Embed Size (px)
DESCRIPTION
细菌耐药性及其变异趋势. 山 东 大 学 医 学 院. 临床医学七年制 四班 四组 董平 黄珊珊 牟慧珺 史雪 张忠伟 刘倩 李宾 许春阳. 细菌的耐药性又称抗药性,一般是指细菌与药物多次接触后,对药物的敏感性下降甚至消失,致使药物对耐药菌的疗效降低或无效。. - PowerPoint PPT Presentation
Citation preview

临床医学七年制 四班 四组 董平 黄珊珊 牟慧珺 史雪张忠伟 刘倩 李宾 许春阳
细菌耐药性及其变异趋势
山东大学医学院
细菌的耐药性又称抗药性,一般是指细菌与药物多次接触后,对药物的敏感性下降甚至消失,致使药物对耐药菌的疗效降低或无效。
1992 年美国疾病控制中心( CDC )的资料表明,有 13300 例住院患者,是因为对所使用的抗菌药物耐药,细菌感染得不到控制而死亡。 几十年来,人类发明了大量的抗菌药物,目前应用于临床的已不下 200 种之多,而仍以平均每年 10 种以上新的抗菌药物问世的速度在增长。一方面多种抗菌药物投入临床应用,另一方面则是耐药菌株的不断产生。耐药细菌的产生,很大程度上应归结为几十年来,人类大量使用抗菌药物的结果。细菌在大范围选择性压力下,保留了它们中最耐药的菌株,而这种耐药性可以在细菌间传播,耐药细菌又能在全世界范围内播散,因而多重耐药菌株的感染导致患者死亡的报道在逐渐增多。
1. 产生灭活酶 灭活酶有两种, 一是水解酶,如 β- 内酰胺酶可水解青霉
素或头孢菌素。该酶可由染色体或质粒介导,某些酶的产生为体质性(组构酶);某些则可经诱导产生(诱导酶) .

。二是 钝化酶又称合成酶,可催化某些基团结合到抗生素的 OH 基或 NH2 基上,使抗生素失活。多数对氨基甙类抗生素耐药的革兰阴性杆菌能产生质粒介导的钝化酶,如乙酰转移酶作用于 NH2 基上,磷酸转移酶及核苷转移酶作用于 OH 基上。上述酶位于胞浆膜外间隙,氨基甙类被上述酶钝化后不易与细菌体内的核蛋白体结合,从而引起耐药性。
2. 改变细菌胞浆膜通透性 细菌可通过各种途径使抗菌药物不易进入菌体,如革兰阴性杆菌的细胞外膜对青霉素 G 等有天然屏障作用;绿脓杆菌和其他革兰阴性杆菌细胞壁水孔或外膜非特异性通道功能改变引起细菌对一些广谱青霉素类、头孢菌素类包括某些第三代头孢菌素的耐药;细菌对四环素耐药主要由于所带的耐药质粒可诱导产生三种新的蛋白,阻塞了细胞壁水孔,使药物无法进入。革兰阴性杆菌对氨基甙类耐药除前述产生钝化酶外,也可由于细胞壁水孔改变,使药物不易渗透至细菌体内。
3. 细菌体内靶位结构的改变 链霉素耐药株的细菌核蛋白体 30S 亚基上链霉素作用靶位 P10 蛋白质发生改变;利福平的耐药性是细菌 RNA 多聚酶的 β' 亚基发生改变,使其与药物的结合力降低而耐药。由质粒介导的对林可霉素和红霉素的耐药性,系细菌核蛋白体 23S 亚基的腺嘌呤甲基化,使药物不能与细菌结合所致。某些肺炎球菌、淋球菌对青霉素 G 耐药,以及金葡菌对甲氧苯青霉素耐药,乃因经突变引起青霉素结合蛋白( PBPs )改变,使药物不易与之结合。这种耐药株往往对其他青霉素(如苯唑或邻氯青霉素)和头孢菌素类也都耐药。
4. 其他 细菌对磺胺类的耐药,可由对药物具拮抗作用的底物 PABA (对氨基苯甲酸)的产生增多所致;也可能通过改变对代谢物的需要等途径。
耐药细菌的变迁有以下 6个主要表现: ( 1 )耐甲氧西林的金葡菌(MRSA)感染率增高; ( 2 )凝固酶阴性葡萄球菌(CNS)引起感染增多; ( 3 )耐青霉素肺炎球菌( PRP)在世界范围,包
括许多国家和地区传播; ( 4 )出现耐万古霉素屎肠球菌( VRE)感染; ( 5 )耐青霉素和耐头孢菌素的草绿色链球菌( PRS)的出现;
( 6 )产生超广谱 β-内酰胺酶( ESBL)耐药细菌变异。

