Embed Size (px)
Citation preview

1
ВАНЯ КОЛЕВА
ОБЩА ГЕНЕТИКАЛЕКЦИОНЕН КУРС
Шумен2014
ИЗДАТЕЛСТВО

2
Настоящото учебно помагало Обща генетика (Лекционен курс) е предназначено за студентите от специалност Екология и опазване на околната среда в Шуменски университет „Епископ Константин Преславски“. Съставено е в съответствие с част от учебната програма на дисциплината „Обща генетика и теория на еволюционното учение“. Включва 10 раздела, в които е представен съвременният прочит на класическата генетика.
Помагалото може да се използва и от студентите от специалност „Педагогика на обучението по ...“ при подготовката им за изпита по дисциплината „Генетика и еволюционни теории“.
Рецензенти: доц. д-р Ивона Димитровадоц. д-р Цветеслава Игнатова-Иванова
© Ваня Колева© Обща генетика. Лекционен курс
ISBN 978-954-9775-95-2
Художествено оформление и предпечатна подготовка: Илиян Петков
печат: “Химера” ООД - ШуменИздателство: “Химера” ООД, тел.: 054/ 830 585e-mail: [email protected], www.himraltd.com

3
СЪДЪРЖАНИЕ
Tема 1. Наследствена информация
Организация на ДНК, гени, хромозоми. Митоза. Мейоза .......................................... 5
Tема 2. Менделова генетика
Монохибридно и дихибридно кръстосване. Закони на Мендел. ................................8
Тема 3. Свързано с пола унаследяване
Унаследяване на признаци скачени с Х-хромозомата. Унаследяване на признаци
скачени с У-хромозомата. Псевдоавтозомно унаследяване. Ограничени от пола
признаци. Зависими от пола признаци ....................................................................... 12
Tема 4. Взаимодействия между гените
Видове доминантност. Пълно доминиране. Непълно доминиране. Кодоминиране.
Свръхдоминиране. Плейотропия. Леталност ........................................................... 15
Взаимодействие между гени от различни метаболитни пътища. Взаимодействие
между гени от един метаболитен път ........................................................................ 19
Тест за комплементарност ........................................................................................... 21
Комплементарно взаимодействие. Епистаза. Супресия. Полигенно взаимодействие.
Синтетичните летали. Гени-модификатори ............................................................... 24
Експресивност. Пенетрантност ................................................................................... 29
Tема 5. Унаследяване на скачени гени
Пълно скачване. Непълно скачване. Кросинговър .................................................... 29
Рекомбинантни и физични карти на хромозомите. Картиране на гените ............... 33
Двоен кросинговър. Интерференция .......................................................................... 34
Tема 6. Изменчивост на генетичния материал
Подвижни генетични елементи ................................................................................... 37
Мутации. Точкови мутации ......................................................................................... 39
Хромозомни мутации – класификация. Моноплоидия ............................................. 41
Хромозомни мутации – полиплоидия ........................................................................ 42
Хромозомни мутации – анеуплоидия. Генен баланс ................................................ 45
Хромозомни мутации – промени в структурата на хромозомите ........................... 48

4
Тема 7. Бактериална генетика
Конюгация при бактерии. F плазмид и полови типове бактерии. Hfr-клетки ........ 50
Картиране на бактериалните хромозоми ................................................................... 54
F плазмиди, носещи геномни фрагменти. F` плазмиди. R плазмиди ..................... 57
Трансдукция. Обща трансдукция. Котрансдукция. Специализирана трансдукция ..58
Трансформация ............................................................................................................ 61
Тема 8. Реализиране на генетичната информация. Репликация ............................. 62
Генетичен код ............................................................................................................... 66
Транскрипция и транслация – същност, основни етапи .......................................... 70
Tема 9. Извънядрени гени
Пластидна и митохондриална наследственост ......................................................... 73
Цитоплазмени мутации при човека. Цитоплазмена мъжка стерилност ................ 77
Тема 10. Количествена генетика
Унаследяване и генетична архитектура на количествени белези. Видове
количествени белези. Влияние на околната среда ................................................... 78
ЛИТЕРАТУРА .............................................................................................................. 81

5
Tема 1. Наследствена информация
Организация на ДНК, гени, хромозоми. Митоза. Мейоза
Науката генетика се занимава с изучаване на гените и на основните закономерности на наследствеността и изменчивостта при живите организми. Основоположник на класическата генетика е августинския монах Грегор Мендел, който през 1865 г. доказва, че белезите се предават в поколенията и изказва предположението, че това става чрез наследствени фактори. През 1933 г. Томас Хънт Морган доказва съществуването на наследствените фактори, които днес наричаме гени. Генът е най-малката единица за наследственост. Представлява структурен и функционален участък от молекулата на ДНК (Фигура 1.1). Пълната генетична информация на всеки организъм е кодирана в неговата ДНК и се нарича геном.
Фигура 1.1. Генът е структурен и функционален участък от молекулата на ДНК (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York.).
Хромозомният набор в соматичните клетки на диплоидните индивиди се състои от двойки хомоложни хромозоми (2n), поради това и гените са по двойки. Двойките хромозоми са хомоложни, а не еднакви, защото имат един и същ генен състав, но алелното състояние на гените от всяка двойка може да е различно (Фигура 1.2).
Фигура 1.2. Двойките хромозоми са хомоложни, защото алелното състояние на всяка двойка гени може да е различно.
Всяка хромозома е изградена от две части, наречени сестрински хроматиди, свързани в областта на центромерата (Фигура 1.3). Двете хроматиди съдържат две

6
идентични ДНК молекули. По време на клетъчното делене сестринските хроматиди се разделят и се превръщат в самостоятелни хромозоми (Фигура. 1.4). Митозата е етап от клетъчното делене, чрез който генетичният материал се разделя по равно между двете дъщерни клетки. Генетичната информация в родителската клетка и в дъщерните клетки е идентична.
Фигура 1.3. Морфологични особености на метафазна хромозома (по Г. Генова, Н. Харизанова, М. Пешева, Т. Декова, О. Конова-Кръстанова, П. Иванова, М. Китанова. 2012. ГЕНЕТИКА, Ръководство за упражнения, ГЕРА АРТ).

7
Фигура. 1.4. По време на мейоза се осъществява преход от диплоиден хромозомен набор към хаплоиден. (по Griffiths, A., J.H. Miller, D.T. Suzuki, R.S. Lewontin, W.M. Gelbart. 2004. An introduction to genetic analysis. 8 ed, W. H. Freeman and Company, New York.).
Клетките, които участват в процеса на размножаване на организмитe се наричат полови клетки. Хромозомният набор на зрелите полови клетки (гаметите) е хаплоиден (n). По време на мейоза се осъществява преход от диплоиден хромозомен набор към хаплоиден (Фигура. 1.4). Процесът мейоза се състои от две последователни клетъчни деления. Първото, наречено редукционно, намалява наполовина хромозомния набор. Второто, наречено еквационно (изравнително), не променя хромозомния набор, но намалява двукратно количеството на ДНК.

8
Интерфазата между двете деления е кратка и протича без да се осъществи репликация на ДНК. Поради това мейозата приключва с формирането на 4 дъщерни клетки с хаплоиден хромозомен набор (n). По време на оплождането ядрата на мъжката и женска гамета се сливат и се възстановява диплоидния хромозомен набор. Мейотичното клетъчно делене е важен процес за живите организми, защото осигурява запазване на диплоидния (2n) хромозомен набор в поколенията.
Tема 2. Менделова генетика
Монохибридно и дихибридно кръстосване. Закони на Мендел.
Генетичният подход за разбиране на всеки биологичен признак е да бъде установен набора от гени, който контролира формирането на този признак. Процесът се нарича генетично изследване. Един от генетичните методи на изследване е изолиране на всички мутантни форми на даден признак и определяне начина на унаследяване на този признак – чрез един или няколко гена и дали съответният ген се намира в автозома или в полова хромозома.
Първите изследвания на унаследяването на признаци са проведени от Грегор Мендел. Той не се е интересувал от признаците на градинския грах Pisum sativum, а от начина, по който се предават в поколенията. Кръстосване, при което се анализира унаследяването на една двойка автозомни гени и съответно на една двойка признаци се нарича монохибридно кръстосване. Това е най-простият вариант на едногенно (моногенно) унаследяване.
Пример за монохибридно кръстосване е унаследяването на признакът цвят на граховите семена (Фигура 2.1). Жълтият цвят на граховите семена е доминантен признак, а зеленият цвят – рецесивен. Растение, което е доминантно хомозиготно по генотип (АА), образува само жълти семена. Всичките му гамети носят само доминантен ген (А). Растение, което е рецесивно хомозиготно (аа), образува само зелени семена. Всичките му гамети носят рецесивен алеа (а). При кръстосване на такива хомозиготни (наречени още чисти линии) грахови растения, всички индивиди във F1 са с доминантен фенотип (образуват жълти семена) и са хетерозиготни (Аа). Тези хибриди образуват два вида гамети. По време на мейоза двойката алели на гена за цвят на граховите семена се разделя, като всеки алел попада в отделна гамета. Половината гамети носят доминантен алел (А), а другата половина – рецесивен алел (а). Това разпределяне на алелите е известно като първи закон на Мендел или закон за разпределянето по равно.

9
Фигура 2.1. Монохибридно кръстосване. Унаследяване на признакът цвят на семената на градинския грах.
При самоопрашване на хибридите от F1, във F2 ¾ (75%) от потомството е с доминантен фенотип (жълти семена) и ¼ (25%) – с рецесивен фенотип (зелени семена). Във F2 разпадането по фенопит е в съотношение 3:1, а по генотип – 1:2:1 (Фигура 2.1).
Кръстосване, при което едновременно се анализира унаследяването на две двойки нескачени автозомни гени и съответно на две двойки признаци се нарича дихибридно кръстосване. В опитите си да установи закономерностите в унаследяването на белезите Мендел кръстосва растения от две чисти линии (Фигура 2.2). Граховите растения от едната линия (ААВВ) са с доминантен фенотип – образуват жълти гладки семена. Растенията от втората линия (аавв) са с рецесивен фенотип – образуват зелени грапави семена. Родителските растения образуват по един вид гамети – носещи съответно алелите АВ и ав. Потомството във F1 е дихибридно (АаВв) и фенотипа му е семена с жълт цвят и гладка форма. То формира 4 вида гамети: АВ, Ав, аВ и ав. При самоопрашване на дихибридите, фенотипното разпадане във F2 е в съотношение 9 : 3 : 3 : 1 . Формират се 4 фенотипни класа: 9/16 от индивидите от F2 образуват жълти гладки семена (А_В_); 3/16 – жълти грапави

10
семена (А_вв); 3/16 – зелени гладки семена (ааВ_) и 1/16 – зелени грапави семена (аавв).
Фигура 2.2. Дихибридно кръстосване. Унаследяване на признаците цвят на семената и форма на семената на градинския грах.
Дихибридното кръстосване може да се разглежда като резултат от две независими монохибридни кръстоски. Разпадането във F2 само по признака цвят на граховите семена е 12/16 жълти : 4/16 зелени т.е. 12:4 = 3:1. Същото съотношение се наблюдава и за признака форма на граховите семена. Поради това независимо

11
унаследяване на двата признака Мендел прави предположението, че наследствените фактори за признака цвят на семената се комбинират независимо с наследствените фактори за признака форма на семената. Поради това съвременният вариант на втори закон на Мендел или закон за независимото комбиниране гласи: През мейоза двойките гени от различни двойки хомоложни хромозоми се групират независимо. Законът е в сила само, когата двойката гени, които детерминират анализираните признаци се намират в различни хомоложни двойки хромозоми.
Изучаването на едногенни признаци има място не само в експерименталната, но и в приложната генетика. Например, в селското стопанство са установени някои полезни признаци, като устойчивост към вредители или по-добра хранителни качества на някои сортове растения и породи животни.
Анализиращо кръстосване. Кръстосване, при което едната родителска форма е с доминантен фенотип, а другата е с рецесивен, се нарича анализиращо кръстосване. Родителската форма с рецесивен фенотип се нарича анализатор, а полученото потомство – анализиращо (Fb). Анализът на Fb позволява да се установи генотипа на родителската форма с доминантен признак (Фигура 2.3). Ако всички индивиди от Fb имат доминантен фенотип (няма фенотипно разпадане) извода е, че анализираният родител е хомозиготен по доминантните алели (АА). Ако се наблюдава фенотипно разпадане (в съотношение 1:1) извода е, че анализираният родител е хетерозиготен. Анализиращото кръстосване се прилага както при анализ на унаследяването на един, така и на два и повече признака. При анализиращо кръстосване на дихибрид фенотипното разпадане във Fb е в съотношение 1:1:1:1, т.е. броят на фенотипните класове е равен на броя на типовете гамети, които хетерозиготният родител образува.
Фигура 2.3. Анализиращо кръстосване. А – анализираната родителска форма градински грах е доминантна хомозиготна по генотип; Б – анализираната родителска форма градински грах е хетерозиготна по генотип.

12
Тема 3. Свързано с пола унаследяване
Унаследяване на признаци скачени с Х-хромозомата. Унаследяване на признаци скачени с У-хромозомата. Псевдоавтозомно унаследяване.
Ограничени от пола признаци. Зависими от пола признаци
Половият диморфизъм е характерен за повечето животински и за много от растителните видове. Полът се дефинира като съвкупност от специфични признаци и в повечето случаи се определя от половите хромозоми. При хората половите хромозоми (Х и Y) в по-голямата част от дължината си са нехомоложни (Фигура 3.1). Белтък кодиращите гени в Х храмозомата са 1098, а в Y хромозомата – 86. Броят на белтък кодиращите гени, установени и в Х и в Y хромозомата е само 54. Половите хромозоми произлизат от автозомна двойка хромозоми. Преди около 300 милиона години е започнал еволюционен процес, при който Y хромозомата губи голяма част от функционалните си участъци. Тази липса на гени в Y хромозомата се обозначава като хемизиготност. Днес Х и Y хромозомите са хомоложни само в малки участъци разположени в краищата им. Обозначават се като псевдоавтозомен район 1 и псевдоавтозомен район 2. През мейоза тези хомоложни участъци се свързват и така половите хромозоми подобно на автозомните хромозоми се разпределят в различни гамети.
Фигура 3.1. При човека половите хромозоми Х и Y са нехомоложни в по-голямата част от дължината си. (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York.).

13
При птиците и при някои видове риби мъжкият пол е хомогаметен (ZZ), а женският е хетерогаметен (ZW). Техните полови хромозоми се обозначават с Z и W, а не с Х и Y. Подобно на Х и Y, Z и W хромозомите на птиците произлизат от автозомни хромозоми. Половите хромозоми на насекомите се обозначават с Х и Y, като женският пол е хомогаметен (ХХ), а мъжкият е хетерогаметен (ХY). При някои насекоми механизмът за определяне на пола включва системата Х0. Мъжките насекоми имат само една полова хромозома (Х0), а женските имат две – ХХ.
При плодовата мушица Drosophila melanogaster, за разлика от бозайниците, детерминирането на пола зависи от съотношението между броя на Х хромозомите и автозомния набор (хаплоиден, диплоиден, триплоиден) (Х:А). Например от зигота с XХ полови хромозоми се развива нормален женски индивид, защото съотношението Х:А е 1 (2Х:2А). Нормална женска плодова муха се развива и от зигота с XXY полови хромозоми, защото съотношението Х:А също е 1 (2Х:2А). Y хромозомата носи само десетина гена и повечето са свързани с мъжката фертилност. Х и Y хромозомите са хомоложни само в един малък участък, съдържащ гените за рРНК. Този участък се нарича място за образуване на двойка. През мейоза тези хомоложни участъци се свързват, в резултат на което половите хромозоми на мъжките дрозофили се разпределят в различни гамети.
Унаследяването на гени разположени извън псевдоавтозомните райони на половите хромозоми, се нарича унаследяване, скачено с пола. То се отличава от унаследяване на автозомните гени. Скаченото с пола унаследяване дава различни фенотипни разпадания в реципрочните кръстоски.
Унаследяването на признаци, чиито гени са разположени в участъка, специфичен за Х хромозомата, се нарича унаследяване, скачено с Х хромозомата. Още в първите десетилетия на ХХ век Томас Морган демонстрира унаследяването на признак, скачен с Х хромозомата. Той изследва унаследяването на признака цвят на очите при плодовата мушица Drosophila melanogaster. В дрозофилната колекция на Т. Морган има линия мухи с червен цвят на очите и линия с бели (white) очи. Двата фенотипа се определят от двата алела на ген, разположен в участъка, специфичен за Х хромозомата. Кръстосването на женски дрозофили с червени очи (Хw+Хw+) с мъжки дрозофили с бели очи (ХwУ) дава потомство F1, в което всички индивиди са с червени очи, т.е. с фенотипа на майката (Фигура 3.2-А). Това показва, че червеният цвят на очите е доминантен признак, а белият – рецесивен. При кръстосването на индивидите от F1 помежду им, разпадането по фенотип във F2 е 3:1. Разпадането по пол е 1♀ : 1♂. Всички женски мухи (Хw+Хw+ и Хw+Хw) и половината от мъжките (Хw+У) са с червени очи. Другата половина мъжки мухи са с бели очи (ХwУ).

