Embed Size (px)
Citation preview

76 Журнал вушних, носових і горлових хвороб, №2, 2016
УДК 616.28-888.14:616-018
Г.Е. ТІМЕН, І.І. САПІЖАК, Л.А. КУДЬ
СЕНСОНЕВРАЛЬНА ПРИГЛУХУВАТІСТЬ ТА СТОВБУРОВІ КЛІТИНИ
Від. ЛОР-патології дитячого віку (керівник – чл.-кор. НАМН України, проф. Г.Е. Тімен) ДУ «Інститут отоларингології ім. проф. О.С. Коломійченка
НАМН України» (директор – акад. НАМН України, проф. Д.І. Заболотний)
Проблема лікування хворих з сенсо-
невральною приглухуватістю посідає чільне місце в оториноларингології, зокрема в клі-нічній аудіології. За даними ВООЗ, 5-8 % населення має зниження слуху, з них 65- 93 % – сенсоневральну приглухуватість. В Україні нараховується більше 270 тисяч пацієнтів з різними формами сенсоневраль-ної приглухуватості (0,6 % від загальної кількості населення), з яких майже у 100 тис. діагностовано глухоту. В той же час відсутність ефективного медикаментозного лікування цих хворих, незважаючи на вели-ку кількість запропонованих методів, зму-шує шукати нові шляхи вирішення даної проблеми.
Головну роль у патогенезі сенсонев-ральної приглухуватості відіграє вплив різ-них патологічних факторів на волоскові клітини спірального органа. До найбільш розповсюджених пошкоджуючих факторів, які часто призводять до загибелі волоскових клітин (ВК) внутрішнього вуха, відносяться генні порушення, шум, вібрація, акустичне перевантаження, наприклад, надмірна сила звука, аутоімунні процеси, бактеріальна та вірусна інфекція, ототоксичні лікарські за-соби, травми та ін.
Спіральний орган (орган Корті) роз-міщується на базилярній мембрані перетин-частого лабіринту в завитці і являє собою сукупність нейроепiтелiальних клітин, се-ред яких найважливішими є рецепторні сен-сори, так звані волоскові клітини, завдяки яким акустичне подразнення трансформу-ється і перетворюється на фізіологічний акт звукового сприйняття. У людини нарахову-
ється приблизно 3,5-4 тис. внутрішніх та від 12 до 20 тис. зовнішніх волоскових клітин. Вважається, що від функції останніх, в ос-новному, залежить сприйняття звуку, його сила та комфортність.
Визначено, що у людей та інших сса-вців пошкоджені і ті, що загинули, волоско-ві клітини не відновлюються, а продуку-ються тільки до народження [25]. Проте у птахів, риб, і амфібій вони зберігають мож-ливість регенерації майже протягом життя. Дослідження Baird, Torres продемонстрова-но, що у риб та амфібій волоскові клітини можуть заміщуватись при пошкодженні і, окрім того, відновлюватись після загибелі [18]. Між тим було показано, що навіть у ссавців слухові волоскові клітини іноді (дуже рідко) можуть регенерувати за раху-нок викликаної їх загибеллю індукції мітозу чи трансдиференціації підтримуючих клі-тин, що заміщують волоскові у місці пош-кодження [38].
Почнемо зі світу тварин. Знайдено, що підтримуючі клітини, які широко представ-лені в слуховому та вестибулярному епітелії хребетних, можуть ділитися. Нові клітини, що з’являються в результаті поділу, мають здатність надалі диференціюватися у волос-кові клітини слухової і вестибулярної сис-тем ссавців, що веде до повернення слуху і рівноваги [38]. Цей процес має два механіз-ми: пряме трансдиференціювання та міто-тичну регенерацію. Перший полягає у зміні частини клітин, коли підтримуючі клітини перетворюються у волоскові без поділу клі-тин. Мітотична регенерація відбувається шляхом ділення підтримуючої клітини на

Журнал вушних, носових і горлових хвороб, №2, 2016 77
дві, а далі одна або обидві дочірні клітини перетворюються у волоскові. Такі повідом-лення не поодинокі.
Чимала активність в дослідженнях спонтанної регенерації сенсорних (волоско-вих) клітин припадає на два останні десяти-річчя. Однак слід зазначити, що Sоbkowicz та співавтори ще в 1975 р. отримали і про-понували для експериментів культуру клі-тин органа Корті. Нас же більше цікавлять дослідження, проведені безпосередньо на завитках ссавців, які можна вважати в біль-шому чи в меншому ступені, наближеними до людини [38].
Дуже цікаві досліди провели Oshima, Beisel, Bermingham та співавтори, які дове-ли, що внутрішнє вухо ссавців має стовбу-рові плюропотентні клітини, які в подаль-шому можуть перетворюватись у волоскові [19, 38].
Треба підкреслити, що процес перехо-ду попередників, зокрема підтримуючих клітин, у волоскові потребує значної кіль-кості особливих умов, навіть у новонаро-джених тварин (мишей). Так, Li і співавтори (2003) продемонстрували, що у новонаро-дженої мишки в спіральному органі, крім волоскових клітин, є їх попередники, які під дією в основному природних факторів – Sonic bedgehog, епідермального фактору росту, ретинової кислоти і нейротрофінів, репродуктивних факторів (Sox E) дозволя-ють їм переходити чи диференціюватись в інші клітини завитки, в тому числі і у воло-скові. Цих допоміжників (різноманітних факторів, сполук, речовин) досить багато. Наприклад, необхідна присутність b-HLH-транскрипційного фактору, Math (мишачий AFoh1) та інших. Трансформація поперед-ників у волоскові клітини поліпшується в присутності Math1, а негативно на цей про-цес впливає HeS1. Репродукція підтримую-чих клітин у волоскові шляхом генної інже-нерії здійснюється також в присутності Math1 [27].
Nіshimura, Nakagava визначили, що через 7-8 днів після введення плюропотент-них стовбурових клітин в завитку тварин в деяких трансплантованих клітинах спосте-рігається поява маркера глутаматергічних нейронів, що, по-перше, підтверджує факт можливого переходу плюропотентних стов-
бурових клітин в нейрональні, а по-друге, доцільність лікування хворих з сенсоневра-льною приглухуватістю (а можливо, і глу-хотою) з додаванням до терапевтичного комплексу глутаргіну [9, 32].
В експерименті з фетальною нерво-вою системою мишей, отримано незрілі нейронні попередники волоскових клітин, яким притаманно диференціюватись у воло-скові чи підтримуючі клітини внутрішнього вуха [27].
Що таке стовбурові клітини взагалі та який стан експериментальних досліджень стосовно їх використання для лікування пацієнтів з сенсоневральною приглухуватіс-тю?
Стовбурові клітини (СК) – недифере-нційовані клітини, які мають здатність до самовідновлення та можливість трансміто-тичної диференціації. Термін “стовбурова клітина” ввів петербурзький гематолог А. Максимов (1908).
Ці клітини мають унікальні властиво-сті, характерні тільки для ранніх стадій ци-тогенетичного розвитку. Конкретно, вони виглядають так:
1. Високі адаптаційні можливості, які реалізуються за рахунок росту, міграції, здатності утворювати міжклітинні контакти, пов’язані з підвищеною кількістю бластних клітин.