1 )耐甲氧西林的金葡菌 (MRSA) MRSA 是多重耐药株,早期就发现它对
青霉素、四环素、链霉素、红霉素、林可霉素及氨基糖甙类等耐药。 90 年代后,全世界各个国家MRSA 已上升为 62% 。而且在耐药谱方面新的 MRSA更为广泛,包括了广谱、超广谱 β- 内酰胺类、氟喹诺酮类、氨基糖甙类、碳青霉烯类等。
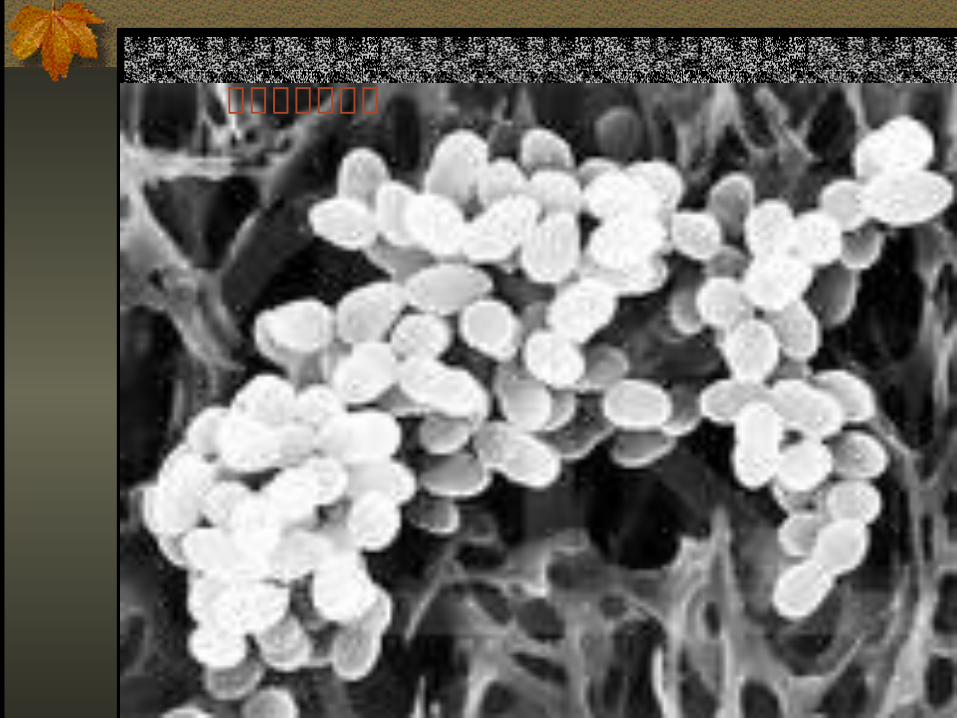
金黄色葡萄球菌
2 凝固酶阴性葡萄球菌( CNS ) 迄今为止,报道最多的凝固酶阴性的葡萄球菌
( CNS )仍是表皮葡萄球菌,同时它也是医院内感染败血症的常见细菌。由于该菌可产生大量细胞间脂多糖粘附素 (polysaccharide Intercellular Adhesion , PIA) 与细胞外粘液样物质 (Extracellular Slime Substance , ESS) ,可通过不同途径吸附在导管表面,引致导管所致的感染。 CNS 中耐甲氧西林菌株十分常见,在治疗上造成了较大困难 .
另外,在动物实验中已证实 PIA 、 ESS具有干扰宿主防御功能, ESS 可抑制 B淋巴细胞的遗传性质、使合成免疫球蛋白减少;还可干扰吞噬细胞;可保护包埋在其中的葡萄球菌免受抗生素的作用,由于膜状物形成的生物膜( Biofilm ),使得感染灶中细菌可以不断释放,从而引起败血症。
凝固酶阴性葡萄球菌
3 耐青霉素肺炎球菌( PRP ) 肺炎球菌是引起危及生命的院外感染的最主要
的病原菌。该菌引起的全球病死率与结核病相似,每年约 300~ 500万人死亡。长期以来该菌一直对青霉素高度敏感,其 MIC 为 0.005~0.001μg/ml 。自 1967 年首次发现耐青霉素肺炎球菌( MIC 0.1μg/ml )后, 1997 年又发现多重耐药肺炎球菌流行。该菌除对青霉素耐药外,对四环素、红霉素、氯霉素、克林霉素、利福平等多种抗生素耐药。 PRP 目前增加也十分迅速,各国报告,分别已占肺炎球菌的 20%~ 50% 。

