Embed Size (px)
Citation preview

ΠΑΝΕΠΙΣΤΗΜΙΟ ΚΡΗΤΗΣ
ΤΜΗΜΑ ΒΙΟΛΟΓΙΑΣ
Καθηγητής Ν. Τσιµενίδης
ΘΑΛΑΣΣΙΑ ΒΙΟΛΟΓΙΑ (Σηµειώσεις)
ΗΡΑΚΛΕΙΟ, ΝΟΕΜΒΡΙΟΣ 2000

2
ΠΕΡΙΕΧΟΜΕΝΑ
1. Η ΘΑΛΛΑΣΑ ΩΣ ΦΥΣΙΚΟ ΠΕΡΙΒΑΛΛΟΝ σελ. 3
1.1. Κατανοµή ξηράς και θάλασσας σελ. 3
1.2. Βάθη, έκταση και χαρακτηριστικά γνωρίσµατα των διαφόρων περιοχών σελ. 4
θαλασσών
1.3. Πυθµένας των ωκεανών σελ. 8
1.4 Ωκεάνια ιζήµατα σελ. 19
1.5. Κατανοµή των ωκεάνιων ιζηµάτων σελ. 32
1.6. ∆ειγµατοληψία ιζηµάτων σελ. 34
1.7. Τα περιθώρια ηπείρων σελ. 35
2. ΧΗΜΙΚΕΣ Ι∆ΙΟΤΗΤΕΣ ΤΟΥ ΘΑΛΑΣΣΙΝΟΥ ΝΕΡΟΥ σελ. 42
2.1. Συστατικά του θαλασσινού νερού σελ. 42
2.2. ∆ιαλυµένα άλατα σελ. 43
2.3. Αλατότητα σελ. 45
2.4. ∆ιαφοροποιήσεις και ανακατανοµη΄των συστατικών του θαλασσινού νερού σελ. 48
2.5. Η αλατότητα ως οικολογικός παράγοντας σελ. 50
2.6. Πυκνότητα θαλασσινού νερού και πίεση σελ. 55
2.7 ∆ιαλυµένα αέρια στο θαλασσινό νερό σελ. 58
2.8 Θρεπτικά σελ. 67
3. Η ΘΑΛΑΣΣΑ ΩΣ ΒΙΟΛΟΓΙΚΟ ΠΕΡΙΒΑΛΛΟΝ σελ. 73
4. ΝΗΚΤΟΝ σελ. 77
4.1. Κεφαλόποδα σελ. 77
4.2. Ψάρια σελ. 82
4.3. Θαλάσσια θηλαστικά σελ. 99
5. Ο ΑΝΘΡΩΠΟΣ ΚΑΙ Η ΘΑΛΑΣΣΑ σελ. 104
ΒΙΒΛΙΟΓΡΑΦΙΚΕΣ ΠΗΓΕΣ σελ. 110

3
1. Η ΘΑΛΑΣΣΑ ΩΣ ΦΥΣΙΚΟ ΠΕΡΙΒΑΛΛΟΝ
1.1. Κατανοµή ξηράς και θάλασσας
Ο πλανήτης Γη κυριαρχείται από ωκεανούς. Οι ωκεανοί καλύπτουν περίπου 71%
της επιφάνειας της Γης. Περίπου 80% της επιφάνειας του Νότιου Ηµισφαιρίου
καλύπτεται από ωκεανούς, ενώ στο Βόρειο Ηµισφαίριο οι ωκεανοί αποτελούν το
61%. Οι περισσότεροι ωκεανοί είναι βαθείς και υπολογίζεται ότι 84% του πυθµένος
των ωκεανών βρίσκεται σε βάθη µεγαλύτερα από 2.000 µ. Το µέσο βάθος των
ωκεανών είναι 3.865 µ ενώ µερικοί τάφροι φθάνουν σε βάθος 11.000 µ περίπου.
∆εδοµένου ότι οι γνώσεις µας για τους θαλάσσιους οργανισµούς περιορίζονται, για
πρακτικούς λόγους κυρίως, σ' εκείνους που ζουν σε βάθη µέχρι 200 µ, η άγνοιά µας
για το συντριπτικά µεγαλύτερο τµήµα των ωκεανών είναι προφανής.
Η εποχή των πάγων της πλειστοκαίνου περιόδου είχε δραµατικές επιδράσεις στη
θαλάσσια ζωή. Η περιοδική προώθηση των ηπειρωτικών πάγων κατά τη διάρκεια
των κρύων περιόδων χαµήλωσε τη στάθµη της θάλασσας εξαιτίας της µεταφοράς και
µετατροπής µέρους του νερού της Γης σε πάγο. Μέχρι τώρα έχουν πραγµατοποιηθεί
διακυµάνσεις της στάθµης των θαλασσών µέχρι και 100 µ (τώρα είµαστε σε περίοδο
σχετικά υψηλής στάθµης της θάλασσας). Κατά τη διάρκεια της τελευταίας
παγετώδους περιόδου, που χρονολογείται στα 11.000 χρόνια πριν, οι ηπειρωτικές
υφαλοκρηπίδες σε ολόκληρο τον κόσµο ήταν κατά το µεγαλύτερο µέρος τους
εκτεθειµένες στον αέρα. Ηµίκλειστοι κόλποι, όπως του Long Island, ήταν λίµνες
γλυκού νερού ή χέρσα γη. Υποχωρήσεις και προωθήσεις των πάγων άλλαξαν
επίσης σηµαντικά την κατανοµή των υδάτινων µαζών και τις κλιµατικές ζώνες των
ωκεανών. Κατά συνέπεια, η πρόσφατη εξελικτική ιστορία της θαλάσσιας ζωής
υπήρξε µέσα στα πλαίσια διακυµάνσεων των κλιµατικών συνθηκών του πλανήτη.
Αυτό το πρότυπο δεν είναι µοναδικό της πλειστοκαίνου περιόδου αλλά εκτείνεται
πίσω δια µέσου της ιστορίας των ωκεανών και της θαλάσσιας ζωής. Παγετώδεις
περίοδοι υπήρξαν επίσης και κατά τον Παλαιοζωικό αιώνα (Ορδοβίκιος και Πέρµιος).
Κατά τις περιόδους αυτές υπήρξαν επίσης µεταβολές κλιµατικές, οι οποίες έχουν
αποδεικτικά στοιχεία. Αυτά τα πρότυπα πρέπει να έχουν επηρεάσει την εξέλιξη της
θαλάσσιας ζωής.

4
1.2. Βάθη, έκταση και χαρακτηριστικά γνωρίσµατα των διαφόρων περιοχών
των ωκεανών και θαλασσών
Πώς τα βάθη των ωκεανών ποικίλουν από τόπο σε τόπο και πώς αυτές οι
διαφοροποιήσεις συνδέονται µε την ξηρά; Πρώτον πρέπει να σηµειωθεί ότι το
µεγαλύτερο τµήµα της επιφάνειας της Γης συγκεντρώνεται σε δύο επίπεδα: τα
επίπεδα εκείνα που βρίσκονται µεταξύ 6.000 και 3.000 µ κάτω από την επιφάνεια της
θάλασσας, και από 1.000 µ κάτω από την επιφάνεια της θάλασσας µέχρι 1.000 µ
πάνω από την επιφάνεια της θάλασσας. (Εικ.1.1). Το πρώτο περιλαµβάνει τις κύριες
ωκεάνιες λεκάνες και το δεύτερο ένα µεγάλο µέρος των περισσότερων ηπείρων που
περιβάλλονται από ρηχές θάλασσες. Μεταξύ αυτών των δύο κύριων επιπέδων
υπάρχει σηµαντική περιπλοκότητα.
Εικ. 1.1 Η µέση ανύψωση της ξηράς πάνω από την επιφάνεια της θάκασσας και το µέσο βάθος των
ωκεανών (τροποποιηµένη, από McLennan, 1965 and Weyl, 1970)
1.2.1. Ωκεάνια λεκάνη - Ήπειρος
Ως ωκεάνια λεκάνη ορίζεται µια µεγάλη περιοχή του πυθµένα των ωκεανών που
βρίσκεται σε βάθος µεγαλύτερο από 2.000 µ. Ως ήπειρος ορίζεται µια µεγάλη
συνεχής περιοχή της ξηράς. Το αόριστο σ' αυτούς τους ορισµούς είναι η λέξη
"µεγάλη". Γιατί η Αυστραλία θεωρείται ήπειρος και η Γροιλανδία (περίπου 2 εκατοµ.
Km2) νησί; Γιατί η Καραϊβική λέγεται θάλασσα και ο Αρκτικός λέγεται Ωκεανός; Τα
ονόµατα φαίνεται µερικές φορές ότι είναι αυθαίρετα. Ακόµη µε την χρήση τους επί
αιώνες τα ονόµατα εντάσσονται στη γλώσσα. Στην Ωκεανογραφία ο όρος "θάλασσα"

5
σήµερα αναφέρεται σε ένα µεγάλο, αλµυρό, σχετικά κλειστό όγκο νερού ή σε µια
µάζα νερού µε φυσικά και χηµικά χαρακτηριστικά τα οποία την διακρίνουν από τον
πλησίον Ωκεανό. Σε σύγκριση µε τον ωκεανό, είναι µικρότερη σε έκταση, έχει
µεγαλύτερη επικοινωνία µε την ξηρά και έχει µικρότερο βάθος. Το ∆ιεθνές
Υδρογραφικό Γραφείο αναγνωρίζει πενήντα τέσσερις θάλασσες αλλά µόνον έξι
Ωκεανούς: τον Αρκτικό, το Βόρειο Ατλαντικό, το Νότιο Ατλαντικό, το Βόρειο Ειρηνικό,
το Νότιο Ειρηνικό και τον Ινδικό.
Οι θάλασσες διακρίνονται συνήθως σε τέσσερις κατηγορίες: Παράκτιοι
(επικοινωνούν ευρέως µε τους ωκεανούς - π.χ. Βόρεια Θάλασσα), Μεσόγειοι
(περιορισµένη επικοινωνία µε τους ωκεανούς -π.χ. Μεσόγειος Θάλασσα, Ερυθρά
Θάλασσα), Εσωτερικές (περιβάλλονται κατά το πλείστον από ξηρά - π.χ. Βαλτική
Θάλασσα, Μαύρη Θάλασσα) και Κλειστές (περιβάλλονται τελείως από ξηρά - π.χ
Κασπία Θάλασσα)
Όπως ακριβώς οι ήπειροι έχουν διακριτικά τοπογραφικά χαρακτηριστικά, π.χ. βουνά,
κοιλάδες, φαράγγια κλπ., έτσι έχουν και οι ωκεάνιες λεκάνες. Μεταξύ των
χαρακτηριστικών των ωκεάνιων λεκανών θα αναφέρουµε τα εξής σπουδαιότερα:
Ωκεάνιες ράχες (oceanic ridges), αβυσσαίες πεδιάδες (abyssal plains),
κοραλλιογενείς ύφαλοι (coral reefs), νησιωτικά τόξα (island arcs), τάφροι (trenches),
θαλάσσια βουνά (seamounts) και νησιωτικές αλυσίδες (island chains). Η βαθύτερη
περιοχή είναι το Challenger Deep στην τάφρο Mariana στο ∆υτικό Βόρειο Ειρηνικό
Ωκεανό, που έχει βάθος πλέον των 11 χλµ.
1.2.2 Σύνθεση των πετρωµάτων
Η διαφορά µεταξύ των ωκεάνιων λεκανών και ηπείρων επεκτείνεται πέρα από
µεγάλους τοπογραφικούς χαρακτήρες. Υπάρχουν σηµαντικές διαφορές στη σύνθεση
των πετρωµάτων και την πυκνότητά τους, το πάχος των ιζηµάτων και τις σχετικές
ιδιότητες. Ο ωκεάνιος φλοιός (oceanic crust) αποτελείται από ένα στρώµα, το οποίο
συνίσταται κυρίως από πετρώµατα βασάλτη και γαύρου, που βρίσκονται πάνω σε
στρώµα περιδοτίτη. Όλα αυτά αποτελούνται από πυριγενή ορυκτά δηλ. είναι
πετρώµατα τα οποία αποτελούνται από λειωµένα υλικά που έχουν κρυσταλλωθεί. Ο
βασάλτης και ο γαύρος διαφέρουν στο µέγεθος των κρυστάλλων αλλά όχι στην
σύνθεσή τους. Και οι δύο περιέχουν τα ορυκτά "πυρόξενοι" και"olivine", και είναι
πλούσια σε ασβεστιούχους άστριους. Ο περιδοτίτης περιέχει πυρόξενους και olivine
όχι όµως άστριους. Όλα είναι πλούσια σε σίδηρο και µαγνήσιο.
Ο ηπειρωτικός φλοιός είναι λιγότερο οµογενής. Αποτελείται από πυριγενή
πετρώµατα τα οποία είναι κυρίως γρανιτικής σύνθεσης. Ένας τυπικός γρανίτης

6
αποτελείται από χαλαζία, κάλλιο και άστριους πλούσιους σε νάτριο, ελάχιστα ποσά
σιδήρου και ορυκτά που περιέχουν µαγνήσιο. Ο ηπειρωτικός φλοιός περιέχει
λιγότερο σίδηρο, µαγνήσιο και ασβέστιο από τον ωκεάνιο φλοιό, αλλά περισσότερο
πυρίτιο, νάτριο και κάλιο.
1.2.3. Πυκνότητα
Εξαιτίας των διαφορών στη σύνθεση µεταξύ του ηπειρωτικού φλοιού και του
ωκεάνιου φλοιού, οι πυκνότητές τους διαφέρουν. Η µέση πυκνότητα του
ηπειρωτικού φλοιού είναι περίπου 2,7g/cm3, ενώ του ωκεάνιου περισσότερο από 3.0
g/cm3. Eξαιτίας της διαφοράς αυτής παρατηρείται διαφορά µεταξύ του ύψους των
εξάρσεων στην ξηρά και τη θάλασσα (Εικ. 1.2).
Εικ.1.2 ∆ιαφορές µεταξύ του ηπειρωτικού και ωκεάνιου φλοιού (τροποποιηµένη, από D. E. Ingmason
and W. J. Wallace, 1989)
1.2.4. Ισοστασία
Ισοστασία (isostacy) σηµαίνει ότι όλες οι µεγάλες περιοχές του φλοιού της γης είναι
σε ισορροπία σαν να επιπλέουν πάνω σ' ένα πυκνότερο στρώµα. Περιοχές µε
λιγότερο πυκνό φλοιό επιπλέουν τοπογραφικά ψηλότερα από περιοχές µε πυκνότερο
φλοιό, εφόσον είναι του ιδίου πάχους. Αυτό εξηγεί γιατί ήπειροι, οι οποίοι

7
αποτελούνται από σχετικά χαµηλότερης πυκνότητας πετρώµατα, υψώνονται
τοπογραφικά υψηλότερα από τις ωκεάνιες λεκάνες (Εικ. 1.3).
Εικ. 1.3 Σχηµατική απεικόνιση της έννοιας ¨ισοστασία¨ του γήινου φλοιού (τροποποιηµένη, από D. E.
Ingmason and W. J. Wallace, 1989)
Ας υποθέσουµε ότι ένα τµήµα του φλοιού - π.χ. µια ήπειρος -επιπλέει σε ισοστασία,
σύµφωνα µε την πυκνότητά της. Αν προστεθεί βάρος στο τµήµα αυτό ή αφαιρεθεί
βάρος, η ισορροπία καταστρέφεται. Ας υποθέσουµε ότι ο Καναδάς επέπλεε σε
ισοστασική ισορροπία. Αν ένα πυκνό κάλυµµα πάγου κάλυπτε την περιοχή, όπως
πράγµατι συνέβαινε κατά την περίοδο των παγετώνων, το επιπλέον βάρος θα έκανε
την ξηρά να βυθισθεί. Μόλις ο πάγος έλειωνε το τµήµα αυτό του φλοιού θα
ανυψώνονταν και πάλι. Το νερό που περιείχετο στα τεράστια παγόβουνα
προέρχονταν φυσικά από τούς ωκεανούς. Συνεπώς, καθώς σχηµατίζονταν πάγος, ο
όγκος του νερού στους ωκεανούς και το βάρος του νερού επί του πυθµένος των
ωκεανών µειώνονταν και κατά συνέπεια ο ωκεάνιος πυθµένας αναγκάζονταν να
ανυψωθεί. Κατά την περίοδο των παγετώνων ο Καναδάς καταποντίσθηκε 200 µ
περίπου κάτω από το σηµερινό επίπεδο, υπό την πίεση στρώµατος πάγου, πάχους
2-3 χλµ και ο ωκεάνιος πυθµένας ανυψώθηκε κατά 40 µ περίπου. Όταν ο πάγος

8
έλειωσε και το νερό επέστρεψε στους ωκεανούς, ο ωκεάνιος πυθµένας
καταποντίσθηκε πάλι.
1.3. Πυθµένας των ωκεανών
Ο πυθµένας των ωκεανών, κατά το µεγαλύτερο µέρος, αποτελείται από στρώµατα
ιζηµάτων τα οποία έχουν σχηµατιστεί σιγά-σιγά σε εκατοµµύρια χρόνια. Ο
χαρακτήρας της ζωής στα υπερκείµενα νερά, το βάθος, η απόσταση από την ακτή, τα
ρεύµατα, αλλά και άλλοι παράγοντες, επηρεάζουν τη φύση των ιζηµάτων σε µια
περιοχή. Μετά το πέρασµα των χρόνων, αλλαγές του κλίµατος, της θερµοκρασίας,
των ρευµάτων και της ηφαιστειογενούς δραστηριότητας, καθώς και η ισορροπία του
γήινου φλοιού, όλα αυτά µαζί µε αλλαγές της πανίδας και χλωρίδας των θαλασσών,
διαµορφώνουν τη φύση των ιζηµάτων και τα απολιθώµατα µέσα σ' αυτά.
Θαλάσσια ιζήµατα από παράκτιες περιοχές και πρώην αβαθείς θάλασσες, που τώρα
είναι πάνω από τη στάθµη της θάλασσας, είναι άµεσα διαθέσιµα για µελέτη και
παρέχουν στους γεωλόγους και παλαιοντολόγους τις περισσότερες πληροφορίες για
την ιστορία της Γης. Η µελέτη των ωκεάνιων ιζηµάτων µε τη χρησιµοποίηση
σύγχρονων ηχοβολιστικών, ηλεκτρονικών και µαγνητικών µεθόδων, για τη λήψη
πληροφοριών σχετικά µε τον πυθµένα των θαλασσών, έχει προσθέσει µια καινούργια
διάσταση στην έρευνα της ιστορίας της Γης.
1.3.1 Χαρακτηριστικά του Ωκεάνιου πυθµένα
(α) Ωκεάνιες ράχες (οροσειρές)
Για περισσότερο από δύο αιώνες, οι Γεωλόγοι είχαν αναγνωρίσει ότι µερικά νησιά,
όπως αυτό της Ισλανδίας, είναι κορυφές υποθαλάσσιων οροσειρών. Οι µεσο-
ωκεάνιες ράχες (mid-oceanic ridges) αποτελούν µια γιγαντιαία και συνεχή
υποθαλάσσια οροσειρά, η οποία έχει συνολικό µήκος 65.000 χλµ ή περίπου 1,5
φορά το µήκος της περιφέρειας της Γης. Επίσης έχουν µέσο πλάτος µεγαλύτερο των
1.000 χλµ, καλύπτουν περίπου το 23% της επιφάνειας της Γης και είναι σηµαντικά
µορφολογικά στοιχεία του πλανήτη µας. Τα µεγαλύτερα πλάτη παρατηρούνται εκεί
όπου οι ράχες είναι περισσότερο ενεργές. Η µεσο-ωκεάνια ράχη ανυψώνεται δύο
χιλιόµετρα, ή ακόµη και εξέρχεται από την επιφάνεια της θάλασσας, όπως συµβαίνει
στην περίπτωση της Ισλανδίας. Η Μεσο-Ατλαντική Ράχη (Mid - Atlantic Ridge )
διατρέχει το µήκος ολόκληρου του Ατλαντικού Ωκεανού από 55ο Νότια µέχρι 80ο
Βόρεια ( Εικ. 1.4 ).

9
Στο Νότιο Ατλαντικό, η οροσειρά (ράχη) στρέφει ανατολικά προς τον Ινδικό Ωκεανό,
όπου καθίσταται η Μεσο-Ινδική Ράχη (Mid - Indian Ridge), η οποία στη συνέχεια
διακλαδίζεται στις 25ο Νότια. Ένας κλάδος πηγαίνει προς Βορρά στην Ερυθρά
Θάλασσα και ο άλλος προς Νότο, µεταξύ Αυστραλίας και Ανταρκτικής. Εκεί γίνεται η
Ειρηνικό - Ανταρκτική Ράχη (Pacific - Antarctic Ridge). Το σύστηµα συνεχίζει δια
µέσου του Ανατολικού Ειρηνικού Ωκεανού, όπου εκεί ονοµάζεται έξαρση του
Ανατολικού Ειρηνικού (East Pacific Rise), καθώς και προς τον Κόλπο της
Καλιφόρνια. Εκεί εξαφανίζεται για να εµφανισθεί δυτικά της πολιτείας Όρεγκον.
Αυτό το εκτεταµένο σύστηµα ωκεάνιων οροσειρών είναι ένας κύριος τοπογραφικός
χαρακτήρας του φλοιού της Γης. (Εικ. 1.5).
Εικ. 1.4 Χάρτης ενός τµήµατος του πυθµένα στον Ατλαντικό, που παρουσιάζει µερικά κύρια
χαρακτηριστικά του ωκεάνιου πυθµένα ωκεάνια ράχη, εγκάρσια ρήγµατα, φαράγγια, θαλάσσια βουνά,
εξάρσεις, τάφροι και αβυσσαίες πεδιάδες (τροποποιηµένη, από Heezen, Tharp and Ewing,19..)
Ολόκληρη η µεσο-ωκεάνια ράχη διατρέχεται κατά µήκος του κεντρικού της άξονα
από µια τεκτονική τάφρο (rift valley). Η τεκτονική τάφρος, η οποία περιβάλλεται από
τεκτονικά όρη (rift mountains) χαρακτηρίζεται από απότοµα τοιχώµατα και έχει βάθος
από 0,5 µέχρι 1,5 χλµ και πλάτος από 12 µέχρι 48 χλµ περίπου. Την τεκτονική τάφρο
τέµνουν κάθετα προς αυτήν και παράλληλα µεταξύ τους τα ρήγµατα τα οποία είναι
γνωστά ως ρήγµατα µετασχηµατισµού (transform faults) κατά µήκος των οποίων η
τεκτονική τάφρος είναι πλευρικά µετατοπισµένη (Εικ.1.6).

10
Εικ. 1.5 Σχηµατικός παγκόσµιος χάρτης των ωκεάνιων οροσειρών (µεσο-ωκεάνιες ράχες) (από S.
Thorarinson “Surtsey”, 1966 in D. E. Ingmanson and W. J. Wallace, 1989)
Εικ. 1.6 Οι µεσο-ωκεάνιες ράχες είναι µετατοπισµένες από ρήγµατα γνωστά ως ρήγµατα
µετασχηµατισµού. Τα πετρώµατα µεταξύ των σηµείων Α και Β του ρήγµατος µετακινούνται σε αντίθετες
κατευθύνσεις (από Α. Θεοδώρου, 1996)
Οι ωκεάνιες τάφροι, σε αντίθεση µε τις µεσο-ωκεάνιες ράχες, καλύπτουν πολύ
µικρότερο µέρος της γήινης επιφάνειας.

11
Το σύστηµα των ωκεάνιων οροσειρών χαρακτηρίζεται από ρήγµατα, συχνούς
σεισµούς µικρού βάθους (συνήθως 20-30 χλµ) και ηφαίστεια. Όταν ο φλοιός
ανυψώνεται πιεζόµενος από κάτω για λόγους ισοστασικής ισορροπίας, θραύεται µε
αποτέλεσµα την εκροή λάβας η οποία χύνεται στις ρωγµές του φλοιού. Εξαιτίας του
γεγονότος ότι η ψύξη εµποδίζει τη λάβα να ανέλθει σε όλο το µήκος της σχισµής του
φλοιού µέχρι την επιφάνεια, σχηµατίζεται ένα κάλυµµα πάνω από τη σχισµή και κάτω
από το γειτονικό θαλάσσιο πυθµένα. Καθώς ο φλοιός εξακολουθεί να ανυψώνεται,
πιεζόµενος από κάτω, σχηµατίζονται καινούργιες σχισµές, πληρούνται µε λάβα,
ψύχονται και καλύπτονται όπως προαναφέρθηκε. Αυτή η διαδικασία σχηµατίζει µια
κοιλάδα µε ρωγµές στο φλοιό της γης (rift valleys), των οποίων η γωνία πλησιάζει τις
90ο σε σχέση µε την κατεύθυνση της οροσειράς (Εικ.1.4). Η Ισλανδία και τα γειτονικά
νησιά είναι σχεδόν εξολοκλήρου ηφαιστειογενούς προέλευσης και ταυτόχρονα
ηφαίστεια εν ενεργεία εξακολουθούν να υπάρχουν εκεί.
Τόσο η ηφαιστειακή όσο και η σεισµική δράση µειώνονται µε την απόσταση από την
τεκτονική τάφρο ενώ το βαθυµετρικό ανάγλυφο γίνεται οµαλότερο. Το τελευταίο
συµβαίνει,τόσο διότι αυξάνεται το πάχος των ιζηµάτων που καλύπτουν τον πυθµένα
όσο και λόγω της ψύξης του µάγµατος που εκχύνεται και επικάθεται επί του ωκεάνιου
πυθµένα. Στον Ατλαντικό και τον Ινδικό Ωκεανό, ο δηµιουργούµενος στο κέντρο της
µεσο-ωκεάνειας ράχης νέος πυθµένας, επεκτεινόµενος από τις δύο πλευρές της
ράχης, συµπαρασύρει µαζί του τις γειτονικές ηπείρους. Κατά συνέπεια και οι δύο
αυτοί ωκεανοί αυξάνονται σε έκταση. Ιδιαίτερα στον Ινδικό ωκεανό, η ωκεάνια ράχη
διεισδύει στον κόλπο του Αντεν και την Ερυθρά Θάλασσα, προκαλώντας βαθµιαία
την διαπλάτυνσή τους. Στον Ατλαντικό ωκεανό, η Αµερική και η Γροιλανδία αφενός,
και αφετέρου η Ευρώπη και η Αφρική, µεταφέρονται σε αντίθετες κατευθύνσεις από
την Μεσο-Ατλαντική Ράχη και προς τις δύο πλευρές της. Η Μεσο-Ατλαντική Ράχη
βρίσκεται ακόµη και σήµερα στο µέσο του Ατλαντικού Ωκεανού, γεγονός που
σηµαίνει ότι ο ρυθµός εξάπλωσης του πυθµένα παραµένει ο ίδιος και προς τις δύο
κατευθύνσεις επέκτασης.
Τα ηφαιστειογενή πετρώµατα, τα οποία σχηµατίζουν την οροσειρά είναι βασάλτης.
Φωτογραφίες και παρατηρήσεις µπορούν να αποκαλύψουν θερµές κηλίδες (hot
spots) και πρόσφατη λάβα χυµένη πάνω στις ρωγµώδεις κοιλάδες (Εικ. 1.7).
Σπάνια εµφανίζονται ιζήµατα πάνω στα πυριγενή πετρώµατα τα οποία σχηµατίζουν
τις ωκεάνιες οροσειρές. Όταν εµφανίζονται έχουν πάντοτε πάχος λιγότερο από 100µ.

12
Εικ. 1.7 Ηφαιστειακές θερµές πηγές, όπως αυτές στο νησί της Χαβάης (τροποποιηµένη, από Van Andel,
1985)
Ο ωκεάνιος φλοιός έχει σχετικά µικρή γεωλογική ηλικία, η οποία αυξάνεται µε την
απόσταση από την κορυφή της µεσο-ωκεάνιας ράχης. Έτσι στην περιοχή της
κορυφής της µεσο-ωκεάνιας οροσειράς συναντάται ο νεαρότερος φλοιός, ενώ όσο
αποµακρυνόµαστε από την κορυφή αυξάνεται η ηλικία του φλοιού, µε ρυθµό περίπου
ενός εκατοµµυρίου ετών για κάθε 20 έως 40 χλµ απόσταση. Το γεγονός αυτό
δηλώνει ότι ο νέος φλοιός δηµιουργείται στην κορυφή της ωκεάνιας ράχης και στη
συνέχεια αποµακρύνεται αργά απ’ αυτήν.
(β) Ωκεάνιες Λεκάνες
Η περιοχή του ωκεάνιου πυθµένα, που εκτείνεται από τη µεσο-ωκεάνια ράχη έως την
ωκεάνια τάφρο ή το ηπειρωτικό πρανές, ονοµάζεται ωκεάνια λεκάνη. Οι ωκεάνιες
λεκάνες καλύπτουν συνολικά το 30% της γήινης επιφάνειας και είναι σχεδόν
επίπεδες επιφάνειες µε διάσπαρτους µικρούς λόφους που ονοµάζονται αβυσσαίοι
λόφοι και σποραδικά από µεγαλύτερου µεγέθους ηφαιστειακά υποθαλάσσια βουνά.
Το ανάγλυφο των ωκεάνιων λεκανών γίνεται οµαλότερο µε την απόσταση από τη
ράχη –λόγω µεγαλύτερης ηλικίας και κατά συνέπεια µεγαλύτερου πάχους
ιζηµατογενούς καλύµµατος. Επίσης κοντά στο ηπειρωτικό πρανές πολλά
υποθαλάσσια φαράγγια κόβουν το ηπειρωτικό υφαλοπρανές εγκάρσια και
µεταφέρουν τεράστιες ποσότητες ιζηµάτων, µέσω των τουρβιδιτικών ρευµάτων
(πυκνά, µετακινούµενα λόγω βαρύτητας ρεύµατα, περιέχοντα ιζήµατα που
αιωρούνται), τα οποία διασπείρονται σχηµατίζοντας τεράστιες επίπεδες εκτάσεις οι
οποίες ονοµάζονται αβυσσαίες πεδιάδες. Οι αβυσσαίες πεδιάδες είναι πλατιές
εκτεταµένες περιοχές που βρίσκονται σε όλους του ωκεανούς σε βάθη που
κυµαίνονται από 4 µέχρι 5 χλµ. Οι πεδιάδες αυτές συνορεύουν µε ωκεάνιες
οροσειρές (oceanic ridges) ή ηπείρους. Ορισµένες είναι πολύ καλά σχηµατισµένες

13
ανοικτά των εκβολών ποταµών, όπως αυτές ανοικτά των εκβολών του Γάγγη και
Ινδού ποταµού, που εκβάλλουν στον Ινδικό ωκεανό. Αυτές οι αβυσσαίες πεδιάδες,
λόγω σχήµατος ονοµάζονται ριπιδοειδείς λεκάνες (deep sea fans).
Οι περισσότερες αβυσσαίες πεδιάδες είναι περιοχές µε οµαλή κλίση, καλυπτόµενες
συχνά µε σχετικά µη συµπυκνωµένα ιζήµατα τα οποία προέρχονται από τουρβιδιτικά
ρεύµατα. Τα ιζήµατα στις αβυσσαίες πεδιάδες τείνουν να είναι πολύ
περισσότερο λεπτά σε σχέση µε εκείνα επί των ηπείρων, κυµαινόµενα σε πάχος από
µερικές εκατοντάδες µέχρι 1000 περίπου µέτρα.
Έχοντας σχηµατισθεί πλησίον ωκεάνιων οροσειρών, το στρώµα βασάλτη που
βρίσκεται κάτω από το ωκεάνιο ίζηµα, είναι πολύ ακανόνιστο. Καθώς ο ωκεάνιος
φλοιός αποµακρύνεται από τις µεσο-ωκεάνιες ράχες, προς την πλευρά της
ηπειρωτικής υφαλοκρηπίδας, τα τουρβιδιτικά ρεύµατα µεταφέρουν ίζηµα από τις
ηπείρους, το οποίο θάβει το ακανόνιστο στρώµα του βασάλτη και σχηµατίζει την
αβυσσαία πεδιάδα. Συνεπώς οι ακανόνιστες περιοχές του στρώµατος του βασάλτη,
της πλευράς της τάφρου προς την θάλασσα, δέχονται λίγο ίζηµα σχηµατίζοντας τους
αβυσσαίους λόφους (abyssal hills). Οι αβυσσαίες πεδιάδες και οι αβυσσαίοι λόφοι
καλύπτονται από λεπτό στρώµα πελαγικών ιζηµάτων, τα οποία αποτελούνται κυρίως
από βιογενή λάσπη (biogenic ooze) µε λίγο πηλό. Η βιογενής λάσπη αποτελείται
από ασβεστολιθικούς ή πυριτικούς σκελετούς µικροσκοπικών οργανισµών οι οποίοι
ζουν κοντά στην επιφάνεια του ωκεανού. Μερική από την λάσπη προσλαµβάνεται
από ισχυρά ρεύµατα, κοντά στην ακτή σε εκβολές ποταµών, συγκρατείται εν
αιωρήσει και καταπίπτει µακριά προς την θάλασσα. Άλλα σωµατίδια επίσης
µεταφέρονται µε τον άνεµο από ερηµικές περιοχές, όπως τη Σαχάρα, και αποτίθενται
στην ανοικτή θάλασσα, όπου σιγά-σιγά βυθίζονται στον πυθµένα. Πλέον αυτών,
άλλα σωµατίδια, που αποτελούνται από ηφαιστειογενή στάχτη, µεταφέρονται µε τον
άνεµο πάνω από την θάλασσα. Τέλος, τεράστιες περιοχές των αβυσσαίων
πεδιάδων καλύπτονται από κονδύλους µαγγανίου, οι οποίοι σε µερικές περιοχές είναι
τόσο πυκνά τοποθετηµένοι ώστε εµποδίζουν να φανεί το υποκείµενο ίζηµα.
(γ) Θαλάσσια βουνά
Στον ωκεάνιο πυθµένα είναι εµφανή αρκετά µορφολογικά χαρακτηριστικά που
προέρχονται από ηφαιστειακή δραστηριότητα. Πολλά από τα ενεργά ηφαίστεια της
Γης βρίσκονται στην ηπειρωτική πλευρά των ζωνών καταβύθισης, επάνω από την
περιοχή όπου βυθίζεται η ωκεάνια πλάκα. Ολόκληρη σχεδόν η υπόλοιπη
ηφαιστειακή δραστηριότητα σχετίζεται µε τις ωκεάνιες ράχες. Υπάρχουν όµως
ορισµένες µεµονωµένες περιοχές ηφαιστειακής δράσης, που δεν σχετίζονται ούτε µε

14
τις ζώνες καταβύθισης ούτε µε τις ωκεάνιες ράχες αλλά µε τις θερµές κηλίδες στον
µανδύα της Γης, από τις οποίες εκτοξεύεται κατά διαστήµατα µάγµα.
Εάν το ηφαίστειο εξέρχεται από την επιφάνεια της θάλασσας δηµιουργείται
ηφαιστειακό βουνό (νησί). Τα υπάρχοντα σήµερα ηφαιστειακά νησιά, όπως π.χ.
Χαβάη, βυθίζονται σιγά-σιγά και προοδευτικά καθίστανται υποθαλάσσια. Τα
υποθαλάσσια βουνά ανυψώνονται πάνω από τις αβυσσαίες πεδιάδες. Τα βουνά
αυτά είναι δύο τύπων: θαλάσσια βουνά τα οποία έχουν κυκλικές ή ακανόνιστες
κορυφές και τα τραπεζοειδή βουνά (guyots) τα οποία έχουν επίπεδες κορυφές. Και
οι δύο τύποι βουνών είναι σβησµένα ηφαίστεια. Αν και οι κορυφές τους µπορεί να
είναι σηµαντικά κάτω από τη θάλασσα, απολιθωµένοι οργανισµοί ρηχών θαλασσών,
όπως κοράλλια και δίθυρα µαλάκια, έχουν βρεθεί πάνω στις επιφάνειές τους. Οι
επίπεδες κορυφές των τραπεζοειδών βουνών φαίνεται να είναι αποτέλεσµα
διάβρωσης από κυµατισµό που έχει πραγµατοποιηθεί πριν γίνουν υποθαλάσσια.
(δ) Κοραλλιογενείς ύφαλοι
Οι κοραλλιογενείς ύφαλοι είναι ύφαλοι στερεοί, πορώδεις και ανθεκτικοί στη δράση
των κυµάτων και έχουν προέλθει από την συσσωµάτωση και στερεοποίηση
σκελετικών υπολειµµάτων πολλών γενεών κοραλλίων και µαλακίων. Η βάση της
τροφικής αλυσίδας σε µιά αποικία κοραλλίων είναι η φωτοσυνθετική δραστηριότητα
ενός φύκους που ζεί στον ιστό των κοραλλίων. Η ανάγκη πρόσληψης ηλιακής
ακτινοβολίας απαιτεί την παρουσία των κοραλλίων κοντά στην επιφάνεια της
θάλασσας. Επειδή τα κοράλλια είναι διηθηµατοφάγοι οργανισµοί, εκµεταλεύονται την
κινητικότητα του θαλάσσιου νερού η οποία προκαλεί την προσκοµιδή τροφικών
σωµατιδίων.
Σε τροπικές περιοχές, ιδιαίτερα στον Ειρηνικό Ωκεανό, διασκορπισµένα νησιά
περιβάλλονται στο χείλος τους από κοραλλιογενείς υφάλους και ανυψώνονται µέσα
από τον ωκεάνιο πυθµένα. Οι ύφαλοι αυτοί καλούνται κροσωτοί κοραλλιογενείς
ύφαλοι (fringing reefs) και αποτελούνται κυρίως από κοχύλια, φύκια και τα
υπολείµµατα αποικιών κοραλλίων (Εικ. 1.8α).
Η ανάπτυξη των κοραλλίων είναι ταχύτερη στη θαλάσσια πλευρά της αποικίας, όπου
η δράση των κυµάτων είναι µέγιστη και η επίδραση πιθανών χερσαίων απορροών
ελάχιστη. Αυξανόµενη η κοραλλιογενής αποικία εγκαταλείπει πίσω της µια
λιµνοθάλασσα, και στο στάδιο αυτό της ανάπτυξής της ονοµάζεται κοραλλιογενές
φράγµα (barrier reef) (Εικ. 1.8β). Σε ορισµένες περιπτώσεις οι κοραλλιογενείς ύφαλοι
σχηµατίζουν επιµήκη φράγµατα µε υπολείµµατα κοραλλίων, που είναι τοποθετηµένα
γύρω, µακριά από τις ακτές του νησιού, όπως αυτό συµβαίνει στις ανατολικές ακτές

15
της Αυστραλίας. Η συνεχής συσσώρευση σκελετικών υπολειµµάτων συµβάλλει στην
προς την θάλασσα επέκταση του κοραλλιογενούς φράγµατος και στην διεύρυνση της
λιµνοθάλασσας.
Καθώς το νησί βυθίζεται, οι κοραλλιογενείς ύφαλοι αυξάνονται σε ύψος µε τον ίδιο
ρυθµό, δηµιουργώντας στη θέση του νησιού µια λιµνοθάλασσα, που περιβάλλεται
από µια σειρά κοραλλιογενών νησιών τοποθετηµένων κυκλικά. Ο σχηµατισµός αυτό
καλείται ατόλλη (atoll) (Εικ. 1.8γ).
Εικ. 1.8 Σχηµατική παράσταση σειράς διαδοχικών φάσεων σχηµατισµού (α) κροσωτού κοραλλιογενούς
υφάλου, (β) κοραλλιογενούς φράγµατος και (γ) ατόλλης (τροποποιηµένη, από D. E. Ingmason and W. J.
Wallace, 1989)
Ο ∆αρβίνος εξετάζοντας τους σχηµατισµούς των κοραλλίων παρατήρησε ότι υπάρχει
στενή σχέση µεταξύ των ηφαιστειογενών νησιών και των κοραλλιογενών
σχηµατισµών. Ειδικότερα στο βιβλίο "The structure and Distribution of Coral Reefs"
από παρατηρήσεις που έκανε στα 1830, γράφει τα εξής: "Τα κύρια είδη των
κοραλλιογενών υφάλων βρέθηκαν να διαφέρουν λίγο σε ό,τι αφορά στην πραγµατική
επιφάνεια του υφάλου. Ο σχηµατισµός ατόλλης διαφέρει από τον σχηµατισµό
κοραλλιογενούς φράγµατος µόνο ως προς την απουσία ξηράς στο κέντρο του
σχηµατισµού. Ο σχηµατισµός του κοραλλιογενούς φράγµατος διαφέρει από τον
κροσωτό κοραλλιογενή ύφαλο στο γεγονός ότι σχηµατίζεται σε πολύ µεγαλύτερη

16
απόσταση από την ακτή, σε σχέση µε την πιθανή κλίση της υποθαλάσσιας έναρξης
σχηµατισµού του, καθώς και στην παρουσία µιας βαθιάς λιµνοθάλασσας. Στους
σχηµατισµούς αυτούς είναι σύνηθες να βλέπει κανείς τεράστιες περιοχές από τις
οποίες πολύ συχνά ανυψώνεται ηφαιστειακό υλικό. Επίσης µπορεί να παρατηρήσει
κανείς µεγάλες περιοχές να βυθίζονται σιγά-σιγά χωρίς να έχει προηγηθεί έκρηξη
ηφαιστείου. Βλέποντας αυτά αισθάνεται κανείς βέβαιος ότι αυτή η καταβύθιση
πρέπει να υπήρξε τεράστια σε ποσότητα και έκταση, έτσι ώστε να έχει θάψει κάτω
από την επιφάνεια της θάλασσας ολόκληρα νησιά πάνω από τα οποία παραµένουν
οι ατόλλες, σαν µνηµεία, σηµειώνοντας την θέση όπου υπήρχαν τα βουνά πριν την
καταβύθισή τους’’.
Περίπου έναν αιώνα αργότερα ο γεωλόγος και γεωφυσικός Harry Hess επρότεινε την
πραγµατοποίηση πυρηνοληψίας στις ατόλλες για να δοκιµασθεί η θεωρία του
∆αρβίνου. Οι πυρηνοληψίες αυτές απέδειξαν ότι κάθε ατόλλη έχει µια ηφαιστειακή
ρίζα θαµµένη κάτω από ασβεστόλιθο µέχρι βάθος 800 µ, γεγονός που επιβεβαίωσε
βασικά την θεωρία του ∆αρβίνου.
(ε) Αλυσίδες ηφαιστειακών νησιών
Αλυσίδες ηφαιστειακών νησιών δηµιουργούνται όταν ο επεκτεινόµενος ωκεάνιος
πυθµένας διέρχεται πάνω από µια στάσιµη θερµή κηλίδα. Τα πολύ νέα σε ηλικία
ηφαίστεια µιας αλυσίδας εξέρχονται από την επιφάνεια της θάλασσας και αποτελούν
ηφαιστειακά νησιά, ενώ τα µεγαλύτερης ηλικίας έχουν βυθισθεί. Ο τύπος αυτός
ηφαιστειακής δραστηριότητας είναι ιδιαίτερα εµφανής στον κεντρικό και δυτικό
Ειρηνικό Ωκεανό. Kαθώς ο ωκεάνιος πυθµένας επεκτείνεται προς τα βορειοδυτικά
από το ύψωµα του ανατολικού Ειρηνικού, σχηµατίζονται µακριές αλυσίδες
ηφαιστειακών νησιών, στις οποίες τα µεγαλύτερα σε ηλικία ηφαίστεια βρίσκονται στο
βορειοδυτικό άκρο κάθε αλυσίδας. Tα νησιά Χαβάη, η Γαλλική Πολυνησία τα νησιά
Μαρσάλ και πολλά άλλα είναι απλός τα νεώτερα µέλη µακρών αλυσίδων
ηφαιστειακών υποθαλάσσιων νησιών, που εκτείνονται χιλιάδες χιλιόµετρα προς τα
βορειοδυτικά των νησιών αυτών. Στο µακρινό µέλλον τα παραπάνω νησιά θα
µετακινηθούν µαζί µε τον ωκεάνιο πυθµένα προς τα βορειοδυτικά και βαθµιαία θα
βυθιστούν κάτω από την επιφάνεια της θάλασσας.
1.3.2 Νησιωτικά τόξα και Τάφροι
Πολλά από τα ενεργά ηφαίστεια στον κόσµο βρίσκονται σε νησιωτικά τόξα τα οποία
έχουν σχηµατισθεί κοντά σε βαθιές τάφρους. Η µορφή, που τυπικά φαίνεται σ' αυτές
τις περιοχές, είναι µια σειρά από ηφαιστειογενή νησιά επί ενός τόξου και µια βαθιά
τάφρος ευρισκόµενη από την πλευρά των νησιών προς την ωκεάνιο λεκάνη (Εικ.

17
1.9). Οι τάφροι αυτοί καθαυτοί είναι στενά παράλληλοι προς το τόξο των νησιών.
Σπάνια είναι φαρδύτεροι από 130 χλµ αλλά το µήκος τους µπορεί να είναι µέχρι 1500
χλµ και το βάθος τους περισσότερο από 11.000 µ.
Εικ. 1.9 Παγκόσµιος χάρτης περιοχών τάφρων (τροποποιηµένη, από B. Gutenberg and C. F. Richter,
1954)
Οι περισσότεροι σχηµατισµοί του τύπου νησιωτικό-τόξο / τάφρος περιβάλλουν τον
Ειρηνικό ή διαχωρίζουν τον Ειρηνικό από τον Ινδικό Ωκεανό. Εξαιρέσεις
περιλαµβάνουν τις ∆υτικές Ινδίες όπου ένα νησιωτικό τόξο διαχωρίζει την Καραϊβική
θάλασσα από τον Ατλαντικό Ωκεανό, καθώς και το νησιωτικό τόξο των νησιών
Sandwich ανοικτά και Ν.Α του άκρου της Ν. Αµερικής. ∆οθέντος ότι πολλά από τα
νησιά αυτά είναι ηφαιστειακά ενεργά, τα όρια του Ειρηνικού Ωκεανού καλούνται
«πύρινος κύκλος» (the ring of fire) (Εικ. 1.10).

18
Εικ. 1.10 Παγκόσµιος χάρτης κατανοµής σεισµικών επίκεντρων για την περίοδο 1963-1977
(τροποποιηµένη, από D. E. Ingmason and W. J. Wallace, 1989)
Σπάνια περνάει χρόνος χωρίς να συµβεί µια σηµαντική έκρηξη ηφαιστείου σ' ένα
τουλάχιστον από τα νησιά που οριοθετούν τον Ειρηνικό. Συχνά οι ηφαιστειακές
εκτινάξεις είναι εκρηκτικές, παράγοντας τεράστιες ποσότητες λάβας και σύννεφα από
ατµούς ύδατος και άλλα αέρια.
Τα νησιωτικά τόξα και οι τάφροι είναι θέσεις συχνών σεισµών καθώς και εκρήξεων
ηφαιστείων. Η συχνότητα αυτών των συµβάντων είναι η ίδια περίπου µε εκείνη επί
των ωκεάνιων οροσειρών.
1.3.3 Υποθαλάσσια υψίπεδα
Ο σχετικά επίπεδος ωκεάνιος πυθµένας διακόπτεται συχνά από ανυψωµένες
περιοχές που ανέρχονται σε ύψη 1 έως 2 χλµ. Οι περιοχές αυτές δεν φθάνουν στην
επιφάνεια της θάλασσας και δεν αποτελούν µέρος της ωκεάνιας ράχης. Τα
υποθαλάσσια αυτά υψίπεδα (plateaus) αποτελούνται από υλικά που έχουν
µεγαλύτερη άνωση από εκείνα του τοπικού ωκεάνιου φλοιού. Μερικά υψίπεδα είναι
ηπειρωτικής ενώ άλλα είναι ηφαιστειακής προέλευσης. Τα υψίπεδα µεταφερόµενα
µαζί µε τον ωκεάνιο πυθµένα στις ζώνες καταβύθισης δεν βυθίζονται, όπως εκείνος
αλλά «επιπλέουν» προσαρτηµένα στο γειτονικό περιθώριο (Εικ.1.11).

19
Εικ. 1.11 Τα ωκεάνια υψίπεδα που µεταφέρονται µαζί µε τον ωκεάνιο φλοιό στις ζώνες καταβύθισης,
δεν βυθίζονται αλλά συγκρούονται µε τον ηπειρωτικό φλοιό , στον οποίο και προσαρτώνται (από Α.
Θεοδώρου, 1989)
1.4 Ωκεάνια ιζήµατα
Τα υλικά που αποτελούν τον φλοιό και την υδρόσφαιρα οφείλονται στην γεωλογική
δραστηριότητα της Γης. Μεταξύ υδρόσφαιρας και φλοιού υπάρχει συνεχής και έντονη
αλληλεπίδραση, µε αποτέλεσµα την αποσάθρωση της επιφάνειας του φλοιού και την
κάλυψή του µε ιζήµατα. Τα ιζήµατα καλύπτουν το σύνολο της επιφάνειας της γης
(Πίνακας 1.1).

