Varijacije u broju
kopija u genomu
čoveka

Koje su razlike između individua ?
Koje su razlike između vrsta ?
Projekat “Genom čoveka”
Genomika
Komparativna genomika
Komparativna
genomika
• Sajt http:// https://gold.jgi.doe.gov/
• 11. januar 2017. godine - sekvencirano 10.625 (kompletno)
+ 72.007 genoma (draft) + 45.678 započetih projekata
• 28. februar 2018. godine - sekvencirano 12.832 (kompletno)
+ 107.124 genoma (draft) + 66.370 započetih projekata
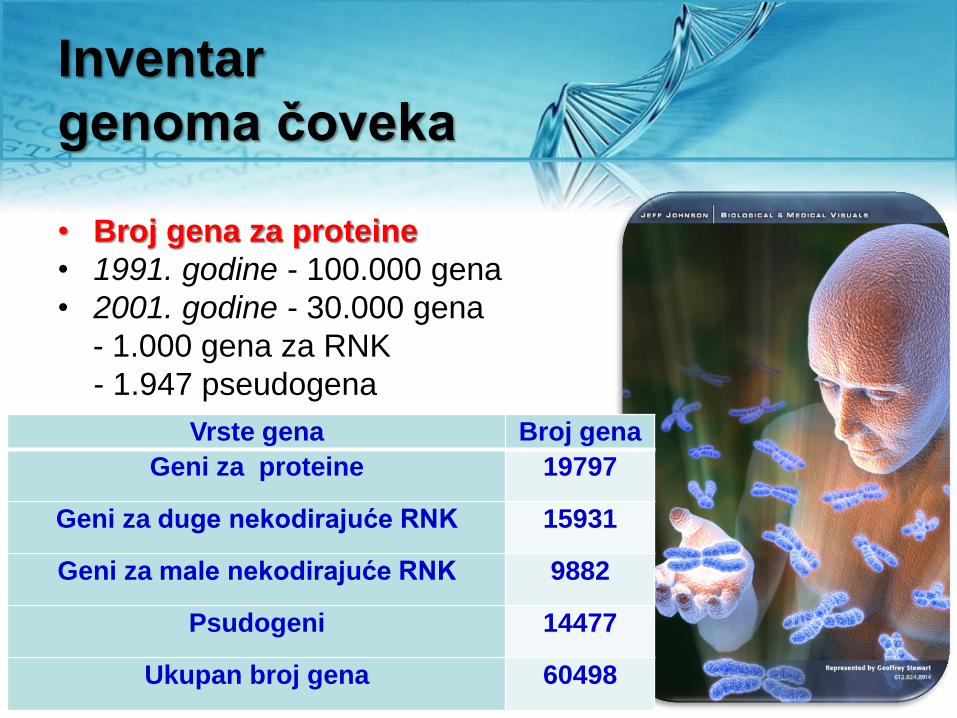
Inventar
genoma čoveka
• Broj gena za proteine
• 1991. godine - 100.000 gena
• 2001. godine - 30.000 gena
- 1.000 gena za RNK
- 1.947 pseudogena
Vrste gena Broj gena
Geni za proteine 19797
Geni za duge nekodirajuće RNK 15931
Geni za male nekodirajuće RNK 9882
Psudogeni 14477
Ukupan broj gena 60498

The Encyclopedia of DNA Elements
(ENCODE)
• Cilj projekta – registrovati sve funkcionalne elemente genoma čoveka:
• Segmenti genoma koji kodiraju za definisani produkt (RNK i/ili protein)
• Elementi genoma koji pokazuju “reproducible biochemical signature" (sekvence za vezivanje proteina ili sekvence odgovorne za specifične strukture hromatina)

ENCODE
funkcije genoma čoveka
• 399.124 regiona koji po svojim karakteristikama
odgovaraju pojačivačima i 70.292 regiona koji po svojim
karakterisitkama odgovaraju promotorima.
• Procena da se dve trećine genoma čoveka prepisuje.

Projekat “Genom čoveka“
(Human Genome Project) Preliminarna procena - Homo sapiens 99.9% identični

Razlike u genomima ljudi
• Prvi eksperimentalni dokazi o postojanju značajnih
razlika u genomima (zdravih) individua dobijeni su
zahvaljujući razvoju novih tehnika…..
• Iafrate et al, 2004
• Sebat et al., 2004

Metodologija istraživanja
odnosno detekcije CNV
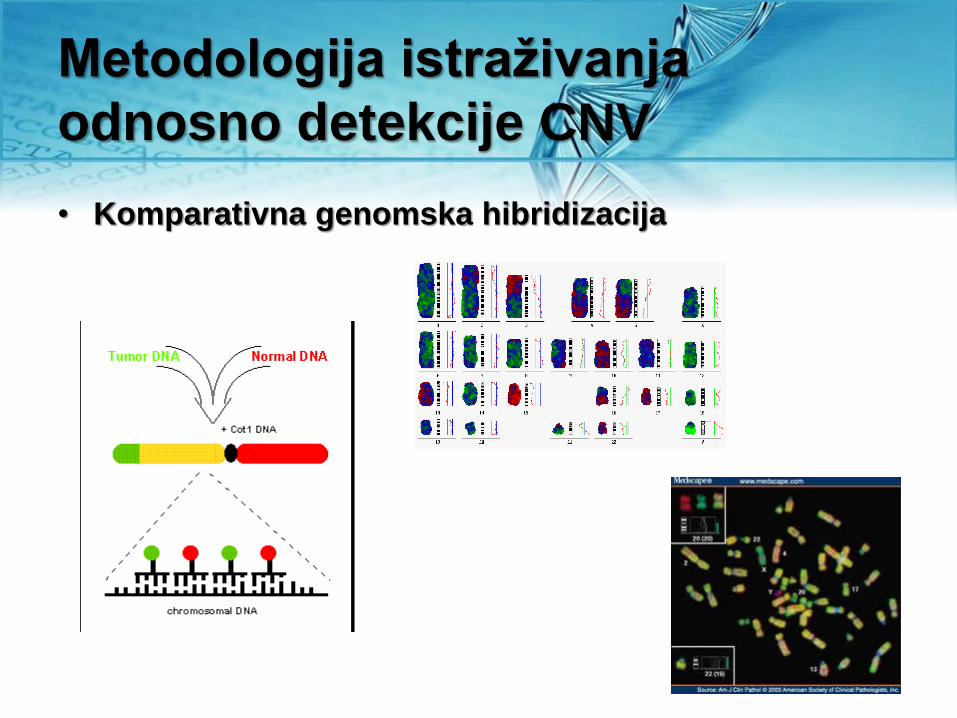
Metodologija istraživanja
odnosno detekcije CNV
• Komparativna genomska hibridizacija

