Embed Size (px)
Citation preview

UNIVERSIDAD TECNICA DE BABAHOYO
FACULTAD CIENCIAS DE LA SALUD
BIOLOGIA
DOCENTE:DR. HERMAN ROMERO

CITOESQUELETO
Concepto El citoesqueleto es una estructura proteica que permite soportar la membrana plasmática, la formación de estructuras como los lamelipodios, el movimiento de vesículas y otros elementosintracelulares. La polimerización controlada de actina y tubulina es responsable de la movilidad de las células eucariotas y de la forma de éstas. El movimiento de las células eucariotas es el resultado de la acción coordinada de formación de extensiones, adherencias y retracciones de la membrana, en donde la red de actina y las interacciones entre estas y los motores moleculares juegan un papel fundamental.

MICROFILAMENTOS - CITOESQUELETO DE ACTINA
La actina es la proteína intracelular mas abundante en eucariotes. Puede llegar a representar hasta el 10% del peso total de proteína. Pesa alrededor de 43 kD y está conservada evolutivamente. Algunos organismos tienen un solo gen (levaduras) mientras que otros tienen múltiples genes. Por ejemplo en humanos existen 6 genes diferentes y en algunas plantas puede haber hasta 60. Existe como un monómero globular llamado G-actina y como polímero filamentoso, F-actina. Cada molécula de actina tiene un ión de Mg+2 que forma complejo bien con ATP o con ADP, existiendo por lo tanto cuatro formas diferentes de actina. El plegamiento de la proteína permite la formación de dos lóbulos con una hendidura en la mitad que permite la unión del ATP y el Mg+2, y un cambio de conformación.
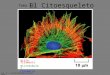
MICROTÚBULOS Y PROTEÍNAS MOTORAS Los microtúbulos son polímeros de la proteína tubulina, un heterodímero de a y b tubulina de unos 55 kD, de secuencias igualmente muy conservadas. Estas proteínas guardan una homología grande con la proteína bacteriana FtsZ que juega un papel importante en la división celular.
Las proteínas globulares pueden también agruparse en diminutos túbulos huecos que actúan como entramado estructural de las células y, al mismo tiempo, transportan sustancias de una parte de la célula a otra. Cada uno de estos microtúbulos está formado por dos tipos de moléculas proteicas casi esféricas que se disponen por parejas y se unen en el extremo creciente del microtúbulo y aumentan su longitud en función de las necesidades.
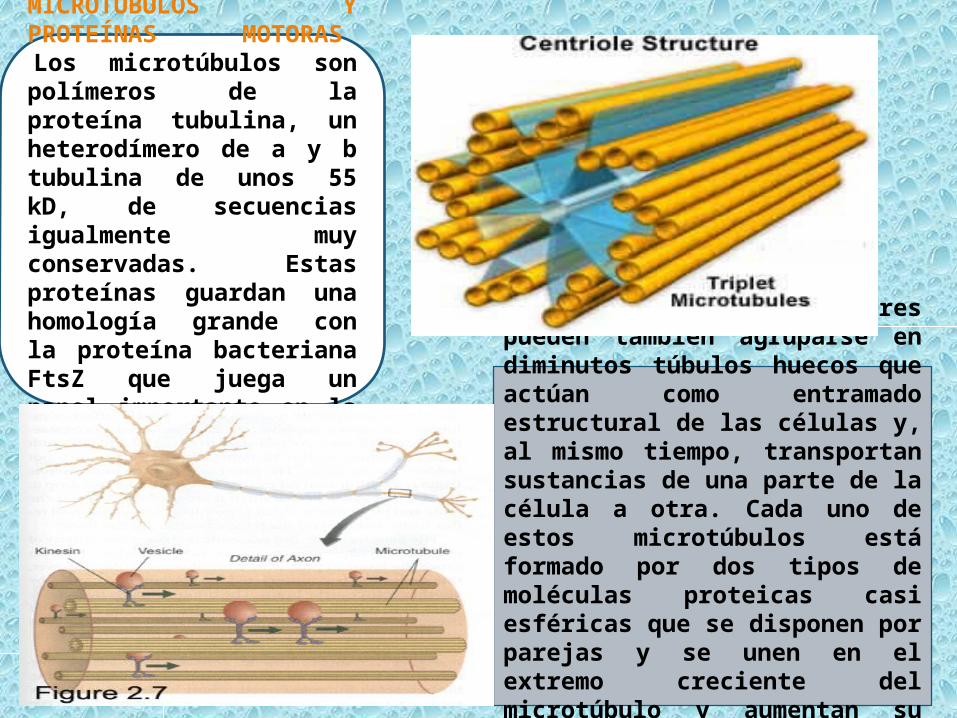
LAS PROTEINAS MICROTUBULARESLas proteínas microtubulares asociadas (MAPs) estabilizan a los microtúbulos y a estos con los organelos y membrana. Las proteínas motoras o ATPasas asociadas a microtúbulos (un subtipo de MAPs) movilizan organelos y otros elementos sobre los microtúbulos. * Quinesinas * Dineínas citoplasmáticas * Dineína ciliar / flagelar * Dinamina

