Embed Size (px)
DESCRIPTION
Biosel
Citation preview

APOPTOSIS, NEKROSIS, KANKER DAN AGING
Resume
Disusun untuk memenuhi tugas Matakuliah Biologi Sel yang dibina oleh
Prof. Dr. Agr . Mohamad Amin, S.Pd
Oleh
Kelompok 2 / Kelas A
Candra Hermawan (130341818685)
Dewi Sagita (130341818675)
Fendy Hardian Permana (130341816927)
“The Learning University”
UNIVERSITAS NEGERI MALANG
FAKULTAS PASCA SARJANA
PENDIDIKAN BIOLOGI
November, 2013

MEKANISME DAN REGULASI APOPTOSIS
A. Pengertian Apoptosis
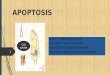
Apoptosis adalah mekanisme kematian sel yang terprogram yang penting dalam berbagai
proses biologi. Berbeda dengan nekrosis, yang merupakan bentuk kematian sel sebagai akibat sel
yang terluka akut, apoptosis terjadi dalam proses yang diatur sedemikian rupa yang secara umum
memberi keuntungan selama siklus kehidupan suatu organisme (Gambar 1). Contohnya adalah
pada diferensiasi jari manusia selama perkembangan embrio membutuhkan sel-sel di antara jari-
jari untuk apoptosis sehingga jari-jari dapat terpisah.
B. Peranan Apoptosis
Apoptosis memiliki peranan penting dalam fenomena biologis, proses apoptosis yang
tidak sempurna dapat menyebabkan timbulnya penyakit yang sangat bervariasi. Terlalu banyak
apoptosis menyebabkan sel mengalami kekacauan, sebagaimana terlalu sedikit apoptosis juga
menyebabkan proliferasi sel yang tidak terkontrol (kanker). Beberapa contoh penyakit yang
ditimbulkan karena apoptosis yang tidak sempurna antara lain:
1. Penyakit autoimun disebabkan karena sel T/B yang autoreaktif terus menerus.
2. Neurodegeneration, seperti pada penyakit Alzheimer dan Parkinson, akibat dari apoptosis
prematur yang berlebihan pada neuron di otak. Neuron yang tersisa tidak mempunyai
kemampuan untuk meregenerasi sel yang hilang.
3. Stroke iskemik, aliran darah ke bagian-bagian tertentu dari otak dibatasi sehingga dapat
menyebabkan kematian sel saraf melalui peningkatan apoptosis.
4. Kanker, sel tumor kehilangan kemampuannya untuk melaksanakan apoptosis sehingga
proliferasi sel meningkat.
C. Fungsi Apoptosis
1. Sel yang rusak atau terinfeksi
Apoptosis dapat terjadi secara langsung ketika sel yang rusak tidak bisa diperbaiki lagi
atau terinfeksi oleh virus. Keputusan untuk melakukan apoptosis dapat berasal dari sel itu
sendiri, dari jaringan di sekitarnya, atau dari sel yang merupakan bagian sistem imun.
Jika kemampuan sel untuk ber-apoptosis rusak atau jika inisiasi apotosis dihambat, sel
yang rusak dapat terus membelah tanpa batas, berkembang menjadi kanker.
2. Respon terhadap stress atau kerusakan DNA
Kondisi stress sebagaimana kerusakan DNA sel yang disebabkan senyawa toksik atau
pemaparan sinar ultraviolet atau radiasi ionisasi (sinar gamma atau sinar X), dapat
menginduksi sel untuk memulai proses apoptosis. Contohnya pada kerusakan genom
dalam inti sel, adanya enzim PARP-1 memacu terjadinya apoptosis. Enzim ini memiliki
peranan penting dalam menjaga integritas genom, tetapi aktivasinya secara berlebihan
dapat menghabiskan ATP, sehingga dapat mengubah proses kematian sel menjadi
nekrosis (kematian sel yang tidak terprogram).
D. Mekanisme Apoptosis
Mekanisme apoptosis sangat kompleks dan rumit. Secara garis besarnya apoptosis dibagi
menjadi 4 tahap, yaitu:
1. Adanya signal kematian (penginduksi apoptosis)
2. Tahap integrasi atau pengaturan (transduksi signal, induksi gen apoptosis yang
berhubungan, dll)

3. Tahap pelaksanaan apoptosis (degradasi DNA, pembongkaran sel, dll)
4. Fagositosis.
E. Signal Penginduksi Apoptosis
Apoptosis tidak memerlukan suatu proses transkripsi atau translasi. Molecular machine
yang dibutuhkan untuk kematian sel dianggap mengalami dormansi dan hanya memerlukan
aktivasi yang cepat. Signal yang menginduksi apoptosis bisa berasal dari ekstraseluler dan
intraseluler. Signal ekstraseluler contohnya hormon hormon. Hormon tiroksin menginduksi
apoptosis pada ekor tadpole. Apoptosis juga bisa dipicu oleh kurangnya signal yang dibutuhkan
sel untuk bertahan hidup seperti growth factor. Sel lain, sel berhubungan dengan sel yang
berdekatan juga bisa memberikan signal untuk apoptosis.
Signal intraseluler misalnya radiasi ionisasi, kerusakan karena oksidasi radikal bebas,
dan gangguan pada siklus sel. Kedua jalur penginduksi tersebut bertemu di dalam sel, berubah
menjadi famili protein pengeksekusi utama yang dikenal sebagai caspase. Sel yang berbeda
memberikan respon yang berbeda terhadap penginduksi apoptosis. Misalnya sel splenic limfosit
akan mengalami apoptosis saat terpapar radiasi ionisasi, sedangkan sel myocyte tidak mengalami
apoptosis untuk pemaparan yang sama. Signal kematian dihubungkan dengan pelaksanaan
apoptosis oleh tahap integrasi atau pengaturan. Pada tahap ini terdapat molekul regulator positif
atau negatif yang dapat menghambat, memacu, mencegah apoptosis sehingga menentukan
apakah sel tetap hidup atau mengalami apoptosis (mati).
Apoptosis diperantarai oleh famili protease yang disebut caspase, yang diaktifkan melalui
proteolisis dari bentuk prekursor inaktifnya (zymogen). Caspase merupakan endoprotease yang
memiliki sisi aktif Cys (C) dan membelah pada terminal C pada residu Asp, oleh karena itu
dikenal sebagai Caspases (Cys containing Asp specific protease). Saat ini telah ditemukan 13
anggota famili caspases pada manusia. Beberapa anggota famili caspase yang terlibat dalam
apoptosis dibedakan menjadi 2 golongan. Golongan yang pertama terdiri dari caspase 8, 9,10
yang mengandung prodomain yang panjang pada terminal N, fungsinya sebagai inisiator dalam
proses kematian sel. Golongan yang kedua terdiri dari caspase 3, 6, 7 yang mengandung
prodomain yang pendek dan berfungsi sebagai efektor, membelah berbagai substrat yang mati
yang pada akhirnya menyebabkan perubahan morfologi dan biokimia yang tampak pada sel yang
mengalami apoptosis. Molekul efektor lain dalam apoptosis adalah Apaf-1 (apoptotic protease