Genski čip
• Na osnovu količine probe koja hibridizuje za svaku tačku
(“spot”) na array-u dobija se informacija o specifičnom
sastavu nk u ispitivanom uzorku.
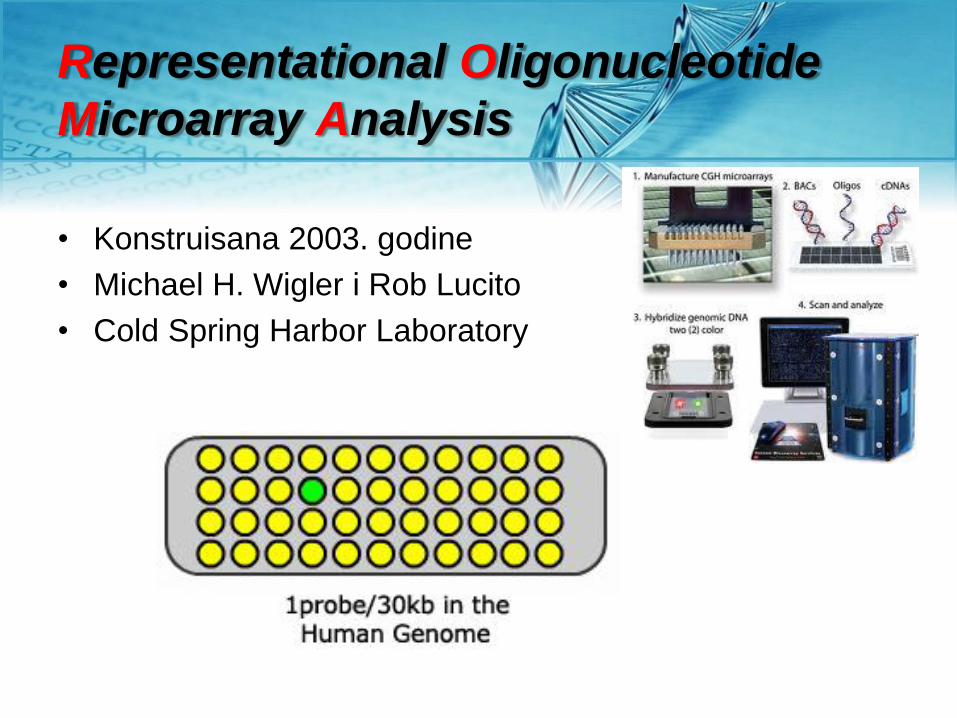
Representational Oligonucleotide
Microarray Analysis
• Konstruisana 2003. godine
• Michael H. Wigler i Rob Lucito
• Cold Spring Harbor Laboratory
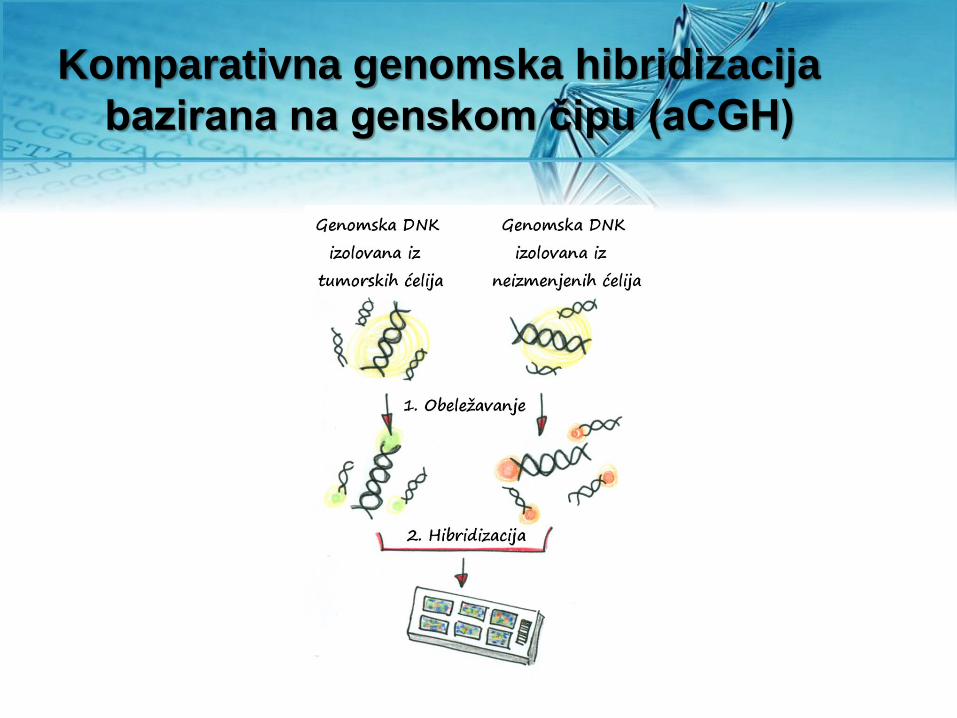
Komparativna genomska hibridizacija
bazirana na genskom čipu (aCGH)

aCGH
• Metodološke kritike
- Šta je referentni genom
- Kako je “istačkan” genski čip
- Analiza dobijenih rezultata
- Tehnika se definiše kao “prljava”

Metode dubinskog sekvenciranja
• Danas najmanje 12 platformi za analizu CNV
Provera rezultata
• FISH
• qPCR
• Metoda multipleks
amplifikacije
ligiranih proba (MLPA)
• Sekvenciranje po Sangeru

MLPA
• Jedan oligonukleotid svake
MLPA probe na 3’ kraju sadrži
sekvencu komplementarnu
specifičnoj ciljnoj sekvenci,
dok na 5’ kraju sadrži sekvencu
koja je zajednička za sve MLPA
probe i identična je
fluorescentno obeleženom
PCR prajmeru.