2. Наявність та продукування великої кількості біологічно активних речовин: фа-кторів росту, a-фетопротеїнів, антиоксидан-тів, адаптогенів, протизапальних та бактері-остатичних сполук, пептидів, які стимулю-ють імунокомпетентні клітини.
3. Високу стійкість до гіпоксії за ра-хунок використання гліколізу.
4. Відсутність імунної реакції відторг-нення, оскільки в І та ІІ триместрах гестації ще не експресуються білки головного ком-плексу гістосумісності І та ІІ класів.
5. Стерильність (плід, знаходячись у матці, захищений від дії мікроорганізмів).
Лікування за допомогою ембріональ-них тканин включає в себе специфічні (за-міщуючі) та неспецифічні механізми, які ґрунтуються на модуляції процесів регене-рації (репарації), проліферації та диференці-ації, і реалізуються на генетичному та епі-геномному рівнях. Розкриття цих механіз-

78 Журнал вушних, носових і горлових хвороб, №2, 2016
мів може бути вирішальною умовою для розробки нових методів терапії хворих з патологічними станами, пов’язаними з по-рушенням морфогенезу.
Трансплантація стовбурових клітин в Україні регламентується Законом України «Про трансплантацію органів та інших ана-томічних матеріалів людини» від 16.07.1999 р. Цим законом, а також Постановою кабі-нету міністрів України від 5 вересня 2007 р. № 1100 «Про заходи щодо організації дія-льності закладів охорони здоров’я та науко-вих установ, пов’язаної з трансплантацією органів, тканин і клітин» покладено відпо-відальність за вилучення та трансплантацію ембріональної стовбурової тканини на Ко-ординаційний центр трансплантації органів.
До захворювань, при яких в Україні вже можливе, а в деяких країнах вже засто-совуються ембріональні клітини та тканини зрілих СК належать:
- хронічні хвороби центральної та пе-риферичної нервової системи (паркінсонізм, хвороба Альцгеймера, розсіяний склероз та таке інше);
- тяжка епілепсія, дитячий церебраль-ний параліч, пігментна дегенерація сітківки;
- важка патологія печінки та підшлун-кової залози (гепатити, цукровий діабет та ін.);
- злоякісні пухлини. На початку стовбурові клітини (СК)
стали виділятись в особливу категорію, зважаючи на їх здатність до самопідтри-мання (асиметричного мітозу) і можливість давати початок клонам диференційованих клітин. Пізніше виявились й інші відмінні властивості СК, що послужило поштовхом до наступних шляхів застосування СК в клініці. Відмінність системи стовбурових клітин від інших систем організму, їх уніка-льність та пластичність, полягають у:
- просторово-часовій моделі функціо-нування (фаза спокою і фаза симетричного або асиметричного спокою);
- неспеціалізованості; - потентності; - здатності до самовідновлення; - здатності до міграції, диференціації,
трансформації; - солітарній дисемінації в тканинах; - ієрархічній підкорюваності;
- тривалості життя [4]. Розрізняється декілька типів стовбу-
рових клітин. Перш за все, це ембріональні та дорослі (від дорослого організму) стов-бурові клітини. Дорослі стовбурові клітини (реґіонарні стовбурові клітини) – недифере-нційовані клітини, які знаходяться серед зрілих диференційованих клітин тканини чи організму, які мають здатність до самовід-новлення та диференціювання, даючи поча-ток спеціалізованим типам клітин даної тканини чи органа [4]. Основна функція дорослих СК в організмі полягає тільки у відновленні тканини, в якій вони знаходять-ся.
Дорослі СК в малій кількості іденти-фіковані в головному мозку, в кістковому мозку, кровоносних судинах, периферичній крові, скелетних м’язах, шкірі, печінці.
Ембріональні стовбурові клітини (ЕСК) – це клітини, похідні від бластоцис-тів. На відміну від спеціалізованих сомати-чних клітин, ЕСК зберігають унікальну ге-нетичну потенцію без спеціалізації. Ці клі-тини мають так званий мінімальний фено-тип та мінімум рецепторів і програм для взаємодії з мікрооточенням, оскільки тільки 5 % з 500 генів транссигналізації експресо-вані в проліферуючих ЕСК [3].
Нейрогенна стовбурова клітина (НСК), або стовбурова клітина ЦНС – це клітина, яка має потенцію диференціюва-тись в нейрони, астроцити і олігодендроци-ти та самовідновлюватись в достатній кіль-кості для забезпечення мозку [29].
Bodson та співавтори в лінійних екс-периментах in vitro показали, що нейрони і глія можуть бути отримані із загального ембріонального попередника; ЦНС дорос-лих також містить мультипотентні поперед-ники для нейронів, астроцитів, олігоденд-роцитів [20].
Термін «прогеніторна клітина» відно-ситься до клітин з більш обмеженим потен-ціалом, ніж стовбурова. Термін «клітина-попередник» відноситься до будь-якої клі-тини, яка знаходиться на більш ранній ста-дії, ніж попередня.
Мозок плодів І триместру вагітності на 90 % складається з ембріональних проге-ніторних клітин [8]. СК присутні у вентри-кулярній зоні і корі ембріонального мозку

Журнал вушних, носових і горлових хвороб, №2, 2016 79
[36]. Лінії нейрогенних СК, виділених з мо-зку плодів, вбудовуються в нейронні спле-тення мозку дорослого щура. Прогеніторні нейробласти здатні повторно інтегруватись в нейронні мережі ембріона після транспла-нтації.
Р. А. Полтавцева і співавтори [6] ви-даляли головний мозок 8-12 тижневих пло-дів людини (абортивний матеріал) і перево-дили його в суспензію, фенотипували мето-дом проточної флюорометрії, використову-ючи антитіла з такими маркерами: нестинп, віментин, алексин, b-тибулін ІІІ, GFAP, CD56, CD34, N-Cad. Результати цитофлюо-рометричного аналізу первинних суспензій НСК людини продемонстрували значну гетерогенність ембріонального матеріалу. В зв’язку з цим для культивування були відіб-рані суспензії з максимальною часткою сто-вбурових і мультипотентних попередників нейтрального ряду з мінімальною долею зрілих клітин в їх селективному середовищі, на 7-у добу утворились справжні нейросфе-ри. Через 14 діб в культурах були присутні як недиференційовані НСК, так і клітини, які диференціювались за нейтральним та гліальним типом.
Нейрональні прогеніторні клітини присутні в постнатальному мозку людини і ссавців, однак кількість зон мозку, в яких відбувається генерація нейронів, лімітована.
Джерелами НСК у дорослих особин є: - cубвентрикулярна зона головного
мозку (нейрональні та гліальні попередни-ки);
- епітелій нюхового тракту і нюхова цибулина;
- зубчасте ядро гіпокампа; - асоціативна кора (мавпи); - мозочок; - голосові центри кори стріатума (пта-
хи). Найбільш вивченою нейрогенератор-
ною ділянкою мозку ссавців є субвентрику-лярна зона латеральних шлуночків. Серед клітинних елементів проліферативних ост-рівців виділяються центрально розташовані проліферуючі нейробласти (А-клітини), слабопроліферуючі великі клітини з астро-цитарними маркерами (В-клітини), а також активні ампліфаєрні прогенітори (С-клітини) [40].