FILAMENTOS INTERMEDIOS Proteínas fuertes, estables y poco solubles. Diámetro de aprox. 10 nm. Compuestas por proteínas fibrosas que se combinan en dímeros helicoidales, que se asocian para formar tetrámeros alargados (protofibrillas). Cuatro protofibrillas conforman un filamento intermedio. Son apolares y tienen como funciones mantener la fuerza de tensión celular (principal) y como soporte mecánico.
Clasificación de las proteínas de los filamentos intermedios: Tipo I: Queratinas ácidas EpitelioTipo II: Queratinas básicas EpitelioTipo III: Vimentina MesenquimaDesmina MúsculoPeriferina NeuronasTipo IV: NF (L,M,H) Neuronas Internexina S. Nervioso en formación.Tipo V: Lamininas A,B,C Núcleo todas las células.Tipo VI
ADHESIONES INTERCELULARES. Se producen antes de que pueda ser organizada una unión de anclaje. Indispensables para el desarrollo de tejidos en los que participa la migración celular.Se requiere:1. Un mecanismo que dirija las células hasta su destino final (quimiotaxis) 2. Extendido de moléculas adhesivas en la matriz extracelular o sobre la superficie de determinadas células guiando las células migradoras (orientación de vía).

UNIONES DE ANCLAJE Ampliamente distribuidas en los tejidos animales. Se constituyen como unidades
estructurales resistentes, conectando los elementos citoesqueleticos de una
célula a los esqueletos de sus vecinas o a la matriz extracelular
PLASMODESMOS Las células vegetales están aisladas entre sí por paredes celulares rígidas y gruesas, de manera que no tienen necesidad de desmosomas para fines de resistencia. Es común que estas paredes estén impregnadas de materiales impermeables lo que también hace que no requieran uniones estrechas.

CENTR
IOLO
S CILI
OS Y FL
AJEL
OS

MICROTÚBULOS.ESTRUCTURA.• Los microtúbulos los principales componentes del citoesqueleto de las células eucariotas.• Pueden estar dispersos en la célula o formando estructuras definidas: cilios, flagelos, centriolos.• Están formados por dímeros de alfa- y beta- tubulina que se organizan formando un tubo alargado. El SNC es el tejido del organismo del que se aísla (10-20% del total de proteínas).• Las tubulinas están codificadas por una familia de genes estrechamente relacionados entre sí y muy conservados en la filogenia, como la actina; porque tienen que interaccionar con otras proteínas estructurales.• Son tubos largos y relativamente rígidos. Sus paredes están formados por unas subunidades proteicas globulares denominadas tubulinas. Éstas se asocian en dímeros compuestos de dos tipos de tubulinas: alfa y beta.• Estas parejas se alinean ordenadamente, mediante enlaces no covalentes, en filas longitudinales que se denominan PROTOFILAMENTO.