MLPA
• Drugi oligonukleotid svake MLPA
probe na 5’ kraju poseduje
sekvencu komplementarnu
specifičnoj ciljnoj sekvenci, na 3’
kraju sadrži sekvencu zajedničku za
sve MLPA probe dužine koja je
komplementarna neobeleženom
prajmeru.
• Između ova dva kraja nalazi se
insert ili “stuffer” sekvenca različite
dužine.

MLPA
• Ligacijom oligonukleotida MLPA probe formiraju se
uspešno ligirane probe koje imaju identične sekvence na
5’ i 3’ krajevima.
• Zahvaljujući ovoj njihovoj osobini omogućena je
istovremena amplifikacija i do 50 različitih MLPA proba
korišćenjem samo jednog para prajmera.
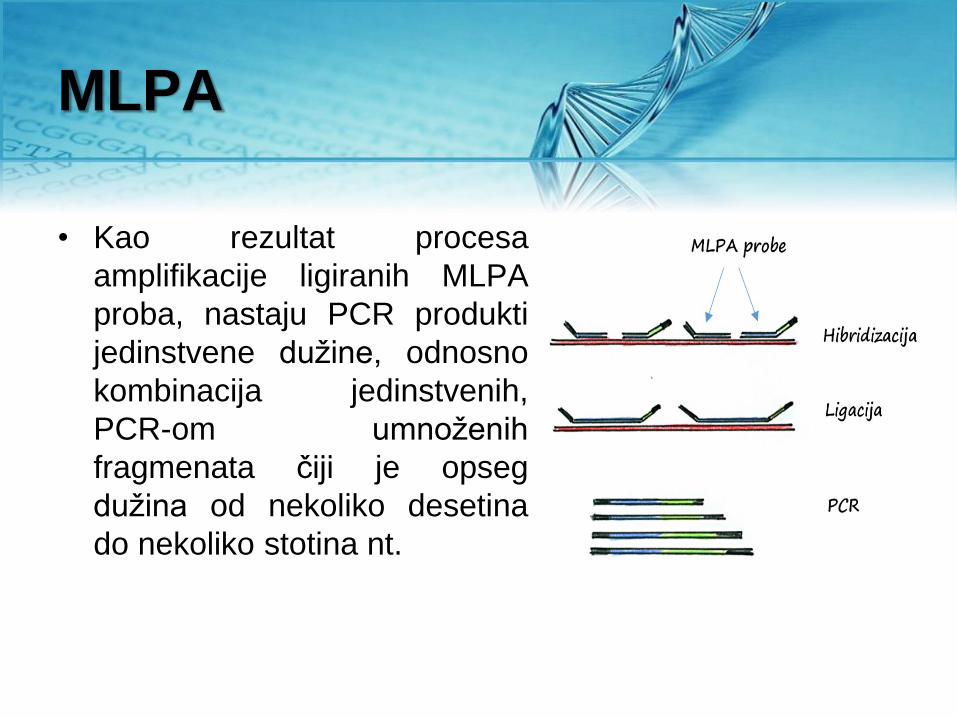
MLPA
• Kao rezultat procesa
amplifikacije ligiranih MLPA
proba, nastaju PCR produkti
jedinstvene dužine, odnosno
kombinacija jedinstvenih,
PCR-om umnoženih
fragmenata čiji je opseg
dužina od nekoliko desetina
do nekoliko stotina nt.
MLPA
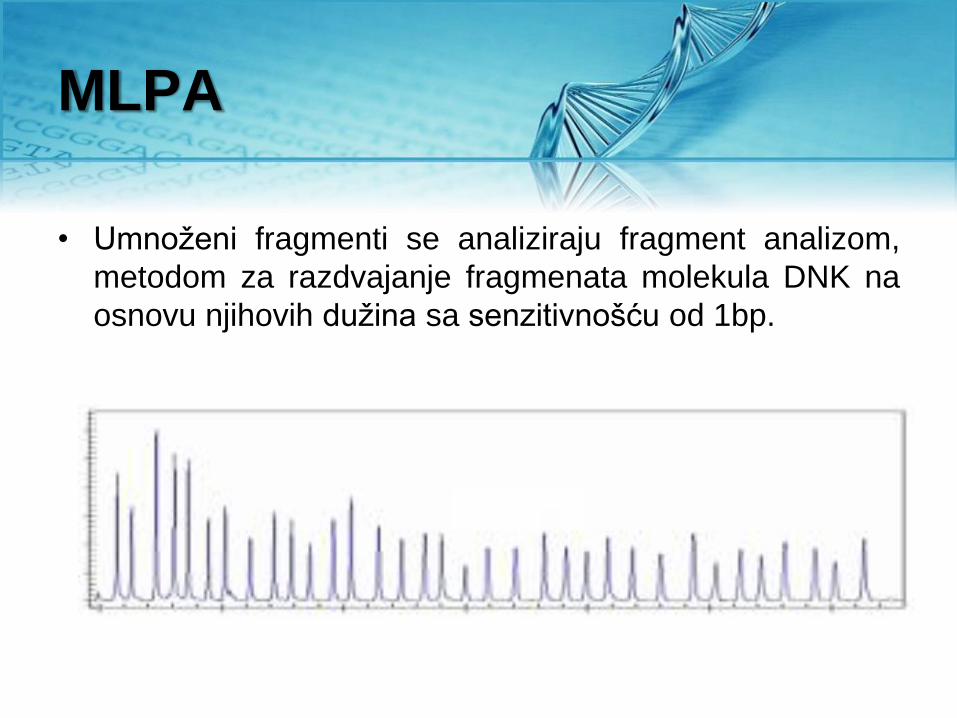
• Umnoženi fragmenti se analiziraju fragment analizom,
metodom za razdvajanje fragmenata molekula DNK na
osnovu njihovih dužina sa senzitivnošću od 1bp.
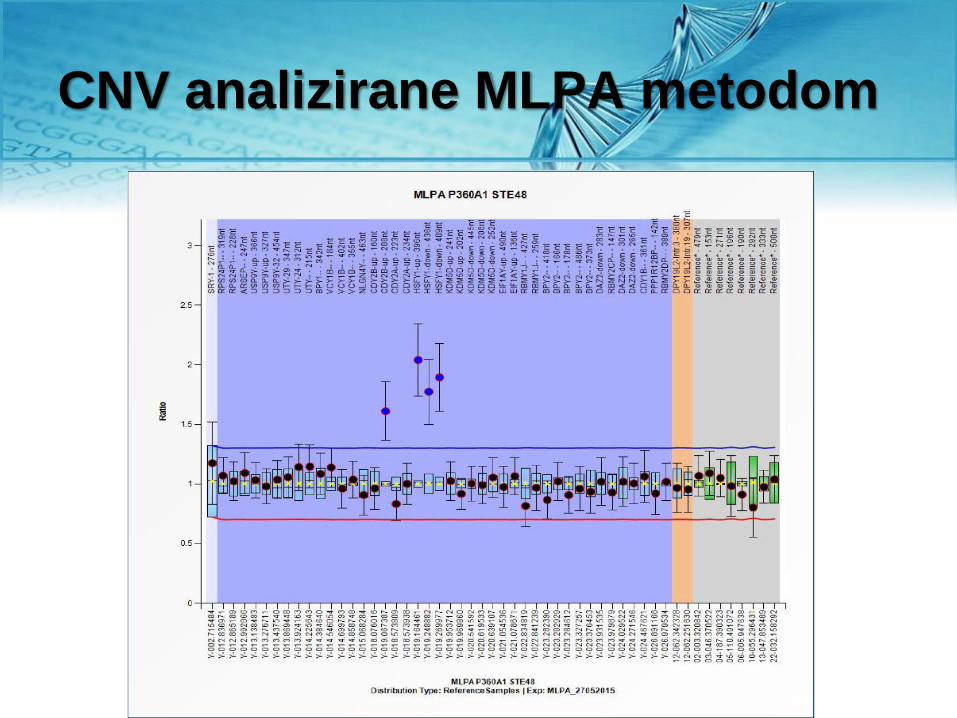
CNV analizirane MLPA metodom
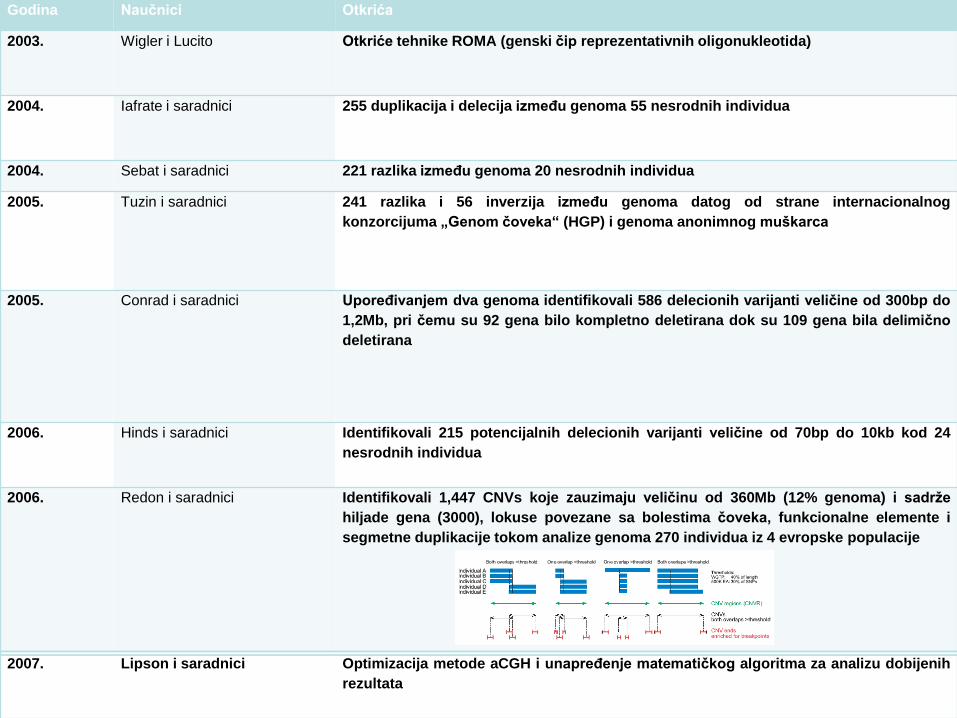
