
1
UNIVERSIDADE FEDERAL DO RIO GRANDE DO NORTE
DEPARTAMENTO DE MORFOLOGIA
CURSO DE CIÊNCIAS BIOLÓGICAS
MILENA DE OLIVEIRA SANTOS
BULBO OLFATÓRIO DO CÃO E DO HOMEM E SEUS ASPECTOS
MORFOFUNCIONAIS: UM ESTUDO DE REVISÃO
NATAL/RN
2017

2
MILENA DE OLIVEIRA SANTOS
BULBO LFATÓRIO DO CÃO E DO HOMEM E SEUS ASPECTOS
MORFOFUNCIONAIS: UM ESTUDO DE REVISÃO
Trabalho de Conclusão de Curso apresentado à
Universidade Federal do Rio Grande do Norte,
para obtenção do título de Bacharel em
Ciências Biológicas.
Orientador: Juarez e Silva Chagas
NATAL/RN
2017

3
MILENA DE OLIVEIRA SANTOS
BULBO OLFATÓRIO DO CÃO E DO HOMEM E SEUS ASPECTOS
MORFOFUNCIONAIS: UM ESTUDO DE REVISÃO
Trabalho de conclusão do curso de graduação em Ciências
Biológicas, Centro de Biociências da Universidade
Federal do Rio Grande do Norte, como requisito parcial
para a obtenção do título de Bacharel em Ciências
Biológicas, sob orientação do Professor Dr. Juarez e Silva
Chagas.
APROVADA EM: 01/12/2017
BANCA EXAMINADORA
_______________________________________________
Prof. Dr. Carlos Eduardo Bezerra de Moura
Universidade Federal Rural do Semi-árido
_______________________________________________
Prof. Msc. Aldo Fonseca de Souza
Universidade Federal do Rio Grande do Norte
_______________________________________________
Prof. Dr. Juarez e Silva Chagas
Universidade Federal do rio Grande do Norte
(Orientador)

4
Dedico esse trabalho aos meus pais Reilta e Sebastião, que, com muito carinho е apoio, não
mediram esforços para que eu chegasse até esta etapa da minha vida.

5
Agradecimentos
Agradeço ao meu orientador, Juarez Chagas, por suas críticas, incentivo, dedicação e
paciência comigo até mesmo nas horas mais difíceis da minha vida acadêmica.
Aos meus colegas de curso Glauce Campelo e Arrilton Araújo por se tornarem verdadeiros
amigos. Agradeço pela amizade, pelo incentivo, pelo companheirismo e pelo nervosismo, na
hora de provas e trabalhos.
Ao meu irmão Rayre e à minha cunhada Jailma, pelo apoio e por me darem os presentes mais
lindos da minha vida, meus sobrinhos Marcos e Miguel.
À Janine Karla, pelo incentivo e apoio para a realização desse trabalho.
Aos professores que me aceitaram para estágio em seus laboratórios Veríssimo, Kátia e
Elizeu.
A todos os professores do Centro de Biociências que contribuíram para minha formação
acadêmica.
Enfim, agradeço a todos que participaram direta ou indiretamente da minha vida acadêmica e
acima de tudo a Deus por permitir essas pessoas em minha vida.

6
“Não são as espécies mais fortes que
sobrevivem, nem as mais inteligentes, e sim as
mais suscetíveis a mudanças”.
Charles Darwin

7
RESUMO
O sentido do olfato tem sido ao longo do tempo, um dos principais sentidos dos Vertebrados
e, especialmente dos mamíferos, os quais dele dependem para sobrevivência, reprodução e
outros comportamentos inerentes à sua evolução no âmbito da filogenia. O Nervo Olfatório,
também anatomicamente conhecido como I par de nervo(s) craniano(s), tem sido motivo de
incontáveis trabalhos científicos e pesquisas, dado sua importância e relação com um dos
mais importantes “sentidos”, que é o olfato na identificação dos odores. Não obstante e,
embora pareça contraditório, o mecanismo do sentido do olfato, do ponto de vista funcional,
ainda carece de explicações no âmbito de sua neurofisiologia, apesar dos estudos realizados
nesta área. Portanto, ainda hoje merece releituras científicas, comparações morfofuncionais e,
sobretudo, estudos comparativos, no sentido de estabelecer ou esclarecer, cada vez mais, suas
importantes funções no que diz respeito ao comportamento animal, sua sobrevivência e,
sobretudo, através do olfato, muitas vezes associado ao paladar e também a importantes
centros neurais, estabelecendo ao indivíduo, seja animal ou humano, em sua devida posição
evolutiva, dentro da escala zoológica ou filogenética. Contudo, o presente trabalho consiste
em apresentar uma revisão bibliográfica, onde alguns aspectos morfofuncionais possam ser
apresentados evolutiva e cronologicamente por alguns importantes pesquisadores desta área
de estudo, com o intuito comparativo mais abrangente, além de comparar aspectos funcionais
evolutivos entre as duas espécies de Mamíferos, em questão, o Cão (Canis familiaris) e o
Homem (Homo sapiens).
Palavras-chaves: Nervo olfatório, bulbo olfatório, lâmina crivosa, olfato.

8
ABSTRACT
The sense of smell has been, over time, one of the main senses of vertebrates, and especially
of mammals, which depend on them for survival, reproduction and other behavior inherent to
its evolution in the context of phylogeny. The Olfactory Nerve, also anatomically known as I
pair of cranial nerves, has been the subject of countless scientific studies and researches,
because of its importance and relationship with one of the most important "senses", that is the
smell. Nevertheless, and although it may seem contradictory, the mechanism of the sense of
smell, from the functional point of view, still lacks explanations in the context of its
neurophysiology, in spite of studies undertaken in this area. So, even nowadays some
reinterpretations of science, morphological e functional comparisons and, above all,
comparative studies, are important to establish and clarify, increasingly, its important
functions with regard to animal behavior, their survival and, above all, through the smell and
odor, often associated with the taste and also the important neural, centers establishing the
individual, whether animal or human, in their proper position, within the scale or zoological
phylogenetic. Therefore, the main goal of the present study is to present a literature review,
where some morphological and functional aspects can be demonstrated evolutionary and
chronologically by some important researchers in this area of study, with the aim of a
comparative study more comprehensively, in addition to compare functional aspects of
evolution between the two studied species of mammals, the dog (Canis familiaris).
Keywords: Olfactory Nerve, Olfactory bulb, cribriform plate, smell.

9
Lista de Figuras
Figura 1. Crânio em corte sagital no qual está sendo evidenciado o Osso etmóide do cão.
(Foto da autora).........................................................................................................................14
Figura 2. Localização do osso etmóide no homem. Fonte: Wikipédia.....................................18
Figura 3. Vista superior do osso etmóide humano (SOBOTTA; BECHER, 1977)..................18
Figura 4. Superfície caudal da LC do cão. Adaptado de Alvites (2015)..................................19
Figura 5. Representação da Lâmina Crivosa no homem, adaptado de Sobotta & Becher
(1977)........................................................................................................................................21
Figura 6. (A). Lâmina Crivosa. (B). Crista Galli. Humano (Foto da autora)............................21
Figura 7. Encéfalo de um cão no qual se evidencia o rinencéfalo. Fonte: Blog da monitoria de
anatomia topográfica do curso de veterinária da Universidade de Uberaba (adaptado).......... 22
Figura 8. Bulbo Olfatório do cão. (Foto da autora)..................................................................24
Figura 9. Bulbo olfatório humano. (Foto da autora).................................................................27
Figura 10. Representação das células sensoriais do bulbo olfativo (adaptado de Lent,
2010).........................................................................................................................................34

10
Lista de abreviações e siglas
BO – Bulbo olfatório
CG – Crista Galli
LC – Lâmina crivosa
MO – mucosa olfatória
NO – Nervo Olfatório
OE – Osso etmóide
OF – osso frontal
CEU - Comissão de Ética
DMOR – Departamento de Morfologia

11
SUMÁRIO
1 INTRODUÇÃO ............................................................................................................... 11
2 OBJETIVOS .................................................................................................................... 14
2.1 OBJETIVOS ESPECÍFICOS ............................................................................................ 14
3 METODOLOGIA ........................................................................................................... 15
4 ASPECTOS ANATÔMICOS OSSO ETMÓIDE DO CÃO E DO HOMEM ............ 16
4.1 ASPECTOS ANATÔMICOS DA LÂMINA CRIVOSA DO OSSO ETMÓIDE DO CÃO
E DO HOMEM ................................................................................................................. 19
5 RINENCÉFALO ............................................................................................................. 22
6 BULBO OLFATÓRIO .................................................................................................... 23
6.1 BULBO OLFATÓRIO DO CÃO E DO HOMEM ........................................................... 24
7 ÓRGÃO VOMERONASAL ........................................................................................... 28
8 ALTERAÇÕES NO SISTEMA OLFATÓRIO NO CÃO E NO HOMEM ............... 30
9 FISIOLOGIA DO OLFATO .......................................................................................... 32
9.1 PLASTICIDADE NEURAL ............................................................................................. 34
10 ASPECTOS FILOGENÉTICOS ................................................................................... 35
11 DISCUSSÃO .................................................................................................................... 36
12 CONCLUSÃO ................................................................................................................. 39
REFERÊNCIAS .............................................................................................................. 40