activating factor) bersama sitokrom c mengambil pro-caspase 9 di ATP-dependent manner, dan
menstimulasi proses perubahan pro-caspase 9 menjadi caspase 9.
Regulator apoptosis yang lain adalah anggota famili Bcl-2. Saat ini ada 18 anggota famili
Bcl-2 yang telah diidentifikasi, dan dibagi ke dalam 3 grup berdasarkan strukturnya. Anggota
grup pertama diwakili oleh Bcl-2 dan Bcl-xL yang berfungsi sebagai anti-apoptosis. Anggota
grup kedua diwakili oleh Bax dan Bak (Bcl-2 associated killer), sebagaimana anggota grup yang
ketiga yaitu Bid (a novel BH3 domain-only death agonist) dan Bad (the Bcl-2 associated death
molecule), merupakan molekul pro-apoptosis (Gambar 2).
ICE (Interleukin Converting Enzim) secara normal tidak terlibat dalam apoptosis, tetapi
aktivasi tiruannya dalam sel mamalia, dapat mendorong ke arah tersebut. Masing-masing caspase
mempunyai urutan yang sama, dirancang untuk membelah, maka menjadi jelas caspase
membelah satu sama lain dalam suatu jalur mekanisme pengaktifan. Dua rangkaian caspase
saling melibatkan. Yang satunya menginisiasi proses aktivasi caspase lainnya. Pertanyaannya
siapa yang mengaktifkan caspase yang pertama? Tampak meragukan, sampai peneliti
menemukan bahwa caspase dapat diaktifkan jika mereka mengumpul pada konsentrasi kritik. Ini
bisa terjadi oleh ikatan molekul signal bunuh diri di permukaan sel. Perubahan konformasi
reseptor dapat mendorong ke arah agregasi dari molekul reseptor permukaan dengan serentak
dengan agregasi caspases intraseluler reseptor agregasi.

F. Target Caspase
Apoptosis melibatkan:
1. memadatkan inti sel
2. memadatkan dan membagi-bagi sitoplasma ke dalam selaput ikat badan apoptotis
3. rusaknya kromosom ke dalam fragmen yang berisi berbagai nukleosom
Target protein pada umumnya harus protein lain, suatu DNA endonuklease. Ketika
protein target pecah, DNase bebas untuk berpindah tempat ke inti dan mulai pelaksanaan.
Perubahan dalam apoptosis terjadi ketika caspase 3 membelah gelsolin, suatu protein dilibatkan
dalam pemeliharaan morfologi sel. Gelsolin yang dibelah membelah actin filamen di dalam sel.
Protein yang lain diperlukan untuk membentuk badan apopotic: suatu kinase yang disebut p21-
activated kinase 2 (PAK-2). Kinase ini diaktifkan oleh caspase-3 dengan proteolisis terbatas.
G. Tahap Pelaksanaan Apoptosis
Sinyal apoptosis bisa terjadi secara intraseluler dan ekstraseluler. Jalur ekstrinsik
(ekstraseluler) diinisiasi melalui stimulasi dari reseptor kematian (death receptor) sedangkan
jalur intrinsik diinisiasi melalui pelepasan faktor signal dari mitokondria dalam sel. Peristiwa
apoptosis jalur ekstrinsik dimulai dari adanya pelepasan molekul signal yang disebut ligan oleh
sel lain tetapi bukan berasal dari sel yang akan mengalami apoptosis. Ligan tersebut berikatan
dengan death receptor yang terletak pada transmembran sel target yang menginduksi apoptosis.
Death receptor yang terletak di permukaan sel adalah famili reseptor TNF (Tumor Necrosis
Factor), yang meliputi TNF-R1, CD 95 (Fas), dan TNF-Related Apoptosis Inducing Ligan
(TRAIL)-R1 dan R2.
Ligan yang berikatan dengan reseptor tersebut akan mengakibatkan caspase inisiator 8
setelah membentuk trimer dengan adaptor FADD (Fas Associeted Death Domain). Kompleks
yang terbentuk antara ligan-reseptor dan FADD disebut DISC (Death Inducing Signaling
Complex). CD 95, TRAIL-R1 dan R2 terikat dengan FADD, sedangkan TNF-R1 terikat secara
tidak langsung melalui molekul adaptor lain, yaitu: TNF-Reseptor Associeted Death Domain
protein (TRADD).
Stress mitokondria yang menginduksi apoptosis jalur intrinsik disebabkan oleh senyawa
kimia atau kehilangan faktor pertumbuhan, sehingga menyebabkan gangguan pada mitokondria
dan terjadi pelepasan sitokrom c dari intermembran mitokondria. Protein capcase-8 akan

