Universidade Estadual de Campinas
Instituto de Física “Gleb Wataghin”
Departamento de Raios Cósmicos e Cronologia
Grupo de Neurofísica
Mapeamento do Córtex Visual Humano Através de uma Abordagem Multimodal Integrando
Eletroencefalografia e Espectroscopia Óptica na Região do Infravermelho Próximo
Enrique Porfirio Uceda Otero
Tese apresentada ao Instituto de Física da Universidade Estadual de Campinas, para a obtenção do título de Doutor em Física.
Orientador: Prof. Dr. Roberto J. M. Covolan
Campinas – SP
2009

ii

iii

iv
“La vida es uma pequeña rama del bosque inmenso de los siglos”

v
Dedicatórias:
A mi madre, Graciela: porque soy todo por ti. Te amo madre mia, Dios ilumine siempre tu vida. A mi Hermano, Rolando: por ser el mejor de todos, que Dios te ayude siempre hermano. A mi hijo, Enrique Jair: que estás aún en los brazos de tu madre, estás creciendo hijo, Dios te cuidará hijo.

vi
Agradecimentos
Gostaria de agradecer às inúmeras pessoas que me ajudaram no decorrer do
programa de doutorado, em especial:
Ao meu orientador, o Prof. Dr. Roberto José Maria Covolan, entre outras coisas pela
orientação principalmente nas etapas mais difíceis deste trabalho.
À minha mãe Graciela, e os meus irmãos pela força constante.
À minha esposa Zulema Maria, pela paciência, e carinho.
À Dra. Helka F. B. Ozelo, pela ajuda toda e pela amizade.
Ao mestre Elvis Lira da Silva, pela força nos momentos difíceis. Caro amigo, um
muito obrigado para você.
Ao mestre Carlos Alessandro da Silva dos Anjos, pelo direcionamento nas análises
ópticas, pela ajuda no árduo trabalho "braçal", pela amizade.
Ao mestre Edgard P. M. Amorim, pela força, pela ajuda e pela amizade. Caro amigo,
um muito obrigado para você.
Aos colegas do grupo Andréa, Márcia e Rickson.

vii
A todos os meus colegas de grupo, que direta ou indiretamente, participaram deste
trabalho.
Aos amigos e colegas do Instituto de Física do Prédio D, e a toda a galera da
Colômbia, em especial ao Hernán, Diego, Dario, e a todos aqueles de um modo o outro
colaboraram direta o indiretamente com o desenvolvimento deste trabalho.
À secretária da pós-graduação do IFGW, em especial a Sra. Maria Ignez S. R.
Mokarzel.
À secretária do Instituto de Física “Gleb Wataghin”, a Sra. Lucia Regina Rio, pela
força, e pela amizade.
À UNICAMP e a todos os funcionários do IFGW.
À Capes e à FAPESP, pelo apoio financeiro.
A todos, meus sinceros agradecimentos.

viii
Resumo
A ativação cerebral envolve um complexo arranjo de processos neuronais,
metabólicos e vasculares, que se estende do nível molecular e celular ao nível de extensas
zonas corticais. O processo de disparo neuronal requer a restauração de gradientes iônicos e
a reciclagem de neurotransmissores, o que implica em um custo energético suprido na
forma de trifosfato de adenosina (ATP). A principal via de síntese do ATP é a via aeróbica
e se dá pelo metabolismo oxidativo da glicose. Para tanto, o metabolismo cerebral depende
de um constante suprimento de glicose e oxigênio, que é mantido pela circulação sanguínea
através de uma complexa rede de vasos, que compõe o sistema vascular cerebral.
A regulação desse complexo sistema neurovascular-metabólico é objeto de intensa
investigação e está no centro do trabalho aqui apresentado, que visa o mapeamento do
córtex visual através de uma abordagem multimodal envolvendo eletroencefalografia
(EEG) e espectroscopia óptica na região do infravermelho próximo (NIRS - near infrared
spectroscopy). O objetivo central deste trabalho foi o desenvolvimento de uma metodologia
que permitisse a realização de medidas simultâneas da atividade neuronal, via EEG, e das
alterações vasculares associadas a essas, via NIRS. Através desta técnica foi estudado o
córtex visual de indivíduos adultos saudáveis, através da apresentação de estímulos
modulados em frequência e em contraste. Esses experimentos foram realizados utilizando
visão tanto binocular quanto monocular, sendo esta última para cada olho separadamente.
Os estudos de EEG, realizados com eletrodos posicionados em O1 e O2, permitiram
registrar claras alterações dos ritmos cerebrais alfa e beta em correlação com as variações
de contraste e frequência do estímulo visual. As medidas ópticas, realizadas através do
escalpo com optodos colocados estímulo dos eletrodos, permitiram registrar respostas
hemodinâmicas bastante enfáticas, que mostraram alguma variabilidade em correlação com
o contraste e a frequência dos estímulos visuais utilizados. Os resultados obtidos
demonstram a viabilidade de se estudar o acoplamento neurovascular-metabólico em
humanos através de uma abordagem multimodal não-invasiva, utilizando-se um sistema
conjugado NIRS-EEG.

ix
Abstract
Brain activation involves a complex arrangement of neuronal, metabolic and
vascular processes, which goes from molecular and cellular level to the level of extensive
cortical and subcortical zones. The process of neuronal firing requires the restoration of
ionic gradients and neurotransmitter recycling, which implies supplying energy in the form
of Adenosine Triphosphate (ATP). ATP synthesis follows mainly the aerobic way and
occurs by the oxidative metabolism of glucose. Therefore, the cerebral metabolism depends
on a constant supply of glucose and oxygen, which is maintained by the blood circulation
through the complex networks of blood vessels that compose the cerebral vascular system.
The regulation of this complex neurovascular-metabolic system is object of intense
investigation and is in the center of the work presented here, that aims to investigate the
human visual system through a multimodal boarding integrating electroencephalography
(EEG) and near infrared spectroscopy (NIRS). The central objective of this work was the
development of a methodology that would allow simultaneous measurements of the
neuronal activity, via EEG, and of the vascular changes associated to these, via NIRS.
Through this technique, we studied the visual cortex of healthy adults, through the
presentation of stimuli modulated alternatively in frequency and contrast. These
experiments were performed for both binocular and monocular vision, being the latter for
both eyes. The studies of EEG were performed with electrodes positioned in O1 and O2
and allowed to register clear alterations of alpha and beta brain rhythms in correlation with
the contrast and frequency variations of the visual stimulus. The optical measurements were
performed through the skull with optodes placed around the electrodes and allowed to
record hemodynamic responses whose variability was also related to the contrast and
frequency modulations of the visual stimuli. The obtained results demonstrate the
feasibility of applying a conjugated NIRS-EEG system as a multimodal approach to study
the neurovascular-metabolic coupling in humans.

x

xi
Sumário
Agradecimentos ................................................................................................................................................. vi Abstract ............................................................................................................................................................. ix
Sumário ............................................................................................................................................................. xi Capítulo 1 ........................................................................................................................................................... 1
O Cérebro e o Sistema Visual Humano .............................................................................................................. 1
1.1 O Cérebro Humano .................................................................................................................................... 1
1.1.1 Anatomia Cerebral ........................................................................................................................ 2
1.1.2 Neurônios ..................................................................................................................................... 5
1.1.3 Córtex cerebral ............................................................................................................................. 7
1.2 Funções cerebrais ...................................................................................................................................... 9
1.2.1 Áreas motoras ..............................................................................................................................11
1.2.2 Áreas sensitivas ...........................................................................................................................11
1.3 O Sistema Visual Humano .................................................................................................................... 12
1.3.1 O Olho .........................................................................................................................................13
1.3.2 A retina ........................................................................................................................................14
1.3.3 Campos receptivos retinais ..........................................................................................................15
1.3.4 Trajetória das Vias Visuais ..........................................................................................................17
Capítulo 2 ..........................................................................................................................................................19
Eletroencefalografia .............................................................................................................................................. 19
2.1 Introdução ................................................................................................................................................. 19
2.2 O Eletroencefalograma .......................................................................................................................... 20
2.2.1 Sinais elétricos do cérebro ...........................................................................................................20
2.2.2 Fontes neurais do sinal EEG ........................................................................................................21
2.2.3 A natureza do EEG ......................................................................................................................23
2.2.4 Potenciais relacionados a eventos (ERP) .....................................................................................25
2.2.5 Resolução espacial e temporal do EEG .......................................................................................26
2.2.7 Aquisição de dados do EEG ........................................................................................................29
2.2.8 Propriedades do EEG ...................................................................................................................31
2.2.9 Aplicações do EEG ......................................................................................................................32
2.3 O equipamento de EEG .......................................................................................................................... 34
2.3.1 Eletrodos ......................................................................................................................................34
2.3.2 Amplificadores ............................................................................................................................34
2.3.3 Configuração dos eletrodos .........................................................................................................35
2.4 Problemas do EEG ................................................................................................................................... 36
Capítulo 3 ..........................................................................................................................................................37
Espectroscopia no Infravermelho Próximo .................................................................................................... 37
3.1 Introdução ................................................................................................................................................. 37
3.2 Conceitos básicos: absorção e Lei de Beer-Lambert .................................................................. 40
3.3 Espalhamento da luz .............................................................................................................................. 43
3.4 Métodos ópticos aplicados ao estudo cerebral ............................................................................ 44
3.4.1 Janela óptica para estudos não-invasivos .....................................................................................45
3.4.2 Absorvedores biológicos: absorção da água ................................................................................48
3.4.3 Absorvedores biológicos: absorção da hemoglobina ...................................................................49
3.5 Propagação da luz em tecido biológico ................................................................................................ 50
3.5.1 Computando variações nas concentrações de cromóforos ...........................................................52
Capítulo 4 ..........................................................................................................................................................54

xii
Objetivos .................................................................................................................................................................... 54
Capítulo 5 ..........................................................................................................................................................55
Sujeitos e Métodos ................................................................................................................................................. 55
5.1 Sujeitos ........................................................................................................................................................ 55
5.2 Materiais e Métodos ............................................................................................................................... 56
5.2.1 Paradigma ....................................................................................................................................56
5.3 Aquisição e análise dos registros de EEG ....................................................................................... 59
5.4 Aquisição e análise dos dados de NIRS ............................................................................................ 62
Capítulo 6 ..........................................................................................................................................................66
Resultados e discussão .......................................................................................................................................... 66
6.1 Resultados de um único indivíduo ........................................................................................................... 67
6.2 Análises de Grupo ........................................................................................................................................... 70
6.2.1. Resultados de NIRS: experimento binocular ...........................................................................70
6.2.2. Resultados de NIRS: experimento monocular .............................................................................78
6.3 Grupos para o EEG, resultados e discussão ........................................................................................... 82
6.3.1 Resultados e discussão para o experimento binocular ..........................................................82
6.3.2. Resultados e discussão para o experimento monocular .......................................................85
6.4 Discussão sobre os ritmos resultantes alfa e beta ............................................................................. 89
Capítulo 7 .........................................................................................................................................................91
Conclusões e Perspectivas .................................................................................................................................. 91
BIBLIOGRAFIA ................................................................................................................................................93
Anexo: Protocolo de aprovação do projeto pesquisa pelo Comitê de Ética da Unicamp...........................104

xiii
Lista de Figuras 1.1 Divisão do Sistema Nervoso Central (SNC) Humano. . . . . . . . . . . . . . . . . . . . . . . . . ...2
1.2 Visão Lateral do Cérebro, mostrando a divisão dos lobos (Figura retirada de
http://www.neuroeducacao.com.br/neurociencias.asp). . . . . . . . . . . . . . . . . . . . . . . . . .3
1.3 Disposição de neurônios e células gliais (astrócitos, oligodendrócitos e microglia) no
córtex (Figura retirada de Scientific American do Brasil, edição 24, maio 2004, pp.49-
53). No canto superior direito em destaque uma sinapse. . . . . . . . . . . . . . . . . . . . . . ... 4
1.4 Estrutura básica de um neurônio. O corpo celular ou pericário possui o núcleo que
contém o material genético (DNA), e as organelas citoplasmáticas responsáveis pela
manutenção da célula. Os dendritos, terminações nervosas, que recebem os sinais de
outros neurônios. Os axônios que transportam o sinal processado no neurônio para
outros neurônios (Figura adaptada de: Histologia Básica, Luis C. Junqueira, capitulo 9,
p. 155). Em destaque (elipses) as partes fundamentais do neurônio . . . . . . .. . . . . . . . . 5
1.5 Principais tipos de neurônios. A morfologia dessas células é muito complexa. Todos os
neurônios mostrados, exceto os dois neurônios bipolares e o pseudo-unipolar, que não
são muito numerosos no tecido nervoso, são neurônios de tipo multipolar. (Figura
retirada de Histologia Básica, Luis C. Junqueira, capitulo 9, p. 156 ). . . . . . . . . . . . . 6
1.6 À esquerda: Esquema de um corte do cérebro, mostrando o córtex (substância cinzenta)
e a substância branca (Fonte: www.guia.heu.nom.br/cortex_cerebral.htm). À direita:
divisão das camadas corticais, mostrando a morfologia e distribuição dos neurônios
piramidais corticais. Note a variabilidade no tamanho das células e a arborização
dendrítica, assim como a presença dos axônios colaterais, dependentes da localização
laminar (I–VI) do neurônio. Alem disso, diferentes tipos de neurônios piramidais com

xiv
uma precisa distribuição laminar projetada sobre as diferentes regiões do cérebro
(Adaptado de Squire, 2002) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 8
1.7 Áreas de Brodmann, mostrando a atribuição funcional do córtex (Figura adaptada de
http://spot.colorado.edu/~dubin/talks/brodmann/brodmann.html). . . . . . . . . . . . . . . . . . . 10
1.8 Áreas de associação (branco) e áreas de projeção, divididas em sensitivas (vermelho) e
motoras (preto) (Figura adaptada de http://medicalartlibrary.com). . . . . . . . . . . . . . . . . . . 11
1.9 Corte estrutural do olho humano (Figura adaptada de
http://www.vision.ime.usp.br/~ronaldo/mac0417-3/aula_02). . . . . . . . . . . . . . . . . . . . . . . 13
1.10 Diagrama esquemático da secção transversal das camadas plexiformes de um
macaco rhesus (Adaptado de Squire, 2002). . . . . . . . . . . . . . . . . . . . ... . . . . . . . . . . . . . 16
1.11. Trajetória das vias visuais (figura adaptada de Color Atlas of Neuroscience
Greenstein, 2000, pag. 283). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
2.1 A célula nervosa (figura retirada de http://infook.blogspot.com/2007/08/como-funciona-
o-crebro.html). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
2.2 Geração de potencias sobre o escalpo pela somatória de correntes.. . . . . . . . . . . . . . . .23 2.3 Representação esquemática de um neurônio piramidal cortical (Figura retirada de:
Epilepsy as a Dynamic Disease, J. Milton et al, editors, capítulo 5, p. 55) . . . . . . . . . . . . 23
2.4 Padrão do fluxo de corrente elétrica para um PPSE sobre um dendrito apical de um
neurônio piramidal no córtex cerebral (Figura extraída de Principles of Neural Science de
Kandel, 2000, Fourth edition, chapter 46, pag. 914) . . . . . . . . . . . . . . . . . . . . . . . . . . . . .24
2.5 Polaridade dependente da posição da sinapse . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ...25

xv
2.6 Diagrama comparando a performance espacial e temporal de quatro técnicas de
neuroimagem minimamente invasivas. A diagonal “inclinada” representa o
melhoramento na resolução espacial com o tempo crescente da medida, uma
característica das quatro modalidades [Strangman, 2002]. . . . . . . . . . . . . . . . . . . . . . .27
2.7 Diagramas das diferentes etapas de um registro EEG e sistema quantitativo
[Thakor,2004]. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 30
2.8 Posicionamento dos eletrodos no Sistema Internacional 10-20,para registros EEG
[Malmivuo, 2008]. A=auricular, Pg=nasofaríngeo, C=central,P=parietal,
T=temporal,F=frontal, Fp=fronto-polar e O=occipital. . . . . . . . . . . . . . . . . . . . . . . . . 35
3.1. Posição Fonte-Detetor na coleta de dados NIRS (extraído de Izzetoglu et al., 2007)
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .45 3.2. Trajetória de “fótons infravermelhos individuais” na interaçãoluz-tecido biológico
(extraído de Boas et al., 2001). . . . . . . . . . . . . . . ... . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46 3.3. Espectro de absorção da oxihemoglobina, desoxihemoglina e água ((extraído de
Izzetoglu et al., 2007) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .47
3.4 Coeficiente de absorção da água (extraído de Hale et al., 1973) . . . . . . . . . . . . . . . . . .48 3.5 Modelo da banana, que representa a forma geral da trajetóriados fótons infravermelhos
(extraído de Bunce et al., 2006) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 50
5.1 Montagem experimental. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58

xvi
5.2 . Estímulo visual: série de 21 épocas, alternando repouso e ativação. . . . . . . . . . . . . . .59 5.3 Estímulos apresentados, variando se a freqüência e o contraste. . . . . . . . . . . . . . . . . . 59 5.4 Estímulo total: série de 15 épocas (8 épocas de repouso e 7 de ativação), para o
experimento monocular. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60
5.5 Posicionamento dos eletrodos O1 e O2 (em azul) sobre cada um dos lobos occipitais,
na região do córtex visual primário. Em amarelo, duas formas semilunares, indicam os
registros de NIRS. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 61
5.6. Layout do software EEGLAB. Acima: à esquerda barra de rolagem e painel de
entrada dos sinais EEG à direita. Abaixo: mapa de intensidade de uma componente
independente (IC) à esquerda e mapas de um canal específico à direita (Figura retirada de
http://scnn.ucsd.edu/eeglab/). . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 62
5.7. Uma das etapas do processamento dos nossos dados EEG, por médio do software
EEGLAB: a esquerda a barra de rolagem com a entrada dos sinais EEG, e a direita os sinais
de EEG cru dos canais O1 e O1. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
5.8. Geometria dos optodos utilizada para os registros de NIRS. As letras (A, B, C, D)
indicam as posições das fontes de luz NIR enquanto que os números (1, 2, 3, 4) indicam as
posições dos detectores. Os eletrodos O1 e O2 foram colocados ao centro de cada um
desses arranjos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64
5.9. Geometria dos optodos e eletrodos. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 64
5.10. Equipamento NIRS , CW6, canto superior esquerdo e acessórios(canto superior
direito e abaixo), . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 65
6.1. Resposta hemodinâmica registrada através do NIRS para um sujeito. . . . . . . . . . . . . 68

xvii
6.2. Resposta hemodinâmica registrada através do NIRS para um sujeito variando-se a
freqüência do estímulo. A barra horizontal indica o período em que o estímulo visual foi
aplicado. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .69
6.3. Resultado da resposta EEG para um sujeito quando o estímulo visual aplicado no caso
binocular. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .70
6.4 Resultados de NIRS (experimento binocular) para a análise de grupo para cada um dos
canais, variando-se a freqüência de estímulo: 4, 8 e 12 Hz . . . . . . . . . . . . . . . . . . . . . . . . .71
6.5 Resultados de NIRS (experimento binocular) para a análise de grupo para cada um dos
canais, variando-se a freqüência de estímulo: 1, 2 e 3 Hz . . . . . . . . . . . . . . . . . . . . . . . . . .72
6.6 Resultados de NIRS (experimento binocular) para a análise de grupo para cada um dos
canais, variando-se o contraste do estímulo: 30, 60 e 100 %. . . . . . . . . . . . . . . . . . . . . . . .73
6.7 Resultados de NIRS (experimento binocular) para a análise de grupo para cada um dos
canais, variando-se o contraste do estímulo: 3, 10 e 20 %. . . . . . . . . . . . . . . . . . . . . . . . . 74
6.8 Resultados de NIRS (experimento binocular) para a análise de grupo para cada um dos
canais, e para todas as freqüências de estímulo:1, 2, 3, 4, 8 e 12 Hz (contraste 100%).. . . 75
6.9 Resultados de NIRS (experimento binocular) para a análise de grupo para cada um dos
canais, e para todos os níveis de contraste: 3, 10, 20, 30, 60 e 100 % (frequência = 4 Hz). . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 76
6.10 Áreas sob as curvas de HbO (em azul) e dHb (em vermelho) em função da freqüência do estímulo (experimento binocular) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .77

