Embed Size (px)
Citation preview

Contents lists available at ScienceDirect
Brain Research Bulletin
journal homepage: www.elsevier.com/locate/brainresbull
Regulation of HDAC1 and HDAC2 during consolidation and extinction offear memory
Sarfraj Ahmad Siddiquia,1, Sanjay Singha,1, Rajesh Ugaleb, Vandana Ranjanc, Rohit Kanojiaa,Sudipta Sahaf, Sukanya Tripathya, Shiv Kumard, Sudhir Mehrotrad, Dinesh Raj Modia,Anand Prakasha,e,⁎
a Department of Biotechnology, Babasaheb Bhimrao Ambedkar University, Lucknow, IndiabDepartment of Pharmaceutical Sciences, RTM Nagpur University, Nagpur, Indiac Department of Biochemistry, RML University, Faizabad, IndiadDepartment of Biochemistry, University of Lucknow, Lucknow, Indiae Department of Biotech, Mahatma Gandhi Central University, Motihari, Bihar, IndiafDepartment of Pharmaceutical Science, Babasaheb Bhimrao Ambedkar University, Lucknow, India
A R T I C L E I N F O
Keywords:HDACFear memoryExtinctionPFCAmygdala
A B S T R A C T
Histone deacetylases (HDACs) regulate gene expression epigenetically through synchronized removal of acetylgroups from histones required towards memory consolidation. Moreover, dysregulated epigenetic machineryduring fear or extinction learning may result in altered expression of some of these genes and result in PostTraumatic Stress Disorder (PTSD). In the present study, region-specific expression of Histone deacetylase 1(HDAC1) and Histone deacetylase 2 (HDAC2) was correlated to the acetylation of histones H3 and H4 and theresultant conditioned response, in rats undergone fear and extinction learning. The neuronal activation, histoneacetylation at H3/H4 and expression of HDAC1/HDAC2 in centrolateral amygdala (CeL) and centromedialamygdala (CeM) of central Amygdala (CeA) and prelimbic (PL) and infralimbic (IL) of Prefrontal cortex (PFC)were found to be associated in a differential manner following fear and extinction learning. Moreover in CeM,the main output of the fear circuitry, the level of HDAC1 was down-regulated following conditioning and up-regulated following extinction as opposed to which HDAC2 was down-regulated in CeM following conditioningbut not following extinction. Furthermore, in CeL the HDAC1 was upregulated and HDAC2 was downregulatedfollowing conditioning and extinction. This has important implications in speculating of the role of HDACs infear memory consolidation and its extinction.
1. Introduction
The molecular processes controlling the synaptic plasticity in thefear circuitry are well known (Pare and Duvarci, 2012; Pape and Pare,2010; Sah et al., 2003, Marek et al., 2013). It is the outcome of syn-chronised activity of certain genes in relation to changing environ-mental contingencies. Of late the involvement of epigenetics duringsynaptic plasticity has gained importance and significant data sug-gesting the importance of histone acetylation (Karpova et al., 2017;Sultan and Day, 2011) and deacetylation in the fear and extinctionlearning is available (Stafford et al., 2012, Siddiqui et al., 2017; Bredyand Barad, 2008; Valiati et al., 2017). Increased histone acetylation hasbeen found to be associated with conditioning and extinction in
different subregions of amygdala in a number of rodent studies (Monseyet al., 2011; Ranjan et al., 2015, Blouin et al., 2016; Siddiqui et al.,2017, Itzhak et al., 2013). Likewise, altered histone acetylation hasbeen found to be involved with fear memory consolidation and ex-tinction in prefrontal cortex of rodents (Siddiqui et al., 2017, Staffordet al., 2012; Vieira et al., 2014, Marek et al., 2011). However, the de-tailed region specific role of epigenetics towards neuronal activity re-quired for fear memory consolidation and its extinction needs betterunderstanding (Zovkic and Sweatt, 2013; Jarome and Lubin, 2014; Diaset al., 2015).
The histone deacetylation (Peixoto and Abel, 2013; Kim and Kaang,2017; Sun et al., 2013, Feng et al., 2007) is catalyzed by certain class ofenzymes histone deacetylases (HDACs) which negatively regulates
https://doi.org/10.1016/j.brainresbull.2019.05.011Received 26 November 2018; Received in revised form 2 April 2019; Accepted 15 May 2019
⁎ Corresponding author at: Department of Biotechnology, Babasaheb Bhimrao Ambedkar University, Lucknow, India.E-mail address: [email protected] (A. Prakash).
1 Both have equal authorship.
Brain Research Bulletin 150 (2019) 86–101
Available online 17 May 20190361-9230/ © 2019 Published by Elsevier Inc.
T
cognition in multiple brain regions (Gallinari et al., 2007, Chen et al.,2015; Korzus et al., 2004, Guan et al., 2009; Schneider et al., 2013;Penney and Tsai, 2014). Different HDAC subtypes (HDAC1, HDAC2,HDAC3, HDAC4 etc.) have been shown to control gene silencing indifferent brain functions (Gregoretti et al., 2004; Fischer et al., 2010).Class I HDACs (HDAC1, HDAC2, HDAC3 and HDAC8) play an im-portant role in consolidation of fear memory (Bowers et al., 2015).HDAC1 expression has been associated with various types of neurolo-gical disorders (Bahari-Javan et al., 2017). It also regulates the ex-pression of c-fos (Yang et al., 2001; Usenko et al., 2003; Renthal et al.,2008), an IEG whose expression is upregulated following fear con-ditioning (Radulovic et al., 1998; Peleg et al., 2010), and down regu-lated following extinction in CA1 subregion of ventral hippocampus(Tronson et al., 2009). A study by Bahari-Javan et al., 2012 showshippocampal HDAC1 to be a positive regulator of fear extinction inmice. Another study by Guan et al., 2009 in HDAC2 knockout miceshow enhanced context and cued fear memory. In other rodent studies,HDAC2 activity has been found to be negatively correlated to con-solidation of fear extinction (Morris et al., 2013; Gräff et al., 2012).HDAC inhibitors (e. g. TSA, SAHA) have been found to enhance ex-tinction of both the contextual and cued fear memory (Itzhak et al.,2012; Maddox et al., 2013a, 2013b), suggestive of their application inprolonging extinction. Moreover targeting HDACs may provide an ex-cellent target for the treatment of PTSD (Singewald et al., 2015; Zovkicand Sweatt, 2013).
In the present study, we looked at the roles of HDACs (HDAC1 andHDAC2) in consolidation and extinction of fear memories. We workedtowards the hypothesis that histone acetylation required towards con-solidation of fear and extinction memories may be intricately balancedby the HDAC1 and HDAC2 in a region or subregion specific manner andmay be controlling the behavioral outcomes observed following fearand extinction learning. Any dysregulation in this circuitry may resultin improper histone acetylation and abnormal disinhibition of fear infear related anxiety disorders.
2. Materials and methods
2.1. Animals
All the experiments were performed on 2–3 months old healthyadult male SD (Sprague–Dawley) rats. The animals were housed understandard conditions of 12-h light/dark cycle with the temperaturemaintained at 23 °C, water, and food available ad libitum. All the ex-periments in the study were approved by Institutional Animal EthicsCommittee and performed as per the CPCSEA guidelines (853/AC/04/CPCSEA), Govt. of India, New Delhi.
2.2. Apparatus for behavioral study
For behavior training, two identical sound proof chambers made upof plexiglass (VJ instrument) were used for fear and extinction learning.The floor of the chamber consisted of stainless steel rods (4 mm, dia-meter), 1.5 cm spaced apart with arrangement for delivery of footshock(US). Context A, was used for conditioning, and Context B (havingdifferent olfaction, flooring and appearance) was used for extinction.Acoustic CS was delivered by a speaker fitted outside the chamber.Ventilation fans provided background noise in the acoustic chambers.
2.3. Fear learning
Fear learning in rats was performed by pairing tone (CS) with anoxious mild shock (US). On day 1, rats were exposed to context A for3min followed by fear conditioning which consisted of five consecutivetrials of CS (tone 80 dB, total duration 10 s) co-terminating with the US(1 s footshock 0.7 mA, intertrial interval : 60 s, Chang et al., 2009)(Fig. 1.A). The conditioning was measured as freezing percent (Freezingis defined as absence of all body movement except respiration).Freezing response was videotaped as well as calculated offline by re-cording the overall time which was spent in 10-second tone CS and 60 sITI. All animals were then returned back to their home cages following1min post-experiment. Additional group of same-aged rats which did
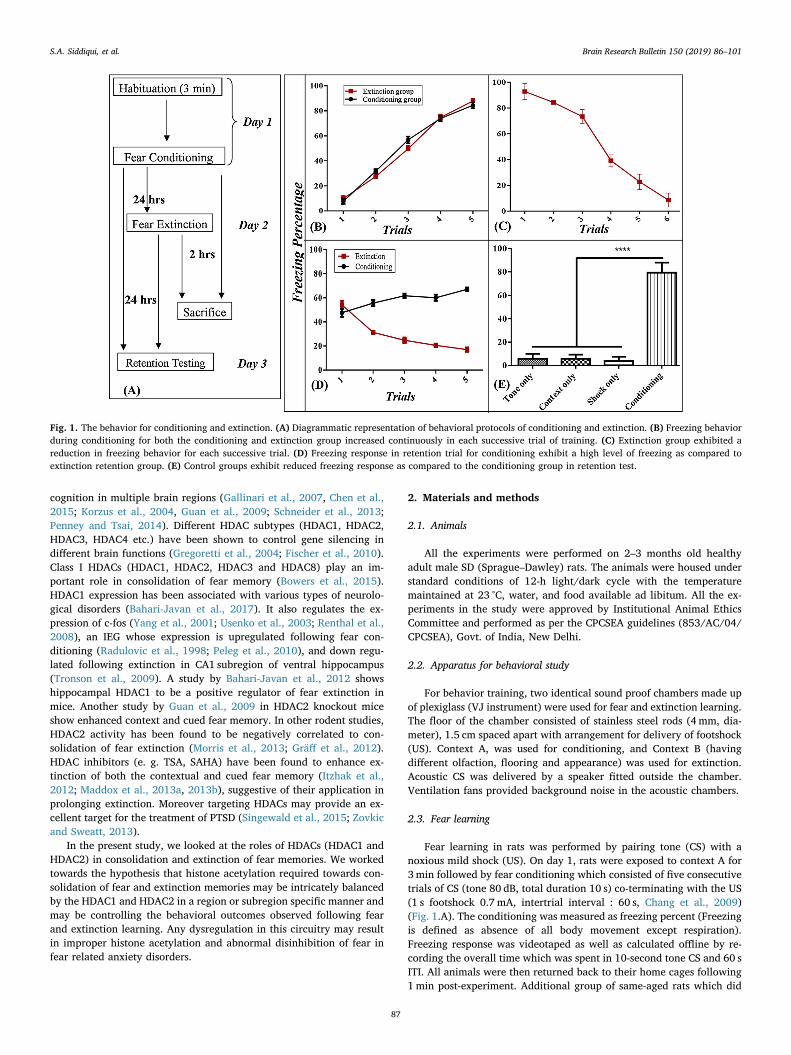
Fig. 1. The behavior for conditioning and extinction. (A) Diagrammatic representation of behavioral protocols of conditioning and extinction. (B) Freezing behaviorduring conditioning for both the conditioning and extinction group increased continuously in each successive trial of training. (C) Extinction group exhibited areduction in freezing behavior for each successive trial. (D) Freezing response in retention trial for conditioning exhibit a high level of freezing as compared toextinction retention group. (E) Control groups exhibit reduced freezing response as compared to the conditioning group in retention test.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
87
not undergo any experimental manipulations, were used as a naïvecontrol. To remove any confounding effect; tone only, shock only andcontext only groups were included for behavioral study (n=8–10).Animals from each group were sacrificed 2 h following experiment.(Fig. 1.A)
2.4. Fear extinction
24 h following fear learning, fear extinction was performed in anovel context (context B). Following 3min of acclimatization period,extinction training was performed by administration of 30 trials of CS(tone, 80 dB, duration 10 s, intertrial interval: 10 s) without presenta-tion of US (Fig. 1A). The ITI for extinction was set to 10 s, as it results ina strong extinction learning (Teichner and Holder, 1952; Laborda and
Miller, 2013). Rats (n=10–12 per group) were sacrificed for im-munohistochemical analysis 2 h after extinction training.