Якщо взяти до уваги дані про функці-онування системи НСК в зрілому організмі, доцільним здається активне вивчення та розробка методів впливу НСК при невроло-гічних чи нейрохірургічних захворюваннях [41], а також їх імплантації в пошкоджені ділянки нервової системи [39]. В організм людини НСК вводяться як системно, розра-ховуючи на хоумінг, так і локально (що, на нашу думку, на сьогодні здається більш доцільним з огляду на можливості трансди-ференціації між окремими представниками СК).
Для вирощування культури та отри-мання необхідних СК з певною потентністю можна використовувати не лише ембріона-льний (клонований) аутогенний матеріал, а й отримані пункційним методом власні (ау-тогенні) СК, не обов’язково нейронального типу. Це створює можливість анонкогенної трансформації культивованих НСК і напів-диференційованих клітин, а також генеру-вання допоміжних клітин – вказівників спрямованого росту закінчень імплантова-них нейроцитів. Таким чином, відкривають-ся багатообіцяючі перспективи отримання і подальшого використання НСК для терапії ряду хворих.
В Україні перші спроби застосування трансплантації нейрогенних клітин при нас-лідках ішемічного ураження головного моз-ку проводились при вивченні відновлюючо-го потенціалу ембріональної нервової тка-нини (ЕНТ) [16].
ЕНТ після трансплантації в зону іше-мії сприяла зменшенню її вираженості, виявила протекторну дію на нейрони реци-пієнта та потенціювала регенераційні про-цеси В.І. Цимбалюк, А.Т. Носов і співавто-ри [12, 16]. Вони, в експерименті на щурах, у яких моделювався гострий ішемічний інсульт, показали, що трансплантована ембріональна нервова тканина (ТЕНТ) в ранній період експериментального ішеміч-ного інсульту сприяє швидкому і більш повному регресу неврологічного дефіциту. Алотрансплантація ЕНТ виконувалась шляхом інтракортикального введення чи аплікації в безсудинну ділянку ішемізова-ної кори півкулі великого мозку. Важливо, що у тварин з ішемією легкого ступеня ТЕНТ не показала суттєвого впливу на від-

80 Журнал вушних, носових і горлових хвороб, №2, 2016
новлювальні процеси. В той же час у тва-рин з ішемією середнього та тяжкого сту-пеня ТЕНТ сприяла більш інтенсивному регресу неврологічних розладів.
Макроскопічно ділянки транспланта-ції характеризувались наявністю інтенсив-ного росту судин в напрямку ЕНТ, при цьо-му розміри самого трансплантату поступово зменшувались протягом 1-3 тижнів після трансплантації. Повне приживлення донор-ської тканини спостерігалось лише у 1 з 56 тварин. При морфологічному дослідженні визначено, що на 7-й день після моделю-вання ішемічного інсульту зменшення кіль-кості клітин тканини пошкодженої півкулі великого мозку досягає максимуму. Через місяць після ТЕНТ загальна кількість клітин в порівнянні з такою на 7-у добу після іше-мії збільшувалась в 1,2 рази, тоді як без ТЕНТ цей показник залишався на поперед-ньому рівні [ 13 ].
З кінця 90-х років минулого століття ведуться активні дослідження ефективності трансплантації ЕНТ та її похідних при ней-родегенеративних захворюваннях [13, 14, 17, 24]. До цього часу проводяться дискусії відносно переваги методу клітинної трансп-лантації над методом ТЕНТ в плані ефекти-вності відновлення популяції дофамінергіч-них нейронів головного мозку. Yureki, Fletcher-Turner провели порівняльне дослі-дження ефективності трансплантації дофа-мінергічних нейронів, які були отримані з ЕСК і ЕНТ вентральних відділів незрілого середнього мозку. За допомогою мафітамі-нового тесту автори показали, що алотранс-плантація ЕНТ в даному випадку більш ефективна, ніж ксенотрансплантація дофа-мінергічних нейронів, похідних ЕСК. При цьому і ТН-позитивні клітини, отримані з ЕСК, і ТН-позитивні клітини ЕНТ після трансплантації створювали зв’язки з нейро-нами стріатума [42].
Особливий інтерес для нас являє дос-лідження вивчення СК для визначення мо-жливості відновлення функції внутрішнього вуха. Основу відтворення клітин спірально-го органа було закладено ще в 1975 р., коли Sobrowicz і співавтори описали довгостро-кову органотипову культуру клітин органа Корті у новонародженої миші з детальним викладенням методів секції і вирощування
спірального органа разом із спіральним ган-глієм [38].
На сьогоднішній день з’явилась гіпо-теза, що внутрішнє вухо, в обмеженій кіль-кості, включає стовбурові слухові і вести-булярні клітини [19, 34]. Ці здатні до утво-рення нейросфер і в подальшому з викорис-танням маркерів клітин-попередників мож-на диференціювати волоскові і нервові клі-тини вестибулярного і завиткового лабірин-ту [21, 30, 31]. Стовбурові клітини спіраль-ного ганглію є плюрипотентними з потенці-алом диференціювання на волоскові кліти-ни та нейрони кохлеарного осередку, втрата яких викликає глухоту [27].
Про мультипотентні кохлеарні кліти-ни – попередники нейронів та умови їх ди-ференціації існує обмежена кількість малоі-нформативних відомостей Зокрема, Lin та співавтори (2009) повідомляють, що відразу після народження у мишей в спіральному органі є клітини, здатні до самовідновлення та диференціації у нейроно- та волоскоподі-бні клітини. Диференціація цих клітин у волоскові залежала від сигнального шляху [28].
Zhong (2010) дослідив проліфератив-ний потенціал клітин, отриманих із завитки новонароджених щурів різного віку [43]. Проліферативні клітини були ізольовані від завитки щурят віком 1, 7, 14 днів та ідентифіковані на підставі даних ультра-структури клітин з кожної вікової групи методами проточної цитометрії та трансмі-сійної електронної мікроскопії. Протягом перших двох тижнів після народження кі-лькість проліферативних клітин поступово зменшилась до нуля. Це зниження відбу-лось паралельно з погіршенням стану про-ліферативного потенціалу клітин. Автор підкреслює, що під час диференціювання кохлеарних стовбурових клітин в них з віком поступово зменшується рівень цик-ліну А2, якому належить не абияка роль у модуляції клітинного циклу.
Nishimura K. (2009), вивчав можли-вість використання індукованих плюрипо-тентних стовбурових клітин як джерела трансплантатів для відновлення слухових нейронів спірального ганглію [32]. Дослід-ник простежив за зростанням аксонів від індукованих клітин-попередниць до волос-

Журнал вушних, носових і горлових хвороб, №2, 2016 81
кових. Нейрони, отримані з плюрипотент-них клітин, скеровували аксони до кохлеар-них волоскових. Факт свідчить про те, що плюрипотентні клітини можуть бути вико-ристані як джерела трансплантатів для ре-генерації нейронів спірального ганглію про-тягом тижня.
У деяких трансплантатах Nishimurа виявив глутаматний транспортер 1, який є маркером глутаматергічних нейронів. Це дуже цікаво з точки зору обґрунтування використання глютаматів в комплексному лікування хворих з сенсоневральною приг-лухуватістю.