14
Фигура 3.2. Унаследяване на признак, скачен с Х хромозомата – цвят на очите при Drosophila melanogaster.
Реципрочната кръстоска дава различни резултати. Кръстосването на женска дрозофила (ХwХw) с бели очи с мъжка с червени очи (Хw+У) дава потомство (F1) от червенооки женски (Хw+Хw) и белооки мъжки дрозофили (ХwУ) (Фигура 3.2-Б). Признакът цвят на очите се е предал от майката на синовете и от бащата на дъщерите. Във F2 се получават мухи с червени очи и бели очи в съотношение 1:1. Това фенотипно разпадане се наблюдава във всеки от двата пола – женски: ½ червенооки (Хw+Хw) и ½ белооки (Хw Хw) и мъжки: ½ червенооки (Хw+У) и ½ белооки (ХwУ).
Унаследяване, при което признакът се предава от майката на синовете и от бащата на дъщерите, се нарича крис-крос или унаследяване на кръст. То е валидно само в случаите, когато рецесивният признак се внася от хомогаметния пол, независимо дали е женски или мъжки.
При човека хемофилията, далтонизмът, мускулната дистрофия на Дюшен и други заболявания са рецесивни, скачени с Х хромозомата признаци. Хипофосфатемията (устойчив на витамин Д рахит) се унаследява като доминантно, скачено с Х хромозомата заболяване.
Унаследяването на признаци, чиито гени са разположени в района,

15
специфичен за Y хромозомата, се нарича унаследяване, скачено с Y хромозомата. Такива признаци се унаследяват по права мъжка линия – от баща на синове. По този начин се унаследява една от формите на двустранна глухота, която е резултат от мутация в гена POU3F4, наличието на косми по ушната мида при мъже и др.
Унаследяването на признаци, чиито гени са разположени в псевдоавтозомен район на половите хромозоми, се нарича псевдоавтозомно унаследяване. При него фенотипното разпадане и при двете реципрочни кръстоски е в съотношение 3:1, т.е. наподобява автозомно унаследяване.
Ограничени от пола признаци се наричат признаците, които се формират само в единия пол. Гените, контролиращи ограничени от пола признаци могат да се намират в автозомна или в полова хромозома. Например, гените за млечност при говедата, гените за носливост при птици.
Зависими от пола признаци са признаци, чието формиране зависи от пола. Например, генът за плешивост при човека. Мъжете и жените с генотип АА са плешиви. Мъжете с генотип Аа също са плешиви, но хетерозиготните жени не са. По подобен начин се унаследява признакът наличие на рога при говедата.
Tема 4. Взаимодействия между гените
Видове доминантност. Пълно доминиране. Непълно доминиране.
Кодоминиране. Свръхдоминиране. Плейотропия. Леталност
Гените функционират като контролират химичните процеси, протичащи в клетката. Гените могат да са от един и същ или от различни метаболитни пътища. Във формирането на определен фенотип участват различен брой гени, които си взаимодействат. Взаимодействията са две големи групи. Първата група включва всички видове взаимодействия между алелите на един ген, наречени видове доминантност. Втората група са взаимодействията между алели на различни гени – взаимодействия между гени от един или от различни метаболитни пътища. Всеки метаболитен път представлява последователност от взаимосвързани стъпки (метаболитни стъпала).
Пълното доминиране е взаимодействие между алелите на един ген, при което доминантният алел напълно подтиска фенотипната проява на рецесивния алел. Поради това, фенотипът на хетерозиготните индивиди (Аа) не се отличава от този на доминантните хомозиготни (АА). Фенилкетонурията е автозомно рецесивно заболяване при хората. Започва от ранна детска възраст и може да доведе до

16
изоставане в умственото развитие. Резултат е от рецесивна мутация в гена, който кодира чернодробния ензим фенилаланин хидроксилаза (ФАХ). ФАХ преобразува фенилаланина, приеман с белтъчната храна в тирозин. При липса на нормален ензим се превръща във фенилпируватна киселина, която се натрупва в мозъка и забавя умственото развитие. Хетерозиготните индивиди имат фенотип от див тип (здрави), защото количеството ФАХ осигурено от единствения доминантен алел е достатъчно за нормалното функциониране на клетката.
Взаимодействие от типа пълно доминиране между алелите на един ген формира признаците цвят и форма на граховите зърна, височина на стъблото на граховите растения, цвят на козината при морските свинчета и др.
Непълното доминиране е взаимодействие между алелите на един ген, при което доминантният алел не подтиска напълно фенотипната проява на рецесивния алел. Поради това фенотипът на хетерозиготните индивиди (Аа) е междинен между фенотипа на доминантните хомозиготни (АА) и фенотипа на рецесивните хомозиготи (аа). По този начин се формира окраската на цветовете на нощната красавица (Фигура 4.1). Доминантните хомозитни растения имат червени цветове, рецесивните хомозиготни – бели, а фенотипът на хетерозитните е междинен – розови цветове.
Фигура 4.1. Фенотиповете на растението нощна красавица (Hesperis matronalis) са червен, розов и бял. (по Griffiths, A., J.H. Miller, D.T. Suzuki, R.S. Lewontin, W.M. Gelbart. 2004. An introduction to genetic analysis. 8 ed, W. H. Freeman and Company, New York.).
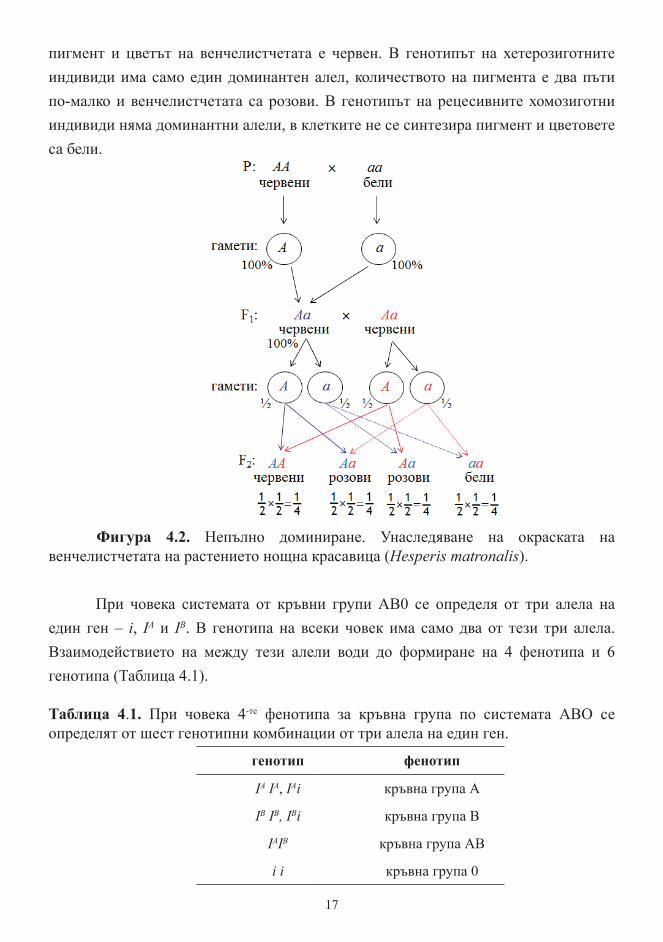
Непълното доминиране води до отклонения от менделовите разпадания. При самоопрашване на хибридите от F1, във F2 разпадането по фенотип съвпада с разпадането по генотип: 1:2:1 (Фигура 4.2). Всеки доминантен алел кодира образуването на определено количество пигмент. Наличието на два доминантни алела в генотипа на растението нощна красавица води до синтез на двойно количество
17
пигмент и цветът на венчелистчетата е червен. В генотипът на хетерозиготните индивиди има само един доминантен алел, количеството на пигмента е два пъти по-малко и венчелистчетата са розови. В генотипът на рецесивните хомозиготни индивиди няма доминантни алели, в клетките не се синтезира пигмент и цветовете са бели.
Фигура 4.2. Непълно доминиране. Унаследяване на окраската на венчелистчетата на растението нощна красавица (Hesperis matronalis).
При човека системата от кръвни групи АВ0 се определя от три алела на един ген – i, IA и IB. В генотипа на всеки човек има само два от тези три алела. Взаимодействието на между тези алели води до формиране на 4 фенотипа и 6 генотипа (Таблица 4.1).
Таблица 4.1. При човека 4-те фенотипа за кръвна група по системата АВО се определят от шест генотипни комбинации от три алела на един ген.
генотип фенотип
IA IA, IAi кръвна група А
IB IB, IBi кръвна група В
IAIB кръвна група АВ
i i кръвна група 0

18
Алелите от тази серия определят наличието и формата на антиген по повърхността на кръвната клетка. Антигенът е комплексна молекула, която се разпознава от имунната система. Алелът i не определя наличие на антиген. Алелите IA и IB определят наличие на две различни форми на този антиген. Те доминират пълно спрямо i. Поради това генотиповете IAi и IBi детерминират съответно фенотиповете кръвна група А и кръвна група В. В генотипът IAIB всеки от алелите продуцира собствен антиген и поради това по повърхността на кръвните клетки има по един от два вида антиген. Това взаимодействие между алелите IA и IB се нарича кодоминантно.
Типът на взаимодействие между алелите на един ген се определя от молекулярните функции на алелите и от фенотипното ниво на изследване. Заболяването сърповидно-клетъчна анемия е интересен пример за няколко вида взаимодействия. Доминантният алел HbА кодира синтеза на нормален хемоглобин А, а рецесивният алел HbS – на хемоглобин S. Доминантните хомозиготи (HbАHbА) имат червени кръвни клетки (еритроцити) с нормална форма (двойно вдлъбнат диск). В полипептидната верига на хемоглобин S липсва една аминокиселина. Поради това рецесивните хомозиготи (HbSHbS) имат сърповидна форма на еритроцитите.
Червените кръвни клетки със сърповидна форма имат ниска ефективност в пренасянето на кислород. От анемия страдат рецесивните хомозиготни индивиди (HbSHbS). Хетерозиготните индивиди (HbАHbS) са здрави, защото единственият доминантен алел (HbА) осигурява синтеза на достатъчно количество функционален хемоглобин. Следователно взаимодействието между доминантния HbА и рецесивния HbS алел по отношение на фенотипа страдащ от анемия е от типа пълно доминиране. По отношение на формата на клетките взаимодействието е два вида:непълното доминиране, защото много от клетките на хетерозиготните
индивиди (HbАHbS) имат леко сърповидна форма;кодоминиране, защото хетерозиготните индивиди (HbАHbS) имат червени
кръвни клетки с нормална форма и със сърповидна (Фигура 4.3).
Фигура 4.3. Червени кръвни клетки на човека. А – нормална форма, Б – сърповидна форма. (Clancy, S. 2008. Genetic mutation. Nature Education 1(1):187).

19
Свъхдоминирането е взаимодействие между доминантния и рецесивния алел, което води до по-голяма приспособеност на хетерозиготните индивиди. Маларията е изключително смъртоносна болест, причинявана от паразити. Сърповидните еритроцити имат способността да се струпват около паразитите и да ги извеждат от кръвния поток. Поради това хетерозиготните индивиди (HbАHbS) са по-устойчиви на малария, отколкото доминантните хомозиготи, които нямат еритроцити със сърповидна форма.
Генът кодиращ хемоглобина при човека е с плейотропно действие, защото определя повече от един признак. Например, при мишките има доминантен алел, който обуславя формирането на жълт цвят на козината. Хомозиготните по този алел зародиши имат сериозни нарушения в развитието и загиват преди раждането. Този алел се обозначава с AY и се нарича летален. Рецесивният алел се обозначава с а. При кръстосване на хетерозиготни (AYа) мишки помежду им в потомството се образуват зиготи в съотношение 1AYAY : 2AYа : 1аа (Фигура 4.4). До раждането оцеляват само зародишите с генотиповете AYа и аа, затова фенотипното разпадане във F1 е 2:1.
Фигура 4.4. Летални алели. Унаследяване на признакът наличие на опашка
при котки (по Попов П., Т. Ирикова. 2004. Обща генетика, Ун. изд. „Паисий Хилендарски“, Пловдив).
Взаимодействие между гени от различни метаболитни пътища. Взаимодействие между гени от един метаболитен път
Взаимодействие между гени от различни метаболитни пътища. При този тип взаимодействие фенотипът се формира от крайните продукти на два отделни метаболитни пътя. Например, окраската на царевичната змия е повтаряща се черно-оранжева камуфлажна шарка (див фенотип) (Фигура 4.5). Черният и оранжевият пигмент са крайни продукти от два отделни метаболитни пътя и оцветяват различни участъци от кожата. Генът А определя синтеза на оранжев пигмент, а генът В на черен пигмент. Генотипът на царевичните змии с див фенотип е А_B_. Фенотипът на змиите с генотип ааB_ е черен, защото не се синтезира оранжев пигмент. Змиите

20
с генотип А_bb имат оранжева окраска, защото в клетките няма черен пигмент. Двойно рецесивните хомозиготи (ааbb) са албиноси. Техният фенотип е резултат от мутации в двата независими метаболитни пътя. Бледорозовият цвят на албиносите се дължи на хемоглобина в кръвта, който прозира през кожата. Светлият мотив, около който се натрупват оранжевия и черен пигмент, се кодира от трети ген. Двата гена (А и В) са независими – намират се в две различни автозомни хромозоми. Поради това, фенотипното разпадане във F2 съответства на менделовото разпадане при дихибридно кръстосване 9/16 с камуфлажна окраска : 3/16 с черна окраска : 3/16 с оранжева окраска : 1/16 албиноси.
Фигура 4.5. Унаследяване на признака окраска при царевичната змия (по Griffiths A., S.R. Wessler, S.B. Carroll, J. Doebley. Introduction to genetic analysis. 2012, 10 ed, W. H. Freeman and Company, New York).
Взаимодействие между гени от един метаболитен път. При този тип взаимодействие фенотипът се формира в резултат от експресията на гени от един метаболитен път. Мутация във всеки един от гените в метаболитния път спира синтеза на крайния продукт и води до формиране на един и същ мутантен фенотип. Дивият тип гъби Neurospora синтезират необходимите им клетъчни компоненти като преработват неорганичните и органичните вещества от околната среда. В синтезът на аргинин участват три ензима, кодирани от три различни гена. Ако в генът arg-2+

21
има мутация няма да се синтезира нормален ензим 2, който превръща орнитина в цитрулин (Фигура 4.6). В хранителната среда трябва да се добави аргинин, защото клетката не може сама да го синтезира. Ако в генът arg-1+ или в генът arg-3+ има мутация, съответните клетки няма да могат да синтезират аргинин, поради липсата на ензим 1 или ензим 3.
Фигура 4.6. Взаимодействие между гени от един метаболитен път – синтез на аргинин в клетките на Neurospora.
Джордж Бидъл и Едуард Тейтъм получават Нобелова награда за изследването на този метаболитен път. Тяхната работа допринася за изясняване функциите на гените и формулирането на хипотезата един ген – един ензим. Всички белтъци, независимо дали са ензими, се кодират от гени. Поради това днес тази хипотеза е променена на един ген – една полипептидна верига. Това уточнение показва, че генът кодира физическата структура на белтъка, която от своя страна определя неговата функция. Трябва да отбележим, че не всички гени кодират белтъци. Някои гени кодират функционални РНК, които имат специфични функции. При транскрипцията на всички гени се синтезира РНК, но само при транскрипцията на белтък кодиращите гени се синтезира и РНК, и полипептидна верига.
Тест за комплементарност
При изследване на генетичните основи на даден признак или биологичен процес се използват мутантни индивиди с подобни фенотипове. Тестът за комплементарност позволява да се установи дали две рецесивни мутации са възникнали в различни участъци на един и същ ген или в различни гени.
В диплоидните организми тестът се провежда като се кръстосват индивиди, които са хомозиготни по две рецесивни мутации. Ако двете рецесивни мутации са в два различни гена (а1 и b1), всички индивиди в потомството ще притежават фенотип от див тип, защото мутантните алели комплементират (допълват се). (Фигура 4.7). Алелите комплeментират независимо дали се намират в една (Фигура 4.7-А) или в различни (Фигура 4.7-Б) хромозоми. Ако мутациите са в различни участъци на един ген, това води до поява на два различни рецесивни алела (а1 и а2) на този ген. Двата

22
алела не могат да комплeментират. Поради това при кръстосване на индивиди, които са хомозиготни по тези алели цялото потомство притежава мутантен фенотип (Фигура 4.8).
Фигура 4.7. Тест за комплементарност при диплоидни организми. Комплементарност на две рецесивни мутации в два различни гена разположени в: А – една хромозома; Б – две различни хромозоми.
Фигура 4.8. Тест за комплементарност при диплоидни организми. Комплементарност по две рецесивни мутации в един ген.
При хаплоидните организми комплементарността на гените не може да бъде изследванa чрез кръстосване. Способността на гъбите да образуват хетерокарион предоставя подходяща алтернатива. Хетерокарионите могат да се разглеждат като своеобразна диплоидна форма на гъбите, защото се формират чрез сливане на цитоплазмите на две клетки и запазване целостта на хаплоидните им ядра. Въз основа на фенотипът на хетерокарионът може да бъде установено дали два

23
гена комплeментират. Например, при Neurospora гените arg-А и arg-B кодират два различни ензима от метаболитния път за синтез на аргинин. Ако щам А е носител на рецесивна мутация в ген А (arg-а1), а щам B – в ген B (arg-b1), то генотиповете на двата щама са съответно arg-а1 arg-B и arg-А arg-b1 (Фигура 4.9). Фенотиповете и на двата щама са ауксотрофност по аргинин – клетките не могат да синтезират аргинин. Ако щам А и щам B се слеят, то генотипът на образувалия се хетерокарион е: arg-А arg-а1 arg-B arg-b1 (Фигура 4.9-А). Генните продукти се формират в общата цитоплазма, поради това алелите arg-А и arg-B проявяват съвместно своя доминантен ефект и формират хетерокарион с фенотип от див тип, т.е. прототрофност по аргинин. Рецесивна мутация в друг участък на гена А води до поява на нов алел (arg-а2) и поява на нов щам (щам D), с фенотип ауксотрофност по аргинин (Фигура 4.9-Б). Хетерокарионът на щам D и щам B е с генотип arg-А arg-а2 arg-B arg-b1. Поради комплементирането на алелите arg-А и arg-В, фенотипът е прототрофност по аргинин. Хетерокарионът на щам А и щам D е с генотип arg-а1 arg-а2 arg-B arg-B (Фигура 4.9-В). Неговият фенотип e ауксотрофност по аргинин, защото в генотипът няма алел arg-А, с който алелът arg-B да комплементира.
Фигура 4.9. Тест за комплементарност при хаплоидни организми. Комплементарност на две рецесивни мутации в гени от метаболитния път за синтез на аргинин при Neurospora: А – хаплоидни ядра с рецесивни алели arg-а1 и arg-b1 комплeментират до прототрофност на хетерокариона; Б – хаплоидни ядра с рецесивни алели arg-а2 и arg-b1 комплeментират до прототрофност на хетерокариона; В – хаплоидни ядра с рецесивни алели arg-а1 и arg-а2 не комплeментират и хетерокариона е ауксотроф.

