Embed Size (px)
Citation preview

UNIVERSIDAD DE COSTA RICA
FACULTAD DE CIENCIAS AGROALIMENTARIAS
Escuela de Agronomía
Segundo semestre 2011
Curso Reguladores de Crecimiento Vegetal
(AF5408)
“Triacontanol (TRIA)”
Estudiante
Carolina Ramírez Víquez A64673

Triacontanol (TRIA)
TRIA es un componentes de las ceras epicuticulares de alfalfa y muchos otras plantas (Chibnall et al, 1983). La estimulación del crecimiento en las plantas por parte de las propiedades del TRIA se demostró por primera vez por Crosby y Vlitos en 1959, y más tarde Ries et al en 1977 quienes lo describen como una sustancia activa de crecimiento que en concentraciones nanomolares aumenta el crecimiento y rendimiento de cultivos.
Entre los estudios realizados por Ries et al en1977, se dilucida el (TRIA) como un alcohol primario saturado (n-C30-H61-OH), además manifiesta que TRIA es muy lipofílica y difíciles de formular. La mejor método de formulación es una dispersión coloidal con el sebo surfactante alquilo sulfato como un dispersante (Ries, 1985). Este formulación es efectiva a concentraciones de aproximadamente dos órdenes de magnitud más bajo que otros que utilizan solventes y surfactantes para formar emulsiones.
TRIA se utiliza para aumentar las cosechas en millones de hectáreas, particularmente en Asia. Muchos investigadores han demostrado que afecta al varios procesos metabólicos básicos, como la fotosíntesis, los nutrientes la captación y la actividad enzimática. Sin embargo, el sitio inicial de la acción no ha sido aclarado. Se han hecho esfuerzos para dilucidar el mecanismo de acción de TRIA, que lleva a pensar que tiene un efecto en cascada (Ries y houtz, 1983). Sin embargo, poco se sabe sobre el mecanismo molecular de esas respuestas. El aislamiento y la caracterización de genes regulados por TRIA es un paso hacia la comprensión de la acción TRIA ya que puede dar pistas sobre las vías bioquímicas y procesos fisiológicos que el TRIA regula, y de tal forma revelar la componentes implicados en la señalización TRIA.
A partir de las investigaciones realizadas se ha podido describir que la utilización de TRIA provoca rápidamente un segundo mensajero identificado como 9- β-L (+)-adenosina (TRIM) del arroz (Oryza sativa L.), que en concentraciones nanomolares hace que las plantas responden de manera similar a TRIA (Ries, 1991).
El uso de estos compuestos naturales TRIA y TRIM, para mejorar el rendimiento de los cultivos o la calidad debe tener poco o ningún efecto sobre el medio ambiente, ya que sólo 0,1 a 10 mg es aplicada por hectárea (Ries, 1991).

Caracterización del TRIA
Características físicas y químicas: Cristal blanco a escala. Es estable en condiciones normales y puede ser almacenado durante mucho tiempo. Es un regulador de crecimiento en la planta de amplio espectro. Es un producto biológico natural que se purifica a partir del extracto de cera de abejas. Es inofensivo para el ser humano, los animales y amigable con el medio ambiente. (Casa comercial: Zhengzhou Sun Rain Biological Products Co., Ltd).
Presentaciones:
1 2 3
.
Figura 1. Presentaciones en las que se encuentra el Triacontanol (TRIA) según la casa comercial Zhengzhou Sun Rain Biological Products Co., Ltd

Figura 2. Caracterización del Triacontanol (TRIA) según Global Chemical Exchange, China Chemical Network and ChemNet.com are services of Zhejiang NetSun Co., Ltd.
Según las indicaciones de la casa comercial del TRIA se deben tener en cuenta las siguientes precauciones:
Diluir sólo cuando se usa. Temperatura de aplicación debe estar por encima de 20 C. Necesidad de hacer un adicional de pulverización del producto en caso de
lluvia en seis horas después de la aplicación.
