Embed Size (px)
DESCRIPTION
esporas
Citation preview
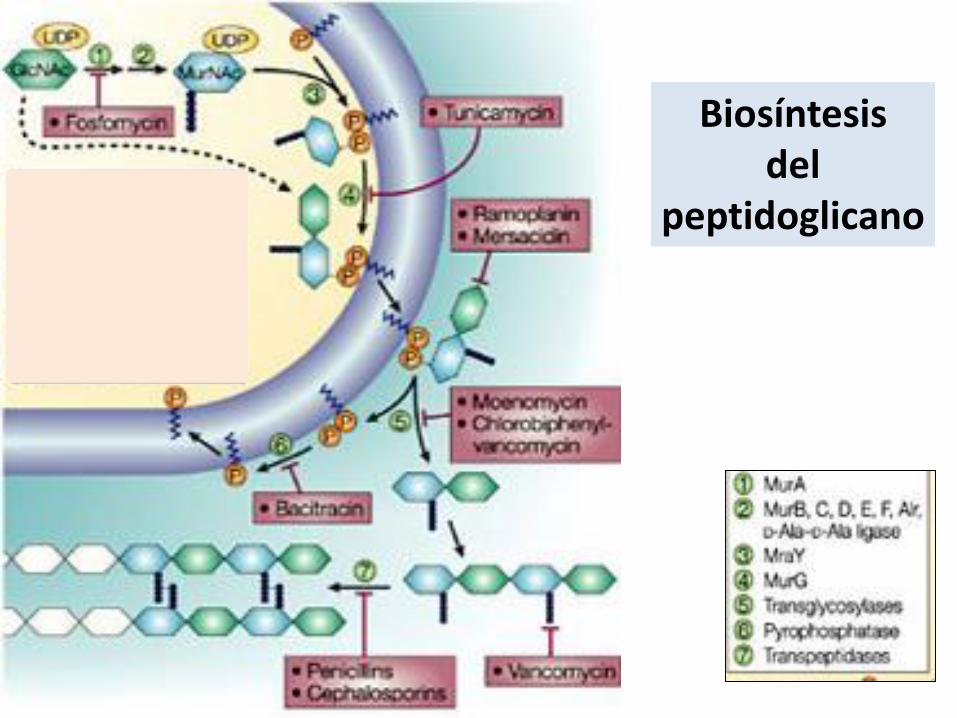
Biosíntesis del
peptidoglicano

Paredes bacterianas
Por que las estudiamos ?
• Es una característica definitoria de la célula bacteriana
• Están constituídas por componentes químicos únicos.
• Algunos de estos componentes pueden causar enfermedad.
• La biosíntesis es el sitio de acción de los antibióticos más
importantes
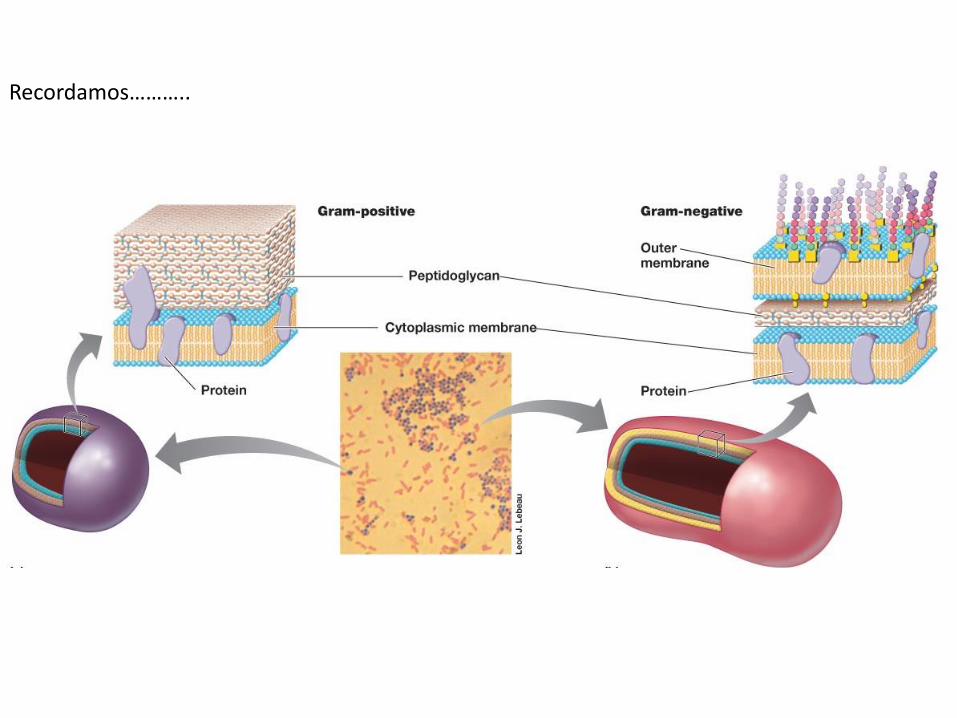
Recordamos………..
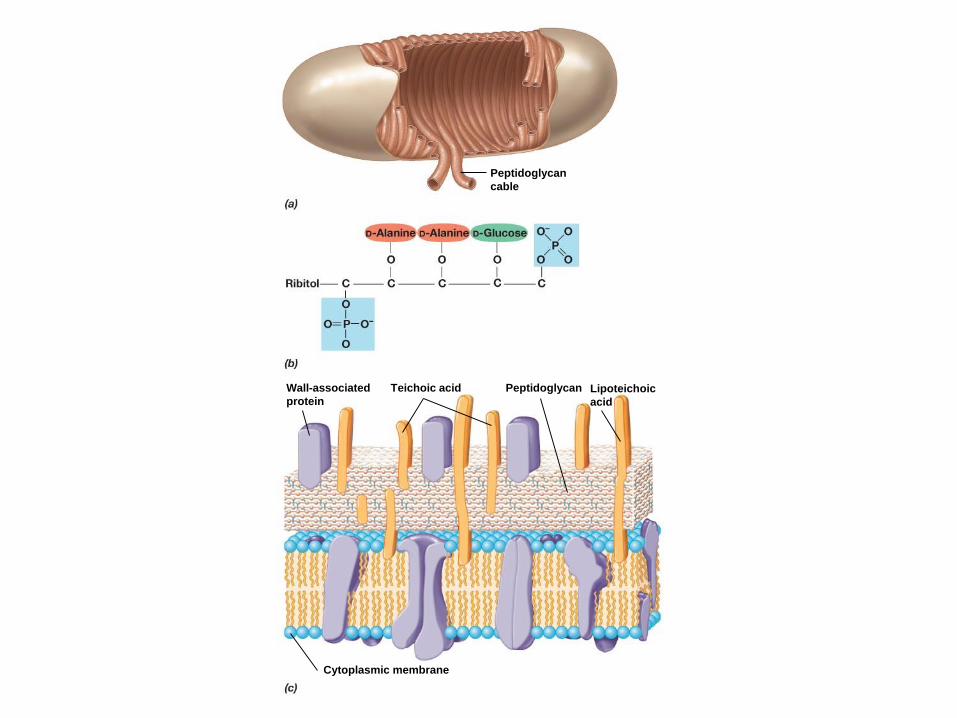
Peptidoglycan
cable
Teichoic acid Peptidoglycan Lipoteichoic
acid
Cytoplasmic membrane
Wall-associated
protein
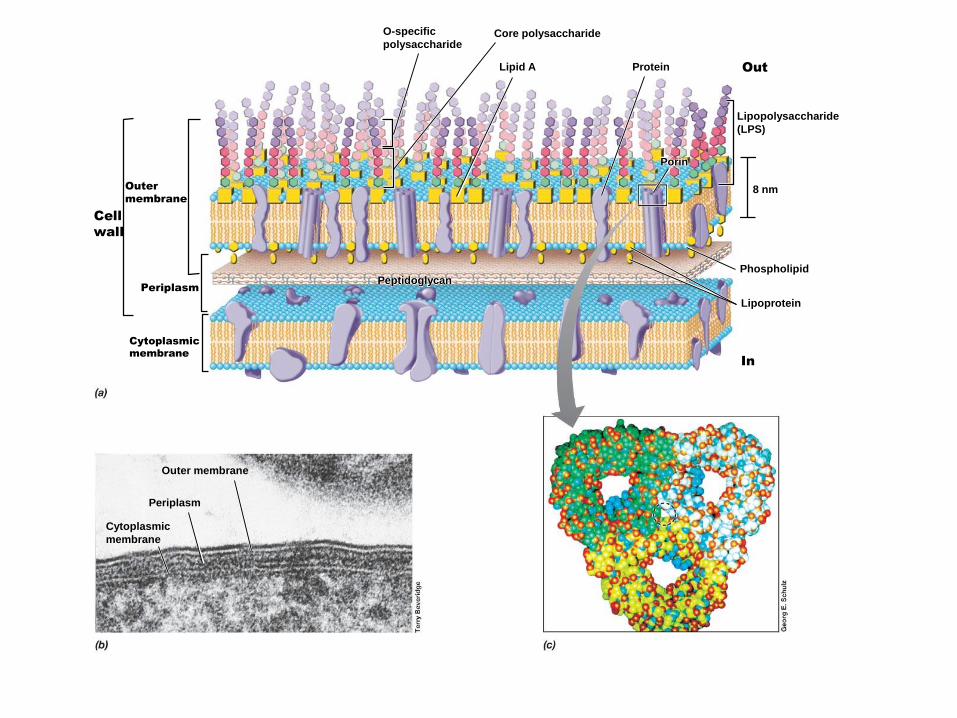
O-specific
polysaccharideCore polysaccharide
Lipid A Protein
Lipopolysaccharide
(LPS)
Phospholipid
Porin
Lipoprotein
Outer membrane
Periplasm
Cytoplasmic
membrane
8 nm
Out
Cell
wall
Outer
membrane
Periplasm
Cytoplasmic
membrane
Peptidoglycan
In
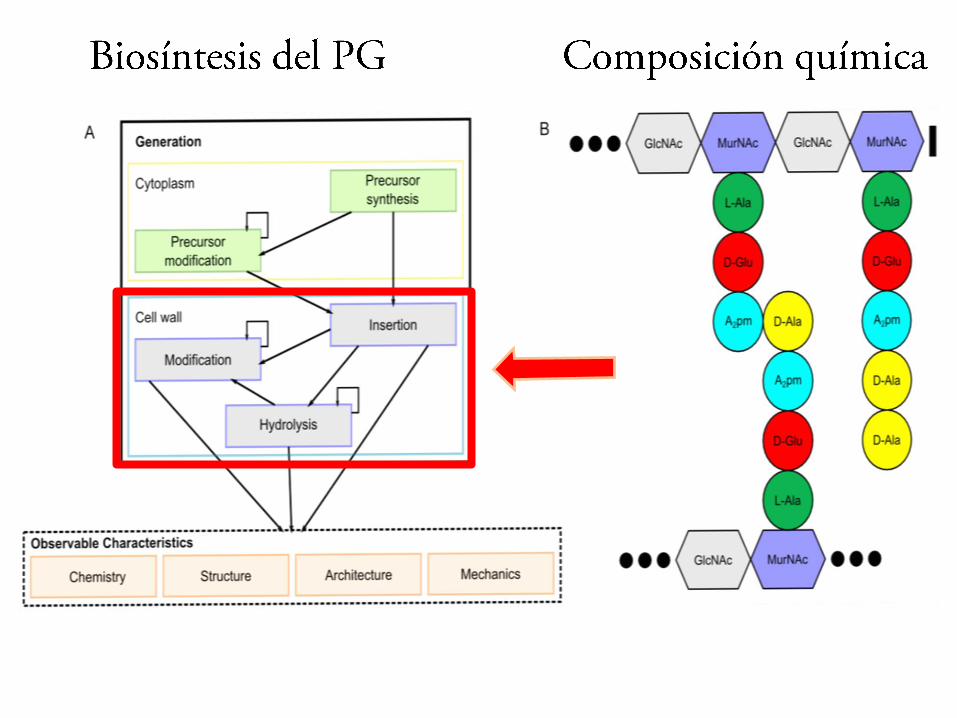

•Síntesis de precursores en el citoplasma
•Ensamblaje parcial en la membrana
•Modificaciones del precursor
•Transporte a la cara externa de la membrana??
•Ensamblaje final en el exterior??
• Reciclado del undecaprenol fosfato
Etapas de la síntesis