24
Комплементарно взаимодействие. Епистаза. Супресия. Полигенно взаимодействие. Синтетичните летали. Гени - модификатори
При комплeментарното взаимодействие, доминантните алели на два гена от един метаболитен път се допълват (комплeментират) и формират фенотип от див тип. При загуба на функционалността на единия и/или на двата гена в резултат на рецесивна мутация, крайния продукт на метаболитния път не се синтезира. Например, дивият фенотип на венчелистчетата на растението камбанка (род Campanula) е синя окраска. Синият цвят се дължи на пигмента антоциан, който е краен продукт от серия биохимични преобразувания на няколко безцветни съединения (предшественици). Във всеки етап от тези преобразувания участват специфични ензими, кодирани от доминантни гени (Фигура 4.10).
Фигура 4.10. Взаимодействие между гени от един метаболитен път – синтез на син антоциан в клетките на венчелистчетата на растението камбанка.
Мутации във всеки един от тези гени води до натрупване на предшественик 1 или на предшественик 2 и не се синтезира син антоциан. Поради това камбанки с генотип А_bb, ааB_ и ааbb имат бели венчелистчетата. Двата гена се намират в две различни хромозоми. Кръстосването на растения с генотип ААbb и ааBB, дава потомство с генотип АаBb (Фигура 4.11). Тези дихетерозиготни камбанки имат сини венчелистчетата, защото доминантните алели А и B комплементират. Във F2 9/16 от камбанките имат сини венчелистчета, а 7/16 – бели (9:7).
Фигура 4.11. Комплeментарно взаимодействие. Унаследяване на признакът цвят на венчелистчетата на растението камбанка.

25
Епистаза е взаимодействие, при което един мутантен алел (а) маскира фенотипната проява на друг мутантен алел (b). Мутацията, чийто фенотип остава скрит, се нарича хипостатична, а съответният мутантен алел (b), се нарича хипостатичен. Мутацията, маскираща хипостатичния фенотип, се нарича епистатична, а мутантният алел (а) – епистатичен. Окраската на венчелистчетата на растението синеоката Мери (Collinsia parviflora) се формира чрез епистатично взаимодействие между два независими гена. Мутантният алел b определя формирането на цикламени венчелистчета, а мутантният алел а – бели (Фигура 4.12). Участието на епистатичният ген в метаболитния път на синтез на син антоциан е преди това на хипостатичния. Кръстосването на растения с генотип ААbb и ааBB, дава потомство с генотип АаBb. Тези дихетерозиготни растения имат сини венчелистчетата, защото доминантните алели А и B комплементират. Фенотипното разпадане в F2 e 9/16 растения със сини венчелистчета, 3/16 с цикламени венчелистчета и 4/16 с бели венчелистчета (9:3:4). Фенотипът на двойните мутанти (ааbb) съвпада с единия от двата мутантни фенотипа – бели венчелистчета. Поради това, фенотипният клас от 4/16 растения с бели цветчета е съставен от две групи индивиди – 1/16 с генотип ааbb и 3/16 ааB_. Този тип взаимодействие се нарича рецесивна епистаза.
Фигура 4.12. Рецесивна епистаза. Унаследяване на признакът цвят на венчелистчетата на растението синеоката Мери.
Епистатичен ефект могат да имат и доминантните мутантни алели. Взаимодействието се нарича доминантната епистаза. Признакът окраска на венчелистчетата на растението напръсник (Digitalis purpurea) се формира в резултат от взаимодействието на два независими гена (Фигура 4.13). Единият ген определя интензивността на червения пигмент в цветчетата: дивият фенотип е светло червена окраска (а), а мутантния фенотип е тъмно червена окраска (А). Другият ген е епистатичен и определя в кои клетки да се синтезира пигмент: b –

26
синтезира се пигмент и венчелистчета са оцветени, B – не се синтезира пигмент и венчелистчета са бели (моделът е опростен). Кръстосването на растения с генотип ААbb и ааBB, дава дихетерозиготно потомство (АаBb). Всички растения във F1 имат бели венчелистчетата, защото доминантният алел B не позволява синтез на червен пигмент. В потомството на дихибридните форми (АаBb) се наблюдават три фенотипа: 12/16 имат бяла окраска на венчелистчетата; 3/16 – тъмно червена (ааB_) и 1/16 – светло червена (ааbb). Фенотипният клас от 12/16 растения с бели венчелистчетата включва и двойномутантните форми (9/16 A_B_).
Фигура 4.13. Доминантна епистаза. Унаследяване на признакът цвят на венчелистчетата на растението напръстник.
Супресията е взаимодействие между мутантните алели на два независими гена, при което единия алел неутрализира ефекта от другия мутантен алел. В резултат от супресията се формира фенотип от див тип. Цветът на очите на Drosophila се определя от двойка алели – мутантният алел а кодира пурпурен цвят, а доминантният А – червен (Фигура 4.14). Мутантният алел b няма собствена фенотипна изява, но супресира фенотипната изява на алела а. Поради това фенотипът на двойните мутанти (ааbb) не е пурпурен цвят на очите, а див тип – червени очи. Кръстосването на дрозофили с генотип ААbb и ааBB, дава потомство с генотип АаBb и фенотип червени очи. Сред потомството на дихибридните форми (АаBb) от F1 се срещат два фенотипа: 13/16 от дрозофилите имат червен цвят на очите, а 3/16 – пурпурен. Фенотипното разпадане 13:3 се наблюдава, когато супресорът е

27
рецесивен и няма собствен фенотип. Ако рецесивният супресор и супресираният ген имат еднакъв фенотип, то разпадането във F2 е 10:6 (див:мутантен). Сепресията трудно се различава от епистазата. Ключовата разлика е, че супресорът спира еспресията на мутантния алел, а епистатичният алел маскира фенотипната проява на хипостатичния алел.
Фигура 4.14. Супресия. Унаследяване на признакът цвят на очите на Drosophila.
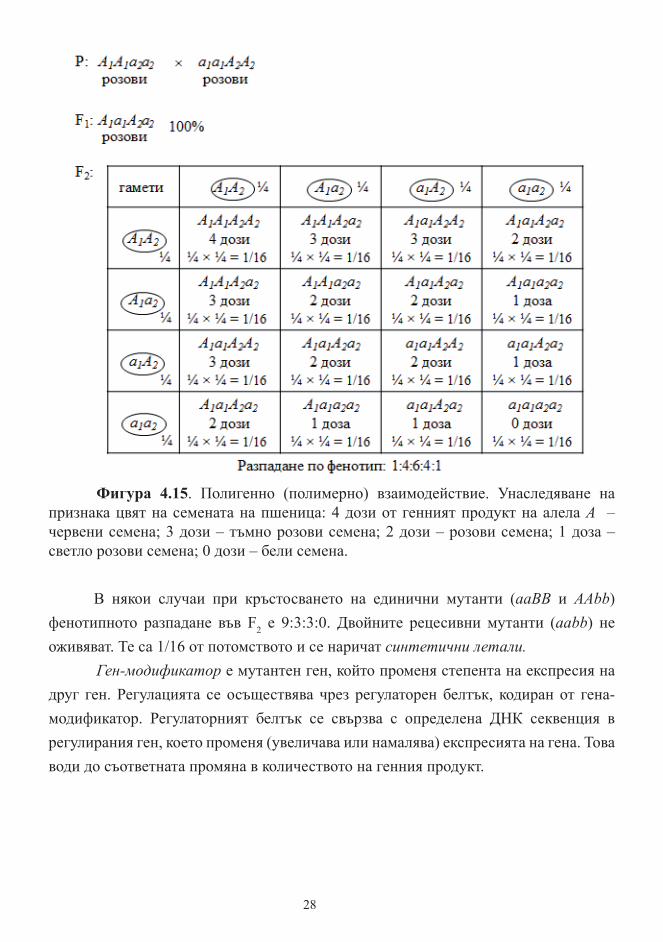
Полигенно (полимерно) взаимодействие е взаимодействие, при което степентта на изява на признака зависи от броя на доминантните алели. В метаболитният път за червен цвят на семената на пшеница участват два гена, кодиращи два ензима (моделът е опростен). Двата гена се обозначават с един и същ символ (А), защото кодираните от тях ензими използват един и същ предшественик, за да синтезират крайния продукт от този метаболитен път. Гените са различни, защото имат различни нуклеотидни секвенции и кодират различни ензими. Поради това се обозначават с различни индекси – А1 и А2. Всеки доминантен алел има определен количествен принос (доза) за интензивността на червения цвят. С добавянето на една доза от генния продукт на А1 или А2, интензивността на червения цвят на семената нараства пропорционално (Фигура 4.15). В генотиповете на потомството броя на доминантните алели варира от 4 (А1А1А2А2) до 0 (а1а1а2а2), а интензивността на окраската на зърната на пшеницата варира от червена до бяла.
28
Фигура 4.15. Полигенно (полимерно) взаимодействие. Унаследяване на признака цвят на семената на пшеница: 4 дози от генният продукт на алела А – червени семена; 3 дози – тъмно розови семена; 2 дози – розови семена; 1 доза – светло розови семена; 0 дози – бели семена.
В някои случаи при кръстосването на единични мутанти (ааBB и ААbb) фенотипното разпадане във F2 е 9:3:3:0. Двойните рецесивни мутанти (ааbb) не оживяват. Те са 1/16 от потомството и се наричат синтетични летали.
Ген-модификатор е мутантен ген, който променя степента на експресия на друг ген. Регулацията се осъществява чрез регулаторен белтък, кодиран от гена-модификатор. Регулаторният белтък се свързва с определена ДНК секвенция в регулирания ген, което променя (увеличава или намалява) експресията на гена. Това води до съответната промяна в количеството на генния продукт.

29
Експресивност. Пенетрантност
Експресивността е степента на развитие на фенотипа при определен генотип. Например, някои породи кучета с генотип аа имат кафява козина. Често окраската на отделните индивиди е с различна интензивност – от светло кафява (аа) до тъмно кафява (аа). Друг показател за оценка на фенотипната проява на гените е пенетрантността – процента на индивидите, които притежават фенотип, съответстващ на генотипа. Пенетрантността и експресивността са резултат от действието на различни условия в околната среда или на действието на други гени (модификатори, супресори или епистатични гени). Те са количествен израз на генната експресия в зависимост от околната среда и генния фон. Пенетрантността изразява процента на индивидите, в които гена се експресира, а експресивността – степента на експресия на гена.
Непълната пенетрантност и променливата експресивност затрудняват генетичните анализи, включително анализът на човешкото родословие и прогнозите в генетичните консултации. Много заболявания се детерминират от алели с непълна пенетрантност. Поради това при някои от носителите на такъв алел не се наблюдават признаци на заболяването. От друга страна е възможно при анализ на родословни схеми да бъдат идентифицирани носителите на такъв алел.
Tема 5. Унаследяване на скачени гениПълно скачване. Непълно скачване. Кросинговър
Гени, които са разположени в една и съща хромозома се наричат скачени гени и образуват една група на скачване. Броят на групите на скачване съответства на броя на хомоложните двойки хромозоми. При човекът хомоложните хромозоми са 23 двойки и групите на скачване са 23. Унаследяването на белези, детерминирани от скачени гени се нарича скачено унаследяване, а самото явление – скаченост на гените.
Скачените гени на дихетерозиготите могат да са в две комбинации: двата доминантни алела са разположени в един и същ хомолог (АB) на двойката хомоложни хромозоми или в различните хомолози (Аb и аB). Тези комбинации се наричат съответно цис структура и транс структура:

30
Пълно скачване на гени. Скачеността на гените възпрепятства независимото комбиниране на гените. При миризливото секирче признаците виолетови венчелистчета (А) и продълговати поленови зърна (B) са доминантни. Рецесивните признаци са червени венчелистчета (а) и кръгли поленови зърна (b) (Фигура 5.1). Двата гена са скачени. При анализиращо кръстосване на дихетерозитно растение с цис структура, фенотипните класове във Fb са два. Съотношението е 1:1, което показва че скачеността на гените е пълна. Фенотипните класове, наблюдавани във Fb се наричат родителски (нерекомбинантен) тип класове, защото повтарят комбинацията от признаци на родителските форми.
Фигура 5.1. Пълно скачване на гени. Унаследяване на признакът цвят на венчелистчетата на растението миризливо секирче.
Непълно скачване на гени. Скачването на гените може да е и непълно. Дължи се на процесът кросинговър – размяна на хомоложни участъци между двойка хомоложни хромозоми. Протича през мейоза по време на конюгацията на хомоложните двойки хромозоми. Най-простият случай на кросинговър е с участието на една хроматида от всяка хомоложна хромозома (Фигура 5.2).

31
Фигура 5.2. Кросинговър. Размяна на хомоложни участъци между двойка хомоложни хромозоми.
При дрозофила закърнелите крила са резултат от рецесивната мутация vestigial wings (vg), а пурпурният цвят на очите – от рецесивната мутация purple eye color (pr). Томас Морган установява, че при анализиращо кръстосване на дихетерозиготна женска във Fb 4
-те фенотипни класа не са в очакваното съотношение 1:1:1:1 (Фигура 5.3). Два от фенотипните класове са родителски тип и включват почти равен брой индивиди. Те са по-многобройни и са общо 87% от Fb. Получени са при оплождане на гамети, които не са претърпели кросинговър в участъците с интересуващите ни гени. Тези гамети се наричат гамети от родителски тип или гамети от нерекомбинантен тип.

32
Фигура 5.3. Непълно скачване на гени. Унаследяване на признака цвят на очите на плодовата муха.
Комбинацията от признаци на другите два фенотипни класа е съответно червени очи, нормални крила и пурпурни очи, закърнели крила (Фигура 5.3). Те не повтарят комбинацията от признаци в родителските форми (червени очи, закърнели крила и пурпурни очи, нормални крила). Поради това се наричат рекомбинантен (неродителски) тип класове. Рекомбинантният тип фенотипни класове са с нова комбинация на признаците. Получени са при оплождане на гамети, които са претърпели кросинговър в участък с един от интересуващите ни гени (vg или pr). Тези гамети се наричат гамети от неродителски тип или гамети от рекомбинантен тип. Фенотипните класове от рекомбинантен тип са по-малобройни и общата им честота е 13%. Следователно, вероятността да протече кросинговър между тези два гена, т.е. честотата на рекомбинация между двата гена (Rf – Recombinant frequency) е 13%. Rf се изчислява по формулата:

33
При анализиращо кръстосване на дихетерозигота във Fb се получават резултати, които се характеризират с общ модел:
Честотите на двата нерекомбинантни класа са почти равни и общо са повече от 50%.
Честотите на двата рекомбинантни класа са почти равнии общо са по-малко от 50%.
Ако по време на мейоза протече кросинговър между хромозомни участъци носещи една двойка гени, то само 50% от гаметите ще са от рекомбинантен тип (Фигура 5.2). Поради това честотата на кросовърните индивиди по отношение на тази двойка гени не може да е повече от 50%. На практика не всяко мейотично делене завършва с образуване на кросовърни гамети.
Рекомбинантни и физични карти на хромозомите. Картиране на гените
Хромозомната карта представя последователността на гените в хромозомата и разстоянието между гените. Хромозомните карти са два вида: физични (цитологични) и рекомбинантни (генетични). Физичните карти представят гените, като участъци от ДНК молекулата, а рекомбинантните карти – като точка. В генетичната карта всеки ген заема строго определено място, наречено локус.
Честотата на рекомбинация между два гена е основен метод за съставяне на хромозомни карти. Рекомбинантните карти включват само гени с известни мутантни алели и мутантни фенотипове. Разстоянието между два гена в рекомбинационната карта съответства на Rf на тези два гена. В чест на Томас Морган единицата за разстояние между два локуса в генетичната карта е наречена сантморган (сМ). Един сантморган съответства на 1% честота на рекомбинация. Установено е, че колкото е честотата на кросинговър между два гена, толкова е разстоянието между тези два гена. За гените vg и pr на дрозофила Rfvg-pr е 13% (Фигура 5.3). Следователно, разстоянието между тези два гена е 13 сМ: vg – pr = 13 сМ.
Физичните карти са карти на действителната геномна ДНК. Те представят гените като участък от ДНК. Единицата за разстояние на физичната карта е броя на базовите двойки (б.дв.). За удобство разстоянията често се представят в килобази (кб): 1кб = 1 000 б.дв. от ДНК. Физичната карта на една ДНК молекула се съставя чрез секвениране на голям брой малки фрагменти. Чрез компютърен анализ се установяват специфични базови последователности. След това, чрез сравняване с база от данни за други организми се идентифицират гените, съдържащи тези

34
секвенции. Отделните индивиди в една популация се различават по някои базови последователности. Тази молекулярна хетерозиготност служи като ориентир (молекулярен маркер) във физичната карта. Някои от гените на физичната карта не са отразени в рекомбинантната. Такива гени се наричат кандит-гени. Част от тях се наричат гени, обект на интерес, защото тяхната функция все още не е установена.
Рекомбинантните и физичните карти отразяват различни страни на генома (Фигура 5.4). Те предоставят възможност да бъде разбрана функцията на гена на молекулно ниво и значението й за фенотипа на индивида. Физичната карта дава информация за вероятното действие на гените на ниво клетка. Рекомбинантната карта дава информация за ефекта от действието на гена на ниво фенотип.
Фигура 5.4. Физична карта и генетична карта на хромозома 2 на гъбата Magnaporthe grisea (http://nar.oxfordjournals.org/content/30/1/121/F1.expansion).
Двоен кросинговър. Интерференция
Броят на гените във всяка ДНК молекула е различен: от десетки до няколко стотици. Поради това е възможно да се осъществят едновременно два (двоен кросинговър) или повече кросинговъра (множествен кросинговър) по дължината на хромозомата (Фигура 5.5).

