Embed Size (px)
Citation preview

SR
M
p
R
cwT
Csrdr
cp
�-Arrestin2 Regulates Cannabinoid CB1 Receptorignaling and Adaptation in a Central Nervous Systemegion–Dependent Manner
Peter T. Nguyen, Cullen L. Schmid, Kirsten M. Raehal, Dana E. Selley, Laura M. Bohn, andLaura J. Sim-Selley
Background: Cannabinoid CB1 receptors (CB1Rs) mediate the effects of �9-tetrahydrocannabinol (THC), the psychoactive component inmarijuana. Repeated THC administration produces tolerance and dependence, which limit therapeutic development. Moreover, THCproduces motor and psychoactive side effects. �-arrestin2 mediates receptor desensitization, internalization, and signaling, but its role inthese CB1R effects and receptor regulation is unclear.
ethods: CB1R signaling and behaviors (antinociception, hypothermia, catalepsy) were assessed in �-arrestin2-knockout (�arr2-KO) andwild-type mice after THC administration. Cannabinoid-stimulated [35S]GTP�S and [3H]ligand autoradiography were assessed by statistical
arametric mapping and region-of-interest analysis.
esults: �-arrestin2 deletion increased CB1R-mediated G-protein activity in subregions of the cortex but did not affect CB1R binding, invehicle-treated mice. �arr2-KO mice exhibited enhanced acute THC-mediated antinociception and hypothermia, with no difference incatalepsy. After repeated THC administration, �arr2-KO mice showed reduced CB1R desensitization and/or downregulation in cerebellum,
audal periaqueductal gray, and spinal cord and attenuated tolerance to THC-mediated antinociception. In contrast, greater desensitizationas found in hypothalamus, cortex, globus pallidus, and substantia nigra of �arr2-KO compared with wild-type mice. Enhanced tolerance to
HC-induced catalepsy was observed in �arr2-KO mice.
onclusions: �-arrestin2 regulation of CB1R signaling following acute and repeated THC administration was region-specific, and resultsuggest that multiple, overlapping mechanisms regulate CB1Rs. The observations that �arr2-KO mice display enhanced antinociceptiveesponses to acute THC and decreased tolerance to the antinociceptive effects of the drug, yet enhanced tolerance to catalepsy, suggest thatevelopment of cannabinoid drugs that minimize CB1R interactions with �-arrestin2 might produce improved cannabinoid analgesics with
educed motor suppression.
cpb(CqrSdgctm
�ittrr
[ic�l�wu
Key Words: Beta-arrestin, cannabinoid receptors, GPCR, statisticalparametric mapping, tolerance
C B1 receptors (CB1Rs) are widely distributed in the centralnervous system (CNS) (1) and mediate the central effects of
�9-tetrahydrocannabinol (THC) and cannabinoids (2). Theendocannabinoid system is implicated in numerous physiologicalprocesses and is a potential therapeutic target for disorders includ-ing neurodegenerative and neuropsychiatric diseases and chronicpain (3). However, therapeutic use is limited by psychoactive andmotor side effects. Moreover, repeated cannabinoid treatment pro-duces tolerance to cannabinoid-mediated in vivo effects (4). Re-gion-specific CB1R desensitization and downregulation occur inconjunction with tolerance (5), but the molecular mechanisms thatunderlie CB1R adaptations and tolerance are not well defined.
CB1Rs primarily activate Gi/o-proteins, which regulate adenylylyclases, ion channels, and kinases (6). Persistent cannabinoid ex-osure induces CB1R uncoupling from G-proteins (desensitization)
(7), with subsequent receptor internalization (8) and degradation(downregulation) (9,10). One mechanism for these adaptations oc-
From the Department of Pharmacology and Toxicology and Institute forDrug and Alcohol Studies (PTN, DES, LJS-S), Virginia CommonwealthUniversity, Richmond, Virginia; and Departments of Molecular Thera-peutics and Neuroscience (CLS, KMR, LMB), The Scripps Research Insti-tute, Jupiter, Florida.
Address correspondence to Laura J. Sim-Selley, Ph.D., Box 980524, 1112 E.Clay Street, Richmond, VA 23298; E-mail: [email protected]
iReceived Aug 4, 2011; revised Oct 25, 2011; accepted Nov 30, 2011.
0006-3223/$36.00doi:10.1016/j.biopsych.2011.11.027
urs by G-protein-coupled receptor (GPCR) kinase (GRK)-mediatedhosphorylation of activated receptors and subsequent �-arrestininding (11). �-arrestin2 is one of two arrestin isoforms in the brain
12), and findings in cell models support a role for �-arrestin2 inB1R adaptations. Coexpression of GRK3 and �-arrestin2 was re-uired for rapid desensitization of CB1R-mediated potassium cur-
ents following exposure to WIN55,212-2 in Xenopus oocytes (8).imilarly, expression of dominant negative �-arrestin2 attenuatedesensitization of WIN55,212-2-mediated inhibition of glutamater-ic neurotransmission in hippocampal neurons (13). Immunohisto-hemical studies show that CB1Rs are codistributed with �-arres-in2 in certain CNS regions (12,14), suggesting that �-arrestin2
ight regulate CB1R signaling in the CNS.Because there are no pharmacological �-arrestin inhibitors,
-arrestin2 knockout (�arr2-KO) mice (15) provide a model to studyts role in regulating GPCRs in vivo (16). Acute administration of THCo �arr2-KO mice revealed enhanced sensitivity to its antinocicep-ive and hypothermic effects (17). However, direct evidence for theole of �-arrestin2 in CB1R adaptations and tolerance followingepeated THC is lacking.
We adapted statistical parametric mapping (SPM) to analyze35S]GTP�S autoradiography (18). SPM has the advantage of assess-ng changes in G-protein activation in an unbiased and anatomi-ally inclusive manner. We applied SPM to examine the role of-arrestin2 in CB1R regulation by comparing cannabinoid-stimu-
ated [35S]GTP�S binding in brains from vehicle and THC-treatedarr2-KO and wild-type (WT) littermates. Combining this approachith behavioral assessment following THC administration alloweds to compare CB R signaling with behavioral responses observed
1n the �arr2-KO mice. We demonstrate region-specific regulation of
BIOL PSYCHIATRY 2012;71:714–724© 2012 Society of Biological Psychiatry

i
isaC
[
P(
dC
([(
�emav
R
T
tbwl.psmFTboieccc
iduptiabFa5FtpTs
�dcFtst3t.glHc[sTd
�
ws
P.T. Nguyen et al. BIOL PSYCHIATRY 2012;71:714–724 715
CB1Rs by �-arrestin2 that parallel changes in THC-mediated behav-or and tolerance.
Methods and Materials
Detailed Methods are provided in Supplement 1.
MiceMale WT and �arr2-KO mice (littermates, 4 months) (15) were
injected intraperitoneally with THC (10 mg/kg) or vehicle (1:1:18ethanol:cremaphor: .9% saline) twice daily for 6.5 days (subchronictreatment). Twenty-four hours after the final injection, mice werechallenged with increasing doses of THC (3, 7, 20, 26, and 44 mg/kg,intraperitoneally) every 40 min, with responses assessed 30 minafter each injection. Studies followed the National Institutes ofHealth Guidelines for the Care and Use of Laboratory Animals.
BehaviorAntinociception was assessed using the warm-water (52°C) tail-
immersion assay (19). Duplicate measurements determined base-line responses, but mice were assessed only once following eachinjection to minimize tissue damage. A trained observer assessedimmobility by determining the time mice spent motionless on ametal ring-stand over 5 min (20). Mice were gently restrained, andbody temperature was measured using a rectal probe thermometer(15). Mice were sacrificed by decapitation 24 hours after testing. Thespinal cord and brain were extracted, frozen, and stored at �80°C.For antinociception and catalepsy, data are presented as the per-centage of the maximum possible effect (%MPE) � 100% � [(exper-mental response latency – basal response latency)/(maximal pos-ible response – basal response latency)]. Nonlinear regressionnalysis was calculated using GraphPad Prism software (La Jolla,alifornia).
35S]GTP�S and [3H]SR141716A BindingWhole spinal cord was collected (Supplement 1), tissue was
homogenized, and agonist-stimulated [35S]GTP�S binding wasconducted as published (10). Concentration-effect curves weregenerated using .01–3 �mol/L CP55,940 or .03–10 �mol/LWIN55,212-2. Percent stimulation � [(agonist-stimulated – basal)/basal] � 100%. Curves were fit using nonlinear regression in Graph-
ad Prism. [3H]SR141716A binding was performed as published21) using [3H]SR141716A (.1–2.5 nmol/L), and nonspecific binding
was measured with 5 �mol/L SR141716A. Data were fit to a one-sitemodel in GraphPad Prism. Statistical comparisons used Student-Newman-Keuls post hoc test.
