Upload
barrioam
View
363
Download
9
Embed Size (px)
Citation preview
Whyshouldwelookatthegenomein3D
!BMC,Nov27th2015lvaroMartnezBarrio,[email protected]/in/ambarrio@ambarrio
-log 1
0P
-log 1
0P
AB1NS*
Atlantic Ocean
SBSkagerrak
BH*BSBV
Baltic Sea
0.08
BAS30
BAH60
BAS16
NS56
BAS39
BAH30
SB40
BAV33
SB14
NS8
BAH51
NS34
SB50
NS7
BAH39
BAH8
NS57
BAH53
BAS55
BAH5
AB4
SB20
SB5
AB56
BAS1
BAS21
BAV58
BAH10
NS13
BAV38
AB25
BAS7
SB41
BAH19
BAH59
SB30
BAH43
NS22
AB29
BAS28
SB16
AB28
NS43
SB44
SB48
SB18
SB38
AB10
AB31
BAV47
NS54
SB60
NS44
SB33
AB18
BAS35
BAV6
BAS43
SB4
BAV22
SB55
NS47
BAH13
SB47
BAH3
BAH42
BAH28
AB6
NS46
SB2
AB26
BAS47
SB22
BAH32
AB49
BAH14
AB48
NS39
BAS14
NS2
BAV10
AB50
NS15
BAS3
NS53
BAV12
BAH52
NS1AB1
BAH35
BAH33
BAH26
AB13 NS49
NS5
BAS6
BAS20
BAH54
BAH48
BAH56
BAS36
BAV18
SB27
NS17
BAV28
BAH38
BAS40
NS3
AB35
AB9
BAS49
SB42
BAH22
AB8
BAV14
BAS45
BAS53
BAH2
BAH12
NS42
BAV27
NS9
NS19
BAS37
NS37
BAS25
SB28
AB14
BAS58
NS59
BAS46
NS30
AB45
BAV4
BAH11
NS55
AB2
AB47NS52
BAS60
BAS52
SB54
BAV16
AB19
NS32
NS45
BAV34
BAS38
BAH45
BAS11
SB29
SB1
SB13
AB43
AB11
NS12
BAS10
NS40
NS33
SB19
NS16
BAV40
BAS54
SB26
BAH57
BAV56
BAH29
BAS56
BAS5
BAH18BAV43
NS14
BAH44
SB15
BAV37
SB8
NS27
BAV45
BAV36
BAS32
NS41
BAS34
BAV55
BAH37
AB42
AB55
BAH24
SB37
BAV8
BAH55
BAS4
BAV24
SB56
NS50
BAV30
NS35
BAV17
SB3
NS60
NS24
AB51
NS6
SB43
SB12
NS23
BAH17
NS38
NS11
BAV49
AB34
BAV52
BAH23
BAS19BAS27
AB40
SB45SB11
BAH47
SB53
NS48
BAH4
BAV59
AB21
BAS33
AB38AB20
BAV48
BAV9
SB31
BAV2
BAH21
BAH36
BAV29BAV35
BAH20
BAV11
NS25
NS21
BAS9
SB52SB10
SB9
NS26
BAV26
NS10
BAH46
BAS57
SB17
SB25
BAV32
BAS41
AB59
NS31
AB30
BAH9BAH49
AB54
SB49
BAV1
AB27
BAV5
BAS42
BAV39
AB22
NS51
BAV50
AB12
AB32
AB39
SB34
AB41
BAV15
BAS15
SB6
AB24
BAV53
SB35
AB60
BAS13
AB44AB57
BAS18
BAS50
BAV13
BAV54
AB15
AB3
BAS17BAV23
SB59
BAV51
BAH41
AB46
SB58
BAS22
BAH27
BAS24
BAV3
NS36
BAS51
NS28
BAH34
AB36
BAH7
SB36
SB21
BAS48
AB17
BAS12
BAV57
NS58
AB16
BAV41
BAH40
AB7
AB37
BAH25
NS20
SB24
BAV19
BAV20
BAH31
BAV21
BAS8
NS18
BAV60
BAV44SB46
BAS26
AB33
SB7
SB23
BAV42
AB53
SB57
BAS2
BAV25
AB58
BAH58
SB32
BAH6
AB52
NS29
BAV31
BAH16
BAH15
BAS23
BAH50
BAS29
BAS44
BAV46
BAS59
SB51
AB23
AB5
BAS31
BAV7
SB39
NS4
A
s218
s1523
s2123
s273s899
0
50
100
150
200
250
300
-log
10
(P)
SNP position
s346
0.05
.
NS*AB1
AI
SBSH
BF*
BH*KB
KTBA
BRBK
BUBSBV
BHBGBVBC
PH
B
D
Salin
ity(
)
1.836 Mb 1.842 Mb
Normalized copy number
2 10020 40 60 80
1 8361 8361 836 MbMbMb
3
6
6
7
7
12
20
25
35
35
Pop
s
AI
AB1
NS*
SH
SB
KB
KT
BF*
BC
BR
BA
BG
BV
BH
BH*
BS
BV
BU
BK
PH
7
6
6
6
6
8
23
25
35
35
High choriolytic enzyme 2
Atlantic Ocean
C
FBX
W7
FHD
C1
AR
FIP
1
ND
UFA
F2
TMEM
2
PG
F5
FOX
D5
NR
N1
PR
LR
HFE
MH
C-I
LRR
C8
C
RR
EB1
AB1NS*
BH*BSBV
s218119.4 kb
s152333.58 kb
s89910.93 kb
s212366.51 kb
s27332.66 kb
NR
N1
s152333.58 kb
PR
LRs89910.93 kb
FBX
W7
FHD
C1
AR
FIP
1
ND
UFA
F2
TMEM
2
PG
F5
FOX
D5
s218119.4 kb
HFE
MH
C-I
LRR
C8
C
s212366.51 kb
RR
EB1
s27332.66 kb
Baltic Sea
SkagerrakSB
-log
10
(P)
50
100
0
150
200
00.20.40.6
0.81
Fst
119.4 kb
scaffold331
-log
10
(P)
Gap
E
KLHL33 SLC12A3CBLN3 KLHL33
C1QL4
Gap
~ 65 kb
Fig. 2
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
0
Alle
le f
req
uen
cy
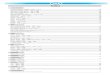
Figure 2 Genetic differentiation between Atlantic and Baltic herring. (A) Manhattan plotof significance values testing for allele frequency differences between pools of herring frommarine waters (Kattegat, Skagerrak, Atlantic Ocean) versus the brackish Baltic Sea. Lowerpanel, corresponding plot for scaffold 218 only; both P- and FST - values are shown. (B)Neighbor-joining phylogenetic tree based on all SNPs showing genetic differentiation in thiscomparison (P < 10-20). (C) Comparison of allele frequencies in five strongly differentiatedregions. The major allele in the AB1 sample (Atlantic Ocean) was used as reference at eachSNP. Lower panel, neighbor-joining tree based on haplotypes formed by 128 differentiatedSNPs from scaffold 218. (D) Heat map showing copy number variation partially overlappingthe HCE gene. Orientation of transcription is marked with an arrow. Population samples andsalinity at sampling locations are indicated to the right; abbreviations are explained in Table 2. (E) Strong genetic differentiation between Atlantic and Baltic herring in a region downstream of SLC12A3; statistical significance based on the 2 test is indicated.
-log 1
0P
Whyshouldwelookatthegenomein3D
http://webvideomarketingportugal.com/httpthenextweb-commedia20130920the-future-of-cinemas/
http://webvideomarketingportugal.com/httpthenextweb-commedia20130920the-future-of-cinemas/
-log 1
0P
WhereismycausativeSNP? Candidategeneapproach(100kbwindows) Inferpathwaysthatarecommon
12
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer potentialIf active enhancers have promoter activities, it follows thatwhat we classically refer to as promoters could also func-tion as enhancers in certain situations. Although in gener-al a neglected topic of research, we find some clues tosupport this in studies of larger chromatin architectures.
Regulatory elements often interact in close physicalproximity in RNAPII foci that are sometimes referred toas transcription factories [37], and the activity of eachregulatory element within these foci can be influencedby other regulatory elements involved (Figure 1C). Al-though the regulatory mechanisms remain poorly charac-terized, a common model posits that regulatory sequencessynergistically achieve high levels of transcriptional activ-ity by increasing the local concentration of factors neededfor transcription [3741] (Figure 1C). Studies based onchromatin conformation capture have observed RNAPIIfoci involving multiple gene promoters, with few or noenhancers [29,42]. In such arrangements, the degree towhich each gene promoter acts as a promoter or enhancermay depend on the local context of chromatin [42],expressed TFs, and sequence context (e.g., the occurrenceand strength of core promoter elements and TF bindingsites). Using in vitro reporter assays, Li et al. found thatcertain gene promoters in such constellations in MCF7cells had enhancer potential [42], suggesting that they
Box 2. Regulatory elements and associated chromatin
modification states
The N-terminal tails of histone proteins H2A, H2B, H3, and H4 withinnucleosomes can be chemically modified. In particular, amino acidresidues can be methylated or acetylated, which correlates withfunctional properties of the chromatin. For instance, H3K27ac (acet-ylation of lysine 27 in histone H3) is correlated with transcriptionalactivity. The locations of histones marked with a certain histonemodification can be measured genome wide using chromatin im-munoprecipitation (ChIP) coupled with either tiling microarrays(ChIP-chip) or sequencing (ChIP-seq) [38]. Profiling of histone mod-ifications in different cell types has enabled systematic inference ofdifferent types of functional genomic entities [12,53,77,78]. Suchstudies have shown that, typically, the nucleosomes flanking activegene promoters are enriched in H3K27ac, H3K4me3 (trimethylationof lysine 4 in histone H3), and, to some degree, H3K4me1 (mono-methylation of lysine 4 in histone H3), whereas enhancers are en-riched in H3K27ac and H3K4me1, but not H3K4me3, or relatively lowlevels thereof. In addition, several marks have been found to localizeat both enhancers and promoters. Most notably, H3K27ac marksactive promoters and a subset of enhancers that appear to be activelyregulating gene transcription, leading to the notion of classifyingregulatory elements into active and poised classes [15,79]. Poisedregulatory elements sometimes carry bivalent marks associated withactivity and repression at the same time, for instance promotershaving H3K4me3 and H3K27me3 (the latter mark is usually found atinactive gene promoters). Collectively, these studies have arguedthat functional elements can be effectively annotated by their distinctpatterns of histone modifications.