Some animals from conditioning and extinction groups underwentretention test 24 h after training. Retention test was performed with thepresentation of the CS (5 tones, duration 10 s, 80 dB, inter-trial interval:10 s) in context B in absence of foot shock. The rats, which were usedfor IHC (immunohistochemistry), did not experience retention test. Atotal of N= 80–96 rats were used in this study with 10–12 animals ineach group. (Fig. 1.A)
2.5. Details of brain subregions under study
In the present study we looked at the region specific changes withinamygdala and prefrontal cortex. These brain regions have been
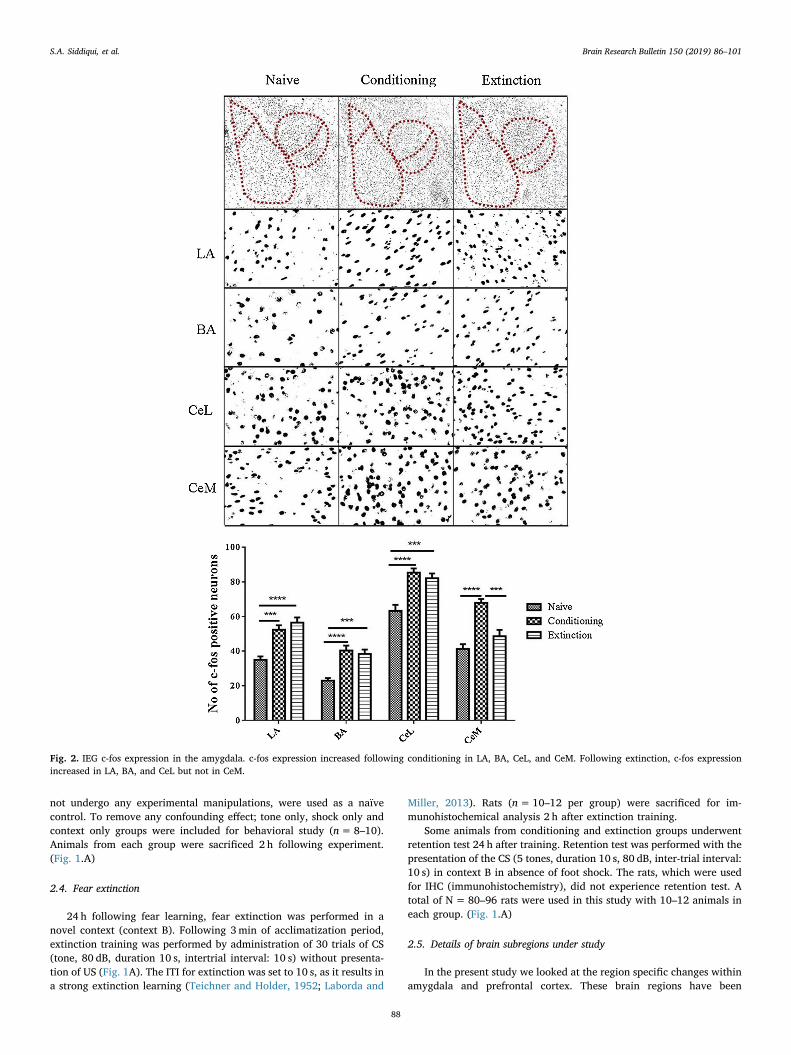
Fig. 2. IEG c-fos expression in the amygdala. c-fos expression increased following conditioning in LA, BA, CeL, and CeM. Following extinction, c-fos expressionincreased in LA, BA, and CeL but not in CeM.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
88
implicated in consolidation and extinction of fear memory. Amygdalaconsists of LA (lateral amygdala), BA (basal amygdala), CeL (cen-trolateral amygdala), and CeM (centromedial amygdala) (McDonald,1998; Turner and Herkenham, 1991; Krettek and Price, 1978; Petrovichand Swanson, 1997; Veening et al., 1984). The Prefrontal cortex (PFC)includes PL (Prelimbic prefrontal cortex) and IL (Infralimbic prefrontalcortex), regulating the expression and suppression of fear memory inrodents, respectively (Giustino and Maren, 2015).
2.6. Tissue preparation for immunohistochemistry
Two hours after fear or extinction learning, rats were anesthetizedwith pentobarbital (60mg/kg, i.p.), perfused transcardially with n-saline, followed by ice-cold 4% paraformaldehyde (in 0.1 M phosphatebuffer, pH 7.4) decapitated and brains removed. These brains were thenpost-fixed in 4% paraformaldehyde for 24 h and then in 10%, 20% and30% sucrose solution (in 0.1 M phosphate buffer, pH 7.4) serially tillthey settled. Brains were then frozen in isopentane at -300 to -350C for30min and kept at -800C for immunohistochemistry.
2.7. Immunohistochemistry
20 μm coronal brain sections containing PFC and Amygdala werecollected serially using cryostat (Microm HM 525, Germany) from thecontrols as well as from the conditioned and the extinction groups foreach antibody. These sections after being washed with 0.01M PBS wereblocked in PBST (0.01M, Phosphate buffer saline and 0.25% tween 20)containing 1% normal horse serum (NHS Vecta-stain Elite ABC kit,Vector Laboratories, Burlingame, CA, USA), 0.25% tween 20. Thesesections were then incubated overnight at room temperature with antic-fos (ab7963, Abcam), anti acetyl H3K9 (ab10812, Abcam), anti acetylH4K5 (H5110-15E2, US Biologicals), anti HDAC1 (H1827-51 J, USBiologicals) and anti HDAC2 (H5109-47E, US Biologicals) primaryantibodies (rabbit monoclonal, 1:500, 1:1000, 1:1000, 1:200 and
1:1000 dilutions respectively). Next day these sections were incubatedwith biotinylated secondary antibody (anti-rabbit IgG, 1:500 dilution,Vector Laboratories) for 2 h at room temperature. Finally the sectionsincubated with avidin-biotinylated-peroxidase complex (PK-6200,Vectastain Elite ABC Kit; Vector Laboratories) and followed by DABstaining (ab64238, DAB peroxidase substrate, Abcam). Stained sectionswere mounted on glass slides and images from the sections were ac-quired using a Nikon Eclipse Ni microscope (Nikon, Tokyo, Japan).Expression was analyzed as number of positive nuclei in amygdala andPFC subregions of the rat brain using the NIS-Basic Research imageanalysis system (Nikon, Tokyo).
2.8. Data analysis
The data was expressed as means and standard error of the means(± SEM), of no. of positive neurons counted in each area in matchedsections and analyzed by one-way ANOVA, two-way ANOVA or stu-dent’s t-test using GraphPad Prism 6 statistical software.
3. Results
3.1. Behavior
There was a similar increment in freezing response after each con-secutive trial during fear learning in both the conditioned and extinc-tion groups. Two-way ANOVA analysis revealed a significant effect oftrials [F(4,72)= 374.4 (p < 0.0001)] with no significant effect ofgroup [F(1,18) = 2.091 (p > 0.05)]. Bonferroni’s post-hoc test con-firmed significant differences between the consecutive trials (allp < 0.001). Freezing response during the last trial was similar(p > 0.05) and significantly higher than freezing response during theinitial trial (p < 0.001) in both the groups (Fig. 1.B).
During extinction learning there was a decrement in freezing re-sponse following each successive trial block. One-way ANOVA analysis
Table 1Correlation of Conditioning with the molecular profiles.
Acetyl H3K9 Acetyl H4K5 HDAC1 HDAC2 c-fos
LA r=0.877 (**)p < 0.001
r= 0.453 (*)p < 0.05
r= 0.898 (**)p < 0.001
r=− 0.696 (**)p < 0.001
r=0.817 (**)p < 0.001
BA r=0.897 (**)p < 0.001
– r= 0.703 (**)p < 0.001
r=− 0.614 (*)p < 0.05
r=0.655 (**)p < 0.05
CeL r=0.755 (**)p < 0.01
r= 0.498 (*)p < 0.05
r= 0.881 (**)p < 0.001
r=− 0.813 (**)p < 0.001
r=0.633 (**)p < 0.05
CeM r=0.904 (**)p < 0.001
r= 0.789 (**)p < 0.001
r=− 0.710 (**)p < 0.001
r=− 0.753 (**)p < 0.001
r= 0.827 (**)p < 0.001
PL r=0.854 (**)p < 0.001
r= 0.723 (**)p < 0.001
r=− 0.819 (**)p < 0.001
r=− 0.574 (**)p < 0.01
r=0.839 (**)p < 0.001
IL – – r= 0.620 (**)p < 0.01
r=− 0.684 (**)p < 0.001
–
Table 2Correlation of Extinction learning with the molecular profiles.
Acetyl H3K9 Acetyl H4K5 HDAC1 HDAC2 c-fos
LA r=− 0.659 (**)p < 0.01
r=− 0.523 (*)p < 0.05
r=− 0.907 (**)p < 0.001
r= 0.748 (**)p < 0.001
r=− 0.730 (**)p < 0.001
BA r=−0.831 (**)p < 0.001
– r=− 0.579 (**)p < 0.01
r= 0.743 (**)p < 0.001
r=− 0.776 (**)p < 0.001
CeL r=− 0.734 (**)p < 0.001
r=−0.717 (**)p < 0.001
r=− 0.794 (**)p < 0.001
r= 0.655 (**)p < 0.01
r=−0.612 (**)p < 0.01
CeM – – r=− 0.822 (**)p < 0.001
– –
PL – – r=− 0.786 (**)p < 0.001
r=−0.647 (**)p < 0.01
–
IL r=− 0.874 (**)p < 0.001
r=−0.542 (*)p < 0.05
r= 0.896 (**)p < 0.001
r=−0.816 (**)p < 0.001
r=− 0.600 (**)p < 0.01
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
89
showed significant difference between trials [F (4, 25)= 299(p < 0.0001)]. Bonferroni’s post-hoc test confirmed significant differ-ences between trials (all p < 0.05). Minimal freezing was observed inthe last trial and it was significantly lower as compared to the freezingobserved in the first trial block during extinction learning (p < 0.0001;Fig. 1.C).
During the retention test both the groups (conditioning and ex-tinction) exhibited similar freezing during initial trail. The next 4 trialswere different from each other (Fig. 1.D). Two-way ANOVA revealed asignificant main effect of group [F (1, 18)= 510.3 (p < 0.0001)] andtrails [F (4, 72) = 10.92 (p < 0.0001)]. Bonferroni’s post hoc com-parison confirmed a significant difference between the trials (allp < 0.001, trial 2–5). The percent freezing was significantly higher inthe conditioning group than the extinction group (p < 0.0001). Thetone only, the shock only and context only control groups exhibitedvery low freezing response as compared to the conditioning group foreach trial (Fig. 1. E). One-way ANOVA showed a significant effect ofgroup [F (4, 45)= 157.9 (p < 0.0001)]. Bonferroni’s post hoc com-parison showed a significant difference in percent freezing between the
conditioning and the control groups including the extinction group (allp < 0.0001).