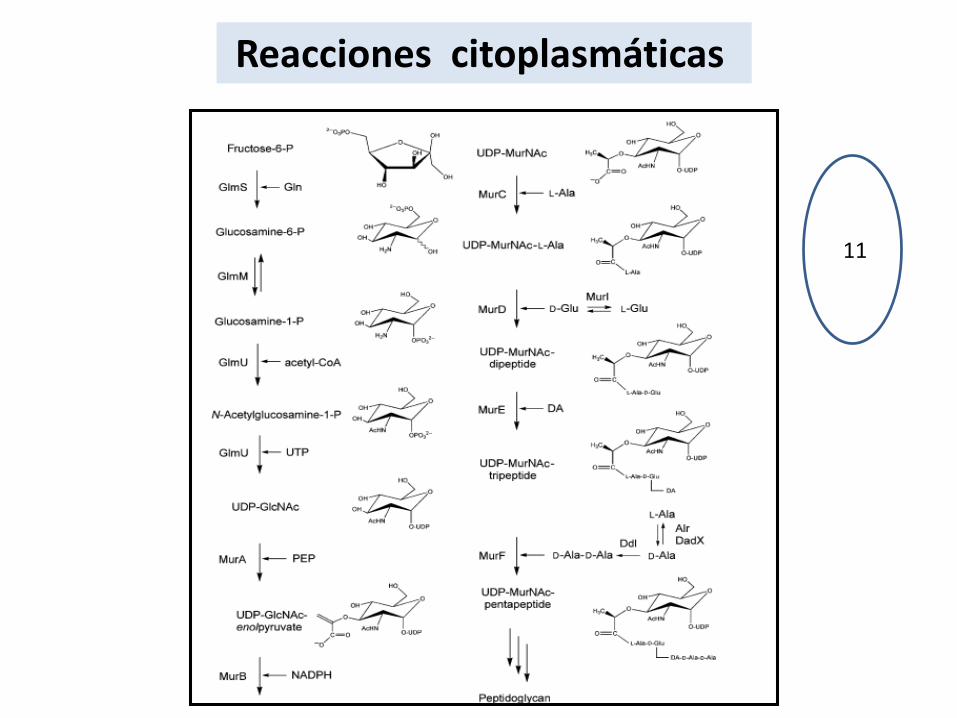
Reacciones citoplasmáticas
11
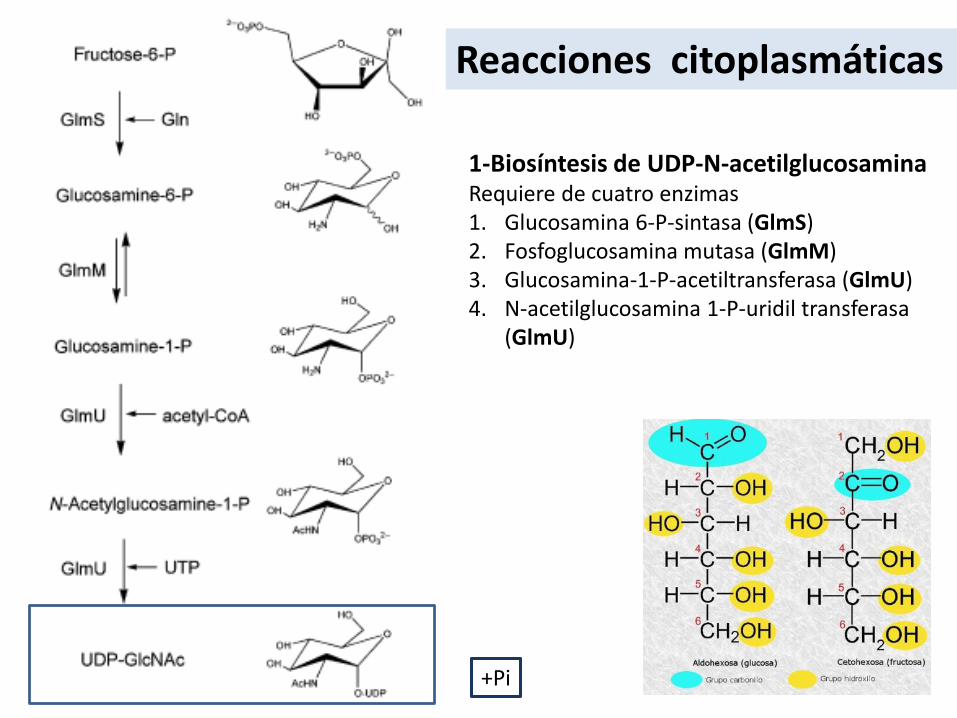
Reacciones citoplasmáticas
1-Biosíntesis de UDP-N-acetilglucosaminaRequiere de cuatro enzimas1. Glucosamina 6-P-sintasa (GlmS)2. Fosfoglucosamina mutasa (GlmM)3. Glucosamina-1-P-acetiltransferasa (GlmU)4. N-acetilglucosamina 1-P-uridil transferasa
(GlmU)
+Pi
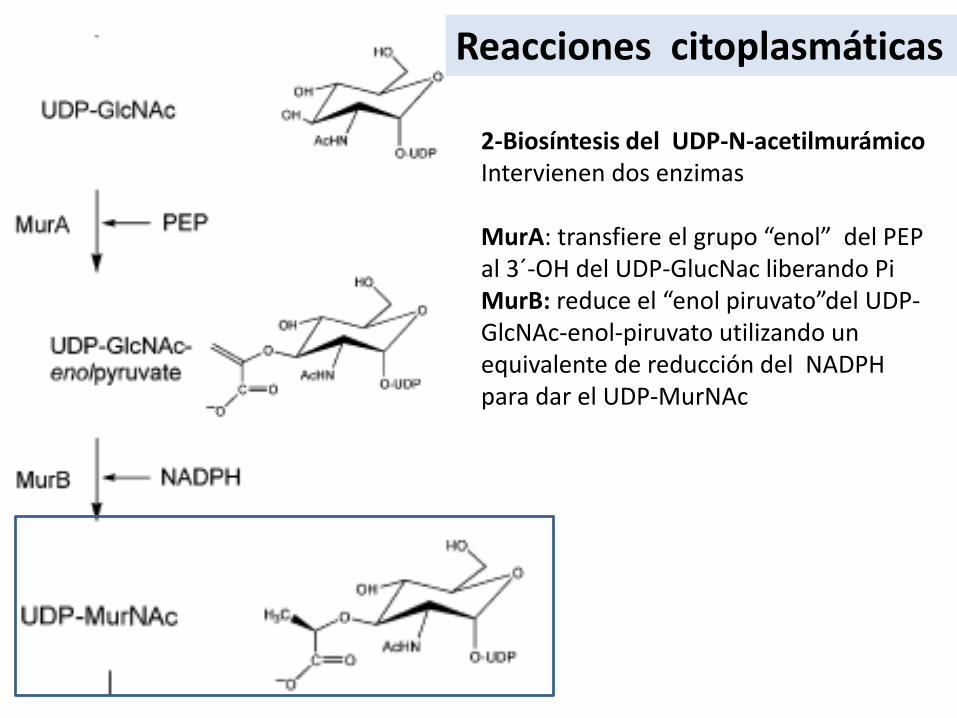
2-Biosíntesis del UDP-N-acetilmurámicoIntervienen dos enzimas
MurA: transfiere el grupo “enol” del PEP al 3´-OH del UDP-GlucNac liberando PiMurB: reduce el “enol piruvato”del UDP-GlcNAc-enol-piruvato utilizando un equivalente de reducción del NADPHpara dar el UDP-MurNAc
Reacciones citoplasmáticas
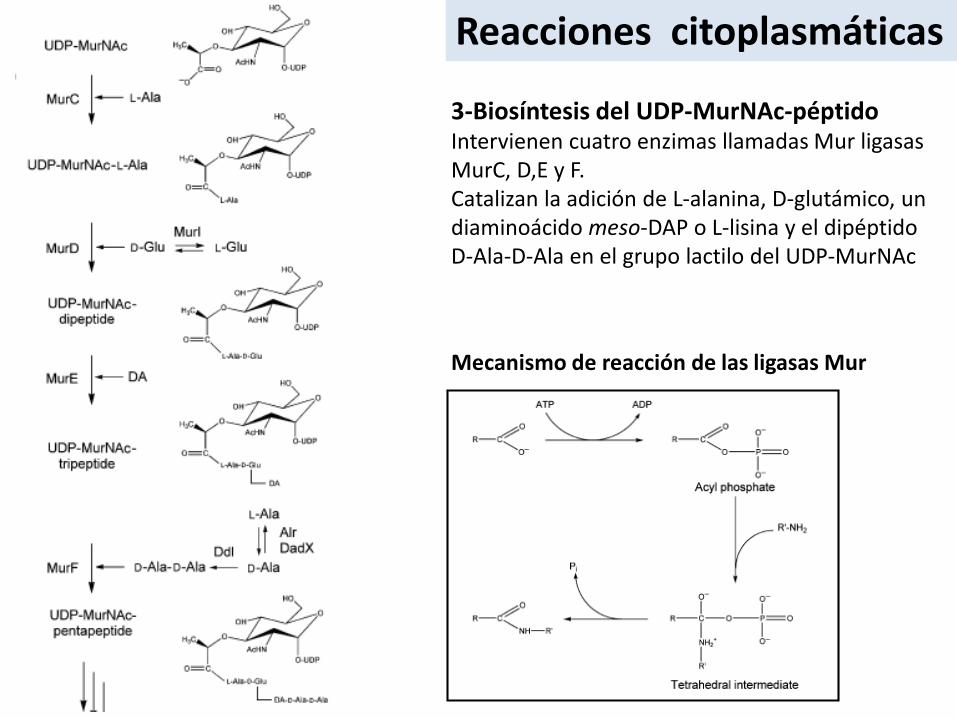
3-Biosíntesis del UDP-MurNAc-péptidoIntervienen cuatro enzimas llamadas Mur ligasasMurC, D,E y F.Catalizan la adición de L-alanina, D-glutámico, undiaminoácido meso-DAP o L-lisina y el dipéptidoD-Ala-D-Ala en el grupo lactilo del UDP-MurNAc
Mecanismo de reacción de las ligasas Mur
Reacciones citoplasmáticas
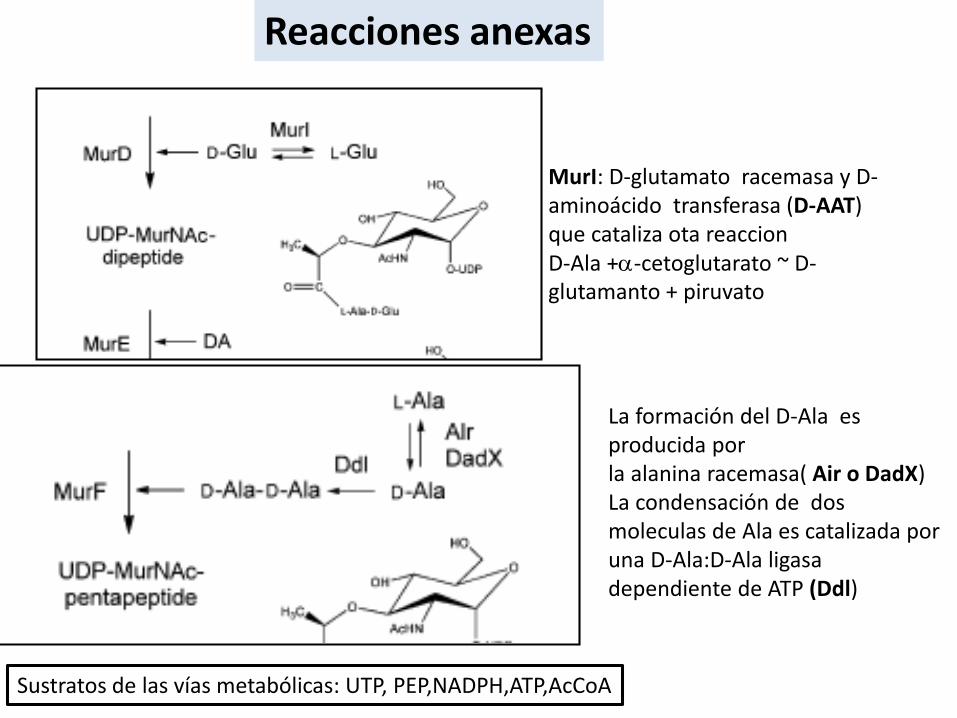
Reacciones anexas
La formación del D-Ala es producida porla alanina racemasa( Air o DadX)La condensación de dos moleculas de Ala es catalizada por una D-Ala:D-Ala ligasadependiente de ATP (Ddl)
MurI: D-glutamato racemasa y D-aminoácido transferasa (D-AAT) que cataliza ota reaccionD-Ala +a-cetoglutarato ~ D-glutamanto + piruvato
Sustratos de las vías metabólicas: UTP, PEP,NADPH,ATP,AcCoA
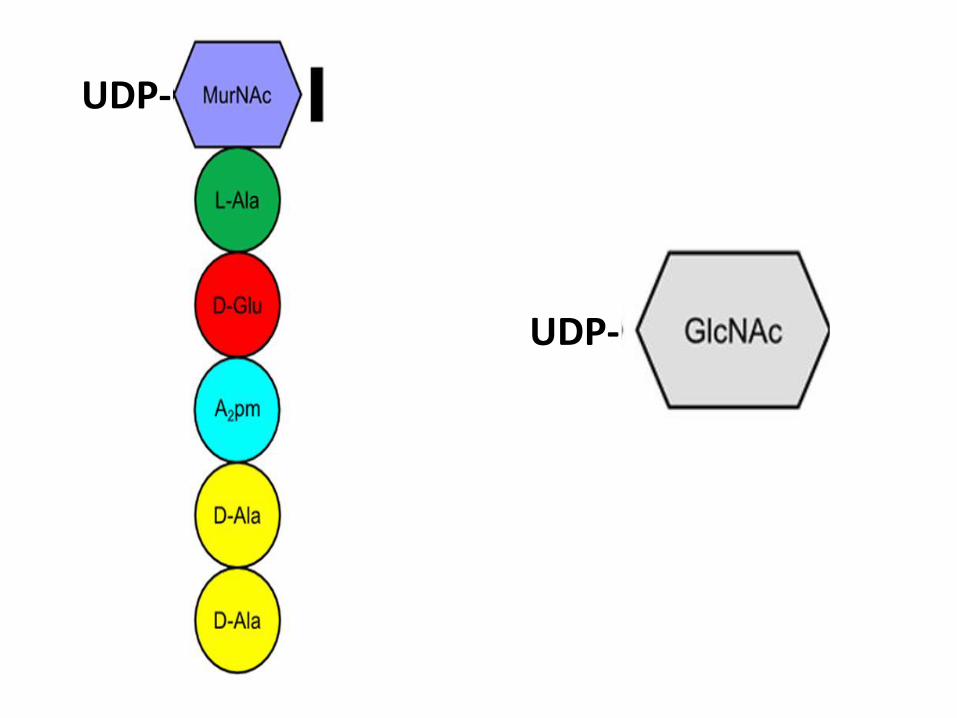
UDP-
UDP-
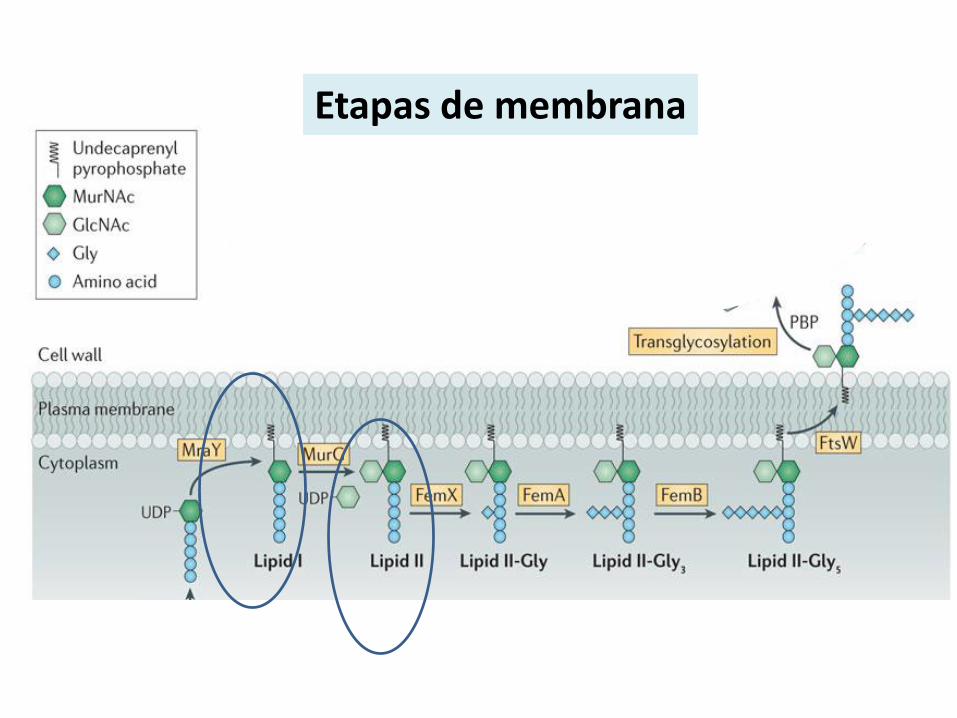
Etapas de membrana
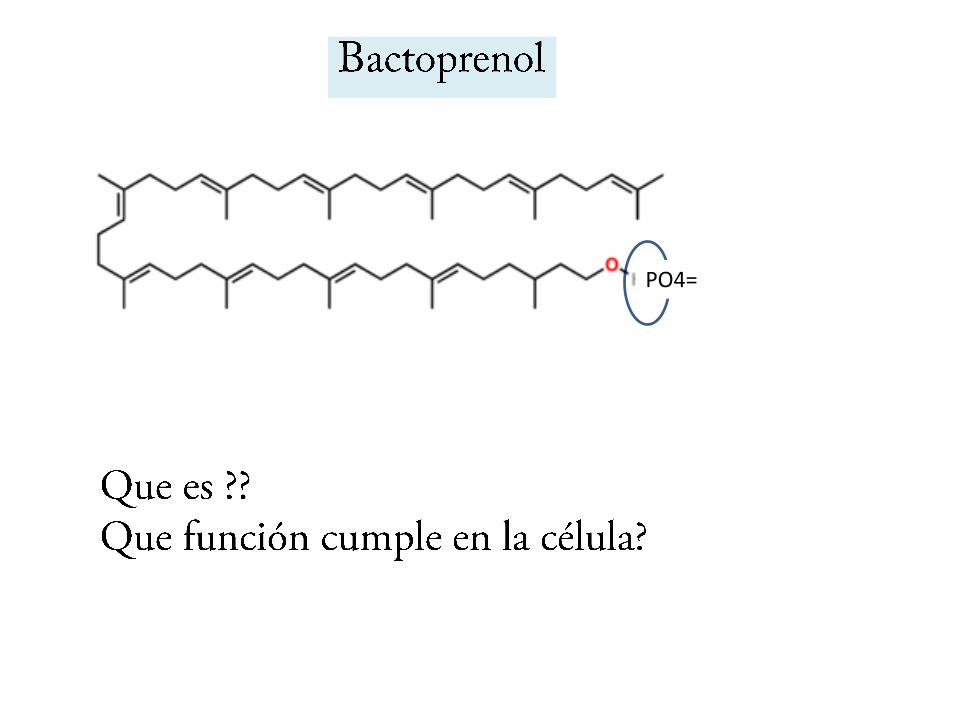
PPP
PO4=
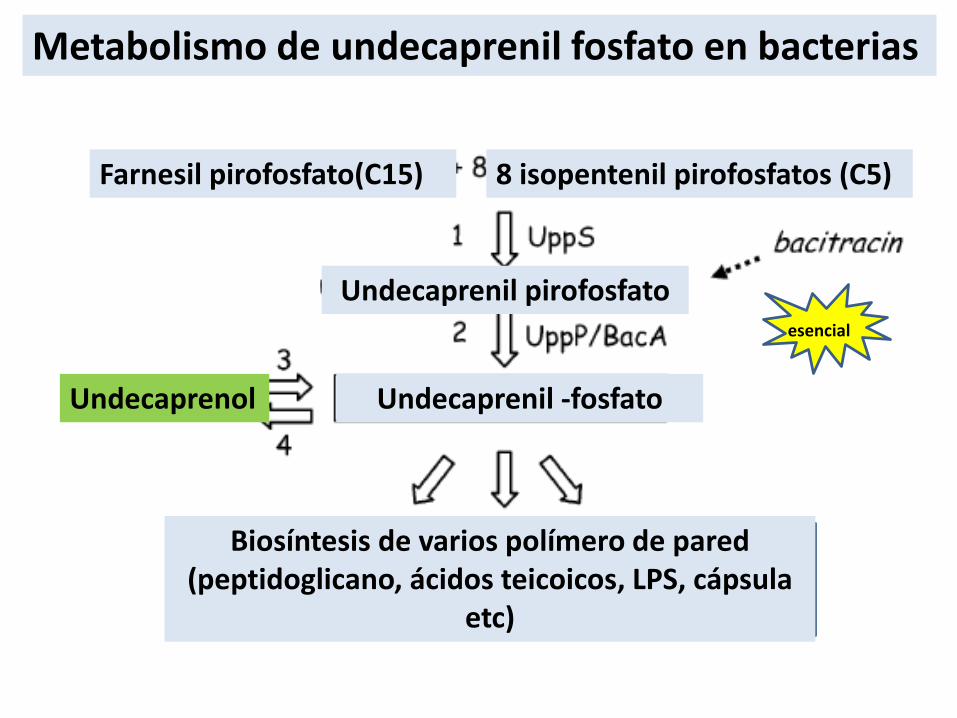
Metabolismo de undecaprenil fosfato en bacterias
Farnesil pirofosfato(C15) 8 isopentenil pirofosfatos (C5)
Undecaprenil pirofosfato
Undecaprenil -fosfatoUndecaprenol
Biosíntesis de varios polímero de pared(peptidoglicano, ácidos teicoicos, LPS, cápsula
etc)
esencial
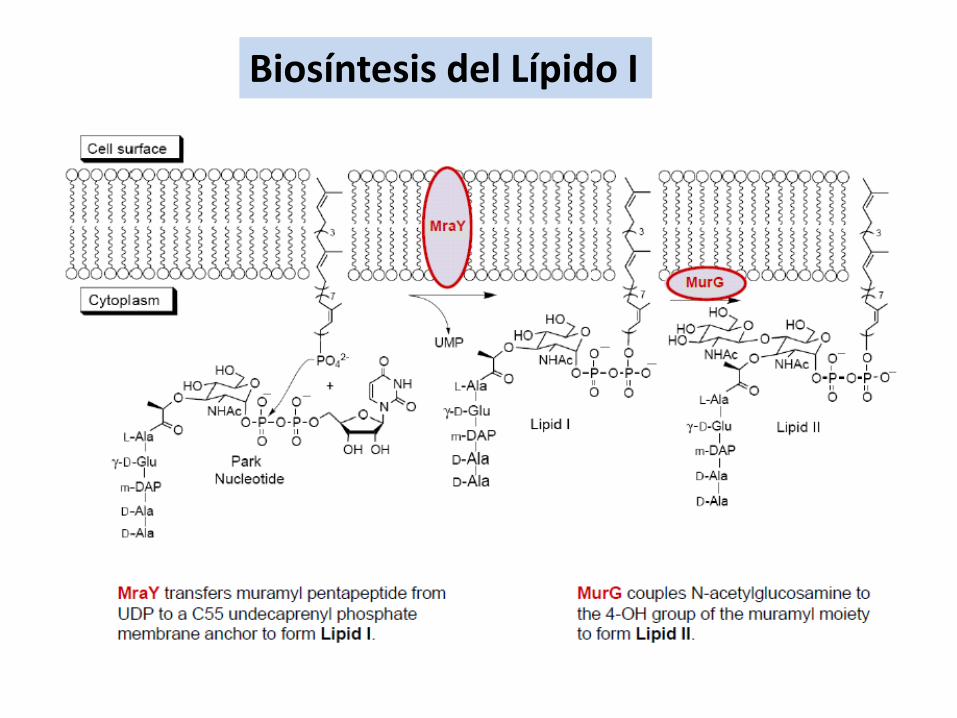
Biosíntesis del Lípido I
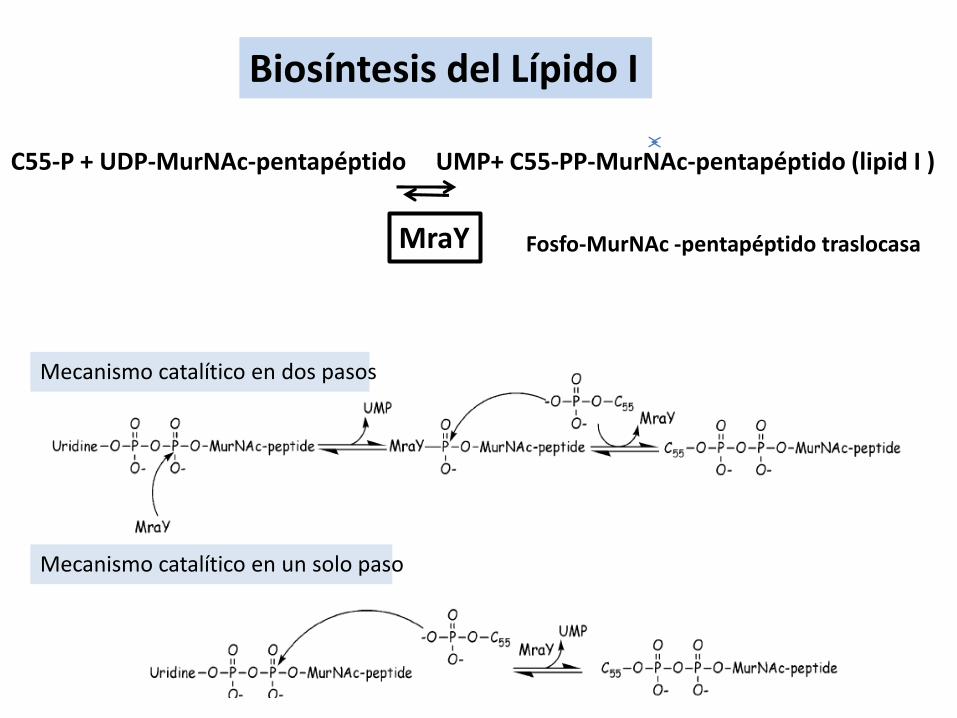
Biosíntesis del Lípido I
Mecanismo catalítico en dos pasos
Mecanismo catalítico en un solo paso
C55-P + UDP-MurNAc-pentapéptido UMP+ C55-PP-MurNAc-pentapéptido (lipid I )
MraY Fosfo-MurNAc -pentapéptido traslocasa
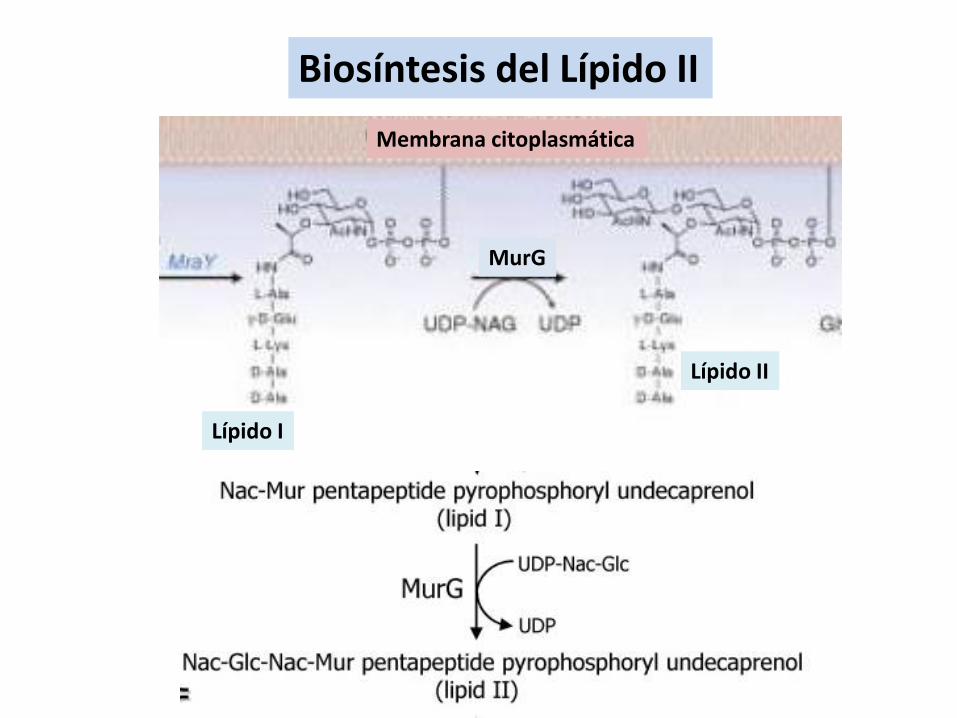
Biosíntesis del Lípido II
Lípido I
Lípido II
MurG
Membrana citoplasmática
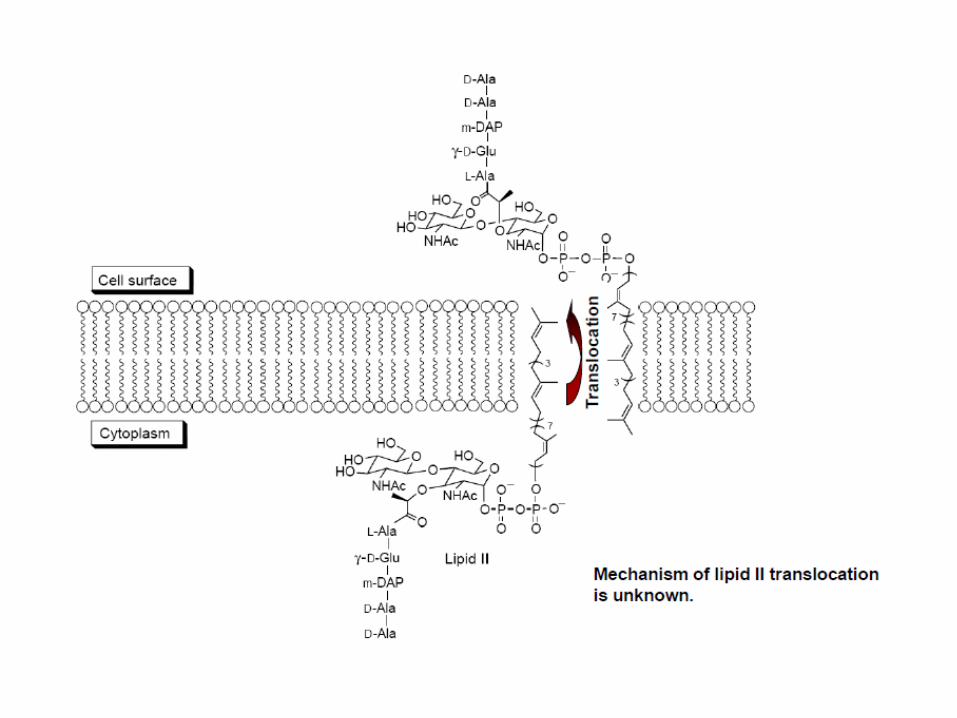
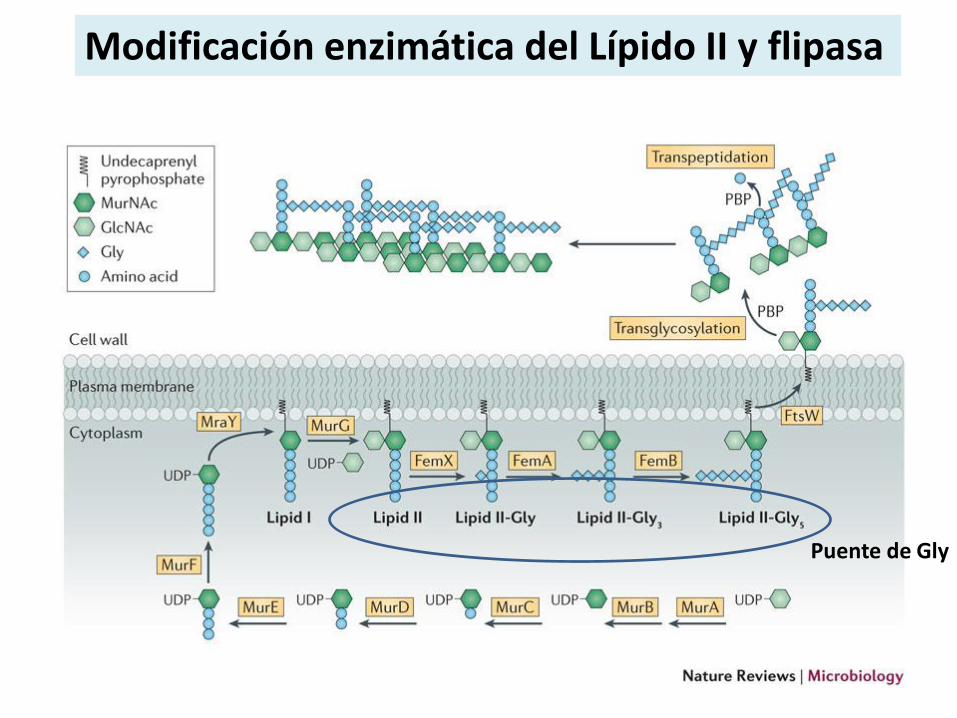
Modificación enzimática del Lípido II y flipasa
Puente de Gly
Peptidoglycan Transglycosylase
activity
Autolysin
activity
Cytoplasmic
membrane
Out
Bactoprenol
Pentapeptide
Growing point
of cell wall
In
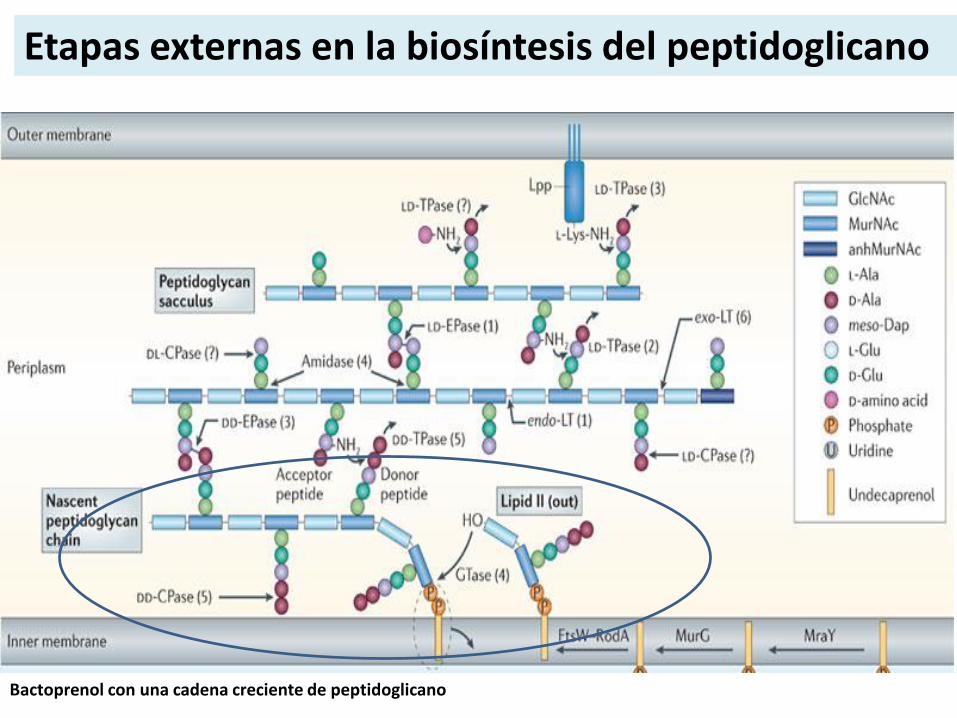
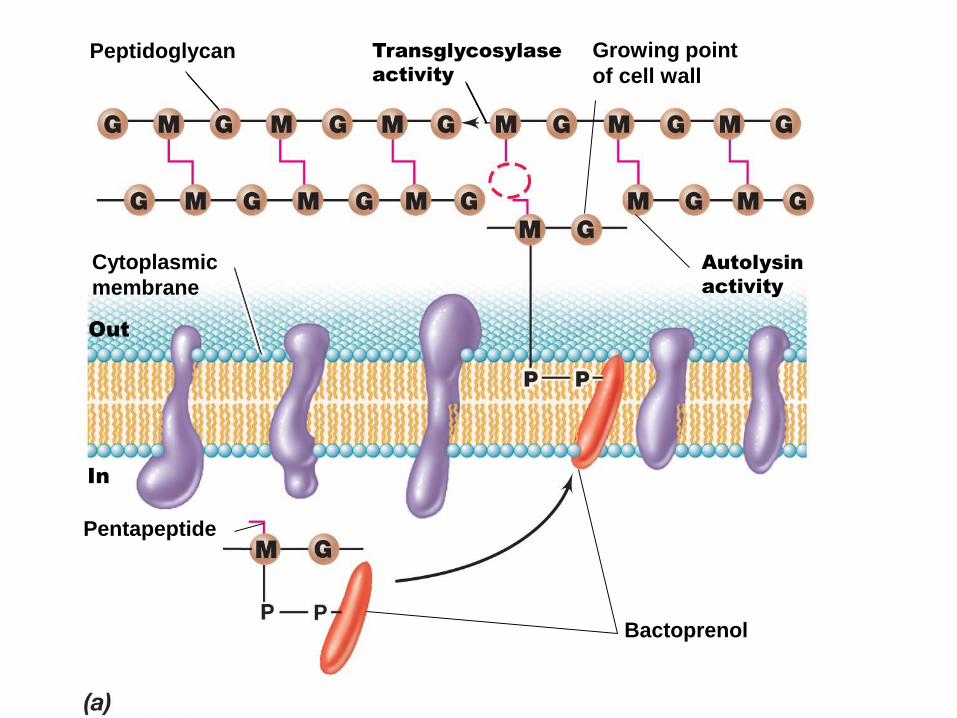
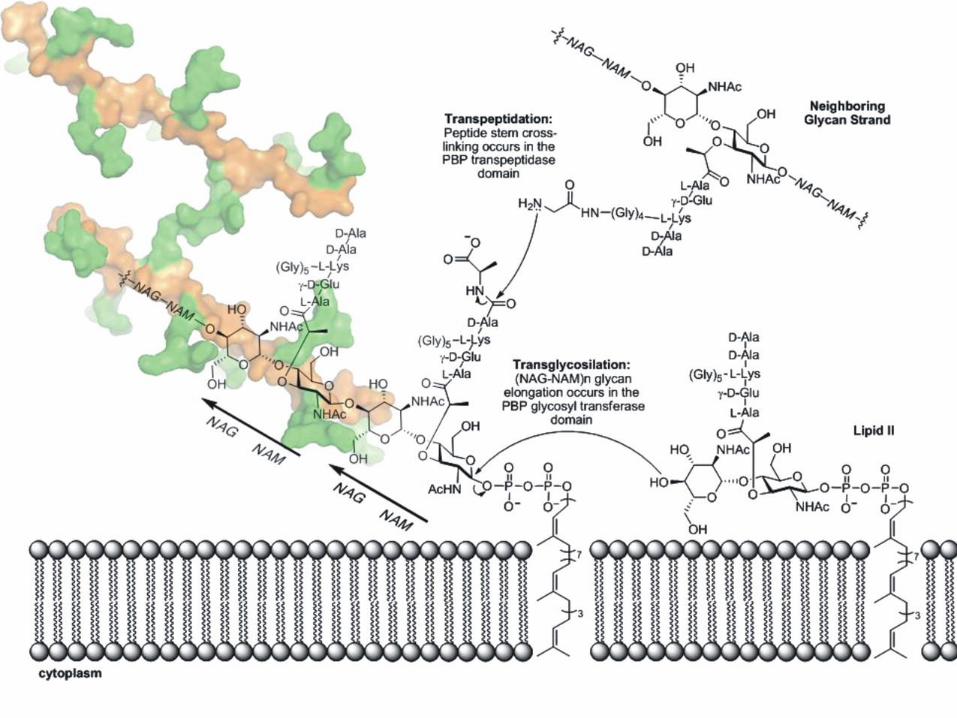
Etapas externas en la biosíntesis del peptidoglicano
Bactoprenol con una cadena creciente de peptidoglicano