PROPIEDADES DE LA POLIMERIZACIÓN DE LA TUBULINA.Resumen global de dichas propiedades:• A concentraciones de alfa/beta-tubulina superiores a la Cc los dímeros se polimerizan para formar microtúbulos; por debajo de la Cc, los microtúbulos se despolimerizan.• El nucleótido, GTP o GDP, unido a la beta-tubulina hace que la Cc para el ensamblaje en los extremos (+) y (-) de un microtúbulo sea diferente; por analogía con el ensamblaje de actina filamentosa, se define el extremo (+) como el preferido por el ensamblaje.• Con concentraciones superiores de alfa/beta-tubulina a la Cc para la polimerización, los dímeros se agregan en mayor cantidad al extremo (+).• Cuando la concentración de alfa/beta-tubulina es más elevada que la Cc del extremo (+) pero menor que la Cc del (-), se puede dar un crecimiento en una sola dirección agregando subunidades a un extremo y disociando subunidades del extremo opuesto.
. INESTABILIDAD DINÁMICA.Una vez se ha producido el comienzo de la formación de un microtúbulo, la incorporación de nuevos dímeros de tubulina hace que el microtúbulo crezca en longitud. Este crecimiento a veces se detiene repentinamente y el microtúbulo comienza a despolimerizarse, llegando a veces incluso a desaparecer, o más frecuentemente reinicia el proceso de polimerización.
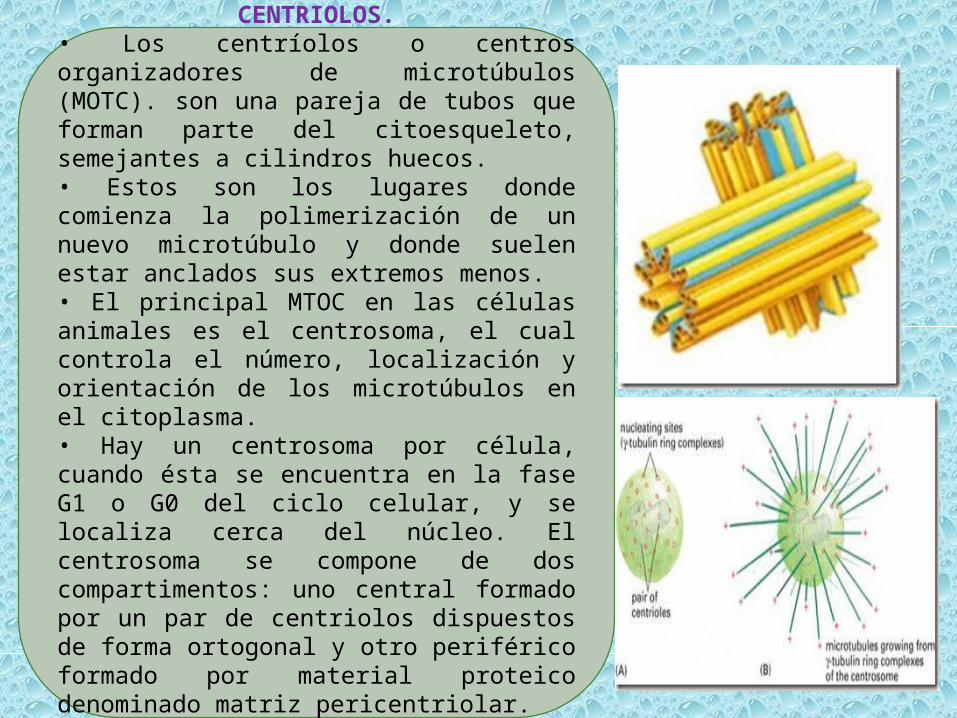
CENTRIOLOS.• Los centríolos o centros organizadores de microtúbulos (MOTC). son una pareja de tubos que forman parte del citoesqueleto, semejantes a cilindros huecos.• Estos son los lugares donde comienza la polimerización de un nuevo microtúbulo y donde suelen estar anclados sus extremos menos.• El principal MTOC en las células animales es el centrosoma, el cual controla el número, localización y orientación de los microtúbulos en el citoplasma.• Hay un centrosoma por célula, cuando ésta se encuentra en la fase G1 o G0 del ciclo celular, y se localiza cerca del núcleo. El centrosoma se compone de dos compartimentos: uno central formado por un par de centriolos dispuestos de forma ortogonal y otro periférico formado por material proteico denominado matriz pericentriolar.