[35S]GTP�S and [3H]CP55,940 AutoradiographyAgonist-stimulated [35S]GTP�S autoradiography was con-
ucted on duplicate serial sections as published (7,18). Basal andP55,940-stimulated [35S]GTP�S binding were conducted in ad-
jacent sections. CP55,940 is a high-efficacy agonist for G-proteinactivation but does not activate non-CB1 sites in brain sections18). Net stimulation (nCi/g) � (agonist-stimulated – basal).3H]CP55,940 autoradiography was modified from Herkenham et al.1) and Moise et al. (22). Total binding was assessed with 3 nmol/L
[3H]CP55,940 and nonspecific binding was measured using 10mol/L CP55,940. Image reconstructions, SPM, and region-of-inter-st (ROI) analysis were conducted as published (18,23). The ROIeasurements were made on original unprocessed images, aver-
ged across hemispheres, and analyzed by two-way analysis ofariance (significance p � .05) and Student-Newman-Keuls post
hoc comparisons. s
esults
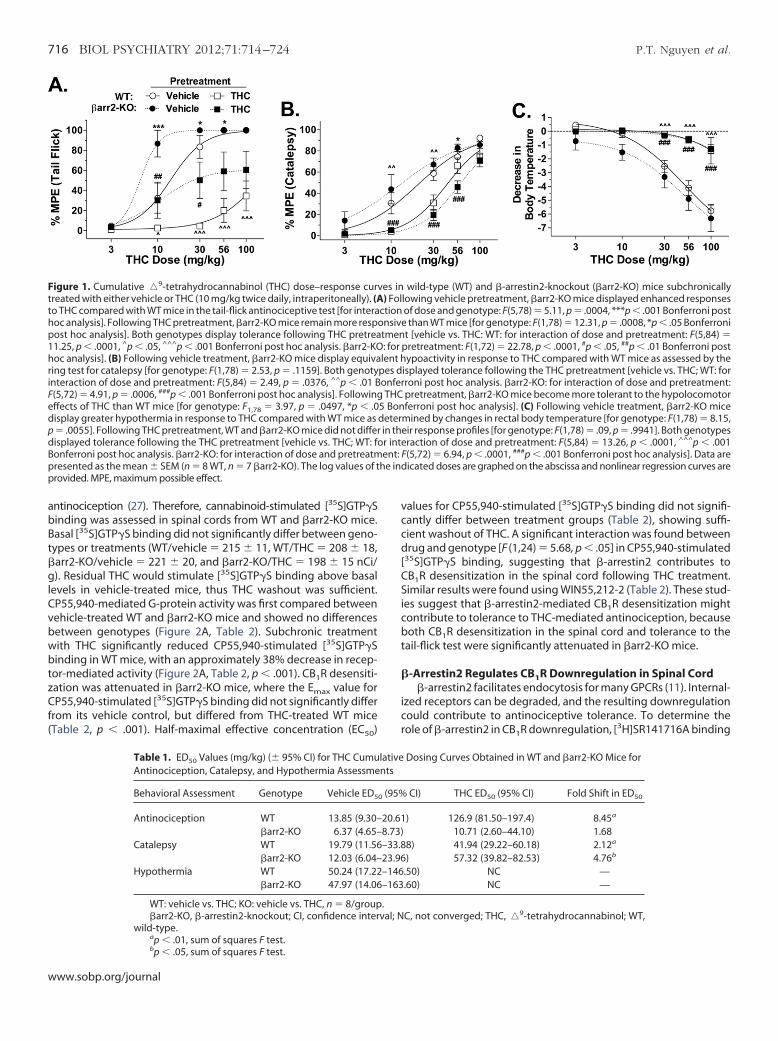
HC-Induced Responses in �arr2-KO MiceTHC-mediated antinociception was assessed in vehicle- or THC-
reated WT and �arr2-KO mice by cumulative dosing of THC. Theasal latencies in WT and �arr2-KO mice subchronically treatedith vehicle were 1.650 � .176 and 1.625 � .251, respectively. Basal
atencies for WT and �arr2-KO mice treated with THC were 1.438 �092 and 1.788 � .210, respectively. Cumulative dosing of THCroduced a greater degree of antinociception in �arr2-KO miceubchronically treated with vehicle, compared with their WT litter-
ates [Figure 1A; for genotype: F (1,78) � 8.95, p � .0037; for dose:(5,78) � 92.00, p � .0001; for interaction: F (5,78) � 5.11, p � .0004].his difference was due to a difference in potency (ED50 value)etween WT and �arr2-KO mice (Table 1). To determine the degreef antinociceptive tolerance, the ED50 of THC and 95% confidence
ntervals were calculated using nonlinear regression analysis ofach curve. Comparison between genotypes revealed that sub-hronic THC treatment shifted the antinociceptive dose-responseurve to the right to a much greater extent in WT (8.45-fold shift)ompared to �arr2-KO mice (1.68-fold shift; Table 1).
Cannabinoids decrease rodent motility measured as time spentn a cataleptic state. Cumulative dosing of THC induced a similaregree of catalepsy in vehicle-treated WT and �arr2-KO mice [Fig-re 1B; genotype: F (1,78) � 2.53, p � .1159; dose: F (5,78) � 65.19,� .0001; interaction: F (5,78) � .71, p � .6193]. Subchronic THC
reatment reduced the time that both WT and �arr2-KO mice spentmmobile compared with vehicle-treated mice given the samecute dose of THC, indicating that both WT and �arr2-KO miceecome tolerant to THC-mediated catalepsy [WT, treatment:(1,84) � 18.13, p � .0001; dose: F (5,84) � 84.00, p � .0001; inter-ction: F (5,84) � 2.49, p � .0376; �arr2-KO, treatment: F (1,72) �3.27, p � .0001; dose: F (5,72) � 54.97, p � .0001; interaction:(5,72) � 4.91, p � .0006]. Comparison of the THC-induced shift in
he ED50 for each genotype revealed that, in contrast to antinocice-tion, �arr2-KO mice displayed a greater degree of tolerance toHC-induced catalepsy (4.76-fold shift) than WT mice [2.12-foldhift; (F (1,70) � 7.873; p � .01 sum of least squares F test; Table 1].
THC-induced hypothermia was also assessed in WT andarr2-KO mice. Vehicle-treated �arr2-KO mice displayed greaterecreases in body temperature across the THC dosing regimen,ompared with vehicle-treated WT mice [Figure 1C; genotype:(1,78) � 8.15, p � .0055; dose: F (5,78) � 41.96, p � .0001; interac-
ion: F (5,78) � .16, p � .9775]. Subchronic THC treatment inducedignificant tolerance to THC-mediated hypothermia in both geno-ypes [WT, treatment: F (1,84) � 51.15, p � .0001; dose: F (5,84) �2.38, p � .0001; interaction: F (5,84) � 13.26, p � .0001; �arr2-KO,reatment: F (1,72) � 65.02, p � .0001; dose: F (5,72) � 14.76, p �0001; interaction: F (5,72) � 6.94, p � .0001]. The data for THCroups did not converge, therefore ED50 values could not be calcu-
ated, and comparison of the degree of tolerance was not possible.owever, statistical analysis of the two curves revealed no signifi-ant difference between THC-pretreated WT and �arr2-KO micegenotype: F (1,78) � .18, p � .6765]. Collectively these studiesuggest region-specific CB1R regulation by �-arrestin2, becauseHC-induced antinociception, catalepsy, and hypothermia are me-iated by different neuronal populations in the CNS (24 –26).
-Arrestin2 Regulates CB1R Desensitization in Spinal CordThe finding that THC-mediated antinociception is enhanced,
hereas development of tolerance is reduced, in �arr2-KO miceuggests that �-arrestin2 might negatively regulate CB Rs in the
1pinal cord because CB1Rs in this region contribute to tail-flick
www.sobp.org/journal
bBt�glCvbw
f(
vccd[CSicbt
�
icr
t
hr
Fe
dBp he ind
716 BIOL PSYCHIATRY 2012;71:714–724 P.T. Nguyen et al.
w
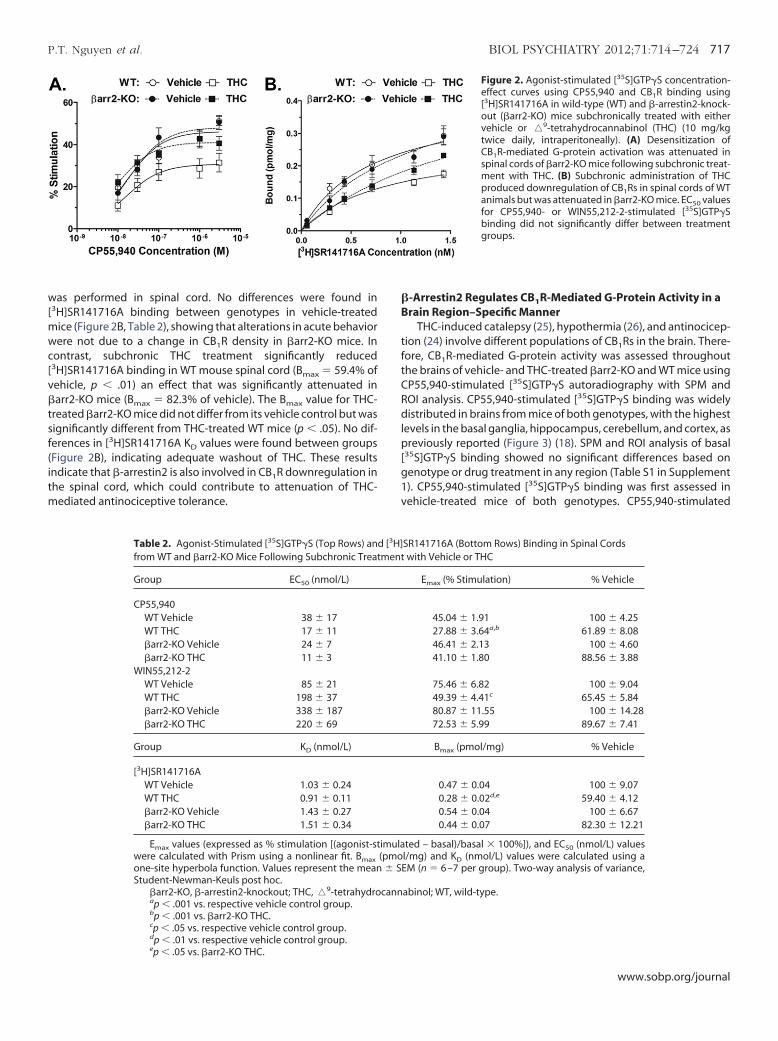
antinociception (27). Therefore, cannabinoid-stimulated [35S]GTP�Sinding was assessed in spinal cords from WT and �arr2-KO mice.asal [35S]GTP�S binding did not significantly differ between geno-ypes or treatments (WT/vehicle � 215 � 11, WT/THC � 208 � 18,arr2-KO/vehicle � 221 � 20, and �arr2-KO/THC � 198 � 15 nCi/). Residual THC would stimulate [35S]GTP�S binding above basal
evels in vehicle-treated mice, thus THC washout was sufficient.P55,940-mediated G-protein activity was first compared betweenehicle-treated WT and �arr2-KO mice and showed no differencesetween genotypes (Figure 2A, Table 2). Subchronic treatmentith THC significantly reduced CP55,940-stimulated [35S]GTP�S
binding in WT mice, with an approximately 38% decrease in recep-tor-mediated activity (Figure 2A, Table 2, p � .001). CB1R desensiti-zation was attenuated in �arr2-KO mice, where the Emax value forCP55,940-stimulated [35S]GTP�S binding did not significantly differrom its vehicle control, but differed from THC-treated WT miceTable 2, p � .001). Half-maximal effective concentration (EC50)
Figure 1. Cumulative �9-tetrahydrocannabinol (THC) dose–response curvreated with either vehicle or THC (10 mg/kg twice daily, intraperitoneally). (A
to THC compared with WT mice in the tail-flick antinociceptive test [for interahoc analysis]. Following THC pretreatment, �arr2-KO mice remain more resppost hoc analysis]. Both genotypes display tolerance following THC pretrea11.25, p � .0001, ^p � .05, ^^^p � .001 Bonferroni post hoc analysis. �arr2-KO
oc analysis]. (B) Following vehicle treatment, �arr2-KO mice display equivaing test for catalepsy [for genotype: F(1,78) � 2.53, p � .1159]. Both genoty
interaction of dose and pretreatment: F(5,84) � 2.49, p � .0376, ^^p � .01 B(5,72) � 4.91, p � .0006, ###p � .001 Bonferroni post hoc analysis]. Followingffects of THC than WT mice [for genotype: F1,78 � 3.97, p � .0497, *p � .0
display greater hypothermia in response to THC compared with WT mice asp � .0055]. Following THC pretreatment, WT and �arr2-KO mice did not differ
isplayed tolerance following the THC pretreatment [vehicle vs. THC; WT: foonferroni post hoc analysis. �arr2-KO: for interaction of dose and pretreatmresented as the mean � SEM (n � 8 WT, n � 7 �arr2-KO). The log values of t
provided. MPE, maximum possible effect.