20
ΠΙΝΑΚΑΣ 1.1 Κατανοµή κατά πάχος των ιζηµάτων
Περιοχή Επιφάνεια Γης (%) Ιζήµατα (%) Μέσο πάχος (χλµ)
Ήπειροι 29 8 0,3
Υφαλοκρηπίδες 6 14 2,5
Κατωφέρειες 4 38 9,0
Υψώµατα 4 28 8,0
Πυθµένας 56 12 0,2
(Πηγή : Stowe,1987)
Από γεωτρήσεις και σεισµικές διασκοπήσεις (seismic echo profiles) έχει υπολογισθεί
ότι το µέσο πάχος των ιζηµάτων είναι περίπου 1 χλµ. Τα ιζήµατα παρουσιάζουν
ελάχιστο πάχος στον πυθµένα των ωκεανών, λόγω της νεαράς ηλικίας του ωκεάνιου
φλοιού, και µέγιστο πάχος στην ηπειρωτική υφαλοκρηπίδα και κατωφέρεια, στις
οποίες συσσωρεύονται τα ιζήµατα που προέρχονται από την διάβρωση της
παρακείµενης Ηπείρου. Ιζήµατα σπανίζουν ή και απουσιάζουν εντελώς από περιοχές
νεαρών σε ηλικία και µε απότοµο ανάγλυφο βουνών. Σε αντίθεση, στις
υφαλοκρηπίδες – που βρίσκονται στην οπίσθια πλευρά των µετακινούµενων µαζί µε
τον ωκεάνιο πυθµένα ηπείρων - το πάχος των ιζηµάτων συχνά υπερβαίνει τα 15
χλµ.. Εντός των ιζηµάτων υπάρχουν κοιτάσµατα υδρογονανθράκων και άλλων
ορυκτών κοιτασµάτων.
1.4.1 Φυσικοί παράγοντες ιζηµατογένεσης
Ο χαρακτήρας των θαλάσσιων ιζηµάτων στον πυθµένα των ωκεανών δεν
καθορίζονται µόνο από την πηγή προέλευσής τους. Οι φυσικοί παράγοντες που
επηρεάζουν την ιζηµατογένεση έχουν σχέση µε τη φυσική κατάσταση του ωκεάνιου
πυθµένα, τη φυσική κατάσταση του νερού και τις κλιµατικές συνθήκες που
επικρατούν κατά την διάρκεια του έτους. Η κλίση, το σχήµα και το µέγεθος της
ωκεάνιας λεκάνης έχουν µια απ’ ευθείας επίδραση στην κατεύθυνση, ταχύτητα και
όγκο του νερού που µεταφέρεται µε τα ρεύµατα και κατά συνέπεια επί των υλικών
που καθιζάνουν. Η θερµοκρασία και η πίεση του θαλασσινού νερού είναι σηµαντικοί
παράγοντες διότι καθορίζουν την ποσότητα αλάτων που κατακρατούνται σε διάλυση.
Οι ατµοσφαιρικές συνθήκες επηρεάζουν τις φυσικές ιδιότητες του θαλασσινού νερού
καθώς και την ωκεάνια κυκλοφορία, η οποία µε τη σειρά της καθορίζει την µεταφορά
των ιζηµάτων στον ωκεανό.

21
Ο µεγαλύτερος όγκος υλικών µεταφέρεται από την ξηρά µε τα ρυάκια, ποτάµια και
παγετώνες. Άλλος σηµαντικός µηχανισµός µεταφοράς είναι η συνδυασµένη ενέργεια
των κυµάτων και ρευµάτων. Αυτοί οι δύο φυσικοί παράγοντες µεταφέρουν υλικά
κάπως διαφορετικά από εκείνα που µεταφέρουν οι άλλοι φυσικοί παράγοντες.
Ειδικότερα, τα υλικά που µεταφέρουν βρίσκονται µάλλον µέσα στον ωκεανό παρά
έξω απ’ αυτόν.
1.4.2 Υφή των ιζηµάτων
Τα ιζήµατα παρουσιάζουν µεγάλη ποικιλία υφής και σύστασης. Το µέγεθος των
κόκκων τους κυµαίνεται από γιγαντιαίους ογκόλιθους µέχρι λεπτόκοκκο άργιλο. Το
χρώµα των ιζηµάτων παρουσιάζει µεγάλη ποικιλία από την λευκή ή κίτρινη άµµο έως
τη σκουροκόκκινη ή καφετιά ιλύ. Η υφή τους χαρακτηρίζεται από εκπληκτική ποικιλία
µεγεθών, σχηµάτων και χρωµάτων.
Τα ιζήµατα υποβάλλονται σε διάφορες φυσικές διεργασίες, όπως διαλογή, δηλαδή σε
διαχωρισµό ανάλογα µε το µέγεθος των κόκκων και την σύσταση τους. Οι περιοχές
όπου συµβαίνει αυτό είναι συνήθως οι ακτές. Εκεί η δράση των κυµάτων συνεχώς
διαχωρίζει τα υλικά µιας ακτής. Βραχώδεις ακτές συναντώνται σε περιοχές
εκτεθειµένες σε ισχυρό κυµατισµό, όπως π.χ. ακρωτήρια ή περιοχές µεγάλων
γεωγραφικών πλατών µε συχνές χειµερινές θύελλες. Σε περιοχές µε µέτριο
κυµατισµό, όπως π.χ. στα µεσαία γεωγραφικά πλάτη, απαντώνται αµµώδεις ακτές µε
ενδιάµεσο µέγεθος κόκκων. Σε ακτές προστατευµένες από τον κυµατισµό, όπως π.χ.
κλειστοί κόλποι και λιµνοθάλασσες, συναντούνται λασπώδεις αποθέσεις
λεπτόκοκκου αργίλου. Οι περισσότερες ακτές χαρακτηρίζονται από οµοιόµορφη
άµµο, που είναι αποτέλεσµα της συνεχούς διαλογής και επεξεργασίας των κόκκων
από τα κύµατα.
Τα ιζήµατα βαθέων υδάτων προέρχονται από πολλές πηγές, όπως κοχύλια και
σκελετοί οργανισµών, που βυθίζονται από τα επιφανειακά νερά, ηφαιστειογενή υλικά,
τόσο χερσαίας προέλευσης όσο και υποθαλάσσιων ηφαιστείων, αυτογενείς
µεταλλικές αποθέσεις ή κατακρηµνίσεις, που σχηµατίζονται από αλληεπιδρώντα
υλικά διαλυµένα στο θαλασσινό νερό, και ακόµη εξωγήινα υλικά που προέρχονται
από τη συνεχή βροχή µετεωριτών κατά τη διάρκεια των αιώνων. Κοντά στην ξηρά,
ειδικότερα κοντά σε ακτές µε εκβολές µεγάλων ποταµών ή πάγων, τα ιζήµατα είναι
κυρίως χερσογενούς χαρακτήρα.
Υπήρχε µέχρι τώρα η σκέψη ότι τα ιζήµατα των βαθέων υδάτων είχαν καθ’
ολοκληρίαν σχηµατισθεί από υλικά που καταπίπτουν από πάνω. Εντούτοις, µερικά
από τα υλικά, όπως σκελετικοί σχηµατισµοί σπόγγων, αγκάθια εχινοδέρµων,

22
ασβεστολιθικά ή πυριτιούχα κοχύλια από τρηµατοφόρα, τεµάχια κοχυλιών και
ακτινόζωα, ακόµη και κελύφη διατόµων, µπορεί να είναι αυτόχθονα υλικά που έχουν
συγκεντρωθεί από οργανισµούς που ζουν στο ίζηµα από υλικά διαλυµένα στο νερό.
Ακόµη µορφές που κανονικά ζουν φωτοσυνθετικά, όπως τα διάτοµα, µπορεί να ζουν
ετεροτροφικά σε βαθιά ιζήµατα. Ο Woοd (1965) κατέγραψε 10 γένη διατόµων που
βρέθηκαν σε βάθη 7.000 µέχρι 10.000 µ. και αναφέρει ότι πειραµατικά έχουν
αναπτυχθεί διάτοµα στο σκοτάδι κάτω από πίεση 500 ατµόσφαιρες, όπου
αναπτύσσονται και αναπαράγονται, παρουσία γλυκόζης, κάτω απ' αυτές τις
συνθήκες. Μικροβιακές διαδικασίες, µε εξαίρεση τη φωτοσύνθεση, µπορούν να
λαµβάνουν χώρα ακόµη και στα µεγαλύτερα βάθη.
1.4.3 Χαρακτηριστικά των ωκεάνιων ιζηµάτων
Οι Ωκεανογράφοι χρησιµοποιούν διάφορες µεταβλητές για την κατάταξη των
ωκεάνιων ιζηµάτων: µέγεθος και σχήµα των κόκκων, πυκνότητα, χρώµα, σύνθεση
και χαρακτήρες εναπόθεσης επί του πυθµένος. Με τα δεδοµένα αυτά οι
ωκεανογράφοι µπορούν να γνωρίζουν τον τρόπο διασποράς και την ηλικία των
ιζηµάτων και απ' αυτά την ιστορία των ωκεάνιων λεκανών.
(α) Μέγεθος κόκκων
Οι Γεωλόγοι κατατάσσουν τα ιζήµατα σύµφωνα µε το µέγεθος των κόκκων που τα
αποτελούν. Χρησιµοποιούν την κλίµακα Wentwoth (Πίνακας 1.3) για να κατατάξουν
τόσο τα χερσαία όσο και τα ωκεάνια ιζήµατα. Τα πιο κοινά συστατικά των
θαλάσσιων ιζηµάτων είναι η άµµος, η λάσπη και ο πηλός.
Η κατανοµή των διαφόρων µεγεθών κόκκων επιτρέπει στους ερευνητές την
αναγνώριση τόσο της καταγωγής του ιζήµατος όσο και του τρόπου µεταφοράς του
στην παρούσα θέση. Π.χ. θαλάσσια ιζήµατα µε διάφορα µεγέθη κόκκων, τα οποία
δεν είναι καλά διαχωρισµένα ανάλογα µε το µέγεθός τους, δίδουν σαφή ένδειξη ότι
µεταφέρθηκαν µε τουρβιδιτικά ρεύµατα ή µε παγετώνες. Ιζήµατα των οποίων οι
κόκκοι είναι του ιδίου µεγέθους αποδεικνύουν µεταφορά µε άνεµο ή κύµατα.

23
ΠΙΝΑΚΑΣ 1.2 Κατάταξη κατά µέγεθος των σωµατιδίων ιζηµάτων σύµφωνα µε την
κλίµακα Wentworth
Είδος Σωµατιδίου ∆ιάµετρος Σωµατιδίου (χλστ)
Τρόχαλο > 256
Κροκάλη, βότσαλο 64-256
Χαλίκι 4-64
Κόκκος 2-4
Αµµος 0,062-2 (62-2.000 µm)
Ιλύς 0,004-0,062 (4-62 µm)
Πηλός, άργιλος <0,004 (<4 µm)
(β) Πυκνότητα και σχήµα
Η πυκνότητα και το σχήµα των κόκκων παρέχουν ενδείξεις για την καταγωγή και την
δύναµη του µέσου µεταφοράς. Ορυκτά διαφορετικής σύνθεσης έχουν διαφορετικές
πυκνότητες. Η παρουσία κόκκων διαφορετικών πυκνοτήτων αποκαλύπτει ότι
προέρχονται από διαφορετικές πηγές και έχουν µεταφερθεί µε µέσα διαφορετικού
βαθµού ενέργειας και ότι έχουν καταπέσει στον πυθµένα µε διαφορετικό βαθµό
πτώσης.
Το σχήµα των κόκκων επίσης φανερώνει σηµαντικές πληροφορίες. Π.χ. ένας
κυκλικός κόκκος και ένας επιµήκης κόκκος, και οι δύο της ίδιας σύνθεσης, θα πέσουν
στον πυθµένα µε διαφορετικούς βαθµούς καταβύθισης και έχουν µεταφερθεί από
ρεύµατα από διαφορετικές αποστάσεις. Οµοιόµορφα κυκλικά σωµατίδια
φανερώνουν ότι έχουν µεταφερθεί από µεγάλες αποστάσεις και έχουν διαχωριστεί
από τον άνεµο ή τα κύµατα.
(γ) Χρώµα
Τα θαλάσσια ιζήµατα είναι διαφόρων χρωµάτων ανάλογα µε την σύνθεση των
ορυκτών που τα αποτελούν. Λευκό γενικά φανερώνει ανθρακικό ασβέστιο. Μαύρο
φανερώνει πολύ συχνά την παρουσία θειούχων ενώσεων σιδήρου ή µαγγανίου.
Κίτρινο φανερώνει παρουσία σιδήρου σε µορφή λεµονίτη ή υδροξειδίου. Καφετί
προς το κόκκινο, φανερώνει την παρουσία οξειδίου του σιδήρου στη µορφή του
αιµατίτη. Τέλος µαύρο συνήθως φανερώνει περιβάλλον µε παντελή έλλειψη

24
οξυγόνου, ενώ κίτρινο, καφέ και κόκκινο συνήθως φανερώνουν περιβάλλον που έχει
οξειδωθεί.
(δ) Ορυκτολογική σύνθεση
Τα ορυκτά που συνήθως συναντώνται στα θαλάσσια ιζήµατα κατά σειρά αφθονίας
είναι: πυριτικά, ανθρακικά, οξείδια και υδροξείδια, φωσφορικά και θειικά.
Τα πυριτικά ορυκτά περιέχουν πυρίτιο και οξυγόνο καθώς και άλλα στοιχεία σε
µικρές ποσότητες. Τα πιο κοινά πυριτικά ορυκτά είναι οι άστριοι, αµφιβολίτες,
πυρόξενοι, olivine, χαλαζίας, οπάλλιον, µαρµαρυγίας, αργίλλιον και ζεόλιθος.
Τα ανθρακικά ορυκτά περιέχουν άνθρακα και οξυγόνο. Στα ωκεάνια ιζήµατα, τα
ανθρακικά ορυκτά συχνά περιέχουν ασβέστιο ή και µαγνήσιο για να σχηµατίσουν τα
ορυκτά ασβεστίτη, αραγωνίτη και δολοµίτη. Τα οξείδια και υδροξείδια µετάλλων είναι
ορυκτά τα οποία περιέχουν αρνητικά φορτισµένες ενότητες αποτελούµενες από Ο-2 ή
ΟΗ-1. Τα πιο κοινά από τα ορυκτά αυτά είναι ο αιµατίτης (οξείδιον του σιδήρου),
γκοεθίτης (ένυδρον οξείδιο του σιδήρου) και υδροξείδια µαγνησίου-σιδήρου. Τα πιο
συνηθισµένα φωσφορικά ορυκτά στα θαλάσσια ιζήµατα είναι ο απατίτης. Τα θειικά
ορυκτά είναι λιγότερο κοινά στα θαλάσσια ιζήµατα διότι η θειική οµάδα SO4-2 έχει την
τάση να διαλύεται στο θαλασσινό νερό.
(ε) Πάχος ιζηµάτων και βαθµός εναπόθεσης
Το πάχος των ιζηµάτων προσδίδει ενδείξεις καταγωγής και ηλικίας ακόµη και της
ιστορίας της ωκεάνιας λεκάνης. Κοντά στις ηπείρους τα ιζήµατα µπορεί να έχουν
πάχος και µέχρι 20 χλµ. Σε περιοχές κοντά σε εκβολές µεγάλων ποταµών, τα
ιζήµατα µπορεί να συσσωρεύονται µε βαθµό αρκετών µέτρων ανά έτος.
Το στροβιλικό ρεύµα είναι ροή νερού φορτωµένου µε ιζήµατα, που συµβαίνει όταν,
µια ασταθής µάζα ιζήµατος στο άνω µέρος µιας σχετικά απότοµης κλίσης
υποθαλάσσιου νερού, αραιώνει πέφτοντας και γλιστράει κατά µήκος της κλίσης.
Καθώς η ολίσθηση συνεχίζει κατά µήκος της κλίσης συνεχώς περισσότερο νερό
αναµιγνύεται µε τη µάζα αυξάνοντας έτσι τη ρευστότητα της αρχικής µάζας νερού και
ιζήµατος. Τα ρεύµατα αυτά φθάνουν σε υψηλές ταχύτητες επιτρέποντας έτσι να
διασπείρονται σε τεράστιες αποστάσεις κατά µήκος των αβυσσαίων πεδιάδων.
Στροβιλικά ρεύµατα προκαλούνται από σεισµούς ή θύελλες. Καθώς τα µεγάλα
στροβιλικά ρεύµατα χάνουν ενέργεια, το ίζηµα αποτίθεται επί του θαλάσσιου
πυθµένος και σε λίγες ηµέρες είναι δυνατόν να σχηµατισθούν στρώµατα ιζήµατος
πάχους αρκετών µέτρων.

25
Σε βαθύτερα νερά, µακριά από το σηµείο που φθάνουν τα στροβιλικά ρεύµατα,
ανόργανη λάσπη µαζί µε κελύφη πλαγκτικών οργανισµών, µεταφερόµενα µε τους
ανέµους και τα ρεύµατα, βυθίζονται σιγά-σιγά στον ωκεάνιο πυθµένα για να
σχηµατίσουν ιζήµατα των βαθιών θαλασσών. Η λάσπη αυτή συγκεντρώνεται πολύ
αργά µε βαθµούς απόθεσης λιγότερο από 2 χλστµ κάθε 1000 χρόνια. Σε περιοχές
όπου το επιφανειακό πλαγκτόν είναι άφθονο, κελύφη πλαγκτού µπορεί να
συσσωρεύονται µε βαθµό 20 χλστµ κάθε 1000 χρόνια.
Σε µεγάλες περιοχές του θαλάσσιου πυθµένα, ο βαθµός εναπόθεσης έχει παραµείνει
σταθερός για εκατοµµύρια χρόνια. Έτσι, το πάχος αυτών των ιζηµάτων δίνει µια
ένδειξη της ηλικίας τους.
1.4.4 Κατάταξη των ωκεάνιων ιζηµάτων
Στα 1891, ο Sir John Murray και ο A.E. Renard, στην πρώτη κύρια µελέτη των
ωκεάνιων ιζηµάτων, δηµοσίευσαν έκθεση για τα δείγµατα που συνέλεξαν κατά τη
διάρκεια της ερευνητικής αποστολής του Challenger (1872 - 1876). Στην έκθεση
αυτή οι ανωτέρω ερευνητές πρότειναν το παρακάτω σύστηµα κατάταξης των
ωκεάνιων ιζηµάτων, το οποίο παραµένει χρήσιµο µέχρι και σήµερα: Τα πελαγικά
ιζήµατα, προέρχονται από διαδικασίες που συµβαίνουν µέσα στον ωκεανό και
µακριά από τις ηπείρους, τα χερσαία ιζήµατα, προέρχονται από πηγές ξηράς και τα
κοσµογενή ιζήµατα είναι εξωγήινης προέλευσης.
(α) Πελαγικά ιζήµατα
Ανάλογα µε την προέλευσή τους διακρίνονται σε Βιογενή, Πελαγική Λάσπη,
Αυτόχθονα ιζήµατα και Ηφαιστειακές Αποθέσεις.
Βιογενή : Μια σηµαντική πηγή ιζηµάτων των µεγάλων βαθών των ωκεανών είναι τα
σκελετικά υπολείµµατα µικροσκοπικών θαλάσσιων οργανισµών. Τα ιζήµατα αυτά
ονοµάζονται Βιογενή (Εικ.1.12). Στα βαθιά και σκοτεινά ωκεάνια νερά υπάρχει
σχετικά µικρή βιολογική δραστηριότητα αλλά το ίδιο ισχύει και για τις κεντρικές
περιοχές των ωκεανών, που φωτίζονται από τις ηλιακές ακτίνες. Γενικά τα νερά των
ανοικτών ωκεανών είναι πολύ λιγότερο παραγωγικά από εκείνα των ηπειρωτικών
υφαλοκρηπίδων. Έτσι η συσσώρευση των βιογενών ιζηµάτων στα ωκεάνια βάθη
γίνεται µε πολύ βραδείς ρυθµούς. Επιπλέον, τα σκελετικά υπολείµµατα διαλύονται
µερικώς στο θαλάσσιο νερό, µειώνοντας έτσι ακόµη περισσότερο το ρυθµό
συσσώρευσης τους σε ορισµένες περιοχές, ενώ σε άλλες δεν αποτίθενται καθόλου.

26
Εικ. 1,12 Βιογενή ιζήµατα: (α) κελύφη ακτινοζώων από βαθιές θάλασσες (β) τρηµατοφόρα (γ)
ακτινόζωα (τροποποιηµένη, από D. E. Ingmason and W. J. Wallace, 1989)
Παρ’ όλα αυτά επειδή οι ποσότητες των προερχόµενων από άλλες πηγές ιζηµάτων
είναι πολύ µικρές, τα βιογενή ιζήµατα αποτελούν µια σηµαντική συνιστώσα των
ωκεάνιων ιζηµάτων.
Σε µεγάλες περιοχές του ωκεάνιου πυθµένα, τα ιζήµατα περιέχουν σκελετικό υλικό.
∆ισεκατοµµύρια µικροοργανισµών ζουν κοντά στην επιφάνεια των ωκεανών και µέσα
στη µάζα του θαλασσινού νερού. Με την πάροδο αιώνων αύξησης, αναπαραγωγής
και θανάτων, τα κελύφη και άλλα σκληρά µέρη αυτών των οργανισµών, σιγά-σιγά
συσσωρεύονται στην επιφάνεια του ωκεάνιου πυθµένα. Οι πιο κοινοί τύποι τέτοιων
οργανισµών είναι τα Τρηµατοφόρα, Ακτινόζωα, Πτερόποδα, ∆ιάτοµα και
Κοκκολιθοφόρα.
Τρηµατοφόρα : Είναι µονοκύτταρα µέλη του βασιλείου των Πρωτίστων και έχουν
µήκος µέχρι 2 χλστµ. Αν και συγγενεύουν µε τις αµοιβάδες έχουν προστατευτικά
κελύφη. Για την παγίδευση της τροφής τους προβάλλουν, µέσα από τις µικρές
τρύπες των κελυφών τους, µικρές κολλώδεις προεκτάσεις (Εικ.1.12β). Οι οργανισµοί
αυτοί εκκρίνουν ανθρακικό ασβέστιο για το σχηµατισµό των κελυφών τους. Τα
περισσότερα από τα τρηµατοφόρα κελύφη, που βρίσκονται στα ιζήµατα των
αβυσσαίων πεδιάδων, είναι κελύφη πλαγκτού, µικροσκοπικών οργανισµών που
επιπλέουν στα επιφανειακά 100 µ των ωκεανών και µετακινούνται µε επιφανειακά
ρεύµατα. Μερικοί πλαγκτικοί οργανισµοί επίσης ζουν επί της υφαλοκρηπίδας από
την οποία τουρβιδιτικά ρεύµατα τους µεταφέρουν κάτω στα υποθαλάσσια φαράγγια
και από εκεί στις αβυσσαίες πεδιάδες. Αυτά δεν αποτελούν κάποιο σηµαντικό

27
συστατικό των ιζηµάτων επί της υφαλοκρηπίδας δεδοµένου ότι στις περιοχές αυτές
αφθονούν τα χερσαία ιζήµατα.
Εικ. 1.13 (α) Ακτινόζωο. Μοντέλο από πυριτιούχο σκελετό µιας αποικίας ακτινοζώων (τροποποιηµένη,
από D. E. Ingmason and W. J. Wallace, 1989) (β) Ποικιλία διατόµων (τροποποιηµένη, από Weihaupt,
1979)
Ακτινόζωα : Είναι πλαγκτικοί οργανισµοί, οι οποίοι έχουν κελύφη από πυρίτιο στη
µορφή του οπαλλίου (ένυδρη ποικιλία του χαλαζία). Τα κελύφη των ακτινόζωων
έχουν επιµήκεις εύθραυστες βελόνες ακτινοειδώς προβαλλόµενες από ένα κεντρικό
χώρο (Εικ. 1.12γ και 1.13α). Κι αυτά συσσωρεύονται επί των αβυσσαίων πεδιάδων
µε τον ίδιο τρόπο όπως και τα τρηµατοφόρα.
∆ιάτοµα: Μονοκύτταροι φωτοσυνθετικοί οργανισµοί µε πυριτιούχα σκληρά τµήµατα.
Συγκαταλέγονται µεταξύ των πιο κοινών φυτοπλαγκτικών οργανισµών τόσο στο
γλυκό όσο και στο θαλασσινό νερό (Εικ.1.13β).
Κοκκολιθοφόρα : Μικροσκοπικά φυτά τα οποία είναι πολύ κοινά στα ιζήµατα των
βαθιών θαλασσών (Εικ. 1.14α). Τα σκληρά τους, κελύφη αποτελούνται από
ανθρακικό ασβέστιο.
Πτερόποδα: Γαστερόποδα τα οποία ζουν σε πελαγικό περιβάλλον (Εικ. 1.14β ).
Επειδή τα περισσότερα των πτεροπόδων εκκρίνουν µια πολύ διαλυτή µορφή
ανθρακικού υλικού (περισσότερο αραγωνίτης και λιγότερο ασβεστίτης) η ποσότητα
της ιλύος των πτεροπόδων είναι περιορισµένη σε σχετικά όχι πολύ βαθιά νερά (µέσο
βάθος 2.070µ) σε σε τµήµα της Μεσο-Ατλαντικής Ράχης κοντά στις 20ο Ν. Πλάτος,
την Ερυθρά Θάλασσα, τον Ισηµερινό, Ν. Ειρηνικό και Καραιβική Θάλασσα. Μικρές
α

28
ποσότητες υπολειµµάτων εχινοδέρµων, οστρακοειδών και αννελιδών µπορεί να
συµµετέχουν επίσης στο ανθρακικό υλικό αυτών των ιζηµάτων.
Εικ. 1.14 (α) Κοκκολιθοφόρα (τροποποιηµένη από Grasse, 1952) (β) πτερόποδα (τροποποιηµένη, από
Hardy, The Open Sea)
Πελαγική Λάσπη : Η πελαγική λάσπη αποτελείται από µικρά (µικρότερα από 20 µm)
ορυκτά σωµατίδια. Μερικά από τα σωµατίδια µεταφέρονται µε τον άνεµο από την
ξηρά. Είναι συνήθως άργιλος ή χαλαζίας. Τα ιζήµατα αυτά περιέχουν επίσης
αυτόχθονα ορυκτά, κοσµογενή σκόνη σιδήρου-νικελίου και µερικές φορές δόντια
ψαριών.
Αυτόχθονα Ιζήµατα : Τα αυτόχθονα ιζήµατα είναι κοινά στις βαθιές θάλασσες.
Σχηµατίζονται στο θαλασσινό νερό ή µέσα στο ίζηµα. Παραδείγµατα τέτοιων
ιζηµάτων είναι οξείδια του σιδήρου και οι ζεόλιθοι. Οι ζεόλιθοι είναι ένυδροι πυριτικοί
κρύσταλλοι για τους οποίους υπάρχει η άποψη ότι έχουν προέλθει από ηφαιστειακό
γυαλί που περιέχει µεγάλες ποσότητες µαγνησίου και σιδήρου και έχει κρυσταλλωθεί
στο θαλασσινό νερό, το οποίο µετέβαλε το γυαλί σε ζεόλιθο. Είναι ιδιαίτερα άφθονοι
στον Ειρηνικό Ωκεανό. Άλλα αυτόχθονα ορυκτά είναι το οξείδιον του βαρίου (βαρία),
οι κόνδυλοι µαγγανίου, οι φωσφορίτες και οι θειούχες λάσπες των υποθαλάσσιων
πηγών. Τα αυτόχθονα ή αυτογενή ή υδρογενή ιζήµατα δηµιουργούνται κατά
διάφορους τρόπους, οι οποίοι δεν είναι απολύτως γνωστοί. Σχηµατίζονται κυρίως
όταν λόγω εξάτµισης αποµακρυνθεί ένα σηµαντικό µέρος νερού, προκαλώντας
αύξηση της συγκέντρωσης των διαλυµένων αλάτων, προκαλώντας έτσι την καθίζηση

29
µερικών απ’ αυτά. Τέτοια ιζήµατα είναι οι εβαπορίτες που σχηµατίζονται σε ρηχές
θάλασσες µε περιορισµένη επικοινωνία µε τον ανοικτό ωκεανό ή και σε λίµνες. Τέτοιο
ίζηµα π.χ. είναι το επιτραπέζιο αλάτι. Ειδικότερα οι κόνδυλοι µαγγανίου (Εικ.1.15)
είναι πολύ ευρέως διασκορπισµένοι επί του θαλάσσιου πυθµένα. Οι κόνδυλοι αυτοί
περιέχουν περίπου 65% διοξείδιο του µαγγανίου, 30% οξείδιο του σιδήρου, λιγότερο
από 5% οξείδια του νικελίου, χαλκού και κοβαλτίου καθώς και ίχνη άλλων ορυκτών.
Οι κόνδυλοι του µαγγανίου παρουσιάζουν εξαιρετικό ενδιαφέρον από οικονοµική
άποψη. Υπάρχουν αρκετές υποθέσεις σχετικά µε τη δηµιουργία τους. Μια θεωρία
υποστηρίζει ότι τα συστατικά τους προέρχονται από χερσαία πετρώµατα και ότι
έχουν µεταφερθεί στη θάλασσα ως διάλυµα για να κατακαθίσουν τελικά επί του
θαλάσσιου πυθµένα. Άλλη θεωρία υποστηρίζει ότι ο σίδηρος, το µαγγάνιο, ο χαλκός
και το νικέλιο που περιέχουν, έχουν προέλθει από θαλάσσια ιζήµατα κάτω από
συνθήκες αναγωγής των ιζηµάτων και µετά µετανάστευσαν στον θαλάσσιο πυθµένα
όπου οξειδώθηκαν και κατακάθισαν. Μια τρίτη θεωρία προτείνει την άποψη ότι τα
συστατικά των κονδύλων έχουν ελευθερωθεί από µεταλλιούχες θερµές πηγές σε
µεσο-ωκεάνιες οροσειρές και ότι έχουν µεταφερθεί από ρεύµατα κατά µήκος του
θαλάσσιου πυθµένα όπου σιγά-σιγά οξειδώνονται και κατακαθίζουν. Πρόσφατες
έρευνες υποστηρίζουν αυτή την άποψη. Μια πιθανή εξήγηση είναι ότι µικροί βενθικοί
οργανισµοί χρησιµοποιούν τους κονδύλους µαγγανίου για προστασία, ζώντας κάτω
απ’ αυτούς και διατηρώντας τους στην επιφάνεια του πυθµένα. Επίσης έχει
διατυπωθεί η άποψη ότι καθοριστικό ρόλο στον σχηµατισµό κονδύλων µαγγανίου
παίζουν µικροοργανισµοί και κυρίως βακτήρια κοντά σε θερµές πηγές. Από την
κατανοµή των κοιτασµάτων κονδύλων µαγγανίου στον παγκόσµιο ωκεανό, φαίνεται
ότι η περιοχή µε την µεγαλύτερη οικονοµική σηµασία βρίσκεται στον Βόρειο
Ισηµερινό Ειρηνικό.
Ηφαιστειακές Αποθέσεις : Εκρήξεις ηφαιστείων αποβάλλουν τεράστιες ποσότητες
στάχτης στην ατµόσφαιρα απ' όπου οι άνεµοι τις µεταφέρουν γύρω από τη γη. Η
στάχτη καθώς πέφτει πάνω στην επιφάνεια των ωκεανών και κατακαθίζει πάνω στον
πυθµένα µετατρέπεται σε πυριτιούχα λάσπη. Τέτοιες αποθέσεις βρίσκονται κυρίως
προς την κατεύθυνση του ανέµου από το σηµείο έκρηξης του ηφαιστείου. Άλλες
αποθέσεις είναι αποτέλεσµα της ροής υποθαλάσσιας λάβας η οποία πήγνυται πάνω
στο θαλάσσιο πυθµένα. Οι ηφαιστειακές αυτές αποθέσεις είναι ιδιαίτερα κοινές στις
µεσο-ωκεάνιες ράχες και κατά µήκος των υποθαλάσσιων πλευρών των
ηφαιστειογενών νησιών, όπως τα νησιά της Χαβάης.

30
Εικ. 1.15 Κύκλοι αύξησης κονδύλου µαγγανίου σε µεγέθυνση περίπου πέντε φορές (από παρατηρήσεις
στο Scripps Institution of Oceanography, University of California)
(β) Χερσογενή ιζήµατα
Τα ιζήµατα κοντά στην ξηρά και οι βαθµοί µε τους οποίους τα ιζήµατα αυτά
συσσωρεύονται είναι περισσότερο ποικιλόµορφα από εκείνα των πραγµατικά
ωκεάνιων περιοχών. Αυτά τα ιζήµατα, τα οποία είναι κυρίως χερσαίας προέλευσης,
αποτελούνται από ιλύ, άµµο, χαλίκια, στάχτη ηφαιστείων και άλλα υλικά που
εκβάλλουν από την ξηρά σε γειτονικές θάλασσες και συχνά µεταφέρονται κατά µήκος
της ακτογραµµής µε ρεύµατα, ανέµους και τη δράση των κυµάτων. Τα ανωτέρω
ιζήµατα αναµιγνύονται κατά διάφορους βαθµούς µε υλικά ωκεάνιας προέλευσης. Τα
µοντέλα ιζηµατογένεσης επηρεάζονται πάρα πολύ από ρεύµατα και άλλες κινήσεις
του νερού, οι οποίες επίσης ασκούν και µια σηµαντική επίδραση στη διαλογή των
κόκκων του ιζήµατος. Τα βαρύτερα αντικείµενα και τα υλικά σε µεγάλους κόκκους
εναποτίθενται πλησιέστερα στα σηµεία προέλευσής τους και µεταφέρονται γενικά σε
µικρότερες αποστάσεις και περισσότερο αργά, ενώ ελαφρότερα αντικείµενα και υλικά
σε µικρούς κόκκους µπορεί να µεταφερθούν σε µεγαλύτερες αποστάσεις και σε
διαφορετικές διευθύνσεις, ανάλογα µε τα ρεύµατα, τις πυκνότητες του νερού στα
διάφορα βάθη και το µέγεθος, σχήµα και πυκνότητα των κόκκων. Σε µερικές
περιοχές µεγαλύτερα και βαριά χερσαία αντικείµενα, όπως π.χ. τρόχαλα ή χαλίκια
µπορεί να ενσωµατώνονται στους πάγους, οι οποίοι µε την σειρά τους συµµετέχουν
στο σχηµατισµό παγόβουνων µέσα στο νερό και µεταφέρονται µε τα παγόβουνα

31
µέχρι του σηµείου εκείνου που λειώνουν οι πάγοι και ελευθερώνουν τα δεσµευµένα
υλικά.
Το µεγαλύτερο µέρος των χερσογενών ιζηµάτων µεταφέρεται στο θαλάσσιο
περιβάλλον από ποτάµια και στη συνέχεια αποτίθεται κυρίως στις ηπειρωτικές
παρυφές (ηπειρωτικά περιθώρια). Έτσι τυπικές τιµές ιζηµατοπόθεσης για τα
ηπειρωτικά περιθώρια είναι 5 – 20 cm ανά 1000 χρόνια ενώ για τις βαθιές ωκεάνιες
περιοχές οι αντίστοιχες τιµές είναι µόνο 0,2 –1 cm ανά 1000 χρόνια. Είναι λοιπόν
σαφές ότι τα χερσογενή ιζήµατα συσσωρεύονται 100 φορές ταχύτερα στα ηπειρωτικά
παρυφές από ότι στο βαθύ ωκεάνιο πυθµένα. Έτσι µολονότι τα χερσογενή ιζήµατα
αποτελούν το 75% του συνολικού όγκου των ιζηµάτων, εντούτοις καλύπτουν µόνο το
20% της επιφάνειας των ωκεανών, αποτιθέµενα κυρίως στα ηπειρωτικά περιθώρια.
Ωστόσο, µερικά φθάνουν στα µεγάλα ωκεάνια βάθη και απαντώνται στα τουρβιδιτικά
αποθέµατα ιζηµάτων των αβυσσαίων πεδιάδων.
Λάσπη : Οι λάσπες χερσαίας προέλευσης αποτελούνται από ιλύ και πηλό. Τα κύρια
ορυκτά σ' αυτά τα ιζήµατα είναι χαλαζίας, άστριοι, καολίνης και χλωρίτης. Οι ποταµοί
µεταφέρουν µικρά σωµατίδια αυτών των ορυκτών στους ωκεανούς, όπου, αν τα
ρεύµατα είναι αρκετά ισχυρά, παραµένουν σε αιώρηση. Στη συνέχεια τα σωµατίδια
µπορεί να µεταφέρονται µακριά µέσα στη θάλασσα πριν κατακαθίσουν πάνω στο
θαλάσσιο πυθµένα. Αυτό το σε αιώρηση ίζηµα αντιπροσωπεύει µικρό τµήµα του
ιζήµατος επί του θαλάσσιου πυθµένα διότι τα περισσότερα παράκτια ρεύµατα
κινούνται κατά µήκος µάλλον της ακτής παρά αποµακρύνονται από την ακτή.
Η µεγαλύτερη ποσότητα χερσαίας λάσπης µεταφέρεται στο θαλάσσιο πυθµένα µε τα
τουρβιδιτικά ρεύµατα. Πολλή απ' αυτήν την λάσπη µεταφέρεται κάτω στα
υποθαλάσσια φαράγγια και διασπείρεται επί του θαλάσσιου πυθµένα. Εκεί όπου οι
υφαλοκρηπίδες είναι φαρδιές (περισσότερο από 10 χλµ) ή όπου δεν υπάρχουν
φαράγγια, τα ιζήµατα δεν έχουν εύκολη δίοδο προς τις ωκεάνιες λεκάνες.
Οι ετήσιοι άνεµοι µεταφέρουν σκόνη από τις τροπικές ερήµους στους ωκεανούς,
όπου η σκόνη καταπίπτει σιγά-σιγά για να καταστεί λάσπη επί του θαλάσσιου
πυθµένα. Σκόνη από την έρηµο Σαχάρα µεταφέρεται κατά µήκος του Ατλαντικού
Ωκεανού προς την Καραϊβική θάλασσα. Θύελλες σε µεγαλύτερα πλάτη µεταφέρουν
επίσης σκόνη από άγονες περιοχές. Σκόνη από τη Μογγολία µεταφέρεται προς το Β.
Ειρηνικό και σκόνη από την Αυστραλία µεταφέρεται προς τον Ινδικό Ωκεανό.
Αποθέσεις παγετώνων : Ιζήµατα προερχόµενα από αποθέσεις παγετώνων
θεωρούνται χερσαία λόγω της συµµετοχής σ' αυτά χερσαίου υλικού. Τέτοια ιζήµατα
κοντά στην Ανταρκτική ήπειρο και τη Γροιλανδία αποτελούνται από υλικά τα οποία

32
προήλθαν από το λειώσιµο παγετώνων. Καθώς ο παγετώνας κινείται στην
κατωφέρεια προς τη θάλασσα, τα χαµηλότερα τµήµατα λειώνουν περιοδικά και
ξαναπαγώνουν εξαιτίας της πίεσης και της θερµοκρασίας τριβής. Καθώς ο πάγος
ταξιδεύει, διαβρώνει τα υποκείµενα τµήµατα στεριάς και µεταφέρει άµµο, ιλύ και
χαλίκια, τα οποία ενσωµατώνονται στον πάγο. Όταν τελικά ο πάγος φθάσει στη
θάλασσα, µεγάλα τεµάχια σπάζουν, µετατρεπόµενα σε παγόβουνα τα οποία
µεταφέρονται προς τη θάλασσα µε τα ωκεάνια ρεύµατα. Καθώς τα παγόβουνα
λειώνουν, αποβάλλουν τα φορτία τους στον ωκεάνιο πυθµένα.
(γ) Κοσµογενή ιζήµατα
Εκτιµάται ότι περίπου 5.000 τόνοι µετεωριτών πέφτουν στην επιφάνεια της Γης κάθε
χρόνο. Τα περισσότερα απ' αυτά τα εξωγήινα αντικείµενα αποτελούνται από µικρά
σωµατίδια τα οποία πέφτουν στους ωκεανούς και ενσωµατώνονται µε άλλα ιζήµατα.
Κάθε µέρα µερικές χιλιάδες τόνοι τέτοιου µετεωρικού υλικού προστίθενται στη Γη και
κυρίως στους ωκεανούς. Όµως µερικές χιλιάδες τόνοι επιµερισµένοι σε ολόκληρη την
επιφάνεια της γης, αποτελούν µια αµελητέα προσθήκη στο κάλυµµα των ιζηµάτων.
Μπορούν να αναγνωρισθούν από τη χαρακτηριστική σύνθεσή τους και το
µαγνητισµό τους. Εκτίµηση της ηλικίας των ιζηµάτων αυτών µε ραδιενεργά στοιχεία
φανερώνει ότι είναι ηλικίας 4,5 δισεκατοµ. ετών,γεγονός που αποτελεί σαφή ένδειξη
ότι δεν έχουν σχηµατισθεί στον ωκεανό.
Τα κοσµογενή συστατικά, τα οποία καλούνται τεκτίτες, µερικές φορές βρίσκονται στα
ιζήµατα των ωκεανών. Αυτά είναι πολυάριθµα στον ανατολικό Ινδικό Ωκεανό. Οι
τεκτίτες συνίστανται από 70% µέχρι 90% SiO2 (υαλώδες πυρίτιον). Είναι διαµέτρου
µικρότερης από 1,5 χλστµ και τυπικά είναι επιµήκεις. Αν και θεωρούνται ότι είναι
εξωγήινης προέλευσης, η ακριβής προέλευσή τους είναι άγνωστη. Μερικοί
επιστήµονες υποστηρίζουν ότι προήλθαν από κοµήτες, πλανητοειδείς ή από τη
σελήνη, όλοι όµως συµφωνούν ότι οι τεκτίτες έχουν πέσει στην επιφάνεια της γης δια
µέσου της γήινης ατµόσφαιρας.
1.5 Κατανοµή των ωκεάνιων ιζηµάτων
Η παγκόσµια κατανοµή των ανθρακικών και πυριτικών αποθέσεων και των ιζηµάτων
µεγέθους αργίλου µέχρι λάσπης ακολουθούν χαρακτηριστικά πρότυπα. Οι
ανθρακικές αποθέσεις είναι περισσότερο κοινές στις τροπικές περιοχές ενώ οι
πυριτικές αποθέσεις στα υψηλότερα νότια πλάτη. Οι άργιλοι (πηλός) είναι άφθονοι
σε ολόκληρο τον Β. Ειρηνικό κοντά στις εκβολές µεγάλων ποταµών. Οι αποθέσεις

33
από παγετώνες βρίσκονται κυκλικά της Ανταρκτικής. Ανθρακικά ιζήµατα είναι
σχετικά σπάνια σε βάθη µεγαλύτερα από 4.000 µ.
Η συσσώρευση των ιζηµάτων στον πυθµένα των θαλασσών είναι µια αργή
διαδικασία που εκτιµάται σε 0,0015 χλστµ καινούργιου ιζήµατος κατά χρόνο. Ο
Kuenen (1941, 1950), χρησιµοποιώντας διάφορες ανεξάρτητες µεθόδους, εκτίµησε
ότι τα ιζήµατα συσσωρεύονται σ' ένα βάθος 3 χλµ κάθε 2 δισεκατοµµύρια χρόνια.
Λαµβάνοντας υπόψη τη µέχρι σήµερα ισχύουσα γνώση ότι η Γη είναι ηλικίας 4
δισεκατοµµυρίων ετών, µπορεί να εκτιµηθεί ότι το µέσο βάθος των πελαγικών
ιζηµάτων είναι τουλάχιστον 5 ή 6 χλµ. Στην πραγµατικότητα όµως υπάρχει µόνο 0,3
χλµ ιζήµατος επί του πυθµένος των βαθιών θαλασσών, όπως έχει διαπιστωθεί µε τη
χρήση των σύγχρονων τεχνικών της Θαλάσσιας Γεωλογίας. Συνολικά περίπου το
14% όλων των βαθιών ωκεάνιων ιζηµάτων είναι πυριτική ιλύς, 48% είναι
ασβεστολιθική ιλύς και το υπόλοιπο 38% είναι αβυσσαίοι άργιλοι και χερσογενή
ιζήµατα.
Ο βαθµός ιζηµατογένεσης δεν είναι οµοιόµορφος στις διάφορες περιοχές των
ωκεανών και δεν είναι ο ίδιος σε µια δεδοµένη περιοχή κατά τη διάρκεια των ετών.
Είναι ταχύτερος στις αβαθείς θάλασσες και στις περιοχές της υφαλοκρηπίδας και
βραδύτερος στα αβυσσαία βάθη της ανοικτής θάλασσας.
Πολλά ζώα ζουν επί ή κοντά στο ενδιάµεσο µεταξύ των ιζηµάτων του πυθµένος και
του κρύου, βραδέως κινούµενου νερού, πάνω από τον πυθµένα. Μερικά σκάπτουν ή
χώνονται τα ίδια µέσα στο ίζηµα. Άλλα προσκολλώνται µε διαδικασίες παρόµοιες µ'
εκείνες των ριζών. Πολλά έρπουν επί ή δια µέσου του ιζήµατος, τρεφόµενα από
οργανικό υλικό που βρίσκεται αναµειγµένο µε το ίζηµα, ή από άλλα ζώα που τα
βρίσκουν εκεί. Επίσης πολλά βενθικά ψάρια και µερικά κεφαλόποδα µετακινούνται
πάνω στο θαλάσσιο πυθµένα, τρεφόµενα από βενθικά ζώα.
Ο χαρακτήρας των πληθυσµών των ζώων πάνω στον πυθµένα καθορίζεται από το
είδος και βάθος του πυθµένα των ωκεανών. Ο πυθµένας των ωκεανών κατοικείται
από ζώα σε όλα του τα βάθη, εντούτοις η ποσότητα της ζωής µειώνεται σηµαντικά
στα µεγάλα βάθη. Μερικές οµάδες ζώων εξαφανίζονται εντελώς στα βαθιά νερά ενώ
άλλες δεν εµφανίζονται καθόλου στα αβαθή.
Ο Zenkevitch (1963) αναφερόµενος στα αποτελέσµατα της ερευνητικής αποστολής
των Ρώσων στο Β.∆. Ειρηνικό διαπιστώνει ότι µεταξύ 1.000 και 5.000 µ υπάρχει µια
απότοµη µείωση της σχετικής σπουδαιότητας των σπόγγων. Μεταξύ 4.000 και 6.000
µ οι αστερίες, οι θαλάσσιες ανεµώνες και οι πολύχαιτοι γίνονται πολύ λιγότερο
εµφανείς. Τα δεκάποδα καρκινοειδή δεν βρέθηκαν σε βάθη µεγαλύτερα από 5.000 µ

34
ενώ τα ολοθούρια έγιναν πολύ περισσότερο σηµαντικά. Κάτω από τα 8.000 µ δεν
υπάρχουν αστερίες και κοράλλια, ενώ τα αµφίποδα και ισόποδα γίνονται σπάνια
όπως και οι ανεµώνες και τα µαλάκια στα 9.000 µ. Κάτω από τα βάθη αυτά το 90%
των ζώων αποτελούνται από µερικούς πολύχαιτους, πωγωνοφόρα και ολοθούρια.
Σε δειγµατοληψία στα 1.000 µ µια απλή σύρση δράγας περιείχε 120 είδη, ενώ σε
βάθη µεταξύ 8.000 και 9.000 µ τρεις σύρσεις δράγας απέδωσαν 6,7 και 17 είδη
αντίστοιχα.
Ένας σπουδαίος λόγος για τον οποίο στα µεγάλα βάθη παρατηρείται µεγάλη µείωση
της βιοµάζας είναι η µη διαθεσιµότητα τροφής.
1.6 δειγµατοληψία ιζηµάτων
Η δειγµατοληψία επιφάνειας των ιζηµάτων γίνεται συνήθως µε ∆ράγα (dredge) (Εικ.
1.16α) ή µε δειγµατολήπτη µε σιαγόνες (δειγµατολήπτης Petersen) (Εικ.1.16β) Οι
∆ράγες χρησιµοποιούνται κυρίως για τη συλλογή αδροµερών ιζηµάτων, όπως οι
κόνδυλοι µαγγανίου. Οι δειγµατολήπτες µε σιαγόνες χρησιµοποιούνται για τη
δειγµατοληψία πιο λεπτόκοκκων ιζηµάτων. Υποεπιφανειακά δείγµατα ιζηµάτων -
υπό µορφή πυρήνων γνωστών ως καρώτων µήκους έως και 20 µ – συλλέγονται µε
πυρηνοδειγµατολήπτες (Εικ. 1.16γ).
Εικ. 1.16 ∆ιάφοροι δειγµατολήπτες ιζηµάτων (α) Κλασσικός τύπος δράγας (β) ∆ειγµατολήπτης µε
σιαγόνες (PETERSEN) (γ) Κλασσικός τύπος πυρηνοδειγµατολήπτη

35
1.7 Τα περιθώρια των ηπείρων
Το περιθώριο των ηπείρων περιλαµβάνει την ακτή, την ηπειρωτική υφαλοκρηπίδα, το
ηπειρωτικό υφαλοπρανές και την ηπειρωτική έξαρση. Αποτελεί το υποθαλάσσιο
τµήµα των ηπείρων και είναι η µεταβατική ζώνη µεταξύ ηπείρων και της ωκεάνιας
λεκάνης.
Τα περιθώρια των ηπείρων αποτελούν αντικείµενο µεγάλου ενδιαφέροντος από όλες
τις βρεχόµενες από θάλασσα χώρες οι οποίες επιθυµούν να αναπτύξουν
δυνατότητες εκµετάλλευσης των θαλάσσιων πόρων. Το ενδιαφέρον των χωρών για
τα περιθώρια των ηπείρων οφείλεται σε δύο λόγους κυρίως: ψάρια και πετρέλαιο.
Το µεγαλύτερο µέρος της παγκόσµιας αλιείας συγκεντρώνεται στα περιθώρια των
ηπείρων, όπου τα νερά είναι πλούσια σε θρεπτικά συστατικά. Πλέον αυτού,
ορισµένες περιοχές των ηπειρωτικών περιθωρίων περιέχουν άφθονα αποθέµατα
πετρελαίου, αερίων, αµµοχάλικου, φωσφορικών αλάτων, άνθρακος και άλλων µη
ζώντων πόρων. Τα ηπειρωτικά περιθώρια και ιδιαίτερα τα υφαλοπρανή και οι
εξάρσεις, περιλαµβάνουν τεράστιες φυσικές πηγές, µερικές από τις οποίες
παραµένουν ακόµη αχρησιµοποίητες.
Καθώς οι φυσικές πηγές επί της ξηράς έχουν αρχίσει να λιγοστεύουν, ασκούνται
κοινωνικές και πολιτικές πιέσεις από εκβιοµηχανοποιηµένα έθνη για την επικέντρωση
του ενδιαφέροντος στην πλουτοπαραγωγική δυναµικότητα αυτών των περιοχών.
Τα ηπειρωτικά περιθώρια είναι τριών κυρίων τύπων. Τα περιθώρια κατά µήκος των
δυτικών ακτών της Βόρειας και Νότιας Αµερικής, τα οποία χαρακτηρίζονται από
συχνούς σεισµούς, ενεργά ηφαίστεια και εν ενεργεία ρήγµατα. Οι περιοχές αυτές
καλούνται ενεργά περιθώρια (active margins). Τα Ανατολικά περιθώρια αυτών των
ηπείρων χαρακτηρίζονται από σχετικά λιγότερους σεισµούς, ενεργά ηφαίστεια και εν
ενεργεία ρήγµατα (Εικ.1.17). Τα περιθώρια αυτά καλούνται παθητικά περιθώρια
(passive margins). Ένας τρίτος τύπος περιθωρίων καλούνται περιθώρια παράλληλης
µετατόπισης (translation margins) και χαρακτηρίζονται από ρήγµατα µε µεγάλες
οριζόντιες µετατοπίσεις και από σεισµούς µε επίκεντρο σε µικρό βάθος. Η ακτή της
Νότιας Καλιφόρνια είναι παράδειγµα αυτού του τύπου ηπειρωτικών περιθωρίων.