35
Фигура 5.5. Двоен кросинговър. Размяна на два хомоложни участъка между двойка хомоложни хромозоми.
Вероятността да протече двоен кросинговър е произведение от вероятностите за двата единични кросинговъра. Например, в първа хромозома на доматите последователно са разположени три гена: за форма на плода (А – кръгла, а – овална), височина на растението (B – нормален ръст, b – джудже) и наличие на петна по листата (D – без петна, d – с петна). Разстоянието между гените е А-B = 21сМ и B-D = 12сМ. Поради това вероятността за единичен кросинговър между тези гени е съответно RfА-B = 21% и RfB-D = 12%. Следователно вероятността за двоен кросинговър в този участък е 2.52%:
тRfА-B-D = RfА-B × RfB-D = 21% × 12% = 21% × 0.12 = 2.52%
тRfА-B-D е теоретично очакваната честота за двоен кросинговър в участъка, в който са локализирани гените A, B и D.
Доматено растение, което е трихетерозиготно по отношение на тези признаци може да образува 8 типа гамети (Фигура 5.6).
Фенотипните класове във Fb формират 4 класа, като всеки клас се състои от 2 групи. Четирите класа представляват различен процент от Fb. Най-голям е делът на родителския тип индивиди. Най-малоброен е фенотипния клас на двойно кросовърните (двойно рекомбинантните) индивиди. Той е резултат от реално протекъл двоен кросинговър, наречен експериментален двоен кросинговър. Останалите два фенотипни класа са резултат от единичен кросинговър. Броят на индивидите в тези класове е междинен – по-малък от броя на индивидите във фенотипния клас с родителски тип индивиди и по-голям от броя на индивидите във фенотипния клас на двойнокросовърния тип индивиди. Честотата на единичния кросинговър между гените A и B (RfА-B) се изчислява по формулата:

36
Честотата на експерименталния двоен кросинговър се изчислява по формулата:
Често двойният кросинговър се осъществява с по-малка честота от теоретично очакваната. Поради това, делът на експериментално получените двойно кросовърни индивиди е по-малък от теоретично очаквания, а стойността на еRfА-B-D е по-малка от стойността на тRfА-B-D. Този феномен се нарича интерференция (И). Силата на интерференцията се определя чрез коефициента на коинциденция (с) или коефициента на съвпадение. Коефициентът се изчислява по формулата:
Стойността на коефициента на съвпадение варира между 0 и 1 и показва, какъв е дела на реално осъществените от теоретично очакваните двойно кросовърни обмени. Например, ако с = 0.65, то реално са се осъществили 65% от теоретично очакваните 2.52% двойни кросинговъри. Интерференцията се изчислява по формулата:
И = 1 – с
Стойността на И варира между 0 и 1. Силата на интерференцията е различна в различните участъци на една и съща хромозома и в различните хромозоми. Установено е, че колкото е по-малко разстоянието между двата единични кросинговъра, толкова интерференцията в този район е по-силна и по-рядко се осъществява двоен кросинговър. В някои райони от хромозомата не протича двоен кросинговър. В тези случаи интерференцията е пълна: с = 0, а И = 1.

37
Фигура 5.6. Анализиращо кръстосване растение домати, което е трихетерозиготно по отношение на признаците форма на плода (А – кръгла, а – овална), височина на растението (B – нормален ръст, b – джудже) и наличие на петна по листата (D – без петна, d – с петна).
Tема 6. Изменчивост на генетичния материал
Подвижни генетични елементи
Подвижните ДНК елементи (транспозонни елементи) притежават уникалната способност да се “движат” в генома. Процесът се нарича транспозиция. Осъществява се с участието на ензима транспозаза. Тези елементи са пръснати из целия геном и са около 45% от човешката ДНК.
В зависимост от начина на преместване подвижните ДНК елементи са две основни групи:
1. ДНК транспозони – преместват се директно като ДНК; 2. Ретротранспозони – преместват се с участието на междинна РНК.
В зависимост от вида на клетката са:І. Бактериални подвижни елементи:
1. IS елементи;2. Транспозони.
ІІ. Еукариотни подвижни елементи:1. ДНК транспозони:
1.1. Ас и Ds елементите; 1.2. Р-елементи.

38
2. Ретротранспозони:2.1. Ретротранспозонните с дълги терминални повтори;2.2. Ретротранспозони без дълги терминални повтори:
2.2.1. Дълги разпръснати елементи;2.2.2. Къси разпръснати елементи.
І. Бактериални подвижни елементи. Повечето подвижни елементи в бактериите се преместват директно като ДНК и се наричат вмъкната секвенция (IS – Insertion Sequences). Установени са повече от 20 различни IS елемента. Подвижните ДНК елементи имат секвенция, кодираща ензима транспозаза. Ензимът извършва три действия: 1) Изрязва IS елемента от донорната ДНК; 2) Срязва приемащата ДНК и 3) Свързва IS елемента с приемаща ДНК. IS се местят в случайни места.
Вторият вид подвижни елементи в бактериите са транспозоните. Те имат дълги кодиращи секвенции и носят различен брой гени. Генът за устойчивост към някои антибиотици се намира в транспозон.
ІІ. Еукариотни подвижни елементи. 1. ДНК транспозони. Ас и Ds елементите (Ас – Activator/активатор, Ds –
Dissociation/разделяне) са еукариотни ДНК транспозони. За пръв път са описани през 1940 г. от Барбара МакКлинток, която изследва оцветяването на семената на царевица. Ас елементите са автономни, защото кодират ензима транспозаза. Ds елементите възникват в резултат на отпадане на участъци от секвенцията на Ас елементите, кодираща транспозаза. Поради това Ds са неавтономни – те се местят само с участието на кодираната от Ас транспозаза. Преместването на Ас/Ds елемент може да доведе до поява на нов алел с нови функции.
Р-елементите са автономни ДНК транспозони – кодират транспозаза. Преместването им е на принципа “изрязване и поставяне”. Съществуват и неавтономни Р-елементи. Преместването им е възможно само, ако в генома на клетката има поне един автономен Р-елемент, който да осигури транспозаза.
2. Ретротранспозони. В еукариотите повечето подвижни елементи са ретротранспозони. Подобно на ретровирусите, при преместването им се синтезира междинна РНК. Те са две групи: с дълги терминални повтори (LTRs – Long Terminal Repeats) и без дълги терминални повтори.
Ретротранспозонните с LTRs. В двата края на тези ретротранспозони има LTRs. Наричат се вирусни ретротранспозони, защото кодират всички белтъци на ретровирусите (с изключение на белтъците необходими за обвивката им). Поради това LTR ретротранспозоните могат да се преместват в нови участъци от ДНК на клетката-гостоприемник, но не могат да се преместват в други клетки.

39
Ретротранспозони без LTR. Наричат се още невирусни ретротранспозони. Те са много разпространени в генома на бозайниците. Наблюдават се и в протозои, насекоми и растения. По време на еволюцията чрез повторно копиране в същите места и в нови места от генома на висшите еукариоти са акумулирани голям брой ретротранспозони без LTR.
Подвижните ДНК елементи нямат основни функции за клетката, но вероятно имат голямо значение в еволюцията на днешните организми. Около 50% от спонтанните мутации в дрозофила са резултат от преместване на подвижни ДНК елементи в рамките на ген или около ген. При мишките този процент е около 10, а при човекът е само 0.1-0.2%. Някои генетични болести при човека са свързани с мутантни алели, в които са включени подвижни елементи.
Транспозицията на еукариотни подвижни елементи в половите клетки е механизъм, чрез който преместените секвенции се предават в следващото поколение. По този начин по време на еволюцията подвижните елементи се мултиплицират и бавно се акумулират в еукариотния геном. Транспозиция протича и в соматичните клетки, но подвижните елементи се предават само в дъщерните клетки.
Мутации. Точкови мутации
Двата основни източника на генетична изменчивост са мутациите и рекомбинациите. Мутациите са изменения в молекулата на ДНК. Могат да представляват промени в базовите двойки, промени в структурата на хромозомата или промени в броя на хромозомите. Възникват спонтанно или в резултат от въздействие с радиация или химични съединения. Повечето мутации се отстраняват от репариращите системи на клетката, които идентифицират и поправят уврежданията в ДНК молекулите. Към ниските нива на мутации съществува известна толерантност, защото те са източник на генетично разнообразие.
Точкови мутации. Мутациите, които възникват в рамките на един ген се наричат генни мутации или точкови мутации. Терминът се отнася основно за мутации, които представляват промяна в една базова двойка или в малък брой базови двойки. Точковите мутации са две основни групи: замяна на бази и промяна в броя на базите.
1. Замяната на бази са мутации, при които една базова двойка в молекулата на ДНК се заменя от друга. Замените са два вида: транзиции и трансверзии. Транзицията е замяна с база от същата химична категория. Например, замяна на една пуринова база с друга пуринова (A с Г) или на пиримидинова с пиримидинова (Ц с Т). Трансверзията е замяна с база от друга химична категория. Например,

40
замяна на пуринова база с пиримидинова (А с Ц) или на пиримидинова с пуринова (Ц с А). Тези замени могат да засягат базови двойки и от двете ДНК вериги.
2. Промяната в броя на базите е отпадане или вмъкване на нуклеотидни двойки. Наричат се индел-мутации (insertion–deletion). Тези мутации променят рамката на четене на иРНК – новата рамка на четене е от мястото на мутацията до новия стоп-кодон. Промяната в рамката на четене се нарича фреймшифт мутация. Резултатът е, че след мястото на тази мутация АК последователност на белтъка е много различна от оригиналната АК последователност. Поради това обикновено фреймшифт мутациите водят до пълна загуба на нормалната структура и функция на кодирания белтък.
Фенотипните последици от точковите мутации зависят от това в кой участък от гена възникват. Ако са в кодиращия район на гена, могат да доведът до дегенериране на генетичния код или до поява на стоп-кодони. В зависимост от това се делят на три основни вида: Тихите мутации (мутации-синоними) превръщат кодон, кодиращ определена аминокиселина в друг кодон, кодиращ същата аминокиселина (АК). Те не променят АК последователност на полипептидните вериги и поради това нямат фенотипен ефект. Мутациите с промяна на смисъла (missense) превръщат кодон, кодиращ определена аминокиселина в кодон, кодиращ друга аминокиселина. Част от тези мутации са неутрални, защото замяната е с химически еквивалентна АК. Не се променят функциите на синтезирания белтък и нямат фенотипен ефект. Мутациите без смисъл (nonsense) превръщат кодон за аминокиселина в стоп-кодон. Поради това се синтезира по-къса полипептидна верига и се променят функциите на синтезирания белтък. Тези мутации имат фенотипен ефект (появява се нов вариант на фенотипа).
Точковите мутации могат да възникнат и в регулаторни или други некодиращи секвенции. Некодиращият участък на гена съдържа много ключови секвенции – места за свързване с РНК полимеразата, белтъци регулиращи транскрипцията и други белтъчни фактори. Мутациите, които разрушават тези места не променят структурата на кодирания белтък. Те променят експресията на гена в определен момент или в определена тъкан, или променят отговора към сигнали от околната среда. Мутациите, възникнали в местата за свързване нарушават експресията на гена и синтеза на генния продукт. Поради това е трудно да се предскажат последиците от мутации в тези участъци.

41
Хромозомни мутации – класификация. Моноплоидия
Хромозомните мутации представляват промени в броя и структурата на хромозомите. Класификация:
І. Промени в броя на хромозомите:1. Аберантна еуплоидия:
1.1.Моноплоидия;1.2.Полиплоидия:
а) автополиплоидия; б) алополиплоидия.
2. Анеуплоидия. ІІ. Промени в структурата на хромозомите:
1. Делеции;2. Дупликации;3. Инверсии;4. Транслокации.
І. Промените в броя на хромозомите представляват промени в броя на ДНК молекулите в клетката. Аберантна еуплоидия (І.1.) е промяна в броя на хромозомните набори. Аберантните еуплоиди имат хромозомен брой, който е по-голям или по-малък от нормалния за вида хромозомен набор. Моноплоидите (І.1.1.) имат само един хромозомен набор (n). Те са индивиди на диплоидни видове. Намират приложение при създаване на нови линии растения с желани белези. Например, нови, полезни за селското стопанство рецесивни мутации, се откриват само в рецесивни хомозиготни диплоиди. Моноплоидни растения могат да бъдат получени чрез тъканни култури. В гаметите на диплоид, за който се предполага, че притежава полезна рецесивна мутация, се предизвиква митотично делене до образуване на ембриоид (маса от моноплоидни клетки). След това от ембриоида се отглежда моноплоидно растение, а от него се получава фертилен диплоид (2n). Така от 2n родителска форма с генотип АаБб може да бъде създаден моноплоид с генотип аб, а след това – диплоид с генотип аабб (Фигура 6.1).
Фигура 6.1. Създаване на моноплоидно растение чрез методите на тъканните култури. (по Griffiths, A., J.H. Miller, D.T. Suzuki, R.S. Lewontin, W.M. Gelbart. 2004. An introduction to genetic analysis. 8 ed, W. H. Freeman and Company, New York).

42
По този начин са създадени растения, устойчиви към хербициди или към токсичните съединения на растителни паразити. Принципът на метода е следния: Моноплоидни клетки се поставят в среда, съдържаща хербицид. Само от тези клетки, които са резистентни към хербицида прорастват ембриоиди. От тях се получават резистентни моноплоидни, а след това – резистентни диплоидни растения. Този метод е много по-бърз и по-ефективен от методите на традиционната селекция. Чрез методите на тъканните култури са създадени някои ценни линии соя, тютюн и захарно цвекло.
В индивидуалното развитие на някои растения, животни и гъби се наблюдават периоди, през които всички клетки или част от клетките имат един хромозомен набор (хаплоиден, n) и периоди, през които имат два хромозомни набора (диплоиден, 2n). За тези видове редуването на хаплофаза и диплофаза е част от нормалното развитие. Техните индивиди с един хромозомен брой (n) също се наричат моноплоиди. Например, моноплоиди са мъжките пчели, осите и мравките. В нормалният жизнен цикъл на тези видове мъжките се развиват чрез партеногенеза (ембриона се развива от специален вид неоплодени яйцеклетки). Гаметите на мъжките индивиди се образуват чрез митоза.
Хромозомни мутации – полиплоидия
Диплоидните индивиди имат два хромозомни набора (2n). Полиплоидите (І.1.2) са аберантни еуплоиди, които имат повече от два хромозомни набора. Те могат да бъдат с 3n (триплоиди), 4n (тетраплоиди), 5n (пентаплоиди) и др. Полиплоидията се среща често при растенията и много рядко при животните. Увеличаването на плоидността е важен начин за поява на нови растителни видове. Често се свързва с увеличаване размера на организма, като се запазват пропорциите на тялото. Например, тетраплоидите са много по-големи от съответните диплоиди (Фигура 6.2).
Фигура 6.2. Млади растения захарно цвекло, създадени чрез методите на тъканните култури. А – диплоид, Б – тетраплоид. (по Yildiz, M. 2013. Current Progress in Biological Research, Publisher: InTech.).