3.2. Immunohistochemistry
3.2.1. IEG c-fos expression in amygdalaThere was a differential expression of the IEG c-fos in LA, BA, CeL
and CeM following fear and extinction learning when compared to thenaive control. The one-way ANOVA analysis confirmed the effect ofgroup on c-fos expression in LA [F(2,27)= 19.96, p < 0.0001], BA [F(2,27) = 16.42, p < 0.0001], CeL [F(2,27) = 16.79, p < 0.0001] andCeM [F(2,27) = 21.36, p < 0.0001]. Bonferroni’s post hoc analysisconfirmed a significant increase in the number of c-fos positive neuronsin LA (p < 0.001, p < 0.0001), BA (p < 0.0001, p < 0.001) and CeL(p < 0.0001, p < 0.001) following both fear and extinction learningand in CeM (p < 0.0001) following fear learning only (Fig. 2).
Furthermore the c-fos expression LA (p < 0.001), BA (p < 0.05),CeL (p < 0.05) and CeM (p < 0.001) in the conditioning group posi-tively correlated to percent freezing following fear learning and the c-
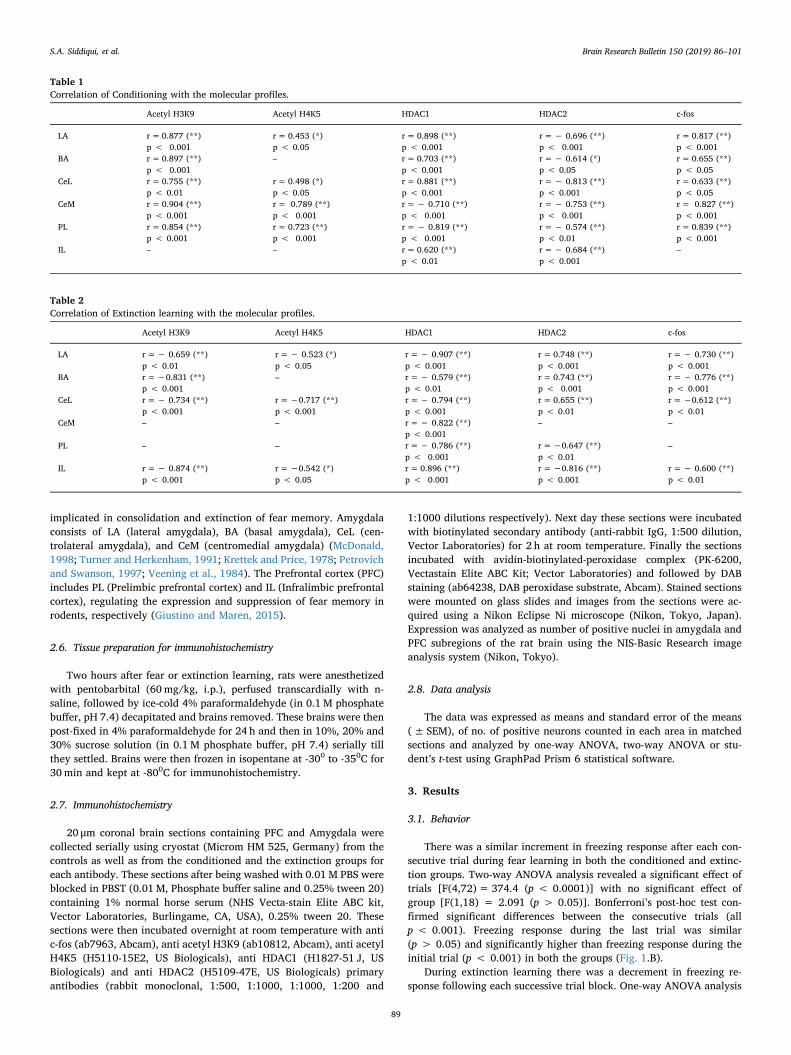
Fig. 3. Acetyl H3K9 level in the amygdala. A significant increase in acetyl H3K9 observed in LA, BA, CeL, and CeM following conditioning while following extinctiona significantly increased expression of acetyl H3K9 observed in LA, BA and CeL but not in CeM.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
90
fos expression in LA (p < 0.001), BA (p < 0.001) and CeL (p < 0.01)negatively correlated with the percent freezing during extinctionlearning (Table 1 and 2 ).
3.2.2. Histone acetylation in amygdalaThe acetylation of histone H3 at K9 was analyzed to find out any
association of freezing response observed following fear and extinctionlearning. One-way ANOVA analysis confirmed the effect of group onacetylation of H3 in LA [F (2, 24)= 19.55, p < 0.0001], BA [F(2, 27)
= 31.61, p < 0.0001], CeL [F(2, 33) = 15.04, p < 0.0001] and CeM[F(2,33) = 74.94, p < 0.0001]. Bonferroni’s post hoc analysis con-firmed that both fear and extinction learning induced a significant in-crease in the acetylation of H3 following both fear and extinctionlearning in LA (p < 0.0001, p < 0.001), BA (p < 0.0001,p < 0.0001) and CeL (p < 0.001, p < 0.001). However a significantAcetylation of H3 in CeM (p < 0.0001) was observed only followingfear learning only. (Fig. 3)
The histone acetylation of H3 at K9 in LA (p < 0.001), BA
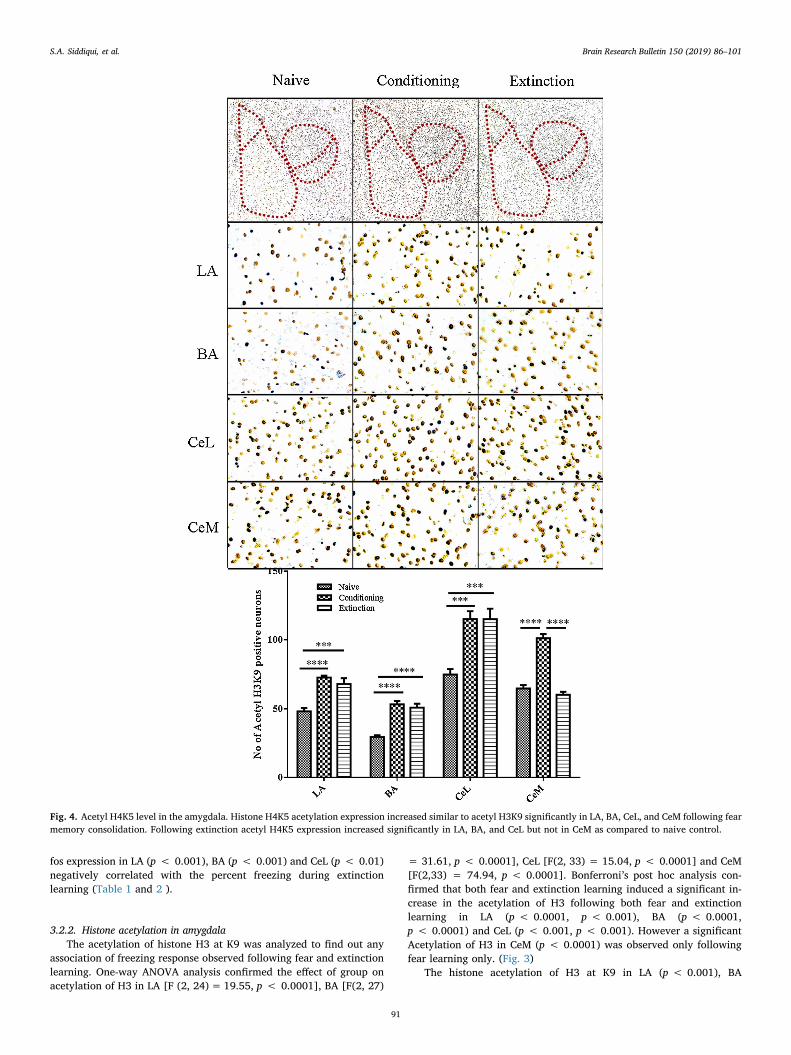
Fig. 4. Acetyl H4K5 level in the amygdala. Histone H4K5 acetylation expression increased similar to acetyl H3K9 significantly in LA, BA, CeL, and CeM following fearmemory consolidation. Following extinction acetyl H4K5 expression increased significantly in LA, BA, and CeL but not in CeM as compared to naive control.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
91
(p < 0.001), CeL (p < 0.01) and CeM (p < 0.001) was positivelycorrelated with the percent freezing during fear learning (Table 1). Theextinction induced acetylation of H3 at k9 in LA (p < 0.01), BA(p < 0.001) and CeL (p < 0.001) showed a negative correlation withthe freezing behavior, whereas the acetylation in CeM (p > 0.05) hadno correlation with the freezing response during extinction (Table 2).
Similar to the acetylation of H3 at K9, there was a significant changein the acetylation of H4 at K5 in LA, BA, CeL and CeM in conditioningand extinction group as compared to the naive control. One-wayANOVA analysis revealed effect of group on acetylation of H4K5 in LA[F(2, 45)= 24.58, p < 0.0001], BA [F(2, 44) = 6.251, p < 0.01], CeL[F(2, 44) = 17.33, p < 0.0001] and CeM [F(2, 35) = 40.45,p < 0.0001]. Bonferroni’s post hoc analysis confirmed that the acet-ylation of H4 at K5 in LA (p < 0.001, p < 0.001), BA (p < 0.05,p < 0.01) and CeL (p < 0.0001, p < 0.0001) increased in both con-ditioning and extinction groups while in CeM (p < 0.0001) increasedonly in conditioning group. (Fig. 4)
Following fear learning histone acetylation at H4K5 was positivelycorrelated with the freezing response during conditioning in LA(p < 0.05), CeL (p < 0.05) and CeM (p < 0.001) (Table 1). On theother hand, during extinction the LA (p < 0.05) and CeL (p < 0.001)exhibited a negative correlation with the freezing response (Table 2)following extinction.
