Embed Size (px)
Citation preview

BIOQUIMICA DE PROTEINAS
PLEGAMIENTO Y ESTABILIDAD DE PROTEINAS Y TECNICAS
ESPECTROSCÓPICAS PARA SU ANÁLISIS
PROF. LUCIA B. CHEMES

BIBLIOGRAFIA
JR LAKOWICS. Principles of fluorescence spectroscopy. 3rd Edition (2006). Capítulos 1 y 16
FLUORESCENCIA
DICROISMO CIRCULARGD FASMAN, Ed. Circular Dichroism and the conformational analysis of biomolecules Springer 1996. Cap 2, 3 y 4
PLEGAMIENTO Y ESTABILIDAD DE PROTEINAS
FERSHT, A. STRUCTURE AND MECHANISM IN PROTEIN SCIENCE (1999) FREEMAN PRESS, CAP 1, 11, 17 18 Y 19
BRANDEN AND TOOZE. INTRODUCTION TO PROTEIN STRUCTURE (1999) GARLAND PUBLISHING
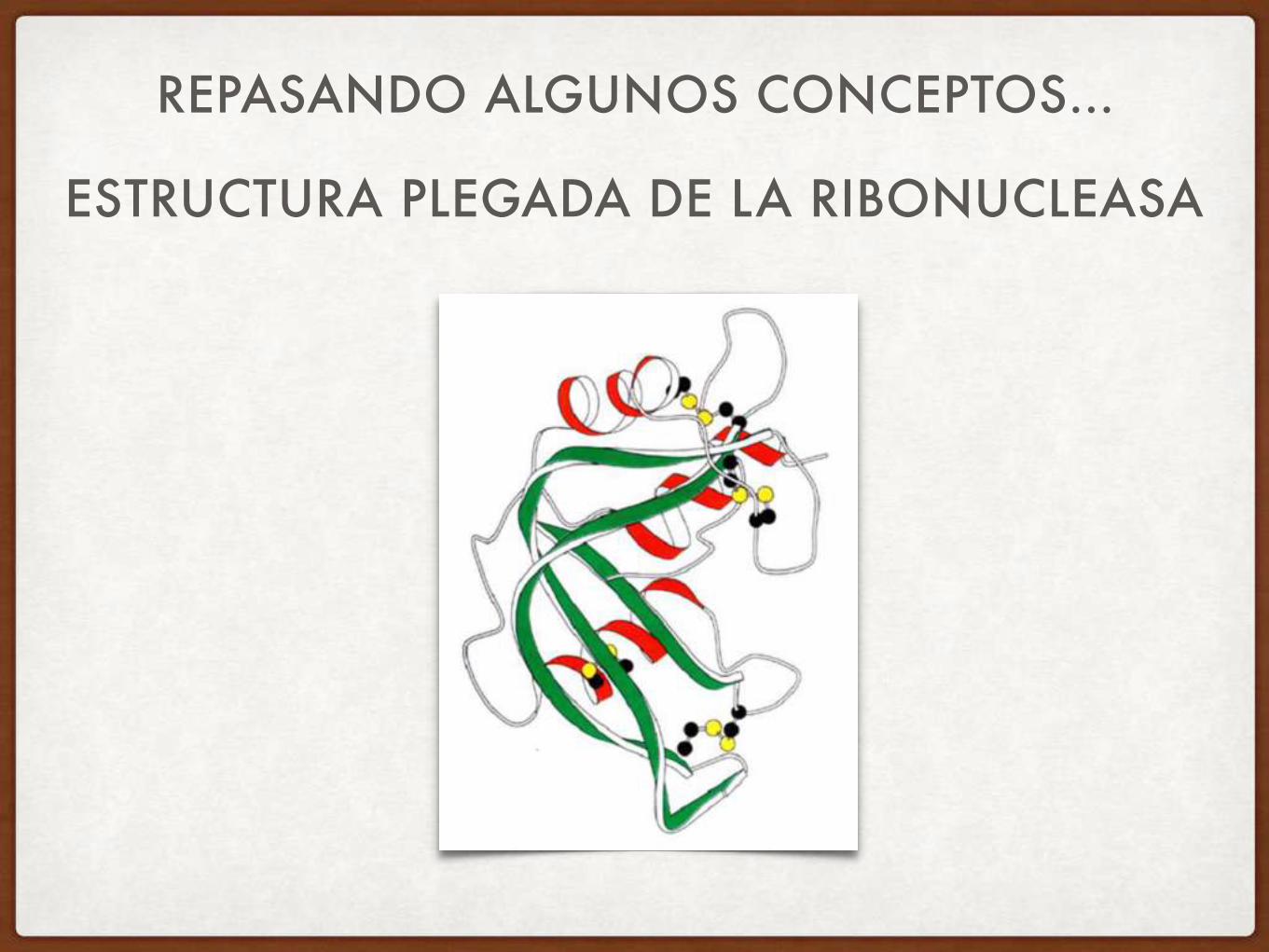
REPASANDO ALGUNOS CONCEPTOS…
ESTRUCTURA PLEGADA DE LA RIBONUCLEASA
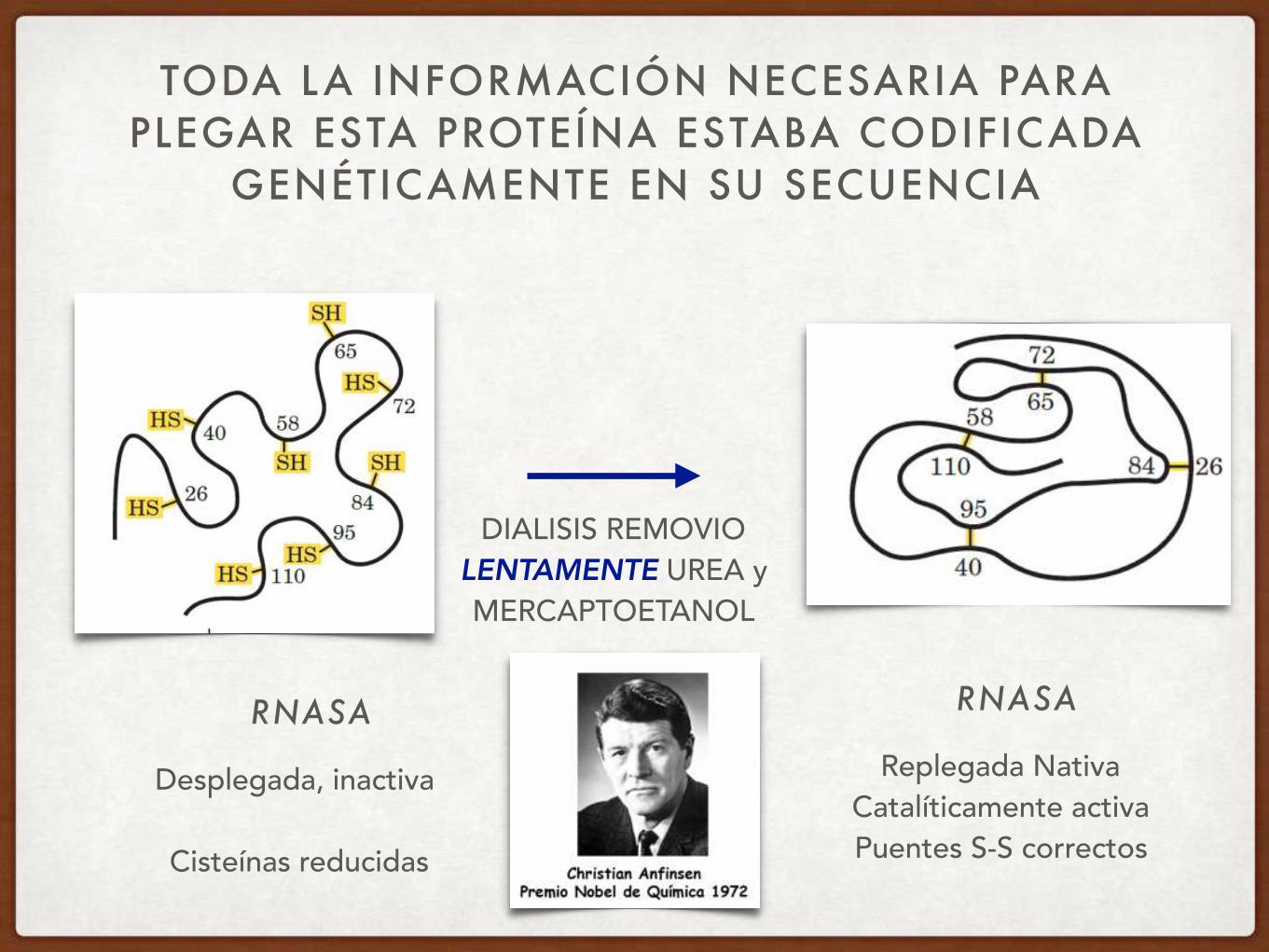
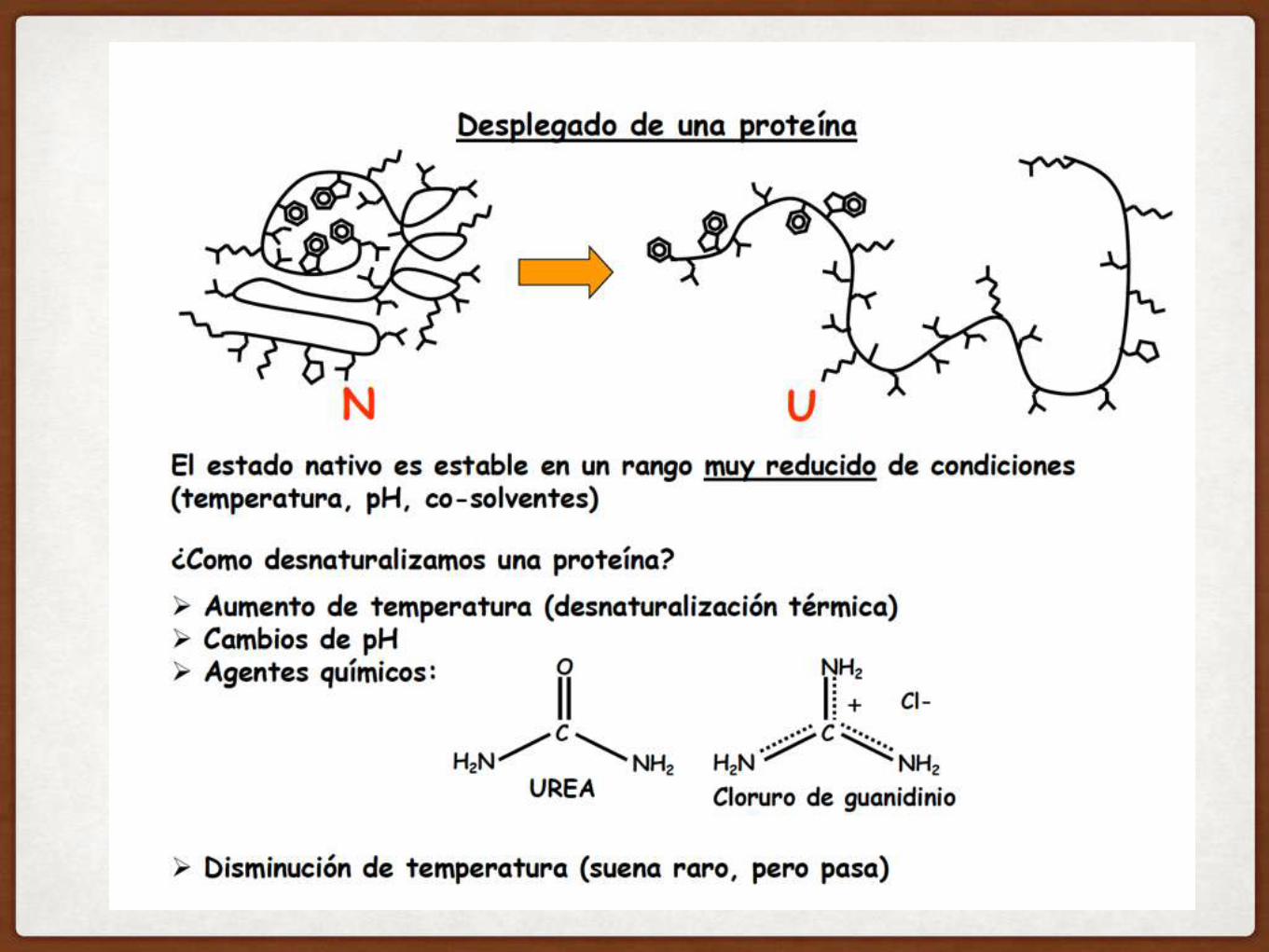
TODA LA INFORMACIÓN NECESARIA PARA PLEGAR ESTA PROTEÍNA ESTABA CODIFICADA
GENÉTICAMENTE EN SU SECUENCIA
RNASA
Desplegada, inactiva
Cisteínas reducidas
DIALISIS REMOVIO LENTAMENTE UREA y MERCAPTOETANOL
RNASA