Godina Naučnici Otkrića
2003. Wigler i Lucito Otkriće tehnike ROMA (genski čip reprezentativnih oligonukleotida)
2004. Iafrate i saradnici 255 duplikacija i delecija između genoma 55 nesrodnih individua
2004. Sebat i saradnici 221 razlika između genoma 20 nesrodnih individua
2005. Tuzin i saradnici 241 razlika i 56 inverzija između genoma datog od strane internacionalnog
konzorcijuma „Genom čoveka“ (HGP) i genoma anonimnog muškarca
2005. Conrad i saradnici Upoređivanjem dva genoma identifikovali 586 delecionih varijanti veličine od 300bp do
1,2Mb, pri čemu su 92 gena bilo kompletno deletirana dok su 109 gena bila delimično
deletirana
2006. Hinds i saradnici Identifikovali 215 potencijalnih delecionih varijanti veličine od 70bp do 10kb kod 24
nesrodnih individua
2006. Redon i saradnici Identifikovali 1,447 CNVs koje zauzimaju veličinu od 360Mb (12% genoma) i sadrže
hiljade gena (3000), lokuse povezane sa bolestima čoveka, funkcionalne elemente i
segmetne duplikacije tokom analize genoma 270 individua iz 4 evropske populacije
2007. Lipson i saradnici Optimizacija metode aCGH i unapređenje matematičkog algoritma za analizu dobijenih
rezultata