12
1 INTRODUÇÃO
Os animais terrestres vivem em um ambiente repleto de moléculas as quais não podem
ser vistas a olho nu. Essas moléculas estão presentes no ar e são advindas dos mais diversos
fatores e podem ser sentidas através do olfato. A capacidade de detectar as substâncias
presentes no ar é uma característica primitiva que alerta os animais - inclusive o ser humano -
sobre as mais diversas moléculas que podem estar no ambiente, por exemplo, alimento, perigo
e até mesmo parceiro sexual (MARIEB; HOEHN, 2009). O nariz é o órgão responsável pela
captação dos odores que são conduzidos pelos quimiorreceptores (existentes na mucosa nasal)
até o bulbo olfatório (LENT, 2010). A percepção dos químicos que estão a certa distância do
corpo ocorre por intermédio das células quimiorreceptoras que constituem o sistema olfatório
dos Vertebrados (MOYES; SCHULTE, 2010).
Estudos comparativos e evolutivos no mundo dos Vertebrados, em especial dos
Mamíferos, mostram que apesar do olfato do ser humano tem assumido, ao longo do tempo,
características não somente adaptativas como também seletivas, quando analisados diversos
tipos de cheiros e odores considerados, passam a ser escolhas pessoais e coletivas. Bem como
suas necessidades e prazeres, especialmente para aqueles que, circunstancialmente, tenham
perdido algum dos outros órgãos dos sentidos. Quando comparada essa capacidade olfatória
do homem (Homo sapiens) com as de outros mamíferos, especialmente a do cão (Canis lupus
familiaris), percebe-se uma maior eficácia do olfato desse animal em relação à do homem
(LOURENÇO; FURLAN, 2007).
O sentido do olfato no ser humano, provavelmente é o menos compreendido, pois
podem existir anormalidades entre o olfato e o paladar. Porque a superfície receptora dos
aromas está localizada na região superior da cavidade nasal e, geralmente, sua área mede
apenas 2,4 cm2 (PALHETA NETO et al., 2011).
O bulbo olfatório do cão é bem maior em comparação ao do homem. No entanto,
apenas este, não pode ser o único aspecto levado em consideração em relação às habilidades
olfativas do cão. Atualmente, se discute a questão do olfato sob vários aspectos científicos,
tais quais fisiológicos e bioquímicos. Não obstante, é inegável que, o devido conhecimento
estrutural anátomofuncional seja imprescindível para quaisquer outros estudos posteriores,
uma vez que, sem o conhecimento estrutural e orgânico, não seria possível analisar seus
aspectos funcionais e evolutivos.

13
Contudo, este trabalho desenvolveu uma de revisão bibliográfica sobre o bulbo
olfatório do cão, comparado ao do homem, em seus aspectos morfofuncionais, do ponto de
vista estrutural e orgânico, observando seus aspectos evolutivos e sua primordial importância
para os cães e para os humanos.

14
2 OBJETIVOS
Dado a importância de estudos comparativos sobre aspectos evolutivos morfofuncionais do
bulbo olfatório do cão e do homem, o principal objetivo deste trabalho é apresentar uma
consistente revisão bibliográfica para avaliar relações anátomofuncionais da estrutura em
discussão com sua função olfativa e sua importância evolutiva.
2.1 OBJETIVOS ESPECÍFICOS
Observar os aspectos anatômicos do osso etmóide do cão e do homem;
Verificar os aspectos anatômicos da lâmina crivosa do osso etmóide;
Mostrar o rinencéfalo;
Evidenciar a importância do órgão vomeronasal;
Considerar algumas alterações do sistema olfatório do cão e do homem;
Analisar a fisiologia do olfato;
Perceber os aspectos filogenéticos.

15
3 METODOLOGIA
O presente trabalho teve como meta a revisão bibliográfica para constar os aspectos
comparativos do BO. Foram utilizadas busca por periódicos informatizados para a coleta de
dados, usando as palavras “bulbo olfatório”, “bulbo olfatório do cão” e “olfato no cão” em
Português e “olfactory bulb” em Inglês. Realizou-se também busca em publicações científicas
e livros acadêmicos.
Apesar de ser um trabalho de revisão foi indispensável à realização de um estudo
prático com o método tradicional, apenas para serem fotografadas as estruturas
correlacionadas com o bulbo olfatório do cão e do homem, inclusive o próprio bulbo
olfatório. Portanto, foram utilizados um crânio e três encéfalos de cães, de médio porte, de
raça indefinida e, igualmente um crânio e três encéfalos dissecados de cadáveres
desconhecidos, anteriormente doados para o DMOR para fins de estudos didáticos.
É importante observar que os encéfalos, tanto de cães como de humanos, utilizados
neste trabalho eram peças do acervo cedidas pelas Disciplinas de Anatomia Comparada dos
Vertebrados e Anatomia Humana, do Departamento de Morfologia do Centro de Biociências.
Portanto, não foi necessário à aprovação pela CEU- Comissão de Ética no uso de animais
local, uma vez que não houve utilização de animais de laboratório, para este fim.

16
4 ASPECTOS ANATÔMICOS OSSO ETMÓIDE DO CÃO E DO HOMEM
Para estudo, pesquisa e compreensão do Bulbo Olfatório (BO), é necessário
reconhecimento da anatomia, topografia e suas associações estruturais, assim como uma breve
revisão teórico-prática desta estrutura, uma vez que a relação crânio-encefálica tem grande
importância morfofuncionalmente. Portanto, visto que o BO tem suas relações estruturais,
especialmente com o Osso Etmóide (OE), sobre o qual repousa, a identificação destas
estruturas se faz necessário, caso as mesmas sejam acometidas de agressões físicas, patologias
ou quaisquer outros danos que possam repercutir diretamente na atuação e função olfatórias.
Situado entre as regiões facial e cranial o OE (Figura 1) é uma estrutura complexa a
qual não pode ser visualizada, em um crânio intacto, portanto, seu estudo ocorre com peças
desarticuladas do crânio. Apesar de ser um osso único, suas porções se desenvolvem em
cortes e ele se encontra organizado entre as paredes das duas órbitas sendo margeado
posteriormente pelo osso frontal, pelo maxilar na região lateral e, pelos ossos vômer e
palatino na parte rostral. Também podemos dizer que ele está composto por quatro porções:
lâmina perpendicular mediana, duas partes laterais cobertas pela lâmina externa e a lâmina
crivosa (EVANS, 1993).
Figura 1. Crânio em corte sagital no qual está sendo evidenciado o Osso etmóide do cão. (Foto da autora).

17
A lâmina perpendicular é uma camada vertical mediana, tendo o vômer abaixo e os
processos dos ossos frontal e nasal, logo acima, formam o septo do osso nasal. Este septo
ósseo é continuado rostralmente pelo septo da cartilagem nasal. Caudalmente, ele se fusiona
com a lâmina crivosa, porém normalmente se continua para formar a Crista Galli (CG),
formando ventralmente parte do septo nasal como as lâminas dos ossos frontal e nasal
imergindo parcialmente para baixo, à medida que as lâminas dos septos dos ossos frontal e
nasal continuam parcialmente para baixo e se fusionam ao mesmo. A lâmina externa
desenvolve-se da porção linear do fundo nasal. Ela é extremamente fina e, é deficiente nas
áreas quando reveste a superfície interior dos ossos mais pesados que formam esta parte da
face. Esta lamina é dividida em partes dorsal, lateral e ventral, comumente chamada de teto,
lado e soalho, das massas laterais respectivamente. De sua origem na lâmina perpendicular
externa dirige-se dorsalmente em contato com as partes frontal e nasal do septo, gira
lateralmente sobre o labirinto etmoidal, formando as lâminas do teto e desce cada lado,
lateralmente. Esta porção da lamina é muito fina, incompleta em algumas áreas e porosa. Sua
parte rostrolateral é canalizada para formar o processo uncinado, a qual é parte da primeira
endoturbinada, tão bem quanto a lamina orbital. A incisura uncinada no meato entre as duas
primeiras endoturbinadas, está localizada dorsocaudal ao processo uncinado. A área com
depressão da lâmina da órbita, a fossa maxilar, forma a parede medial do recesso maxilar
(EVANS, 1993).
A lâmina externa é caudalmente deficiente, atuando apenas como uma fina camada,
placas irregulares que permanecem atadas a lâmina basal das linhas do osso. As turbinas
individuais dirigem-se do teto e da porção lateral desta delicada cobertura. A lâmina ventral
ou transversa, a qual forma o soalho da lâmina, pode ser isolada como uma folha fina e macia
fusionada à superfície medial da maxila. Ela continua da parte ventral da lâmina orbital
medialmente para o osso vômer num arco transverso e dorsalmente convexo. Ela está
proximalmente junta à parte horizontal do vômer. O labirinto etmoidal forma a maior parte da
massa lateral. Ela é composta a partir de robustas linhas ósseas, as quais se aderem à lâmina
externa através da lamina basal e adere-se caudalmente à lâmina crivosa. Da mesma forma, a
lâmina crivosa adere-se dorsalmente à plataforma mais interna do osso frontal (OF), o qual
em crânios longos e velhos é separado da plataforma exterior por mais de 2 cm.
Anatomicamente considerando, fica muito claro que, várias estruturas similares entre o
homem e o cão, apresentam suas especificidades decorrentes, principalmente, das diferentes
posições anatômicas peculiares a ambos, uma vez que a posição bípede e a quadrúpede,

18
embora seccionadas pelos mesmos eixos e planos, mantém não apenas diferenças em alguns
termos, porém muitas variações anatômicas decorrentes de suas adaptações às suas posições
específicas. Podemos falar em relação à gravidade para cujo centro não apenas os corpos dos
indivíduos convergem, mas principalmente a distribuição do sentido das estruturas orgânicas
se ajusta, fazendo com que ocorram muitas adaptações estruturais, principalmente levando-se
em conta o meio ambiente e o tipo de vida dos indivíduos (EVANS, 1993).
No ser humano, o OE é um osso extremamente esponjoso e leve (Figura 2 e 3). Está
localizado anteriormente na base do crânio, entre as duas órbitas, formando a parte das
paredes da região superior da cavidade nasal. O OE é constituído de quatro partes: lâmina
crivosa, lâmina perpendicular, e duas massas laterais (GRAY, 1977).
Figura 2. Localização do osso etmóide no homem. Fonte: Wikipédia.
Figura 3. Vista superior do osso etmóide humano (SOBOTTA; BECHER, 1977).
Lâmina crivosa
com forames