Replegada Nativa Catalíticamente activa Puentes S-S correctos

PLEGAMIENTO DE LAS PROTEINASLA PARADOJA DE LEVINTHAL
Para una cadena de 100 aminoácidos…. Si hay sólo 3 conformaciones para cada aminoácidoCombinaciones azarosas que deben ser exploradas 3100
Si cada transición lleva 10-13 seg (muy poco!)
5 x 1047~
5 x 1034seg 1.6 x 1027 años!
CUANTAS COMBINATORIAS DEBE EXPLORAR UNA SECUENCIA PARA PODER PLEGARSE?
CUANTO TIEMPO LLEVA HACERLO?
EL PLEGAMIENTO DE LAS PROTEÍNAS NO OCURRE POR UN PROCESO DE EXPLORACIÓN AZAROSA DEL ESPACIO DE CONFORMACIONES !!!!!!
🤔
"ni corriendo se llega…no es compatible con la vida!
ENTONCES: CÓMO OCURRE?

LA ANALOGIA DEL MONO DE SHAKESPEARE
Hacen falta 1040 combinaciones!!!! para que un mono tipee el siguiente texto de Shakespeare al azar
Peeero….si se retienen las posiciones correctas
LA ESENCIA DEL PLEGAMIENTO DE PROTEINAS ES LA RETENCION DE INTERMEDIARIOS CORRECTOS DE PLEGADO
Sólo hacen falta ~3000 tipeosal azar = 2326 COMBINACIONES
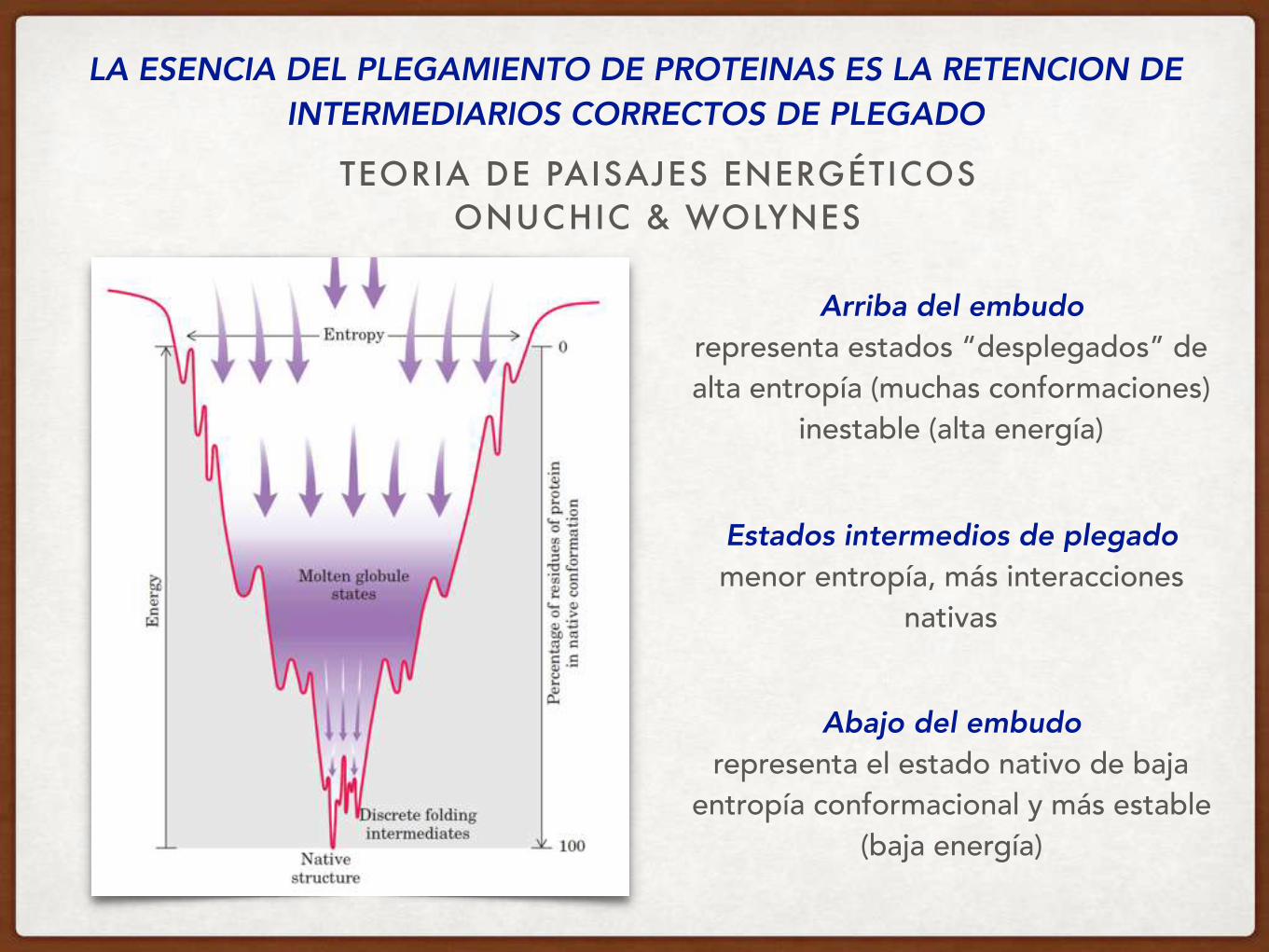
TEORIA DE PAISAJES ENERGÉTICOS ONUCHIC & WOLYNES