43
Полиплоидите са два вида – автополиплоиди (І.1.2.1) и алополиплоиди (І.1.2.2). Автополиплоидите произлизат от собствения си вид. Удвояването може да е спонтанно или да бъде предизвикано. Въздействието с химични съединения, като колхицин и колцемид, се спира формирането на нишките на делителното вретено. Това води до удвояване на хромозомния брой и на всички гени в генотипа (Фигура 6.3). Например, ако 2n клетка е с генотип АаВв, то 4n ще бъде с генотип ААааВВвв, а 8n – с ААААааааВВВВвввв. Хромозомните набори на автополиплоидите могат да са и нечетен брой – 3n, 5n и др. Триплоидите се появяват спонтанно в природата, но могат да бъдат създадени и чрез кръстосване на 2n и 4n. По правило 3n са стерилни. Например, бананите които са широко разпространени в търговската мрежа са стерилни 3n (n=11), защото не образуват семена.
Фигура 6.3. Създаване на тетраплоидна клетка чрез въздействие с колхицин.
(по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).
Алополиплоидите са полиплоиди, произлезли от различни видове. Имат две или повече копия на хромозомния набор на всеки родителски вид. Например, зелето (Brassica) и ряпата (Raphanus) са диплоиди с 2n=18 (Фигура 6.4). Гаметите им съответно са n1 = 9 и n2 = 9. Техният хибрид (n1 + n2 = 2n = 18) е стерилен алополиплоид, защото 9-те хромозоми на двете родителски форми не са хомоложни хромозоми и не конюгират през мейоза. В резултат на спонтанно хромозомно удвояване една част от тези растения могат да образуват фертилни семена, от които възникват нови тетраплоидни растения (4n = 36). По този начин от стерилния хибрид на ряпата и зелето е получен Raphanobrassica. Тези алополиплоидни растения са фертилни и понякога се наричат амфидиплоиди или удвоени диплоиди.

44
Фигура 6.4. Създаване на Raphanobrassica – алополиплоид от зеле и ряпа.
В природата алополиплоидията е един от основните начини за поява на нови видове растения. Алополиплоидите могат да бъдат създадени и чрез методите на тъканни култури. Принципът е следния: Първо се разрушават клетъчните стени на две диплоидни клетки от два различни вида. След това се сливат клетъчните им мембрани и се образува една клетка с две ядра. Това често води до сливане и на ядрата. Формираната 4n клетка се поставя в хранителна среда, която е обогатена с подходящи хормони и минерали. При тези условия 4n клетката дава начало на ново растение. Алополиплоидните растения са важни за селското стопанство. Памукът и пшеницата са естествени алополиплоиди, а Triticale е изкуствен амфидиплоид. Triticale е получен от пшеница (Triticum, 6n = 42) и ръж (Secale, 2n = 14). Хромозомният му брой е 2n = 2×(21 + 7) = 56. Това ново растение комбинира високия добив на пшеницата и устойчивостта на ръжта.
Полиплоидията е често срещана при растенията. Също така е сравнително обичайно явление за земноводни и влечуги. При бозайниците се среща много рядко и в повечето случаи полиплоидните индивиди загиват преди или скоро след раждането. Описани са няколко случая на деца с полиплоидия, които оцеляват до една година след раждането. Описан е случай на момиче на възраст от 26 месеца,

45
чийто кариотип е от 92 автозомни хромозоми и четири Х хромозоми (Фигура 6.5). Основните клинични признаци са лицев дисморфизъм, силно изоставане в растежа и интелектуалното развитие.
Фигура 6.5. Кариотип на момиче с тетраплоидия. (по Guc-Scekic, M, J. Milasin, M. Stevanovic, L.J. Stojanov, М. Djordjevic. 2002. Tetraploidy in a 26-month-old girl (cytogenetic and molecular studies). CLINICAL GENETICS, 61:62-65).
Хромозомни мутации – анеуплоидия. Генен баланс
Анеуплоидите (І.2.) са индивиди, чийто хромозомен набор се отличава от нормалния за вида с отделни хромозоми. Хромозомният брой на анеуплоидите може да е както по-голям, така и по-малък. Класификацията на диплоидните анеуплоиди се основава на броя на копията на хромозомите. Индивид в хромозомния набор, на който липсва една хромозома (2n – 1) се нарича монозомик. Хромозомният набор на нулизомикът е с липсваща двойка хомоложни хромозоми (2n – 2), а в набора на двойния монозомик липсват две хромозоми от две различни хомоложни двойки (2n – 1 – 1). Анеуплоидите с допълнителни хромозоми са съответно тризомик (2n + 1) и двоен тризомик (2n+1+1). Хаплоидите с (n + 1) се наричат дизомик. Отбелязването на анеуплоидията по отношение на половите хромозоми е чрез изброяване на копията на всяка полова хромозома. Например XXY, XYY или XXX. Липсата на втора полова хромозома се означава с 0 (нула) – Х0.
Анеуплоидията е спонтанен процес на неразделяне на хроматидите на хромозомите по време на мейоза или митоза. Неразделянето през митоза води

46
до поява на анеуплоидни участъци в тялото на индивида (мозаичен анеуплоид). Неразделянето на хомоложни хромозоми в гаметите на родителските форми води до образуване на анеуплоидни гамети (Фигура 6.6) и поява на анеуплоидни индивиди в потомството. Анеуплоидията може да е резултат и от протичането на кросинговър, когато формираните хиазми се запазят.
Фигура 6.6. Образуване на анеуплоидни гамети – неразделяне при първо мейотично делене и неразделяне при второ мейотично делене. (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).
Анеуплоидията променя формата и основните характеристики на тялото. При човека монозомията с една липсваща Х хромозома (2n = 44 + Х0) води до състояние известно като Синдром на Търнър. Засегнатите хора имат характерен фенотип – стерилни жени, нисък ръст, наличие на кожна ивица между врата и рамената. Някои от когнитивните им функции са нарушени. Известни са и други форми на монозомия, като всички са свързани с неблагоприятни последици за индивида. Синдромът на Клайнфелд е тризомия 2n=44+ХХY, а синдромът на

47
Даун е тризомия с допълнителна 21-ва хромозома (2n+1). За разлика от аберантната еуплоидия, анеуплоидията променя формата и основните характеристики на тялото на индивида. Анеуплоидите се характеризират с много повече аномалии от полиплоидите. Например, татула (Datura stramonium) има 12 хромозомни двойки. Полиплоидната форма татул има по-големи размери от диплоидната форма, но е със същите пропорции. Известни са и всичките 12 тризомии на татула (Фигура 6.7). Всяка тризомия дава специфична форма на семенната кутийка. Дванадесетте монозомии на татула се различават, както помежду си, така и от съответните тризомии.
Фигура 6.7. Всяка от 12-те тризомии на татула дава специфична форма на семенната кутийка (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).
Различната степен на наблюдавани аномалии в полиплоиди, тризомии и монозомии вероятно се дължи на промените в генния баланс. Генният баланс е съотношение между генните продукти в клетката, което осигурява нейното нормално функциониране. Например, ако генът А се намира в хромозома 1, а гена В в хромозома 2, то съотношението между двата гена в 2n индивид е 2А:2В = 2:2 т.е. А:В = 50%:50%. За 4n индивид съотношението също е А:В = 50%:50% (4А:4В = 4:4). За тризомик по хромозома 2 съотношението е 2А:3В = 2:3, а за монозомик по същата хромозома – 2А:1В = 2:1. Количеството на генния продукт на даден ген е правопропорционално на броя на копията на този ген. С увеличаване на броя на копията на един ген, се увеличава и количеството на белтъка кодиран от този ген. Тази зависимост се нарича ефект ген-доза.

48
Хромозомни мутации – промени в структурата на хромозомите
Хромозомните мутации, които променят структурата на хромозомите се наричат хромозомни аберации или хромозомни преустройства. Възникването им е свързано с разкъсване на двойната верига на ДНК и неточно възстановяване или невъзстановяване на разкъсването от репариращите (поправящите) системи на клетката. Ако тези системи правилно свържат двата разкъсани края, не се осъществява хромозомна аберация. Хромозомните аберации са 4 основни групи:
1. Делеция – загуба на участък от хромозомата;2. Дупликация – удвояване на участък от хромозомата;3. Инверсия – завъртане на вътрешен участък от хромозомата на 180 градуса;4. Транслокация – преместване на участък от една хромозома в друга,
нехомоложна хромозома (Фигура 6.8).Оцеляват само хромозомни аберации, в резултат на които се получават
хромозоми с една центромера и два теломера. Ако се образува хромозома без центромера (ацентромерна хромозома), през анафаза тя не се придвижва към полюсите на делителното вретено и не се включва в нито едно от двете дъщерни ядра. Поради това, ацентромерните хромозоми не се унаследяват. Когато се образува хромозома с две центромери (дицентрична), в повечето случаи нишките на делителното вретено издърпват центромерите към двата срещуположни полюса. Това води до формиране на хроматиден мост (Фигура 6.9), който се разкъсва на случайно място по дължината на хромозомата. Делециите (Фигура 6.10) и разкъсването на хроматидните мостове обикновено са свързани със загуба на теломерен участък.
Хромозомните аберации свързани със загуба или удвояване на голям участък нарушават генния баланс в клетката и са причина за фенотипни аномалии. Двойно верижното разкъсване на ДНК (ДВР-ДНК) в нуклеотидната последователност на даден ген нарушава функционалността на този ген. Хибридните гени са съставени от участъци от два съседни гена. Формират се в местата на възстановени разкъсвания и са нефункционални. Делециите и дупликациите са мутации, които променят броя на гените и нарушават генния баланс на клетката.
В някои групи организми има последователности на ДНК, които са повторени в същата или в друга хромозома. По време на мейоза е възможно нехомоложни хромозоми с такива хомоложни участъци да конюгират частично. Протичането на кросинговър в тези райони на конюгация се нарича неалелна хомоложна рекомбинация Неалелната хомоложна рекомбинация е причина за поява на хромозомни аберации в конюгиралите хромозоми.

49
Фигура 6.8. Хромозомни аберации – промени в структурата на хромозомите.
Фигура 6.9. Хроматиден мост в анафазна клетка от коренова меристема на Allium cepa L. (Kalcheva, V., A. Dragoeva, K. Kalchev, D. Enchev. 2009. Cytotoxic and genotoxic effects of Br-containing oxaphosphole on Allium cepa L. root tip cells and mouse bone marrow cells. Gen. Mol. Biol. 32(2):389-393).

50
Фигура 6.10. Делеция в клетка от костен мозък на бели лабораторни мишки третирани с Br-oxph: триизмерено изображение на част от метафазна пластинка. (Koleva, V., A. Dragoeva, A. Andreeva, M. Burova, S. Georgiev, D. Enchev. 2013. Comparative analysis of clastogen-induced chromosome aberrations observed with light microscopy and by means of atomic force microscopy. Mutation Research, 753(1):29-35).
Тема 7. Бактериална генетика
Конюгация при бактерии. F плазмид и полови типове бактерии. Hfr - клетки
Бактериалната генетика изучава организацията и функционирането на бактериалния геном. Тя дава началото на най-новото направление в науката генетика – съвременната молекулярна генетика. Технологиите, разработени чрез бактериите и техните вируси са в основата на постижения, като установяването на пълната последователност на генома на хората и успешното приложение на ДНК анализите в криминалистиката.
Бактериите са най-многобройните организми на планетата. Отличителната им черта е, че нямат обособено клетъчно ядро и тяхната ДНК се намира в цитоплазмата. Поради това се класифицират като прокариоти. Геномът на повечето бактерии е една кръгова двойно верижна ДНК молекула (Фигура 7.1). Бактериите имат и допълнителни ДНК молекули, наречени плазмиди. Повечето плазмиди също са кръгови ДНК, но са с много по-малък размер от геномната ДНК. Бактериите могат да паразитират чрез специфични вируси наречени бактериофаги или фаги. Генетичният материал на фагите е къса хромозома от ДНК или РНК. Вирусите се репродуцират, като използват молекулните механизми на клетката-гостоприемник.

51
Фигура 7.1. ДНК на бактерия и на бактериофаг.
Мутационният процес в бактериалната ДНК не се отличава от мутационния процес в еукариотните организми. Бактериите са много удобни моделни организми, защото са многобройни, лесно се отглеждат и сравнително лесно се изследват редките генетични събития. Размножават се чрез безполово клетъчно делене, което се предшества от репликация на ДНК. При бактериите и фагите няма мейоза, но протича конюгация. Бактериалната конюгация е процес, подобен на половото размножаване и на рекомбинацията: клетка-донор предава генетичен материал на клетка-реципиент.
В бактериалната генетика най-често използваният тест-обект е Escherichia coli. Размножава се чрез просто делене, но въпреки това клетките се обозначават като мъжки или женски тип. Някои бактерии имат в цитоплазмата си кръгов плазмид, наречен F плазмид. Те се приемат за мъжки тип и се обозначават с F+ (Фигура 7.2). Клетките, които нямат F плазмид се приемат за женски тип и се обозначават с F–. При бактериалната конюгация F– клетка може да се превърне в F+. Механизмът е следния: F– и F+ бактерия се притеглят една към друга чрез пили (тънки тръбести структури). Синтезира се едноверижно копие на кръговия плазмид на F+ бактерията. Плазмидът се завърта и насочва копието, което през пилата преминава в F– клетката. Втората верига на едноверижното копие се синтезира в приемащата клетка. Така чрез репликация се осигурява по едно копие на F плазмида за донорната и за приемащата бактерия.

52
Фигура 7.2. Бактериална конюгация – превръщане на F- клетка във F+ клетка.
В хромозомата и в F плазмидът има няколко копия на IS елементи. Например, F плазмидът на E. coli К12 има по едно копие на IS2 и Tn1000 и две копия на IS3 (Фигура 7.3). Това осигурява райони на хомоложност между ДНК на F плазмида и геномната ДНК на бактериалната клетка и възможност за хомоложна рекомбинация. Хомоложната рекомбинация между IS в ДНК на F плазмида и съответният IS в геномната ДНК, води до интегриране на F плазмида в хромозомата на приемащата клетка. Бактерии с интегриран F плазмид се обозначават като Hfr бактерии, защото се характеризират с висока честота на рекомбинация (Hfr – High Frequency of Recombination). Интегрираният плазмид не се реплицира самотоятелно. Когато Hfr бактерия конюгира с F– бактерия, репликацията на F плазмида започва от място за начало на трансфер (oriT – origin of transfer) (Фигура 7.3). Районът, обозначаван като

53
tra район (tra – transfer), включва oriT и всички гени на F плазмида, необходими за осъществяване на преноса. В някои случаи се реплицират и гени на геномната ДНК, които са разположени след интегрирания F плазмид. Синтезираното едноверижно ДНК копие се избутва през пилата. В приемащата клетка чрез репликация се изгражда комплементарна верига (Фигура 7.4). Двойноверижната ДНК на F плазмида има райони, които са хомоложни и с геномната ДНК на приемащата клетка. Това дава възможност F плазмида да рекомбинира с ДНК на приемащата F– бактерия чрез двоен кросинговър. По този начин част от гените на геномната ДНК на Hfr-клетката стават част и от геномната ДНК на приемащата клетка. Такава клетка се нарича ексконюгат. Тази клетка има две копия на един участък от ДНК – едно копие от F– клетката и едно копие от Hfr клетката. В този стадий ексконюгатът е частичен диплоид и се нарича мерозигота. Бактериалната генетика е генетика на мерозиготите.
Фигура 7.3. F плазмид на E. coli К12 (по Madigan, M.T., J.M. Martinko, D. Stahl, D.P. Clark. 2010. Brock Biology of Microorganisms, 13th Edition, Pearson Inc.).