Table 1. ED50 Values (mg/kg) (� 95% CI) for THC CumuAntinociception, Catalepsy, and Hypothermia Assessme
Behavioral Assessment Genotype Vehicle ED5
Antinociception WT 13.85 (9.30–�arr2-KO 6.37 (4.65–
Catalepsy WT 19.79 (11.56�arr2-KO 12.03 (6.04–
Hypothermia WT 50.24 (17.22�arr2-KO 47.97 (14.06
WT: vehicle vs. THC; KO: vehicle vs. THC, n � 8/group�arr2-KO, �-arrestin2-knockout; CI, confidence inter
wild-type.a
p � .01, sum of squares F test.bp � .05, sum of squares F test.ww.sobp.org/journal
alues for CP55,940-stimulated [35S]GTP�S binding did not signifi-antly differ between treatment groups (Table 2), showing suffi-ient washout of THC. A significant interaction was found betweenrug and genotype [F (1,24) � 5.68, p � .05] in CP55,940-stimulated
35S]GTP�S binding, suggesting that �-arrestin2 contributes toB1R desensitization in the spinal cord following THC treatment.imilar results were found using WIN55,212-2 (Table 2). These stud-
es suggest that �-arrestin2-mediated CB1R desensitization mightontribute to tolerance to THC-mediated antinociception, becauseoth CB1R desensitization in the spinal cord and tolerance to the
ail-flick test were significantly attenuated in �arr2-KO mice.
-Arrestin2 Regulates CB1R Downregulation in Spinal Cord�-arrestin2 facilitates endocytosis for many GPCRs (11). Internal-
zed receptors can be degraded, and the resulting downregulationould contribute to antinociceptive tolerance. To determine theole of �-arrestin2 in CB1R downregulation, [3H]SR141716A binding
wild-type (WT) and �-arrestin2-knockout (�arr2-KO) mice subchronicallylowing vehicle pretreatment, �arr2-KO mice displayed enhanced responsesof dose and genotype: F(5,78) � 5.11, p � .0004, ***p � .001 Bonferroni postthan WT mice [for genotype: F(1,78) � 12.31, p � .0008, *p � .05 Bonferroni
t [vehicle vs. THC: WT: for interaction of dose and pretreatment: F(5,84) �retreatment: F(1,72) � 22.78, p � .0001, #p � .05, ##p � .01 Bonferroni postypoactivity in response to THC compared with WT mice as assessed by the
isplayed tolerance following the THC pretreatment [vehicle vs. THC; WT: forroni post hoc analysis. �arr2-KO: for interaction of dose and pretreatment:pretreatment, �arr2-KO mice become more tolerant to the hypolocomotorferroni post hoc analysis]. (C) Following vehicle treatment, �arr2-KO mice
mined by changes in rectal body temperature [for genotype: F(1,78) � 8.15,ir response profiles [for genotype: F(1,78) � .09, p � .9941]. Both genotypesraction of dose and pretreatment: F(5,84) � 13.26, p � .0001, ^^^p � .001(5,72) � 6.94, p � .0001, ###p � .001 Bonferroni post hoc analysis]. Data areicated doses are graphed on the abscissa and nonlinear regression curves are
Dosing Curves Obtained in WT and �arr2-KO Mice for
CI) THC ED50 (95% CI) Fold Shift in ED50
) 126.9 (81.50–197.4) 8.45a
10.71 (2.60–44.10) 1.688) 41.94 (29.22–60.18) 2.12a
) 57.32 (39.82–82.53) 4.76b
.50) NC —
.60) NC —
C, not converged; THC, �9-tetrahydrocannabinol; WT,
es in) Fol
ctiononsivetmen: for plent h
pes donferTHC
5 Bondeterin ther inteent: F
lativents
0 (95%
20.618.73)–33.823.96–146–163
.val; N
mw
v�
(itm
�B
tftCRdlp[g1v
P.T. Nguyen et al. BIOL PSYCHIATRY 2012;71:714–724 717
was performed in spinal cord. No differences were found in[3H]SR141716A binding between genotypes in vehicle-treated
ice (Figure 2B, Table 2), showing that alterations in acute behaviorere not due to a change in CB1R density in �arr2-KO mice. In
contrast, subchronic THC treatment significantly reduced[3H]SR141716A binding in WT mouse spinal cord (Bmax � 59.4% of
ehicle, p � .01) an effect that was significantly attenuated inarr2-KO mice (Bmax � 82.3% of vehicle). The Bmax value for THC-
treated �arr2-KO mice did not differ from its vehicle control but wassignificantly different from THC-treated WT mice (p � .05). No dif-ferences in [3H]SR141716A KD values were found between groupsFigure 2B), indicating adequate washout of THC. These resultsndicate that �-arrestin2 is also involved in CB1R downregulation inhe spinal cord, which could contribute to attenuation of THC-
ediated antinociceptive tolerance.
Table 2. Agonist-Stimulated [35S]GTP�S (Top Rows) andfrom WT and �arr2-KO Mice Following Subchronic Treat
Group EC50 (nmol/L)
CP55,940WT Vehicle 38 � 17WT THC 17 � 11�arr2-KO Vehicle 24 � 7�arr2-KO THC 11 � 3
WIN55,212-2WT Vehicle 85 � 21WT THC 198 � 37�arr2-KO Vehicle 338 � 187�arr2-KO THC 220 � 69
Group KD (nmol/L)
[3H]SR141716AWT Vehicle 1.03 � 0.24WT THC 0.91 � 0.11�arr2-KO Vehicle 1.43 � 0.27�arr2-KO THC 1.51 � 0.34
Emax values (expressed as % stimulation [(agonist-stwere calculated with Prism using a nonlinear fit. Bmax
one-site hyperbola function. Values represent the meanStudent-Newman-Keuls post hoc.
�arr2-KO, �-arrestin2-knockout; THC, �9-tetrahydroap � .001 vs. respective vehicle control group.bp � .001 vs. �arr2-KO THC.cp � .05 vs. respective vehicle control group.d
p � .01 vs. respective vehicle control group.ep � .05 vs. �arr2-KO THC.-Arrestin2 Regulates CB1R-Mediated G-Protein Activity in arain Region–Specific Manner
THC-induced catalepsy (25), hypothermia (26), and antinocicep-ion (24) involve different populations of CB1Rs in the brain. There-ore, CB1R-mediated G-protein activity was assessed throughouthe brains of vehicle- and THC-treated �arr2-KO and WT mice usingP55,940-stimulated [35S]GTP�S autoradiography with SPM andOI analysis. CP55,940-stimulated [35S]GTP�S binding was widelyistributed in brains from mice of both genotypes, with the highest
evels in the basal ganglia, hippocampus, cerebellum, and cortex, asreviously reported (Figure 3) (18). SPM and ROI analysis of basal
35S]GTP�S binding showed no significant differences based onenotype or drug treatment in any region (Table S1 in Supplement). CP55,940-stimulated [35S]GTP�S binding was first assessed inehicle-treated mice of both genotypes. CP55,940-stimulated
Figure 2. Agonist-stimulated [35S]GTP�S concentration-effect curves using CP55,940 and CB1R binding using[3H]SR141716A in wild-type (WT) and �-arrestin2-knock-out (�arr2-KO) mice subchronically treated with eithervehicle or �9-tetrahydrocannabinol (THC) (10 mg/kgtwice daily, intraperitoneally). (A) Desensitization ofCB1R-mediated G-protein activation was attenuated inspinal cords of �arr2-KO mice following subchronic treat-ment with THC. (B) Subchronic administration of THCproduced downregulation of CB1Rs in spinal cords of WTanimals but was attenuated in �arr2-KO mice. EC50 valuesfor CP55,940- or WIN55,212-2-stimulated [35S]GTP�Sbinding did not significantly differ between treatmentgroups.