Para su dilución es importante tomar en cuenta que es casi insoluble en agua (10 mg/L a 20 °C), ligeramente soluble en alcohol frío y en benceno. Soluble en sulfúrico de etilo, cloroformo, hexano, cloruro de metileno y benceno caliente.
Aplicación: Usualmente se aplica como un spray foliar.

Usos del TRIA
La mejora de la actividad de la enzima. Aumentar el contenido de la clorofila. La promoción de la fotosíntesis. Promover la acumulación de materia seca. La promoción en la absorción de los elementos minerales de las plantas. Aumentar el contenido de proteína y azúcar. Promover el agua absorbida de las plantas. La disminución de agua evaporada. Promover la germinación de la semilla. Promover la formación y crecimiento de raíces, tallos y hojas. Promover la división de las yemas y flores. La promoción de principios de madurez. La protección de flores y frutos. Mejorar la capacidad de las plantas para resistir al frio y la sequía, a fin de
aumentar la calidad de las cosechas y frutos.
Respuestas de crecimiento
TRIA se ha demostrado que aumenta el crecimiento y/o rendimiento de vegetales anuales más importantes y de las especies agronómicas de los cultivos así como el té (Camellia sinensis L.) y varias especies forestales (Ries, 1985; Kawashima et al. 1987; Shanghai Institute of Plant Physiology, 1987).
El incremento en el rendimiento cuando el crecimiento vegetativo es el producto cosechado se debe a que el rápido aumento de la red la asimilación de la tasa después de tratamientos con TRIA. Por lo tanto, incluso aunque la tasa relativa de crecimiento durante un período prolongado se mantiene constante, las plantas tratadas se hacen más grandes, por el aumento inicial de tamaño de la planta. Los aumentos en el rendimiento de cereales y frutas debido al tratamiento de TRIA se deben a un aumento en el número de frutos por planta, y/o un aumento en el tamaño de la cosecha de productos (Ries, 1985).
TRIA aumenta muchos procesos bioquímicos y fisiológicos. El problema, como con muchas hormonas de las plantas y el crecimiento sustancias, es la delimitación de la primera acción en las plantas, que al parecer desencadena una cascada de eventos metabólicos que resulta en el aumento de materia seca o rendimiento de los cultivos (Ries, 1985).
TRIA ha sido utilizado para entender las respuestas a muchas de las plantas. Algunas de las respuestas obtenidas son difíciles de explicar con nuestro conocimiento actual de los procesos fisiológicos, por ejemplo, TRIA aumenta

rápidamente el peso seco de las plantas tanto en la luz como en la oscuridad (Ries, 1985). A la luz, TRIA tiene un aumento significativo del peso seco de los brotes de maíz 8% menos de 1 min y más del 20% en 1 h. Respuestas similares se medida con las plántulas de arroz (Ries y Wert, 1988).
¿Qué factores son responsables del aumento de peso en seco?
Una hipótesis que podría explicar en parte el aumento en el peso seco es que el agua es incorporada a los productos de almidón y de hidrólisis de proteínas en respuesta a la TRIA. Además, TRIA produjo un aumento simultáneo de la proteína soluble, lo que reduce azúcares y aminoácidos libres (Ries, 1985; Shanghai Institute of Plant Physiology, 1987). Estos cambios se debieron a TRIA causa un aumento en el peso seco total (Ries, 1985).