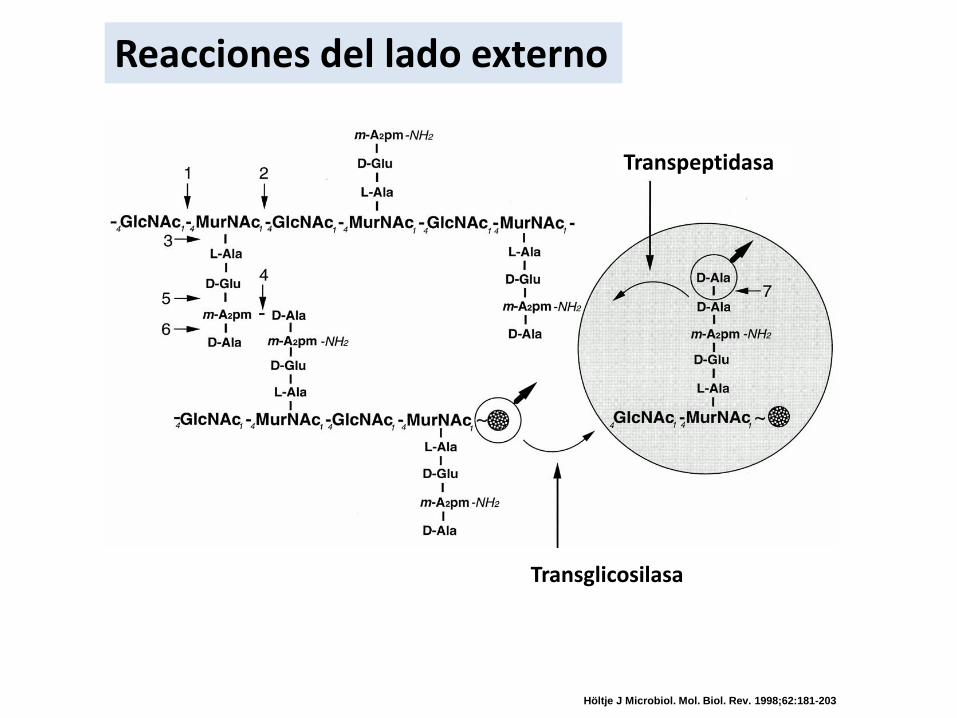
Höltje J Microbiol. Mol. Biol. Rev. 1998;62:181-203
Reacciones del lado externo
Transglicosilasa
Transpeptidasa
Scheffers D , and Pinho M G Microbiol. Mol. Biol. Rev. 2005;69:585-607
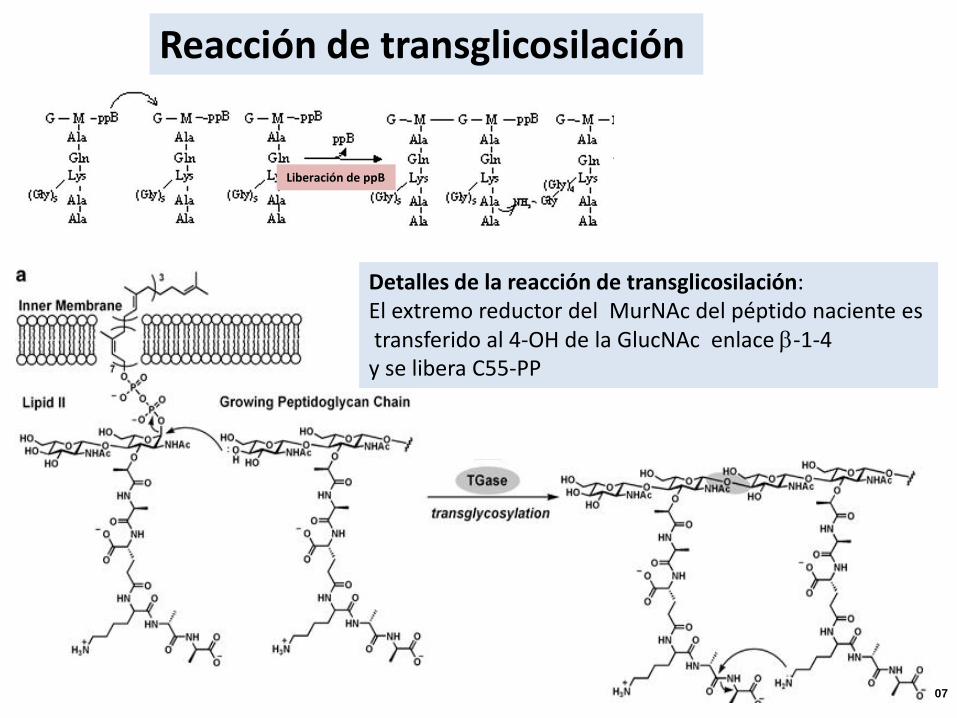
Reacción de transglicosilación
Liberación de ppB
Detalles de la reacción de transglicosilación:El extremo reductor del MurNAc del péptido naciente estransferido al 4-OH de la GlucNAc enlace b-1-4
y se libera C55-PP
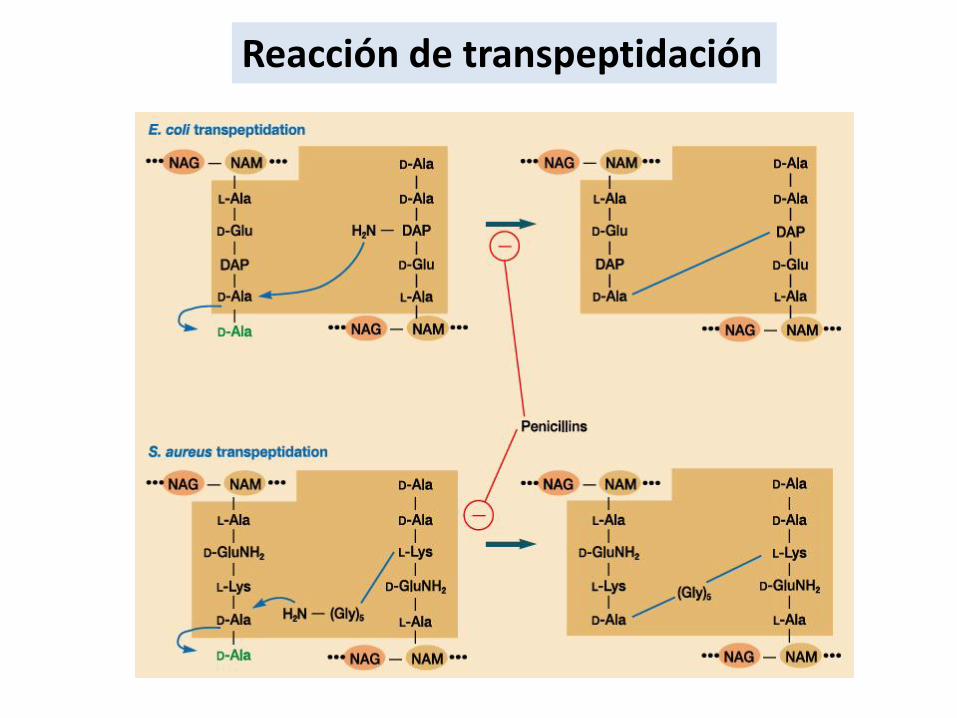
Reacción de transpeptidación
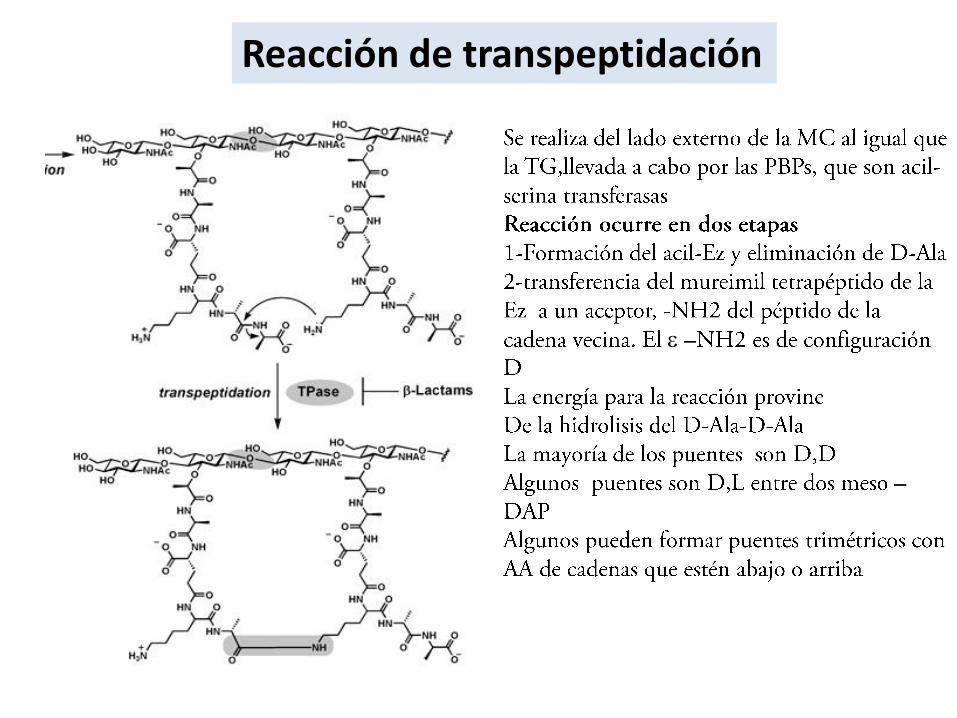
Reacción de transpeptidación
e
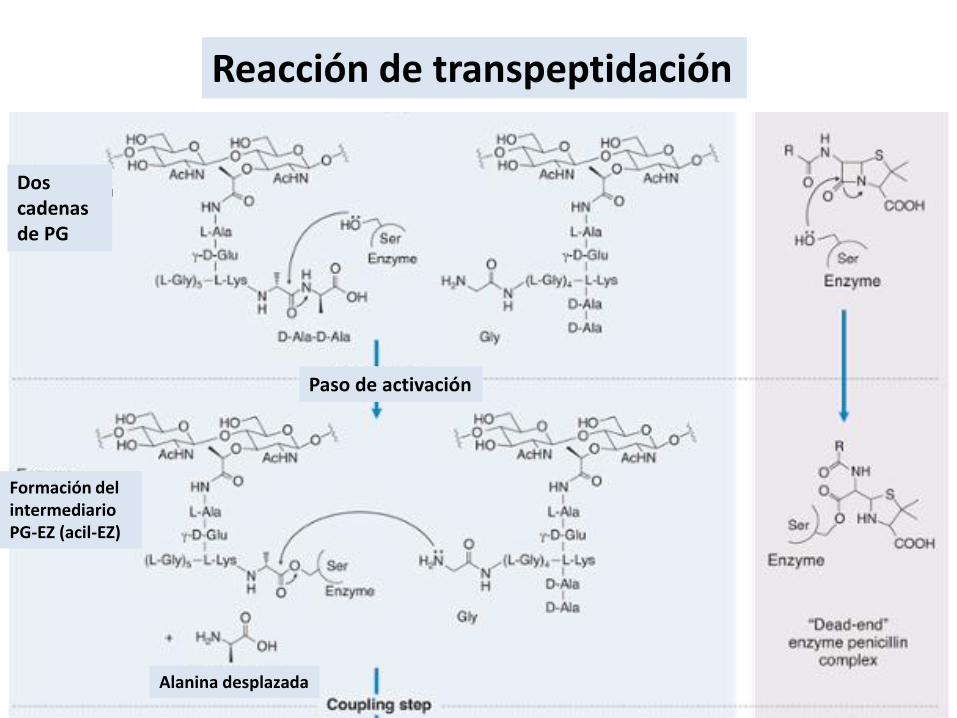
Reacción de transpeptidación
Formación del intermediarioPG-EZ (acil-EZ)
Dos cadenas de PG
Paso de activación
Alanina desplazada
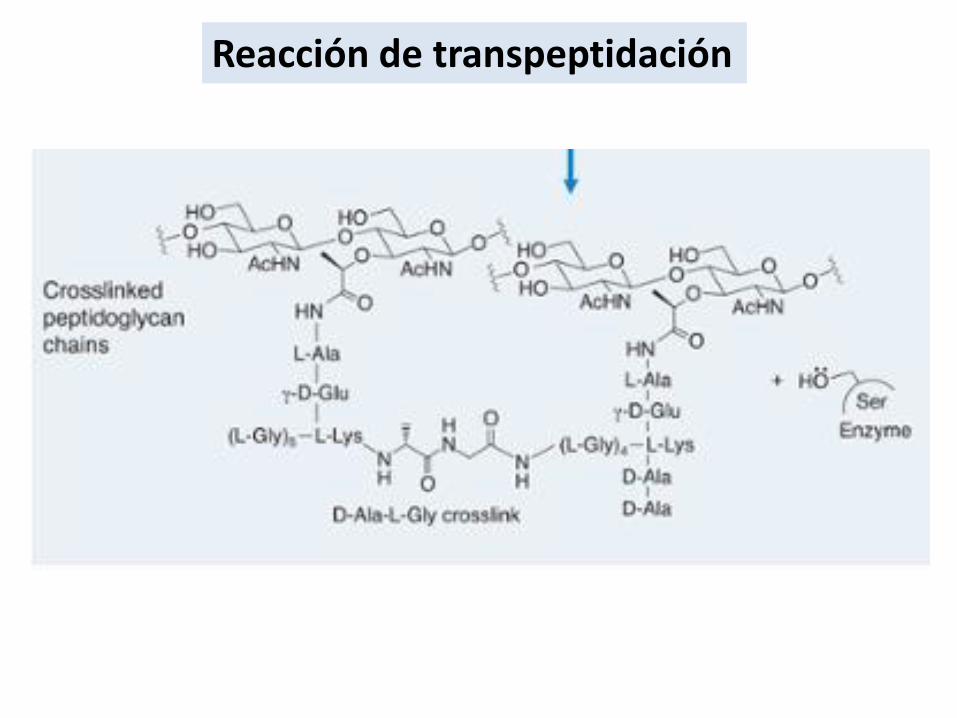
Reacción de transpeptidación
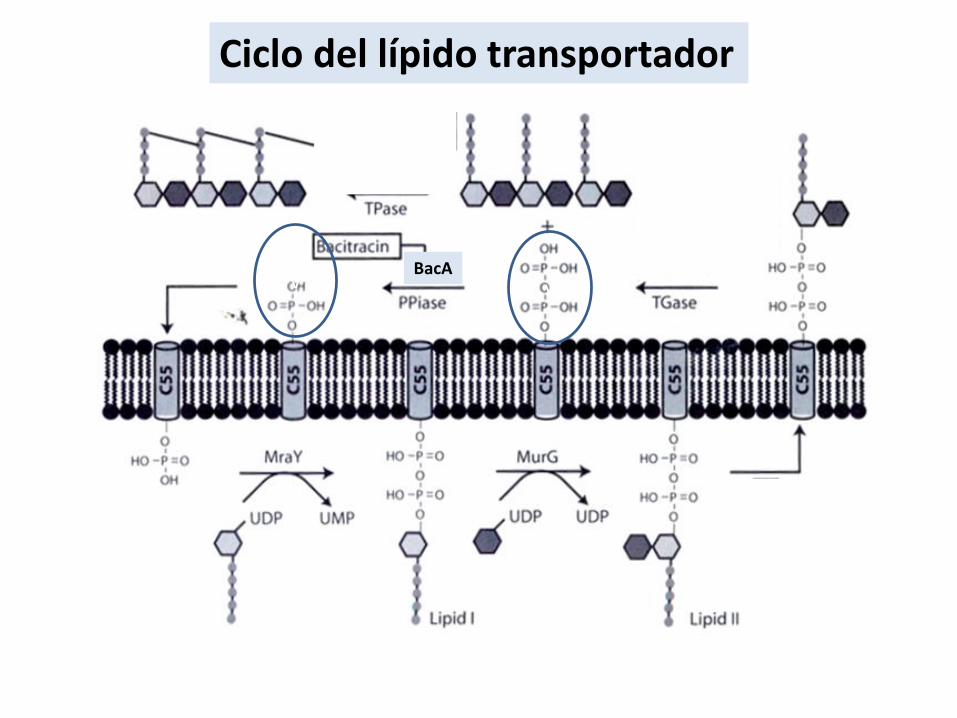
Reciclado del bactoprenol
Ciclo del lípido transportador
vvBacA