Rak та співавтори (2011) дослідили стовбурові клітини з нейросфер завитки, які отримувались у щурів віком в один день. Шляхом гістологічних досліджень вони довели їх здатність до самооновлення і ди-ференціювання до попередників нейронної лінії. Стовбурові клітини виявлялись мето-дом підрахунку та аналізу після їх специфі-чного визначення. Нейронні клітини-попередники позначались двома різними маркерами: Nestin і Atoh1. Автори зробили висновок, що ізольовані із спірального ган-глію клітини мають усі ознаки нейронних стовбурових клітин. Прогнозується, що во-ни можуть стати найкращими транспланта-тами для заміни втрачених сенсорних клі-тин у дорослих з подальшим відновленням слуху [35].
Такахасі і Яманакі в 2006, вивчаючи можливості ембріональних та дорослих сто-вбурових клітин для заміни пошкоджених сенсорних елементів завитки, на основі нау-кових досягнень запропонували стовбурові та індуковані плюропотентні клітини вико-ристовувати з терапевтичною метою. Зруч-ний шлях індукції стовбурових плюрипоте-нтних клітин у глухих було використано під час кохлеарної імплантації. Цим доступом, в подальшому, на нашу думку зможуть ко-ристуватись отоларингологи для трансплан-тації та лікувального введення СК з їх пода-льшою диференціацією та трансформацією у волоскові [22].
Стовбурова терапія швидко розвива-ється і, на наше переконання, має великий потенціал застосування при сенсоневраль-них ураженнях, в першу чергу, у хворих з приглухуватістю та глухотою. Перші спро-
би вивчення можливості СК при функціо-нальних порушеннях внутрішнього вуха були проведені Nakagawa [32]. Він показав, що трансплантовані у внутрішнє вухо НСК можуть вижити і диференціюватись у фено-типи нейронних клітин, гліальних та волос-кових.
Han та співавтори (2009) дослідили, що трансплантація СК у внутрішнє вухо ссавців попереджає ототоксичну дію ген-таміцина. Подібні дані були отримані нами Г.Е. Тімен та співавт. [9, 10, 11], причому ця властивість притаманна не тільки СК, а й витяжці з них (трофіну). Морфологічні дані свідчали, що багаторазове введення глутаргіну та трофіну (після останньої ін’єкції гентаміцину) як одразу, так і через 2 тижня, нейтралізує ототоксичну дію ген-таміцину.
Штучно викликаний у ссавців (морсь-кі свинки) гентаміцином гострий аміноглі-козидний ототоксикоз практично у 100 % тварин створював сенсоневральні порушен-ня. Надалі метою роботи було вивчення дії нейрональних стовбурових клітин (НСК) при різноманітних методах їх введення на морфо - функціональний стан внутрішнього вуха у морських свинок при гострій експе-риментально викликаній сенсоневральній приглухуватості.
Експеримент проведено на 60 статево-зрілих морських свинках масою тіла 250-600 г, які були розподілені на дев’ять груп:
1-а група (контрольна) – тваринам вводився фізіологічний розчин; через 14 та на 22-й день досліджувався слух;
2-а група – вводились НСК інтрапери-тонеально в перший день чотирнадцятиден-ного застосування гентаміцину;
3-я група – НСК вводились інтрапери-тонеально на 8-й день чотирнадцятиденного використання гентаміцину.
В 5-й групі вивчалось інтратимпана-льне введення НСК в перший день чотирна-дцятиденного призначення гентаміцину, та в 6-й групі – на 8-й день з наступним семи-денним відпочинком .
До 4 та 7-ї груп входили тварини, яким вводився 14 днів гентаміцин внутріш-ньом’язово. Об’єктивне дослідження слуху в 4-й групі проводилось до та після 14 днів введення, а у 7-й – після 21-ї доби.
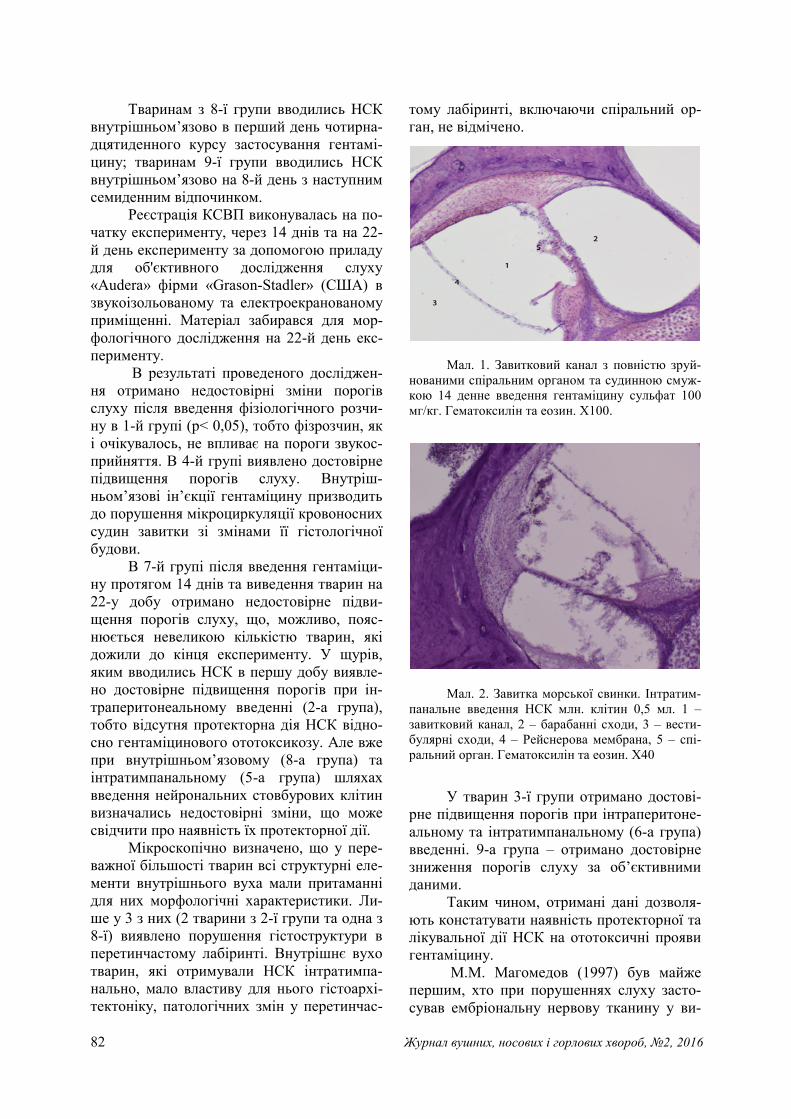
82 Журнал вушних, носових і горлових хвороб, №2, 2016
Тваринам з 8-ї групи вводились НСК внутрішньом’язово в перший день чотирна-дцятиденного курсу застосування гентамі-цину; тваринам 9-ї групи вводились НСК внутрішньом’язово на 8-й день з наступним семиденним відпочинком.
Реєстрація КСВП виконувалась на по-чатку експерименту, через 14 днів та на 22-й день експерименту за допомогою приладу для об'єктивного дослідження слуху «Audera» фірми «Grason-Stadler» (США) в звукоізольованому та електроекранованому приміщенні. Матеріал забирався для мор-фологічного дослідження на 22-й день екс-перименту.
