM EDT A, 0,4 M glycine) pour la migration (Schneider, Gander et al. 1986). Les
gels ont été séchés et les complexes ADN/protéine ont été révélés par autoradiographie.
Pour les compétitions, des oligonucléotides non-marqués ont été ajoutés au mélange

56
réactionnel en excès molaire à raison de I DOX ou 250X par rapport à la sonde. Des super
rétentions ont également été réalisées par l'ajout d'un anticorps spécifique dirigé contre
chacun des facteurs de transcription (SpI , Sp3, NFI et AP-I; Santa Cruz Biotechnology).

57
3.3 Résultats
3.3.1 Potentiel tu morigène et capacité des cellules 'à générer des colonies
en agar mou
Les images générées par microscopie en contraste de phase des différentes lignées
cancéreuses ont permis de déterminer à quel type morphologique (épithélial, fusiforme ou
mixte) appartenaient les cellules cancéreuses. En effet, en comparaison aux mélanocytes
normaux de la choroïde (MeN) (Figure 3.1), les lignées cancéreuses T97, T98, TI08 et
T115 (Figure 3.1) présentent toutes un patron mixte contenant néanmoins beaucoup de
cellules de type épithélioïde.
Figure 3.1 - Morphologie cellulaire. Microscopie en contraste de phase de cultures en monocouche des lignées cellulaires T97, T98, TI08 et Tl15 dérivées de mélanomes uvéaux primaires humains. MeN: Mélanocytes non-néoplasiques. Grossissement: 100X.

58
Les essais d'indépendance à l'ancrage réalisés en agar mou ont permis de déterminer si
les lignées cellulaires utilisées (T97, T98, Tl 08 et T 115) étaient réellement des cellules
cancéreuses. En effet, les cellules dites normales sont incapables de s'ancrer à ce type de
substrat, celles-ci entrant en apoptose, tandis que les cellules cancéreuses prolifèrent de
façon à former de petites colonies. L'expérience a été réalisée avec des mélanocytes
normaux de l' œil (MCN) en guise de contrôle négatif ainsi qu'avec les quatre lignées
provenant de tumeurs uvéales primaires (T97, T98, T108 et Tl15). Nous avons pu observer
l'absence complète de colonie avec les MCN (contrôle négatif) (Figure 3.2).
MeN
l00x 200x
'.
97
'. T108
Figure 3.2 - Indépendance à l'ancrage sur substrat d'agar mou. Les mélanocytes non-néoplasiques de l'œil humain (A) et les lignées dérivées de mélanomes uvéaux primaires T97 (B) et TI08 (C) ont été cultivés sur agar mou. Les cultures ont été photographiées 7 jours plus tard en microscopie en contraste de phase. Grossissement: lOOX et 200X.

59
Cependant, bien que les quatre lignées cancéreuses présentaient plusieurs colonies sur
agar mou, nous avons choisi d'illustrer les pétris ensemencés avec les lignées T97 (Figure
3.2 B) et T108 (Figure 3.2 C) puisque ces deux lignées représentent à la fois une lignée qui
sous-exprime la sous-unité a5 à sa surface cellulaire (T97) et une lignée qui exprime
davantage cette même sous-unité (T108) (Figure 1.15). Ces r~sultats confirment ainsi le
phénotype cancéreux des lignées de mélanome T97, T98, T108 et Tl15.
3.3.2 Activité transcriptionnelle du promoteur du gène aS
Des essais en transfections transitoires ont été réalisés afin de définir les nIveaux
d'activité transcriptionnelle dirigés par le promoteur du gène a5 dans les lignées T97, T98,
T108 et T 115. Des plasmides recombinants portant le gène rapporteur CAT fusionné à
différents segments du promoteur a5 (-92a5CAT, -132a5CAT et -954a5CAT) ont été
transfectés afin d'observer les différences entre ces lignées. Nous avons remarqué une plus
forte activité du promoteur a5 dans les lignées T108 et Tl15 comparativement aux lignées
T97 et T98 et ce, pour tous les segments du promoteur utilisées (Figure 3.3). Toutefois,
cette différence est davantage marquée lorsque les cellules sont transfectées avec le
plasmide portant le plus long segment du promoteur a5 (-954a5CAT). En effet, il est
possible d'observer une différence 'd'au moins dix fois dans l'activité du promoteur a5 dans
les lignées T108 et Tl15 comparativement aux lignées T97 et T98 lorsque le plus court
segment (-92a5CAT) est utilisé. Cette différence s'accroît à près de dix-huit fois avec le
plasmide -954a5CAT par rapport aux résultats obtenus dans la T98. Qui plus est, il s'agit
du segment du promoteur se rapprochant le plus de la version endogène du gène a5. Ceci
confirme donc que le gène a5 est plus fortement transcrit dans les lignées T108 et Tl15 que
dans les lignées T97 et T98.
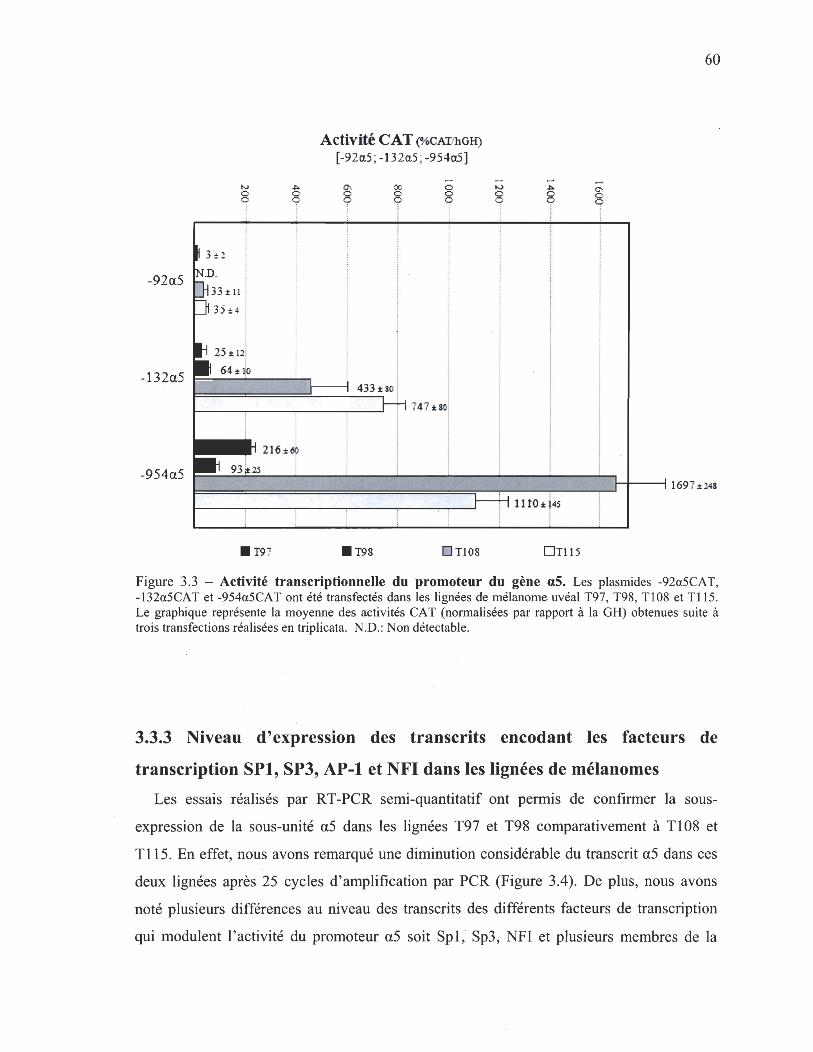
-92(15
-132u5
-954(15
Act ivité CAT ( oCAT/hGR)
[-9 2a5 --13 0.5 ~ -950.5]
...... 00 0
0 0 0
f f 9 0 0
......
.t>.o o
60
~================================~~ ____ -. ____ -.~+---~ 169 ±~
.T9 .T98 DT1 0B DTl1 5
Figure 3.3 - Activité transcriptionnelle du promoteur du gène aS. Les plasmides -92a5CAT, -I32a5CAT et -954a5CAT ont été transfectés dans les lignées de mélanome uvéal T97, T98, TI08 et T II5. Le graphique représente la moyenne des activités CA T (normalisées par rapport à la GH) obtenues suite à trois transfections réalisées en triplicata. N.D.: Non détectable.
3.3.3 Niveau d'expression des transcrits encodant les facteurs de
transcription SPI, SP3, AP-I et NFI dans les lignées de mélanomes
Les essais réalisés par RT -PCR semi-quantitatif ont permis de confirmer la sous
expression de la sous-unité a5 dans les lignées T97 et T98 comparativement à T108 et
T 115. En effet, nous avons remarqué une diI?inution considérable du transcrit a5 dans ces
deux lignées après 25 cycles d'amplification par PCR (Figure 3.4). De plus, nous avons
noté plusieurs différences au niveau des transcrits des différents facteurs de transcription
qui modulent l'activité du promoteur a5 soit SpI ,' Sp3, NFI et plusieurs membres de la

61
famille AP-l. Ainsi, des différences ont été observées au niveau du transcrit Sp 1, plus
abondant dans la T97, et le transcrit Sp3 qui, au contraire, semble plus abondant dans les
lignées TI08 et T115 (Figure 3.4).
- - -----
5 ~ ............
\....; 1 1 j
- - -
Sp1 ' p
c Jun
JunB, 1 ~
-------
Fa 1 ~
Fr -1 ........... ~ -.. --. "
NFI B
N tC
F X
GAPD
- - - - -
'-. ...... l---.j'
-- -- -----
~~ . - - -- -- -- --- - --
L.) II. 1. \
--Fl
Cris
Figure 3.4 - Niveaux des transcrits endogènes encodant la sous-unité aS et les facteurs de transcription Spl, Ap-l et NFI. L'ADN complémentaire a été produit par transcription inverse de l'ARN total isolé de différentes lignées dérivées de mélanomes uvéaux primaires et ensuite amplifié par PCR en utilisant des amorces spécifiques pour a5 (25 cycles); pour SpI (27 cycles) et Sp3 (35 cycles); pour les sousunités AP-l: membres Jun, FosB et Fra-I (40 cycles), c-Fos et Fra-2 (27 cycles); pour NF! (27 cycles); et pour les contrôles GAPD (25 cycles) et HMBS ( 40 cycles).
Les transcrits pour les sous-unités Jun du FT AP-I (c-Jun, JunB et Jun D) semblent
également plus fortement exprimés dans les lignées TI08 et Tl15 (Figure 3.4). Cependant,
les autres membres de la famille AP-I ne semblent pas suivre la même tendancé, exception
faite du transcrit Fra-l, qui est également plus élevé dans les lignées TI08 et Tl15 (Figure
3.4). Le transcrit de c-Fos semble moins abondant dans ces deux lignées (TI08 et T115)