xviii
6.11 Áreas sob as curvas de HbO (em azul) e dHb (em vermelho) em função dos níveis de contraste (experimento binocular) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 77 Fig. 6.12. Áreas sob as curvas de HbO (em azul) e dHb (em vermelho) em função da freqüência do estímulo (experimento binocular) . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . 78 Fig. 6.13. Áreas sob as curvas de HbO (em azul) e dHb (em vermelho) em função dos níveis de contraste (experimento binocular) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . 79 6.14. Experimento monocular: variação de frequência . . . . . . . . . . . . . . . . . . . . . . . . . . . 80 6.15. Experimento monocular: variação de contraste. . . . . . . . . . . . . . . . . . . . . . . . . . . . . 81 6.16. Figura retirada de Zhang et al ( 2006),Rev.Sci. Instrum, 77 (11) 114301. . . . . . . . . 82 6.17. Figura retirada de Hoge et al (2005), Neuroimage 25 , 701-707. . . . . . . . . . . . . . . . 82 6.18. Resultados do EEG binocular, variação de frequência do estímulo. . . . . . . . . . . . . . 84 6.19. Resultados do EEG binocular, variação de contraste do estímulo. . . . . . . . . . . . . . . 85 6.20. Resultados do EEG monocular, variação de freqüência do estímulo. . . . . . . . . . . . . 86 6.21. Resultados do EEG monocular, variação de contraste do estímulo . . . . . . . . . . . . .. 88 6.22. Resultados globais EEG, variação de frequência do estímulo . . . . . . . . . . . . . . . . . . 89 6.23. Resultados globais EEG, variação de contraste do estímulo. . . . . . . . . . . . . . . . . . . .89

xix

xx
LISTA DE ABREVIATURAS E SIGLAS
ASL – Arterial Spin-Labeling, ou Marcação de Spin Arterial
ATP – Adenosina Trifosfato
BOLD – Blood Oxygention Level Dependent (Effect)
CBF – Cerebral Blood Flow, ou Fluxo Sangüíneo Cerebral
CBS - Cerebral Blood Saturation, ou Saturação de Oxigênio Sangüíneo Cerebral
CBV – Cerebral Blood Volume, ou Volume Sangüíneo Cerebral
CGL – Corpo Geniculado Lateral
CMRO2 – Cerebral Metabolic Rate of Oxygen, ou Consumo Metábolico de Oxigênio
CT – Computerized Tomography, ou Tomografia computarizada
CW - Continuous Wave
DOT – Diffuse Optical Tomography, ou Tomografia Óptica de Difusão
DDP - Diferença de Potencial
DPF - Differential Pathlength Factor
EEG – Eletroencefalografia
EDF – European Date Format
EPSPs / IPSPs – Excitatory/ Inhibitory Postsinaptic Potential, ou Potenciais pós-
sinápticos excitatórios e inibitórios
ERP – Event Related Potential, ou Potencial Relacionado a Eventos
EP – Evoked Potential, ou Potencial Evocado
fMRI – functional Magnetic Resonance Imaging, ou Ressonância Magnética funcional
HbO – Oxihemoglobina
dHb – Desoxihemoglobina
HbT – Hemoglobina total
HRF – Hemodynamic Response Function, ou Resposta Hemodinâmica
HOMER - Hemodynamic Evoked Response
LDF - Laser Doppler Fluxometry

xxi
MBLL - Modified Beer-Lambert Law, ou Lei de Beer Lambert modificada
MEG - Magnetoencefalograma
MRI – Magnetic Resonance Imaging, ou Imagem por Ressonância Magnética
NIRS – Near Infrared Spectroscopy, ou Espectroscopia no Infravermelho Próximo
OD – Optical Density, ou Densidade Óptica
PET – Positron Emission Tomography, ou Tomografia por Emissão de Pósitrons
PA – Potencial de Ação
PPS - Potenciais Pós-Sinápticos
PS – Pressão Sangüínea
RTE – Radiative Transport Equation, ou Equação de Transporte Radiativa
SNC – Sistema Nervoso Central
SNP – Sistema Nervoso Periférico
SNR – Signal-to-Noise Ratio, ou Relação Sinal-Ruído
SPECT - Single Photon Emission Computed Tomography
VEP – Visual Event-related Potencial

xxii

xxiii

1
Capítulo 1
O Cérebro e o Sistema Visual Humano
1.1 O Cérebro Humano
O cérebro humano, um dos mais fascinantes e maiores mistérios da ciência, é um órgão
complexo e altamente especializado. É o mais intrincado sistema dos seres viventes que
existem sobre a Terra. Tem por finalidade propiciar controle motor, fisiológico e perceptivo
do mundo ao nosso redor, nos permite executar atividades mentais de alto nível, tais como
o pensamento racional, sentimentos e emoções, raciocínio criativo, ordenado e intencional,
Fig. 1.8 e, além disso, nos dá os meios para o estabelecimento das mais variadas formas de
comunicação.
O estudo do cérebro tem gerado grandes enigmas, que têm sido tratados através de
muitas disciplinas, tais como a filosofia, a psicologia e a psicanálise. Desde a antiga Grécia
até hoje, seguem perguntas abertas tais como: Como lembramos? O que é a memória?
Quais são os mecanismos que geram a memória? Como prestamos atenção?, entre outras.
Todos estes processos ocorrem em nosso cérebro, e conhecer os mecanismos e/ou as
funções envolvidas nesses processos é o objetivo principal dos neurocientistas.
Sabe-se que áreas específicas do cérebro desempenham funções determinadas [Kandel,
2000]. No entanto, podem ser encontradas áreas comuns para diferentes atividades, ou
várias áreas responsáveis por uma única atividade. Na verdade, ainda não se conhecem
profundamente as funções de cada parte do cérebro. Quando se trata de abordagens não
invasivas, o que é possível observar desde há duas décadas, é a distribuição espacial do

2
consumo de energia no cérebro de um voluntário envolvido em uma determinada tarefa,
para estimar quais regiões estão mais envolvidas nessa tarefa específica. Para os
neurocientistas, por mais simples que seja uma tarefa, as regiões “envolvidas” do cérebro
não são apenas uma pequena parte, pois muitas outras áreas, de função ainda não
determinada, são também “ativadas”. Nestes casos, a diferença pode estar na intrincada
circuitaria de milhões de conexões estabelecidas pelos neurônios.
1.1.1 Anatomia Cerebral
Anatomicamente, o sistema nervoso humano se divide em sistema nervoso central
(SNC) e sistema nervoso periférico (SNP) [Waxman, 2000].
O SNC compreende a medula espinhal, alargada, e o encéfalo, de grande
complexidade, que é concebido como uma super-rede de redes interconectadas de células
nervosas. O encéfalo por sua vez se divide em cérebro, talo encefálico e cerebelo (Fig. 1.1).
Fig. 1.1. Divisão do Sistema Nervoso Central (SNC) Humano.
O cérebro, a parte mais importante e desenvolvida do encéfalo, é dividido em duas
metades, os hemisférios cerebrais direito e esquerdo, que se conectam por um feixe de

3
fibras nervosas, através do corpo caloso (900 milhões de fibras). Cada hemisfério é
dividido em quatro lobos: o frontal, o parietal, o occipital e o temporal (Fig. 1.2).
Os lobos frontais são dominantes em cognição social e emoção. Eles também
controlam o comportamento motor especializado, na parte direita; e as funções relacionadas
com a linguagem, na parte esquerda [Miller et al., 2006].
Os lobos parietais, localizados na parte superior do cérebro, são responsáveis pela
interpretação dos estímulos provenientes do corpo, o que possibilita controlar os
movimentos corporais. Eles regulam funções corticais tais como noção espacial e de
reconhecimento dos objetos [Machado, 2003].
Os lobos temporais estão associados a habilidades de memória, permitindo que os
sujeitos reconheçam outros sujeitos e objetos, processem e lembrem-se de eventos
distantes, e estabeleçam comunicação entre eles.
Os lobos occipitais abrangem mais a porção posterior do córtex cerebral humano,
sendo responsáveis principalmente pela visão [DeYoe, 2002].
Fig. 1.2. Visão Lateral do Cérebro, mostrando a divisão dos lobos (Figura retirada de
http://www.neuroeducacao.com.br/neurociencias.asp)
Na base do cérebro, estão localizados agrupamentos de células nervosas em estruturas
denominadas gânglios basais, tálamo e o hipotálamo. Os gânglios basais colaboram na
realização de movimentos suaves. O tálamo geralmente organiza as mensagens sensoriais

4
de entrada e saída dos níveis superiores do cérebro (córtex cerebral), e o hipotálamo
coordena algumas das funções mais automáticas do corpo, como o controle do sono e da
vigília, a manutenção da temperatura corporal e a regulação do equilíbrio hídrico do
organismo.
Os componentes estruturais ou unidades de construção básicos do cérebro são as
células nervosas: os neurônios e as células gliais (macroglia: oligodendrócitos, células de
Schwann e astrócitos; e microglia: fagócitos). Os neurônios são células altamente
especializadas, que geram sinais elétricos em resposta a umas substâncias chamadas de
neurotransmissores e os transmitem a outras células. São responsáveis pelo processamento
de informações, em quanto que as células gliais têm funções de dar sustentação aos
neurônios, ajudando a manter as concentrações iônicas adequadas e transportar substâncias
entre os vasos sangüíneos e o tecido cerebral (Fig. 1.3).
Fig.1.3. Disposição de neurônios e células gliais (astrócitos, oligodendrócitos e microglia) no córtex
(Figura retirada de Scientific American do Brasil, edição 24, maio 2004, pp.49-53). No canto superior
direito em destaque uma sinapse.

5
1.1.2 Neurônios
Os neurônios são células nervosas estruturais, funcional e anatomicamente complexas,
altamente especializadas, que recebem, geram, processam e transmitem informação, através
de impulsos elétricos [Mattews, 2003; Squire, 2003; Aidley, 2001; Kandel, 2000]. O
cérebro humano possui cerca de 1010 neurônios, cada um com aproximadamente 104
interconexões. Portanto, estima-se em 1014 o numero de conexões existentes no cérebro
humano [Guyton, 2000; Waxman, 2000; Lent,2005].
Morfologicamente, cada neurônio possui quatro partes bem definidas: o pericário ou
corpo celular, os dendritos, o axônio e suas terminações axônicas ou pré-sinápticas (Fig.
1.4).
Fig. 1.4. Estrutura básica de um neurônio. O corpo celular ou pericário possui o núcleo que contém o
material genético (DNA), e as organelas citoplasmáticas responsáveis pela manutenção da célula. Os
dendritos são terminações nervosas que recebem os sinais de outros neurônios. Os axônios transportam o
sinal processado no neurônio para outros neurônios (Figura adaptada de: Histologia Básica, Luis C.
Junqueira, capítulo 9, p. 155). Em destaque (elipses) as partes fundamentais do neurônio.

6
O corpo celular é o centro metabólico do neurônio, a árvore dendrítica é o principal
lugar para receber o sinal de entrada, e o axônio é a principal unidade de condução para
transmitir sinais, chamados de potenciais de ação, entre os neurônios e suas terminações
pré-sinápticas, lugares onde acontecem as sinapses [Kandel, 2000].
Existem diferentes tipos de neurônios, de forma que podemos classificá-los do ponto
de vista morfológico e funcional. Morfologicamente temos principalmente neurônios
unipolares, neurônios bipolares, neurônios multipolares (predominantes no sistema nervoso
dos vertebrados, por exemplo: a célula de Purkinje do cerebelo). Funcionalmente, temos os
neurônios sensoriais, que carregam informação para o sistema nervoso periférico; os
neurônios motores, que carregam comandos do cérebro ou medula espinal até músculos e
glândulas; e os interneurônios de projeção e interneurônios locais. No entanto, no cérebro e
principalmente no córtex cerebral, podem ser divididos em dois principais grupos:
neurônios piramidais e não-piramidais (Fig. 1.5).
Figura 1.5. Principais tipos de neurônios. A morfologia dessas células é muito complexa. Todos os
neurônios mostrados, exceto os dois neurônios bipolares e o pseudo-unipolar, que não são muito
numerosos no tecido nervoso, são neurônios de tipo multipolar. (Figura retirada de Histologia Básica, Luis
C. Junqueira, capítulo 9, p. 156).

7
O mecanismo de acoplamento entre o axônio de uma célula e o neurônio seguinte pode
ser de vários tipos, sendo os mais comuns: os axodendríticos (cerca de 75% dos casos),
axosomáticos (cerca de 20% dos casos) e os axoaxônicos (cerca de 5%). Outros contatos
tais como os dendrodendríticos e somatosomáticos também são encontrados, mas são raros.
O impulso elétrico que se propaga pelo neurônio ativo é transformado em sinal químico na
sinapse, a região de comunicação entre as células nervosas.
A fenda sináptica compreende o espaço (30nm) entre o elemento pré-sináptico, que
armazena e libera o neurotransmissor, e o elemento pós-sináptico, que contém os
neuroreceptores. Quando o impulso nervoso, ou potencial de ação, atinge a membrana do
terminal pré-sináptico, origina-se uma pequena alteração do potencial de membrana, capaz
de abrir os canais de cálcio, determinando a entrada desse íon. O aumento dos íons de
cálcio no interior do elemento pré-sináptico provoca uma série de fenômenos. Alguns deles
culminam com a fusão de vesículas sinápticas com a membrana pré-sináptica, ocorrendo,
assim, a liberação de neurotransmissores na fenda sináptica, que se difundem até se ligarem
aos neuroreceptores. Esta ação origina alterações no equilíbrio da célula receptora,
tornando-a mais ou menos susceptível à propagação de um potencial de ação, o potencial
pós-sináptico.
A diferença de concentração iônica entre o interior e o exterior da célula nervosa dá
origem a uma diferença de potencial (ddp) entre as paredes da membrana. Em estado de
repouso, essa diferença é mantida constante a aproximadamente -70mV. O impulso elétrico
de uma célula nervosa provoca o surgimento de correntes iônicas na membrana celular
seguinte, alterando sua ddp. Dessa forma, uma célula comunica informação à outra.
1.1.3 Córtex cerebral
O córtex cerebral é uma fina lâmina de células que recobre o cérebro humano, com
aproximadamente 250000mm2, sendo composta de aproximadamente 1010 neurônios [Jirsa
et al, 2007; Cechetto et al, 2002]. Sua espessura varia de 2-7 mm. O córtex constitui a
substância cinzenta que envolve os hemisférios. Nos humanos, o cérebro apresenta uma

8
superfície irregular, com áreas mais protuberantes, os giros cerebrais, intercalada com
pequenos vales, os sulcos (Fig. 1.6).
Fig. 1.6. À esquerda: Esquema de um corte do cérebro, mostrando o córtex (substância cinzenta) e a
substância branca (Fonte: www.guia.heu.nom.br/cortex_cerebral.htm). À direita: divisão das camadas
corticais, mostrando a morfologia e distribuição dos neurônios piramidais corticais. Note a variabilidade
no tamanho das células e a arborização dendrítica, assim como a presença dos axônios colaterais,
dependentes da localização laminar (I–VI) do neurônio. Além disso note diferentes tipos de neurônios
piramidais com uma precisa distribuição laminar projetada sobre as diferentes regiões do cérebro
(Adaptado de Squire, 2002).
O córtex é uma das partes mais importantes do sistema nervoso. Nele chegam os
impulsos provenientes de todas as vias de sensibilidade que aí se tornam conscientes e são
interpretadas, e dele saem os impulsos nervosos que iniciam e comandam os movimentos
voluntários. Com ele estão relacionados todos os fenômenos psíquicos. O neocórtex é sua
estrutura mais complexa: possui estrutura laminar formada por seis camadas diferentes e
contém várias áreas sensoriais e motoras.
Diferentes tipos de neurônios e fibras se organizam ao longo das seis camadas
corticais e são ligados entre si por circuitos locais, através de diferentes tipos dos
interneurônios. Esses circuitos ligam diferentes áreas corticais a outras regiões do próprio
córtex e a inúmeras estruturas subcorticais.

9
As informações sensoriais chegam ao córtex através das fibras aferentes, que partem
do tálamo, transitam pelas diferentes camadas através de microcircuitos formados por
células piramidais e interneurônios, e são dirigidas; através das fibras eferentes, para a
medula espinhal, tálamo e diferentes regiões do tronco encefálico, entre outras.
As camadas infragranulares (V e VI) são responsáveis pela saída extracortical,
processando informações e as enviando às estruturas subcorticais, como o núcleo estriado e
a medula espinhal. A camada granular (IV) recebe as aferências talâmicas, ou seja,
informações sensoriais do corpo o do mundo externo, que são recebidas primeiramente pelo
tálamo. Já as camadas supragranulares (I, II e III) são responsáveis pela comunicação
intercortical.
Além da organização horizontal em camadas, o córtex também é organizado
verticalmente em colunas. Pequenas porções verticais de neurônios, localizadas em
diferentes regiões do córtex, são responsáveis pelo processamento de funções distintas.
Assim, as espessuras das camadas, no interior de uma coluna, variam de acordo com a
especialização da região onde a coluna se encontra [Kandel et al., 2000].
O córtex compreende ainda outras duas partes. Com uma porção dos lobos frontal,
parietal e occipital forma-se o lobo límbico, também conhecido como sistema límbico.
Além dele, outro constituinte importante do córtex cerebral é o córtex insular, que se
localiza na profundidade do sulco lateral, coberto por partes dos lobos frontal e parietal.
O sistema límbico tem forma de um anel cortical contínuo, que contorna as formações
inter-hemisféricas. Ele compreende um grupo de estruturas que inclui o tálamo, o
hipotálamo, a amígdala, o hipocampo, os corpos mamilares e o giro do cíngulo. Esse
sistema é muito importante para a emoção e reações emocionais.
1.2 Funções cerebrais
O córtex cerebral não é homogêneo em toda sua extensão, o que permite individualizar
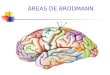
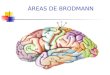
várias áreas. Na Fig. 1.7, podemos observar as àreas de Brodmann e sua atribuição
funcional, pois diferentes regiões têm diferentes conexões e funções. Existe uma
especialização funcional de cada região do córtex, ainda que não em termos absolutos; isto

10
é, diferentes áreas podem contribuir para a execução de uma mesma função, sem contar o
envolvimento de algumas estruturas subcorticais. Dessa forma, é difícil falar em centros
funcionais, sendo preferível considerar a existência de sistemas funcionais envolvendo
várias áreas que funcionam harmonicamente.
Fig. 1.7. Áreas de Brodmann, mostrando a atribuição funcional do córtex (Figura
adaptada de http://spot.colorado.edu/~dubin/talks/brodmann/brodmann.html)
Atualmente, as funções cerebrais podem se dividir em dois grandes grupos: áreas de
projeção - áreas primárias - que recebem e dão origem às fibras relacionadas diretamente
com a sensibilidade e com a motricidade, sendo divididas em áreas sensitivas e motoras; e
as áreas de associação - áreas secundárias e terciárias – que estão diretamente relacionadas
com funções cerebrais complexas (Fig. 1.8).

11
Fig. 1.8. Áreas de associação (branco) e áreas de projeção, divididas em sensitivas (vermelho) e
motoras (preto) (Figura adaptada de http://medicalartlibrary.com).
1.2.1 Áreas motoras
A área de projeção motora se localiza no lobo frontal, sendo responsável pelos
movimentos das regiões contralaterais do corpo, tais como pé, mão e lábios. O córtex motor
possui três subáreas: o córtex motor primário: área 4 de Brodmann, que ordena a região do
movimento e que se localiza na região posterior do giro pré-central; o córtex pré-motor e o
córtex motor suplementar que corresponde à área 6 de Brodmann, que se encarrega do
planejamento.
1.2.2 Áreas sensitivas
A área de projeção sensitiva situa-se na região do giro pós-central e corresponde às
áreas 1 - 3 de Brodmann. Esta área recebe fibras de neurônios situados no tálamo, que
trazem as informações de temperatura, pressão, dor, tato, cheiro, e outras, da metade oposta
do corpo e da cabeça.
As áreas de associação ocupam a maior parte da superfície do cérebro humano. Ao
longo do processo evolutivo, o aumento da superfície cortical se fez através da expansão do

12
córtex de associação, permitindo o aparecimento de funções no homem não encontradas em
outras espécies, com a linguagem verbal e autoconsciência. Podem ser divididas em áreas
secundárias e terciárias. As áreas secundárias estão diretamente conectadas às áreas de
projeção e são unimodais, ou seja, estão ainda relacionadas com uma modalidade sensorial
ou com a motricidade. As áreas terciárias são áreas integradoras, estão conectadas
basicamente com as áreas secundárias e com as áreas límbicas e são multimodais, ou seja,
não se ocupam mais do processamento sensorial ou motor, mas estão envolvidas com as
atividades superiores, como o pensamento abstrato e/ou processos que permitem a
simbolização.
A área auditiva primária situa-se no giro temporal transverso anterior (áreas 41 e 42 de
Brodmann). Sons de diferentes freqüências chegam a partes diferentes do córtex auditivo,
onde ocorre então uma tonotopia. Estimulações na área auditiva primária provocam
sensação auditiva mal-definida, como zumbidos. Lesões nesta área dificilmente causam
surdez, porque as vias auditivas, apesar de cruzarem a linha média, têm um grande
componente ipsilateral (do mesmo lado), ou seja, fibras que não se cruzam e que irão
atingir o córtex auditivo do mesmo lado.
A área visual primária situa-se na área 17 de Brodmann, nas bordas do sulco calcarino
no lobo occipital. O córtex visual primário de cada hemisfério cerebral recebe informações
procedentes do campo visual contralateral. Cada ponto do campo visual encontra um
correspondente no córtex visual. Há uma retinotopia, já que cada parte da retina se projeta
para uma parte específica do córtex cerebral. Estimulação na área 17 faz com que o
indivíduo relate estar vendo clarões ou pontos luminosos nas regiões correspondentes do
campo visual.
1.3 O Sistema Visual Humano
A compreensão da visão humana começa com o estudo da anatomia e da fisiologia
visual básica. É importante estudar o hardware do sistema visual porque este pode dar
introspecções nos tipos da informação que podem ser codificados pelos mecanismos
visuais.