Varijacije u broju kopija (eng. Copy number variations, CNVs)
• Termin CNV objedinjuje i pojmove:
• LSV – large–scale CNV (Iafrate et al.,2004)
• CNP – copy number polymorphism (Sebat et
al, 2004)
• ISV – intermediate – sized variants (Tuzun et
al., 2005)

Polimorfizmi DNK lokusa
• Razlike u sekvenci DNK koje postoje u određenom DNK
lokusu između individua jedne vrste označene su kao
polimorfizmi DNK.
• Postoje dva tipa DNK polimorfizama:
1. Polimorfizmi nukleotidne sekvence, koji su vezani za
- varijabilnost tipa bazne zamene (tačkasti polimorfizmi)
- za inserciju ili deleciju određenog niza nukleotida
(inserciono-delecioni prolimorfizmi)
2. Polimorfizmi dužina sekvence, koji su vezani za različiti
broj ponovaka u mini- i mikro-satelitskim lokusima.
Varijacije u broju kopija - CNVs
• Termin polimorfizam zamenjen
terminom varijanta jer se
polimorfizam definiše kao genetička
varijanta sa učestalošću ređeg alela
≥ 1%.
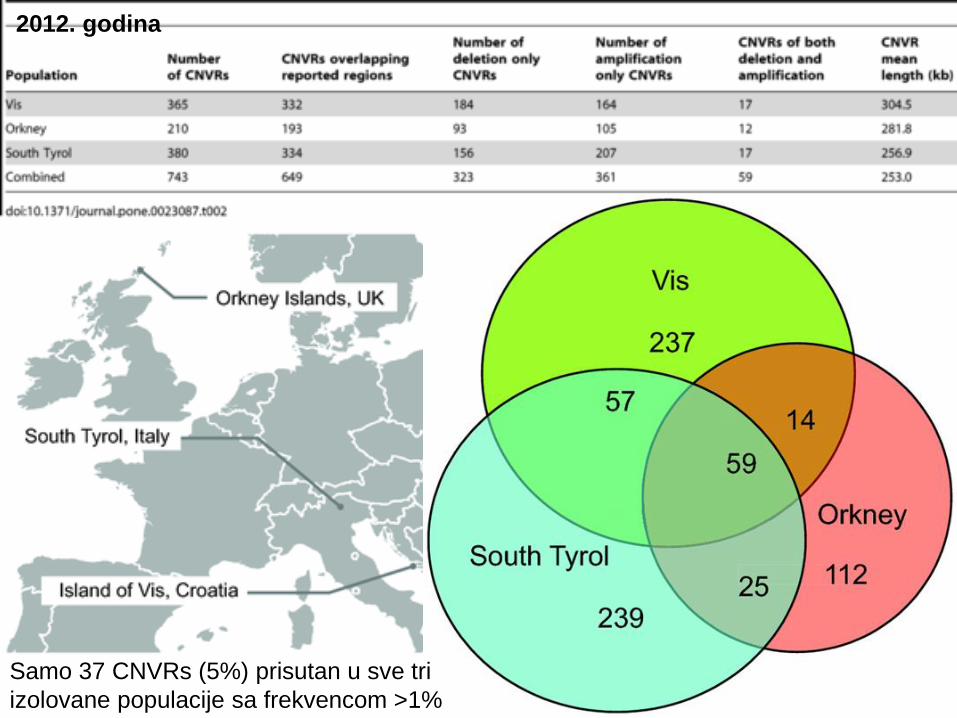
Samo 37 CNVRs (5%) prisutan u sve tri
izolovane populacije sa frekvencom >1%
2012. godina

Novo pitanje
• Genom čoveka ili genomi ljudi

Genomi
ljudi
Jedna vrsta jedan genom
Genom čoveka NE genomi ljudi
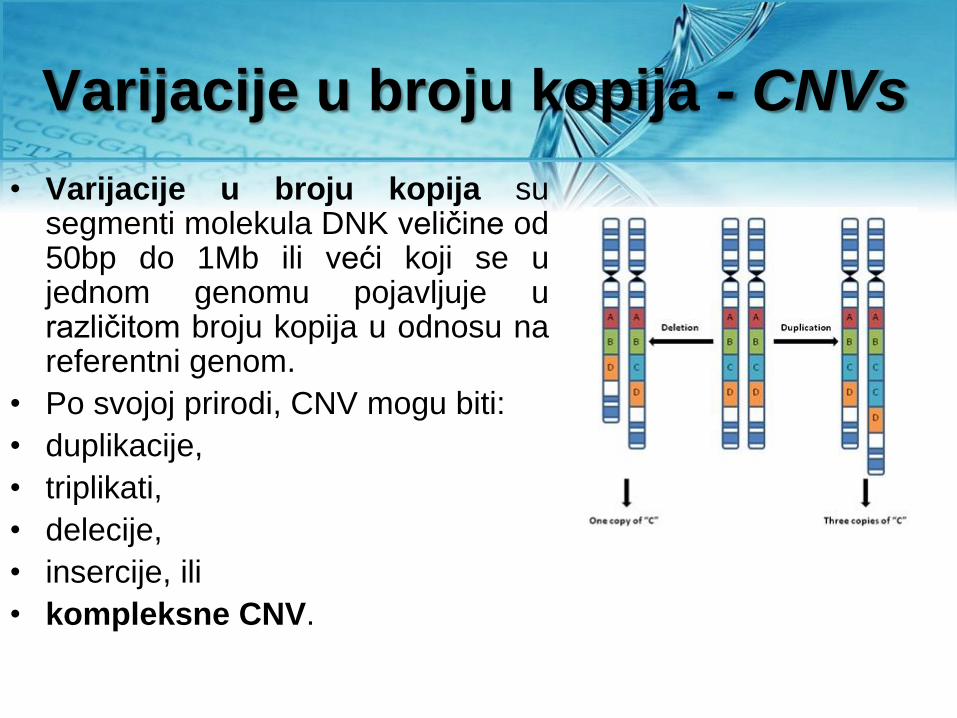
Varijacije u broju kopija - CNVs
• Varijacije u broju kopija su segmenti molekula DNK veličine od 50bp do 1Mb ili veći koji se u jednom genomu pojavljuje u različitom broju kopija u odnosu na referentni genom.
• Po svojoj prirodi, CNV mogu biti:
• duplikacije,
• triplikati,
• delecije,
• insercije, ili
• kompleksne CNV.
Varijacije u broju kopija - CNVs
• STRUKTURNE VARIJANTE GENOMA
• CNV su kvantitativne strukturne varijante genoma.
• Pored kvantitativnih, razlikuju se i pozicione strukturne varijante (translokacije) i orijentacione strukturne varijante (inverzije).
Varijacije u broju kopija - CNVs
• CNV treba razlikovati od mikro-delecija i mikro-duplikacija hromozoma koje su vidljive tehnikom bojenja traka i koje se, kao takve, ubrajaju u hromozomske aberacije.
• CNV po tipu duplikacija treba razlikovati od mikro-satelita i mini-satelita kao i triplikate od trinukleotidnih ponovaka.