Hidrolasas del peptidoglicano
Tienen roles relevantes en el crecimiento del PG, división celular , espesor del PG y forma de la bacteria.
Las enzimas encargadas son llamadas autolisinas: rompen enlaces glicosídicosy amida.
En E. coli algunas son•D,D-Carboxipeptidasas (PBP 5, PBP4B, PBP6, PBP6B)•D,D-Endopeptidasas (PBP 4 y PBP 7)•Amidasas•LT Transglicosilasas líticas
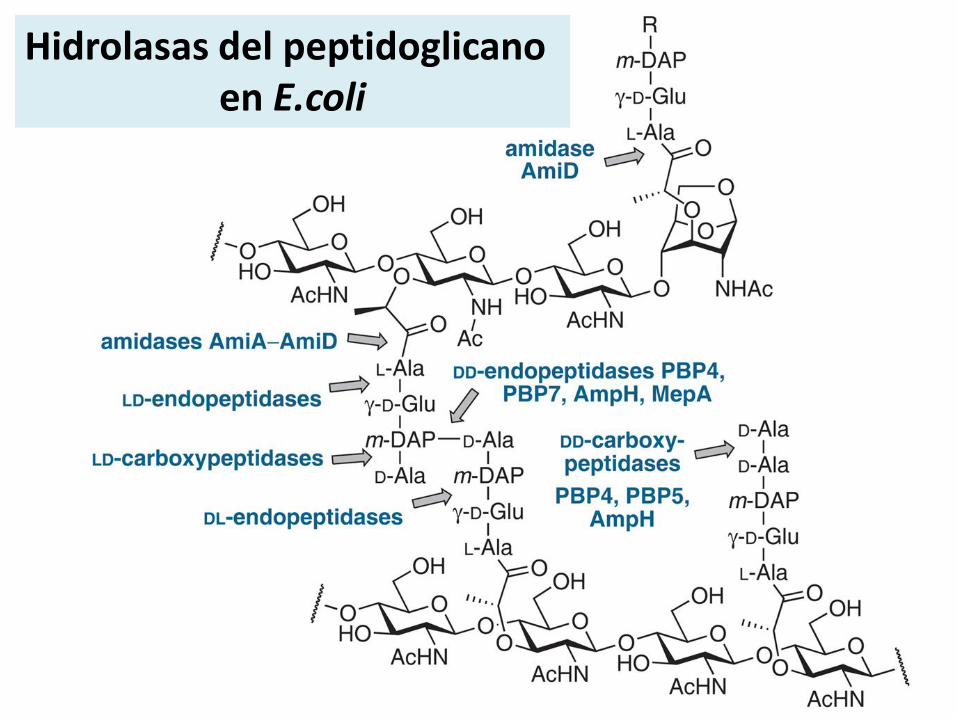
Hidrolasas del peptidoglicanoen E.coli
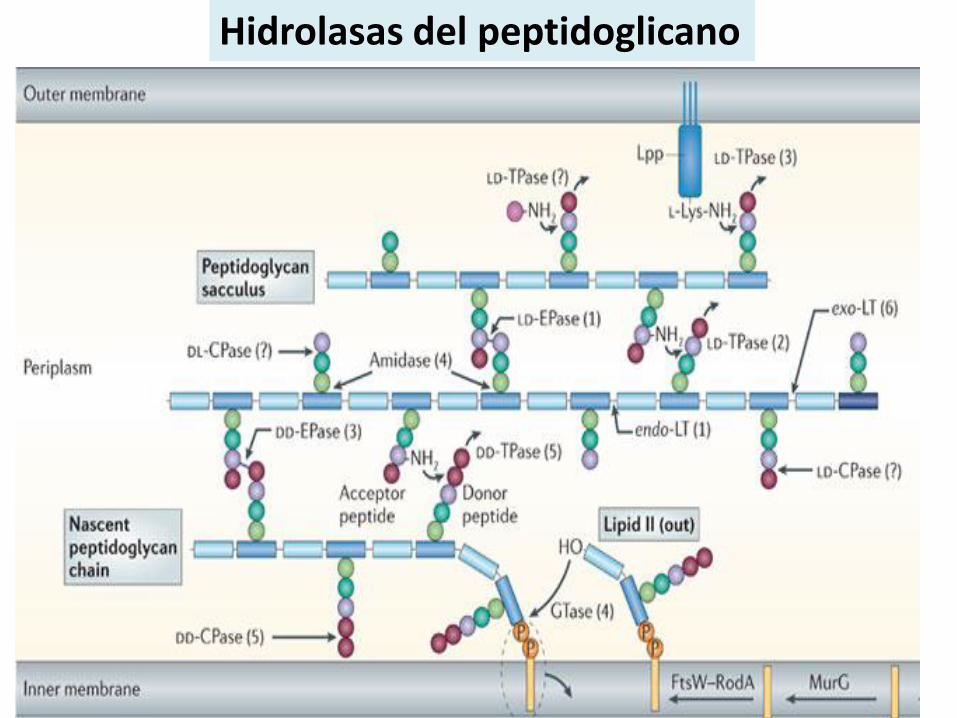
Hidrolasas del peptidoglicano
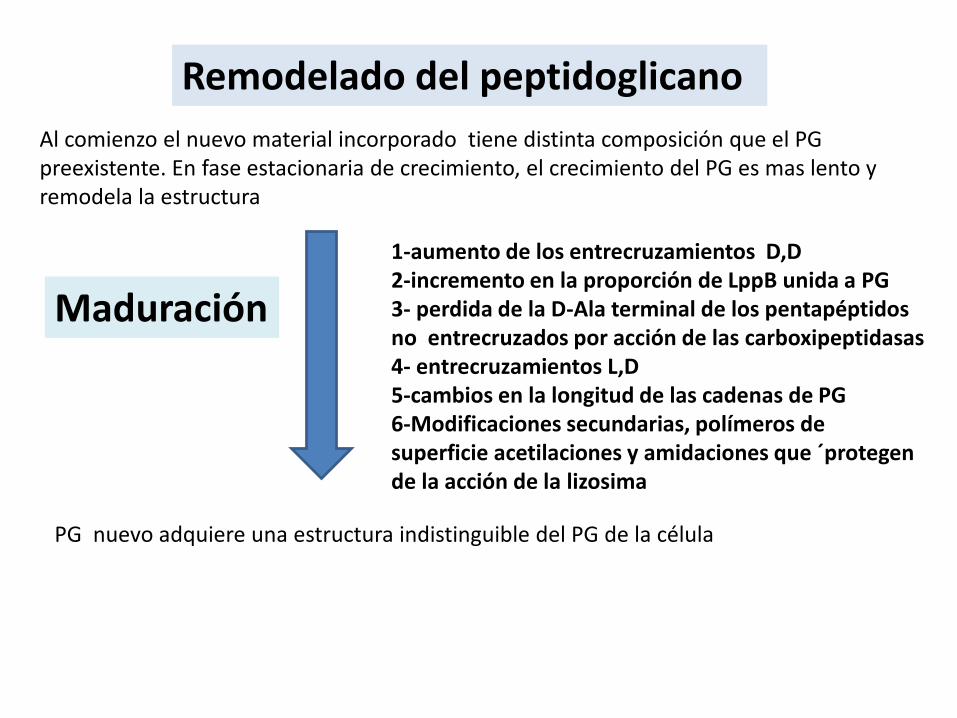
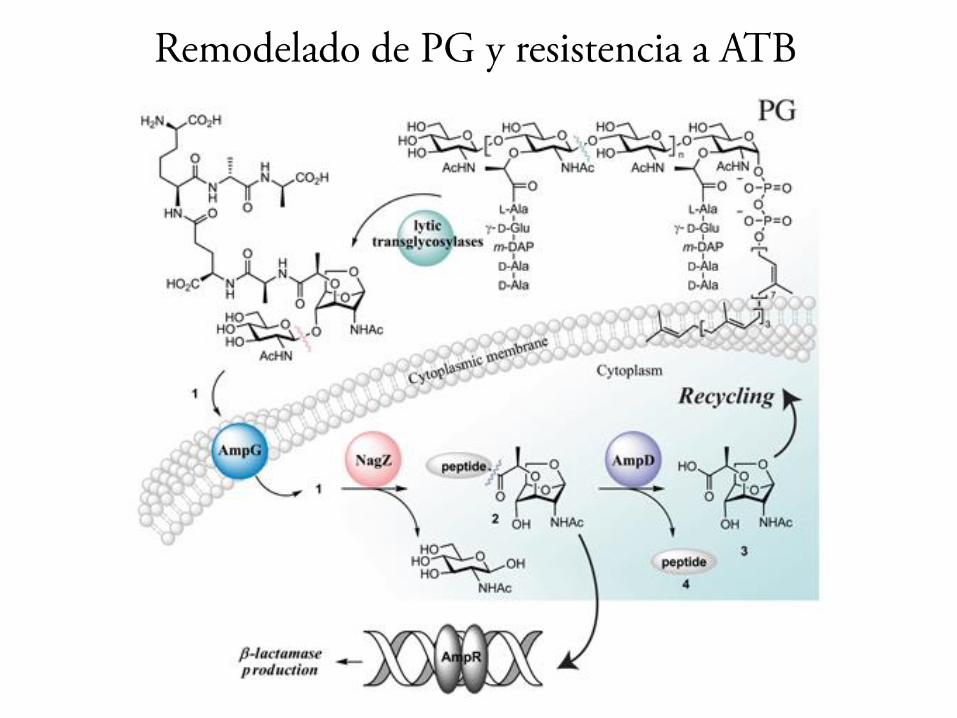
Remodelado del peptidoglicano
1-aumento de los entrecruzamientos D,D 2-incremento en la proporción de LppB unida a PG3- perdida de la D-Ala terminal de los pentapéptidosno entrecruzados por acción de las carboxipeptidasas4- entrecruzamientos L,D 5-cambios en la longitud de las cadenas de PG6-Modificaciones secundarias, polímeros de superficie acetilaciones y amidaciones que ´protegen de la acción de la lizosima
PG nuevo adquiere una estructura indistinguible del PG de la célula
Al comienzo el nuevo material incorporado tiene distinta composición que el PG preexistente. En fase estacionaria de crecimiento, el crecimiento del PG es mas lento y remodela la estructura
Maduración
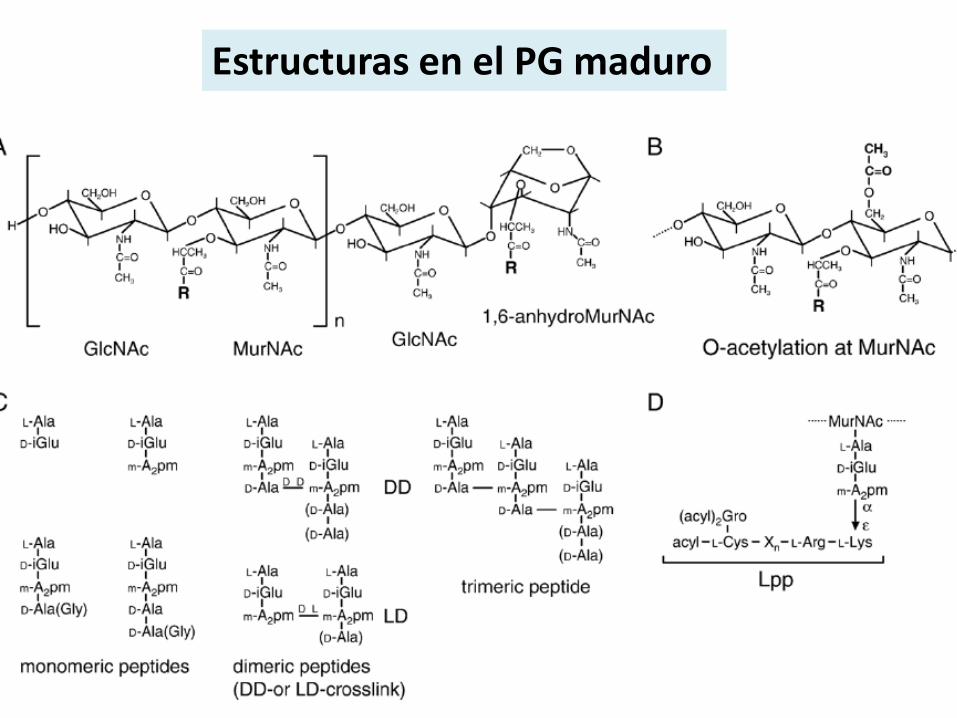
Estructuras en el PG maduro
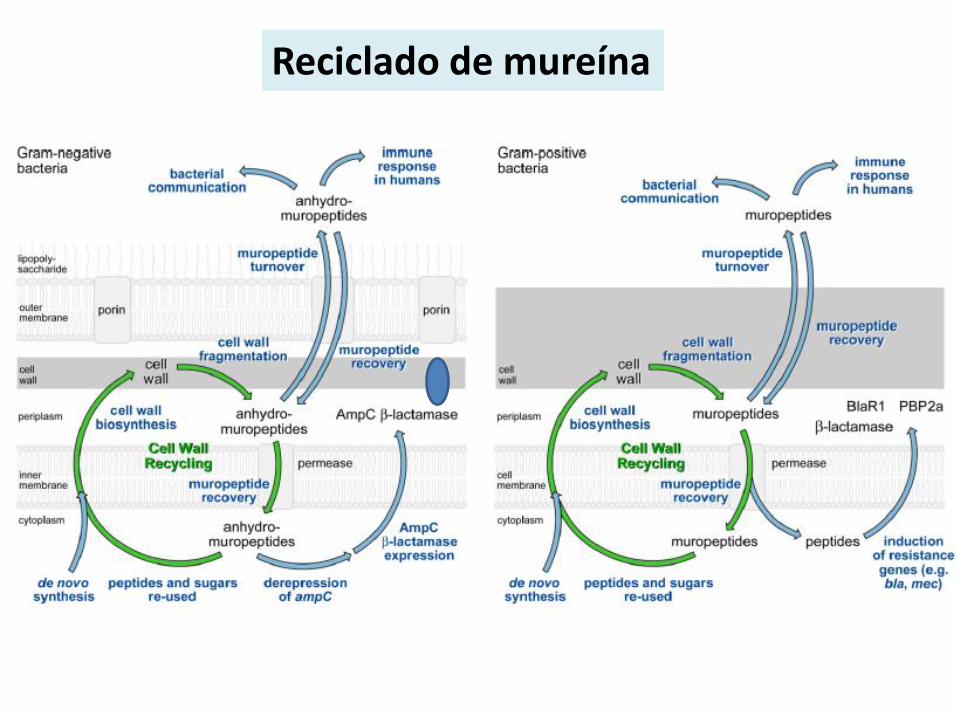
Reciclado de mureína
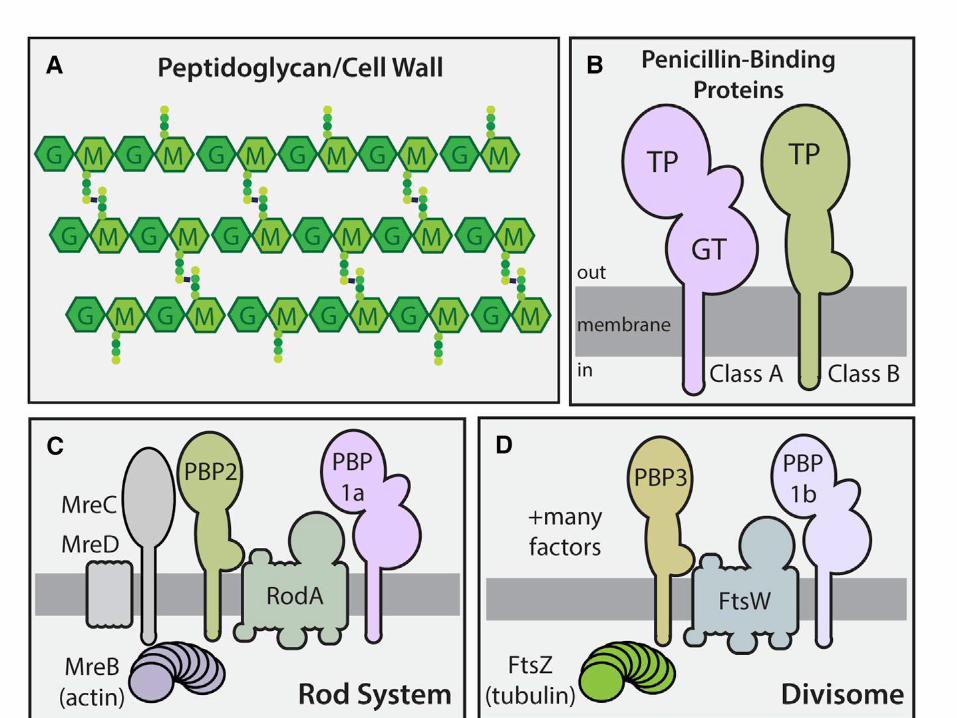
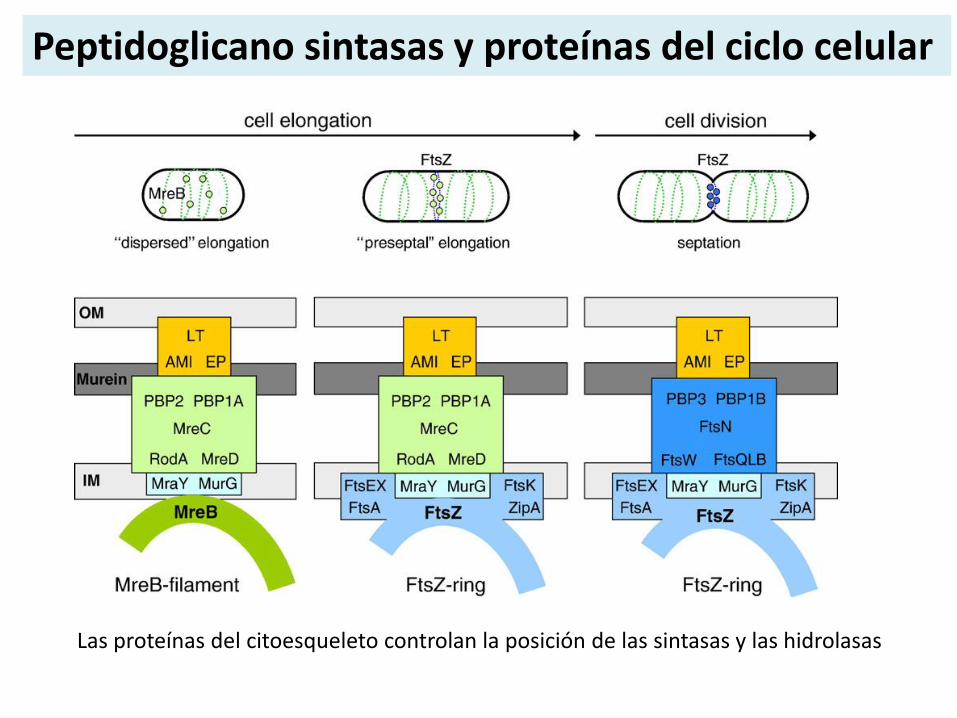
Peptidoglicano sintasas y proteínas del ciclo celular
Las proteínas del citoesqueleto controlan la posición de las sintasas y las hidrolasas
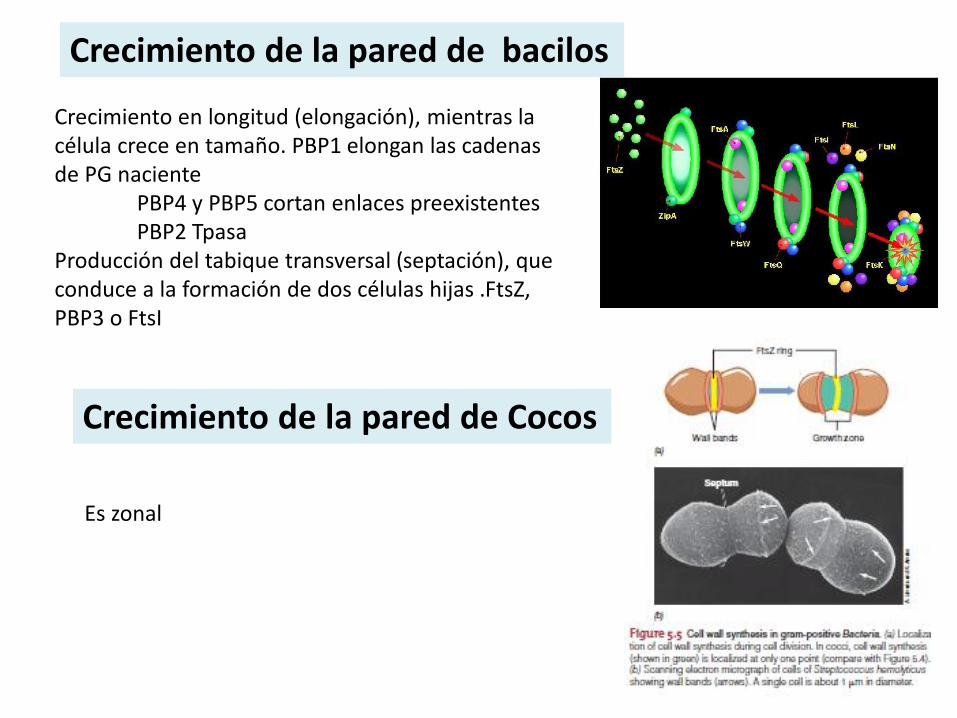
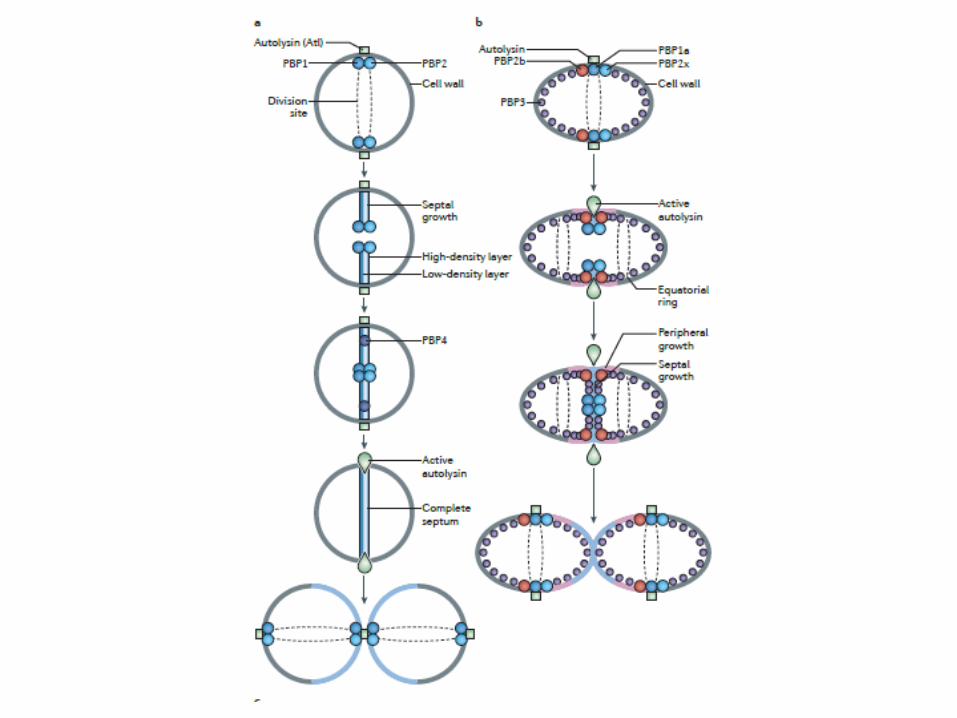
Crecimiento de la pared de bacilos
Crecimiento en longitud (elongación), mientras la célula crece en tamaño. PBP1 elongan las cadenas de PG naciente
PBP4 y PBP5 cortan enlaces preexistentesPBP2 Tpasa
Producción del tabique transversal (septación), que conduce a la formación de dos células hijas .FtsZ, PBP3 o FtsI
Crecimiento de la pared de Cocos
Es zonal

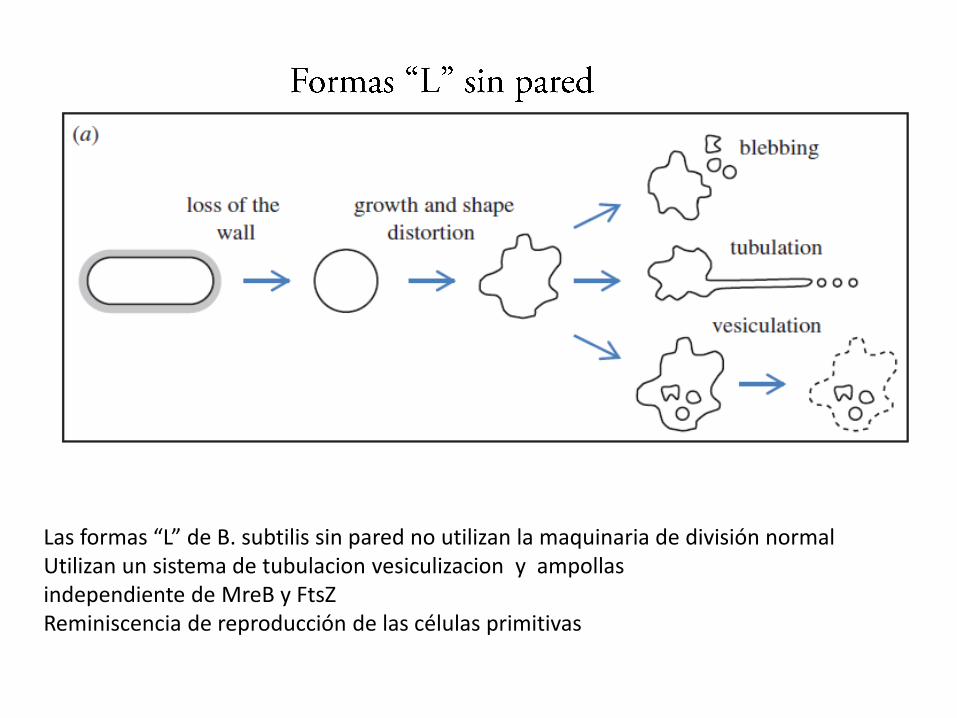
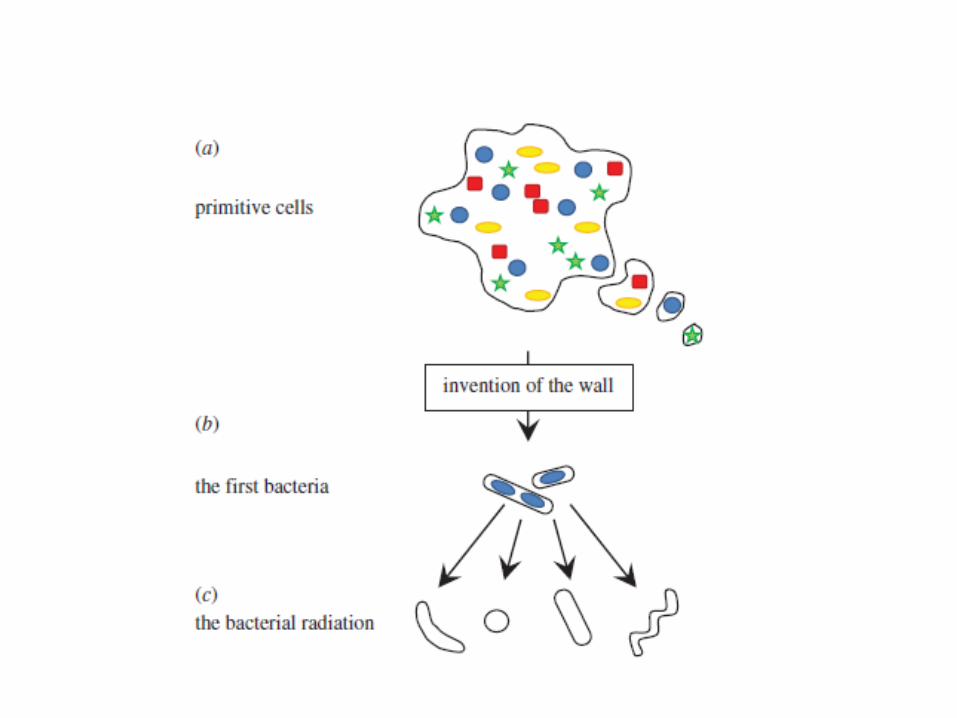
Las formas “L” de B. subtilis sin pared no utilizan la maquinaria de división normalUtilizan un sistema de tubulacion vesiculizacion y ampollasindependiente de MreB y FtsZReminiscencia de reproducción de las células primitivas
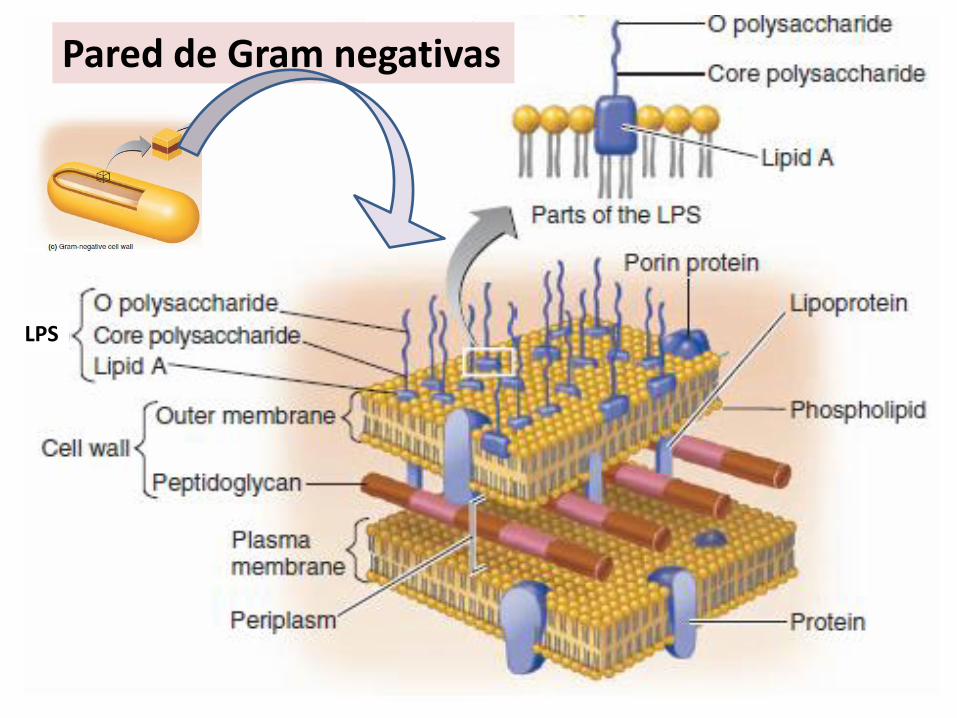
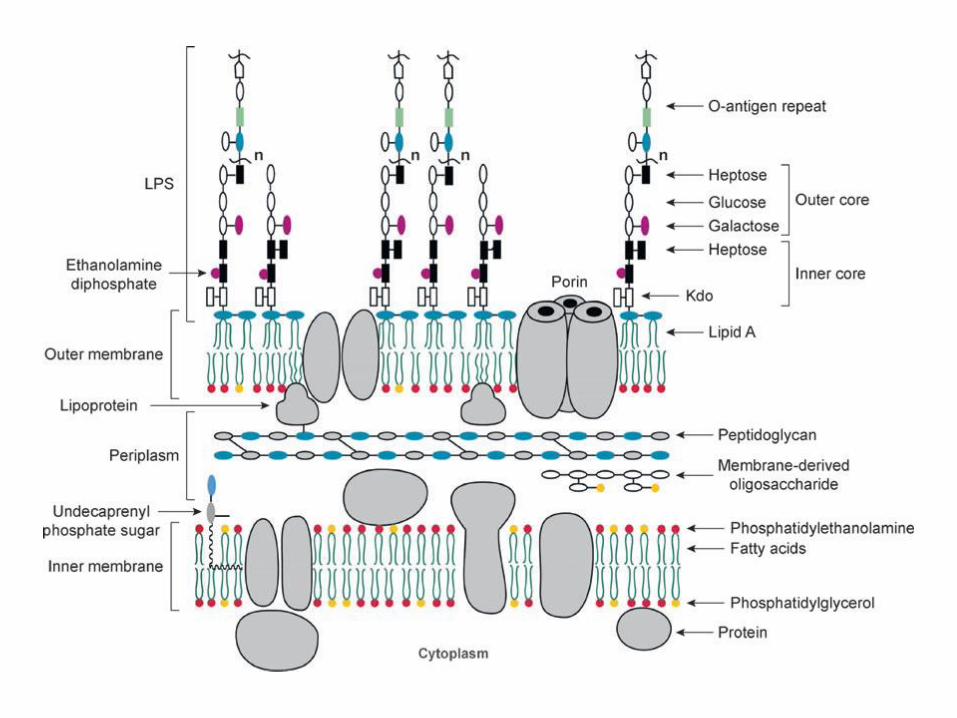
LPS
Pared de Gram negativas
Glucosamina-
ß(16)-
glucosamina, con
–OH en 1
sustituido con –P-
etanolamina
A.G. saturados
(C-14): beta-
hidroximirístico
Núcleo internoNúcleo externoUnidad repetitiva de
la cadena lateral
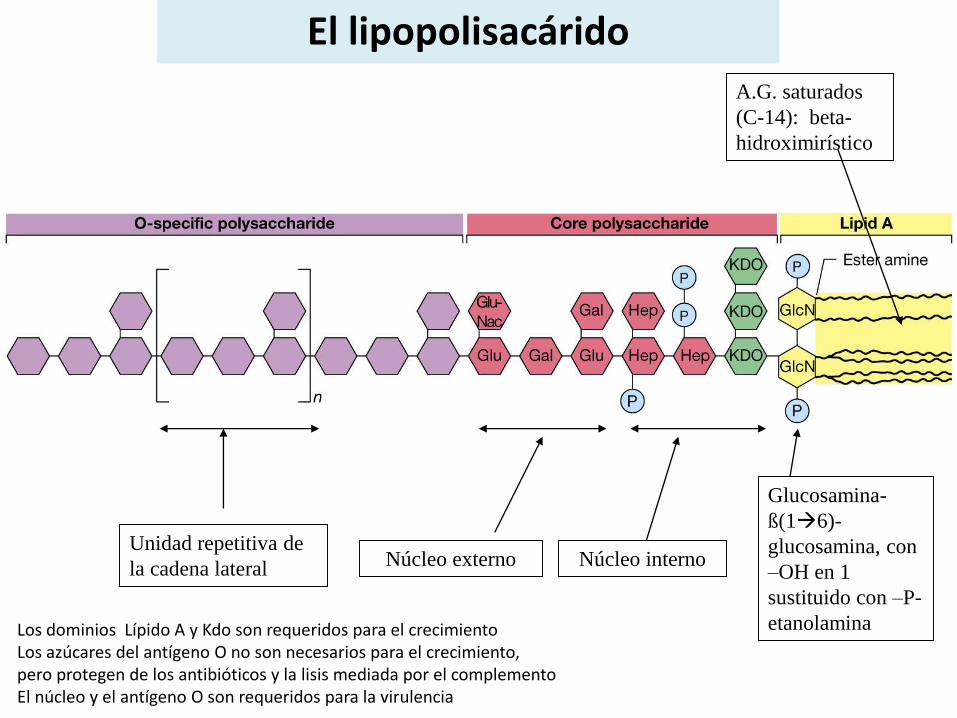
El lipopolisacárido
Los dominios Lípido A y Kdo son requeridos para el crecimientoLos azúcares del antígeno O no son necesarios para el crecimiento,pero protegen de los antibióticos y la lisis mediada por el complementoEl núcleo y el antígeno O son requeridos para la virulencia

Composición del lipopolisacárido (LPS)
Lípido A: endotoxina, estructura conservada
Región intermedia: núcleo del polisacárido, estructura conservada, se divide en NI y NE
Región distal o antígeno O: cadena lateral específica, polisacarídica, son repeticiones depocos azúcares, muy variable.