13
1.3.1 O Olho
O sistema visual começa no olho. A Fig. 1.9 mostra uma secção transversal através de
um esquema do olho humano. A seção anterior do olho humano contém o sistema óptico do
olho cujas estruturas principais são a córnea, a íris e a lente ou cristalino. A córnea
proporciona aproximadamente duas terças partes do poder refrativo do olho, mas a lente
fornece controle focal fino para alvos em distâncias de 10 cm até cerca de 6 m. A íris se
situa na frente da lente e possui uma abertura central conhecida como a pupila que admite a
luz à cavidade central do olho. O espaço entre a córnea e a lente é preenchido de um líquido
conhecido como humor aquoso. A cavidade central do globo ocular contém um líquido
gelatinoso, conhecido como humor vítreo.
Fig. 1.9 Corte estrutural do olho humano (Figura adaptada de
http://www.vision.ime.usp.br/~ronaldo/mac0417-3/aula_02)

14
A seção posterior do olho tem três camadas. A esclera é uma coberta exterior
resistente, que protege o interior de dano e ajuda a manter o olho aproximadamente na
forma esférica. A coróide é uma camada média que fornece a fonte de sangue às estruturas
celulares do olho. A retina é a camada interior que contém células fotorreceptoras e seus
tecidos neurais associados.
1.3.2 A retina
A retina é composta de duas classes principais de células fotorreceptoras conhecidas
como bastonetes e cones por causa das formas de seus segmentos externos. Cada retina
possui entre 100 a 120 milhão de bastonetes e 7 a 8 milhões de cones. Os bastonetes são
extremamente sensíveis à luz e fornecem visão acromática a baixos níveis da iluminação
(escotópicos). Os cones são menos sensíveis do que os bastonetes, mas fornecem a visão de
cor a níveis elevados (fotópicos). Os segmentos fotosensitivos dos bastonetes e dos cones
jazem mais próximos à camada coróide. Isto significa que a luz que bate a retina deve
primeiramente passar por diversas camadas de tecido neural antes de alcançar os
fotorreceptores. Somente uma pequena área de 1.5mm de diâmetro, próxima do eixo óptico,
chamada fóvea, é que é uma superfície fotorreceptora exposta diretamente à luz.
Os sistemas dos bastonetes e dos cones são sensíveis aos comprimentos de onda de luz
aproximadamente entre 400 e 700nm. Os bastonetes têm seu pico de sensibilidade máxima
em aproximadamente 498nm. Três tipos de cones têm características de resposta espectral
passa-banda. O comprimento de onda curto ou os cones “azuis” têm seu pico máximo em
420nm, o comprimento de onda médio ou os cones “verdes” têm seu pico máximo em
534nm, e em comprimentos de onda longos os cones “vermelhos” têm seu pico máximo em
564nm.
Existe uma sobreposição significativa entre as escalas da resposta das diferentes
classes de cones, o qual significa que espectralmente os estímulos de faixa larga
simultaneamente ativam múltiplos tipos de cones.

15
Os bastonetes e os cones não são distribuídos igualmente sobre a superfície retinal. A
fóvea é mais densa em cones, mas é de população quase nula em bastonetes.
1.3.3 Campos receptivos retinais
As sinapses dos bastonetes e dos cones ocorrem numa rede de neurônios nas camadas
plexiformes interna e externa da retina. A Fig. 1.10 mostra um diagrama esquemático de
uma secção transversal das camadas plexiformes de um macaco rhesus. As células nas
camadas plexiformes conectam grupos de bastonetes e de cones às células ganglionares
cujas fibras neurais formam o nervo óptico. O grupo localizado de fotorreceptores que
serve uma particular célula ganglionar é chamado de campo receptivo da célula.
Os campos receptivos das células ganglionares são as unidades básicas de codificação
visual. Estudos eletrofisiológicos em gatos mostraram que muitos campos receptores têm
uma organização antagônica centro-periferia [Kuffler, 1953]. A ativação produzida por
estimulação no centro de um campo receptivo tende a ser suprimida pela estimulação na
periferia anular. Estimulação uniforme sobre todo o campo receptivo produz tipicamente
uma resposta fraca.
Os pesquisadores identificaram duas classes de campos receptivos de células
ganglionares. As células centralizadas (on-center) aumentam sua taxa de disparo em
resposta aos incrementos da luz nos centros de seus campos, e as células descentralizadas
(off-center) aumentam sua taxa de disparo em resposta aos decréscimos de luz. A
organização antagonista dos campos receptivos significa que em princípio no sistema
visual, informação sobre a intensidade absoluta da luz é em sua maior parte perdida e
primeiramente o contraste é sinalizado a estados avançados de processamento visual.

16
Fig. 1.10 Diagrama esquemático da secção transversal das camadas
plexiformes de um macaco rhesus (Adaptado de Squire, 2002).
Isto tem implicações significativas nas teorias da percepção de superfície da
luminosidade e da iluminação.
As células ganglionares podem ser igualmente classificadas por padrões e pela
duração de suas respostas às mudanças na luz em seus campos receptivos. As células X
mostram uma resposta estacionária aos incrementos ou aos decréscimos nos centros de seus
campos. As células Y mostram uma breve mudança transiente na resposta e logo retornam
a sua taxa de limiar fundamental.
Aproximadamente metade de todas as células ganglionares retinais têm campos
receptores que mostram oposição espectral assim como espacial. As células oponentes
verde-vermelhas tomam sua entrada preliminar de comprimentos de ondas médio e longos.
Células oponentes azul-amarelo tomam sua entrada de todos os três tipos de cones, com

17
oposição entre a soma dos comprimentos de onda longos e médios dos cones e dos
comprimentos de onda curtos dos cones. A descoberta das células com propriedades
opostas espectralmente tem sido bem utilizada para dar suporte fisiológico às teorias da
percepção da cor.
1.3.4 Trajetória das Vias Visuais
A Fig. 1.11 mostra os principais caminhos neurais no sistema visual. Os axônios
longos das células ganglionares retinais formam o nervo óptico, que contém
aproximadamente um milhão de fibras, das quais 100.000 servem à fóvea. O feixe de fibras
do nervo óptico sai do globo ocular em aproximadamente 17 graus ao lado nasal da linha
do eixo óptico. Não existem fotorreceptores nesta área conhecida geralmente como ponto
cego.
As fibras do nervo óptico projetam-se ao quiasma óptico. Nesta junção, fibras das
parcelas nasais de cada retina se cruzam, dirigindo-se ao lado oposto da cabeça. Estas fibras
do cruzamento juntam-se com fibras das partes temporais da retina oposta e se projetam até
os corpos (ou núcleos) geniculados laterais (CGL) em cada hemisfério.
As seis camadas do CGL recebem entradas especializadas das fibras do nervo óptico
dos olhos de cada hemisfério. As duas camadas mais inferiores, chamadas de camadas
magno-celulares, tomam as entradas principais da retina periférica, onde células
ganglionares espectralmente não-oponentes com grandes campos receptores e
características temporais transitórias são dominantes. As camadas parvo-celulares restantes
tomam entradas principalmente da região da fóvea onde as células espectralmente
oponentes com campos receptores pequenos e características estacionárias são dominantes.
As diferenças impressionantes nas propriedades funcionais das camadas magno e parvo-
celulares sugerem que os olhos podem, de fato, servir como dois sistemas de
processamento visuais. Um é de resposta rápida, sistema acromático, sensível ao
movimento, mas com baixa definição espacial. O outro é de resposta lenta, sistema
tricromático, relativamente insensível ao movimento, mas com alta resolução.

18
Fig 1.11 Trajetória das vias visuais (figura adaptada de Color Atlas of Neuroscience Greenstein, 2000, pag. 283)
Do CGL, as fibras projetam-se ao córtex visual. O córtex visual primário é conhecido
também como V1, área 17 de Brodmann ou córtex estriado. As células no córtex visual têm
sensibilidades distintas. Algumas células são sensíveis a um alvo de cor ou contraste, mas
não a forma nem a movimento. Outras são seletivas à orientação, mas insensíveis a cor e a
movimento. Além disso, outras células são seletivas à orientação e ao sentido do
movimento, mas não a cor. A especificidade funcional observada em V1 e em outras áreas
do córtex visual conduziram à especulação que o sistema visual está dividido em sistemas
de identificação e de localização. Diversos estudos de caso sustentam esta conjetura,
mostrando que danos cerebrais podem produzir perdas em um tipo de função sem afetar a
outra.

19
Capítulo 2
Eletroencefalografia
2.1 Introdução
O cérebro é um órgão complexo. A especialização biológica dele é receber, analisar,
processar, armazenar, recuperar, sintetizar e enviar informações, desde e até os mais
distantes ou sutis locais do corpo humano. Estas informações são transmitidas entre as
unidades funcionais do cérebro, os neurônios, por processos eletroquímicos. A cada
instante, o somatório das diferenças de energia elétrica entre os neurônios emerge da caixa
óssea que contém o cérebro, o crânio, e determina diferenças de potencial elétrico entre
pontos diferentes do escalpo ou entre estes e um ponto neutro de referência. A
eletroencefalografia reflete as populações de oscilações sincronizadas e dessincronizadas da
atividade elétrica em curso no cérebro, principalmente atividade dendrítico-cortical. As
diferentes bandas de freqüência da eletroencefalografia (delta, teta, alfa, beta)
proporcionam um índice de diferentes níveis de excitação e de ativação [Gordon, 1999].
Os potenciais relacionados a eventos (ERPs) - potenciais exógenos e endógenos -
refletem os potenciais elétricos transientes fixados no tempo (time-locked) ante a repetida
apresentação de estímulos discretos. Consistem de uma atividade elétrica produzida pelo
cérebro em resposta a um estímulo sensorial associado a tarefas motoras, cognitivas ou
psicofisiológicas. Essas são ondas discretas que estão relacionadas com algum estímulo ou
evento motor, e que se encontram presentes no eletroencefalograma (EEG) [Durand-Rivera,
2004; Van Boxtel, 1998; Handy, 2004].
As medidas de eletroencefalografia e dos ERP são facilmente accessíveis e não
invasivas, e os potenciais são registrados através de eletrodos colocados no couro cabeludo
(relativo a um ponto de referência na cabeça). As correntes elétricas medidas são

20
principalmente geradas perpendicularmente ao crânio e parcialmente amortecidas pelas
resistências do crânio e do couro cabeludo [Gordon, 1999, Van Boxtel, 1998; Handy,
2004].
2.2 O Eletroencefalograma
O Eletroencefalograma (EEG) é um registro da atividade elétrica cerebral, o qual é
feito com ajuda de um arranjo de eletrodos colocados sobre o escalpo [Thakor, 2004]. Estes
eletrodos de metal colocados sobre o couro cabeludo transformam a atividade elétrica em
padrões comumente chamados de ondas ou ritmos cerebrais, os quais são formas de ondas
recorrentes de forma e duração semelhantes [Nunez, 2005]. Após a amplificação, o sinal é
gravado num formato digital ou gráfico (transmitindo-se a uma tela de um computador).
O primeiro EEG humano foi registrado por Hans Berger, em 1929. Berger foi capaz
de registrar os traços de EEG de um canal fronto-occipital usando a técnica de registro
bipolar. Os registros duraram somente uns minutos e foram feitos em papel fotográfico com
um galvanômetro de dupla bobina [Niedermeyer, 1999].
2.2.1 Sinais elétricos do cérebro
O sinal EEG reflete o somatório espaço-temporal dos potenciais pós-sinápticos na
vizinhança de um eletrodo de registro [Medvedev, 2002]. O EEG do escalpo proporciona
uma medida em grande escala da função dinâmica neocortical. Um único eletrodo fornece
uma média estimada da ação sináptica de uma massa tecidual contendo entre 10 milhões e
1 bilhão de neurônios [Nunez, 2000]. A base de todos os sinais bioelétricos é a
transformação dos sinais não elétricos para um sinal elétrico na membrana celular ativa.
Um processo químico, que envolve o fluxo de entrada e saída de íons de potássio (K+) e
sódio (Na+), inicia-se com o potencial de ação (PA) (Fig. 2.1). Estes sinais chegam ao
neurônio através de um mecanismo químico que ocorre nos dendritos. Se um neurônio
dispara, o potencial de ação viaja através do axônio ao terminal extremo deste para ativar

21
outros neurônios. A junção entre um axônio e o adjacente chama-se sinapse. Esta é o canal
que o neurônio usa para comunicar-se com outro. A informação desde o corpo celular viaja
através do axônio como um potencial de ação elétrico, logo alcança o fim de um axônio
pré-sináptico, liberando as chamadas moléculas de neurotransmissores no espaço sináptico,
que atravessam este para a próxima célula nervosa ou célula muscular [Malmivuo, 1995].
A parte da sinapse sobre o axônio é chamado de terminal pré-sináptico, e a parte do
lado receptor é chamado de terminal pós-sináptico. O espaço entre estes dois lados, a fenda
sináptica, e suas propriedades químicas são responsáveis para que a informação na sinapse
viaje numa única direção [Malmivuo, 1995].
Fig 2.1 A célula nervosa (figura retirada de
http://infook.blogspot.com/2007/08/como-funciona-o-crebro.html)
2.2.2 Fontes neurais do sinal EEG
O cérebro é um sistema extremamente complexo, realizando constantemente
transferência e processamento de informação. Os trabalhos do sistema neural são realizados
através das interações entre os conjuntos grandes dos neurônios no sistema nervoso central
(CNS) e no sistema neural periférico. Ao nível celular, os neurônios transferem e

22
processam a informação através dos potenciais de ação e dos disparos neurais (conhecidos
também como spikes). Quando este tipo de atividade elétrica se transmite à superfície do
córtex e à superfície do escalpo, nós podemos registrar isto com o EEG.
Os potencias de ação são demasiado rápidos (duram somente entre 1 e 2 ms) e são
incapazes de se somarem de forma coerente no intervalo de tempo de EEG com suficiência
para serem diretamente observados no EEG. No entanto, os PA causam potenciais pós-
sinápticos (PPS) em diferentes células. Comparados com os PA, os PPS são muito mais
demorados ou longos, aproximadamente entre 10-250 ms [Lopes da Silva , 1999]. Os PPS
são a fonte primária dos campos potenciais extracelulares, i.e., os potenciais que são
medíveis com métodos não invasivos [Speckmann, 1999]. Eles podem se somar espacial e
temporalmente, o que torna possível registrá-los usando EEG. Os ritmos de EEG
registrados no couro cabeludo são o resultado do efeito da soma de muitos potenciais pós-
sinápticos excitatórios e inibitórios (EPSPs e IPSPs) produzidos na capa piramidal do
córtex cerebral.
Nos seres humanos, o tálamo poderia ser o sítio principal da origem das atividades de
EEG (bandas alfa e beta) [Hughes, 1999]. As oscilações talâmicas ativam o disparo dos
neurônios corticais. A despolarização (principalmente na camada IV) cria um dipolo com
negatividade na camada IV e positividade nas camadas mais superficiais. Os eletrodos do
couro cabeludo detectam um pequeno, mas perceptível potencial de campo distante, que
representa as flutuações potenciais somadas [Misulis, 1997]. Em condições clínicas e
experimentais, EEG é o registro da diferença de potencial entre dois eletrodos (EEG
bipolar) ou um eletrodo do couro cabeludo e outro de ouvido, como referência (EEG
unipolar). Os eletrodos do couro cabeludo não podem detectar cargas fora de 6 cm2 da área
superficial cortical correspondente, e a profundidade eficaz do registro é de vários
milímetros.
Severas limitações afetam a detecção dos sinais EEG. Primeiro os potenciais têm
que ser sincrônicos. Segundo, os neurônios têm que estar alinhados para os potenciais se
somarem, pois de outro modo os potenciais se cancelam uns com outros (Fig. 2.2)
[Niedermeyer, 2004].

23
Fig 2.2. Geração de potenciais sobre o escalpo pela somatória de correntes.
2.2.3 A natureza do EEG
Em primeira aproximação, o EEG é gerado por uma grande população de neurônios
piramidais (Fig. 2.2) orientados perpendicularmente ao córtex cerebral, localizados nas
camadas II, III, V e VI (Fig. 1.6). Estes neurônios piramidais separam espacialmente os
inputs excitatórios e inibitórios sobre sua superfície, sendo os potenciais pós-sinápticos
excitatórios e inibitórios que, somando-se sobre os neurônios piramidais, dão origem ao
sinal EEG [Ebersole, 2003].
Fig 2.3 Representação esquemática de um neurônio piramidal cortical (Figura retirada de: Epilepsy as a Dynamic Disease, J. Milton et al, editors, capítulo 5, p. 55).

24
Para melhor entender a natureza deste fenômeno, consideraremos a contribuição da
atividade de um único neurônio para o EEG, examinado um circuito cortical simplificado e
alguns princípios elétricos básicos (Fig. 2.3). Neurônios piramidais são os neurônios de
principal projeção sobre o córtex. Os dendritos apicais das células piramidais, os quais
estão situados perpendicularmente à superfície celular, recebem vários inputs sinápticos. A
atividade sináptica das células piramidais é a principal fonte da atividade EEG.
Para entender a contribuição de um único neurônio para o EEG, considere o fluxo
de corrente produzido por um EPSP (excitatory postsinaptic potencial) sobre o dendrito
apical de um neurônio piramidal cortical (Fig. 2.4). As correntes fluem no dendrito no local
da geração do EPSP, criando uma corrente de vazamento. Então, deveria completar um laço
fluindo abaixo do dendrito e retirar-se através da membrana para outros sítios, criando uma
fonte de corrente. A intensidade da voltagem criada pela corrente sináptica é dada
aproximadamente pela Lei de Ohm (V = IR, V: voltagem, I: corrente e R: Resistência). A
resistência da membrana (Rm) é muito maior do que a da solução salina, que constitui o
meio extracelular (Re). Por esse motivo, a voltagem registrada através da membrana com
um eletrodo intracelular (1) é também maior que num eletrodo extracelular (2).
Fig. 2.4 Padrão do fluxo de corrente elétrica para um EPSP sobre um dendrito apical de um
neurônio piramidal no córtex cerebral (Figura extraída de Principles of Neural Science
de Kandel, 2000, Fourth edition, chapter 46, pag. 914)

25
No lugar da geração do EPSP, o eletrodo extracelular detecta correntes fluindo longe
do eletrodo dentro do citoplasma como uma deflexão para baixo. No entanto, um eletrodo
extracelular perto da fonte tem polaridade oposta ( compare os eletrodos 2 e 3, Fig. 2.4). A
situação é inversa se o lugar da geração do EPSP está sobre um dendrito proximal. No
córtex, inputs excitatórios do hemisfério contralateral contactam (interagem com) os
neurônios piramidais, principalmente sobre as partes distais do dendrito nas camadas 2 e 3.
No entanto, inputs talamo-corticais terminam na camada 4. A atividade medida na
superfície do eletrodo do EEG terá polaridades opostas para estes dois inputs (Fig. 2.5)
[Kandel, 2000].
Fig. 2.5 Polaridade dependente da posição da sinapse
(http://www.acm.org/conferences/sac/sac2000/Proceed/FinalPapers/BC-07/)
2.2.4 Potenciais relacionados a eventos (ERP)
Estudos de EEG geralmente são focados ou sobre ritmos espontâneos cerebrais ou
sobre potenciais relacionados a eventos, ERPs (ou também chamados potenciais evocados
EP). A banda de freqüência do sinal EEG está numa faixa que se estende aproximadamente
de 0-70 Hz. Enquanto os EP e ERP têm amplitudes próximas de dezenas de microvolts

26
(µV), os ritmos espontâneos podem atingir centenas de microvolts [ Handy, 2004; Thakor,
2004].
Os ritmos espontâneos incluem descargas epilépticas e outros eventos que ocorrem
aperiodicamente, assim como ritmos periódicos definidos [Cooper et al., 1980]. Por
exemplo, ondas alfa oscilando entre 8 e 13 Hz são geradas na maioria de adultos sadios
quando eles têm seus olhos fechados. Estas ondas são observadas na parte posterior do
cérebro, que corresponde às áreas visuais [Nunez, 2001].
Os EP e ERP ocorrem quando o cérebro responde a estímulos, os quais podem ser
de origem exógena (EP) ou endógena (ERP). No entanto, a resposta de um evento
individual num sinal EEG é muito pequena para ser confiavelmente detectada, devido ao
ruído e ritmos espontâneos do EEG. Conseqüentemente, o estímulo é repetido várias vezes,
entre 100-200, e os sinais EEG são promediados com respeito ao início do estímulo para
recuperar o EP obtido.
Os ERP podem ser obtidos por diversos tipos de estímulos, por exemplo um flash de
luz. Este tipo de potencial é chamado de potencial evocado visual (VEP). Um exemplo de
estímulo auditivo é o chamado potencial evocado auditivo (AEP), apresentado como um
estímulo sonoro [Cooper,1980].
2.2.5 Resolução espacial e temporal do EEG
No domínio do tempo, o sinal EEG é visto como uma função do tempo, o que permite
uma análise visual da forma, amplitude e período das ondas nele encontradas. É a forma
clássica de registro e leitura do EEG, desde Hans Berger, permanecendo como base
indispensável para qualquer investigação de EEG até os dias de hoje.
No domínio da freqüência, o sinal EEG é visto como uma função da freqüência, o que
permite uma análise quantitativa do espectro de freqüências de que é composto. É uma
nova forma de leitura do EEG, só tornada possível pelo computador, devido ao grande
número de cálculos necessários em curto tempo.
O sinal EEG tem baixa resolução espacial, mas ele tem excelente resolução
temporal da ordem de milissegundos. No entanto, novas técnicas de imageamento cerebral,