memotong anggota famili Bcl-2 yaitu Bid. Kemudian Bid yang terpotong pada bagian ujungnya
akan menginduksi insersi Bax dalam membran mitokondria dan melepaskan molekul
proapoptotik seperti sitokrom c, Samc/Diablo, Apoptosis Inducing Factor (AIF), dan omi/Htr2.
dengan adanya dATP akan terbentuk kompleks antara sitokrom c, APAF1 dan caspase 9 yang
disebut apoptosom. Selanjutnya, capcase 9 akan mengaktifkan downstream procaspase-3. Protein
caspase 3 yang aktif memecah berbagai macam substrat, diantaranya enzim DNA repair seperti
poly-ADP Ribose Polymerase (PARP) dan DNA protein kinase yaitu protein struktural seluler
dan nukleus, termasuk aparatus mitotik inti, lamina nukleus, dan aktin serta endonuklease,
seperti Caspase-Aktivated Deoxyribonuklease Inhibitor (ICAD) dan konstituen seluler lainnya.
Selain itu, caspase 3 juga mempunyai kemampuan untuk mengaktifkan caspese lainnya, seperti
procaspase-6 dan procaspase-7 yang memberikan amplifikasi terhadap kerusakan seluler.
Adanya seluler stres meningkatkan ekspresi dari protein p53 yang mengakibatkan
terjadinya GI arrest atau apoptosis. Anggota dari apoptosis Stimulating Protein p53 (ASPP) yaitu
ASPP 1 dan ASPP 2 secara spesifik menstimulasi fungsi transsktivasi p53 pada promotor gen
proapoptotik seperti Bax dan p53 Inducible Gene 3 (PIG 3), tapi tidak pada promotor gen yang
menyebabkan cell cycle arrest, yaitu p21 dan MDM2.
H. Tahapan Apoptosis Jalur Ekstrinsik/Deatch Receptor Pathway
Jalur ini khas pada sistem imun dan digunakan untuk menghilangkan sel T yang aktif
pada akhir dari respon imun. Jalur ini terutama diperantarai oleh perforin / granzyme. Tahap-
tahap apoptosis dalam death receptor pathway:
1. Ikatan antara FasL, suatu TNF (Tumor Necrosis Factor) dengan reseptornya. TNF adalah
molekul penginduksi interseluler yang berupa asam amino-157, dihasilkan terutama oleh
makrofag yang teraktivasi, merupakan mediator apoptosis ekstrinsik utama. Ada 2
macam reseptor untuk TNF yaitu TNFR-1 dan TNFR-2. TNF yang berikatan dengan
TNFR-1 yang dapat menginisiasi jalur aktivasi caspase. Fas (Apo-1 atau CD 95) adalah
reseptor untuk signal apoptosis ekstrinsik lain pada membran sel, dan termasuk famili
reseptor TNF. FasL (Fas ligan) adalah protein yang berikatan dengan Fas untuk
mengaktifkan jalur Fas. Fas merupakan protein transmembran yang juga termasuk famili
TNF.

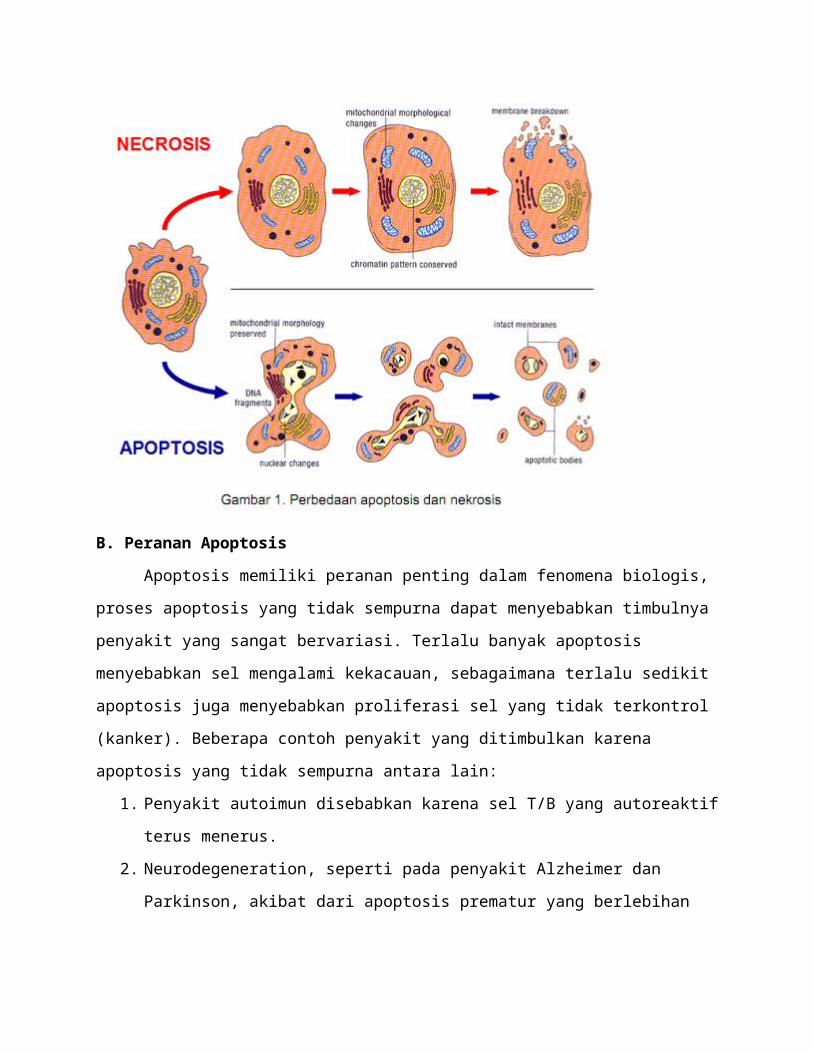
2. Ikatan FasL dengan Fas menginduksi reseptor untuk mengelompok (trimerisasi)
Gambar Ikatan FasL dengan Fas menyebabkan trimerisasi reseptor
3. Pengikatan FADD (Fas associated death domain protein) pada domain kematian (death
domain).
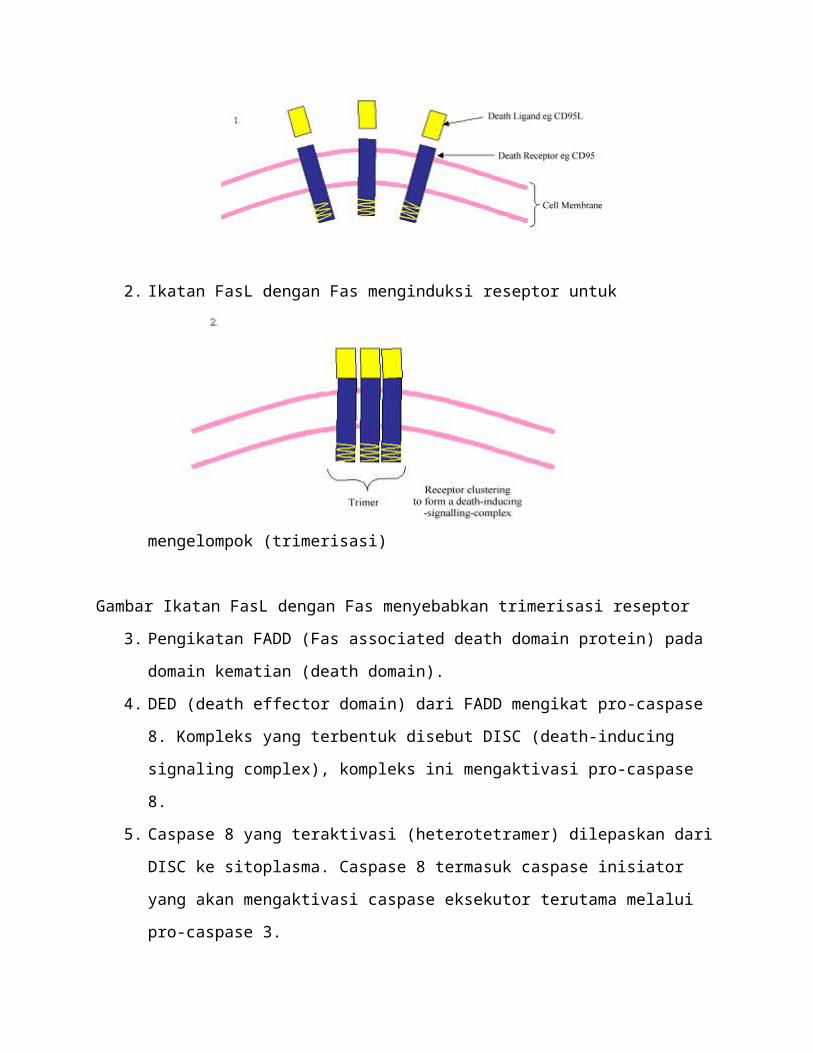
4. DED (death effector domain) dari FADD mengikat pro-caspase 8. Kompleks yang
terbentuk disebut DISC (death-inducing signaling complex), kompleks ini mengaktivasi
pro-caspase 8.
5. Caspase 8 yang teraktivasi (heterotetramer) dilepaskan dari DISC ke sitoplasma. Caspase
8 termasuk caspase inisiator yang akan mengaktivasi caspase eksekutor terutama melalui
pro-caspase 3.