LA ESENCIA DEL PLEGAMIENTO DE PROTEINAS ES LA RETENCION DE INTERMEDIARIOS CORRECTOS DE PLEGADO
Arriba del embudo representa estados “desplegados” de alta entropía (muchas conformaciones)
inestable (alta energía)
Abajo del embudo representa el estado nativo de baja
entropía conformacional y más estable (baja energía)
Estados intermedios de plegado menor entropía, más interacciones
nativas
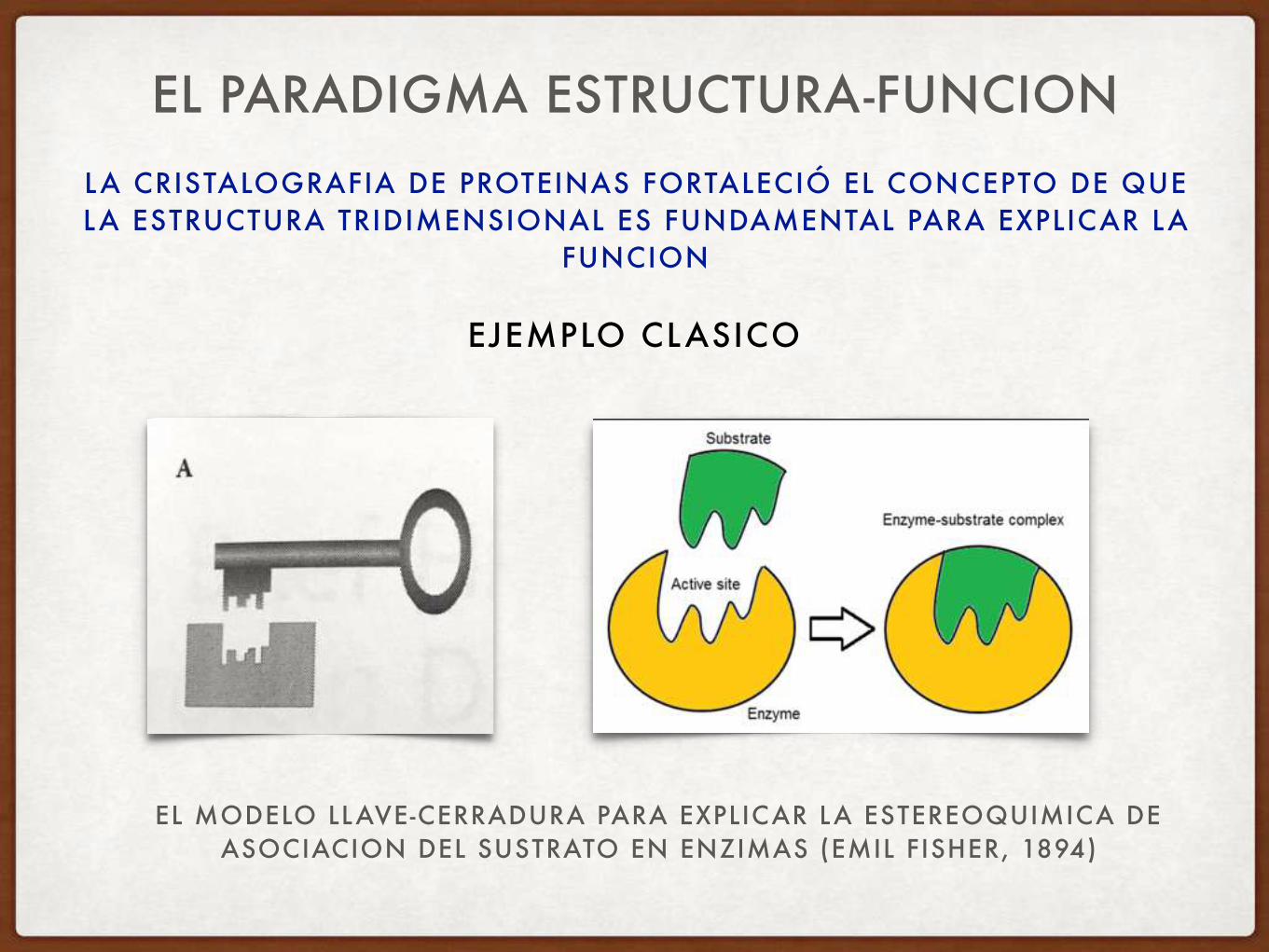
LA CRISTALOGRAFIA DE PROTEINAS FORTALECIÓ EL CONCEPTO DE QUE LA ESTRUCTURA TRIDIMENSIONAL ES FUNDAMENTAL PARA EXPLICAR LA
FUNCION
EL PARADIGMA ESTRUCTURA-FUNCION
EJEMPLO CLASICO
EL MODELO LLAVE-CERRADURA PARA EXPLICAR LA ESTEREOQUIMICA DE ASOCIACION DEL SUSTRATO EN ENZIMAS (EMIL FISHER, 1894)

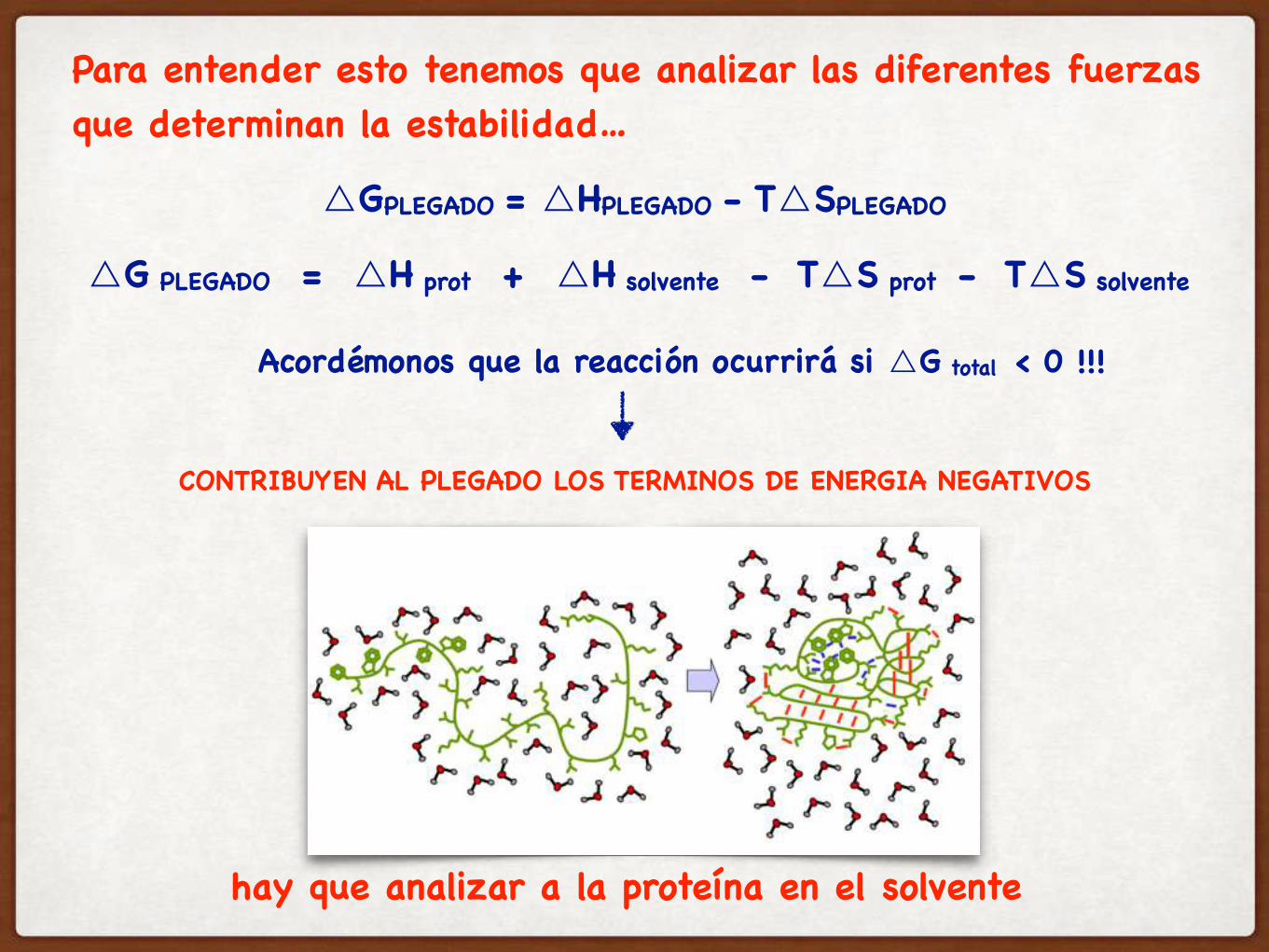
△G PLEGADO = △H prot + △H solvente - T△S prot - T△S solvente
CONTRIBUYEN AL PLEGADO LOS TERMINOS DE ENERGIA NEGATIVOS
△GPLEGADO = △HPLEGADO - T△SPLEGADO
Para entender esto tenemos que analizar las diferentes fuerzas que determinan la estabilidad…
Acordémonos que la reacción ocurrirá si △G total < 0 !!!