19
4.1 ASPECTOS ANATÔMICOS DA LÂMINA CRIVOSA DO OSSO ETMÓIDE DO CÃO
E DO HOMEM
No cão os aspectos anatômicos, estruturais e orgânicos básicos, assemelham-se de
modo geral, aos do crânio humano, devendo, portanto considerar as variações anatômicas
pertinentes e, em razão, principalmente da posição anatômica do quadrúpede. É importante
também observar o crânio como um todo, inclusive sua cavidade craniana, forma e situação
topográfica, uma vez que os bulbos olfatórios se projetam para a Lâmina Crivosa (LC) do
osso etmóide, tal qual ocorre no ser humano (EVANS, 1993).
Por outro lado, diferentes raças caninas apresentam grandes variações crânio-
encefálicas, há aquelas que têm um crânio longo e estreito que são denominados
dolicocéfalicos. Outros dispõem de um crânio largo e curto, chamados de braquiocefálicos. E
os crânios com uma formação intermediária a estes são os mesaticefálicos. Normalmente a
extensão do crânio é medida a partir da crista nucal até o limite anterior da sutura
interincisiva, e a largura entre os ápices dos arcos zigomáticos (GETTY, 1975). Na pesquisa
de Alvites (2015) observou seis crânios de cães com idade, sexo e raça desconhecidos e
percebeu que nenhuma das LC analisadas, apresentou total simetria entre as duas metades e
que de fato as duas partes da lâmina se distinguem tanto pela localização e arranjo dos
forames entre si quanto pelo número total dos mesmos. O lado direito da lâmina tende a
possuir um número maior de forames, em consequência do seu tamanho que se torna superior
ao lado esquerdo (Figura 4).
Figura 4. Superfície caudal da LC do cão. Adaptado de Alvites (2015).

20
Na cavidade craniana é encontrado o encéfalo com seus envoltórios e vasos. Na
porção denominada fossa craniana está situado o BO e os seus tratos, e as estruturas restantes
dos lobos frontais do cérebro. Essa fossa repousa numa área mais alta e mais estreita que o
seu soalho craniocaudal e seguido anteriormente pela fossa etmoidal profunda. Assim como
no homem, o OE do cão também é provido da LC, que pode ser equiparada a uma peneira por
conter cerca de 300 forames cribiformes, alguns medem até 15mm de diâmetro, tais orifícios
permitem que os feixes de fibras nervosas, dos nervos olfatórios, atravessem a lâmina até a
mucosa olfatória (MO). Nas bordas laterais situadas mais perto do plano sagital mediano do
que do plano transverso, se forma uma depressão profunda. A LC é uma estrutura côncava e
bem proeminente na parte rostral que se articula posteriormente com as fissuras do osso
frontal e, ventral e dorsalmente com o osso pré-esfenoide. As duas metades da LC se
delimitam em planos sagitais e se apresentam na porção anterior num ângulo de 45°.
Diferentemente dos humanos, no cão a CG aparece apenas em cães idosos e sua crista vertical
mediana geralmente se encontra na metade da porção rostral da fossa etmóide. Grande parte
dos espécimes possui uma linha que aponta a borda caudal da lâmina perpendicular do osso
etmóide, substituindo a CG (EVANS, 1993).
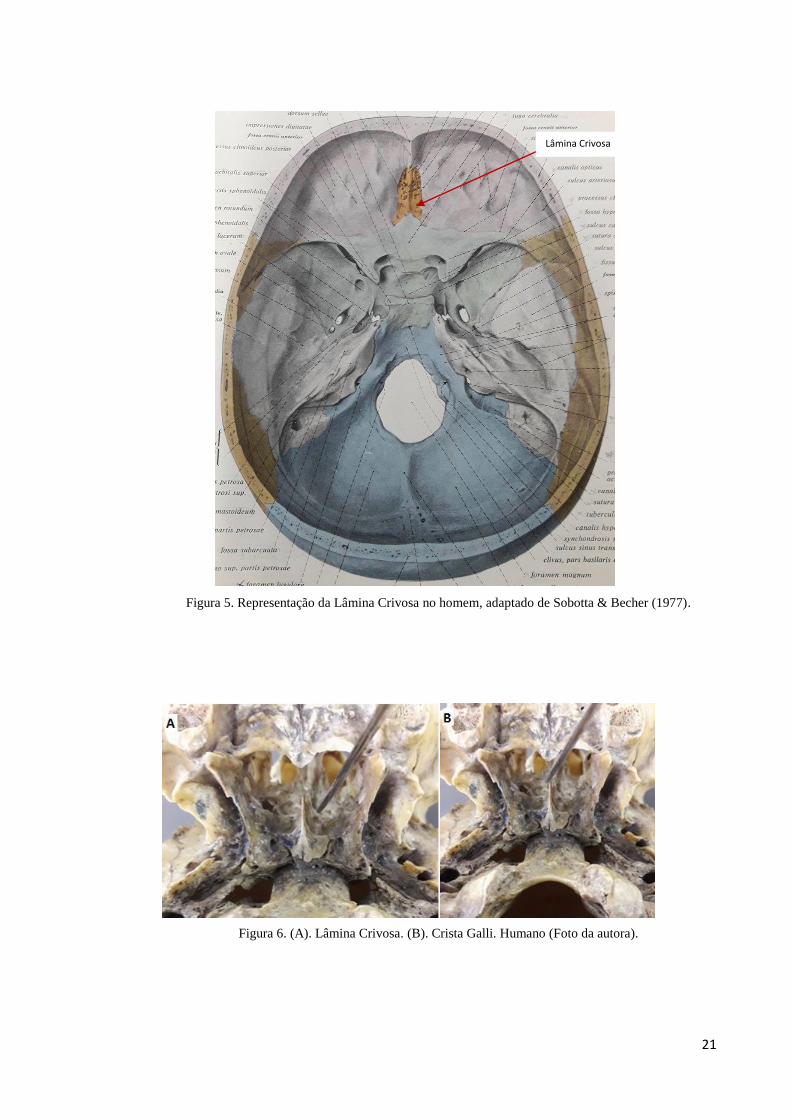
No homem, a LC está situada na incisura etmoidal do osso frontal (OF) formando o
teto da cavidade nasal (Figuras 5 e 6). Localiza-se na região superior interna da fossa
craniana, nela se observa um seguimento triangular na linha mediana dessa placa, que é a CG
(Figura 6). Por sua vez, essa crista possui uma borda posterior longa e fina que dá suporte a
inserção da foice do cérebro, já a borda anterior se apresenta curta e espessa, ela se articula
com o OF, predispondo duas asas pequenas, as quais fazem ligação em depressões
correspondentes do OF completando o forame cego. A LC se torna estreita nas laterais da CG,
apresentando vários sulcos, ele dá suporte ao BO e permite a passagem dos nervos olfatórios
através dos orifícios chamados de forames cribiformes (GRAY, 1977).
21
Figura 5. Representação da Lâmina Crivosa no homem, adaptado de Sobotta & Becher (1977).
Figura 6. (A). Lâmina Crivosa. (B). Crista Galli. Humano (Foto da autora).
Lâmina Crivosa
22
5 RINENCÉFALO
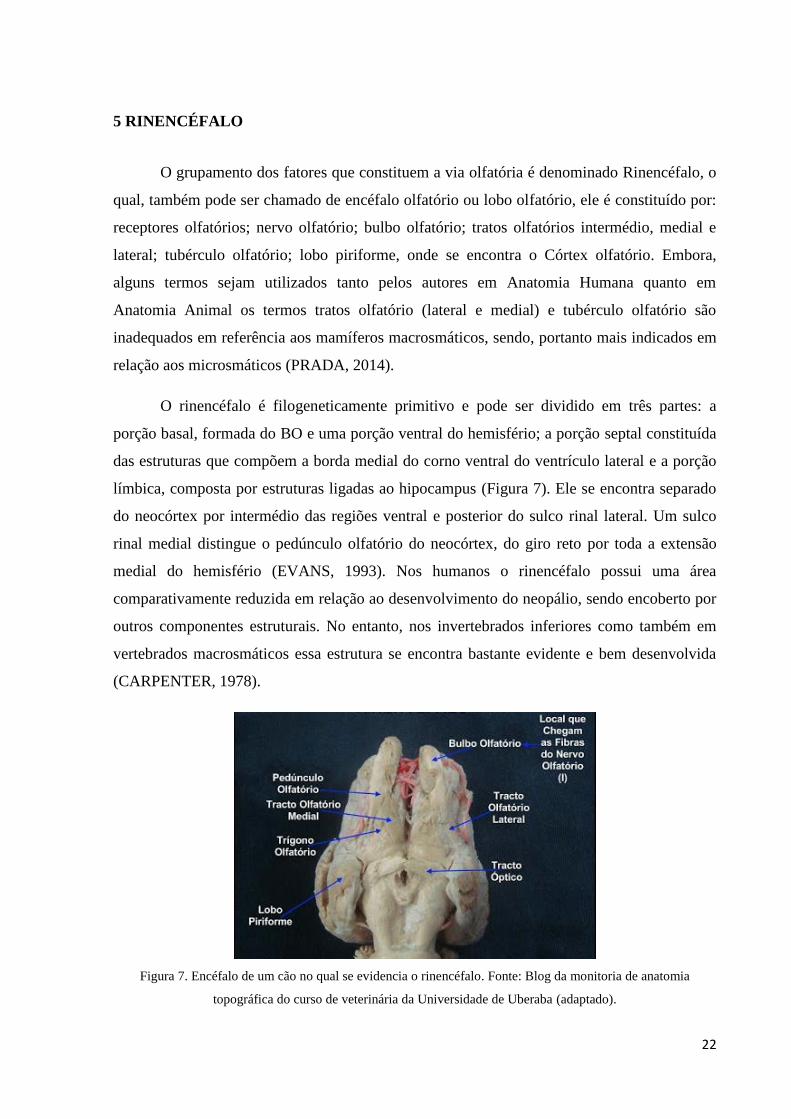
O grupamento dos fatores que constituem a via olfatória é denominado Rinencéfalo, o
qual, também pode ser chamado de encéfalo olfatório ou lobo olfatório, ele é constituído por:
receptores olfatórios; nervo olfatório; bulbo olfatório; tratos olfatórios intermédio, medial e
lateral; tubérculo olfatório; lobo piriforme, onde se encontra o Córtex olfatório. Embora,
alguns termos sejam utilizados tanto pelos autores em Anatomia Humana quanto em
Anatomia Animal os termos tratos olfatório (lateral e medial) e tubérculo olfatório são
inadequados em referência aos mamíferos macrosmáticos, sendo, portanto mais indicados em
relação aos microsmáticos (PRADA, 2014).
O rinencéfalo é filogeneticamente primitivo e pode ser dividido em três partes: a
porção basal, formada do BO e uma porção ventral do hemisfério; a porção septal constituída
das estruturas que compõem a borda medial do corno ventral do ventrículo lateral e a porção
límbica, composta por estruturas ligadas ao hipocampus (Figura 7). Ele se encontra separado
do neocórtex por intermédio das regiões ventral e posterior do sulco rinal lateral. Um sulco
rinal medial distingue o pedúnculo olfatório do neocórtex, do giro reto por toda a extensão
medial do hemisfério (EVANS, 1993). Nos humanos o rinencéfalo possui uma área
comparativamente reduzida em relação ao desenvolvimento do neopálio, sendo encoberto por
outros componentes estruturais. No entanto, nos invertebrados inferiores como também em
vertebrados macrosmáticos essa estrutura se encontra bastante evidente e bem desenvolvida
(CARPENTER, 1978).
Figura 7. Encéfalo de um cão no qual se evidencia o rinencéfalo. Fonte: Blog da monitoria de anatomia
topográfica do curso de veterinária da Universidade de Uberaba (adaptado).