TFs
Regulatory ac!vegene promoters and/or enhancers
Core promoter Core promoterNDR
GTFs
RNAPII RNAPII
(A)
(C)
(B)(i) Silent state
Enhancer
RNAPII
Transcrip!on
Gene promoter
(ii) S!mulus-induced enhancer ac!vity
Enhancer
Enhancement
Gene promoter
(iii) Lagged gene ac!va!on
(i) (ii)
Enhancer
Enhancement
Gene promoter
Low
RNAPIItranscrip!on
Low abundanceof factors
Inac!ve regulatoryelement
Ac!ve regulatory elementPromoter strength and/or transcrip!onal level
High
High abundanceof factors
TRENDS in Genetics
Figure 1. Active regulatory elements are divergently transcribed. (A) Both regulatory active gene promoters and gene-distal enhancers are transcribed. RNA polymerase II(RNAPII) recruitment and transcription initiation are mediated by general transcription factors (GTFs) binding core promoter regions in close proximity to flankingnucleosomes. This is facilitated by transcription factors (TFs), which often bind proximal to core promoters. Transcription often initiates divergently and at the boundary ofthe nucleosome-depleted region (NDR). (B) Gene expression is often preceded by, or changes concurrently with, changes in enhancer transcription. In a silent(nonexpressed) state (i), enhancers and promoters may, or may not, bind RNAPII. Upon stimulus (ii), transcriptional activity at enhancers marks regulatory enhancer activitywith local transcription and increases in RNAPII recruitment at the target gene promoter. (iii) Gene expression may lag behind transcriptional activation at enhancers. (C)Chromatin interactions place regulatory elements in close physical proximity. The individual properties of regulatory elements (chromatin characteristics as well as TF andRNAPII recruitment strengths) as well as context-dependent properties (such as promoter competition, insulation, and core promoter specificity) jointly determine theformation of multiple regulatory interactions (Box 1). Via regulatory cooperation, multiple regulatory elements may increase the local concentration of factors (TFs, GTFs,co-activators, and RNAPII) needed for transcription in RNAPII-enriched foci (i) and thereby achieve in aggregate different levels of transcriptional activity compared withRNAPII foci, including fewer regulatory elements (ii). Nucleosome illustrations in (A) reproduced, with permission, from [38]; (C) modified, with permission, from [38].
Opinion Trends in Genetics August 2015, Vol. 31, No. 8
428
13
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer potentialIf active enhancers have promoter activities, it follows thatwhat we classically refer to as promoters could also func-tion as enhancers in certain situations. Although in gener-al a neglected topic of research, we find some clues tosupport this in studies of larger chromatin architectures.
Regulatory elements often interact in close physicalproximity in RNAPII foci that are sometimes referred toas transcription factories [37], and the activity of eachregulatory element within these foci can be influencedby other regulatory elements involved (Figure 1C). Al-though the regulatory mechanisms remain poorly charac-terized, a common model posits that regulatory sequencessynergistically achieve high levels of transcriptional activ-ity by increasing the local concentration of factors neededfor transcription [3741] (Figure 1C). Studies based onchromatin conformation capture have observed RNAPIIfoci involving multiple gene promoters, with few or noenhancers [29,42]. In such arrangements, the degree towhich each gene promoter acts as a promoter or enhancermay depend on the local context of chromatin [42],expressed TFs, and sequence context (e.g., the occurrenceand strength of core promoter elements and TF bindingsites). Using in vitro reporter assays, Li et al. found thatcertain gene promoters in such constellations in MCF7cells had enhancer potential [42], suggesting that they
Box 2. Regulatory elements and associated chromatin
modification states
The N-terminal tails of histone proteins H2A, H2B, H3, and H4 withinnucleosomes can be chemically modified. In particular, amino acidresidues can be methylated or acetylated, which correlates withfunctional properties of the chromatin. For instance, H3K27ac (acet-ylation of lysine 27 in histone H3) is correlated with transcriptionalactivity. The locations of histones marked with a certain histonemodification can be measured genome wide using chromatin im-munoprecipitation (ChIP) coupled with either tiling microarrays(ChIP-chip) or sequencing (ChIP-seq) [38]. Profiling of histone mod-ifications in different cell types has enabled systematic inference ofdifferent types of functional genomic entities [12,53,77,78]. Suchstudies have shown that, typically, the nucleosomes flanking activegene promoters are enriched in H3K27ac, H3K4me3 (trimethylationof lysine 4 in histone H3), and, to some degree, H3K4me1 (mono-methylation of lysine 4 in histone H3), whereas enhancers are en-riched in H3K27ac and H3K4me1, but not H3K4me3, or relatively lowlevels thereof. In addition, several marks have been found to localizeat both enhancers and promoters. Most notably, H3K27ac marksactive promoters and a subset of enhancers that appear to be activelyregulating gene transcription, leading to the notion of classifyingregulatory elements into active and poised classes [15,79]. Poisedregulatory elements sometimes carry bivalent marks associated withactivity and repression at the same time, for instance promotershaving H3K4me3 and H3K27me3 (the latter mark is usually found atinactive gene promoters). Collectively, these studies have arguedthat functional elements can be effectively annotated by their distinctpatterns of histone modifications.
TFs
Regulatory ac!vegene promoters and/or enhancers
Core promoter Core promoterNDR
GTFs
RNAPII RNAPII
(A)
(C)
(B)(i) Silent state
Enhancer
RNAPII
Transcrip!on
Gene promoter
(ii) S!mulus-induced enhancer ac!vity
Enhancer
Enhancement
Gene promoter
(iii) Lagged gene ac!va!on
(i) (ii)
Enhancer
Enhancement
Gene promoter
Low
RNAPIItranscrip!on
Low abundanceof factors
Inac!ve regulatoryelement
Ac!ve regulatory elementPromoter strength and/or transcrip!onal level
High
High abundanceof factors
TRENDS in Genetics
Figure 1. Active regulatory elements are divergently transcribed. (A) Both regulatory active gene promoters and gene-distal enhancers are transcribed. RNA polymerase II(RNAPII) recruitment and transcription initiation are mediated by general transcription factors (GTFs) binding core promoter regions in close proximity to flankingnucleosomes. This is facilitated by transcription factors (TFs), which often bind proximal to core promoters. Transcription often initiates divergently and at the boundary ofthe nucleosome-depleted region (NDR). (B) Gene expression is often preceded by, or changes concurrently with, changes in enhancer transcription. In a silent(nonexpressed) state (i), enhancers and promoters may, or may not, bind RNAPII. Upon stimulus (ii), transcriptional activity at enhancers marks regulatory enhancer activitywith local transcription and increases in RNAPII recruitment at the target gene promoter. (iii) Gene expression may lag behind transcriptional activation at enhancers. (C)Chromatin interactions place regulatory elements in close physical proximity. The individual properties of regulatory elements (chromatin characteristics as well as TF andRNAPII recruitment strengths) as well as context-dependent properties (such as promoter competition, insulation, and core promoter specificity) jointly determine theformation of multiple regulatory interactions (Box 1). Via regulatory cooperation, multiple regulatory elements may increase the local concentration of factors (TFs, GTFs,co-activators, and RNAPII) needed for transcription in RNAPII-enriched foci (i) and thereby achieve in aggregate different levels of transcriptional activity compared withRNAPII foci, including fewer regulatory elements (ii). Nucleosome illustrations in (A) reproduced, with permission, from [38]; (C) modified, with permission, from [38].
Opinion Trends in Genetics August 2015, Vol. 31, No. 8
428
14
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer potentialIf active enhancers have promoter activities, it follows thatwhat we classically refer to as promoters could also func-tion as enhancers in certain situations. Although in gener-al a neglected topic of research, we find some clues tosupport this in studies of larger chromatin architectures.