hay que analizar a la proteína en el solvente
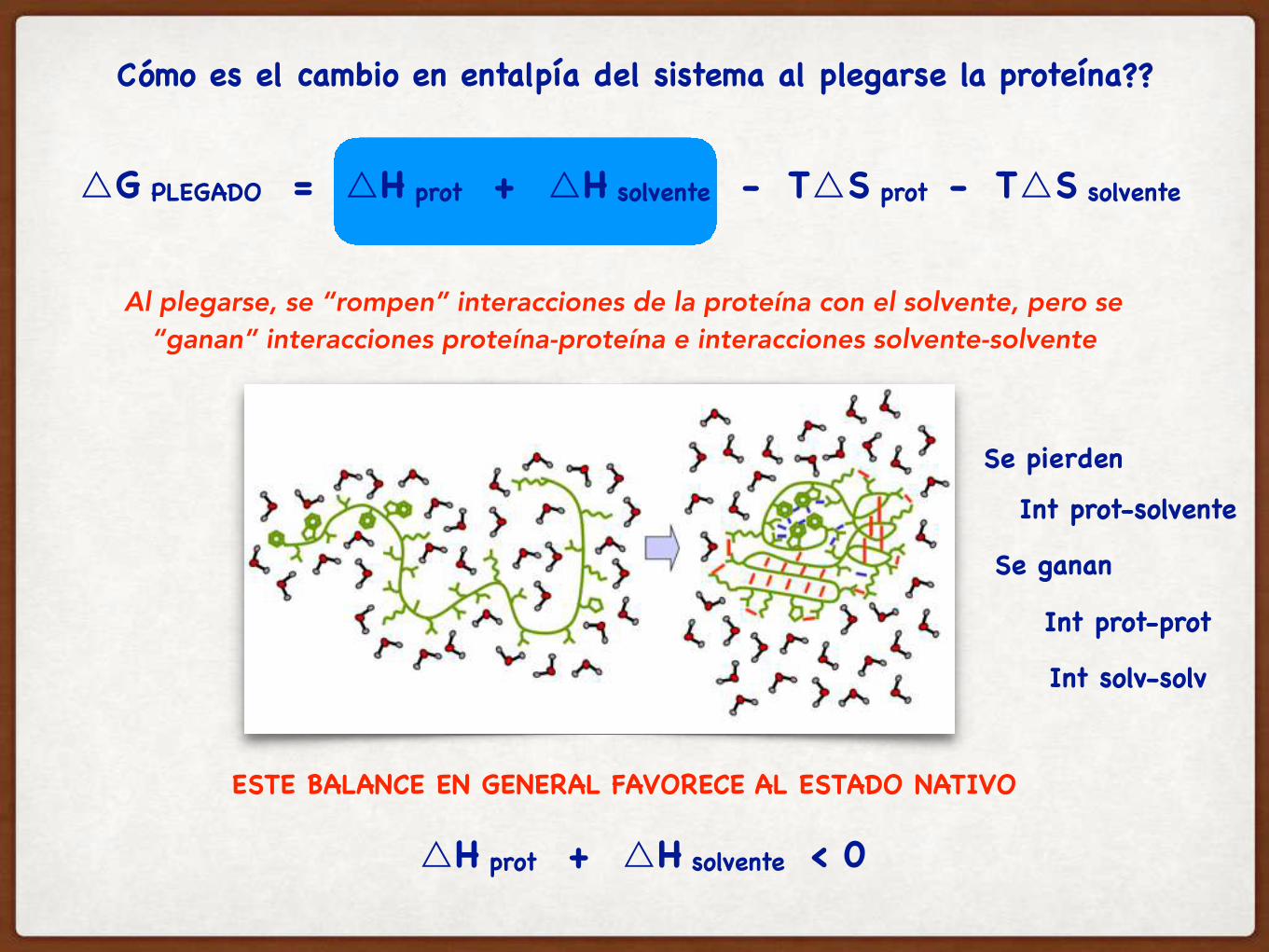
Al plegarse, se “rompen” interacciones de la proteína con el solvente, pero se “ganan” interacciones proteína-proteína e interacciones solvente-solvente
ESTE BALANCE EN GENERAL FAVORECE AL ESTADO NATIVO
Cómo es el cambio en entalpía del sistema al plegarse la proteína??