62
alors que ceux de FosB ét Fra-2 ne démontrent aucune différence entre les quatre lignées
(Figure 3.4).
Finalement, il importe de souligner que des différences importantes dans l'expression de
certains isoformes de NFI, et plus particulièrement NFI-A et NFI-X ont été observées entre
ces lignées. En effet, nous avons noté l'absence complète du transcrit NFI-A ·dans la lignée
T98 et une forte expression de celui-ci dans la lignée 1'108 (Figure 3.4). Le transcrit de
NFI-X est plus fortement exprimé dans les lignées T97 et T98 que dans les lignées Tl08 et
Tl15 (Figure 3.4). La transcription des isoforriles NFI-B et NFI-C semble comparable entre
les quatre lignées de mélanome uvéal (Figure 3.4). En conclusion, plusieurs différences
dans l'expression des transcrits de certains facteurs de transcription importants pour
l'expression du gène a5 ont été .observées, ce qui pourrait contribuer à conférer les
propriétés qui caractérisent les lignées T97-T98 et Tl08-Tl15.
3.3.4 Niveau d'expression des facteurs de transcription SPI; SP3, AP-I et
NFI dans les lig~ées T97, T98, TI08 et Tl15
Par des essais d'immunobuvardage de type Western, nous avons évalué les niveaux
d'expression des FTs SpI, Sp3, AP-l et NFI dans les lignées cancéreuses T97, T98, Tl08
et T 115 . Nous avons ainsi ·observé que le niveau d'expression des facteurs de transcription
ciblés diffère d'une lignée cancéreuse à l'autre (Figure 3.5). On observe généralement un
pairage entre les lignées T97 et T98 par rapport aux lignées ·Tl08 et Tl15. En effet, le
niveau d'expression des facteurs semble varier selon le duo (T97-T98 ou Tl08-Tl15), mais
demeure toutefois identique au sein du même duo. En premier lieu, les différences notées
pour les FTs SpI et Sp3, bien que minimes, semblent indiquer une plus forte expression de
ces protéines dans les cellules Tl08 et Tl15 (Figure 3.5 A). Une variation contraire
survient cependant au niveau de l'expression de NFI, qui semble beaucoup plus fortement
exprimé dans les lignées T97 et T98 (Figure 3.5 B). Finalement, pour les membres de la
famille AP-l, tous semblent varier, mais demeurent néanmoins semblables au sein d'un
même duo. En effet, pour la famille Jun, nous avons remarqué que c-Jun et JunB sont
exprimés plus fortement dans les cellules T97 et T98, JunB étant très fortement exprimé

63
dans ces deux lignées (Figure 3.5 C). Cependant, JunD démontre une plus forte expression
dans les lignées TI08 et Tl15 que dans les T97 et T98 (Figure 3.5 C). Pour les autres
membres de la famille AP-I , nous avons observé une plus faible expression de tous les
facteurs chez T97 et T98 sauf pour Fra-2 qui semble, au contraire, beaucoup plus fortement
exprimé chez ces lignées (Figure 3.5 C). En effet, c-Fos, FosB et Fra-l sont tous les trois
moins exprimés chez T97 et T98 que dans les lignées TI08 et TIl5 (Figure 3.5 C). Les
différences observées chez chacun des duos pourraient également influencer de façon
différentielle l'activité du promoteur a5 et, par conséquent, le niveau d'expression de cette
sous-unité à la surface de la cellule.
A. c.
B.
1
Actine Actine
Figure 3.5 - Niveau d'expression des facteurs de transcription SpI, NFI et AP-I dans les lignées de mélanomes. Analyse d'immunobuvardage de type western réalisée avec 15Jlg de protéines nucléaires provenant de chacune des lignées. Les extraits ont été fractionnés sur gel, transférés sur membrane de nitrocellulose et incubés avec des anticorps dirigés contre les facteurs de transcription Sp 1, Sp3 (A), NFI (B), c-Jun, JunB, JunD, c-Fos, FosB, Fra-l ~ Fra-2 (C) et contre l'actine. La position des marqueurs de poids moléculaire (kDa) est indiquée sur la figure.

64
3.3.5 Capacité de liaison des facteurs de transcription SPI, AP-I et NFI
dans les lignées T97, T98, TI08 et Tl15
Afin de vérifier la capacité de liaison des facteurs de transcription dans les lignées de
mélanomes, nous avons réalisé des essais de retard sur gel. Dans un premier temps, des
essais ont été réalisés afin de déterminer la capacité de liaison à l'ADN de Sp 1 dans nos
quatre lignées cancéreuses: T97, T98, T108 et Tl15 (Figure 3.6). Ces résultats permettent
d'évaluer, par le fait même, la capacité de liaison à l'ADN de Sp3 puisque ce dernier
possède la capacité de se lier à la séquence çible également reconnue par Sp 1 (Philipsen et
Suske 1999; Scohy, Gabant et al. 2000). Nous avons observé que Sp3 possède une capacité
de liaison nettement supérieure dans les lignées T108 et Tl15 comparativement aux
cellules T97 et T98 (Figure 3.6). Nous avons également noté la formation d'un complexe
Sp1/Sp3 plus prononcé dans les lignées T108 et Tl15. Nous avons ensuite réalisé des
analyses en compétition et de super-rétention en gel afin de s'assurer de la spécificité des
signaux obtenus. Les essais de compétitions en gel ont permis de démontrer clairement la
spécificité du signal dans la lignée T108 puisque l'ajout de 250X en excès molaire d'un
oligonucléotide SpI non-marqué empêche complètement la formation du complexe
Sp1/Sp3 et réduit considérablement la formation du complexe Sp3 (Figure 3.6). Les
analyses de super-rétentions ont permis d'identifier les composantes protéiques du
complexe Sp1/Sp3 et de s'assurer de sa spécificité. On remarque que l'ajout d'une quantité
déterminée d'anticorps SpI ou Sp3, selon le cas, entraîne la formation d'un super-complexe
(SSC) autant dans la lignée T97 que dans la lignée TI08 (Figure 3.6) démontrant ainsi que
les anticorps spécifiques correspondants à ces facteurs se sont accrochés aux complexes
protéines-sonde. Il est important de noter qu'en dépit de l'ajout de la même quantité d'un
anticorps dirigés contre NFI, ,aucun super-complexe n'est produit dans les lignées T97 et
T108 (Figure 3.6). Nous sommes donc assurés de la spécificité de nos signaux etpouvons
confirmer que bien que Sp 1 ne semble pas varier beaucoup entre les lignées, Sp3 apparaît
avoir une plus forte capacité de liaison à l'ADN dans les lignées Tl 08 et T 115.

65
SpI
1
Figure 3.6 - Capacité de liaison des facteurs de transcription SpI et Sp3 dans les lignées de mélanomes. Analyses de retard sur gel réalisées avec 5,0 f..lg de protéines nucléaires obtenues de chacune des lignées. Ces extraits ont été incubés avec une sonde radiomarquée correspondant à un oligonucléotide consensus pour SpI. Afin de vérifier l'identité et la spécificité des complexes obtenus, des oligonucléotides non-marqués portant des sites à haute affinité pour Spi et AP-I ont été ajoutés (250x d'excès molaire) en guise de compétiteurs. Des anticorps dirigés contre les FTs Sp l, Sp3 et NFI (4 fll) ont également été utilisés en combinaison avec une sonde Sp 1 dans des essais de super-rétention en gel. Les complexes ADN/protéines ont été séparés sur gel de polyacrylamide (8%) et révélés par autoradiographie. U : sonde libre; SSC: Supercomplexe, SpllSp3: complexe Spl-Sp3.
Dans un deuxième temps, nous avons aussi réalisé des essais de retard sur gel afin de
vérifier la capacité de liaison à l'ADN du FT AP-l dans nos quatre lignées cancéreuses
(Figure 3.7).

66
ssc
Figure 3.7 - Capacité de liaison du facteur de transcription Ap-l dans les lignées de mélanomes. Analyses de retard sur gel réalisées avec 5,0 f..lg de protéines nucléaires préparées de chacune des lignées. Ces extraits ont été incubés avec une sonde radiomarquée correspondant à un oligonucléotide consensus pour Ap-l. Afin de vérifier l'identité et la spédficité des complexes obtenus, des oligonucléotides non-marqués portant des sites à haute affinité pour AP-I et pour NFI ont été ajoutés (IOOx ou 250x d'excès molaire) en guise de compétiteurs. Des anticorps dirigés contre les membres de la famille Jun (JunB, c-Jun, JunD) et NFI (4 J..lI) ont également été utilisés en combinaison avec une sonde Ap-I dans des essais de superrétention en gel. Les complexes ADN/protéines ont été séparés sur gel de polyacrylamide (8%) et révélés par autoradiographie. U : sonde libre; SSC: Super-complexe; NS: Non-spécifique.
Ainsi, nous avons observé des différences dans la liaison dè ce facteur entre les lignées
T97 et T98 versus TI08 et Tl15. En effet, AP-l possède la capacité de lier l'ADN
seulement dans les lignées T97 et T98 (Figure 3.7). Par l' ajout de compétiteurs (de 100X .à
250X d'excès molaire d'un oligo AP-l non-marqué), il a été possible de distinguer les
complexes ADN/protéine dont la formation est spécifique c'est-à-dire, ceux produits par

67
AP-1, de ceux non-spécifiques (signal observé dans les lignées T108 et Tl15) (Figure 3.7).
De plus, nous avons réalisé des essais de super-rétention à l'aide d'anticorps dirigés contre
certains membres spécifiques de la famille AP-1 dont JunB, c-Jun et JunD et que l'on
soupçonnait varier entre les lignées de rriélanome (l'anticorps NFI étant utilisé comme
contrôle négatif). Les résultats semblent démontrer que le signal observé correspond
davantage à JunB et à JunD puisque l'addition individuelle de ces anticorps entraîne la
fomation d'u? super-complexe, mais n'empêche que partiellement la formation du
complexe AP-1 alors qu'en combinaison, les anticorps JunB et JunD empêchent
complètement sa formation (Figure 3.7). Il est important de noter. que le super-complexe ne
représente pas la bande très intense observée à la limite supérieure des puits, mais une
petite bande plutôt diffuse tout juste en dessous. Ce super-complexe n'apparaît que lors de
l'ajout des anticorps JunE et JunD et non avec les anticorps c-Jun et NFI (Figure 3.7). Ces
données confirment que AP-1 possède la capacité de lier l'ADN seulement dans la lignée
T97 et aussi dans la lignée T98 (non montré) et que les membres JunB et JunD semblent les
comp<?santes Jun majeures de ce facteur.
En dernier lieu, nous avons également effectué des essais de retard 'sur gel afin de
vérifier, cette fois-ci, la capacité de liaison à l'ADN de NFI dans ces mêmes lignées. Nous
avons encore une fois observé une nette différence entre les lignées T97 et T98 et les
lignées Tl 08 et TIl 5'. En effet, un signal beaucoup plus intense est observé pour NFI avec
les extraits nucléaires des lignées T97 et T98, ce signal étant à peine détectable dan·s les
extraits provenant des deux autres lignées (T108 et Tl15) (Figure 3.8). Les essais de
compétition dans des essais de retard sur gel nous ont permis de confirmer que la protéine
produisant le complexe apparaissant à la figure 3.8 appartient à la famille NF!. En effet,
l' oligonucléotide non-marqué portant la séquence cible NFI empêche totalement la
formation de ce complexe alors qu'un oligonucléotide portant la séquence cible AP-1 en est
incapable. Les essais de super-rétentions confirment également la spécificité du signal
correspondant à NF!. En effet, la formation d'un super-complexe n'est observée qu'avec
l'ajout d'un anticorps dirigé contre NFI, autant dans la lignée T97 que dans la lignée T108
(Figure 3.8) et non avec l'anticorps contrôle dirigé contre SpI. Nous sommes donc assurés
de la spécificité du signal observé dans nos quatre lignées cancéreuses et avons pu ainsi

68
identifier le signal correspondant à NF!. Nous pouvons également affirmer que NFI
possède une plus grande capacité de liaison à l'ADN dans les lignées T97 et T98
comparativement aux lignées Tl 08 et T 115.
Figure 3.8 Capacité de liaison du facteur de transcription NFI dans les lignées de mélanomes. Analyses de retard sur gel réalisées avec 5,0 /-lg de protéines nuCléaires préparées de chacune des lignées. Ces extraits ont été incubés avec une sonde radiomarquée correspondant à un oligonuc1éotide consensus pour NF!. Afin de vérifier l'identité et la spécificité des complexes obtenus, des oligonuc1éotides non-marqués portant des sites à haute affmité pour NFI et AP-I ont été ajoutés (IOOX ou 250X d'excès molaire) en guise de compétiteurs. Des anticorps dirigés contre NFI et Sp 1 (4 /-lI) ont également été utilisés en
. combinaison avec une sonde NFI dans des essais de super-rétention en gel. Les complexes ADN/protéines ont été séparés sur gel de polyacrylamide (8%) et révélés par autoradiographie. U : sonde libre; SSC: Supercomplexe; NS: Non-spécifique.