I. Mitocondrial Pathway
Riset mengindikasi keterlibatan mitokondria dalam jalur apoptotis. Sitokrom c, suatu
heme protein yang bertindak sebagai suatu pembawa elektron dalam fosforilasi oksidasi
mitokondria, pemberhenti elektron cytochrome C oxidase atau kompleks IV, keluar
intermembran dan mengikat protein sitoplasmik yang disebut Apaf-1. Yang kemudian
mengaktikan suatu inisiator caspase-9 di sitoplasma. Protein ini keluar mitokondria setelah
perubahan potensiasi eletrokimia di membran. Perubahan potensial menyebabkan terbukanya
suatu kanal yang non-spesifik dalam membran yang permeabel, terdiri atas dua protein selaput
bagian dalam (adenine nucleotide translocator-ANT) dan suatu protein bagian luar (porin, yang
voltage-gated-kanal anion VDAC). Protein ini bertindak bersama-sama, kemungkinan pada sisi
luar dan sisi dalam terjadi kontak. Saluran ini dapat dilewati zat yang memiliki bobot molekular
kurang dari 1500. Perubahan gradien proton menyebabkan oksidasi dan foforilasi di mitokondria
perubahan kekuatan ion menyebabkan pembekakan matriks. Karena sisi bagian dalam sangat
kusut dan memilki luas permukaan jauh lebih besar dibanding selaput yang luar, bengkak pada
matriks mengarah rusaknya sisi luar, sehingga sitokrom c dan Apaf-1 keluar masuk sitoplasma.
Jalur ini biasa diaktifkan dalam respon stimulus letal yang lain seperti pengrusakan DNA, stress
oksidatif, dan hipoksia. Mitokondria mengandung faktor pro-apoptosis seperti sitokrom c dan