3.2.3. HDAC1 expression in amygdalaFurther, the HDAC1 expression was analyzed in amygdala to find
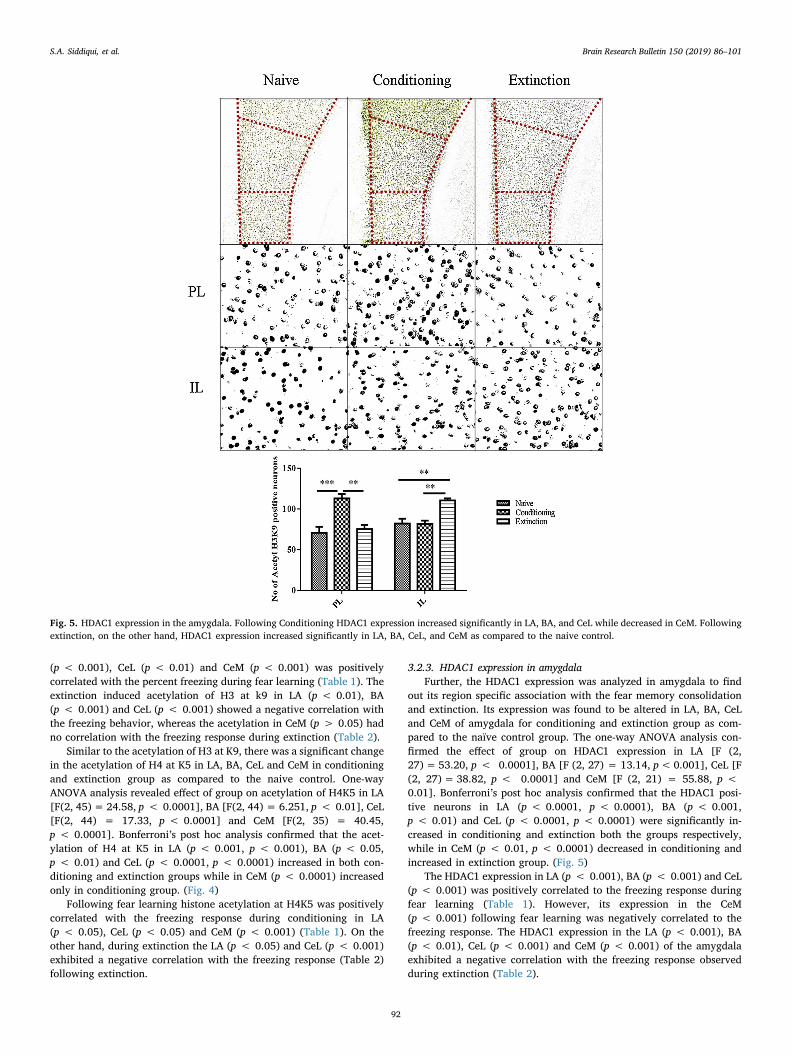
out its region specific association with the fear memory consolidationand extinction. Its expression was found to be altered in LA, BA, CeLand CeM of amygdala for conditioning and extinction group as com-pared to the naïve control group. The one-way ANOVA analysis con-firmed the effect of group on HDAC1 expression in LA [F (2,27)= 53.20, p < 0.0001], BA [F (2, 27) = 13.14, p<0.001], CeL [F(2, 27)= 38.82, p < 0.0001] and CeM [F (2, 21) = 55.88, p <0.01]. Bonferroni’s post hoc analysis confirmed that the HDAC1 posi-tive neurons in LA (p < 0.0001, p < 0.0001), BA (p < 0.001,p < 0.01) and CeL (p < 0.0001, p < 0.0001) were significantly in-creased in conditioning and extinction both the groups respectively,while in CeM (p < 0.01, p < 0.0001) decreased in conditioning andincreased in extinction group. (Fig. 5)
The HDAC1 expression in LA (p < 0.001), BA (p < 0.001) and CeL(p < 0.001) was positively correlated to the freezing response duringfear learning (Table 1). However, its expression in the CeM(p < 0.001) following fear learning was negatively correlated to thefreezing response. The HDAC1 expression in the LA (p < 0.001), BA(p < 0.01), CeL (p < 0.001) and CeM (p < 0.001) of the amygdalaexhibited a negative correlation with the freezing response observedduring extinction (Table 2).
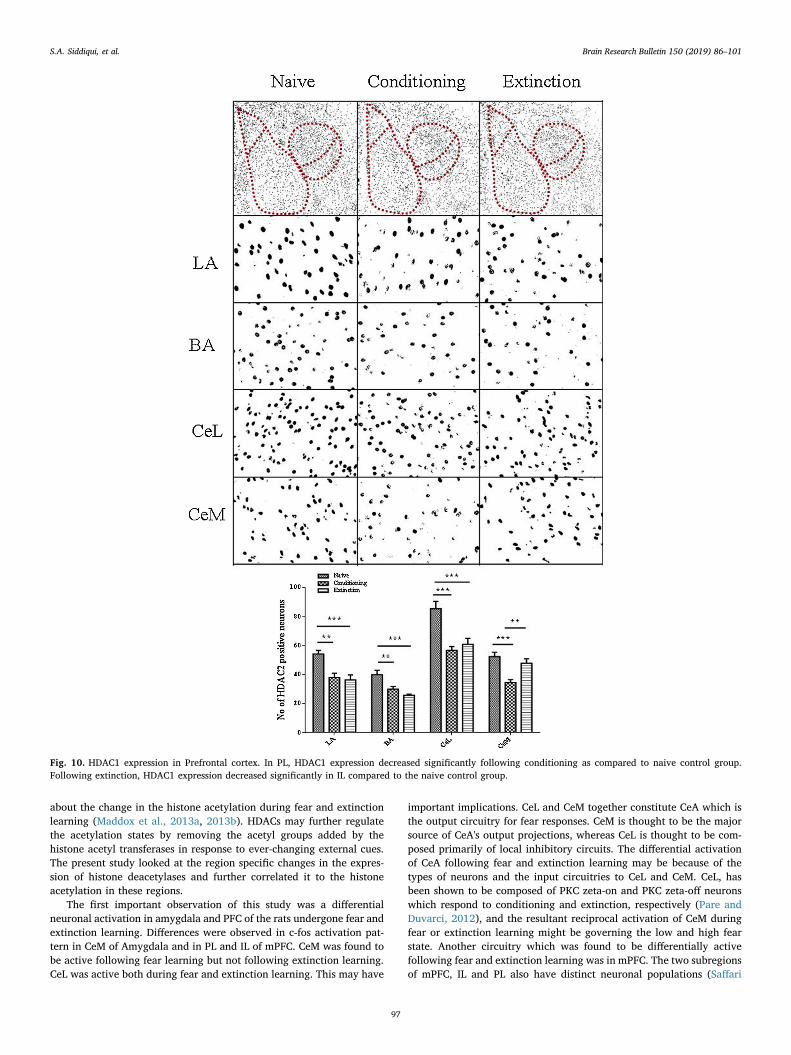
Fig. 5. HDAC1 expression in the amygdala. Following Conditioning HDAC1 expression increased significantly in LA, BA, and CeL while decreased in CeM. Followingextinction, on the other hand, HDAC1 expression increased significantly in LA, BA, CeL, and CeM as compared to the naive control.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
92
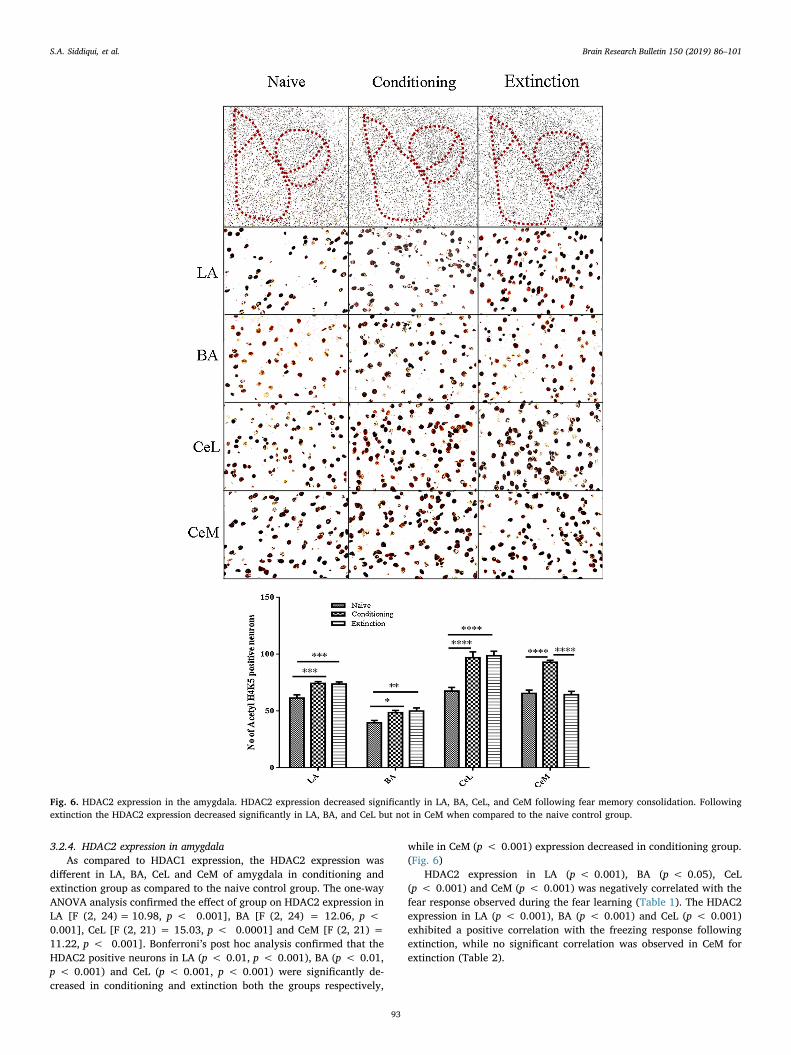
3.2.4. HDAC2 expression in amygdalaAs compared to HDAC1 expression, the HDAC2 expression was
different in LA, BA, CeL and CeM of amygdala in conditioning andextinction group as compared to the naive control group. The one-wayANOVA analysis confirmed the effect of group on HDAC2 expression inLA [F (2, 24)= 10.98, p < 0.001], BA [F (2, 24) = 12.06, p <0.001], CeL [F (2, 21) = 15.03, p < 0.0001] and CeM [F (2, 21) =11.22, p < 0.001]. Bonferroni’s post hoc analysis confirmed that theHDAC2 positive neurons in LA (p < 0.01, p < 0.001), BA (p < 0.01,p < 0.001) and CeL (p < 0.001, p < 0.001) were significantly de-creased in conditioning and extinction both the groups respectively,
while in CeM (p < 0.001) expression decreased in conditioning group.(Fig. 6)
HDAC2 expression in LA (p < 0.001), BA (p < 0.05), CeL(p < 0.001) and CeM (p < 0.001) was negatively correlated with thefear response observed during the fear learning (Table 1). The HDAC2expression in LA (p < 0.001), BA (p < 0.001) and CeL (p < 0.001)exhibited a positive correlation with the freezing response followingextinction, while no significant correlation was observed in CeM forextinction (Table 2).