△H prot + △H solvente < 0
Se pierden
Int prot-solvente
Se ganan
Int prot-prot
Int solv-solv
△G PLEGADO = △H prot + △H solvente - T△S prot - T△S solvente
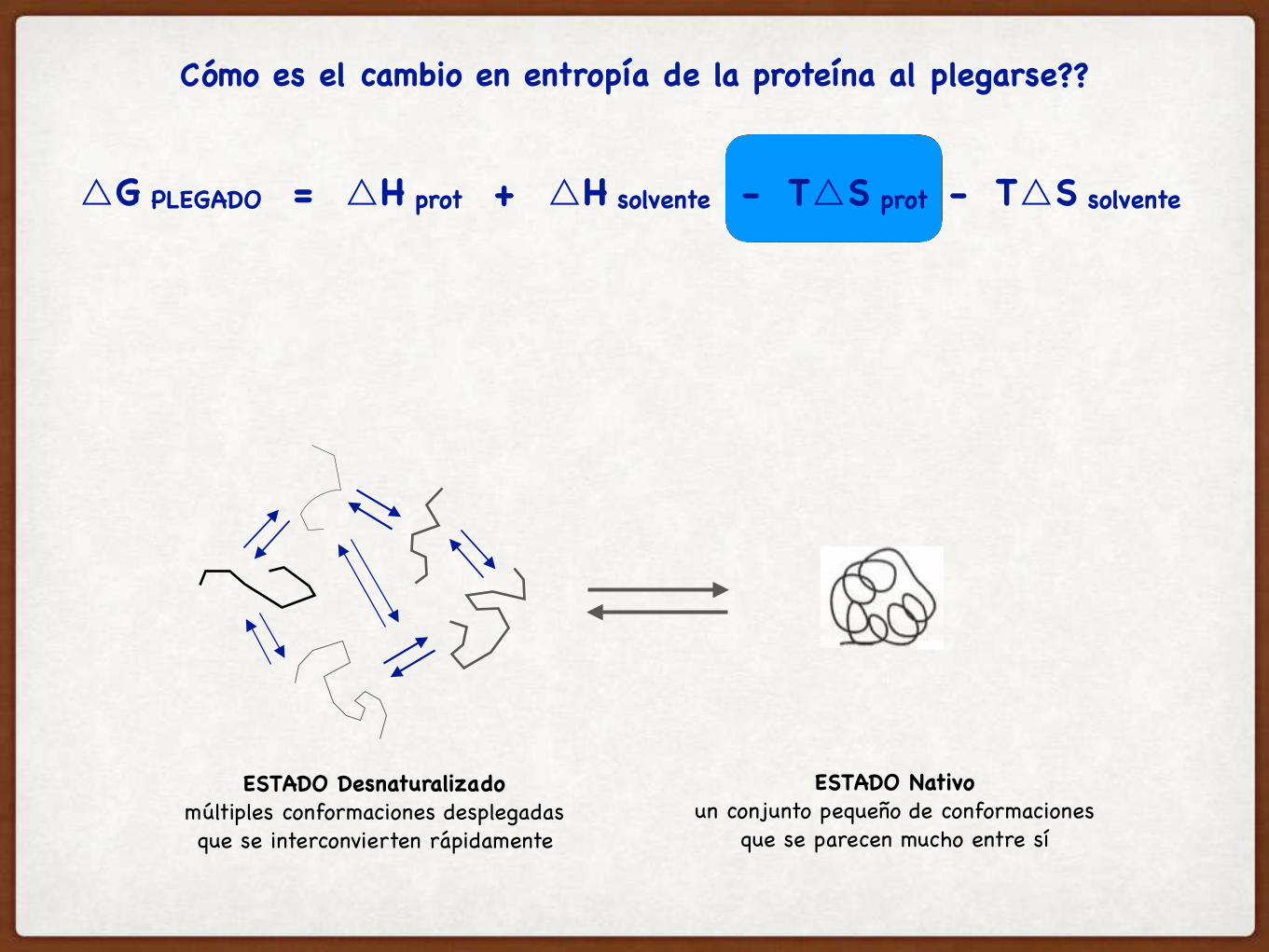
ESTADO Desnaturalizadomúltiples conformaciones desplegadasque se interconvierten rápidamente
ESTADO Nativoun conjunto pequeño de conformaciones
que se parecen mucho entre sí
Cómo es el cambio en entropía de la proteína al plegarse??
△G PLEGADO = △H prot + △H solvente - T△S prot - T△S solvente
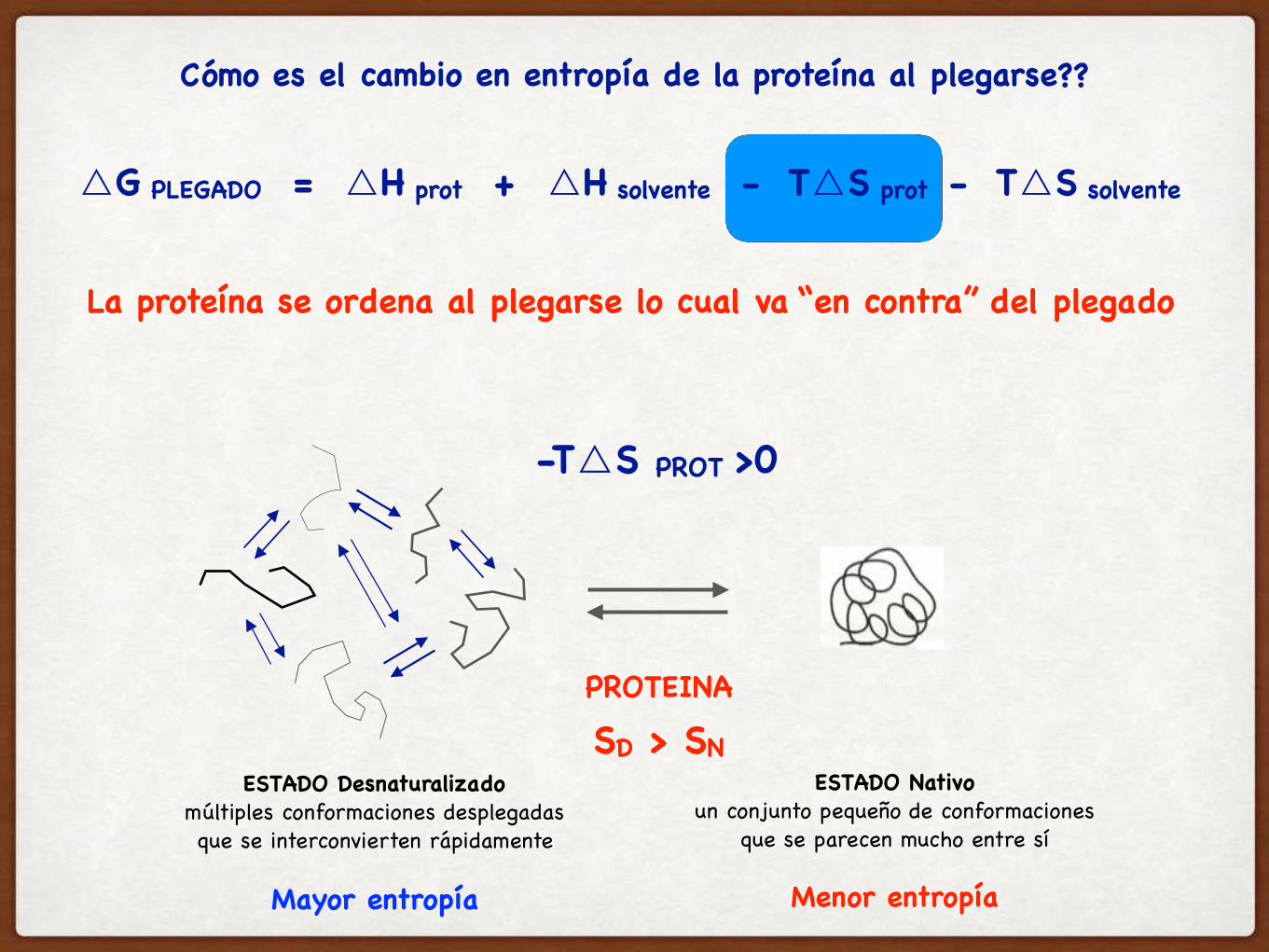
La proteína se ordena al plegarse lo cual va “en contra” del plegado
ESTADO Desnaturalizadomúltiples conformaciones desplegadasque se interconvierten rápidamente
ESTADO Nativoun conjunto pequeño de conformaciones
que se parecen mucho entre sí
Mayor entropía Menor entropía
PROTEINA
SD > SN
-T△S PROT >0
Cómo es el cambio en entropía de la proteína al plegarse??
△G PLEGADO = △H prot + △H solvente - T△S prot - T△S solvente

Aquí tenemos que recordar el “efecto hidrofóbico”….
Cómo es el cambio en entropía del solvente al plegarse??