~~---- --- ~-~ ~--
69
3.3.6 Influence des facteurs SPI, AP-I et NFI dans les lignées T97 et TI08
Les transfections transitoires réalisées ont permis de déterminer plus précisément
l'influence des facteurs de transcription SpI , AP.-1 et NFI sur l'activité transcriptionnelle
du promoteur de gène de la sous-unité a5. À cette fin, des mutations ont été introduites, par
mutagénèse dirigée, dans les sites pour ces FTs dans le promoteur a5. Une première série
de transfections a été réalisée avec le promoteur -132a5 afin de déterminer séparément
l'influence spécifique de chacun des sites Sp 1 : proximal (situé entre les positions -60 et -
52) et distal (situé entre les positions -117 et -101). Les plasmides recombinants contenant
la séquence du promoteur et portant ou non un des deux sites Sp 1 muté (ou la combinaison
des deux sites mutés) ont été transfectés dans les lignées T97 et TI08 (Figure 3.9).
L'activité CAT du promoteur a5 a été rapportée en activité relative en référence à l 'activité
de base obtenue avec le plasmide -132a5CAT ne portant aucune mutation dans le
promoteur a5 (correspondant à 100%) (Figure 3.9). Lorsque les deux sites SpI sont mutés,
une chute considérable d~ l'activité du promoteur est obtenue autant dans la lignée T97 que
dans la lignée T108 (baisse de 65% et de 75%, respectivement) (Figure 3.9) démontrant
clairement que SpI est un activateur du promoteur a5. Toutefois, la mutation individuelle
des sites Sp 1 n' entaîne aucune modification statistiquement valable à l'exception d'une
légère augmentation dans la lignée T108 observée lorsque le site Sp 1 distal est muté
(Figure 3.9). Les résultats obtenus par mutation des sites Sp 1 individuellement pourraient
s'expliquer par la redondance des sites Sp 1. En effet, lorsqu'un site est muté, Sp 1 se lie au
second site afin de maintenir l'activité de base du promoteur a5 (Bergeron et al. 1997). En
conclusion, on peut affirmer . que SpI est un activateur de l'activité du promoteur a5 dans
les lignées T97 et T108.
Nous avons ensuite réalisé des transfections avec un plasmide recombinant portant un
segment plus court du promoteur a5 (-92a5) et dans lequel nous avons introduit des
mutations dans l~s sites Sp 1, AP-1 et NFI, individuellement ou en combinaison. Ces
plasmides recombinants ont été transfectés dans les lignées T97 et T108. Étant donné que le
promoteur -92a5 possède une très faible activité dans la lignée T97 (Figure 3.3), il a été
difficile de mettre les résultats en perspective d'autant plus que les lignées T97 et Tl 08
semblent varier différemment. Il a néanmoins été possible. de tirer certaines conclusions

70
intéressantes pour la suite du projet. Tout d'abord, comme on s'y attendait, nous n 'avons
observé aucune activité du promoteur a5 dans la lignée T97 et une faible activité, mais
détectable, dans la lignée T108 lorsque le promoteur est intact (sans mutation) (Figure
3.10).
Activité relative CAT (%) [-132a5 ]
Vl Q
o o
-1 32u5CAT 1----""""------1
-1 32u5/Mp I-----...J.....---..., I
Vl
?
99 ±8 1.
-1 32u51Md I-----.....:....-------i..---.l---.,
-1 32a 5/Mp+d f 19 *
tv o
?
167 ±33 *
D TI 08
Figure 3.9 - Influence des sites Spl proximal et distal sur le promoteur a5. Le plasmide -132a5CAT portant ou non des mutations dans le site SpI proximal et/ou distal a été transfecté dans les lignées cancéreuses T97 et TI08. Le graphique représente la valeur relative de" la moyenne des activités CAT (normalisées par rapport à la GR) obtenues suite à trois transfections réalisées en triplicat. Mp: mutation dans le site proximal; Md: mutation dans le site distal; Mp+d: mutation dans les sites distal et proximal.

-92a5 CAT
-92a5 /SpIm+APlm . j
o 7 t O,l: *
Activité CAT (%CAT GH)
[-92a5]
N o
w o
-9 a5/API m+NFIm t----....:......-------,
VI o
-92a5/NFIm Spim t--__ ....l.-______ .l...-__ "'"""--__ ~ __ ___.:.....,
71
t----...,.-------,----.,..----....,....-------~I---+-----I 61 t 15 *
N .D . -92a5/Splm APlm NF1m t----....i-.------'----"'------,
!----'----i 37 t 11 *
D T108
Figure 3.10 - Influence des FTs SpI, AP-I et NFI sur le promoteur basal aS. Le plasmide -92a5CAT portant ou non des mutations dans les sites SpI, AP-I ou NFI a été transfecté dans les lignées cancéreuses T97 et TI08. Le graphique représente la valeur moyenne des activités CAT (normalisées par rapport à la GH) obtenues suite à trois transfections réalisées en triplicat. Splm+APlm: mutations dans les sites SpI et AP-I ; APlm+NFIm: mutations dans les sites AP-I et NFI; NFIm+Splm: mutations dans les sites NFI et SpI du promoteur -92a5; N.D.: Non détectable.
Lorsque seul le site NFI est intact (dans le plasmide -92a5/SpIm+AP-Im), une chute
presque complète de l'activité dans la lignée TI08 est observée (la lignée T97 ne présentant
aucune activité notable) (Figure 3.10). Ces résultats indiquent que NFI serait
vraisemblablement un répresseur de l'activité du promoteur a5, ce qui est en accord avec
d'autres résultats obtenus précédemment dans notre laboratoire (Gingras, Larouche et al.
2003). Lorsque seul le site SpI est intact (dans le plasmide -92a5/AP-Im+NFIm), une
augmentation de près de trois fois de l'activité initiale du promoteur est observée dans la
lignée TI08 (Figure 3.10). Ces résultats semblent en accord avec ceux obtenus avec la
mutation du site SpI. pro~imal (Figure 3.9). Toutefois, aucun changem~nt n'a été observé
dans l'activité du promoteur lorsque transfecté dans la lignée T97 (Figure 3.10). Cependant,
lorsque le seul site AP-I est conservé intact (dans le plasmide -92a5/NFIm+SPIm), une
augmentation considérable de l'activité du promoteur est observée dans les lignées T97 et

72
Tl08 (augmentation de plus de 9 fois) (Figure 3.10). AP-l semble donc être un FT très
important dans la lignée T97 et représente sans aucun doute un activateur puissant du
promoteur a5 autant dans la lignée T97 que dans la lignée Tl08. Finalement, lorsque les
séquences cibles des trois FTs sont mutées simultanément (dans le plasmide
-92a5/SPlm+AP-lm+NFlm), aucune activité n 'est observée dans la lignée T97 tandis
qu'une hausse de plus de 5 fois dans l' activité du promoteur est mesurée dans la lignée
Tl08 (Figure 3.10). Ces résultats portent à croire qu'un autre facteur, un activateur, pourrait
contribuer à l'augmentation de l'activité du promoteur a5 observée dans la lignée Tl08.
Prenant en considération que la lignée T98 suit la même tendance que la lignée T97 et Tl15
avec Tl08, on pourrait penser que cette observation expliquerait en partie la dichotomie qui
existe entre les paires de lignées cancéreuses (T97 -T98 versus Tl 08-T 115).

73
3.4 Discussion
Les données cliniques que nous avons répertoriées provenant des patients atteints d'un
mélanome uvéal semblent indiquer une certaine tendance suggérant que les patients qui
expriment plus fortement la sous-unité d'intégrine aS soient encore en vie ou du moins
qu'ils ne soient pas décédés des suites du mélanome uvéal. Toutefois, plusieurs patients
. dont les lignées cellulaires dérivées des tumeurs primaires démontrent une plus faible
expression à leur surface de la sous-unité aS sont décédés des suites de l'apparition de
métastases hépatiques. Nous avons concentré' notre étude sur quatre lignées dérivées de
tumeurs primaires de patients atteints d'un mélanome uvéal, T97, T98, T108 et T11S. Nous
avons choisi ces lignées puisque nous possédions des informations sur le suivi de ces quatre
patients, toujours en vie, et qu'elles semblaient agir en paire bien qu'elles proviennent
toutes de tumeurs uvéales différentes. En effet, les lignées T97 et T98 semblent réagir de la
même manière et expriment peu la sous-unité d'intégrine a5 (Figure 1.15). Les lignées
T108 et T 115 partagent également plusieurs de leurs propriétés et à la différence .des
lignées T97 et T98, elles expriment plus fortement la sous-unité a5 (Figure 1.15). Ces
nouvelles lignées non encore caractérisées ont donc fait l'objet de plusieurs expériences
afin d'établir leurs caractéristiques respectives.
Ainsi, ces lignées démontrent plusieurs des propriétés typiques des cellules cancéreuses :
elles présentent non seulement une morphologie épithélioïde très distincte de la
morphologie fusiforme des mélanocytes normaux de l'œil (Figure 3.1), mais présentent
également, la capacité de croître en agar mou (Figure 3.2), une caractéristique que ne
partagent pas les cellules non-transformées.
Les premiers résultats obtenus en transfection (Figure 3.3) ainsi que par RT-PCR semi
quantitatif réalisés avec les amorces a5 (Figure 3.4) confirment également que l'activité du
promoteur a5 est plus élevée dans les lignées Tl 08 et T 115 et que le transcrit endogène a5
y est également plus abondant. Suivant cette confirmation, nous avons déterminé plus
p~écisément qu' elle( s) pourrait( ent) être la ou les causee s) de la diminution d'expression de
la sous-unité a5 ou de son augmentation selon le cas. Les facteurs de transcription SpI, Ap·

74
1 et NFI ont été ciblés pour la présente étude. En effet, d'autres travaux réalisés dans notre
laboratoire avaient permis d'identifier ces facteurs de transcription comme jouant un rôle
prépondérant dans l'activation ou la répression de l'activité du promoteur a5 (Gingras,
Larouche et al. 2003; Gingras et al. 2008). Des différences ont été observées au niveau de
ces facteu~s chez les lignées et pourraient potentiellement expliquer pourquoi la sous-unité
a5 est sous-exprimée dans les lignées T97 et T98.
De celles-ci, certaines s'inscrivent premièrement au niveau de Sp 1. On remarque de
légères différences au niveau du transcrit SpI (Figure 3.4) qui semble plus fortement
exprimé dans la lignée T97. Ce résultat va à l'encontre des résultats obtenus en ·
immunobuvardage western où la protéine Sp 1 semble plus fortement exprimée dans les
lignées T108 et Tl15 (Figure 3.5). Cette différence n'est cependant que très faible et
l'incohérence qui semble survenir entre ces résultats peut s'expliquer notamment par le fait
qu'il y a une très grande nuance à faire entre l'expression d'un ARNm et l'expression de sa
protéine. En effet, il se peut que l'ARNm SpI soit plus instable dans les lignées T98, T108
et T 115 ce qui pourrait expliquer cette légère variation. Il faut également noter que .la
technique utilisée, le RT-PCR, demeure une technique semi-quantitative. Les résultats
obtenus avec les essais de retard sur .gel ne démontrent pas non plus une différence
importante entre les lignées (Figure 3.6) et suggèrent que SpI aurait la même capacité à lier
l'ADN dans les quatre lignées. Finalement, des différences entre les lignées T97 et Tl 08
ont été observées suite aux transfections réalisées. Les résultats obtenus démontrent que .
Sp 1 est un activateur du promoteur a5 dans nos lignées. Il est cependant important de noter
que Sp3 peut également lier la même séquence que SpI sur le promoteur a5. Nous validons
présentement ces résultats par la transfection de petits ARN interférents qui ciblent
spécifiquement l'un ou l'autre des FTs (Sp 1 ou Sp3) afin de vérifier lequel peut avoir le
plus d'influence sur l'activité du promoteur. De plus, étant donné les minimes différences
qui existent entre les lignées (immunobuvardage western ou RT-PCR), il est peu probable
que Sp 1 soit le principal FT responsable de la forte expression de la sous-unité a5 dans les
lignées Tl 08 et T115.