SR141716A (Bottom Rows) Binding in Spinal Cordswith Vehicle or THC
Emax (% Stimulation) % Vehicle
45.04 � 1.91 100 � 4.2527.88 � 3.64a,b 61.89 � 8.0846.41 � 2.13 100 � 4.6041.10 � 1.80 88.56 � 3.88
75.46 � 6.82 100 � 9.0449.39 � 4.41c 65.45 � 5.8480.87 � 11.55 100 � 14.2872.53 � 5.99 89.67 � 7.41
Bmax (pmol/mg) % Vehicle
0.47 � 0.04 100 � 9.070.28 � 0.02d,e 59.40 � 4.120.54 � 0.04 100 � 6.670.44 � 0.07 82.30 � 12.21
ted – basal)/basal � 100%]), and EC50 (nmol/L) valuesl/mg) and KD (nmol/L) values were calculated using aM (n � 6 –7 per group). Two-way analysis of variance,
abinol; WT, wild-type.
[3H]ment
imula(pmo� SE
cann
www.sobp.org/journal
cobhwstww
T
r
aWss
cbmG7st�s[dats(tw(uslovTnv
aaboTbh(
tbt5p
cgrtmps
FbScgkp
718 BIOL PSYCHIATRY 2012;71:714–724 P.T. Nguyen et al.
w
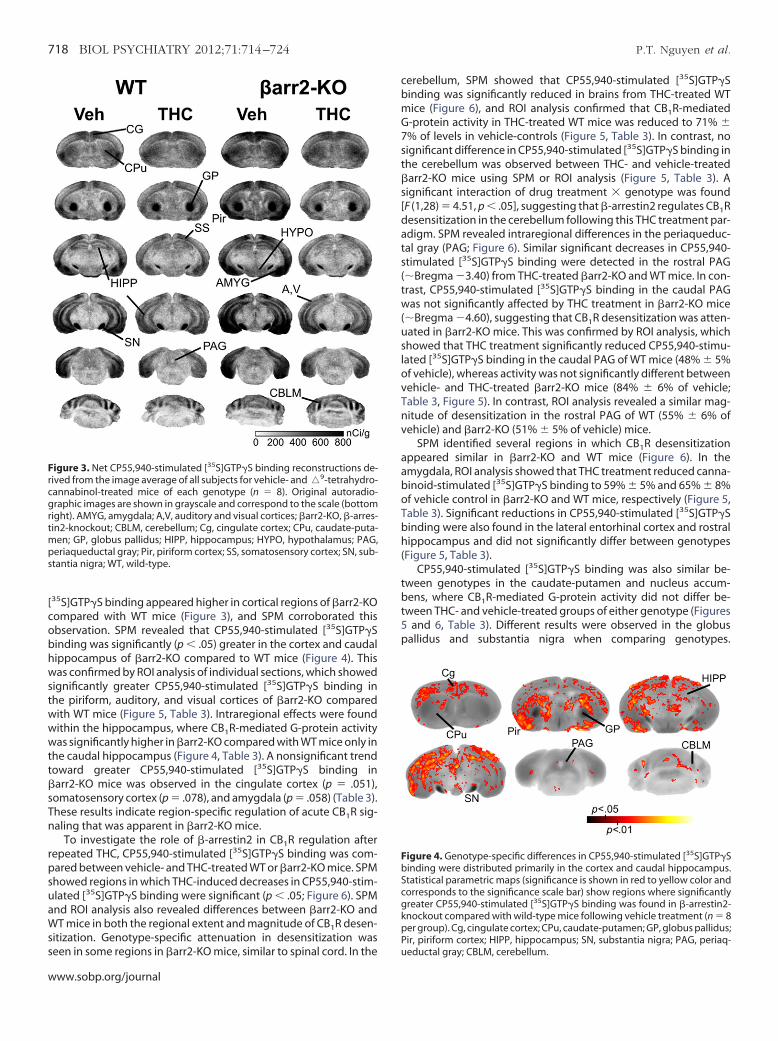
[35S]GTP�S binding appeared higher in cortical regions of �arr2-KOompared with WT mice (Figure 3), and SPM corroborated thisbservation. SPM revealed that CP55,940-stimulated [35S]GTP�Sinding was significantly (p � .05) greater in the cortex and caudalippocampus of �arr2-KO compared to WT mice (Figure 4). Thisas confirmed by ROI analysis of individual sections, which showed
ignificantly greater CP55,940-stimulated [35S]GTP�S binding inhe piriform, auditory, and visual cortices of �arr2-KO comparedith WT mice (Figure 5, Table 3). Intraregional effects were foundithin the hippocampus, where CB1R-mediated G-protein activity
was significantly higher in �arr2-KO compared with WT mice only inthe caudal hippocampus (Figure 4, Table 3). A nonsignificant trendtoward greater CP55,940-stimulated [35S]GTP�S binding in�arr2-KO mice was observed in the cingulate cortex (p � .051),somatosensory cortex (p � .078), and amygdala (p � .058) (Table 3).
hese results indicate region-specific regulation of acute CB1R sig-naling that was apparent in �arr2-KO mice.
To investigate the role of �-arrestin2 in CB1R regulation afterepeated THC, CP55,940-stimulated [35S]GTP�S binding was com-
pared between vehicle- and THC-treated WT or �arr2-KO mice. SPMshowed regions in which THC-induced decreases in CP55,940-stim-ulated [35S]GTP�S binding were significant (p � .05; Figure 6). SPM
nd ROI analysis also revealed differences between �arr2-KO andT mice in both the regional extent and magnitude of CB1R desen-
itization. Genotype-specific attenuation in desensitization was
Figure 3. Net CP55,940-stimulated [35S]GTP�S binding reconstructions de-rived from the image average of all subjects for vehicle- and�9-tetrahydro-
annabinol-treated mice of each genotype (n � 8). Original autoradio-raphic images are shown in grayscale and correspond to the scale (bottom
ight). AMYG, amygdala; A,V, auditory and visual cortices; �arr2-KO, �-arres-in2-knockout; CBLM, cerebellum; Cg, cingulate cortex; CPu, caudate-puta-
en; GP, globus pallidus; HIPP, hippocampus; HYPO, hypothalamus; PAG,eriaqueductal gray; Pir, piriform cortex; SS, somatosensory cortex; SN, sub-tantia nigra; WT, wild-type.
een in some regions in �arr2-KO mice, similar to spinal cord. In thePu
ww.sobp.org/journal
erebellum, SPM showed that CP55,940-stimulated [35S]GTP�Sinding was significantly reduced in brains from THC-treated WTice (Figure 6), and ROI analysis confirmed that CB1R-mediated-protein activity in THC-treated WT mice was reduced to 71% �% of levels in vehicle-controls (Figure 5, Table 3). In contrast, noignificant difference in CP55,940-stimulated [35S]GTP�S binding inhe cerebellum was observed between THC- and vehicle-treatedarr2-KO mice using SPM or ROI analysis (Figure 5, Table 3). A
ignificant interaction of drug treatment � genotype was foundF (1,28) � 4.51, p � .05], suggesting that �-arrestin2 regulates CB1Resensitization in the cerebellum following this THC treatment par-digm. SPM revealed intraregional differences in the periaqueduc-al gray (PAG; Figure 6). Similar significant decreases in CP55,940-timulated [35S]GTP�S binding were detected in the rostral PAGBregma �3.40) from THC-treated �arr2-KO and WT mice. In con-rast, CP55,940-stimulated [35S]GTP�S binding in the caudal PAGas not significantly affected by THC treatment in �arr2-KO mice
Bregma �4.60), suggesting that CB1R desensitization was atten-ated in �arr2-KO mice. This was confirmed by ROI analysis, whichhowed that THC treatment significantly reduced CP55,940-stimu-ated [35S]GTP�S binding in the caudal PAG of WT mice (48% � 5%f vehicle), whereas activity was not significantly different betweenehicle- and THC-treated �arr2-KO mice (84% � 6% of vehicle;able 3, Figure 5). In contrast, ROI analysis revealed a similar mag-itude of desensitization in the rostral PAG of WT (55% � 6% ofehicle) and �arr2-KO (51% � 5% of vehicle) mice.
SPM identified several regions in which CB1R desensitizationppeared similar in �arr2-KO and WT mice (Figure 6). In themygdala, ROI analysis showed that THC treatment reduced canna-inoid-stimulated [35S]GTP�S binding to 59% � 5% and 65% � 8%f vehicle control in �arr2-KO and WT mice, respectively (Figure 5,able 3). Significant reductions in CP55,940-stimulated [35S]GTP�Sinding were also found in the lateral entorhinal cortex and rostralippocampus and did not significantly differ between genotypes
Figure 5, Table 3).CP55,940-stimulated [35S]GTP�S binding was also similar be-
ween genotypes in the caudate-putamen and nucleus accum-ens, where CB1R-mediated G-protein activity did not differ be-
ween THC- and vehicle-treated groups of either genotype (Figuresand 6, Table 3). Different results were observed in the globus
allidus and substantia nigra when comparing genotypes.