В результаті проведеного досліджен-ня отримано недостовірні зміни порогів слуху після введення фізіологічного розчи-ну в 1-й групі (p< 0,05), тобто фізрозчин, як і очікувалось, не впливає на пороги звукос-прийняття. В 4-й групі виявлено достовірне підвищення порогів слуху. Внутріш-ньом’язові ін’єкції гентаміцину призводить до порушення мікроциркуляції кровоносних судин завитки зі змінами її гістологічної будови.
В 7-й групі після введення гентаміци-ну протягом 14 днів та виведення тварин на 22-у добу отримано недостовірне підви-щення порогів слуху, що, можливо, пояс-нюється невеликою кількістю тварин, які дожили до кінця експерименту. У щурів, яким вводились НСК в першу добу виявле-но достовірне підвищення порогів при ін-траперитонеальному введенні (2-а група), тобто відсутня протекторна дія НСК відно-сно гентаміцинового ототоксикозу. Але вже при внутрішньом’язовому (8-а група) та інтратимпанальному (5-а група) шляхах введення нейрональних стовбурових клітин визначались недостовірні зміни, що може свідчити про наявність їх протекторної дії.
Мікроскопічно визначено, що у пере-важної більшості тварин всі структурні еле-менти внутрішнього вуха мали притаманні для них морфологічні характеристики. Ли-ше у 3 з них (2 тварини з 2-ї групи та одна з 8-ї) виявлено порушення гістоструктури в перетинчастому лабіринті. Внутрішнє вухо тварин, які отримували НСК iнтратимпа-нально, мало властиву для нього гістоархі-тектоніку, патологічних змін у перетинчас-
тому лабіринті, включаючи спіральний ор-ган, не відмічено.
Мал. 1. Завитковий канал з повністю зруй-
нованими спіральним органом та судинною смуж-кою 14 денне введення гентаміцину сульфат 100 мг/кг. Гематоксилін та еозин. Х100.
Мал. 2. Завитка морської свинки. Інтратим-
панальне введення НСК млн. клітин 0,5 мл. 1 – завитковий канал, 2 – барабанні сходи, 3 – вести-булярні сходи, 4 – Рейснерова мембрана, 5 – спі-ральний орган. Гематоксилін та еозин. Х40
У тварин 3-ї групи отримано достові-рне підвищення порогів при інтраперитоне-альному та інтратимпанальному (6-а група) введенні. 9-а група – отримано достовірне зниження порогів слуху за об’єктивними даними.
Таким чином, отримані дані дозволя-ють констатувати наявність протекторної та лікувальної дії НСК на ототоксичні прояви гентаміцину.
М.М. Магомедов (1997) був майже першим, хто при порушеннях слуху засто-сував ембріональну нервову тканину у ви-

Журнал вушних, носових і горлових хвороб, №2, 2016 83
гляді гомогенату ембріональних клітин. Спостереження проведені у 14 хворих з нейросенсорною приглухуватістю ІІ-ІІІ сту-пенів та давністю захворювання від 6 міс до 3 років. Гомогенат вводився ендолюмбаль-но (у 3), меатотимпанально (у 4), ендоназа-льно (у 6), в одному випадку – в перилімфа-тичний простір крізь основу стремена. Піс-ля одноразового застосування відбулося покращання слуху за даними тональної по-рогової аудіометрії, в середньому на 10 дБ, та аудіометрії в розширеному діапазоні час-тот, на 5 дБ. Але акустичний рефлекс та отоакустична емісія в процесі лікування не змінилися. При порівнянні ефективності терапії (тривалість спостереження – 1 рік) методи ендолюмбальної та інтракохлеарної трансплантації, на думку автора, виявилися найбільш ефективними. На жаль, мабуть, сам М.М. Магомедов не знав, що за клітини були в так званому гомогенаті (були там СК і скільки?), з одного боку, а з другого, ре-зультати, наведені автором, не є достовір-ними. Може тому ніхто, та і він сам, не на-важився повторювати запропонований ме-тод лікування, але перша проба введення гомогенату ембріональних клітин безпосе-редньо ендолюмбально відбулася, що, як факт, не може заперечуватись.
Дослідження посттрансплантаційної диференціації та інтеграції НСК в структу-рах внутрішнього вуха виконали Faijoo і співавтори (2011). Вони проводили ало-трансплантацію НСК у внутрішнє вухо ми-ші, при цьому спостерігали інтеграцію НСК тільки в складі сенсорного епітелію вестибу-лярного апарата, але не спірального органа. Але в тому ж році в експериментах на твари-нах доведено, що трансплантація НСК у вну-трішнє вухо викликає їх диференціацію. В подальшому автори інтегрували мічені клі-тини в ділянки внутрішнього вуха, порівню-вались наслідки алотрансплантації мічених НСК в порожнину бокового півколового каналу (перший шлях) і безпосередньо в по-рожнину кохлеарного каналу через бокову стінку завитки (другий шлях). Через 2 тижні автори вивчали функцію слухового аналіза-тора (електрична активність нейронів стов-бура у відповідь на подразнення звуковим стимулом периферичних відділів аналізато-ра). Трансплантація НСК першим шляхом –
мічені клітини виживали і виявлялись в клі-тинах scala tympani і scala vestibulаe. При цьому не відбувалось погіршення функції слухового аналізатора. В той же час введен-ня НСК через бокову стінку завитки приво-дило до значної втрати слуху в дослідних тварин, (мабуть, причиною була травма?), не дивлячись на те, що трансплантовані клітини розташовувались всередині scala media. Igushi вважає, що переважаючим методом трансплантації НСК в лабіринт є введення їх суспензії в порожнину бокового півколового каналу. Ми думаємо, що обидва шляхи в клініці навряд чи знайдуть застосування.
В лабораторії культивування тканин Інституту нейрохірургії НАМН України ім. А.П. Ромоданова було проведено експери-ментальне дослідження впливу ембріональ-ної нервової тканини на пошкоджений сен-сорний епітелій внутрішнього вуха щурів. Експерименти проводились на 22 білих не-лінійних щурах віком 1-2 тижні з середньою масою тіла 50 г. Спіральний орган щурів видалявся з внутрішнього вуха в стериль-них умовах методом Я.А. Віннікова, Л.К. Титової [2]. Ембріональна нервова тканина (ЕНТ) отримувалась з тканини головного мозку у ембріонів щурів пренатального пе-ріоду. Дисоціація ЕНТ виконувалась за ме-тодом Mosconа у модифікації Вікторова.
Досліди показали, що в такому варіа-нті експериментів епітеліальний компонент значною мірою зберігає типове розташу-вання та побудову більшості клітин. Резуль-тати проведених експериментів з експлан-тацією клітинного матеріалу з внутрішнього вуха щурів свідчили про можливість його приживлення протягом 7 діб зі збереженням життєздатності експлантатів та характерної морфології, що відповідає органотиповим особливостям.