CNV po tipu duplikacija
• Tandemske duplikacije podrazumevaju duplikacije
određenog segmenta molekula DNK koji se u svim
analiziranim genomima dupliciraju na isti način.
• Multialelske duplikacije podrazumevaju duplikacije čiji
se broj dupliciranih segmenata molekula DNK razlikuje
između analiziranih genoma.
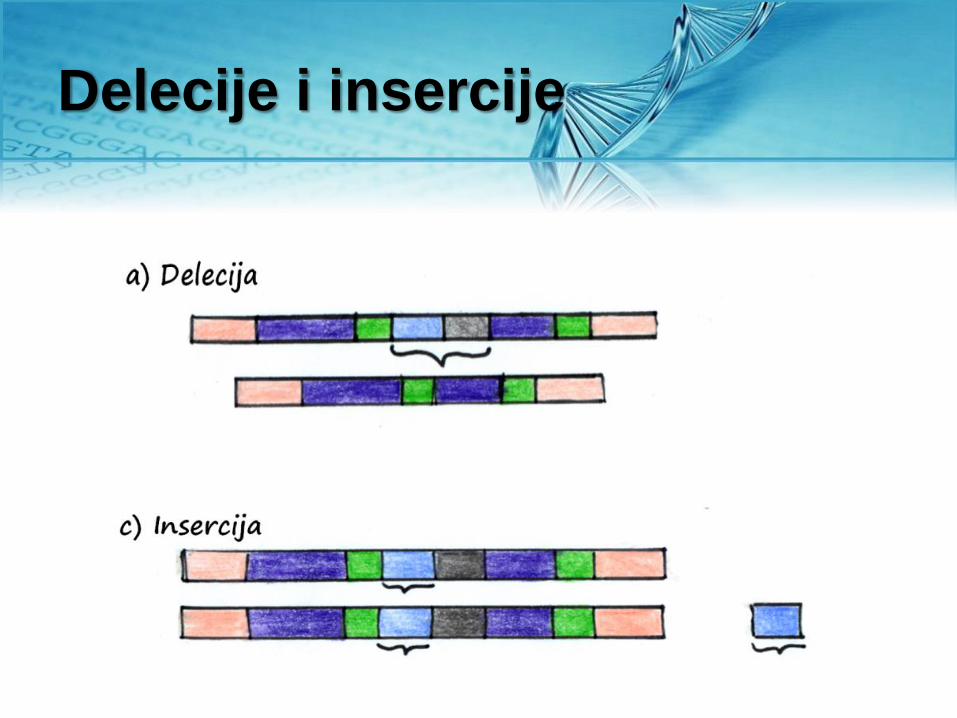
Delecije i insercije

Kompleksne CNV
• Kompleksna CNV podrazumeva segment molekula
DNK koji u jednom genomu može biti dupliciran a u
drugom deletiran.
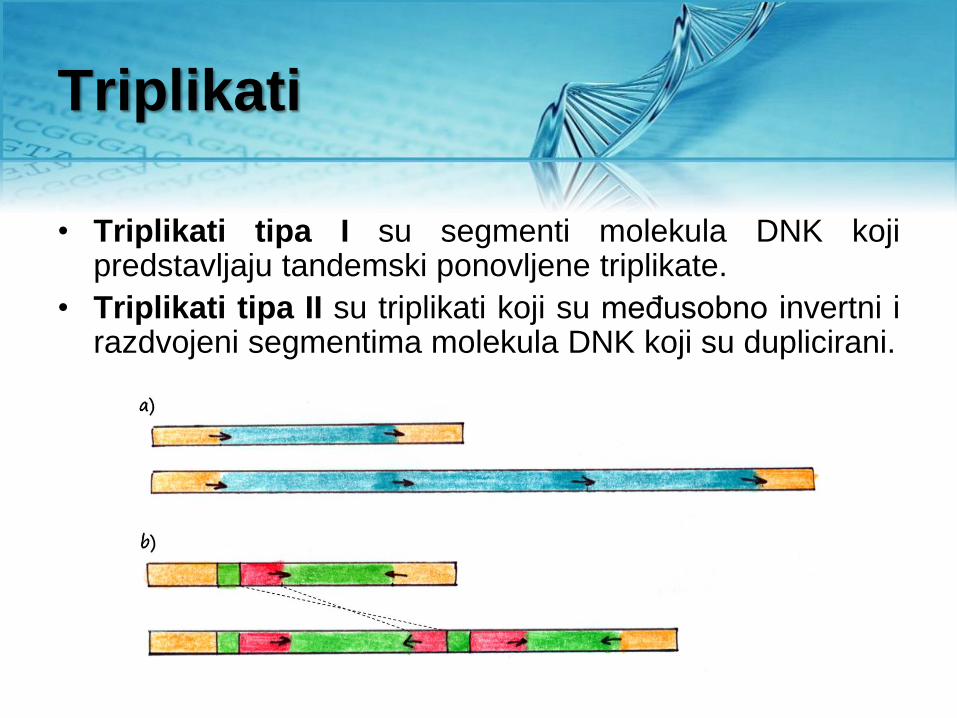
Triplikati
• Triplikati tipa I su segmenti molekula DNK koji predstavljaju tandemski ponovljene triplikate.
• Triplikati tipa II su triplikati koji su međusobno invertni i razdvojeni segmentima molekula DNK koji su duplicirani.