ESTRUCTURA.Cada centríolo está formado por nueve tripletes de microtúbulos que forman todos estos juntos y unidos entre si un círculo. El más interno se llama microtúbulo A y está completo (compuesto de trece protofilamentos). A él se unen dos microtúbulos: el microtúbulo B que comparte tres protofilamentos con el A y el microtúbulo C, el más externo, que comparte tres protofilamentos.
Su pared contiene 9 tripletes de túbulos que se disponen de forma regular y están inclinados formando un ángulo de 40 grados con el radio. Tienen una estructura de 9 + 0.– A lo largo de la pared que pone en contacto los microtúbulos del doblete del microtúbulo corre un filamento delgado formado por la proteína TECTINA que parece estar relacionada con los filamentos intermedios. Parece colaborar en la formación de la pared compartida de los túbulos A y B.– Los tripletes se unen entre sí por puentes de nexina.

POLIMERIZACIÓN DE LOS MICROTÚBULOSLos microtúbulos que emanan desde el centrosoma terminan en el material pericentriolar, no en los centriolos, y es el material pericentriolar el que inicia el montaje de los microtúbulos. Centriolina, y sobre todo la Gamma-tubulina (en realidad un complejo de proteínas en anillo asociado llamado (Gamma-TuRc) uniéndose al extremo “menos” (-) de los microtúbulos, tiene un papel clave en el cebado de la nucleación del ensamblaje de los microtubulos que crecen alargándose a partir de ahí por la adición de protómeros de alfa/beta-tubulina libres del citosol a su extremo “más” (+), así como en el anclaje de los microtubulos al centrosoma (otras proteínas como la nineina están también involucrada).
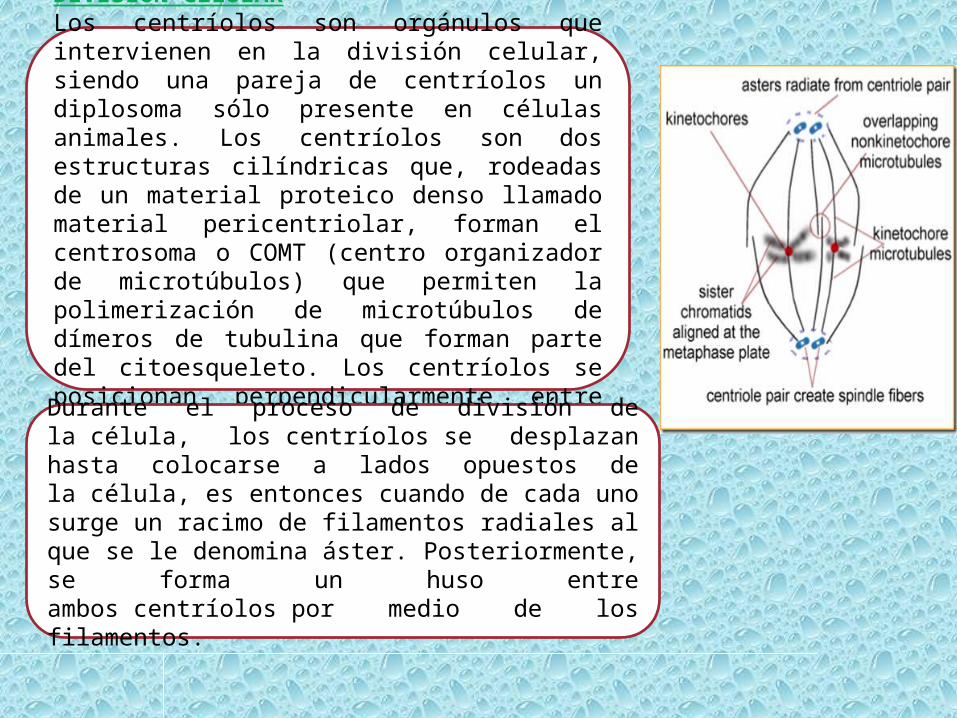
DIVISIÓN CELULARLos centríolos son orgánulos que intervienen en la división celular, siendo una pareja de centríolos un diplosoma sólo presente en células animales. Los centríolos son dos estructuras cilíndricas que, rodeadas de un material proteico denso llamado material pericentriolar, forman el centrosoma o COMT (centro organizador de microtúbulos) que permiten la polimerización de microtúbulos de dímeros de tubulina que forman parte del citoesqueleto. Los centríolos se posicionan perpendicularmente entre sí.