27
tais como CT e MRI, podem prover alta resolução espacial. Assim, a combinação do EEG
com outras técnicas de imageamento cerebral pode oferecer ambas resoluções temporal e
espacial (Fig. 2.6).
Fig 2.6. Diagrama comparando a performance espacial e temporal de quatro técnicas de neuroimagem
minimamente invasivas. A diagonal “inclinada” representa o melhoramento na resolução espacial com o
tempo crescente da medida, uma característica das quatro modalidades [Strangman, 2002].
2.2.6 Os ritmos do EEG
Os ritmos cerebrais são formas de ondas eletromagnéticas produzidas pela
atividade elétrica das células cerebrais (neurônios). Sabe-se que as ondas cerebrais mudam
freqüentemente em função da atividade elétrica dos neurônios. Essas alterações estão
relacionadas com mudanças de estados de consciência (concentração, relaxamento,
meditação, etc.).
Cada indivíduo tem a sua própria característica de atividade de ritmos cerebrais,
possui um padrão, conforme as suas atividades diárias. Com a interação de determinadas
situações físicas ou emocionais, esses ritmos podem variar. Os principais ritmos cerebrais
são:

28
Delta
Delta é a mais baixa de todas as freqüências de ondas cerebrais. Está associada com
o sono profundo, e também ocorre na infância. Algumas freqüências na faixa Delta liberam
o hormônio do crescimento humano (HGH), que é muito benéfico para a regeneração
celular. Acredita-se que se originam no córtex e no tálamo. A faixa das ondas Delta está
entre 0-4 Hz, e sua amplitude varia entre 10-50µV.
Teta
São ondas rítmicas ou arrítmicas. Relaciona-se a processos de criatividade e
memória. O estado Teta propicia flashes de imagens do inconsciente, criatividade e acesso
a memórias há muito tempo esquecidas. Quando um indivíduo entra num estado sonolento
a predominância de ondas teta aumenta. A excessiva presença de ondas Teta está
relacionada a um déficit de atenção. A faixa das ondas Teta está entre 4-7 Hz e sua
amplitude varia entre 50-100µV.
Alfa
São encontradas em quase todas as pessoas adultas normais quando elas estão
acordadas o no estado de repouso. Relaciona-se a processos de relaxamento, visualização,
meditação. Quando alguém está relaxado, sua atividade cerebral baixa do padrão beta, que
é rápido, para as ondas alfa que são mais lentas, e experimenta uma sensação de paz e
bem-estar.
O estado Alfa funciona como um portal para estados de consciência mais profundos.
A faixa de ondas Alfa está entre 7-13 Hz e sua amplitude varia entre 15-45µV [Evans,
2009; Kropotov, 2009]. Esta faixa de freqüência teoricamente é gerada pelo córtex
principalmente, mas também se discute e assume-se a possibilidade de estarem envolvidos
os sistemas: corticotalâmico e corticocortical. “Nenhuma teoria neurofisiológica ou
psicofisiológica do ritmo alfa têm ainda uma aceitação geral. Ainda existem incertezas

29
sobre a origem e o significado psicofisiológico deste notável fenômeno. No entanto, nossas
introspecções da natureza do ritmo alfa têm-se aprofundado” [Niedermeyer, 2005].
Beta
Relaciona-se a processos de atenção, concentração, cognição. No estado beta, os
neurônios transmitem as informações rapidamente, permitindo atingir estados de
concentração. O ritmo beta é usado por terapeutas de biofeedback para tratar um problema
de aprendizagem e concentração chamado de transtorno de déficit de atenção (TDA)
[Evans, 2009; Kropotov, 2009].
A faixa de ondas Beta está entre 13-30 Hz e sua amplitude varia entre 5-30µV. O
estado Beta está associado com concentração, atenção aumentada, melhor acuidade visual e
coordenação.
As freqüências Beta 13 Hz e 18 Hz, Gama 40 Hz (30-80 Hz, 3-10µV) e Alta Gama
(80-150 Hz) atuam em funções cognitivas complexas.
2.2.7 Aquisição de dados do EEG
Para realizar a aquisição e a análise quantitativa do EEG (qEEG), os eletrodos são
posicionados em pontos definidos sobre o couro cabeludo. Durante a fase de aquisição de
dados, cada eletrodo coleta sinais elétricos do CNS. O registro do sistema inclui (I)
eletrodos e head stage, (II) pré-processamento e qEEG, e (III) armazenagem de dados e
resultados (Fig 2.7):
1. Eletrodos: O eletrodo do EEG é um sensor de potencial elétrico.
2. Aquisição e amplificação: A largura de banda do sinal de EEG é 0,5 – 100Hz em
freqüência (mais importantes as menores de 30Hz) e as amplitudes típicas estão no
intervalo de 10-300 µV.
3. Filtragem: Um EEG de rotina é geralmente amostrado a uma freqüência de
aproximadamente 250Hz ou 500Hz, o qual teoricamente cobre a banda de 0 a 125Hz,

30
podendo ser um pouco maior para obter uma melhor resolução do sinal. Eventualmente o
sistema EEG pode necessitar um filtro especial para remover os artefatos da linha de
potência (50 ou 60 Hz).
4. Armazenagem: Antigamente o EEG era registrado num papel. Atualmente, se
armazenam os dados do registro EEG num computador para sua posterior análise.
Fig 2.7 Diagramas das diferentes etapas de um registro EEG e sistema quantitativo [Thakor,2004].
Um exame EEG básico dura ao redor de 45 minutos, com um raio de ação de 30 a 90
minutos. O EEG pode ser utilizado para avaliar pessoas com crises de epilepsia, confusão,
lesões na cabeça e outras condições que podem surgir por causa de uma anormalidade no
cérebro. O EEG pode ajudar a diagnosticar certos tipos de doenças do cérebro que levem a
piorar a debilidade mental (demência) ou alguma disfunção do cérebro, por exemplo, a
encefalopatia, causada por doença severa do fígado ou dos rins.
Na maioria dos casos, não há preparação especial necessária. No entanto, em algumas
pessoas, um EEG dá melhores resultados se o registro é obtido após um período de
privação do sono. Isto torna mais provável que ocorram crises epilépticas ou outras

31
anormalidades. Em caso de necessidade, o médico dará instruções específicas sobre como e
quando limitar o sono antes do EEG.
Um dos mais significativos problemas da implementação do EEG é a avaliação e
quantificação de ondas. O método clínico convencional de observação da forma de onda é
subjetivo e tedioso, razão pela qual os resultados dependem da experiência e habilidade dos
técnicos. O desenvolvimento do EEG quantitativo foi motivado pela necessidade de
medidas objetivas assim como pelo grau de automatização do sistema.
2.2.8 Propriedades do EEG
O sinal EEG é complexo. A complexidade do EEG tem a sua origem no intrincado
sistema neural. Tradicionalmente, o EEG espontâneo é caracterizado como um processo
estocástico linear com grandes semelhanças ao ruído.
Do ponto de vista do processamento de sinais, o EEG tem as seguintes propriedades:
(a) É ruidoso e pseudo-estático: O EEG varia usualmente entre 10-300µV, sendo
facilmente afetado pelos ruídos fisiológico e elétrico. Sempre um registro de EEG mostra
um alto grau de aleatoriedade e não-estacionaridade;
(b) Variabilidade no tempo e não-estacionaridade: O EEG é um processo não
estacionário, i.e. varia com os estados fisiológicos. As formas das ondas incluem um
complexo de ondas regulares, spikes/polyspikes irregulares, ou spindles/polyspindles. Em
condições patológicas, tais como em crises de epilepsia, o EEG pode mostrar evidentes
singularidades ou não-estacionaridade. Na prática, considera-se o EEG como um processo
estacionário apenas num período relativamente curto de tempo (~3.5s para um EEG
espontâneo de rotina);
(c) alta não-linearidade: embora os modelos lineares do EEG tenham ainda um papel
importante no diagnóstico e na análise do EEG, o EEG é um processo não-linear.

32
2.2.9 Aplicações do EEG
Por causa dos métodos experimentais comumente adotados nos laboratórios, os dados
obtidos (sem refinar) são usualmente contaminados com várias fontes de ruído e artefatos.
O pré-processamento do EEG lida principalmente com estes artefatos e interferências. A
atividade neural varia no nível de 10 até 300 µV, assim é facilmente afetada pelos vários
fatores internos e externos. Os artefatos mais comuns incluem (a) movimento dos pacientes
durante o registro, tais como movimento dos olhos ou queixo, língua, ou movimento do
corpo ou cabeça; (b) artefatos de músculo, e (c) ondas de pulso ou batimentos cardíacos.
Fisiologicamente, sabe-se que as características das ondas elétricas cerebrais variam
conforme o funcionamento (situação funcional) do órgão. As maiores variações se
observam entre os estados de vigília, ou seja, entre o estar acordado, dormindo, sonolento,
ou em coma.
O eletroencefalograma é usado em neurologia e psiquiatria principalmente para
auxiliar no diagnóstico de doenças do cérebro, tais como a epilepsia, as desordens do sono
e alguns tipos de tumores cerebrais. De 1930 até bem pouco tempo atrás, a
eletroencefalografia esteve quase estagnada e com aplicação médica perdendo terreno
seguidamente para outros métodos de diagnóstico e de exames. Nas últimas décadas,
entretanto, a informática foi acoplada ao método eletroencefalográfico e novos horizontes
foram descobertos.
Uma dessas novas aplicações do EEG é tentar localizar com exatidão os focos
epilépticos ou tumores cerebrais. Os focos epilépticos são pequenas regiões no cérebro
onde a atividade elétrica se apresenta anormal. Pela observação dos traçados dos canais, o
neurologista que interpreta o EEG é capaz de deduzir onde exatamente esta anormalidade
está situada. Entretanto, a interpretação pessoal dos traçados é muito difícil quando o
número de canais é grande ou a natureza da anomalia é complexa. Vem daí a necessidade
de se acoplar ao processo de análise de sinal as ferramentas da informática. Com isso, foi
possível a elaboração de um mapeamento cerebral eletricamente determinado [Niedermeyer
et al., 2004].

33
A informática, através de softwares próprios e de cálculos matemáticos complexos,
tem sido usada para realizar mapeamentos cerebrais coloridos. Este tipo de exame é
chamado de EEG quantitativo (qEEG), em contrapartida à avaliação qualitativa da
eletroencefalografia tradicional. A topografia cerebral oferecida pelo EEG foi tornada
possível devido ao grande número de eletrodos colocados na cabeça e da resolução dos
computadores. O mapeamento cerebral gerado pelos computadores avalia a quantidade da
atividade elétrica de uma determinada região através das diversas tonalidades de cor. O
EEG Quantitativo proporciona uma avaliação mais precisa da atividade cerebral, dando
uma visão gráfica mais acurada da localização de alterações elétricas.
A informática também proporciona animações dinâmicas das imagens cerebrais,
facilitando o estudo da função cerebral e do cérebro em ação. Atualmente, a principal
indicação do EEG Quantitativo é determinar a localização precisa de tumores cerebrais,
bem como a localização precisa de doenças focais do cérebro, incluindo entre elas a
epilepsia, as alterações vasculares e derrames [Thakor, 2004].
Em psiquiatria, o EEG Quantitativo tem sido usado para estabelecer diferenças entre
vários diagnósticos, tais como a hiperatividade e os distúrbios da atenção em crianças, as
demências (senis ou não), a atrofia cerebral, a esquizofrenia, e até alguns casos de
depressão.
Em neurologia, o EEG Quantitativo, além de ser utilizado na determinação de focos
epilépticos, é útil na monitoração da abstinência de drogas, em infecções do cérebro, nos
estados de coma, de narcolepsia e no acompanhamento pós-operatório de pacientes que
foram submetidos à cirurgia cerebral.
O futuro do EEG Quantitativo será proporcional ao futuro acoplamento de métodos
digitais de análise de sinais e de processamento de imagens pelos computadores futuros,
assim como o acoplamento de métodos não lineares para series temporais estocásticas que
permitam explorar mais detalhes [Thakor, 2004].

34
2.3 O equipamento de EEG
2.3.1 Eletrodos
A montagem dos eletrodos do EEG pode ser bipolar ou unipolar. Numa
montagem bipolar, a diferença de potencial entre um par de eletrodos é usada. Nas
montagens unipolares, o potencial de cada eletrodo é comparado com um outro eletrodo
neutro (eletrodo de referência) ou com a média de todos os eletrodos (média comum)
[Medvedev, 2002]. A exibição do curso temporal de um só eletrodo é também referida
como canal.
Eletrodos de superfície são anéis metálicos, com diâmetro de aproximadamente 1
cm. Quando usamos eletrodos de superfície, o material dos eletrodos é importante. O
eletrodo deve proporcionar valores estáveis, i.e., as propriedades elétricas do mesmo não
devem mudar no tempo. Além disso, o eletrodo deve proporcionar um bom e permanente
contato elétrico, e que o faça de forma insensível aos movimentos do eletrodo. Eletrodos de
superfície bons são de metais nobres (ouro, platino, prata) e eletrodos revestidos com sal,
como Ag-AgCl.
2.3.2 Amplificadores
O sinal do EEG de escalpo vai desde o eletrodo através de um fio até o amplificador.
Os fios estão expostos a distúrbios eletromagnéticos, assim os comprimentos destes devem
conservar-se mínimos. Do amplificador, o sinal é transferido para a transformação posterior
usando um cabo blindado, como um cabo coaxial, ou, como se faz mais recentemente, via
fibra óptica.
Para reduzir os efeitos de distúrbios de saída, o amplificador diferencial é,
freqüentemente, usado para amplificar os sinais do EEG. O amplificador diferencial
somente amplifica a diferença entre dois eletrodos, a fim de eliminar o ruído comum (com
freqüência de 50-60 Hz). Na prática, isto não é totalmente possível.

35
2.3.3 Configuração dos eletrodos
A configuração de eletrodos mais amplamente usada é a standard internacional,
chamada Sistema Internacional 10-20. Neste sistema, a posição é determinada por quatro
pontos de referência: o nasion (na profundidade do topo do nariz), o inion (na base
posterior do crânio na cabeça) e os outros nas bordas dos canais auditivos de ambos
ouvidos. Após medir as distâncias entre esses pontos (linha central e ouvido-ouvido), as
linhas se dividem até o 10% e em intervalos de 20%, e esta informação é usada para colocar
outros eletrodos. A Figura 2.8. mostra este sistema na prática, mostrando também a
nomenclatura dos eletrodos. A vantagem de se usar o sistema 10-20 é que os resultados de
diversos laboratórios e de temas relacionados são comparáveis.
(a) (b)
Fig 2.8. Posicionamento dos eletrodos no Sistema Internacional 10-20, para registros EEG
[Malmivuo, 2008]. A=auricular, Pg=nasofaríngeo, C=central,P=parietal, T=temporal,F=frontal,
Fp=fronto-polar e O=occipital

36
2.4 Problemas do EEG
O EEG tem uma excelente resolução temporal [Strangman, 2002], da ordem de 1ms,
mas sua resolução espacial é muito pobre, tornando-o assim uma fonte de estimação
inadequada. Mas, a precisão espacial pode ser melhorada incrementando o número de
eletrodos e usando uma estimação laplaciana. Este sistema de usar o EEG pode atingir a
precisão espacial do magnetoencefalograma (MEG). Esta melhora tem seus limites porque
os tecidos subjacentes do escalpo atenuam e apagam (velam) o sinal.
Modelos teóricos e dados experimentais de EEG têm sido utilizados para quantificar a
resolução espacial dos registros da superfície do escalpo no EEG. Os resultados sugerem
que os eletrodos do escalpo são geralmente sensíveis à atividade correlacionada sobre
grandes áreas de superfície neocortical, com pequenas contribuições de fontes mais
profundas [Srinivasan, 1999]. Os estudos de resolução espacial do EEG realizados nos
últimos anos levam a concluir que a resolução espacial do sistema de eletrodos 10-20 é
insuficiente para as atuais pesquisas cerebrais. No presente, existem sistemas de medidas de
256 eletrodos ou mais, existindo o interesse geral para avaliar o incremento da resolução
espacial com estes sistemas. Baseado na referência [Ryynänen, 2004] se afirma que, com
sistemas mais densos, se obtém uma melhor resolução espacial. Assim mesmo, as
vantagens de um equipamento EEG de alta resolução são grandemente dependentes da
quantidade de ruído na medida.

37
Capítulo 3
Espectroscopia no Infravermelho Próximo
3.1 Introdução
Nos últimos anos, têm surgido muitas técnicas neurofisiológicas de imageamento não-
invasivas que nos permitem “penetrar” e “olhar” no cérebro, contribuindo muito ao nosso
entendimento da ativação funcional cerebral. Com essas técnicas podemos explorar as
respostas funcionais cerebrais ao nível metabólico e neurovascular, o que nos dá uma idéia
da dinâmica subjacente durante a ativação.
As técnicas utilizadas, tais como a eletroencefalografia (EEG), potenciais relacionados
a eventos (ERPs) e magnetoeletroencefalografia (MEG), medem respostas
eletrofisiológicas relativas à atividade neural com excelente resolução temporal, mas com
uma pobre resolução espacial. A ressonância magnética funcional (fMRI: functional
Magnetic Resonance Imaging), tomografia por emissão de pósitrons (PET: positron
emission tomography), arterial spin labeling (ASL) e outras técnicas medem respostas
hemodinâmicas e metabólicas, com excelente resolução espacial (por exemplo o fMRI, cuja
resolução espacial é estímulo de 1 mm3). Esta resolução espacial, porém, é alcançada às
custas de uma perda de resolução temporal e de informação neural direta.
Em meio a esse cenário, aparece a espectroscopia no infravermelho próximo (NIRS:
Near InfraRed Spectroscopy), a qual possui atributos que a tornam viável para estudos de
neuroimageamento do córtex cerebral humano e de outros seres vivos. NIRS é uma técnica
óptica segura, não-invasiva, de baixo custo, portátil, e pode trabalhar sem fios [Bunce,

38
2006]. Ela aproveita mudanças das propriedades ópticas dos tecidos, utilizando níveis
seguros de luz, cujos comprimentos de onda variam aproximadamente entre 650-900nm.
Esta técnica mede mudanças neurofisiológicas através da penetração dessa luz no córtex,
até alguns centímetros, permitindo a interação com a oxi- e desoxi-hemoglobina do fluxo
sangüíneo cerebral.
A técnica NIRS tem se convertido em uma tecnologia que pode lançar mais luz sobre o
funcionamento cerebral, provendo informação sobre a oxigenação no tecido de forma não-
invasiva, sem o uso de energia ionizante. É uma técnica com potencial para o diagnóstico e
tratamento da depressão, esquizofrenia e doença de Alzheimer, assim como na reabilitação
de Stroke.[Boas, 2004]. NIRS tem se aplicado ao monitoramento não invasivo da
oxigenação cerebral em crianças recém nascidas [Cope, 1988; Cope, 1991; Hebden, 2003].
A técnica NIRS, como tecnologia emergente na área de imageamento cerebral, tem
sido empregada para diversos estudos em neurociências e em neurodiagnóstico, em
particular, em alguns casos de epilepsia. Dentro deste tema, podemos destacar dois
trabalhos. Watanabe e colaboradores utilizaram um sistema NIRS de multicanais acoplado
a video-EEG para a avaliação pré-operatória de 32 pacientes com epilepsia resistente a
tratamento medicamentoso, a fim de aferir a sua eficácia em comparação com uma técnica
muito usada em medicina nuclear, o SPECT (single photon emission computed
tomography) [Watanabe et al, 2004]. Em um outro estudo, Buchheim e colaboradores
utilizaram NIRS ictal concomitantemente com telemetria através de video-EEG para
examinar três pacientes com crises de ausência [Buchheim et al., 2004]. Outros estudos
bem sucedidos como esses têm indicado o potencial para o uso desta técnica na área de
epilepsia.
Comparado com outros métodos de neuroimagem funcional não-invasivos, tais como
tomografia por emissão de pósitrons (PET), imagens de ressonância magnética funcional
(fMRI) e magnetoencefalografia (MEG), NIRS supera estas técnicas em (1) flexibilidade,
(2) especificidade bioquímica, e (3) alta sensibilidade para detecção de baixas
concentrações de substâncias. Além disso, (4) o uso da técnica NIRS pode se ampliar,
potencialmente permitindo a aquisição de imagens do metabolismo mitocondrial e até o
registro direto da atividade neuronal. Vamos, então, comentar cada um desses pontos.