23
6 BULBO OLFATÓRIO
O bulbo olfatório (BO) em mamíferos varia consideravelmente em forma e tamanho
relativo. É muito grande em monotrômetros e insetívoros, de tamanho moderado na maioria
dos amalhados, pequenos e pedunculados em primatas e ausentes em alguns cetáceos
(ALLISON, 1953). O BO possui um formato ovoide achatado e, encontra-se prostrado em
cima da Lâmina Crivosa do Osso Etmóide (CARPENTER, 1978).
O Nariz dos mamíferos em geral possui uma anatomia bem adaptada para que o ar
inspirado seja bem conduzido dentro da cavidade nasal, que é revestida por uma mucosa, na
qual ficam inseridos os quimiorreceptores olfatórios. Na mucosa existem glândulas
produtoras de uma secreção serosa, esta secreção é denominada muco olfatório e nele se
dissociam previamente as moléculas aromáticas para posterior contato com os neurônios
receptores presentes no trato olfatório. Os quimiorreceptores olfatórios penetram na cavidade
craniana pelos orifícios da LC e seguem formando filetes nervosos que constituem o Nervo
Olfatório (NO) e dão origem ao BO (LENT, 2010).
O BO corresponde ao segmento final do NO, ele integra o Sistema Nervoso Central e
reflete evaginações do Telencéfalo (CARPENTER, 1978). Esse Bulbo é ainda o responsável
por transportar os estímulos iniciados nos quimiorreceptores nasais, esses estímulos, passam
por todo o trato olfatório e seguem até a seção medial do lobo temporal do cérebro. Tornando
assim, o sentido do olfato depende totalmente da funcionalidade do NO (LUNDY-EKMAN,
2000). Anatomicamente, o NO, está diretamente ligado ao telencéfalo e tem sua origem
aparente do encéfalo no BO e, em relação à área óssea, no crânio, e mais precisamente na LC.
O NO é constituído por fibras aferentes que se formam fora do encéfalo, a partir de
grupos de células neurais que geralmente incidem em um gânglio ao longo do nervo. Assim,
os processos centrais das células neurais adentram no encéfalo, formando grupos celulares
denominados núcleos sensitivos (O’RAHILLY, 1985). Esse nervo ainda pertence ao grupo
dos órgãos dos sentidos especiais, por possuir receptores denominados “especiais” que se
diferenciam dos outros receptores delimitados pelo corpo chamados de gerais. A partir de
receptores gustativos e olfatórios classificados como viscerais, nascem às fibras aferentes
viscerais especiais, as quais são os elementos “especiais”, que se conceituam em viscerais, por
estarem situados nos sistemas viscerais, como por exemplo, o digestório (MACHADO, 1998).

24
O I Par de Nervos Cranianos é classificado como um nervo sensorial específico, uma vez que
possui uma função específica que é o sentido do olfato (LUNDY-EKMAN, 2000).
6.1 BULBO OLFATÓRIO DO CÃO E DO HOMEM
O bulbo olfatório do cão é bem evidente, ele aparece como um prolongamento ventral
do pedúnculo olfatório, ambos são rasos e contêm um ventrículo olfatório que ocasionalmente
retém uma ligação com os cornos ventral e lateral do ventrículo, e o pedúnculo é o
responsável em fazer a ligação do BO com o cérebro (Figura 8). A parede do BO é de fora
para dentro, constituída, por sete camadas histológicas entre as quais: a primeira camada é o
NO; a segunda é uma camada glomerular; a terceira é a camada plexiforme externa com
células em tufos; a quarta é a camada de células mitrais consistindo de células cônicas
organizadas em fila; a quinta é a camada plexiforme interna delgada; a sexta é a camada de
células granulares; e a sétima é uma camada perivascular formada por axônios de projeção
que entram ou saem do bulbo (EVANS, 1993).
Figura 8. Bulbo Olfatório do cão. (Foto da autora).
A partir de um entrelaçamento das terminações das células olfativas e dos dendritos
das células mitrais formam-se os glomérulos, sendo independentes uns dos outros, não
ocorrendo anastomose. As fibrilas olfativas do cão se encontram livres no glomérulo, e cada
glomérulo recebe uma grande quantidade de células mitrais. Tanto no homem quanto no cão,
um grande número de neurônios bipolares está conectado a uma célula mitral, mas cada célula

25
olfativa da mucosa está ligada apenas uma, e raramente a duas células mitrais, e em geral no
glomérulo cada célula olfativa termina com apenas uma célula mitral. Em todos os animais
em que o sentido do olfato é maior e cada célula bipolar pode estar em contato com várias
células mitrais, não pela bifurcação da fibra que se conecta a células distintas, mas porque
podem ser encontrados dendritos de várias células mitrais no mesmo glomérulo (GOLGI,
1875 apud READ, 1908).
As glândulas olfativas também conhecidas por glândulas de Bowman, foram
reconhecidas como um dos traços característicos da mucosa olfativa em todos os vertebrados
terrestres. Essas glândulas se mostram como uma extensão do epitélio olfatório e desaparecem
abruptamente no início da mucosa respiratória. Elas têm um formato simples túbulo-alveolar
brevemente ramificada e com ductos curtos que se abrem para a superfície do epitélio olfativo
em intervalos regulares. As células que constituem as glândulas possuem um formato
triangular ou colunar e são coradas pelo azul de metileno. A secreção das glândulas é liberada
uniformemente em todo o epitélio olfativo e em nenhum outro lugar, ela banha as expansões
terminais das células receptoras, com seus prolongamentos minuciosos. E consequentemente,
as partículas que envolvem odor se dissolvem nessa secreção, e estimulam a ação das células
sensoriais (ALLISON, 1953).
No interior do bulbo olfatório existe uma lacuna que diz respeito à expansão do corno
anterior do ventrículo lateral correspondente. Em sua parede rostral, mais espessa, é
encontrada unicamente substância cinzenta e, ao mesmo tempo, sua parede dorsal, mais
delgada, é provida apenas de substância branca composta por fibras advindas das laterais do
bulbo olfatório e seguem no trato olfatório intermédio. Assim como no humano, o caminho da
olfação tem início com as células olfatórias viscerais aferentes da mucosa, estas células são
neurônios do tipo bipolares presentes na mucosa olfatória, com fibras centrais desprovidas de
mielina que cruzam os forames da lâmina crivosa do osso etmóide chegando ao bulbo
olfatório (PRADA, 2014).
Segundo Read (1908) os nervos olfatórios podem ser visualizados claramente de
forma irregular sobre a lâmpada olfativa, em secções transversais e sagitais do bulbo olfatório
e da mucosa olfatória do cão. As fibras individuais podem ser rastreadas por uma distância
considerável e, em alguns casos quase através da lâmina crivosa. O autor não conseguiu
visualizar os nervos bifurcados na camada, porém descreve que estes permanecem como

26
fibras individuais próximas ao glomérulo, mas a sua entrada se subdividem formando muitos
ramos que se entrelaçam, eles não se anastomosam com outros tipos de fibras lá encontradas.
O nervo olfatório do cão está posicionado ventralmente ao diencéfalo. Os filetes
olfatórios conectam-se com o hipotálamo e também com outras regiões do sistema límbico
para gerar um feedback comportamental adequado para o olfato (CHRISMAN, 1985).
Axônios de neurônios mitrais e em tufos deixam o bulbo como tratos olfatórios (células
mitrais ativam aminoácidos excitatórios como neurotransmissores. A camada de células
granulares recebe feedback do núcleo ao longo do trajeto e do bulbo olfatório contralateral.
Células granulares inibem as células em tufos, as quais excitam reciprocamente as células
granulares (EVANS, 1993).
A lâmpada olfativa é constituída pelas camadas de fibras nervosas olfativas, pelos
glomérulos e pelos feixes de fibras olfatórias sendo delimitada no interior por meio das
células mitrais. A célula mitral é grande, possui apenas um dendrito principal, que atinge um
único glomérulo, e uma série de dendritos acessórios que terminam na camada plexiforme. Os
feixes são de tamanho médio e, assim como a célula mitral, possuem um único dendrito
principal que se finaliza em um glomérulo e um ou mais dendritos acessórios na camada
plexiforme externa. Na camada plexiforme interna os axônios de células mitrais e dos feixes
passam de volta ao caminho olfatório secundário, e um grande número de dendritos
recorrentes viajam horizontalmente por uma curta distância antes de virar para fora para
terminar na camada plexiforme externa. Na camada granular encontram-se as células
granulares, envolvidas em um denso plexo de fibras finas que, como na camada
periventricular, parece derivar em grande parte de fontes extrabulbar. Os axônios das células
granulares e algumas células grandes na quinta camada termina na camada plexiforme
externa, juntamente com colaterais recorrentes de células mitrais e tufadas, em relação aos
dendritos de células mitrais e tufadas. Os glomérulos nas proximidades são relacionados por
células periglomerulares de axônio curto (ALLISON, 1953).
As fibras nervosas olfativas terminam em corpos esféricos bem definidos, os
glomérulos. Essas estruturas podem ser consideradas como a característica mais diferenciada
da lâmpada olfativa, formando um estrato claramente definido perto da periferia da lâmpada.
Além disso, a sinapse de alta entre as séries primária e secundária de neurônios olfativos
ocorre nos glomérulos, e isso por si só deve indicar que têm um significado fisiológico
considerável. Eles são limitados e parecem ter a capacidade de atuar de maneira independente.