75
Pour le FT Sp3, les résultats indiquent un meilleur potentiel en tant que FT candidat
dans la régulation de l'expression du gène a5. En effet, les résultats obtenus en R T -PCR
ont révélé une légère augmentation du transcrit Sp3 dans les lignées T108 et Tl15 (Figure
3.4). Il serait intéressant de reprendre cette analyse en PCR quantitatif ou PCR en temps
réel afin de confirmer ce résultat. Les analyses en immunobuvardage de type western
indiquent un niveau protéique de Sp3 plus élevé dans les lignées T108 et Tl15. De plus,
suite aux essais de retard sur gel réalisés, nous avons observé une plus forte activité de
liaison de Sp3 dans ces mêmes lignées (T108 et Tl15). Dans les transfections· réalisées
avec la version courte du promoteur a5 (-92a5CAT), une très forte augmentation de
l'activité CAT (Figure 3.10) a été observée dans la lignée T108 lorsque le site Sp1/Sp3 est
intact et que les sites NFI et AP-1 sont mutés. Plusieurs études démontrent notamment que
Sp3 peut agir en tant qu'activateur de la transcription (Gingras, Larouche et al. 2003;
Oguariri, Brann et al. 2007; Xu et Shu 2007; Xu, Zhang et al. 2007). Les transfections
réalisées avec le plasmide -132a5CAT vont également dans le même sens: que ce soit Sp 1
ou Sp3 qui se lie sur le site du promoteur a5, ce dernier ag~t en tant qu'activateur de la
transcription autant dans la lignée T97 que dans la lignée Tl 08 puisque la mutation des
deux sites Sp 1/Sp3 (sites proximal et distal), entraîne une diminution considérable de
l'activité du promoteur a5 (Figure 3.9). L'absence d'effet négatif observable suite aux
transfections réalisées avec les mutations simples (-132a5CAT/Mp et -132a5CAT/Md) est
probablement due à la répétition des sites de liaison. En effet, lorsque les sites Sp 1/Sp3 sont
mutés individuellement, le site intact compense afin de maintenir l'activité du promoteur a5
(Bergeron et al. 1997; Gaudreault et al. 2007). Finalement, étant donné que Sp3 semble
plus abondant dans les lignées T108 et Tl15, il pourrait être responsable de l'expression
différentielle de la sous-unité a5 dans ces lignées.
On remarque également des différences au niveau de NF!. Ce facteur, reconnu comme
un répresseur de l'activité du promoteur a5, notamment dans le contexte de la cicatrisation
cornéenne (Gingras, Larouche et al. 2003), pourrait également s'avérer être un répresseur
dans nos lignées cancéreuses. En premier lieu, des différences au niveau des transcrits
encodant les isoformes NFI-A et NFI-X ont été observées (Figure 3.4). En effet, nous avons
noté une plus forte expression de NFI-A dans les lignées TI08 et TII5 et une absence

-- -------- -------- ----,
76
complète du transcrit correspondant dans la lignée T98. De plus, le transcrit NFI-X semble
à première vue, plus abondant dans les lignées T97 et T98. En effet, selon les résultats
obtenus en immunobuvardage western, qui ne fait aucune distinction entre les isoformes de
NFI puisque l'anticorps utilisé est dirigé contre un épitope du domaine de liaison à l'ADN
partagé par les quatre isoformes, nous avons noté une très forte expression de ce FT dans
les lignées T97 et T98 comparativement aux lignées T108 et Tl15 (Figure 3.5). Cette
même tendance se remarque aussi dans les essais de retard sur gel. En effet, NFI semble
avoir une plus forte capacité à lier l'ADN dans les lignées T97 et T98, un très faible signal
étant néanmoins généré par les extraits nucléaires provenant des lignées Tl 08 et T 115
(Figure 3.7). En dernier lieu, les résultats obtenus suite aux transfections réalisées avec le
plasmide portant la version courte du promoteur a5 (-92a5), ont révélé une influence
négative exercée par NFI sur l'activité du promoteur a5 dans la lignée T108 lorsque le site
NFI est intact alors qu'une augmentation de l'activité du promoteur est observée chaque
fO,is que ce site est muté (Figure 3.10). Bien que la fonction de NFI n'ait pu être confirmée '
dans la lignée T97 étant donné que l'activité du promoteur basal mesurée est nulle, les
résultats obtenus dans la lignée T108 démontrent clairement l'influence négative que peut
avoir NFI sur l'activité du promoteur aS. Néanmoins, on peut supposer qu'étant beaucoup
plus abondant dans les lignées T97 et T98, NFI demeure le candidat le plus probable pour
jouer le rôle de répresseur de l'activité du promoteur a5 dans ces lignées, NFI contribuant
ainsi à maintenir un niveau très faible de la sous-unité a5 dans ces lignées. De surcroît, les
différents isoformes de NFI ont été rapportées pour agir en, tant que répresseur ou activateur
selon le type et le contexte cellulaire (Gronostajski 2000). Ainsi, il est possible que
l'isoforme NFI-A, qui semble plus abondant dans les lignées T108 et Tl15, ne soit pas un
répresseur, mais bien un activateur de l'activité du promoteur aS et que l'isoforme NFI-X
soit, quant à lui, un répresseur dans les lignées T97 et T98. Afin de confirmer cette
hypothèse, il serait intéressant de réaliser des transfections transitoires à l'aide de petits
ARN interférents qui cibleraient chacun des isoformes dans le but de vérifier l'influence
qu'aurait l'inhibition d'un isoforme particulier sur l'activité du promoteur a5.
Le FT AP-1 est plus complexe que les FTs Sp 1 et NFI puisque celui-ci est un dimère
résultant de l'association entre une sous-unité de la famille Jun (c-Jun, JunB et JunD) et

77
d'une sous-unité de la famille Fos (c-Fos, FosB, Fra-1 et Fra-2). Des différences au niveau
du transcrit ont été observées pour JunB, JunD et Fra-1 qui semblent exprimés à un niveau
plus élevé dans les lignées Tl 08 et T 115" alors que c-Fos est plus fortement exprimé dans
les lignées T97 et T98 (Figure 3.4). Ces résultats ne s'accordent cependant pas tout à fait
avec ceux obtenus en immunobuvardage western, l'instabilité des ARNs AP-1 pouvant
expliquer en partie ces différences. Du point de vue de la protéine, nous avons remarqué
que certains facteurs semblent prépondérants chez les lignées T97 et T98. En effet, c-Jun,
JunB et Fra-2, qui sont fortement exprimés dans ces lignées, ne sont pas (JunB et Fra-2) ou
peu (c-Jun) exprimés dans les T108 et Tl15 (Figure 3.5). D'autres sous-unités semblent
cependant prépondérantes dans les lignées Tl 08 et T 115. De la famille Jun, seul JunD est
plus fortement exprimé dans ces lignées alors que la majorité des membres de la famille
Fos l'est, à l'exception de Fra-2 (Figure 3.5). L'absence d'une sous-unité peut empêcher
notamment AP-I d'être actif et de pouvoir lier l'ADN. Cette possibilité pourrait expliquer
pourquoi, malgré la présence de plusieurs sous-unités, aucun signal AP-I n'a pu être
observé suite aux essais de retard sur gel dans les lignées TI08 et Tl15 (Figure 3.7). Ce
signal est cependant présent dans les lignées T97 et T98 et nous indique que l 'hétérodimère
AP-I exprimé dans ces lignées (probablement constitué des sous-unités JunB/Fra-2)
possède la capacité de lier l'ADN. De plus, les analyses de super-rétention nous informent
que la sous-unité JunB est présente dans le complexe AP-I (Figure 3.7). Finalement,
sachant que AP-I est très souvent reconnu pour être un activateur de la transcription
(Bames 2006; Rosskothen, Hirschmuller-Ohmes et al. 2008), les résultats obtenus en
transfection confirment · également une telle fonction pour ce FT dans nos lignées
cancéreuses. En effet, autant dans la lignée T97 que dans la lignée Tl 08, l'activité
transcriptionnelle du promoteur a5 augmente considérablement lorsque seul le site AP-I est
intact (Figure 3.10). Toutefois, les résultats obtenus en retard sur gel n'indiquaient aucun
signal AP-I dans les TI08 et Tl15 alors que le FT est actif dans ces lignées (résultats
obtenus en transfections lorsque seul le site AP-I est intact). Ceci peut s'expliquer par la
quantité de protéines utilisées lors des essais en retard sur gel. Il est possible que la quantité
de protéines utilisée (5Jlg) était trop faible pour voir le complexe malgré qu'il soit tout de
même présent dans les lignées TI08 et T115, mais en très faible quantité. De plus, les
résultats obtenus en transfection suggèrent que cet hétérodimère AP-! n'est pas un

78
activateur suffisamment puissant pour lever la répression observée dans la lignée T97
lorsque le promoteur ne contient aucune mutation. Nous pouvons ainsi conclure que malgré
qu'AP-I soit présent dans les lignées T97 et T98, il ne semble pas suffisant pour activer la
transcription du promoteur aS peut-être parce qu'il entre en compétition avec un autre FT
(un répresseur fort) qui l'empêcherait de se lier sur le promoteur et d'en activer la
transcription ou encore, l 'hétérodimère JunB/Fra-2 est peut-être un faible activateur de la
transcription du promoteur aS.
En guise de · conclusion, il est certain que ces facteurs agissent de façon dynamique et
dans un contexte cellulaire. On doit donc tenir compte non seulement de l'environnement
cellulaire, mais également du rôle que peuvent exercer plusieurs facteurs agissant de
concert ou en compétition. Certaines études réalisées précédemment ont permis de
démontrer que certains FTs étaient capables de se lier simultanément sur le promoteur aS
alors que d'autres en étaient incapables (Gingras et al. 2008). En effet, les FTs SpI et AP-I
semblent aptes à se lier simultanément sur le promoteur tout comme NFI et AP-I.
Cependant, lorsque NFI se lie sur le promoteur, il empêche SpI de s'y lier (Gingras et al. .
2008). Suite aux différences observées entre les lignées T97-98 et TI08-IIS, il est possible
de formuler une hypothèse plausible pour expliquer la sous-expression de la sous-unité aS
dans' les lignées T97 et T98 qui pourrait, rappelons-le, être la cause de leur comportement
plus agressif. En premier lieu, Sp l ne semble pas constituer un élément déterminant,
contrairement à Sp3. En effet, ce dernier, étant abondant dans les lignées TI08 et Tl15,
pourrait entrer en compétition avec NFI, qui lui, présent en faible quantité dans ces lignées,
ne pourrait lier le promoteur pour en réprimer sa transcription et permettrait à Sp3 de s'y
lier pour l'activer. Le phénomène contraire pourrait survenir dans les lignées T97 et T98.
En effet, puisque NFI est très abondant dans ces lignées, il pourrait notamment empêcher la .
liaison de Sp3 sur le promoteur et inhiber l'effet activateur que pourrait avoir AP-I. Il en
résulterait une répression très forte de l'activité du promoteur dans les lignées T97 et T98.
Un FT autre que NFI, AP-l, Sp l ou Sp3 pourrait également lier le promoteur aS et agir de
concert avec NFI dans la répression du promoteur aS observée dans les lignées T97 et T98,
ce qui expliquerait qu'en dépit de la mutation du site NFI, il n'y ait aucune augmentation
dans l'activité du promoteur en transfections dans la lignée T97. En effet, on remarque la