Таким чином, експериментальні дос-лідження останніх десятиріч стосовно дії СК та можливості їх трансплантування у внутрішнє вухо з метою заміщення ушко-джених структур або доставки лікарських засобів безпосередньо у внутрішнє вухо, стимуляції трансдифференціації чи мітозів клітин-попередників, з нашого погляду є перспективними напрямками в наукових розробках для попередження і, головне, лікування пацієнтів з сенсоневральними

84 Журнал вушних, носових і горлових хвороб, №2, 2016
хворобами внутрішнього вуха. Але серед інших умов успішного клінічного застосу-вання ЕНТ необхідним після всебічного експериментального обґрунтування будуть
критерії відбору хворих до трансплантації з урахуванням зворотності захворювання та глибини патологічних змін у сенсоневраль-ному епітелії внутрішнього вуха.
Література 1. Викторов И.В., Божкова В.П. Руководство по куль-
тивированию нервной ткани: Методы. Техника. Проблемы // Наука. – 1988. – С. 315.
2. Винников Я.А., Титова Л.К. Кортиев орган. – Ле-нинград: Изд. Акад. наук СССР, 1961. – 260 с.
3. Гордеева О.Ф., Мануилова Е.С., Гуляева Д.В. и со-авт. Морфологические аспекты дифференцировки эмбриональных стволовых клеток в культуре // Ци-тология. Cytology. – 2011. – Т.43, №9. – С. 851.
4. Запорожан В.Н., Бажора Ю.И. Стволовые клетки. – Одесса: Одес. гос. мед. университет, 2004. – С. 228.
5. Магомедов М.М. Применение фетальных тканей в терапии хронической нейросенсорной тугоухости // Матеріали міжнародної науково-практичної конфе-ренції, присвяченої 75-річчю кафедри та клініки отоларингології Дніпропетровської медичної ака-демії. – Дніпропетровськ, 1997. – С. 140-141.
6. Полтавцева Р.А., Ржанинова А.А., Ревищин А.В., Александрова М.А., Корочкин Л.И., Репин В.С., Сухих Г.Т. Развитие in vitro нейтральных прогени-торных клеток эмбрионов человека // Бюл. экспе-рим. биол. и медицины. – 2001. – №9. – С. 290-293.
7. Репин В.С. Медицинская клеточная биология: но-вые фундаментальные и прикладные исследования // Трансплантация фетальных тканей и клеток чело-века. – М., 1996. – С. 19-27.
8. Сухих Г.Т. Трансплантация фетальных клеток: на-стоящее и будущее // Бюл. эксперим. биол. – 1998. – Т.126, прил. 1. – С. 3-13.
9. Тімен Г.Е, Цимбалюк В.І., Бєлоусова А.О, Вінничук П.В., Чубко С.П., Руденька К.Л. Вивчення дії тро-фіну та глутаргіну на функціональний стан внутрі-шнього вуха при аміноглікозидному ототоксикозі // Журн. вушних, носових і горлових хвороб. – 2011. – № 4с. – С. 123.
10. Тімен Г.Е, Цимбалюк В.І., Бєлоусова А.О, Вінничук П.В., Сапіжак І.І. Вивчення впливу нейрональних стовбурових клітин на морфо-функціональний стан внутрішнього вуха морських свинок після експери-ментально викликаного аміноглікозидного ототок-сикозу // Журн. вушних, носових і горлових хвороб. – 2015. - №3с. - С.137.
11. Тімен Г.Е, Цимбалюк В.І., Семенова В.М., Вінни-чук П.В., Сапіжак І.І. Протекторна дія нейрональ-них стовбурових клітин при гострому аміногліко-зидному ототоксикозі // Журн. вушних, носових і горлових хвороб. – 2014. – №5с. – С.137.
12. Цимбалюк В.І. Нейротрансплантація // Лікування та діагностика. – 2000. – № 3. – С. 15-19.
13. Цимбалюк В.І. Нейротрансплантація // Лікування та діагностика, перспективи розвитку. – 2004. – №2. – С. 19-23.
14. Цимбалюк В.І., Носов А.Т., Боднар Л.В., Васлович В.В. Ефективність нейротрансплантації при лікуванні гострої недостатності мозкового кровообігу в експерименті // Бюл. УАН. – 1998. – № 6. – С. 13-14.
15. Цимбалюк В.І., Пічкур Л.Д., Вербовська С.А. Ре-зультати нейротрансплантації у хворих на дитячий церебральний параліч // Укр. нейрохірург. журн. – 2001. – № 2(14). – С. 90.
16. Чехонин В.П., Баклушев В.П., Белопасов В.В., Дмитриева Т.Б. Трансплантация эмбриональной нервной ткани в терапии паркинсонизма: совре-менное состояние проблемы // Журн. неврологи и психиатрии. – 1999. – Т.99, № 11. – С. 60-66.
17. Baird R.A., Torres M.A. Hair cell regeneration in the bullfrog vestibular otolith organs foiiwing aminoglyco-side toxity / Ann. NY Acad. – 1993. – P. 1-12.
18. Berminham-McDonogh O. Regulated reprogramming in the regeneration of sensory receptor cells / O. Ber-minham-McDonogh, T.A. Reh // Neuron. – 2011. – Vol. 71(3). – P. 389-405.
19. Bodson M. Hair cell progenitors: identification and regulatory genes / M. Bodson, I. Breuskin, P. Lefebvre [et al.] // Acta Otolaryngol. – 2010. – Vol. 130(3). – P. 312-317.
20. Groves A.K. The challenge of hair cells regeneration / A.K. Groves // Exp. Biol. Med. (Maywood). – 2010. – Vol. 235(4). – P. 434-446.
21. Gunewarde N. The convergence of cochlear implanta-tion with induced pluripotent stem cell therapy / N. Gunewarde, M. Dottori, B.A. Nayagam // Stem Cell Rev. – 2012. – Vol. 8(3). – P. 741-754.
22. Han Y. The expression of NOB 1 in spiral ganglion cells of guinea pig / Y. Han, L. Hong // Int. J. Pediatr. Otorhinolaryngol. – 2009. – Vol. 73(2). – P. 315-319.
23. Isacson O. The production and use of cells as therapeu-tic agents in neurotic generative diseases // LancetNeu-rol. – 2003. – V.2. – P. 417-424.
24. Katayama A., Corvin J.T. Cell production in chicken cochlea // J. Comp. Neurol. – 1989. – 281. – P. 129-135.
25. Kelley M.W. Cellular commitment and differentiation in the organ of Corti / M.W. Kelley // Int. J. Dev. Biol. – 2007. – Vol. 51(6-7). P. 571-583.
26. Li H.J. Pluripotent stem cells from the adult mouse in-ner ear / H. Li, H. Liu, S. Heller // Nat. Med. – 2003. – Vol. 9(10). – P. 1293-1299.
27. Lin J. Directed differentiation of mouse cochlear neural progenitors in vitro / J. Lin, L. Feng // Am. J. Phisiol. Cell Physiol. – 2009. – Vol. 296(3). – P. 441-452.
28. Malgrange B. Differentiation, protection and regenera-tion of hair cells and auditory neurons in mammals / B. Malgrange // Bull. Mem. Acad. R. Med. Belg. – 2005. – Vol. 160(5-6). – P. 276-286.
29. Martinez-Monedero R. Differentiation of inner ear stem cells to functional sensory neurons / R. 30. Marti-nez-Monedero, E. Yi, K. Oshima [et al.] // Dev. Neuro-biol. – 2008. – Vol. 68(5). – P. 669-684.