igure 4. Genotype-specific differences in CP55,940-stimulated [35S]GTP�Sinding were distributed primarily in the cortex and caudal hippocampus.tatistical parametric maps (significance is shown in red to yellow color andorresponds to the significance scale bar) show regions where significantlyreater CP55,940-stimulated [35S]GTP�S binding was found in �-arrestin2-nockout compared with wild-type mice following vehicle treatment (n � 8er group). Cg, cingulate cortex; CPu, caudate-putamen; GP, globus pallidus;
ir, piriform cortex; HIPP, hippocampus; SN, substantia nigra; PAG, periaq-eductal gray; CBLM, cerebellum.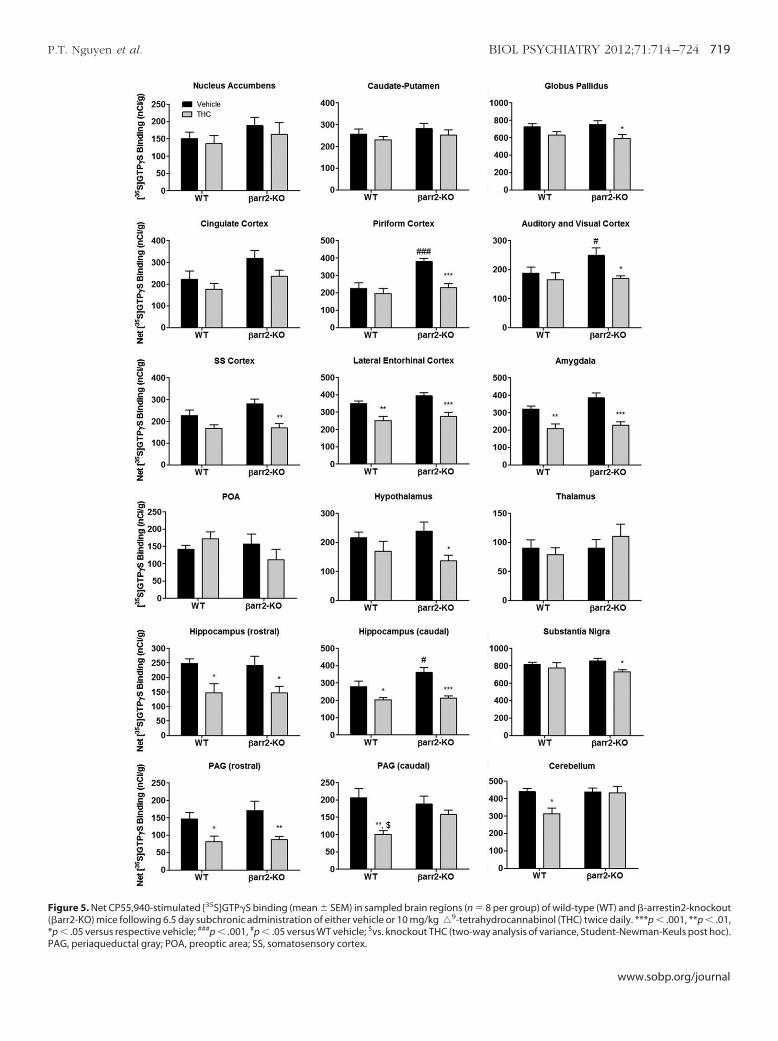
*P
P.T. Nguyen et al. BIOL PSYCHIATRY 2012;71:714–724 719
Figure 5. Net CP55,940-stimulated [35S]GTP�S binding (mean � SEM) in sampled brain regions (n � 8 per group) of wild-type (WT) and �-arrestin2-knockout(�arr2-KO) mice following 6.5 day subchronic administration of either vehicle or 10 mg/kg �9-tetrahydrocannabinol (THC) twice daily. ***p � .001, **p � .01,
### # $
p � .05 versus respective vehicle; p � .001, p � .05 versus WT vehicle; vs. knockout THC (two-way analysis of variance, Student-Newman-Keuls post hoc).AG, periaqueductal gray; POA, preoptic area; SS, somatosensory cortex.www.sobp.org/journal

Tobvp
mmgS
emTCpm
�C
udgl[lwgdad4d
D
pd�
720 BIOL PSYCHIATRY 2012;71:714–724 P.T. Nguyen et al.
w
CP55,940-stimulated [35S]GTP�S binding did not differ betweenHC- and vehicle-treated WT mice in these regions, showing a lackf CB1R desensitization. However, CP55,940-stimulated [35S]GTP�Sinding was significantly reduced in the substantia nigra (85% ofehicle) and globus pallidus (79% of vehicle) of THC-treated com-ared with vehicle-treated �arr2-KO mice (Figures 5 and 6, Table 3).
Therefore, deletion of �-arrestin2 appeared to enhance CB1R de-sensitization in the globus pallidus and substantia nigra, withoutaltering acute CB1R-mediated G-protein activity in vehicle-treatedmice.
SPM analysis revealed enhanced desensitization in several otherregions of �arr2-KO compared to WT mice (Figure 6). In the hypo-thalamus, CP55,940-stimulated [35S]GTP�S binding in THC-treated
ice was 57% � 8% and 79% � 16% of vehicle in �arr2-KO and WTice, respectively (Figure 5, Table 3) and occurred in the absence of
enotype-dependent differences in vehicle-treated mice (Table 3).ubchronic THC also induced CB1R desensitization in the piriform,
auditory, and visual cortices of �arr2-KO, but not WT, mice (Figure 5,Table 3). SPM revealed intraregional differences in desensitizationwithin the hippocampus, wherein desensitization was significantlygreater in magnitude in the caudal hippocampus of �arr2-KO mice(Figures 6). ROI analysis showed a significantly greater reduction inCP55,940-stimulated [35S]GTP�S binding in the caudal hippocam-pus in �arr2-KO mice (58.6% � 4% of vehicle), compared with WTmice (72.6% � 4% of vehicle; Figure 5, Table 3).
Increases in the apparent efficacy of CP55,940-stimulated[35S]GTP�S binding in some regions of �arr2-KO brains appeared tobe associated with the enhancement or unmasking of CB1R desen-sitization in these same regions following THC administration. Therelative magnitude of desensitization in �arr2-KO mice when nor-
Table 3. Net CP55,940-Stimulated [35S]GTP�S Binding iDay Administration of Either Vehicle or THC (n � 8 per G
Region
Veh
WT
Nucleus Accumbens 150 � 20Caudate-Putamen 256 � 23Globus Pallidus 726 � 36Cingulate Cortex 221 � 39Piriform Cortex 225 � 32Auditory and Visual Cortices 188 � 21Somatosensory Cortex 226 � 25Lateral Entorhinal Cortex 350 � 16Amygdala 321 � 17Hippocampus (Rostral) 248 � 17Hippocampus (Caudal) 279 � 31Preoptic area 141 � 12Hypothalamus 216 � 21Thalamus 90 � 15Substantia Nigra 814 � 26PAG (Rostral) 146 � 19PAG (Caudal) 207 � 26Cerebellum 440 � 19
[35S]GTP�S binding is expressed as net agonist-stimuSEM, two-way analysis of variance, Student-Newman Ke
�arr2-KO, �-arrestin2-knockout; PAG, periaqueductaap � .05 vs. respective vehicle.bp � .001 vs. WT vehicle.cp � .001 vs. respective vehicle.dp � .05 vs. WT vehicle.ep � .01 vs. respective vehicle.fp � .05 vs. knockout THC.
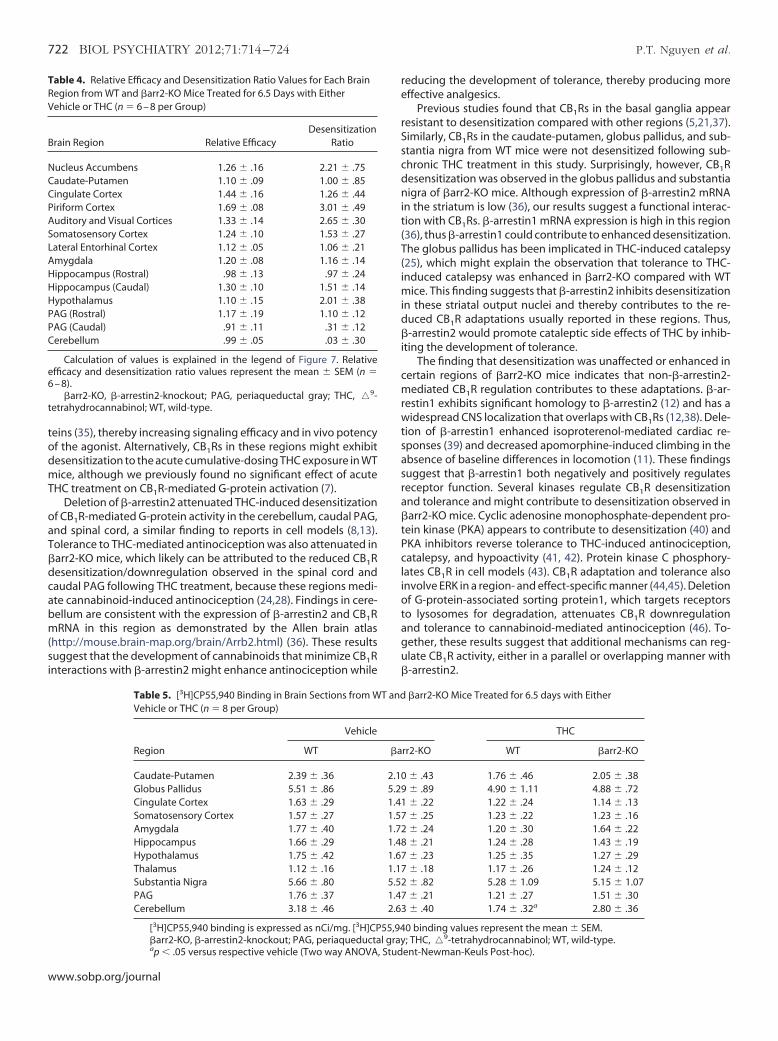
malized to WT animals was positively correlated with the relative a
ww.sobp.org/journal
fficacy of CP55,940-stimulated [35S]GTP�S binding in �arr2-KOice when normalized to WT animals (p � .0011, R2 � .60; Figure 7,
able 4). For example, a 1.5-fold greater relative efficacy ofP55,940-stimulated [35S]GTP�S binding in �arr2-KO mice com-ared to WT mice corresponded to an approximately 2-fold greateragnitude of desensitization in �arr2-KO relative to WT mice.