39
(1) Com respeito à flexibilidade, fMRI, PET, e MEG requerem instrumentos de
enorme porte, que interferem com o monitoramento clínico dos pacientes. Além disso, é
impossível examinar situações normais do ponto de vista fisiológico tais como o estar em
pé ou caminhando. Indivíduos que não podem colaborar completamente (crianças, pessoas
com demência, e outros) têm dificuldades para serem examinados através desses
equipamentos. Em contraste, os métodos ópticos precisam apenas de uma conexão com luz
através de fibra óptica, algo semelhante à tecnologia do EEG. O grau muito mais elevado
de flexibilidade permite o uso de NIRS para estudos em pacientes adultos que não
colaboraram como o exame, assim como em bebês.
(2) Outro defeito, especialmente do fMRI, é a falta de uma relação claramente
definida do sinal observado (i.e. do sinal BOLD) com um parâmetro fisiológico ou
bioquímico quantificável. Métodos ópticos oferecem alta especificidade bioquímica do
sinal, e o método para quantificação de concentrações de oxigênio é mais direto (veja
abaixo considerações sobre a Lei de Beer-Lambert modificada).
(3) Os métodos ópticos são sensíveis a baixas concentrações de substâncias
utilizando métodos de fluorescência. Em teoria, a sensitividade pode chegar até o nível de
moléculas individuais [Harms et al., 2001]. A aplicação de NIRS para estudos não-
invasivos em humanos pode oferecer um potencial semelhante ao do PET, cuja
desvantagem óbvia é a necessidade de traçadores radioativos.
(4) Finalmente, as tecnologias existentes medem um sinal vascular, tal como em
fMRI e H2O-PET; um sinal metabólico intracelular, tal como em FDG-PET; ou um sinal
mais diretamente representativo da atividade neuronal, tal como em MEG ou EEG. Ao
produzir imagens de diferentes aspectos da função cerebral, as técnicas se diferenciam em
termos de resolução espacial e temporal. Alta resolução temporal é o domínio dos métodos
eletrofisiológicos (EEG/MEG). Por outro lado, excelente resolução espacial é lograda pelas
técnicas baseadas em hemodinâmica. A combinação das vantagens respectivas tem sido
pesquisada, no entanto, por métodos combinados. Contudo, os requisitos técnicos são
complexos e a relação sinal-ruído é mais baixa para os tais métodos. Métodos ópticos
oferecem a opção para monitorar respostas vascular, metabólica celular e neuronal.

40
Ademais, quando NIRS é utilizado em métodos combinados com fMRI, PET ou
MEG/EEG, ele não interfere com a biofísica dessas outras técnicas.
3.2 Conceitos básicos: absorção e Lei de Beer-Lambert
Quando a luz atinge uma amostra de matéria, o campo elétrico incidente induz um
movimento oscilatório nas cargas que a constituem. Na maior parte dos casos, são
induzidas colisões que aumentam a energia cinética das partículas envolvidas. Esta energia
de oscilação associada ao campo incidente é na sua maior parte dissipada como calor no
meio. A este processo chama-se absorção. Portanto, o efeito final da absorção é da
diminuição da intensidade do feixe de luz incidente que atravessa uma amostra [Bohren,
1983; Ishimaru,1978, Tuchin, 2007]. O coeficiente de absorção (mm-1) é definido pela
lei de Lambert-Bouguer como:
(3.1)
onde dI é a mudança infinitesimal da intensidade I que um feixe de luz sofre ao percorrer a
distância dx num meio com coeficiente de absorção . Da expressão acima, temos:
(3.2)
O coeficiente de absorção representa a probabilidade da absorção por unidade de
comprimento para um fóton dado. O comprimento de absorção é definido como o inverso
de e representa a distância necessária para que a intensidade do feixe incidente
diminua para 1/e da intensidade inicial. Expressando (3.2) em log de base 10 temos:
(3.3)

41
onde a constante K representa o coeficiente de extinção. Por outro lado, a absorbância da
amostra é definida como:
(3.4)
Portanto, neste caso os coeficientes de extinção e de absorção são equivalentes.
Em 1852, Beer achou uma relação linear que determina a relação entre e a
concentração C de um material diluído num meio não-absorvedor como:
(3.5)
onde é o coeficiente de absorção específico. Substituindo em (3.2), se obtém:
(3.6)
A equação acima é conhecida como Lei de Beer-Lambert. Esta lei é valida para um
meio não-espalhador, iluminado com um feixe monocromático e homogêneo [Tsuchiya,
2001]. Ela permite determinar quantitativamente a concentração de cromóforos
absorventes. Indica que a atenuação da luz (A) é proporcional à concentração da molécula
absorvente (C), onde (coeficiente extinção especifico) é um fator de proporcionalidade, e
pode ser expressa como:
(3.7)
Ambas as equações, (3.4) e (3.7), são equivalentes. Entretanto, a Eq. (3.7) supõe
concentrações infinitesimais e não estima o espalhamento. Esta suposição não ajuda à
espectroscopia de tecidos. O espalhamento aumenta o comprimento da trajetória da luz, a

42
distância real entre a fonte e o detector. Nos estudos cerebrais, o comprimento de caminho
médio da luz é estímulo de seis vezes a distância entre fonte e detector (Duncan, 1995).
Para levar em conta o comprimento do caminho da luz, que é maior que a distância fonte-
detector, modifica-se a Lei de Beer-Lambert e se introduz o DPF, differential pathlength
factor, B. Além disso, uma segunda modificação na Lei de Beer-Lambert é necessária já
que luz pode ser perdida devido ao espalhamento, i.e. há fótons que não atingem o detector.
Por este motivo, um termo G é introduzido [Cope, 1991; Toga, 2002; Boas, 2004]. Este
termo depende do tamanho do detector e da geometria do sistema. Tendo em conta estas
considerações, chegamos ao que se chama na literatura de Lei de Beer-Lambert modificada
(MBLL - Modified Beer-Lambert Law):
. (3.8)
Assumindo que B e G sejam dados, em forma diferencial, a MBLL é dada por:
(3.9)
que é a equação geralmente utilizada para se determinar variações de concentração em
tecidos biológicos através de técnicas ópticas.
O coeficiente de absorção de um meio pode-se dever a muitas substâncias
absorventes (cromóforos) todas misturadas. Os coeficientes de extinção individuais de cada
cromóforo representam sua absorção a uma concentração particular. Portanto, o coeficiente
de absorção de uma mistura de cromóforos pode-se expressar como uma combinação linear
das concentrações de cada cromóforo Cn com o seu respectivo coeficiente de extinção ϵn, µa
(λ) = Σ ϵn(λ)Cn.

43
3.3 Espalhamento da luz
O espalhamento da luz acontece quando, ao atravessar um dado material, esta encontra
heterogeneidades, que podem ser contínuas ou discretas. O problema do espalhamento de
luz por uma partícula qualquer não tem uma solução analítica exata geral. No entanto,
existem aproximações para vários limites da relação entre o comprimento de onda (ou
geometrias) e o diâmetro das partículas, soluções exatas para formas particulares e métodos
numéricos para formas arbitrárias [Ishimaru, 1978; Ishimaru, 1991]. Além disso, existem
dois regimes distintos: espalhamento simples e espalhamento múltiplo.
No espalhamento simples, assume-se que a luz é espalhada apenas uma vez. Em outras
palavras, a distância entre as partículas é grande o suficiente para garantir que a luz apenas
interage uma vez antes de sair do volume considerado. O espalhamento simples por
partículas de pequenos diâmetros quando comparado com o comprimento de onda, λ, é
também conhecido por espalhamento de Rayleigh. As soluções para este tipo de
espalhamento têm uma dependência do tipo 1/λ4 e são bem conhecidas. Este é o
mecanismo responsável pelo espalhamento da luz solar na atmosfera e que dá origem à cor
azul ao céu. No que diz respeito à determinação do campo espalhado por partículas da
mesma ordem de grandeza ou superiores ao comprimento de onda, a complexidade
aumenta bastante. Uma das teorias utilizadas para descrever este tipo de situação é a teoria
de Mie, que é válida apenas para partículas esféricas. A utilização das soluções exatas das
equações de Maxwell, e as condições de fronteira adequadas são utilizadas de maneira a
descrever os processos de espalhamento e de absorção [Bohren, 1983; Ishimaru, 1978]. O
tecido biológico é um meio altamente espalhador da luz, não é homogêneo e é constituído
de vários componentes que têm cada um suas propriedades ópticas individuais, de forma
que os fótons, ao atravessá-lo, percorrem um caminho aleatório [Tsuchiya, 2001].
O coeficiente de espalhamento, µs , que ora se introduz, descreve as propriedades de
espalhamento do meio. Este coeficiente de espalhamento é o produto de um número denso
de partículas espalhadoras e a seção transversal espalhadora destas partículas. Portanto, µs,
representa a probabilidade de um fóton ser espalhado por unidade de comprimento. Se o

44
meio é espalhador, os caminhos feitos pelos fótons passando através do tecido não são
diretos. Isto tem dois efeitos: a) não podemos detectar muito longe todos os fótons
emergentes, a menos que o detector pudesse coletar sobre todos os ângulos e em todos os
pontos da superfície do meio, e b) os fótons terão viajado distâncias variáveis através do
meio turvo.
Se o meio é absorvente (tecido biológico), as distâncias viajadas pelos fótons
(conhecidas como comprimentos de trajetória = pathlengths) serão atenuadas em
concordância com (3.2). As distâncias viajadas por cada fóton dependerão de quantos
eventos de espalhamento o fóton encontrar e isto será corrigido pelo fator de comprimento
de trajetória diferencial, DPF, já mencionado. O DPF é uma função do coeficiente de
espalhamento, do coeficiente de anisotropia (g), da absorção do meio e da geometria do
meio.
A quantificação da absorção óptica (i.e. as mudanças da atenuação da luz em meios
materiais altamente espalhadores) como resultado das mudanças nas concentrações dos
cromóforos presentes, tal como ocorre no tecido cerebral, pode ser computada pela Lei de
Beer Lambert modificada (MBLL), a qual fornece um simples e razoável modelo para
absorção da luz em meios altamente espalhadores [Boas, 2001; Stragman, 2003]. A MBLL
é a base da espectroscopia contínua de tecidos no infravermelho próximo [Kocsis, 2006]. A
forma diferencial da MBLL afirma que mudanças na atenuação da luz são proporcionais às
mudanças nas concentrações nos cromóforos no tecido [Villringer, 2002; Kocsis, 2006;
Kim, 2007]. Isso será detalhado mais adiante.
3.4 Métodos ópticos aplicados ao estudo cerebral
O uso de técnicas ópticas utilizando a região do infravermelho próximo cresceu
bastante na última década. Nos principais laboratórios de neuroimagem do mundo, se usa a
técnica NIRS. A interação da luz com o tecido biológico é o princípio básico desta técnica
emergente.
A região de comprimentos de onda referida como infravermelho próximo corresponde
a uma banda do espectro eletromagnético que se estende aproximadamente desde 780nm

45
até 2.5µm (12.820 – 4.000cm-1) [Hecht, 1998]. Esta faixa do espectro eletromagnético
possui energia suficiente para excitar harmônicos e combinações de vibrações moleculares
a níveis de energia maiores [Workman, 2008].
NIRS é uma medida da intensidade e comprimento de onda absorvida de radiação NIR
por uma amostra. NIRS fornece resultados rápidos, é um método não-destrutivo, e
apresenta simplicidade na preparação de amostras, sendo que a maior desvantagem da
técnica é provavelmente a baixa sensitividade as constituintes em baixas concentrações
[Burns, 2001; Skoog, 2002] (ver adiante observação sobre fluorescência e fosforescência).
3.4.1 Janela óptica para estudos não-invasivos
Como já foi observado, devido ao espalhamento pronunciado, a luz não viaja ao longo
de uma reta do emissor ao receptor, mas a trajetória dos fótons é representada pelo produto
da distância geométrica (d) e de um fator diferencial de trajetória de caminho (DPF). Este
fato tem que ser considerado quando realizamos medidas de concentração assim como ao
aplicar algoritmos de reconstrução de imagens.
Fig. 3.1. Posição Fonte-Detetor na coleta de dados NIRS
(extraído de Izzetoglu et al., 2007) .
Estudos em adultos têm sido realizados colocando-se o optodo fonte de luz diretamente

46
sobre o escalpo do indivíduo a uma distância de cerca de 3 cm do optodo receptor,
colocado também sobre a cabeça dele. A amostra de tecido cerebral examinada corresponde
a um volume de forma semilunar plana sob o escalpo do indivíduo e entre os dois optodos
(Fig. 3.1). Para o modelo de um sistema simples, os efeitos do espalhamento e absorção na
trajetória de “fótons individuais” é esquematizada e ilustrada na Fig. 3.2.
Fig. 3.2. Trajetória de “fótons infravermelhos individuais” na interação
luz-tecido biológico (extraído de Boas et al., 2001)
A profundidade de penetração da luz, no entanto, depende do comprimento de onda. A
profundidade de penetração da luz com comprimento de onda na faixa do visível (400-
700nm) é limitada pela alta absorção devido à hemoglobina. Para a faixa além dos 950nm,
a luz penetra pobremente no tecido uma vez que a água é um forte absorvedor nesta faixa
espectral. O intervalo entre 700-900nm, que é parte da faixa NIR, pode ser chamado de
“janela óptica” biológica, limitada pela absorção da hemoglobina e da água (Fig. 3.3).
Nesta faixa, podemos inclusive “olhar” através do cérebro de um humano adulto. A luz
penetra até o córtex, o que permite a realização de estudos não-invasivos do cérebro
humano. Esta janela óptica é usada para estudos não-invasivos na faixa do NIR.
Em geral, a energia (do fóton) a qual é tomada por uma molécula absorvente (i.e.
hemoglobina ou água) é convertida em energia térmica. No entanto, dependendo da
molécula, após certa demora, a emissão de luz com um comprimento de onda maior pode

47
ocorrer. Quando o atraso é muito menor que 10-8 s, o fenômeno é chamado de
fluorescência, quando este está entre 10-8 e 10-6 s é chamado fluorescência atrasada, e
quando é maior que 10-6 s, é chamado fosforescência. Baseados na fluorescência e
fosforescência, moléculas podem ser detectadas em baixas concentrações. A maioria dos
traçadores fluorescentes são excitados pela luz visível. No entanto, traçadores para estudos
na faixa NIR estão sendo desenhados, permitindo assim estudos de fluorescência não-
invasiva em humanos.
Há um outro parâmetro óptico que representa um caso especial de espalhamento da luz,
que ocorre quando a luz é espalhada por mudanças ligeiras nas freqüências do movimento
das partículas (efeito Doppler). Este deslocamento da freqüência depende da velocidade do
movimento da partícula, da direção do movimento, e do número de interações do
movimento da partícula. Assim o deslocamento Doppler oferece a oportunidade para
determinar o movimento das partículas no tecido. Este método tem sido bem implementado
através de laser Doppler fluxometry (LDF) para medidas do fluxo sanguíneo cerebral na
superfície do cérebro.
Figura 3.3. Espectro de absorção da oxihemoglobina, desoxihemoglina e água
(extraído de Izzetoglu et al., 2007) .

48
3.4.2 Absorvedores biológicos: absorção da água
O maior constituinte dos tecidos biológicos é a água, sendo que o conteúdo médio
desta no cérebro do neonato é de 90% [Fillerup, 1967] e no cérebro do adulto é de 80%
[Woodard, 1986; Bronzino, 2000; Vander, 1990]. Embora a água não seja muito absorvente
no intervalo de freqüências do NIRS, sua dependência com o comprimento de onda pode
afetar medidas espectroscópicas devido à sua abundância. A Fig. 3.4 mostra o espectro de
absorção da água na faixa do NIRS [Izzetoglu et al., 2007].
O fato de a água ser muito abundante no cérebro implica que qualquer banda de
absorção desta terá um grande efeito óptico. Dados sobre o espectro do coeficiente de
absorção da água são achados sob um amplo intervalo do espectro eletromagnético que vai
desde os 200nm até os 200µm, o qual inclui a região visível e infravermelha, como se
mostra na Fig. 3.4 [Hale et al, 1973].
Figura 3.4. Coeficiente de absorção da água (extraído de Hale et al., 1973) .
O coeficiente de absorção da água mostra uma tendência ao crescimento quando o
comprimento de onda cresce. Nos comprimentos de onda menores que 600nm e acima da

49
região ultravioleta, o coeficiente de extinção da água é menor que 0.001 cm-1 e é
insignificante. Entre os 600 nm até 1.35 µm, a água teria uma absorção mensurável (que
depende do número de centímetros do tecido cerebral iluminado e da relação sinal-ruído do
espectrofotômetro). Nos comprimentos de onda entre os 900nm e 1.35µm, as perdas serão
tão elevadas que medidas somente através de alguns centímetros do tecido cerebral são
possíveis.
3.4.3 Absorvedores biológicos: absorção da hemoglobina
Absorvedores dinâmicos são cromóforos cuja concentração varia dentro da faixa do
tempo das medidas do NIRS, sendo, portanto, os principais responsáveis pela geração das
mudanças nas intensidades vistas com esta técnica. A oxihemoglobina (HbO) e a
desoxihemoglobina (dHb) são fundamentais, no entanto outros, tais como hemoglobina
(Hi) e carboxihemoglobina (HbCO), não fornecem informação funcional, mas estão
presentes em quantidades desconhecidas, introduzindo erros na quantificação da
oxigenação e do volume sanguíneo.
A hemoglobina é o maior absorvedor dinâmico no tecido. Está presente no tecido
cerebral a uma concentração nominal ao redor de 84 µmol/litro [Vander, 1990].
Interpretando a hemoglobina como oxigenação sanguínea in vivo, podemos considerar
a soma das concentrações da oxi e desoxihemoglobina ([HbO] + [dHb]) como uma medida
do volume sanguíneo cerebral (CBV, do inglês cerebral blood volume), enquanto que a
diferença ([HbO] – [dHb]) pode ser interpretada como uma medida da saturação de
oxigênio sanguíneo cerebral (CBS, do inglês cerebral blood saturation) [Cope, 1991].
No intervalo entre 700 até 900 nm, temos a “janela óptica”, já mencionada, limitada
pela absorção da hemoglobina e da água. Nesta faixa de frequências de luz, pode-se “olhar”
através do crânio, pois os três absorvedores principais, HbO, dHb e água, apresentam uma
fraca absorção da luz [Villringer, 2002; Boas, 2001].
Os espectros da oxi e desoxigehemoglobina são mostrados na Fig. 3.3, onde podemos
observar o ponto isobéstico a (798 ± 1.5)nm, ponto que indica a presença de equilíbrio
entre as duas substâncias e que possuem idêntica absorvância.

50
As ações conjuntas da absorção da hemoglobina e o espalhamento da mielina dão
conta da profundidade da penetração da luz no tecido cerebral do adulto, sendo que esta é
dominada fortemente pelo grau de mielinização do tecido [Svaasand, 1983].
3.5 Propagação da luz em tecido biológico
Como vimos, na interação luz NIR-tecido, os fenômenos predominantes são os de
espalhamento e de absorção. Os fótons infravermelhos injetados experimentam milhares de
eventos de espalhamento elástico enquanto, algumas vezes, outros são absorvidos neste
processo. O padrão migratório de um fóton individual é parecido a um caminho randômico,
com cada trajetória do fóton composta de segmentos retilíneos e interrupções súbitas que
mudam randomicamente a direção de propagação dos fótons, ilustrado pela Fig.3.2.
Frequentemente, a trajetória retilínea é chamada de caminho livre médio dos fótons
migrantes. Este caminho, que é da ordem de 28µm, possui um tempo de propagação de
aproximadamente 0.13ps (Wang & Wu, 2007).
O conjunto das trajetórias individuais dos fótons, que são emitidos por uma fonte de luz
NIR e que chegam a um detector a alguns centímetros de distância, apresenta uma
formasemelhante à de uma banana, conforme ilustrado na Fig. 3.5.
Fig. 3.5. Modelo da banana, que representa a forma geral da trajetória
dos fótons infravermelhos (extraído de Bunce et al., 2006) .