Regulatory elements often interact in close physicalproximity in RNAPII foci that are sometimes referred toas transcription factories [37], and the activity of eachregulatory element within these foci can be influencedby other regulatory elements involved (Figure 1C). Al-though the regulatory mechanisms remain poorly charac-terized, a common model posits that regulatory sequencessynergistically achieve high levels of transcriptional activ-ity by increasing the local concentration of factors neededfor transcription [3741] (Figure 1C). Studies based onchromatin conformation capture have observed RNAPIIfoci involving multiple gene promoters, with few or noenhancers [29,42]. In such arrangements, the degree towhich each gene promoter acts as a promoter or enhancermay depend on the local context of chromatin [42],expressed TFs, and sequence context (e.g., the occurrenceand strength of core promoter elements and TF bindingsites). Using in vitro reporter assays, Li et al. found thatcertain gene promoters in such constellations in MCF7cells had enhancer potential [42], suggesting that they
Box 2. Regulatory elements and associated chromatin
modification states
The N-terminal tails of histone proteins H2A, H2B, H3, and H4 withinnucleosomes can be chemically modified. In particular, amino acidresidues can be methylated or acetylated, which correlates withfunctional properties of the chromatin. For instance, H3K27ac (acet-ylation of lysine 27 in histone H3) is correlated with transcriptionalactivity. The locations of histones marked with a certain histonemodification can be measured genome wide using chromatin im-munoprecipitation (ChIP) coupled with either tiling microarrays(ChIP-chip) or sequencing (ChIP-seq) [38]. Profiling of histone mod-ifications in different cell types has enabled systematic inference ofdifferent types of functional genomic entities [12,53,77,78]. Suchstudies have shown that, typically, the nucleosomes flanking activegene promoters are enriched in H3K27ac, H3K4me3 (trimethylationof lysine 4 in histone H3), and, to some degree, H3K4me1 (mono-methylation of lysine 4 in histone H3), whereas enhancers are en-riched in H3K27ac and H3K4me1, but not H3K4me3, or relatively lowlevels thereof. In addition, several marks have been found to localizeat both enhancers and promoters. Most notably, H3K27ac marksactive promoters and a subset of enhancers that appear to be activelyregulating gene transcription, leading to the notion of classifyingregulatory elements into active and poised classes [15,79]. Poisedregulatory elements sometimes carry bivalent marks associated withactivity and repression at the same time, for instance promotershaving H3K4me3 and H3K27me3 (the latter mark is usually found atinactive gene promoters). Collectively, these studies have arguedthat functional elements can be effectively annotated by their distinctpatterns of histone modifications.
TFs
Regulatory ac!vegene promoters and/or enhancers
Core promoter Core promoterNDR
GTFs
RNAPII RNAPII
(A)
(C)
(B)(i) Silent state
Enhancer
RNAPII
Transcrip!on
Gene promoter
(ii) S!mulus-induced enhancer ac!vity
Enhancer
Enhancement
Gene promoter
(iii) Lagged gene ac!va!on
(i) (ii)
Enhancer
Enhancement
Gene promoter
Low
RNAPIItranscrip!on
Low abundanceof factors
Inac!ve regulatoryelement
Ac!ve regulatory elementPromoter strength and/or transcrip!onal level
High
High abundanceof factors
TRENDS in Genetics
Figure 1. Active regulatory elements are divergently transcribed. (A) Both regulatory active gene promoters and gene-distal enhancers are transcribed. RNA polymerase II(RNAPII) recruitment and transcription initiation are mediated by general transcription factors (GTFs) binding core promoter regions in close proximity to flankingnucleosomes. This is facilitated by transcription factors (TFs), which often bind proximal to core promoters. Transcription often initiates divergently and at the boundary ofthe nucleosome-depleted region (NDR). (B) Gene expression is often preceded by, or changes concurrently with, changes in enhancer transcription. In a silent(nonexpressed) state (i), enhancers and promoters may, or may not, bind RNAPII. Upon stimulus (ii), transcriptional activity at enhancers marks regulatory enhancer activitywith local transcription and increases in RNAPII recruitment at the target gene promoter. (iii) Gene expression may lag behind transcriptional activation at enhancers. (C)Chromatin interactions place regulatory elements in close physical proximity. The individual properties of regulatory elements (chromatin characteristics as well as TF andRNAPII recruitment strengths) as well as context-dependent properties (such as promoter competition, insulation, and core promoter specificity) jointly determine theformation of multiple regulatory interactions (Box 1). Via regulatory cooperation, multiple regulatory elements may increase the local concentration of factors (TFs, GTFs,co-activators, and RNAPII) needed for transcription in RNAPII-enriched foci (i) and thereby achieve in aggregate different levels of transcriptional activity compared withRNAPII foci, including fewer regulatory elements (ii). Nucleosome illustrations in (A) reproduced, with permission, from [38]; (C) modified, with permission, from [38].
Opinion Trends in Genetics August 2015, Vol. 31, No. 8
428
15
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer potentialIf active enhancers have promoter activities, it follows thatwhat we classically refer to as promoters could also func-tion as enhancers in certain situations. Although in gener-al a neglected topic of research, we find some clues tosupport this in studies of larger chromatin architectures.
Regulatory elements often interact in close physicalproximity in RNAPII foci that are sometimes referred toas transcription factories [37], and the activity of eachregulatory element within these foci can be influencedby other regulatory elements involved (Figure 1C). Al-though the regulatory mechanisms remain poorly charac-terized, a common model posits that regulatory sequencessynergistically achieve high levels of transcriptional activ-ity by increasing the local concentration of factors neededfor transcription [3741] (Figure 1C). Studies based onchromatin conformation capture have observed RNAPIIfoci involving multiple gene promoters, with few or noenhancers [29,42]. In such arrangements, the degree towhich each gene promoter acts as a promoter or enhancermay depend on the local context of chromatin [42],expressed TFs, and sequence context (e.g., the occurrenceand strength of core promoter elements and TF bindingsites). Using in vitro reporter assays, Li et al. found thatcertain gene promoters in such constellations in MCF7cells had enhancer potential [42], suggesting that they
Box 2. Regulatory elements and associated chromatin
modification states
The N-terminal tails of histone proteins H2A, H2B, H3, and H4 withinnucleosomes can be chemically modified. In particular, amino acidresidues can be methylated or acetylated, which correlates withfunctional properties of the chromatin. For instance, H3K27ac (acet-ylation of lysine 27 in histone H3) is correlated with transcriptionalactivity. The locations of histones marked with a certain histonemodification can be measured genome wide using chromatin im-munoprecipitation (ChIP) coupled with either tiling microarrays(ChIP-chip) or sequencing (ChIP-seq) [38]. Profiling of histone mod-ifications in different cell types has enabled systematic inference ofdifferent types of functional genomic entities [12,53,77,78]. Suchstudies have shown that, typically, the nucleosomes flanking activegene promoters are enriched in H3K27ac, H3K4me3 (trimethylationof lysine 4 in histone H3), and, to some degree, H3K4me1 (mono-methylation of lysine 4 in histone H3), whereas enhancers are en-riched in H3K27ac and H3K4me1, but not H3K4me3, or relatively lowlevels thereof. In addition, several marks have been found to localizeat both enhancers and promoters. Most notably, H3K27ac marksactive promoters and a subset of enhancers that appear to be activelyregulating gene transcription, leading to the notion of classifyingregulatory elements into active and poised classes [15,79]. Poisedregulatory elements sometimes carry bivalent marks associated withactivity and repression at the same time, for instance promotershaving H3K4me3 and H3K27me3 (the latter mark is usually found atinactive gene promoters). Collectively, these studies have arguedthat functional elements can be effectively annotated by their distinctpatterns of histone modifications.
TFs
Regulatory ac!vegene promoters and/or enhancers
Core promoter Core promoterNDR
GTFs
RNAPII RNAPII
(A)
(C)
(B)(i) Silent state
Enhancer
RNAPII
Transcrip!on
Gene promoter
(ii) S!mulus-induced enhancer ac!vity
Enhancer
Enhancement
Gene promoter
(iii) Lagged gene ac!va!on
(i) (ii)
Enhancer
Enhancement
Gene promoter
Low
RNAPIItranscrip!on
Low abundanceof factors
Inac!ve regulatoryelement
Ac!ve regulatory elementPromoter strength and/or transcrip!onal level
High
High abundanceof factors
TRENDS in Genetics
Figure 1. Active regulatory elements are divergently transcribed. (A) Both regulatory active gene promoters and gene-distal enhancers are transcribed. RNA polymerase II(RNAPII) recruitment and transcription initiation are mediated by general transcription factors (GTFs) binding core promoter regions in close proximity to flankingnucleosomes. This is facilitated by transcription factors (TFs), which often bind proximal to core promoters. Transcription often initiates divergently and at the boundary ofthe nucleosome-depleted region (NDR). (B) Gene expression is often preceded by, or changes concurrently with, changes in enhancer transcription. In a silent(nonexpressed) state (i), enhancers and promoters may, or may not, bind RNAPII. Upon stimulus (ii), transcriptional activity at enhancers marks regulatory enhancer activitywith local transcription and increases in RNAPII recruitment at the target gene promoter. (iii) Gene expression may lag behind transcriptional activation at enhancers. (C)Chromatin interactions place regulatory elements in close physical proximity. The individual properties of regulatory elements (chromatin characteristics as well as TF andRNAPII recruitment strengths) as well as context-dependent properties (such as promoter competition, insulation, and core promoter specificity) jointly determine theformation of multiple regulatory interactions (Box 1). Via regulatory cooperation, multiple regulatory elements may increase the local concentration of factors (TFs, GTFs,co-activators, and RNAPII) needed for transcription in RNAPII-enriched foci (i) and thereby achieve in aggregate different levels of transcriptional activity compared withRNAPII foci, including fewer regulatory elements (ii). Nucleosome illustrations in (A) reproduced, with permission, from [38]; (C) modified, with permission, from [38].