Modo de acción
El descubrimiento de que TRIA provocó la formación o liberación de 9- β-L (+)-adenosina (TRIM) en los tejidos de las raíces de las plántulas de arroz dentro de un min de aplicación a los brotes pueden haber aclarado la primera paso en el modo de acción TRIA (Ries et al. 1990). Se postula que TRIA aumenta rápidamente la proporción de L (+) - a D (-)-adenosina, probablemente en el tonoplasto. Esta idea es apoyada por la investigación que muestran que (10 -8 y 10-
9M) TRIA aumento la actividad de la ATPasa , en las vesículas de la cebada enriquecidas de membrana plasmática de (Hordeum vulgare L.) en presencia de la calmodulina (Lesniak et al. 1989). Queda el problema de cómo TRIA provoca L (+) - adenosina y cuál es la fuente de L (+)-adenosina en las plantas. Debido a la rapidez de la respuesta,la síntesis de novo es poco probable, sobre la base de los procesos metabólicos conocidos. La mayoría de la fuente probable de la adenosina es AMP derivada a partir de ADP y ATP (Olsson et al. 1990). Se necesita más información acerca de los protocolos para garantizar unos resultados coherentes con los TRIA y TRIM en los estudios de cámara de crecimiento, invernadero y campo. Sin embargo, la evidencia es abrumadora de que TRIA aplicado en concentraciones nanomolares pueden aumentar el rendimiento o la mejora de la calidad de un grupo diverso de especies de cultivo (Ries, 1985).
Está claro que también es un desafío importante para descubrir cómo tanto TRIA y TRIM aumentan rápidamente metabolismo de las plantas y cómo TRIA provoca la salida de TRIM. La identificación de cómo se inhibe la actividad de TRIA puede conducir al descubrimiento del sitio de acción TRIA y/o TRIM (Ries, 1985).
Sin embargo, los efectos de la TRIA ha sido a menudo inconsistente en algunas plantas (Hoagland, 1980; Eriksen et al, 1982). Las razones de esta falta de coherencia se han atribuido a los rendimientos de los cultivos no siempre son las

mejoras mediante el uso de las proporciones recomendadas de los nutrientes principales, sino por la influencia ambiental y el adecuado manejo del cultivo.
Usos del TRIA en experimentos
Su efecto sobre los cultivos de tejidos se probado en Oryza sativa (Yun y Kim, 1986), las plantas leñosas (Tantos et al, 2001). Knight y Mitchell en 1987 informaron el estimular la productividad de lechuga hidropónica en un ambiente controlado con TRIA. Ma et al, en 1990 encontraron que TRIA tuvo un efecto estimulante significativo en el en in vitro del crecimiento de la planta Malus domestica cv Fugi. Moorthy y Kathiresan en 1993 estudiaron las respuestas fisiológicas en plántulas de los manglares con TRIA. Por otra parte, TRIA aumenta la producción de metabolitos secundarios en la Artemisia annua (Yaseen y Tajuddin, 1998). TRIA ha demostrado que el aumento de peso de semillas y contenido de clorofila de las hojas, y para mejorar la fotosíntesis, ramificación, longitud del tallo y las raíces (Debata y Murthy, 1981).
Reddy et al en el 2002 informaron que TRIA podría ser utilizado como un efectivo crecimiento promotor en la micropropagación de Capsicum frutescens y Decalepis hamiltonii. Debido a que el TRIA aumenta la velocidad de varios procesos bioquímicos y fisiológicos (Ries y Houtz 1983, Ries, 1991; Naeem et al, 2009) ha sido utilizado en la micropropagación (Reddy et al. 2002), la embriogénesis somática (Giridhar et al. 2004) y la producción de la metabolitos secundarios (Giridhar et al, 2005).
Se demuestra que TRIA promueve vegetativo el crecimiento en el algodón (Gossypium hirsutum L.) y aumenta el nivel de monogalactodiacylglycerol (MGDG) (Shripathi y Swamy, 1994), un galacto-lípido que parece estar implicadas en el envasado de las proteínas del fotosistema-I (Dominy y Williams, 1987).

Demostraciones en experimentos con TRIA
Efecto del TRIA en el crecimiento de las plántulas de arroz de 15 días de edad, se vio estimulada por la aplicación foliar de TRIA en una concentración muy baja (10mg/l), donde se observó un mayor nivel de peso seco en las plantas tratadas con TRIA en comparación con el tratamiento control 4h después. Los Contenidos de proteínas y clorofila incrementaron significativamente en un 16% y 25% respectivamente en las plantas tratadas. (Chen et al. 2002).