Durante el proceso de división de la célula, los centríolos se desplazan hasta colocarse a lados opuestos de la célula, es entonces cuando de cada uno surge un racimo de filamentos radiales al que se le denomina áster. Posteriormente, se forma un huso entre ambos centríolos por medio de los filamentos.

CILIOS, FLAGELOSLos cilios y los flagelos son apéndices móviles existentes en la superficie celular. Los flagelos y cilios son estructuras microtubulares, que se extienden hacia afuera en algunas células y funcionan para darles movimiento. Contienen una estructura central altamente ordenada, constituida generalmente por más de 600 tipos de proteínas, envuelta por el citosol y la membrana plasmática.
CILIOS: son cortos y en alto número (menos de 10 micras). Son estructuras digitiformes que pueden moverse en sincronía. Los cilios se encuentran en epitelios especializados en eucariontes. Por ejemplo, cilios barren los fluidos sobre células estacionarias en el epitelio de la traquea y tubos del oviducto femenino.
FLAGELOS: Son más largos que los cilios (200 micras) y están en bajo número. Son apéndices como látigos que ondulan para mover las células.
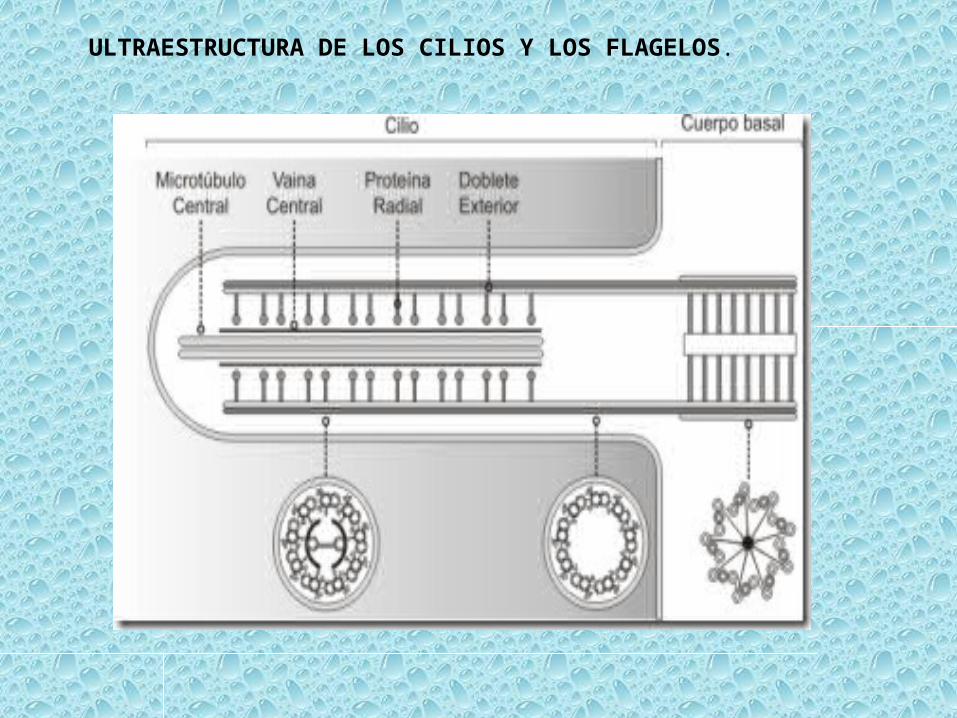
ULTRAESTRUCTURA DE LOS CILIOS Y LOS FLAGELOS.