36
Εικ. 1.17 Παγκόσµιος χάρτης παρουσίας της σηµερινής θέσης των παθητικών ηπειρωτικών περιθωρίων
(τροποποιηµένη από D. E. Ingmason and W. J. Wallace, 1989)
∆ιαφορές στην τεκτονική δραστηριότητα δηµιουργούν διαφορές στη δοµή των
πετρωµάτων και την ιζηµατογένηση κατά µήκος των περιθωρίων. Εναλλασσόµενες
αβαθείς ακτές, κοιλώµατα, ρήγµατα και σχετικά στενές υφαλοκρηπίδες είναι τυπικά
περιθώρια παράλληλης µετατόπισης.
Βαθιές χαράδρες και ηφαιστειογενούς προέλευσης νησιωτικά τόξα είναι κοινά κατά
µήκος των ενεργών περιθωρίων. Τέλος τα παθητικά περιθώρια, τυπικά έχουν
πλατειές ηπειρωτικές υφαλοκρηπίδες (ακτές των ανατολικών ακτών της Β. Αµερικής),
µεγάλα ∆έλτα (ποταµός Μισσισσιπής) ή πλατειά διασπαρµένους κοραλλιογενείς
υφάλους (Β.Α. Αυστραλία) (Εικ. 1.16). ∆εν παρουσιάζουν εν ενεργεία ρήγµατα ή
ηφαίστεια και οι ηπειρωτικές υφαλοκρηπίδες κυµαίνονται σε πλάτος από περίπου 20
χλµ µέχρι πάνω από 200 χλµ.
1.7.1 Ακτές
Η παράκτια ζώνη είναι η πιο καλά αντιληπτή από τις τεσσερις κύριες φυσιογραφικές
υποδιαιρέσεις των ηπεορωτικών περιθωρίων. Η σχετικά πλήρης γνώση της
παράκτιας ζώνης προέρχεται από το γεγονός ότι είναι ορατή σε αντίθεση µε την
υποθαλάσσια ηπειρωτική υφαλοκρηπίδα, ηπειρωτικό υφαλοπρανές και ηπειρωτική

37
έξαρση. Επιπλέον, οι παράκτιες ζώνες του κόσµου είναι πάντοτε οι πιο
πυκνοκατοικηµένες περιοχές στις οποίες αναπτύσσονται πλήθος δραστηριοτήτων.
Ακτή καλείται το τµήµα της στεριάς σε επαφή µε την υδάτινη µάζα. Οι ακτές δεν
έχουν σταθερότητα σε έκταση και µορφή. Αιτίες των µεταβολών τους είναι:
(α)µεταβολές της στάθµης της θάλασσας (β) παράκτια διάβρωση (οριζόντια και
κάθετη) και (γ) προσχώσεις.
Φυσιογραφικά διακρίνονται σε χαµηλές, υψηλές, οµοιόµορφες και διακεκοµµένες,
ενώ από την άποψη προσβολής τους από τους ανέµους διακρίνονται σε προσήνεµες
και υπήνεµες. Χαρακτηριστικοί τύποι χαµηλών ακτών αποτελούν οι χερσονησίδες, οι
λιµνοθάλασσες, τα τενάγη κλπ. Οι υψηλές ακτές, ανάλογα µε τη γωνία κλίσης τους
ως προς τη επιφάνεια της θάλασσας, διακρίνονται σε οµαλές (γωνία κλίσης µέχρι
15ο), υπόκρηµνες (γωνία κλίσης 15ο-25ο), κρηµνώδεις (γωνία κλίσης 25ο-45ο),
απόκρηµνες (γωνία κλίσης µέχρι 75ο) και κάθετες (γωνία κλίσης 90ο) Από
µορφολογική άποψη διακρίνονται σε βραχώδεις, χαλικώδεις, αµµώδεις και
λασπώδεις.
1.7.2 Ηπειρωτικές υφαλοκρηπίδες
Ηπειρωτική υφαλοκρηπίδα είναι τα υπό τη θάλασσα τµήµατα των ηπείρων. Αν και οι
περισσότερες υφαλοκρηπίδες βρίσκονται στα όρια µεταξύ ηπείρου και ωκεανού,
µερικές σχεδόν περιβάλλονται από ξηρά, όπως αυτή του κόλπου Hudson στον
Καναδά. Οι ηπειρωτικές υφαλοκρηπίδες αποτελούν 7,5% της συνολικής επιφάνειας
του θαλάσσιου πυθµένα. Μπορεί να είναι πλατειές µέχρι 1500 χλµ αλλά το µέσο
πλάτος τους είναι µεταξύ 50 και 100 χλµ.
Το προς τη θάλασσα τµήµα της ηπειρωτικής υφαλοκρηπίδας είναι τυπικά σε βάθος
130 µέχρι 200 µ, µετά το οποίο η κλίση του θαλάσσιου πυθµένα αυξάνει απότοµα.
Από την ακτή η ηπειρωτική υφαλοκρηπίδα βαθαίνει κατά 1 µ περίπου ανά 1000 µ και
φθάνει στα 10 µ περίπου ανά 1000 µ στο ηπειρωτικό υφαλοπρανές. Υπάρχουν
µεγάλες διαφορές σε ό,τι αφορά την κλίση της υφαλοκρηπίδας. Π.χ. υφαλοκρηπίδες
που είναι διαβρωµένες από παγετώνες εκτείνονται γενικά σε βάθος σχεδόν µέχρι 500
µ πριν φθάσουν στο σηµείο έναρξης του ηπειρωτικού υφαλοπρανούς ενώ µερικές
υφαλοκρηπίδες κοραλλιών φθάνουν σε βάθος µόνο 20µ.
Οι χαρακτήρες των ηπειρωτικών υφαλοκρηπίδων δύνανται να αποδοθούν σε επτά
διαδικασίες γένεσής τους: παγετώνες, αλλαγές της στάθµης της θάλασσας, κύµατα
και ρεύµατα, ιζηµατογένεση χερσαίου υλικού, αποθέσεις ανθρακικών, ρήγµατα και
ηφαίστεια .

38
Παγετώνες: Κατά τη διάρκεια των τελευταίων δύο εκατοµ. ετών, παγετώνες είχαν
συνεχώς καλυµµένο το περισσότερο τµήµα της Ανταρκτικής και Γροιλανδίας. Κατά
την περίοδο εκείνη, η Β. Αµερική, Ευρώπη και Ασία είχαν περιοδικά καλυφθεί από
παγετώνες αρκετά µεγάλου πάχους και πλάτους ώστε να ονοµάζονται σεντόνια
πάγου. Μεταξύ αυτών των επεισοδίων µαζικού παγώµατος, ενδιάµεσα συνέβη
λιώσιµο των πάγων εξαιτίας µαλακού κλίµατος παρόµοιου µ' αυτό που υπάρχει
σήµερα. Η έκταση του παγώµατος ήταν διάφορη ανάλογα µε την κατά περιοχή
αλλαγή του κλίµατος και της τοπογραφίας.
Οι εναλλαγές των περιόδων ψύξης και ηπιότερου κλίµατος, είχε αναµφίβολα
σηµαντική επίδραση επί των ωκεανών, την ατµόσφαιρα, τους θαλάσσιους
οργανισµούς και τις ηπείρους αυτές καθαυτές. Όταν η στάθµη της θάλασσας ήταν
100 έως 125 µ κάτω από τη σηµερινή στάθµη, µεγάλα τµήµατα αυτού που καλούµε
ηπειρωτική υφαλοκρηπίδα ήταν εκτεθειµένα. Τεράστια δόντια απολιθωµένα, που
έχουν ανακαλυφθεί κατά την βυθοκόρηση της ηπειρωτικής υφαλοκρηπίδας ανοικτά
της Νέας Αγγλίας (Ν.Α. των ΗΠΑ), παρέχουν ενδείξεις, οι οποίες πείθουν ότι υπήρξε
τέτοια έκθεση εκτός υδάτων. Πλέον αυτών, δείγµατα ιζηµάτων πυθµένος παρέχουν
ενδείξεις κατάκλισης µε ύδατα πεδιάδων τύρφης πλούσιων σε απολιθώµατα φυτών
και γυρεόκοκκων.
Τα κελύφη πολλών θαλάσσιων µικροοργανισµών, όπως τα τρηµατοφόρα,
αποτελούνται από CaC03. Ερευνητές έχουν χρησιµοποιήσει τα απολιθώµατα αυτών
των οργανισµών για να εκτιµήσουν την ηλικία ωκεάνιων ιζηµάτων και να βγάλουν
συµπεράσµατα σχετικά µε τις θερµοκρασίες του νερού στις διάφορες περιόδους των
παγετώνων. Μελετώντας τα απολιθώµατα τα οποία αποτελούνται από CaCO3
διαπιστώνεται ότι αυτά έχουν κοινή ιστορία µε την ιστορία των παγετώνων.
Κατά τη διάρκεια των περιόδων αύξησης και ελάττωσης των παγετώνων, τµήµατα
της ηπειρωτικής υφαλοκρηπίδας στα µεγάλα γεωγραφικά πλάτη διαβρώθηκαν από
την τριβή µεταξύ των βράχων και των εγκλεισµένων στους παγετώνες τροχάλων. Σε
µερικές περιοχές οι παγετώνες απέθεσαν τεράστιες ποσότητες ιζηµάτων. Σε άλλες
περιοχές, οι µεταβολές της στάθµης της θάλασσας αντίστοιχα µε τις µετακινήσεις των
πάγων οδήγησαν σε διάβρωση του χείλους των ηπείρων από την ενέργεια των
κυµάτων.
Αλλαγές της στάθµης της θάλασσας: Η επίδραση των παγετώνων επί των ηπείρων
υπήρξε η κύρια αιτία των διακυµάνσεων της στάθµης της θάλασσας, δηµιουργώντας
ανύψωση και πτώση 130 µ από τη σηµερινή στάθµη της. Οι µεταβολές της στάθµης
της θάλασσας εξαιτίας των παγετώνων έχουν κυριαρχήσει στην πρόσφατη

39
γεωλογική ιστορία, αλλά σηµαντικές µεταβολές στο παρελθόν ήταν το αποτέλεσµα
άλλων διεργασιών. Περιοδικές µεταβολές µέχρι µερικές εκατοντάδες µέτρα έλαβαν
χώρα µέσα σε εκατοµµύρια χρόνια, οι αιτίες των οποίων δεν είναι γνωστές.
Κατά τις περιόδους που η στάθµη της θάλασσας ήταν χαµηλή, οι ήπειροι ήταν
σχεδόν 25% µεγαλύτεροι από όσο είναι σήµερα . Ποταµοί έρεαν δια µέσου αυτών
που σήµερα είναι ηπειρωτικές υφαλοκρηπίδες και απέθεταν ιζήµατα κατευθείαν επί
του θαλάσσιου πυθµένα. Σε άλλες εποχές, όπως κατά την Κρητιδική περίοδο, όταν η
στάθµη της θάλασσας ήταν υψηλή, 40% των ηπείρων ήταν πληµµυρισµένες.
Αβαθείς θάλασσες κυριαρχούσαν και οι βαθιές θάλασσες ήταν µακριά από την ξηρά.
Κύµατα και ρεύµατα: ∆ιάφορες δυνάµεις επηρέασαν τα χαρακτηριστικά της
ηπειρωτικής υφαλοκρηπίδας στη διάρκεια του χρόνου. Χαρακτηριστικά που ήταν
υποθαλάσσια για πολύ χρόνο έχουν αποκαλυφθεί εξαιτίας των µεταβολών της
στάθµης της θάλασσας. Κύµατα, ποταµοί και ρεύµατα είχαν την έντονη επίδραση επί
των χαρακτηριστικών της ηπειρωτικής υφαλοκρηπίδας. Σε µικρότερη έκταση
επέδρασαν επίσης οι παλίρροιες, τα κύµατα εξαιτίας σεισµών και τα κύµατα εξαιτίας
καταιγίδων.
Σε µερικές περιοχές παλιρροϊκά ρεύµατα, πλέοντα µεταξύ χερσαίων µαζών
σκουπίζουν την υφαλοκρηπίδα και εµποδίζουν τη συσσώρευση φερτών υλικών επί
της υφαλοκρηπίδας. Επίσης τουρβιδιτικά ρεύµατα, ρέοντα δια µέσου φαραγγιών,
µεταφέρουν ιζήµατα από την υφαλοκρηπίδα στο βαθύ θαλάσσιο πυθµένα που
βρίσκεται έξω από τα φαράγγια. Τα ρεύµατα αυτά ρέουν κατερχόµενα την πλαγιά
µέχρι να χάσουν την ενέργειά τους στη βαθιά θάλασσα.
Τόσο τα παλιρροϊκά όσο και τα σεισµικά θαλάσσια κύµατα επηρεάζουν το ωκεάνιο
νερό από την επιφάνεια µέχρι τον πυθµένα, ακόµη και στη βαθιά θάλασσα. Η
έκταση της επίδρασής τους επί της υφαλοκρηπίδας δεν είναι γνωστή, παρά το
γεγονός ότι γνωρίζουµε ότι τα θαλασσινά κύµατα µπορούν να µεταφέρουν µεγάλες
ποσότητες µη συµπυκνωµένου ιζήµατος σε µεγάλες αποστάσεις µέσα σε µικρή
χρονική περίοδο. Το ίδιο αποτέλεσµα φαίνεται ότι έχουν και οι κυµατισµοί εξαιτίας
καταιγίδων.
Ιζηµατογένεση χερσαίου υλικού: Για την ιζηµατογένεση χερσαίου υλικού βλέπε Κεφ.
1.4.4 (β) Χερσογενή ιζήµατα
Αποθέσεις ανθρακικών: Οι ανθρακικές αποθέσεις είναι περισσότερο κοινές στις
τροπικές περιοχές.
Ρήγµατα και ηφαίστεια : ∆εδοµένου ότι οι ηπειρωτικές υφαλοκρηπίδες είναι η έκθεση
της επιφάνειας της µεταβατικής ζώνης µεταξύ ηπείρων και ωκεάνιων λεκανών, δεν

40
πρέπει να εκπλήσσει το γεγονός ότι παρουσιάζουν κάποια αστάθεια. Όπως φαίνεται
στο χάρτη των σεισµών πολλά ηπειρωτικά περιθώρια είναι ασταθή. Πιέσεις µέσα
στη γη σπάζουν τα πετρώµατα και προκαλούν κίνηση κατά µήκος των
προκαλούµενων ρηγµάτων, µια διεργασία που είναι γνωστή ως faulting (δηµιουργία
ρηγµάτων). Βρίσκουµε ενδείξεις για την ύπαρξη ρηγµάτων σε όλες τις ηπειρωτικές
υφαλοκρηπίδες που έχουν ερευνηθεί µέχρι σήµερα. Η περιοχή ανοικτά της νότιας
Καλιφόρνια καλείται µεθόριος περιοχή (borderland). Η περιοχή αυτή περιέχει
ηπειρωτικά πετρώµατα τα οποία είναι κοµµατιασµένα από ρήγµατα, ώστε υπάρχει
µια στενή υφαλοκρηπίδα η οποία συνεχίζεται προς την πλευρά της θάλασσας από
µια σειρά κοιλωµάτων και εξάρσεων πριν φθάσουν στο υφαλοπρανές το οποίο
βρίσκεται στα 165 χλµ ανοικτά της περιοχής.
1.7.3 Ηπειρωτικές πλαγιές (πρανή)
Η ηπειρωτική πλαγιά ή ηπειρωτικό υφαλοπρανές αρχίζει εκεί όπου η ελαφρά κλίση
της ηπειρωτικής υφαλοκρηπίδας δίνει τη θέση της σε µια ζώνη απότοµης αύξησης
της κλίσης. Η κατωφέρεια της ηπειρωτικής πλαγιάς διαδοχικά ελαττώνεται,
διατηρώντας µια µέση γωνία κλίσης 4ο µέχρι που συναντάει την ηπειρωτική έξαρση.
Οι ηπειρωτικές πλαγιές αποτελούν περίπου 8,5% της ολικής επιφάνειας του
ωκεάνιου πυθµένα και περιέχουν περίπου 40% των ιζηµάτων της γης.
Στο σύστηµα των τάφρων του Ειρηνικού Ωκεανού το ηπειρωτικό υφαλοπρανές
φθάνει σε βάθη µεταξύ 5 και 10 χλµ, σε άλλους ωκεανούς όµως σπάνια φθάνει σε
τέτοια βάθη. Μη συµπαγή ιζήµατα επί των υφαλοπρανών τείνουν να είναι πιο
λεπτόκοκκα σε σχέση µε εκείνα επί των υφαλοκρηπίδων και περιέχουν λιγότερα
απολιθώµατα.
1.7.4 Ηπειρωτικές εξάρσεις
Οι ηπειρωτικές εξάρσεις, οι οποίες αποτελούν την τελική µεταβατική ζώνη µεταξύ των
ηπείρων και της ωκεάνιας λεκάνης, αποτελούν περίπου 3% του θαλάσσιου πυθµένα
και περιέχουν περίπου 30% των ιζηµάτων επί της γης. Στις περισσότερες περιοχές,
η ηπειρωτική έξαρση αποτελείται από µη συµπαγή ιζήµατα λάσπης, ιλύος και άµµου,
τα οποία έχουν µεταφερθεί από την ηπειρωτική υφαλοκρηπίδα και τα υφαλοπρανή
µε τουρβιδιτικά ρεύµατα, υποθαλάσσιες κατολισθήσεις και άλλες διαδικασίες.
Στον Ειρηνικό Ωκεανό, ηπειρωτικές εξάρσεις υπάρχουν µόνο κατά µήκος του χείλους
της Ανταρκτικής ηπείρου και του Β.Α. Ειρηνικού, ανοικτά των ακτών της Β. Αµερικής.
Σε άλλους ωκεανούς, εξάρσεις είναι περισσότερο συνήθεις κυµαινόµενες από βάθη 3
µέχρι 4 χλµ. Ο τύπος των ηπειρωτικών εξάρσεων φαίνεται καθαρά ότι σχετίζεται µε
τις κινήσεις των τεκτονικών πλακών. Σε ενεργά περιθώρια (active margins) τα

41
ιζήµατα εναλλάσσονται και µεταφέρονται στον ηπειρωτικό φλοιό της γης µε καθίζηση.
Σε παθητικά περιθώρια, τα ιζήµατα διατηρούνται και εκτείνονται ανοικτά προς τον
ωκεάνιο πυθµένα.

42
2. ΧΗΜΙΚΕΣ Ι∆ΙΟΤΗΤΕΣ ΤΟΥ ΘΑΛΑΣΣΙΝΟΥ ΝΕΡΟΥ
2.1 Συστατικά του θαλασσινού νερού
Το θαλασσινό νερό είναι διάλυµα µιας µεγάλης ποικιλίας χηµικών στοιχείων. Είναι το µέσο το
οποίο υποστηρίζει όλη τη θαλάσσια ζωή, και η πηγή διατροφής για το µικροσκοπικό
πλαγκτόν, το οποίο είναι η βασική τροφή των ζώων της θάλασσας. Παρά το γεγονός ότι τα
βασικά χηµικά χαρακτηριστικά του θαλασσινού νερού είναι συγκριτικά σταθερά, οι φυσικές
του ιδιότητες δεν είναι σταθερές. Ειδικότερα µεταβάλλονται σε όλες τις διαστάσεις – οριζόντια
κάθετα, γεωγραφικά και ως συνάρητηση των ρευµάτων και της διαφοροποίησης των µαζών
του νερού, ανάλογα µε τις φυσικο – χηµικές ιδιότητες του.
Το θαλασσινό νερό είναι ένα πλήρες χηµικό µέσον για ζωή δεδοµένου ότι περιέχει όλες τις
χηµικές ουσίες που είναι απαραίτητες για την ανάπτυξη και τη διατήρηση των ιστών φυτών
και ζώων. Τα στοιχεία Mg και Ca καθώς και τα οξέα ανθρακικό και πυριτικό είναι σπουδαία
συστατικά των σκληρών σκελετικών µερών των θαλάσσιων οργανισµών. Τα νιτρικά και
φωσφορικά άλατα, εξάλλου, είναι απαραίτητα για την σύνθεση οργανικού υλικού από τα
φυτά. Εκτός απ' αυτό, µια ζωτική οµοιότητα υπάρχει µεταξύ της χηµικής σύνθεσης του
θαλασσινού νερού και εκείνης των υγρών του σώµατος των θαλασσινών φυτών και ζώων.
Τα περισσότερα από τα πλέον άφθονα ιόντα που αναφέρονται στον Πίνακας 2.1 είναι
σπουδαία συστατικά των υγρών του σώµατος όλων των θαλασσίων οργανισµών (ζώα και
φυτά).
Το θαλασσινό νερό είναι το προϊόν της διαλυτικής ενέργειας του νερού πάνω σε βράχους, στο
έδαφος, στους οργανισµούς και στην ατµόσφαιρα για εκατοµµύρια χρόνια. Περίπου 3,5%
του θαλασσινού νερού συνίσταται από διαλυµένα χηµικά συστατικά προερχόµενα από τις
πηγές αυτές. Το άλλο 96,5% είναι καθαρό νερό. Ίχνη, πιθανώς από όλα τα υπάρχοντα στον
πλανήτη µας φυσικά υλικά υπάρχουν στο θαλασσινό νερό. Τα υλικά αυτά µπορούν να
διαιρεθούν σε 3 γενικές κατηγορίες:
(α) Ανόργανες ουσίες, αναφερόµενες συνήθως ως άλατα, που περιλαµβάνουν θρεπτικά
συστατικά απαραίτητα για την ανάπτυξη των φυτών.
(β) ∆ιαλυµένα αέρια και
(γ) Οργανικά στοιχεία προερχόµενα συνήθως από ζωντανούς οργανισµούς. Στοιχεία αυτής
της κατηγορίας είναι διαλυµένα στο θαλασσινό νερό και περιλαµβάνουν λίπη, έλαια,
υδρογονάνθρακες, βιταµίνες, αµινοξέα, πρωτείνες και άλλες ουσίες. Αυτά τα στοιχεία
αποτελούν µια σπουδαία πηγή διατροφής των θαλασσινών βακτηρίων και πολλών άλλων
τύπων οργανισµών. Σύγχρονες έρευνες απέδειξαν ότι άλλες οργανικές συνθέσεις, όπως το
DDT και άλλοι χλωριωµένοι υδρογονάνθρακες, έχουν καταστρεπτικές συνέπειες επί
ορισµένων τύπων θαλάσσιας ζωής.
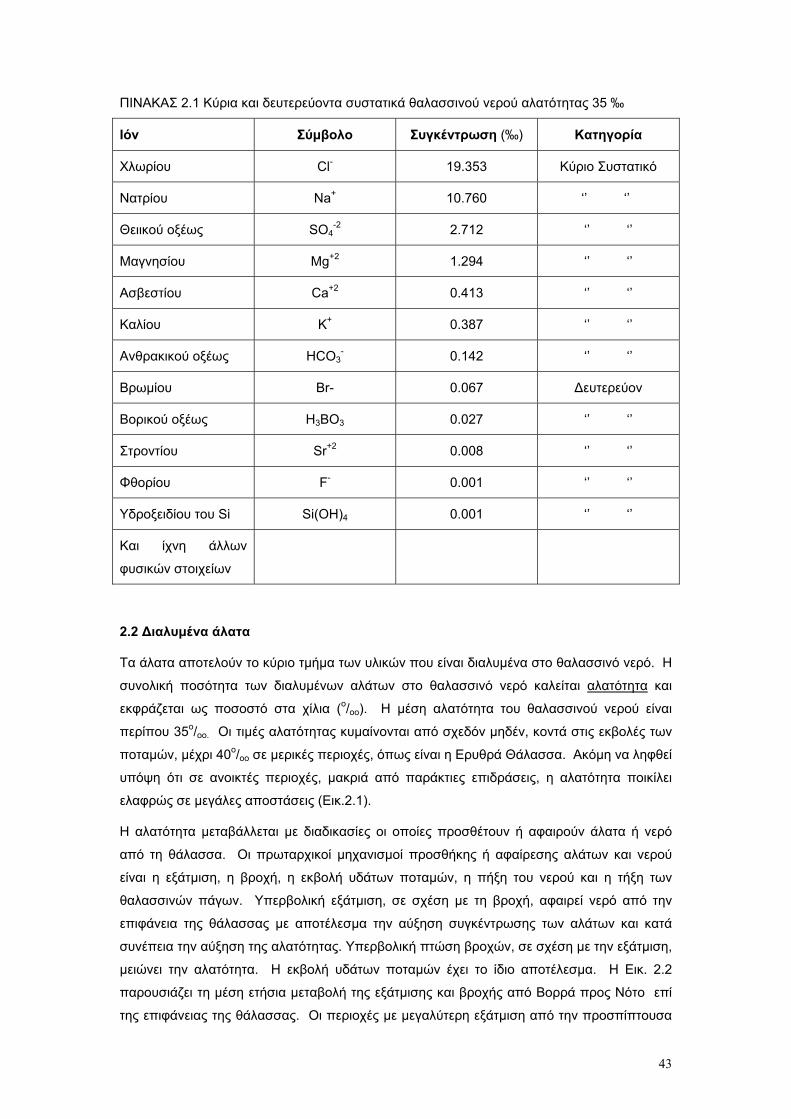
43
ΠΙΝΑΚΑΣ 2.1 Κύρια και δευτερεύοντα συστατικά θαλασσινού νερού αλατότητας 35 ‰
Ιόν Σύµβολο Συγκέντρωση (‰) Κατηγορία
Χλωρίου Cl- 19.353 Κύριο Συστατικό
Νατρίου Na+ 10.760 ‘’ ‘’
Θειικού οξέως SO4-2 2.712 ‘’ ‘’
Μαγνησίου Mg+2 1.294 ‘’ ‘’
Ασβεστίου Ca+2 0.413 ‘’ ‘’
Καλίου K+ 0.387 ‘’ ‘’
Ανθρακικού οξέως ΗCO3- 0.142 ‘’ ‘’
Βρωµίου Br- 0.067 ∆ευτερεύον
Βορικού οξέως H3BO3 0.027 ‘’ ‘’
Στροντίου Sr+2 0.008 ‘’ ‘’
Φθορίου F- 0.001 ‘’ ‘’
Υδροξειδίου του Si Si(OH)4 0.001 ‘’ ‘’
Και ίχνη άλλων
φυσικών στοιχείων
2.2 ∆ιαλυµένα άλατα
Τα άλατα αποτελούν το κύριο τµήµα των υλικών που είναι διαλυµένα στο θαλασσινό νερό. Η
συνολική ποσότητα των διαλυµένων αλάτων στο θαλασσινό νερό καλείται αλατότητα και
εκφράζεται ως ποσοστό στα χίλια (ο/οο). Η µέση αλατότητα του θαλασσινού νερού είναι
περίπου 35ο/οο. Οι τιµές αλατότητας κυµαίνονται από σχεδόν µηδέν, κοντά στις εκβολές των
ποταµών, µέχρι 40ο/οο σε µερικές περιοχές, όπως είναι η Ερυθρά Θάλασσα. Ακόµη να ληφθεί
υπόψη ότι σε ανοικτές περιοχές, µακριά από παράκτιες επιδράσεις, η αλατότητα ποικίλει
ελαφρώς σε µεγάλες αποστάσεις (Εικ.2.1).
Η αλατότητα µεταβάλλεται µε διαδικασίες οι οποίες προσθέτουν ή αφαιρούν άλατα ή νερό
από τη θάλασσα. Οι πρωταρχικοί µηχανισµοί προσθήκης ή αφαίρεσης αλάτων και νερού
είναι η εξάτµιση, η βροχή, η εκβολή υδάτων ποταµών, η πήξη του νερού και η τήξη των
θαλασσινών πάγων. Υπερβολική εξάτµιση, σε σχέση µε τη βροχή, αφαιρεί νερό από την
επιφάνεια της θάλασσας µε αποτέλεσµα την αύξηση συγκέντρωσης των αλάτων και κατά
συνέπεια την αύξηση της αλατότητας. Υπερβολική πτώση βροχών, σε σχέση µε την εξάτµιση,
µειώνει την αλατότητα. Η εκβολή υδάτων ποταµών έχει το ίδιο αποτέλεσµα. Η Εικ. 2.2
παρουσιάζει τη µέση ετήσια µεταβολή της εξάτµισης και βροχής από Βορρά προς Νότο επί
της επιφάνειας της θάλασσας. Οι περιοχές µε µεγαλύτερη εξάτµιση από την προσπίπτουσα
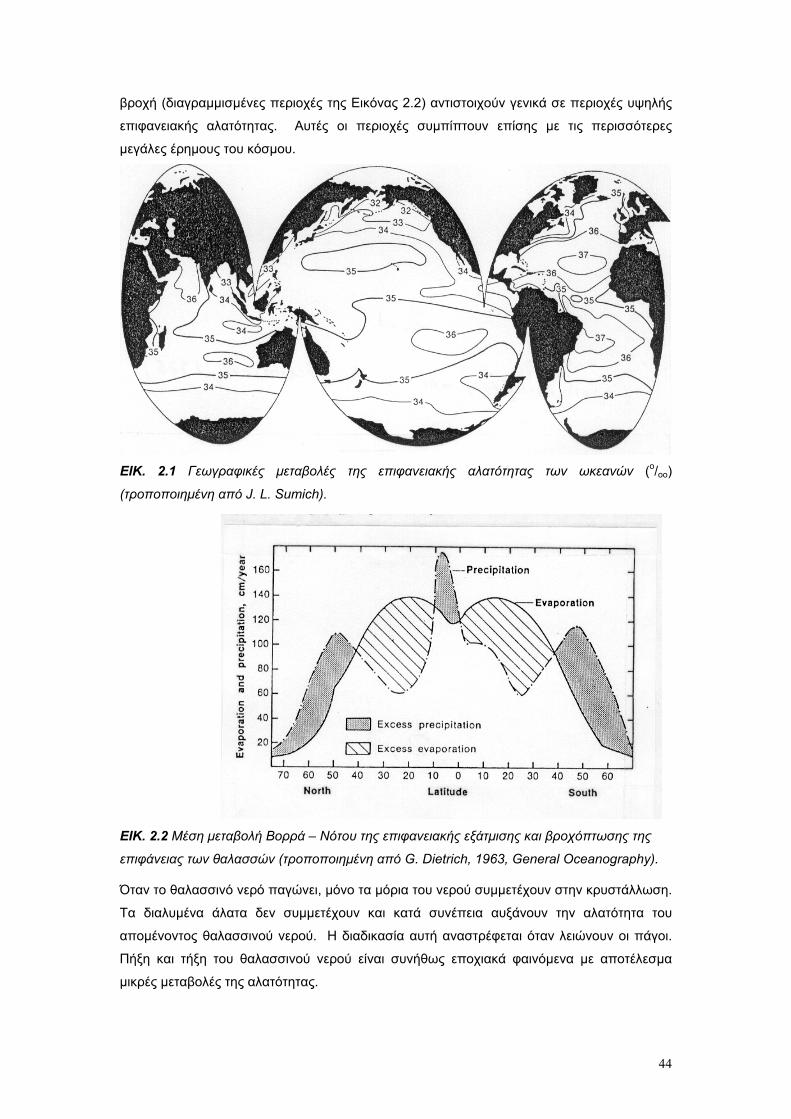
44
βροχή (διαγραµµισµένες περιοχές της Εικόνας 2.2) αντιστοιχούν γενικά σε περιοχές υψηλής
επιφανειακής αλατότητας. Αυτές οι περιοχές συµπίπτουν επίσης µε τις περισσότερες
µεγάλες έρηµους του κόσµου.
ΕΙΚ. 2.1 Γεωγραφικές µεταβολές της επιφανειακής αλατότητας των ωκεανών (ο/οο)
(τροποποιηµένη από J. L. Sumich).
ΕΙΚ. 2.2 Μέση µεταβολή Βορρά – Νότου της επιφανειακής εξάτµισης και βροχόπτωσης της
επιφάνειας των θαλασσών (τροποποιηµένη από G. Dietrich, 1963, General Oceanography).
Όταν το θαλασσινό νερό παγώνει, µόνο τα µόρια του νερού συµµετέχουν στην κρυστάλλωση.
Τα διαλυµένα άλατα δεν συµµετέχουν και κατά συνέπεια αυξάνουν την αλατότητα του
αποµένοντος θαλασσινού νερού. Η διαδικασία αυτή αναστρέφεται όταν λειώνουν οι πάγοι.
Πήξη και τήξη του θαλασσινού νερού είναι συνήθως εποχιακά φαινόµενα µε αποτέλεσµα
µικρές µεταβολές της αλατότητας.

45
Όταν διαλύονται τα άλατα στο νερό σχηµατίζουν θετικά και αρνητικά ιόντα. Για παράδειγµα,
αν διαλύσουµε το κοινό µαγειρικό αλάτι διασπάται για να σχηµατίσει θετικά ιόντα Νατρίου
(Na+) και αρνητικά ιόντα χλωρίου (Cl-). Τα περισσότερο κοινά ιόντα που βρίσκονται στο
θαλασσινό νερό φαίνονται στον Πίνακα 2.1 και οµαδοποιούνται σε κύρια ή δευτερεύοντα
συστατικά ανάλογα µε την αφθονία τους.
Τα κύρια συστατικά αποτελούν πάνω από το 98% της συνολικής συγκέντρωσης αλάτων στο
θαλασσινό νερό. Η σχέση συγκέντρωσης ιόντων, µεταξύ των κύριων συστατικών, παραµένει
αξιοσηµείωτα σταθερή, ακόµη και όταν η ολική αφθονία τους διαφέρει από τόπο σε τόπο.
Πολύ νωρίς ερευνητές όπως ο Forchhammer, Natterer και ο Dittmar ανακάλυψαν ότι οι
αναλογίες µερικών από τα άλατα στο θαλασσινό νερό παραµένουν σταθερές, ανεξάρτητα
από την συνολική συγκέντρωσή τους στο δείγµα του θαλασσινού νερού. Στα 1884 ο Dittmar
ήταν ο πρώτος που καθόρισε την εντυπωσιακή οµοιοµορφία στις αναλογίες των κύριων
στοιχείων του θαλασσινού νερού αναλύοντας 77 δείγµατα νερού κατά τη διάρκεια της
αποστολής του Challenger: π.χ. το ιόν του Χλωρίου πάντοτε αποτελεί 55,25% όλων των
διαλυµένων στερών σε «κανονικό» θαλασσινό νερό, και το ιόν του Μαγνησίου αποτελεί
πάντοτε 3,75% των διαλυµένων στερών. Η σχετική αφθονία όλων των συστατικών που
αναφέρονται στον Πιν. 2.1 είναι σταθερή σε όλους σχεδόν τους ωκεανούς της γης,
ανεξάρτητα από τις συνολικές αλλαγές στην ολική συγκέντρωση στερεών συστατικών. Αυτή
η σχεδόν σταθερή αναλογία συγκέντρωσης των σπουδαιότερων ιόντων είναι γνωστή ως
Αρχή του Marcet ή Αρχή του Forchhammer.
2.3 Αλατότητα
Αλατότητα (S) είναι ο αριθµός των γραµµαρίων διαλυµένων αλάτων σε 1000 γραµµάρια
θαλασσινού νερού (αφού αντικατασταθεί όλο το βρώµιο από το χλώριο, όλο το ανθρακικό
οξύ µετατραπεί σε οξείδιο και καταστραφεί όλη η οργανική ύλη). Η αλατότητα στην ανοιχτή
θάλασσα κυµαίνεται από 33 µέχρι 39ο/οο.
Εξαιτίας της σταθερότητας των αναλογιών των κύριων συστατικών του θαλασσινού νερού, το
Χλώριο χρησιµοποιείται ως δείκτης αλατότητας. Χλωριότητα (Cl) είναι ο αριθµός των γραµ.
ιόντων Χλωρίου σε 1000 γραµ. θαλασσινού νερού. Η χλωριότητα µετριέται µε τιτλοδότηση µε
νιτρικό Άργυρο (µέθοδος Knudsen) και σχετίζεται µε την αλατότητα µε την εξής συσχέτιση:
S(ο/οο) = 1.80655 Χ Cl(ο/οο)
Άλλη µέθοδος µέτρησης της αλατότητας είναι δια µέσου της αγωγιµότητας του θαλασσινού
νερού. Το θαλασσινό νερό είναι ηλεκτρολύτης και κατά συνέπεια µπορεί να είναι αγωγός
ηλεκτρικού ρεύµατος. Συνεπώς η αγωγιµότητα του θαλασσινού νερού µπορεί να
χρησιµοποιηθεί ως δείκτης αλατότητας. Σε µια συγκεκριµένη θερµοκρασία η ηλεκτρική
αγωγιµότητα αυξάνεται µε την αλατότητα. Οι τεχνικές χρησιµοποίησης της αγωγιµότητας για
την εκτίµηση της αλατότητας είναι πολύ πιο ακριβείς (± 0,003% για αλατότητες 32 έως 38ο/οο)

46
από τις µετρήσεις της χλωριότητας (± 0,02%). Η χρήση των σύγχρονων οργάνων γνωστών
ως CTD βασίζεται στην ιδιότητα της αγωγιµότητας.
Επίσης η αλατότητα µπορεί να µετρηθεί µε τη χρήση των διαθλασιµέτρων, δεδοµένου ότι ο
δείκτης διάθλασης του νερού σχετίζεται άµεσα µε την αλατότητα. Σύγχρονα διαθλασίµετρα
αποτελούν καλή εναλλακτική µέθοδο µέτρησης της αλατότητας στο πεδίο.
Στην ανοικτή θάλασσα η αλατότητα ελέγχεται από την εξάτµιση και το σχηµατισµό θαλάσσιου
πάγου, παράγοντες οι οποίοι αυξάνουν την αλατότητα, και από διαδικασίες όπως η βροχή και
οι εκβολές των ποταµών, που ελαττώνουν την αλατότητα. Μεταβολές των βαθµών εξάτµισης
και βροχοπτώσεων, σε σχέση µε το γεωγραφικό πλάτος, οδηγούν σε δύο µέγιστα αλατότητας
(Εικ. 2.3), που είναι 35,5ο/οο στα πλάτη µεταξύ 30οΒ και 30οΝ.
Ελαφρά υπέρβαση των βροχοπτώσεων, σε σχέση µε την εξάτµιση, έχει ως αποτέλεσµα τα
ελάχιστα της αλατότητας στον Ισηµερινό (34,5ο/οο) και σε πλάτη υψηλότερα των 40ο η
αλατότητα κυµαίνεται µεταξύ 34,0 και 34,5ο/οο. Κατά µήκος των ακτών είναι αντιληπτές
περισσότερο δραµατικές µεταβολές αλατότητας εξαιτίας της επίδρασης των εκβολών των
ποταµών.
Η αλατότητα αυξάνεται σε υδάτινες µάζες όταν περιορισµένη κυκλοφορία των υδάτων
συνοδεύεται από υπερβολική εξάτµιση. Έντονες αλλαγές αλατότητας συµβαίνουν επίσης σε
λιµνοθάλασσες που δηµιουργούνται από τις διακυµάνσεις της στάθµης της θάλασσας
εξαιτίας των παλιρροιών, όπου οι βροχοπτώσεις ή η εξάτµιση µπορούν να µεταβάλλουν το
θαλασσινό νερό από 0,0ο/οο µέχρι πολύ υψηλές συγκεντρώσεις αλάτων.
Σε τροπικές και ηµιτροπικές περιοχές, οι τιµές της αλατότητας είναι υψηλές εξαιτίας του
συνδιασµού των παραγόντων ανέµων, εξάτµισης και της υψηλής ηλιοφάνειας, καθώς και των
χαµηλών βροχοπτώσεων.
Αν και η σταθερή αναλογία των κυρίων στοιχείων στο θαλασσινό νερό παραµένει σταθερή για
τις περισσότερες ανοικτές θάλασσες, οποιαδήποτε προσθήκη ή αφαίρεση υλικών τείνει να
τροποποιήσει την χηµική σύσταση των ωκεανών. Συγκεκριµένα οι κύριοι παράγοντες που
επηρεάζουν την συνολική µάζα διαλυµένων στερεών ή την σύνθεση των διαλυµένων στερεών
στο θαλασσινό νερό είναι : α) εξάτµιση και βροχόπτωση, β) πάγωµα και λιώσιµο του χιονιού
ή των πάγων, γ) εκβολή υδάτων ποταµών, δ) χηµική ανταλλαγή µε την ατµόσφαιρα, ε)
ηφαιστειακή δραστηριότητα, βιολογική δραστηριότητα, και ζ) διάσπαση ραδιενεργών υλικών.
Για τους παράγοντες (α), (β) και (γ) έχουν αναφερθεί στο κεφάλαιο αυτό και θα αναφερθούν
στο επόµενο κεφάλαιο ορισµένα χαρακτηριστικά της επίδρασης τους.
Η χηµική ανταλλαγή µε την ατµόσφαιρα µπορεί να λάβει χώρα δια µέσου της απευθείας
ανταλλαγής ατµοσφαιρικών αερίων µε την θάλασσα, ανάλογα µε τις επικρατούσες φυσικές
συνθήκες της ατµόσφαιρας και της θάλασσας (περισσότερα για το θέµα αυτό δίδονται στο
κεφάλαιο «διαλυµένα αέρια στο θαλασσινό νερό»). Ανταλλαγή µε την ατµόσφαιρα µπορεί να
πραγµατοποιηθεί µε την αποµάκρυνση αλάτων από την επιφάνεια της θάλασσας µέσο του
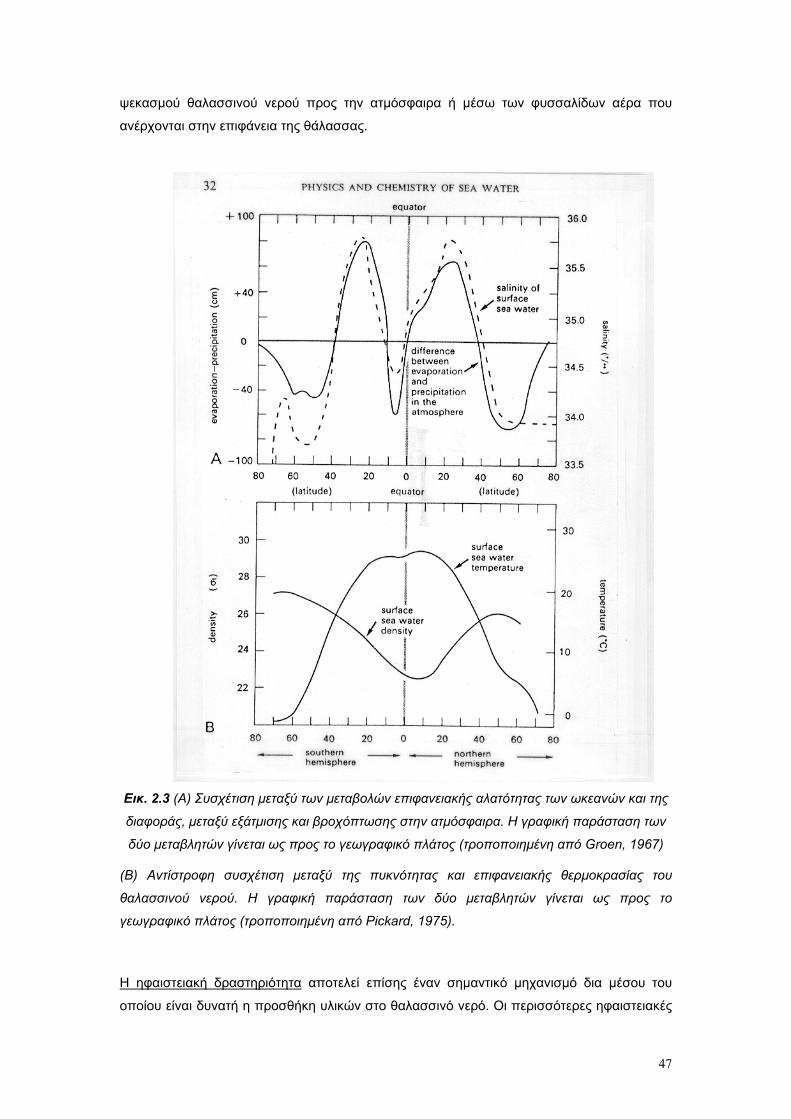
47
ψεκασµού θαλασσινού νερού προς την ατµόσφαιρα ή µέσω των φυσσαλίδων αέρα που
ανέρχονται στην επιφάνεια της θάλασσας.
Εικ. 2.3 (Α) Συσχέτιση µεταξύ των µεταβολών επιφανειακής αλατότητας των ωκεανών και της
διαφοράς, µεταξύ εξάτµισης και βροχόπτωσης στην ατµόσφαιρα. Η γραφική παράσταση των
δύο µεταβλητών γίνεται ως προς το γεωγραφικό πλάτος (τροποποιηµένη από Groen, 1967)
(B) Αντίστροφη συσχέτιση µεταξύ της πυκνότητας και επιφανειακής θερµοκρασίας του
θαλασσινού νερού. Η γραφική παράσταση των δύο µεταβλητών γίνεται ως προς το
γεωγραφικό πλάτος (τροποποιηµένη από Pickard, 1975).
Η ηφαιστειακή δραστηριότητα αποτελεί επίσης έναν σηµαντικό µηχανισµό δια µέσου του
οποίου είναι δυνατή η προσθήκη υλικών στο θαλασσινό νερό. Οι περισσότερες ηφαιστειακές