Opinion Trends in Genetics August 2015, Vol. 31, No. 8
428
16
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer potentialIf active enhancers have promoter activities, it follows thatwhat we classically refer to as promoters could also func-tion as enhancers in certain situations. Although in gener-al a neglected topic of research, we find some clues tosupport this in studies of larger chromatin architectures.
Regulatory elements often interact in close physicalproximity in RNAPII foci that are sometimes referred toas transcription factories [37], and the activity of eachregulatory element within these foci can be influencedby other regulatory elements involved (Figure 1C). Al-though the regulatory mechanisms remain poorly charac-terized, a common model posits that regulatory sequencessynergistically achieve high levels of transcriptional activ-ity by increasing the local concentration of factors neededfor transcription [3741] (Figure 1C). Studies based onchromatin conformation capture have observed RNAPIIfoci involving multiple gene promoters, with few or noenhancers [29,42]. In such arrangements, the degree towhich each gene promoter acts as a promoter or enhancermay depend on the local context of chromatin [42],expressed TFs, and sequence context (e.g., the occurrenceand strength of core promoter elements and TF bindingsites). Using in vitro reporter assays, Li et al. found thatcertain gene promoters in such constellations in MCF7cells had enhancer potential [42], suggesting that they
Box 2. Regulatory elements and associated chromatin
modification states
The N-terminal tails of histone proteins H2A, H2B, H3, and H4 withinnucleosomes can be chemically modified. In particular, amino acidresidues can be methylated or acetylated, which correlates withfunctional properties of the chromatin. For instance, H3K27ac (acet-ylation of lysine 27 in histone H3) is correlated with transcriptionalactivity. The locations of histones marked with a certain histonemodification can be measured genome wide using chromatin im-munoprecipitation (ChIP) coupled with either tiling microarrays(ChIP-chip) or sequencing (ChIP-seq) [38]. Profiling of histone mod-ifications in different cell types has enabled systematic inference ofdifferent types of functional genomic entities [12,53,77,78]. Suchstudies have shown that, typically, the nucleosomes flanking activegene promoters are enriched in H3K27ac, H3K4me3 (trimethylationof lysine 4 in histone H3), and, to some degree, H3K4me1 (mono-methylation of lysine 4 in histone H3), whereas enhancers are en-riched in H3K27ac and H3K4me1, but not H3K4me3, or relatively lowlevels thereof. In addition, several marks have been found to localizeat both enhancers and promoters. Most notably, H3K27ac marksactive promoters and a subset of enhancers that appear to be activelyregulating gene transcription, leading to the notion of classifyingregulatory elements into active and poised classes [15,79]. Poisedregulatory elements sometimes carry bivalent marks associated withactivity and repression at the same time, for instance promotershaving H3K4me3 and H3K27me3 (the latter mark is usually found atinactive gene promoters). Collectively, these studies have arguedthat functional elements can be effectively annotated by their distinctpatterns of histone modifications.
TFs
Regulatory ac!vegene promoters and/or enhancers
Core promoter Core promoterNDR
GTFs
RNAPII RNAPII
(A)
(C)
(B)(i) Silent state
Enhancer
RNAPII
Transcrip!on
Gene promoter
(ii) S!mulus-induced enhancer ac!vity
Enhancer
Enhancement
Gene promoter
(iii) Lagged gene ac!va!on
(i) (ii)
Enhancer
Enhancement
Gene promoter
Low
RNAPIItranscrip!on
Low abundanceof factors
Inac!ve regulatoryelement
Ac!ve regulatory elementPromoter strength and/or transcrip!onal level
High
High abundanceof factors
TRENDS in Genetics
Figure 1. Active regulatory elements are divergently transcribed. (A) Both regulatory active gene promoters and gene-distal enhancers are transcribed. RNA polymerase II(RNAPII) recruitment and transcription initiation are mediated by general transcription factors (GTFs) binding core promoter regions in close proximity to flankingnucleosomes. This is facilitated by transcription factors (TFs), which often bind proximal to core promoters. Transcription often initiates divergently and at the boundary ofthe nucleosome-depleted region (NDR). (B) Gene expression is often preceded by, or changes concurrently with, changes in enhancer transcription. In a silent(nonexpressed) state (i), enhancers and promoters may, or may not, bind RNAPII. Upon stimulus (ii), transcriptional activity at enhancers marks regulatory enhancer activitywith local transcription and increases in RNAPII recruitment at the target gene promoter. (iii) Gene expression may lag behind transcriptional activation at enhancers. (C)Chromatin interactions place regulatory elements in close physical proximity. The individual properties of regulatory elements (chromatin characteristics as well as TF andRNAPII recruitment strengths) as well as context-dependent properties (such as promoter competition, insulation, and core promoter specificity) jointly determine theformation of multiple regulatory interactions (Box 1). Via regulatory cooperation, multiple regulatory elements may increase the local concentration of factors (TFs, GTFs,co-activators, and RNAPII) needed for transcription in RNAPII-enriched foci (i) and thereby achieve in aggregate different levels of transcriptional activity compared withRNAPII foci, including fewer regulatory elements (ii). Nucleosome illustrations in (A) reproduced, with permission, from [38]; (C) modified, with permission, from [38].
Opinion Trends in Genetics August 2015, Vol. 31, No. 8
428
Stormo,G.D.etal.NucleicAcidsResearch(1982)
17
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer potentialIf active enhancers have promoter activities, it follows thatwhat we classically refer to as promoters could also func-tion as enhancers in certain situations. Although in gener-al a neglected topic of research, we find some clues tosupport this in studies of larger chromatin architectures.
Regulatory elements often interact in close physicalproximity in RNAPII foci that are sometimes referred toas transcription factories [37], and the activity of eachregulatory element within these foci can be influencedby other regulatory elements involved (Figure 1C). Al-though the regulatory mechanisms remain poorly charac-terized, a common model posits that regulatory sequencessynergistically achieve high levels of transcriptional activ-ity by increasing the local concentration of factors neededfor transcription [3741] (Figure 1C). Studies based onchromatin conformation capture have observed RNAPIIfoci involving multiple gene promoters, with few or noenhancers [29,42]. In such arrangements, the degree towhich each gene promoter acts as a promoter or enhancermay depend on the local context of chromatin [42],expressed TFs, and sequence context (e.g., the occurrenceand strength of core promoter elements and TF bindingsites). Using in vitro reporter assays, Li et al. found thatcertain gene promoters in such constellations in MCF7cells had enhancer potential [42], suggesting that they
Box 2. Regulatory elements and associated chromatin
modification states
The N-terminal tails of histone proteins H2A, H2B, H3, and H4 withinnucleosomes can be chemically modified. In particular, amino acidresidues can be methylated or acetylated, which correlates withfunctional properties of the chromatin. For instance, H3K27ac (acet-ylation of lysine 27 in histone H3) is correlated with transcriptionalactivity. The locations of histones marked with a certain histonemodification can be measured genome wide using chromatin im-munoprecipitation (ChIP) coupled with either tiling microarrays(ChIP-chip) or sequencing (ChIP-seq) [38]. Profiling of histone mod-ifications in different cell types has enabled systematic inference ofdifferent types of functional genomic entities [12,53,77,78]. Suchstudies have shown that, typically, the nucleosomes flanking activegene promoters are enriched in H3K27ac, H3K4me3 (trimethylationof lysine 4 in histone H3), and, to some degree, H3K4me1 (mono-methylation of lysine 4 in histone H3), whereas enhancers are en-riched in H3K27ac and H3K4me1, but not H3K4me3, or relatively lowlevels thereof. In addition, several marks have been found to localizeat both enhancers and promoters. Most notably, H3K27ac marksactive promoters and a subset of enhancers that appear to be activelyregulating gene transcription, leading to the notion of classifyingregulatory elements into active and poised classes [15,79]. Poisedregulatory elements sometimes carry bivalent marks associated withactivity and repression at the same time, for instance promotershaving H3K4me3 and H3K27me3 (the latter mark is usually found atinactive gene promoters). Collectively, these studies have arguedthat functional elements can be effectively annotated by their distinctpatterns of histone modifications.