CNV u genomu čoveka
• 300.000 CNV regiona
• Pericentromerni i subtelomerni regioni
• Prosečna veličina CNV iznosi 2,9 kb
• Oko 100 gena deletirano bez efekta na fenotip
• CNV se nasleđuju
• De novo CNV registrovane kod jednojajčanih blizanaca
• Česte (eng. common CNV) čine preko 80% svih CNV
• Retke (eng. rare CNV) koje su obično povezuju sa
bolestima čoveka

CNV češće pogađaju 3 kategorije
gena za proteine
1) Proteine signalnih puteve imunskog sistema i međućelijske
komunikacije
2) Proteine uključene u interakcija sa spoljašnom sredinom
(imunski odgovor i čulo mirisa, npr.)
3) Proteine retrovirusa i proteine povezane sa mehanizmima
transpozicije.
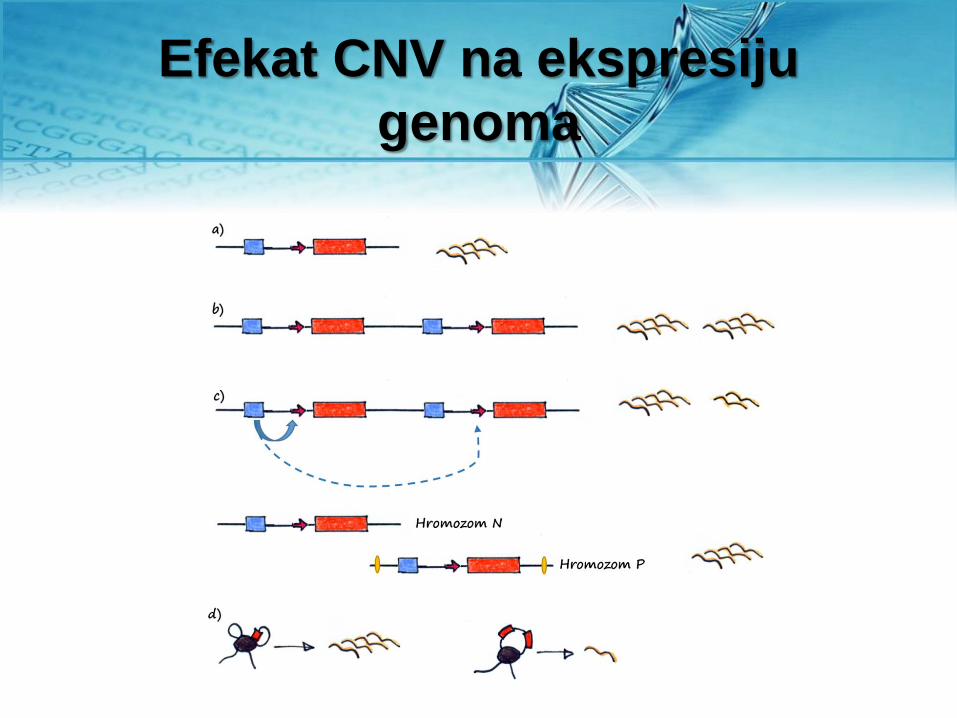
Efekat CNV na ekspresiju
genoma

CNV u drugim vrstama
• Primati, psi, miševi, konji, krave,
kokoške, kukuruz, Arabidopsis
thaliana, Vinska mušica, pivski
kvasac, C. Elegans....
• 331 CNVs kod 20 šimpanza od
toga 74 CNVs detektovane u istim
regionima kao i kod čoveka
• Distribucija CNV kod šimpanze i
čoveka u prilog hipotezi o
dupliciranju gena kao mehanizmu
povećanja genoma tokom
evolucije
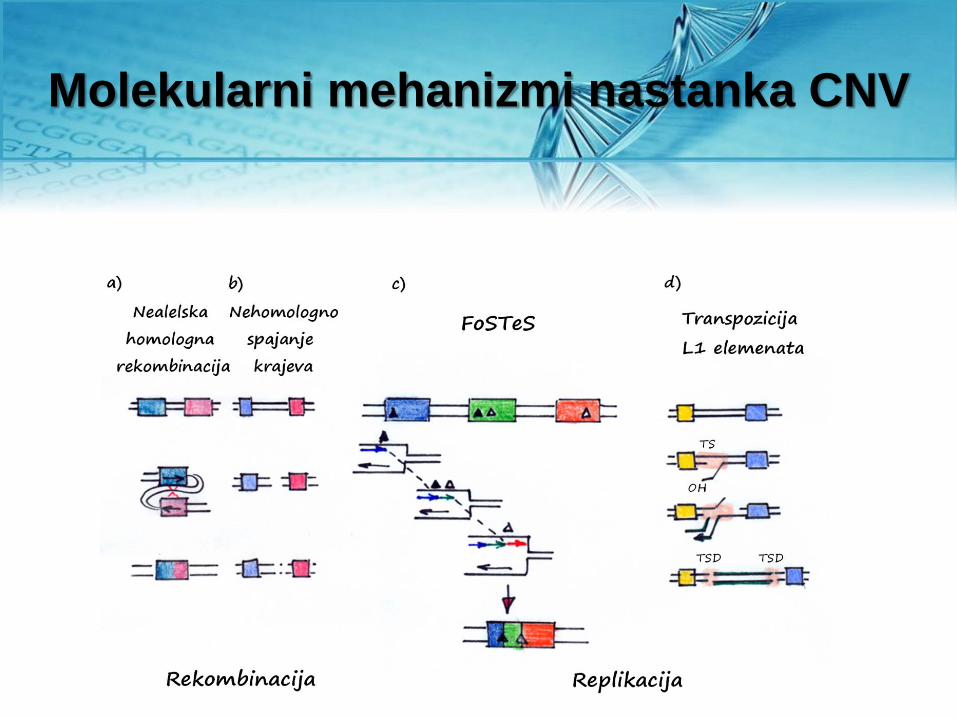
Molekularni mehanizmi nastanka CNV
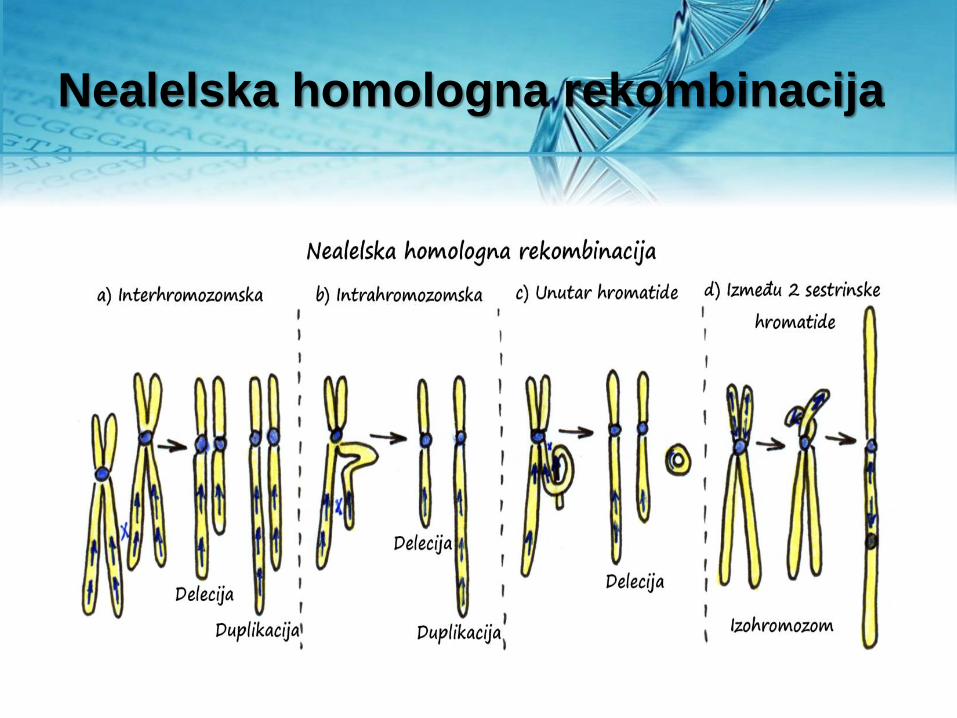
Nealelska homologna rekombinacija