Tabla 1. Efectos del TRIA en el crecimiento de plántulas con 15 días de edad.

Así mismo en el estudio realizado por Chen et al. 2002, uno de los clones que fue identificado por el regulamiento en genes, es el que induce a las proteínas en la producción y señalamiento del ABA (OsAsrl) el cual es reportado como el que regula el ABA exógeno y el estrés salino (Vaidyanathan et al 1999).
Tabla 2. Identificación de genes regulados en arroz (Oryza sativa) por TRIA.

De igual forma Chen et al 2002 observaron que el peso seco, proteínas y contenido de clorofila en las plantas de arroz se incrementaron con la aplicación foliar. Tasa de fotosíntesis neta de la hoja (Pn) se incrementó muy rápidamente y persistentemente a una densidad de flujo de fotones dado (PFD).
Figura 3. Tasa de fotosíntesis neta de la hoja (Pn) (µmol m-2s-1) y curva de respuesta a la luz (Pn) (µmol m-2s-1) en plántulas de arroz tratadas con TRIA y plántulas control. (A) Tasa de fotosíntesis neta de la hoja (Pn) a través del tiempo en horas (h), (B) Tasa de fotosíntesis neta de la hoja (Pn) según una densidad de flujo de fotones dado (PFD).

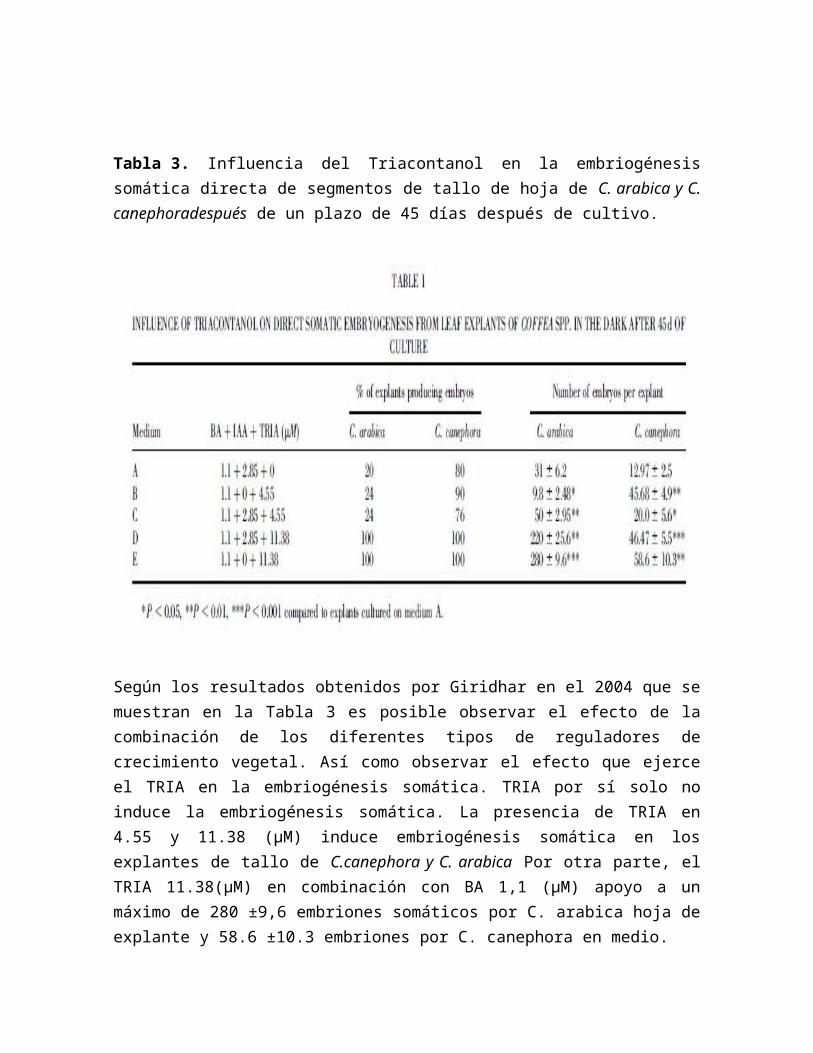
Tabla 3. Influencia del Triacontanol en la embriogénesis somática directa de segmentos de tallo de hoja de C. arabica y C. canephoradespués de un plazo de 45 días después de cultivo.