79
présence d'un site de liaison pour le FT CEBP dans la région promotrice a5 (Figure 1.14).
Selon certaines études, ce FT peut agir en tant que répresseur, notamment sur le promoteur
de l' intégrine a Il (Lopez-Rodriguez, Botella et al. 1997) et sur le promoteur de CIIT A
(transactivateur du complexe majeur d'histocompatibilité classe II) (Pennini et al. 2007). Il
se pourrait que son influence s'avère également négative sur l'activité transcriptionnelle du
promoteur a5 dans nos lignées cancéreuses. Il serait donc intéressant d'approfondir l'étude
de ce FT afin de déterminer également son influence sur l'activité du promoteur a5.
Les résultats obtenus lors de cette étude ont fourni un certain nombre de pistes qUI
permettraient d'expliquer la faible expression de la sous-unité a5 dans les lignées T97 et
T98. De plus, certains résultats préliminaires non présentés lors de cette étude démontrent
qu'effectivement, ces lignées seraient plus agressives que les lignées T108 et Tl15. En
effet, nous avons injecté sous-cutané des cellules de chacune des lignées (T97, T98, T108
et Tl15) dans des souris immunosupprimées (CD1 : nude). Après seulement deux semaines
post-injections, nous avons pu observer la présence de tumeurs d'un diamètre de plus de
1cm dans les souris injectées avec les lignées T97 et T98 ·.alors qu'aucune tumeur n 'était
détectable dans les souris injectées avec les lignées T108 et Tl15. Il semble donc que les
lignées sous-exprimant la sous-unité a5 soient ainsi plus agressives in vivo que celles
. exprimant un haut niveau de cette intégrine. Ces résultats doivent cependant être confirmés
afin de s'assurer de leur reproductibilité. Pour donner suite à ce projet, nous avons prévu
réaliser des expériences de transfections stables avec des lentivirus exprimant des shARN
visant à réduire l'expression des FTs NFI, Sp 1, Sp3 et AP-1 dans les lignées T97, T98,
T108 et Tl15. Ces transfections auront pour but d'évaluer l'effet exercé par l'inhibition de
ces FTs sur l'activité du promoteur a5. Ces cellules seront ensuite injectées dans les flancs
de souris immunosupprimées dans le but de vérifier si la progression tumorale est modifiée
selon le type cellulaire en réponse à une modification dans l'expression des niveaux
endogènes de a5. Ces expériences permettront de vérifier s'il existe une corrélation entre la
sous-expression de la sous-unité a5 et l'agressivité tumorale des lignées utilisées.
Finalement, des expériences d'immunoprécipitation de la chromatine pourraient également
permettre de comparer le recrutement · ~es différents facteurs de transcription aux
promoteurs des sous-unités des intégrines, entre les différentes lignées cellulaires.

80
4. CONCLUSION
4.1 Résumé
Les objectifs de ce mémoire consistaient, dans un premier temps, à déterminer si le FT
Pax-6 est responsable de l'expression aberrante de la sous-unité d'intégrine a4 dans une
lignée dérivée de métastases secondaires provenant d'un patient atteint d'un mélanome
uvéal afin de déterminer si une corrélation existe entre l'expression de ce facteur et la
capacité qu'ont les cellules à migrer vers d'autres organes. Dans un deuxième temps, et
dans un tout autre ordre d'idées, nous avons voulu déterminer si la sous-expression de la
sous-unité d'intégrine a5 était reliée à un phénotype plus agressif des cellules cancéreuses
et définir quelles pouvaient être les conséquences de cette sous-expression dans des cellules
dérivées de tumeurs primaires de patients atteints d'un mélanome uvéa!.
4.1.1 Influence de Pax-6 sur le promoteur du gène «4
En guise de conclusion pour la première partie de l'étude, nous avons démontré que
l'activité du promoteur a4 est effectivement plus élevée dans la lignée H79 dérivée de
métastases hépatiques d'un patient atteint d'un mélanome uvéa!. Toutefois, nous avons
également démontré que Pax -6 n'est pas l'élément déterminant dans l'expression aberrante
de cette sous-unité dans cette lignée. En effet, ce dernier semble davantage réprimer
l'activité transcriptionnelle du promoteur a4 chez les lignées de mélanome étudiées. Des
candidats potentiels pourraient quant à eux être responsables de la trans-activation du
-promoteur a4, notamment certains membres de la famille Ets reconnue pour être une
famille ·d' activateurs puissants de la transcription génique.
Cependant, étant donné que cette intégrine est très impliquée dans la migration
cellulaire, il demeure encore à déterminer si ces observations pourraient corréler avec le
pouvoir métastatique de la lignée H79 par, et ceci n'est qu'un exemple, des essais
. d'implantation in vivo de tumeurs intraoculaires des cellules cancéreuses des différentes

81
lignées marquées au GFP chez des souris immunosupprimées afin de suivre l'atteinte du
foie (et des a~tres organes) par les métastases associées à ce type de cancer.
4.1.2 Influence de la sous-unité aS dans les propriétés tumorigènes du
mélanome uvéal
Les conclusions obtenues suite à la seconde partie de l'étude ont également ouvert des
portes pour des études a posteriori. Tout d'abord, nous avons confirmé le potentiel et le
phénotype cancéreux de nos quatre lignées utilisées pour l'étude: T97, T98, TI08 et TII5.
Nous avons ensuite démontré que l'activité du promoteur a5 était plus faible dans les
!ignées T97 et T98 comparativement aux lignées TI08 et TII5. Ces résultats étaient en
accord avec les résultats obtenus précédemment par analyse en cytométrie de flux (Figure
1.15). De plus, nous avons observé plusieurs différences au niveau de l'expression des FTs
SpI, Sp3, AP-I et NF! et avons pu discriminer ceux qui pourraient influencer (soit de façon
positive ou de façon négative) l'expression de la sous-unité a5 de ceux qui ne semblent
exercer aucune influence importante dans cette régulation. Selon les résultats obtenus, les
conclusions possibles suite à cette étude démontrent que NF! pourrait être le candidat
responsable de la répression de l'expression du gène a5 dans les lignées T97 et T98 tandis
que Sp3 pourrait être le candidat responsable de son expression dans les lignées TI08 et
TII5. Les résultats préliminaires obtenus suite à l'injection d'une quantité déterminée de
ces cellules cancéreuses dans des souris immunodéficientes permettent clairement
d'associer les lignées T97 et T98 à des phénotypes cancéreux beaucoup plus agressifs que
les lignées TI08 et TII5.
Finalement, afin de vérifier les nouvelles hypothèses formulées suite à cette étude,
l'injection de cellules cancéreuses ayant préalablement été transfectées de façon stable par
des lentivirus exprimant, par exemple, des shARN dans le but d'inhiber le FT NF! ou
encore, en sur-exprimant ce même FT selon le type cellulaire, pourrait permettre de
confirmer le rôle de NF! in vivo et démontrer s'il est indirectement responsable du
phénotype plus agressif chez certains types de cancers . . En dernier lieu, nos résultats
semblent appuyer l'hypothèse de départ qui voulait que les lignées qui sous-expriment la

82
sous-unité a5 à leur surface cellulaire, en l'occurrence T97 et T98, présentent un potentiel
plus agressif. Le suivi de l'expression de la sous-unité a5, dans les cellules dérivées des
tumeurs primaires de patients atteints de cette forme de cancer, pourrait se révéler être un
marqueur de dépistage intéressant afin d'identifier les patients à haut risque de développer
des métastases secondaires au foie dans le but d'assurer un meilleur suivi de ces individus.

83
4.2 Perspectives de recherche
Une somme de travail considérable demeure encore à réaliser afm de déterminer quel
facteur est responsable de l'expression aberrante de la sous-unité a4 dans la lignée
métastatique H79 ou afin d'évaluer si le FT NFI est le seul répresseur responsable de la
faible expression d' a5 dans les lignées T97 et T98 résultant ainsi d'une augmentation de
leur potentiel d'agressivité. La perspective la plus intéressante dans la suite logique du
projet est sans l'ombre d'un doute les analyses in vivo dans des souris immunosupprimées,
car elles permettront d'étudier le comportep1ent de ces cel~ules dans un contexte cellulaire
plus près de la réalité. Ces analyses, présentement en cours de réalisation, permettront donc
de confirmer in vivo les résultats obtenus in vitro, assurant ainsi la suite tant du projet a4
que du projet a5.
Il serait également intéressant d'établir un lien entre la recherche clinique et la recherche
fondamental~ chez les patients atteints d'un mélanome uvéa!. Ainsi, un suivi plus strict et
plus complet du patient à des intervalles réguliers serait non seulement bénéfique pour le
patient, mais fournirait également un complément d'informations très riche pour nos
futures études. En effet, s'il était possible de relier nos conclusions et nos observations, tel
que le profil cellulaire de la tumeur (épithélioïde ou fusiforme) et le profil d'expression des
intrégrines à la surface des cellules cancéreuses, aux données cliniques récoltées, c'est-à
dire l'âge au diagnostic de la tumeur uvéale, l'âge à l'apparition des métastases hépatiques
s'il y a lieu, l'âge et la cause du décès, la présence de boucles vas~ulogéniques etc., il en
ressortirait une quantité d'informations précieuses quant au choix du traitement à
privilégier et de la stratégie à adopter pour tenter d'augmenter le plus possible la qualité de
vie des patients atteints et éventuellement, permettre de sauver la vie d'un plus grand
nombres de ceux-ci.
Pour conclure, bien que ces deux parties du projet soient de natures complètement
différentes, elles ont en commun la compréhension de certaines complications qui peuvent
survenir dans les cas de mélano~e uvéa!. Connaître davantage les mécanismes sous-jacents
à l'agressivité tumorale et les processus qui modulent la progression du cancer vers la

84
métastasie donneront au médecin les outils nécessaires afin d'adapter les soins prodigués
selon le profil personnalisé ou plutôt « cellularisé » de chaque patient, ce qui augmentera
les chances de réussite du traitement et finalement, d'extrapoler les connaissances acquises
chez d'autres cancers à plus forte incidence.