27
Portanto, o glomérulo atua para efetuar soma espacial, e isso o torna responsável pela
funcionalidade do sentido do olfato. Ao mesmo tempo, o sistema glomerular pode funcionar
de maneira independente, desempenhando um papel importante no mecanismo central da
olfação, porque eles são capazes de funcionar em caminhos fechados. Apesar de essas
considerações serem generalizadas, podem ser muito limitadas para os mamíferos, em que se
sugere que o tamanho dos glomérulos varia em proporção quase direta ao tamanho da
lâmpada olfativa, de modo que o número total de glomérulos permanece bastante constante
(ALLISON, 1953).
O bulbo olfatório humano tem basicamente o formato de um gânglio e está
estrategicamente posicionado na base do encéfalo, sobrepondo-se sobre a lâmina crivosa ou
placa crivosa do osso etmóide a qual seus orifícios permitem a passagem das fibras do nervo
olfatório até a mucosa olfatória (Figura 9). As informações que chegam ao bulbo olfatório
seguem diretamente para o córtex cerebral sem passar pelo tálamo. A área inervada é extensa
e forma o chamado córtex piriforme um tipo de córtex primitivo, que por sua simplicidade é
denominado paleocórtex o qual se comunica com o tálamo, e este posteriormente, faz ligação
com o lobo frontal do neocórtex. Contudo, considera-se essa rota como a promotora dos
aspectos conscientes da via sensorial olfatória (LENT, 2010).
Figura 9. Bulbo olfatório humano. (Foto da autora).
No encéfalo humano o NO projeta-se em ambos os lados da cavidade nasal por meio
dos inúmeros feixes de fibras nervosas que são a extensão dos neurônios da mucosa olfatória
do nariz (GRAY, 1977). Read (1908) afirma que as células sensoriais presentes na mucosa
são as células nervosas os seus prolongamentos centrais são os NO, que se estendem para

28
formar glomérulos olfativos no bulbo olfatório. Ele ainda descreve que, em humanos, os NO
se estendem por uma área que mede cerca de um terço do septo e quase toda concha superior,
e que os nervos se prolongam em feixes não anastomosantes para a lâmpada olfativa.
Os quimiorreceptores do olfato são neurônios do tipo bipolar, que se estendem até a
superfície do epitélio presente na cavidade nasal, e se ramificam em cílios olfatórios que se
encontram no muco da mucosa olfatória. As extensões centrais dos neurônios são mais
delgadas e centenas delas formam os filetes olfatórios, os quais são pequenos feixes de fibras
nervosas amielínicas. Existem aproximadamente vinte filetes de cada lado que se prolongam
em glomérulos do bulbo olfatório, uma estrutura oval medindo cerca de 3 mm x 15 mm, que
repousa sobre a lâmina crivosa do osso etmóide. Esses filetes olfatórios acabam formando o
nervo olfatório propriamente dito (GRAY, 1977). Portanto, é impossível falar do bulbo
olfatório sem citar o nervo olfatório, pois essas estruturas estão totalmente interligadas.
Read (1908) verificou a lâmpada olfatória no homem, apenas em preparações
grosseiras. Foram feitas análises dos nervos olfatórios da lâmina crivosa do osso etmoide para
a camada externa do bulbo olfatório e, observou-se que os glomérulos se formam a partir de
um cruzamento dos axônios dos neurônios olfativos e os dendritos das células mitrais, do
mesmo modo que nos animais inferiores. Cada axônio advém de uma célula olfativa
individual, mas os dendritos podem surgir de um único axônio ou de várias células mitrais.
Foi observado que os dendritos de diferentes células mitrais no cão, migram para um
glomérulo e um único dendrito se ramificou para três glomérulos diferentes.
7 ÓRGÃO VOMERONASAL
Pode se observar que na maioria dos mamíferos existe a presença de um bulbo olfativo
acessório, denominado órgão vomeronasal (OV), como parte do seu sistema olfatório, essa
estrutura é um tanto similar em suas conexões periféricas para o principal BO, mesmo sendo
claramente bem menor quando separado dele (McCOTTER, 1912). Essa estrutura foi
descoberta pelo cirurgião dinamarquês Jacobson, em 1811, é um órgão pequeno e atraiu o
interesse de muitos anatomistas comparativos na época (ALLISON, 1953). É possível
perceber que esse órgão possui uma função olfativa e promove a formação de filamentos
nervosos, como os da mucosa olfatória, que formam feixes que atravessam a LC para
alcançarem a região do NO (McCOTTER, 1912).

29
Read (1908) observou as estruturas olfativas do gato, do cão e do humano e nas suas
pesquisas percebeu que as inervações do OV se encontraram apenas nos epitélios do cão e do
gato. Allison (1953) afirmou que esse órgão geralmente tem a forma de um tubo longo e fino
deitado no sobre o assoalho da cavidade nasal ao lado do septo nasal e protegido por osso ou
cartilagem e ele é parcialmente revestido pelo epitélio sensorial. Essa estrutura está ausente ou
é apenas vestigial em humanos.
Após remover cuidadosamente a pia-máter se observa os nervos vomeronasais que
surgem em grande número de filamentos da superfície mediana do órgão vomeronasal. Esses
filamentos se unem e formam oito cordas nervosas que deixam a borda dorsal do órgão. Num
breve afastamento, se unem gerando quatro grandes feixes. Os quatro feixes passam
caudalmente e pela submucosa do septo nasal até a lâmina crivosa do osso etmóide. Eles
entram no crânio por duas aberturas e depois se unem em um único tronco de nervo, que
passa em torno da margem espessa mediana da lâmpada olfativa, onde, girando lateralmente
em sua superfície dorsal, do nervo sai uma rede de fibras que se inserem no bulbo olfativo
acessório. Na superfície dorsal do bulbo olfativo, os nervos vomeronasais são intimamente
aplicados na camada de fila olfatória (McCOTTER, 1912).
SALAZAR; CIFUENTES; SÁNCHEZ-QUINTEIRO (2013) ainda afirmam que o OV
integra o ducto vomeronasal que está rodeado de tecido mole anexado pelo tecido conjuntivo,
juntamente com uma considerável quantidade de glândulas, vasos e nervos. Tais estruturas
estão cobertas por uma lâmina cartilaginosa incompleta, que é envolvida externamente pela
mucosa da cavidade nasal. O ducto vomeronasal pode ser isolado por meio de dissecção e
microdissecção, visto que mede aproximadamente 80mm e sua extremidade anterior se abre
para o ducto incisivo. Esse ducto se comunica com as cavidades nasais e bucais. Esta
comunicação se torna bem evidente por causa da papila incisiva que é uma proeminência
específica da cavidade oral. Em contraste, na cavidade nasal, o ducto incisivo é difuso e pode
ser considerado como a projeção da mucosa, que cobre as bordas da fissura palatina. Além
disso, a parte anterior da cartilagem vomeronasal está associada à mesma fissura, e sua
extensão por meio da papila incisiva promove um reforço incompleto do ducto incisivo.
McCotter (1912) afirma que no cão, a grande lâmpada olfativa possui um formato de
quadrilátero e que ela é comprimida pela região frontal do cérebro de um lado e a estrutura do
nasal do outro. A superfície dorsocaudal é côncava e moldada no limite anterior do lobo
frontal. O pedúnculo olfativo se estende para a região posterior a partir da parte ventrolateral

30
desta superfície. A região anteroventral situa-se contra a lâmina crivosa de maneira oblíqua.
Ela recebe os nervos olfatórios que se espalham em uma rede espessa sobre a superfície da
lâmpada olfativa. A margem mediana espessada da lâmpada olfativa é separada do bulbo
oposto pela extensão dorsal da placa perpendicular. A lâmpada olfativa acessória é menor no
cão do que no coelho, rato e cobaia. Formando uma pequena elevação oval na superfície
mediana do pedúnculo olfativo.
8 ALTERAÇÕES NO SISTEMA OLFATÓRIO NO CÃO E NO HOMEM
A cavidade nasal humana se encontra revestida por um epitélio ciliado não sensorial
que limpa, aquece e umidifica parcialmente o ar inspirado. Na cavidade nasal posterior, se
observa uma mucosa olfativa sensorial. Essa mucosa apresenta um neuroepitélio avascular
constituído especialmente de quatro tipos de células (células receptoras olfativas, células de
apoio, células basais e células microvíricas) e uma lâmina própria de tecido conjuntivo
subjacente, contendo feixes de nervos, glândulas de Bowman e vasos sanguíneos. Arranjos de
tecido não sensorial surgem constantemente na região na qual acreditava-se ser puramente
olfativa. O significado desses arranjos não foi encontrado, mas podem ser considerados
consequências da exposição a agentes ambientais ou mudanças que ocorrem durante o
processo normal de envelhecimento (MORRISON; COSTANZO, 1992).
Jafek et al. (1989 apud MORRISON; COSTANZO, 1992, p.59) observaram estruturas
resultantes de biópsia de pacientes anósmicos com lesão na cabeça. Os pesquisadores
verificaram que o neuroepitélio olfatório teria sofrido um bloqueio celular, visto que, o os
seus núcleos se encontravam totalmente espalhados em todo o epitélio, alguns bem próximos
à mucosa. Outro fator encontrado foi uma extinção quase que total dos cílios nos botões
olfativos, mesmo na presença dos corpos basais. Próximos à lâmina basal estavam os axônios
olfatórios dispersados por todo o epitélio formando feixes no interior da lâmina própria.
Contudo, foi possível constatar que houve a possibilidade substituição neuronal do epitélio
olfatório humano e formação de estruturas neurais. Porém, os axônios das células olfatórias
não estabeleceram um crescimento central, assim, não foi possível chegar ao bulbo olfatório
em virtude da fibrose da lâmina crivosa.
Oliveira et al. (2014) observaram áreas do tronco encefálico que estão relacionadas
com doenças neurodegenerativas, prevalentes em idosos, como Alzheimer e Parkinson. Eles
selecionaram as seguintes regiões específicas de interesse: o mesencéfalo (substância cinzenta