Fig. 6. HDAC2 expression in the amygdala. HDAC2 expression decreased significantly in LA, BA, CeL, and CeM following fear memory consolidation. Followingextinction the HDAC2 expression decreased significantly in LA, BA, and CeL but not in CeM when compared to the naive control group.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
93
3.2.5. IEG c-fos expression in prefrontal cortexPL and IL exhibited a differential activation pattern for c-fos ex-
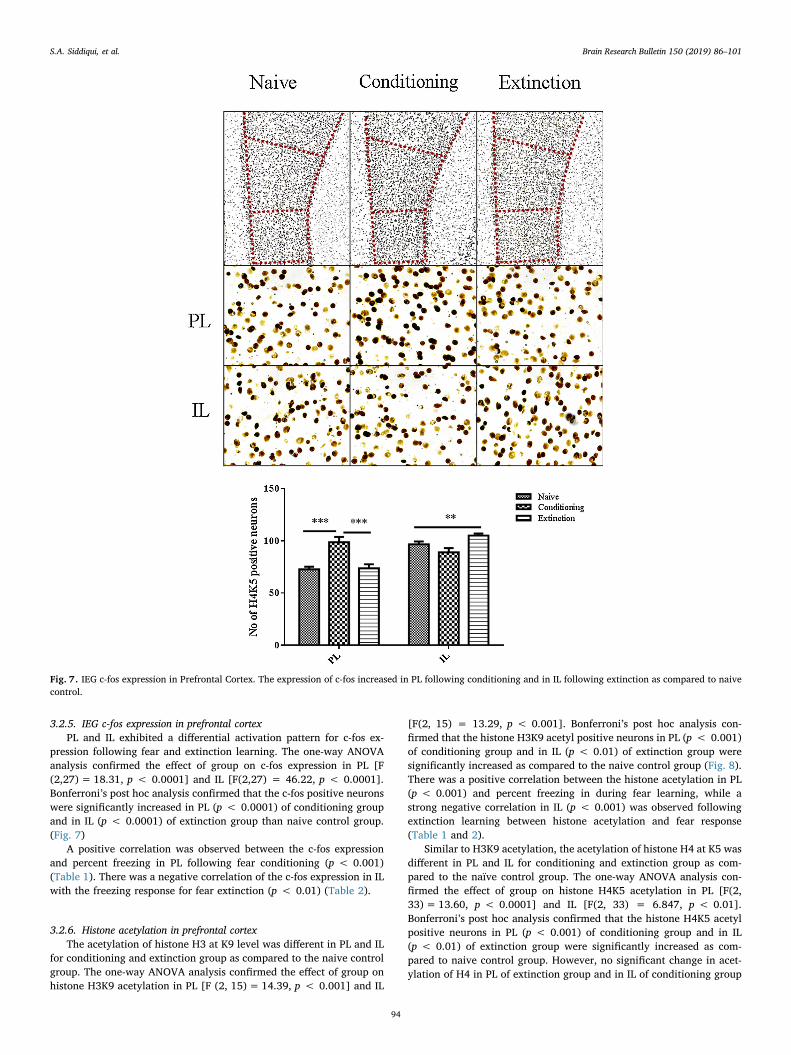
pression following fear and extinction learning. The one-way ANOVAanalysis confirmed the effect of group on c-fos expression in PL [F(2,27)= 18.31, p < 0.0001] and IL [F(2,27) = 46.22, p < 0.0001].Bonferroni’s post hoc analysis confirmed that the c-fos positive neuronswere significantly increased in PL (p < 0.0001) of conditioning groupand in IL (p < 0.0001) of extinction group than naive control group.(Fig. 7)
A positive correlation was observed between the c-fos expressionand percent freezing in PL following fear conditioning (p < 0.001)(Table 1). There was a negative correlation of the c-fos expression in ILwith the freezing response for fear extinction (p < 0.01) (Table 2).
3.2.6. Histone acetylation in prefrontal cortexThe acetylation of histone H3 at K9 level was different in PL and IL
for conditioning and extinction group as compared to the naive controlgroup. The one-way ANOVA analysis confirmed the effect of group onhistone H3K9 acetylation in PL [F (2, 15)= 14.39, p < 0.001] and IL
[F(2, 15) = 13.29, p < 0.001]. Bonferroni’s post hoc analysis con-firmed that the histone H3K9 acetyl positive neurons in PL (p < 0.001)of conditioning group and in IL (p < 0.01) of extinction group weresignificantly increased as compared to the naive control group (Fig. 8).There was a positive correlation between the histone acetylation in PL(p < 0.001) and percent freezing in during fear learning, while astrong negative correlation in IL (p < 0.001) was observed followingextinction learning between histone acetylation and fear response(Table 1 and 2).
Similar to H3K9 acetylation, the acetylation of histone H4 at K5 wasdifferent in PL and IL for conditioning and extinction group as com-pared to the naïve control group. The one-way ANOVA analysis con-firmed the effect of group on histone H4K5 acetylation in PL [F(2,33)= 13.60, p < 0.0001] and IL [F(2, 33) = 6.847, p < 0.01].Bonferroni’s post hoc analysis confirmed that the histone H4K5 acetylpositive neurons in PL (p < 0.001) of conditioning group and in IL(p < 0.01) of extinction group were significantly increased as com-pared to naive control group. However, no significant change in acet-ylation of H4 in PL of extinction group and in IL of conditioning group
Fig. 7. IEG c-fos expression in Prefrontal Cortex. The expression of c-fos increased in PL following conditioning and in IL following extinction as compared to naivecontrol.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
94
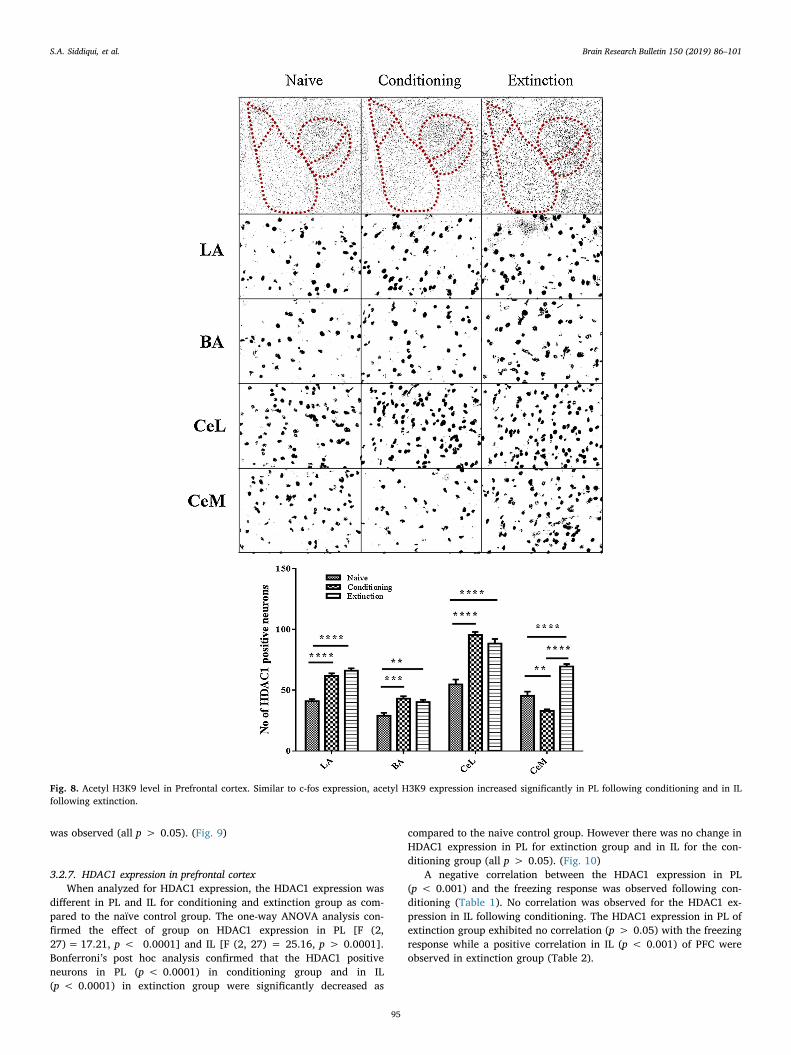
was observed (all p > 0.05). (Fig. 9)
3.2.7. HDAC1 expression in prefrontal cortexWhen analyzed for HDAC1 expression, the HDAC1 expression was
different in PL and IL for conditioning and extinction group as com-pared to the naïve control group. The one-way ANOVA analysis con-firmed the effect of group on HDAC1 expression in PL [F (2,27)= 17.21, p < 0.0001] and IL [F (2, 27) = 25.16, p > 0.0001].Bonferroni’s post hoc analysis confirmed that the HDAC1 positiveneurons in PL (p < 0.0001) in conditioning group and in IL(p < 0.0001) in extinction group were significantly decreased as
compared to the naive control group. However there was no change inHDAC1 expression in PL for extinction group and in IL for the con-ditioning group (all p > 0.05). (Fig. 10)
A negative correlation between the HDAC1 expression in PL(p < 0.001) and the freezing response was observed following con-ditioning (Table 1). No correlation was observed for the HDAC1 ex-pression in IL following conditioning. The HDAC1 expression in PL ofextinction group exhibited no correlation (p > 0.05) with the freezingresponse while a positive correlation in IL (p < 0.001) of PFC wereobserved in extinction group (Table 2).
Fig. 8. Acetyl H3K9 level in Prefrontal cortex. Similar to c-fos expression, acetyl H3K9 expression increased significantly in PL following conditioning and in ILfollowing extinction.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
95
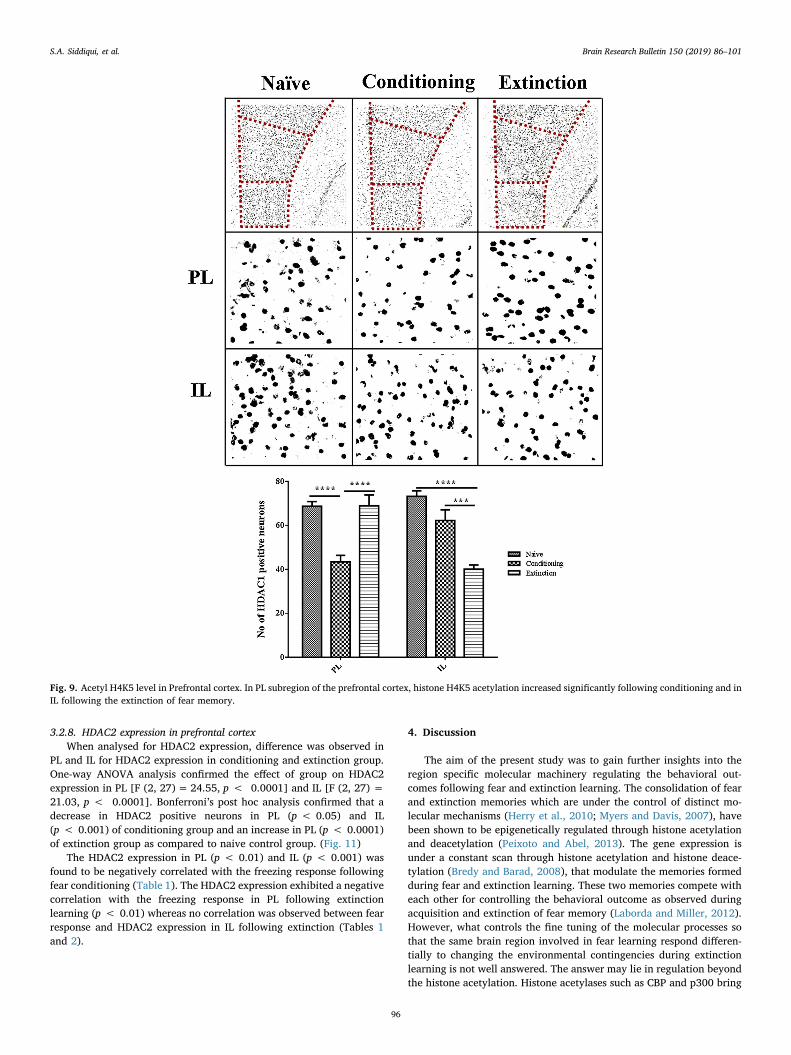
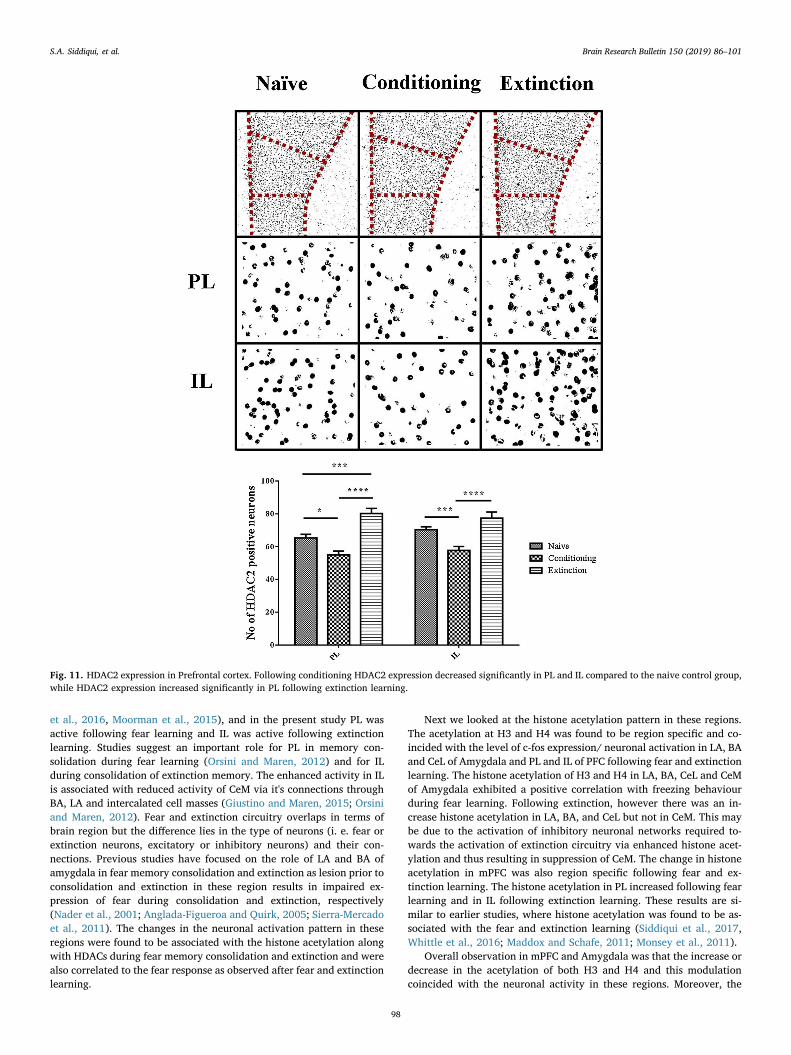
3.2.8. HDAC2 expression in prefrontal cortexWhen analysed for HDAC2 expression, difference was observed in
PL and IL for HDAC2 expression in conditioning and extinction group.One-way ANOVA analysis confirmed the effect of group on HDAC2expression in PL [F (2, 27)= 24.55, p < 0.0001] and IL [F (2, 27) =21.03, p < 0.0001]. Bonferroni’s post hoc analysis confirmed that adecrease in HDAC2 positive neurons in PL (p < 0.05) and IL(p < 0.001) of conditioning group and an increase in PL (p < 0.0001)of extinction group as compared to naive control group. (Fig. 11)
The HDAC2 expression in PL (p < 0.01) and IL (p < 0.001) wasfound to be negatively correlated with the freezing response followingfear conditioning (Table 1). The HDAC2 expression exhibited a negativecorrelation with the freezing response in PL following extinctionlearning (p < 0.01) whereas no correlation was observed between fearresponse and HDAC2 expression in IL following extinction (Tables 1and 2).