85
Références bibliographiques
Adams, J. C. et F. M. Watt (1993). "Regulation of development and differentiation by the extracellular matrix." Development 117(4): 1183-98.
Ager, A. et M. J. Humphries (1991). "Integrin alpha 4 beta 1: its structure, ligand-binding specificity and role in lymphocyte-endothelial cell interactions." Chem Immunol 50: 55-74.
Ahmad, A. et I. R. Hart (1997). "Mechanisms of metastasis." Crit Rev Oncol Hematol 26(3): 163-73.
Akiyama, S. K., S. S. Yamada, et al. (1989). "Analysis of fibronectin receptor function with monoclonal antibodies: roles in cell adhesion, migration, matrix assembly, and cytoskeletal organization." J Cell Biol 109(2): 863-75.
Albelda, S. M. (1991). "Endothelial and epithelial cell adhesion molecules." Am J Respir Cell Mol Biol 4(3): 195-203.
Allavena, P., C. Paganin, et al. (1991). "Molecules and structures involved in the adhesion of natural killer cells to vascular endothelium." J Exp Med 173(2): 439-48.
Aplin, A. E., A. Howe, et al. (1998). "Signal transduction and signal modulation by cell adhesion receptors: the role of integrins, cadherins, immunoglobulin-cell adhesion molecules, and selectins." Pharmacol Rev 50(2): 197-263.
Arnaout, M. A., B. Mahalingam, et al. (2005). "Integrin structure, allostery, and bidirectional signaling." Annu Rev Cell Dev Biol 21: 381-410.
Audet, J. F., J. Y. Masson, et al. (1994). "Multiple regulatory elements control the basal promoter activity of the human alpha 4 integrin gene." DNA Cell Biol 13(11): 1071-85.
Bao, L., R. Pigott, et al. (1993). "Correlation of VLA-4 integrin expression with metastatic potential in various human tumour celllines." Differentiation 52(3): 239-46.
Bames, P. 1. (2006). "Transcription factors in airway diseases." Lab Invest 86(9): 867-72. Bechrakis, N. E. et W. R. Lee (1991). "Dedifferentiation potential of iris melanomas."
Fortschr OphthalmoI88(6): 651-6. Beckner, M. E., H. C. Krutzsch, et al. (1995). "Identification of a new immunoglobulin
superfamily protein expressed in blood vessels with a heparin-binding consensus sequence." Cancer Res 55(10): 2140-9.
Beliveau, A., M. Bérubé, et al. (2000). "Expression of integrin alpha5betal and MMPs associated with epithelioid morphology and malignancy of uveal melanoma." Invest Ophthalmol Vis Sci 41(8): 2363-72.
Bergeron, MJ et al. (1997). "Transcriptional regulation of the rat Pbly(ADP-ribose) polymerase gene by Sp1." Eur J Biochem 250(2):342-53.
Berman, A. E. et N. I. Kozlova (2000). "Integrins: structure and functions." Membr Cell , Biol 13(2): 207-44.
Bérubé, M., M. Talbot, et al. (2005). "Role of the extracellular matrix proteins in the resistance of SP6.5 uveal melanoma cells toward cisplatin." Int J Oncol 26(2): 405-13. '
Birkenmeier, T. M., 1. 1. McQuillan, et al. (1991). "The alpha 5 beta 1 fibronectin receptor. Characterization of the alpha 5 gene promoter." J Biol Chem 266(30): 20544-9.
Bock, E. (1991). "Cell-cell adhesion molecules." Biochem Soc Trans 19(4): 1076-80.

86
Boissy, R. E. (1988). "The melanocyte. Its structure, function, and subpopulations in skin, eyes, and hair." Dermatol Clin 6(2): 161-73.
Bopp, D., E. Jamet, et al. (1989). "Isolation of two tissue-specific Drosophila paired box genes, Pox mesa and Pox neuro." Embo J 8(11): 3447-57.
Bosman, F. T. et 1. Stamenkovic (2003). "Functional structure and composition of the extracellular matrix." J Pathol 200(4): 423-8.
Brem, R.B., S.G. Robbins et al. (1994). "Immunolocalization of integrins in the human retina." Invest Ophtalmol Vis Sci 35: 3466-74.
Brodt, P., L. Fallavollita, et al. (1997). "Liver endothelial E-selectin mediates carcinoma cell adhesion and promotes liver metastasis." Int J Cancer 71(4): 612-9.
Callaerts, P., G. Halder, et al. (1997). "PAX-6 in development and evolution." Annu Rev Neurosci 20: 483-532.
Carriere, C., S. Plaza, et al. (1993). "Characterization of quail Pax-6 (Pax-QNR) proteins expressed in the neuroretina." Mol Cell Biol 13(12): 7257-66.
Char, D. H. (1989). "Current treatments and trials in uveal melanoma." Oncology eWilliston Park) 3(9): 113-21; discussion 122, 124, 127-8.
Chauhan, B. K., N. A. Reed, et al. (2002). "Identification of genes downstream of Pax6 in the mouse lens using cDNA microarrays." J Biol Chem 277(13): 11539-48.
Corbi, A. L., U. B. Jensen, et al. (2000). "The alpha2 and alpha5 integrin genes: identification of transcription factors that regulate promoter activity in epidermal keratinocytes." FEBS Lett 474(2-3): 201-7.
Damato, B. (2006). "Treatment of primary intraocular melanoma." Expert Rev Anticancer Ther 6(4): 493-506.
Danen, E. H. et A. Sonnenberg (2003). "Integrins in regulation of tissue development and function." J PathoI201(4): 632-41.
Davidorf, F. H. et J. R. Lang (1975). "The natural history of malignant melanoma of the choroid: small vs large tumors." Trans Sect Ophthalmol Am Acad Ophthalmol Otolaryngol 79(2): OP31 0-20.
Davis, L. S., N. Oppenheimer-Marks, et al. (1990). "Fibronectin promotes proliferation of naive and memory T cells by signaling through both the VLA-4 and VLA-5 integrin molecules." J ImmunoI145(3): 785-93.
De Potter, P., C. L. Shields, et al. (1996). "New treatment modalities for uveal melanoma." Curr Opin Ophthalmol 7(3): 27-32.
de Vries, E., W. van Driel, et al. (1987). "Contactpoint analysis of the HeLa nuclear factor l recognition site reveals symmetrical binding at one side of the DNA helix." Embo J 6(1): 161-8.
Dedhar, S. et G. E. Hannigan (1996). "Integrin cytoplasmic interactions and bidirectional transmembrane signalling." Curr Opin Cell Biol 8(5): 657-69.
DeSimone, D. W. (1994). "Adhesion and matrix in vertebrate development." Curr Opin Cell Biol 6(5): 747-51.
Desprez, P. Y., C. Q. Lin, et al. (1998). "A novel pathway for mammary epithelial cell invasion induced by the helix-Ioop-helix protein Id-1." Mol Cell Biol 18(8): 4577-88.
Diener-West, M., B. S. Hawkins, et al. (1992). "A review of mortality from choroidal melanoma. II. A meta-analysis of 5-year mortality rates following enucleation, 1966 through 1988." Arch Ophthalmoll10(2): 245-50.

87
Dolin, P. J., A. J. Foss, et al. (1994). "Uveal Il)elanoma: is solar ultraviolet radiation a risk factor?" Ophthalmic Epidemiol1(1): 27-30.
Dufour, S., J. L. Duband, et al. (1988). "Attachment, spreading and locomotion of avian neural crest cells are mediated by multiple adhesion sites on fibronectin molecules. " Embo J 7(9): 2661-71.
Duncan, M. K., A. Cvekl, et al. (2000) .' "Truncated forms ofPax-6 disrupt lens morphology in transgenic mice." Invest Ophthalmol Vis Sci 41(2): 464-73.
Duncan, M. K., Z. Kozmik, et al. (2000). "Overexpression of PAX6(5a) in lens fiber cells results in cataract and upregulation of (alpha)5(beta)1 integrin expression." J Cell Sci 113 ( Pt 18): 3173-85.
Duncan, M.K., JI Haynes et al. (1998). "Dual roles for Pax-6: a transcriptional repressor of lens fiber cell-specific beta -crystallin genes." Mol Cell Biol 18(9):5579-86.
Dynan, W. S. et R. Tjian (1983). "The promoter-specific transcription factor Sp 1 binds to upstream sequences in the SV40 ear1y promoter." Ce1l35(1): 79-87.
Elner, S. G. et V. M. Elner (1996). "The integrin superfamily and the eye." Invest Ophthalmol Vis Sci 37(5): 696-701.
Epstein, J. A., T. Glaser, et al. (1994). "Two independent and interactive DNA-binding subdomains of the Pax6 paired domain are regulated by alternative splicing." Genes Dev 8(17): 2022-34.
Eskelin, S., S. Pyrhonen, et al. (2000). "Tumor doubling times in metastatic malignant melanoma of the uvea: tumor progression before and after treatment." Ophthalmology 107(8): 1443-9.
Ferguson, T. A., H. Mizutani, et al. (1991). "Two integrin-binding peptides abrogate T cellmediated immune responses in vivo." Proc Nad Acad Sci USA 88(18): 8072-6.
Fidler,I. J. (1991). "Cancer metastasis." Br Med Bull 47(1): 157-77. Flocks, M., J. H. Gerende, et al. (1955). "The size and shape of malignant melanomas of
the choroid and ciliary body in relation to prognosis and histologic characteristics; a statistical study of 210 tumors." Trans Am Acad Ophthalmol Otolaryngol 59(6): 740-58.
Fraunfelder, F. T., F. W. Boozman, et al. (1977). "No-touch technique for intraocular malignant melanomas." Arch OphthalmoI95(9): 1616-20.
Frenette, P. S. èt D. D. Wagner (1996). "Adhesion molecules--Part 1." N Engl J Med 334(23): 1526-9.
Frisch, S. M. et E. Ruoslahti (1997). "Integrins and anoikis." CUIT Opin Cell Biol 9(5): 701-6.
Gailit, 1., M. Pierschbacher, et al. (1993). "Expression of functional alpha 4 beta 1 integrin by human dermal fibroblasts." J Invest Dermatol100(3): 323-8.
Gaudreault, M. et al. (2007). "Laminin reduces expression of the human alpha6 integrin subunit gene by altering the level of the transcription factors Sp 1 and Sp3." Invest Ophtalmol Vis Sci 48(8):3490-505.
Giancotti, F. G. (2003). "A structural view of integrin activation and signaling." Dev Cell 4(2): 149-51.
Giancotti, F. G. et F. Mainiero (1994). "Integrin-mediated adhesion and signaling in tumorigenesis." Biochim Biophys Acta 1198(1): 47-64.
Giancotti, F. G. et E. Ruoslahti (1999). "Integrin signaling." Science 285(5430): 1028-32.

88
Gingras, M.E., et al. (2008). "The Transcriptional Activity Directed by the Human alpha5 Integrin Gene Promoter is Detennined through an Spl/NFI/AP-l Composite Element." Invest Ophtalmol Vis Sci (en préparation).
Gingras, M. E., K. Larouche, et al. (2003). "Regulation of the integrin subunit alpha5 gene promoter by the transcription factors Sp I/Sp3 is influenced by the cell density in rabbit corneal epithelial cells." Invest Ophthalmol Vis Sci 44(9): 3742-55.
Gonder, J. R., J. A. Shields, et al. (1982). "Uveal malignant melanoma associated with ocular and oculodennal melanocytosis." Ophthalmology 89(8): 953-60.
Gong, 1., D. Wang, et al. (1997). "Role of alpha 5 beta 1 integrin in determining malignant properties of colon carcinoma cells." Cell Growth Differ 8(1): 83-90.
Gronostajski, R. M. (2000). "Roles of the NFI/CTF gene family in transcription and development." Gene 249(1-2): 31-45.
Guenel, P., L. Laforest, et al. (2001). "Occupational risk factors, ultraviolet radiation, and ocular melanoma: a case-control study in France." Cancer Causes Control 12(5): 451-9.
Gumbiner, B. M. et P. D. McCrea (1993). "Catenins as mediators of the cytoplasmic functions of cadherins." J Cell Sci Suppl 17: 155-8.
Hakulinen, 'T., L. Teppo, et ~l. (1978)~ "Cancer of the eye, a review of trends and differentials." World Health Stat 0 31(2): 143-58.
Hanahan, D. et R. A. Weinberg (2000). "The hallmarks of cancer." Cell100(1): 57-70. Hatta, K., S. Takagi, et al. (1987). "Spatial and temporal expression pattern of N-cadherin
cell adhesion molecules correlated with morphogenetic processes of chicken embryos." Dev Biol 120(1): 215-27.
Hemler, M. E., M. J. Elices, et al. (1990). "Structure of the integrin VLA-4 and its cell-cell and cell-matrix adhesion functions." Immunol Rev 114: 45-65.
Hotchin, N. A. et A. Hall (1995). "The assembly of integrin adhesion complexes requires both extracellular matrix and intracellular rho/rac GTPases." J Cell Biol 131(6 Pt 2): 1857-65.
Humphries, M. J., P. A. McEwan, et al. (2003). "Integrin structure: heady advances in ligand binding, but activation still makes the knees wobble." Trends Biochem Sci 28(6): 313-20.
HungerJord, J. L. (1995). "Management of ocular melanoma." Br Med Bull 51(3): 694-716. Hynes, R. O. (1992). "Integrins: versatility, modulation, and signaling in cell adhesion."
CeIl69(1): 11-25. Hynes, R. O. (2002). "Integrins: bidirectional, allosteric signaling machines." Cell 110(6):
673-87. lozzo, R. V. (1998). "Matrix proteo.glycans: from molecular design to cellular function."
Annu Rev Biochem 67: 609-52. Jaspers, M., R. R. Wu, et al. (1995). "Localization of alpha 4m integrin at sites of
mesenchyme condensation during embryonic mouse development." Differentiation 59(2): 79-86.
Jockusch, B. M., P. Bubeck, et al. (1995). "The molecular architecture of focal adhesions." Annu Rev Cell Dev Biol 11: 379-416.
Johnson, J. P., B. G. Stade, et al. (1989). "De nova expression of intercellular-adhesion mole cule 1 in melanoma correlates with increased risk of metastasis." Proc Nad Acad Sci USA 86(2): 641-4.