TALLO O AXONEMA.– Se encuentra rodeado por una membrana plasmática y sitúa en su interior dos microtúbulos centrales y nueve pares de microtúbulos periféricos que están orientados paralelamente al eje principal. Se dice que su estructura es 9 + 2.– Los dos microtúbulos centrales son completos (13 protofibrillas) y se hallan rodeados por una vaina central.– Cada par de microtúbulos periféricos (dobletes) consta de dos túbulos, uno de ellas completo (A) y otro con solo 10 u 11 protofilamentos (B).– El túbulo A presenta dos brazos formados por DINEINA, que se dirigen hacia el túbulo B del par adyacente. Forma un brazo interno y otro externo que son curvados y acaban en una cabeza globular. Los brazos están espaciados a lo largo del túbulo A (24 nm).– Cada túbulo A del doblete se une al adyacente mediante la NEXINA que funciona como una cinta elástica. Desde cada doblete, y proyectándose hacia el interior, se proyectan las fibras radiales que se extienden hasta una vaina interna que rodea a el par central de microtúbulos.– De la vaina central salen unas proyecciones que junto a las fibras radiales regulan el batido de los cilios.
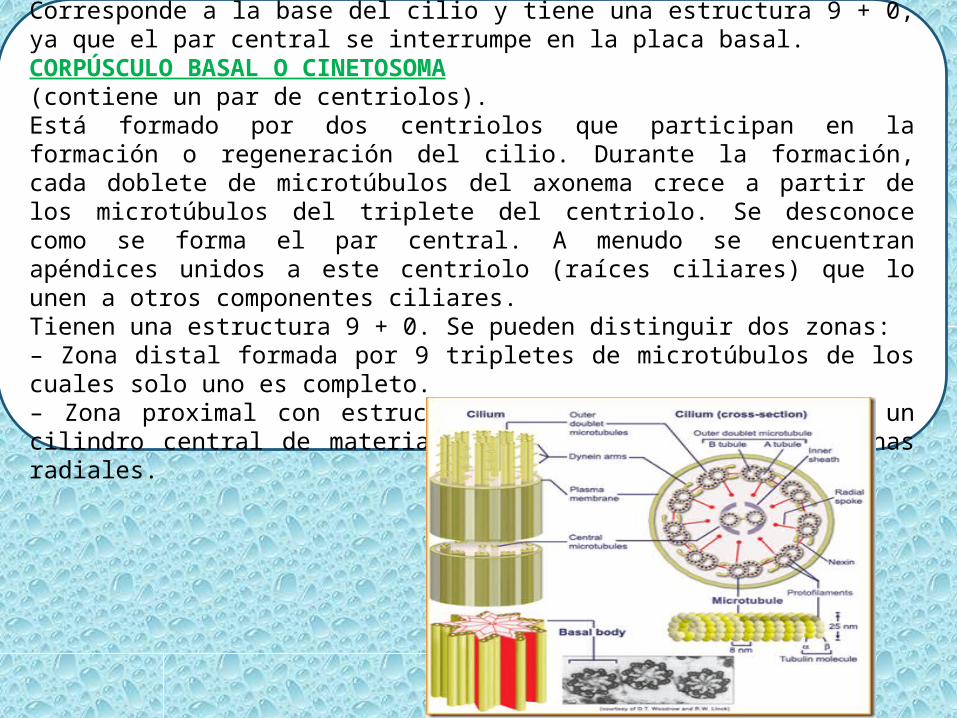
ZONA DE TRANSICIÓN.Corresponde a la base del cilio y tiene una estructura 9 + 0, ya que el par central se interrumpe en la placa basal.CORPÚSCULO BASAL O CINETOSOMA (contiene un par de centriolos).Está formado por dos centriolos que participan en la formación o regeneración del cilio. Durante la formación, cada doblete de microtúbulos del axonema crece a partir de los microtúbulos del triplete del centriolo. Se desconoce como se forma el par central. A menudo se encuentran apéndices unidos a este centriolo (raíces ciliares) que lo unen a otros componentes ciliares.Tienen una estructura 9 + 0. Se pueden distinguir dos zonas:– Zona distal formada por 9 tripletes de microtúbulos de los cuales solo uno es completo.– Zona proximal con estructura en rueda de carro y con un cilindro central de material opaco del que parten 9 laminas radiales.