31
ao nível do surgimento do nervo oculomotor), Ponte (seção que inclui o segmento do lócus
cerúleos) e bulbo (ao nível do núcleo motor dorsal do vago). Além dessas regiões, ainda
avaliaram o bulbo olfatório, que foi examinado por completo e impregnado na parafina na
orientação craniocaudal. Os pesquisadores analisaram 14 cérebros de cadáveres com uma
média de 72 anos de idade. Seis desses 14 cérebros analisados apresentaram alterações
macroscópicas. Os autores encontraram a presença de atrofia frontal leve com expansão do
nervo frontal dos ventrículos laterais como o achado mais frequente (encontrado em 3 casos).
Detectaram atrofia temporal com redução volumétrica do hipocampo no cérebro número de
10. O cérebro número 3 era de um paciente com diagnóstico clínico de atrofia de sistema
múltiplo e apresentava atrofia cerebelar grave, predominantemente na substância branca,
putamen, base da ponte e pedúnculo cerebelar, bem como um pequeno clareamento da
substância cinzenta. Em 11 dos 14 casos, não foram identificadas alterações histológicas no
bulbo olfatório. No entanto, para a análise com a proteína tau fosforilada, três destes
apresentaram positividade difusa e forte ao anticorpo tau, sob a forma de neurite distrófica,
principalmente na camada plexiforme externa. Toda via os pesquisadores não encontraram
sinais clínicos de Doença neurodegenerativa precoce associação entre os depósitos de
proteínas e as alterações clínicas.
Hirai et al. (1996) estudaram as alterações do sistema olfatório do cão em relação à
idade, ele examinou 22 cães com idade entre 10 e 19 anos, sendo que 10 eram do sexo
masculino e 12 do sexo feminino, alguns foram eutanasiados e outros morreram de alguma
doença, incluindo dirofilariose, insuficiência renal e tumor nos órgãos genitais. Cães jovens e
maduros ainda foram usados como grupo controle. E amostras da mucosa olfatória, bulbo
olfatório e cérebro foram coletadas rapidamente após a necropsia. As análises foram
realizadas com imuno-histoquímica e microscopia eletrônica. Na técnica de imuno-
histoquímica utilizaram o anticorpo anti-carnosina como marcador para as células olfativas, e
evidenciaram uma redução da quantidade de células. Na microscopia eletrônica, observaram
uma diminuição no número de cílios das células olfativas e microvilos de células basais. Em
animais com idade acima de 17 anos considerados idosos, observaram atrofias com aspectos
regenerativos apareceram raramente, e que grânulos de um tipo de lipofuscina no epitélio
olfatório se tornaram volumosos com a idade. Eles ainda afirmam que alterações relacionadas
com a idade foram igualmente encontradas no epitélio olfativo de homens e ratos.

32
9 FISIOLOGIA DO OLFATO
Na camada mucosa que reveste a cavidade nasal se encontram os receptores de
proteínas ligantes de odorantes. Proteínas presentes no muco se ligam as moléculas
aromáticas que adentram a cavidade nasal, essas proteínas permitem a locomoção das
substâncias lipofílicas dissolvidas na mucosa. As células receptoras dessas proteínas são
neurônios bipolares do epitélio olfatório que se prolongam e fazem sinapses com os
interneurônios presentes no BO (MOYES; SCHULTE, 2010). Então, esses neurônios fazem
sinapses com os neurônios de segunda ordem presente nos glomérulos, em que cada
glomérulo caracteriza um ponto que capta um sinal específico de odorífero e são ativados
vários grupos de glomérulos, que convergem para as células mitrais as quais ampliam e
difundem o sinal. As células granulares que liberam GABA e inibem a ação das células
mitrais (MARIEB; HOEHN, 2009).
As proteínas receptoras de odorantes são receptores acoplados à proteína G, que
possuem características muito semelhantes à ação hormonal. A proteína G se torna ativa
quando uma molécula odorante se une a um receptor, acarretando uma alteração na
configuração do próprio receptor. Depois de ativada essa proteína aciona uma via de
transdução do sinal envolvendo adenilato-ciclase e consequentemente gera um potencial
despolarizante (MOYES; SCHULTE, 2010). Uma subunidade alfa desprende-se da proteína
G, sempre que o receptor é excitado, alfa ativa a adenilciclase. Ao ser ativada a ciclase
transforma as moléculas de trifosfato de adenosina intracelular em monofosfato cíclico de
adenosina (AMPc). O AMPc age com outra proteína de membrana próxima, permitindo a
entrada de uma grande quantidade de íons de sódio no citoplasma da célula receptora, estes
íons de sódio contribuem positivamente para o interior da membrana celular, agindo assim
sobre neurônios olfatórios que transmitem potencial de ação para dentro do SNC, através do
próprio nervo olfatório. Este mecanismo é importante porque ativa os nervos olfatórios
fazendo com que o efeito excitatório se multiplique enormemente na mais fraca partícula
odorífera (GUYTON; HALL, 1996).
Outro mecanismo é o do epitélio do órgão vomeronasal que apresenta
quimiorreceptores os quais detectam moléculas químicas ambientais, os feromônios, que são
substâncias liberadas e reconhecidas por animais da mesma espécie. Distintamente do epitélio
olfatório, os receptores dos feromônios estimulam um sistema de transdução de sinal
agregado à PLC (Phospholipase C), que hidrolisa o fosfatidilinositol-4-5-bisfofato na

33
membrana plasmática, formando trifosfato de inositol e diacilglicerol, que acarretam um
aumento do Ca2+
intracelular e abre os canais de Cl- na membrana plasmática (MOYES;
SCHULTE, 2010). Segundo Allison (1953) a maioria dos sistemas sensoriais consiste
principalmente do nervo medulado de fibras de diâmetro espesso que é comparativamente
fácil detectar degenerações em experimentos. No entanto, as fibras nervosas olfativas, são
extremamente finas e de difícil identificação histológica, e era impossível observar potenciais
elétricos com os métodos de amplificação existentes na época. Os centros olfativos estão
situados na base do telencéfalo e na maioria dos mamíferos, eles se encontram sob uma
grande massa de tecido cortical e, também não são facilmente abordados experimentalmente.
No caso da formação olfativa acessória ou órgão vomeronasal de Jacobson e suas conexões
centrais, a situação é ainda mais complicada, porque o órgão é vestigial ou ausente no
homem, e o observador é obrigado a confiar em experimentos realizados em animais
inferiores para obter evidências indiretas quanto à sua importância.
Os receptores olfatórios são neurônios bipolares, especializados na detecção de
moléculas aromáticas. Eles são encontrados no revestimento do neuroepitélio na porção
anterior da cavidade nasal. Na região apical do corpo da célula surge um dendrito único e não
ramificado que se prolonga até a superfície epitelial. Entre as células de suporte adjacentes ou
ao longo do percurso olfatório, os dendritos podem seguir diretamente ou seguir um caminho
torto, eles alcançam a região epitelial, eventualmente entrelaçados com dendritos de neurônios
adjacentes. Os processos dendríticos geralmente medem de 1-2µm de diâmetro diminuindo
ligeiramente à medida que se estendem para a superfície epitelial. Os dendritos finalizam em
intumescências perto da porção epitelial, essas intumescências são os denominados
glomérulos, os quais normalmente se estendem acima da superfície epitelial, processo
observado na figura 10 (MORRISON; COSTANZO, 1992).
O epitélio olfatório está projetado sobre a lâmpada olfativa, os axônios das células
receptoras olfativas se unem e constituem os folhetos do nervo olfativo que seguem um curso
paralelo nas camadas mais profundas da mucosa que seguem em direção ao osso etmoide
(Figura 8.). Eles passam para a fossa craniana anterior, e os feixes são entrelaçados de uma
forma mais complexa na camada mais externa da lâmpada olfativa. Nas seções histológicas da
lâmpada olfativa corados com prata, os feixes de fibras são vistos entrando em cada
glomérulo de vários ângulos. Portanto, é impossível determinar uma localização topográfica

34
da projeção do epitélio olfatório na lâmpada olfativa sem o auxílio de experimentos
(ALLISON, 1953).
Na pesquisa de Hirai et al. (1996), podemos destacar os achados do BO em cães, eles
observaram a frequente presença de amiloidose cerebrovascular nas meninges e às vezes no
parênquima tanto pela imuno-histoquímica P-amilóide quanto pela coloração com tioflavina-
S. Em quase todas as camadas da lâmpada olfativa encontrou-se gliose astrocítica. Arranjos
de ubiquitina que estão atrelados à idade foram achados principalmente na substância branca.
Todos esses apanhados estão relacionados e se tornaram protuberantes de acordo com o
tempo de vida de cada animal. Não foram encontrados placa senil, nem emaranhados
neurofibrilares no bulbo olfativo, por meio das técnicas utilizadas, até mesmo em animais
com um grande número de placas senis no córtex cerebral.
Figura 10. Representação das células sensoriais do bulbo olfativo. (adaptado de Lent, 2010).
9.1 PLASTICIDADE NEURAL
Geralmente, no sistema nervoso central, não ocorre substituição dos neurônios quando
eles morrem após uma lesão ou uma doença. Em relação a essa característica, o epitélio
olfatório, possui uma grande capacidade de reposição neuronal regularmente e após uma
lesão. Essa possibilidade existe por causa da provável capacidade de neurogênese de um
grupo de células-tronco neuroblásticas e de células basais. Essa eficácia não é encontrada em
outros neurônios do SNC vertebrado. A longevidade dos neurônios olfatórios pode ser