89
Joseph-Silverstein, J. et R. L. Silverstein (1998). "Cell adhesion molecules: an overview." Cancer Invest 16(3): 176-82.
Journee-de Korver, J. G. et J. E. Keunen (2002). "Thermotherapy in the management of choroidal melanoma." Prog Retin Eye Res 21(3): 303-17.
Juliano, R. L. et S. Haskill (1993). "Signal transduction from the extracellular matrix." l Cell Biol 120(3): 577-85.
Juliano, R. L., SK. AlaHari et al. (2002. "Biological aspects of signal transduction by cell adhesion receptors." Int Rev Cytol 220: 145-84.
Kassner, P. D., J. Teixido, et al. (1992). "Analyses ofVLA-4 structure and function." Adv Exp Med Biol 323: 163-70.
Kersten, R. C., D. T. Tse, et al. (1985). "Iris melanoma. Nevus or malignancy?" SUry OphthalmoI29(6): 423-33.
Khatib, A. M., L. Fallavollita, et al. (2002). "Inhibition of hepatic endothelial E-selectin expression by C-raf antisense oligonucleotides blocks colorectal carcinoma liver metastasis." Cancer Res 62(19): 5393-8.
Kim, J. P., K. Zhang, et al. (1992). "Mechanism of human keratinocyte migration on fibronectin: unique roles of RGD site and integrins." J Cell PhysioI151(3): 443-50.
Kincaid, M. C. (1998). "Uveal Melanoma." Cancer Control 5(4): 299-309. Kita, D., T. Takino, et al. (2001). "Expression of dominant-negative form of Ets-l
suppresses fibronectin-stimulated cell adhesion and migration through downregulation of integrin alphaS expression in U251 glioma cell line." Cancer Res 61(21): 7985-91.
Kozmik, Z., T. Czerny, et al. (1997). "Alternatively spliced insertions in the paired domain restrict the DNA sequence specificity ofPax6 and Pax8." Embo J 16(22): 6793-803.
Kuphal, S., R. Bauer, et al. (2005). "Integrin signaling in malignant melanoma." Cancer Metastasis Rev 24(2): 195-222.
Laffon, A., R. Garcia-Vicuna, et al. (1991). "Upregulated expression and function ofVLA-4 fibronectin receptors on human activated T cells in rheumatoid arthritis." J Clin Invest 88(2): 546-52.
Larouche, K., S. Leclerc, et al. (1996). "Multiple nuclear regulatory proteins bind a single cis-acting promoter element to control basal transcription of the human alpha 4 integrin gene in comeal epithelial cells." DNA Cell Biol 15(9): 779-92.
Larouche, K., S. Leclerc, et al. (2000). "Expression of the alpha 5 integrin subunit gene promoter is positively regulated by the extracellular matrix component fibronectin through the transcription factor Sp 1 in corneal epithelial cells in vitro." J Biol Chem 275(50): 39182-92.
Larouche, N., K. Larouche, et al. (1998). "Transcriptional regulation of the alpha 4 integrin subunit gene in the metastatic spread of uveal melanoma." Anticancer Res 18(5A): 3539-47.
Lauweryns, B., J. J. van den Oord, et al. (1991). "Distribution of very late activation integrins in the human comea. An immunohistochemical study using monoclonal antibodies." Invest Ophthalmol Vis Sci 32(7): 2079-85.
Lee, J. W. et R. Juliano "(2004). "Mitogenic signal transduction by integrin- and growth factor receptor-mediated pathways." Mol Cells 17(2): 188-202.
Lefebvre, T., N. Planque, et al. (2002). "O-glycosylation of the nuclear forms of Pax-6 products in ·quail neuroretina cells." J Cell Biochem 85(1): 208-18.

90
Leyvraz, S., V. Spataro, et al. (1997). "Treatment of ocular melanoma metastatic to the liver by hepatic arterial chemotherapy." J Clin OncoI15(7): 2589-95.
Liddington, R. C. et M. H. Ginsberg (2002). "Integrin activation takes shape." J Cell Biol 158(5): 833-9.
Lobb, R. R., W. M. Abraham, et al. (1996). "Pathophysiologic role of alpha 4 integrins in the lung." Ann N Y Acad Sci 796: 113-23.
Loeser, R. F. (1994). "Modulation of integrin-mediated attachment of chondrocytes to extracellular matrix proteins by cations, retinoic acid, and transforming growth factor beta~" Exp Cell Res 211(1): 17-23.
Lopez-Rodriguez, C., L. Botella, et al. (1997). "CCAA T -enhancer-binding proteins (C/EBP) regulate the tissue specific activity of the CD Il c integrin gene promoter through functional interactions with SpI proteins." J Biol Chem 272(46): 29120-6.
Lutz, 1. M., 1. Cree, et al. (2005). "Occupational risks for uveal melanoma results from a case-control study in nine European countries." Cancer Causes Control 16(4): 437-47.
Macdonald, R. et S. W. Wilson (1996). "Pax proteins and eye development." CUIT Opin NeurobioI6(1): 49-56.
Maio, M., A. Pinto, et al. (1990). "DifferentiaI expression of CD54/intercellular adhesion molecule-l in myeloid leukemias and in lymphoproliferative disorders." Blood 76(4): 783-90.
Mareel,· M. et A. Leroy (2003). "Clinical, cellular, and molecular aspects of cancer invasion." Physiol Rev 83(2): 337-76.
Marshall, 1. F., D. C. Rutherford, et al. (1995). "Alpha v beta 1 is a receptor for vitronectin and fibrinogen, and acts with alpha 5 beta 1 to mediate spreading on fibronectin." l Cell Sci 108 ( Pt 3): 1227-38.
Mathers, P. H. et M. Jarnrich (2000). "Regulation of eye formation by the Rx and pax6 homeobox genes." Cell Mol Life Sci 57(2): 186-94.
McCartney, A. C. (1995). "Pathology of ocular melanomas." Br Med Bull 51(3): 678-93. McLean,L W., D~ Berd, et al. (1990). "A randomized study of methanol-extraction residue
of bacille Calmette-Guerin as postsurgical adjuvant therapy of uveal melanoma." Am J Ophthalmolll0(5): 522-6.
McLean, M. J., W. D. Foster, et al. (1977). "Prognostic factors in small malignant melanomas of choroid and ciliary body." Arch Ophthalmol 95( 1): 48-58.
Meenan, J., J. Spaans, et al. (1997). "Altered expression of alpha 4 beta 7; a gut homing integrin, by circulating and mucosal T cells in colonic mucosal inflammation." Gut 40(2): 241-6.
Mikkola, L, J. A. Bruun, et al. (1999). "Phosphorylation of the transactivation domain of Pax6 by extracellular signal-regulated kinase and p38 mitogen-activated protein kinase." J Biol Chem 274(21): 15115-26.
Mizejewski, G. J. (1999). "Role of integrins in cancer: survey of expressionpattems." Proc Soc Exp Biol Med 222(2): 124-38.
Moan, J., A. C. Porojnicu, et al. (2008). "Ultraviolet radiation and malignant melanoma." Adv Exp Med Biol 624: 104-16.
Moles, J. P. et F. M. Watt (1997). "The epidermal stem cell compartment: variation in expression levels of E-cadherin and catenins within the basal layer of human . epidermis." J Histochem Cytochem 45(6): 867-74.

91
Nobes, C. D. et A. Hall (1995). "Rho, rac, anq cdc42 GTPases regulate the assembly of multimolecular focal complexes associated with actin stress fibers, lamellipodia, and filopodia." Ce1l81(1): 53-62.
Nojima, Y., M. J. Humphries, et al. (1990). "VLA-4 mediates CD3-dependent CD4+ T cell activation via the CS 1 altematively spliced domain of fibronectÏn." J Exp Med 172(4): 1185-92.
O'Toole, T. E., Y. Katagiri, et al. (1994). "Integrin cytoplasmic domains mediate inside-out signal transduction." J Cell Biol 124(6): 1047-59.
Oguariri, R. M., T. W. Brann, et al. (2007). "Hydroxyurea and interleukin-6 synergistically reactivate HIV -1 replication in a latently infected promonocytic cell line via SP1/SP3 transcription factors." J Biol Chem 282(6): 3594-604. .
Ohene-Abuakwa, Y. et M. Pignatelli (2000). "Adhesion Molecules as Diagnostic Toois in Tumor Pathology." Int J Surg PathoI8(3): 191-200.
Olsen, B. R. (1995). "New insights into the function of collagens from genetic analysis." Curr Ooin Cell Biol 7(5): 720-7.
Oosterhuis, J. A., L. N. Went, et al. (1982). "Primary choroidal and cutaneous melanomas, bilateral choroidal melanomas, and familial occurrence of melanomas." Br J Ophthalmo166( 4): 230-3.
Park, C. C., M. J. Bissell, et al. (2000). "The influence of the microenvironment on the malignant phenotype." Mol Med Today 6(8): 324-9.
i Passlick, B., J. R. Izbicki, et al. (1994). "Expression of major histocompatibility class 1 and class II antigens and intercellular adhesion molecule-Ion operable non-small cell lung carcinomas: frequency and prognostic significance." Eur J Cancer 30A(3): 376-81.
Pennini ME., Y. Liu et al. (2007). "CCAAT/enhancer-binding protein beta and delta binding to CIIT A promoters is associated with the inhibition of CIIT A expression in response to Mycobacteriu~ tuberculosis 19-kDa lipoprotein." J ImmunoI179(10): 6910-8.
Petruzzelli, . L., M. Takami, et al. (1999). "Structure and function of cell adhesion molecules." Am J Med 106(4): 467-76.
Philipsen, S. et G. Suske (1999). "A tale of three fingers: the family of mammalian Sp/XKLF transcription factors." Nucleic Acids Res 27(15): 2991-3000.
Planque, N., L. Leconte, et al. (2001). "Specific Pax-6/microphthalmia transcription factor interactions involve their DNA-binding domains and inhibit transcriptional properties ofboth proteins." J Biol Chem 276(31): 29330-7.
Plow, E. F., T. A. Haas, et al. (2000). "Ligand binding to integrins." J Biol Chem 275(29): 21785-8.
Pothier, F., M. Ouellet, et al. (1992). "An improved CAT assay for promoter analysis in either transgenic mice or tissue culture cells." DNA Cell Biol 11(1): 83-90.
Przybylo, M., E. Pochec, et al. (2008). "Betal-6 branching of cell surface glycoproteins may contribute to uveal melanoma progression by up-regulating cell motility." Mol Vis 14: 625-36.
Pulai, 1. I., M. Del Carlo, Jr., et al. (2002). "The alpha5betal integrin provides matrix survival signaIs for normal and osteoarthritic hum an articular chondrocytes in vitro." Arthritis Rheum 46(6): 1528-35.
Robbins, S. G., R. B. Brem, et al. (1994). "Immunolocalization of integrins in proliferative retinal membranes." Invest Ophthalmol Vis Sci 35(9): 3475-85.