TFs
Regulatory ac!vegene promoters and/or enhancers
Core promoter Core promoterNDR
GTFs
RNAPII RNAPII
(A)
(C)
(B)(i) Silent state
Enhancer
RNAPII
Transcrip!on
Gene promoter
(ii) S!mulus-induced enhancer ac!vity
Enhancer
Enhancement
Gene promoter
(iii) Lagged gene ac!va!on
(i) (ii)
Enhancer
Enhancement
Gene promoter
Low
RNAPIItranscrip!on
Low abundanceof factors
Inac!ve regulatoryelement
Ac!ve regulatory elementPromoter strength and/or transcrip!onal level
High
High abundanceof factors
TRENDS in Genetics
Figure 1. Active regulatory elements are divergently transcribed. (A) Both regulatory active gene promoters and gene-distal enhancers are transcribed. RNA polymerase II(RNAPII) recruitment and transcription initiation are mediated by general transcription factors (GTFs) binding core promoter regions in close proximity to flankingnucleosomes. This is facilitated by transcription factors (TFs), which often bind proximal to core promoters. Transcription often initiates divergently and at the boundary ofthe nucleosome-depleted region (NDR). (B) Gene expression is often preceded by, or changes concurrently with, changes in enhancer transcription. In a silent(nonexpressed) state (i), enhancers and promoters may, or may not, bind RNAPII. Upon stimulus (ii), transcriptional activity at enhancers marks regulatory enhancer activitywith local transcription and increases in RNAPII recruitment at the target gene promoter. (iii) Gene expression may lag behind transcriptional activation at enhancers. (C)Chromatin interactions place regulatory elements in close physical proximity. The individual properties of regulatory elements (chromatin characteristics as well as TF andRNAPII recruitment strengths) as well as context-dependent properties (such as promoter competition, insulation, and core promoter specificity) jointly determine theformation of multiple regulatory interactions (Box 1). Via regulatory cooperation, multiple regulatory elements may increase the local concentration of factors (TFs, GTFs,co-activators, and RNAPII) needed for transcription in RNAPII-enriched foci (i) and thereby achieve in aggregate different levels of transcriptional activity compared withRNAPII foci, including fewer regulatory elements (ii). Nucleosome illustrations in (A) reproduced, with permission, from [38]; (C) modified, with permission, from [38].
Opinion Trends in Genetics August 2015, Vol. 31, No. 8
428
Stormo,G.D.etal.NucleicAcidsResearch(1982)
ChIP-exoAn extension of chromatin immunoprecipitation followed by sequencing (ChIPseq) that includes exonuclease trimming after ChIP to increase the resolution of the mapped transcription factor bound sites.
regulating transcription to establish cell-lineage-specific programmes. Experiments usingthe ChIP-exo technique uncovered a 52-bp CTCF-binding motif that contains four CTCF-binding modules15,16 (FIG.1).
The presence of CpGs in the DNA consensus sequence of the CTCF-binding site supports the notion that methylation of cytosine residues at carbon 5 of the base to form 5-methylcytosine (5mC) in CpG-containing sites may, at least partly, underlie CTCF tar-get selectivity in different cell types17. Recent studies indicate that DNA methylation has a widespread role in regulating CTCF occupancy at many genes, includ-ing CDKN2A (which encodes INK4A and ARF)18, B-cell CLL/lymphoma 6 (BCL6)19 and brain-derived neurotrophic factor (BDNF)20. One study has mapped the occupancy of CTCF in 19 human cell types; by comparing this information with DNA methylation data from parallel reduced representation bisulphite sequencing, it was found that 41% of cell-type-specific CTCF-binding sites are linked to differential DNA methylation21 (FIG.2). Conversely, at 67% of sites that showed variability in DNA methylation, the presence of 5mC was associated with a concomitant downregula-tion of cell-type-specific CTCF occupancy. CTCF can also affect the methylation status of DNA by forming a complex with poly(ADP-ribose) polymerase 1 (PARP1) and DNA (cytosine-5)-methyltransferase 1 (DNMT1). CTCF activates PARP1, which can then inacti-vate DNMT1 by poly(ADP-ribosyl)ation, and thus
maintains methyl-free CpGs in the DNA22,23. An addi-tional level of complexity in the interaction between CTCF and its target sequence can arise from the oxida-tion of 5mC to 5-hydroxymethylcytosine (5hmC)24,25, 5-formylcytosine (5fC) and 5-carboxylcytosine (5caC)26 by ten-eleven translocation (TET) enzymes. Genome-wide profiling analyses of 5hmC have shown that this modification and, to a lesser extent, 5fC are enriched genomic locations that contain CTCF-binding sites27,28. Furthermore, identification of proteins that bind to different oxidized derivatives of 5mC discov-ered CTCF as a 5caC-specific binder29. These results underscore the complexity and possible importance of the relationship between DNA methylation status and plasticity of CTCF occupancy. However, the presence of cell-type-specific CTCF-binding sites that are not differentially methylated suggests the existence of other mechanisms by which the DNA occupancy of this protein is regulated (FIG.2).
One such mechanism is post-translational cova-lent modification of CTCF, such as sumolyation30 and poly(ADP-ribosy)lation31. In breast cancer cells, defec-tive poly(ADP-ribosyl)ation of CTCF leads to its dissoci-ation from the CDKN2A locus, which results in aberrant silencing of this tumour suppressor gene32. In Drosophila melanogaster, poly(ADP-ribosy)lation of Centrosomal protein 190 kDa (Cp190) and CTCF facilitates their interaction, tethering to the nuclear matrix and intrachromosomal contacts33.
Figure 1 | Features of CTCF-binding sites in the genome. Binding sites of CCCTC-binding factor (CTCF) are associated with different genetic elements. The majority of these sites are intergenic and colocalize with cohesin. In
genes and short interspersed nuclear elements (SINEs)) and extra-TFIIIC (ETC) loci, which suggests that TFIIIC and CTCF
cooperate in some nuclear processes. The 12-bp consensus sequence of CTCF-binding sites is embedded within binding
modules 2 and 3 as determined by ChIP-exo experiments. DNA methylation (represented by red circles) of cytosine
residues occurs at positions 2 and 12 of the consensus sequence in a subset of CTCF-binding sites.
Nature Reviews | Genetics
TFIIIC
ETC locus
B box
Promoter
Cohesin
Pol III
Enhancer
Gene
Condensin
tRNA geneand SINE
CTCFCTCF
CTCF CTCF
Module 2Module 1 Module 3 Module 4
CTCF
ETC locus
REVIEWS
NATURE REVIEWS | GENETICS VOLUME 15 | APRIL 2014 | 235
2014 Macmillan Publishers Limited. All rights reserved
OngC-tandCorcesV.G.NatureReviewGenetics2014
18
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer potentialIf active enhancers have promoter activities, it follows thatwhat we classically refer to as promoters could also func-tion as enhancers in certain situations. Although in gener-al a neglected topic of research, we find some clues tosupport this in studies of larger chromatin architectures.
Regulatory elements often interact in close physicalproximity in RNAPII foci that are sometimes referred toas transcription factories [37], and the activity of eachregulatory element within these foci can be influencedby other regulatory elements involved (Figure 1C). Al-though the regulatory mechanisms remain poorly charac-terized, a common model posits that regulatory sequencessynergistically achieve high levels of transcriptional activ-ity by increasing the local concentration of factors neededfor transcription [3741] (Figure 1C). Studies based onchromatin conformation capture have observed RNAPIIfoci involving multiple gene promoters, with few or noenhancers [29,42]. In such arrangements, the degree towhich each gene promoter acts as a promoter or enhancermay depend on the local context of chromatin [42],expressed TFs, and sequence context (e.g., the occurrenceand strength of core promoter elements and TF bindingsites). Using in vitro reporter assays, Li et al. found thatcertain gene promoters in such constellations in MCF7cells had enhancer potential [42], suggesting that they
Box 2. Regulatory elements and associated chromatin
modification states
The N-terminal tails of histone proteins H2A, H2B, H3, and H4 withinnucleosomes can be chemically modified. In particular, amino acidresidues can be methylated or acetylated, which correlates withfunctional properties of the chromatin. For instance, H3K27ac (acet-ylation of lysine 27 in histone H3) is correlated with transcriptionalactivity. The locations of histones marked with a certain histonemodification can be measured genome wide using chromatin im-munoprecipitation (ChIP) coupled with either tiling microarrays(ChIP-chip) or sequencing (ChIP-seq) [38]. Profiling of histone mod-ifications in different cell types has enabled systematic inference ofdifferent types of functional genomic entities [12,53,77,78]. Suchstudies have shown that, typically, the nucleosomes flanking activegene promoters are enriched in H3K27ac, H3K4me3 (trimethylationof lysine 4 in histone H3), and, to some degree, H3K4me1 (mono-methylation of lysine 4 in histone H3), whereas enhancers are en-riched in H3K27ac and H3K4me1, but not H3K4me3, or relatively lowlevels thereof. In addition, several marks have been found to localizeat both enhancers and promoters. Most notably, H3K27ac marksactive promoters and a subset of enhancers that appear to be activelyregulating gene transcription, leading to the notion of classifyingregulatory elements into active and poised classes [15,79]. Poisedregulatory elements sometimes carry bivalent marks associated withactivity and repression at the same time, for instance promotershaving H3K4me3 and H3K27me3 (the latter mark is usually found atinactive gene promoters). Collectively, these studies have arguedthat functional elements can be effectively annotated by their distinctpatterns of histone modifications.