51
Dado que é impossível monitorar individualmente as trajetórias dos fótons viajantes,
quantificamos o problema de transporte da luz no tecido em termos de observáveis
macroscópicos mais facilmente mensuráveis, tais como a densidade de energia do fóton
dentro da amostra. Portanto, uma modelagem desse transporte em termos de movimento
aleatório dos fótons entre as moléculas do meio torna-se mais adequada. Por outro lado, a
teoria de difusão é amplamente utilizada para descrever o transporte das partículas e ondas
em meios densos, turvos e randômicos. Neste modelo, somente as interações entre as
partículas de luz e o meio são levadas em conta, sendo desprezados os efeitos de
polarização. Uma completa descrição da propagação dos fótons no tecido é fornecida pela
equação de transporte radiativo (RTE; equação (3.10)) [Ishimaru, 1989], que é também
denominada equação de transporte de Boltzmann:
(3.10)
Nesta equação, é a radiância (número de fótons por unidade de volume) para
os fótons que se encontram na posição r, na direção s, no tempo t; v é a velocidade do fóton
no tecido, é o coeficiente de transporte (onde , é o coeficiente de
absorção, e , é o coeficiente de espalhamento), e é a fonte de luz na posição r e
tempo t na direção s; é a função de fase do espalhamento. A RTE (Eq. 3.10) é uma
equação de conservação de energia, que estabelece que, para um qualquer instante de
tempo t dado, a quantidade de fótons saindo de um certo volume do meio é igual à
quantidade dos mesmos entrando nesse mesmo volume.
Existem muitas, boas e claras deduções desta equação que podem ser encontradas na
literatura [Arrigde, 2005]. Soluções analíticas da RTE só existem para geometrias simples.
Assim, se expandimos a radiância, a fonte e a função de fase em harmônicos esféricos,

52
obtemos uma série infinita de equações diferenciais de derivadas parciais, as quais
aproximam a RTE. Tomando os N primeiros harmônicos, se obtém (N + 1)2 equações
diferenciais parciais acopladas. Está é a chamada aproximação PN, sendo que um
incremento de N nesta aproximação modela a RTE com maior precisão, mas a expensas de
um maior custo computacional.
Para o caso de N = 1, ou primeira aproximação, obtemos quatro equações diferenciais
parciais elípticas acopladas. Considerando nesta aproximação que a função de fase
independe do ângulo, que o fluxo dos fótons é constante (o qual se justifica pela condição
« ), onde é o coeficiente de espalhamento reduzido, e a isotropia das fontes,
obtemos a equação de difusão de fótons:
(3.11)
Onde Φ é densidade de fótons:
(3.12)
e κ é o coeficiente de difusão:
(3.13)
onde e é o fator de anisotropia do meio.
3.5.1 Computando variações nas concentrações de cromóforos
Como vimos, a atividade cerebral pode ser monitorada usando técnicas ópticas e luz
NIR no comprimento de onda 700 a 900nm, especialmente através das alterações nas
concentrações de oxihemoglobina (HbO) e desoxihemoglobina (dHb), cromóforos
importantes do tecido cerebral, que possuem propriedades características nesta faixa de
freqüências. Outros cromóforos de significância nesses comprimentos de onda são a água,

53
lipídios e o citocromo oxidase aa3, os quais não são considerados por terem absorvância
insignificante comparada à da hemoglobina nesta faixa de freqüência, não sendo facilmente
mensuráveis sem o uso de, ao menos, seis comprimentos de onda. Para achar as mudanças
de concentração desses dois principais cromóforos, HbO e dHb, aplicamos a Teoria de
Beer-Lambert Modificada (MBLL: Modified Beer-Lambert Law).
A solução geral da equação de difusão de fótons pode ser usada para predizer a
fluência do fóton ou intensidade detectada para típicas medidas de difusão. Assumindo que
as mudanças de concentração são globais e pequenas, a solução para a equação de difusão
de fótons (3.11) para um meio semi-infinito pode ser expressa em termos de variações da
densidade óptica OD:
(3.14)
onde:
.
Portanto, para um comprimento de onda, λ, em particular:
(3.15)
Dados os coeficientes de extinção, é possível quantificar as mudanças na concentração dos
cromóforos, por:
(3.16a)
(3.16b)

54
Capítulo 4
Objetivos
O objetivo geral deste trabalho é realizar medidas simultâneas de Eletroencefalografia
(EEG) e de Espectroscopia no Infravermelho Próximo (NIRS, do inglês Near Infrared
Spectroscopy) visando a detecção e a caracterização de respostas neurais e hemodinâmicas
do córtex visual humano, tendo em vista obter informações complementares acerca do
acoplamento neurovascular para uma dada função sensorial, neste caso, a visão, em pessoas
sadias com visão normal.
Os objetivos específicos são:
1. Adequar tecnicamente os equipamentos de EEG e NIRS para o estudo dinâmico do
córtex visual.
2. Desenvolver um paradigma de estímulo visual que se adéqüe às aquisições de EEG e
NIRS em tempo real.
3. Otimizar parâmetros de análise espectral nas duas técnicas para este tipo de função
sensorial.
4. Verificar a distribuição regional nos hemisférios do córtex das ativações e
desativações cerebrais associadas à estimulação visual no grupo de indivíduos em estudo.
5. Estudar a dependência das respostas neuronal e hemodinâmica quanto ao estímulo
visual aplicado variando-se o contraste e a freqüência com que esse estímulo é apresentado.

55
Capítulo 5
Sujeitos e Métodos
5.1 Sujeitos
Fizeram parte deste estudo de EEG-NIRS um total de 13 voluntários, todos homens,
embora nem todos tenham participado de todos os experimentos.
Critérios de inclusão
a. Foram incluídos neste estudo 13 voluntários destros, saudáveis, com idade entre 20
e 42 anos (com idade média de 33 anos).
b. Eram voluntários sem qualquer histórico de distúrbios oftalmológicos e livres de
qualquer impedimento para a realização de exames em equipamentos de EEG e NIRS.
Critérios de exclusão
a. Indivíduos com história prévia de doenças oftalmológicas, neurológicas e/ou
psiquiátricas.
b. Foram excluídos do estudo quaisquer sujeitos com contra-indicação aos exames
mencionados.
Todos os indivíduos participantes concordaram em participar voluntariamente,
receberam um termo de consentimento livre e esclarecido de livre consentimento,
previamente aprovado pelo Comitê de Ética em Pesquisa da Unicamp, pelo qual tomaram
ciência de todos os aspectos que envolveriam procedimentos a que seriam submetidos e o
assinaram.

56
A segurança desses indivíduos foi assegurada através da utilização de materiais e
dispositivos apropriados para aquisição de imagens ópticas e de EEG.
O estudo visou à aplicação de duas técnicas que fazem uso dos princípios da Física
para caracterizar a atividade cerebral, e melhor compreender que estruturas funcionais se
encontram envolvidas nos diferentes processos da atividade de estímulo visual.
Local da pesquisa: Os dados foram coletados no Laboratório de Neurofísica do IFGW.
O processamento e análise dos mesmos foram realizados no Depto. de Raios Cósmicos e
Cronologia, IFGW, UNICAMP
5.2 Materiais e Métodos
5.2.1 Paradigma
De acordo com a literatura, os paradigmas em bloco são o principal método que se
utiliza tipicamente em experimentos onde a meta fundamental é a detecção da ativação
[Liu, 2004]. Os paradigmas em bloco são os que apresentam o maior poder de detecção da
atividade cerebral, pois cada bloco resulta na sobreposição das respostas hemodinâmicas
referentes aos vários eventos de um mesmo tipo, ou seja, relacionados a uma mesma
condição.
Os experimentos foram de dois tipos, binocular (dois olhos abertos) e monocular
(apenas um olho aberto, uma vez o olho direito aberto e na outra vez o olho esquerdo
aberto).
Nestes experimentos, o paradigma de blocos foi baseado na alternância de períodos de
ativação e desativação, sendo que os períodos de ativação consistiram na apresentação de
um estímulo visual do tipo xadrez circular (radial checkerboard). Trata-se do principal
método utilizado para obter ativações consistentes na região dos lobos occipitais [Rovati,
2007].
A apresentação dos estímulos foi gerada através de um software livre GLstim
(www.nmr.mgh.harvard.edu/~rhoge/GLstim/GLstim.html), ferramenta desenvolvida para
produzir estímulos visuais.

57
Cada estímulo consistiu na apresentação de um radial checkerboard em branco e preto e de
campo uniforme, numa tela de computador de 15 polegadas, tela quase cheia (Fig. 5.1). Os
experimentos foram realizados em uma sala silenciosa, com luminosidade ambiente muito
baixa (quase às escuras), e a uma distância tela-sujeito de 80 cm.
Fig. 5.1. Montagem experimental.
No caso do experimento binocular, o paradigma foi aplicado durante aquisições
simultâneas de EEG e NIRS por um tempo total de 7 minutos. Cada série, neste
experimento, foi dividida em 21 épocas de 20s cada, sendo 10 épocas de ativação e 11
épocas de repouso (Fig. 5.2).

58
Fig. 5.2. Estímulo visual: série de 21 épocas, alternando repouso e ativação.
Para cada indivíduo, foram feitas duas séries de tomadas de dados. Uma delas em que
variou-se a freqüência do radial checkerboard: 4 Hz, 8 Hz e 12 Hz (4 Hz quer dizer que o
estímulo visual apresentou-se 4 vezes, e assim sucessivamente) , mantendo-se o contraste
fixo em 100%. Na outra série, variou-se o contraste: 30, 60, 100%, mantendo-se a
freqüência fixa em 4Hz (Fig. 5.3). Foram feitos 13 registros.
Outro experimento utilizando estímulos de freqüências de 1, 2 e 3 Hz ( contraste fixo
100%), assim como também de contraste usando 3%, 10% e 20% (freqüência fixa de 4 Hz).
Fig. 5.3. Estímulos apresentados, variando-se a freqüência e o contraste.
No experimento monocular, o tempo total (de aquisição simultânea EEG/NIRS) de
cada série ou registro foi de 5 minutos, sendo que cada série foi dividida em 15 épocas de
20s cada: 7 épocas de ativação e 8 épocas de repouso (Fig.5.4). Nesse caso também variou-

59
se a freqüência: 4 , 8 , 12 Hz (contraste fixo: 100%) e o contraste: 30 , 60, 100%
(freqüência fixa 4Hz). Foram feitos 10 registros.
Fig. 5.4. Estímulo total: série de 15 épocas (8 épocas de repouso e 7 de ativação),
para o experimento monocular.
Em ambos os casos, medidas binoculares e monoculares, a definição do experimento
como um todo, e em especial aspectos como a duração total e a escolha do número de
estímulos de freqüências e contrastes aplicados, obedeceu a um fator limitante de caráter
geral que era o conforto dos indivíduos que estavam sendo monitorados. Ou seja, os
experimentos não se estenderam para além de um limite dentro do qual os voluntários se
sentiam confortáveis (ou minimamente incomodados) com o experimento.
5.3 Aquisição e análise dos registros de EEG
A preparação para o registro de EEG consistiu em posicionar os eletrodos O1 e O2 de
acordo com o Sistema Internacional 10-20 da Sociedade Internacional de Neurofisiologia
Clínica. Estes eletrodos foram colocados no couro cabeludo para topografia de córtex
visual primário, na região dos lobos occipitais, um eletrodo em cada lobo occipital de cada

60
voluntário, com a ajuda de uma pasta condutora, separados por uma distância de 6 cm. Veja
figura de posicionamento dos eletrodos ( Figura 5.5).
Os dados foram coletados com uma taxa de amostragem de 1000 Hz, e convertidos
para o formato EDF( do inglês, European Data Format), pelo software v-32EEG da
Nicolet (EUA), para o seu posterior processamento através do software EEGLAB.
Fig. 5.5. Posicionamento dos eletrodos O1 e O2 (em azul) sobre cada um dos lobos occipitais, na
região do córtex visual primário. Em amarelo, as duas formas semilunares, indicam os registros de NIRS.
Ao início de cada experimento, foi coletado um traçado EEG em base fundamental,
i.e., em ausência total de estímulo (apenas o individuo olhava a tela cinza) durante 7
minutos no experimento binocular e 5 minutos no experimento monocular. Nesta fase,
manteve-se a impedância dos dois canais em 5 kΩ. Estes registros não foram usados na
análise EEG, mas sim na analise NIRS (devemos lembrar que os registros são simultâneos
EEG/NIRS).
A análise de dados foi feita através do software EEGLAB
(http://sccn.ucsd.edu/eeglab), uma ferramenta interativa desenvolvida sobre a plataforma
MatLab (The Mathworks, Inc.), preparada para processar dados eletrofisiológicos
multicanais. Esse processamento usa análise de componentes independentes (Independent

61
Component Analysis, ICA), análise de tempo/frequência e métodos que incluem a rejeição
de artefatos.
Entre as funções disponíveis estão a importação de dados de EEG em diversos
formatos, a importação de arquivos com informações de eventos relacionados ao
experimento, a visualização dos dados (rolagem e mapas de escalpo com modelo de
dipolos), o pré-processamento (rejeição de artefatos, filtros, médias dos sinais e seleção de
intervalos específicos - épocas) e a decomposição por ICA e time/frequency analysis. (Fig.
5.6).
Fig.5.6. Layout do software EEGLAB. Acima: à esquerda barra de rolagem e painel de entrada dos
sinais EEG à direita. Abaixo: mapa de intensidade de uma componente independente (IC) à esquerda e
mapas de um canal específico à direita (Figura retirada de http://scnn.ucsd.edu/eeglab/).
Uma vez convertido o registro EEG para o formato EDF, foi processado no software
EEGLAB. Manteve-se a taxa de amostragem de 1000 Hz. Os dados contínuos de cada
canal foram processados separadamente para cada condição do estímulo aplicado. Aplicou-
se um filtro passa-alta de 1 Hz, para eliminar os ruídos da baixa freqüência e alta amplitude,

62
como as piscadas de olhos, e um filtro passa-baixa de 50 Hz, para eliminar artefatos de
músculos (>50Hz) e ruídos da rede elétrica (60Hz) (Fig.5.7).
Foram extraídos os intervalos temporais de cada arquivo, referentes aos períodos de
repouso e atividade. Aos dados de cada canal, aplicou-se um fitting interpolante para
suavizar estes espectros. Todos os espectros relativos a uma mesma condição de estímulo
foram promediados para se obter um sinal final como média relativa a cada condição de
estímulo. Isso foi feito mediante programas próprios, escritos em MatLab.
Fig. 5.7. Uma das etapas do processamento dos nossos dados EEG, por meio do software EEGLAB: à
esquerda a barra de rolagem com a entrada dos sinais EEG, e à direita os
sinais de EEG cru dos canais O1 e O1.
5.4 Aquisição e análise dos dados de NIRS
O equipamento de NIRS utilizado possui 20 detectores e 20 fontes transmissoras de
laser, permitindo o uso de até 400 pares fonte-detector independentes. Neste estudo, se

63
utilizaram quatro fontes e quatro detectores, de acordo com a geometria esquematizada nas
Figs. 5.8 e 5.9. Dois comprimentos de onda foram utilizados, λ1 = 690nm e λ2 = 830nm,
através das fontes, os quais são os comprimentos de onda que o CW6 somente fornece.
Fig. 5.8. Geometria dos optodos (optodo: par fonte-detector) utilizada para os registros de NIRS. As
letras (A, B, C, D) indicam as posições das fontes de luz NIR enquanto que os números (1, 2, 3, 4) indicam
as posições dos detectores. Os eletrodos O1 e O2 foram colocados ao centro de cada um desses arranjos.
Fig. 5.9. Geometria dos optodos e eletrodos.

64
Este equipamento, denominado CW6, construído pela empresa TechEn e acessórios
(Figura 5.10), sediada em Boston (EUA), foi desenvolvido por pesquisadores do Photon
Migration Lab, do MGH Martinos Center for Biomedical Imaging e da Harvard Medical
School (Boston, EUA), com os quais o Grupo de Neurofísica mantém colaboração
científica. Sobre este equipamento, cabe notar que se trata do primeiro protótipo de NIRS
em utilização no Brasil. Até o presente, não se sabe de nenhum projeto brasileiro que tenha
utilizado técnicas ópticas para monitorar diretamente a atividade cerebral.
Fig. 5.10.Equipamento NIRS , CW6, canto superior esquerdo e
Acessórios (canto superior direito e abaixo).
Os sinais ópticos foram analisados através de um programa desenvolvido pelo Photon
Migration Lab, de distribuição gratuita. Trata-se do programa HOMER (Hemodynamic
Evoked Response), uma interface gráfica, que roda na plataforma do MatLab e que permite

65
a visualização e análise dos dados de NIRS, incluindo diversas operações de processamento
de sinais e reconstrução de imagens.
Os optodos (sondas de NIRS) foram colocados ao redor dos eletrodos, de forma a
coletar sinais ópticos associados a cada canal de EEG em operação (Fig. 5.8). Para cada
experimento o ganho dos detectores foi calibrado. O CW6 tem uma faixa dinâmica de
operação que varia de 60 dB a 140 dB. Usamos um valor próximo de 100 dB por estar no
meio da faixa, o que evita pegar ruído (se estivermos muito próximo de 60 dB) e evitamos
saturar o detector (se estivermos muito próximos de 140 dB).
Montamos o estímulo no MatLab para uso posterior no HOMER. O processamento dos
sinais ópticos envolveu ajuste dos valores de DPF, Fator de caminho diferencial ou do
inglês Differential Pathlength Factor, (6.27 para 690nm e 5.64 para 830nm) no HOMer,
uso de um filtro passa banda (DAB), passa alta de 0.02Hz e passa baixa de 0.5Hz.
Os sinais assim processados foram utilizados para o cálculo das variações de
concentração de HbO (Oxihemoglobina) e de dHb (Desoxihemoglobina) via MBLL, de
acordo com o formalismo apresentado no Cap.3.

66
Capítulo 6 Resultados e discussão
No presente estudo, respostas hemodinâmicas e eletrofisiológicas nos córtices visuais
primários de voluntários, obtidas pela apresentação de um estímulo visual apropriado,
foram registradas simultaneamente através de NIRS e EEG.
Com respeito às respostas ópticas em particular, esses resultados correspondem aos
pares fonte-detector A-2, B-1, C-4 e D-3. Lembramos que os pares A-2 e B-1 estão
colocados em forma de cruz, tendo como centro o eletrodo O1, enquanto que os pares C-4 e
D-3 formam uma outra cruz, mas com o eletrodo O2 como centro. Lembramos também que
os pares A-2 e D-3 estão na posição vertical, enquanto que os pares B-1 e C-4 estão na
posição horizontal (cf. Fig.5.9). Daqui para frente, esses pares fonte-detector serão referidos
simplesmente como A2, B1, C4 e D3.
A Fig. 6.1 apresenta uma resposta hemodinâmica típica como resultado da aplicação de
um estímulo visual. Nesta resposta, se observa um aumento na concentração de HbO e uma
diminuição na concentração de dHb, o que implica em um aumento efetivo na concentração
da hemoglobina total (HbT), já que [HbT] = [HbO] + [dHb]. Nessa figura são mostrados
também erros experimentais típicos para este tipo de medida. Nas demais figuras, os erros
experimentais não serão mostrados para que os resultados possam ser visualizados com
maior clareza.
O aumento na concentração de HbO e a diminuição simultânea na concentração de
dHb durante a estimulação sugerem um aumento na circulação sanguínea cerebral (CBF do
inglês Cerebral Blood Flow) e correspondem a um excesso na oferta de O2 já que o
aumento do consumo de oxigênio (CMRO2, do inglês Cerebral Metabolic Rate of Oxygen)
na área cortical ativada não corresponde ao que é oferecido.

67
Fig. 6.1. Resposta hemodinâmica registrada através do NIRS para um sujeito. Em azul, dados experimentais de HbO; em vermelho, dados de dHb.
6.1 Resultados de um único indivíduo
Nas Figs. 6.2 e 6.3 são mostrados resultados de NIRS e EEG, respectivamente, para
um único indivíduo variando-se as propriedades do estímulo, neste caso variações na
freqüência e no contraste do estímulo visual aplicado.
A Figura 6.2 mostra os achados de NIRS para um indivíduo nos períodos de
apresentação do estímulo visual e repouso, num intervalo de 40s. A barra horizontal

68
amarela clara indica o período de 20s durante o qual o estímulo é apresentado. Neste caso,
variamos a freqüência do estímulo em 4 Hz, 8 Hz e 12 Hz.
Fig. 6.2. Resposta hemodinâmica registrada através do NIRS para um sujeito variando-se a freqüência do estímulo num experimento binocular. A barra horizontal indica o período em que o estímulo visual foi aplicado.
.
Ao início da aplicação do estímulo, pode-se observar claramente o surgimento da
resposta hemodinâmica, manifesta no incremento da concentração de HbO e na diminuição
de dHb, que vai desde t= 0 até aproximadamente t=20s, decaindo lentamente a partir deste
instante. Respostas desse tipo foram detectadas por sensores ópticos (detectores) colocados
na região do córtex visual primário nos lobos occipitais, esquerdo e direito. Em geral, elas
mostram esse mesmo aspecto: uma amplitude máxima de t = 20 s, seguida de uma
diminuição do sinal nos seguintes 20s, até o retorno ao estado basal.
No caso do EEG, o ritmo alfa é usado como um marcador do estado da excitabilidade
do córtex visual. A Figura 6.3 mostra os achados de EEG para o experimento binocular de
um indivíduo nos períodos de repouso e apresentação do estímulo visual, dos eletrodos O1

69
e O2 respectivamente. Quanto ao estímulo aplicado, variou-se a freqüência em 4 Hz, 8 Hz
e 12 Hz, correspondente aos períodos de estímulo (E).
De acordo com a análise espectral, pode-se observar o surgimento de oscilações alfa
em torno de 10 Hz na região dos lobos occipitais, com amplitude máxima no eletrodo O1
em 8 Hz durante a ativação, e com amplitude máxima no eletrodo O2 em torno de 10 Hz,
também durante o período de ativação. Além disso, observamos aqui a presença de uma
onda alfa bem definida estímulo da freqüência de 10 Hz, assim como também observamos
o surgimento do ritmo beta (13-30 Hz). De acordo com a literatura [Lopes da Silva, 1999],
o ritmo beta é freqüentemente encontrado nessa região e está relacionado com o ritmo alfa
achado quando aplicado um estímulo visual.
Fig. 6.3. Resultado da resposta EEG para um sujeito durante o estímulo visual
aplicado no caso binocular.