4. Discussion
The aim of the present study was to gain further insights into theregion specific molecular machinery regulating the behavioral out-comes following fear and extinction learning. The consolidation of fearand extinction memories which are under the control of distinct mo-lecular mechanisms (Herry et al., 2010; Myers and Davis, 2007), havebeen shown to be epigenetically regulated through histone acetylationand deacetylation (Peixoto and Abel, 2013). The gene expression isunder a constant scan through histone acetylation and histone deace-tylation (Bredy and Barad, 2008), that modulate the memories formedduring fear and extinction learning. These two memories compete witheach other for controlling the behavioral outcome as observed duringacquisition and extinction of fear memory (Laborda and Miller, 2012).However, what controls the fine tuning of the molecular processes sothat the same brain region involved in fear learning respond differen-tially to changing the environmental contingencies during extinctionlearning is not well answered. The answer may lie in regulation beyondthe histone acetylation. Histone acetylases such as CBP and p300 bring
Fig. 9. Acetyl H4K5 level in Prefrontal cortex. In PL subregion of the prefrontal cortex, histone H4K5 acetylation increased significantly following conditioning and inIL following the extinction of fear memory.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
96
about the change in the histone acetylation during fear and extinctionlearning (Maddox et al., 2013a, 2013b). HDACs may further regulatethe acetylation states by removing the acetyl groups added by thehistone acetyl transferases in response to ever-changing external cues.The present study looked at the region specific changes in the expres-sion of histone deacetylases and further correlated it to the histoneacetylation in these regions.
The first important observation of this study was a differentialneuronal activation in amygdala and PFC of the rats undergone fear andextinction learning. Differences were observed in c-fos activation pat-tern in CeM of Amygdala and in PL and IL of mPFC. CeM was found tobe active following fear learning but not following extinction learning.CeL was active both during fear and extinction learning. This may have
important implications. CeL and CeM together constitute CeA which isthe output circuitry for fear responses. CeM is thought to be the majorsource of CeA's output projections, whereas CeL is thought to be com-posed primarily of local inhibitory circuits. The differential activationof CeA following fear and extinction learning may be because of thetypes of neurons and the input circuitries to CeL and CeM. CeL, hasbeen shown to be composed of PKC zeta-on and PKC zeta-off neuronswhich respond to conditioning and extinction, respectively (Pare andDuvarci, 2012), and the resultant reciprocal activation of CeM duringfear or extinction learning might be governing the low and high fearstate. Another circuitry which was found to be differentially activefollowing fear and extinction learning was in mPFC. The two subregionsof mPFC, IL and PL also have distinct neuronal populations (Saffari
Fig. 10. HDAC1 expression in Prefrontal cortex. In PL, HDAC1 expression decreased significantly following conditioning as compared to naive control group.Following extinction, HDAC1 expression decreased significantly in IL compared to the naive control group.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
97
et al., 2016, Moorman et al., 2015), and in the present study PL wasactive following fear learning and IL was active following extinctionlearning. Studies suggest an important role for PL in memory con-solidation during fear learning (Orsini and Maren, 2012) and for ILduring consolidation of extinction memory. The enhanced activity in ILis associated with reduced activity of CeM via it's connections throughBA, LA and intercalated cell masses (Giustino and Maren, 2015; Orsiniand Maren, 2012). Fear and extinction circuitry overlaps in terms ofbrain region but the difference lies in the type of neurons (i. e. fear orextinction neurons, excitatory or inhibitory neurons) and their con-nections. Previous studies have focused on the role of LA and BA ofamygdala in fear memory consolidation and extinction as lesion prior toconsolidation and extinction in these region results in impaired ex-pression of fear during consolidation and extinction, respectively(Nader et al., 2001; Anglada-Figueroa and Quirk, 2005; Sierra-Mercadoet al., 2011). The changes in the neuronal activation pattern in theseregions were found to be associated with the histone acetylation alongwith HDACs during fear memory consolidation and extinction and werealso correlated to the fear response as observed after fear and extinctionlearning.
Next we looked at the histone acetylation pattern in these regions.The acetylation at H3 and H4 was found to be region specific and co-incided with the level of c-fos expression/ neuronal activation in LA, BAand CeL of Amygdala and PL and IL of PFC following fear and extinctionlearning. The histone acetylation of H3 and H4 in LA, BA, CeL and CeMof Amygdala exhibited a positive correlation with freezing behaviourduring fear learning. Following extinction, however there was an in-crease histone acetylation in LA, BA, and CeL but not in CeM. This maybe due to the activation of inhibitory neuronal networks required to-wards the activation of extinction circuitry via enhanced histone acet-ylation and thus resulting in suppression of CeM. The change in histoneacetylation in mPFC was also region specific following fear and ex-tinction learning. The histone acetylation in PL increased following fearlearning and in IL following extinction learning. These results are si-milar to earlier studies, where histone acetylation was found to be as-sociated with the fear and extinction learning (Siddiqui et al., 2017,Whittle et al., 2016; Maddox and Schafe, 2011; Monsey et al., 2011).
Overall observation in mPFC and Amygdala was that the increase ordecrease in the acetylation of both H3 and H4 and this modulationcoincided with the neuronal activity in these regions. Moreover, the
Fig. 11. HDAC2 expression in Prefrontal cortex. Following conditioning HDAC2 expression decreased significantly in PL and IL compared to the naive control group,while HDAC2 expression increased significantly in PL following extinction learning.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
98

enhanced neuronal activity in PL was associated with increased fearresponse while the activity of IL with the fear extinction (Peters et al.,2009; Knapska and Maren, 2009). As expected the HDAC expressionwas not always negatively correlated with histone acetylation, theHDAC1 and HDAC2 expression was different during fear and extinctionlearning and expressed differentially in a region specific manner. Itcould be therefore speculated that initially during fear learning theincrease in the acetylation required for synaptic plasticity might havebeen under the control of HATs but once the environmental con-tingencies change during fear learning followed by extinction learning,the HDACs may taking over as the main players.
The most important finding of this study was the differential ex-pression of HDAC1 and HDAC2 in amygdala and PFC following fear andextinction learning. Their expression was differently associated with thehistone acetylation in a region specific manner. The HDAC1 expressionwas significantly high in BA, LA and CeL following fear and extinctionlearning. However, it was down regulated in CeM and PL following fearlearning and in IL following extinction learning. The increase and de-crease in the acetylation observed in CeM following conditioning andextinction, respectively seems to be associated with the HDAC1 ex-pression. Similarly, the increased acetylation in PL following fearlearning and in IL following extinction also seem to be associated withthe HDAC1 expression. The changes in Histone acetylation in the othersubregions such as BA, LA and CeL seem to be independent of theHDAC1 activity and HDAC1 might be associated with the suppressionof inhibitory components in these regions that affects conditioning andextinction. Earlier studies where the contextual and cued fear con-ditioning has been shown to be dependent on region specific changesfor the HDAC expression in the subregions of hippocampus andamygdala are in line with present study (Guan et al., 2009, Morris et al.,2013; Kwapis et al., 2017, Alaghband et al., 2017).
The HDAC2 expression which was significantly down regulated inLA, BA, CeM, CeL and PL following fear learning and in LA, BA, CeL andIL following extinction learning seems to be mostly associated with theregulation of histone acetylation in these regions. Guan et al., 2009have shown that HDAC2 binds to memory related genes in hippo-campus and its reduction is required for cued and contextual learning.However, another study by Bahari-Javan et al. (2012) confirmed thatthe binding of HDAC1 enhances extinction by reducing the expressionof memory related genes in hippocampus during contextual learning,leaving fear memory intact. In the current study while HDAC1 pro-moted extinction and conditioning through subregion specific neuronalactivation, the HDAC2 promoted conditioning and extinction throughsuppression in subregion specific neuronal population. The positiveassociation between HDAC1 expression and histone acetylation/ neu-ronal activation as exemplified by c-fos expression might be associatedwith the suppression of other genes that are involved in suppression ofmemory related genes (i.e. suppressing them) (e.g. PP1) (Graff et al.,2010; Waddell, 2003) or it is suppressing the inhibitory components offear circuitry and a negative association with the HDAC2 might be as-sociated with the suppression of memory related genes. At fear circuitrylevel the results suggests that the activation of LA, BA, CeL and CeM ofamygdala and PL, IL of PFC may be associated with the histone acet-ylation. This may be further regulated by the suppression of HDAC2 andactivation of HDAC1 in amygdala to accompany fear learning throughdifferential histone acetylation.