TFs
Regulatory ac!vegene promoters and/or enhancers
Core promoter Core promoterNDR
GTFs
RNAPII RNAPII
(A)
(C)
(B)(i) Silent state
Enhancer
RNAPII
Transcrip!on
Gene promoter
(ii) S!mulus-induced enhancer ac!vity
Enhancer
Enhancement
Gene promoter
(iii) Lagged gene ac!va!on
(i) (ii)
Enhancer
Enhancement
Gene promoter
Low
RNAPIItranscrip!on
Low abundanceof factors
Inac!ve regulatoryelement
Ac!ve regulatory elementPromoter strength and/or transcrip!onal level
High
High abundanceof factors
TRENDS in Genetics
Figure 1. Active regulatory elements are divergently transcribed. (A) Both regulatory active gene promoters and gene-distal enhancers are transcribed. RNA polymerase II(RNAPII) recruitment and transcription initiation are mediated by general transcription factors (GTFs) binding core promoter regions in close proximity to flankingnucleosomes. This is facilitated by transcription factors (TFs), which often bind proximal to core promoters. Transcription often initiates divergently and at the boundary ofthe nucleosome-depleted region (NDR). (B) Gene expression is often preceded by, or changes concurrently with, changes in enhancer transcription. In a silent(nonexpressed) state (i), enhancers and promoters may, or may not, bind RNAPII. Upon stimulus (ii), transcriptional activity at enhancers marks regulatory enhancer activitywith local transcription and increases in RNAPII recruitment at the target gene promoter. (iii) Gene expression may lag behind transcriptional activation at enhancers. (C)Chromatin interactions place regulatory elements in close physical proximity. The individual properties of regulatory elements (chromatin characteristics as well as TF andRNAPII recruitment strengths) as well as context-dependent properties (such as promoter competition, insulation, and core promoter specificity) jointly determine theformation of multiple regulatory interactions (Box 1). Via regulatory cooperation, multiple regulatory elements may increase the local concentration of factors (TFs, GTFs,co-activators, and RNAPII) needed for transcription in RNAPII-enriched foci (i) and thereby achieve in aggregate different levels of transcriptional activity compared withRNAPII foci, including fewer regulatory elements (ii). Nucleosome illustrations in (A) reproduced, with permission, from [38]; (C) modified, with permission, from [38].
Opinion Trends in Genetics August 2015, Vol. 31, No. 8
428
19
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer potentialIf active enhancers have promoter activities, it follows thatwhat we classically refer to as promoters could also func-tion as enhancers in certain situations. Although in gener-al a neglected topic of research, we find some clues tosupport this in studies of larger chromatin architectures.
Regulatory elements often interact in close physicalproximity in RNAPII foci that are sometimes referred toas transcription factories [37], and the activity of eachregulatory element within these foci can be influencedby other regulatory elements involved (Figure 1C). Al-though the regulatory mechanisms remain poorly charac-terized, a common model posits that regulatory sequencessynergistically achieve high levels of transcriptional activ-ity by increasing the local concentration of factors neededfor transcription [3741] (Figure 1C). Studies based onchromatin conformation capture have observed RNAPIIfoci involving multiple gene promoters, with few or noenhancers [29,42]. In such arrangements, the degree towhich each gene promoter acts as a promoter or enhancermay depend on the local context of chromatin [42],expressed TFs, and sequence context (e.g., the occurrenceand strength of core promoter elements and TF bindingsites). Using in vitro reporter assays, Li et al. found thatcertain gene promoters in such constellations in MCF7cells had enhancer potential [42], suggesting that they
Box 2. Regulatory elements and associated chromatin
modification states
The N-terminal tails of histone proteins H2A, H2B, H3, and H4 withinnucleosomes can be chemically modified. In particular, amino acidresidues can be methylated or acetylated, which correlates withfunctional properties of the chromatin. For instance, H3K27ac (acet-ylation of lysine 27 in histone H3) is correlated with transcriptionalactivity. The locations of histones marked with a certain histonemodification can be measured genome wide using chromatin im-munoprecipitation (ChIP) coupled with either tiling microarrays(ChIP-chip) or sequencing (ChIP-seq) [38]. Profiling of histone mod-ifications in different cell types has enabled systematic inference ofdifferent types of functional genomic entities [12,53,77,78]. Suchstudies have shown that, typically, the nucleosomes flanking activegene promoters are enriched in H3K27ac, H3K4me3 (trimethylationof lysine 4 in histone H3), and, to some degree, H3K4me1 (mono-methylation of lysine 4 in histone H3), whereas enhancers are en-riched in H3K27ac and H3K4me1, but not H3K4me3, or relatively lowlevels thereof. In addition, several marks have been found to localizeat both enhancers and promoters. Most notably, H3K27ac marksactive promoters and a subset of enhancers that appear to be activelyregulating gene transcription, leading to the notion of classifyingregulatory elements into active and poised classes [15,79]. Poisedregulatory elements sometimes carry bivalent marks associated withactivity and repression at the same time, for instance promotershaving H3K4me3 and H3K27me3 (the latter mark is usually found atinactive gene promoters). Collectively, these studies have arguedthat functional elements can be effectively annotated by their distinctpatterns of histone modifications.
TFs
Regulatory ac!vegene promoters and/or enhancers
Core promoter Core promoterNDR
GTFs
RNAPII RNAPII
(A)
(C)
(B)(i) Silent state
Enhancer
RNAPII
Transcrip!on
Gene promoter
(ii) S!mulus-induced enhancer ac!vity
Enhancer
Enhancement
Gene promoter
(iii) Lagged gene ac!va!on
(i) (ii)
Enhancer
Enhancement
Gene promoter
Low
RNAPIItranscrip!on
Low abundanceof factors
Inac!ve regulatoryelement
Ac!ve regulatory elementPromoter strength and/or transcrip!onal level
High
High abundanceof factors
TRENDS in Genetics
Figure 1. Active regulatory elements are divergently transcribed. (A) Both regulatory active gene promoters and gene-distal enhancers are transcribed. RNA polymerase II(RNAPII) recruitment and transcription initiation are mediated by general transcription factors (GTFs) binding core promoter regions in close proximity to flankingnucleosomes. This is facilitated by transcription factors (TFs), which often bind proximal to core promoters. Transcription often initiates divergently and at the boundary ofthe nucleosome-depleted region (NDR). (B) Gene expression is often preceded by, or changes concurrently with, changes in enhancer transcription. In a silent(nonexpressed) state (i), enhancers and promoters may, or may not, bind RNAPII. Upon stimulus (ii), transcriptional activity at enhancers marks regulatory enhancer activitywith local transcription and increases in RNAPII recruitment at the target gene promoter. (iii) Gene expression may lag behind transcriptional activation at enhancers. (C)Chromatin interactions place regulatory elements in close physical proximity. The individual properties of regulatory elements (chromatin characteristics as well as TF andRNAPII recruitment strengths) as well as context-dependent properties (such as promoter competition, insulation, and core promoter specificity) jointly determine theformation of multiple regulatory interactions (Box 1). Via regulatory cooperation, multiple regulatory elements may increase the local concentration of factors (TFs, GTFs,co-activators, and RNAPII) needed for transcription in RNAPII-enriched foci (i) and thereby achieve in aggregate different levels of transcriptional activity compared withRNAPII foci, including fewer regulatory elements (ii). Nucleosome illustrations in (A) reproduced, with permission, from [38]; (C) modified, with permission, from [38].
Opinion Trends in Genetics August 2015, Vol. 31, No. 8
428
Lindblad-Toh,K.etal.Nature(2011)
20
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer potentialIf active enhancers have promoter activities, it follows thatwhat we classically refer to as promoters could also func-tion as enhancers in certain situations. Although in gener-al a neglected topic of research, we find some clues tosupport this in studies of larger chromatin architectures.
Regulatory elements often interact in close physicalproximity in RNAPII foci that are sometimes referred toas transcription factories [37], and the activity of eachregulatory element within these foci can be influencedby other regulatory elements involved (Figure 1C). Al-though the regulatory mechanisms remain poorly charac-terized, a common model posits that regulatory sequencessynergistically achieve high levels of transcriptional activ-ity by increasing the local concentration of factors neededfor transcription [3741] (Figure 1C). Studies based onchromatin conformation capture have observed RNAPIIfoci involving multiple gene promoters, with few or noenhancers [29,42]. In such arrangements, the degree towhich each gene promoter acts as a promoter or enhancermay depend on the local context of chromatin [42],expressed TFs, and sequence context (e.g., the occurrenceand strength of core promoter elements and TF bindingsites). Using in vitro reporter assays, Li et al. found thatcertain gene promoters in such constellations in MCF7cells had enhancer potential [42], suggesting that they
Box 2. Regulatory elements and associated chromatin
modification states
The N-terminal tails of histone proteins H2A, H2B, H3, and H4 withinnucleosomes can be chemically modified. In particular, amino acidresidues can be methylated or acetylated, which correlates withfunctional properties of the chromatin. For instance, H3K27ac (acet-ylation of lysine 27 in histone H3) is correlated with transcriptionalactivity. The locations of histones marked with a certain histonemodification can be measured genome wide using chromatin im-munoprecipitation (ChIP) coupled with either tiling microarrays(ChIP-chip) or sequencing (ChIP-seq) [38]. Profiling of histone mod-ifications in different cell types has enabled systematic inference ofdifferent types of functional genomic entities [12,53,77,78]. Suchstudies have shown that, typically, the nucleosomes flanking activegene promoters are enriched in H3K27ac, H3K4me3 (trimethylationof lysine 4 in histone H3), and, to some degree, H3K4me1 (mono-methylation of lysine 4 in histone H3), whereas enhancers are en-riched in H3K27ac and H3K4me1, but not H3K4me3, or relatively lowlevels thereof. In addition, several marks have been found to localizeat both enhancers and promoters. Most notably, H3K27ac marksactive promoters and a subset of enhancers that appear to be activelyregulating gene transcription, leading to the notion of classifyingregulatory elements into active and poised classes [15,79]. Poisedregulatory elements sometimes carry bivalent marks associated withactivity and repression at the same time, for instance promotershaving H3K4me3 and H3K27me3 (the latter mark is usually found atinactive gene promoters). Collectively, these studies have arguedthat functional elements can be effectively annotated by their distinctpatterns of histone modifications.