48
εκροές είναι κυρίως νερό, πολλές όµως περιέχουν υδροχλώριο (HCl), µοριακό υδρογόνο
(Η2), υδροφθόριο (Η2S), µονοξείδιο του άνθρακα (CO), µεθάνιο (CH4), χλωριούχο νάτριο
(NaCl), χλωριούχο σίδηρο (FeCl2), χλωριούχο κάλλιο (KCl), χλωριούχο µαγνήσιο (MgCl2).
Ίσως τα πιο εντυπωσιακά παραδείγµατα επίδρασης αυτού του τύπου µηχανισµού µεταβολής
της σύνθεσης του θαλασσινού νερού είναι η δηµιουργία των θερµών εστιών αλµυρού νερού
στα βαθιά της Ερυθρής θάλασσας. Πολλές θαλάσσιες λεκάνες βρέθηκαν να έχουν στον
πυθµένα τους µάζες νερού ασυνήθιστα υψηλής θερµοκρασίας (22 °C µέχρι 56 °C). Οι υψηλές
αυτές θερµοκρασίες στα βαθιά νερά της Ερυθρής θάλασσας συνοδεύονται από εξαιρετικά
υψηλές αλατότητες που κυµαίνονται από 74‰ µέχρι 255‰. Αυτές οι θερµές µάζες αλµυρού
νερού φαίνεται να ελευθερώνονται περιοδικά από τον θαλάσσιο πυθµένα όπως ακριβώς οι
πίδακες θερµού νερού που βρίσκονται σε ορισµένες περιοχές στην ξηρά.
Η βιολογική δραστηριότητα είναι σηµαντική αιτία µεταβολής της σύνθεσης του θαλασσινού
νερού εξαιτίας των προϊόντων µεταβολισµού των θαλάσσιων οργανισµών. Στις περισσότερες
περιπτώσεις τα στοιχεία που προσλαµβάνονται από τους οργανισµούς επιστρέφουν στο
θαλασσινό νερό όταν πεθαίνουν οι οργανισµοί, ιδιαίτερα όταν τα στοιχεία συγκεντρώνονται
στα µαλακά µέρη των οργανισµών. Εκεί που τα στοιχεία συγκεντρώνονται στα σκληρά µέρη,
αυτά µπορούν είτε να επιστρέψουν στο θαλασσινό νερό ή µπορεί να κατακαθίσουν στον
πυθµένα. Στη δεύτερη περίπτωση αυτές οι σκελετικές κατασκευές µπορούν να αποτελέσουν
συστατικά θαλάσσιων ιζηµάτων και κατά συνέπεια προκαλούν µια σταθερή απώλεια από το
θαλασσινό νερό.
Αν και ποσοτικά φαίνονται µη σηµαντικές οι ποσότητες υλικού που εισάγονται στο θαλασσινό
νερό από άλλες πηγές, τα προϊόντα της ραδιενεργούς αποσύνθεσης για εκατοµµύρια χρόνια
συνεχίζουν να συµβάλλουν στην τελική διαµόρφωση της σύνθεσης των στοιχείων στο
θαλασσινό νερό,
2.4 ∆ιαφοροποιήσεις και ανακατανοµή των συστατικών του θαλασσινού νερού
Περίπου 2.730 εκατοµµύρια µετρικοί τόνοι διαλυµένων στερεών εισρέουν στις θάλασσες και
τους ωκεανούς, από τα ποτάµια κάθε χρόνο. Αυτός ο ρυθµός εισροής αλάτων σε διάλυση
είναι περισσότερο από αρκετός, στην ιστορία της Γης, για να δικαιολογήσει την παρουσία
όλων των διαλυµένων υλικών στους ωκεανούς σήµερα, Όµως, αυτή η πρόταση βασίζεται στις
προϋποθέσεις ότι ο βαθµός εισροής υπήρξε σταθερός, ότι ο φλοιός της Γης αποτελεί την
µόνη πηγή εισροής αλάτων στους ωκεανούς, ότι κανένα από τα υλικά που έφθασαν στην
θάλασσα δεν αποµακρύνθηκε αργότερα. Όµως όταν η σύνθεση του θαλασσινού νερού
συγκρίνεται µε την σύνθεση του φλοιού της Γης, ορισµένες αξιοσηµείωτες διαφορές γίνονται
αντιληπτές. Τα περισσότερα από τα στοιχεία είναι παρόντα σε αναλογίες παρόµοιες µε αυτές
του φλοιού της Γης, όµως, οι σχετικές συγκεντρώσεις ορισµένων στοιχείων στο θαλασσινό
νερό, όπως π.χ. χλωρίου, ιωδίου, βρωµίου, βορίου και µαγνησίου είναι πολύ µεγαλύτερες

49
από όσο στο νερό των ποταµών, οι οποίοι µεταφέρουν διαλυµένα άλατα στη θάλασσα.
Μήπως, υπάρχει κάποια άλλη πηγή αυτών των διαλυµένων αλάτων στo θαλασσινό νερό;
Οι µέσο-ωκεάνιες ράχες πιθανόν να δίνουν απάντηση στο ερώτηµα αυτό. Φρέσκος βασάλτης
ξεχύνεται από τον µανδύα της Γης στα µέσο-ωκεάνια ρήγµατα. Αυτό το βασαλτικό υλικό
συνοδεύεται από «καινούργιο» νερό, δηλαδή νερό που δεν υπήρξε ποτέ πριν στους
ωκεανούς, τα ποτάµια, τις λίµνες ή την ατµόσφαιρα της Γης. Αυτό το καινούργιο νερό
περιέχει, σε διάλυση, πολλά από τα στοιχεία που βρίσκονται στο θαλασσινό νερό, όπως
χλώριο, βρώµιο, ιώδιο, άνθρακα, βόριο, άζωτο και ορισµένα ίχνη στοιχείων. Τα στοιχεία αυτά
που βρίσκονται σε υψηλότατες συγκεντρώσεις σε ηφαιστειακά νερά είναι τα στοιχεία τα οποία
δεν υπολογίζονται στα διαλυµένα υλικά, που µεταφέρονται στους ωκεανούς µε τα ποτάµια.
Εξαιτίας της συνεχούς εισροής και αποµάκρυνσης ορισµένων στοιχείων από το θαλασσινό
νερό, είναι δυνατό να καθορισθεί ο κύκλος ορισµένων από αυτά τα στοιχεία. Ένα τµήµα του
κύκλου αντιπροσωπεύει το χρόνο κατά τον οποίο το στοιχείο παραµένει στο θαλασσινό νερό.
Η χρονική περίοδος που περιέχεται το στοιχείο στη θάλασσα αναφέρεται ως «χρόνος
παραµονής» (residence time). Οι χρόνοι παραµονής των στοιχείων ποικίλουν, ανάλογα µε
τον βαθµό στον οποίο τα στοιχεία αυτά εισέρχονται στις βιολογικές ή φυσικές διαδικασίες οι
οποίες τείνουν να τα αποµακρύνουν από το διάλυµα και τη θάλασσα. Όπου οι χρόνοι
παραµονής είναι µεγάλοι, τα στοιχεία παρουσιάζουν µικρή µεταβολή στη σχετική
συγκέντρωση τους στο θαλασσινό νερό. Εκεί που οι χρόνοι παραµονής είναι µικροί, τα
στοιχεία τείνουν να εµφανίζουν µια πιο σποραδική κατανοµή, ως συνάρτηση της συµµετοχής
τους στις διαδικασίες αποµάκρυνσης. Π.χ. ο χρόνος παραµονής του νατρίου είναι 21 – 26
εκατοµµύρια χρόνια, είναι ο µεγαλύτερος χρόνος παραµονής από όλα τα διαλυµένα στοιχεία
στο θαλασσινό νερό. Σε αντίθεση ο χρόνος παραµονής του αλουµινίου είναι 100 χρόνια, που
πιθανόν είναι ο συντοµότερος όλων των χηµικών στοιχείων.
Ένας άλλος όρος που χρησιµοποιείται στην ωκεανογραφία είναι ο «χρόνος ανάµειξης». Ο
όρος αναφέρεται στον χρόνο που χρειάζεται ώστε τα συστατικά του παγκόσµιου ωκεανού να
αναµειχθούν πλήρως. Είναι δηλαδή ο χρόνος που περνάει ώστε, ένα σωµατίδιο νερού να
περάσει από την επιφάνεια του ωκεανού, στα ενδιάµεσα βάθη και να επανέλθει πάλι στην
επιφάνεια. Έτσι ένα «τυπικό» σωµατίδιο νερού το οποίο εγκαταλείπει την επιφάνεια του
ωκεανού χρειάζεται περίπου 1.000 χρόνια για να επανέλθει στην επιφάνεια και πάλι. Κατά
συνέπεια µπορούµε να πούµε ότι ο «χρόνος ανάµειξης» του ωκεανού είναι περίπου 1.000
χρόνια.
Η συγκέντρωση πολλών από τα δευτερεύοντα στοιχεία του θαλασσινού νερού ποικίλουν σε
συνάρτηση µε τις φυσικές και βιολογικές διεργασίες, όπως οι διάφορες θερµοκρασίας ή η
κατανοµή των θαλασσινών ειδών. Π.χ. η ποσότητα του ανθρακικού ασβεστίου και του
πυριτίου στο θαλασσινό νερό ρυθµίζεται από οργανισµούς όπως τα δισεκατοµµύρια του
µικροσκοπικού πλαγκτού τα οποία προσλαµβάνουν αυτά τα υλικά και προκαλούν την
συγκέντρωση µεγάλων ποσοτήτων ανθρακικών και πυριτικών ιζηµάτων στον πυθµένα των
θαλασσών. Τα περισσότερα από τα υλικά που συγκεντρώνονται στα µαλακά τµήµατα των

50
θαλασσινών οργανισµών, συνήθως επιστρέφουν σε διαφορετική θέση από εκείνη από την
οποία προήλθαν, προκαλώντας έτσι ανακατανοµή στη θάλασσα. ∆εδοµένου ότι το πλαγκτόν
και άλλοι οργανισµοί βυθίζονται µετά τον θάνατό τους, η ανακατανοµή αυτών των θρεπτικών
γίνεται συνήθως προς τα κάτω.
Τα µέγιστα και ελάχιστα των νιτρικών και φωσφορικών σε ορισµένες περιοχές συµβαίνουν
κατά τους ίδιους µήνες. Ο φωσφόρος επίσης βρίσκεται στην πιο υψηλή συγκέντρωση κατά τη
διάρκεια των χειµερινών µηνών και στη χαµηλότερη κατά την διάρκεια των εαρινών και
θερινών µηνών. Η διαθεσιµότητα, κατά συνέπεια, των νιτρικών και του φωσφόρου είναι
σηµαντική προϋπόθεση για την αύξηση ενός µεγάλου πληθυσµού πλαγκτού. Αλλαγές στην
ωκεάνια κυκλοφορία, οι οποίες τροποποιούν τις ποσότητες αυτών των θρεπτικών που
µεταφέρονται σε µια περιοχή, µπορούν αν καθορίσουν σε µεγάλο ποσοστό την πυκνότητα
των οργανισµών του εν λόγω πληθυσµού πλαγκτού για µια συγκεκριµένη εποχή.
ΟΙ εποχιακές διαφοροποιήσεις που προκύπτουν από την βιολογική δραστηριότητα είναι πιο
έντονες στην εύφωτη ζώνη (0 – 80 µ βάθος) όπου το φυτοπλαγκτόν είναι άφθονο.
Η κατανοµή των φωσφορικών στους ωκεανούς ποικίλει όχι µόνο µε την γεωγραφική θέση
αλλά επίσης και µε το βάθος. Τα επάνω στρώµατα στους ωκεανούς κατά µέσον όρο
στερούνται φωσφόρου διότι οι οργανισµοί αυτοί, και ιδιαίτερα το πλαγκτόν χρησιµοποιούν τα
φωσφορικά. Οι µεγαλύτερες συγκεντρώσεις στον Ατλαντικό, Ειρηνικό και Ινδικό ωκεανό
παρατηρούνται στα µέσα πλάτη και σε βάθη τα οποία κυµαίνονται από 1.000 µ µέχρι τον
πυθµένα.
Οι τιµές του πυριτίου επίσης κυµαίνονται µε την εποχή, και συγκεκριµένα εµφανίζονται
υψηλές συγκεντρώσεις κατά τη διάρκεια του χειµώνα και χαµηλότερες συγκεντρώσεις κατά τη
διάρκεια του θέρους. Η έλλειψη πυριτίου στα επιφανειακά νερά κατά τη διάρκεια του θέρους,
εξαρτάται όχι µόνο από τις µεταβολικές διαδικασίες του πλαγκτού αλλά επίσης και από το
γεγονός ότι πολλά πυριτιούχα κελύφη βυθίζονται και κατά συνέπεια χάνονται από την
επιφανειακή ζώνη. Μερικά κελύφη καθιζάνουν και ενσωµατώνονται µε την πυριτιούχα λάσπη
ή άλλα διαλύονται σε βαθύτερα νερά πριν φθάσουν στον πυθµένα. Όπου τα στοιχεία πυρίτιο,
άζωτο, και φωσφόρος ελλείπουν από το θαλασσινό νερό, τα φυτά που εξαρτώνται από αυτά
σταµατούν να ανθίζουν και τα ζώα που είναι υψηλότερα στην τροφική αλυσίδα, τα οποία
κανονικά τρέφονται από τα φυτά, ελλείπουν.
2.5 Η αλατότητα ως οικολογικός παράγοντας
Η αλατότητα στις περιοχές παλιρροιών και εκβολών ποταµών µπορεί να παρουσιάζει
προβλήµατα στους θαλάσσιους οργανισµούς εξαιτίας των φυσικών διαδικασιών της διάχυσης
και όσµωσης. ∆ιάχυση δηµιουργείται όταν οι συγκεντρώσεις µιας διαλυµένης ουσίας είναι
άνισες µεταξύ των δυο πλευρών µιας µεµβράνης εκλεκτικής διαπερατότητας. Τα ιόντα
Νατρίου διαχέονται δια µέσου της µεµβράνης µέχρι να γίνει εξίσωση των συγκεντρώσεων.
Ορισµένοι θαλάσσιοι οργανισµοί όµως πρέπει να ενεργούν δραστικά προκειµένου να
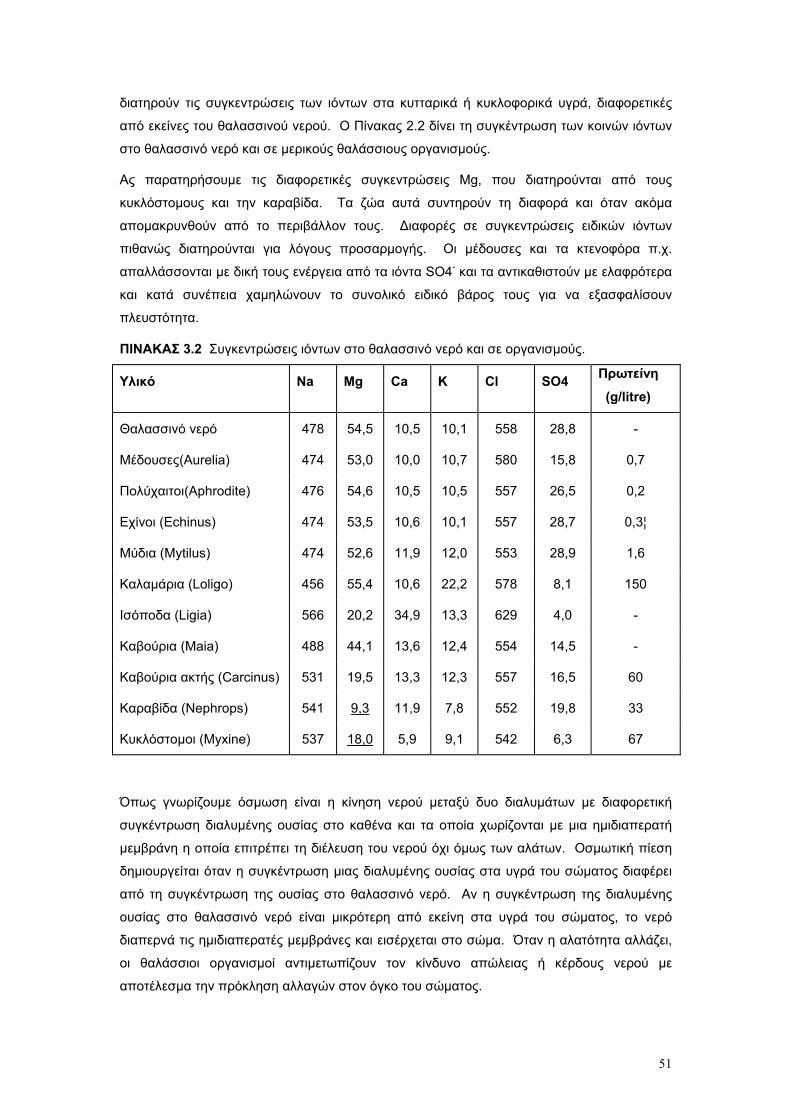
51
διατηρούν τις συγκεντρώσεις των ιόντων στα κυτταρικά ή κυκλοφορικά υγρά, διαφορετικές
από εκείνες του θαλασσινού νερού. Ο Πίνακας 2.2 δίνει τη συγκέντρωση των κοινών ιόντων
στο θαλασσινό νερό και σε µερικούς θαλάσσιους οργανισµούς.
Ας παρατηρήσουµε τις διαφορετικές συγκεντρώσεις Mg, που διατηρούνται από τους
κυκλόστοµους και την καραβίδα. Τα ζώα αυτά συντηρούν τη διαφορά και όταν ακόµα
αποµακρυνθούν από το περιβάλλον τους. ∆ιαφορές σε συγκεντρώσεις ειδικών ιόντων
πιθανώς διατηρούνται για λόγους προσαρµογής. Οι µέδουσες και τα κτενοφόρα π.χ.
απαλλάσσονται µε δική τους ενέργεια από τα ιόντα SO4- και τα αντικαθιστούν µε ελαφρότερα
και κατά συνέπεια χαµηλώνουν το συνολικό ειδικό βάρος τους για να εξασφαλίσουν
πλευστότητα.
ΠΙΝΑΚΑΣ 3.2 Συγκεντρώσεις ιόντων στο θαλασσινό νερό και σε οργανισµούς.
Υλικό Na Mg Ca K Cl SO4 Πρωτείνη
(g/litre)
Θαλασσινό νερό 478 54,5 10,5 10,1 558 28,8 -
Μέδουσες(Aurelia) 474 53,0 10,0 10,7 580 15,8 0,7
Πολύχαιτοι(Aphrodite) 476 54,6 10,5 10,5 557 26,5 0,2
Εχίνοι (Echinus) 474 53,5 10,6 10,1 557 28,7 0,3¦
Μύδια (Mytilus) 474 52,6 11,9 12,0 553 28,9 1,6
Καλαµάρια (Loligo) 456 55,4 10,6 22,2 578 8,1 150
Ισόποδα (Ligia) 566 20,2 34,9 13,3 629 4,0 -
Καβούρια (Maia) 488 44,1 13,6 12,4 554 14,5 -
Kαβούρια ακτής (Carcinus) 531 19,5 13,3 12,3 557 16,5 60
Kαραβίδα (Nephrops) 541 9,3 11,9 7,8 552 19,8 33
Κυκλόστοµοι (Myxine) 537 18,0 5,9 9,1 542 6,3 67
Όπως γνωρίζουµε όσµωση είναι η κίνηση νερού µεταξύ δυο διαλυµάτων µε διαφορετική
συγκέντρωση διαλυµένης ουσίας στο καθένα και τα οποία χωρίζονται µε µια ηµιδιαπερατή
µεµβράνη η οποία επιτρέπει τη διέλευση του νερού όχι όµως των αλάτων. Οσµωτική πίεση
δηµιουργείται όταν η συγκέντρωση µιας διαλυµένης ουσίας στα υγρά του σώµατος διαφέρει
από τη συγκέντρωση της ουσίας στο θαλασσινό νερό. Αν η συγκέντρωση της διαλυµένης
ουσίας στο θαλασσινό νερό είναι µικρότερη από εκείνη στα υγρά του σώµατος, το νερό
διαπερνά τις ηµιδιαπερατές µεµβράνες και εισέρχεται στο σώµα. Όταν η αλατότητα αλλάζει,
οι θαλάσσιοι οργανισµοί αντιµετωπίζουν τον κίνδυνο απώλειας ή κέρδους νερού µε
αποτέλεσµα την πρόκληση αλλαγών στον όγκο του σώµατος.

52
Ένα διάλυµα λέγεται ότι είναι υπεροσµωτικό όταν νερό ρέει προς το διάλυµα δια µέσου µιας
ηµιδιαπερατής µεµβράνης. Το διάλυµα αντίθετα είναι υποοσµωτικό όταν το νερό φεύγει από
το διάλυµα προς τα έξω. Το αποτέλεσµα της όσµωσης, που δηµιουργείται από διαλυµένα
συστατικά στα υγρά του σώµατος, µπορεί να εκτιµηθεί από τον υποβιβασµό του σηµείου
πήξεως κάτω του 0oC.
Οι µηχανισµοί προσαρµογής στις αλλαγές των συνθηκών όσµωσης και ιοντικών µεταβολών
ποικίλουν πάρα πολύ µεταξύ των θαλάσσιων οργανισµών. Μερικά θαλάσσια ζώα είναι
σταθεροοσµωτικά και άλλα είναι ποικιλοοσµωτικά. Τα σταθεροοσµωτικά δεν ρυθµίζουν ούτε
τον όγκο των κυττάρων ή του σώµατος ούτε τα ιοντικά συστατικά. Εδώ θα πρέπει να τονιστεί
η διάκριση µεταξύ οσµωτικών προβληµάτων και της ανάγκης να ρυθµίζονται οι
συγκεντρώσεις ειδικών ιόντων. Οι οργανισµοί που διατηρούν τη σύνθεση των ιόντων ριζικά
διαφορετική από εκείνη του θαλασσινού νερού (π.χ. οι τελεόστεοι ιχθείς) πρέπει να
προσαρµόζονται και στα δυο προβλήµατα. Οι περισσότεροι οργανισµοί εµφανίζουν
µεταβαλλόµενους βαθµούς οσµωτικής ρύθµισης και ιοντικής ρύθµισης. Μερικοί οργανισµοί,
όπως τα καρκινοειδή, έχουν σχετικά αδιαπέραστο τοίχωµα σώµατος και ρυθµίζουν έντονα τις
ανταλλαγές νερού και ιόντων σε διαπερατά µόνον σηµεία, όπως είναι τα βράγχια και τα
όργανα απέκκρισης.
Για να διαφωτίσουµε τα βασικά προβλήµατα των αλάτων και της ισορροπίας νερού σε
θαλάσσιους οργανισµούς ας εξετάσουµε δύο αντιπροσωπευτικά ζώα: ένα ολοθούριο και ένα
σολοµό. (Εικ. 2.4).
ΕΙΚ. 2.4 Σύγκριση των προβληµάτων ώσµωσης ενός θαλάσσιου Ολοθούριου και ενός
σολοµού που µεταφέρονται από το θαλασσινό στο γλυκό νερό (τροποποιηµένη από J. L.
Sumich, 1976).

53
Τα θαλάσσια ολοθούρια αποφεύγουν προβλήµατα άλατος και ισορροπίας νερού διατηρώντας
απλά ένα εσωτερικό υγρό το οποίο είναι χηµικά παρόµοιο προς εκείνο του θαλασσινού νερού
(περίπου 35ο/οο διαλυµένα άλατα). ∆εν υπάρχει πρόβληµα για τα ολοθούρια όταν οι
συγκεντρώσεις των υγρών στις δύο πλευρές µεταξύ των διαχωριστικών µεµβρανών είναι ίσες
και δεν παρατηρείται κλίση διαφοράς συγκέντρωσης (αυτό είναι γνωστό ως φαινόµενο
ισοτονικών συνθηκών). Η κατάσταση ισορροπίας διατηρείται όταν το νερό διαχέεται έξω από
το ολοθούριο τόσο γρήγορα όσο γρήγορα εισέρχεται και η συγκέντρωση αλάτων των
εσωτερικών υγρών παραµένει ίση προς εκείνη του θαλασσινού νερού έξω από το ζώο.
Όταν το ολοθούριο µεταφερθεί εκτός του θαλασσινού νερού και τοποθετηθεί σε γλυκό νερό
παρατηρούνται τα εξής : Η συγκέντρωση αλάτων στην περίπτωση αυτή είναι µεγαλύτερη
εσωτερικά του ζώου (35ο/οο) από εκείνη εξωτερικά. Τα υγρά του σώµατος είναι τώρα
υπερτονικά σε σχέση µε το περιβάλλον. Τα µόρια του νερού, ακολουθώντας την κλίση
συγκέντρωσης διαχέονται δια µέσου των επιλεκτικά διαπερατών µεµβρανών του ολοθούριου.
Η κίνηση του νερού δια µέσου αυτών των µεµβρανών είναι το γνωστό φαινόµενο της
όσµωσης. Τα διαλυµένα άλατα τώρα, περισσότερο συγκεντρωµένα µέσα στο ζώο από όσο
έξω, δεν µπορούν να διαχέονται έξω από το ολοθούριο δεδοµένου ότι µια τέτοια κίνηση
µπλοκάρεται από την ηµιδιαπερατότητα των µεµβρανών σ' αυτά τα άλατα. Το καθαρό
αποτέλεσµα είναι αύξηση του ποσού του νερού µέσα στο ολοθούριο. Το επιπλέον νερό
δηµιουργεί µια εσωτερική οσµωτική πίεση, η οποία είναι δυναµικά επιβλαβής εφόσον το ζώο
δεν έχει την δυνατότητα να αποβάλλει την περίσσεια του νερού. Τα περισσότερα θαλασσινά
ασπόνδυλα και πολλά θαλασσινά φυτά έχουν λίγη ή καµία δυνατότητα να αντιµετωπίζουν
τέτοιο οσµωτικό στρες. Κατά συνέπεια τέτοιοι οργανισµοί περιορίζονται σε περιοχές όπου η
αλατότητα διακυµαίνεται λίγο από τις συνθήκες της ανοικτής θάλασσας.
Σε αντίθεση µε τον περιορισµένο έλεγχο τον οποίο έχει η οσµωτική κατάσταση των
ολοθουρίων, οι τελεόστεοι και µερικά άλλα θαλασσινά ζώα και φυτά έχουν καλά
αναπτυγµένους µηχανισµούς οσµωτικής ρύθµισης. Ως ένα δώρο της φύσης µερικοί απ'
αυτούς τους οργανισµούς είναι ελεύθεροι να µετακινούνται µεταξύ περιοχών διαφορετικής
αλατότητας µη εµποδιζόµενοι από οσµωτικές µεταβολές. Για παράδειγµα αναφέρουµε το
σολοµό, ο οποίος περνάει µέρος της ζωής του στο θαλασσινό νερό και το υπόλοιπο στο
γλυκό νερό, για να διευκρινισθεί πώς µερικοί οργανισµοί διατηρούν ένα οµοιοστατικό
εσωτερικό σύστηµα υγρών, ανεξάρτητα από τις συνθήκες του περιβάλλοντος.
Η συγκέντρωση των αλάτων στα υγρά του σώµατος του σολοµού, όπως και στους
περισσότερους οστειχθύες, είναι ενδιάµεση µεταξύ του γλυκού και του θαλασσινού νερού
(περίπου 18ο/οο). Συνεπώς τα υγρά του σώµατος είναι υποτονικά σε σχέση µε το θαλασσινό
νερό. Έτσι τα ψάρια αυτά ποτέ δεν επιτυγχάνουν οσµωτική ισορροπία µε το εξωτερικό τους
περιβάλλον. Αντίθετα πρέπει να ξοδεύουν συνεχώς ενέργεια για να διατηρούν σταθερή
εσωτερική οσµωτική κατάσταση αρκετά διαφορετική και από τον ποταµό και από τον ωκεανό.
Στο θαλασσινό νερό ο σολοµός χάνει νερό από το σώµα του δια της οσµώσεως και έτσι τα
ψάρια αυτά αντιµετωπίζουν συνεχώς προβλήµατα αφυδάτωσης, αν και περιβάλλονται από

54
ωκεάνιο νερό. Για να αντιµετωπίσει αυτό ο σολοµός πίνει µεγάλες ποσότητες θαλασσινού
νερού που απορροφώνται από τον πεπτικό αγωγό και έτσι το νερό διατηρείται στους ιστούς
του σώµατος. Η περίσσεια των αλάτων εκκρίνεται από ειδικά χλωρο-κύτταρα, που είναι
τοποθετηµένα στα βράγχια. Τα νεφρά του σολοµού δεν µπορούν να παράγουν ουρία µε
συγκέντρωση αλάτων µεγαλύτερη από εκείνη των υγρών του σώµατος, κατά συνέπεια δε
χρησιµοποιούνται για την απαλλαγή της περίσσειας αλάτων. Για το λόγο αυτόν όταν ο
σολοµός βρίσκεται στα ποτάµια και στις λίµνες τα οσµωτικά του προβλήµατα αντιστρέφονται
εντελώς. Εδώ το πρόβληµα είναι η πρόσληψη οσµωτικού νερού δια µέσου των βραγχίων και
των µεµβρανών του πεπτικού συστήµατος και µια σταθερή απώλεια αλάτων προς το
περιβάλλον νερό. Ο σολοµός πίνει πολύ λίγο γλυκό νερό, αν και µερικό καταπίνεται και κατά
την λήψη της τροφής. Για να ισορροπήσουν την είσοδο του νερού, τα νεφρά παράγουν
άφθονες ποσότητες αραιάς ουρίας µετά από αποτελεσµατική επαναπρόσληψη των
περισσότερων από τα άλατα της ουρίας. Τα αναγκαία άλατα προσλαµβάνονται από την
τροφή, άλατα όµως απορροφώνται δραστικά και από το περιβάλλον νερό µέσω ειδικών
κυττάρων στα βράγχια. Έτσι µε κατανάλωση σηµαντικής ενέργειας, ο σολοµός διατηρεί ένα
οµοιοστατικό εσωτερικό περιβάλλον υγρών τόσο στο ποτάµι όσο και στον ωκεανό.
Αν βυθίσοµε κοµµάτια θαλλού του θαλασσινού φύκους Porphyra tenera σε αραιωµένο
θαλασσινό νερό τα κυτταρικά τοιχώµατα παίρνουν νερό και έτσι προκαλείται επιµήκυνση των
θαλλών. Ένα παρόµοιο πείραµα που πραγµατοποιήθηκε µε το σκουλήκι Confingia gouldii
(Sipunculida) είχε ως αποτέλεσµα αρχικά να φουσκώσει το σώµα αλλά µετά να ακολουθήσει
µείωση του όγκου, αρχίζοντας µία ηµέρα µετά. Η ρύθµιση του όγκου επιτυγχάνεται µε την
αποβολή αλάτων δια µέσου του νεφρικού συστήµατος. Καθώς τα άλατα αποµακρύνονται, το
νερό αποβάλλεται από τα τοιχώµατα του σώµατος προς το θαλασσινό νερό.
Τα δίθυρα µαλάκια, µεταξύ των άλλων οµάδων ασπόνδυλων, δεν ρυθµίζουν οσµωτικά
εξωκυτταρικά υγρά ρυθµίζουν όµως τον οσµωτικό χαρακτήρα των ενδοκυτταρικών υγρών.
Αυτά επιτυγχάνουν σταθερό όγκο κυττάρων ρυθµίζοντας τη συγκέντρωση των διαλυµένων
αµινοξέων. Το περιεχόµενο των ελεύθερων αµινοξέων αυξάνεται και µειώνεται για να
προσαρµοστεί σε αντίστοιχες αυξήσεις και µειώσεις της αλατότητας.
Αν και η ανταπόκριση στις µεταβολές του όγκου µπορεί να είναι αρκετά γρήγορη,
εγκλιµατισµός στις µεταβολές της αλατότητας είναι µια βραδεία διαδικασία συγκριτικά µε τον
εγκλιµατισµό στις µεταβολές της θερµοκρασίας. Όταν µύδια (Mytilus edulis) εµβαπτίζονται σε
αραιωµένο θαλασσινό νερό, τα κελύφη τους αρχικά κλείνουν και ο ρυθµός της αναπνοής
µειώνεται. Μετά, καθώς τα κελύφη ανοίγουν, επιτυγχάνεται καινούριος ρυθµός αναπνοής όχι
όµως πριν περάσουν 4-7 εβδοµάδες. Εξάλλου η ανταπόκριση ενός οργανισµού στις αλλαγές
της αλατότητας ποικίλει πάρα πολύ, ακόµη και στο ίδιο είδος. Αν ένα µύδι τοποθετηθεί σ' ένα
πειραµατικό µέσον µε αλατότητα διαφορετική από εκείνη του περιβάλλοντος από το οποίο
προήλθε, η κατανάλωση οξυγόνου µειώνεται. Εντούτοις όταν οι ρυθµοί κατανάλωσης
οξυγόνου µετρούνται σε φυσικούς πληθυσµούς, που συλλέγονται µέσα από ένα ευρύ πεδίο

55
φυσικών αλατοτήτων (5-30o/oo) διαπιστώνεται ότι οι ρυθµοί είναι το ίδιο ποικίλοι. Μια γενετική
πιθανώς διαφορά ή µια διαδικασία εγκλιµατισµού θα µπορούσε να εξηγήσει το φαινόµενο.
Οι τελεόστεοι ιχθείς εµφανίζουν οσµωτική ρύθµιση διατηρώντας τα υγρά του σώµατός τους
σε συγκεντρώσεις ίσες προς το 1/3 έως 1/4 εκείνων του φυσικού θαλασσινού νερού.
Τα χέλια Anguilla anguilla (Ευρωπαϊκό χέλι) και A. rostrata (Αµερικανικό χέλι) αναπαράγονται
στη θάλασσα των Σαργασών, ενώ τα νεαρά άτοµα επιστρέφουν σε αλµυρούς υγρότοπους και
άλλες παράκτιες περιοχές για να καταλήξουν σε γλυκά νερά όπου και διαβιούν µέχρι να
αρχίσουν την προετοιµασία της αναπαραγωγής.
Το ψάρι Fundulus heteroclitus µπορεί να ζήσει τόσο στο γλυκό νερό όσο και στο θαλασσινό.
Πολλά είδη της οικογένειας των Salmonidae γεννιούνται στο γλυκό νερό, µεταναστεύουν στο
θαλασσινό νερό και επιστρέφουν στο γλυκό νερό για την αναπαραγωγή τους. Στα
ελασµαβράγχια, όπως οι καρχαρίες και οι ράγιες, η υψηλή συγκέντρωση ουρίας
χρησιµοποιείται για να διατηρείται η οσµωτική ισορροπία, όπως στην περίπτωση των
ελεύθερων αµινοξέων στα δίθυρα µαλάκια. Οι καρχαρίες και οι ράγιες επίσης αποβάλλουν
ιόντα όπως Na+
Oι συγκεντρώσεις αλάτων στο αίµα επηρεάζονται από το στέγνωµα του ζώου καθώς επίσης
και από την αλατότητα του θαλασσινού νερού. Αυτό είναι ένα σοβαρό πρόβληµα για τους
οργανισµούς που ζουν στο ανώτερο σηµείο της µεσοπαλιρροικής ζώνης, όπου η επί µακρόν
έκθεσή τους µπορεί να έχει ως αποτέλεσµα υπερβολική εξάτµιση των υγρών τους. Με την
απώλεια νερού, οι συγκεντρώσεις ιόντων αυξάνονται και το γεγονός αυτό θέτει σε κίνδυνο τις
βιοχηµικές διεργασίες, οι οποίες επηρεάζονται από την ιοντική συγκέντρωση. Οι οργανισµοί
που ζουν στην ανωτέρα µεσοπαλιρροική ζώνη εµφανίζουν µειωµένη απώλεια νερού και
αυξηµένη ανθεκτικότητα στην ξηρασία, συγκριτικά µε τους οργανισµούς της κατώτερης
µεσοπαλιρροικής ζώνης.
2.6 Πυκνότητα του θαλασσινού νερού
∆εδοµένου ότι η πυκνότητα είναι συνάρτηση κυρίως της αλατότητας, θεωρήθηκε σκόπιµο στο
κεφάλαιο αυτό να συµπεριληφθεί και η ανάλυση της έννοιας της πυκνότητας του θαλασσινού
νερού. Ο όρος «πυκνότητα» συνήθως χρησιµοποιείται στην ωκεανογραφία στη θέση του
όρου «ειδικού βάρους» για τη σύγκριση δειγµάτων θαλασσινού νερού. Κατά συνέπεια η
πυκνότητα του θαλασσινού νερού αναφέρεται στον λόγο της µάζας ανά µονάδα όγκου
θαλασσινού νερού σε δεδοµένη θερµοκρασία προς την µάζα ανά µονάδα όγκου
αποσταγµένου νερού 4 °C (στην ατµοσφαιρική πίεσης). ∆εδοµένου ότι τόσο η θερµοκρασία
όσο και η αλατότητα ποικίλουν, η πυκνότητα του θαλασσινού νερού µπορεί να κυµαίνεται σε
ένα εύρος από 1,0000 στους 4°C και αλατότητα σχεδόν 0,0 ‰ µέχρι 1,0322 στους 0 °C και
αλατότητα 40 ‰. Αυτή η ευρεία διαφοροποίηση των θερµοκρασιών και της αλατότητας σε
διαφορετικά τµήµατα του ωκεανού υποστηρίζουν την ιδέα ότι η τιµή της πυκνότητας µπορεί

56
να χρησιµοποιείται για την ταυτοποίηση συγκεκριµένων µαζών νερού. Όµως, αφού η
πυκνότητα είναι συνάρτηση της θερµοκρασίας και αλατότητας, νερά µε παρόµοιες
πυκνότητες, µπορεί στην πραγµατικότητα, να έχουν τελείως διαφορετικές αλατότητες και
θερµοκρασίες. Για το λόγο αυτό, εκείνο που καθορίζει την ταυτότητα µιας µάζας νερού δεν
είναι η πυκνότητα αλλά είναι η θερµοκρασία και η αλατότητα.
Για το απεσταγµένο νερό η πυκνότητα αυξάνει µε την µείωση της θερµοκρασίας στην µέγιστη
τιµή 1,000 στους 4 °C µετά µειώνεται µέχρι του σηµείου πήξεως στους 0 °C (Εικ 2.5). Μια
παρόµοια αλλαγή παρατηρείται στο θαλασσινό νερό µε αλατότητα π.χ. 15 ‰. Αλλά εφόσον
σύµφωνα µε το διάγραµµα της Εικ 2.5 η θερµοκρασία της µέγιστης πυκνότητας του
θαλασσινού νερού µειώνεται ταχύτερα από όσο κατέρχεται το σηµείο πήξεως καθώς η
αλατότητα αυξάνει, η θερµοκρασία της µέγιστης πυκνότητας και το σηµείο πήξεως
συµπίπτουν στους -1,322 °C στην κρίσιµη τιµή αλατότητας 24,7‰. Σε αλατότητες
µεγαλύτερες από 24,7 ‰ η πυκνότητα του θαλασσινού νερού µειώνεται καθώς η
θερµοκρασία αυξάνει. Γενικά, κρύο αλµυρό νερό έχει συγκριτικά υψηλή πυκνότητα ενώ θερµό
ελαφρώς αλµυρό νερό έχει συγκριτικά χαµηλή πυκνότητα.
Για ευκολία οι ωκεανογράφοι χρησιµοποιούν διεθνώς το γράµµα «ρ» για να δηλώσουν την
πυκνότητα του θαλασσινού νερού. ∆εδοµένου ότι η πυκνότητα του θαλασσινού νερού είναι
συνάρτηση της πίεσης (P) , της αλατότητας (S) και της θερµοκρασίας (T) το σύµβολο αυτό
γράφεται πολύ συχνά ως pS.T.P
Παρά το γεγονός ότι η πυκνότητα εκφράζεται σε γραµµάρια ανά κυβικό εκτµ. (οι τιµές
κυµαίνονται από 1,024 µέχρι 1,030 g cm-3 στην ανοικτή θάλασσα) η παρακάτω εξίσωση είναι
η περισσότερο χρησιµοποιούµενη διότι έχει έννοια ωκεανογραφική:
σS.T.P = (density -1) x 103
Εποµένως, νερό µε πυκνότητα ίση προς 1,024 θα έχει σS.T.P ίση προς 24. Η παράµετρος
αυτή εκφράζει την πυκνότητα σε δεδοµένη αλατότητα (S), θερµοκρασία (Τ) και πίεση (P) και
συνήθως γράφεται ως σΤ και διεθνώς καλείται "σίγµα τ ". To σΤ, αν δεν καθορίζεται µε
κάποιο πρόσθετο χαρακτηριστικό, αναφέρεται στην πυκνότητα του νερού, µιας δεδοµένης
αλατότητας και θερµοκρασίας, σε πίεση µιας (1) ατµόσφαιρας.
Η πυκνότητα αυξάνεται µε την αύξηση της αλατότητας και µε τη µείωση της θερµοκρασίας.
Σε υψηλότερες θερµοκρασίες οι αλλαγές της θερµοκρασίας έχουν περισσότερο έντονη
επίδραση στην πυκνότητα από όση έχουν στις χαµηλότερες θερµοκρασίες. ∆εδοµένου ότι
τόσο η θερµοκρασία όσο και η αλατότητα µεταβάλλονται µε το βάθος, είναι συχνά χρήσιµο να
γίνεται γραφική παράσταση της «σ» ως συνάρτηση του βάθους (Εικ. 2.6.Γ).
Ο συνδυασµός των µεταβολών της αλατότητας και θερµοκρασίας, λόγω γεωγραφικού
πλάτους, έχει ως αποτέλεσµα µια ελάχιστη πυκνότητα στην επιφάνεια των υδάτων κοντά
στον Ισηµερινό, που αυξάνεται προς Βορρά και προς Νότο (Εικ.2.3.Β). Οι ζώνες βάθους
ταχείας µεταβολής της θερµοκρασίας (θερµοκλινές) και της αλατότητας (αλοκλινές) πρέπει να
είναι επίσης περιοχές µεταβολών της πυκνότητας (πυκνοκλινές). Όταν νερό χαµηλής
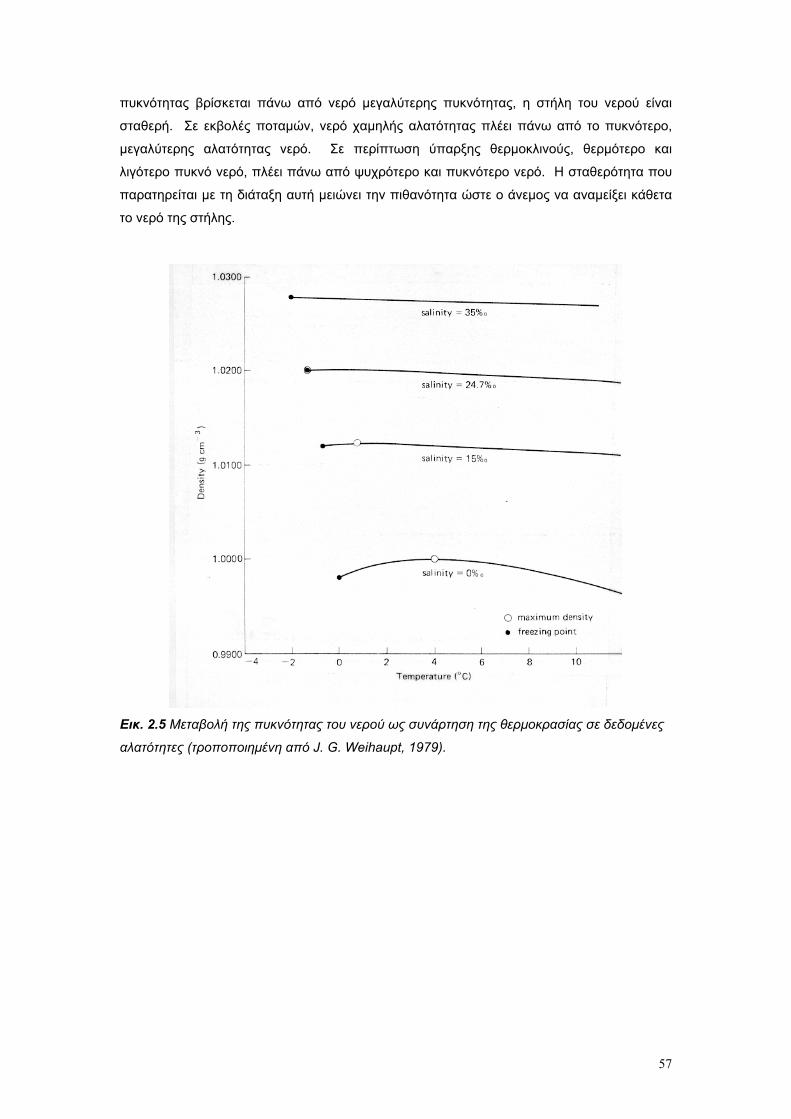
57
πυκνότητας βρίσκεται πάνω από νερό µεγαλύτερης πυκνότητας, η στήλη του νερού είναι
σταθερή. Σε εκβολές ποταµών, νερό χαµηλής αλατότητας πλέει πάνω από το πυκνότερο,
µεγαλύτερης αλατότητας νερό. Σε περίπτωση ύπαρξης θερµοκλινούς, θερµότερο και
λιγότερο πυκνό νερό, πλέει πάνω από ψυχρότερο και πυκνότερο νερό. Η σταθερότητα που
παρατηρείται µε τη διάταξη αυτή µειώνει την πιθανότητα ώστε ο άνεµος να αναµείξει κάθετα
το νερό της στήλης.
Εικ. 2.5 Μεταβολή της πυκνότητας του νερού ως συνάρτηση της θερµοκρασίας σε δεδοµένες
αλατότητες (τροποποιηµένη από J. G. Weihaupt, 1979).
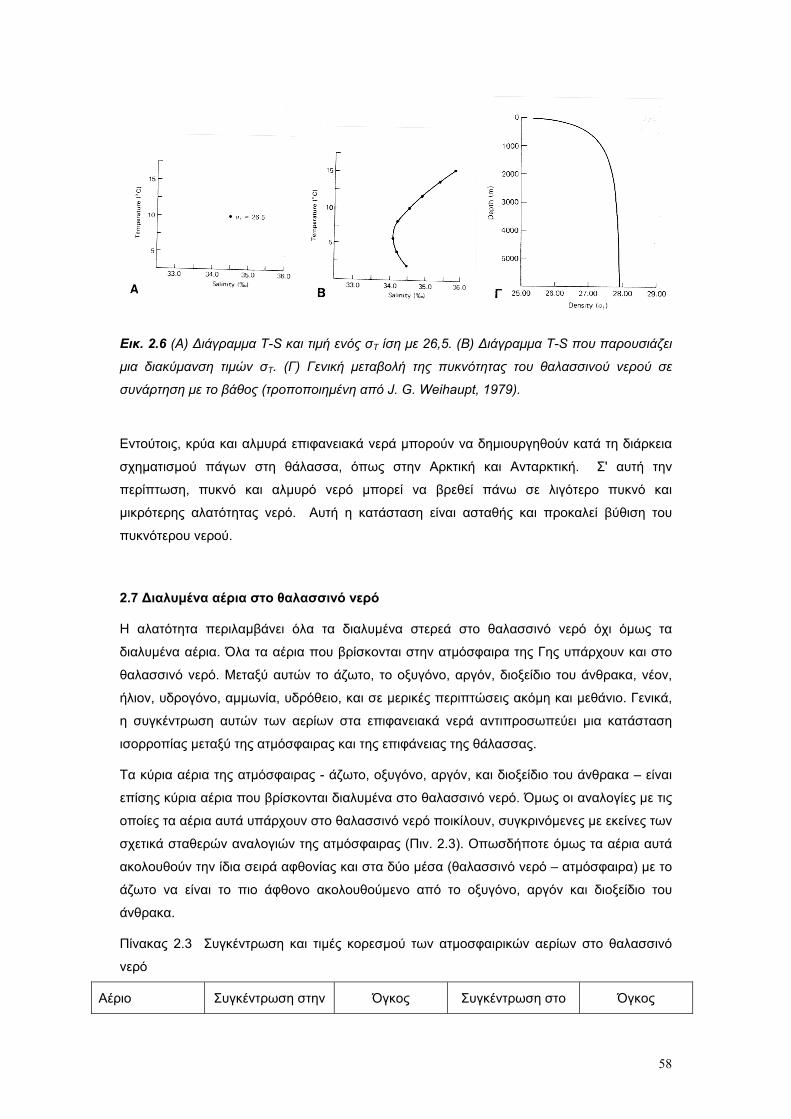
58
Εικ. 2.6 (Α) ∆ιάγραµµα Τ-S και τιµή ενός σΤ ίση µε 26,5. (Β) ∆ιάγραµµα Τ-S που παρουσιάζει
µια διακύµανση τιµών σΤ. (Γ) Γενική µεταβολή της πυκνότητας του θαλασσινού νερού σε
συνάρτηση µε το βάθος (τροποποιηµένη από J. G. Weihaupt, 1979).
Εντούτοις, κρύα και αλµυρά επιφανειακά νερά µπορούν να δηµιουργηθούν κατά τη διάρκεια
σχηµατισµού πάγων στη θάλασσα, όπως στην Αρκτική και Ανταρκτική. Σ' αυτή την
περίπτωση, πυκνό και αλµυρό νερό µπορεί να βρεθεί πάνω σε λιγότερο πυκνό και
µικρότερης αλατότητας νερό. Αυτή η κατάσταση είναι ασταθής και προκαλεί βύθιση του
πυκνότερου νερού.
2.7 ∆ιαλυµένα αέρια στο θαλασσινό νερό
Η αλατότητα περιλαµβάνει όλα τα διαλυµένα στερεά στο θαλασσινό νερό όχι όµως τα
διαλυµένα αέρια. Όλα τα αέρια που βρίσκονται στην ατµόσφαιρα της Γης υπάρχουν και στο
θαλασσινό νερό. Μεταξύ αυτών το άζωτο, το οξυγόνο, αργόν, διοξείδιο του άνθρακα, νέον,
ήλιον, υδρογόνο, αµµωνία, υδρόθειο, και σε µερικές περιπτώσεις ακόµη και µεθάνιο. Γενικά,
η συγκέντρωση αυτών των αερίων στα επιφανειακά νερά αντιπροσωπεύει µια κατάσταση
ισορροπίας µεταξύ της ατµόσφαιρας και της επιφάνειας της θάλασσας.
Τα κύρια αέρια της ατµόσφαιρας - άζωτο, οξυγόνο, αργόν, και διοξείδιο του άνθρακα – είναι
επίσης κύρια αέρια που βρίσκονται διαλυµένα στο θαλασσινό νερό. Όµως οι αναλογίες µε τις
οποίες τα αέρια αυτά υπάρχουν στο θαλασσινό νερό ποικίλουν, συγκρινόµενες µε εκείνες των
σχετικά σταθερών αναλογιών της ατµόσφαιρας (Πιν. 2.3). Οπωσδήποτε όµως τα αέρια αυτά
ακολουθούν την ίδια σειρά αφθονίας και στα δύο µέσα (θαλασσινό νερό – ατµόσφαιρα) µε το
άζωτο να είναι το πιο άφθονο ακολουθούµενο από το οξυγόνο, αργόν και διοξείδιο του
άνθρακα.
Πίνακας 2.3 Συγκέντρωση και τιµές κορεσµού των ατµοσφαιρικών αερίων στο θαλασσινό
νερό
Αέριο Συγκέντρωση στην Όγκος Συγκέντρωση στο Όγκος