Según los resultados obtenidos por Giridhar en el 2004 que se muestran en la Tabla 3 es posible observar el efecto de la combinación de los diferentes tipos de reguladores de crecimiento vegetal. Así como observar el efecto que ejerce el TRIA en la embriogénesis somática. TRIA por sí solo no induce la embriogénesis somática. La presencia de TRIA en 4.55 y 11.38 (µM) induce embriogénesis somática en los explantes de tallo de C.canephora y C. arabica Por otra parte, el TRIA 11.38(µM) en combinación con BA 1,1 (µM) apoyo a un máximo de 280 ±9,6 embriones somáticos por C. arabica hoja de explante y 58.6 ±10.3 embriones por C. canephora en medio.
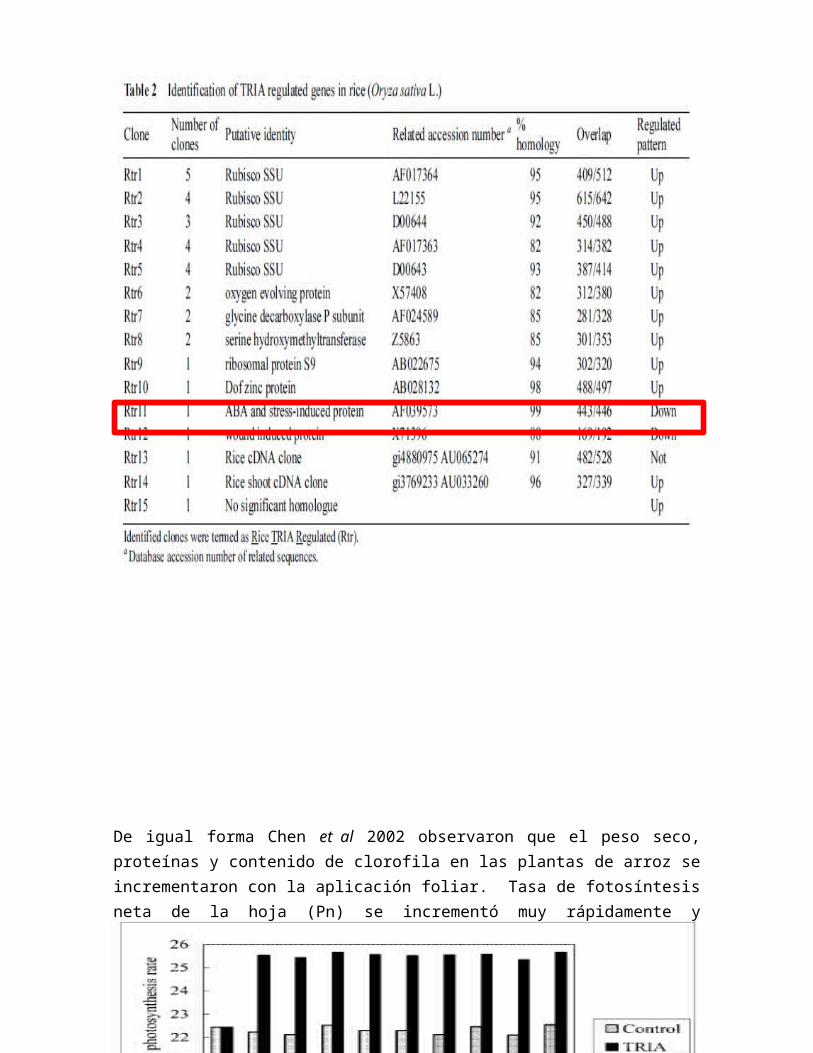