54
Фигура 7.4. Hfr бактерия. (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).
Рекомбинацията при бактериите се отличава с някои особености. При двойният кросинговър се получава цяла кръгова ДНК и фрагмент, който се губи при следващото делене на бактерията. Клетките са хаплоидни и поради това при бактериалната рекомбинация няма реципрочна размяна на генетичен материал. Пренасянето на гени и кросинговърът дават възможност бактериалните хромозоми да бъдат картирани.
Картиране на бактериалните хромозоми
Картиране чрез прекъсване на конюгацията. Това е метод за картиране на гените в хромозомата на Hfr бактерии. Основава се на прекъсване през определен период от време на конюгацията на Hfr и F– клетки. Поради това единицата за разстояние между гените е минута. Колкото по-рано се прекъсне конюгацията, толкова по-малко са пренесените гени. Ексконюгатите се изследват, за да се установи кои гени от Hfr са рекомбинирали с хромозомата на F– клетката. Ако генът а+ премине в ексконюгата 10 мин. след генът b+, това означава че разстоянието между двата гена в бактериалната хромозома е 10 мин. (Фигура 7.5). След 17 мин. ще бъде пренесен и генът c+. Следователно генът b+ се намира на разстояние 7 мин. от гена c+. Съставената по този начин карта представлява генетична карта на

55
бактериалната хромозома.
Фигура 7.5. Картиране на бактериална хромозома чрез прекъсване на конюгацията. (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).
Картиране въз основа на честотата на рекомбинация. Генетичната карта на хромозомата на Hfr клетка може да бъде съставена и въз основа на честотата на рекомбинация между непълното геномно ДНК копие от Hfr клетка (екзогеном) и геномната ДНК на F– клетката (ендогеном). F– клетка с ендогеном и екзогеном е мерозигота, защото е частично диплоидна. След двоен кросинговър в мерозигота оцелява само рекомбинантния продукт, който е включен в бактериалната хромозома. Това позволява лесно идентифициране на рекомбиниралите гени. Въз основа на честотата на рекомбинация на гените се установява последователността им и разстоянието между тях. Тази генетична карта е по-прецизна от генетичната карта построена чрез прекъсване на конюгацията.
В резултат на мутации някои клетки са устойчиви към определени антибиотици или други инхибитори. Тези бактерии могат да се размножават в среда с инхибитор. Наричат се резистентни мутанти, а мутантните алели – генетични маркери или маркерни алели. Честотата на рекомбинация на гените се определя с помощта на генетични маркери. Например, генетичната карта на участък от
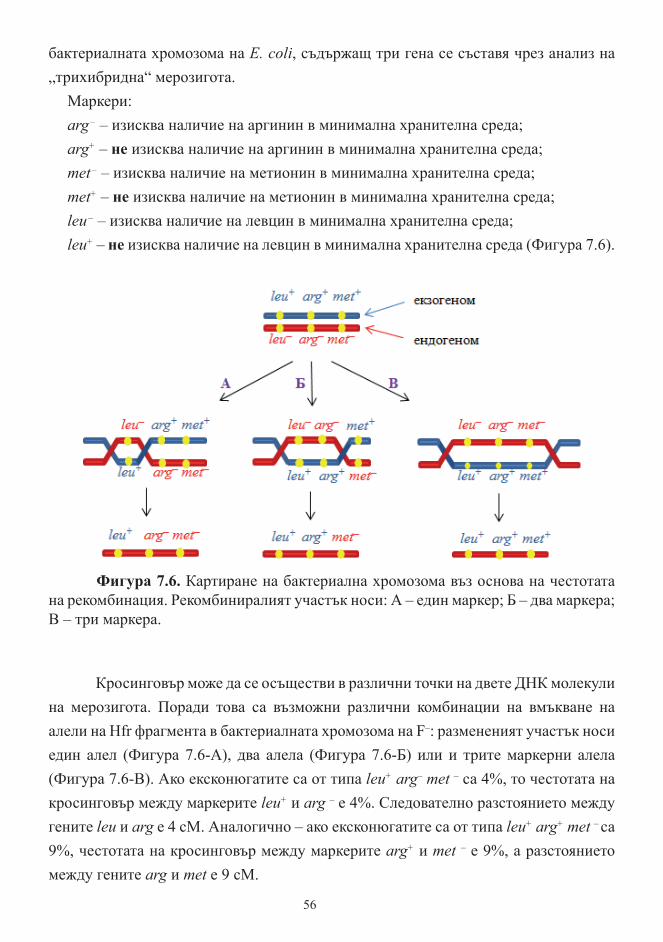
56
бактериалната хромозома на E. coli, съдържащ три гена се съставя чрез анализ на „трихибридна“ мерозигота.
Маркери:arg– – изисква наличие на аргинин в минимална хранителна среда;arg+ – не изисква наличие на аргинин в минимална хранителна среда;met– – изисква наличие на метионин в минимална хранителна среда;met+ – не изисква наличие на метионин в минимална хранителна среда;leu– – изисква наличие на левцин в минимална хранителна среда;leu+ – не изисква наличие на левцин в минимална хранителна среда (Фигура 7.6).
Фигура 7.6. Картиране на бактериална хромозома въз основа на честотата на рекомбинация. Рекомбиниралият участък носи: А – един маркер; Б – два маркера; В – три маркера.
Кросинговър може да се осъществи в различни точки на двете ДНК молекули на мерозигота. Поради това са възможни различни комбинации на вмъкване на алели на Hfr фрагмента в бактериалната хромозома на F–: размененият участък носи един алел (Фигура 7.6-А), два алела (Фигура 7.6-Б) или и трите маркерни алела (Фигура 7.6-В). Ако ексконюгатите са от типа leu+ arg– met – са 4%, то честотата на кросинговър между маркерите leu+ и arg – е 4%. Следователно разстоянието между гените leu и arg е 4 сМ. Аналогично – ако ексконюгатите са от типа leu+ arg+ met – са 9%, честотата на кросинговър между маркерите arg+ и met – е 9%, а разстоянието между гените arg и met е 9 сМ.

57
F плазмиди, носещи геномни фрагменти. F` плазмиди. R плазмиди
Понякога, заедно с F плазмидът от бактериалната хромозома се отделя и част от геномната ДНК на Hfr (Фигура 7.7). Такъв плазмид се нарича F` плазмид. Неговата ДНК е съставена от ДНК на F плазмида и част от геномната ДНК. F` плазмидите са резултат от неправилно изрязване на F плазмид. Наименованието на F` плазмидите се определя от носеният геномен ген. Например, F` плазмидът носещ геномния ген lac се нарича F` lac. F` плазмидите са частично диплоидни. Поради това намират приложение при изследване на алелното състояние на нови мутации. Някои F` плазмиди носят до ¼ от бактериалната хромозома.
Фигура 7.7. Формиране на F` lac - F` плазмид, носещ геномния ген lac. (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).
R плазмидите са група плазмиди, в които са включени гени за резистентност. Плазмидът R6 носи гена за резистентност към тежки метали, а Тi – гена за канцерогенност. Agrobacterium tumefaciens е Грам-отрицателна бактерия, която паразитира по двусемеделни растения (Фигура 7.8-А). Фенолните съединения секретирани от наранените растения активират гените в Тi плазмида (Тi – тумор индуциращ). Механизмът, чрез който бактерията паразитира включва интегриране

58
на част от бактериалната ДНК в геномната ДНК на растителната клетка. Това води до промени в растителния метаболизъм и развитието на тумори в растителната тъкан (Фигура 7.8-Б). R плазмидите, подобно на F плазмидите, се пренасят чрез бактериална конюгация. R плазмидите носещи алели за резистентност към антибиотик често са част от транспозони. pBR 322 и pUC са R плазмиди от E. coli, които намират широко приложение в клонирането на ДНК.
Фигура 7.8. Плазмидът Ti участва в механизма, чрез който бактерията Agrobacterium tumefaciens паразитира по растения. А – Agrobacterium tumefaciens; Б – туморно образование по растение заразено с Agrobacterium tumefaciens. (http://www.cals.ncsu.edu/course/pp728/Agrobacterium/Alyssa_Collins_profile.htm).
Трансдукция. Обща трансдукция. Котрансдукция. Специализирана трансдукция
Бактериофагите или фагите са вируси, които паразитират в бактериите (Фигура 7.1). Техният генетичен материал е ДНК или РНК. Обозначават се със символи – Т4, λ, Р1 и др. Механизмите на размножаване на бактериофагите са два. Наричат се литичен и лизогенен цикъл.
При литичният цикъл фагите атакуват бактерията и инжектират генетичния си материал в цитоплазмата. Бактериалният клетъчен апарат спира синтеза на бактериалните компоненти и започва репликация и експресия на генетичния материал на фагите. Изграждат се много нови фаги, които разрушават бактериалната клетъчната стена и клетката загива. Процесът на разрушаване на бактериалната клетка се нарича лизис, а популацията от нови фаги отделени в околната среда – фагов лизат. Фагите които се размножават чрез литичен цикъл се наричат вирулентни фаги. Те веднага лизират клетката-гостоприемник и атакуват други бактерии.

59
При лизогенният цикъл фагите инфектират клетката без да я убиват. Тези фаги се наричат умерени фаги. Тяхната ДНК се интегрира в бактериалната ДНК или остава в цитоплазмата като плазмид. Интегрираната фагова ДНК се нарича профаг. Фаговата ДНК се реплицира едновременно с бактериалната хромозома и при деленето на бактерията се предава в дъщерните клетки. Такива бактерии се описват като лизогенни. Понякога умереният фаг, носен от лизогенна бактерия се активира и започва многократна репликация и експресия на фаговата ДНК. Накрая изградените многобройни нови фаги лизират бактерията и атакуват други клетки.
Бактериофагите могат да пренасят гени от донорна бактерия в реципиентна. Процесът на пренос на гени с посредничеството на бактериофаги се нарича трансдукция. Тя е два вида – обща и специализирана. При общата трансдукция фагът носи случайна част от бактериалната хромозома (Фигура 7.9). Например
Фигура 7.9. Обща трансдукция. (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).
фагът Р1 разкъсва бактериалната хромозома на E. сoli на малки фрагменти. В някои от новоформираните фаги вместо фагова ДНК погрешно се включват фрагменти от бактериалната ДНК. Образувалият се бактериофаг се нарича трансдуциращ

60
фаг. Той атакува друга бактерия и е възможно чрез рекомбинация носената от него ДНК да се включи в ДНК на приемащата клетка. Възможно е два близко разположени бактериални гена да бъдат включени в един трансдуциращ фаг. Те могат да бъдат интегрирани в хромозомата на друга бактерия. Този процес се нарича котрансдукция. Колкото по-близко са разположени гените, толкова по-висока е честотата на котрансдукция. Поради това котрансдукцията намира приложение при картиране на бактериалните гени.
При специализираната трансдукция фагът пренася точно определена част от бактериалната хромозома на клетката-гостоприемник в хромозомата на клетката-реципиент. Този фаг се нарича специализиран трансдуктор. Фагът λ е най-добре изследваният специализиран трансдуктор на E. coli. Винаги се включва в специфичен участък между гените gal и bio, които кодират галактоза и биотин (Фигура 7.10). Включването е чрез рекомбинация между специфичните свързващи места в λ фага и в бактериалната хромозома. Процесът се катализира от кодирани от фага ензими. Изрязването често е неправилно (Фигура 7.11-А). В този случай кръговата ДНК на фага носи бактериален ген (gal или bio), а част от фаговите гени остават в бактериалната хромозома (Фигура 7.11-Б). Фагът, носещ бактериалния ген gal се означава като λdgal (λ-defective gal).
Фигура 7.10. Специализирана трансдукция – включване на λ фага в бактериалната хромозома на E. coli. (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).

61
Фигура 7.11. Специализирана трансдукция – изрязване на λ фага от бактериалната хромозома на E. coli. А – правилно изрязване на λ фага; Б – погрешно изрязване на λ фага. (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).
Трансформация
Трансформацията е процес, при който бактериалната клетка приема ДНК от околната среда (трансформираща ДНК). В някои случаи ДНК е отделена от мъртва бактериална клетка, а в други – от жива. Източник на ДНК може да е клетка от същия или от друг вид. Генотипът на приемащата бактерия се променя (трансформира), когато трансформиращата ДНК е от клетка с генотип, различен от генотипът на приемащата бактерия. Трансформацията се осъществява в три основни стъпки:
1. ДНК-свързващ комплекс, който е разположен в клетъчната стена, захваща трансформиращата ДНК;
2. Бактериални ензими разкъсват едната верига на трансформиращата ДНК до нуклеотиди;
3. Втората верига на трансформиращата ДНК (цялата или част от нея) се включва в бактериалната хромозома чрез процес, който е подобен на двойния кросинговър (Фигура 7.12).
Трансформацията намира широко приложение в генното инженерство и в генната терапия, защото е изключително удобен метод за работа. Тя дава възможност бактериалния генотип да бъде променян съобразно целите на изследването.
62
Фигура 7.12. Трансформация. (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).
Тема 8. Реализиране на генетичната информация
Репликация
Репликацията е механизъм, чрез който клетката удвоява ДНК молекулите си и осигурява по едно копие от генетичната си информация за двете дъщерни клетки. Удвояването на ядрения геном е сложен процес, който се осъществява от комплекс от ензими. Те осигуряват изключително точно и еднократно копиране на всяка ДНК молекула. ДНК репликацията се осъществява по полуконсервативен механизъм както в прокариотните клетки, така и в еукариотните. Поради това съществува голямо сходство в процеса на удвоява ДНК молекулите им.
Репликацията започва от консервативна секвенция на ДНК, наречена репликационно начало. Прокариотните клетки имат една хромозома с едно репликационно начало (Фигура 8.1-А). Повечето еукариотните клетки имат няколко линейни хромозоми като по дължината на всяка една от тях има много места с репликационно начало. Големият брой точки за начало на репликация позволява бързо удвояване на големия еукариотен геном (Фигура 8.1-Б).

63
Фигура 8.1. Начало на репликацията. А – еукариотни клетки, Б – прокариотни клетки. (по O‘Donnell, M., L. Langston, B. Stillman. 2013. Principles and Concepts of DNA Replication in Bacteria, Archaea, and Eukarya. Cold Spring Harb Perspect Biol, doi: 10.1101/cshperspect.a010108).
Реплизома е комплексът от ензими, който осъществява репликацията. Включва ДНК полимерази, хеликаза, праймаза и SSB белтъци. Реплизомата има три основни функции:
разрушава нуклеозомите в родителските вериги на ДНК чрез отделяне на хистоните от ДНК;
копира родителските вериги на ДНК;пакетира новосинтезираните ДНК вериги в нуклеозоми.
Репликацията започва, когато към репликационното начало на всяка от двете вериги на ДНК молекулата се свърже по една хеликаза (Фигура 8.2). Хеликазите могат да се местят само в посока 5`→3` по дължината на ДНК. Поради това се движат в срещуположни посоки, а двуверижната ДНК се разплита двупосочно. Формират се две репликационни вилки със срещуположна посока на движение. Към всяка от разделените вериги се захващат SSB белтъци. Те не позволяват на едноверижните участъци на ДНК молекулата отново да се свържат, предпазват двуверижните участъци от нуклеази и улесняват удължаването на новосинтезираните едноверижни ДНК. Локалното разплитане на ДНК предизвиква суперспирализиране на молекулата, което се отстранява от ензима топоизомераза.

64
Фигура 8.2. Формиране на репликационни вилки.
Родителските ДНК вериги служат като матрици, към които в прокариотните клетки ДНК полимераза ІІІ свързва комплeментарни бази: тимин с аденин и гуанин с цитозин. Репликацията винаги протича в посока 5`→3`: кислорода от 3`ОН групата на добавяния нуклеотид се свързва към 5`фосфатния край на последния нуклеотид. В активността на ДНК полимераза ІІІ има две ограничения: 1) не може да инициира синтеза на нова ДНК верига. Поради това синтезът започва със синтез на РНК праймер (къси РНК, комплементарни на ДНК) от ензима РНК полимераза и 2) може да добавя нуклеотиди само към 3`края, но не и към 5`края на съществуваща едноверижна ДНК. Поради това ДНК полимераза ІІІ синтезира двете дъщерни вериги едновременно, но по различен начин (Фигура 8.3). Едната верига нараства в посоката на движение на репликационната вилка. Нарича се водеща, защото синтезът й е непрекъснат и бърз процес. Другата верига се нарича изоставаща. Нейният синтез протича в посока обратна на посоката на движение на репликационната вилка. Това е бавен процес с многократни прекъсвания. Изоставащата верига се синтезира на фрагменти: ДНК полимераза ІІІ изгражда малък участък (фрагмент на Оказаки) и се връща обратно в началото на репликационната вилка, където към следващия РНК-праймер добавя нуклеотиди и изгражда нов фрагмент на Оказаки.
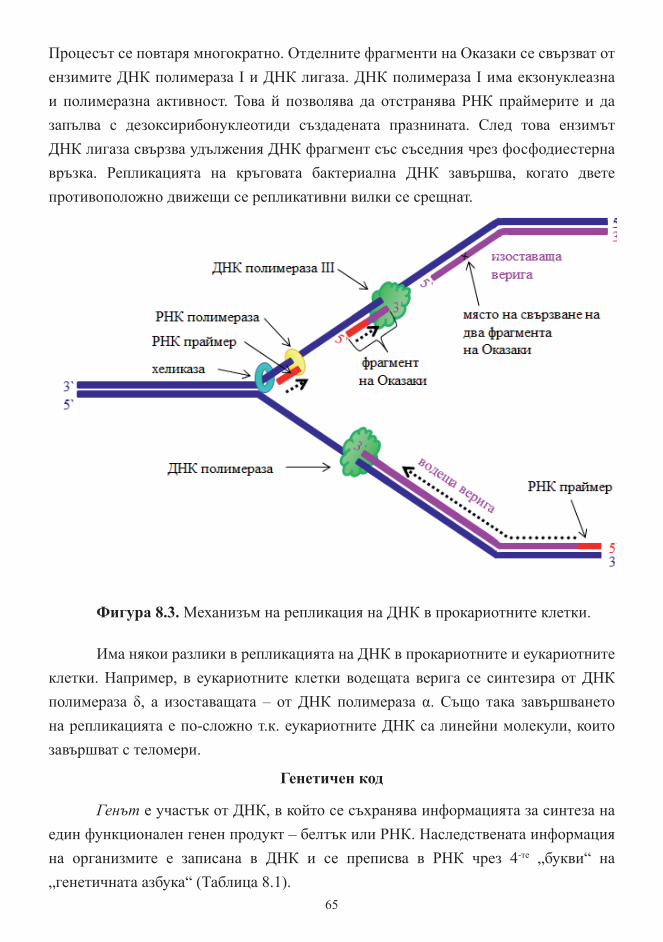
65
Процесът се повтаря многократно. Отделните фрагменти на Оказаки се свързват от ензимите ДНК полимераза І и ДНК лигаза. ДНК полимераза І има екзонуклеазна и полимеразна активност. Това й позволява да отстранява РНК праймерите и да запълва с дезоксирибонуклеотиди създадената празнината. След това ензимът ДНК лигаза свързва удължения ДНК фрагмент със съседния чрез фосфодиестерна връзка. Репликацията на кръговата бактериална ДНК завършва, когато двете противоположно движещи се репликативни вилки се срещнат.
Фигура 8.3. Механизъм на репликация на ДНК в прокариотните клетки.
Има някои разлики в репликацията на ДНК в прокариотните и еукариотните клетки. Например, в еукариотните клетки водещата верига се синтезира от ДНК полимераза δ, а изоставащата – от ДНК полимераза α. Също така завършването на репликацията е по-сложно т.к. еукариотните ДНК са линейни молекули, които завършват с теломери.
Генетичен код
Генът е участък от ДНК, в който се съхранява информацията за синтеза на един функционален генен продукт – белтък или РНК. Наследствената информация на организмите е записана в ДНК и се преписва в РНК чрез 4-те „букви“ на „генетичната азбука“ (Таблица 8.1).
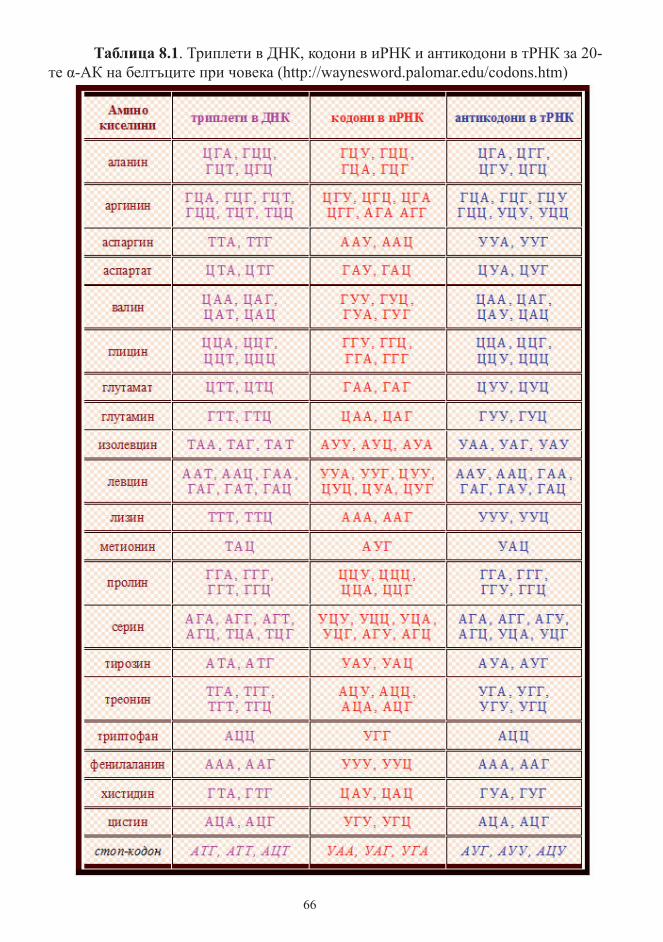
66
Таблица 8.1. Триплети в ДНК, кодони в иРНК и антикодони в тРНК за 20-те α-АК на белтъците при човека (http://waynesword.palomar.edu/codons.htm)