AIF (apoptosis inducing factors). Keduanya merupakan substrat yang berbahaya, akan tetapi
tersimpan aman dalam mitokondria. Saat keduanya dilepaskan ke sitoplasma dapat mengaktifkan
jalur aktivasi caspase. Pelepasannya diatur oleh famili Bcl-2 yang terikat dengan mitokondria,
yaitu Bax dan Bad.
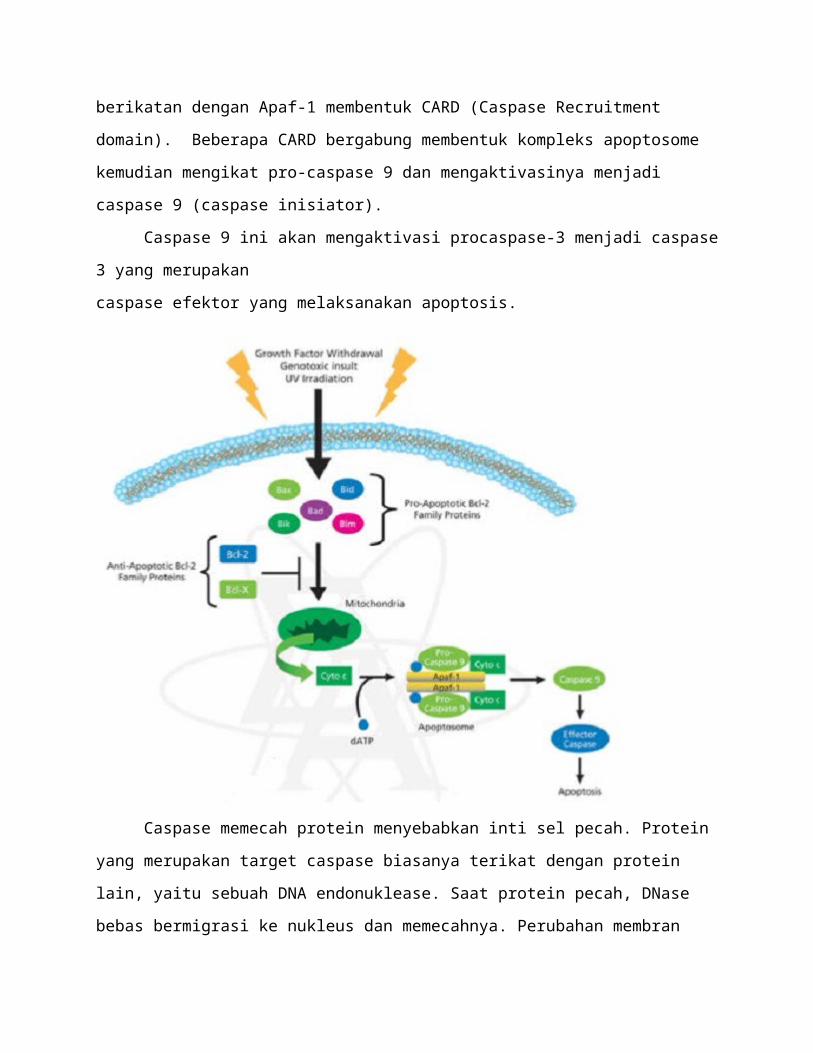
Sitokrom c adalah protein heme yang berperan sebagai pembawa elektron yang larut
dalam air dalam fosforilasi oksidatif mitokondria. Bila terjadi kumparan elektron melalui
sitokrom c oxidase atau kompleks IV, adanya perubahan kekuatan ion menyebabkan gelombang
matriks. Saat membran dalam mitokondria memiliki permukaan yang lebih luas dibanding
membran luar maka gelombang matriks menyebabkan nonspecific inner membrane permeability
transition pore terbuka sehingga sitokrom c keluar ke sitoplasma. Sitokrom c yang keluar ke
sitoplasma kemudian berikatan dengan Apaf-1 membentuk CARD (Caspase Recruitment
domain). Beberapa CARD bergabung membentuk kompleks apoptosome kemudian mengikat
pro-caspase 9 dan mengaktivasinya menjadi caspase 9 (caspase inisiator).
Caspase 9 ini akan mengaktivasi procaspase-3 menjadi caspase 3 yang merupakan

caspase efektor yang melaksanakan apoptosis.
Caspase memecah protein menyebabkan inti sel pecah. Protein yang merupakan target
caspase biasanya terikat dengan protein lain, yaitu sebuah DNA endonuklease. Saat protein
pecah, DNase bebas bermigrasi ke nukleus dan memecahnya. Perubahan membran terjadi saat
caspase 3 memecah gelsolin, suatu protein yang terlibat dalam pemeliharaan morfologi sel.
Gelsolin yang terpecah akan membelah filamen aktin di dalam sel. Caspase 3 juga mengaktivasi
kinase yang disebut p21-activated kinase 2 (PAK 2) melalui proteolisis. PAK2 termasuk protein
yang dibutuhkan dalam membentuk apoptotic body. Selama apoptosis mitokondria mengalami
perubahan yang disebabkan oleh:
a. Gangguan oksidasi-fosforilasi dan transport elektron karena radiasi dan adanya second
messenger tertentu seperti ceramide.
b. Perubahan dalam potensial redoks sel dan turunan Reactive Oxygen Species (ROS).
c. Kerusakan DNA.
d. Kerusakan DNA memacu ekspresi protein yang dikenal sebagai p53. Protein ini
menyebabkan penghambatan pembelahan sel atau apoptosis, dimana keduanya akan
mnjaga sel dari menjadi sel tumor. Oleh karena itu gen p53 adalah gen tumor suppressor.
e. Peningkatan ion Ca2+ intraseluler melalui tranduksi signal. Death Receptor Pathway dan
Mitocondrial Pathway bertemu saat caspase inisiator (caspase 8, 9, 10) menghasilkan
aktivasi caspase efektor (caspase 3, 6, 7).
Gambar Pertemuan Death ReceptorPathway dan Mitocondrial Pathway
J. Tahap Fagositosis
Sel yang terfragmentasi menjadi apoptotic body mengeluarkan signal “eat me”
yang dikenali oleh fagosit. Ada 2 macam fagosit, yaitu:
1. Fagosit professional, contohnya sel makrofag.
2. Fagosit semiprofesional, sel tetangga dari sel yang mengalani apoptosis.
Adanya sel-sel fagosit ini dapat menjamin tidak timbulnya respon inflamasi setelah terjadinya
apoptosis. Sel fagosit juga harus dihilangkan setelah aktif bekerja. Sel imun aktif mulai
mengekspresikan Fas beberapa hari setelah aktivasi, mentargetkannya untuk eliminasi. Beberapa
sel yang stress dapat mengekspresikan Fas dan FasL lalu digunakan untuk bunuh diri. Akan
tetapi sebagian besar hanya dapat mengekspresikan Fas, sedangkan FasL diekspresikan terutama
oleh sel T aktif.
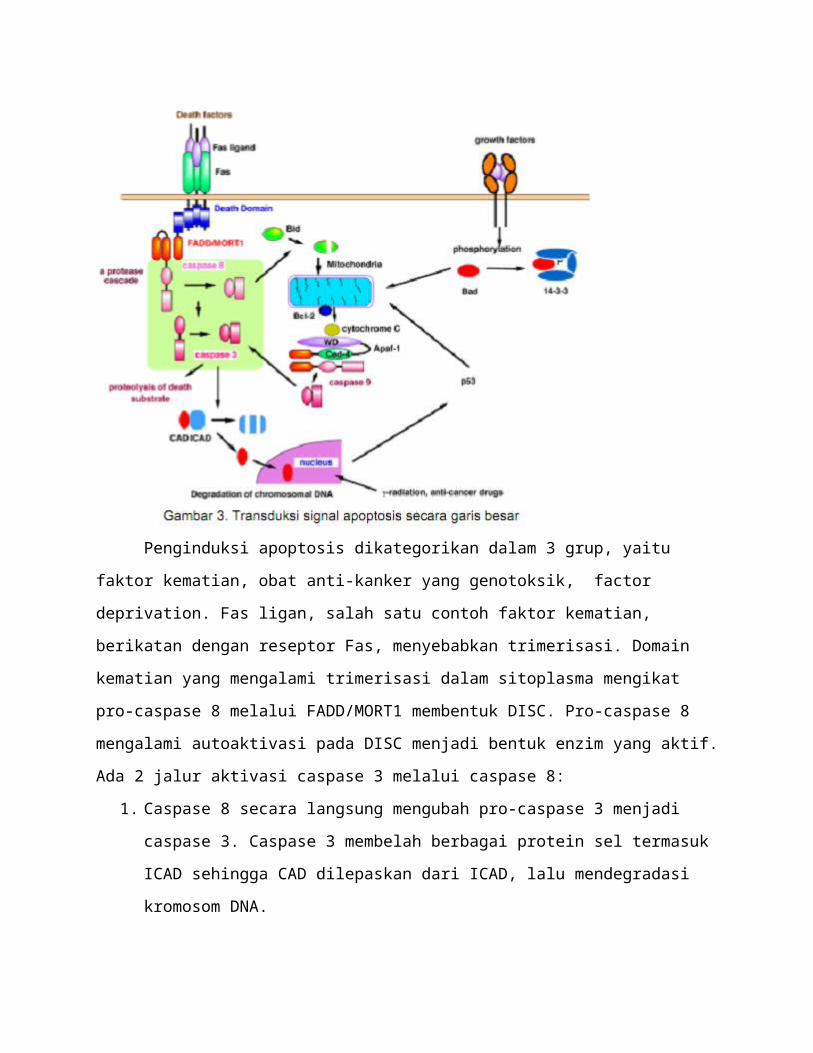
Penginduksi apoptosis dikategorikan dalam 3 grup, yaitu faktor kematian, obat anti-
kanker yang genotoksik, factor deprivation. Fas ligan, salah satu contoh faktor kematian,
berikatan dengan reseptor Fas, menyebabkan trimerisasi. Domain kematian yang mengalami
trimerisasi dalam sitoplasma mengikat pro-caspase 8 melalui FADD/MORT1 membentuk DISC.
Pro-caspase 8 mengalami autoaktivasi pada DISC menjadi bentuk enzim yang aktif. Ada 2 jalur
aktivasi caspase 3 melalui caspase 8:
1. Caspase 8 secara langsung mengubah pro-caspase 3 menjadi caspase 3. Caspase 3
membelah berbagai protein sel termasuk ICAD sehingga CAD dilepaskan dari ICAD,
lalu mendegradasi kromosom DNA.
2. Caspase 8 membelah Bid, molekul pro-apoptosis yang termasuk famili Bcl-2, yang
kemudian ditranslokasikan ke mitokondria untuk melepaskan sitokrom c ke sitosol. Bcl-2
atau Bcl-xl, molekul anti-apoptosis, dapat menghambat pelepasan sitokrom c dengan
mekanisme yang belum diketahui dengan pasti. Sitokrom c bersama Apaf-1
mengaktifkan Caspase 9, dimana caspase 9 kemudian mengaktifkan caspase 3. Caspase 3