70
6.2 Análises de Grupo
6.2.1. Resultados de NIRS: experimento binocular
A Fig. 6.4 mostra os achados de NIRS para um grupo de 10 indivíduos nos períodos de
repouso e apresentação do estímulo visual, num intervalo temporal de 40s, para os
experimentos em que variou-se a freqüência do estímulo em 4 Hz, 8 Hz e 12 Hz, mantendo-
se o contraste em 100%. As respostas hemodinâmicas apresentadas correspondem ao
resultado médio obtido para este grupo de indivíduos. Na Fig. 6.5, são mostrados resultados
do mesmo tipo, mas obtidos da série suplementar de medidas, para um grupo menor de
indivíduos, 6 , para as freqüências de estímulos 1 Hz, 2 Hz e 3 Hz.
Fig. 6.4 Resultados de NIRS (experimento binocular) para a análise de grupo para cada um dos
canais, variando-se a freqüência de estímulo: 4, 8 e 12 Hz.

71
Fig. 6.5 Resultados de NIRS (experimento binocular) para a análise de grupo para cada um dos canais, variando-se a freqüência de estímulo: 1, 2 e 3 Hz.
Analisando-se os resultados das Figs. 6.4 e 6.5, nota-se que, para cada canal, as
respostas ópticas possuem um aspecto semelhante entre si, com alguma variabilidade com
respeito à freqüência do estímulo aplicado, porém não sendo possível encontrar um padrão
definido para esta variabilidade. No entanto, das figuras pode-se observar uma clara
resposta hemodinâmica como resposta ao estímulo aplicado. Pode-se notar também que os
canais verticais, A2 e D3, são menos ruidosos do que os canais horizontais, B1 e C4. Isso
muito provavelmente está ligado à qualidade dos dados coletados, pois, pela própria
montagem experimental, foi possível conseguir um melhor acoplamento ao escalpo dos
optodos colocados verticalmente em relação ao que ocorreu com os optodos colocados
horizontalmente.

72
As Figs. 6.6 e 6.7 mostram também resultados hemodinâmicos para o experimento
binocular obtidos com o NIRS, mas variando-se agora o nível do contraste do estímulo em
3%, 10%, 20%, 30%, 60% e 100%, e mantendo-se a freqüência fixa em 4 Hz.
Fig. 6.6 Resultados de NIRS (experimento binocular) para a análise de grupo para cada um dos canais, variando-se o contraste do estímulo: 30, 60 e 100 %.
Para os resultados da Fig. 6.6, em que se tomou dados para os contrastes de estímulo
30%, 60% e 100 %, pode-se fazer as mesmas observações já feitas anteriormente: 1) as
respostas hemodinâmicas tem um aspecto semelhante entre si, com alguma variabilidade
frente à variação de contraste, mas sem um padrão definido. Novamente observou-se que os
canais horizontais foram mais ruidosos. As mesmas observações valem para os resultados
da Fig.6.7 no que diz respeito à variação com o nível de contraste, a qual se variou em 3%,
10% e 20%, sendo que neste caso B1 é que foi o canal menos ruidoso.

73
Fig. 6.7 Resultados de NIRS (experimento binocular) para a análise de grupo para cada um dos canais, variando-se o contraste do estímulo: 3, 10 e 20 %.
Avaliações globais das respostas ópticas obtidas para todas as freqüências e para
todos os níveis de contraste utilizados no experimento binocular podem ser feitas a partir
das Figs. 6.8 e 6.9, em que esses resultados são respectivamente apresentados.

74
Fig. 6.8 Resultados de NIRS (experimento binocular) para a análise de grupo para cada um dos canais, e para todas as freqüências de estímulo:1, 2, 3, 4, 8 e 12 Hz (contraste 100%).
Pouco pode ser acrescentado ao que já foi dito. Por um lado, observa-se nessas figuras
padrões de respostas bem definidos tanto para HbO quanto para dHb. No entanto, não se
observa facilmente nenhum padrão de variabilidade associado à variação dos parâmetros de
freqüência e de contraste.
Nas Figs. 6.10 e 6.11 são mostradas curvas correspondentes a ajustes quadráticos, com
a intenção de mostrar a tendência média do conjunto dos pontos. O que se observa na Fig.
6.10, é que, tanto para HbO quanto para dHb, a tendência de quase todas as curvas é
praticamente uma constante, com variações muito suaves. Apenas a curva de C4 para HbO
é que apresenta uma tendência inicial de crescimento e depois de estabilização.

75
Fig. 6.9 Resultados de NIRS (experimento binocular) para a análise de grupo para cada um dos canais, e para todos os níveis de contraste: 3, 10, 20, 30, 60 e 100 % (frequência = 4 Hz).
As curvas referentes ao nível de contraste é que apresentam uma variação um pouco
mais bem definida, sobretudo para o caso da HbO (Fig. 6.11). Essas curvas indicam um
crescimento das áreas de HbO até cerca 50-60% de contraste, decrescendo em seguida.

76
Fig. 6.10 Áreas sob as curvas de HbO (em azul) e dHb (em vermelho) em função da freqüência do estímulo (experimento binocular).
Fig. 6.11 Áreas sob as curvas de HbO (em azul) e dHb (em vermelho) em função dos níveis de contraste (experimento binocular).

77
Por outro lado, nas Figs. 6.12 e 6.13 são mostradas as áreas sob as curvas de HbO (em
azul) e dHb (em vermelho) em função da freqüência do estímulo (experimento binocular) e
em função dos níveis de contraste (experimento binocular) com a finalidade de comparar
os nossos resultados com aqueles resultados de Ozus et al. (2001) , e com os resultados de
Emir et al. (2008).
Na Fig. 6.12 (canto superior esquerdo), podemos observar uma mudança máxima
percentagem média no sinal BOLD de um grupo de sujeitos submetidos a um estimulo de
freqüências. Podemos observar uma correlação linear entre as mudanças máximas do sinal
BOLD e os flashes de freqüências abaixo de 4 Hz. Logo esses resultados alcançam um
maximo em 6Hz para depois atingir um “dip” em 12 Hz. Este resultado pode ser
comparado com as áreas baixo as curvas dos nossos resultados (curva azul do HbO) dos
nossos estímulos de freqüências para o canal C4 do experimento Binocular, por um lado.
Por outro lado temos na Fig. 6.12 (canto inferior esquerdo) o trabalho de Emir et al que
nos apresenta os seus resultados do sinal BOLD positivo (PBOLD) os quais podem ser
comparáveis com os nossos resultados das áreas baixo as curvas dos nossos resultados
(curva azul do HbO) dos nossos estímulos de freqüências para o canal C4 do experimento
binocular.
Fig. 6.12. Áreas sob as curvas de HbO (em azul) e dHb (em vermelho) em função da freqüência do estímulo (experimento binocular).

78
Na Fig. 6.13, são apresentados os resultados de Rovati et al. (à direita) para um
experimento em que houve variação de contraste do estímulo visual utilizado. Observa-se
uma forte correlação da resposta hemodinâmica tanto para HbO como para dHb frente à
variação de contraste quando aos dados são aplicados um fitting logarítmico. No entanto, os
nossos resultados de áreas sob as curvas de HbO (em azul) e dHb (em vermelho) em
função dos níveis de contraste do estímulo (experimento binocular) refletem um
comportamento parabólico.
Fig. 6.13. Áreas sob as curvas de HbO (em azul) e dHb (em vermelho) em função dos níveis de contraste (experimento binocular).
6.2.2. Resultados de NIRS: experimento monocular
De uma maneira geral, os experimentos monoculares apresentaram resultados com
qualidade de sinal bem inferior à obtida para os experimentos binoculares. Apenas os
canais verticais A2 e D3 apresentaram alguns conjuntos de dados com um nível de ruído
aceitável. Os resultados obtidos para esses canais são apresentados nas Figs. 6.14 e 6.15,

79
que correspondem às variações de freqüência e de contraste, respectivamente. Infelizmente
esses dados foram insuficientes (4 indivíduos) para se extrair alguma informação fidedigna.
No entanto podemos observar no canal D3 (para ambos os olhos) uma resposta com maior
intensidade, que no canal A2 (para ambos os olhos).
Fig. 6.14 Experimento monocular: variação de freqüência

80
Fig. 6.15 Experimento monocular: variação de contraste
As respostas hemodinâmicas encontradas representam o efeito neto dos respectivos
eventos vasculares, e são concordantes com os resultados de potenciais evocados visuais e
respostas hemodinâmicas obtidos por Obrig e colaboradores [Obrig et al., 2001].
Outros resultados que estão de acordo com os nossos são os de Zhang et al. (2006),
apresentados na Fig. 6.16, que obtiveram respostas à estímulação visual aplicando
simultaneamente ressonância magnética funcional e espectroscopia no infravermelho
próximo, e o trabalho de Hoge et al. (2005), cujos resultados são mostrados na Fig. 6.17 e
correspondem a uma tarefa motora sendo avaliada pela técnica simultânea de
espectroscopia no infravermelho próximo e ASL-MRI (Arterial Spin Labeling MRI). Por
último, tanto para a variação da freqüência do estímulo como para a variação do contraste,
nossos resultados mostram um maior aumento na mudança da concentração da HbO que a
diminuição correspondente na concentração da dHb, o que dá como resultado um aumento
no sinal da HbT, coerente com a literatura.

81
Fig. 6.16 Figura retirada de Zhang et a.l ( 2006), Rev. Sci. Instrum, 77 (11) 114301.
Fig. 6.17. Figura retirada de Hoge et al (2005),
Neuroimage 25 , 701-707.

82
6.3 Grupos para o EEG, resultados e discussão
6.3.1 Resultados e discussão para o experimento binocular
Aplicou-se um estímulo visual nas medidas simultâneas de EEG e NIRS. O estímulo
consistiu na apresentação de um ‘radial checkerboard’ (em branco e preto) e de campo
uniforme, numa tela de computador de 15 polegadas, tela cheia, mudando as propriedades
de freqüência e contraste, neste caso variou-se a freqüência (4 Hz, 8 Hz e 12 Hz) e variou-
se o contraste (30, 60 e 100%), em uma sala escura.
A análise final representa a média das médias das medidas no grupo de 10 indivíduos
normais, análise feita através do software livre EEGLAB, e através de programas feitos por
mim. Optou-se pelo uso da média das médias na análise de grupo porque se acredita que a
média das médias pode melhor refletir a ativação de cada grupo, pois reduz o efeito de
diferenças individuais. O estímulo visual foi aplicado de forma a ativar a região dos lobos
occipitais no córtex visual primário.
A Figura 6.18 mostra os achados de EEG para o experimento binocular de 10
indivíduos, variando a freqüência do estímulo nos períodos de atividade (períodos de
repouso e apresentação do estímulo visual) dos eletrodos O1 e O2, respectivamente. Do
lado esquerdo da Fig. 6.18, temos as médias finais dos registros correspondentes à período
de repouso para os eletrodos O1 e O2, e no lado direito temos as medias finais dos registros
que correspondem ao período de estímulo nos eletrodos O1 e O2 respectivamente.
Nos períodos de repouso, de acordo com a análise espectral pode-se observar
claramente um padrão de igual intensidade e fase dos ritmos alfa devido à atividade
neuronal fundamental dos neurônios das camadas corticais e subcorticais nos lobos
occipitais do córtex visual primário, com amplitudes máximas (picos máximos)
semelhantes aproximadamente de 6µV para os eletrodos O1 e O2 de 9 Hz; e nos períodos
de estímulo pode-se observar uma clara mudança dando lugar ao aparecimento de ondas
alfa com freqüências maiores e menores que 10 Hz, devido à atividade neuronal, produto
do estímulo visual aplicado, dos neurônios das camadas corticais nos lobos occipitais do
córtex visual primário. Além disso, houve também a clara aparição de um pico em torno de

83
16 Hz e de outro de 24 Hz, isto é, o surgimento de um ritmo beta (13-30 Hz) na região
occipital no hemisfério esquerdo e direito, envolvendo os eletrodos occipitais O1 e O2,
ritmo este condizente com a tarefa de atenção visual, de acordo com a literatura [Foxe et
al., 1998].
Fig. 6.18. Resultados do EEG binocular, variação de freqüência do estímulo.
Esquerda: Período de repouso. Direita: Período de estímulo. Encima:
Hemisfério esquerdo. Em baixo: Hemisfério direito.
A Figura 6.19 mostra o sinal EEG para o experimento binocular de 10 indivíduos,
variando a propriedade de contraste nos períodos de repouso e nos períodos apresentação
do estímulo visual, dos eletrodos O1 e O2 respectivamente. Do lado esquerdo da Fig. 6.19,
temos as médias finais dos registros correspondentes ao período de repouso para os
eletrodos O1 e O2, e no lado direito temos as médias finais dos registros que correspondem
ao período de estímulo nos mesmos eletrodos.

84
Nos períodos de repouso, a análise espectral mostra claramente um padrão quase em
fase de ritmos alfa para os eletrodos O1 e O2 de 9 Hz; e nos períodos de estímulo pode-se
observar uma clara mudança dando lugar ao aparecimento de ondas alfa com freqüências
maiores e menores que 10 Hz, devido à atividade neuronal, produto do estímulo visual
aplicado, dos neurônios das camadas corticais dos lobos occipitais no córtex visual
primário com freqüências diferentes em O1 e O2 na freqüência de 10 Hz. Novamente,
achamos picos em torno de 16 e 24 Hz, indicando o surgimento de um ritmo beta (13-30
Hz), na região occipital nos dois hemisférios, envolvendo os eletrodos occipitais O1 e O2.
Fig. 6.19. Resultados do EEG binocular, variação de contraste do estímulo.
Esquerda: Período de repouso. Direita: Período de estímulo. Encima:
Hemisfério esquerdo. Em baixo: Hemisfério direito.

85
6.3.2. Resultados e discussão para o experimento monocular
Os resultados dos sinais EEG para o experimento monocular de 10 indivíduos nos
períodos de repouso e apresentação do estímulo visual (variando-se a freqüência do
estímulo) dos eletrodos O1 e O2, são mostrados na Fig. 6.20. Nesta figura são apresentados
os gráficos correspondentes às médias finais dos períodos de estímulo para os eletrodos O1
e O2, e para o olho esquerdo e direito correspondentemente.
Fig. 6.20. Resultados do EEG monocular, variação de freqüência do estímulo.
Os gráficos mostram os resultados para os períodos de estímulo. Esquerda:
Olho esquerdo. Direita: Olho direito. Encima: Hemisfério esquerdo.
Em baixo: Hemisfério direito.

86
Aplicado o estímulo, a análise espectral dos registros EEG do olho esquerdo, mostra
claramente um padrão de picos ao redor da freqüência de 8 Hz correspondentes a ritmos
alfa para o eletrodo O1 e O2; e do lado direito, isto é, no olho direito, os períodos de
estímulo mostram ondas alfa com freqüências maiores e menores que 10 Hz, assim como
observamos diferentes amplitudes desta, devido à atividade neuronal. Além disso, houve
também o aparecimento de picos em torno de 16 e 24 Hz, isto é, o surgimento de um ritmo
beta (13-30 Hz), na região occipital em ambos hemisférios, envolvendo os eletrodos
occipitais O1 e O2.
A Fig. 6.21 mostra os resultados dos sinais EEG para o experimento monocular de
10 indivíduos nos períodos de repouso e apresentação do estímulo visual (variando o
contraste do estímulo) dos eletrodos O1 e O2. Do lado esquerdo da figura temos as médias
finais dos registros do olho esquerdo correspondentes ao período de estímulo para os
eletrodos O1 e O2, e no lado direito temos as médias finais dos registros do olho direito que
correspondem ao período de estímulo nos eletrodos O1 e O2 respectivamente. A análise
espectral dos registros EEG do olho esquerdo, mostram claramente um padrão de picos por
volta da freqüência de 8 Hz correspondentes a ritmos alfa para os eletrodos O1 e O2 e no
olho direito, podem-se observar ondas alfa com freqüências também por volta de 8 Hz.
Além disso, houve também o aparecimento de um pico em torno de 16 Hz, isto é, o
surgimento de um ritmo beta (13-30 Hz), na região occipital em ambos hemisférios,
envolvendo os eletrodos occipitais O1 e O2.
A Figura 6.22 mostra os sinais EEG globais, registros: binocular e monocular, de 10
indivíduos nos períodos de repouso e apresentação do estímulo visual, dos eletrodos O1 e
O2 respectivamente, em cada uma das freqüências de estímulo aplicadas (4 Hz, 8 Hz, e 12
Hz). Das médias finais resultantes podemos observar uma clara diferença dos sinais entre
os registros monoculares vs binoculares, observamos também ondas alfa (em círculo) e
ondas beta (em elipses).

87
A Fig. 6.23 mostra os sinais EEG globais, registros: binocular e monocular, de 10
indivíduos nos períodos de repouso e apresentação do estímulo visual, dos eletrodos O1 e
O2 respectivamente, em cada um dos contrastes do estímulo aplicado (30%, 60% e 100 %).
Das médias finais resultantes podemos observar uma clara diferença dos sinais entre os
registros monoculares vs binoculares, observamos também ondas alfa (em circulo) e ondas
beta (em elipses).
Fig. 6.21. Resultados do EEG monocular, variação de contraste do estímulo, para os
períodos de estímulo. Esquerda: Olho esquerdo. Direita: Olho direito. Encima:
Hemisfério esquerdo. Em baixo: Hemisfério direito.

88
Fig. 6.22. Resultados globais do EEG, variação de freqüência do estímulo
Fig. 6.23. Resultados globais do EEG, variação de contraste do estímulo

89
6.4 Discussão sobre os ritmos resultantes alfa e beta
Em todos os nossos resultados do EEG obtivemos ritmos alfa e beta, produto do
estímulo visual aplicado na região do córtex visual primário. Segundo Nunez et al. (2001),
acredita-se que os ritmos alfa representem oscilações de potenciais pós-sinápticos no
neocórtex, produzidos pelo estado de excitabilidade do córtex. Portanto ao excitar o córtex
visual primário com estímulos visuais é possível obter ondas alfa. Também obtivemos
ondas alfa baixas (com freqüências menores que 10 Hz) e ondas alfa altas (com freqüências
maiores que 10 Hz). Além disso, os nossos resultados mostraram ondas alfa com diferentes
amplitudes, refletindo um estado de dessincronização freqüentemente associado com a
diminuição de amplitude no ritmo alfa (Lopes da Silva et al, 1999), o que geralmente
acontece quando registrado com os olhos abertos (Kirschfeld, 2005).
Uma teoria completamente satisfatória do ritmo alfa deveria levar em conta o
problema de como as propriedades espaço-temporais estão mudando com o input do
estímulo ou com a mudança do estado cerebral, ou seja; como resultado da influência
neuromoduladora.
Diversos fatores determinam as propriedades das oscilações do EEG:
1. As propriedades intrínsecas da membrana neuronal e a dinâmica dos processos
sinápticos.
2. A intensidade e a extensão das interconexões entre elementos das redes neuronais, a
maioria formadas por feedback loops. Diferentes tipos de feedback podem ser
distinguidos, envolvendo circuitos tálamo-corticais e/ ou córtico-corticais a curtas
ou longas distâncias.
3. A influência neuromoduladora de sistemas neuronais locais ou globais.
Os ritmos alfa são uma correlação neural importante dos processos de integração
córtico-cortical e tálamo-cortical (Nunez, 1995). Os neurônios corticais piramidais geram
um padrão de ritmos alfa que mudam de amplitude (alfa ERD/ ERS na faixa de 8-12 Hz)

90
em associação com a abertura ou fechamento das conexões bidirecionais entre as regiões
corticais e os núcleos talâmicos (Pfurtscheller & Lopes da Silva, 1999).
Por outro lado, qualquer ritmicidade sobre a atividade do EEG variando acima de 13
Hz e inferior de 30 Hz pode ser considerada um ritmo beta. Historicamente, a descoberta do
ritmo beta está intimamente ligada à descoberta do ritmo alfa, já descrito pelo pesquisador
Hans Berger no ano de 1930 [Niedermeyer &Lopes da Silva, 2004]. Em nosso estudo,
todos os resultados do EEG mostram a aparição de ondas beta, indicando que todos os
sujeitos participantes nesta pesquisa foram pessoas saudáveis [Niedermeyer & Lopes da
Silva, 2004].
A atividade beta é altamente correlacionada com o surgimento de ondas alfa, e
deve-se ao processo de atenção inerente ao experimento do estímulo visual aplicado
[Wrobel, 2000]. No entanto, o papel funcional da banda beta ainda não está claro [Steriade,
1993].
Apesar do amplo uso do EEG na clínica diária e na pesquisa, os geradores
individuais do EEG de escalpo não têm sido claramente identificados. A regra de ouro para
localizar geradores neuronais de potenciais de escalpo, registros numa única célula, não é
praticável no ser humano (Spileers e et al, 1994). No ser humano, muitos estudos foram
dedicados à investigação dos geradores do EEG por meio dos registros de potenciais de
escalpo.