67
Тези 4 букви са 4-те нуклеотида, изграждащи нуклеиновите киселини. Нуклеотидите се различават само по азотните си бази – А, Г, Ц и Т в ДНК и А, Г, Ц и У в РНК. Нуклеотидната последователност в ДНК определя нуклеотидната последователност в РНК. С помощта на генетичния код иРНК определя последователността на α-аминокиселините (α-АК) в полипептидната верига (ППВ) на белтъците (Фигура 8.4). Генетичният код може да се разглежда като набор от правила, чрез които клетката превежда информацията записана в ДНК или РНК и синтезира съответната ППВ.
Фигура 8.4. иРНК чрез генетичния код определя последователността на α-аминокиселините в полипептидната верига на белтъците.
Генетичният код притежава характеристики, които позволяват генетичната информация за целия организъм да бъде записана в ДНК на всяка една клетка:
Кодът е триплетен. Всяка α-АК се кодира от кодон, съставен от три последователни нуклеотида в РНК. Всеки кодон е комплементарен на определен триплет – 3 последователни нуклеотида в ДНК. Например, различната последователност на три нуклеотида с азотните бази Ц, Ц и А могат да формират два различни кодона в иРНК за две различни АК. Кодонът, в който последователността на нуклеотидите е УГГ кодира α-АК-та триптофан, а кодонът с последователност ГГУ кодира глицин (Фигура 8.4). Тези кодони са комплементарни на триплетите АЦЦ и ЦЦА в ДНК. Често триплетите в ДНК също се наричат кодони.
Кодоните не се препокриват. Един и същи нуклеотид не влиза в състава на два съседни кодона. Например, първите 10 α-АК от ППВ на хемоглобина на човека се кодират от 10 кодона и 30 нуклеотида в РНК (Фигура 8.5-А).

68
Фигура 8.5. ДНК секвенция на кодоните за първите 10 α-АК от β-хемоглобина на човека. А – нормална секвенция, Б – секвенция с делеция, причина за сърповидно клетъчна анемия. (Clancy, S. 2008. Genetic mutation. Nature Education 1(1):187).

69
Кодът е изроден, защото някои от АК се кодират от повече от един кодон. Четирите нуклеотида на РНК могат да формират 64 различни комбинации от по три нуклеотида: 43 = 64 кодона. Повечето от тях (61 броя) кодират α-АК. Например, за аргинин има 6 кодона, а за аланин – 4 (Таблица 8.1). Наричат се изродени кодони. Повечето от тях се различават в третия нуклеотид. Почти всички организми използват този генетичен код. Има само няколко изключения – по-различен е смисъла на някои от кодоните в митохондриалния геном и в генома на някои едноклетъчни.
В генетичният код има само един препинателен знак. Последните три от 64-те кодона са стоп-кодони: УАА, УАГ и УГА (Таблица 8.1). Също така се наричат безсмислени, защото не кодират α-АК. Те имат функционално значение – служат като сигнал за спиране синтеза на ППВ. Поради това стоп-кодонът може да се разглежда като единственият препинатерен знак в генетичния код. Кодонът за метионин служи и като старт-кодон. Изграждането на иРНК започва с този кодон.
Генетичният код се чете без прекъсване. Три последователни нуклеотида от иРНК представляват т.н. „рамка на четене“ на генетичната информация. Някои мутации, като индел мутациите, могат да променят рамката на четене. Например, β0-таласемията (вид анемия) е резултат от делеция, обхващаща само една базова двойка в ДНК (Фигура 8.6). Мутацията е в триплет №44 на гена, кодиращ β-глобина на хемоглобина при хората. Делецията на втората базова двойка на този триплет води до три съществени последици:
1. мястото на отпадналия нуклеотид се заема от следващия и рамката на четене се измества;
2. превръща кодон №60 (за валин) в РНК в стоп-кодон;3. стабилността на иРНК намалява с около 40 пъти.
Фигура 8.6. Част от секвенцията на гена, кодиращ хемоглобина при хората. А – нормална секвенция, Б – секвенция с делеция в кодон №44, водеща до β0-таласемия (Kinniburgh, A.J., L.E. Maquat, T. Schedl, E. Rachmilewtz, J. Ross. 1982. mRNA-deficient β0-thalassemia results from a single nucleotide deletion. Nucleic Acids Research, 10(18):5421-5427.)

70
В ретикулоцитите на периферната кръв на индивиди, които са хомозиготни по тази делеция, не се синтезира β-глобин и те умират в ранна детска възраст.
Мутациите от типа замяна на базови двойки, не променят рамката на четене, но някои от тях оказват значително влияние върху живота и здравето на човека. Например, сърповидно клетъчната анемия при хората е резултат от замяна само на една базова двойка (А-Т заменена от Т-А) в триплет №7 на ДНК молекулата (Фигура 8.5-Б). Поради това, АК №6 (глутамат) в β-глобина в нормалния хемоглобин е заменена с валин в ППВ на мутантния β-глобин. Разликата е само една АК, но се формират твърди снопове от слепени молекули хемоглобин. Това придава сърповидна форма на червените кръвни клетки (Фигура 4.3). Сърповидните клетки се характеризират с ниска ефективност в пренасянето на кислород, често запушват капилярите и нарушават кръвоснабдяването на тъканите. Заболяването често има фатален край за индивидите, които са хомозиготни по тази мутация.
Транскрипция и транслация – същност, основни етапи
Реализирането на наследствената информация е многоетапен процес. В някои от гените е закодирана информация за синтез на функционалните РНК, които са две основни групи: 1) големи функционални РНК – тРНК и всички рРНК, 2) малки функционални РНК като snРНК, miРНК, siРНК, piРНК и др. Повечето гени са белтък-кодиращи. Изграждането на белтъци се осъществява в два етапа – транскрипция и транслация. Транскрипцията е синтез на иРНК, а транслацията е синтез на ППВ.
Транскрипцията е процес, при който информацията кодирана в ДНК се преписва (транскрибира) в РНК (транскрипт). РНК молекулата е комплементарна на матричната ДНК верига. Наследствената информация на организмите е записана в ДНК чрез 4-те дезоксирибонуклеотида и се преписва в РНК чрез 4-те рибонуклеотида (Таблица 8.2). Последователността на дезоксирибонуклеотидите в ДНК определя последователността на свързване на рибонуклетидите в изграждащата се РНК. По време на транскрипцията едната ДНК-верига служи като матрица за синтез на комплементарна РНК, която представлява РНК-копие на белтък-кодиращия ген (Фигура 8.4).

71
Таблица 8.2. Бази на ДНК и РНК и съответните им нуклеотид трифосфатибаза нуклеотид
ДНК
аденин (А) дезоксиаденозин трифосфат (дАТФ)гуанин (Г) дезоксигуанозин трифосфат (дГТФ)цитозин (Ц) дезоксицитидин трифосфат (дЦТФ)тимин (Т) дезокситимидин трифосфат (дТТФ)
РНК
аденин (А) аденозин трифосфат (АТФ)гуанин (Г) гуанозин трифосфат (ГТФ)цитозин (Ц) цитидин трифосфат (ЦТФ)урацил (У) уридин трифосфат (УТФ)
Транскрипцията се осъществява като ДНК се разплита локално и една от веригите служи като матрица за синтеза на РНК. По дължината на цялата хромозома и двете вериги на ДНК служат като матрици, но в един ген само едната верига е матрица. Винаги една и съща верига е матрица при всяка транскрипция на гена (Фигура 8.7). Рибонуклеотидите образуват връзки с комплементарните им бази в матричната ДНК верига. Рибонуклеотида А с Т в ДНК, Г с Ц, Ц с Г, и У с А. Ензимът РНК-полимераза осигурява свързването на рибонуклеотидите към правилните дезоксирибонуклеотиди. РНК-полимеразата се свързва към ДНК и движейки се по дължината й свързва рибонуклеотидите в едноверижна молекула РНК. Синтезът на РНК е от 5`-края към 3`-края т.е. нуклеотидите се добавят към растящия 3`-край.
Фигура 8.7. Два гена с противоположна посока на транскрипция (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).
Прокариотните клетки нямат обособено ядро и транслацията на РНК започва преди да е приключил синтезът й (Фигура 8.8). В еукариотните клетки белтъчният

72
синтез не започва веднага. Транскрипцията протича в ядрото, а транслацията – в цитоплазмата. Синтезираната РНК се нарича първичен транскрипт или иРНК-предшественик (пре-иРНК). пре-иРНК претърпява няколко промени, които общо се обозначават като зреене на иРНК. Транслацията започва едва след като зрялата иРНК бъде пренесена в цитоплазмата.
Фигура 8.8. Място на транскрипция и транслация в прокариотна и еукариотна клетка (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).
Транслацията започва с формирането на рибозомите, които са комплекс от рРНК и белтъци. Те осигуряват свързването на двойката АК-тРНК към иРНК и ръководят удължаването на полипептидната верига на белтъка. По време на белтъчния синтез генетичната информация, записана в иРНК, се превежда с помощта на генетичния код, тРНК и рибозомите. Последователността на кодоните в иРНК определя последователността на 20-те α-АК в ППВ на белтъците (Фигура 8.4). Всеки кодон кодира определена АК и всяка АК има собствена група от тРНК-и. За всяка α-АК има специфичен ензим, наречен аминоацил-тРНК синтетаза. Ролята на ензима е да свържи α-АК към свободния 3`-край на специфичната й тРНК. Транспортната РНК е посредник, който чрез антикодона си дешифрира информацията в иРНК. Антикодонът е триплет в средната бримка на тРНК, който образува базови двойки с комплементарния кодон от иРНК. Ако тРНК носи кодираната от иРНК амино киселина, кодонът и антикодона са комплементарни и се свързват чрез водородни връзки. Носената от тРНК α-АК се включва в изграждащата се ППВ. Ако тРНК не носи кодираната от иРНК аминокиселина, кодонът и антикодона не са комплементарни и не се свързват с водородни връзки.

73
В този случай тРНК напуска рибозомата заедно с α-АК, която носи (Фигура 8.9). Свързването на α-АК при синтеза на белтъците е чрез пептидна връзка между карбоксилната група на последната α-АК и свободната амино група на следващата α-АК.
Фигура 8.9. При транслацията водородни връзки се образуват само между кодон и антикодон, които са комплементарни.
Tема 9. Извънядрени гени
Пластидна и митохондриална наследственост
Еукариотните клетки притежават многокомпонентна генетична система – основна (ядрена) и вторична (органелна). Освен в ядрото, ДНК има в хлоропластите на растенията и в митохондриите на животните, растенията и гъбите. Характерните особености на органелната ДНК са:
1. Унаследява се независимо от ядрената ДНК;2. Осигурява специфичните функции на органелите. В митохондриалният геном
е кодирана информация свързана с осигуряването на енергия за клетката, а хлоропластния геном – с процеса фотосинтезата;
3. Органелната ДНК понякога е пакетирана в нуклеоиди (Фигура 9.1);4. Броят на копията на органелната ДНК в клетката е голям. Всяка клетка
притежава голям брой митохондрии и/или хлоропласти, всеки от който има голям брой органелни ДНК молекули. Поради това броят на органелните хромозоми в една клетка може да достигне стотици или хиляди;

74
5. Броят на копията на органелните хромозомни във всяка клетка е различен, защото броя на органелите е различен.
6. В органелният геном възникват мутации, които се предават в следващото поколение. Наричат се органелни мутации или цитоплазмени мутации.
Фигура 9.1. Митохондриална ДНК, пакетирана в нуклеоиди (Hallberg, B.M., N.-G. Larsson. 2011. TFAM forces mtDNA to make a U-turn. Nature Structural & Molecular Biology, 18:1179-1181. doi:10.1038/nsmb.2167).
Митохондриалната ДНК (мтДНК) се намира в митохондриалния матрикс на митохондриите. мтДНК на растенията е кръгова, линейна или разклонена двуверижна молекула, а на останалите еукариоти – само кръгова. мтДНК се реплицира през интерфаза. По време на цитокинеза дъщерните клетки получават приблизително еднакъв брой митохондрии, без механизъм за точно разпределяне. При оплождането на бозайниците и повечето многоклетъчни организми зиготата получава минимални количества цитоплазма от гаметата на бащата. Поради това почти всички митохондрии на ембриона са от гаметата на майката. При мишките само 0.01% от мтДНК е от бащата.
Човешката мтДНК е напълно секвенирана. Изградена е от 16 569 б.дв. Съдържа 37 гена, които кодират 13 белтъка, 2 рРНК и 22 тРНК. Всички белтъци кодирани в мтДНК се синтезират чрез митохондриални рибозоми, а рРНК и тРНК участват в транслацията на митохондриалната иРНК.
Митохондриалният генетичния код на растенията не се отличава от стандартния ядрен, но се характеризира със специфичност (Таблица 9.1). Тя се дължи на РНК редактиране на митохондриалната пре-иРНК. Например, кодонът ЦГГ кодира

75
аргинин. В резултат на РНК редактиране цитозинът се превъръща в урацил, а кодона ЦГГ – в УГГ за аминокиселините треонин и триптофан. Митохондриалният генетичен код се характеризира със специфичност и при останалине еукариоти (Таблица 9.1).
Таблица 9.1. Митохондриалният генетичен код е различен при различните еукариоти.
кодонядрен митохондриален
бозайници гъби Drosophila растения
УАГ стоп кодон триптофан триптофан триптофан стоп кодон
АГА, АГГ аргинин стоп кодон аргинин серин аргинин
Всички белтъци кодирани в мтДНК са субединици в комплекса, функциониращ в електронния транспорт или в синтеза на АТФ. Заболяванията Наследствена оптична невропатия на Лебер (LHON) и Синдрром на Киърнс-Сейър са резултат от цитоплазмени мутации при човека. LHON е дегенерация на очния нерв, която води до слепота. Резултат е от мутация в базовата двойка 11 778 на мтДНК, което води до замяна на α-АК хистидин с аргинин в ППВ на НАДН дехидрогеназата. Митохондриите с тази мутация имат ниска ефективност в процесите на окислително фосфорилиране и синтез на АТФ. Унаследяването на болестта е само по майчина линия, защото митохондриите се предават основно чрез цитоплазмата на яйцеклетката (Фигура 9.2).
Фигура 9.2. Родословна схема на унаследяването на LHON. Индивидите с LHON са представени със запълнен символ (Wallace, D.C., M.D. Brown, M.T. Lott. 1999. Mitochondrial DNA variation in human evolution and disease. Gene, 238:211-230.).