30. McKay R. Stem cells in the central nervous system // Sciense. – 1997. – V.276. – P. 66-71.
31. Nishimura K. Transplantation of mouse induced pluri-potent stem cells in to the cochlea / K. Nishimura, T.

Журнал вушних, носових і горлових хвороб, №2, 2016 85
Nakagawa, K. Ono [et al.] // Neuroreport. – 2009. – Vol. 20(14). – P. 1250-1254.
32. Ohtsuka T., Sakamoto M., Guillemot F. Roles of the basic helixloop-helix genes Hes1 and Hes5 in expan-sion of neural stem cells of the developing brain // J. Biol. chem. – 2001. – V.276, №32. – P. 30467-30474.
33. Oshima K. Mechanosensitive hair cell-like cells from embryonic and induced pluripotent stem cells / K. Oshima , K. Shin , M. Diensthuuber [et al.] // Cell. – 2010. – Vol. 14(4). – P. 704-716.
34. Rak K. Isolation and characterization of neuronal stem cells from the neonatal rat cochlear nucleus // K. Rak, N.V. Wasielewski, A. Radeloff [et al.] // Cell Tissue Res. – 2011. – Vol. 34(3). – P. 499-508.
35. Reynolds B.A., Weyss S. Generation of neuron and astrocytes from isolated cells of the adult mammalian central nervous system // Science. – 1992. V.255. – P.1707-1710.
36. Rubel E., Dew L.A., Roberson D.W. Mammalian ves-tibular hair cell regeneration // Science. – 1995. – Vol. 267. – P. 701 -703.
37. Sobkowicz H.M. Tissue culture of the organ of Corti / H.M. Sobkowicz, J.M. Loftus, S.M. Slapnick // Acta Otolaringol. Suppl. – 1993. – Vol. 502. – P. 3-36.
38. Svedsen C.N., Smith A.G. New prospects for human stem cell therapy in the nervous system // TNIS.-1999. – V.22, №8. – P.357-364.
39. Temple S. The development of neural stem cells // Na-ture. – 2001. – V.414, №6859. – P.112-117.
40. Wagner J.P., Black I., KiCico-Bloom E. Stimulation of nеonatal and adult brain neurogenesis by subcutaneous ingection of basic Fibroblast growth factor // J. Neuro-sci. – 1999. – V.19, №14. – P. 6006-6016.
41. Yurek D.M., Fletcher-Turner A. Comparison of em-bryonic stem cell-derived dopamine neuron grafts and fetal ventral mesencephalic tissue grafts: morphology and function // Cell Transplant. – 2004; 13 (3):295-306.
42. Zhong C. A comparison o the proliferative capacity and ultrastructure of proliferative cells from the cochleae of newborn rats of different ages / C. Zhong, Y. Han. // Int. J. Pediatr. Otorhinolaringol. – 2010. – Vol. 74(2). – P. 192-197.
References 1. Vyktorov IV, Bozhkova VP. Guide to nervous tissue
cultures: methods. Technology. Problems. Science. 1988:315. Russian.
2. Vynnykov JA, Titova LK. Organ of Corty. Leningrad: Ed. Academy of Sciences USSR; 1961. 260 p. Russian.
3. Gordeev OF, Manuylova ES, Hulyaeva DV. Morpho-logical aspects of stem cell differentiation in culture. Cytology. 2011;43(9):851. Russian.
4. Zaporozhan VN, Bazhora YI. Stem cells. Odessa: Odessa state med University; 2004:228. Russian.
5. Magomedov M. Application of fetal tissue in sensori-neural hearing loss therapy. Proceedings of the interna-tional scientific-practical conference on the 75th anni-versary of the Department and Clinic of Otolaryngolo-gy of Dnepropetrovsk Medical Academy. Dnepro-petrovsk; 1997:140-141. Russian.
6. Poltavtsev RA, Rzhanynova AA, Revyschyn AV, Alexandrov MA, Korochkyn LI, Repin VS, Sukhih GT. The in vitro development of neutral progenitor cells of human embryon. Bulletin experym Biol and medicine. 2001;(9):290-3. Russian.
7. Repin VS. Medical cell biology: New fundamental and applied research. Tissue and organ transplantation of fetal tissues and cells. M., 1996. S. 19-27. Russian.
8. Sukhih GT. Transplantation of fetal cells: Present and Future. Bulletin experimental biol. 1998;126 Suppl 1:3-13. Russian.
9. Timen GE, Tsymbaliuk VI, Belousov IA, Vinnychuk PV, Chubko SP, Rudenka KL. Study of trophin and glutargin action on the functional state of the inner ear with aminoglycoside ototoxity. Zhurnal vushnyh, no-sovyh i gorlovyh hvorob. 2011;(4c):123. Ukrainian.
10. Timen GE, Tsymbaliuk VI, Belousov A.O, Vinnychuk PV Sapizhak II. Study of neuronal stem cells in the morpho functional status of the inner ear of guinea pigs after experimentally induced aminoglycoside ototoxity. Zhurnal vushnyh, nosovyh i gorlovyh hvorob. 2015;(4c):137. Ukrainian.
11. Timen GE, Tsymbaliuk VI, Semenov VN, Vinnychuk PV Sapizhak II. Protective effect of neuronal stem cells in acute aminoglycoside ototoxity. Zhurnal vushnyh, nosovyh i gorlovyh hvorob. 2014;(5c):137. Ukrainian.
12. Tsymbaliuk VI. Neyrotransplantatsiya. Treatment and diagnosis. 2000;(3):15-9. Ukrainian.
13. Tsymbaliuk VI Neyrotransplantatsiya. Treatment and diagnosis prospects. 2004. - №2. - P. 19-23.
14. Tsymbaliuk VI, Nosov AT, Bodnar L, Vaslovych V. Neurotransplantation efficiency in the treatment of acute failure of cerebral circulation in the experiment. Bull UAS. 1998;(6):13-4. Ukrainian.
15. Tsymbaliuk VI, Pichkur LD, Verbovska SA. Results of neurotransplantation in children with cerebral palsy. Ukr neurosurgery Zh. 2001;14(2):90. Ukrainian.
16. Chekhonin VP, VP Baklanov, Belopasov VV, Dmitrieva TB. Transplantation of embryonic neural tissue in the treatment of Parkinson's disease: state of the art. Zh. neurology and psychiatry. 1999;99(11):60-6. Russian
17. Baird RA, Torres MA. Hair cell regeneration in the bullfrog vestibular otolith organs following aminogly-coside toxity. Ann NY Acad; 1993:1-12.
18. Berminham- McDonogh O, Reh TA. Regulated repro-graming in the regeneration of sensory receptor cells. Neuron. 2011;71(3):389-405.
19. Bodson M, Breuskin I, Lefebvre P, Malgrange B. Hair cell progenitors: identification and regulatory genes. Acta Otolaryngol. 2010 Mar;130(3):312-7.
20. Groves A.K. The challenge of hair cells regeneration. Exp Biol Med (Maywood). 2010;235(4):434-46.
21. Gunewarde N. The convergence of cochlear implanta-tion with induced pluripotent stem cell therapy. N. Gunewarde, M. Dottori, B.A. Nayagam. Stem Cell Rev. 2012;8(3):741-754.