The current study only looked at the subregion specific role ofHDACs at fear circuitry level. Further studies to gauge the expression ofindividual HDACs at a single neuron level will be required to assignspecific roles of these epigenetic players. Guan et al., 2009 have alreadyreported different gene targets for the HDAC1 and 2, indicative ofdifferential recruitment of HDACs in fear and extinction learning. Ourresult suggests an overall reduction in the expression of HDAC2 andincreased expression of HDAC1 in the subregions of Amygdala and PFCtowards successful encryption of fear and extinction engram and furtherinsights will be required to find out to what extent both the HDACs
differ in their function and targets at brain subregion level.To our knowledge, this is the first study that directly assesses the
relationship between histone acetylation and its regulation throughHDACs. Our results suggest an alternate mechanism of region specificregulation of Histone Acetylation through differential expression ofHDAC1 and HDAC2, which have important implications for the un-derstanding and treatment of cognitive, psychiatric and neurodegen-erative disorders related to impaired histone acetylation. Future re-search will be helpful to clarify the exact mechanism by which theseHDAC subtypes regulate fear circuitry at cellular and gene level.
Conflict of interest
Author declares no conflict of interest.
Acknowledgement
The funding for the above work was provided by the Department ofBiotechnology, New Delhi, India, (BT/PR14365/MED/30/522/2012).
References
Alaghband, Y., Kwapis, J.L., López, A.J., White, A.O., Aimiuwu, O.V., Al-Kachak, A.,Bodinayake, K.K., Oparaugo, N.C., Dang, R., Astarabadi, M., Matheos, D.P., Wood,M.A., 2017. Distinct roles for the deacetylase domain of HDAC3 in the hippocampusand medial prefrontal cortex in the formation and extinction of memory. Neurobiol.Learn. Mem. 145, 94–104. https://doi.org/10.1016/j.nlm.2017.09.001.
Anglada-Figueroa, D., Quirk, G.J., 2005. Behavioral/Systems/Cognitive Lesions of theBasal Amygdala Block Expression of Conditioned Fear But Not Extinction. https://doi.org/10.1523/JNEUROSCI.2600-05.2005.
Bahari-Javan, S., Maddalena, A., Kerimoglu, C., Wittnam, J., Held, T., Bahr, M.,Burkhardt, S., Delalle, I., Kugler, S., Fischer, A., Sananbenesi, F., 2012. HDAC1 reg-ulates fear extinction in mice. J. Neurosci. 32, 5062–5073. https://doi.org/10.1523/JNEUROSCI.0079-12.2012.
Bahari-Javan, S., Varbanov, H., Halder, R., Benito, E., Kaurani, L., Burkhardt, S.,Anderson-Schmidt, H., Anghelescu, I., Budde, M., Stilling, R.M., Costa, J., Medina, J.,Dietrich, D.E., Figge, C., Folkerts, H., Gade, K., Heilbronner, U., Koller, M., Konrad,C., Nussbeck, S.Y., Scherk, H., Spitzer, C., Stierl, S., Stöckel, J., Thiel, A., von Hagen,M., Zimmermann, J., Zitzelsberger, A., Schulz, S., Schmitt, A., Delalle, I., Falkai, P.,Schulze, T.G., Dityatev, A., Sananbenesi, F., Fischer, A., 2017. HDAC1 links early lifestress to schizophrenia-like phenotypes. Proc. Natl. Acad. Sci. U. S. A. 114,E4686–E4694. https://doi.org/10.1073/pnas.1613842114.
Blouin, A.M., Sillivan, S.E., Joseph, N.F., Miller, C.A., 2016. The potential of epigeneticsin stress-enhanced fear learning models of PTSD. Learn. Mem. 23, 576–586. https://doi.org/10.1101/lm.040485.115.
Bowers, M.E., Xia, B., Carreiro, S., Ressler, K.J., 2015. The Class I HDAC inhibitorRGFP963 enhances consolidation of cued fear extinction. Learn. Mem. 22, 225–231.https://doi.org/10.1101/lm.036699.114.
Bredy, T.W., Barad, M., 2008. The histone deacetylase inhibitor valproic acid enhancesacquisition, extinction, and reconsolidation of conditioned fear. Learn. Mem. 39–45.https://doi.org/10.1101/lm.801108.derstanding.
Chang, C., Knapska, E., Orsini, C.A., Rabinak, C.A., Zimmerman, J.M., Maren, S., 2009.Fear extinction in rodents. Curr. Protoc. Neurosci. https://doi.org/10.1002/0471142301.ns0823s47. Chapter 8, Unit 8.23.
Chen, H.P., Zhao, Y.T., Zhao, T.C., 2015. Histone deacetylases and mechanisms of reg-ulation of gene expression. Crit. Rev. Oncog. 20, 35–47. https://doi.org/10.1615/CritRevOncog.2015012997.
Dias, B.G., Maddox, S., Klengel, T., Ressler, K.J., 2015. Epigenetic mechanisms underlyinglearning and the inheritance of learned behaviors. Trends Neurosci. 38, 96–107.https://doi.org/10.1016/j.tins.2014.12.003.
Feng, J., Fouse, S., Fan, G., 2007. Epigenetic regulation of neural gene expression andneuronal function. Pediatr. Res. 61, 58R–63R. https://doi.org/10.1203/pdr.0b013e3180457635.
Fischer, A., Sananbenesi, F., Mungenast, A., Tsai, L.-H., 2010. Targeting the correctHDAC(s) to treat cognitive disorders. Trends Pharmacol. Sci. 31, 605–617. https://doi.org/10.1016/j.tips.2010.09.003.
Gallinari, P., Di Marco, S., Jones, P., Pallaoro, M., Steinkühler, C., 2007. HDACs, histonedeacetylation and gene transcription: from molecular biology to cancer therapeutics.Cell Res. 17, 195–211. https://doi.org/10.1038/sj.cr.7310149.
Giustino, T.F., Maren, S., 2015. The role of the medial prefrontal cortex in the con-ditioning and extinction of fear. Front. Behav. Neurosci. 9, 298. https://doi.org/10.3389/fnbeh.2015.00298.
Gräff, J., Rei, D., Guan, J.-S., Wang, W.-Y., Seo, J., Hennig, K.M., Nieland, T.J.F., Fass,D.M., Kao, P.F., Kahn, M., Su, S.C., Samiei, A., Joseph, N., Haggarty, S.J., Delalle, I.,Tsai, L.-H., 2012. An epigenetic blockade of cognitive functions in the neurodegen-erating brain. Nature 483, 222–226. https://doi.org/10.1038/nature10849.
Graff, J., Koshibu, K., Jouvenceau, A., Dutar, P., Mansuy, I.M., 2010. Protein phosphatase1-dependent transcriptional programs for long-term memory and plasticity. Learn.Mem. 355–363. https://doi.org/10.1101/lm.1766510.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
99

Gregoretti, I., Lee, Y.-M., Goodson, H.V., 2004. Molecular evolution of the histone dea-cetylase family: functional implications of phylogenetic analysis. J. Mol. Biol. 338,17–31. https://doi.org/10.1016/j.jmb.2004.02.006.
Guan, J.-S., Haggarty, S.J., Giacometti, E., Dannenberg, J.-H., Joseph, N., Gao, J.,Nieland, T.J.F., Zhou, Y., Wang, X., Mazitschek, R., Bradner, J.E., DePinho, R.A.,Jaenisch, R., Tsai, L.-H., 2009. HDAC2 negatively regulates memory formation andsynaptic plasticity. Nature 459, 55–60. https://doi.org/10.1038/nature07925.
Herry, C., Ferraguti, F., Singewald, N., Letzkus, J.J., Ehrlich, I., Lüthi, A., Lü Thi, A., 2010.Neuronal circuits of fear extinction. Eur. J. Neurosci. 31, 599–612. https://doi.org/10.1111/j.1460-9568.2010.07101.x.
Itzhak, Y., Anderson, K.L., Kelley, J.B., Petkov, M., 2012. Histone acetylation rescuescontextual fear conditioning in nNOS KO mice and accelerates extinction of cued fearconditioning in wild type mice. Neurobiol. Learn. Mem. 97, 409–417. https://doi.org/10.1016/j.nlm.2012.03.005.
Itzhak, Y., Liddie, S., Anderson, K.L., 2013. Sodium butyrate-induced histone acetylationstrengthens the expression of cocaine-associated contextual memory. Neurobiol.Learn. Mem. 102, 34–42. https://doi.org/10.1016/j.nlm.2013.03.007.
Jarome, T.J., Lubin, F.D., 2014. Epigenetic mechanisms of memory formation and re-consolidation. Neurobiol. Learn. Mem. 115, 116–127. https://doi.org/10.1016/j.nlm.2014.08.002.
Karpova, N.N., Sales, A.J., Joca, S.R., 2017. Epigenetic basis of neuronal and synapticplasticity. Curr. Top. Med. Chem. 17, 771–793.
Kim, S., Kaang, B.-K., 2017. Epigenetic regulation and chromatin remodeling in learningand memory. Exp. Mol. Med. 49, e281. https://doi.org/10.1038/emm.2016.140.
Knapska, E., Maren, S., 2009. Reciprocal patterns of c-Fos expression in the medial pre-frontal cortex and amygdala after extinction and renewal of conditioned fear. Learn.Mem. 16, 486–493. https://doi.org/10.1101/lm.1463909.
Korzus, E., Rosenfeld, M.G., Mayford, M., 2004. CBP histone acetyltransferase activity is acritical component of memory consolidation. Neuron 42, 961–972. https://doi.org/10.1016/j.neuron.2004.06.002.
Krettek, J.E., Price, J.L., 1978. A description of the amygdaloid complex in the rat and catwith observations on intra-amygdaloid axonal connections. J. Comp. Neurol. 178,255–280. https://doi.org/10.1002/cne.901780205.
Kwapis, J.L., Alaghband, Y., López, A.J., White, A.O., Campbell, R.R., Dang, R.T., Rhee,D., Tran, A.V., Carl, A.E., Matheos, D.P., Wood, M.A., 2017. Context and auditory fearare differentially regulated by HDAC3 activity in the lateral and basal subnuclei ofthe amygdala. Neuropsychopharmacology 42, 1284–1294. https://doi.org/10.1038/npp.2016.274.
Laborda, M.A., Miller, R.R., 2012. Reactivated memories compete for expression afterPavlovian extinction. Behav. Processes 90, 20–27. https://doi.org/10.1016/J.BEPROC.2012.01.012.
Laborda, M.A., Miller, R.R., 2013. Preventing return of fear in an animal model of an-xiety: additive effects of massive extinction and extinction in multiple contexts.Behav. Ther. 44, 249. https://doi.org/10.1016/J.BETH.2012.11.001.
Maddox, S.A., Schafe, G.E., 2011. Epigenetic alterations in the lateral amygdala are re-quired for reconsolidation of a Pavlovian fear memory. Learn. Mem. 18, 579–593.https://doi.org/10.1101/lm.2243411.
Maddox, S.A., Schafe, G.E., Ressler, K.J., 2013a. Exploring epigenetic regulation of fearmemory and biomarkers associated with post-traumatic stress disorder. Front.Psychiatry 4, 62. https://doi.org/10.3389/fpsyt.2013.00062.
Maddox, S.A., Watts, C.S., Schafe, G.E., 2013b. p300/CBP histone acetyltransferase ac-tivity is required for newly acquired and reactivated fear memories in the lateralamygdala. Learn. Mem. 20, 109–119. https://doi.org/10.1101/lm.029157.112.