△G total = △H prot + △H solvente - T△S prot - T△S solvente
EL PLEGADO LLEVA A UN AUMENTO EN LA ENTROPIA DEL SOLVENTE
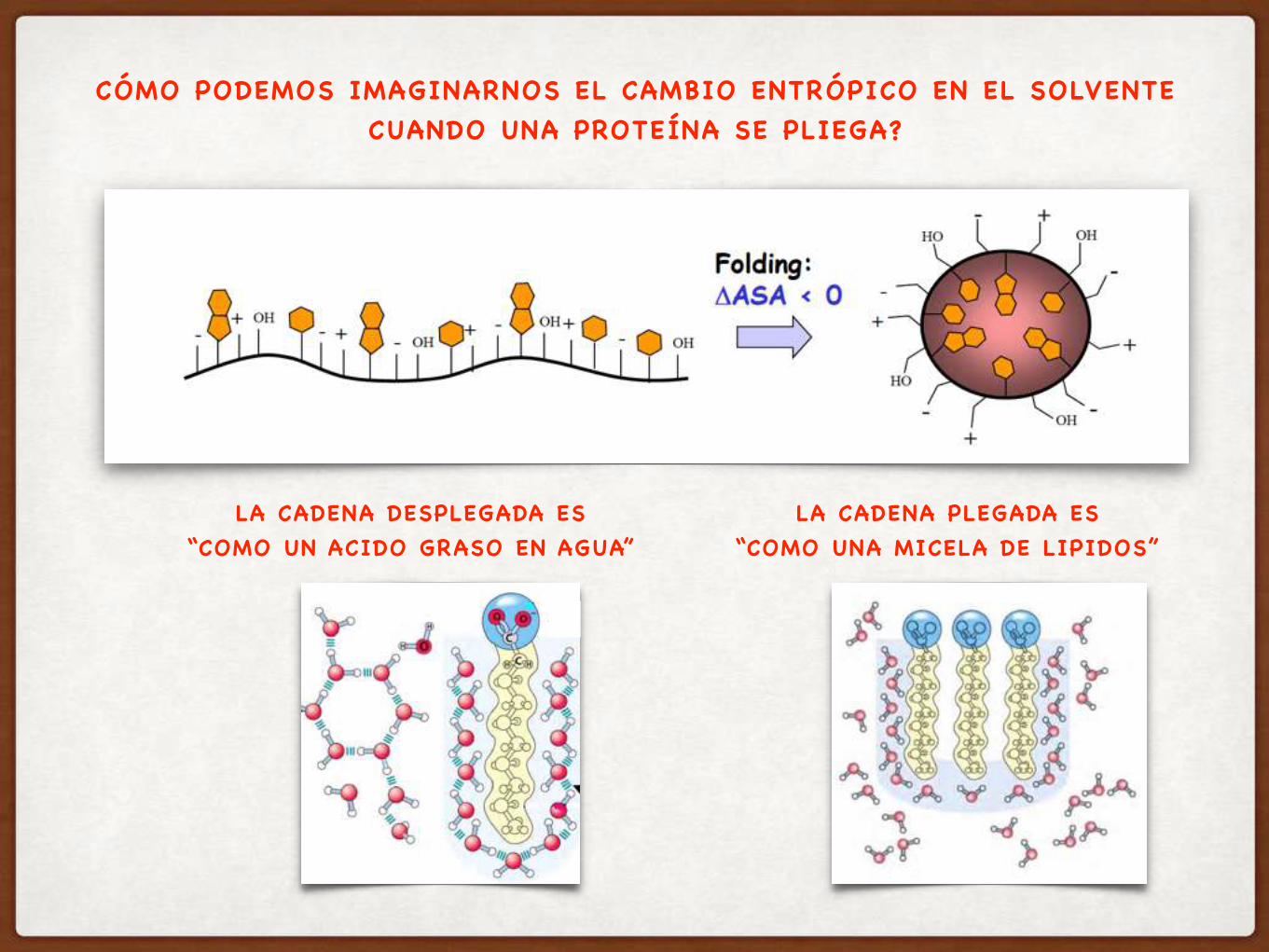
CÓMO PODEMOS IMAGINARNOS EL CAMBIO ENTRÓPICO EN EL SOLVENTE CUANDO UNA PROTEÍNA SE PLIEGA?
LA CADENA DESPLEGADA ES “COMO UN ACIDO GRASO EN AGUA”
LA CADENA PLEGADA ES “COMO UNA MICELA DE LIPIDOS”
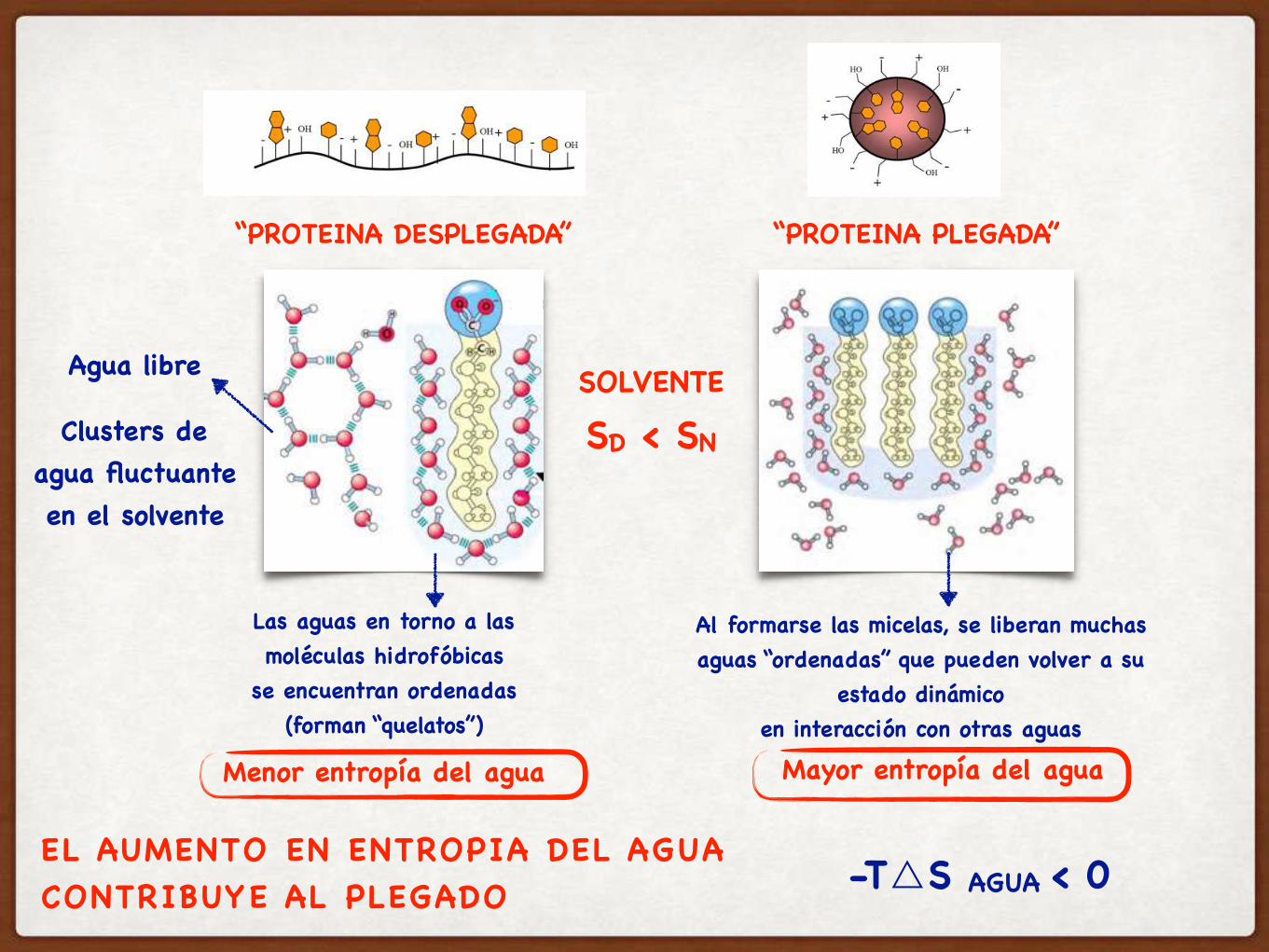
Las aguas en torno a las moléculas hidrofóbicas
se encuentran ordenadas(forman “quelatos”)
Menor entropía del agua
Agua libre
Al formarse las micelas, se liberan muchas aguas “ordenadas” que pueden volver a su
estado dinámicoen interacción con otras aguas
Clusters de agua fluctuante en el solvente