membelah berbagai protein sel termasuk ICAD sehingga CAD dilepaskan dari ICAD lalu
mendegradasi kromosom DNA.
Obat anti-kanker yang genotoksik seperti etoposida dan radiasi γ menyebabkan kerusakan
kromosom DNA. Signal tersebut ditransfer ke mitokondria oleh p53 melalui mekanisme yang
belum diketahui. Hal ini dapat menyebabkan pelepasan sitokrom c dari mitokondria dan
mengaktifkan caspase 9 seperti dijelaskan di atas. Apoptosis yang diinduksi oleh factor
deprivation dapat dipelajari dengan baik menggunakan IL-3 dependent myeloid cell lines.
Dengan keberadaan IL-3, signal dari reseptor IL-3 menyebabkan fosforilasi Bad, molekul pro-
apoptosis famili Bcl-2. Bad yang terfosforilasi tertangkap oleh adaptor 14-3-3. Bila IL-3 sudah
tidak ada lagi maka Bad yang tak terfosforilasi dilepaskan dari 14-3-3, lalu ditranslokasikan ke
mitokondria untuk melepaskan sitokrom c untuk mengaktifkan caspase 9.
K. Pengendalian Apoptosis
Haruslah jelas sel menjaga kontrol caspases. Dua spesies untuk menginhibisi
apoptosis adalah protein mitochondrial Bcl-2 dan Bcl-xL, yang dapat menghalangi pelepasan
sitokrom c dari mitokondria. Protein keluarga Bcl mempunyai suatu gugus hidrofob dan terikat
di sisi luar permukaan mitokondria dan organel lain seperti inti dan retikulum endoplasma.
Protein ini mampu membentuk kanal ion di liposom. Sejauh ini 15 anggota keluarga ini (ced-9
yang dihubungkan dengan C. elegans) telah ditemukan di manusia. Bcl-2 dapat juga mengikat
Apaf-1 dan menghalangi pengaktifan inisiasi caspase 9. Bcl-2 diatur oleh perubahan ekspresi gen
Bcl-2, dengan post-translational fosforilasi oleh kinase, atau oleh pecahnya caspase. Kelebihan
ekpresi Bcl-2 dapat menyebabkan suatu sel menjadi suatu sel tumor. Anggota lain keluarga,
BAX dan BAD yang mengikat mitokondria dan memfasilitasi apoptosis dengan menstimulasi
pelepasan sitokrom C. Sebagai tambahan, protein lain yang disebut IAPS (inhibitor of apoptosis)
dapat menghalangi caspase atau protein apoptotis lainnya.