92
Robinson, E. E., K. M. Zazzali, et al. (2003). "Alpha5beta1 integrin mediates strong tissue cohesion." J Cell Sci 116(Pt 2): 377-86.
Romer, L. H., K. G. Birukov, et al. (20.06). "Focal adhesions: paradigm for a signaling nexus." Circ Res 98(5): 606-16.
Rosen, G. D., 1. L. Barks, et al. (1994). "An intricate arrangement of binding sites for -the Ets family of transcription factors regulates activity of the alpha 4 integrin gene promoter." 1 Biol Chem 269(22): 15652-60.
Rosen, G. D., T. M. Birkenmeier, et al. (1991). "Characterization of the alpha 4 integrin gene promoter." Proc Nat! Acad Sci USA 88(10): 4094-8.
Roskelley, C. D., A. Srebrow, et al. (1995). "A hierarchy of ECM-mediated signalling regulates tissue-specific gene expression." Curr Opin Cell Biol 7(5): 736-47.
Rosskothen, N., 1. Hirschmuller-Ohmes, et al. (2008). "AP-1 activity rises by stimulationdependent c-Fos expression in auditory neurons." Neuroreport 19(11): 1091-3.
Roy, R. J., P. Gosselin, et al. (1991). "A short protocol for micro-purification of nuclear ' proteins from whole animal tissue." Biotechnigues 11(6): 770-7.
Sanders, R. 1., F. Mainiero, et al. (1998). "The role of integrins in tumorigenesis and metastasis." Cancer Invest 16(5): 329-44.
Schaller, M. D. et 1. T. Parsons (1994). "Focal adhesion kinase and associated proteins." CUIT Opin Cell Biol 6(5): 705-10.
Schatzmann, F., R. Marlow, et al. (2003). "lntegrin signaling and mammary cell function." J Mammary Gland Biol Neoplasia 8(4): 395-408.
Schneider, R., 1. Gander, et al. (1986). "A sensitive and rapid gel retenti on assay for nuclear factor 1 and other DNA-binding proteins in crude nuclear extracts." Nucleic Acids Res 14(3): 1303-17.
Schraermeyer, U., K. Addicks, et al. (1998). "Capillaries are present in Bruch's membrane at the ora serrata in the human eye." Invest Ophthalmol Vis Sci 39(7): 1076-84.
Scohy, S., P. Gabant, et al. (2000). "Identification of KLF13 and KLF14 (SP6), novel members of the SP/XKLF transcription factor family." Ge~omics 70(1): 93-101.
Scotto, 1., J. F. Fraumeni, Ir., et al. (1976). "Melanomas of the eye and other noncutaneous sites: epidemiologic aspects." J Nat! Cancer Inst 56(3): 489-91.
Seftor, R. E., E. A. Seftor, et al. (1999). "Molecular role(s) for integrins in human melanoma invasion." Cancer Metastasis Rev 18(3): 359-75.
Seregard, S. (1996). "Posterior uveal melanoma. The Swedish perspective." Acta Ophthalmol Scand 74(4): 315-29.
Seregard, S. (1999). "Long-term survival after ruthenium plaque radiotherapy for uveal melanoma. A meta-analysis of studies including 1,066 patients." Acta Ophthalmol Scand 77(4): 414-7.
Shammas, H. F. et F. C. Blodi (1977). "Prognostic factors in choroidal and ciliary , body melanomas." Arch OphthalmoI95(1): 63-9.
Sherry, D.M., P.A. Proske (2001). "Localization 'of alpha integrin subunits in the neural retina of the tiger salamander." Graefes Arch Clin Exp Ophtalmol239: 278-87.
Shields, C. L., 1. A. Shields, et al. (1998). "Transpupillary thermotherapy for choroidal melanoma: tumor control ,and visual results in' 100 consecutive cases." Ophthalmology 105(4): 581-90.
Shields, J. A., W. H. Annesley, Jr., et al. (1977). "Necrotic melanocytoma of iris with secondary glaucoma." Am J Ophthalmo184(6): 826-9.

93
Shields, J. A., C. L. Shields, et al. (1991). "Management of posterior uveal melanoma." SUry OphthalmoI36(3): 161-95.
Shimaoka, M. et T. A. Springer (2003). "Therapeutic antagonists and conformational regulation of integrin function. " Nat Rev Drug Discov 2(9): 703-16.
Shimaoka, M., J. Takagi, et al. (2002). "Conformational regulation of integrin structure and function." Annu Rev Biophys Biomol Struct 31: 485-516.
Shimizu, Y. , G. A. van Seventer, et al. (1990). "Costimulation of proliferative responses of resting CD4+ T cel~s by the interaction of VLA-4 and VLA-5 with fibronectin or VLA-6 with laminin." J ImmunoI145(1): 59-67.
Sincock, P. M., G. Mayrhofer, et al. (1997). "Localization of the transmembrane 4 superfamily (TM4SF) member PETA-3 (CD151) in normal human tissues: comparison with CD9, CD63, and alpha5beta1 integrin." J Histochem Cytochem 45(4): 515-25.
Singh, A. D. (2001). "Uveal melanoma: implications of tumor doubling time." Ophthalmology 108(5): 829-31.
Singh, A. D. et A. Topham (2003). "Incidence of uveal melanoma in the United States: 1973-1997." Ophthalmology 110(5): 956-61.
Sisley, K., D. W. Cottam, et al. (1992). "Non-random abnormalities of chromosomes 3, 6, and 8 associated with ·posterior uveal melanoma." Genes Chromosomes Cancer 5(3): 197-200.
Sorensen, F. B., 1. W. Gamel, et al. (1993). "Prognostic value of nucleolar size and size pleomorphism in choroidal mela~omas." Apmis 101(5): 358-68.
Stallmach, A., B. von Lampe, et al. (1992). "Diminished expression of integrin adhesion molecules on human colonic epithelial cells during the benign to malign tumour transformation." Gut 33(3): 342-6.
Stepp, M. A., S. SpuIT-Michaud, et al. (1993). "Integrins in the wounded and unwounded stratified squamous epithelium of the comea." Invest Ophthalmol Vis Sci 34(5): 1829-44.
Strachan, T. et A. P. Read (1994). "PAX genes." CUIT Opin Genet Dev 4(3): 427-38. Streuli, C. (1999). "Extracellular matrix remodelling and cellular differentiation." CUIT
Opin Cell Biol 11(5): 634-40. Striz, 1. et U. Costabel (1992). "The role of integrins in the immune response.!1 Sarcoidosis
9(2): 88-94. Stupack, D. G. et D. A. Cheresh (2002). "Get a ligand, get a life: integrins, signaling and
cell survival." J Cell Sci 115(Pt 19): 3729-38. Symington, B. E. (1995). "Growth signalling through the alpha 5 beta 1 fibronectin
receptor." Biochem Biophys Res Commun 208(1): 126-34. Tsonis, P. A. et E. J. Fuentes (2006). "Focus on molecules: Pax-6, the eye master." Exp Eye
Res 83(2): 233-4. Urbich, C., E. Dembach, et al. (2002). "Shear stress-induced endothelial cell migration
involves integrin signaling via the fibronectin receptor subunits alpha( 5) and beta(l)." Arterioscler Thromb Vasc Biol 22(1): 69-75.
van der Flier, A. et A. Sonnenberg (2001). "Function and interactions of integrins." Cell Tissue Res 305(3): 285-98.
van Heyningen, V. et K. A. Williamson (2002). "PAX6 in sensory development." Hum Mol . Genet 11(10): 1161-7.

94
Vidal, 1. L. , F. Bacin, et al. (1995). "["Melanoma 92". epidemiological study of uveal melanoma in France J." J Fr OphtalmoI18(8-9): 520-8.
Vigneault, F. , K. Zaniolo, et al. (2007). "Control of integrin genes expression in the eye." Prog Retin Eye Res 26(2): 99-161.
Walsh, L. 1. et G. F. Murphy (1992). "Role of adhesion molecules in cutaneous inflammation and neoplasia." J Cutan PathoI19(3): 161-71.
Wayner, E. A., A. Garcia-Pardo, et al. (1989). "Identification and characterization of the T lymphocyte adhesion receptor for an altem"ative ceU attachment domain (CS-1) in plasma fibronectin." J CeU Biol 109(3): 1321-30.
Wehrle-HaUer, B. et B. A. Imhof (2003). "Actin, microtubules and focal adhesion dynamics during cell migration." Int J Biochem Cell Biol 35(1): 39-50.
Wiesner, S. , K. R. Legate, et al. (2005). "Integrin-actin interactions." Cell Mol Life Sci 62(10): 1081-99.
Williams, D. A. , M. Rios, et al. (1991). "Fibronectin and VLA-4 in haematopoietic stem cell-microenvironment interactions." Nature 352(6334): 438-41.
Wolff, J. M., R. N. Stephenson, et al. (1995). "Levels of circulating intercellular adhesion molecule-1 in patients with metastatic cancer of the prostate and benign prostatic hyperplasia." Eur J Cancer 31A(3): 339-41.
Woodward, J. K. , 1. G. Rennie, et al. (2005). "Invasive and noninvasive uveal melanomas have different adhesive properties." Eye 19(3): 342-8.
Xu, H. E., M. A. Rould, et al. (1999). "Crystal structure of the hum an Pax6 paired domainDNA complex reveals specific roles for the linker region and carboxy-terminal subdomain in DNA binding." Genes Dev 13(10): 1263-75.
Xu, K. et H. K. Shu (2007). "EGFR activation results in enhanced cyclooxygenase-2 expression through p38 mitogen-activated protein kinase-dependent activation of the Sp1/Sp3 transcription factors in hum an gliomas." Cancer Res 67(13): 6121-9.
Xu, R., P. Zhang, et al. (2007). "SpI and Sp3 regulate bas,}! transcription of the survivin gene." Biochem Biophys Res Commun 356(1): 286-92.
Yan, Q., W. B. Liu, et al. (2007). "Prote in phosphatase-1 modulates the function of Pax-6, a transcription factor controlling brain and eye development." J Biol Chem 282(19): 13954-65.
Yap, A. S., C. M. Niessen, et al. (1998). "The juxtamembrane region of the cadherin cytoplasmic tail supports lateral clustering, adhesive strengthening, and interaction with p120ctn." J Cell Biol 141(3): 779-89.
Young, RW. (1976). "Visual cells and the concept ofrenewal." Invest OphtalmoI15:700-25.
Zagris, N. (2001). "Extracellular matrix in development of the early embryo." Micron 32(4): 427-38.
Zaniolo, K., S. Leclerc, et al. (2004). "Expression of the alpha4 integrin subunit gene promo ter is modulated by the transcription factor Pax-6 in comeal epithelial ceUs." Invest Ophthalmol Vis Sci 45(6): 1692-704.
Zimmerman, L. E., 1. W. McLean, et al. (1978). "Does enucleation of the eye containing a malignant melanoma prevent or accelerate the dissemination of tumour cells." Br J OphthalmoI62(6): 420-5.
Zhang, Z.H., S. Vekemans et al. (1991). "The gene for the alpha4 subunit of the VLA-4 integrin maps to chromosome 2Q31-32. Blood 78: 2396-99.
Recommended

