La síntesis del LPS se divide en dos procesos claramente diferenciados:
• la formación del lípido A y del núcleo del LPS, y
•la síntesis del antígeno O.
Una vez sintetizados estos dos componentes tiene lugar la unión de los mismos, su modificación y transporte hacia la membrana externa
Biosíntesis del LPS
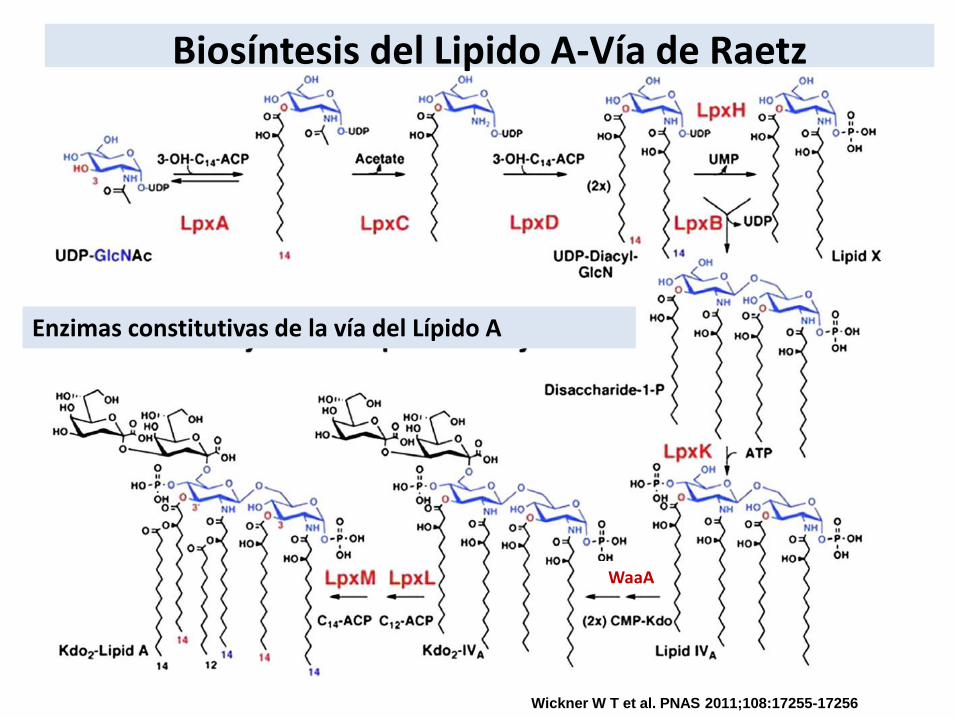
Biosíntesis del Lipido A-Vía de Raetz
Wickner W T et al. PNAS 2011;108:17255-17256
Enzimas constitutivas de la vía del Lípido A
WaaA
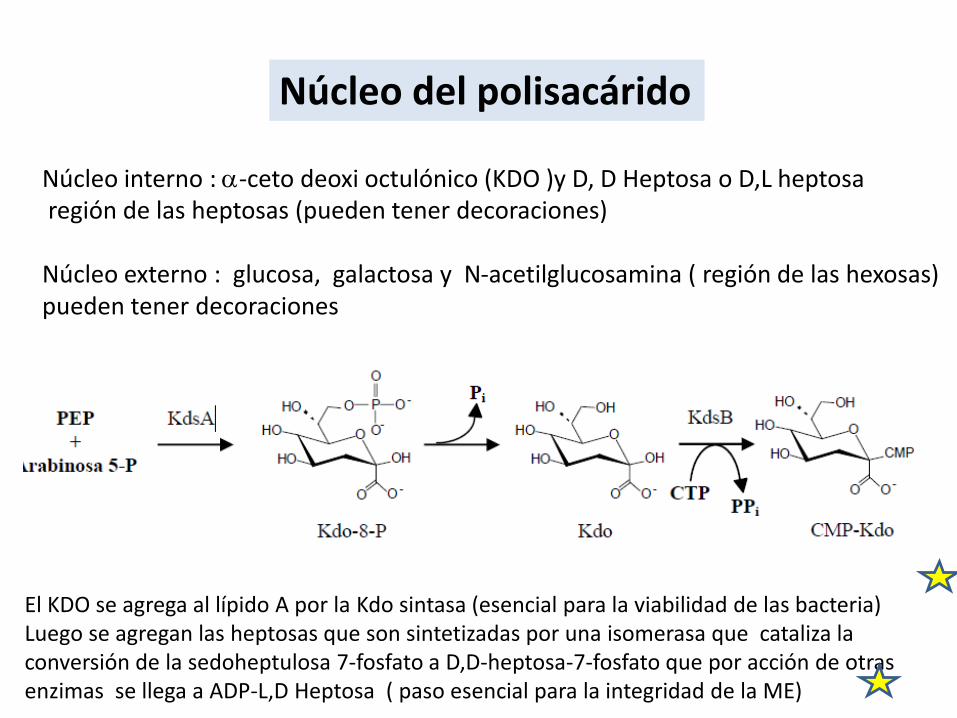
Núcleo del polisacárido
Núcleo interno : a-ceto deoxi octulónico (KDO )y D, D Heptosa o D,L heptosaregión de las heptosas (pueden tener decoraciones)
Núcleo externo : glucosa, galactosa y N-acetilglucosamina ( región de las hexosas)pueden tener decoraciones
El KDO se agrega al lípido A por la Kdo sintasa (esencial para la viabilidad de las bacteria)Luego se agregan las heptosas que son sintetizadas por una isomerasa que cataliza laconversión de la sedoheptulosa 7-fosfato a D,D-heptosa-7-fosfato que por acción de otras enzimas se llega a ADP-L,D Heptosa ( paso esencial para la integridad de la ME)
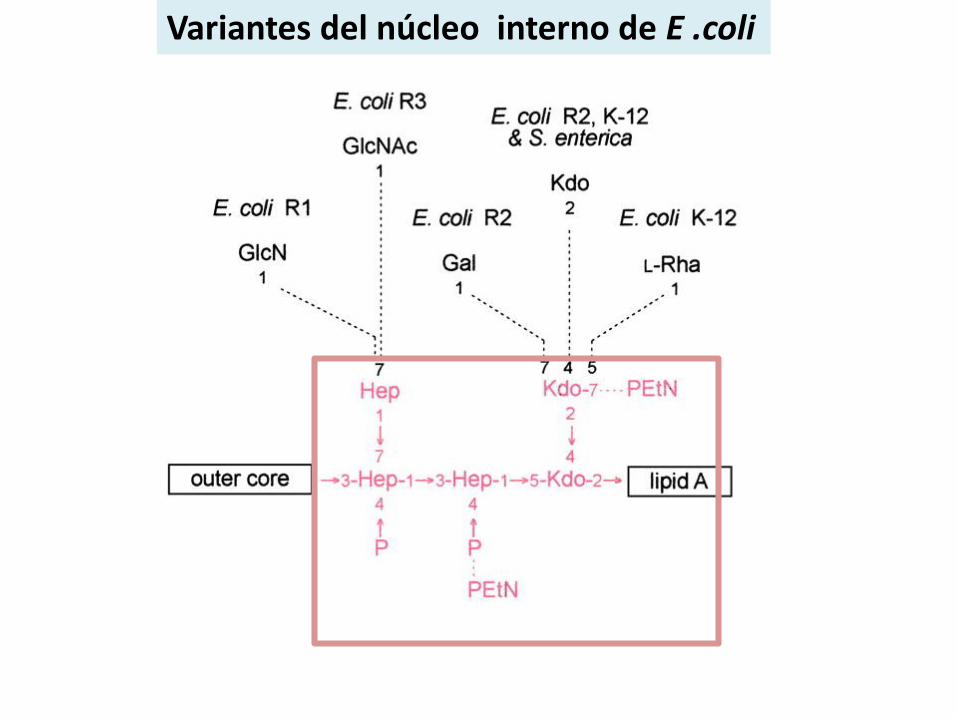
Variantes del núcleo interno de E .coli
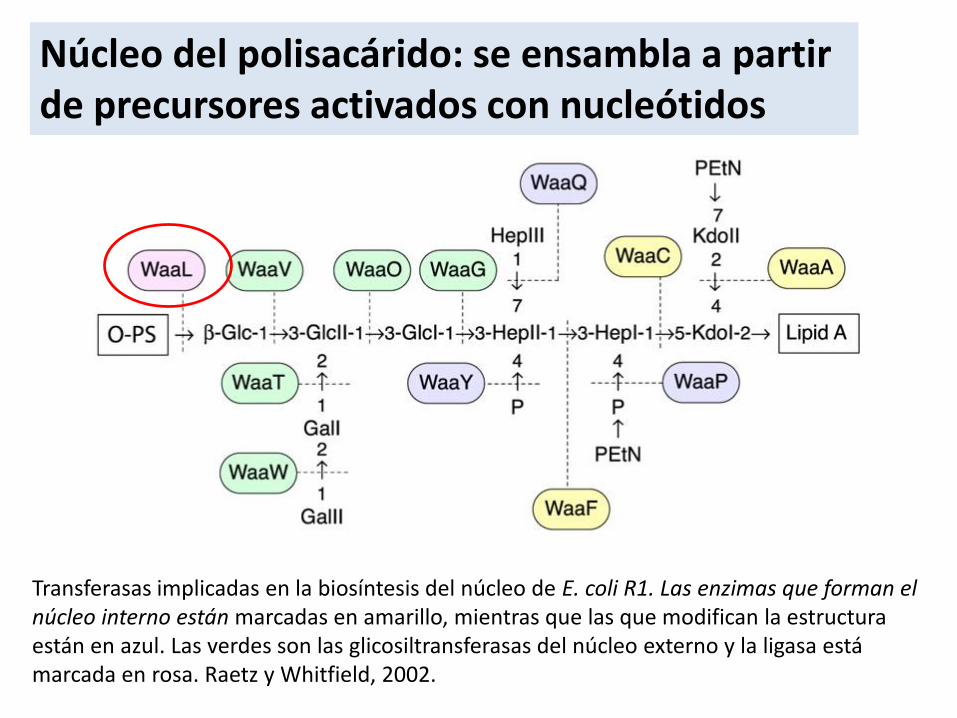
Transferasas implicadas en la biosíntesis del núcleo de E. coli R1. Las enzimas que forman el núcleo interno están marcadas en amarillo, mientras que las que modifican la estructura están en azul. Las verdes son las glicosiltransferasas del núcleo externo y la ligasa está marcada en rosa. Raetz y Whitfield, 2002.
Núcleo del polisacárido: se ensambla a partir de precursores activados con nucleótidos

Biosíntesis del antígeno O
El antígeno O es la parte más externa del LPS y a su vez la más inmunogénica yvariable. Algunos precursores se obtienen del metabolismo central de la bacteria.UDP-glucosa, de la UDP-galactosa o de la UDP-N-acetilglucosamina, que seincorporan directamente al polímero en formación, o son intermediariosDe GDPmanosa, TDP-ramnosa y otros azúcares presentes exclusivamente en los diversosantígenos O.Se forma sobre el undecaprenil-PIntervienen glicosiltransferasasLos oligosacáridos se forman en la cara interna de la MI y luego son traslocados.El antígeno O se ensambla separadamente sobre el undecaprenil-P y es traslocado por un transportador putativo Wzx. Los oligosacáridos son polimerizados en la cara periplásmica de la membrana interna por Wzy y Wzz y luego trasferidos al lípido A por WaaL..
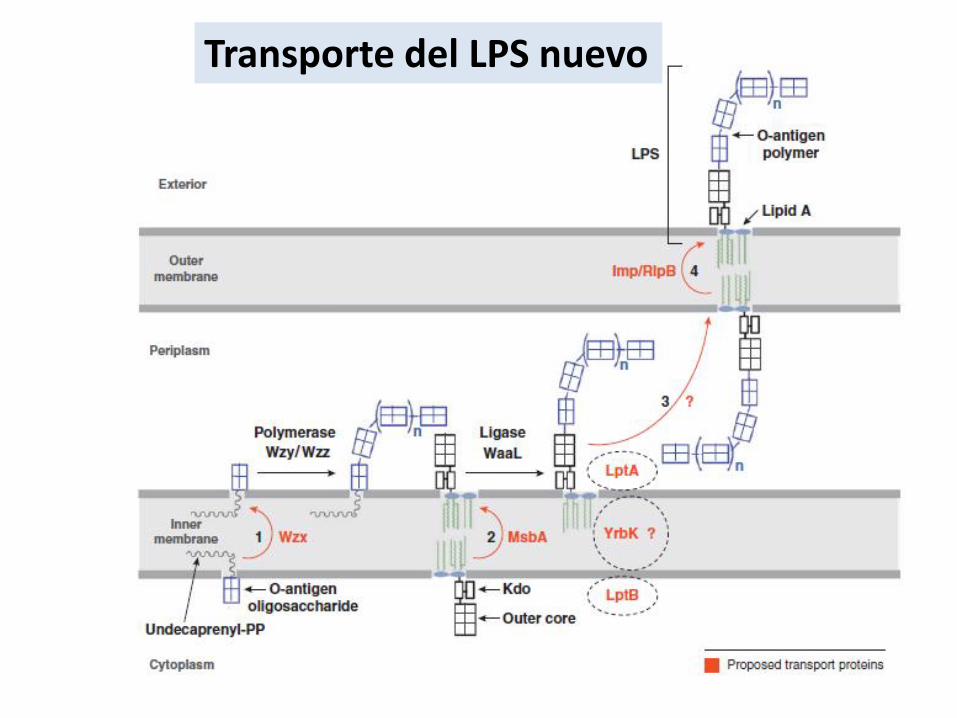
Transporte del LPS nuevo

Diferenciaciones en la célula procariota
1-Diversidad2- Esporas bacterianasObservaciónComposición química y estructuraEl proceso de eporulaciónPropiedades biológicas de las esporasGerminación3- Otras células diferenciadasQuistes bacterianosMixosporasAcinetos
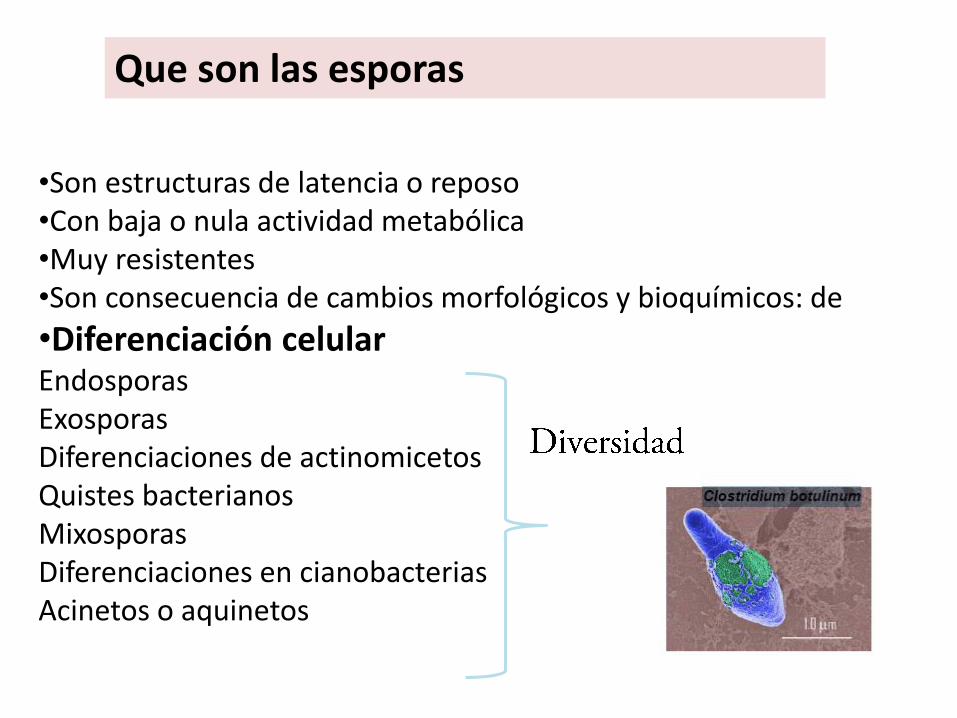
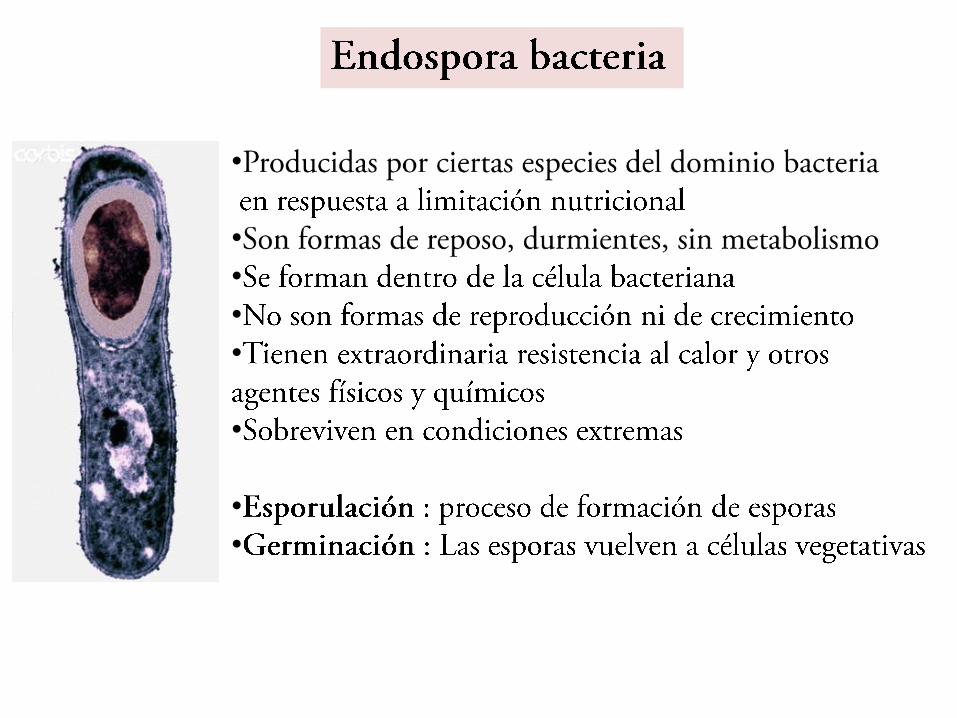
Que son las esporas
•Son estructuras de latencia o reposo•Con baja o nula actividad metabólica•Muy resistentes•Son consecuencia de cambios morfológicos y bioquímicos: de
•Diferenciación celularEndosporasExosporasDiferenciaciones de actinomicetosQuistes bacterianosMixosporasDiferenciaciones en cianobacteriasAcinetos o aquinetos
•
•
•
•
•
•
•
•

Bacilos• Bacillus• Clostridium
Cocos• Sporosarcina
°
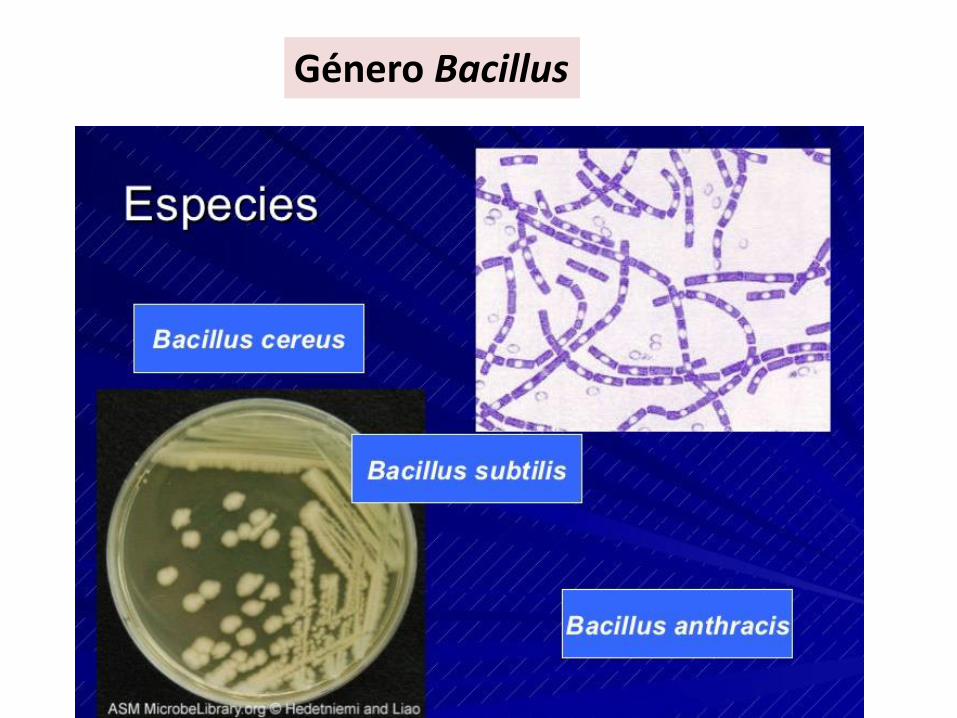
Género Bacillus
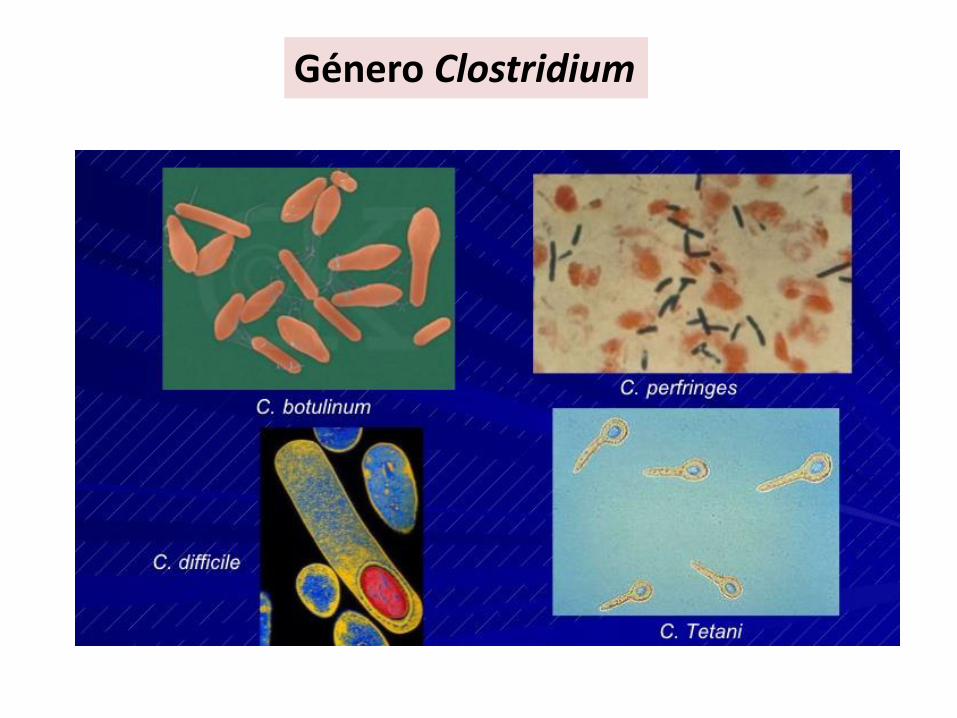
Género Clostridium
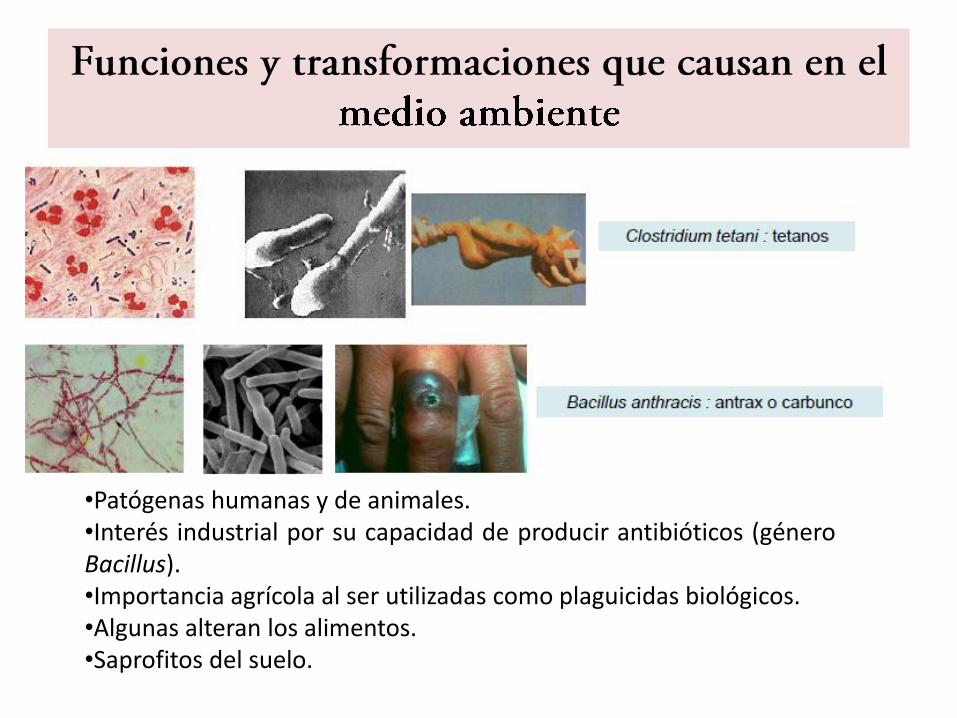
•Patógenas humanas y de animales.•Interés industrial por su capacidad de producir antibióticos (géneroBacillus).•Importancia agrícola al ser utilizadas como plaguicidas biológicos.•Algunas alteran los alimentos.•Saprofitos del suelo.
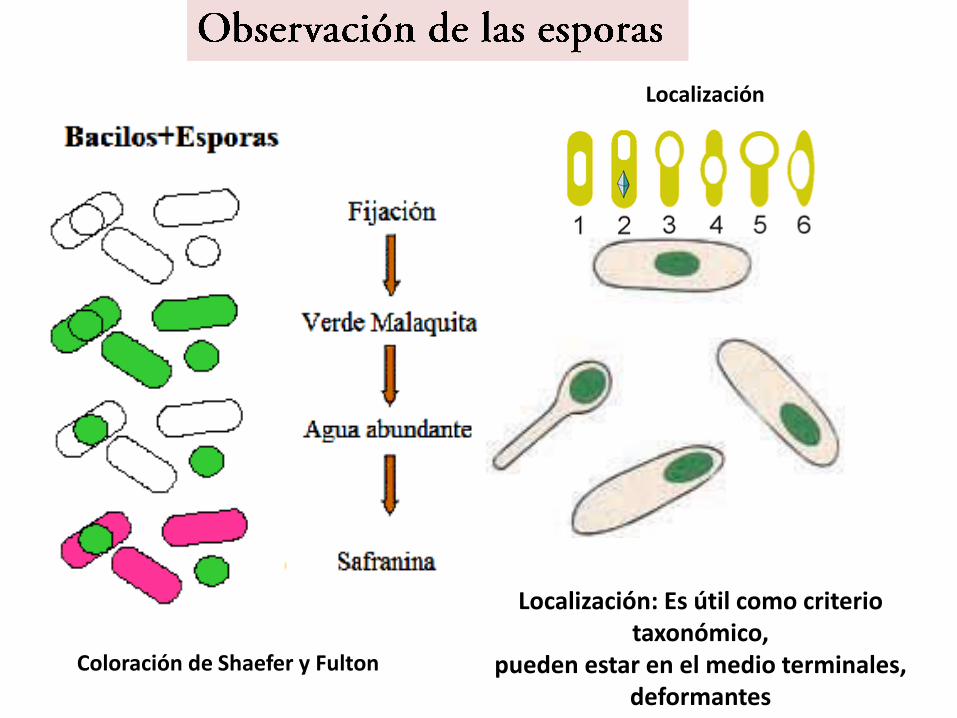
Localización
Coloración de Shaefer y Fulton
Localización: Es útil como criterio taxonómico,
pueden estar en el medio terminales, deformantes
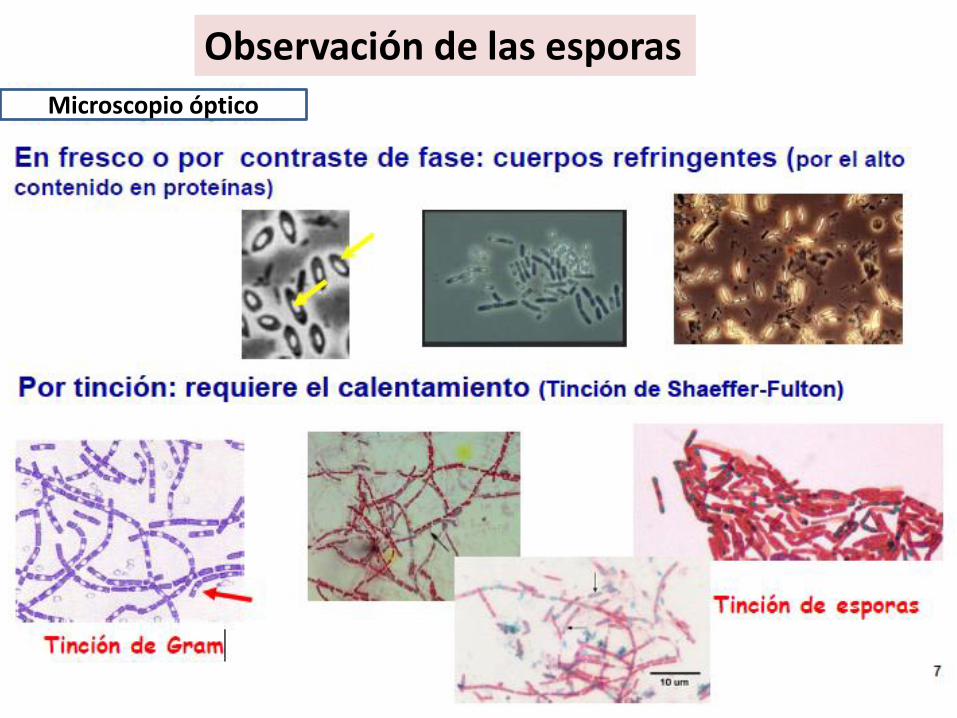
Observación de las esporas
Microscopio óptico