MEMBRANA CILIAR.La membrana del cilio se continúa a través de la zona apical de la membrana plasmática. Se forma por fusión de vesículas alrededor del centriolo o de compartimentos especializados, y consta de una serie de dominios que se distinguen entre sí por su composición de proteínas y probablemente también de lípidos, adaptada ésta última en cada caso al medio donde se desenvuelven.Se puede distinguir los siguientes dominios de membrana:· Dominio periciliar· Base de la membrana ciliar· Membrana del tallo ciliar.
MECANISMO DE DESLIZAMIENTO DEL AXONEMA.– La fuerza de flexión se produce por el deslizamiento de los microtúbulos. La dineina es una ATPasa que permite el deslizamiento de un doblete sobre otro, hasta alcanzar 9 veces más su longitud.-Los brazos de dineina en presencia de ATP contactan con el doblete vecino. La dineina es un gran complejo proteico que tiene dos o tres cabezas globulares unidas a una raíz común a través de cadenas delgadas y flexibles. Cada cabeza globular tiene actividad ATPasa que se potencia unas 6 veces cuando se une a un microtúbulo.– El proceso de deslizamiento de la dineina es similar al de las cadenas de miosina sobre la actina. Este movimiento genera una fuerza que impulsa los dobletes microtubulares adyacentes hacia el extremo del axonema.– Para poder producir la flexión local y que se propague desde la base hasta el polo apical, han de existir controles que coordinen el movimiento de la dineina. Se cree que no depende de calcio y sí de interacciones proteína-proteína.
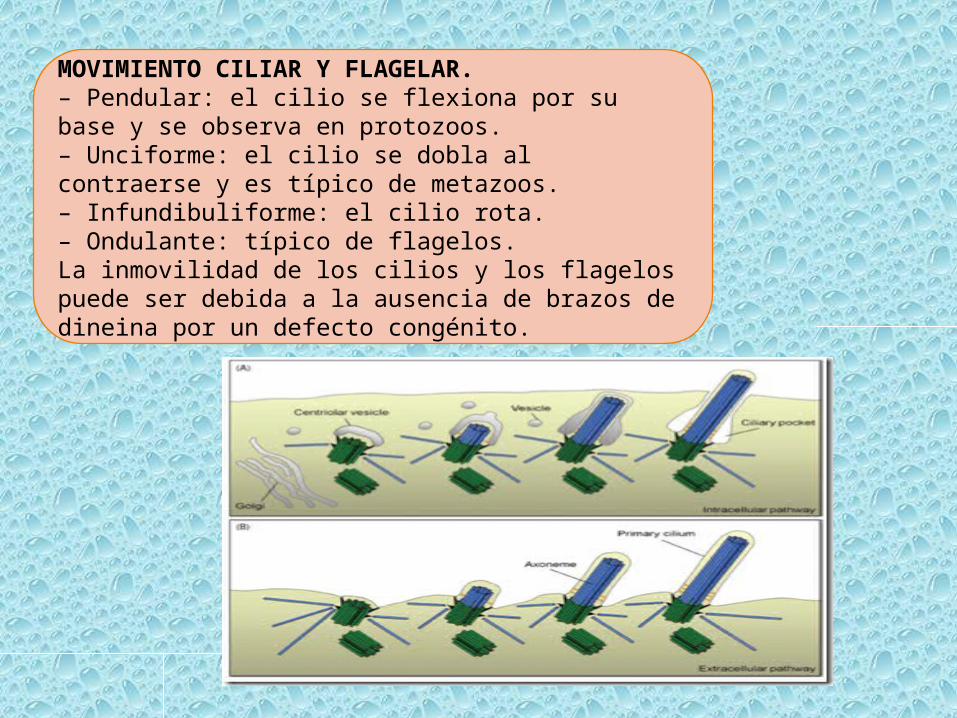
MOVIMIENTO CILIAR Y FLAGELAR.– Pendular: el cilio se flexiona por su base y se observa en protozoos.– Unciforme: el cilio se dobla al contraerse y es típico de metazoos.– Infundibuliforme: el cilio rota.– Ondulante: típico de flagelos.La inmovilidad de los cilios y los flagelos puede ser debida a la ausencia de brazos de dineina por un defecto congénito.