76
Хлоропластната ДНК (хлДНК) е кръгова двуверижна молекула. В зависимост от вида на клетката броя на хлоропластните хромозоми варира. В клетките на семената са няколко, а в клетките на младите бързо растящи листа достигат до над 10 хил. Големият им брой е резултат от многократна репликация на хлДНК, която не е последвана от делене на хлоропласта. С излизане на растенията от фазата на бърз растеж, количеството на ДНК драстично намалява. Репликация на хлДНК е процес, който протича независимо от клетъчния цикъл и от репликацията на ядрената ДНК.
хлДНК е изградена от 120 хиляди до 160 хиляди базови двойки и включва 120 гена. От тях 60 са свързани с транскрипцията и транслацията на хлРНК, включително за рРНК, тРНК, субединиците на РНК полимеразата, и рибозомни белтъци. Около 20 гена кодират субединиците на комплекса за фотосинтетичния транспорт на електрони и АТФ-азния комплекс. В генома на хлоропластите е кодирана и голямата субединица на ензима рибулозо 1,5-дифосфат карбоксилаза, участващ в цикъла на Калвин.
Въпреки, че цялостната организация на хлДНК на различните видове е подобна, има някои различия в състава на гените. Например, хлДНК на растението гълъбови очички има някои гени, които липсват в хлДНК на тютюна и обратно. Хлоропластите на двете растения имат едни и същи белтъци като някои от тези гени са в хлДНК на единия растителен вид, а на другия вид са в ядрената ДНК. Генетичният код на хлоропластите не се отличава от стандартния ядрен код. Хлоропластните рибозоми синтезират хлоропласт-кодираните белтъци. Останалите хлоропластни белтъци се синтезират в цитоплазмени рибозоми и след това се внасят в хлоропластите.
Мутационният процес може да засегне и хлДНК. Например, мутантен алел в хлДНК не позволява синтеза на хлорофил. Засегнатите клетки са бели, защото хлоропластите са бели. Мутациите могат да са в ДНК само на отделни хлоропласти. Такива клетки се наричат хетероплазмони или цитохети. В повечето случаи органелите се разпределят в дъщерните клетки неравномерно и на случаен принцип. Поради това в отделните клетки броят на засегнатите органели е различен. Понякога се наблюдават изключения. Например, при клетъчното делене на хетероплазмон с мутантни (бели) хлоропласти в едната дъщерна клетка се събират бели, а в другата – зелени хлоропласти (Фигура 9.3). Двата вида хлоропласти се разпределят в двете дъщерни клетки. Процесът се нарича цитоплазмено разделяне. Пъстрите петна по листата са мозайка от зелени и бели клетки. Белите листни петна се дължат на по-нататъшното делене на белите клетки. Колкото по-рано в индивидуалното развитие протече цитоплазменото разделяне, толкова по-големи ще бъдат зелените и белите участъци по растението.

77
Цитоплазмени мутации при човека. Цитоплазмена мъжка стерилност
Цитоплазмени мутации при човека. Някои рядко срещани заболявания при хората се предават само по майчина линия, защото са резултат от мутации в мтДНК. Например, заболяването MERRF (MERRF – myoclonic epilepsy and ragged red fiber: миоклонична епилепсия и дрипави червени влакна) е резултат от точкова мутация в мтДНК. Заболяването засяга мускулите, но симптомите включват отклонения в дейността на очите и слуха. Други подобни примери са Наследствената оптична невропатия на Лебер (LHON) (Фигура 9.2) и Синдромът на Киърнс-Сейър. Във всички тези случаи клетките съдържат смес от нормални и мутантни мтДНК. В резултат на цитоплазменото разделяне съотношението им в клетките е различно: във времето, в тъканите на един и същ индивид и в клетките на отделните потомци. Натрупването на митохондриални мутации във времето е една от възможните причини за нарушаване функциите на клетките и стареенето на организмите.
Цитоплазмената мъжка стерилност (ЦМС) се наблюдава при растенията. Представлява формиране на нефункционален полен, поради изменения в цитоплазмата (Фигура 9.4.). Цитоплазмата е два типа – нормална (N) и стерилна (S). ЦМС е белег, унаследяващ се по майчина линия. Резултат е от мутации в мтДНК или на други цитоплазмени фактори. Често се свързва с необичайна рамка на четене в мт геном. В много случаи мъжката фертилност може да бъде възстановена от доминантни ядрени гени: Rf – fertility restorer т.е. възстановител на фертилността. ЦМС се използва за производството на хибриди при царевица, ориз, пшеница и др.
Фигура 9.3. Цитоплазмено разделяне на хлоропластите на хетероплазмон (по Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York).

78
Тема 10. Количествена генетика
Унаследяване и генетична архитектура на количествени белези. Видове количествени белези. Влияние на околната среда
Количествената генетика е направление в генетиката, което изследва унаследяването на количествените признаци. Качествени са признаци, които не могат да бъдат измерени и факторите на околната среда имат незначително влияние върху формирането им. Например, цвят на граховите зърна. Количествени са признаци, които могат да бъдат измерени и се представят с числови стойности. Например, млечност, масленост, височина на тялото и др. Характеризират се с изменчивост, която варира в генетично определени граници. Формирането na количествените признаци е резултат от взаимодействието на няколко гена, наречени локуси определящи количествени признаци (QTL – Quantitative Trait Loci) или полигени. В повечето случаи полигените се намират в различни хромозоми. Гените за всеки количествен признак се унаследяват съгласно законите на Мендел и имат различна степен на влияние върху фенотипа на индивида.
От генетична гледна точка, терминът „среда“ включва всички влияния, които са различни от генотипа. Те може да са външни като семейство, приятели, колеги и др. или биологични като хранене, хормонален баланс, токсини и др. Според многофункционалната хипотеза количествените признаци се формират в резултат от комплексното влияние на факторите на околната среда и на голям брой гени. Поради това по-точният термин е комплексни признаци. Те са от първостепенно значение за медицинската и селскостопанската генетика. Смята се, че унаследяването на атеросклерозата (удебеляване на артериалната стена в резултат от натрупване на мастни материали като холестерол) и хипертонията (високо кръвно налягане), телесното тегло, податливостта на депресия, развитието на рак са признаци с полигенен характер. Комплексни признаци, които са значими
Фигура 9.4. Цитоплазмена мъжка стерилност при ориз. Нефункционалните поленови зърна са безцветни (Eckardt. N.A. 2014. Cytoplasmic Male Sterility and Fertility Restoration. Plant Cell 2006;18;515-517. DOI 10.1105/tpc.106.041830).

79
за селското стопанство са добивът, устойчивостта на патогени или на засушаване, ефективността в усвояването на торовете, и дори вкусът на житните растения.
Основната цел на количествената генетика е да се установи генетичната архитектура на комплексните признаци. Генетичната архитектура се описва с всички гени и фактори на околната среда, които влияят върху формирането на даден комплексен признак. Тя е свойство, което може да е различно за различните популации на един вид. Например, в някои популации високото кръвно налягане се дължи основно на начина на живот (факторите на околната среда), а в други е генетично обусловено. Унаследяването на комплексните белези се изучава с използването на някои основни статистически характеристики като средна стойност и дисперсия. Те позволяват да се установи до колко наблюдаваните вариации се дължат на генотипът и до колко на факторите на околната среда.
Влиянието на наследствеността се измерва чрез анализ на различията в количествените белези на генетично идентични индивиди, които са поставени при различни условия на средата. Генетично идентични индивиди са чистите линии растения и животни и еднояйчните близнаци при хората.
Взаимодействието между генотип и околна среда е феномен, при който в индивиди с един и същ генотип се формират различни фенотипове при различни условия на околната среда. Взаимодействието се бележи с G×E (Genotype × Еnvironment: генотип × околна среда). Границите на вариране на признака при различни условия на средата са генотипно детерминирани и се наричат норма на реакция. Поради това фенотипната изява на определен генотип може да е различна при различни условия. Например, взаимовръзката G×E по отношение на затлъстяването при човека е силно изразена (Фигура 10.1). Фенотипът (степента на затлъстяване) зависи както от генотипа, така и от факторите на средата (градски или селски район).
Често отделните алели на QTL имат сравнително малък количествен принос за фенотипа. Взаимодействието се бележи с Q×Е (QTL × Еnvironment). Силата на това взаимодействие за определен локус се измерва чрез отклонението в средните стойности на QTL фенотипа при специфични условия на околната среда в сравнение със средната стойност на признака.
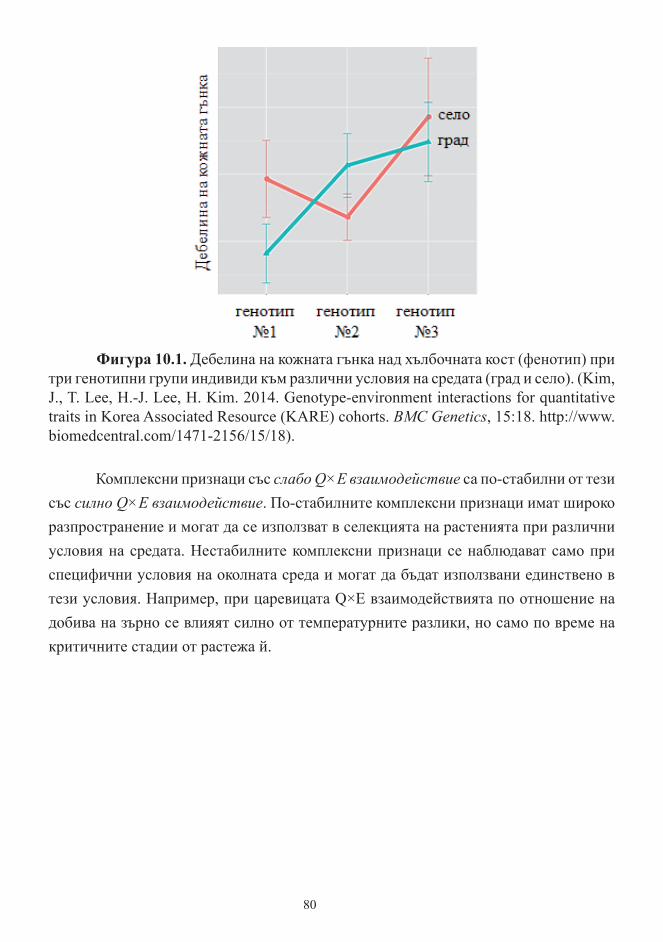
80
Фигура 10.1. Дебелина на кожната гънка над хълбочната кост (фенотип) при три генотипни групи индивиди към различни условия на средата (град и село). (Kim, J., T. Lee, H.-J. Lee, H. Kim. 2014. Genotype-environment interactions for quantitative traits in Korea Associated Resource (KARE) cohorts. BMC Genetics, 15:18. http://www.biomedcentral.com/1471-2156/15/18).
Комплексни признаци със слабо Q×Е взаимодействие са по-стабилни от тези със силно Q×Е взаимодействие. По-стабилните комплексни признаци имат широко разпространение и могат да се използват в селекцията на растенията при различни условия на средата. Нестабилните комплексни признаци се наблюдават само при специфични условия на околната среда и могат да бъдат използвани единствено в тези условия. Например, при царевицата Q×Е взаимодействията по отношение на добива на зърно се влияят силно от температурните разлики, но само по време на критичните стадии от растежа й.

81
ЛИТЕРАТУРА
1. Драгоева, А., О. Димитров. 2005. Структура и функции на клетките. Изд. „Светлана“, Шумен
2. Драганов, М., Н. Попов. 2010. Клетъчна биология. Ун. изд. „Паисий Хилендарски“, Пловдив
3. Генова, Г., Н. Харизанова, М. Пешева, Т. Декова, О. Конова-Кръстанова, П. Иванова, М. Китанова. 2012. Генетика, Ръководство за упражнения, Изд. Гера Арт.
4. Попов П., Т. Ирикова. 2004. Обща генетика, Ун. изд. „Паисий Хилендарски“, Пловдив.
5. Chial, H., Craig, J. 2008. mtDNA and mitochondrial diseases. Nature Education, 1(1):217 http://www.nature.com/scitable/topicpage/mtdna-and-mitochondrial-diseases-903
6. Clancy, S. 2008. Genetic mutation. Nature Education 1(1):187 http://www.nature.com/scitable/topicpage/genetic-mutation-441
7. Drouaud, J., R. Mercier, L. Chelysheva, A. Bérard, M. Falque, O. Martin, V. Zanni, D. Brunel, C. Mézard. 2007. Sex-Specific Crossover Distributions and Variations in Interference Level along Arabidopsis thaliana Chromosome 4. PLoS Genet, 3(6):e106. doi:10.1371/journal.pgen.0030106
8. Eckardt., N.A. 2006. Cytoplasmic Male Sterility and Fertility Restoration. Plant Cell, 18(3):515-517. DOI 10.1105/tpc.106.041830
9. Zhao, F., S. Xu. 2012. Genotype by Environment Interaction of Quantitative Traits: A Case Study in Barley. G3 (Genes, Genomes, Genetics), 2(7):779-788.
10. Guc-Scekic, M, J. Milasin, M. Stevanovic, L.J. Stojanov, М. Djordjevic. 2002. Tetraploidy in a 26-month-old girl (cytogenetic and molecular studies). CLINICAL GENETICS, 61:62-65.
11. Griffiths, A., J.H. Miller, D.T. Suzuki, R.S. Lewontin, W.M. Gelbart. 2004. An introduction to genetic analysis. 8 ed, W. H. Freeman and Company, New York.
12. Griffiths, A., S.R. Wessler, S.B. Carroll, J. Doebley. 2012. Introduction to genetic analysis. 10 ed, W. H. Freeman and Company, New York.
13. Hallberg, B.M., N.-G. Larsson. 2011. TFAM forces mtDNA to make a U-turn. Nature Structural & Molecular Biology, 18:1179-1181. doi:10.1038/nsmb.2167
14. Kalcheva V., A. Dragoeva, K. Kalchev, D. Enchev. 2009. Cytotoxic and genotoxic effects of Br-containing oxaphosphole on Allium cepa L. root tip cells and mouse bone marrow cells. Genetics and Molecular Biology, 32(2):389-393.

82
15. Karthikeyan, P.P., P.T.V. Lakshmi, C.K. Dwibedi, A. Annamalai. 2009. A database for human Y chromosome protein data. Bioinformation, 4(5):184-186.
16. Kim, J., T. Lee, H.-J. Lee, H. Kim. 2014. Genotype-environment interactions for quantitative traits in Korea Associated Resource (KARE) cohorts. BMC Genetics, 15:18. http://www.biomedcentral.com/1471-2156/15/18
17. Kinniburgh, A.J., L.E. Maquat, T. Schedl, E. Rachmilewtz, J. Ross. 1982. mRNA-deficient β0-thalassemia results from a single nucleotide deletion. Nucleic Acids Research, 10(18):5421-5427.
18. Koleva, V., A. Dragoeva, A. Andreeva, M. Burova, S. Georgiev, D. Enchev. 2013. Comparative analysis of clastogen-induced chromosome aberrations observed with light microscopy and by means of atomic force microscopy. Mutation Research/Genetic Toxicology and Environmental Mutagenesis, 753(1):29-35.
19. Lodish, H., A Berk, S. Zipursky, P Matsudaira, D. Baltimore, J. Darnell. 2004. Molecular cell biology, 5th ed, Freeman and Co.
20. Madigan, M.T., J.M. Martinko, D. Stahl, D.P. Clark. 2010. Brock Biology of Microorganisms, 13th Edition, Pearson Inc.
21. O‘Donnell, M., L. Langston, B. Stillman. 2013. Principles and Concepts of DNA Replication in Bacteria, Archaea, and Eukarya. Cold Spring Harb Perspect Biol, doi: 10.1101/cshperspect.a010108
22. Tsai, J.-H., B.D. McKee. 2011. Homologous pairing and the role of pairing centers in meiosis. Journal of Cell Science, 124:1955-1963. doi:10.1242/jcs.006387
23. Wallace, D.C., M.D. Brown, M.T. Lott. 1999. Mitochondrial DNA variation in human evolution and disease. Gene, 238:211-230.
24. Veluthambi, K., M. Krishnan, Jean H. Gould, Roberta H. Smith, Stanton B. Gelvin. 1989. Opines Stimulate Induction of the vir Genes of the Agrobacterium tumefaciens Ti Plasmid. Journal of Bacteriology, 171 (7):3696-3703.
25. Yildiz, M. 2013. Current Progress in Biological Research, Publisher: InTech.
26. http://waynesword.palomar.edu/codons.htm
27. http://www.cals.ncsu.edu/course/pp728/Agrobacterium/Alyssa_Collins_profile.htm
28. http://nar.oxfordjournals.org/content/30/1/121/F1.expansion

83

84
© Ваня Колева© Обща генетика. Лекционен курс
Българска
І издание
Снимков материал: Ваня Колева
Художествено оформление и предпечатна подготовка: Илиян Петков
печат: “Химера” ООД - ШуменИздателство: “Химера” ООД,тел.: 054/ 830 585www.himeraltd.com
Формат 1/16 от 70/100. Обем 5.25 п. к.Дадена за печат септември 2014 г.
Излязла от печат октомври 2014 г.





