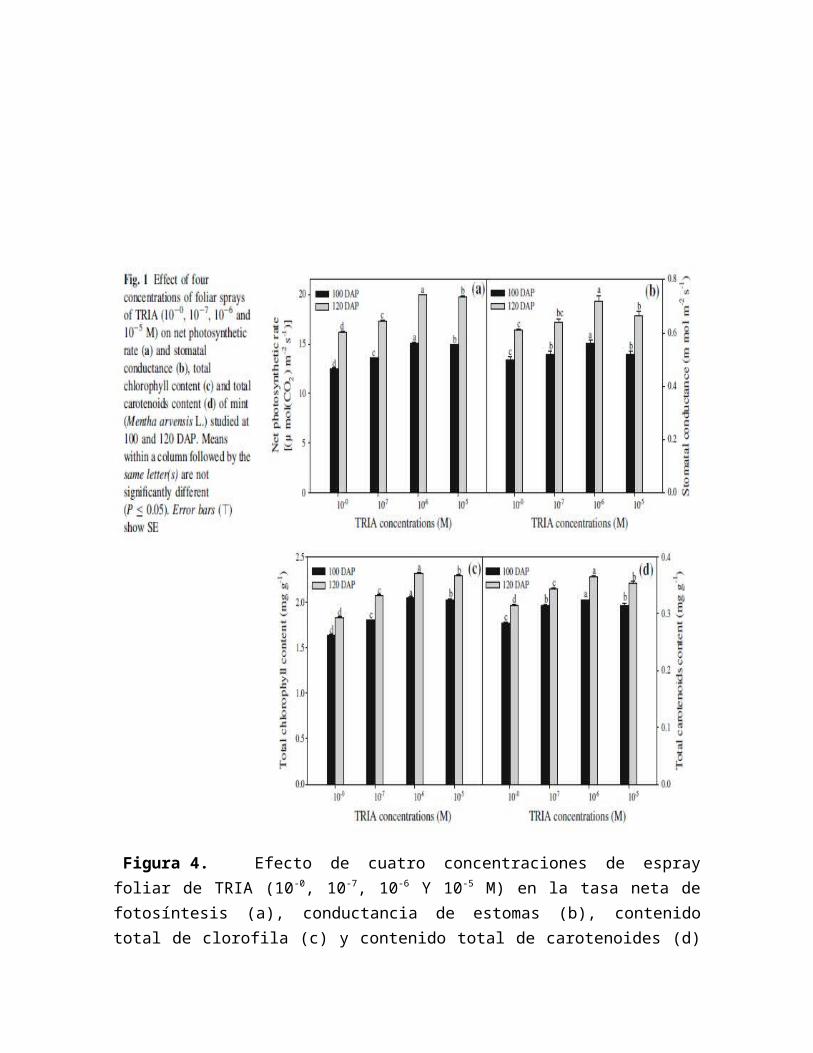
Figura 4. Efecto de cuatro concentraciones de espray foliar de TRIA (10 -0, 10-7, 10-6 Y 10-5 M) en la tasa neta de fotosíntesis (a), conductancia de estomas (b), contenido total de clorofila (c) y contenido total de carotenoides (d) en Menta (Mentha arvensis L.) estudiadas a los 100 y 120 días después de plantadas (DAP), (Naeem et al, 2011).
Aquí se ve demostrado que las aplicaciones de TRIA ejercen un efecto positivo en algunos de los parámetros de crecimiento mas importantes, donde el contenido total de clorofila aumenta en una concentración de 10-7 en 100 DAP, igualando el contenido de clorofila que obtiene el tratamiento control de 120 DAP.