59
ατµόσφαιρα
(ml / l αέρα)
(%) βαθύ θαλασσινό
νερό
(ml / l αέρα)
(%)
Nitrogen 780,9 78,08 13 63,6
Oxygen 209,5 20,94 2 – 8 33,4
Argon 9,32 0,93 0,32 1,6
Carbon dioxide 0,3 0,03 50 1,4
Neon 182 x 10-4 1,8 x 10-4
Hellium 52 x 10-4 5,0 x 10-4
Krypton 10 x 10-4 6,0 x 10-4
Xenon 8 x 10-5 7,0 x 10-4
H κατανοµή των κύριων ατµοσφαιρικών αερίων στο θαλασσινό νερό, ελέγχεται από έναν
αριθµό παραγόντων όπως η θερµοκρασία, η αλατότητα, η ατµοσφαιρική πίεση, η
υδροστατική πίεση, η διαφορά θερµοκρασίας θαλάσσης – αέρα, κατάσταση της επιφάνειας
της θάλασσας, ανάµειξη, βιολογική δραστηριότητα, ηλιακή ακτινοβολία, ραδιενεργός
αποσύνθεση και ηφαιστειακή δραστηριότητα.
Η δέσµευση ή διάλυση αυτών είναι αποτέλεσµα µιας διαδικασίας που πραγµατοποιείται στα
επιφανειακά στρώµατα των θαλασσών. Γενικά η διαλυτότητα των ατµοσφαιρικών αερίων στο
νερό ποτέ δεν είναι µεγάλη. Άζωτο, διοξείδιο του άνθρακα και οξυγόνο είναι τα περισσότερο
άφθονα αέρια διαλυµένα στο νερό. Το άζωτο στη µοριακή του µορφή είναι συγκριτικά
αδρανές και κατά συνέπεια δεν συµµετέχει στις βασικές διαδικασίες της ζωής των
περισσότερων οργανισµών. Σε αντίθεση, το διοξείδιο του άνθρακα και το νερό
χρησιµοποιούνται από τα πράσινα φυτά στη φωτοσύνθεση για να παράγουν οξυγόνο και
υψηλής ενέργειας οργανικά συστατικά, όπως π.χ. γλυκόζη. Η αναπνοή από φυτά και ζώα
διατηρεί τη φωτοσυνθετική διαδικασία για να ελευθερώνει την χρησιµοποιούµενη ενέργεια
που είναι ενσωµατωµένη στα οργανικά συστατικά της τροφής των οργανισµών. Το τρίτο
κατά σειρά αφθονίας αέριο, το οξυγόνο, χρησιµοποιείται στην αναπνοή και αποβάλλεται ως
διοξείδιο του άνθρακα, που είναι και το αφθονότερο διαλυµένο αέριο στο θαλασσινό νερό.
2.7.1 Οξυγόνο
Το οξυγόνο στη µοριακή του µορφή Ο2 είναι απαραίτητο για την επιβίωση των περισσοτέρων
οργανισµών. Μόνη εξαίρεση αποτελούν µερικά είδη αναερόβιων οργανισµών. Η αφθονία ή
έλλειψη Ο2 στο θαλασσινό νερό επηρεάζει έντονα την κατανοµή της θαλάσσιας ζωής.
Οξυγόνο χρησιµοποιείται από τους οργανισµούς σε όλες τις περιοχές του θαλάσσιου
περιβάλλοντος συµπεριλαµβανοµένων των πολύ βαθιών τάφρων. Εντούτοις, η µεταφορά
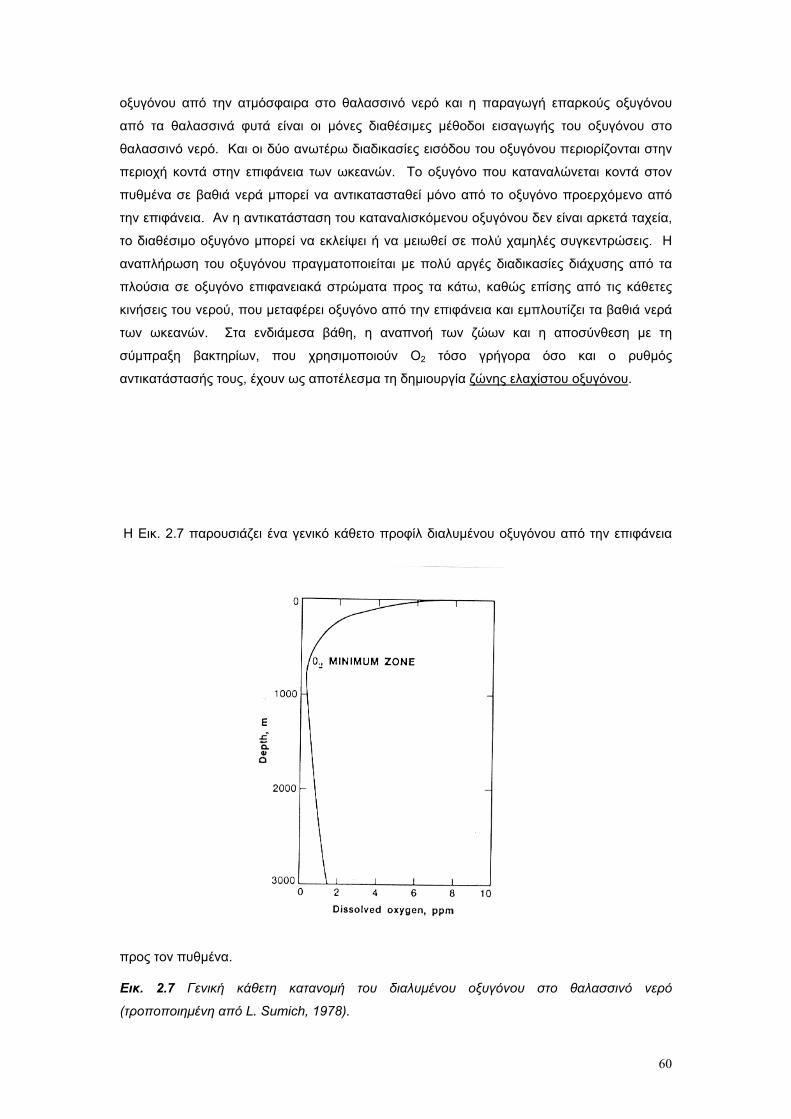
60
οξυγόνου από την ατµόσφαιρα στο θαλασσινό νερό και η παραγωγή επαρκούς οξυγόνου
από τα θαλασσινά φυτά είναι οι µόνες διαθέσιµες µέθοδοι εισαγωγής του οξυγόνου στο
θαλασσινό νερό. Και οι δύο ανωτέρω διαδικασίες εισόδου του οξυγόνου περιορίζονται στην
περιοχή κοντά στην επιφάνεια των ωκεανών. Το οξυγόνο που καταναλώνεται κοντά στον
πυθµένα σε βαθιά νερά µπορεί να αντικατασταθεί µόνο από το οξυγόνο προερχόµενο από
την επιφάνεια. Αν η αντικατάσταση του καταναλισκόµενου οξυγόνου δεν είναι αρκετά ταχεία,
το διαθέσιµο οξυγόνο µπορεί να εκλείψει ή να µειωθεί σε πολύ χαµηλές συγκεντρώσεις. Η
αναπλήρωση του οξυγόνου πραγµατοποιείται µε πολύ αργές διαδικασίες διάχυσης από τα
πλούσια σε οξυγόνο επιφανειακά στρώµατα προς τα κάτω, καθώς επίσης από τις κάθετες
κινήσεις του νερού, που µεταφέρει οξυγόνο από την επιφάνεια και εµπλουτίζει τα βαθιά νερά
των ωκεανών. Στα ενδιάµεσα βάθη, η αναπνοή των ζώων και η αποσύνθεση µε τη
σύµπραξη βακτηρίων, που χρησιµοποιούν Ο2 τόσο γρήγορα όσο και ο ρυθµός
αντικατάστασής τους, έχουν ως αποτέλεσµα τη δηµιουργία ζώνης ελαχίστου οξυγόνου.
Η Εικ. 2.7 παρουσιάζει ένα γενικό κάθετο προφίλ διαλυµένου οξυγόνου από την επιφάνεια
προς τον πυθµένα.
Εικ. 2.7 Γενική κάθετη κατανοµή του διαλυµένου οξυγόνου στο θαλασσινό νερό
(τροποποιηµένη από L. Sumich, 1978).

61
Η κατανοµή του οξυγόνου στους ωκεανούς ελέγχεται δια µέσου: (α) της ανταλλαγής µε την
ατµόσφαιρα και (β) των βιολογικών διαδικασιών της φυτοσύνθεσης και αναπνοής.
Όπως προαναφέρθηκε, το οξυγόνο της ατµόσφαιρας διαλύεται στην επιφάνεια των ωκεανών.
Το ποσό το οποίο µπορεί να διαλυθεί ελαττώνεται πολύ µε την αύξηση της θερµοκρασίας και
σε µικρότερο βαθµό µε την αύξηση της αλατότητας. Μάζες νερού, οι οποίες συγκεντρώνονται
στον πυθµένα της Ανταρκτικής, σχηµατίζονται στην επιφάνεια, βυθίζονται και µεταφέρουν
µαζί τους µια συγκέντρωση οξυγόνου που αποκτήθηκε δια µέσου της αλληλοεπίδρασης
ατµόσφαιρας και ωκεανού. Οι αναπνέοντες οργανισµοί όµως καταναλώνουν οξυγόνο καθώς
το νερό βυθίζεται και κινείται. Έτσι, αν και το µεγαλύτερο τµήµα των βαθιών θαλάσσιων
µαζών είναι οξυγονωµένο εξαιτίας της επιφανειακής προέλευσής τους, µάζες νερού, όπως
π.χ. συµβαίνει στα βαθιά νερά του Ειρηνικού, µπορεί να έχουν χαµηλή συγκέντρωση
οξυγόνου µετά από ταξίδι µερικών εκατοντάδων ετών κατά µήκος του πυθµένα.
Φωτοσύνθεση και αναπνοή των άφθονων πλαγκτικών οργανισµών στα επιφανειακά νερά
µπορούν να µεταβάλλουν σηµαντικά τη συγκέντρωση του οξυγόνου σε µικρές σχετικά
χρονικές περιόδους. Φωτοσυνθετικό πλαγκτόν σε αβαθή νερά µπορεί να κάνει
υπερκορεσµένο το νερό σε οξυγόνο. Η διαδικασία της αναπνοής αντιστρέφει την κατεύθυνση
της µεταβολής του οξυγόνου.
Κατά τη διάρκεια φυτοπλαγκτικών εξάρσεων (phytoplakton blooms) οργανικό υλικό
κατακερµατισµένο, υπό µορφή αποσυντεθηµένων πτωµάτων πλαγκτικών οργανισµών,
βυθίζεται και συγκεντρώνεται επί της ζώνης µεταβολής της πυκνότητας που δηµιουργείται
από το θερµοκλινές. Τα βακτήρια αποσυνθέτουν τα κατάλοιπα και κατά τη διαδικασία αυτή
καταναλώνουν οξυγόνο µε συνέπεια τη δηµιουργία ζωνών ελάχιστου οξυγόνου σε βάθη 100 -
200 µέτρων.
Σε µερικές θαλάσσιες λεκάνες η παροχή οργανικών καταλοίπων και ο ρόλος τους στην
εξάντληση του οξυγόνου προχωρεί γρηγορότερα από την ανάµιξη των υδάτων του πυθµένα
της λεκάνης µε άλλα οξυγονωµένα νερά. Έτσι τα νερά του πυθµένα της Μαύρης Θάλασσας,
το βαθύ Cariaco της Καραϊβικής και µερικά Νορβηγικά φιόρντ έχουν παντελή έλλειψη
οξυγόνου. Σε άλλες περιπτώσεις η ισορροπία κλίνει υπέρ της εποχιακής εξάντλησης, όπως
κατά τη διάρκεια του θέρους σε µερικές βαθιές περιοχές της Βαλτικής και ανά µερικά χρόνια
σε πολλές περιορισµένες λεκάνες. Σε µικρές µάζες νερού, όπως στις λίµνες που
σχηµατίζονται προσωρινά εξαιτίας των παλιρροιών (tidal pools) αυτή η ισορροπία µπορεί να
µεταβάλλεται δραµατικά κατά την διάρκεια της ηµέρας.
Οξυγόνο και ζωή
Όλοι σχεδόν οι ευκαρυωτικοί οργανισµοί έχουν ανάγκη οξυγόνου για το µεταβολισµό. Η
συνεχιζόµενη απουσία ή εξάντληση διαλυµένου οξυγόνου έχει ως αποτέλεσµα τη µείωση της
µεταβολικής δραστηριότητας. Η κατανάλωση οξυγόνου εκφράζεται συνήθως σε ml που
καταναλώνονται ανά µονάδα βάρους σώµατος στη µονάδα του χρόνου (ml * g-1 * hr-1). Παρά

62
το γεγονός ότι µεγάλα άτοµα καταναλώνουν περισσότερο οξυγόνο από όσο µικροί
αντιπρόσωποι του ίδιου είδους, η κατανάλωση οξυγόνου κατά µονάδα βάρους σώµατος στη
µονάδα του χρόνου είναι αντιστρόφως ανάλογη προς το µέγεθος του σώµατος. Έτσι, µε
βάση τη µονάδα βάρους του σώµατος, τα πρωτόζωα καταναλώνουν περισσότερο οξυγόνο
από τα καβούρια. Η συσχέτιση του βαθµού κατανάλωσης οξυγόνου προς το µέγεθος του
σώµατος µπορεί να εκφρασθεί ως εξής:
ml O2 = kWb
όπου b = εκθέτης υπολογιζόµενος από γραφική παράσταση, W = βάρος του σώµατος και k =
σταθερά. Τα περισσότερα θαλάσσια ποικιλόθερµα έχουν τιµές b µικρότερες από 1,0 (0,66 -
1,0).
Ο βαθµός µεταβολισµού δεν αυξάνει γραµµικά µε την αύξηση του βάρους. Υπάρχουν
αρκετοί πιθανοί λόγοι για το γεγονός ότι ο µεταβολικός ρυθµός δεν αυξάνεται ανάλογα µε το
βάρος του σώµατος. Στα Πρωτόζωα η µείωση της αναλογίας της επιφάνειας του σώµατος
προς τον όγκο, µε την αύξηση του µεγέθους του σώµατος είναι πιθανώς ένας περιοριστικός
µηχανισµός. Σε άλλους οργανισµούς µε αναπνευστικό και κυκλοφορικό σύστηµα µπορεί να
είναι σηµαντικοί άλλοι µηχανισµοί. Π.χ. αύξηση της αναλογίας της µάζας των συστηµάτων
που δε σχετίζονται µε την αναπνοή (σκελετός, λίπος και συνδετικός ιστός), µεταβολές της
ενεργητικότητας και των προτύπων ανάπτυξης καθώς και περιορισµός των αναπνευστικών
οργάνων, όπως είναι τα βράγχια, µπορεί να είναι µερικοί σηµαντικοί παράγοντες.
Σπόγγοι, ασκίδια και τα περισσότερα δίθυρα µαλάκια καταναλώνουν πολύ πιο λίγο οξυγόνο
από τα δεκάποδα, καρκινοειδή, τα κεφαλόποδα και τους τελεόστεους. Η ενεργητικότητα
µπορεί ν' αλλάξει κατά τη διάρκεια της ζωής του ατόµου. Έτσι, είδη που τρέφονται δραστικά
κατά την διάρκεια της ηµέρας, έχουν κατά συνέπεια ανάγκη περισσότερου οξυγόνου για τη
χρονική αυτή διάρκεια.
Οι µηχανισµοί της λήψης οξυγόνου ποικίλουν µε το µέγεθος του σώµατος, φυλετική
καταγωγή, χώρο διαβίωσης, ηλικία και ενεργητικότητα. Πολλοί µικροί οργανισµοί, όπως
πρωτόζωα, νηµατώδεις, έµβρυα και προνύµφες, βασίζουν την αναπνοή τους πάνω στη
διάχυση του οξυγόνου σε όλη την επιφάνεια του σώµατος. Οι µεγαλύτεροι πολύχαιτοι,
µαλάκια και τα περισσότερα καρκινοειδή χρησιµοποιούν τα βράγχια και τα κυκλοφορικά
συστήµατα για τις αναπνευστικές ανταλλαγές. Τα βράγχια έχουν µεγάλη αναπνευστική
επιφάνεια και οι οργανισµοί που φέρουν βράγχια είτε µετακινούνται γρήγορα είτε υπόκεινται
σε ρεύµατα νερού που διέρχονται πάνω απ' αυτά για ανταλλαγή οξυγόνου. Είδη που ζουν
φωλιασµένα στην άµµο, όπως οι πολύχαιτοι, δηµιουργούν πάντοτε ένα ρεύµα νερού για να
καταιονίζουν τη φωλιά τους µε οξυγονωµένο νερό. Τα ψάρια χρησιµοποιούν τα βράγχια
αναρτηµένα επί βραγχιακών τόξων. Σε άλλες περιπτώσεις όµως τα βράγχια παίζουν και
άλλους ρόλους, εκτός της αναπνοής, όπως τον ρόλο πρόσληψης τροφής από ορισµένους
οργανισµούς (π.χ. τα µαλάκια), της ανταλλαγής ιόντων (π.χ. τα καρκινοειδή, ψάρια).

63
Η διαθεσιµότητα οξυγόνου µπορεί περιοδικά να µειώνεται και να επηρεάζει το µεταβολισµό
στα θαλάσσια ασπόνδυλα. Στη ζώνη χαµηλής παλιρροιακής στάθµης, τα ζώα που έχουν
ανάγκη να βυθίζονται κάτω από το νερό για να λαµβάνουν οξυγόνο, υπόκεινται σε
παρατεταµένη περίοδο έλλειψης οξυγόνου.
Σε υψηλά γεωγραφικά πλάτη, κατά τη διάρκεια του χειµώνα, η αναπνοή πιέζεται προς
χαµηλούς ρυθµούς και η χαµηλή θερµοκρασία προκαλεί µειωµένους ρυθµούς µεταφοράς
οξυγόνου στα κύτταρα. Κατά τη διάρκεια αυτών των περιόδων πολλά θαλάσσια ασπόνδυλα
χρησιµοποιούν αναερόβια διαδικασία στο µεταβολισµό. Κατά τις περιόδους χαµηλής
παλίρροιας τα τελικά προϊόντα του αναερόβιου µεταβολισµού (αλανίνη και κρυσταλλικό
δικαρβοξυλικό οξύ) οικοδοµούνται στους ιστούς. Στα µαλάκια, ένα µέρος του δικαρβοξυλικού
οξέος εξουδετερώνεται από διάλυµα CaCΟ3.
Τα ζώα που έχουν ανάγκη µεγάλων ποσοτήτων οξυγόνου ή ζουν σε περιβάλλον όπου το
οξυγόνο είναι µειωµένο, µπορεί να έχουν αιµατικά χρωµατοφόρα και πολύ αυξηµένες
ικανότητες µεταφοράς οξυγόνου.
Η δέσµευση και ελευθέρωση οξυγόνου από µία χρωστική, όπως η αιµοσφαιρίνη (Hb), µπορεί
να περιγραφεί µε την παρακάτω εξίσωση:
Hb + O2 <========> HbO2
Αυξανόµενης της µερικής πίεσης του οξυγόνου, συνεχώς περισσότερο οξυγόνο δεσµεύεται
µε την αιµοσφαιρίνη µέχρι κορεσµού. Σε µια δεδοµένη πίεση οξυγόνου, η ποσότητα του
οξυγόνου που δεσµεύεται µε την αιµοσφαιρίνη είναι καθορισµένη. Κατά συνέπεια µπορεί να
κατασκευασθεί η καµπύλη δέσµευσης οξυγόνου (Εικ. 2.8), η οποία εµφανίζει το ποσοστό
κορεσµού ως συνάρτηση της πίεσης του οξυγόνου. Το pH του αίµατος και των κοιλωµατικών
υγρών επηρεάζει επίσης τα χαρακτηριστικά της αιµοσφαιρίνης στη δέσµευση του οξυγόνου.
Γενικά πτώση του pH τείνει να µετατοπίσει την καµπύλη οξυγόνου προς τα δεξιά (the Bohr
effect). Έτσι: (α) όπου το CO2 είναι άφθονο, γεγονός που οδηγεί στη µείωση του pH και (β)
όπου το Ο2 είναι σε χαµηλή συγκέντρωση, η οξυγονωµένη αιµοσφαιρίνη τείνει να
ελευθερώσει οξυγόνο. Αυτός ο παράγοντας είναι σηµαντικός από άποψη προσαρµογής
διότι, καθώς το οξυγονωµένο αίµα φθάνει στους ιστούς οι οποίοι είναι φτωχοί σε Ο2 και
πλούσιοι σε CO2, το οξυγόνο ελευθερώνεται και γίνεται διαθέσιµο στους ιστούς. Είναι
προφανές ότι δραστήρια ζώα, που έχουν κατά συνέπεια ανάγκη γρήγορης παροχής
οξυγόνου, η καµπύλη οξυγόνου µετατίθεται προς τα δεξιά ενώ για τα λιγότερο δραστήρια ζώα
η καµπύλη µετατίθεται προς τα αριστερά (Εικ. 2.8). Αυτό φαίνεται να ισχύει και για τα
ασπόνδυλα που περιέχουν την χρωστική αιµοκυανίνη. Τα κεφαλόποδα έχουν υψηλή στάθµη
αιµοκυανίνης και βασίζονται σ' αυτήν για την µεταφορά του οξυγόνου.

64
Εικ. 2.8 Καµπύλη δέσµευσης – αποδέσµευσης του οξυγόνου µε την ατµόσφαιρα
Οι χρωστικές του αίµατος µπορεί να εξυπηρετούν ως δεξαµενές αποθήκευσης οξυγόνου για
ζώα που σκάβουν και ζουν µέσα σε ιζήµατα µε λίγο οξυγόνο. ∆ίθυρα, που ζουν σε τροπικά
µαλακά ιζήµατα µε λίγο ή καθόλου οξυγόνο, τείνουν να έχουν χρωστικές αίµατος. Σε
αντίθεση, οι χρωστικές ελλείπουν από τα είδη τα οποία ζουν σε περισσότερο οξυγονωµένα
περιβάλλοντα.
Ζώα της υψηλής µεσοπαλιρροικής ζώνης περνούν περισσότερο χρόνο εκτεθειµένα στον αέρα
παρά βυθισµένα στο νερό. Αρκετά είδη διθύρων ανοίγουν τις θυρίδες τους όταν εκτίθενται
στον αέρα. Το είδος Mytilus californianus καταναλώνει οξυγόνο στον αέρα σε συγκρίσιµες
αναλογίες µε την αναπνοή τους στο νερό.
2.7.2 ∆ιοξείδιο του άνθρακα
Το διοξείδιο του άνθρακα (CO2) είναι άφθονο στις περισσότερες περιοχές της θάλασσας και
γενικά δεν παρατηρούνται συγκεντρώσεις τόσο µικρές ώστε να εµποδίζουν την ανάπτυξη των
φυτών.
Το θαλασσινό νερό έχει µια ασυνήθιστα µεγάλη ικανότητα να απορροφά CO2 διότι το
περισσότερο CO2 δεν παραµένει ως αέριο. Μάλλον πολύ απ' αυτό συνδυάζεται µε το νερό
για να παράγει ένα αδύνατο οξύ, το ανθρακικό οξύ (H2CO3). Γενικά το ανθρακικό οξύ
διασπάται για να σχηµατίσει ένα ιόν υδρογόνου (Η+) και ένα ιόν όξινου ανθρακικού οξέος
(ΗCO3-) ή δυο ιόντα Η+ και ένα ιόν ανθρακικού οξέος (CO3
-2) Αυτές οι αντιδράσεις µπορούν
να γραφούν περιληπτικά ως κατωτέρω:
1. CO2 + H2O <=====> H2CO3
2.H2CO3 <=====> H+ + HCO3-
3. HCO3- <=====> H+ + CO3
-2
Η αφθονία των ιόντων Η+ στα υδατικά διαλύµατα ελέγχει την οξύτητα ή αλκαλικότητα κάθε
υδατικού διαλύµατος και µετρείται σε µια κλίµακα από 1 µέχρι 14 µονάδες pH. Ειδικότερα η

65
οξύτητα ή το pH του νερού καθορίζεται ως ο αρνητικός log10 της δραστικότητας των ιόντων
Η+. Γι' αυτό, επειδή η δραστικότητα των ιόντων Η+ του απεσταγµένου νερού είναι 10-7 , το pH
του είναι 7. Οι µονάδες pH είναι µια µέτρηση της συγκέντρωσης ιόντων Η+. Το 1 της
κλίµακας pH δηλώνει µεγάλη οξύτητα και αντιπροσωπεύει µια µεγάλη συγκέντρωση ιόντων
Η+. Το 14 δηλώνει ότι το διάλυµα είναι πολύ βασικό ή αλκαλικό και αντιπροσωπεύει µια πολύ
µικρή συγκέντρωση ιόντων Η+. Ουδέτερο pH (το pH του καθαρού νερού) είναι το 7 της
κλίµακας.
Το σύστηµα του ανθρακικού οξέος (ιόντα όξινου ανθρακικού και ανθρακικού οξέος
αντίστοιχα) στο θαλασσινό νερό ενεργεί στην εξουδετέρωση της περίσσειας οξέων ή βάσεων
(buffer) ή και περιορίζει µεταβολές του pH στο θαλασσινό νερό. Αν υπάρχει περίσσεια ιόντων
Η+ οι ανωτέρω αντιδράσεις δρουν προς τα αριστερά γεγονός που προκαλεί την
αποµάκρυνση των ιόντων αυτών από το διάλυµα. Αν είναι παρόντα πολύ λίγα ιόντα Η+ τότε
τα ιόντα αυτά γίνονται περισσότερα από την µετατροπή του H2CO3 σε HCO3- και του HCO3-
σε CO3-2. Σε συνθήκες ανοικτής θάλασσας αυτό το σύστηµα εξουδετέρωσης είναι πολύ
αποτελεσµατικό µε αποτέλεσµα να περιορίζονται οι τιµές του pH µεταξύ 7,5 και 8,4. Αν
διατηρείται µια κατάσταση ισορροπίας µεταξύ διαλύµατος και κατακάθισης του κρυσταλλικού
ανθρακικού οξέος, των ενδιάµεσων ιόντων και της πίεσης του CO2 στην ατµόσφαιρα, τότε το
pH πρέπει να είναι γύρω στην τιµή 8. Σε περιπτώσεις όπου διαλυµένο CO2 παρέχεται σε
µεγάλες ποσότητες, όπως στην ζώνη του ελάχιστου O2, το pH µπορεί να ελαττωθεί µέχρι 7,5.
Αντίθετα σε υψηλούς βαθµούς φωτοσύνθεσης, σε ρηχά νερά, καταναλίσκεται πολύ CO2 µε
συνέπεια το pH να φθάνει µέχρι την τιµή 9.
2.7.3 Άζωτο
Το άζωτο είναι ένα σχετικά αδρανές αέριο του οποίου η συγκέντρωση στο θαλασσινό νερό
κυµαίνεται από 8 µέχρι 15 ml/l. Εκτός όµως από το διαλυµένο µοριακό άζωτο, το θαλασσινό
νερό περιέχει µικρή αλλά εξαιρετικά µεγάλης σηµασίας συγκέντρωση ανόργανων και
οργανικών αζωτούχων ενώσεων, το συνολικό βάρος των οποίων είναι περίπου το 1/10 του
βάρους του αερίου µοριακού αζώτου.
Το άζωτο που απαιτείται για την σύνθεση των πρωτεϊνών υπάρχει διαλυµένο σε τρεις κύριες
ανόργανες µορφές: αµµωνία, νιτρώδη και νιτρικά. Το άζωτο επίσης βρίσκεται διαλυµένο σε
οργανικές µορφές, όπως ουρία, αµινοξέα και πεπτίδια.
Η αµµωνία είναι συνήθως η περισσότερο διαδεδοµένη µορφή του αζώτου, επειδή έτσι µπορεί
να χρησιµοποιηθεί απευθείας για τη σύνθεση αµινοξέων χωρίς καµία αλλαγή στην κατάσταση
οξείδωσής του. Πλέον αυτού η αµµωνία µπορεί να δεσµεύεται περισσότερο αποτελεσµατικά,
σε χαµηλά επίπεδα φωτισµού, από όσο τα νιτρικά.
Οι υψηλότερες συγκεντρώσεις διαλυµένου αζώτου που παρατηρούνται στους ωκεανούς είναι
τα νιτρικά τα οποία είναι συχνά η περισσότερο άφθονη µορφή του αζώτου στα εύστροφα
παράκτια ύδατα. Αναβλύσεις υδάτων (upwelling) και αναταράξεις προερχόµενες από
στροβίλους φέρουν νιτρικά στην εύφωτη ζώνη. Τα νιτρώδη είναι γενικά τα σπανιότερα από

66
τις τρεις µορφές του αζώτου και συµπεριφέρονται µε τον ίδιο τρόπο όπως τα νιτρικά στην
ανακύκλωση των θρεπτικών στο φυτοπλαγκτόν. Κάτω από ορισµένες περιστάσεις η
αµµωνία µπορεί να ξεπεράσει σε αφθονία τα νιτρικά (συνήθως όταν χρησιµοποιούνται
νιτρικά). Στον κόλπο του Main η αµµωνία βρέθηκε να είναι 25 µέχρι 100 φορές αφθονότερη
από τα νιτρικά. Οι διαλυµένες συγκεντρώσεις και των τριών µορφών του αζώτου αυξάνονται
κατά την περίοδο των ευκράτων χειµώνων του Βορρά και µειώνονται κατά την άνοιξη και
καλοκαίρι όταν αναπτύσσονται οι πληθυσµοί του φυτοπλαγκτού.
Σε αβαθείς παράκτιους κόλπους και εκβολές ποταµών η σύζευξη µε το βενθικό σύστηµα
µπορεί να επηρεάσει τη δυναµική των θρεπτικών του φυτοπλαγκτού. Έχει αποδειχθεί ότι
άζωτο σε µορφή αµµωνίας µεταφέρεται εκτεταµένα από τον πυθµένα προς τα υπερκείµενα
ύδατα. Αµµωνία ελευθερώνεται κατά τη διάρκεια της διαδικασίας µικροβιακής αποσύνθεσης
καταλοίπων στον πυθµένα. Το µισό από το άζωτο που επιστρέφει από τον πυθµένα στη
στήλη του νερού είναι πιθανώς σε µορφή οργανικού αζώτου όπως η ουρία. Μερικά είδη
φυτοπλαγκτού είναι ικανά να προσλαµβάνουν ουρία, ουρικό οξύ και αµινοξέα.
Η ανακύκληση των διαφόρων µορφών αζώτου εξαρτάται από τον τόπο και την φύση του
κύκλου αναγέννησης του θρεπτικού. Σε παράκτιες περιοχές µε υψηλή ανάβλυση υδάτων,
όπως ανοικτά των ακτών του Περού, νιτρικά αναγεννώνται από τον πυθµένα και είναι η κύρια
οδός δια µέσου της οποίας το άζωτο εισέρχεται στο σύστηµα του πλαγκτού. Έρευνες µε Ν-
15 απέδειξαν ότι περισσότερο από το µισό άζωτο, που αναδύεται στις περιοχές αναβλύσεων,
είναι στη µορφή του νιτρικού. Το υπόλοιπο είναι στη µορφή αµµωνίας, η οποία
ανακυκλώνεται από τα περιττώµατα του ζωοπλαγκτού και την αποσύνθεσή τους πίσω προς
το φυτοπλαγκτόν. Αντίθετα, στις φτωχές σε θρεπτικά περιοχές, λιγότερο από 10% της
µετρούµενης πρόσληψης αζώτου είναι στην µορφή των νιτρικών. Κατά συνέπεια η
µεγαλύτερη πρόσληψη αζώτου πρέπει να περιλαµβάνει αποτελεσµατική ανακύκληση
αµµωνίας και οργανικού αζώτου µεταξύ ζωοπλαγκτού και φυτοπλαγκτού. Το άζωτο µπορεί
να ενσωµατώνεται στις θαλάσσιες τροφικές αλυσίδες δια µέσου της διαδικασίας δέσµευσής
του, που πραγµατοποιείται από µερικά βακτήρια, µπλε-πράσινα φύκη και µύκητες. Πολλά
διάτοµα, τα οποία ζουν σε πτωχά νερά από άποψη θρεπτικών, σχηµατίζουν συµβιωτικές
κοινωνίες µε ένα µπλε-πράσινο φύκος που φαίνεται ότι δεσµεύει άζωτο. Αλλά και το
αντίθετο, δηλ. απονιτρωποιητικά βακτήρια συντελούν στην επιστροφή µεγάλων ποσοτήτων
αζώτου στην ατµόσφαιρα.
Κύκλος του αζώτου (Εικ. 2.9)
Το ζωοπλαγκτόν τρέφεται µε οργανικό και ανόργανο άζωτο που βρίσκεται στο φυτοπλαγκτόν
και παράλληλα αποβάλλει άζωτο σε διαλυτή µορφή, όπως αµµώνιο ή οργανικό άζωτο.
Φυτοπλαγκτόν και µικροοργανισµοί προσλαµβάνουν και αποβάλλουν διαλυτό ανόργανο και
οργανικό άζωτο. Υπάρχει επίσης ανταλλαγή µε το ίζηµα δια µέσου της διαδικασίας
διατροφής και αποβολής περιττωµάτων των βενθικών µικροοργανισµών και των
ασπόνδυλων. Το φυτοπλαγκτόν µπορεί να προσλαµβάνει NH4 και νιτρώδη ιόντα καθώς

67
επίσης και νιτρικά, αλλά µπορεί επίσης να αποβάλλει νιτρώδη κατά τη διάρκεια της
πρόσληψης νιτρικών.
Εικ. 2.9 Κύκλος του αζώτου στη θάλασσα (τροποποιηµένη από P.S. Meadous and J. J.
Campbell, 1987).
Τα µπλε-πράσινα φύκη και µερικά βακτήρια σχηµατίζουν µοριακό άζωτο, ειδικότερα στις
τροπικές περιοχές. Στις εύκρατες περιοχές τα νιτρικά µπορεί να εξαντλούνται πλήρως, ενώ
τα φωσφορικά είναι ακόµη παρόντα σε χαµηλά επίπεδα. Τα βακτήρια παίζουν βασικό ρόλο
στον κύκλο του αζώτου. Μπορούν να σχηµατίζουν µοριακό άζωτο και συγχρόνως να
µετατρέπουν διαλυτό οργανικό άζωτο, το οποίο έχει παραχθεί από ζώντας φυτο- και ζωο-
πλαγκτικούς οργανισµούς σε αµµωνία. Οξειδώνουν το NH4 σε νιτρώδη και τα νιτρώδη σε
νιτρικά (nitrification). Τα βακτήρια αυτά βρίσκονται κυρίως µέσα και ακριβώς επάνω στα
ιζήµατα του πυθµένα και στο πλαγκτόν.
Το µέγιστο των νιτρωδών, που µερικές φορές αναπτύσσεται επί του πυθµένος στη
φωτοσυνθετική ζώνη, µπορεί να οφείλεται µερικώς στην οξείδωση του NH4 σε νιτρώδη από
βακτήρια και µερικώς στην αναγωγή των νιτρικών σε νιτρώδη από το φυτοπλαγκτόν. Τα
βακτήρια µπορούν να ανάγουν τα νιτρικά σε νιτρώδη και τα νιτρώδη σε µοριακό άζωτο
(denitrification).
Το τελευταίο δεν είναι πιθανώς µεγάλης σπουδαιότητας, αν και είναι κοινά στο θαλασσινό
νερό και στα ιζήµατα τα βακτήρια τα οποία οξειδώνουν τα νιτρικά σε νιτρώδη.
2.8 Θρεπτικά
Θρεπτικά είναι όλα εκείνα τα συστατικά που χρειάζονται τα φυτά. Θα µπορούσαµε να τα
ονοµάσουµε θρεπτικά συστατικά ή θρεπτικά στοιχεία (π.χ. άζωτο), που υπάρχουν σε

68
διαλυµένη ανόργανη µορφή (π.χ. αµµωνία, νιτρικά άλατα) ή σε οργανική µορφή (π.χ.
αµινοξέα). Μπορούν να διαιρεθούν σε κύρια θρεπτικά στοιχεία, σε ίχνη θρεπτικών στοιχείων
και σε οργανικά θρεπτικά συστατικά.
Τα κύρια θρεπτικά στοιχεία απαιτούνται σε µεγάλες ποσότητες και περιλαµβάνουν τον
άνθρακα, άζωτο, φωσφόρο, οξυγόνο, πυρίτιο, µαγνήσιο, κάλιο και ασβέστιο.
Τα ίχνη θρεπτικών στοιχείων χρειάζονται σε πολύ µικρότερες ποσότητες και περιλαµβάνουν
το σίδηρο, χαλκό και βανάδιο.
Τα οργανικά θρεπτικά συστατικά, στα οποία συµπεριλαµβάνονται και οι βιταµίνες,
σχετίζονται άµεσα µε το φαινόµενο της αυξοτροφίας. Ειδικότερα, οι ορισµοί αυτότροφος και
αυξότροφος, είναι γνωστό ότι αναφέρονται ο µεν πρώτος στην πρόσληψη µόνο ανόργανων
ο δε δεύτερος στην πρόσληψη ανόργανων και οργανικών θρεπτικών συστατικών, σε
συνδυασµό µε τη φωτοσύνθεση. Ο ορισµός ετερότροφος εξάλλου, αναφέρεται στη
διεργασία πρόσληψης οργανικών συστατικών για διατροφή όταν απουσιάζει η φωτοσύνθεση.
Πολλά συστατικά τα οποία είναι ουσιώδη για την διατροφή του φυτοπλαγκτού µπορούν να
βρίσκονται σε αδιάλυτες ή διαλυµένες µορφές. Ο άνθρακας, το άζωτο και ο φωσφόρος
µπορούν να βρίσκονται σε πτώµατα φυτοπλαγκτού και ζωοπλαγκτού βυθιζόµενα στην στήλη
του νερού. Μικροστροβιλώδη κίνηση του νερού µπορεί επίσης να δηµιουργεί τεµαχίδια
προερχόµενα από τη σύγκρουση διαλυµένων οργανικών µορίων. Το αποτέλεσµα αυτής της
συγκέντρωσης εύθραυστου οργανικού υλικού είναι γνωστό ως θαλασσινό χιόνι και συχνά
βρίσκεται εµπλουτισµένο µε ποικιλία πλαγκτικών οργανισµών και προϊόντων θρυµµατισµού
πλαγκτικών οργανισµών. Τέλος, εισροές από ποτάµια και θαλασσινά λιβάδια (seaweed
beds) συµβάλλουν επίσης στη συγκέντρωση θρυµµατισµένου οργανικού υλικού.
∆ιαλυµένα οργανικά συστατικά, θρυµµατισµένα οργανικά υλικά και ζώσα βιοµάζα υπάρχουν
κατά µέσο όρο στην αναλογία περίπου 100:10:2 Η θάλασσα είναι ένα διάλυµα θρεπτικών
ουσιών σε διαλυµένη µορφή µε τα επιφανειακά στρώµατα εξαντλούµενα συνεχώς εξαιτίας της
πρόσληψης των ουσιών αυτών από τις ζώσες µορφές της εύφωτης ζώνης. Οι διαδικασίες
ανταλλαγής, όπως αναβλύσεις (upwelling) και αναµίξεις υδάτων προκαλούµενες από
ανέµους, µπορούν να ισοσταθµίζουν την απώλεια των επιφανειακών θρεπτικών σε
µεγαλύτερα βάθη δια µέσου των βυθιζόµενων πτωµάτων και απορριµµάτων του
ζωοπλαγκτού. Ο χρόνος ανακύκλησης (turnover) του διαλυµένου στους ωκεανούς οργανικού
υλικού εκτιµάται ότι είναι 30 έως 300 χρόνια, υπάρχουν όµως και µερικά µόρια (π.χ. γλυκόζη)
που έχουν πολύ ταχύτερους ρυθµούς ανακύκλησης, ενώ άλλα βαίνουν πιθανόν πιο σιγά.
Κύρια θρεπτικά στοιχεία
Όπως προαναφέρθηκε, τα στοιχεία O2, N2, P, C, Si, Mg, K και Ca αποτελούν τα κύρια
θρεπτικά συστατικά. Το άζωτο και οξυγόνο, ως αέρια διαλυµένα στο νερό, εξετάσθηκαν
ανωτέρω. Στη συνέχεια δίνονται ορισµένες πληροφορίες για µερικά από τα σπουδαιότερα
µεταλλικά στοιχεία.