¿Cuánto puede vivir una espora?Los datos publicados sobre la longevidad de las endosporas indican que puedenpermanecer viables durante varias décadas y probablemente mucho tiempo mássegún las condiciones a las que esté sometida.
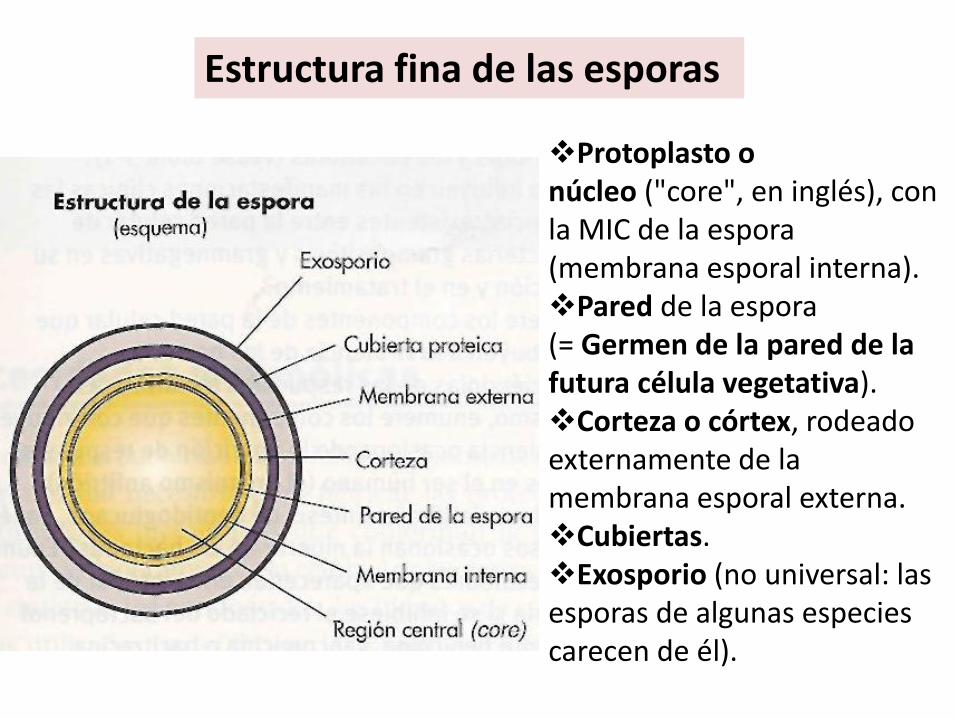
Protoplasto o núcleo ("core", en inglés), con la MIC de la espora (membrana esporal interna).Pared de la espora (= Germen de la pared de la futura célula vegetativa).Corteza o córtex, rodeado externamente de la membrana esporal externa.Cubiertas.Exosporio (no universal: las esporas de algunas especies carecen de él).
Estructura fina de las esporas
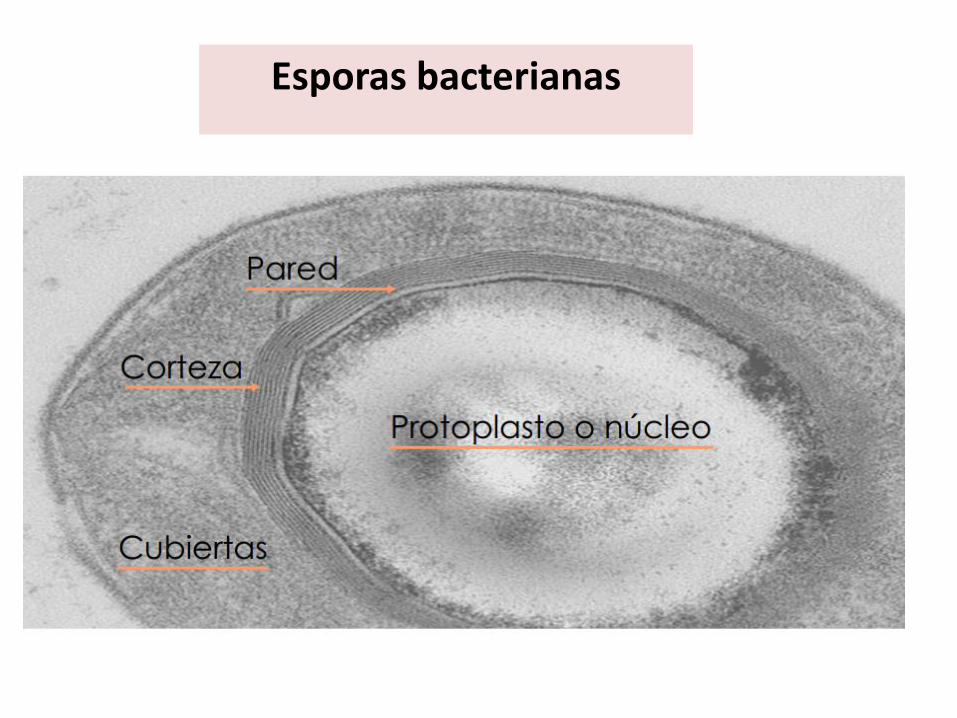
Esporas bacterianas
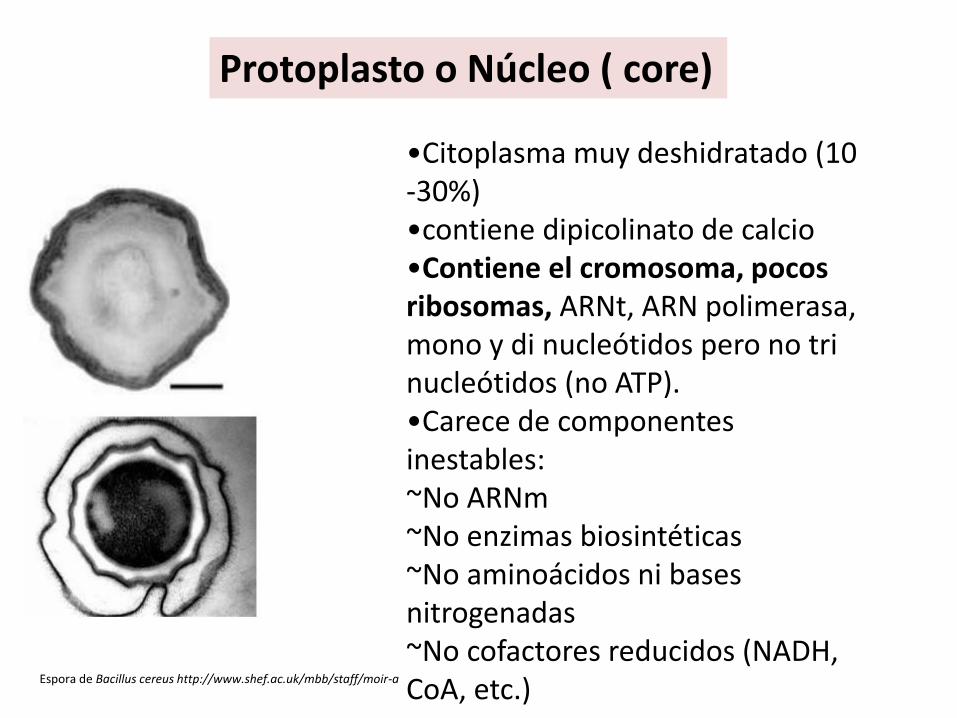
Protoplasto o Núcleo ( core)
Espora de Bacillus cereus http://www.shef.ac.uk/mbb/staff/moir-a
•Citoplasma muy deshidratado (10 -30%)•contiene dipicolinato de calcio•Contiene el cromosoma, pocos ribosomas, ARNt, ARN polimerasa, mono y di nucleótidos pero no trinucleótidos (no ATP).•Carece de componentes inestables: ~No ARNm~No enzimas biosintéticas~No aminoácidos ni bases nitrogenadas ~No cofactores reducidos (NADH, CoA, etc.)
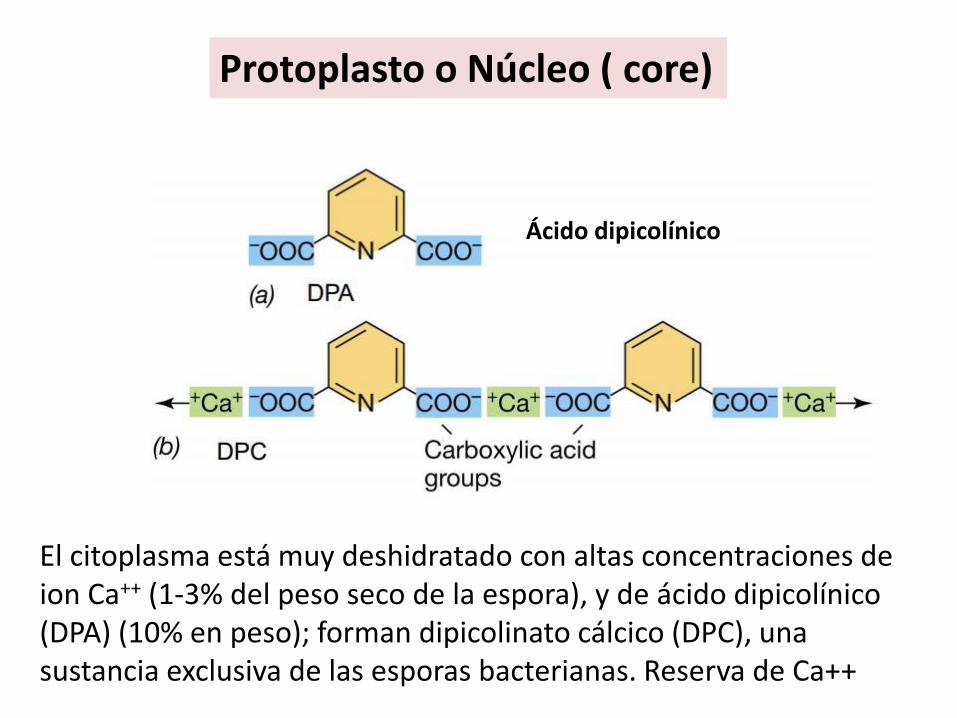
Protoplasto o Núcleo ( core)
El citoplasma está muy deshidratado con altas concentraciones de ion Ca++ (1-3% del peso seco de la espora), y de ácido dipicolínico (DPA) (10% en peso); forman dipicolinato cálcico (DPC), una sustancia exclusiva de las esporas bacterianas. Reserva de Ca++
Ácido dipicolínico

Protoplasto o Núcleo ( core)
•-Gran cantidad de pequeñas proteínas especiales, las pequeñas, ácidas, solubles (SASP) que mantienen le pH más bajo que en la célula vegetativa. -Durante la germinación se usarán como fuente de C, E y AA. -Acomplejan el ADN: protegen de las radiaciones UV.•-Fuente de energía: 3-fosfoglicerato→PEP•pH es una unidad menor que el citoplasma de la cel. vegetativa
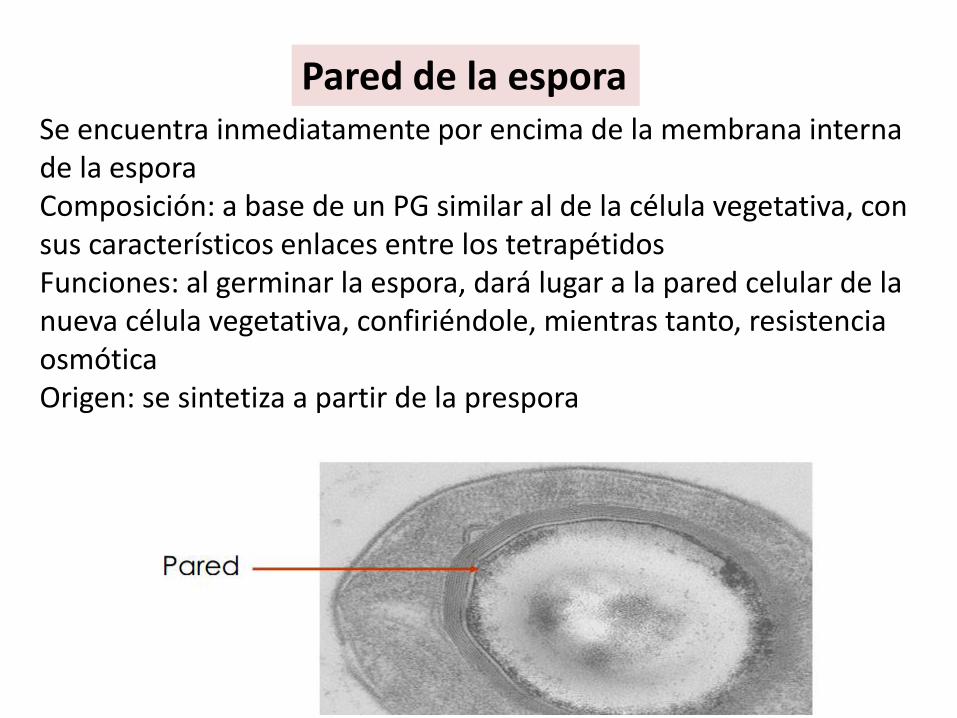
Pared de la esporaSe encuentra inmediatamente por encima de la membrana interna de la esporaComposición: a base de un PG similar al de la célula vegetativa, con sus característicos enlaces entre los tetrapétidosFunciones: al germinar la espora, dará lugar a la pared celular de la nueva célula vegetativa, confiriéndole, mientras tanto, resistencia osmóticaOrigen: se sintetiza a partir de la prespora
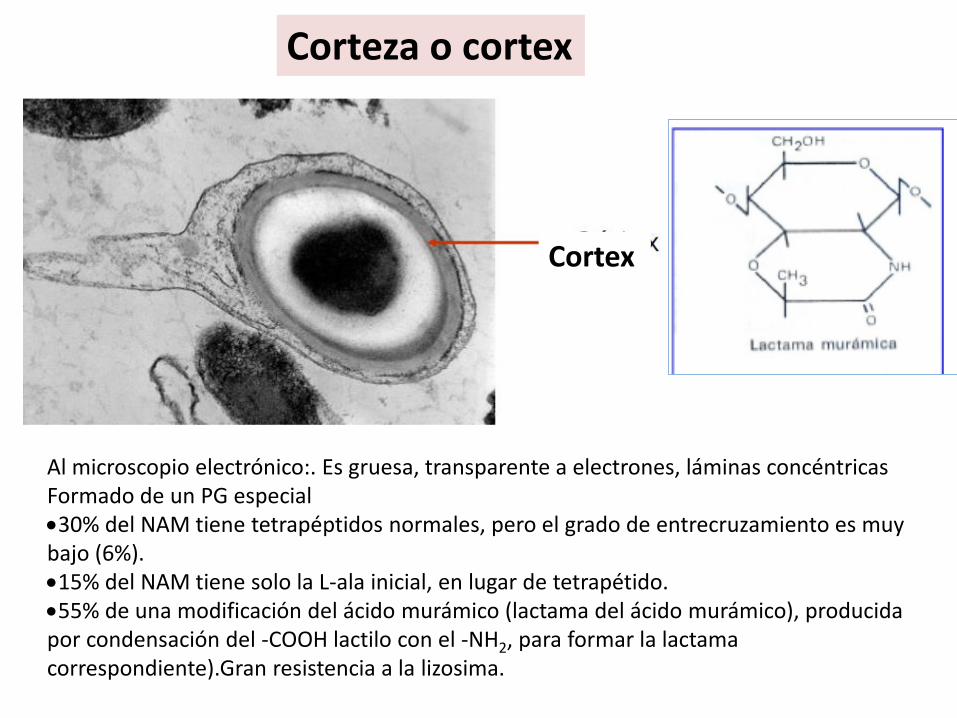
Corteza o cortex
Al microscopio electrónico:. Es gruesa, transparente a electrones, láminas concéntricas Formado de un PG especial30% del NAM tiene tetrapéptidos normales, pero el grado de entrecruzamiento es muy bajo (6%).15% del NAM tiene solo la L-ala inicial, en lugar de tetrapétido.55% de una modificación del ácido murámico (lactama del ácido murámico), producida por condensación del -COOH lactilo con el -NH2, para formar la lactamacorrespondiente).Gran resistencia a la lizosima.
Cortex
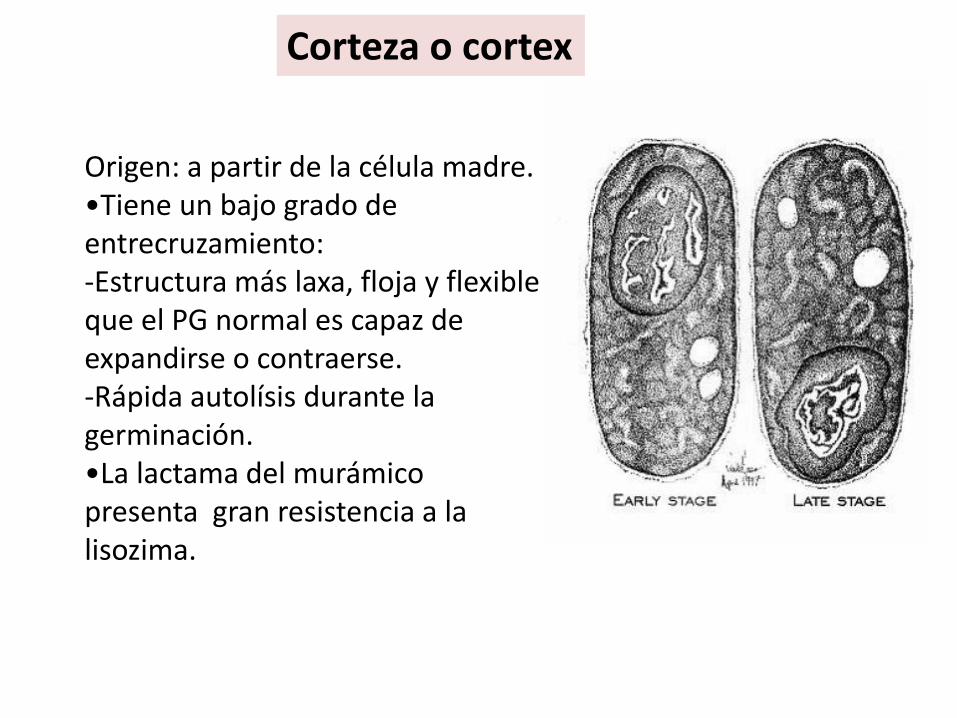
Corteza o cortex
Origen: a partir de la célula madre.•Tiene un bajo grado de entrecruzamiento: -Estructura más laxa, floja y flexible que el PG normal es capaz de expandirse o contraerse.-Rápida autolísis durante la germinación.•La lactama del murámico presenta gran resistencia a la lisozima.
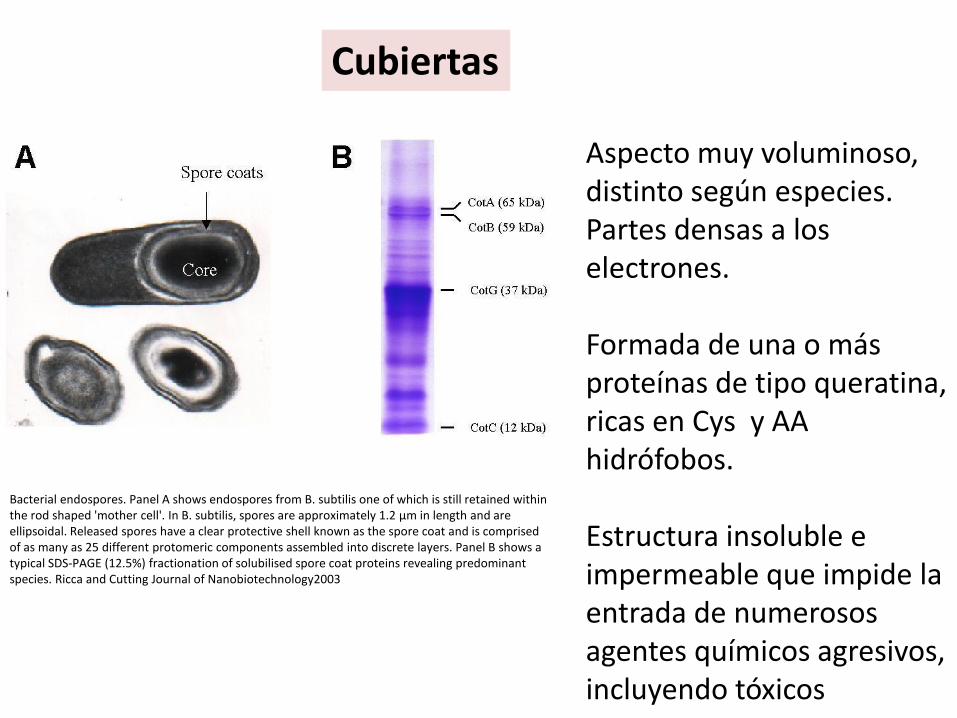
Aspecto muy voluminoso, distinto según especies. Partes densas a los electrones.
Formada de una o más proteínas de tipo queratina, ricas en Cys y AA hidrófobos.
Estructura insoluble e impermeable que impide la entrada de numerosos agentes químicos agresivos, incluyendo tóxicos
Cubiertas
Bacterial endospores. Panel A shows endospores from B. subtilis one of which is still retained within the rod shaped 'mother cell'. In B. subtilis, spores are approximately 1.2 μm in length and are ellipsoidal. Released spores have a clear protective shell known as the spore coat and is comprised of as many as 25 different protomeric components assembled into discrete layers. Panel B shows a typical SDS-PAGE (12.5%) fractionation of solubilised spore coat proteins revealing predominant species. Ricca and Cutting Journal of Nanobiotechnology2003
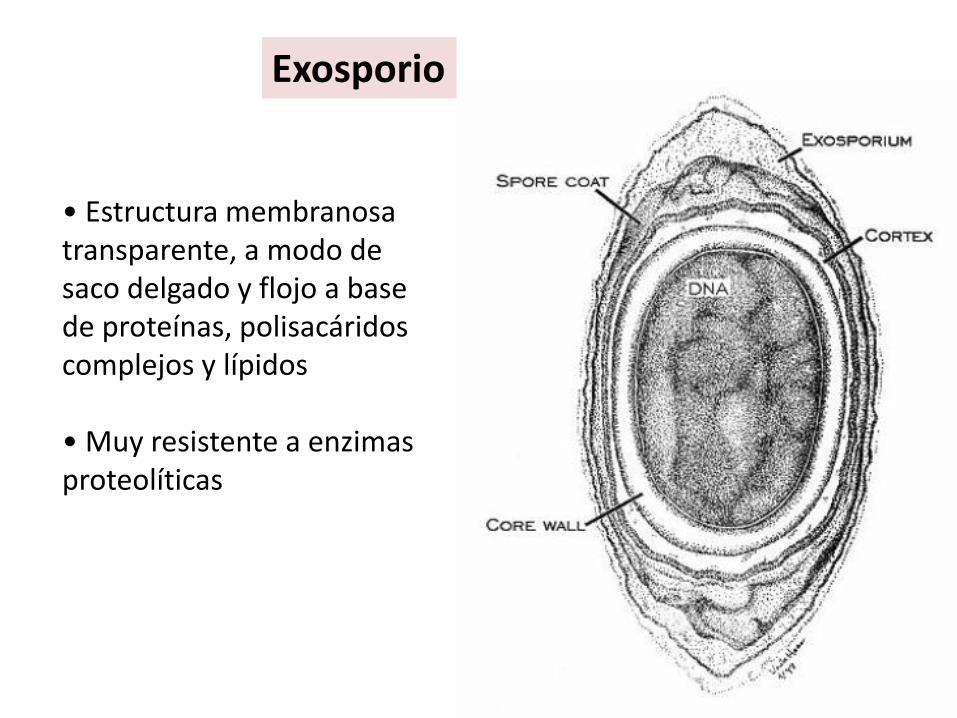
• Estructura membranosa transparente, a modo de saco delgado y flojo a base de proteínas, polisacáridos complejos y lípidos
• Muy resistente a enzimas proteolíticas
Exosporio