Figura 5. Figura 5. Enraizamiento in vitro de tejidos derivados de brotes del cultivo Capsicum frutescens. (A, B) medio MS. (D) medio MS + 2 μg l−1 de triacontanol. (E) medio MS + 5 μg l−1 triacontanol. (C) medio MS + 10 μg l−1
triacontanol (Ready et al, 2002).
En la figura anterior es notable la contribución del TRIA en el adecuado enraizamiento de plantas de Capsicum frutescens, cultivadas de manera in vitro.
Bibliografía
CHEN, X; YUAN, H; CHEN, R; ZHU, L; DU, B; WENG, Q; HE, G. 2002. Isolation and characterization of triacontanol-rugulated genes in rice (Oryza sativa L): Possible role of triacontanol as a plant growth stimulator. Platn Cell Physiol. 43(8): 869-876.
CHIBNALL, A; WILLIAMS, E; LATNER, A; PIPER, S. 1933. The isolation of n-triacontanol from lucerne wax. Biochem J. 27:1885–1888.
CROSBY, D; VLITOS, A. 1959. Growth substances from Maryland Mammoth tobacco: long chain alcohols. Contrib Boyce Thompson Inst. 20:283.
DEBATA, A; MURTHY, K. 1981. Effect of growth regulators on photosynthetic efficiency, translocation and senescence in rice. Ind. J. Exp. Biol. 19:986–987; 1981.
DOMINY, P; WILLIAMS, W. 1987. Is monogalactosyldiacylglycerol involved in the packaging of light harvesting chlorophyll proteins in the thylakoid membrane. E: Stumpf, P.K., Mudd, J.B., Nes, W.D. (Eds.), The Metabolism, Structure and Function of Plant Lipids. Plenum Press, New York. pp. 185-187.
ERIKSEN, A; HAUGSTAD, M; NILSEN, S. 1982. Yield of tomato and maize in response to foliar and root applications of triacontanol. Plant Growth Regul. 1:11–14.
FRATERNALE, D; GIAMPERI, L; RICCI, D; ROCCHI, M; GUIDI, L; EPIFANO, F; MARCOTULLIO, M. 2003. The effect of triacontanol on micropropagation and on secretory system of Thymus mastichina. Plant cell, tissue and organ culture. 74: 87-97.
GIRIDHAR, P; RAJASEKARAN, T; RAVISHANKAR, G. 2005. Improvement of growth and root specific flavor compound 2-hydroxy-4-methoxy benzaldehyde of micropropagated plants of Decalepis hamiltonii Wight & Arn., under triacontanol treatment. - Scientia Hort. 106: 228-236.
GIRIDHAR, P; INDU, E; CHANDRASEKAR, A; RAVISHANKAR, G. 2004. Influence of triacontanol on somatic embryogenesis in Coffea arabica L. and Coffea canephora P.ex.Fr. - In Vitro cell. dev. Biol. Plant. 40: 200-203.
HOAGLAND, R. 1980. Effects of triacontanol on seed germination and early growth. Bot. Gaz. 141:53–55.
KAWASHIMA, S; MURATA, Y; SAKANE, K; NAGOSHI, T; TOI, Y; NAKAMURA, T. 1987. Effect of foliar application of triacontanol on the growth and yield of rice plants. JPN J Crop Sci. 56: 553-562.
KNIGHT, S; MITCHELL, C. 1987. Stimulation productivity of hydroponic lettuce in controlled environments with triacontanol. HortScience 22:1307–1309.
LESNIAK, A; HAUG, A; RIES, S. 1989. Stimulation of ATPase activity in barley (Hordeum vulgare) root plasma membranes after treatment with triacontanol and calmodulin. Physiol Plant. 75: 75-80.
MA, F; WANG, J; RONG, W. 1990. Effect of plant growth regulators on in vitro propagation of apple cultivar Fuji. J. Fruit. Sci. 7:201–206.
MOORTHY, P; KATHIRESAN, K. 1983. Physiological responses of a mangrove seedling to triacontanol. Biol. Plant. 35:577–581.