Molekularni mehanizmi nastanka CNV
• Dvolančani prekidi u molekulu
DNK čoveka najčešće se
repariraju nehomologim
spajanjem krajeva. Ovaj
mehanizam može biti uzrok
delecije određenog segmenta
molekula DNK.
• Tokom transpozicije moguće je
“nastanak” CNV po tipu
duplikacije - retro CNV.

Mehanizam FoSTeS (eng. fork stalling and template switching)
• Kompleksne CNV
• To je mehanizam reparacije
jednolančanih prekida u molekulu DNK
tokom procesa replikacije.
• Zaostajući lanac, na osnovu
mikrohomologije nastavlja replikaciju
korišćenjem druge replikacione viljuške
koja se nalazi u blizini.
• U zavisnosti da li se nova replikaciona
viljuška nalazi uzvodno ili nizvodno,
nastavak replikacije dovodi do delecije ili
duplikacije.

CNV i bolesti ljudi
• Dosadašnja istraživanja pokazala su snažnu asocijaciju
pojedinih CNV koje obuhvataju gene za proteine sa
razvojem psihijatrijskih bolesti (uključujući i autizam),
gojaznosti, psorijaze, šećerne bolesti, zapaljenskih
bolesti creva, itd.

CNV i bolesti ljudi
• Poslednjih godina istražuje se moguća uloga CNV u
procesima neoplastične transformacije koji su
odogovorni za razvoj karicinoma debelog creva, pluća i
prostate.

Oboljenja broja kopija
(eng. copy-number disorder)
• Za neke bolesti čoveka molekularno-biološkim
analizama pokazano je da su jedini do sada utvrđeni
genetički uzročnici razvoja bolesti CNV.
• Određene kongentalne bolesti srca ili bubrega su neke
od njih.
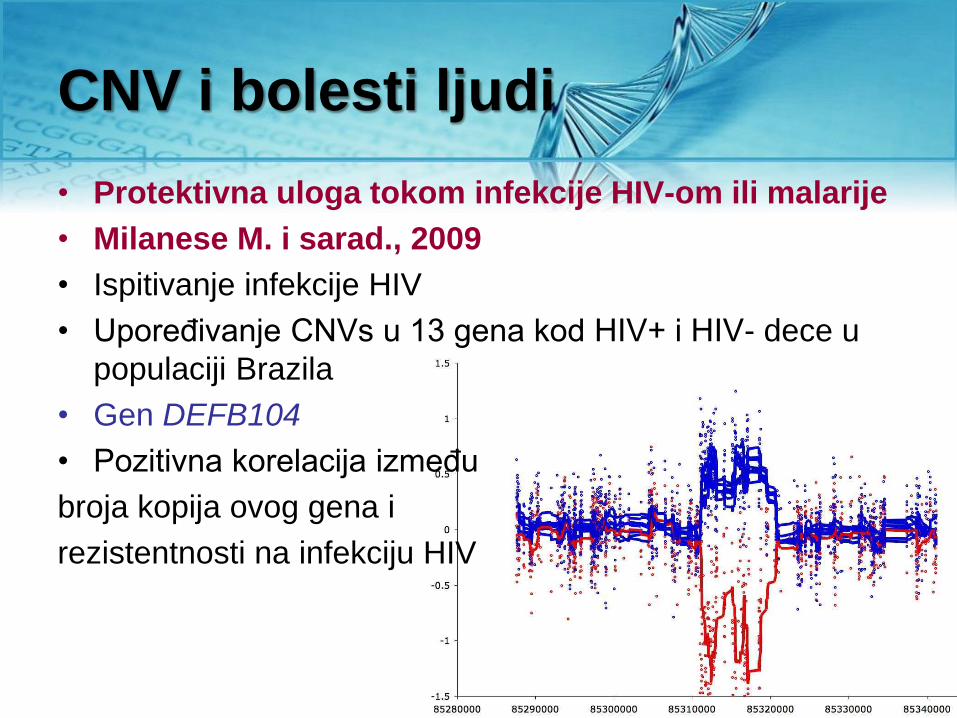
CNV i bolesti ljudi
• Protektivna uloga tokom infekcije HIV-om ili malarije
• Milanese M. i sarad., 2009
• Ispitivanje infekcije HIV
• Upoređivanje CNVs u 13 gena kod HIV+ i HIV- dece u
populaciji Brazila
• Gen DEFB104
• Pozitivna korelacija između
broja kopija ovog gena i
rezistentnosti na infekciju HIV
CNV i bolesti ljudi
• Personalna medicina
• Farmakogenomika
• Uloga CNV u individualnim odgovorima
na primenjenu terapiju u lečenju brojnih
bolesti čoveka

Budućnost
• Copy Number Variation Project
• Analiza genoma 269 nasumično izabranih individua
iz svih krajeva sveta
• Otkriće uloge CNV u fenomenima odgovornim za
ekspresiju
• Ključno pitanje – značaj CNV u evoluciji genoma
Recommended