35
acometida por interferências ambientais. As células maduras com sinapses vivem por muito
tempo e constatou-se que podem superar até um ano de vida. A morte celular foi observada
especialmente em células receptoras jovens que não desenvolveram contato sináptico com a
lâmpada olfativa. Assim, tanto os fatores genéticos quanto os ambientais podem interferir em
ambos os processos (MORRISON; COSTANZO, 1992).
10 ASPECTOS FILOGENÉTICOS
O sistema olfatório é constante em todos os vertebrados por esse motivo, esse sistema
se torna de muita importância para os neurologistas comparativos. Os animais pertencentes ao
Filo Vertebrata apresentam bulbo olfatório e centros similares em estrutura e compõem uma
porção de um cérebro relativamente inalterado, mas que sofreu uma série de
desenvolvimentos complicados. Tais aspectos consideraram que o sentido do olfato é
extremamente primitivo, e que foi determinado no princípio da evolução dos vertebrados,
após a formação das áreas olfativas o prosencélafo teve seu total desenvolvimento. No auge
da neurologia comparativa (entre 1890-1920), considerou-se que para se entender a evolução
do prosencéfalo seria necessário investigar o sistema olfatório (ALLISON, 1953).
“As vias filogeneticamente mais elevadas do Sistema Olfatório são complexas e
sujeitas às influências modificadoras, provenientes de várias fontes. Algumas destas
vias, que envolvem sistemas multissinápticos são submetidas a outros tipos de
influências e perderam completamente sua especificidade olfatória original”
(CARPENTER, 1978).
O epitélio olfatório humano pode ter aspectos análogos a outras espécies de
vertebrados. No entanto, em alguns casos, o arranjo do seu epitélio sensorial não é
homogêneo, pois possui pontos do epitélio não sensorial constantemente percebido nas áreas
olfatórias. Ainda não foi constatado o motivo dessas áreas irregulares, mas existe a hipótese
que podem ser consequências de agentes ambientais, doença ou o resultado do processo
natural de envelhecimento dos seres humanos. Toda via, ainda existem estudos que
pressupõem a capacidade de reposição dos neurônios olfatórios no ser humano, em que um
prolongamento da reativação funcional varia de acordo com o grau da injúria e a
predisposição da regeneração axonal para atingir o bulbo olfatório (MORRISON;
COSTANZO, 1992).

36
11 DISCUSSÃO
Como pode ser visto ao longo deste trabalho pautado nas questões morfofuncionais do
bulbo olfatório do Cão e do Homem e, mais especificamente observações de pesquisas e
trabalho sobre esse órgão do sentido, descrito por vários autores, torna-se possível discutir
estudos comparativos do ponto de vista evolutivo, tanto estrutural quanto funcionalmente,
focando essas duas espécies de mamíferos, cada uma com suas especificidades e estruturas
orgânicas e funcionais adaptadas.
Muitos têm sido os pesquisadores e estudiosos do sentido do olfato, dado sua
importância para os Vertebrados, especialmente os mamíferos, filogeneticamente
considerados mais evoluídos, inclusive o ser humano, que do olfato dependem para manter o
equilíbrio com o ambiente e ter suas funções orgânicas adaptadas, garantindo assim
sobrevivência, reprodução e alimentação, para citar apenas as mais importantes.
Revendo alguns dos mais importantes trabalhos e pesquisas sobre o tema, um dos
artigos a chamar a atenção da comunidade científica, mostra que é notória a importância da
revisão, ora em discussão, uma vez que nos traz dados iniciais sobre aspectos evolutivos, ao
longo do tempo, como oportunamente observou Read (1908) diz que “a região olfativa tem
sido objeto de investigação especial por muitos anos. Foram publicadas várias opiniões sobre
os fins dos nervos olfativos. Alguns pontos de vista foram refutados e, ele ainda ressalta que
em 1856, Max Schultze estabeleceu com uma certeza considerável as condições verdadeiras
das terminações dos nervos olfativos na mucosa nasal. Uma revisão da literatura dará o ponto
de vista atual”. Pautado no trabalho de revisão para o momento atual, teremos uma cronologia
evolutiva importante para rebuscar uma pesquisa comparativa sobre a evolução do bulbo
olfatório e o sentido da olfação, o que permite observar uma desenvoltura comportamental no
mundo animal e humano, como Jia et al. (2014) afirmam que humanos dependem das
habilidades olfativas de cães para difíceis tarefas, como a detecção de bombas, minas
terrestres, produtos químicos perigosos e substâncias ilícitas. Portanto, uma compreensão
mecânica do sistema olfativo em cães é de grande interesse científico.
Retomando um dos trabalhos mais distantes dos de hoje, observamos que pouca coisa
mudou no que se refere aos aspectos anátomofuncionais e, de certa forma, fisiológicos, muito
embora a terminologia anatômica tivesse que ser adaptada, através do tempo, por questões de
nomina anatomica, como podemos ver que McCotter (1912) afirma que no cão, a grande

37
lâmpada olfativa possui um formato de quadrilátero e que ela é comprimida pela região
frontal do cérebro de um lado e a estrutura do nasal do outro. A superfície dorsocaudal é
côncava e moldada no limite anterior do lobo frontal. O pedúnculo olfativo se estende para a
região posterior a partir da parte ventrolateral desta superfície. A região anteroventral situa-se
contra a lâmina crivosa de maneira oblíqua. Ela recebe os nervos olfatórios que se espalham
em uma rede espessa sobre a superfície da lâmpada olfativa. A margem mediana espessada da
lâmpada olfativa é separada do bulbo oposto pela extensão dorsal da placa perpendicular. A
lâmpada olfativa acessória é menor no cão do que no coelho, rato e cobaia. Formando uma
pequena elevação oval na superfície mediana do pedúnculo olfativo.
Enquanto Kavoi & Jameela (2011) realizaram uma análise morfométrica do bulbo,
trato e estria olfativos em animais ecologicamente diversos, com o intuito de apontar
diferenças morfométricas relacionadas à função olfativa. Foram utilizados cérebros inteiros de
humanos e de cães, cinco de cada espécie, fixados por perfusão com 10% de formalina. Os
pesquisadores correlacionaram os volumes e as medidas sequenciais das estruturas olfativas
com os do cérebro e do cérebro inteiro, e observaram que o volume da lâmpada olfativa foi
maior em cães que em seres humanos constituindo 0,31% e 0,01%, respectivamente, do
volume cerebral. Do mesmo modo, a dimensão do volume do bulbo, trato e estria para o
cérebro foi de 1,95% no cão e 0,03% no ser humano. A largura da lâmpada foi maior (p
<0,05) em cães (10,80 ± 1,64 mm) comparada com a do homem (5,50 ± 0,71 mm), e
apresentaram uma amplitude do hemisfério de 42,91% e 8,94%, respectivamente. No entanto,
o comprimento total do bulbo, trato e estrias olfativas aumentou na ordem humano (36,25 ±
1,70 mm) e cão (48,20 ± 1,92 mm) e constituiu 51,87% e 72,30%, respectivamente, do
comprimento do hemisfério. Esses resultados sugerem que as adaptações morfométricas dos
componentes olfativos à função olfativa, podem indicar a variação das necessidades
funcionais olfativas em relação à diversidade ecológica das espécies. Nesse estudo
compararam os cérebros de humanos, cães e cabras. No entanto, descartamos os dados
obtidos para cabras.
Para compreender os substratos neurais de nível cognitivo Jia et al. (2014) usaram a
ressonância magnética para investigar como o comportamento é conduzido por representações
sensoriais filtradas em áreas cognitivas de ordem superior. Para isso, eles construíram um
sistema de entrega de odorantes controlado por um computador adaptado o qual teria sido
sincronizado com a captação de imagens, possibilitando a análise das áreas cerebrais ativadas

38
pelas substâncias aromáticas. Assim, os pesquisadores perceberam que o bulbo olfatório e os
lobos piriformes foram normalmente acionados em cães anestesiados e acordados, enquanto o
córtex frontal foi ativado basicamente em cães conscientes.
Assim sendo, este trabalho traz uma importante observação do ponto de vista
evolutivo, porém independente da sequência e disposição dos vertebrados na escala zoológica,
até então aceita cientificamente, mas que esta evolução não obedece igual padrão de
desenvolvimento no que diz respeito às suas adaptações, estruturas orgânicas seus aspectos
morfofuncionais. Sabemos que a sequência disposta, nos verterbrados filogenéticamente
obedece a seguinte ordem: peixes, anfíbios, répteis, aves e mamíferos (CHAGAS, 1987).
Com essa constatação, observamos que o ser humano, apesar de toda a evolução neural e seu
desenvolvimento mental, em relação ao olfato, não é o ser mais evoluído dentre os
vertebrados, pois a evolução implica também em adaptações e funções altamente complexas e
desenvolvidas de acordo com seu papel e importância no meio ambiente. Para corroborar esta
afirmativa, é suficiente exemplificar que nós humanos vemos menos que as aves de rapina e
certos animais noturnos. Também ouvimos menos do que os morcegos. E temos olfato menos
desenvolvido do que os cães e, inclusive os mamíferos de um modo geral, têm olfato menos
desenvolvido do que os tubarões, que são peixes condricthyies e se acham no início da escala
filogenética.