NAEEM, M; KHAN, M; MOINUDIN; SIDDIQUI, M. 2009. Triacontanol stimulates nitrogen-fixation, enzyme activities, photosynthesis, crop productivity and quality of hyacinth bean (Lablab purpureus L.). Sci Hortic 121:389–396.
NAEEM, M; MASROOR, M; MOINUDDIN, MOHD, I; TARIQ,A. 2011. Triacontanol-mediated regulation of growth and other physiological attributes, active constituents and yield of Mentha arvensis L. Plant Growth Regul. 65:195–206
OLSSON, R; PEARSON, J. 1990. Cardiovascular purinoceptors. Physiol Rev 70: 761-845.
REDDY, B; GIRIDHAR, P; RAVISHANKAR, G. 2002. The effect of triacontanol on micropropagation of Capsicum frutescens and Decalepis hamiltonii W&A. Plant Cell Tiss. Organ Cult. 71:253–258.
RIES, S. 1985. Regulation of plant growth with triacontanol. CRC Crit Rev Plant Sci. 2 :239–285.
RIES S. 1991. TRIA and its second messenger 9-β-L(+)-adenosine as plant growth substances. Plant Physiol. 95: 986-989.
RIES, S; HOUTZ, R. 1983. Triacontanol as a plant growth regulator. Hort Sci. 18: 654–662.
RIES, S; WERT, V; BIENBAUM, J. 1984. Interference of triacontanol activity by chemical constituents present in experimental and field sprayers. J. Am. Soc. Hort. Sci. 109:145–150.
RIES, S; WERT, V; BIERNBAUM, J; GIBSON, T; BARDLEY, W. 1983. Factors altering response of plants to triacontanol. HortScience. 108:917–922.
RIES, S; WERT, V. 1988. Rapid elicitation of second messengers by nanomolar doses of triacontanol and octacosanol. Planta. 173: 79-8.

RIES, S; WERT, V; O'LEARY, D; NAIR, M. 1990. 9-/3-L(+)-Adenosine: a new naturally occurring plant growth substance elicited by triacontanol in rice. Plant Growth Regul. 9: 263-273.
RIES, S; WERT, V; SWEELEY, C; LEAVITT; R. 1977. Triacontanol: a new naturally occurring plant growth regulator. Science 195: 1339–1341.
Shanghai Institute of Plant Physiology, Academia Sinica. 1987. Abstracts, International Symposium on Triacontanol, November. 25-28. Zhenjiang, China.
SHRIPATHI, V; SWAMY, G. 1994. Effect of triacontanol on the lipid composition of cotton (Gossypium hirsutum L.) leaves and its in-teraction with indole-3-acetic acid and benzyl adenine. Plant Growth Regul. 14:45-50.
SUN RAIN, Biological Products Co., Ltd. Zhengzhou. Casa comercial. Consultado el 10 de octubre del 2001. Disponible en:
1. http://www.made-in-china.com/showroom/dengyanzi/product-detaillbCmkLFMnGUo/China-Plant-Growth-Regulator-Triacontanol.html
2. http://young1982.en.made-in-china.com/product/pDQJmlAPzngM/China-Triacontanol.html
3. http://tuchem.en.made-in-china.com/product/UeImvRXMrCkK/China-Triacontanol.html
TANTOS, A; MESZAROS, A; KISSIMON, J; HORVATH, G; FARKAS, T. 2001. Triacontanolsupported micropropagation of woody plants. Plant Cell Rep. 19:88–91.
YASEEN, M; TAJUDDIN, K. 1988. Effect of plant growth regulators on yield, oil composition and artemisinina of Artemisia annua under temperate conditions. J. Med. Aromat. Plant Sci. 20:1038–1041.
YUN, C; KIM, H. 1986. Differences in callus formation from different explants sources and effect of plant growth regulators in plant regeneration in rice (Oryza sativa L.). Res. Rep. Rur. Dev. Admin. 1:113–116.





