Mayor entropía del agua
EL AUMENTO EN ENTROPIA DEL AGUA CONTRIBUYE AL PLEGADO
“PROTEINA DESPLEGADA” “PROTEINA PLEGADA”
SOLVENTE
SD < SN
-T△S AGUA < 0
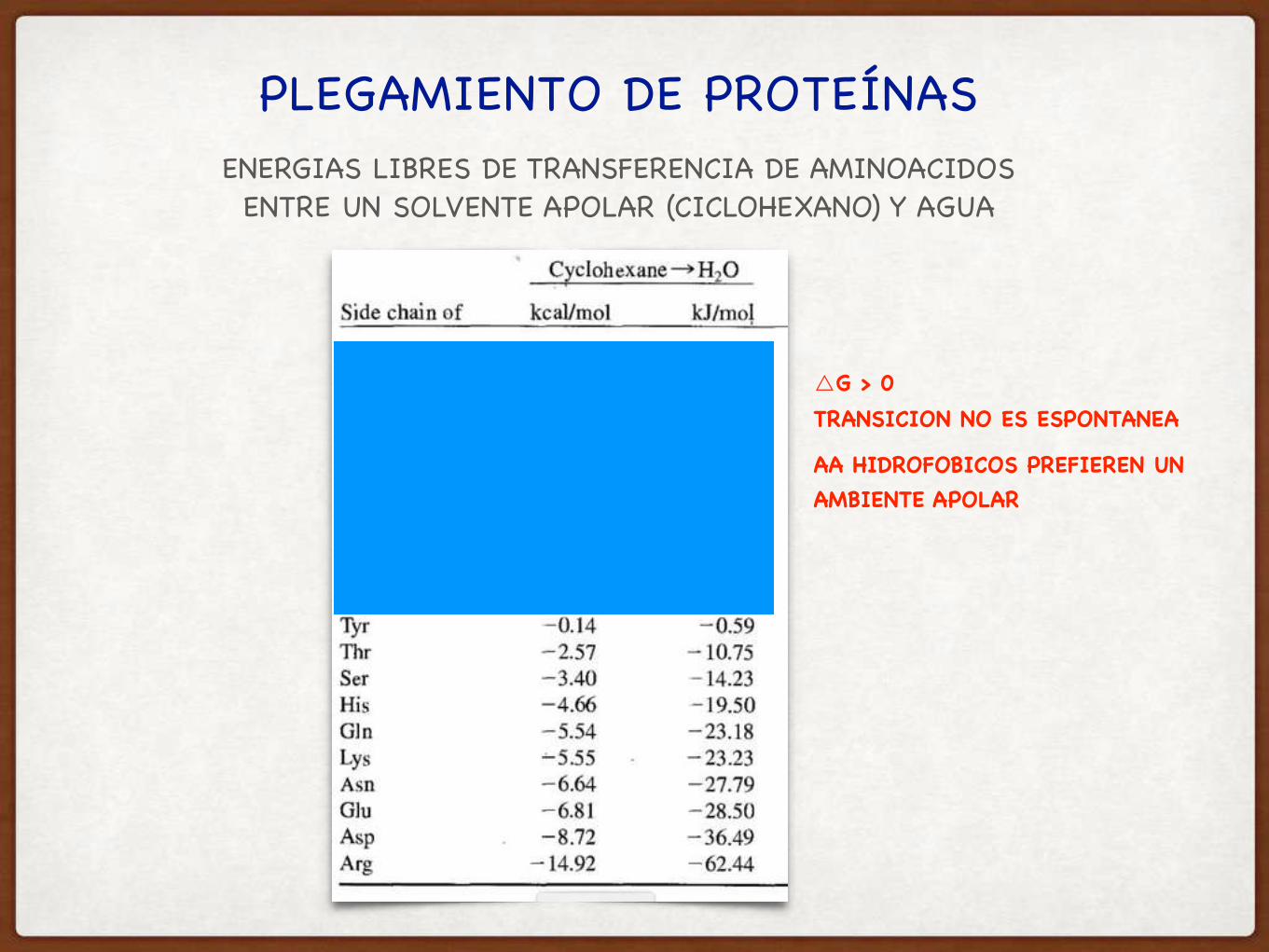
PLEGAMIENTO DE PROTEÍNASENERGIAS LIBRES DE TRANSFERENCIA DE AMINOACIDOSENTRE UN SOLVENTE APOLAR (CICLOHEXANO) Y AGUA
△G > 0TRANSICION NO ES ESPONTANEA
AA HIDROFOBICOS PREFIEREN UNAMBIENTE APOLAR

ENTONCES, RESUMIENDO LAS FUERZAS QUE CONTRIBUYEN AL PLEGADO DE LAS PROTEINAS:

EJEMPLOS DE LOS VALORES TERMODINÁMICOS PARA VARIAS PROTEÍNAS REALES
BIEN…ENTONCES LAS PROTEÍNAS SE PLIEGAN…PERO CÓMO??
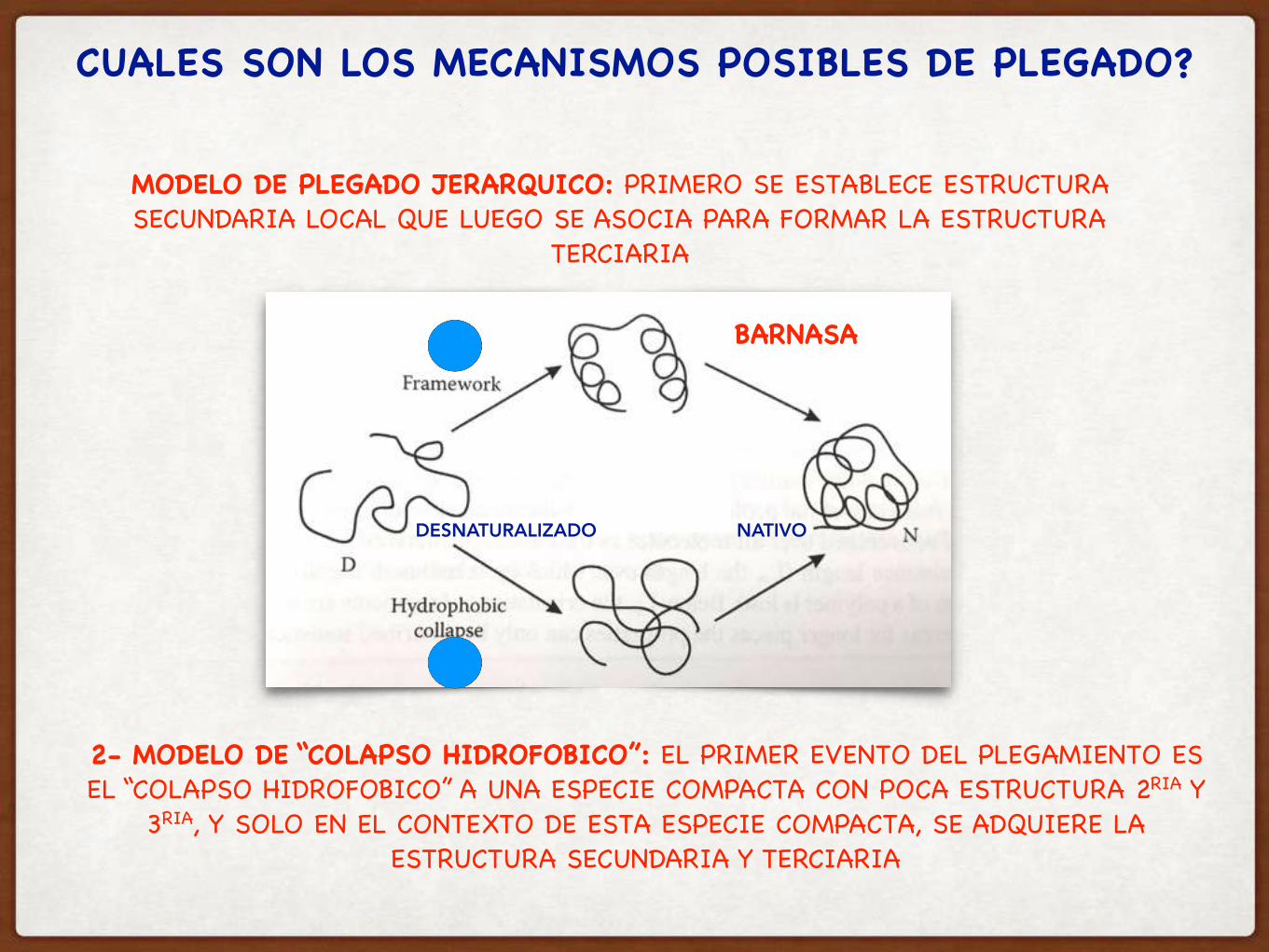
CUALES SON LOS MECANISMOS POSIBLES DE PLEGADO?