69
(α) Φωσφόρος
Ο φωσφόρος αποτελεί ένα από τα πιο σηµαντικά στοιχεία για τη ζωή. Αποτελεί, ως είναι
γνωστό, βασικό µέρος των οργανικών οξέων που βρίσκονται στον πυρήνα των ζωντανών
κυττάρων. Επίσης το στοιχείο αυτό µε την µορφή του ATP και ADP είναι υπεύθυνο για το
µεταβολισµό της ενέργειας στα κύτταρα και συναντάται σε ενώσεις, από τις οποίες
αποτελούνται τα σκελετικά τµήµατα πολλών οργανισµών. Έχει υπολογισθεί ότι κάθε χρόνο
µεταφέρονται στη θάλασσα µε τα χερσαία ρεύµατα, περίπου 15 εκατοµµύρια τόνοι
φωσφόρου.
Ο φωσφόρος υπάρχει στο θαλασσινό νερό σε τρεις µορφές: ανόργανο φωσφορικό άλας (ιόν
ορθοφωσφορικού οξέος), διαλυµένος οργανικός φωσφόρος και αδιάλυτος φωσφόρος υπό
µορφή κολλοειδούς διαλύµατος. Επίσης σε µολυσµένες περιοχές βρίσκεται σε µορφή
πολυφωσφορικών αλάτων, που µπορεί να χρησιµοποιούνται περιστασιακά από το
φυτοπλαγκτόν. Το ορθοφωσφορικό είναι η µορφή που προτιµάται από το φυτοπλαγκτόν και
ανταλλάσσεται πολύ γρήγορα µεταξύ φυτοπλαγκτού και θαλασσινού νερού.
Η απόλυτη ποσότητα του φωσφορικού άλατος στο θαλασσινό νερό περιορίζει το παραγωγικό
δυναµικό τους αν και υπάρχουν ορισµένες αντικρουόµενες απόψεις ως προς το ελάχιστο της
συγκέντρωσής του που είναι αρκετή για να περιορίσει το βαθµό αύξησης. Ο Strickland
(1965) ισχυρίζεται ότι είναι λίγες οι ενδείξεις ότι υπάρχει πράγµατι περιοριστικός βαθµός στο
θαλάσσιο περιβάλλον. Συνήθως η έλλειψη αζώτου πιστεύεται ότι είναι υπεύθυνη για το
σταµάτηµα της ανάπτυξης φυτοπλαγκτικών πληθυσµών.
Η έλλειψη φωσφόρου στο κύτταρο πρέπει να είναι παρά πολύ µεγάλη πριν επηρεασθούν οι
διαδικασίες απελευθέρωσης ενέργειας, όπως οι ενζυµατικές αντιδράσεις και οι φωτοχηµικές
δράσεις στη φωτοσύνθεση. Ο ρόλος του φωσφόρου είναι διαφορετικός από εκείνον του
αζώτου επειδή ο φωσφόρος χρησιµοποιείται κυρίως στον κύκλο ενέργειας του κυττάρου.
Ο κύκλος του φωσφόρου (Εικ. 2.10)
Εικ. 2.10 Κύκλος του φωσφόρου στη θάλασσα (τροποποιηµένη από P.S. Meadous and J.I.
Cambell, 1987)

70
Το ζωοπλαγκτόν χρησιµοποιεί ως τροφή τον οργανικό και ανόργανο φωσφόρο που
βρίσκονται στο φυτοπλαγκτό και µετά τον αποβάλλει σε διαλυτή µορφή. Φυτοπλαγκτόν και
µικροοργανισµοί προσλαµβάνουν και αποβάλλουν ευδιάλυτο ανόργανο και οργανικό
φωσφόρο. Ο οργανικός φωσφόρος υδρολύεται σε ανόργανο φωσφόρο µέσα σε αλκαλικό
θαλασσινό νερό και µε την παρουσία ενζύµων, επί της επιφάνειας µερικών φυτοπλαγκτικών
µορφών. Υπάρχει επίσης ανταλλαγή µε το ίζηµα δια µέσου της διαδικασίας λήψεως τροφής
και απέκκρισης από βενθικούς µικροοργανισµούς και ασπόνδυλα. Ο φωσφόρος µπορεί να
ανακυκλωθεί µέχρι πέντε φορές γρηγορότερα από το άζωτο.
Αναλογία Φωσφόρου - Αζώτου στη θάλασσα
Ο Harvey (1926) παρατήρησε ότι το σταµάτηµα της ανάπτυξης του φυτοπλαγκτού στο κανάλι
της Μάγχης ήταν αποτέλεσµα σύγχρονης εξάντλησης τόσο του αζώτου όσο και του
φωσφόρου. Πέραν αυτού, ο φωσφόρος και το άζωτο είναι διαθέσιµα στο θαλασσινό νερό σε
αναλογία N:P = 15:1, που είναι πολύ κοντά στην αναλογία που συνήθως απαιτείται από το
φυτοπλαγκτόν.
Η σύµπτωση της αναλογίας φωσφόρου και αζώτου στη θάλασσα και στις απαιτήσεις του
φυτοπλαγκτού µπορεί να εξηγηθεί από την υπόθεση ότι η ανάπτυξη των κυττάρων του
φυτοπλαγκτού, που ακολουθείται από βύθιση και αποσύνθεση, ελέγχει την αναλογία N:P στο
φυτοπλαγκτόν και το θαλασσινό νερό. Η ισορροπία του σχηµατισµού αζώτου και της
απονιτροποίησης ρυθµίζει πιθανώς συνολικά την συσχέτιση του αζώτου προς το φωσφόρο.
Παρά την πιθανή ορθότητα αυτής της γενικής υπόθεσης, υπάρχουν πολυάριθµα
παραδείγµατα φυτοπλαγκτικών ειδών τα οπoία αποκλίνουν από την τυπική αναλογία N:P.
(β) Ιώδιο
Το στοιχείο, που η συγκέντρωση του στο θαλασσινό νερό είναι περίπου 50 mg/m3,
συγκεντρώνεται σε µεγάλες ποσότητες από θαλάσσια φύκη, κατά ένα µέρος σε µορφή δι-
ιώδιο-τυροσίνης.
(γ) Πυρίτιο
Το πυρίτιο είναι στοιχείο που βρίσκεται στο θαλασσινό νερό σε µορφή διαλυµένη και
κολλοειδή. Η διαλυµένη µορφή του πυριτίου είναι ορθοπυριτικό οξύ. Η κολλοειδής µορφή
του αποτελείται από µικρού µεγέθους πυριτικά υλικά. Πολλά απ' αυτά προέρχονται από τη
διάβρωση πετρωµάτων της στεριάς και µεταφέρονται στη θάλασσα µε τα ποτάµια και τον
άνεµο. Τα υλικά αυτά βυθίζονται και συντελούν στον σχηµατισµό των θαλάσσιων ιζηµάτων.
Ιζήµατα πλούσια σε πυρίτιο δηµιουργούνται επίσης και από φυτοπλαγκτικούς οργανισµούς
(διάτοµα κ.λ.π), οι οποίοι µετά το θάνατό τους συγκεντρώνονται στον πυθµένα των ωκεανών.
Στα επιφανειακά στρώµατα των θαλάσσιων περιοχών, όπου παρατηρείται αφθονία διατόµων,
η τιµή της συγκέντρωσης του διαλυµένου πυριτίου είναι της τάξεως 10 mg/m3 περίπου. Σε

71
βαθύτερα στρώµατα παρατηρούνται πολύ µεγαλύτερες τιµές, που κυµαίνονται από 1000
mg/m3 (Β. Ατλαντικός) µέχρι 3000 mg/m3 (Ν. Ατλαντικός - περιοχή Ανταρκτικής).
Στις παράκτιες περιοχές παρατηρείται εποχιακή διακύµανση της συγκέντρωσης του πυριτίου
των επιφανειακών υδάτων της οποίας η µικρότερη (10-20 mg/m3) παρατηρείται κατά τους
µήνες Απρίλιο, Μάιο και Ιούνιο. Η χαµηλή αυτή τιµή συγκέντρωσης του πυριτίου οφείλεται
στην αυξηµένη χρησιµοποίησή του από τους φυτοπλαγκτικούς οργανισµούς κατά την
περίοδο αυτή. Η συγκέντρωση πυριτίου παίρνει τη µεγαλύτερη της τιµή (100-150 mg/m3)
κατά τη χειµερινή περίοδο. Εξαιτίας της γρήγορης επαναφοράς του πυριτίου στο θαλασσινό
νερό, µετά το θάνατο των φυτοπλαγκτικών οργανισµών, είναι δυνατόν να παρατηρηθεί υψηλή
τιµή συγκέντρωσης και κατά τον Αύγουστο.
Σηµειώνεται ότι στις θαλάσσιες περιοχές που βρίσκονται πολύ κοντά στις ακτές, η
συγκέντρωση του πυριτίου είναι πολύ µεγαλύτερη από την µέση τυπική τιµή των θαλασσών.
Αυτό οφείλεται στην επίδραση της παρουσίας, στα ύδατα αυτά, ποτάµιων υδάτων.
Σε φυσικά ύδατα η εξάντληση του πυριτίου περιορίζει την ανάπτυξη των φυτοπλαγκτικών
πληθυσµών. Εξάντληση του πυριτίου έχει ως αποτέλεσµα την παρεµπόδιση της διαίρεσης
του κυττάρου και τελικά την καταστροφή της δραστηριότητας του µεταβολισµού του.
Ίχνη θρεπτικών στοιχείων
Ορισµένα στοιχεία όπως ο σίδηρος, το µαγνήσιο και ο ψευδάργυρος υπηρετούν
σπουδαιότατο ρόλο σε συστήµατα οξειδώσεων (ο σίδηρος είναι συµπαράγοντας στη
φωτοσύνθεση), ενώ άλλα στοιχεία, όπως π.χ. ο µόλυβδος, ο ψευδάργυρος, το κοβάλτιο, ο
χαλκός, το βανάδιο κ.α., αποτελούν συµπαράγοντες µε τα ένζυµα που είναι ουσιώδη για την
ανάπτυξη των φυτών.
Το είδος των ενώσεων του σιδήρου στο θαλασσινό νερό εξαρτάται από την ποσότητα του
οξυγόνου που υπάρχει σ' αυτό. Η διαλυµένη ποσότητα σιδήρου στο θαλασσινό νερό είναι
εξαιρετικά µικρή, γεγονός που οφείλεται στην αδιαλυτότητα του υδροξειδίου του σιδήρου, ενώ
η ποσότητα των ιόντων Fe2O3+ που µπορούν να µείνουν διαλυµένα, είναι πολύ περιορισµένη.
Επίσης έχει αποδειχθεί ότι ενώσεις του σιδήρου της µορφής Fe(OH)3, που δηµιουργούν µε το
θαλασσινό νερό κολλοειδές διάλυµα, βρίσκονται σε πολύ µικρή ποσότητα.
Ο σίδηρος έχει αποδειχθεί ότι είναι περιοριστικός παράγοντας στο φυτοπλαγκτό και η έλλειψή
του µειώνει το φωτοσυνθετικό δυναµικό.
Οργανικά θρεπτικά συστατικά
Οργανικά θρεπτικά σε ελάχιστες ποσότητες, ειδικότερα οι βιταµίνες, µπορεί επίσης να είναι
µεγάλης σπουδαιότητας στη θάλασσα. Σχεδόν όλα τα είδη του θαλάσσιου φυτοπλαγκτού
είναι αυξοτροφικά και απαιτούν κοµπαλαµίνη, θειαµίνη ή βιοτίνη. Σε µικτές καλλιέργειες, η
παραγωγή και ελευθέρωση βιταµίνης από ένα είδος µπορεί να διεγείρει την ανάπτυξη ενός
άλλου είδους, αν και η παραγωγή των περισσοτέρων βιταµινών προέρχονται πιθανόν από τα
βακτήρια. Στη θάλασσα των Σαργασσών µικρά διάτοµα, που χρειάζονται τη B12, αυξάνουν

72
την αφθονία τους σε σχέση µε τα κοκκολιθοφόρα (τα οποία χρειάζονται µόνο θειαµίνη) κατά
την περίοδο του χρόνου που η βιταµίνη B12 είναι άφθονη.
Ανακύκληση θρεπτικών από ετερότροφους και χηµειοαυτότροφους οργανισµούς
Ορισµένοι φυτοπλαγκτικοί οργανισµοί µπορούν να αναπτύσσονται ετεροτροφικά πάνω σε
πηγές διαλυµένου οργανικού άνθρακα (σάκχαρα, οργανικά- οξέα- γαλακτικό οξύ, αµινοξέα ή
αλκοόλες). Γενικά πολλά υλικά µπορούν να προσλαµβάνονται από ένα δεδοµένο είδος
πλαγκτού αλλά δεν υποστηρίζουν την αύξηση του οργανισµού αυτού στο σκοτάδι. Είδη τα
οποία δε µπορούν ν' αναπτύσσονται στο σκοτάδι είναι δυνατόν να επισωρεύουν διάφορα
είδη οργανικών µορίων. Για ένα δεδοµένο είδος, ορισµένα οργανικά µόρια που
προσλαµβάνονται µπορεί να ανακόπτουν την ανάπτυξη, ενώ άλλες παρόµοιες ενώσεις (π.χ.
ένα διαφορετικό αµινοξύ) µπορεί να χρησιµοποιούνται ετεροτροφικά. Σε µερικές
περιπτώσεις, το φως εµποδίζει ετεροτροφική πρόσληψη.
Οι ετεροτροφικές δυνατότητες είναι ευρέως διαδεδοµένες µεταξύ των pennate διατόµων και
σπάνιες µεταξύ των centric διατόµων. Τα pennate διάτοµα είναι κυρίως βενθικά αλλά
ετεροτροφικά pennate και centric διάτοµα βρίσκονται σε πλαγκτικά οικοσυστήµατα µε υψηλές
συγκεντρώσεις διαλυµένου οργανικού υλικού. Μερικά δινοµαστιγωτά µπορούν να
καταβροχθίσουν κοµµατιασµένο υλικό και η φαγοκυττάρωση µπορεί να συµπληρώνει τη
φωτοσύνθεση.
Η παρουσία ετεροτροφικής πρόσληψης στο φυτοπλαγκτό συνιστά µια υπερκάλυψη της
θρέψης µε σαπροφυτικά βακτήρια. Σε αβαθείς κόλπους τα βακτήρια µπορεί επίσης να
συναγωνίζονται µε το φυτοπλαγκτόν για θρεπτικά που αναγεννώνται από τον πυθµένα.
Το φυτοπλαγκτόν καταναλώνει το µεγαλύτερο µέρος των διαλυµένων ανόργανων θρεπτικών
σε καλά φωτισµένα επιφανειακά νερά. Το φυτοπλαγκτόν είναι επίσης υπεύθυνο, κυρίως
κοντά στις ακτές, για την ετεροτροφική κατανάλωση των διαλυµένων οργανικών θρεπτικών.
Τα βακτήρια εντούτοις, είναι πιθανόν οι κύριοι ετροτροφικοί καταναλωτές στη στήλη του
νερού. Τα βακτήρια χρησιµοποιούν αποτελεσµατικά τόσο διαλυµένο όσο και κολλοειδές
υλικό συµπεριλαµβανοµένων της χιτίνης και της κυτταρίνης.
Αν και πανταχού παρόντα, τα βακτήρια είναι περισσότερο άφθονα σε συνδυασµό µε το
αιωρούµενο κολλοειδές υλικό. Σε αβαθείς λεκάνες, κατά τη διάρκεια της άνοιξης και του
θέρους, τα βακτήρια συχνά συµπίπτουν µε συγκεντρώσεις κολλοειδούς υλικού στη βάση της
θερµοκλινούς ζώνης. Στην ανοικτή θάλασσα, τα βακτήρια είναι παρόµοια συνδυασµένα µε το
στρώµα του οξυγόνου κάτω από τη θερµοκλινή ζώνη, η οποία αυτή καθαυτή συνδέεται µε
µικροβιακή κατανάλωση οργανικού υλικού. Επίσης αιωρούµενο κολλοειδές υλικό κοντά στον
πυθµένα παρέχει καταφύγιο σε µεγάλους πληθυσµούς βακτηρίων. Τα βακτήρια στη στήλη
του νερού είναι υπεύθυνα για την ταχεία πρόσληψη τέτοιου οργανικού υποστρώµατος, όπως
υδρογονάνθρακες, αµινοξέα και πεπτίδια. Κατά συνέπεια αυτά τα υποστρώµατα µε
διαλυµένα τα παραπάνω υλικά µπορούν να µετατραπούν σε κολλοειδές αδιάλυτο υλικό
διαθέσιµο στους καταναλωτές, όπως πρωτόζωα και σάλπες (tunicates).

73
Τα βακτήρια και τα ετερότροφα φύκη που προσκολλώνται στο κολλοειδές οργανικό υλικό
µπορεί να είναι η κύρια πηγή τροφής για τα κωπήποδα κοντά στις ακτές και στις εκβολές
ποταµών. Μια µεγάλη ποικιλία βακτηρίων εξειδικεύονται στο να χρησιµοποιούν
υποστρώµατα µε διαλυµένο ανόργανο υλικό για να δηµιουργούν την ενέργεια (ATP) που
απαιτείται για τη σύνθεση. Τέτοιοι χηµειοαυτότροφοι οργανισµοί οξειδώνουν υποστρώµατα
όπως NH4 (νιτροποιητικά βακτήρια) και H2S (θειοβακτήρια). Αυτές οι οµάδες βακτηρίων
τείνουν να είναι µη βιώσιµες ή αναποτελεσµατικές για διαβίωση σε οξυγονωµένα νερά. Κατά
συνέπεια η χηµειοσύνθεση περιορίζεται σε ανοξικά ή ελάχιστα οξυγονοµένα νερά. Τα
παραπάνω βακτήρια είναι ενεργά επίσης σε ανοξικά ιζήµατα.

74

75
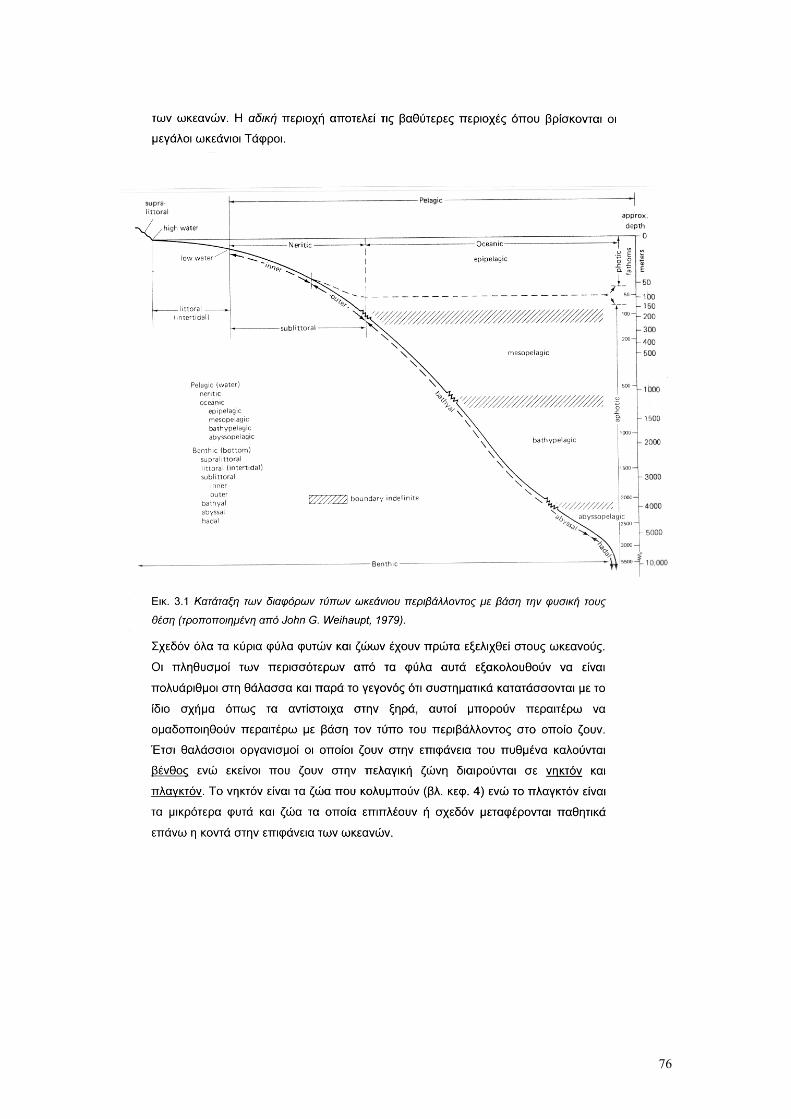
76
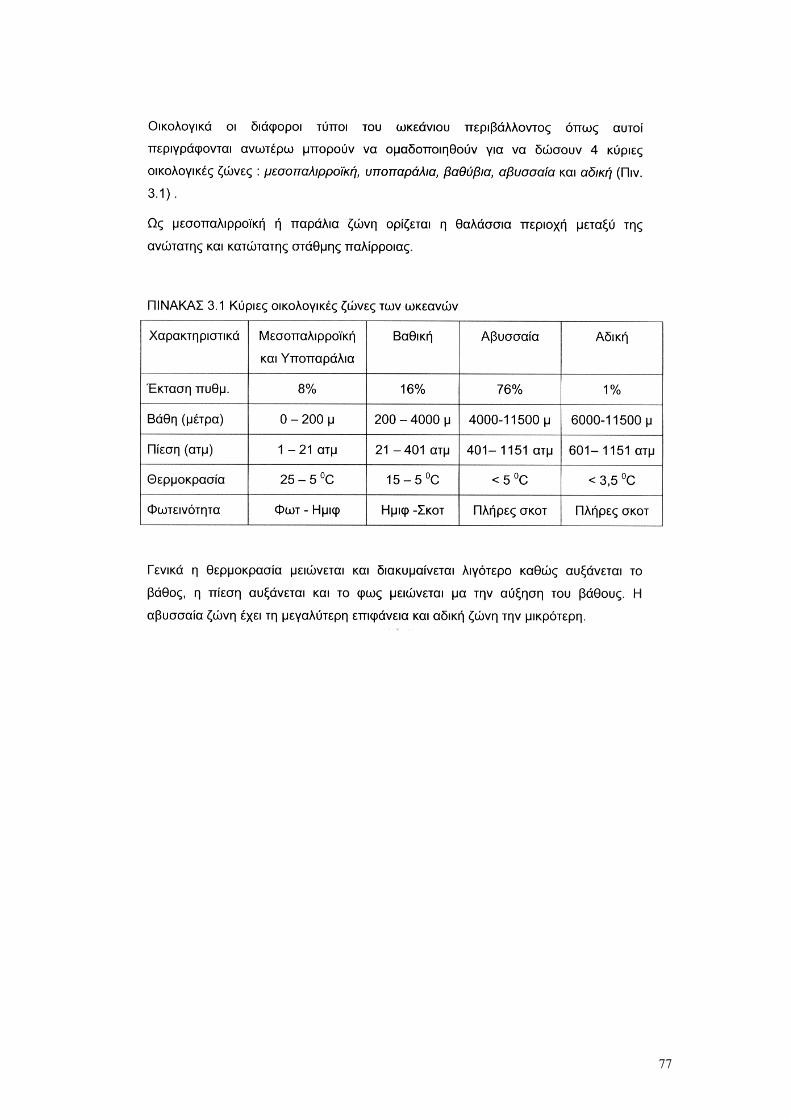
77

78
4. ΝΗΚΤΟΝ
∆εν υπάρχει σαφής διαχωριστική γραµµή µεταξύ πελαγικών ζώων που θεωρούνται
ως πλαγκτόν και εκείνων πού αποτελούν το νηκτόν. Τα περισσότερα ελεύθερα
κολυµβώντα ασπόνδυλα, γενικά, θεωρούνται ότι αποτελούν µέροs του πλαγκτού, αν
και πολλά απ' αυτά, όπως καρκινοειδή σκουλήκια και άλλα έχουν δυνατότητες
κολύµβησης ίσες µε εκείνες µερικών ζώων, τα οποία συνήθως θεωρούνται ως µέρος
του νηκτού. Πολλά µέλη του νηκτού εξάλλου, υπάρχουν επίσης ως πλαγκτόν κατά
τη διάρκεια των νεαρών σταδίων της ζωής τους.
∆ύο οµάδες ζώων -τα κεφαλόποδα (µαλάκια) και τα ψάρια- περιλαµβάνουν
πολυάριθµα, µεγάλα και δραστήρια κολυµβώντα είδη, τα οποία αποτελούν το
µεγαλύτερο ποσοστό των ζώων που γενικά θεωρούνται ως νηκτόν. Στο νηκτόν
συµπεριλαµβάνονται επίσης τα θαλάσσια θηλαστικά. Ούτε µεταξύ του νηκτού και του
βένθους υπάρχει κάποιος σαφής διαχωρισµός. Βενθοπελαγικά ψάρια και χταπόδια
π.χ. µπορούν να θεωρούνται ως µέρος της βενθικής κοινότητας. Εντούτοις τα
βενθοπελαγικά ψάρια και τα χταπόδια περιλαµβάνονται στο νηκτόν διότι κολυµβούν
εκτεταµένα και διότι οι πλησιέστεροι συγγενείς τους αποτελούν το µεγαλύτερο
ποσοστό των ζώων που συγκαταλέγονται στο νηκτόν.
4.1. Κεφαλόποδα
Η κλάση των κεφαλόποδων σήµερα αντιπροσωπεύει ένα αποµεινάρι µιας παλαιάς
και κάποτε κυρίαρχης οµάδας µαλακίων τα οποία διαφοροποιήθηκαν νωρίς από τα
πρωτόγονα αποθέµατα µαλακίων. Έγιναν καταπληκτικά άφθονα σε αριθµό ειδών
κατά τη διάρκεια των περιόδων περί το τέλος του Παλαιοζωικού και Μεσοζωικού,
αριθµούντα περισσότερο από 10.000 είδη για να λιγοστέψουν σήµερα σε 600 ζώντα
είδη περίπου. Μερικές από τις κύριες οµάδες έχουν εξαφανισθεί ολοκληρωτικά, ενώ
άλλα έχουν µερικούς αντιπροσώπους που ζουν και σήµερα. Εντούτοις, αρκετά από
τα σηµερινά κεφαλόποδα είναι πετυχηµένα, ευρισκόµενα σε πολύ µεγάλη αφθονία
και ευρέως κατανεµηµένα δίδοντα την εντύπωση ότι, τα χταπόδια π.χ. ακόµη και
στην παρούσα περίοδο, υπόκεινται σε δραστική εξελικτική διαφοροποίηση.
Σε αντίθεση µε τον εδραίο βενθικό χαρακτήρα των περισσότερων µελών του φύλου
των µαλακίων, τα κεφαλόποδα έχουν εξελιχθεί σε δραστήρια σαρκοφάγα και
πτωµατοφάγα ζώα. Τα πρώτα κεφαλόποδα ήταν βενθικά, διατηρώντας την
πρωτόγονη αµφίπλευρη συµµετρία των υποτιθέµενων προγόνων µαλακίων, µε µία
προς τα πίσω αναπτυγµένη µανδυακή κοιλότητα, διαφέροντα όµως ως προς τη

79
µεγάλη ανάπτυξη της ραχιαίας πλευράς και της σπλαχνικής κοιλότητας, έτσι ώστε ο
ραχοκοιλιακός άξονας εξελικτικά έγινε ο λειτουργικός διαµήκης άξονας του σώµατος.
Οι πολύ εκτεταµένες προς τα εµπρός και προς τα πίσω επιφάνειες, τώρα
εξυπηρετούν ως λειτουργικές ραχιαίες και κοιλιακές πλευρές αντίστοιχα. Το πόδι
έγινε πολυσχιδές προς τα εµπρός και λίγο-πολύ συγχωνεύθηκε µε το κεφάλι
σχηµατίζοντας ένα στοµατικό στεφάνι µε µακριά πλοκάµια γύρω από το στόµα. Ένα
τµήµα του ποδός των µαλακίων έχει τροποποιηθεί σε ένα κοιλιακό σίφωνα δια µέσου
του οποίου το κυκλοφορούν ρεύµα ύδατος στη µανδυακή κοιλότητα εκτινάσσεται
προς τα έξω. Τα πλοκάµια έχουν αναπτύξει βεντούζες προσκολλητικές και σε
µερικές περιπτώσεις µε χιτινώδη αγκίστρια. Η επιµηκυνθείσα κοιλιακή µανδυακή
κοιλότητα, που περιέχει τα ζεύγη των βραγχίων - δύο ζεύγη στα περισσότερα
πρωτόγονα κεφαλόποδα, ένα ζεύγος στα πλέον σύγχρονα είδη - λειτουργεί
αποτελεσµατικά για την κυκλοφορία του νερού µε τη συνεργασία των µυϊκών ιστών
του µανδύα. Το νερό εισέρχεται κατά µήκος όλου του ανοίγµατος του χείλους της
µανδυακής κοιλότητας και αυτό το σχεδόν επίµηκες κοίλωµα κλείνει και εκτινάσσει το
νερό από το σίφωνα. Οι ιστοί του µανδύα έχουν χάσει τα βλεφαριδωτά νηµάτια πού
βρίσκονταν στους ιστούς των µαλακίων και έχουν µετατραπεί σε ισχυρό µυώδες
σύστηµα, µε αρκετά µεγάλη δύναµη ώστε το εκτοξευόµενο νερό να είναι περισσότερο
αποτελεσµατικό για την κίνηση από όσο τα πλοκάµια, τα οποία επιτρέπουν στο ζώο
να σύρεται πάνω στο υπόστρωµα.
Τα πρωτόγονα κεφαλόποδα, όπως τα περισσότερα από τα άλλα µαλάκια, είχαν
µεγάλα εξωτερικά όστρακα, τα οποία σε πολλά είδη είχαν γίνει πολλαπλώς ελικοειδή
και καθώς τα ζώα αναπτύσσονταν, χωρίζονταν σε διαδοχικά διαµερίσµατα. Τα ζώα
ζούσαν στο τελευταίο και µεγαλύτερο διαµέρισµα, ενώ το αµέσως οπίσθιο
διαµέρισµα γέµιζε µε αέρα για να ισορροπήσει το βάρος και όλα τα άλλα
διαµερίσµατα συνδέονταν µε το οπίσθιο µέρος του σώµατος των κεφαλόποδων µε
ένα µεµβρανώδη σωλήνα πλήρη µε σωµατικό ιστό (siphuncle). Ο ιστός αυτός
εκκρίνει αέριο στα άδεια διαµερίσµατα του οστράκου και έτσι το ζώο καθίσταται ικανό
να επιπλέει. Τα τέσσερα ζώντα είδη του γένους Νautilus είναι οι µόνοι ζώντες
αντιπρόσωποι µιας µεγάλης οµάδας των τετραβραγχίων. Τα ζώα αυτά είναι επίσης
πρωτόγονα έχοντα πολυάριθµα ισοµεγέθη πλοκάµια χωρίς βεντούζες, ατελώς
αναπτυγµένα µάτια χωρίς φακούς, χωρίς σάκο µελάνης, καρδία τετράχωρη χωρίς
βραγχιακές καρδίες και άλλα χαρακτηριστικά που τοποθετούν τα ζώα αυτά µακριά
από τα σύγχρονα κεφαλόποδα.
Τα περισσότερα σύγχρονα κεφαλόποδα έχουν όστρακο πολύ µειωµένης ανάπτυξης
πλήρως χωµένο µέσα στο ραχιαίο µανδυακό µύ ή δεν έχουν καθόλου όστρακο. Ο

80
µανδύας είναι πολύ περισσότερο µυώδης από εκείνον του Νautilus, αvαπτυχθείς σε
ένα αποτελεσµατικό όργανο κολύµβησης.
Το νευρικό σύστηµα και τα όργανα αίσθησης είναι άριστα αναπτυγµένα. Τα
εγκεφαλικά, πλευρικά, ποδικά, προποδικά και σπλαχνικά γάγγλια είναι
συγχωνευµένα όλα µαζί σε ένα περίπλοκο εγκέφαλο εγκατεστηµένο µέσα σ' ένα
χονδρώδη κρανίο. Το εντυπωσιακό µάτι των κεφαλoπόδων, ανάλογο σε κάθε
άποψη µε εκείνο των σπονδυλωτών -µε φακούς, εµπρόσθιο και οπίσθιο
διαµερίσµατα, αµφιβληστροειδή χιτώνα και οπτικά νεύρα (ένα από κάθε πλευρά του
εγκεφαλικού γαγγλίου) -έχει προκαλέσει το θαυµασµό και την περιέργεια των
Βιολόγων στη διάρκεια των ετών.
Μια τέτοια εντυπωσιακή κατασκευή πρέπει να έχει εξελιχθεί ανεξάρτητα δύο φορές,
µέσα στα πλαίσια των τόσο διαφορετικών γραµµών εξέλιξης των µαλακίων και των
σπονδυλωτών, σε µια από τις περισσότερο παράξενες εξελικτικές συµπτώσεις ή
περιπτώσεις συγκλίνουσας εξέλιξης. Σε µερικές περιπτώσεις το µάτι των
κεφαλόποδων φαίνεται περισσότερο λογικά κατασκευασµένο από όσο είναι το µάτι
των σπονδυλωτών. Τα αισθητήρια κύτταρα είναι µπροστά, αντί πίσω από τα νεύρα
τα οποία συνδέουν αυτά µε τον οπτικό αγωγό και δεν υπάρχει τυφλό σηµείο στον
αµφιβληστροειδή για το πέρασµα του οπτικού νεύρου.
Πολλά κεφαλόποδα έχουν χρωµατοφόρα δύο ή τριών διαφορετικών χρωµάτων, κάτω
από νευρικό έλεγχο επιτρέποντα στα ζώα αυτά να πραγµατοποιούν εντυπωσιακές
και γρήγορες αλλαγές. Όταν το ζώο παρατηρείται από κοντά, γίνεται αντιληπτή µια
συνεχής αλλαγή χρωµάτων, παρά το γεγονός ότι το συνολικό µέσο χρώµα µπορεί να
παραµένει το ίδιο. Αυτό το παίξιµο των χρωµάτων στο δέρµα µπορεί να συνεχίζεται
για αρκετές ώρες µετά τον θάνατο του κεφαλόποδου.
Το στόµα των κεφαλοπόδων είναι εφοδιασµένο µε κεράτινο ράµφος, κάπως
παρόµοιο µε εκείνο του παπαγάλου. Επίσης είναι εφοδιασµένο µε οδοντικό όργανο.
Με το στόµα συνδέονται µεγάλοι σιελογόνοι αδένες και το δάγκωµα µερικών ειδών
είναι δηλητηριώδες και καµιά φορά θανατηφόρο. Εν τούτοις πρέπει να τονιστεί ότι ο
γενικός φόβος που υπάρχει για τα χταπόδια από αρκετό κόσµο δεν είναι
δικαιολογηµένος, δεδοµένου ότι τα ζώα αυτά κρύβονται και αποφεύγουν τον κόσµο,
όπου είναι δυνατόν. Ακόµη και όταν πιαστούν από κάποιον, σπάνια δαγκώνουν.
Αντίθετα τα καλαµάρια φαίνονται περισσότερο επιθετικά στο να δαγκώνουν όταν
συλλαµβάνονται και τους δίδεται ευκαιρία να δαγκώσουν. Πάντως όλα τα
κεφαλόποδα είναι τελείως ακίνδυνα έξω από το νερό όταν κάποιος τα χειρίζεται µε
προσοχή κρατώντας τα δάκτυλά του µακριά από το στόµα.

81
Ο χαρακτηριστικός αµυντικός µηχανισµός των περισσοτέρων κεφαλοπόδων είναι η
εκτόξευση µελάνης από ένα σάκο, που είναι τοποθετηµένος στην κοιλιακή επιφάνεια
του συκωτιού και ο οποίος αδειάζει µαζί µε ένα πίδακα νερού δια µέσου του σίφωνα.
Η εκτοξευόµενη µελάνη πολλές φορές χρησιµεύει στο ζώο για να ξεφεύγει τη
σύλληψη από τον άνθρωπο πίσω από ένα παραπέτασµα µαύρου σύννεφου, τις
περισσότερες φορές όµως η µελάνη λειτουργεί ως ένα οµοίωµα στο νερό που
µπερδεύει τον θηρευτή. Η πλούσια καφετιά µελάνη της σουπιάς έχει χρησιµοποιηθεί
από καλλιτέχνες πολλά χρόνια πριν. Μερικά βαθυπελαγικά καλαµάρια εντούτοις,
έχουν µελάνη µάλλον για να λαµπιρίζουν παρά για τους λόγους που έχουν τη µελάνη
οι συγγενείς τους, οι οποίοι ζουν σε καλά φωτισµένα νερά.
Πολλά καλαµάρια, ειδικότερα διάφορα oegopsids που ζουν σε βαθιά νερά, έχουν ένα
µεγάλο οπίσθιο κοιλωµατικό χώρο, που είναι γεµάτος µε υλικό σε µορφή ζελέ και
λειτουργεί ως υδροστατικό όργανο. Αυτός ο χώρος έχει ένα πλεονέκτηµα έναντι της
νηκτικής κύστης των ψαριών. Το πλεονέκτηµα αυτό οφείλεται στο γεγονός ότι δεν
αλλάζει σηµαντικά όγκο µε την ταχεία αλλαγή της στάθµης του νερού. Τέτοια
κεφαλόποδα συχνά κρεµούν χαρακτηριστικά το κεφάλι τους προς τα κάτω όταν
αναπαύονται.
Μια παράξενη και χαρακτηριστική τροποποίηση συνδεόµενη µε το µοντέλο κίνησης
είναι ένα ζευγάρι από πτυσσόµενα φύλλα ή βαλβίδες στο τέλος του εντέρου, που
ανοίγουν στη µανδυακή κοιλότητα πίσω από το σίφωνα. Όταν ο µανδύας
συστέλλεται ξαφνικά, δηµιουργώντας τη σηµαντική πίεση στη µανδυακή κοιλότητα
που είναι απαραίτητη για την εκτόξευση του πίδακα, αυτά τα πτυσσόµενα φύλλα
κλείνουν το άνοιγµα για να προστατευθεί το ίδιο το ζώο από την είσοδο κάποιου
ανεπιθύµητου υλικού, όταν το ζώο µετακινείται κάπου βιαστικά.
Μερικά από τα καλαµάρια συναγωνίζονται ή και ξεπερνούν ακόµη στην ταχύτητα και
στη χάρη κολύµβησης τα περισσότερα ψάρια. Τα ζώα αυτά µπορούν να
κατευθύνουν το σίφωνα προς οποιαδήποτε κατεύθυνση. Μερικά απ' αυτά έχουν
παρατηρηθεί να πηδούν από το νερό και να διασχίζουν ιπτάµενα σηµαντικές
αποστάσεις στον αέρα, κατά τον τρόπο που πετούν τα ιπτάµενα ψάρια.
Η ερωτοτροπία και το ζευγάρωµα των κεφαλόποδων είναι επίσης χαρακτηριστικά και
ποικίλα. Μερικά είδη εµφανίζουν ιδιαίτερες δραστηριότητες ερωτοτροπίας,
συµπεριλαµβανοµένης και της αλλαγής του χρώµατος. Το σπέρµα των αρσενικών
περιέχεται σε µεγάλες σπερµατοφόρες θήκες και µεταφέρεται στη µανδυακή
κοιλότητα του θηλυκού ατόµου µε ένα από τα πλοκάµια, το δεξιό ή αριστερό τέταρτο
στα καλαµάρια και στις σουπιές και το αριστερό τρίτο στα χταπόδια, που

82
τροποποιούνται ειδικά γι' αυτό τον σκοπό. Στο Αrgonauta το τελικό τµήµα του
αρσενικού πλοκαµιού, που περιλαµβάνει πολλές βεντούζες οι οποίες συγκρατούν τα
σπερµατοφόρα όργανα, αποκολλάται και αφήνεται στη µανδυακή κοιλότητα του
θηλυκού. Η παρουσία του τµήµατος αυτού µπερδεύτηκε από τον Cuvier µε ένα
χαρακτηριστικό µε πολλές βεντούζες παρασιτικό σκουλήκι και του δόθηκε το όνοµα
hectocotylus. Από τότε το τροποποιηµένο τµήµα του αρσενικού πλοκαµιού έχει
µείνει γνωστό ως ‘’hectocotylus’’. Tα περισσότερα κεφαλόποδα δεν αφήνουν τµήµα
πλοκαµιού στη µανδυακή κοιλότητα του θηλυκού ατόµου αλλά απλά χρησιµοποιούν
το ειδικά τροποποιηµένο πλοκάµι για να µεταφέρουν το σπέρµα.
Το σπερµατοφόρο όργανο αυτό καθαυτό είναι µια εντυπωσιακή κατασκευή. Ένα
ώριµο σπερµατοφόρο συνήθως αποθέτει το σπέρµα αν τοποθετηθεί σε λίγο
θαλασσινό νερό πάνω σε µια αντικειµενοφόρο πλάκα µικροσκοπίου. Αποτελείται
από ένα φουσκωµένο σαν σάκο τµήµα, που περιέχει το σπέρµα και ένα µικρό
σωληνωτό σύστηµα σαν λαιµός µε ένα ακραίο κάλυµµα που συνδέεται µε ένα µακρύ
νηµάτιο. Μέσα στο λαιµό είναι ένας περίπλοκος σπειροειδώς τυλιγµένος σαν
ελατήριο µηχανισµός, ο οποίος, όταν το σπερµατοφόρο εκτίθεται σε θαλασσινό νερό,
ξαφνικά ελευθερώνεται και αρχίζει να ξετυλίγεται τεντώνοντας έτσι το σωληνώδες
τµήµα του σπερµατοφόρου σε ένα πολύ µεγαλύτερο µήκος. Σε µερικά κεφαλόποδα -
π.χ. Rossia- το τελικό τµήµα του σπερµατοφόρου σωλήνα καθώς επιµηκύνεται,
διαπερνά τους ιστούς του θηλυκού µέσα στη µανδυακή κοιλότητα και µετά
εκρήγνυται, όπως ένα µπαλόνι, αγκιστρώνοντας το σπερµατοφόρο σταθερά σε µια
θέση, που µοιάζει σαν παρασιτικό σκουλήκι µε το κεφάλι του θαµµένο µέσα στον
ιστό. Σε µεγάλα χταπόδια οι σπερµατοφόροι σωλήνες µπορεί να είναι µακρείς πάνω
από 35 εκτµ.
Μερικά από τα ζώντα σε κοπάδια καλαµάρια όπως το Loligo µπορεί να βρίσκονται σε
απέραντα µεγάλους αριθµούς, γεγονός που δηµιουργεί τις προϋποθέσεις µιας
πολύτιµης αλιείας. Οι µυώδεις ιστοί του µανδύου και µέρη από τα πλοκάµια είναι
εξαιρετικά για φαγητό. Τα καλαµάρια δεν είναι συνήθως εξαιρετικά πολυάριθµα στα
βόρεια νερά, αλλά καµιά φορά διαπιστώνεται άφθονη η παρουσία τους ακόµη και στο
Βορρά.
Μερικά από τα γιγαντιαία καλαµάρια, όπως τα Architeuthis και Ommastephes
αποτελούν την κύρια τροφή του είδους της φάλαινας sperm whale, ένα είδος το
οποίο ήταν αρκετά άφθονο. Το γεγονός ότι αυτές οι φάλαινες είναι αχόρταγοι
καταβροχθιστές µε γρήγορη χώνευση, αποδεικνύει ότι τα γιγαντιαία καλαµάρια
πρέπει να είναι πολύ πιο άφθονα από όσο υποτίθεται ότι είναι µε βάση τα στοιχεία
παρατηρήσεων αλιείας και περιπτώσεων αποβολής στις ακτές πεθαµένων ατόµων.

83
Τα σύγχρονα κεφαλόποδα τοποθετούνται τόσο µακριά από τα µαλάκια ώστε φαίνεται
να είναι εκτός θέσεως από το φύλο αυτό και για πολλές απόψεις παροµοιάζονται
περισσότερο προς τα σπονδυλωτά. Το εύρος µεγεθών του σώµατος οµοιάζει µε
εκείνο των σπονδυλωτών, µε τα µεγαλύτερα κεφαλόποδα να φθάνουν σε µήκος
µεγαλύτερο από 12 µ, συµπεριλαµβανοµένων και των δύο πολύ µεγάλων πλοκαµιών
τους, γεγονός που καθιστά τα κεφαλόποδα πολύ µεγαλύτερα και πιο ογκώδη από
όλα τα ασπόνδυλα. Ο ιστός του σώµατος είναι µυώδης και έχει ένα σηµαντικά
συνεκτικό ιστό που µοιάζει µε εκείνον των σπονδυλωτών, περιέχοντας ακόµη και
καθαρό χόνδρο. Το αποµεινάρι του οστράκου του καλαµαριού αποτελεί ένα σκληρό
(άκαµπτο) στοιχείο σε µια θέση ανάλογη προς εκείνη της σπονδυλικής στήλης των
σπονδυλωτών. Σε αντίθεση µε τα άλλα µαλάκια, τα κεφαλόποδα έχουν κλειστό
κυκλοφορικό σύστηµα µε τριχοειδή αγγεία. Η ενεργητικότητά τους, η εξυπνάδα τους
και η νευροµυική τους ανάπτυξη συναγωνίζονται µε εκείνα των ψαριών. Πάνω απ'
όλα όµως, τα µάτια τους προσδίδουν στα ζώα αυτά µια ιδιαίτερη εµφάνιση
σπονδυλωτών. Είναι δύσκολο να κοιτάξεις στα µάτια ένα χταπόδι και να πιστέψεις
ότι δε βλέπεις κάτι πιο κοντινό προς τον άνθρωπο, κάτι σχεδόν ανθρώπινο παρά το
µάτι ενός µαλακίου.
4.2. Ψάρια
Τα ψάρια είναι τα περισσότερο άφθονα από όλα τα σπονδυλωτά τόσο σε αριθµό
ειδών όσο και σε αριθµό ατόµων. Εκτιµάται ότι υπάρχουν περισσότερα από 22.000
διαφορετικά είδη στην κλάση Ιχθύες. Το µέγεθός τους κυµαίνεται από 1 εκτµ.
περίπου, σε ώριµα γεννητικά άτοµα, µέχρι 21 µ περίπου µήκος και πολλούς τόνους
βάρος που φθάνει ο γιγάντιος whale shark. Τα ψάρια συνεχίζουν να αναπτύσσονται
σε όλη τη διάρκεια της ζωής τους, αν και ο βαθµός ανάπτυξής τους µειώνεται
σηµαντικά µετά το στάδιο της πρώτης γεννητικής ωριµότητας, έτσι ώστε το µήκος
των ωρίµων ατόµων ενός δεδοµένου είδους είναι τουλάχιστον σε πολλές
περιπτώσεις, λιγότερο αυστηρά καθορισµένο από όσο είναι σε πολλά άλλα ζώα.
Τα ψάρια, ζώα που µπορούν και κολυµπούν ελεύθερα, χωρίς σοβαρό ανταγωνισµό
από άλλες οµάδες, έχουν κυριαρχήσει σε όλα τα υδάτινα περιβάλλοντα. Μόνο τα
καλαµάρια και τα κητώδη έχουν επιτυχώς εισβάλλει στο βασίλειό τους. Ο
ανταγωνισµός µεταξύ των ειδών έχει οδηγήσει σε εξελικτική απόκλιση προς όλες τις
κατευθύνσεις και την επέκτασή τους σχεδόν σε κάθε οικολογικό θώκο. Η ποικιλότητα
µεταξύ των ψαριών είναι πολύ µεγαλύτερη από οποιαδήποτε άλλη µέσα στα
σπονδυλωτά, κάνοντας τα ψάρια ένα εξαιρετικά παραγωγικό πεδίο για την µελέτη της

84
εξέλιξης. Η απεραντοσύνη του υδάτινου περιβάλλοντος συµβάλλει επίσης σ' αυτή
την ποικιλότητα κάνοντας δυνατή την επιβίωση διαφόρων τύπων, οι οποίοι µπορεί να
έχουν εκτοπισθεί από διάφορες άλλες οµάδες στα κέντρα της αφθονίας τους, µπορεί
όµως και να συνεχίζουν να υπάρχουν σε άλλες περιοχές σε αποµονωµένα ή βαθιά
νερά. Η αφθονία των ψαριών είναι η ανταπόκριση της αφθονίας του πλαγκτού σε
τοπικό επίπεδο αλλά επίσης και η ανταπόκριση του τύπου του φυτοπλαγκτού.
∆εδοµένου ότι όλο το φυτοπλαγκτόν και ένα µεγάλο τµήµα του ζωοπλαγκτού
περιορίζονται στα ανώτερα στρώµατα του νερού µέσα ή κοντά στην εύφωτη ζώνη, τα
περισσότερα ψάρια βρίσκονται επίσης εκεί. Οι περιοχές της νηριτικής ζώνης, οι
οποίες δέχονται υλικά πού προέρχονται από τη στεριά και τα οποία υλικά
εµπλουτίζονται µε την αύξηση του στροβιλισµού, ανάβλυσης υδάτων κλπ, είναι
χαρακτηριστικά πλουσιότερες σε ψάρια. Τέτοια νερά παρέχουν ποικιλότητα
οικολογικών θώκων, πολύ µεγαλύτερη από εκείνη που συναντάται στην ανοικτή
θάλασσα και οι περιοχές αυτές των υδάτων περιέχουν επίσης τη µεγαλύτερη
ποικιλότητα στα είδη των ψαριών.
4.2.1. Χρωµατισµός
Ο χρωµατισµός των ψαριών είναι σχεδόν απέραντης ποικιλότητας και οµορφιάς και
είναι σηµαντικής οικολογικής σπουδαιότητας. Τα πιο διαδεδοµένα χαρακτηριστικά
είναι η προσποίηση του χρώµατός τους προς το υπόστρωµα ή το χώρο όπου
διαβιούν και η δι' αντισκιάσεως εξαφάνιση, προσαρµογές που έχουν ουσιαστική
σηµασία επιβίωσης τόσο για το θηρευτή, όσο και για το θύµα. Τα περισσότερα είδη
που κολυµβούν κοντά στην επιφάνεια της θάλασσας, όπως η ρέγγα, ο τόνος, το
σκουµπρί κ.α., έχουν χρώµα πρασινωπό ή προς το µπλε από το επάνω µέρος και
προς το λευκό, το ασηµένιο ή το ελαφρώς χρωµατιστό από κάτω. Από τα είδη εκείνα
που ζουν στα παράκτια νερά τα περισσότερα είναι πράσινα και από εκείνα που ζουν
µακριά από τις ακτές τα περισσότερα είναι µπλε. Έτσι ταυτίζονται µε το χρώµα των
νερών στα οποία ζουν όταν τα βλέπουµε από πάνω ενώ το ελαφρό τους χρώµα από
κάτω αναµιγνύεται µε το χρώµα του ουρανού, όταν τα βλέπουµε από κάτω,
ταυτιζόµενο µε το επιφανειακό φιλµ, το οποίο λάµπει από κάτω σαν καθρέπτης.
Τα βενθοπελαγικά ψάρια, τα οποία ζουν κοντά στον πυθµένα, τείνουν να είναι
οµοιόµορφα χρωµατισµένα µε διάσπαρτα στίγµατα ουδέτερων χρωµάτων, που
ταυτίζονται λίγο ή πολύ µε τον πυθµένα επί του οποίου ζουν. Πολλά απ' αυτά,
ειδικότερα τα πλατύψαρα, µπορούν γρήγορα να αλλάζουν χρώµα και τύπο για να
ταυτιστούν µε το υπόστρωµα.