22. Han Y, Hong L. The expression of NOB 1 in spiral ganglion cells of guinea pig. Int J Pediatr Otorhinolaryngol. 2009;73(2):315-19.
23. Isacson O. The production and use of cells as therapeu-tic agents in neurotic generative diseases. Lancet Neurol. 2003;2:417-24.
24. Katayama A, Corvin JT. Cell production in chicken cochlea. J Comp Neurol. 1989;281:129-35.
25. Kelley MW. Cellular commitment and differentiation in the organ of Corti. Int J Dev Biol. 2007;51(6-7):571-83.

86 Журнал вушних, носових і горлових хвороб, №2, 2016
26. Li HJ, Liu H, Heller S. Pluripotent stem cells from the adult mouse inner ear. H LiNat Med. 2003;9(10):1293-9.
27. Lin J, Feng L. Directed differentiation of mouse coch-lear neural progenitors in vitro. J LinAm J Phisiol. Cell Physiol. 2009;296(3):441-52.
28. Malgrange B. Differentiation, protection and regenera-tion of hair cells and auditory neurons in mammals. Bull Mem Acad R Med Belg. 2005;160(5-6):276-86.
29. Martinez-Monedero R, Yi E, Oshima K, Glowatzki E, Edge AS. Differentiation of inner ear stem cells to functional sensory neurons. Dev Neurobiol. 2008 Apr;68(5):669-84.
30. McKay R. Stem cells in the central nervous system. Sciense. 1997;276:66-71.
31. Nishimura K, Nakagawa T, Ono K, Ogita H, Sakamoto T, Yamamoto N, Okita K, Yamanaka S, Ito J. Trans-plantation of mouse induced pluripotent stem cells into the cochlea. Neuroreport. 2009 Sep 23;20(14):1250-4.
32. Ohtsuka T, Sakamoto M, Guillemot F. Roles of the ba-sic helixloop-helix genes Hes1 and Hes5 in expansion of neural stem cells of the developing brain. J Biol chem. 2001;276(32):30467-74.
33. Oshima K, Shin K, Diensthuber M, Peng AW, Ricci AJ, Heller S. Mechanosensitive hair cell-like cells from embryonic and induced pluripotent stem cells. Cell. 2010 May 14;141(4):704-16.
34. Rak K, Wasielewski NV, Radeloff A, Völkers J, Scherzed A, Jablonka S, Hagen R, Mlynski R. Isolation
and characterization of neural stem cells from the neonatal rat cochlear nucleus. Cell Tissue Res. 2011 Mar;343(3):499-508.
35. Reynolds BA, Weyss S. Generation of neuron and as-trocytes from isolated cells of the adult mammalian central nervous system. Science. 1992;255:1707-10.
36. Rubel E, Dew LA, Roberson DW. Mammalian vestibular hair cell regeneration. Science. 1995;267:701-3.
37. Sobkowicz HM, Loftus JM, Slapnick SM. Tissue cul-ture of the organ of Corti. Acta Otolaringol. Suppl. 1993;502:3-36.
38. Svedsen CN, Smith AG. New prospects for human stem cell therapy in the nervous system. TNIS. 1999;22(8):357-64.
39. Temple S. The development of neural stem cells. Nature. 2001;414(6859):112-7.
40. Wagner JP, Black I, KiCico-Bloom E. Stimulation of nеonatal and adult brain neurogenesis by subcutaneous ingection of basic Fibroblast growth factor. J Neurosci. 1999;19(14):6006-16.
41. Yurek DM, Fletcher-Turner A. Comparison of em-bryonic stem cell-derived dopamine neuron grafts and fetal ventral mesencephalic tissue grafts: morphology and function. Cell Transplant. 2004;13(3):295-306.
42. Zhong C, Han Y. A comparison o the proliferative ca-pacity and ultrastructure of proliferative cells from the cochleae of newborn rats of different ages. Int J Pediatr Otorhinolaringol. 2010;74(2):192-7.
Надійшла до редакції 21.10.15.
© Г.Е. Тімен, І.І. Сапіжак, Л.А. Кудь, 2016
СЕНСОНЕВРАЛЬНАЯ ТУГОУХОСТЬ И СТВОЛОВЫЕ КЛЕТКИ
Тимен Г.Э., Сапижак И.И., Кудь Л.А (Киев)
А н н о т а ц и я
Фактическое отсутствие эффективного медикаментозного лечения при сенсоневральной тугоухо-сти, несмотря на огромное количество предложенных методов, вынуждает искать новые, оригинальные пути решения мировой проблемы. В последнее десятилетие в экспериментальной иммунологии, эмбрио-логии и особенно трансплантологии успешно разрабатываются методы трансплантации эмбриональных тканей и клеток, которые имеют неслыханные уникальные свойства, характерные только для клеток тка-ней, находящихся на ранних стадиях цитогенетического развития. Лечение с помощью эмбриональной ткани, априори, включает в себя специфические (заместительные) и неспецифические механизмы, кото-рые основываются на модуляции процессов регенерации (репарации), пролиферации и трансдифферен-циации, а также реализовываются на генетическом и эпигеномном уровнях. Раскрытие этих механизмов может быть решающим условием для разработки новых методов терапии больных с патологическими состояниями, обусловленными нарушением морфогенеза. Заданием исследования было изучение морфо-логической структуры тканей внутреннего уха и их функциональных возможностей у морских свинок при разных путях введения нейрональных стволовых клеток (НСК) на фоне аминогликозидной ототоксикации. Результаты, полученные при проведении эксперимента, в частности при введении интратимпанальным путем СНК, разрешают надеяться на лечебный и протекторный эффект у людей с ототоксическими и дру-гими поражениями слуха.

Журнал вушних, носових і горлових хвороб, №2, 2016 87
SENSONEVRAL HEARING LOSS AND STEAM CELLS
Timen G.E., Sapizhak I.I., Kud L.A. (Kiev)
State institution «O.S. Kolomiychenko Institute of Otolaryngology of National Academy of Medical Sciences of Ukraine»; e-mail: [email protected]
A b s t r a c t
Absence of efficiency of sensoneural hearing loss treatment, without regard to plenty of the offered me-thods, makes us searching the new ways of decision of this problem. In the last decade in experimental immunol-ogy, embryology and transplantology methods of transplantation of embryo tissues and cells were successfully developed. Treatment with embryo tissues, includes for itself specific substitute and heterospecific mechanisms, that are based on modulation of processes of regeneration, reparation, proliferation and differentiation. They rea-lized on genetic and epygenic levels. Disclosure of these mechanisms can be a milestone for new therapy methods development targeted on morphogenesis disorders. The task of our research was a morphological structure study of guinea-pigs internal ear tissue at the different ways of introduction of neuronal of stem cells on a background of amynoglycoside intoxication. The results obtained in the experiment give hope for healing and protective effect in patients with ototoxic and other types of hearing loss.








![[Skolkovo Robotics 2015 Day 1] Иванова Г.Е. Реабилитация пациентов с сенсорно-перцептивными нарушениями](https://img.pdfslide.tips/doc/110x75/55aa91ee1a28ab496f8b48e5/skolkovo-robotics-2015-day-1-.jpg)


