El proceso de esporulación.. Cuando???
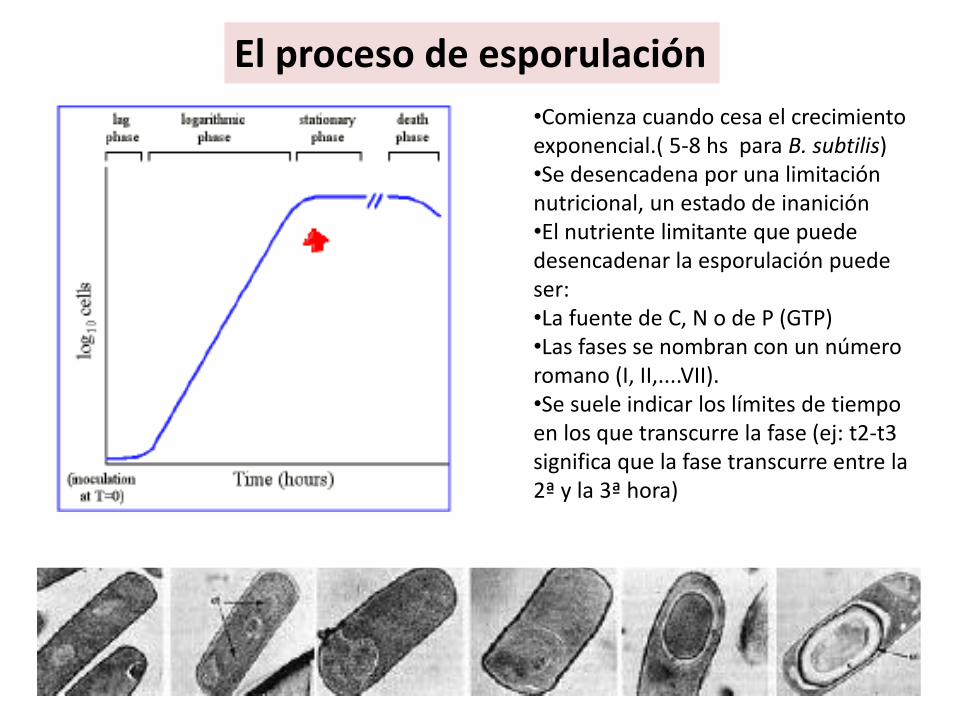
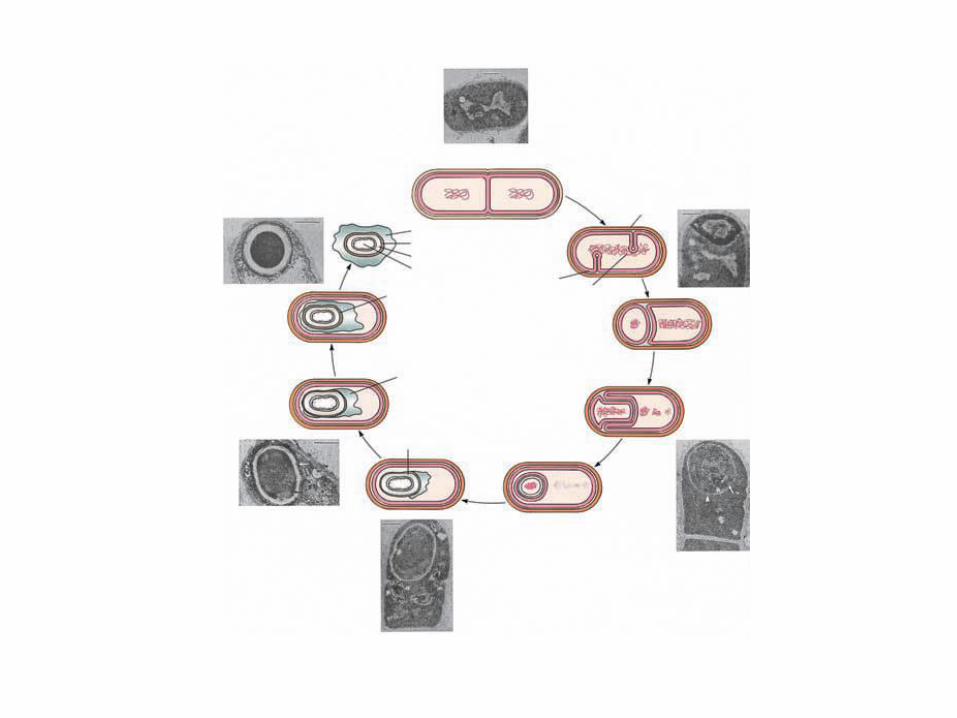
El proceso de esporulación
•Comienza cuando cesa el crecimiento exponencial.( 5-8 hs para B. subtilis)•Se desencadena por una limitación nutricional, un estado de inanición•El nutriente limitante que puede desencadenar la esporulación puede ser: •La fuente de C, N o de P (GTP)•Las fases se nombran con un número romano (I, II,....VII). •Se suele indicar los límites de tiempo en los que transcurre la fase (ej: t2-t3 significa que la fase transcurre entre la 2ª y la 3ª hora)
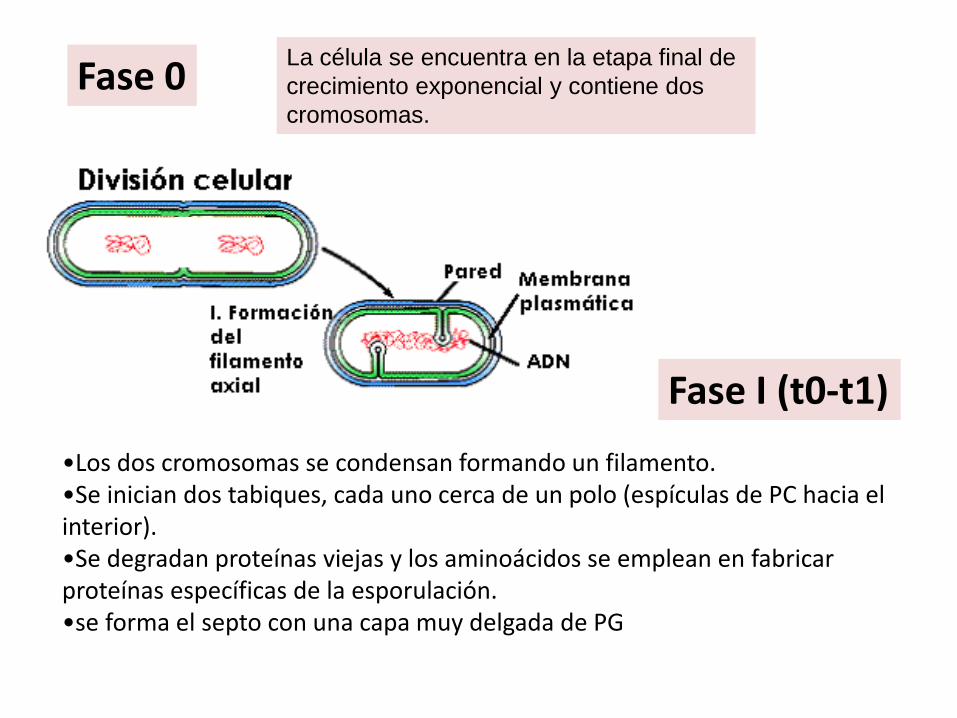
•Los dos cromosomas se condensan formando un filamento. •Se inician dos tabiques, cada uno cerca de un polo (espículas de PC hacia el interior). •Se degradan proteínas viejas y los aminoácidos se emplean en fabricar proteínas específicas de la esporulación. •se forma el septo con una capa muy delgada de PG
La célula se encuentra en la etapa final de
crecimiento exponencial y contiene dos
cromosomas.
Fase 0
Fase I (t0-t1)
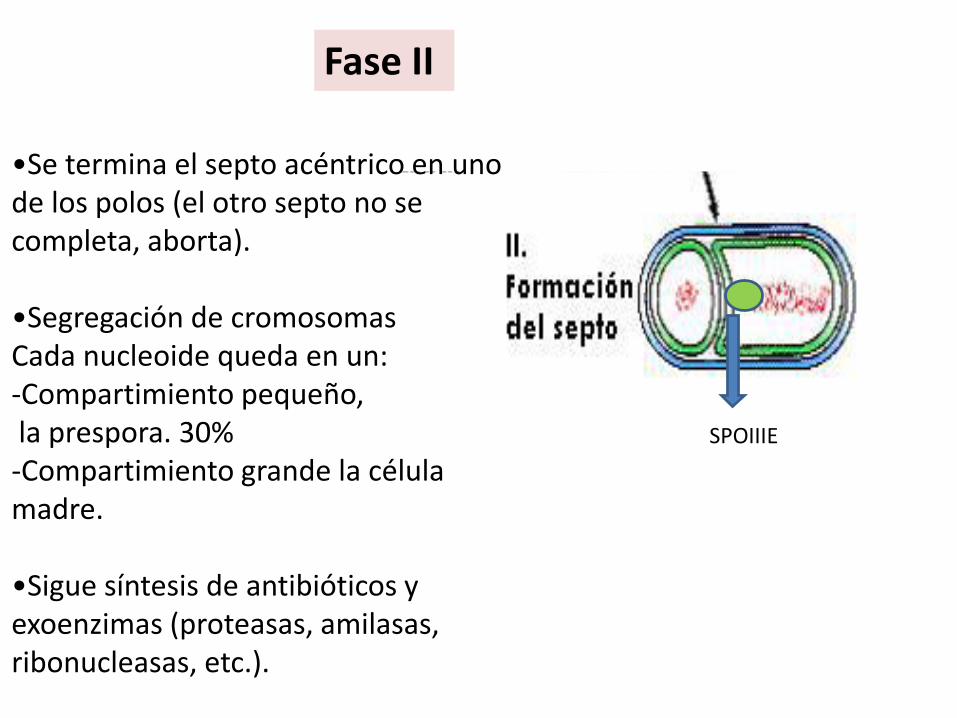
Fase II
•Se termina el septo acéntrico en uno de los polos (el otro septo no se completa, aborta).
•Segregación de cromosomasCada nucleoide queda en un: -Compartimiento pequeño,la prespora. 30%-Compartimiento grande la célula madre.
•Sigue síntesis de antibióticos y exoenzimas (proteasas, amilasas, ribonucleasas, etc.).
SPOIIIE
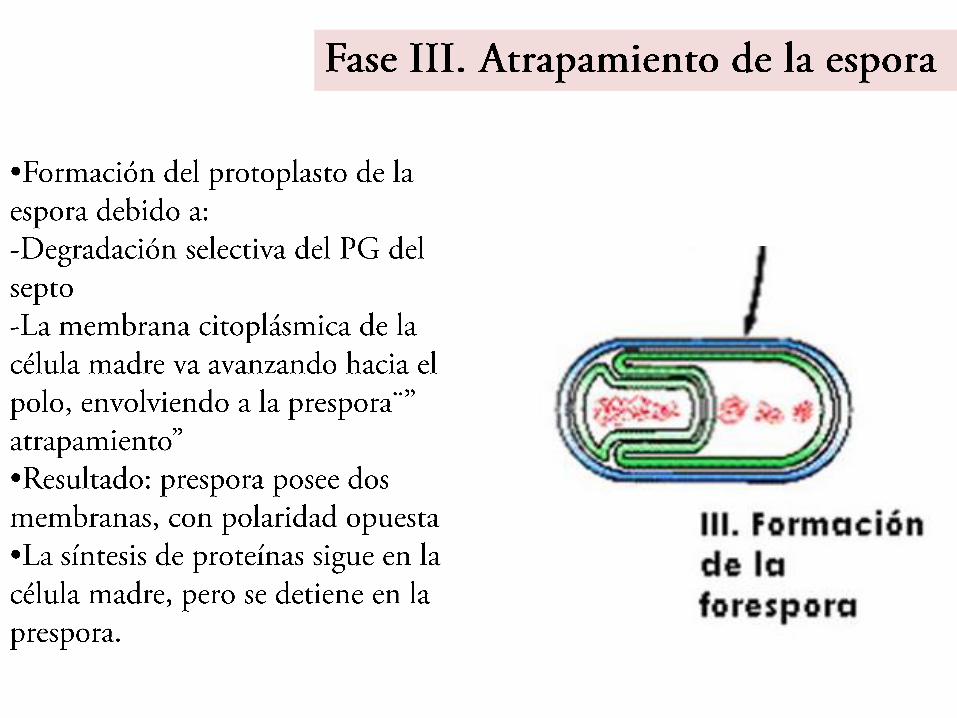
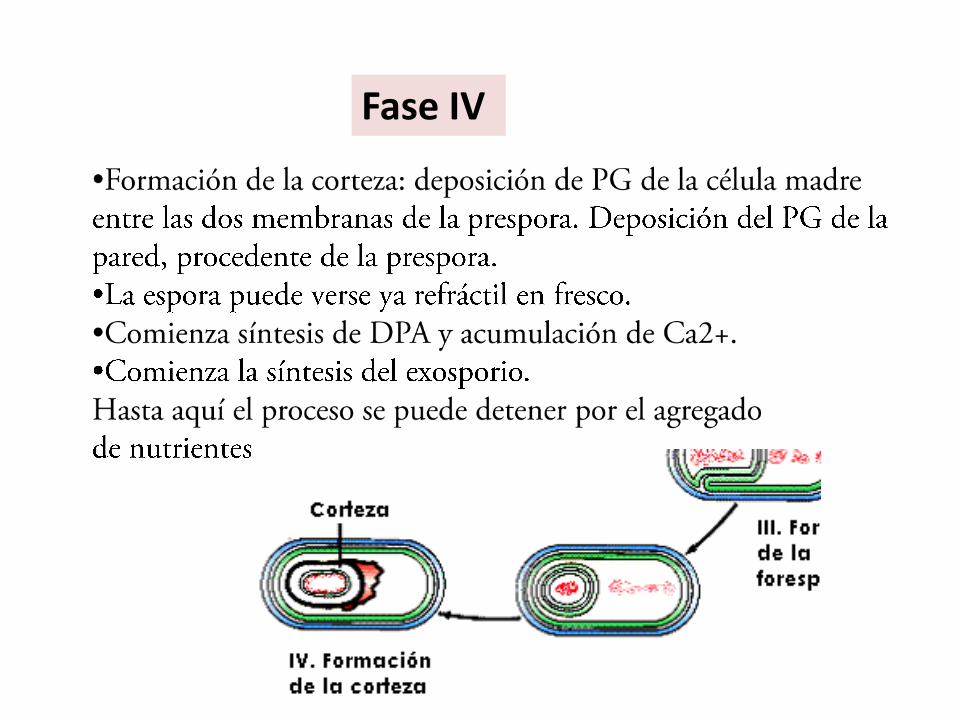
Fase IV
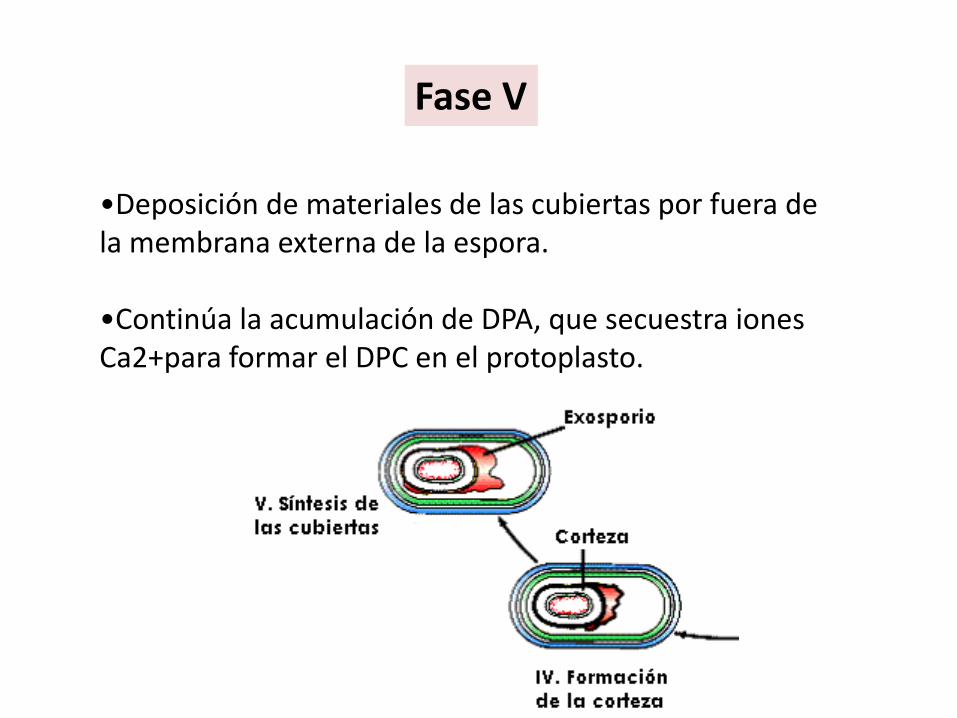
•Deposición de materiales de las cubiertas por fuera de la membrana externa de la espora.
•Continúa la acumulación de DPA, que secuestra iones Ca2+para formar el DPC en el protoplasto.
Fase V
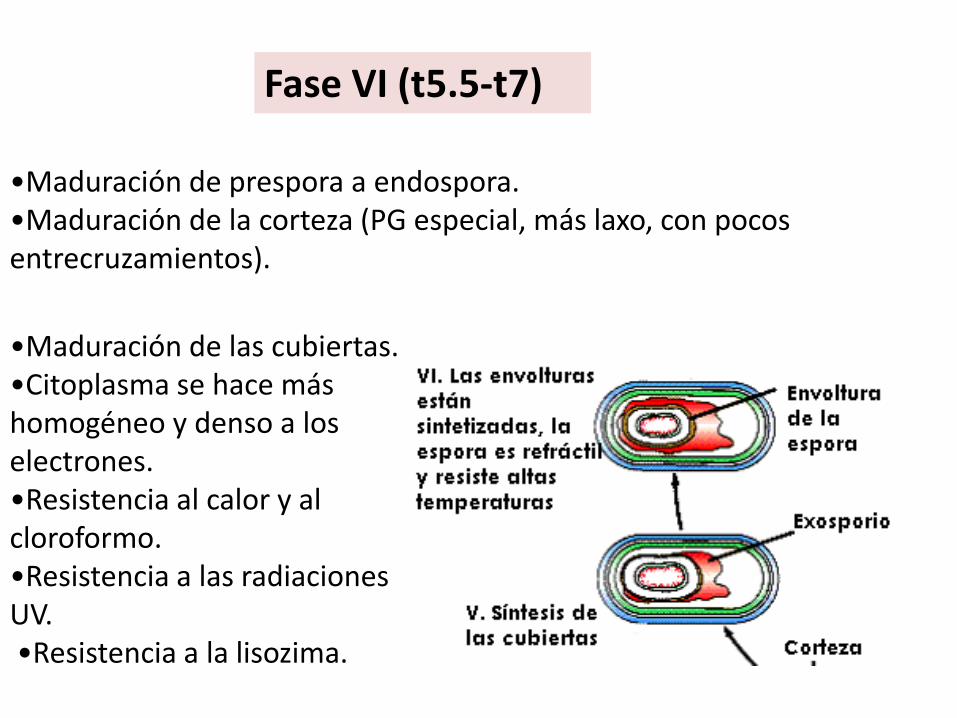
•Maduración de las cubiertas. •Citoplasma se hace más homogéneo y denso a los electrones.•Resistencia al calor y al cloroformo. •Resistencia a las radiaciones UV.•Resistencia a la lisozima.
•Maduración de prespora a endospora. •Maduración de la corteza (PG especial, más laxo, con pocos entrecruzamientos).
Fase VI (t5.5-t7)
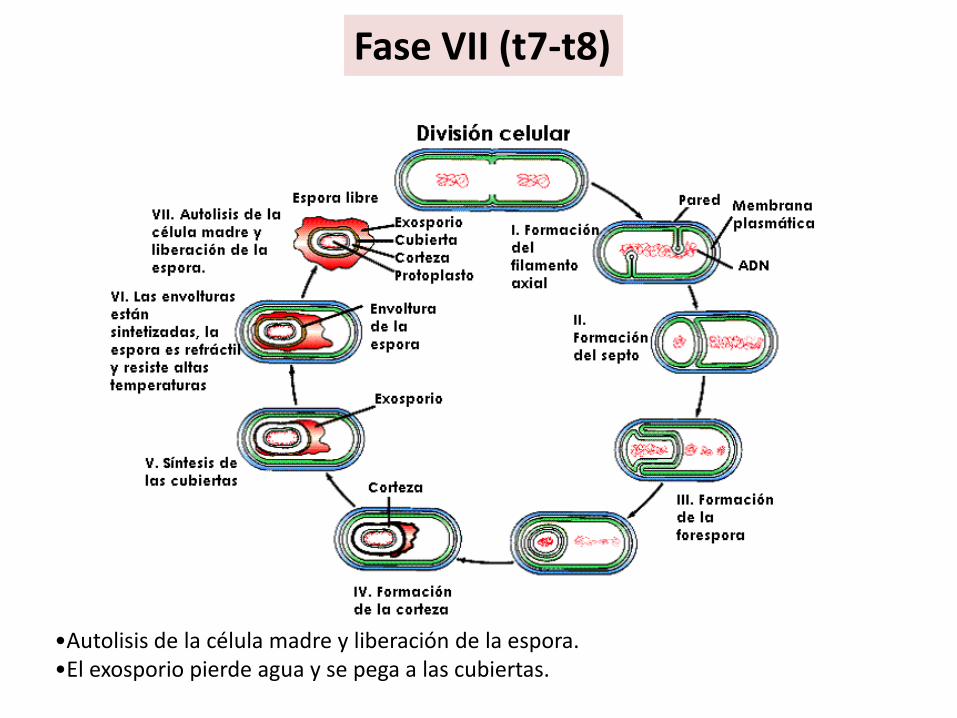
•Autolisis de la célula madre y liberación de la espora. •El exosporio pierde agua y se pega a las cubiertas.
Fase VII (t7-t8)

Propiedades de las endosporas
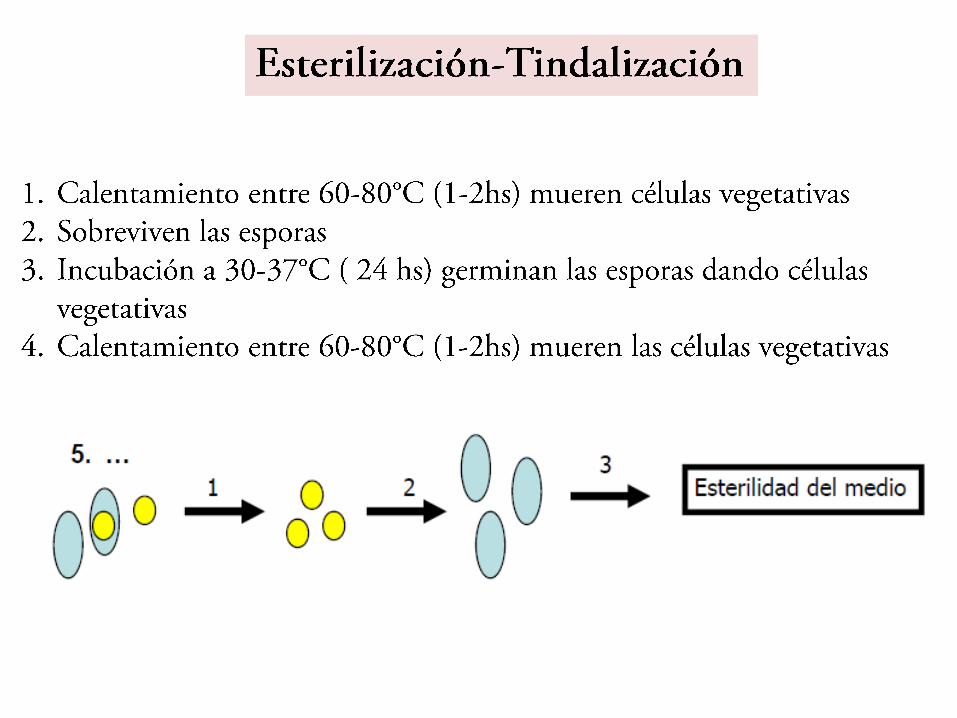
• Resistencia al calor. Es una consecuencia de los cambios que llevan a la deshidratación como medio de lograr la el estado de dormancia y baja tasa metabólica. Algunas resisten 120ºC durante 15 minutos lo que condiciona los parámetros para esterilizar
• La tasa metabólica más baja
• Permite la supervivencia en ambientes desfavorables
• DNA protegido por ácido dipicolínico y proteínas SAPS
• Luego de la activación por stress, la disponibilidad de nutrientes dispara la germinación y el crecimiento
• Dormancia. Gran inercia a los sustratos exógenos, la espora sólo perderá la dormancia cuando se haya activado para la germinación.