85
Στα ψάρια αυτά η ανακλαστική δυνατότητα του περιβάλλοντος υποστρώµατος είναι
περισσότερο σπουδαίος παράγοντας για την ανταπόκριση των χρωµατοφόρων παρά
η ένταση του φωτισµού από πάνω. Είναι περισσότερο σπουδαίο γι' αυτά να
ταυτίζονται µε το υπόστρωµα, σε οποιονδήποτε προσπίπτοντα φωτισµό, παρά να
ανταποκρίνονται απλά στην ένταση του φωτός.
Το ανώτερο και κατώτερο µισό του ορατού πεδίου επηρεάζουν την ανταπόκριση στο
φως µε αντίθετο τρόπο. Το φως που ανακλάται από το υπόστρωµα προσβάλλει
περισσότερο το ανώτερο τµήµα του αµφιβληστροειδούς. Το φως που έρχεται από
πάνω προσβάλλει το κατώτερο τµήµα του αµφιβληστροειδούς. Η αναλογία µεταξύ
των δύο είναι σπουδαίος παράγοντας. Η αύξηση του φωτισµού του ανώτερου µισού
του αµφιβληστροειδούς σε σχέση µε το κατώτερο, οδηγεί στον αποχρωµατισµό ενώ
το αντίθετο οδηγεί στο σκοτείνιασµα.
Αν το υπόστρωµα φωτίζεται από µια πηγή φωτισµού, που δεν τη βλέπει το ψάρι, δε
λαµβάνεται πλέον η συνήθης αναλογία φωτισµού του πάνω µισού και του κάτω
µισού του αµφιβληστροειδούς. Το ψάρι µπορεί να υποθέτει ένα τελείως διαφορετικό
χρωµατισµό από εκείνον που θα υπέθετε κανονικά στο ίδιο υπόστρωµα, φωτιζόµενο
µε την ίδια ένταση από πάνω, µε φωτισµό τον οποίο βλέπει το ψάρι.
Τα ψάρια που ζουν στις λιµνούλες που σχηµατίζονται στις περιοχές παλιρροιών,
ανάµεσα σε ποικιλία έγχρωµων φυτών, κοραλλιογενών υφάλων και διαφόρων ζώων
των περιοχών αυτών. είναι χρωµατισµένα φωτεινά µε ποικιλία χρωµάτων, συχνά
ταυτιζόµενα µε τους βράχους, τα κοραλλιογενή φυτά ή άλλους χαρακτήρες του
περιβάλλοντός τους µε τρόπο εντυπωσιακό.
Η δι' αντισκιάσεως εξαφάνιση είναι ο κανόνας µεταξύ των ψαριών που ζουν σε καλά-
φωτισµένα νερά, µε το ανώτερο τµήµα να είναι αρκετά σκούρο ώστε να εκµηδενίζει
τη σκιά του κατώτερου τµήµατος του ψαριού, προσδίδοντας στο όλο ψάρι µια µάλλον
επίπεδη µη διακρινόµενη φιγούρα. Μερικές αξιοπρόσεκτες εξαιρέσεις σ' αυτό το
πρότυπο επιβεβαιώνουν τον σκοπό τους, όπως το γατόψαρο του Νείλου Synodontis,
το οποίο συνηθίζει να κολυµπάει µε το πάνω µέρος κάτω και έχει την κοιλιακή
πλευρά σκοτεινή, καθώς και είδη του γένους Remora, τα οποία προσκολλώνται
στους καρχαρίες δια µέσου µιας ραχιαίας κολλητικής βεντούζας. Έτσι η ραχιαία
επιφάνεια του ψαριού αυτού, που βρίσκεται στη σκιά του καρχαρία, φωτίζεται ενώ η
κοιλιακή επιφάνεια αντισκιάζεται (countershaded).
Ζώνες, ρίγες ή στίγµατα σε έντονα αντιτιθέµενα χρώµατα, αν και κάνουν ένα ψάρι, σε
περίπτωση που βρίσκεται έξω από το περιβάλλον του, εξαιρετικά εµφανές, µπορεί
να υπηρετούν επίσης προστατευτική και αποκρυπτική λειτουργία µε το να σπάζουν

86
τη φιγούρα του ψαριού. Τέτοια πρότυπα συνήθως περιλαµβάνουν µια σκούρα ταινία
κατά µήκος του µατιού, κάνοντας το ψάρι λιγότερο εµφανές. Επίσης καµιά φορά
µικρά στίγµατα ή σηµάδια σαν µάτια σε άλλα µέρη του σώµατος µπορεί να
χρησιµοποιούνται για να µπερδεύουν τον θηρευτή. Μερικά ψάρια µιµούνται ειδικά
φύκια µε τα οποία συνεργάζονται µε ένα καταπληκτικό τρόπο, που περιλαµβάνει όχι
µόνο το χρώµα αλλά και την κατεύθυνση και το σχήµα του σώµατος καθώς και την
ανάπτυξη εξαρτηµάτων από την επιφάνεια του σώµατος, που µοιάζουν στο σχήµα τα
φύκη.
Ψάρια που ζουν περισσότερο χρόνο στα µεσοπελαγικά δύσφωτα βάθη είναι
συνήθως κόκκινα, όταν τα βλέπουµε σε λευκό φως, αλλά στο δικό τους περιβάλλον,
κάτω από το βάθος διείσδυσης του κόκκινου φωτός, τα ψάρια αυτά τείνουν να
απορροφούν τις πρασινωπές ακτίνες του φωτός που φτάνουν σ' αυτό και
ευρισκόµενα σε περιοχή όπου δεν υπάρχουν κόκκινες ακτίνες, καθίστανται αφανή.
Άλλα τείνουν να είναι χωρίς χρώµα, αργυρώδη ή µερικώς διαφανή.
Κάτω από τα µεσοπελαγικά βάθη, στα βαθιά νερά της βαθυπελαγικής και αβυσσαίας
ζώνης, όπου δεν υπάρχει φως, τα περισσότερα από τα ψάρια είναι οµοιόµορφα
µαύρα, εκτός των οργάνων βιοφωσφορισµού.
Μερικά από τα χρωµατοφόρα των ψαριών πιθανόν ασκούν προστατευτική λειτουργία
παρέχοντας καταφύγιο από το υπερβάλλον φως σε ζωτικά όργανα. Οι
συγκεντρώσεις µελανίνης, η οποία συνήθως είναι επικεντρωµένη πάνω στο µυαλό,
πιθανόν ασκεί µια τέτοια λειτουργία και η µαύρη περιτοναϊκή επένδυση της
κοιλότητας του σώµατος πολλών ψαριών µπορεί να προστατεύει τα εντόσθια ή
µερικά από τα χωνευτικά ένζυµα.
Μερικά είδη ψαριών, που είναι επιθετικοί υποστηρικτές του χώρου τους, πολεµώντας
κάθε εισβολέα, ή εκείνα που εµπλέκονται σε δυνατή αντιζηλία για το σύντροφό τους,
έχουν καταφανή προειδοποιητικά χρώµατα, τόσο για να φοβίζουν τους ανταγωνιστές
όσο και για την αναγνώριση του φύλου. Σε µερικά ψάρια το αρσενικό γίνεται
περισσότερο έντονα ή βαθιά χρωµατισµένο κατά τη διάρκεια της αναπαραγωγής.
Είδη τα οποία διαλέγουν συντρόφους και τα οποία έχουν µια περίπλοκη σεξουαλική
συµπεριφορά, τείνουν να δείχνουν µεγαλύτερη διαφορά µεταξύ των φύλων από ό,τι
κάνει η πλειονότητα των άλλων ψαριών στα οποία τα δύο φύλα µοιάζουν στην
εµφάνιση.
4.2.2. Προσαρµογές για κολύµβηση
Τα πτερύγια των ψαριών πιστεύεται ότι έχουν προέλθει δια µέσου διαίρεσης και
τροποποίησης πτυχών πτερυγίων, όπως εκείνων του Amphioxus, ο οποίος έχει

87
ζεύγη πλευροκοιλιακών και µεταπλευρικών πτυχών, που βαίνουν κατά µήκος του
εµπρόσθιου τµήµατος του σώµατος και συνενούνται πίσω στην περιοχή περίπου της
έδρας σε µια µεσαία πτυχή. Η πτυχή αυτή βαίνει προς τα πίσω, γύρω από το
οπίσθιο τέλος του σώµατος και συνεχίζει προς τα εµπρός κατά µήκος της µέσης
ραχιαίας γραµµής, σαν µια ραχιαία πτυχή πτερυγίου. ∆ιαίρεση του ζεύγους των
πλευρικών πτυχών δηµιούργησε δύο ζεύγη πλευροκοιλιακών πτερυγίων ήτοι ένα
ζεύγος θωρακικών πτερυγίων προς τα εµπρός και ένα ζεύγος κοιλιακών πτερυγίων
προς τα πίσω. Αυτά τα πτερύγια είναι προάγγελοι των ζυγών άκρων όλων των
ανωτέρω σπονδυλωτών. Η µονή πτυχή οδήγησε στη δηµιουργία του ουραίου
πτερυγίου ή της ουράς, ένα ποικίλλοντα αριθµό ραχιαίων πτερυγίων και
πτερυγιωδών αποφύσεων, το εδρικό πτερύγιο και τις πτερυγιωδεις αποφύσεις πίσω
από την έδρα. Στην κολύµβηση, η προώθηση επιτυγχάνεται στα περισσότερα ψάρια
δια µέσου ισχυρών κυµατοειδών κάµψεων του οπίσθιου µέρους του σώµατος,
προκαλώντας το ουραίο πτερύγιο να κτυπάει δυνατά µέσα στο νερό. Το ραχιαίο και
το εδρικό πτερύγιο λειτουργούν αρχικά ως πηδάλια και τα ζυγά πτερύγια λειτουργούν
ως όργανα ισορροπίας και φρεναρίσµατος. Το βαρύ εµπρόσθιο άκρο ενεργεί ως
υποµόχλιο για την κίνηση της ουράς, η οποία ενεργεί µε την σειρά της ως εύκαµπτος
µοχλός.
Παρατεταµένη κολύµβηση σπάνια είναι µεγαλύτερη από 32 χλµ ανά ώρα, αν και
παρατηρούνται µερικές σύντοµες εξάρσεις σε ορισµένες περιπτώσεις που ξεπερνούν
αυτό το όριο. Πάντως η πλειονότητα των ψαριών κολυµπούν µε σηµαντικά
µικρότερη ταχύτητα από την προαναφερθείσα. Ειδικότερα η µέγιστη ταχύτητα στα
µικρά ψάρια µπορεί να φθάσει 10 φορές το µήκος του σώµατoς ανά δευτερόλεπτο
για µικρές εξάρσεις. Ψάρια που κολυµπούν ταχέως έχουν υδροδυναµικό σχήµα
σώµατος. Έχει βρεθεί ότι οι κινήσεις κολύµβησης είναι τέτοιες, ώστε το κενό που
προκαλείται από το στροβιλισµό πίσω από ένα ψάρι που κολυµπάει είναι µικρότερο
από εκείνο του µοντέλου του ψαριού, το οποίο πειραµατικά προωθείται ή σύρεται
δια µέσου του νερού. Έτσι η ενέργεια που χρειάζεται για την κολύµβηση είναι
λιγότερη από εκείνη, που µπορεί κανείς να υποθέσει από τη µελέτη τέτοιων
µοντέλων.
Από όλα τα µεγάλα ψάρια, ο τόνος και τα συγγενικά του είδη είναι µοναδικά
ειδικευµένα για ταχεία κολύµβηση σε µεγάλες αποστάσεις. Το σώµα τους είναι
κοµψά υδροδυναµικό. Μόνα µεταξύ των ψαριών έχουν κόκκινη σάρκα, σηµάδι
µεγάλης µυϊκής ενέργειας. Κατά µήκος κάθε πλευράς υπάρχει µια ζώνη απόχρωσης
σκούρου-µαύρου, µε ένα πλούσιο σε αγγεία και µυοσφαιρίνη µυϊκό σύστηµα. Αυτή η

88
περιοχή διατηρείται σε θερµοκρασία από 3ο µέχρι 12οC πάνω από εκείνη του
περιβάλλοντος νερού και µεγαλύτερη από τη θερµοκρασία του λοιπού σώµατος.
Σε πελώρια πειραµατικά ενυδρεία, ο τόνος έχει µετρηθεί να έχει ταχύτητα πορείας
230 χλµ/ηµέρα. Μαρκαρισµένα άτοµα έχουν συλληφθεί κοντά στο Bergen της
Νορβηγίας, σε λιγότερο χρόνο από 4 µήνες, από τη στιγµή που αφέθηκαν ελεύθερα
µετά το µαρκάρισµα στις Μπαχάµες, αποδεικνύοντας µέση ταχύτητα πορείας από 65
µέχρι 162 χλµ/ηµέρα.
∆εδοµένου ότι ο τόνος δεν προστατεύεται από το χάσιµο θερµότητας από στρώµατα
λίπους κάτω από το δέρµα, όπως συµβαίνει σε µερικά θαλάσσια θηλαστικά και
πτηνά, η διατήρηση θερµοκρασίας µεγαλύτερης από εκείνη του περιβάλλοντος νερού
και η έντονα µυώδης δραστηριότητα συνεπάγονται µεγάλο κόστος ενέργειας και
απαιτούν υψηλό µεταβολικό ρυθµό. Νεαρά άτοµα τόνου πρέπει να τρώνε τόσο πολύ
όσο το 1/4 του βάρους τους για την καθηµερινή κανονική ανάπτυξή τους.
Φαίνεται ότι οι τόνοι καταλαµβάνουν ένα πολύ ειδικευµένο οικολογικό θώκο στο
θαλάσσιο οικοσύστηµα. Αναγκάζονται να µετακινούνται ευρύτατα κυνηγώντας
κοπάδια άλλων ψαριών για να συντηρήσουν τον υψηλό µεταβολικό ρυθµό τους.
4.2.3. Πλεύση και διατήρηση της στάθµης
Σε όλα τα ψάρια, εκτός από τα βενθοπελαγικά τα οποία προτιµούν να ζουν πάνω ή
κοντά στο βυθό, το πρόβληµα της πλεύσης είναι περισσότερο οξύ από όσο είναι στα
περισσότερα από τα ασπόνδυλα και τους πλαγκτικούς οργανισµούς. Υπάρχει µια
πολύ µικρότερη αναλογία επιφάνειας προς τον όγκο του σώµατος, ενώ τα
απαραίτητα για αποτελεσµατική κολύµβηση συστήµατα του µυϊκού ιστού και του
σκελετού κάνουν το ψάρι αναλογικά πυκνότερο. Οι περισσότεροι τελεόστεοι µε
βαρεία σκελετικά και µυϊκά συστήµατα εξουδετερώνουν την πυκνότητά τους µε µια
κύστη αέρα (νηκτική κύστη), η οποία, όπως οι πνεύµονες των ανώτερων ζώων, έχει
προέλθει από µία υπερανάπτυξη του οισοφάγου. Το γεγονός ότι µόνο οι τελεόστεοι
έχουν κύστη αερίου υποστηρίζει κάπως την υπόθεση ότι τόσο οι πνεύµονες όσο και
η νηκτική κύστη πιθανόν να προήλθαν από ένα απλό εκκόλπωµα του οισοφάγου, το
οποίο αρχικά είχε αναπνευστική λειτουργία. Το εκκόλπωµα πιθανόν υπήρξε σε
ψάρια που ζούσαν σε γλυκά νερά, όπου χαµηλές συγκεντρώσεις οξυγόνου είναι
περισσότερο συχνά συναντώµενες και οι ελλείψεις οξυγόνου οδήγησαν στην ανάγκη
αναπλήρωσης µε την κατάποση αέρα. Στους τελεόστεους η νηκτική κύστη βρίσκεται
ραχιαία πάνω από το πεπτικό σύστηµα και λειτουργεί ως υδροστατικό όργανο. Το
αέριο της νηκτικής κύστης είναι ένα µίγµα αερίων διαφορετικό από εκείνο που
βρίσκεται στο νερό και στην ατµόσφαιρα και λαµβάνεται από αδένα που εκκρίνει

89
αέριο (φυσόκλειστοι ιχθείς). Σε άλλη κατηγορία ψαριών (φυσόστοµοι ιχθείς) η
πλήρωση της νηκτικής κύστης µε αέρα όπως και η αποβολή του αέρα γίνεται δια
µέσου του πεπτικού συστήµατος.
Ένα ψάρι µπορεί να ελέγχει το βάθος του στο νερό, ρυθµίζοντας τον όγκο του αέρα
στη νηκτική του κύστη. Ο έλεγχος δεν είναι µια εθελοντική µυϊκή κίνηση αλλά µάλλον
µια µικρής ταχύτητας διαδικασία έκκρισης ή απορρόφησης αερίου µέσα στη νηκτική
κύστη. Το ψάρι µετά ισορροπεί την ολική του πυκνότητα για να την ταυτίσει µε εκείνη
του νερού στο επίπεδο εκείνο στο οποίο ζει. ∆υνατοί κολυµβητές θηρευτές, όπως το
σκουµπρί, µερικοί τόνοι και καρχαρίες, οι οποίοι κολυµπούν σε διάφορα επίπεδα
καταδιώκοντας τη λεία τους, και συχνά πολύ γρήγορα διασχίζουν ένα σηµαντικό
εύρος βαθών, δεν έχουν νηκτική κύστη. Αυτά τα είδη ψαριών θυσιάζουν την
πλευστότητα για την ταχύτητα και τη δυνατότητα να ανεβοκατεβαίνουν. Τα ψάρια
αυτά πρέπει να κολυµπούν για να διατηρούν την επιθυµητή στάθµη µέσα στο νερό.
Βενθικά ψάρια µε νηκτική κύστη, ερχόµενα στην επιφάνεια, κατά την αλιεία τους από
µηχανότρατες, είναι συνήθως νεκρά όταν φθάσουν στην επιφάνεια µε την νηκτική
τους κύστη εκτεταµένη και µερικές φορές βγαλµένη έξω από το στόµα. Οι
αισθητήρες πιέσεως, που συνδέονται µε τη νηκτική κύστη, θεωρούνται επίσης ότι
παίζουν ρόλο στην αντίληψη αλλαγών της υδροστατικής πίεσης σε ψάρια που
διαβιούν σε ενδιάµεσες στάθµες νερού. Πάντως, πολλά είδη ψαριών τα οποία δεν
έχουν νηκτική κύστη ή άλλες συµπιεζόµενες κύστεις παρουσιάζουν επίσης
καθοριστικές αντιδράσεις σε αλλαγές της υδροστατικής πίεσης.
Οι Morris και Kittleman (1967), κατά τη διάρκεια δύο καλοκαιριών καταδυόµενοι και
πιάνοντας ψάρια µε παγίδες στην Ανταρκτική, βρήκαν ότι υπάρχει µια καλώς
καθορισµένη στρωµατοποίηση κατά βάθος πολλών ειδών ψαριών, χωρίς κανένα απ'
αυτά να έχει νηκτική κύστη. Εξάλλου παρατήρησαν ότι σ' εκείνα τα νερά δεν υπάρχει
κλίση θερµοκρασιών η οποία να βοηθά στη διατήρηση των ανωτέρω ειδών σε
δεδοµένες στάθµες βάθους. Η θερµοκρασία ολόκληρης της στήλης του νερού στην
ανωτέρω περιοχή έχει µια οµοιόµορφη κατανοµή γύρω στους -1,90C .
4.2.4. Οσφρητικές και χηµικές αισθήσεις
Σε πολλά ψάρια οι οσφρητικοί λοβοί είναι το κυρίαρχο µέρος του εγκεφάλου, ενώ
όργανα χηµικής αίσθησης δεν είναι µόνο συγκεντρωµένα στη ρινική περιοχή αλλά
απλώνονται πάνω στο κεφάλι και στο σώµα ως µια σύνθετη σειρά οργάνων
πλευρικής γραµµής. Η χηµική ή η οσφρητική αίσθηση χρησιµοποιείται όχι µόνο για
να γίνεται αντιληπτή η τροφή σε πολύ µεγαλύτερες αποστάσεις από όσο τα ψάρια

90
µπορούν να δουν στο νερό - ειδικότερα σε σκοτεινά ή κάπως στροβιλώδη νερά -
αλλά επίσης για επικοινωνία.
H σεξουαλική αναγνώριση, η ρύθµιση της σεξουαλικής συµπεριφοράς και η
συµπεριφορά κυριότητας του χώρου καθώς και η διατήρηση του κοινωνικού status
σε κοινωνίες µε εµφανείς ιεραρχίες έχει αποδειχθεί ότι σε µερικά ψάρια επιτυγχάνεται
µε ειδικές ορµόνες (φεροµόνες). Τέτοιες χηµικές ουσίες καθώς και οι υφιστάµενες
χηµικές αισθήσεις, είναι γνωστό ότι αποτελούν σπουδαία γνωρίσµατα σε πολλές
άλλες οµάδες ζώων, όπως τα καρκινοειδή και τα έντοµα.
4.2.5 ∆ιατροφή - Συνήθειες διατροφής
Ο παράγοντας που πιθανόν επηρεάζει περισσότερο έντονα την εξελικτική
διαφοροποίηση των ψαριών είναι οι συνήθειες διατροφής. Πάνω σ' αυτή τη βάση τα
ψάρια µπορεί να διαιρεθούν χονδρικά σε πλαγκτοφάγα, νηκτοφάγα και βενθοφάγα.
Οι γραµµές διαχωρισµού µεταξύ αυτών των κατηγοριών και ειδικότερα µεταξύ των
δύο τελευταίων δεν είναι σαφείς. Είναι γνωστόν επίσης ότι για τα περισσότερα
ψάρια, οι συνήθειες διατροφής τους µεταβάλλονται ριζικά κατά τη διάρκεια της
ανάπτυξης του νεαρού ατόµου µέχρι να φθάσουν στο στάδιο της ωριµότητας. Τα
νεαρά άτοµα των περισσοτέρων ψαριών τρέφονται µε µικρούς οργανισµούς
(ζωοπλαγκτόν), ή/και κοµµάτια πελαγικών ή βενθικών οργανισµών παρόµοιου
µεγέθους. Καθώς µεγαλώνουν τα µεγαλύτερα αυτά άτοµα αρχίζουν να λαµβάνουν
ως τροφή µεγαλύτερους οργανισµούς και τελικά οποιαδήποτε τροφή είναι
χαρακτηριστική για τα ενήλικα.
Τα πλαγκτοφάγα ψάρια, καθώς και άλλα ψάρια τα οποία τρέφονται µε πολύ
µικροσκοπικά ζώα έχουν γενικά καλά ανεπτυγµένες βραγχιακές άκανθες σε ένα ή
περισσότερα βραγχιακά τόξα επί της εσωτερικής κοίλης πλευράς τους, σε αντίθετη
θέση µε τα βραγχιακά νηµάτια. Καθώς το νερό διέρχεται από το στόµα των ψαριών
και βγαίνει από τις βραγχιακές σχισµές για την αναπνοή, πλαγκτικοί οργανισµοί
ειδικού εύρους µεγεθών φιλτράρονται και συγκρατούνται στο λαιµό από τις
βραγχιακές άκανθες, µέχρι να µεταφερθούν στο άνοιγµα του οισοφάγου και να
καταπωθούν. Το άνοιγµα του οισοφάγου κρατιέται κλειστό κατά τον περισσότερο
χρόνο, εκτός από τη στιγµή κατάποσης της τροφής, έτσι ώστε τα ψάρια αποφεύγουν
να πίνουν πολύ νερό. Σε λίγα µόνο είδη ψαριών οι βραγχιακές άκανθες είναι αρκετά
πολυάριθµες και αρκετά λεπτές ώστε να συγκρατούν αρκετό από το φυτοπλαγκτόν.
Εντούτοις µερικά ψάρια όπως το menhaden (είδος του Ατλαντικού) τρέφονται
απευθείας µε διάτοµα. Η πλειονότητα των πλαγκτοφάγων ψαριών τρέφονται µε
ζωοπλαγκτόν. Η τραχύτητα και ο αριθµός των βραγχιακών ακανθών δίνουν κάποια

91
ένδειξη για την κύρια τροφή τέτοιων ψαριών. Τα ψάρια εκείνα που τρέφονται µε
µακροπλαγκτόν έχουν γενικά λιγότερες και τραχύτερες βραγχιακές άκανθες από
εκείνα τα οποία τρέφονται µε µικρά τεµάχια ζωοπλαγκτού, ενώ εκείνα που τρέφονται
µε φυτοπλαγκτόν έχουν πολύ µεγάλο αριθµό λεπτών βραγχιακών ακανθών. Στο
αντίθετο άκρο της κλίµακας, τα ψάρια που τρέφονται µε άλλα ψάρια έχουν συνήθως
ορισµένες στοιχειώδεις βραγχιακές άκανθες ή δεν έχουν καθόλου. Τα ψάρια που
τρέφονται µε µακροπλαγκτόν αρχίζουν να επιλέγουν και να αρπάζουν άτοµα που
τρέφονται επίσης µε µακροπλαγκτόν. Από το σηµείο αυτό ένα µικρό µόνο βήµα
χωρίζει την κατηγορία αυτή των ψαριών από τους τυπικούς καταβροχθιστές, οι
οποίοι τρέφονται από οργανισµούς των οποίων το µέγεθος συνεχώς αυξάνει. Η
ρέγγα είναι ένα καλό παράδειγµα ενός τυπικού ζωοπλαγκτοφάγου ψαριού τρεφόµενη
κυρίως µε πυκνές µάζες κωπηπόδων, όπως το είδος Calanus finmarchicus. Ακόµη
και κατά το χρόνο πριν ακριβώς από την ωοτοκία, όταν δεν τρέφεται, η ρέγγα τείνει
να παραµένει σε περιοχές πλούσιες σε κωπήποδα παρά σε περιοχές µε πυκνό
φυτοπλαγκτόν.
Αφού το πλαγκτόν είναι αποδεδειγµένα η κύρια πηγή τροφής στους ωκεανούς, τα
πλαγκτοφάγα ψάρια µπορούν να συνθέτουν τη µεγαλύτερη βιοµάζα ψαριών. Τα
ψάρια αυτά αποτελούν την κύρια τροφή όλων των µεγαλύτερων σαρκοφάγων
ψαριών, πουλιών, φωκών κλπ και τη βάση για τις παραγωγικότερες αλιείες.
Τα περισσότερα από τα υπόλοιπα ψάρια είναι ενεργητικά θηρευτές τρεφόµενα µε
άλλα ψάρια ή ασπόνδυλα συγκρινόµενου µεγέθους. Εκείνα που τρώνε άλλα ψάρια ή
καλαµάρια στην ανοικτή θάλασσα είναι γενικά αρκετά µεγάλα, ταχείς κολυµβητές και
συχνά καλά εφοδιασµένα µε κοφτερά ή βελονοειδή δόντια. Εκείνα που ζουν σε
µεσοπελαγικά ή βενθοπελαγικά βάθη τείνουν να είναι µικρότερα από εκείνα των
επιπελαγικών υδάτων και έχουν σχετικά µεγαλύτερα κεφάλια και στόµατα, καθώς και
πιο εξογκωµένα δόντια, πού δίνουν σ' αυτά τα ψάρια µια άγρια µορφή δράκου. Τα
ψάρια θηρευτές καταπίνουν τη λεία τους ολόκληρη ή αν συλλάβουν κάποια λεία πολύ
µεγάλη για να την καταπιούν, τότε την ξεσχίζουν σε µικρότερα κοµµάτια, τα οποία
καταπίνουν ολόκληρα. Πολλά απ' αυτά έχουν ισχυρή την αίσθηση της όσφρησης και
µπορούν να αντιληφθούν την παρουσία κάποιας κατάλληλης τροφής στο νερό πολύ
µακρύτερα από την απόσταση από την οποία µπορούν να ιδούν την εν λόγω τροφή.
Μερικά επίσης φαίνεται ότι προσελκύονται από την αναταραχή στο νερό, όπως αυτή
που δηµιουργείται από την κολύµβηση ενός κοπαδιού ψαριών ή έναν άνθρωπο
κολυµβητή.
Σε βραχώδεις περιοχές όπου υπάρχουν κατάλληλες θέσεις απόκρυψης µεταξύ των
βράχων ή µέσα στα χόρτα ή πάνω στον πυθµένα, πολυάριθµα είδη ψαριών έχουν

92
υιοθετήσει τη συνήθεια της ενέδρας, κρυπτόµενα µέχρι τη στιγµή που κάποιο
κατάλληλο θύµα έλθει µέσα στην περιοχή που ελέγχει ο θηρευτής. Τότε εκτινάσσεται
προς αυτό για να το αρπάξει. Τα ψάρια αυτής της κατηγορίας έχουν συνήθως
µεγάλα κεφάλια και στόµατα και συγχρόνως το στόµα και τα µάτια τους
κατευθύνονται προς τα πάνω.
Τα ψάρια που κατοικούν πάνω στον πυθµένα, όπως τα διάφορα πλατύψαρα, οι
ράγιες και πολυάριθµα είδη της Οικογ. Scorpaenidae είναι συνήθως νωθρά. Έχουν
χαµηλότερους ρυθµούς µεταβολισµού, χαµηλότερους βαθµούς κατανάλωσης
οξυγόνου και µικρότερες αναλογίες επιφάνειας βραγχίων προς το βάρος του
σώµατος από όσο έχουν οι περισσότερο δραστήριοι κολυµβητές. Σε άλλες
περιπτώσεις, όπως στην περίπτωση του µπακαλιάρου (cod), ο οποίος συνήθως
τρέφεται µε µαλάκια και άλλα βενθικά ασπόνδυλα, το συνηθέστερο είναι να έχουν το
στόµα και τα συνδεόµενα µ' αυτό όργανα αίσθησης (µύτη, µουστάκια κλπ)
κατευθυνόµενα κατευθείαν µπροστά ή ακόµη προς την κοιλιακή περιοχή του
σώµατος, όπως στις ράγιες. Τα ψάρια πού τρώνε µαλάκια και καβούρια είναι
συνηθέστερο να έχουν µάλλον χονδρά και πλατειά δόντια για να σπάζουν τα
κοχύλια, σε αντίθεση µε άλλα είδη ψαριών τα οποία έχουν δόντια οξέα σαν βελόνες
για να τρώνε ψάρια. Ειδικού ενδιαφέροντος είναι επίσης µερικά πλατύψαρα, όπως οι
γλώσσες, στις οποίες οι σιαγόνες της πλευράς που βρίσκονται τα µάτια είναι χωρίς
δόντια.
4.2.6 Κοπάδιασµα ψαριών
Τα περισσότερα από τα άφθονα ψάρια, τα οποία τρέφονται µε πλαγκτόν ή µε
ποικιλία µικρών ασπόνδυλων και µικρών ψαριών, σε λιγότερο ή περισσότερο
ανοικτές θάλασσες, παρουσιάζουν το φαινόµενο του κοπαδιάσµατος. Κατά τη
διάρκεια της ηµέρας τα ψάρια αυτά τείνουν να κολυµπούν σε σχετικά πυκνές οµάδες.
Tα ψάρια µέσα στην οµάδα είναι όλα σχεδόν του ίδιου µεγέθους και κατανέµονται σε
ίσες περίπου αποστάσεις µεταξύ τους. Τέτοια κοπάδια συχνά δρουν σχεδόν σαν
σύνολο ατόµων που κολυµπούν και κατευθύνονται όλα µαζί. Άλλα είδη, µερικές
φορές, κοπαδιάζουν για µικρό χρονικό διάστηµα πριν από την ωοτοκία και
µεταναστεύουν στα πεδία ωοτοκίας ως κοπάδια παρά το γεγονός ότι τα ψάρια αυτά
δεν παραµένουν τόσο οργανωµένα κατά την περίοδο της διατροφής τους. Μερικά
τέτοια κοπάδια µπορεί να φθάνουν σε εκατοντάδες χιλιάδες ή εκατοµµύρια ατόµων.
Στη Βόρεια θάλασσα έχουν παρατηρηθεί κοπάδια από ρέγγες 15 έως 17 χιλιοµέτρων
µήκους και 5 χιλιοµέτρων πλάτους.

93
Τα ψάρια που βρίσκονται σε µεγάλα κοπάδια δεν παραµένουν στις ίδιες σχετικές
θέσεις αλλά συνεχώς µετατοπίζονται. Το κοπάδι δεν έχει καθορισµένους ηγήτορες.
Αν το κοπάδι στραφεί ξαφνικά, τα ψάρια που βρίσκονται στη µια πλευρά, απότοµα
βρίσκονται µπροστά και εκείνα που ήταν µπροστά βρίσκονται στην πλευρά. Ψάρια
από τις άκρες του κοπαδιού τείνουν προς το κέντρο αφήνοντας άλλα να εκτίθενται
στις άκρες.
Τα ψάρια που κοπαδιάζουν έχουν ευρυγώνια µάτια µε πλατύ οπτικό πεδίο,
χαρακτηριστικό που βοηθάει το κοπάδι να βρίσκεται συγκεντρωµένο. Κάθε ψάρι
εύκολα µπορεί να κρατάει θέση κοντά σε άλλο ψάρι µπροστά ή πίσω απ' αυτό µέσα
στο οπτικό πεδίο του. Με τη δυνατότητα αυτή των ψαριών που κοπαδιάζουν, να
βλέπουν σε ένα ευρύ σχετικά πεδίο, είναι δύσκολο για τον θηρευτή να τα πλησιάσει
χωρίς να τον αντιληφθούν.
∆εδοµένου ότι στα ψάρια αυτά, τα προς τα εµπρός πτερύγια είναι συνήθως σχετικά
σταθερά και ακίνητα, δεν µπορούν να σταµατούν, να κάνουν προς τα πίσω ή να
περνούν πάνω από τα άλλα, όπως τα µεµονωµένα ψάρια του πυθµένα µε κινητά
εµπρόσθια πτερύγια, αλλά πρέπει να κολυµπούν συνεχώς. Mπορούν όµως να
εκτρέπονται ή να αλλάζουν πορεία. Στα πραγµατικά κοπάδια ο χώρος κολύµβησης
είναι αρκετός, δεν υπάρχει όµως χώρος µεταξύ τους για στροφή. Ένα ψάρι µέσα σε
κοπάδι δε µπορεί µόνο του να στρέψει γύρω απ' αυτό. χωρίς να συγκρουσθεί µε
άλλο ψάρι που κολυµπάει µαζί του.
Το κοπάδιασµα παρέχει πιθανώς προστασία στα άτοµα των ψαριών που το
αποτελούν, τα οποία θα ήταν στο έλεος των µεγαλύτερων θηρευτών αν βρίσκονταν
µόνα τους στην ανοικτή θάλασσα. Τα ψάρια και ειδικότερα εκείνα που βρίσκονται
στο κεντρικό τµήµα του κοπαδιού µπορεί επίσης να προκαλούν σύγχιση στον
θηρευτή, µετακινούµενα µε ορµή κατά τρόπο που κάνει δύσκολο στον θηρευτή να
διαλέξει και να συλλάβει ένα συγκεκριµένο άτοµο. Η ικανότητα των µελών ενός
κοπαδιού να κάνουν ξαφνικές εξορµήσεις ταχύτατης κολύµβησης φαίνεται
περισσότερο σηµαντική για να ξεφεύγουν τους θηρευτές, παρά για τη διατήρηση της
ταχύτητας αυτής καθαυτής .Οι περισσότεροι θηρευτές είναι οπωσδήποτε
µεγαλύτεροι και ταχύτεροι έτσι ώστε η διατήρηση της ταχύτητας δε θα βοηθούσε
πολύ τα µέλη του κοπαδιού να διαφύγουν. Μερικά ψάρια επιτυγχάνουν µέγιστη
ταχύτητα µέσα σε 1/20 του δευτερολέπτου σε απάντηση ενός κατάλληλου
ερεθίσµατος.
Όταν ψάρια που αποτελούν λεία θηρευτών είναι συγκεντρωµένα σε µεγάλα κοπάδια,
υπάρχει µικρότερη ευκαιρία για έναν θηρευτή να τα βρει στο κοπάδι απ' όσο θα

94
εύρισκε µερικά µεµονωµένα άτοµα του είδους της λείας εάν αυτά ήταν
διασκορπισµένα ευρέως στην ανοικτή θάλασσα. Αρκετά παράδοξα, η ικανότητα του
θηρευτή να καταβροχθίσει όλη τη λεία πού θέλει όταν συναντήσει ένα κοπάδι µπορεί
µακροπρόθεσµα να συµβάλλει επίσης στην ασφάλεια της λείας. Ειδικότερα, ο
θηρευτής µπορεί να φάει µόνον µερικά ψάρια σε µια επιδροµή του για διατροφή. Η
πιθανότητα ενός ψαριού σ' ένα κοπάδι να φαγωθεί σε µια συνάντηση ενός θηρευτή
µε το κοπάδι είναι 1 στα χ/α, όπου α = ο αριθµός των ατόµων της λείας που µπορεί
να φάει ένας θηρευτής και χ = ο αριθµός των ατόµων ενός κοπαδιού. Όσο
µεγαλύτερο είναι το κοπάδι τόσο µικρότερη η πιθανότητα για ένα άτοµο του κοπαδιού
να φαγωθεί, σε µια συγκεκριµένη συνάντηση µε έναν θηρευτή. Αν αντίθετα, ο
θηρευτής συναντάει άτοµα της λείας αρκετά κανονικά, σε µεγάλα διαλείµµατα χώρου,
ο θηρευτής αυτός ποτέ δε θα είναι χορτάτος και έτσι κάθε άτοµο λείας που θα
συναντάει θα το καταβροχθίζει. Είναι επίσης αλήθεια ότι µεγάλα κοπάδια ψαριών
προσελκύουν περισσότερους θηρευτές στην περιοχή, από όσους θα ήταν εκεί και
έτσι γίνεται ευκολότερο, τουλάχιστον για µερικούς θηρευτές, να λαµβάνουν ένα
πλήρες γεύµα, αλλά η ισορροπία του πλεονεκτήµατος βρίσκεται προς το µέρος του
κοπαδιού.
Αρκετά είδη µικρών πελαγικών ψαριών που κοπαδιάζουν, όπως η σαρδέλα, ο
γαύρος κλπ, έχουν λέπια που φεύγουν εύκολα, γεγονός που δίνει τη δυνατότητα στα
ψάρια αυτά να διαφεύγουν ελαφρώς καλύτερα όταν συλλαµβάνονται από έναν
θηρευτή.
Η παρουσία του ανθρώπου στη σκηνή µε αποτελεσµατικά αλιευτικά εργαλεία για να
κάνει χρήση του πλεονεκτήµατος του κοπαδιάσµατος των ψαριών, µπορεί εύκολα να
µετατοπίσει την ισορροπία του πλεονεκτήµατος και να επιφέρει καταστροφή των
ιχθυοπληθυσµών σε ορισµένα είδη, όπως προφανώς συνέβη στην περίπτωση της
σαρδέλας της Καλιφόρνια αµέσως µετά τον 2ο παγκόσµιο πόλεµο. Είναι πιθανόν
ένα τέτοιο γεγονός να είναι ακόµη χειρότερο για τους µεγαλύτερους θηρευτές, παρά
για τη λεία που κοπαδιάζει, γιατί ο άνθρωπος επίσης αλιεύει εντατικά τους µεγάλους
θηρευτές, οι οποίοι είναι πολύ λιγότεροι σε αριθµό και επειδή η τροφή τους σε µια
περιοχή µπορεί να µειωθεί κάτω από το επίπεδο στο οποίο ο άνθρωπος ή οι µεγάλοι
θηρευτές θα µπορούσαν αποτελεσµατικά να αλιεύσουν ή να τραφούν αντίστοιχα. Σε
µια τέτοια περίπτωση οι µεγάλοι θηρευτές µπορεί να εξαφανισθούν τελείως, ενώ τα
αποµείναντα ίχνη των πληθυσµών που τρέφονται µε πλαγκτόν, θα επιβιώσουν, και
έχοντας τώρα µικρότερη πίεση από τους θηρευτές είναι δυνατόν να δώσουν
πληθυσµούς µικρών πελαγικών ψαριών σε µεγαλύτερο αριθµό από όσο πριν από
την προαναφερθείσα καταστροφή τους. ∆εδοµένου ότι τα περισσότερα από τα είδη

95
αυτά παράγουν τροµερά µεγάλους αριθµούς αυγών και ωριµάζουν γρηγορότερα από
τα µεγάλα ψάρια, η ανασυγκρότηση του πληθυσµού µπορεί να γίνει σχετικά
γρηγορότερα σε µια δεδοµένη ευνοϊκή ευκαιρία.
Μια πολύ πιο δυσοίωνη πιθανότητα που αντιµετωπίζουν πολλά είδη ψαριών είναι η
καταστροφή των πεδίων ωοτοκίας τους, ειδικότερα όταν αυτά τα πεδία βρίσκονται σε
παράκτια ύδατα και εκβολές ποταµών που υπόκεινται σε µεταβολές και σε ρύπανση.
Το κοπάδιασµα των µεγάλων µεταναστευτικών ειδών, όπως ο τόνος, µπορεί
πρωτίστως να εξυπηρετεί, τη διατήρηση των ψαριών µαζί, για λόγους
αναπαραγωγής, παρά να εξυπηρετεί την ανάγκη προστασίας από µεγαλύτερους
θηρευτές. Αυτή η λειτουργία είναι πλέον των πλεονεκτηµάτων που προαναφέρθηκαν
για τα µικρά ψάρια.
4.2.7 Αναπαραγωγή
Κατά κανόνα τα φύλα στα ψάρια είναι χωρισµένα. Υπάρχουν όµως και αρκετές
περιπτώσεις ψαριών που τα αρσενικά και θηλυκά όργανα αναπαραγωγής βρίσκονται
συγχρόνως ή αναπτύσσονται διαδοχικά στο ίδιο άτοµο. ∆ιαφορετικά φύλα
ευρισκόµενα σε διαφορετικά άτοµα εξαλείφουν οποιαδήποτε πιθανότητα
αυτογονιµοποίησης και της συνεπαγόµενης µείωσης γενετικής διαφοροποίησης.
Ακόµη και στα ερµαφρόδιτα ψάρια, όπως είναι τα Serranus και Serranelus, τα οποία
πράγµατι γονιµοποιούν τα δικά τους αυγά όταν είναι µόνα τους, η συχνότητα
συνάντησης διαφορετικών ατόµων είναι ο κανόνας. Μερικά ψάρια είναι ερµαφρόδιτα,
όχι όµως όπως τα Serranus. Στα ψάρια αυτά ο ερµαφροδιτισµός είναι διαδοχικός,
δηλ. η όλη γονάδα στην αρχή είναι ενός φύλου και στη συνέχεια αλλάζει στο αντίθετο
φύλο, όπως συµβαίνει στο γνωστό λυθρίνι και άλλα Σπαροειδή.
Στους τελεόστεους η γονιµοποίηση είναι εξωτερική, όπου το αρσενικό γονιµοποιεί τα
αυγά τη στιγµή που αποτίθενται από το θηλυκό. Τα ψάρια αυτά λέγονται ωοτόκα.
Υπάρχουν όµως και αρκετές περιπτώσεις ψαριών (πολλοί Σελάχιοι και ελάχιστοι
Τελεόστεοι), στα οποία η γονιµοποίηση είναι εσωτερική. Στις περιπτώσεις αυτές τα
έµβρυα αναπτύσσονται µέσα στο σώµα του θηλυκού ατόµου, το οποίο στη συνέχεια
γεννά ζωντανά άτοµα µε αρκετά πλούσιο όµως λεκιθικό σάκκο. Τα ψάρια αυτά
λέγονται ωοζωοτόκα. Σε Σελάχιους όπου η γονιµοποίηση είναι εσωτερική - στις
περιπτώσεις που το σπέρµα µεταφέρεται µε τη βοήθεια των πτερυγιοπόδων στην
υποδοχή του θηλυκού- αποτίθενται είτε αυγά γονιµοποιηµένα ήδη και µε το έµβρυο
σε ανάπτυξη ή γεννούν ζωντανά άτοµα, που συµπληρώνουν την ανάπτυξή τους
µέσα στο σώµα της µητέρας τους (ζωοτόκα).

96
Σε άλλα είδη Σελαχίων (Whale shark), τα γονιµοποιηµένα αυγά περιβάλλονται από
κερατοειδές ορθογώνιο κέλυφος, πλούσιο σε λέκιθο, εντός του οποίου προστατεύεται
το έµβρυο µέχρι την εκκόλαψή του. Το κέλυφος αυτό αφήνεται από το θηλυκό άτοµο
στον πυθµένα ή στερεώνεται σε φύκια και κοράλλια µε τη βοήθεια νηµατοειδών
προεκτάσεων, που βρίσκονται στις γωνιές του κελύφους και εµπλέκονται πάνω στο
υποστήριγµα όπως οι έλικες της κληµαταριάς. Όταν τα αυγά αυτά εκκολαφθούν,
µερικές φορές ύστερα από αρκετούς µήνες, τα νεαρά ψάρια είναι πολύ καλά
αναπτυγµένα και αρκετά ικανά για να επιβιώσουν. Τα ψάρια αυτά λέγονται ωο-
ωοζωοτόκα.
Στους τελεόστεους ο απλούστερος τρόπος αναπαραγωγής είναι η συγκέντρωση των
ατόµων των δύο φύλων σε κοπάδια και η ταυτόχρονη αποβολή αυγών και
σπέρµατος που τα γονιµοποιεί, όπως π.χ. οι σαρδέλες, ο γαύρος κ.α. πελαγικά
ψάρια τα οποία κοπαδιάζουν. Υπάρχουν όµως πολλές περιπτώσεις ψαριών των
οποίων η συµπεριφορά της αναπαραγωγής είναι περισσότερο εξελιγµένη. Στα είδη
του γένους Μugil (κέφαλοι) π.χ. το κάθε θηλυκό άτοµο περιστοιχίζεται από µερικά
αρσενικά ή σε άλλα είδη (Gasterosteus) σχηµατίζουν ζευγάρια. Σε πολλές
περιπτώσεις τα αρσενικά άτοµα κατά την εποχή της αναπαραγωγής αποκτούν
λαµπερούς χρωµατισµούς (µαρίδα, χέλια κλπ), ενώ σε άλλα παρατηρούνται αληθινές
γαµήλιες τελετές µε µονοµαχίες των αρσενικών για το θηλυκό άτοµο. Το µέγεθος
των αυγών ποικίλει σηµαντικά από είδος σε είδος. Έτσι τα ώριµα αυγά έχουν
διαστάσεις από κλάσµα του χλστµ, σε µερικά κλουπεοειδή και πλατύψαρα, µέχρι 80
χλστµ. στους περισσότερους καρχαρίες και χίµαιρες. Ένα αυγό του ψαριού whale
shark, το µεγαλύτερο σε µέγεθος από την κλάση των ψαριών, έχει συχνά µήκος 30
εκστµ. Στα ευρωπαϊκά νερά, µεγάλα αυγά αποθέτουν οι σολοµοί, οι πέστροφες και
οι σµέρνες, των οποίων τα αυγά ξεπερνούν τα 4-5 χλστµ. Γενικά όµως η διάµετρος
των αυγών των περισσοτέρων ειδών κυµαίνεται γύρω στα 1-2 χλστµ.
Τα αυγά πολλών ειδών ψαριών της θάλασσας είναι ελαφρά και επιπλέουν στο νερό.
Είναι η περίπτωση των πελαγικών αυγών που ανήκουν στο πλαγκτόν, όπως και οι
προνύµφες από τα αυγά αυτά. Τα πελαγικά αυγά δεν τα φροντίζουν καθόλου οι
γονείς τους και για το λόγο αυτό υπόκεινται σε τροµερές απώλειες. Όµως η φυσική
αυτή απώλεια αντισταθµίζεται από την εξαιρετικά υψηλή γονιµότητα των γεννητόρων
τους.
Όλα τα αυγά δεν είναι πελαγικά. Πολλά είδη ψαριών αποθέτουν τα αυγά τους στον
πυθµένα, είτε ελεύθερα στην άµµο, τις πέτρες, τα χαλίκια, ή κολληµένα στα φύκια.
Πολλά πελαγικά ψάρια, όπως οι ρέγγες, οι σαρδελοµάνες (Αlosa sp.), oι σολοµοί κ.α.
ακολουθούν αυτόν τον τρόπο απόθεσης των αυγών τους. Τα αυγά αυτών των ειδών

97
ψαριών µόλις τοποθετηθούν αφήνονται στην τύχη τους χωρίς καµιά επίβλεψη.
Πολλές φορές µάλιστα έχει παρατηρηθεί ότι πολλά άλλα ψάρια ακολουθούν τους
γεννήτορες ή και οι ίδιοι οι γεννήτορες τρώνε τα αυγά που αποτίθενται. Υπάρχουν
όµως και αρκετές περιπτώσεις ψαριών, όπως πολλοί γωβιοί, σαλιάρες κ.α. που τα
αρσενικά άτοµα επιβλέπουν τα αυγά τους µέχρι την εκκόλαψη, διώχνοντας τους
εχθρούς που θα ήθελαν να τα φάνε (καβούρια, αστερίες κλπ). Άλλα πάλι ψάρια
προστατεύουν τα αυγά τους κάνοντας φωλιές (Gasterosteus, Labridae κλπ) όπου και
τα αποθέτουν. Ακόµη πιο χαρακτηριστική είναι η περίπτωση των Sygnathidae
(αλογάκι της θάλασσας) που τα αρσενικά µεταφέρουν τα αυγά µέχρι την εκκόλαψη
µέσα σ' ένα µάρσιπο στην κοιλιά τους. Η απόθεση αυτή συνοδεύεται µερικές φορές
από χαριτωµένες γαµήλιες τελετές. Ύστερα από την εκκόλαψη, σε ορισµένες
περιπτώσεις, τα νεαρά άτοµα µπαινοβγαίνουν στον µάρσιπο. Μερικά άλλα ψάρια,
όπως το γατόψαρο Galeichthys felis, πραγµατοποιούν την εκκόλαψη των αυγών
τους µέσα στη στοµατική κοιλότητα του αρσενικού, όπου τα νεαρά άτοµα µπορούν
να µπαινοβγαίνουν για προστασία. Μέχρι την ανάπτυξη των νεαρών, το αρσενικό
δεν τρέφεται. Το ίδιο φαινόµενο παρατηρείται και σε άλλα ψάρια, όπως στα Cichlidae
(Τilapia) όπου όµως η στοµατική επώαση και η εκκόλαψη γίνεται και στα δύο φύλα.
Τέλος θα πρέπει να αναφερθεί η περίπτωση του Rhodeus sericeus όπου το θηλυκό
φέρει ένα µακρύ ωοαποθετήρα µε τον οποίο αποθέτει τα αυγά του στο εσωτερικό
διθύρων µαλακίων (Αnodonta) όπου και αναπτύσσονται, αφού προηγουµένως
γονιµοποιηθούν από το αρσενικό που τα ραντίζει µε σπέρµα από το άνοιγµα του
σίφωνα.
Η ανάπτυξη του εµβρύου είναι εύκολο να παρατηρηθεί στα αυγά των τελεοστέων
επειδή το περίβληµά τους είναι κατά κανόνα διαφανές. Μετά τη γονιµοποίηση
εµφανίζεται ένας δίσκος πιο διαθλαστικός από το υπόλοιπο του αυγού. Αυτός ο
δίσκος εκτείνεται σιγά-σιγά σαν ένα σκέπασµα και τελικά περιβάλλει ολόκληρη την
επιφάνεια του αυγού. Στη συνέχεια εµφανίζεται ένα στενόµακρο εξόγκωµα σαν
σαµάρι που είναι το έµβρυο. Σ' αυτό εµφανίζονται γρήγορα το κεφάλι µε τα µεγάλα
µάτια, το έσω ους και τα ίχνη των µεταµεριδίων των µυών. Στο µέτρο που το έµβρυο
µεγαλώνει, η µάζα του αυγού ελαττώνεται αφού οι θρεπτικές ουσίες του (λέκιθος)
χρησιµοποιούνται για το σχηµατισµό νέων ιστών. Κατά την εκκόλαψη τα νεαρά άτοµα
είναι διαφανή και εξαιρετικά εύθραυστα. Το κεφάλι τους, ο κορµός τους, η ουρά τους
είναι ευδιάκριτα ενώ στην κοιλιά τους διατηρείται για λίγο µια µάζα λεκίθου, η οποία
δεν έχει απορροφηθεί ακόµη. Τα θωρακικά πτερύγια έχουν ήδη σχηµατισθεί ενώ τα
άζυγα πτερύγια αντιπροσωπεύονται από µια λεπτή συνεχή πτυχή. Σ' αυτό το στάδιο
η χρώση του σώµατος περιορίζεται σε µερικές κηλίδες που σχηµατίζουν ειδικά

98
κύτταρα τα χρωµατοφόρα. Η λέκιθος απορροφάται σύντοµα και τα νεαρά ψαράκια
πρέπει τότε να διατραφούν µόνα τους. Η µορφή που έχει η προνύµφη είναι πολύ
διαφορετική από εκείνη του ενήλικου ατόµου. Ύστερα από ένα χρονικό διάστηµα,
που ποικίλει από µερικές εβδοµάδες (στα περισσότερα είδη) µέχρι 2 χρόνια (στο
χέλι), η προνύµφη υφίσταται αξιοσηµείωτες µεταµορφώσεις, συχνά γρήγορες, που
µετασχηµατίζουν την προνύµφη τελείως και της δίδουν µορφή πολύ κοντινή προς το
ενήλικο. Έτσι από τον λεπτοκέφαλο (στα χέλια) σχηµατίζεται ένα νεαρό χέλι και από
τις προνύµφες της γλώσσας, που έχουν τα µάτια και το στόµα στην αρχή σε κανονική
θέση, για να µεταφερθούν σιγά-σιγά στη δεξιά, σχηµατίζοντας έτσι ένα κανονικό
άτοµο γλώσσας, που έχει την αριστερή της πλευρά µε την οποία στηρίζεται στο βυθό
αχρωµάτιστη και τυφλή.
4.2.8 Γονιµότητα και επιβίωση
Αυξηµένη προσοχή δίδεται στον αριθµό των αυγών που αναπτύσσονται στα θηλυκά
άτοµα και στην επιβίωσή τους καθώς και στα πρώτα στάδια ανάπτυξης µετά την
εκκόλαψη. Τα χαρακτηριστικά αυτά είναι συνδεδεµένα µε τη σταθερότητα ενός
πληθυσµού και τις διακυµάνσεις των ετησίων κλάσεων. Τέτοιες διακυµάνσεις
φτάνουν σε ακραίες αναλογίες σε µερικά είδη, γεγονός που αποτελεί βασικό
παράγοντα ο οποίος καθορίζει τις µεταβολές της παραγωγής από έτος σε έτος. Οι
αριθµοί των αυγών που αποθέτουν τα ψάρια ποικίλουν σηµαντικά, από µερικά
µεγάλα αυγά στους καρχαρίες µέχρι πολλά εκατοµύρια στο φεγγαρόψαρο (Mola
mola). Αλλά και τα ποσοστά επιβίωσης αποτελούν σηµαντικό χαρακτηριστικό των
ψαριών. Στο στουριόνι π.χ. η επιβίωση είναι 0,01%, στο σολοµό του Ατλαντικού από
0,1 µέχρι 0,5% ενώ σε ορισµένα Σπαροειδή (Bream) η επιβίωση είναι ακόµη
µικρότερη (0,006 - 0,020%).
Η γονιµότητα ενός πληθυσµού δυνατόν να υπόκειται σε σηµαντικές διακυµάνσεις ως
συνέπεια του είδους της διατροφής τους. Ένας µικρότερος πληθυσµός ενός είδους,
ο οποίος κατά συνέπεια έχει µεγαλύτερη παροχή τροφής, συνήθως έχει µεγαλύτερη
γονιµότητα από εκείνη που έχει όταν ο πληθυσµός είναι µεγαλύτερος και η τροφή
µοιράζεται σε περισσότερα άτοµα. Έτσι, το είδος του σολοµού Oncorhynchus
gorbuscha στις εκβολές του Αµούρ είναι περισσότερο γόνιµος κατά τις περιόδους
µικρής ανόδου από όσο κατά τις περιόδους µεγαλύτερης ανόδου. Στη Β. Κασπία
έχει παρατηρηθεί ότι η γονιµότητα του είδους "τσιρόνι" ενός συγκεκριµένου µεγέθους
αυξάνεται κατά τις περιόδους καλύτερης παροχής τροφής. Επίσης διαφορές
γονιµότητας συµβαίνουν και σε ψάρια του ίδιου µεγέθους όταν ζουν σε διαφορετικές
συνθήκες. Π.χ. η γονιµότητα του λούτσου µήκους 35-45 εκστµ. στη θάλασσα της
Αράλης είναι κατά µέσο όρο 8.500 αυγά ενώ στη Β. Κασπία θάλασσα 17.500 αυγά.