. FUNCIÓN.Los flagelos pueden propulsar células móviles en un líquido, mientras que los cilios se sitúan normalmente en células estacionarias, y gracias a su impulso mueven líquidos o elementos contenidos en él.
Casi todos los eucariotas poseen células ciliadas, salvo los que tienen pared celular, que carecen habitualmente de ellos. En vertebrados, prácticamente todos los tipos celulares tienen cilios o proceden de células que los tuvieron.
Los cilios móviles intervienen a la propulsión de organismos unicelulares, la limpieza de las vías respiratorias y el desplazamiento de los gametos, pero también contribuyen a regular el balance hídrico en los órganos excretores, la circulación de fluidos en la cavidad celómica, el sistema nervioso, el filtrado de partículas en las branquias.
Los corpúsculos básales y los centriolos son estructuras interconvertibles y originan el organizador microtubular. Al iniciar la mitosis, los flagelos son reabsorbidos y sus corpúsculos migran junto al núcleo para organizar el huso mitótico. Al finalizar la mitosis los centriolos vuelven a formar los cilios.
EVOLUCIÓNLas teorías se pueden clasificar en tres categorías: De origen endosimbionte, de origen viral y de origen en el transporte vesicular o teorías endógenas.
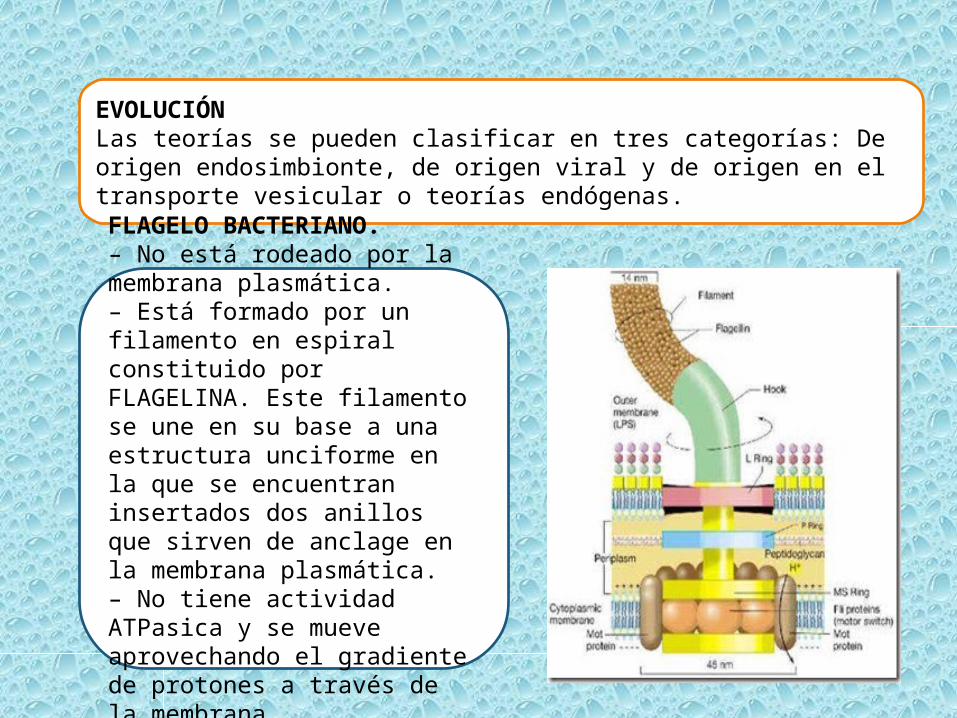
FLAGELO BACTERIANO.– No está rodeado por la membrana plasmática.– Está formado por un filamento en espiral constituido por FLAGELINA. Este filamento se une en su base a una estructura unciforme en la que se encuentran insertados dos anillos que sirven de anclage en la membrana plasmática.– No tiene actividad ATPasica y se mueve aprovechando el gradiente de protones a través de la membrana.