Deshidratación. Mecanismo:~El DPA va entrando al protoplasto de la espora ~El Ca2+entra a la espora se forman quelatos de DPC.~La corteza se queda sin cationes, las cargas negativas del PG cortical se repelen, la corteza se expande, se topa con las cubiertas y hay extracción de agua del protoplasto. ~El protoplasto queda muy deshidratado, con componentes inmovilizados.
Propiedades biológicas de las endospora

• Resistencia a los rayos UV: ~Absorción de UV por cubiertas. ~Presencia del DPC. ~Las proteínas SASPs forman complejos con el ADN. ~Por la deshidratación del protoplasto no se generan dímeros de pirimidina.•Resistencia a los agentes químicos. Debida principalmente a la gran impermeabilidad de las cubiertas(grosor, composición a base de proteínas ricas en aminoácidos hidrófobos y con abundantes puentes disulfuro.
Propiedades biológicas de las endospora
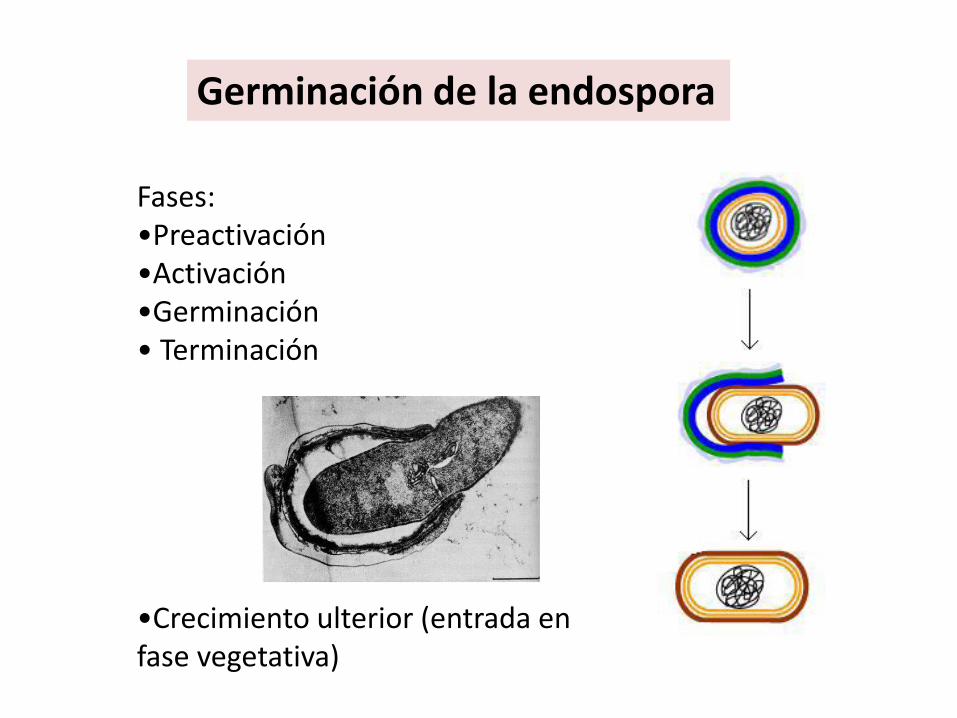
Fases: •Preactivación •Activación •Germinación• Terminación
•Crecimiento ulterior (entrada en fase vegetativa)
Germinación de la endospora
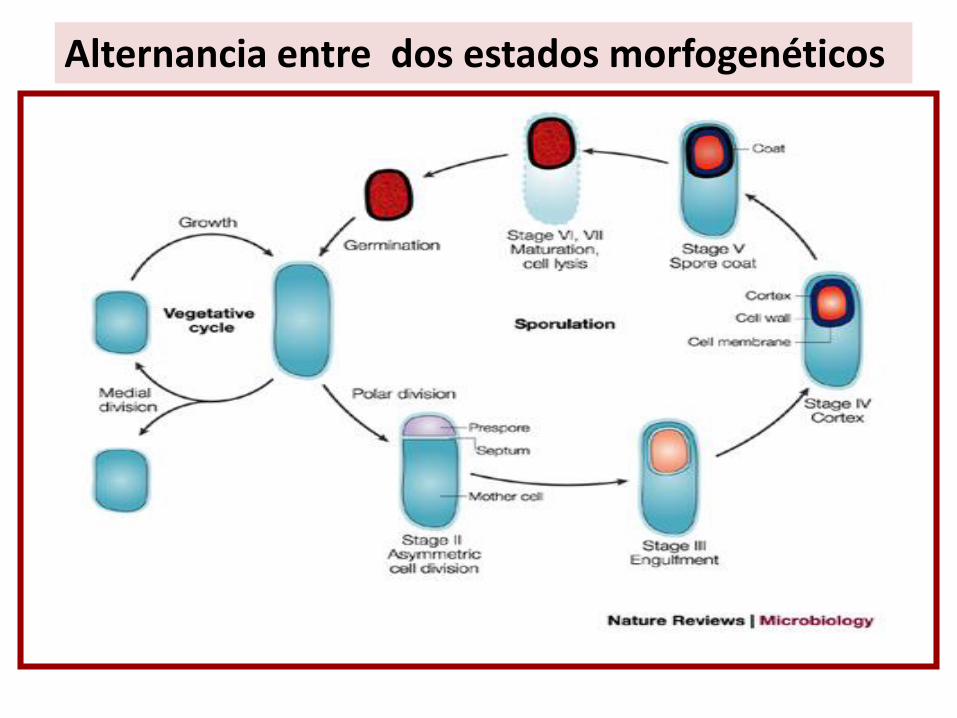
Alternancia entre dos estados morfogenéticos

Fenómenos bioquímicos asociados a la esporulación
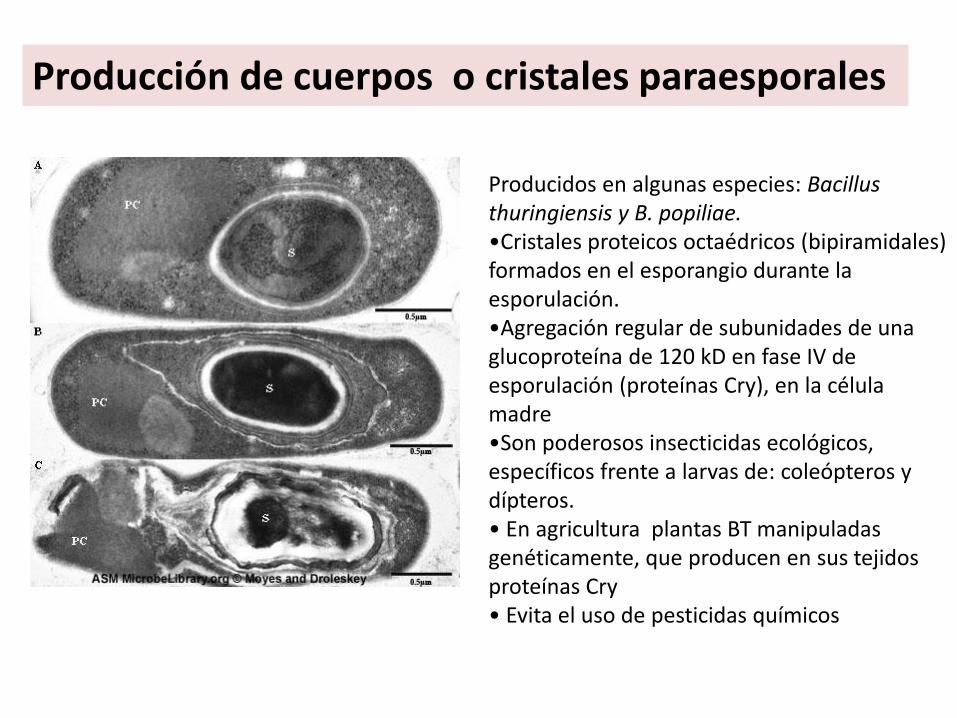
Producción de cuerpos o cristales paraesporales
Producidos en algunas especies: Bacillusthuringiensis y B. popiliae.•Cristales proteicos octaédricos (bipiramidales) formados en el esporangio durante la esporulación. •Agregación regular de subunidades de una glucoproteína de 120 kD en fase IV de esporulación (proteínas Cry), en la célula madre•Son poderosos insecticidas ecológicos, específicos frente a larvas de: coleópteros y dípteros.• En agricultura plantas BT manipuladas genéticamente, que producen en sus tejidos proteínas Cry• Evita el uso de pesticidas químicos
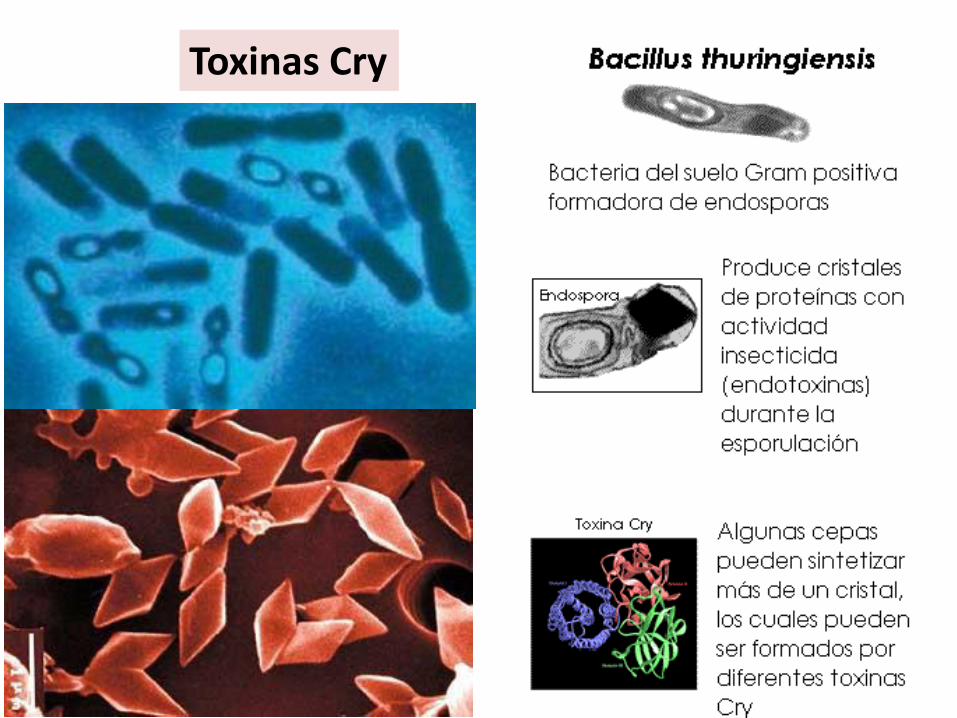
Toxinas Cry

Actúa como insecticida.•Oruga ingiere materia vegetal con bacterias esporuladas que producen Cry. •La proteína Cry se disuelve en el tracto digestivo. El pH alcalino provoca la proteolísis que activa a la toxina. •La toxina altera la permeabilidad del epitelio intestinal, pasa a la hemolinfa lo que provoca la parálisis y muerte de la larva.
Toxinas Cry
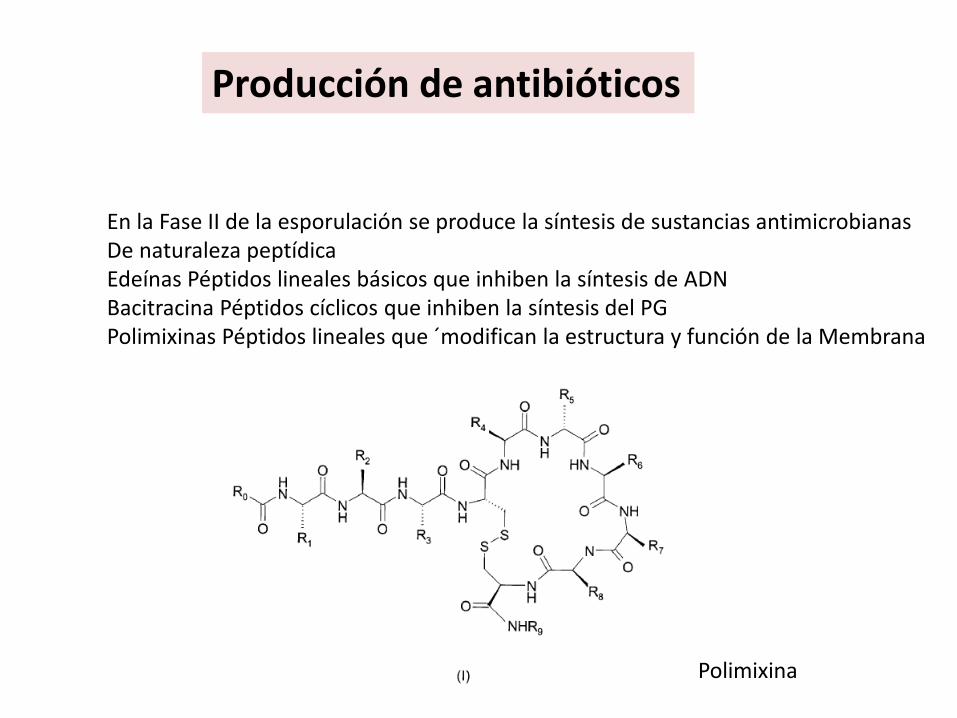
Producción de antibióticos
En la Fase II de la esporulación se produce la síntesis de sustancias antimicrobianas De naturaleza peptídicaEdeínas Péptidos lineales básicos que inhiben la síntesis de ADNBacitracina Péptidos cíclicos que inhiben la síntesis del PGPolimixinas Péptidos lineales que ´modifican la estructura y función de la Membrana
Polimixina

Exosporas
Son esporas externas, son resistentes sólo a calor y desecación y no contienen DPA. Podemos encontrar exoesporas en el género Methylosinum (bacterias que oxidan metano) y Rhodomicrobium. Estas bacterias forman esporas reproductivas por gemaciones sucesivas al final de sus prostecas. Estas exosporas poseen una envuelta a base de pared rodeada de una cápsula o cubierta gruesa.
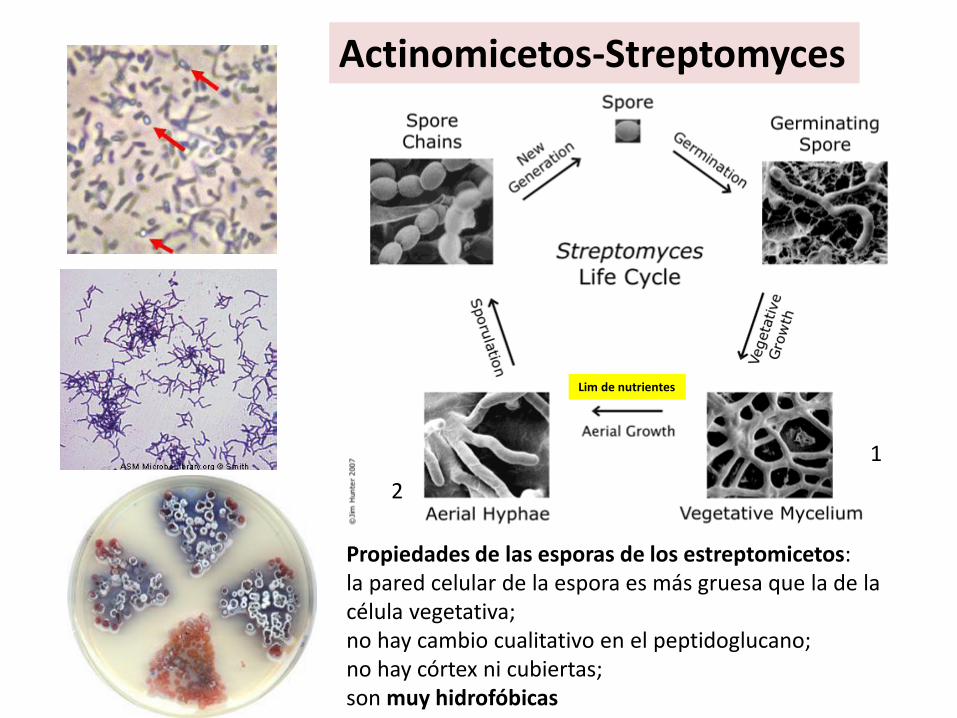
Actinomicetos-Streptomyces
Propiedades de las esporas de los estreptomicetos:la pared celular de la espora es más gruesa que la de la célula vegetativa;no hay cambio cualitativo en el peptidoglucano;no hay córtex ni cubiertas;son muy hidrofóbicas
1
2
Lim de nutrientes
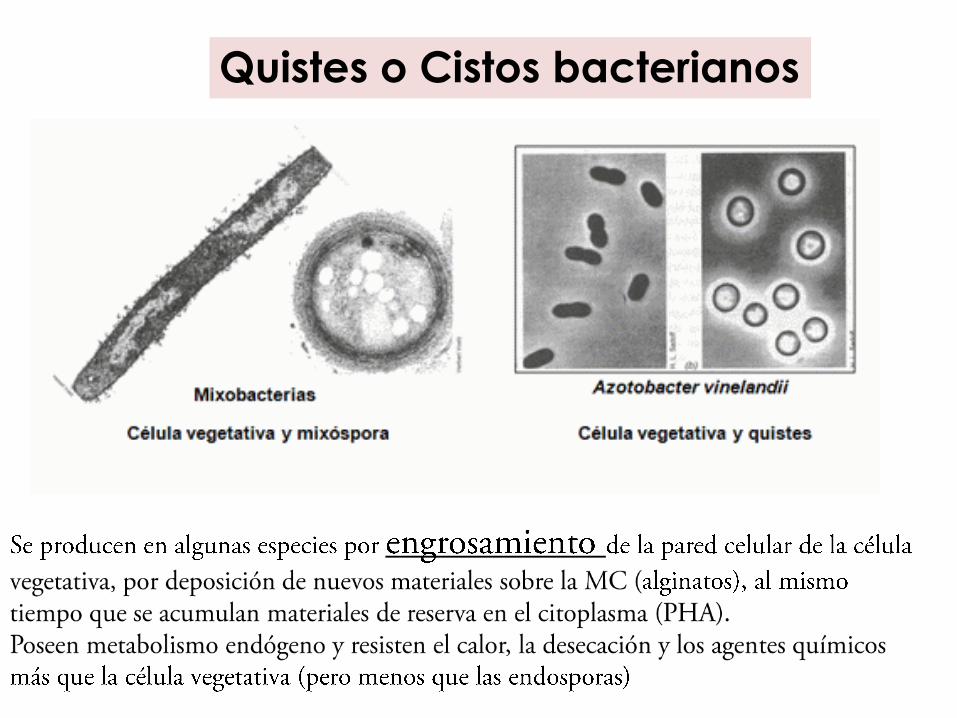
Quistes o Cistos bacterianos
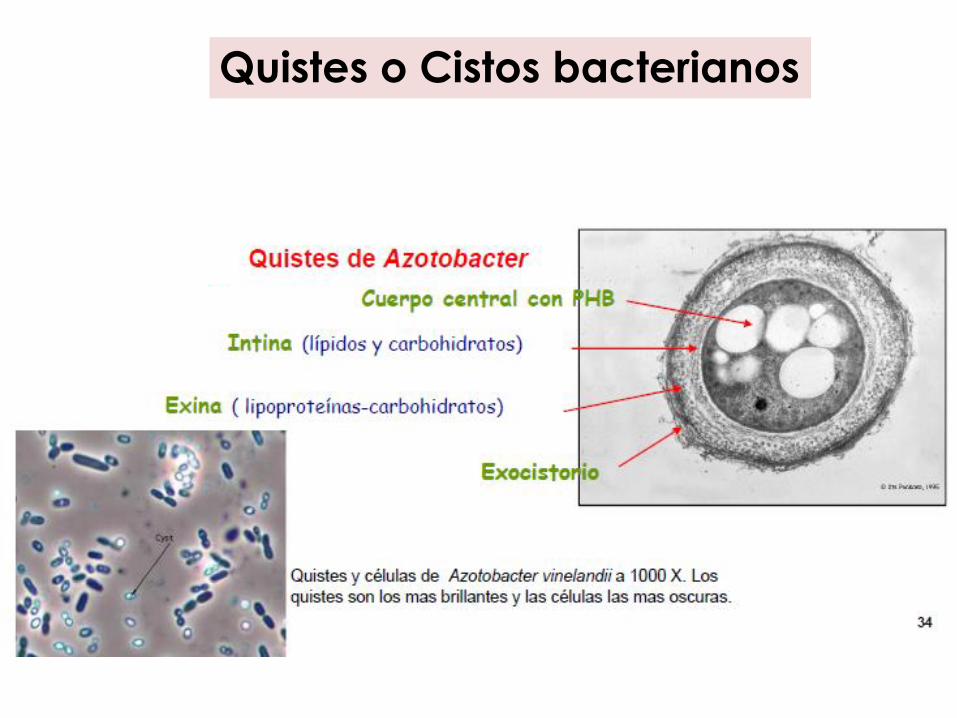
Quistes o Cistos bacterianos
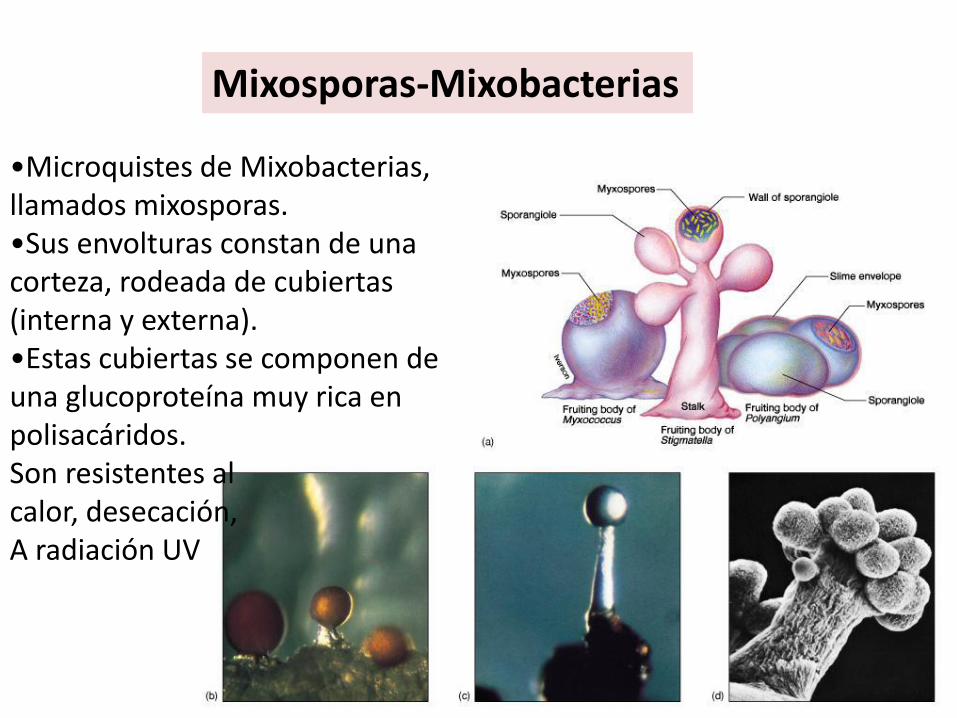
•Microquistes de Mixobacterias, llamados mixosporas. •Sus envolturas constan de una corteza, rodeada de cubiertas (interna y externa). •Estas cubiertas se componen de una glucoproteína muy rica en polisacáridos.Son resistentes alcalor, desecación,A radiación UV
Mixosporas-Mixobacterias
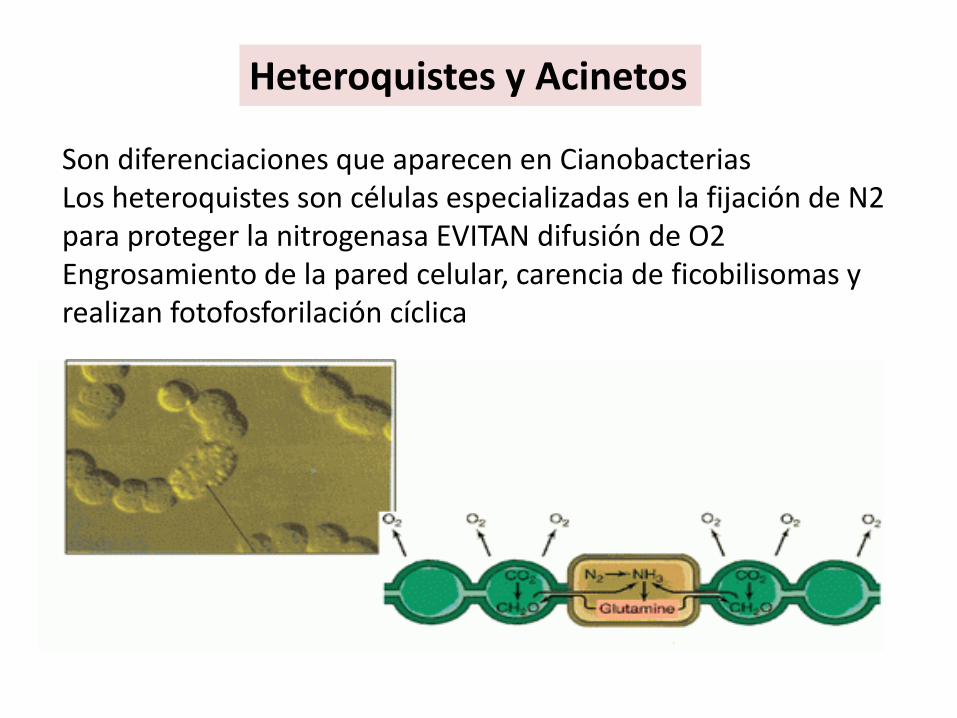
Heteroquistes y Acinetos
Son diferenciaciones que aparecen en CianobacteriasLos heteroquistes son células especializadas en la fijación de N2 para proteger la nitrogenasa EVITAN difusión de O2Engrosamiento de la pared celular, carencia de ficobilisomas y realizan fotofosforilación cíclica

Acinetos
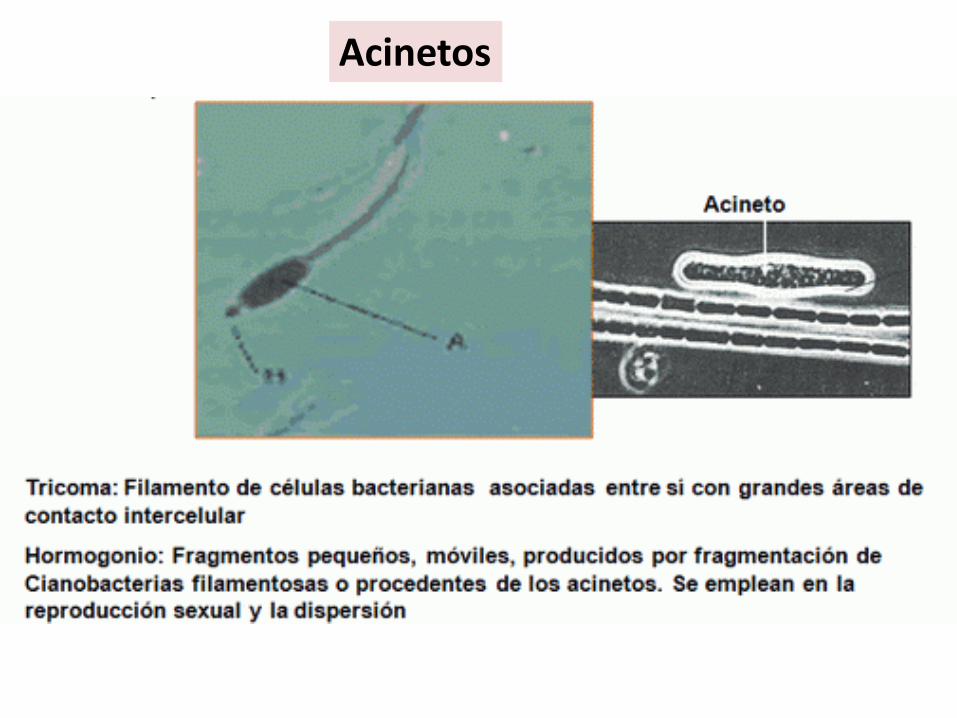
Acinetos





