99
Αύξηση της γονιµότητας στα ψάρια είναι δυνατόν να προκληθεί από διάφορες αιτίες.
Μπορεί να προέλθει από µείωση του αποθέµατος της λεκίθου µε τη συνεπαγόµενη
µείωση της παθητικής περιόδου του εµβρύου, ή από την αύξηση της πυκνότητας της
λεκίθου, ή τελικά από την αύξηση του όγκου της γονάδας. Π.χ. η γονιµότητα του
Huso dauricus (στουριόνι) από τον ποταµό Αµούρ, είναι µεγαλύτερη από εκείνη του
είδους Huso huso στον ίδιο ποταµό, εξαιτίας του µικρότερου όγκου γονάδας, που
συνεπάγεται µικρότερο αριθµό αυγών. Η ατοµική γονιµότητα επίσης αυξάνει µε την
κλασµατική απόθεση αυγών, που συµβαίνει όταν µόνο ένα µέρος των αυγών είναι
ώριµο σε διαδοχικές περιόδους. Η κλασµατική απόθεση αυγών και η παρατεταµένη
περίοδος αναπαραγωγής είναι χαρακτηριστικά κυρίως των τροπικών και
υποτροπικών ειδών. Ο µεγαλύτερος αριθµός ειδών µε κλασµατική απόθεση αυγών
στις τροπικές περιοχές είναι συνέπεια προφανώς του γεγονότος ότι στην τροπική
ζώνη δεν υπάρχει καθαρή εποχιακή προσφορά τροφής, που είναι το πλαγκτόν, µε το
οποίο τρέφονται οι προνύµφες. Αυτό επιτρέπει στις προνύµφες να βρίσκουν τροφή
για µεγάλη χρονική περίοδο.
Γενικά η γονιµότητα των ψαριών είναι πολύ µεγαλύτερη από όλα τα χερσαία
σπονδυλωτά. Τα πιο γόνιµα ψάρια είναι εκείνα που έχουν επιπλέοντα πελαγικά
αυγά. ∆εύτερα στη σειρά είναι εκείνα που αποθέτουν τα αυγά τους στα φυτά, ενώ
εκείνα τα ψάρια που προστατεύουν ή κρύβουν τα αυγά τους έχουν συνήθως
µικρότερη γονιµότητα. Εξάλλου η γονιµότητα των θαλασσινών ψαριών είναι γενικά
µεγαλύτερη από εκείνη των ψαριών του γλυκού νερού.
Η γονιµότητα των ψαριών, όπως και των άλλων ζώων, είναι µια προσαρµογή η
οποία εξασφαλίζει των επιβίωση του είδους κάτω από συνθήκες στις οποίες υπάρχει
ο πληθυσµός. Μεγάλη γονιµότητα συνεπάγεται κάτω από ορισµένες συνθήκες,
υψηλή θνησιµότητα, ειδικά όταν αυτή οφείλεται σε θηρευτές (εκτός της αλιείας).
Επίσης ταχύτερα αναπτυσσόµενα άτοµα έχουν υψηλότερη γονιµότητα από εκείνη
των αναπτυσσοµένων µε βραδύτερο ρυθµό.
4.3. Θαλάσσια θηλαστικά
Υπάρχουν 3 κύριες οµάδες θαλάσσιων θηλαστικών, κάθε µια από τις οποίες
προήλθε ανεξάρτητα από διάφορα θηλαστικά αποθέµατα τα οποία έχουν
προσαρµοσθεί για ζωή στους ωκεανούς. Η ζωή στους ωκεανούς είναι τόσο
διαφορετική από εκείνη στην ξηρά έτσι ώστε έχει εντυπώσει µερικές γενικές τάσεις ή
συγκλίσεις στην εξέλιξη και των τριών οµάδων. Και οι τρεις οµάδες έχουν αποκλίνει

100
σηµαντικά από τα αρχικά αποθέµατα έτσι ώστε δεν είναι πλέον άµεσα εµφανές, από
περιπτώσεις εξέτασης των σηµερινών υπαρχόντων ειδών, να διαπιστωθεί µε ποια
από τις οµάδες των θηλαστικών της στεριάς συγγενεύουν περισσότερο.
Έχοντας βασικά προσαρµοσθεί για τη ζωή στους ωκεανούς και οι τρεις οµάδες έχουν
υποστεί διαφοροποιήσεις και έχουν προσαρµοσθεί στην εκµετάλλευση διαφορετικών
οικολογικών θώκων, που προσφέρονται στο θαλάσσιο περιβάλλον. Κοινοί
χαρακτήρες που βρέθηκαν και στις τρεις οµάδες είναι:
(α) τροποποιηµένα άκρα και σχήµα σώµατος,
(β) θερµική ρύθµιση,
(γ) τροποποιηµένο αναπνευστικό σύστηµα και
(δ) οσµωτικές προσαρµογές.
Οι αλλαγές στα άκρα και στο σχήµα του σώµατος καθιστούν το ζώο ικανό να
προσαρµόζεται σε ζωή κολύµβησης. Αυτές οι αλλαγές περιλαµβάνουν µετατροπή
των άκρων σε πτερύγια, που µοιάζουν µε κουπιά, και σε µερικές περιπτώσεις στην
απώλεια των πίσω άκρων και την ανάπτυξη του πίσω µέρους του σώµατος σε µια
κατασκευή, η οποία µοιάζει µε την ουρά του ψαριού αλλά τοποθετηµένη οριζόντια.
Η θερµική ρύθµιση περιλαµβάνει την πρόβλεψη για τη διατήρηση της θερµοκρασίας
του σώµατος σε ένα περιβάλλον γενικά πολύ ψυχρότερο από τη θερµοκρασία του
σώµατος του ζώου. Αυτό έχει επιτευχθεί µε µια ποικιλία µέσων, στα οποία
συµπεριλαµβάνονται το σκληρό δέρµα µε µονωµένα στρώµατα από γούνα και/ή
λιπώδη ιστό κάτω από το δέρµα, µείωση της περιφερειακής κυκλοφορίας στο δέρµα
και στις προεκτάσεις του σώµατος, µεταβολισµό που να παράγει εσωτερική
θερµότητα και αύξηση του µεγέθους. Ο µεγαλύτερος λόγος της σχέσης του όγκου
προς την επιφάνεια στα µεγάλα ζώα, τα καθιστά ικανά να συγκρατούν τη θερµότητα
περισσότερο αποτελεσµατικά από όσο συµβαίνει στην πλειονότητα των θηλαστικών.
Η διαφοροποίηση του αναπνευστικού συστήµατος καθιστά ικανά τα θαλάσσια
θηλαστικά να αναπνέουν αποτελεσµατικά χωρίς να δέχονται θαλασσινό νερό µέσα
στους πνεύµονες και να συγκρατούν την αναπνοή τους πολύ περισσότερο από τα
περισσότερα θηλαστικά, έτσι τα ζώα αυτά είναι ικανά να πραγµατοποιούν
παρατεταµένες βαθιές καταδύσεις.
Το κράτηµα της αναπνοής στην περίπτωση των βαθιά καταδυοµένων θαλασσίων
θηλαστικών είναι µια διαφορετική διαδικασία από εκείνη του ανθρώπου και άλλων
ζώων της στεριάς.

101
Οι καταδύσεις γίνονται µε τους πνεύµονες άδειους ώστε να αποφεύγεται το δίδυµο
πρόβληµα της κατανάλωσης υπερβολικής ενέργειας για να αντιπαρέλθει την
πλευστότητα, καθώς καταδύεται και της συµπίεσης του αζώτου µέσα στο αίµα στα
µεγάλα βάθη, το οποίο µπορεί να σχηµατίσει φυσαλίδες στο αίµα καθώς η πίεση
µειώνεται κατά τη διάρκεια της ανόδου.
Το αίµα των καταδυοµένων σε βαθιά νερά θηλαστικών είναι πλούσιο σε
αιµοσφαιρίνη ενώ υπάρχει µια επί πλέον αποθήκη αναπνευστικής χρωστικής, η
µυοσφαιρίνη, που βρίσκεται στους µύες και έχει ακόµη µεγαλύτερη συγγένεια µε το
οξυγόνο, δίδοντας στους µύες µια σκούρα κόκκινη, σχεδόν προς το µαύρο
απόχρωση. Η µεταφορά του οξυγόνου από τους πνεύµονες στο αίµα και στους µύες
είναι ταχεία και περισσότερο αποτελεσµατική από εκείνη στα ζώα στης στεριάς. Ο
όγκος του αέρα που αναπνέεται είναι µεγαλύτερος και το σχήµα καθώς και οι
κινήσεις των ζώων είναι τέτοιες ώστε κολυµπούν µε µεγάλη αποτελεσµατικότητα
επιτυγχάνοντας το µέγιστο της κίνησης στην κολύµβηση για την ενέργεια που
χρησιµοποιείται.
Κατά την κατάδυση στα βαθιά νερά, σηµαντική οικονοµία οξυγόνου γίνεται µε τους
σφιγκτήρες στις αρτηρίες που οδηγούν προς τα σπλάχνα και στο δέρµα, έτσι ώστε η
κυκλοφορία προς τα µέρη αυτά να διακόπτεται και όλο το αίµα να πηγαίνει εκεί που
χρειάζεται περισσότερο - στους µύες του σκελετού και στον εγκέφαλο. Ο ρυθµός της
καρδιάς επίσης µειώνεται και οι µύες αυτοί καθαυτοί είναι ικανοί να ανέχονται
µεγαλύτερη έλλειψη οξυγόνου από όση τα ζώα της στεριάς, σε βαθµό ώστε να
λειτουργούν για κάποιο χρόνο αναερόβια. Τα νευρικά κέντρα ελέγχου, τα οποία
τελικά ρυθµίζουν την αναπνοή, δεν είναι ευαίσθητα σε υψηλότερες συγκεντρώσεις
διοξειδίου του άνθρακα και γαλακτικού οξέος στο αίµα, γεγονός που δεν συµβαίνει
στα ζώα της στεριάς. Με την άνοδό του στην επιφάνεια, το ζώο παίρνει έγκαιρα
µεγάλη αναπνοή, πραγµατοποιεί γρήγορη και αποτελεσµατική απαλλαγή από το
διοξείδιο και εκπνέει καθώς αρχίζει άλλη κατάδυση.
Οσµωτικές προσαρµογές προφυλάσσουν το ζώο από την είσοδο υπερβολικής
ποσότητας θαλασσινού νερού, επιτρέποντας τα θαλάσσια θηλαστικά να ζουν χωρίς
την ανάγκη γλυκού νερού. Τα ζώα αυτά δεν κατέχουν ειδικούς αδένας απέκκρισης
αλάτων, όπως οι τελεόστεοι και τα θαλασσινά πουλιά. Τα θαλάσσια θηλαστικά
φαίνεται ότι έχουν πετύχει τη ρύθµιση περιορίζοντας την είσοδο νερού όσο το
δυνατόν περισσότερο -έχοντας υδατοστεγές δέρµα και λαµβάνοντας το περισσότερο
από το νερό που χρειάζονται από την τροφή τους (ψάρια κ.α. ασπόνδυλα)- που
έχουν σωµατικά υγρά περισσότερο αραιωµένα από όσο το θαλασσινό νερό. Η
επαναρρόφηση του νερού δια µέσου των νεφρών είναι αποτελεσµατική και όπως

102
φαίνεται, οι ισχυρές µονωτικές στρώσεις λίπους κάτω από το δέρµα, πιθανόν
παίζουν επίσης και ένα ρόλο αποθήκευσης του νερού µεταβολισµού, όπως βρέθηκε
να συµβαίνει σε µερικά ζώα της ερήµου.
Είναι αξιοσηµείωτο ότι σε σύντοµο χρόνο, µιλώντας γεωλογικά δηλ. σε 50 εκατοµ.
χρόνια περίπου, τα θηλαστικά τα οποία έχουν προσαρµοσθεί στη θαλάσσια ζωή
έχουν επιτύχει ριζικές αλλαγές και έχουν γίνει πολύ πετυχηµένα ζώα στο θαλασσινό
τους περιβάλλον. Οι φάλαινες έχουν πλήρως προσαρµοσθεί στη θαλάσσια ζωή και
ουδέποτε έρχονται έξω στη στεριά, ούτε ακόµη όταν έχουν µαζί τους νεαρούς
απογόνους. Μερικά από τα ζώα αυτά έχουν εξελιχθεί στα µεγαλύτερα ζώα τα οποία
έχουν ποτέ ζήσει, σηµαντικά ξεπερνώντας σε όγκο τους µεγαλύτερους από τους
τροµερούς σε µέγεθος δεινόσαυρους. Οι φώκιες έχουν δηµιουργήσει τους
µεγαλύτερους πληθυσµούς µεγάλων σαρκοφάγων θηλαστικών επί της Γης. Και οι
δύο αυτές οµάδες φαίνεται να κατείχαν ασφαλείς και κυρίαρχες θέσεις στους
ωκεανούς µέχρι την εντυπωσιακή έκρηξη του πληθυσµού και της τεχνολογίας εκ
µέρους του ανθρώπου κατά τις τελευταίες λίγες εκατοντάδες χρόνια.
Τα θαλάσσια θηλαστικά περιλαµβάνουν δύο άφθονες και ευρέως διαδεδοµένες
οµάδες τα Kητώδη (φάλαινες, φώκαινες, και δελφίνια) και τα Σαρκοφάγα (φώκιες,
θαλάσσιοι λέοντες και θαλάσσιοι ελέφαντες), καθώς και άλλες οµάδες λιγότερο κοινές
όµως, όπως τα Sirenia (ντυγκόκ, µανάτοι και θαλάσσιες αγελάδες). Κάθε µια από τις
οµάδες αυτές έχουν προέλθει από έναν χερσαίο θηλαστικό πρόγονο.
Τα Κητώδη (Cetacea) είναι όλα µεγάλα µέχρι πολύ µεγάλα θαλάσσια θηλαστικά στα
οποία έχει επέλθει πλήρης απώλεια ή εξαιρετική µείωση τριχώµατος. Τα εµπρόσθια
άκρα είναι σαν κουπιά, χρησιµοποιούνται ως όργανα ισορροπίας κατά την
κολύµβηση και δείχνουν µια δευτερογενή αύξηση στον αριθµό των οστών στα
δάκτυλα, σε σχέση µε εκείνον στα άλλα θηλαστικά. Τα πίσω άκρα έχουν
εξαφανισθεί, τα κοιλιακά οστά έχουν εκφυλισθεί σε εσωτερικά υποτυπώδη ίχνη. Το
σώµα είναι µονωµένο µε ένα ελαιώδες φουσκωµένο στρώµα κάτω από το δέρµα, το
οποίο επίσης βοηθάει στην επίπλευση του ζώου και πιθανόν στην αποθήκευση
νερού µεταβολισµού. Τα ρουθούνια όλων των συγχρόνων ειδών είναι τοποθετηµένα
στο πάνω µέρος της κεφαλής µάλλον παρά προς το εµπρόσθιο άκρο, όπως
συµβαίνει στα άλλα θηλαστικά και έχουν ειδικές βαλβίδες, οι οποίες, κάνοντας χρήση
της πίεσης του νερού, κλείνουν τον φυσητήρα όταν το ζώο καταδύεται. Ο εγκέφαλος
είναι µεγάλος µε συσπειρωµένα εγκεφαλικά ηµισφαίρια. Το στοµάχι µερικών απ'
αυτά είναι σύνθετο µε 4-5 χώρους. Τα θηλυκά έχουν ένα ζεύγος γαλακτικών αδένων
στη βουβωνική περιοχή. Το πίσω άκρο του σώµατος είναι ισχυρά µυώδες και έχει το
σχήµα ζεύγους οριζόντιων πτερυγίων που µοιάζουν µε την ουρά των ψαριών και

103
δίνουν στο ζώο κίνηση, όπως ακριβώς το πλοίο που προωθείται από την έλικα. Έχει
υπολογισθεί ότι µια φάλαινα 27 µέτρων µπορεί να παράγει ώθηση ίση µε εκείνη που
παράγεται από µια µηχανή 520 ΗΡ. Οι µεγαλύτερες φάλαινες µπορούν να κινηθούν
µε ταχύτητα µέχρι 20 κόµβων και µπορούν να κολυµπούν όλη την ηµέρα µε µέση
ταχύτητα 10 κόµβων και ίσως περισσότερο.
Τα Σαρκοφάγα (Carnivora) περιλαµβάνουν τρεις οικογένειες θαλάσσιων θηλαστικών,
που και οι τρεις εµφανίσθηκαν κατά το Μειόκαινο, οι άµεσοι όµως πρόγονοί τους
είναι αβέβαιοι. Όλα είναι µεγάλα υδρόβια σαρκοβόρα που κοπαδιάζουν και είναι
πολύγαµα. Τα ζώα αυτά προσεγγίζουν τις ακτές ή τους πάγους και πάνω σ' αυτά
γεννούν τους απογόνους τους. Οι θαλάσσιοι ελέφαντες είναι µεγαλύτεροι από τις
θαλάσσιες αγελάδες και τις περισσότερες φορές δηµιουργούν χαρέµια τα οποία
υποστηρίζουν µαχητικά έναντι άλλων ελεφάντων. Κινούνται δύσκολα και άκοµψα
στην ξηρά αλλά είναι έµπειροι κολυµβητές και δύτες. Τα άκρα τους είναι πτερύγια µε
νηκτική µεµβράνη. Τα δόντια τους είναι απλούστερα από εκείνα των θηλαστικών της
ξηράς µε τους προγοµφίους και γοµφίους του ίδιου περίπου χαρακτήρα. Τα
περισσότερα είδη θηρεύονται πολύ εντατικά για το δέρµα, το λάδι και τη σάρκα τους.
Μερικά τείνουν σχεδόν να εκλείψουν. Άλλα έχουν τεθεί κάτω από διεθνές καθεστώς
προστασίας.
Τα Sirenia είναι µεγάλα ογκώδη υδρόβια θηλαστικά τα οποία εκ πρώτης όψεως
µοιάζουν µε τις φάλαινες, στο γενικό σαν οβίδα σχήµα του σώµατός τους, που γίνεται
λεπτότερο προς τα πίσω για να καταλήξει σε οριζόντια πτερύγια. Τα πίσω άκρα
έχουν εξαφανισθεί ενώ τα εµπρόσθια είναι τροποποιηµένα σε πλατειά πτερύγια.
Παρ' όλα αυτά τα χαρακτηριστικά µόνο το 1816 ο De Blainville αναγνώρισε τις
βασικές διαφορές µεταξύ αυτής της οµάδας και των κητωδών και τοποθέτησε τα ζώα
αυτά συστηµατικά. Όλα είναι νωθρά ζώα, µε προτίµηση στα ρηχά νερά και στις
εκβολές ποταµών, τρεφόµενα µε θαλάσσια φυτά και θαλάσσιο γρασίδι. Το κεφάλι
τους είναι αναλογικά µικρότερο από εκείνο των φαλαινών, έχουν πλατύ ρύγχος µε
κινητά κάθετα εκτεινόµενα χείλια και ισχυρά µουστάκια στους ρώθωνες. Τα µάτια
είναι µικρά, οι ρώθωνες µάλλον ραχιαίοι και τα ακουστικά ανοίγµατα χωρίς ακουστικά
πτερύγια. Το κεφάλι ξεπροβάλλει όχι ευδιάκριτα µέσα από το ογκώδες έµπροσθεν
τµήµα του σώµατος. Το δέρµα είναι σκληρό και είναι γυµνό ή έχει αραιό τρίχωµα.
Τα θηλυκά έχουν ένα ζεύγος γαλακτοφόρων αδένων θωρακικά τοποθετηµένους. Τα
οστά του κρανίου και η θωρακική περιοχή είναι εξαιρετικά ογκώδη. ∆εν υπάρχει
οστούν της κλείδας ούτε κοιλιακά οστά εκτός από ένα µικρό ζεύγος υποτυπωδών
οστών εξαρτώµενο σε κάποια απόσταση από τη σπονδυλική στήλη. Ο εγκέφαλος
είναι πολύ λιγότερο σπειροειδής και αναπτυγµένος σε σχέση µε τα κητώδη. Τα

104
Sirenia δείχνουν γενικά λίγο φόβο από την παρουσία του ανθρώπου και εύκολα
πλησιάζονται. Γεννούν ένα άτοµο την φορά, το οποίο φροντίζουν πάρα πολύ.

105
5. Ο ΑΝΘΡΩΠΟΣ ΚΑΙ Η ΘΑΛΑΣΣΑ
∆ύο χιλιάδες χρόνια πριν, ο ανθρώπινος πληθυσµός επί της Γης ήταν πιθανόν
µεταξύ 200 και 300 εκατοµ. ατόµων. Ο υψηλός βαθµός γεννήσεων ισορροπούσε µε
τον υψηλό βαθµό θανάτων και ο πληθυσµός αυξάνονταν σιγά-σιγά. Περί τα 1650 ο
πληθυσµός διπλασιάστηκε στα 500 εκατοµ. Η βιοµηχανική επανάσταση έφερε
προόδους στην ιατρική και οι τεχνολογικές βελτιώσεις στα συστήµατα παραγωγής
τροφής µείωσαν απότοµα τη θνησιµότητα των ανθρώπων. Έτσι, µε τον υψηλό βαθµό
γεννήσεων, που συνεχίστηκε, και µε µειωµένο βαθµό θνησιµότητας, ο πληθυσµός
της Γης διπλασιάστηκε και πάλι (1 δις) στα 1850. Οι µειωµένοι βαθµοί θνησιµότητας
συνεχίστηκαν και σε συνδυασµό µε τους αυξηµένους βαθµούς γεννήσεων, η αύξηση
του πληθυσµού της Γης επιταχύνθηκε µέσα στον 20ο αιώνα. Σήµερα ο πληθυσµός
της Γης υπολογίζεται στα 6 δις και θα φθάσει στα 7 στο τέλος της πρώτης δεκαετίας
του 21ου αιώνα.
Οι ανάγκες για τροφή από τον συνεχώς αυξανόµενο πληθυσµό της Γης, είναι
τροµακτικές και αυξάνονται σε βαθµό ανησυχητικό για το γνωστό µέχρι σήµερα
δυναµικό παραγωγής τροφών από τη Γη. Τα υπάρχοντα συστήµατα παραγωγής
τροφής είναι αδύνατον να ικανοποιήσουν αυτές τις ανάγκες. Αµέτρητα εκατοµµύρια
ανθρώπων υποφέρουν από πείνα µέχρι θανάτου και εκατοντάδες εκατοµµύρια
υποφέρουν από υγεία εξαιτίας της µη ικανοποιητικής διατροφής τους. Πολλοί
άνθρωποι θεωρούν τις θαλάσσιες πηγές τροφής ως ένα κρίσιµο σηµείο για τη λύση
των προβληµάτων της τροφής του αύριο.
Η λήψη τροφής από τη θάλασσα, αποτελεί µια από της αρχαιότερες δραστηριότητες
του ανθρώπου. Παρά το γεγονός ότι η χρησιµοποίηση της θάλασσας ως µέσον
µεταφοράς, ως τόπος αναψυχής και ιδιαίτερα ως πηγή πετρελαίου και µετάλλων,
είναι πολύ σηµαντικές χρήσεις, η αλιεία εξακολουθεί να αποτελεί µια από τις κύριες
δραστηριότητες εκµετάλλευσης της θάλασσας από τον άνθρωπο.
Όταν η αλιεία αρχίζει σε ένα οποιοδήποτε απόθεµα αρχικά η παραγωγή είναι γενικά
υψηλή και περιλαµβάνει µεγάλο ποσοστό µεγάλων ψαριών. Εφόσον όµως η
αλιευτική προσπάθεια συνεχίζεται µε αυξανόµενη ένταση, το µέσο αλιευόµενο
µέγεθος ψαριών τείνει να µειωθεί και µερικές φορές µειώνεται η αφθονία του
αποθέµατος. Αντίθετα αν η αλιευτική προσπάθεια πηγαίνει παράλληλα µε το
δυναµικό αύξησης και αναπαραγωγής του αποθέµατος, τότε είναι δυνατή η αλίευση
µιας µεγίστης επιτρεπόµενης παραγωγής (maximum sustainable yield), η οποία
οδηγεί στην εξαλίευση ενός αποθέµατος χωρίς σηµαντικές διακυµάνσεις στην
αφθονία του. Πολύ συχνά όµως η αλιευτική πίεση πάνω στα αποθέµατα είναι πολύ

106
µεγαλύτερη από εκείνη που το απόθεµα µπορεί να αντέξει. Έτσι η υπερβολική
εξαλίευση σε συνδυασµό µε τη φυσική θνησιµότητα ξεπερνούν το ρυθµό εισαγωγής
νέων ατόµων στο απόθεµα, µε αποτέλεσµα η παραγωγή να µειώνεται.
Παραδείγµατα τέτοιων υπεραλιεύσεων σε αποθέµατα υπάρχουν πάρα πολλά. Η
ρέγγα, η περουβιανή αντσούγα, οι φάλαινες (great whales) είναι µερικά πρόσφατα
οδυνηρά παραδείγµατα αυτής της τακτικής. Και γεννάται το ερώτηµα: γιατί οι
ψαράδες και οι αλιευτικές χώρες γενικά εκµεταλλεύονται τις πηγές, από τις οποίες
εξαρτώνται, σε βαθµό που προκαλούν αρχικά µείωση της παραγωγής και πολύ
συχνά ολοκληρωτική απώλεια των αποθεµάτων; Ακόµη και µε το λαβύρινθο των
νοµικών και οικονοµικών συσχετισµών, οι οποίοι εµπλέκονται στο φαινόµενο αυτό,
δεν είναι δύσκολο να βρούµε τους λόγους, οι οποίοι προφανώς ωθούν σε τέτοιες
καταστροφικές ενέργειες. Τα είδη που υπεραλιεύονται είναι προφανές ότι δεν έχουν
κτήτορα, δεν ανήκουν σε έθνη ή σε άτοµα. Υπάρχουν έξω από τα όρια δικαιοδοσίας
των κρατών και κατά συνέπεια είναι ελεύθερα για πρόσβαση από οποιοδήποτε
έθνος.
Ιστορικά ο όρος ελεύθερη πρόσβαση (open access) στις θάλασσες που δεν ανήκουν
σε χώρες (διεθνή ύδατα - high seas) έχει αναπτυχθεί κατά τους 16ο και 17ο αιώνες,
όταν το δικαίωµα για ελεύθερη ναυσιπλοία ήταν περισσότερο σηµαντικό από την
ελευθερία στην αλιεία. Αλλά καθώς τα παράκτια ιχθυαποθέµατα εξαντλούνταν, οι
ψαράδες γίνονταν µε αυξανόµενο ρυθµό εξαρτώµενοι από αποθέµατα που υπήρχαν
στα διεθνή ύδατα. Έτσι τελικά ανακάλυψαν ότι η ελευθερία αλιείας στα διεθνή ύδατα
ήταν ουσιαστικά διαφορετικό από την ελευθερία στη ναυσιπλοία.
Θεωρητικά υποτίθεται ότι οι θαλάσσιες πηγές δεν έχουν ιδιοκτήτη και είναι ελεύθερες
για πρόσβαση από οποιονδήποτε. Αλλά µε τη σηµερινή εξέλιξη προηγµένης
τεχνολογίας, µερικά έθνη έχουν πετύχει πιο γρήγορα από άλλα την εκµετάλλευση
αυτών των πηγών. Έτσι αν οι ψαράδες ενός έθνους αποτύχουν να εκµεταλλευθούν
αυτές τις χωρίς ιδιοκτήτη πηγές, όσο το δυνατόν πιο γρήγορα, κάποιοι άλλοι θα το
κάνουν γρήγορα. Με την τάση αυτή λοιπόν, από ένα εκµεταλλεύσιµο είδος σ' ένα
άλλο, τα έθνη οδηγούνται σε αναπόφευκτα, προβλεπόµενα όµως αποτελέσµατα, που
είναι αυξηµένος ανταγωνισµός µεταξύ εθνών για περιορισµένες πηγές, αύξηση της
αλιευτικής προσπάθειας, φθίνουσα αλιευτική αποτελεσµατικότητα και φυσικά,
υπεραλίευση.
Όταν ένα κοπάδι από ψάρια βρεθεί µέσα στα δίκτυα τότε πλέον δεν είναι περιουσία
όλων, αλλά ανήκει στον ψαρά που τοποθέτησε το δίχτυ και έσυρε τα ψάρια πάνω
στο πλοίο του.

107
Όλοι οι άνθρωποι και όλα τα έθνη µοιράζονται το κόστος της απώλειας των ψαριών
που εξαφανίζονται, των φαλαινών που µειώνονται δραµατικά, καθώς και άλλων
θαλάσσιων θηλαστικών, τα οποία έχουν σχεδόν εξαφανισθεί εξαιτίας της
υπερεκµετάλλευσης. Αλλά ακόµη και η βραχυπρόθεσµη ωφέλεια που προκύπτει
από την υπερεκµετάλλευση δεν µοιράζεται στον ίδιο βαθµό. Το τραγικό της
κατάστασης είναι ότι το µεγαλύτερο µέρος αυτής της βραχυπρόθεσµης ωφέλειας
κερδίζεται από εκείνον που πιο βαριά εκµεταλλεύτηκε και καταχράστηκε τις
απροστάτευτες ζώσες πηγές των διεθνών υδάτων.
Η αναπτυσσόµενη παγκόσµια εκµετάλλευση των αλιευτικών πηγών, το αυξανόµενο
ενδιαφέρον για τα µεταλλεύµατα του βυθού των θαλασσών, το πετρέλαιο και άλλες
πηγές, καθώς και πιθανές χρήσεις της θάλασσας για στρατιωτικές δραστηριότητες
έχουν γίνει θέµατα διεθνούς ενδιαφέροντος. Συνεχείς προσπάθειες είναι υπό εξέλιξη
για να αναπτυχθεί µέσω των Ηνωµένων Εθνών ένας περιεκτικός Νόµος της
Θάλασσας για να αποφευχθεί το χάος, η σύγχυση και η διαµάχη που µπορούν να
αναπτυχθούν, αν αφεθούν τα θέµατα αυτά στην ανταγωνιστική πάλη µεταξύ των
εθνών.
Το δόγµα της ελευθερίας των θαλασσών, όπως διατυπώθηκε από τον Γερµανό
δικαστή Ηugo Grotius στα 1609, ήταν νόµος παραδεκτός από τα έθνη µέχρι
πρόσφατα, όταν προβλήµατα υπεραλίευσης, ρύπανσης, αυξηµένων εµπορικών και
στρατιωτικών χρήσεων, καθώς και η ανακάλυψη πηγών µετάλλων ανοικτά των
θαλασσών έκαναν την απλή µέχρι πρόσφατα υπόθεση, να µην είναι πλέον πρακτικά
εφαρµόσιµη.
Η βασική σκέψη της ελευθερίας των θαλασσών απλά διαίρεσε τη θάλασσα σε
χωρικά ύδατα (territorial sea), που είναι µια στενή λωρίδα θάλασσας, ελεγχόµενη
από την παράκτιο χώρα και τα διεθνή ύδατα (high seas), όπου υπάρχει πλήρης
ελευθερία πρόσβασης. Στην αρχή η ζώνη των χωρικών υδάτων εκτείνονταν µέχρι τα
τρία µίλια από την ακτή. Στα 1945 οι ΗΠΑ επέκτειναν τη δικαιοδοσία τους επί των
φυσικών πηγών της ηπειρωτικής υφαλοκρηπίδας. Το παράδειγµα των Αµερικανών
ακολούθησαν αµέσως άλλες χώρες και τα Η.Ε. άρχισαν έναν αριθµό διεθνών
συναντήσεων για να κωδικοποιήσουν αυτές τις αλλαγές.
Από το 1958, που πραγµατοποιήθηκε στη Νέα Υόρκη η πρώτη διεθνής συνάντηση
για τον Νόµο των Θαλασσών, έγιναν µέχρι σήµερα περισσότερες από είκοσι τέτοιες
συναντήσεις. Οι διαφορές µεταξύ των εθνών ήταν και θα είναι πάντοτε µεγάλες,
εξαιτίας ποικίλων συµφερόντων, γι' αυτό και οι αποφάσεις µε την απαραίτητη

108
πλειοψηφία υπήρξαν ελάχιστες. Εντούτοις υπάρχουν ορισµένες αποφάσεις οι
οποίες ισχύουν, είτε ως γενικά αποδεκτές είτε ως µονοµερείς ενέργειες.
Ο νέος Νόµος των Θαλασσών, όπως προτείνεται να ισχύσει, χωρίς φυσικά γενική
παραδοχή, διαιρεί τις θάλασσες σε έξι πολύπλοκες και µερικές φορές
υπερκαλύπτουσες η µια την άλλη περιοχές.
1. Εσωτερικά ύδατα (Internal waters). Τέτοιες περιοχές είναι οι κόλποι παράκτιων
χωρών, στους οποίους οι χώρες έχουν πλήρη εθνική κυριαρχία.
2. Χωρικά Ύδατα (Territorial Sea). Κάθε χώρα έχει το δικαίωµα να καθορίσει το
πλάτος της ζώνης των χωρικών της υδάτων µέχρι 12 ναυτικά µίλια από τη βασική
γραµµή που καθορίστηκε σύµφωνα µε απόφαση της συνόδου των Η.Ε.
3. Γειτονική Ζώνη (Contiguous Zone). Η γειτονική ζώνη µπορεί να εκτείνεται 24
ναυτικά µίλια πέρα της βασικής γραµµής. Μέσα σ' αυτή τη ζώνη η παράκτια χώρα
έχει το δικαίωµα να προλαµβάνει τυχόν παραβιάσεις παγίων συνηθειών του
παράκτιου κράτους, καθώς και δεδοµένων της οικονοµικής τους ζωής και θεµάτων
υγιεινής.
4. Αποκλειστική Οικονοµική Ζώνη (Exclusive Economic Zone). H ζώνη αυτή µπορεί
να εκτείνεται µέχρι 200 µίλια. Σ' αυτήν την περιοχή κάθε παράκτια χώρα µπορεί
να έχει αποκλειστικά δικαιώµατα σε όλες τις πηγές, ζώσες και µη ζώσες, καθώς
και την ευθύνη για την εφαρµογή και τον έλεγχο κατάλληλων µέτρων διαχείρισης.
Μπορεί να έχουν επίσης δικαίωµα στο νερό καθώς και στον πυθµένα µε το
υπέδαφος, για τη ναυσιπλοία όµως η περιοχή αυτή θεωρείται ουσιαστικά ως
διεθνή ύδατα (high seas)
5. Ηπειρωτική υφαλοκρηπίδα εκτεινόµενη πέρα των 200 µιλίων (Continental Shelf
Extending Beyond 200 miles). Χώρες µε τέτοια ευρεία ζώνη διεκδικούν
υφαλοκρηπίδα η οποία εκτείνεται σε µερικές περιπτώσεις αρκετές εκατοντάδες
µίλια πέρα από τη ζώνη των 200 µιλίων. Στην περιοχή αυτή οι χώρες έχουν
δικαίωµα επί τού πυθµένος και του υπεδάφους αλλά όχι στο θαλασσινό νερό.
6. ∆ιεθνής περιοχή (International Area). Τέτοια περιοχή είναι ό,τι αποµένει ύστερα
από όλες τις παραπάνω διεκδικήσεις. Στη ζώνη αυτή έχουν δικαίωµα
ναυσιπλοίας και αλιείας όλες οι χώρες. Εδώ βρίσκονται οι περισσότεροι από τους
περίφηµους κονδύλους µαγγανίου, των οποίων η διαχείριση γίνεται κάτω από την
αιγίδα ∆ιεθνούς Αρχής του Θαλάσσιου Πυθµένα (International Seabed Authority).
Οι αντιπροσωπείες των χωρών ύστερα από επανειληµµένες διασκέψεις έχουν
καταλήξει σ' ένα αριθµό σηµείων για τα οποία υπάρχει µια σχετικά µεγάλη αποδοχή:

109
1. Ζώνη 12 µιλίων χωρικών υδάτων, µε άλλα 12 µίλια γειτονικής ζώνης.
2. Αποκλειστική Οικονοµική ζώνη 200 µιλίων στην οποία οι παράκτιες χώρες έχουν
αποκλειστικό δικαίωµα των πηγών της ζώνης.
3. ∆ιεκδίκηση µεριδίου εισοδήµατος από άντληση πετρελαίου πέρα από τη ζώνη των
200 µιλίων.
4. Αποδοχή της αρχής για τη λειτουργία µιας διεθνούς επιτροπής που να
διαχειρίζεται τους βαθιούς θαλάσσιους πυθµένας.
5. Ανεµπόδιστη µετακίνηση στα διεθνή ύδατα και δια µέσου διεθνών πορθµών.
6. Αποδοχή αρχών και ευθυνών σε σχέση µε τη µόλυνση.
7. Πρόσβαση σε θάλασσες χωρών, που είναι περιορισµένες από παντού µε ξηρά.
8. Ανάγκη ίδρυσης ενός ∆ικαστηρίου για τον Νόµο της Θάλασσας, που να εκδικάζει
διαφορές µεταξύ χωρών (Το ∆ιεθνές ∆ικαστήριο της Χάγης).
9. Επιστηµονική Έρευνα επιτρέπεται µε την προϋπόθεση ότι το αντικείµενο της
έρευνας πρέπει να είναι καθορισµένο.
10.Μεταφορά τεχνολογίας σε αναπτυσσόµενες χώρες.
11.Υποχρέωση για διατήρηση του θαλάσσιου περιβάλλοντος.
12.Η επιτυχία µιας συνάντησης για τα θέµατα αυτά εξαρτάται από την επίλυση του
προβλήµατος άντλησης µετάλλων από τον πυθµένα των θαλασσών και από τη
δηµιουργία µιας διεθνούς αρχής ελέγχου.
5.1 ∆ιεθνείς Οργανισµοί Αλιείας
Για τον έλεγχο της πρόσβασης των ψαράδων στις αλιευτικές πηγές των διεθνών
υδάτων, τα ενδιαφερόµενα έθνη έχουν δηµιουργήσει επιτροπές κανονισµού της
αλιείας στις κατά περίπτωση αλιευτικές πηγές. Αυτές οι επιτροπές είναι
επιβαρηµένες µε την ευθύνη του συντονισµού διαχείρισης και εκµετάλλευσης των
διαφόρων αποθεµάτων που υπάγονται στον έλεγχο αυτών των επιτροπών. Π.χ. η
∆ιεθνής Επιτροπή για τις Αλιείες του Βορειοδυτικού Ατλαντικού (ΙCNAF)
περιλαµβάνει τον Καναδά, ∆ανία, Γαλλία, Γερµανία, Ισλανδία, Ιταλία, Πολωνία,
Ισπανία, Ρωσία, ΗΠΑ και Ηνωµένο Βασίλειο. Οι κανονισµοί που έχουν θεσπισθεί
από την Επιτροπή αυτή δεν έχουν την ισχύ διεθνούς νόµου αλλά είναι σηµαντικοί για
τα έθνη µέλη. Παρόλα αυτά, τα έθνη δεν έχουν κατορθώσει να συγκρατήσουν µια

110
σοβαρή υπεραλίευση του µπακαλιάρου και της ωκεάνιας πέρκας του Β.∆.
Ατλαντικού.
Άλλες επιτροπές και ειδικότερα εκείνες που ρυθµίζουν τις αλιείες του Χάλιµπατ (είδος
γλώσσας πολύ µεγαλύτερου µεγέθους) και του Σολοµού στο Β. Ειρηνικό έχουν
µεγαλύτερη επιτυχία. Αλλά πολύ µεγάλος αριθµός αλιευτικών σκαφών από τις ΗΠΑ,
τον Καναδά, την Κορέα, την Ιαπωνία και τη Ρωσία προσελκύονται απ' αυτές τις καλά
διαχειριζόµενες αλιείες. Κάτω απ' αυτές τις συνθήκες αλιευτικής πίεσης, τα
προβλήµατα διαχείρισης µεγεθύνονται, τα οφέλη των ψαράδων µειώνονται και οι
πιθανότητες για υπεραλίευση είναι πολύ έντονα υπαρκτές.
Πολλά έθνη, µόνα τους ή σε συνεργασία µε διεθνείς οργανισµούς, έχουν
πραγµατοποιήσει βήµατα να εµποδίσουν την υπεραλίευση και να προωθήσουν
καλύτερη χρήση των περιορισµένων πηγών τροφής από τη θάλασσα. Μερικά έθνη
συνηγόρησαν για έλεγχο όλων των ειδών πηγών στα διεθνή ύδατα από τα Ηνωµένα
Έθνη, όπως οι πηγές τροφής, µετάλλων και πετρελαίου. Τα µειονεκτήµατα αυτής της
ιδέας βέβαια είναι φανερά σε όλους. Τα Έθνη δεν επιθυµούν να επιτρέψουν στον
ΟΗΕ ή οποιονδήποτε άλλο διεθνή Οργανισµό να λαµβάνουν κοινωνικές, πολιτικές
και νοµικές αποφάσεις για να ρυθµίζουν δίκαια και ισοκατανεµηµένα την
εκµετάλλευση των θαλάσσιων πηγών. Κι αυτό διότι όταν υπάρχει αποτελεσµατικός
διεθνής έλεγχος, µερικά έθνη δρουν για λογαριασµό τους για τη ρύθµιση των
αλιευτικών δραστηριοτήτων τους µέσα σε µια ευρεία περιοχή που βρίσκεται πλησίον
τους. Για παράδειγµα στα 1974, 35 έθνη επεξέτειναν αυθαίρετα τα χωρικά τους
ύδατα ή δηµιούργησαν ελεγχόµενες αλιευτικές ζώνες πέρα από τα 12 µίλια που
αναγνωρίζονται από τον διεθνή κανονισµό. Αρκετά από τα έθνη αυτά διεκδικούν
δικαίωµα ελέγχου πάνω σε παράκτιες ζώνες που εκτείνονται µέχρι 200 και
περισσότερα ακόµη µίλια από τις ακτές τους.

111
ΒΙΒΛΙΟΓΡΑΦΙΚΕΣ ΠΗΓΕΣ
Barnes, R.S.K. and R.N. Hughes. 1988. An Introduction to Mar;ine Ecology .
Blackwell Scientific Publications.
Burke W.T. 1983. Extended Fisheries Juristiction and the New Law of the Sea
In:Global Fisheries. Perspectives for the 1980s. Brian J. Rothshild (Editor),Spring-
Verlag. Θεοδώρου, Αθανάσιος Ι. 1996. Εισαγωγή στο Θαλάσσιο Περιβάλλον. Σηµειώσεις. Βόλος.
Gross, M. G. 1995. Principles of Oceanography. Prentice Hall.
Ingmanson, D. and W. Wallace, 1989. Oceanography. An Introduction.
Wadsworth Publishing Company, Belmont, California.
King Michael, 1995. Fisheries Biology, Assessment and Management.
Fishing News Books.
Λυκάκης, Σ. 1992. Υδροβιολογία: θαλάσσια Βιολογία και Στοιχεία
Λιµνολογίας. Εκδόσεις ΣΥΜΜΕΤΡΙΑ, Πάτρα.
Levinton, S. Jeffrey, 1982. Marine Ecology. Prentice Hall Inc. Englewood
Cliffs, N. Jersey.
McConnaughey, Bayard. H,1978. Introduction to Marine Biology. 3rd Edition.
The C.V. Mosby Company.
Moyle, P.B. and J. J. Cech, Jr. 1982. Fishes: An Introduction to Ichthyology.
Prentice-Hall, Inc.
Meadows, P.S. and J.I. Campbell, 1978. An introduction to Marine science.
John Wiley and Sons.
Παπουτσόγλου, Ε.Σ. 1980. Tο υδάτινο περιβάλλον και οι οργανισµοί του.
Εκδόσεις Καραµπερόπουλος Α.Ε.
Sumich, L. James, 1976. An introduction to the Biology of Marine Life. Un C.
Brown Company publishers, Dubuque Iowa.
Weihaupt, John G.1979. Explorations of the OCEANS. An Introduction to
Oceanography. Macmillan Publishing Co., Inc., New York – Collier Macmillan
Publishers, London.

112





