TFs
Regulatory ac!vegene promoters and/or enhancers
Core promoter Core promoterNDR
GTFs
RNAPII RNAPII
(A)
(C)
(B)(i) Silent state
Enhancer
RNAPII
Transcrip!on
Gene promoter
(ii) S!mulus-induced enhancer ac!vity
Enhancer
Enhancement
Gene promoter
(iii) Lagged gene ac!va!on
(i) (ii)
Enhancer
Enhancement
Gene promoter
Low
RNAPIItranscrip!on
Low abundanceof factors
Inac!ve regulatoryelement
Ac!ve regulatory elementPromoter strength and/or transcrip!onal level
High
High abundanceof factors
TRENDS in Genetics
Figure 1. Active regulatory elements are divergently transcribed. (A) Both regulatory active gene promoters and gene-distal enhancers are transcribed. RNA polymerase II(RNAPII) recruitment and transcription initiation are mediated by general transcription factors (GTFs) binding core promoter regions in close proximity to flankingnucleosomes. This is facilitated by transcription factors (TFs), which often bind proximal to core promoters. Transcription often initiates divergently and at the boundary ofthe nucleosome-depleted region (NDR). (B) Gene expression is often preceded by, or changes concurrently with, changes in enhancer transcription. In a silent(nonexpressed) state (i), enhancers and promoters may, or may not, bind RNAPII. Upon stimulus (ii), transcriptional activity at enhancers marks regulatory enhancer activitywith local transcription and increases in RNAPII recruitment at the target gene promoter. (iii) Gene expression may lag behind transcriptional activation at enhancers. (C)Chromatin interactions place regulatory elements in close physical proximity. The individual properties of regulatory elements (chromatin characteristics as well as TF andRNAPII recruitment strengths) as well as context-dependent properties (such as promoter competition, insulation, and core promoter specificity) jointly determine theformation of multiple regulatory interactions (Box 1). Via regulatory cooperation, multiple regulatory elements may increase the local concentration of factors (TFs, GTFs,co-activators, and RNAPII) needed for transcription in RNAPII-enriched foci (i) and thereby achieve in aggregate different levels of transcriptional activity compared withRNAPII foci, including fewer regulatory elements (ii). Nucleosome illustrations in (A) reproduced, with permission, from [38]; (C) modified, with permission, from [38].
Opinion Trends in Genetics August 2015, Vol. 31, No. 8
428
Kim,T.H.andB.Ren,AnnuRevGenomicsHumGenet,2006
21
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer potentialIf active enhancers have promoter activities, it follows thatwhat we classically refer to as promoters could also func-tion as enhancers in certain situations. Although in gener-al a neglected topic of research, we find some clues tosupport this in studies of larger chromatin architectures.
Regulatory elements often interact in close physicalproximity in RNAPII foci that are sometimes referred toas transcription factories [37], and the activity of eachregulatory element within these foci can be influencedby other regulatory elements involved (Figure 1C). Al-though the regulatory mechanisms remain poorly charac-terized, a common model posits that regulatory sequencessynergistically achieve high levels of transcriptional activ-ity by increasing the local concentration of factors neededfor transcription [3741] (Figure 1C). Studies based onchromatin conformation capture have observed RNAPIIfoci involving multiple gene promoters, with few or noenhancers [29,42]. In such arrangements, the degree towhich each gene promoter acts as a promoter or enhancermay depend on the local context of chromatin [42],expressed TFs, and sequence context (e.g., the occurrenceand strength of core promoter elements and TF bindingsites). Using in vitro reporter assays, Li et al. found thatcertain gene promoters in such constellations in MCF7cells had enhancer potential [42], suggesting that they
Box 2. Regulatory elements and associated chromatin
modification states
The N-terminal tails of histone proteins H2A, H2B, H3, and H4 withinnucleosomes can be chemically modified. In particular, amino acidresidues can be methylated or acetylated, which correlates withfunctional properties of the chromatin. For instance, H3K27ac (acet-ylation of lysine 27 in histone H3) is correlated with transcriptionalactivity. The locations of histones marked with a certain histonemodification can be measured genome wide using chromatin im-munoprecipitation (ChIP) coupled with either tiling microarrays(ChIP-chip) or sequencing (ChIP-seq) [38]. Profiling of histone mod-ifications in different cell types has enabled systematic inference ofdifferent types of functional genomic entities [12,53,77,78]. Suchstudies have shown that, typically, the nucleosomes flanking activegene promoters are enriched in H3K27ac, H3K4me3 (trimethylationof lysine 4 in histone H3), and, to some degree, H3K4me1 (mono-methylation of lysine 4 in histone H3), whereas enhancers are en-riched in H3K27ac and H3K4me1, but not H3K4me3, or relatively lowlevels thereof. In addition, several marks have been found to localizeat both enhancers and promoters. Most notably, H3K27ac marksactive promoters and a subset of enhancers that appear to be activelyregulating gene transcription, leading to the notion of classifyingregulatory elements into active and poised classes [15,79]. Poisedregulatory elements sometimes carry bivalent marks associated withactivity and repression at the same time, for instance promotershaving H3K4me3 and H3K27me3 (the latter mark is usually found atinactive gene promoters). Collectively, these studies have arguedthat functional elements can be effectively annotated by their distinctpatterns of histone modifications.
TFs
Regulatory ac!vegene promoters and/or enhancers
Core promoter Core promoterNDR
GTFs
RNAPII RNAPII
(A)
(C)
(B)(i) Silent state
Enhancer
RNAPII
Transcrip!on
Gene promoter
(ii) S!mulus-induced enhancer ac!vity
Enhancer
Enhancement
Gene promoter
(iii) Lagged gene ac!va!on
(i) (ii)
Enhancer
Enhancement
Gene promoter
Low
RNAPIItranscrip!on
Low abundanceof factors
Inac!ve regulatoryelement
Ac!ve regulatory elementPromoter strength and/or transcrip!onal level
High
High abundanceof factors
TRENDS in Genetics
Figure 1. Active regulatory elements are divergently transcribed. (A) Both regulatory active gene promoters and gene-distal enhancers are transcribed. RNA polymerase II(RNAPII) recruitment and transcription initiation are mediated by general transcription factors (GTFs) binding core promoter regions in close proximity to flankingnucleosomes. This is facilitated by transcription factors (TFs), which often bind proximal to core promoters. Transcription often initiates divergently and at the boundary ofthe nucleosome-depleted region (NDR). (B) Gene expression is often preceded by, or changes concurrently with, changes in enhancer transcription. In a silent(nonexpressed) state (i), enhancers and promoters may, or may not, bind RNAPII. Upon stimulus (ii), transcriptional activity at enhancers marks regulatory enhancer activitywith local transcription and increases in RNAPII recruitment at the target gene promoter. (iii) Gene expression may lag behind transcriptional activation at enhancers. (C)Chromatin interactions place regulatory elements in close physical proximity. The individual properties of regulatory elements (chromatin characteristics as well as TF andRNAPII recruitment strengths) as well as context-dependent properties (such as promoter competition, insulation, and core promoter specificity) jointly determine theformation of multiple regulatory interactions (Box 1). Via regulatory cooperation, multiple regulatory elements may increase the local concentration of factors (TFs, GTFs,co-activators, and RNAPII) needed for transcription in RNAPII-enriched foci (i) and thereby achieve in aggregate different levels of transcriptional activity compared withRNAPII foci, including fewer regulatory elements (ii). Nucleosome illustrations in (A) reproduced, with permission, from [38]; (C) modified, with permission, from [38].
Opinion Trends in Genetics August 2015, Vol. 31, No. 8
428
Kim,T.H.andB.Ren,AnnuRevGenomicsHumGenet,2006
SegalE,Nature2006
22
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer potentialIf active enhancers have promoter activities, it follows thatwhat we classically refer to as promoters could also func-tion as enhancers in certain situations. Although in gener-al a neglected topic of research, we find some clues tosupport this in studies of larger chromatin architectures.
Regulatory elements often interact in close physicalproximity in RNAPII foci that are sometimes referred toas transcription factories [37], and the activity of eachregulatory element within these foci can be influencedby other regulatory elements involved (Figure 1C). Al-though the regulatory mechanisms remain poorly charac-terized, a common model posits that regulatory sequencessynergistically achieve high levels of transcriptional activ-ity by increasing the local concentration of factors neededfor transcription [3741] (Figure 1C). Studies based onchromatin conformation capture have observed RNAPIIfoci involving multiple gene promoters, with few or noenhancers [29,42]. In such arrangements, the degree towhich each gene promoter acts as a promoter or enhancermay depend on the local context of chromatin [42],expressed TFs, and sequence context (e.g., the occurrenceand strength of core promoter elements and TF bindingsites). Using in vitro reporter assays, Li et al. found thatcertain gene promoters in such constellations in MCF7cells had enhancer potential [42], suggesting that they
Box 2. Regulatory elements and associated chromatin
modification states
The N-terminal tails of histone proteins H2A, H2B, H3, and H4 withinnucleosomes can be chemically modified. In particular, amino acidresidues can be methylated or acetylated, which correlates withfunctional properties of the chromatin. For instance, H3K27ac (acet-ylation of lysine 27 in histone H3) is correlated with transcriptionalactivity. The locations of histones marked with a certain histonemodification can be measured genome wide using chromatin im-munoprecipitation (ChIP) coupled with either tiling microarrays(ChIP-chip) or sequencing (ChIP-seq) [38]. Profiling of histone mod-ifications in different cell types has enabled systematic inference ofdifferent types of functional genomic entities [12,53,77,78]. Suchstudies have shown that, typically, the nucleosomes flanking activegene promoters are enriched in H3K27ac, H3K4me3 (trimethylationof lysine 4 in histone H3), and, to some degree, H3K4me1 (mono-methylation of lysine 4 in histone H3), whereas enhancers are en-riched in H3K27ac and H3K4me1, but not H3K4me3, or relatively lowlevels thereof. In addition, several marks have been found to localizeat both enhancers and promoters. Most notably, H3K27ac marksactive promoters and a subset of enhancers that appear to be activelyregulating gene transcription, leading to the notion of classifyingregulatory elements into active and poised classes [15,79]. Poisedregulatory elements sometimes carry bivalent marks associated withactivity and repression at the same time, for instance promotershaving H3K4me3 and H3K27me3 (the latter mark is usually found atinactive gene promoters). Collectively, these studies have arguedthat functional elements can be effectively annotated by their distinctpatterns of histone modifications.