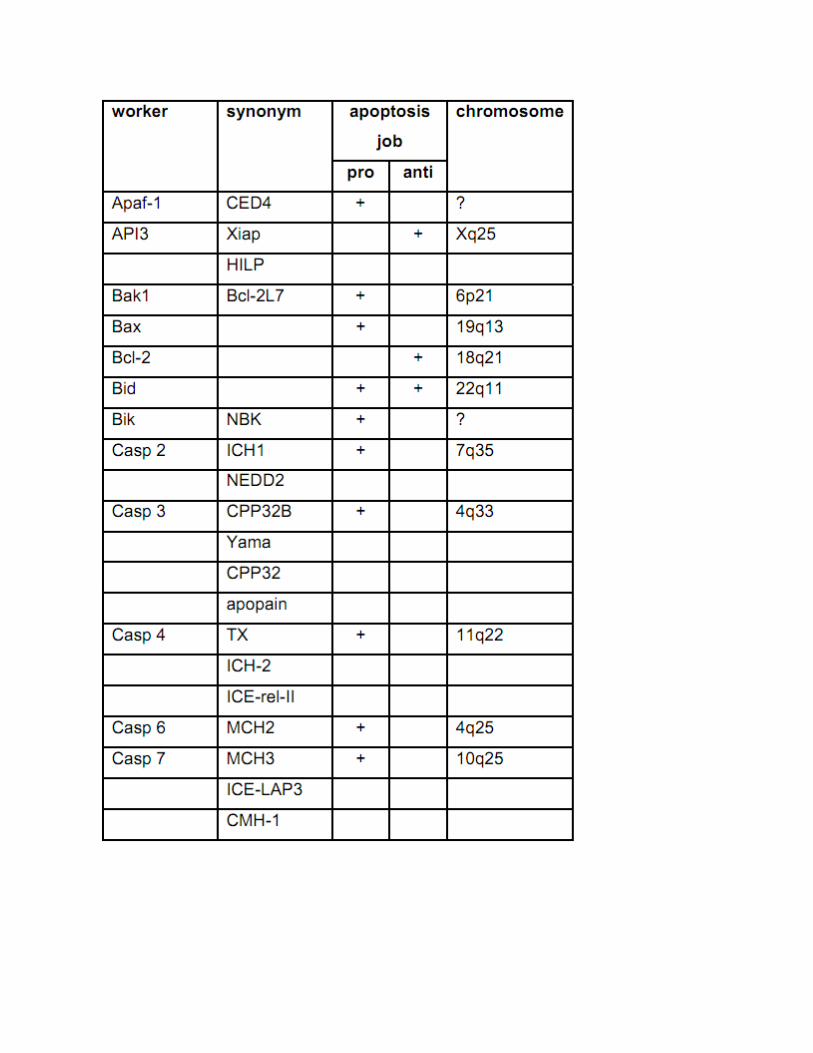
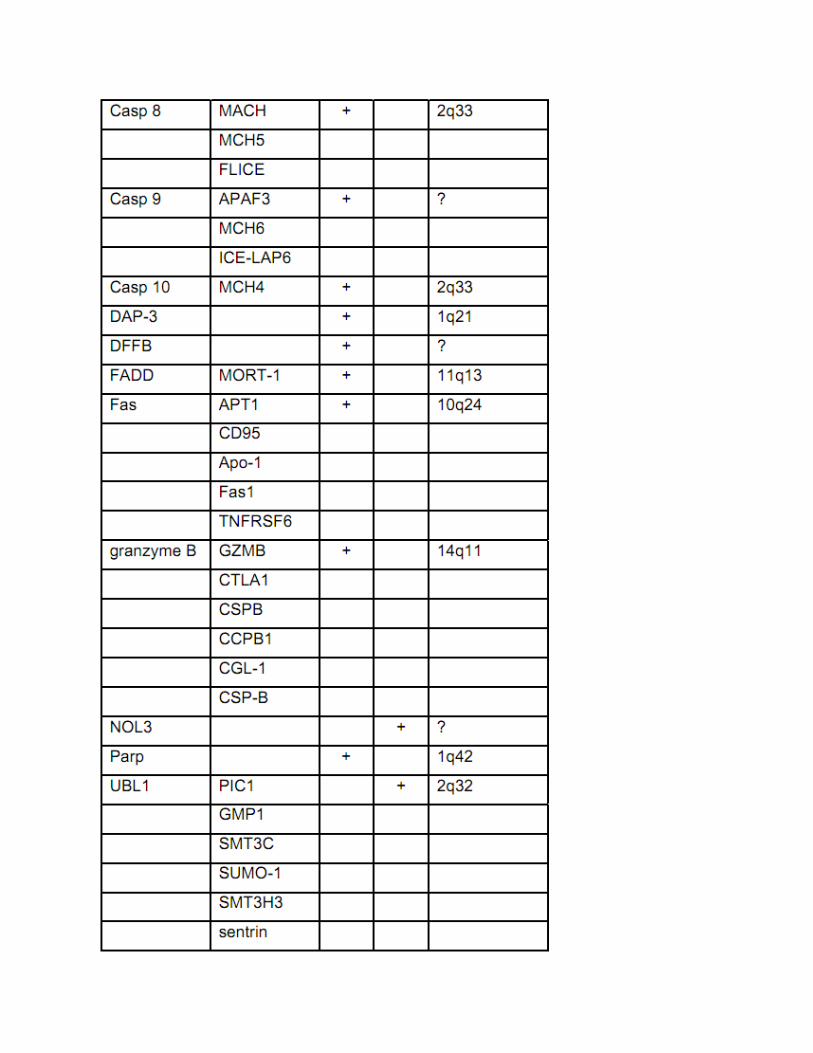
Beberapa molekul regulator yang berperan dalam apotosis