91
Capítulo 7
Conclusões e Perspectivas
Este trabalho teve como objetivo principal estudar a dinâmica neurovascular do
córtex visual humano integrando duas técnicas simultaneamente, EEG e NIRS, e propor
métodos (metodologias, processos, técnicas) para o seu melhor entendimento. Utilizamos
métodos ópticos atualmente usados em neurociências como ferramenta básica para o
desenvolvimento deste estudo. Algumas outras abordagens, tais como fisiológicas,
eletrofisiológicas e de ressonância magnética funcional se fizeram necessárias para o
tratamento das medidas deste trabalho.
O equipamento NIRS (CW6) permite estudar o acoplamento neurovascular através
de experimentos em humanos. Com ele podemos testar a relação entre a resposta
hemodinâmica (HRF do inglês Hemodynamic Response Function) e os diversos graus de
contraste do estímulo aplicado, assim como a resposta hemodinâmica com a variação da
freqüência do estímulo. Aplicamos um estímulo visual com dois paradigmas em bloco, com
20 s de duração, durante um tempo de 7 minutos. Este experimento nos permitiu perceber
as diferenças que o estímulo produz na HRF e, além disso, reforça o uso do equipamento
em humanos. Nossos resultados mostram uma clara resposta hemodinâmica quando
aplicado o estímulo. Esses resultados estão de acordo com as medidas já publicadas (Hoge
et al. 2005) para as duas situações analisadas com NIRS.

92
Por fim, desenvolvemos ao longo deste trabalho ferramentas computacionais para
modelar a resposta hemodinâmica a partir de dados eletro-óptico-fisiológicos medidas
diretamente em humanos sadios. Desenvolvemos um método para utilização em estudos de
acoplamento neuro-vascular e neuro-metabólico durante as épocas de estímulo, quando
aplicado um estímulo visual, variando-se os parâmetros de freqüência e de contraste.
Usamos dados obtidos em nosso laboratório NIRS do IFGW-UNICAMP.
Portanto, através dos experimentos apresentados e analisados acima, acredito que
tenha sido possível contribuir com o desenvolvimento de uma metodologia para o estudo da
dinâmica cerebral, sobretudo do acoplamento neuro-vascular-metabólico, além de
disponibilizar ferramentas úteis para as aplicações clinicas, como o uso dos mapas de
correlação para o estudo das relações vasculares e a obtenção de mapas metabólicos de
consumo de oxigênio.
Dada a complexidade do sistema visual humano, fica muito que pesquisar com o
CW6. No presente trabalho, uma origem cortical principalmente é assumida, desde que O1
e O2 foram analisados considerando pequenas contribuições das camadas cortico-
subcorticais e da área estriada. Para nosso conhecimento seria importante estudar o input
relativo a um estímulo visual de radial chekerboard variando a sua cor, tamanho e forma
aplicando-se esta mesma metodologia.

93
BIBLIOGRAFIA
Aidley D. J., The Physiology of Excitable Cells, 2002, Cambridge University Press,
New York, 2001.
Boas, D. et al., (2001), Imaging the body of Diffuse Optical Tomography, IEEE signal
Processing Magazine, pp. 57- 75.
Boas D.A.; Gaudette T.; Strangman G.; Cheng X.; Marota J.J.A.; Mandeville J.B.,
(2001), The Accuracy of Near Infrared Spectroscopy and Imaging during Focal Changes in
Cerebral Hemodynamics, Neuroimage, 13 (1), 76-90.
Boas, D.A. et al., (2001), Imaging the body with diffuse optical tomography, Signal
Processing Magazine, IEEE, 18 (6), 57-75.
Boas, G., (2004), Noninvasive Imaging of the Brain, Optics & Photonics News,
January, 52-55.
Boas, D. A., Dale A. M., and Franceschini M. A., (2004), Diffuse optical imaging of
brain activation: approaches to optimizing image sensitivity, resolution, and accuracy,
NeuroImage 23, 275-288.
Bohren C., Hu_man D., Absorption and Scattering of light by Small Particles, John
Wiley and Sons, New York, 1983
Bronzino, J. D., The Biomedical Engineering Handbook, Second Edition ed. Boca
Raton, FL: CRC Press LLC, 2000.

94
Burns, D. A., Ciurczak, E. W. Handbook of Near-Infrared Analysis. New York: Marcel
Dekker, 2001.
Buchheim K, Obrig H, Pannwitz W v, et al. (2004), Decrease in haemoglobin
oxygenation during absence seizures in adult humans, Neurosci Lett 354:119–122.
Bunce SC, Izzetoglu M, Izzetoglu K, Onaral B, Pourrezaei K, (2006), Functional
near-infrared spectroscopy, IEEE Eng Medicine and Biology Magazine, 25:54-62.
Cechetto D. F., & Topolovec J. C., Cerebral Cortex in Encyclopedia of the
Human Brain, vol.1, p.663, editor V. S. Ramachandran, 2002.
Cooper, R.; Shaw, JC; Osselton, JW, EEG Technology, Butterworths & Co. Ltd, Third
Edition, 1980.
Cope M. and Delpy D. T., (1988), System for long-term measurement of cerebral blood
and tissue oxygenation on newborn infants by near infrared transillumination, Medical and
Biological Engineering and computing, 26 (3), 289-294.
Cope, Mark, The application of Near Infrared Spectroscopy to Noninvasive Monitoring
of cerebral oxygenation in the Newborn Infant. Thesis. University College London,
London, 1991.
DeYoe, E. A., The Occipital Lobe, in The Encyclopedia of the Human Brain. V. S.
Ramachandran. New York, Academic Press. 2002.
Durand-Rivera, A. et al, Potenciales relacionados con eventos y la lectura, Revista de
Ciencias Clínicas, 5 (1) 31, 2004.

95
Ebersole J & Milton J., The electroencephalogragram (EEG): A measure of Neural
synchrony, Chap. 5, p. 55-56, in Epilepsy as a Dynamic Disease. Editors: Milton, J. & Jung,
P., Springer, Germany, 2003.
Emir U E, Bayraktaroglub Z, Ozturka C, Ademoglua A, Demiralpb T, (2008), Changes
in BOLD transients with visual stimuli across 1-44 Hz, Neuroscience Letters 436(2):185-8
Evans J., Introduction to Quantitative EEG and Neurofeedback, Second edition,
editorial Elsevier, USA, 2009.
Foxe, J.J., Simpson, G.V., Ahlfors, S.P., (1998), Parieto-occipital 10 Hz activity
reflects anticipatory state of visual attention mechanisms, Neuroreport, 9(17):3929-3933.
Fillerup, D. L. and Mead, J. F., (1967), The Lipids of the Aging Human Brain, Lipids,
2(4), 295-298.
Florian E. W. Schmidt, Ph.D. Thesis, (1999), Development of a Time-Resolved Optical
Tomography System for Neonatal Brain Imaging, Department of Medical Physics and
Bioengineering University College London.
Gibson, AP, Hebden, JC, and Arridge, SR, (2005), Recent advances in diffuse optical
imaging, Physics in Medicine and Biology, 50, R1-R43.
Greenstein, Color Atlas of Neuroscience, editorial Thieme,USA, 2000.
Gordon E., (1999), Brain imaging technologies: how, what, when and why?, Australian
& New Zealand Journal of Psychiatry, 33, 187.
Guyton, A. C. & Hall J. E., Textbook of Medical Physiology, editorial Harcover,
USA, 2000.

96
Hale ,G. M. and M. R. Querry, (1973), Optical constants of water in the 200-nm to
200-micrometer wavelength region, Appl. Opt. 12, 555-565.
Handy, T. C., Event-Related Potentials a Methods Handbook, editorial MIT Press,
2004.
Hecht, E., Optics, 3rd. ed. USA: Adisson Wesley Longman, Inc., 1998.
Hebden, J. C., (2003), Advanced in optical imaging of the newborn infant brain,
Psychophysiology 40 (4), pp. 501-510.
Henyey, L. G. & Greenstein, J. L., (1941), Diffuse radiation in the galaxy, Astrophys.
J. 93, 70.
Heino, Jenni et al, (2003), Anisotropic effects in highly scattering media, Physical
Review E 68, 1-8.
Hoge R.D., Franceschini M.A.,. Covolan R.J.M, Huppert T., Mandeville J.B. and Boas
D.A., (2005), Simultaneous recording of task-induced changes in blood oxygenation,
volume, and flow using diffuse optical imaging and arterial spin-labeling MRI,
Neuroimage, 25,701-707.
Hollis, Veronica S et al., (2001), Noninvasive monitoring of brain tissue temperature
by near-infrared spectroscopy, Proc SPIE 4250, pp. 470-481.
Hughes, J. & John, E., (1999), Conventional and Quantitative electroencephalography
in Psychiatry, Journal of Neuropsychiatry and Clinical Neuroscience. 11(2), 190-208.
Jirsa, Viktor K.; McIntosh, A.R., Handbook of Brain Connectivity, Eds Springer, 2007.

97
Junqueira L. C. & Carneiro, J. , Histologia Básica, Décima edição, editorial Guanabara
Koogam, Rio de Janeiro, 2004.
Ishimaru A., Electromagnetic Wave Propagation, Radiation and Scattering, Prentice
Hall, New Jersey, 1991.
Ishimaru A., Diffusion of light in turbid media, Appl. Opt. 28, 2210– 2215, (1989).
Ishimaru A., Wave propagation and scattering in random media, New York: Academic
Press, 1978.
Izzetoglu, M., Bunce S. C., Izzetoglu K, Onaral B, e Pourrezaei K, (2007), Functional
Brain Imaging Using Near-Infrared Technology, IEEE Eng Medicine and Biology
Magazine, 26(4), 38-47.
Joseph D. K., Huppert T. J., Franceschini M. A., and Boas D. A., (2006), Diffuse
optical tomography system to image brain activation with improved spatial resolution and
validation with functional magnetic resonance imagin, Applied Optics, 45(31), 8142-8151.
Kandel, Eric R., et al., Principles of Neural Science, Editorail Harcover, Fourth edition,
U.S.A., 2000.
Kim, J.G., Liu, H., (2007), Variation of haemoglobin extinction coefficients can cause
errors in the determination of haemoglobin concentration measured by near-infrared
spectroscopy, Phys. Med. Biol., 52 (20), 6295-6322.
Kirschfeld K, (2005), The physical basis of alpha waves in the electroencephalogram
and the origin of the "Berger effect", Biol. Cybern., 92(3):177-85.

98
Kocsis L, Herman P, Eke A, (2006), The modified Beer-Lambert law revisited, Phys.
Med. Biol. 51 N91-N98.
Kropotov J. D., Quantitative EEG, Event-Related Potentials and Neurotherapy, First
edition, editorial Elsevier, USA, 2009.
Kuffler S.W., (1953), Discharge Patterns and Functional Organization of the Mammalian
Retina, J. Neurophysiology, 16, 37-68.
Liu, T., (2004), Efficiency, power, and entropy in event-related fMRI with multiple trial
types: Part II: Design of experiments, NeuroImage, 21:401-413.
Lent, R., Cem Bilhões de Neurônios: conceitos fundamentais em neurociências,
Editora Atheneu, 2005.
Lopes da Silva, F., and Van Rotterdam, A., Byophysical aspects of EEG and
magnetoencephalogram generation, in Niedermeyer, E. and Da Silva, F. L., editors,
Electroencephalography: Basic Principles, Clinical Applications, and Related Fields, 1-
14, Lippincontt Williams & Wilkins, fourth edition, 1999.
Machado, A., Neuroanatomia Funcional, editorial Atheneu, 2da edição, Rio Janeiro,
2000.
Mattews, G. G., Cellular physiology of Nerve and Muscle, Fourth edition, 2003.
Medvedev, Andrei V., (2002), Epileptiform spikes desynchronize and diminish fast
(gamma) activity of the brain - An ''anti-binding'' mechanism?, Brain.Research Bulletin, 58
(1), 115.

99
Nunez, Paul L., (2000), Toward a Quantitative Description of Large Scale Neocortical
Dynamic Function and EEG, Behav Brain Sci. 23(3):371– 437.
Malmivuo, J. & Plonsey, R., Bioelectromagnetism, Principles and Applications of
Bioelectric and Biomagnetic Fields, Oxford University Press, arquivo capturado em 23 de
fevereiro de 2008, de http://butler.cc.tut.fi/~malmivuo/bem/bembook/.
Miller B L., Cummings J L., The Human Frontal Lobes, Second Edition: Functions
and Disorders The Guilford Press 2006.
Misulis, K. E., Essentials of Clinical Neurophysiology, 3rd Edition Boston:
Butterworth-Heinemann, 1997.
Niedermeyer, E; Historical aspects, em Niedermeyer, E. and da Silva, F. L., editors,
Electroencephalography: Basic Principles, Clinical Applications, and Related Fields, pags.
1-14, Lippincontt Williams & Wilkins, fourth edition, 1999.
Niedermeyer, E. and Lopes da Silva, F. L., editors, Electroencephalography: Basic
Principles, Clinical Applications, and Related Fields, pags. 1-14, Lippincontt Williams &
Wilkins, fifth edition, 2004.
Nunez, P. L. et al., (2001), Spatial-temporal structures of human alpha rhythms:
Theory, microcurrent sources, multiscale measurements, and global binding of local
networks Human Brain Mapping, 13 (3), pags. 125 – 164,.
Nunez PL, Srinivasan R., Electric Fields of the Brain: The Neurophysics of EEG, 2nd
ed. USA: Oxford University Press, 2005.
Nunez P.L., Neocortical Dynamics and Human EEG Rhythms, Oxford University
Press, USA; 1st edition, 1995.

100
Nuwer M., (1997), Assessment of digital EEG, quantitative EEG, and EEG brain
mapping: report of the American Academy of Neurology and the American Clinical
Neurophysiology Society, Neurology, 49(1), 277- 292.
Obrig H.; Israel H.; Kohl-Bareis M.; Uludag K.; Wenzel R.; Müller B.; Arnold
G.; Villringer A., (2002), Habituation of the Visually Evoked Potential and Its Vascular
Response: Implications for Neurovascular Coupling in the Healthy Adult,
Neuroimage,17,1-18.
Ozus B, Liu H L, Chen L, Iyer M B, Fox P T, Gao J, (2001), Rate dependence of
human visual cortical response due to brief stimulation: an event-related fMRI study,
Magnetic Resonance Imaging, 19: 21–25.
Pfurtscheller G., Lopes da Silva F. H., (1999), Event-related EEG/MEG
synchronization and desynchronization: basic principles. Clinical Neurophysiology, 110
(11), 1842-1857.
Rovati, L et al., (2007), Optical and electrical recording of neural activity evoked by
graded contrast visual stimulus, 2007 biomedical-engineering-online, 6, (28).
Ryynänen O, Hyttinen J, Malmivuo J., (2004), Study on the spatial resolution of EEG--
effect of electrode density and measurement noise, Conf. Proc. IEEE Eng Med Biol Soc.
6:4409-12
Strangman, G et al, (2002), Non-invasive neuroimaging using near-infrared light,
Biological Psychiatry, 52 (7), 679-693.
Speckmann, E. J. and Elger, C., Introduction to the neurophysiological basis of the
EEG and DC potentials, em Niedermeyer, E. and da Silva, F. L., editors,

101
Electroencephalography: Basic Principles, Clinical Applications, and Related Fields, pags.
1-14, Lippincontt Williams & Wilkins, fourth edition, 1999.
Spileers, W., Maes, H., Lagae, L., and Orban, G. A.,( 1994), Contrast modulated
steady-state visual evoked potentials (CMSS VEPs): Recording evoked potentials and
related single cell responses in area 17 of the cat. Electroencephalogr. Clin. Neurophysiol.
92: 64–77.
Strangman G, Boas DA, Sutton JP, (2002), Non-invasive neuroimaging using near-
infrared light, Biol Psychiatry. Oct 1; 52(7):679-93.
Strangman G, Franceschini MA, Boas DA, (2003), Factors affecting the accuracy of
near-infrared spectroscopy concentration calculations for focal changes in oxygenation
parameters, Neuroimage , 18 (4), 865-879.
Srinivasan, R., (1999), Methods to improve the spatial resolution of EEG, International
Journal of Bioelectromagnetism, 1(1):102–111.
Svaasand L. O. & Ellingsen R, (1983), Optical Properties of Human Brain,
Photochemistry and Photobiology, 38 (3), 293-299.
Skoog, D. A.; Holler, F. J.; Nieman, T. A., Princípios de análise instrumental, 5ed. São
Paulo: Bookman, 2002.
Squire L. R., Zigmond M. J.; Bloom F. E.; Landi S. C., and Roberts J. L., Fundamental
Neuroscience, Academic Press, Second edition, 2003.
Steriade M., (1993), Cellularsubstrates of brain rhythms. In: Electroencephalography:
basic principles. clinical application and related fields (Eds. E. Niedermeyer and F. Lopes
da Silva), fourth edition, Williams and Wilkins. Baltimore. p. 27-62.

102
Thakor, N. & Tong, S., (2004), Advances in quantitative electroencephalogram
analysis methods. Annual Review of Biomedical Engineering 6, 453- 495.
Toga, A. W. & Mazziotta J. C., Brain Mapping: The Methods, Second Edition, USA.
2002.
Tsuchiya, Yutaka, (2001), Photon path distribution and optical responses of turbid
media: theoretical analysis based on the microscopic Beer-Lambert law, Phys. Med. Biol.
46 (8), 2067-2084.
Tuchin Valery V., Tissue Optics: Light Scattering Methods and Instruments for
Medical Diagnosis, 2th edition, 2007.
Vaupel P., Schlenger K., Knoop C. and Höckel M., (1991), Oxygenation of Human
Tumors: Evaluation of Tissue Oxygen Distribution in Breast Cancers by Computerized O2
Tension Measurement, Cancer Research, 51, 3316-3322.
Vaupel P, Thews O, Kelleher DK, Hoeckel M, (1998), Current status of knowledge
and critical issues in tumor oxygenation, results from 25 years research in tumor
pathophysiology, Adv Exp Med Biol., 454, 591-602.
Van Boxtel G. J. M., (1998), Computational and statistical methods for analyzing
event-related potential data, Behavior research methods, instruments & computers, 30
(1), 87.
Vander, A. J., Sherman, J. H., and Luciano, D. S., Human Physiology: The
Mechanisms of Body Function, Fifth Edition, editorial New York: Mc Graw-Hill, 1990.

103
Villringer, Arno and Obrig, Hellmuth, Near Infrared Spectroscopy and Imaging, in:
Brain Mapping, The Methods, 2nd edition, Elsevier Science, USA, 2002
Waxman, D. G., Neuroanatomia Correlativa, editorial Manual Moderno, México,
2000.
Wang L. V. and Wu H., Biomedical optics: principles and imaging, First edition, John
Wiley & Sons, Inc, New Jersey, 2007.
Watanabe E, Nagahori Y, and Myanagi Y , (2004), Focus Diagnosis of Epilepsy using
Near Infrared Spectroscopy, Epilepsia 43(Suppl. 9): 50-55,
Weidner N, Folkman J, Pozza F, Bevilacqua P, Allred E. N., Moore D. H., Meli S,
Gasparini G, (1992), Tumor Angiogenesis: A New Significant and Independent Prognostic
Indicator in Early-Stage Breast Carcinoma, Journal of the National Cancer Institute, Vol.
84, No. 24, 1875-1887, December 16, 1992, Oxford University Press.
Wrobel, (2000), Beta activity: a carrier for visual attention, Acta Neurobiol. Exp.60,
247-260.
Woodard, HQ and White, DR., (1986), The composition of body tissues, Br. J. Radiol.,
59 (708), 1209-1218.
Wokrman J Jr. and Weyer L., Practical Guide to Interpretive Near-
InfraredSpectroscopy, CRC Press, Taylor & Francis Group, USA, 2008.
Zhang, Xiaofeng; Toronov, Vladislav Y.; Webb, Andrew G., (2006), Integrated
measurement system for simultaneous functional magnetic resonance imaging and diffuse
optical tomography in human brain mapping, Rev. Sci. Instrum., 77 (11), 114301.

104
ANEXO
Protocolo de aprovação do projeto pesquisa pelo Comitê de Ética da Unicamp

105
.
Recommended


![Hybrido Unreim Brodmann-Areale [39;40] Texte. Löcher ... · 3 Einführung Konzept, Methode, Installation Die Fiktion Brodmann-Areale [39;40]. Texte. Löcher. Projektionen ist eine](https://img.pdfslide.tips/doc/110x75/5d66233e88c993280c8b4f17/hybrido-unreim-brodmann-areale-3940-texte-loecher-3-einfuehrung-konzept.jpg)