TFs
Regulatory ac!vegene promoters and/or enhancers
Core promoter Core promoterNDR
GTFs
RNAPII RNAPII
(A)
(C)
(B)(i) Silent state
Enhancer
RNAPII
Transcrip!on
Gene promoter
(ii) S!mulus-induced enhancer ac!vity
Enhancer
Enhancement
Gene promoter
(iii) Lagged gene ac!va!on
(i) (ii)
Enhancer
Enhancement
Gene promoter
Low
RNAPIItranscrip!on
Low abundanceof factors
Inac!ve regulatoryelement
Ac!ve regulatory elementPromoter strength and/or transcrip!onal level
High
High abundanceof factors
TRENDS in Genetics
Figure 1. Active regulatory elements are divergently transcribed. (A) Both regulatory active gene promoters and gene-distal enhancers are transcribed. RNA polymerase II(RNAPII) recruitment and transcription initiation are mediated by general transcription factors (GTFs) binding core promoter regions in close proximity to flankingnucleosomes. This is facilitated by transcription factors (TFs), which often bind proximal to core promoters. Transcription often initiates divergently and at the boundary ofthe nucleosome-depleted region (NDR). (B) Gene expression is often preceded by, or changes concurrently with, changes in enhancer transcription. In a silent(nonexpressed) state (i), enhancers and promoters may, or may not, bind RNAPII. Upon stimulus (ii), transcriptional activity at enhancers marks regulatory enhancer activitywith local transcription and increases in RNAPII recruitment at the target gene promoter. (iii) Gene expression may lag behind transcriptional activation at enhancers. (C)Chromatin interactions place regulatory elements in close physical proximity. The individual properties of regulatory elements (chromatin characteristics as well as TF andRNAPII recruitment strengths) as well as context-dependent properties (such as promoter competition, insulation, and core promoter specificity) jointly determine theformation of multiple regulatory interactions (Box 1). Via regulatory cooperation, multiple regulatory elements may increase the local concentration of factors (TFs, GTFs,co-activators, and RNAPII) needed for transcription in RNAPII-enriched foci (i) and thereby achieve in aggregate different levels of transcriptional activity compared withRNAPII foci, including fewer regulatory elements (ii). Nucleosome illustrations in (A) reproduced, with permission, from [38]; (C) modified, with permission, from [38].
Opinion Trends in Genetics August 2015, Vol. 31, No. 8
428
Kim,T.H.andB.Ren,AnnuRevGenomicsHumGenet,2006
SegalE,Nature2006
23
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer potentialIf active enhancers have promoter activities, it follows thatwhat we classically refer to as promoters could also func-tion as enhancers in certain situations. Although in gener-al a neglected topic of research, we find some clues tosupport this in studies of larger chromatin architectures.
Regulatory elements often interact in close physicalproximity in RNAPII foci that are sometimes referred toas transcription factories [37], and the activity of eachregulatory element within these foci can be influencedby other regulatory elements involved (Figure 1C). Al-though the regulatory mechanisms remain poorly charac-terized, a common model posits that regulatory sequencessynergistically achieve high levels of transcriptional activ-ity by increasing the local concentration of factors neededfor transcription [3741] (Figure 1C). Studies based onchromatin conformation capture have observed RNAPIIfoci involving multiple gene promoters, with few or noenhancers [29,42]. In such arrangements, the degree towhich each gene promoter acts as a promoter or enhancermay depend on the local context of chromatin [42],expressed TFs, and sequence context (e.g., the occurrenceand strength of core promoter elements and TF bindingsites). Using in vitro reporter assays, Li et al. found thatcertain gene promoters in such constellations in MCF7cells had enhancer potential [42], suggesting that they
Box 2. Regulatory elements and associated chromatin
modification states
The N-terminal tails of histone proteins H2A, H2B, H3, and H4 withinnucleosomes can be chemically modified. In particular, amino acidresidues can be methylated or acetylated, which correlates withfunctional properties of the chromatin. For instance, H3K27ac (acet-ylation of lysine 27 in histone H3) is correlated with transcriptionalactivity. The locations of histones marked with a certain histonemodification can be measured genome wide using chromatin im-munoprecipitation (ChIP) coupled with either tiling microarrays(ChIP-chip) or sequencing (ChIP-seq) [38]. Profiling of histone mod-ifications in different cell types has enabled systematic inference ofdifferent types of functional genomic entities [12,53,77,78]. Suchstudies have shown that, typically, the nucleosomes flanking activegene promoters are enriched in H3K27ac, H3K4me3 (trimethylationof lysine 4 in histone H3), and, to some degree, H3K4me1 (mono-methylation of lysine 4 in histone H3), whereas enhancers are en-riched in H3K27ac and H3K4me1, but not H3K4me3, or relatively lowlevels thereof. In addition, several marks have been found to localizeat both enhancers and promoters. Most notably, H3K27ac marksactive promoters and a subset of enhancers that appear to be activelyregulating gene transcription, leading to the notion of classifyingregulatory elements into active and poised classes [15,79]. Poisedregulatory elements sometimes carry bivalent marks associated withactivity and repression at the same time, for instance promotershaving H3K4me3 and H3K27me3 (the latter mark is usually found atinactive gene promoters). Collectively, these studies have arguedthat functional elements can be effectively annotated by their distinctpatterns of histone modifications.
TFs
Regulatory ac!vegene promoters and/or enhancers
Core promoter Core promoterNDR
GTFs
RNAPII RNAPII
(A)
(C)
(B)(i) Silent state
Enhancer
RNAPII
Transcrip!on
Gene promoter
(ii) S!mulus-induced enhancer ac!vity
Enhancer
Enhancement
Gene promoter
(iii) Lagged gene ac!va!on
(i) (ii)
Enhancer
Enhancement
Gene promoter
Low
RNAPIItranscrip!on
Low abundanceof factors
Inac!ve regulatoryelement
Ac!ve regulatory elementPromoter strength and/or transcrip!onal level
High
High abundanceof factors
TRENDS in Genetics
Figure 1. Active regulatory elements are divergently transcribed. (A) Both regulatory active gene promoters and gene-distal enhancers are transcribed. RNA polymerase II(RNAPII) recruitment and transcription initiation are mediated by general transcription factors (GTFs) binding core promoter regions in close proximity to flankingnucleosomes. This is facilitated by transcription factors (TFs), which often bind proximal to core promoters. Transcription often initiates divergently and at the boundary ofthe nucleosome-depleted region (NDR). (B) Gene expression is often preceded by, or changes concurrently with, changes in enhancer transcription. In a silent(nonexpressed) state (i), enhancers and promoters may, or may not, bind RNAPII. Upon stimulus (ii), transcriptional activity at enhancers marks regulatory enhancer activitywith local transcription and increases in RNAPII recruitment at the target gene promoter. (iii) Gene expression may lag behind transcriptional activation at enhancers. (C)Chromatin interactions place regulatory elements in close physical proximity. The individual properties of regulatory elements (chromatin characteristics as well as TF andRNAPII recruitment strengths) as well as context-dependent properties (such as promoter competition, insulation, and core promoter specificity) jointly determine theformation of multiple regulatory interactions (Box 1). Via regulatory cooperation, multiple regulatory elements may increase the local concentration of factors (TFs, GTFs,co-activators, and RNAPII) needed for transcription in RNAPII-enriched foci (i) and thereby achieve in aggregate different levels of transcriptional activity compared withRNAPII foci, including fewer regulatory elements (ii). Nucleosome illustrations in (A) reproduced, with permission, from [38]; (C) modified, with permission, from [38].
Opinion Trends in Genetics August 2015, Vol. 31, No. 8
428
Kim,T.H.andB.Ren,AnnuRevGenomicsHumGenet,2006
AnderssonRetal.,GenomeResearch2009
24
88% of known enhancers that were tested [31]. Theseresults suggest that the promoter activity of an enhanceracross cells is a proxy of its enhancer activity. This con-clusion is also supported by the observation that transcrip-tion at enhancers often changes in a stimulus-dependent
manner [3236] and correlates with changes in the tran-scriptional output of target genes [18,21], often with dy-namics that match or precede gene activation [20,33,35](Figure 1B).
Gene promoters have enhancer pote