-Arrestin2 Is Involved in CB1R Downregulation in theerebellum
The role of �-arrestin2 in CB1R downregulation was assessedsing [3H]CP55,940 autoradiography. [3H]CP55,940 binding did notiffer between genotypes in vehicle-treated mice (Table 5), sug-esting that genotype-dependent differences in CP55,940-stimu-
ated [35S]GTP�S binding were not due to differences in CB1R levels.3H]CP55,940 binding was significantly decreased in the cerebel-um of WT mice following THC treatment (55% � 10% of vehicle),
hereas [3H]CP55,940 binding was similar in both treatmentroups in �arr2-KO mice. [3H]CP55,940 binding did not significantlyiffer between THC- and vehicle-treated mice of either genotype inny other region examined (Table 5). A significant interaction ofrug treatment � genotype was found in the cerebellum [F (1,28) �.277, p � .05], suggesting that �-arrestin2 is involved in CB1Rownregulation in the cerebellum.
iscussion
This study shows that �-arrestin2 regulates CB1R-mediated G-rotein activity in a CNS region–specific manner and that theseifferences are reflected in THC-mediated responses observed inarr2-KO mice. Genotype-specific differences were observed after
in Sections from WT and �arr2-KO Mice Following 6.5-)
THC
�arr2-KO WT �arr2-KO
189 � 24 136 � 24 163 � 33282 � 24 230 � 17 252 � 25751 � 47 631 � 41 593 � 43a
318 � 36 176 � 29 235 � 29380 � 18b 195 � 29 229 � 25c
249 � 26d 165 � 23 169 � 9a
281 � 22 168 � 15 169 � 20e
393 � 19 252 � 23e 277 � 23c
385 � 27 209 � 26e 228 � 19c
242 � 32 147 � 32a 146 � 23a
362 � 27d 203 � 12a 212 � 14c
157 � 29 172 � 20 111 � 30238 � 32 169 � 35 136 � 20a
90 � 15 79 � 12 110 � 21856 � 30 776 � 58 733 � 24a
171 � 28 81 � 16a 88 � 9e
189 � 22 100 � 12e,f 158 � 12437 � 23 314 � 32a 434 � 38
[35S]GTP�S binding in nCi/g. Values represent mean �st hoc.; THC, �9-tetrahydrocannabinol; WT, wild-type.
n Braroup
icle
lateduls pol gray
cute THC, with �arr2-KO mice displaying enhanced THC-mediated
se
hwehtC
cMacfiiuaPrbnptsat(lhnas
ptifdapogr
gt�iBt
Fp�carmdnvlgp
P.T. Nguyen et al. BIOL PSYCHIATRY 2012;71:714–724 721
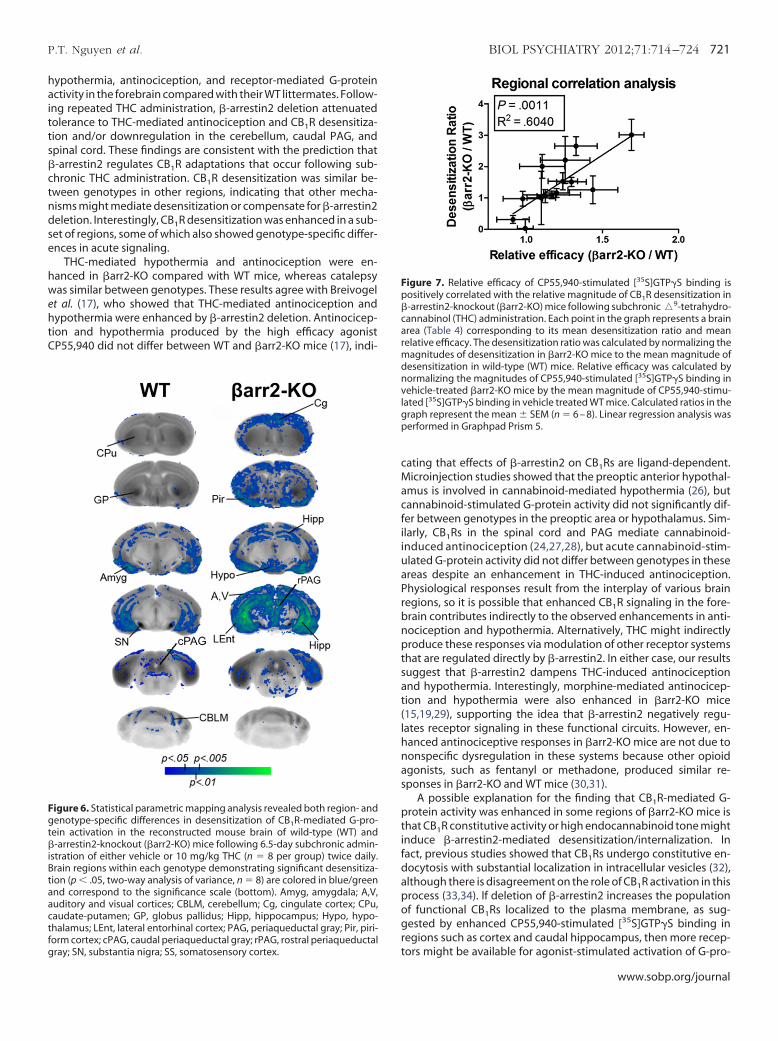
hypothermia, antinociception, and receptor-mediated G-proteinactivity in the forebrain compared with their WT littermates. Follow-ing repeated THC administration, �-arrestin2 deletion attenuatedtolerance to THC-mediated antinociception and CB1R desensitiza-tion and/or downregulation in the cerebellum, caudal PAG, andspinal cord. These findings are consistent with the prediction that�-arrestin2 regulates CB1R adaptations that occur following sub-chronic THC administration. CB1R desensitization was similar be-tween genotypes in other regions, indicating that other mecha-nisms might mediate desensitization or compensate for �-arrestin2deletion. Interestingly, CB1R desensitization was enhanced in a sub-et of regions, some of which also showed genotype-specific differ-nces in acute signaling.
THC-mediated hypothermia and antinociception were en-anced in �arr2-KO compared with WT mice, whereas catalepsyas similar between genotypes. These results agree with Breivogel
t al. (17), who showed that THC-mediated antinociception andypothermia were enhanced by �-arrestin2 deletion. Antinocicep-
ion and hypothermia produced by the high efficacy agonistP55,940 did not differ between WT and �arr2-KO mice (17), indi-
Figure 6. Statistical parametric mapping analysis revealed both region- andenotype-specific differences in desensitization of CB1R-mediated G-pro-
ein activation in the reconstructed mouse brain of wild-type (WT) and-arrestin2-knockout (�arr2-KO) mice following 6.5-day subchronic admin-
stration of either vehicle or 10 mg/kg THC (n � 8 per group) twice daily.rain regions within each genotype demonstrating significant desensitiza-ion (p � .05, two-way analysis of variance, n � 8) are colored in blue/green
and correspond to the significance scale (bottom). Amyg, amygdala; A,V,auditory and visual cortices; CBLM, cerebellum; Cg, cingulate cortex; CPu,caudate-putamen; GP, globus pallidus; Hipp, hippocampus; Hypo, hypo-thalamus; LEnt, lateral entorhinal cortex; PAG, periaqueductal gray; Pir, piri-
tform cortex; cPAG, caudal periaqueductal gray; rPAG, rostral periaqueductalgray; SN, substantia nigra; SS, somatosensory cortex.
ating that effects of �-arrestin2 on CB1Rs are ligand-dependent.icroinjection studies showed that the preoptic anterior hypothal-
mus is involved in cannabinoid-mediated hypothermia (26), butannabinoid-stimulated G-protein activity did not significantly dif-er between genotypes in the preoptic area or hypothalamus. Sim-larly, CB1Rs in the spinal cord and PAG mediate cannabinoid-nduced antinociception (24,27,28), but acute cannabinoid-stim-lated G-protein activity did not differ between genotypes in thesereas despite an enhancement in THC-induced antinociception.hysiological responses result from the interplay of various brainegions, so it is possible that enhanced CB1R signaling in the fore-rain contributes indirectly to the observed enhancements in anti-ociception and hypothermia. Alternatively, THC might indirectlyroduce these responses via modulation of other receptor systems
hat are regulated directly by �-arrestin2. In either case, our resultsuggest that �-arrestin2 dampens THC-induced antinociceptionnd hypothermia. Interestingly, morphine-mediated antinocicep-ion and hypothermia were also enhanced in �arr2-KO mice15,19,29), supporting the idea that �-arrestin2 negatively regu-ates receptor signaling in these functional circuits. However, en-anced antinociceptive responses in �arr2-KO mice are not due toonspecific dysregulation in these systems because other opioidgonists, such as fentanyl or methadone, produced similar re-ponses in �arr2-KO and WT mice (30,31).
A possible explanation for the finding that CB1R-mediated G-rotein activity was enhanced in some regions of �arr2-KO mice is
hat CB1R constitutive activity or high endocannabinoid tone mightnduce �-arrestin2-mediated desensitization/internalization. Inact, previous studies showed that CB1Rs undergo constitutive en-ocytosis with substantial localization in intracellular vesicles (32),lthough there is disagreement on the role of CB1R activation in thisrocess (33,34). If deletion of �-arrestin2 increases the populationf functional CB1Rs localized to the plasma membrane, as sug-ested by enhanced CP55,940-stimulated [35S]GTP�S binding in
egions such as cortex and caudal hippocampus, then more recep-
igure 7. Relative efficacy of CP55,940-stimulated [35S]GTP�S binding isositively correlated with the relative magnitude of CB1R desensitization in-arrestin2-knockout (�arr2-KO) mice following subchronic�9-tetrahydro-annabinol (THC) administration. Each point in the graph represents a brainrea (Table 4) corresponding to its mean desensitization ratio and meanelative efficacy. The desensitization ratio was calculated by normalizing the
agnitudes of desensitization in �arr2-KO mice to the mean magnitude ofesensitization in wild-type (WT) mice. Relative efficacy was calculated byormalizing the magnitudes of CP55,940-stimulated [35S]GTP�S binding inehicle-treated �arr2-KO mice by the mean magnitude of CP55,940-stimu-
ated [35S]GTP�S binding in vehicle treated WT mice. Calculated ratios in theraph represent the mean � SEM (n � 6 – 8). Linear regression analysis waserformed in Graphpad Prism 5.
ors might be available for agonist-stimulated activation of G-pro-
www.sobp.org/journal
aT�
i
re
rSscdnit(T(imid�i
cmrwtsasra�tPcliotagu�
R
CCPASLAHHHPPC
e6
t
722 BIOL PSYCHIATRY 2012;71:714–724 P.T. Nguyen et al.
w
teins (35), thereby increasing signaling efficacy and in vivo potencyof the agonist. Alternatively, CB1Rs in these regions might exhibitdesensitization to the acute cumulative-dosing THC exposure in WTmice, although we previously found no significant effect of acuteTHC treatment on CB1R-mediated G-protein activation (7).