PRP 耐药机制是肺炎球菌的青霉素结合蛋白 PBP(Penicillin binding protein) 发生了改变,使其与青霉素的亲合力减低,研究证实细菌是通过遗传转化、吸收并整合来自细胞外的多聚脱氧核苷酸,使得 PBP 基因中获得异种 DNA片段,多达 4 种 PBP 发生改变。
4 耐万古霉素的屎肠球菌( VRE )万古霉素( Vancomycin )属于多肽类
抗生素。来自东方链霉菌( Streptomyces Orientalis )或土壤丝菌属( Amycolatopsis Orientalis )的糖肽类抗生素。九十年代以来一直被国际抗生素专家誉为“人类对付顽固性耐药菌株的最后一道防线”。
肠球菌对万古霉素耐药初见于 1988 年,国家医院感染监测系统报告:肠球菌对万古霉素的耐药率在美国已从 1989 年的0.8% 增加到 1993 年 13.3%;
目前,肠球菌耐药谱愈来愈广泛,表现为高水平耐青霉素,高水平耐氨基糖甙类及耐万古霉素,屎肠球菌比粪肠球菌要严重得多 .
VRE

肠球菌的多重耐药性也很严重,它可包括因产生过量的 PBP 和合成大量 β- 内酰胺酶,出现高水平耐青霉素。产生各种修饰酶,而耐氨基糖甙类抗菌素及耐万古霉素。肠球菌耐药基因通过质粒、转座子播散。在体外试验中已证明能成功转移给链球菌、乳酸球菌、单核细胞李斯特菌,特别重要的是金黄色葡萄球菌。所以必须努力控制多重耐药株的播散 .

5 耐青霉素和耐头孢菌素的草绿色链球菌 (PRS)
以前,认为它们在大多数情况下是以正常菌群定植在咽部,不引起任何症状和疾病 .
现在发现 , 它们可引起脑膜炎,心内膜炎及手术后的软组织感染等,进一步证明,在患者因疾病或治疗后抵抗力下降情况下,可以引起机会性感染。长期以来,青霉素是治疗链球菌病首选药物。近来有关耐青霉素和耐头孢菌素类的草绿色链球菌感染报道增多.
PRS
6 超广谱 β- 内酰胺酶( ESBL )耐药细菌
大多数革兰阴性杆菌对 β- 内酰胺类的耐药性是由质粒介导的 β- 内酰胺酶引起,这些酶能分解氨苄西林、羧苄西林、头孢噻吩、头孢孟多、哌拉西林等.
最近,从血培养中发现一株阴沟杆菌对第三代头孢菌素高度耐药,因为基因已由诱导型改变成稳定的、不受约束的结构型头孢菌素酶,它对克拉维酸、他唑巴坦等酶抑制剂也不敏感。这种超广谱 β- 内酰胺酶细菌产生,可能对人类健康形成严重威胁。

细菌耐药性的防治措施
1.严格控制抗菌药物的应用,必须严格掌握抗菌药的适应症,合理应用,给予足够的剂量与疗程,避免滥用。要注意必要的联合用药和有计划的轮换供药。
2.在医院内严格执行消毒隔离制度,防止耐药菌如金葡菌、绿脓杆菌、大肠杆菌、肺炎杆菌等引起交叉感染。
3.新抗菌药物的继续寻找和灭活酶拮抗剂的研究,改造化学结构,使其具有耐酶特性或易于透入菌体为解决细菌耐药性的重要措施之一。

细菌耐药性一旦产生并非稳固不变,在停用有关药物一段时间后,敏感性又可逐渐恢复。根据细菌耐药动态和发展趋势,有计划地将抗菌药物分批、分期地交替使用,可能是一项具重要意义的措施。













![p4家長會簡報.ppt [相容模式] - lctms.edu.hk · 級任介紹 中文科 英文科 數學科 4a 陳文冰老師 陳珺珺老師 黃彩霞老師 4b 梁啟妍老師 陳珺珺老師](https://img.pdfslide.tips/doc/110x75/5e097a4477b9674b7a43fddd/p4eoefcppt-c-lctmseduhk-cc-c-ec.jpg)
