39
12 CONCLUSÃO
Baseado nos conteúdos bibliográficos, em especial os de revisão discutidos nesse
trabalho, confirmou-se que, no caso do cão e do homem, o tamanho do bulbo olfatório
corresponde à eficiência de seu olfato, ou seja, quanto maior o bulbo, maior a capacidade
olfatória, entre estes mamíferos. Em consequência disto os cães apresentam uma maior
mucosa olfatória do que os humanos e, portanto uma maior quantidade de receptores que se
projetam para o bulbo olfatório, através do qual as informações seguem para o sistema
nervoso central.
É importante também observar a influência direta dos agentes biológicos, físicos ou
químicos na condição normal ou patológica do nervo olfatório e/ou de seu bulbo, fazendo
com que o sentido da olfação tenha ou não desempenho apropriado na capacidade olfativa
desses indivíduos. Assim sendo, células do trato olfatório são as únicas que se renovam no
sistema nervoso, fazendo com que a capacidade olfatória permaneça ativa por mais tempo em
comparação com outras estruturas neurais, as quais não renovam nem regeneram seus
neurônios.
Em relação à qualidade do sentido olfativo há de se considerar também questões
referentes às posições anatômicas das duas espécies de mamíferos, as quais têm importante
influência do que diz respeito a um melhor desenvolvimento do sentido do olfato. Enquanto o
ser humano, o qual é bípede e, portanto, sua posição ortostática verticalizada, lhe permite
apenas captação do cheiro e do odor melhor absorvido em correntes de ar à altura do seu
sistema olfatório, o cão por sua vez, além de possuir um bulbo olfatório dezenas de vezes
maior do que o do ser humano, tem sua posição quadrúpede, cuja posição encefálica dispõe-se
mais horizontalmente no sentido antero superior, de modo que as correntes de ar, através das
quais permeiam o olfato, chegam com mais liberdade e maior amplitude à área rinencefálica,
conferindo melhor captação aerífera de cheiro ou odor.

40
REFERÊNCIAS
ALLISON, A. C. The morphology of the olfactory system in the vertebrates. Biological
Reviews, v.28, n.2, p. 195-244, May. 1953. Disponível
em:<http://onlinelibrary.wiley.com/doi/10.1111/j.1469-185X.1953.tb01376.x/abstract>.
Acesso em: 02.ago.2017.
ALVITES, R. D. Sistema Olfativo do Cão: Estabelecimento de uma Metodologia de
Abordagem à Lâmina Crivosa do Cão. 2015. 96 f. Dissertação (Mestrado Integrado em
Medicina Veterinária) – Escola de Medicina Veterinária, Universidade de Trás-os-Montes e
Alto Douro, Vila Real, 2015. Disponível
em:<https://repositorio.utad.pt/bitstream/10348/6060/1/msc_rdalvites.pdf >. Acesso em:
25. Jul. 2017.
BLOG DA MONITORIA DE ANATOMIA TOPOGRÁFICA DO CURSO DE
VETERINÁRIA DA UNIVERSIDADE DE UBERABA. Rinencéfalo do cão. Anatomia
Topográfica dos Animais Domésticos. Disponível
em:<http://anatomiadosanimais.blogspot.com.br/2009/09/sistema-nervoso-central-parte-
ii.html>. Acesso em: 01.Nov.2017.
CARPENTER, M. B. Neuroanatomia Humana. 7.ed. Rio de Janeiro: Interamericana, 1978.
CHAGAS, J. S. Tópicos de anatomia Comparativa. Natal: Livraria Universitária, 1987.
CHRISMAN, C. L. Neurologia dos Pequenos Animais. São Paulo: Roca, 1985.
EVANS, H. E. Miller’s anatomy of the dog. Philadelphia: WB Saunders, 1993.
GETTY, R. Anatomia dos Animais Domésticos. 5.ed. Rio de Janeiro: Guanabara Koogan,
1986. v.2.
GOLGI, Sulla fina Struttura dei bulbi olfattorii, Reggio Emilia.1875 apud READ, E. A. A
Contribution to the knowledge of the olfactory apparatus in dog, cat and man. Developmental
Dynanmics, v.8, n.1, p.17-47. 1908. Disponível em:<
http://onlinelibrary.wiley.com/doi/10.1002/aja.1000080103/full>. Acesso em: 02.Ago.2017.
GRAY, H. Anatomia Charles May Goss. 29.ed. Rio de Janeiro: Guanabara Koogan, 1977.
GUYTON, A. C.; HALL, J. E. Tratado de Fisiologia Médica. 9.ed. Rio de Janeiro:
Guanabara Koogan, 1997.
HIRAI, T. et al. Age-related changes in the olfactory system of dogs. Neuropathology and
Applied Neurobiology, v.22, n.6, p.531-539. Dec.1996. disponível
em:<http://onlinelibrary.wiley.com/doi/10.1111/j.1365-2990.1996.tb01132.x/full>. Acesso
em: 02.ago.2017.
JAFEK, B. W. et al. Posttraumatic anosmia. Arch. Neurol. 1989, 46: p.300-304. Apud in
MORRISON, E. E.; COSTANZO, R. M. Morphology of Olfactory Epithelium in Humans
and Other Vertebrates. Microscopy Research & Technique, v.23, n.1, p.49-61, Oct.1992.
Disponível em:<http://onlinelibrary.wiley.com/doi/10.1002/jemt.1070230105/abstract>.
Acesso em: 02.ago.2017.

41
JIA, H. et al. Functional MRI of the Olfactory System in Conscious Dogs. Plos one, v.9, n.1,
p.1-21 , Jan. 2014. Disponível
em:<http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0086362>. Acesso em:
31.Jul.2017.
KAVOI, B. M.; JAMEELA, H. Comparative morphometry of the olfactory bulb, tract and
stria in the human, dog and goat. Int. J. Morphol. Nairobi, 29(3):p.939-946, 2011.
Disponível
em:<https://www.researchgate.net/publication/262585195_Morfometria_Comparada_del_Bul
bo_Tracto_y_Estria_Olfatoria_en_el_Humano_Perro_y_Cabra>. Acesso em: 31.Jul.2017.
LENT, R. Cem Bilhões de Neurônios?: conceitos fundamentais de neurociência. 2. ed.
São Paulo: Atheneu, 2010.
LOURENÇO, F. D.; FURLAN, M. M. D. P. Sensibilidade olfatória em homens e cães: um
estudo comparativo. Arq Mudi., Maringá, v.11, n.2, p.14-9. 2007. Disponível
em:<http://periodicos.uem.br/ojs/index.php/ArqMudi/article/viewFile/19997/pdf>. Acesso
em: 25.Jul.2017.
LUNDY-EKMAN, L. Neurociência: Fundamentos para a Reabilitação. Rio de Janeiro:
Guanabara Koogan, 2000.
MACHADO, A. Neuroanatomia Funcional. 2. ed. São Paulo: Atheneu, 1998.
MARIEB, E. N.; HOEHN, K. Anatomia e Fisiologia. 3. ed. Porto Alegre: Artmed, 2009.
McCOTTER, R. E. The connection of the vomeronasal nerves with the accessory olfactory
bulb in the opossum and other mammals. The Anatomical Record, v.6, n.8, p.299-318, Aug.
1912. Disponível em:<http://onlinelibrary.wiley.com/doi/10.1002/ar.1090060802/abstract>.
Acesso em: 02.ago.2017.
MORRISON, E. E.; COSTANZO, R. M. Morphology of Olfactory Epithelium in Humans
and Other Vertebrates. Microscopy Research & Technique, v.23, n.1, p.49-61, Oct.1992.
Disponível em:<http://onlinelibrary.wiley.com/doi/10.1002/jemt.1070230105/abstract>.
Acesso em: 02.ago.2017.
MOYES, C. D.; SCHULTE, P. M. Princípios de Fisiologia Animal. 2. ed. Porto Alegre:
Artmed, 2010.
O’RAHILLY, R. Anatomia Humana Básica: Um estudo regional da estrutura humana.
Rio de Janeiro: Interamericana, 1985.
OLIVEIRA, F. H. et al. Neurodegenerative changes in the brainstem and olfactory bulb in
people older than 50 years old: a descriptive study. Arq. Neuro Psiquiatr., São Paulo,
vol.73, no.7, p. Jul. 2015. Disponível
em:<http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0004-282X2015000700569>.
Acesso em: 31. Jul.2017.
Osso etmóide do cão. Wikipédia enciclopédia livre. Disponível em:
<https://pt.wikipedia.org/wiki/Osso_etmoide> acesso em: 18.Ago.2017.

42
PALHETA NETO, F. X. et al. Anormalidades sensoriais: olfato e paladar. Arq. Int.
Otorrinolaringol. / Intl. Arch. Otorhinolaryngol. , São Paulo - Brasil, v.15, n.3, p. 350-358,
Jul/Ago/Set. 2011. Disponível em:
<http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1809-48722011000300014>
acesso em: 10 ago. 2017.
PRADA, I. Neuroanatomia Funcional em Medicina Veterinária com correlações clínicas.
1. ed. Jaboticabal: Terra Molhada, 2014.
READ, E. A. A Contribution to the knowledge of the olfactory apparatus in dog, cat and man.
Developmental Dynanmics, v.8, n.1, p.17-47. 1908. Disponível em:<
http://onlinelibrary.wiley.com/doi/10.1002/aja.1000080103/full>. Acesso em: 02.Ago.2017.
SALAZAR, I.; CIFUENTES, J. M.; SÁNCHEZ-QUINTEIRO, P. Morphological and
Immunohistochemical Features of the Vomeronasal System in Dogs. The Anatomical
Record, Santiago de Compostela, v.296, p.146–155, Sept. 2013. Disponível
em:<http://onlinelibrary.wiley.com/doi/10.1002/ar.22617/pdf >. Acesso em: 18.Ago.2017.
SOBOTTA, J.; BECHER, H. Atlas de Anatomia Humana. 17.ed. Rio de Janeiro: Guanabara
Koogan, 1977.
Recommended

