Deletion of �-arrestin2 attenuated THC-induced desensitizationof CB1R-mediated G-protein activity in the cerebellum, caudal PAG,
nd spinal cord, a similar finding to reports in cell models (8,13).olerance to THC-mediated antinociception was also attenuated inarr2-KO mice, which likely can be attributed to the reduced CB1R
desensitization/downregulation observed in the spinal cord andcaudal PAG following THC treatment, because these regions medi-ate cannabinoid-induced antinociception (24,28). Findings in cere-bellum are consistent with the expression of �-arrestin2 and CB1RmRNA in this region as demonstrated by the Allen brain atlas(http://mouse.brain-map.org/brain/Arrb2.html) (36). These resultssuggest that the development of cannabinoids that minimize CB1Rnteractions with �-arrestin2 might enhance antinociception while
Table 4. Relative Efficacy and Desensitization Ratio Values for Each Brainegion from WT and �arr2-KO Mice Treated for 6.5 Days with Either
Vehicle or THC (n � 6 – 8 per Group)
Brain Region Relative EfficacyDesensitization
Ratio
Nucleus Accumbens 1.26 � .16 2.21 � .75audate-Putamen 1.10 � .09 1.00 � .85ingulate Cortex 1.44 � .16 1.26 � .44iriform Cortex 1.69 � .08 3.01 � .49uditory and Visual Cortices 1.33 � .14 2.65 � .30omatosensory Cortex 1.24 � .10 1.53 � .27ateral Entorhinal Cortex 1.12 � .05 1.06 � .21mygdala 1.20 � .08 1.16 � .14ippocampus (Rostral) .98 � .13 .97 � .24ippocampus (Caudal) 1.30 � .10 1.51 � .14ypothalamus 1.10 � .15 2.01 � .38AG (Rostral) 1.17 � .19 1.10 � .12AG (Caudal) .91 � .11 .31 � .12erebellum .99 � .05 .03 � .30
Calculation of values is explained in the legend of Figure 7. Relativefficacy and desensitization ratio values represent the mean � SEM (n �– 8).
�arr2-KO, �-arrestin2-knockout; PAG, periaqueductal gray; THC, �9-etrahydrocannabinol; WT, wild-type.
Table 5. [3H]CP55,940 Binding in Brain Sections from WVehicle or THC (n � 8 per Group)
Region
Vehicle
WT
Caudate-Putamen 2.39 � .36Globus Pallidus 5.51 � .86Cingulate Cortex 1.63 � .29Somatosensory Cortex 1.57 � .27Amygdala 1.77 � .40Hippocampus 1.66 � .29Hypothalamus 1.75 � .42Thalamus 1.12 � .16Substantia Nigra 5.66 � .80PAG 1.76 � .37Cerebellum 3.18 � .46
[3H]CP55,940 binding is expressed as nCi/mg. [3H]CP
�arr2-KO, �-arrestin2-knockout; PAG, periaqueductal grayap � .05 versus respective vehicle (Two way ANOVA, Studww.sobp.org/journal
educing the development of tolerance, thereby producing moreffective analgesics.
Previous studies found that CB1Rs in the basal ganglia appearesistant to desensitization compared with other regions (5,21,37).imilarly, CB1Rs in the caudate-putamen, globus pallidus, and sub-tantia nigra from WT mice were not desensitized following sub-hronic THC treatment in this study. Surprisingly, however, CB1Resensitization was observed in the globus pallidus and substantiaigra of �arr2-KO mice. Although expression of �-arrestin2 mRNA
n the striatum is low (36), our results suggest a functional interac-ion with CB1Rs. �-arrestin1 mRNA expression is high in this region36), thus �-arrestin1 could contribute to enhanced desensitization.he globus pallidus has been implicated in THC-induced catalepsy
25), which might explain the observation that tolerance to THC-nduced catalepsy was enhanced in �arr2-KO compared with WT
ice. This finding suggests that �-arrestin2 inhibits desensitizationn these striatal output nuclei and thereby contributes to the re-uced CB1R adaptations usually reported in these regions. Thus,-arrestin2 would promote cataleptic side effects of THC by inhib-
ting the development of tolerance.The finding that desensitization was unaffected or enhanced in
ertain regions of �arr2-KO mice indicates that non-�-arrestin2-ediated CB1R regulation contributes to these adaptations. �-ar-
estin1 exhibits significant homology to �-arrestin2 (12) and has aidespread CNS localization that overlaps with CB1Rs (12,38). Dele-
ion of �-arrestin1 enhanced isoproterenol-mediated cardiac re-ponses (39) and decreased apomorphine-induced climbing in thebsence of baseline differences in locomotion (11). These findingsuggest that �-arrestin1 both negatively and positively regulateseceptor function. Several kinases regulate CB1R desensitizationnd tolerance and might contribute to desensitization observed inarr2-KO mice. Cyclic adenosine monophosphate-dependent pro-
ein kinase (PKA) appears to contribute to desensitization (40) andKA inhibitors reverse tolerance to THC-induced antinociception,atalepsy, and hypoactivity (41, 42). Protein kinase C phosphory-
ates CB1R in cell models (43). CB1R adaptation and tolerance alsonvolve ERK in a region- and effect-specific manner (44,45). Deletionf G-protein-associated sorting protein1, which targets receptors
o lysosomes for degradation, attenuates CB1R downregulationnd tolerance to cannabinoid-mediated antinociception (46). To-ether, these results suggest that additional mechanisms can reg-late CB1R activity, either in a parallel or overlapping manner with-arrestin2.
�arr2-KO Mice Treated for 6.5 days with Either
THC
rr2-KO WT �arr2-KO
� .43 1.76 � .46 2.05 � .38� .89 4.90 � 1.11 4.88 � .72� .22 1.22 � .24 1.14 � .13� .25 1.23 � .22 1.23 � .16� .24 1.20 � .30 1.64 � .22� .21 1.24 � .28 1.43 � .19� .23 1.25 � .35 1.27 � .29� .18 1.17 � .26 1.24 � .12� .82 5.28 � 1.09 5.15 � 1.07� .21 1.21 � .27 1.51 � .30� .40 1.74 � .32a 2.80 � .36
0 binding values represent the mean � SEM.9
T and
�a
2.105.291.411.571.721.481.671.175.521.472.63
55,94
; THC, � -tetrahydrocannabinol; WT, wild-type.ent-Newman-Keuls Post-hoc).

uMlaede
mapdlmCo
ePD(U
c
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
3
P.T. Nguyen et al. BIOL PSYCHIATRY 2012;71:714–724 723
In summary, the present studies demonstrate multiple effects of�-arrestin2 deletion on CB1R function and confirm its role in atten-
ating acute THC-mediated antinociception and hypothermia.oreover, results demonstrate both positive and negative modu-
atory effects of �-arrestin2 on THC tolerance, such that tolerance tontinociception was reduced whereas tolerance to catalepsy wasnhanced in �arr2-KO mice. These findings corresponded with re-uced CB1R desensitization in the caudal PAG and spinal cord, andnhanced CB1R desensitization in striatal output nuclei, respec-
tively. The findings further suggest that developing cannabinoidagonists that minimize interactions between CB1R and �-arrestin2
ight be therapeutically beneficial, given that data indicate thesegonists might enhance antinociception but attenuate antinocice-tive tolerance. The side effect profile might also improve becauseeletion of �-arrestin2 enhanced tolerance to THC-induced cata-
eptic effects, suggesting that motor side effects might be mini-ized. These findings demonstrate that interactions between
B1Rs and �-arrestin2 should be considered in future drug devel-pment.
We thank James Gillespie and Patraic Lichtman for assistance withxperiments and image processing. This study was supported by U.S.ublic Health Service Grant Nos. DA014277 (to LJS), DA025321 (toES), DA14460 (to LMB), F31-DA021952 (to KMR), and F30-DA023758
to PTN) and by an A.D. Williams Award from Virginia Commonwealthniversity (to LJS).
The authors report no biomedical financial interests or potentialonflicts of interest.
Supplementary material cited in this article is available online.
1. Herkenham M, Lynn AB, Johnson MR, Melvin LS, de Costa BR, Rice KC(1991): Characterization and localization of cannabinoid receptors in ratbrain: A quantitative in vitro autoradiographic study. J Neurosci 11:563–583.
2. Zimmer A, Zimmer AM, Hohmann AG, Herkenham M, Bonner TI (1999):Increased mortality, hypoactivity, and hypoalgesia in cannabinoid CB1receptor knockout mice. Proc Natl Acad Sci U S A 96:5780 –5785.
3. Pertwee RG (2005): The therapeutic potential of drugs that target can-nabinoid receptors or modulate the tissue levels or actions of endocan-nabinoids. AAPS J 7:E625– 654.
4. Lichtman AH, Martin BR (2005): Cannabinoid tolerance and depen-dence. Handb Exp Pharmacol 691–717.
5. Sim-Selley LJ (2003): Regulation of cannabinoid CB1 receptors in thecentral nervous system by chronic cannabinoids. Crit Rev Neurobiol 15:91–119.