Marek, R., Coelho, C.M., Sullivan, R.K.P., Baker-Andresen, D., Li, X., Ratnu, V., Dudley,K.J., Meyers, D., Mukherjee, C., Cole, P.A., Sah, P., Bredy, T.W., 2011. Paradoxicalenhancement of fear extinction memory and synaptic plasticity by inhibition of thehistone acetyltransferase p300. J. Neurosci. 31, 7486–7491. https://doi.org/10.1523/JNEUROSCI.0133-11.2011.
Marek, R., Strobel, C., Bredy, T.W., Sah, P., 2013. The amygdala and medial prefrontalcortex: partners in the fear circuit. J. Physiol. 591, 2381–2391. https://doi.org/10.1113/jphysiol.2012.248575.
McDonald, A.J., 1998. Cortical pathways to the mammalian amygdala. Prog. Neurobiol.55, 257–332.
Monsey, M.S., Ota, K.T., Akingbade, I.F., Hong, E.S., Schafe, G.E., 2011. Epigenetic al-terations are critical for fear memory consolidation and synaptic plasticity in thelateral amygdala. PLoS One 6, e19958. https://doi.org/10.1371/journal.pone.0019958.
Moorman, D.E., James, M.H., McGlinchey, E.M., Aston-Jones, G., 2015. Differential rolesof medial prefrontal subregions in the regulation of drug seeking. Brain Res. 1628,130–146. https://doi.org/10.1016/j.brainres.2014.12.024.
Morris, M.J., Mahgoub, M., Na, E.S., Pranav, H., Monteggia, L.M., 2013. Loss of histonedeacetylase 2 improves working memory and accelerates extinction learning. J.Neurosci. 33, 6401–6411. https://doi.org/10.1523/JNEUROSCI.1001-12.2013.
Myers, K.M., Davis, M., 2007. Mechanisms of fear extinction. Mol. Psychiatry 12,120–150. https://doi.org/10.1038/sj.mp.4001939.
Nader, K., Majidishad, P., Amorapanth, P., LeDoux, J.E., 2001. Damage to the lateral andcentral, but not other, amygdaloid nuclei prevents the acquisition of auditory fearconditioning. Learn. Mem. 8, 156–163. https://doi.org/10.1101/lm.38101.
Orsini, C.A., Maren, S., 2012. Neuroscience and Biobehavioral Reviews Neural and cel-lular mechanisms of fear and extinction memory formation. Neurosci. Biobehav. Rev.36, 1773–1802. https://doi.org/10.1016/j.neubiorev.2011.12.014.
Pape, H.C., Pare, D., 2010. Plastic synaptic networks of the amygdala for the acquisition,expression, and extinction of conditioned fear. Physiol. Rev. 90, 419–463. https://doi.org/10.1152/physrev.00037.2009.
Pare, D., Duvarci, S., 2012. Amygdala microcircuits mediating fear expression and ex-tinction. Curr. Opin. Neurobiol. 22, 717–723. https://doi.org/10.1016/j.conb.2012.
02.014.Peixoto, L., Abel, T., 2013. The role of histone acetylation in memory formation and
cognitive impairments. Neuropsychopharmacology 38, 62–76. https://doi.org/10.1038/npp.2012.86.
Peleg, S., Sananbenesi, F., Zovoilis, A., Burkhardt, S., Bahari-Javan, S., Agis-Balboa, R.C.,Cota, P., Wittnam, J.L., Gogol-Doering, A., Opitz, L., Salinas-Riester, G., Dettenhofer,M., Kang, H., Farinelli, L., Chen, W., Fischer, A., 2010. Altered histone acetylation isassociated with age-dependent memory impairment in mice. Science 328, 753–756.https://doi.org/10.1126/science.1186088.
Penney, J., Tsai, L.-H., 2014. Histone deacetylases in memory and cognition. Sci. Signal.7, re12. https://doi.org/10.1126/scisignal.aaa0069.
Peters, J., Kalivas, P.W., Quirk, G.J., 2009. Extinction circuits for fear and addictionoverlap in prefrontal cortex. Learn. Mem. 16, 279–288. https://doi.org/10.1101/lm.1041309.
Petrovich, G.D., Swanson, L.W., 1997. Projections from the lateral part of the centralamygdalar nucleus to the postulated fear conditioning circuit. Brain Res. 763,247–254.
Radulovic, J., Kammermeier, J., Spiess, J., 1998. Relationship between fos productionand classical fear conditioning: effects of novelty, latent inhibition, and uncondi-tioned stimulus preexposure. J. Neurosci. 18, 7452–7461.
Ranjan, V., Singh, S., Siddiqui, S.A., Khan, M.Y., Prakash, A., 2015. Differential histoneacetylation in the Amygdala leads to fear memory consolidation and extinction. Int.J. Sci. Technol. Soc. 1. https://doi.org/10.18091/ijsts.v1i1.13.
Renthal, W., Carle, T.L., Maze, I., Covington, H.E., Truong, H.-T., Alibhai, I., Kumar, A.,Montgomery, R.L., Olson, E.N., Nestler, E.J., Nestler, E.J., 2008. Delta FosB mediatesepigenetic desensitization of the c-fos gene after chronic amphetamine exposure. J.Neurosci. 28, 7344–7349. https://doi.org/10.1523/JNEUROSCI.1043-08.2008.
Saffari, R., Teng, Z., Zhang, M., Kravchenko, M., Hohoff, C., Ambrée, O., Zhang, W., 2016.NPY+-, but not PV+-GABAergic neurons mediated long-range inhibition from infra-to prelimbic cortex. Transl. Psychiatry 6, e736. https://doi.org/10.1038/tp.2016.7.
Sah, P., Faber, E.S.L., Lopez De Armentia, M., Power, J., 2003. The amygdaloid complex:anatomy and physiology. Physiol. Rev. 83, 803–834. https://doi.org/10.1152/physrev.00002.2003.
Schneider, A., Chatterjee, S., Bousiges, O., Selvi, B.R., Swaminathan, A., Cassel, R., Blanc,F., Kundu, T.K., Boutillier, A.-L., 2013. Acetyltransferases (HATs) as targets forneurological therapeutics. Neurotherapeutics 10, 568–588. https://doi.org/10.1007/s13311-013-0204-7.
Siddiqui, S.A., Singh, S., Ranjan, V., Ugale, R., Saha, S., Prakash, A., 2017. Enhancedhistone acetylation in the infralimbic prefrontal cortex is associated with fear ex-tinction. Cell. Mol. Neurobiol. 37. https://doi.org/10.1007/s10571-017-0464-6.
Sierra-Mercado, D., Padilla-Coreano, N., Quirk, G.J., 2011. Dissociable roles of prelimbicand infralimbic cortices, ventral hippocampus, and basolateral amygdala in the ex-pression and extinction of conditioned fear. Neuropsychopharmacology 36, 529–538.https://doi.org/10.1038/npp.2010.184.
Singewald, N., Schmuckermair, C., Whittle, N., Holmes, A., Ressler, K.J., 2015.Pharmacology of cognitive enhancers for exposure-based therapy of fear, anxiety andtrauma-related disorders. Pharmacol. Ther. 149, 150–190. https://doi.org/10.1016/j.pharmthera.2014.12.004.
Stafford, J.M., Raybuck, J.D., Ryabinin, A.E., Lattal, K.M., 2012. Increasing histoneacetylation in the hippocampus-infralimbic network enhances fear extinction. Biol.Psychiatry 72, 25–33. https://doi.org/10.1016/j.biopsych.2011.12.012.
Sultan, F.A., Day, J.J., 2011. Epigenetic mechanisms in memory and synaptic function.Epigenomics 3, 157–181. https://doi.org/10.2217/epi.11.6.
Sun, H., Kennedy, P.J., Nestler, E.J., 2013. Epigenetics of the depressed brain: role ofhistone acetylation and methylation. Neuropsychopharmacology 38, 124–137.https://doi.org/10.1038/npp.2012.73.
Teichner, W.H., Holder, E., 1952. Reminiscence as a function of the amount of change inthe intertrial interval. J. Exp. Psychol. 44, 347–351.
Tronson, N.C., Schrick, C., Guzman, Y.F., Huh, K.H., Srivastava, D.P., Penzes, P., Guedea,A.L., Gao, C., Radulovic, J., 2009. Segregated populations of hippocampal principalCA1 neurons mediating conditioning and extinction of contextual fear. J. Neurosci.29, 3387–3394. https://doi.org/10.1523/JNEUROSCI.5619-08.2009.
Turner, B.H., Herkenham, M., 1991. Thalamoamygdaloid projections in the rat: a test ofthe amygdala’s role in sensory processing. J. Comp. Neurol. 313, 295–325. https://doi.org/10.1002/cne.903130208.
Usenko, T., Kukushkin, A., Pospelova, T., Pospelov, V., 2003. Transient expression of E1Aand Ras oncogenes causes downregulation of c-fos gene transcription in non-transformed REF52 cells. Oncogene 22, 7661–7666. https://doi.org/10.1038/sj.onc.1206975.
Valiati, F.E., Vasconcelos, M., Lichtenfels, M., Petry, F.S., de Almeida, R.M.M.,Schwartsmann, G., Schröder, N., de Farias, C.B., Roesler, R., 2017. Administration ofa histone deacetylase inhibitor into the basolateral amygdala enhances memoryconsolidation, delays extinction, and increases hippocampal BDNF levels. Front.Pharmacol. 8, 415. https://doi.org/10.3389/fphar.2017.00415.
Veening, J.G., Swanson, L.W., Sawchenko, P.E., 1984. The organization of projectionsfrom the central nucleus of the amygdala to brainstem sites involved in central au-tonomic regulation: a combined retrograde transport-immunohistochemical study.Brain Res. 303, 337–357.
Vieira, P.A., Lovelace, J.W., Corches, A., Rashid, A.J., Josselyn, S.A., Korzus, E., 2014.Prefrontal consolidation supports the attainment of fear memory accuracy. Learn.Mem. 21, 394–405. https://doi.org/10.1101/lm.036087.114.
Waddell, S., 2003. Protein phosphatase 1 and memory: practice makes PP1 imperfect?Trends Neurosci. 26 (3), 117–119. https://doi.org/10.1016/S0166-2236(03)00029-8.
Whittle, N., Maurer, V., Murphy, C., Rainer, J., Bindreither, D., Hauschild, M., Scharinger,A., Oberhauser, M., Keil, T., Brehm, C., Valovka, T., Striessnig, J., Singewald, N.,
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
100

2016. Enhancing dopaminergic signaling and histone acetylation promotes long-termrescue of deficient fear extinction. Transl. Psychiatry 6, e974. https://doi.org/10.1038/tp.2016.231.
Yang, S.H., Vickers, E., Brehm, A., Kouzarides, T., Sharrocks, A.D., 2001. Temporal re-cruitment of the mSin3A-histone deacetylase corepressor complex to the ETS domain
transcription factor Elk-1. Mol. Cell. Biol. 21, 2802–2814. https://doi.org/10.1128/MCB.21.8.2802-2814.2001.
Zovkic, I.B., Sweatt, J.D., 2013. Epigenetic mechanisms in learned fear: implications forPTSD. Neuropsychopharmacology 38, 77–93. https://doi.org/10.1038/npp.2012.79.
S.A. Siddiqui, et al. Brain Research Bulletin 150 (2019) 86–101
101





