L. Mengenali sel yang apoptosis
Sel yang mengalami apoptosis dapat diamati dengan menggunakan mikroskop cahaya
maupun mikroskop elektron melalui ciri-ciri morfologis yang ditampakkan. Ciri-ciri tersebut
antara lain:
a. Sel menjadi bulat (sirkuler). Ini terjadi karena struktur protein yang menyusun
sitoskeleton dicerna oleh enzim peptidase spesifik yang disebut caspaspse yang telah
diaktifkan di dalam sel.
b. Kromatin (DNA dan protein-protein yang terbungkus di dalam inti sel) mulai mengalami
degradasi dan kondensasi.
c. Kromatin mengalami kondensasi lebih lanjut, menjadi semakin memadat. Pada tahap ini,
membran yang mengelilingi inti sel masih tampak utuh, walaupun caspase tertentu telah
melakukan degradasi protein pori inti sel dan mulai mendegradasi lamin yang terletak
dalam lingkungan inti sel.
d. Lingkungan dalam inti sel tampak terputus dan DNA di dalamnya terfragmentasi (proses
ini dikenal dengan karyorrhexis). Inti sel pecah melepaskan berbagai bentuk kromatin
atau unit nukleosom karena disebabkan degradasi DNA.
e. Plasma membran mengalami blebbing.
f. Sel tersebut kemudian di’makan’ atau pecah menjadi gelembung-gelembung yang
disebut apoptotic bodies dan kemudian di’makan’.
Sel yang mengalami apoptosis juga dapat dikenali dengan:
a. Penandaan inti yang mengalami kondensasi dengan pewarna fluorescence Hoechst atau
DAPI.
b. Sel yang mengalami apoptosis mengeluarkan PS (Phosphatidil Serin) pada permukaan
ekstraselulernya, sehingga dapat ditandai dengan annexin V yang dilabeli fluorescence.
PS secara normal terdapat pada cytosolic surface dari membran plasma (di bagian dalam
membran plasma), tetapi diredistribusikan ke permukaan ekstraseluler selama apoptosis
oleh protein hipotetik yang dikenal sebagai scramblase.
c. DNA yang terfagmentasi dapat dideteksi dengan TUNEL (Terminal
deoxynuclotidyltransferase-mediated UTP end labelling) atau elektroforesis DNA yang
diisolasi dalam gel agarosa. TUNEL juga dapat digunakan untuk mendeteksi enzim yang
terlibat dalam pengrusakan inti sel.


NEKROSIS
Nekrosis terjadi karena kerusakan membran, lisososm mengeluarkan enzim ke sitoplasma
dan menghancurkan sel, isi sel keluar dikarenakan kerusakan membran plasma dan
mengakibatkan reaksi inflamatori.
Nekrosis adalah pathway yang secara umum terjadi pada kematian sel yang diakibatkan
oleh:
- Ischemia
- Keracunan
- infeksi dan
- trauma

PERBEDAAN APOPTOSIS DAN NEKROSIS
Gambaran Mikroskopik :
A. Nukleus
Piknosis: nukleus terlihat lebih bundar, ukuran lebih kecil dan gelap
Karioreksis: nukleus mengalami fragmentasi menjadi kecil dan tersebar
Kariolisis: nukleus lisis, tidak terlihat sehingga rongga kosong dibatasi membran nukleus
disebut ghost.
B. Sitoplasma: berwarna asidofilik, struktur tidak jelas, jika melanjut :
1. Tidak terlihat garis besar struktur histologi sel
2. Tidak terlihat adanya pewarnaan
Tipe-tipe morfologik nekrosis jaringan
Secara makroskopik dan dengan pemeriksaan mikroskop dapat dikenali beberapa bentuk
nekrosis.
Bentuk-bentuk tersebut:
- Nekrosis koagulasi
Tidak hanya terjadi denaturasi protein, namun juga berkaitan dengan hambatan enzim-
enzim litik.
Sel tidak mengalami lisis, dengan demikian kerangka luar sel relatif utuh.
Inti menghilang dan sitoplasma yang mengalami asidifikasi menjadi eosinofilik
Nekrosis koagulasi-infrak ginjal
Gambaran makroskopik :
terlihat berwarna putih, keabu-abuan atau kekuning-kuningan dan sedikit
berlemak, padat
Gambaran mikroskopik :
struktur sel dan jaringan masih jelas, inti sel mengalami piknotik (menghilang),
sitoplasma lebih acidophilic
- Nekrosis liquefaktif (mencair)
Ditandai oleh larutnya jaringan akibat lisis enzimatik sel-sel yang mati.
Proses ini biasanya terjadi di otak sewaktu terjadi pelepasan enzim-enzim otokatalitik
dari sel-sel yang mati.
Nekrosis likuefaktif juga terjadi pada peradangan purulen akibat efek heterolitik leukosit
polimorfonuklear pada pus.
Jaringan yang mengalami likuefaksi menjadi lunak, mudah mencair, dan tersusun oleh
sel-sel yang mengalami disintegrasi dan cairan
Nekrosis liquefaktif-infark otak
Gambaran makroskopik :
adanya benjolan berisi cairan dikelilingi kapsula tipis dan ireguler.
Gambaran mikroskopik :
tampak ruang kosong dengan sisa kapsula yang ireguler, terlihat fibrin dan
neutrophil disekitarnya.
- Nekrosis lemak
Terjadi akibat kerja enzim-enzim lipolitik pada jaringan lemak.
Proses ini biasanya terjadi pada nekrosis pankreatik akut dan merupakan konsekuensi
pelepasan lipase pankreas ke jaringan peripankreas.
Lipolisis ditandai oleh hilangnya kontur sel-sel lemak.
Asam-asam lemak yang dibebaskan dari sel lemak mengalami saponifikasi dengan
mengikat natrium, kalium dan kalsium.
Nekrosis lemak-pankreatitis akut

- Nekrosis kaseosa (perkejuan)
Memiliki baik gambaran nekrosis koagulasi maupun likuefaktif.
Biasanya nekrosis ini terjadi di bagian tengah granuloma tuberkolusa, yang mengandung
bahan seperti keju yang putih atau kekuningandan merupakan asal nama nekrosis tipe ini.
Secara histologis, rangka luar sel tidak lagi utuh, tetapi sebaliknya jaringan juga belum
mencair.
Sisa-sisa sel tampak sebagai bahan amorf bergranula halus.
Nekrosis kaseosa-tuberculosis paru
Gambaran makroskopik :
terlihat berwarna putih, keabu-abuan atau kekuning-kuningan dan sedikit
berlemak, padat
Gambaran mikroskopik :
struktur histologi sudah tidak terlihat lagi membentuk masa bergranulasi.
Dengan pengecatan HE berwarna keabu-abuan, dikelilingi oleh epiteloid dan
limfosit.