6. Howlett AC, Barth F, Bonner TI, Cabral G, Casellas P, Devane WA, et al.(2002): International Union of Pharmacology. XXVII. Classification ofcannabinoid receptors. Pharmacol Rev 54:161–202.
7. Sim LJ, Hampson RE, Deadwyler SA, Childers SR (1996): Effects of chronictreatment with delta9-tetrahydrocannabinol on cannabinoid-stimu-lated [35S]GTPgammaS autoradiography in rat brain. J Neurosci 16:8057– 8066.
8. Jin W, Brown S, Roche JP, Hsieh C, Celver JP, Kovoor A, et al. (1999):Distinct domains of the CB1 cannabinoid receptor mediate desensitiza-tion and internalization. J Neurosci 19:3773–3780.
9. Oviedo A, Glowa J, Herkenham M (1993): Chronic cannabinoid adminis-tration alters cannabinoid receptor binding in rat brain: A quantitativeautoradiographic study. Brain Res 616:293–302.
10. Sim-Selley LJ, Schechter NS, Rorrer WK, Dalton GD, Hernandez J, MartinBR, et al. (2006): Prolonged recovery rate of CB1 receptor adaptationafter cessation of long-term cannabinoid administration. Mol Pharma-col 70:986 –996.
11. Gainetdinov RR, Premont RT, Bohn LM, Lefkowitz RJ, Caron MG (2004):Desensitization of G protein-coupled receptors and neuronal functions.Annu Rev Neurosci 27:107–144.
12. Attramadal H, Arriza JL, Aoki C, Dawson TM, Codina J, Kwatra MM, et al.
(1992): Beta-arrestin2, a novel member of the arrestin/beta-arrestingene family. J Biol Chem 267:17882–17890.3. Kouznetsova M, Kelley B, Shen M, Thayer SA (2002): Desensitization ofcannabinoid-mediated presynaptic inhibition of neurotransmission be-tween rat hippocampal neurons in culture. Mol Pharmacol 61:477– 485.
4. Tsou K, Brown S, Sanudo-Pena MC, Mackie K, Walker JM (1998): Immu-nohistochemical distribution of cannabinoid CB1 receptors in the ratcentral nervous system. Neuroscience 83:393– 411.
5. Bohn LM, Lefkowitz RJ, Gainetdinov RR, Peppel K, Caron MG, Lin FT(1999): Enhanced morphine analgesia in mice lacking beta-arrestin 2.Science 286:2495–2498.
6. Schmid CL, Bohn LM (2009): Physiological and pharmacological impli-cations of beta-arrestin regulation. Pharmacol Ther 121:285–293.
7. Breivogel CS, Lambert JM, Gerfin S, Huffman JW, Razdan RK (2008):Sensitivity to delta9-tetrahydrocannabinol is selectively enhanced inbeta-arrestin2 –/– mice. Behavioural pharmacology 19:298 –307.
8. Nguyen PT, Selley DE, Sim-Selley LJ (2010): Statistical parametric map-ping reveals ligand and region-specific activation of G-proteins by CB1receptors and non-CB1 sites in the 3D reconstructed mouse brain. Neu-roimage 52:1243–1251.
9. Bohn LM, Lefkowitz RJ, Caron MG (2002): Differential mechanisms ofmorphine antinociceptive tolerance revealed in (beta)arrestin-2 knock-out mice. J Neurosci 22:10494 –10500.
0. Pertwee RG (1972): The ring test: A quantitative method for assessingthe “cataleptic” effect of cannabis in mice. Br J Pharmacol 46:753–763.
1. McKinney DL, Cassidy MP, Collier LM, Martin BR, Wiley JL, Selley DE, et al.(2008): Dose-related differences in the regional pattern of cannabinoidreceptor adaptation and in vivo tolerance development to delta9-tetra-hydrocannabinol. J Pharmacol Exp Ther 324:664 – 673.
2. Moise AM, Eisenstein SA, Astarita G, Piomelli D, Hohmann AG (2008): Anendocannabinoid signaling system modulates anxiety-like behavior inmale Syrian hamsters. Psychopharmacology (Berl) 200:333–346.
3. Nguyen PT, Holschneider DP, Maarek JM, Yang J, Mandelkern MA (2004):Statistical parametric mapping applied to an autoradiographic study ofcerebral activation during treadmill walking in rats. Neuroimage 23:252–259.
4. Lichtman AH, Cook SA, Martin BR (1996): Investigation of brain sitesmediating cannabinoid-induced antinociception in rats: evidence sup-porting periaqueductal gray involvement. J Pharmacol Exp Ther 276:585–593.
5. Pertwee RG, Wickens AP (1991): Enhancement by chlordiazepoxide ofcatalepsy induced in rats by intravenous or intrapallidal injections ofenantiomeric cannabinoids. Neuropharmacology 30:237–244.
6. Rawls SM, Cabassa J, Geller EB, Adler MW (2002): CB1 receptors in thepreoptic anterior hypothalamus regulate WIN 55212-2 [(4,5-dihydro-2-methyl-4(4-morpholinylmethyl)-1-(1-naphthalenyl-carbonyl)-6H-pyrrolo[3,2,1ij]quinolin-6-one]-induced hypothermia. J PharmacolExp Ther 301:963–968.
7. Smith PB, Martin BR (1992): Spinal mechanisms of delta 9-tetrahydro-cannabinol-induced analgesia. Brain Res 578:8 –12.
8. Lichtman AH, Martin BR (1991): Spinal and supraspinal components ofcannabinoid-induced antinociception. J Pharmacol Exp Ther 258:517–523.
9. Bohn LM, Gainetdinov RR, Lin FT, Lefkowitz RJ, Caron MG (2000): Mu-opioid receptor desensitization by beta-arrestin-2 determines mor-phine tolerance but not dependence. Nature 408:720 –723.
0. Bohn LM, Dykstra LA, Lefkowitz RJ, Caron MG, Barak LS (2004): Relativeopioid efficacy is determined by the complements of the G protein-coupled receptor desensitization machinery. Mol Pharmacol 66:106 –112.
1. Raehal KM, Bohn LM (2011): The role of beta-arrestin2 in the severity ofantinociceptive tolerance and physical dependence induced by differ-ent opioid pain therapeutics. Neuropharmacology 60:58 – 65.
2. Leterrier C, Bonnard D, Carrel D, Rossier J, Lenkei Z (2004): Constitutiveendocytic cycle of the CB1 cannabinoid receptor. J Biol Chem 279:36013–36021.
3. Leterrier C, Laine J, Darmon M, Boudin H, Rossier J, Lenkei Z (2006):Constitutive activation drives compartment-selective endocytosis andaxonal targeting of type 1 cannabinoid receptors. J Neurosci 26:3141–3153.
4. McDonald NA, Henstridge CM, Connolly CN, Irving AJ (2007): An essen-
tial role for constitutive endocytosis, but not activity, in the axonaltargeting of the CB1 cannabinoid receptor. Mol Pharmacol 71:976 –984.www.sobp.org/journal

4
4
4
4
4
4
724 BIOL PSYCHIATRY 2012;71:714–724 P.T. Nguyen et al.
w
35. Grimsey NL, Graham ES, Dragunow M, Glass M Cannabinoid Receptor 1trafficking and the role of the intracellular pool: Implications for thera-peutics. Biochem Pharmacol 80:1050 –1062.
36. Lein ES, Hawrylycz MJ, Ao N, Ayres M, Bensinger A, Bernard A, et al.(2007): Genome-wide atlas of gene expression in the adult mouse brain.Nature 445:168 –176.
37. Breivogel CS, Childers SR, Deadwyler SA, Hampson RE, Vogt LJ, Sim-Selley LJ (1999): Chronic delta9-tetrahydrocannabinol treatment pro-duces a time-dependent loss of cannabinoid receptors and cannabi-noid receptor-activated G proteins in rat brain. J Neurochem 73:2447–2459.
38. Gurevich EV, Benovic JL, Gurevich VV (2002): Arrestin2 and arrestin3 aredifferentially expressed in the rat brain during postnatal development.Neuroscience 109:421– 436.
39. Conner DA, Mathier MA, Mortensen RM, Christe M, Vatner SF, SeidmanCE, et al. (1997): beta-Arrestin1 knockout mice appear normal but dem-onstrate altered cardiac responses to beta-adrenergic stimulation. CircRes 81:1021–1026.
40. Huang CC, Chen YL, Lo SW, Hsu KS (2002): Activation of cAMP-depen-
dent protein kinase suppresses the presynaptic cannabinoid inhibitionww.sobp.org/journal
of glutamatergic transmission at corticostriatal synapses. MolPharmacol 61:578 –585.
1. Bass CE, Welch SP, Martin BR (2004): Reversal of delta 9-tetrahydrocan-nabinol-induced tolerance by specific kinase inhibitors. Eur J Pharmacol496:99 –108.
2. Lee MC, Smith FL, Stevens DL, Welch SP (2003): The role of severalkinases in mice tolerant to delta 9-tetrahydrocannabinol. J PharmacolExp Ther 305:593–599.
3. Garcia DE, Brown S, Hille B, Mackie K (1998): Protein kinase C disruptscannabinoid actions by phosphorylation of the CB1 cannabinoid recep-tor. J Neurosci 18:2834 –2841.
4. Rubino T, Forlani G, Vigano D, Zippel R, Parolaro D (2004): Modulation ofextracellular signal-regulated kinases cascade by chronic delta 9-tetra-hydrocannabinol treatment. Mol Cell Neurosci 25:355–362.
5. Rubino T, Forlani G, Vigano D, Zippel R, Parolaro D (2005): Ras/ERKsignalling in cannabinoid tolerance: From behaviour to cellular aspects.J Neurochem 93:984 –991.
6. Tappe-Theodor A, Agarwal N, Katona I, Rubino T, Martini L, Swiercz J, etal. (2007): A molecular basis of analgesic tolerance to cannabinoids.
J Neurosci 27:4165– 4177.




