MODELO DE PLEGADO JERARQUICO: PRIMERO SE ESTABLECE ESTRUCTURA SECUNDARIA LOCAL QUE LUEGO SE ASOCIA PARA FORMAR LA ESTRUCTURA
TERCIARIA
2- MODELO DE “COLAPSO HIDROFOBICO”: EL PRIMER EVENTO DEL PLEGAMIENTO ES EL “COLAPSO HIDROFOBICO” A UNA ESPECIE COMPACTA CON POCA ESTRUCTURA 2RIA Y
3RIA, Y SOLO EN EL CONTEXTO DE ESTA ESPECIE COMPACTA, SE ADQUIERE LA ESTRUCTURA SECUNDARIA Y TERCIARIA
1
2
DESNATURALIZADO NATIVO
BARNASA
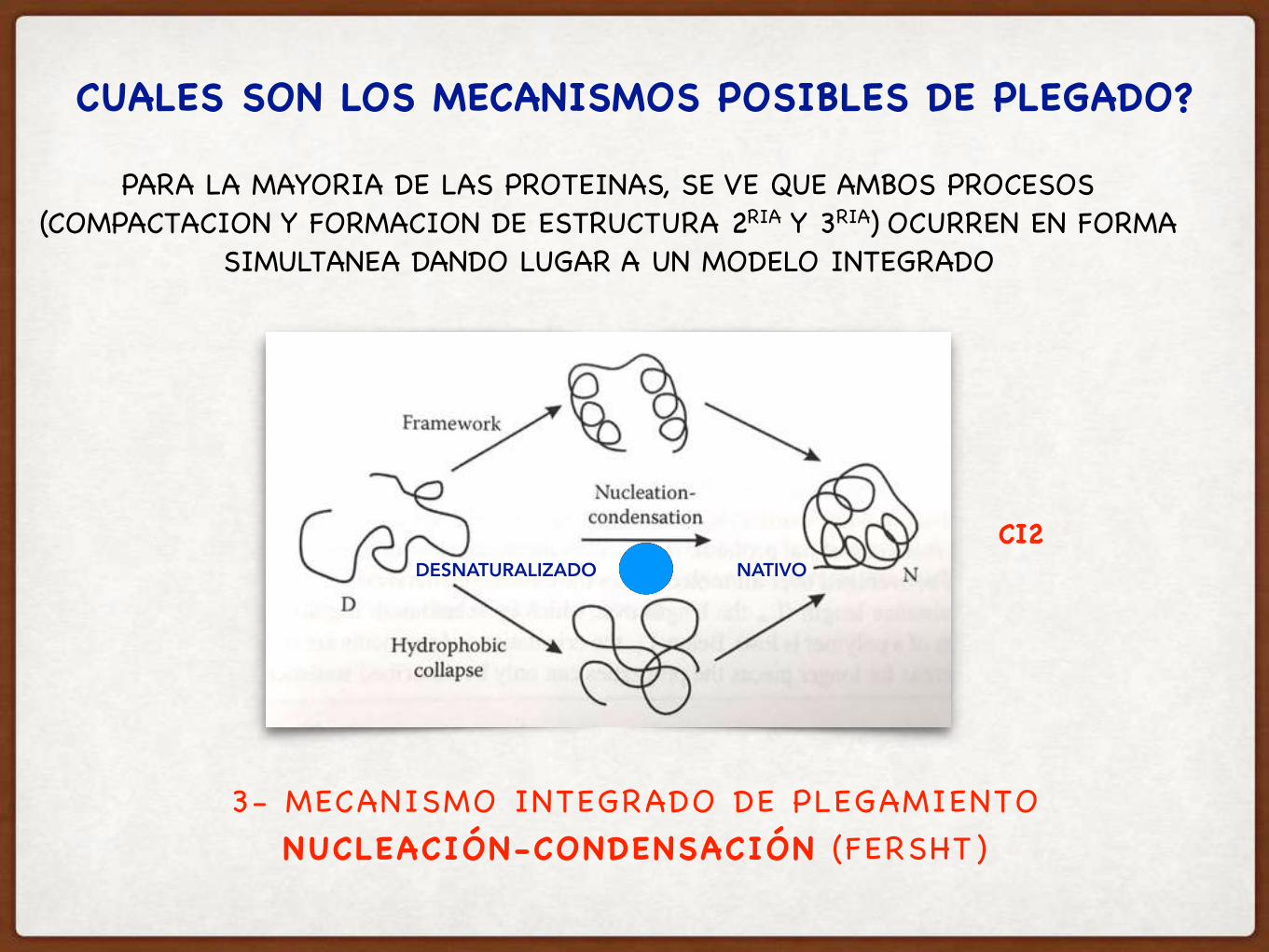
3- MECANISMO INTEGRADO DE PLEGAMIENTONUCLEACIÓN-CONDENSACIÓN (FERSHT )
CUALES SON LOS MECANISMOS POSIBLES DE PLEGADO?
DESNATURALIZADO NATIVO3
PARA LA MAYORIA DE LAS PROTEINAS, SE VE QUE AMBOS PROCESOS (COMPACTACION Y FORMACION DE ESTRUCTURA 2RIA Y 3RIA) OCURREN EN FORMA
SIMULTANEA DANDO LUGAR A UN MODELO INTEGRADO
CI2
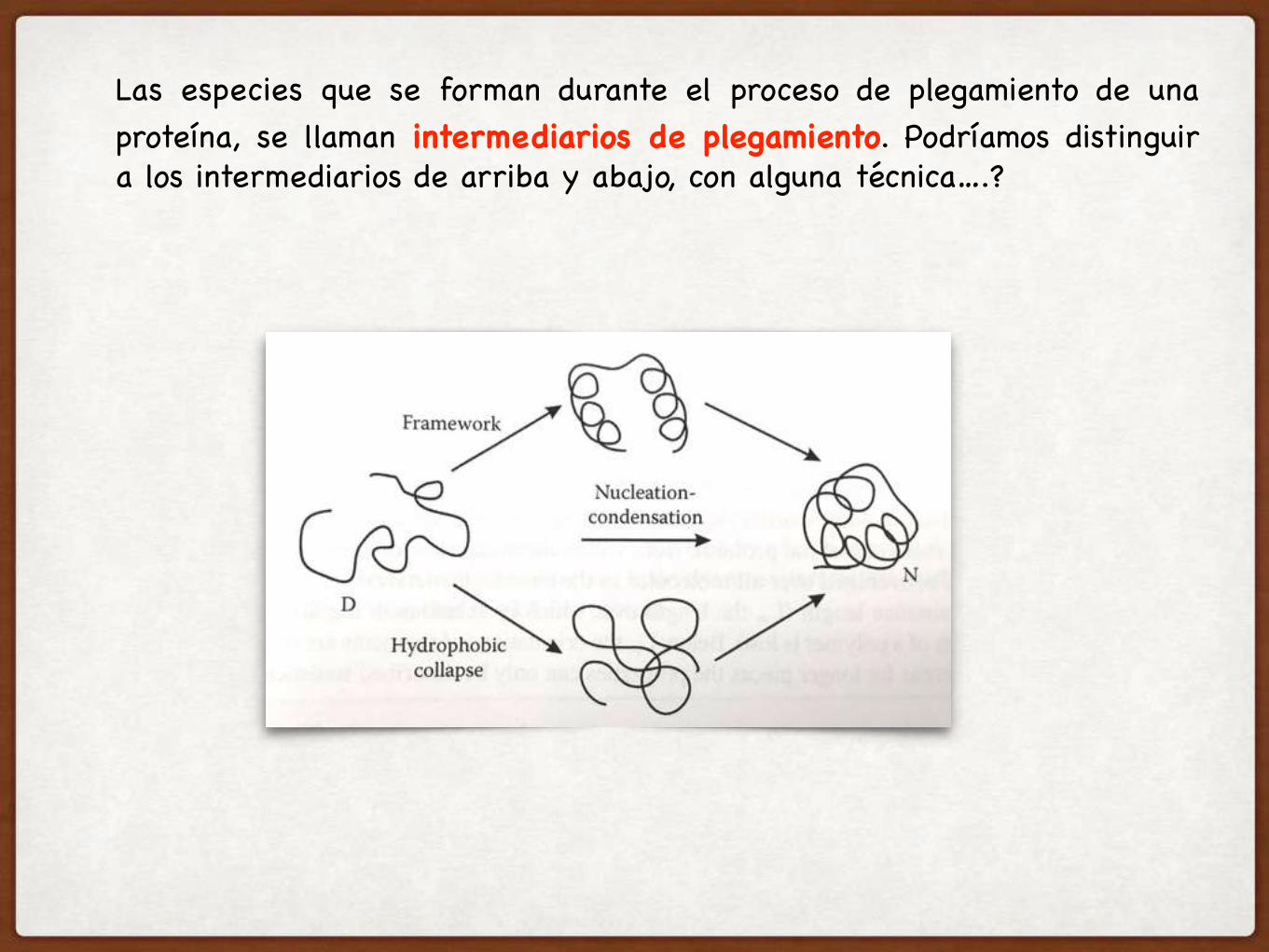
Las especies que se forman durante el proceso de plegamiento de una proteína, se llaman intermediarios de plegamiento. Podríamos distinguir a los intermediarios de arriba y abajo, con alguna técnica….?

POR OTRA PARTE…LA EFICIENCIA DEL PLEGAMIENTO VA A DEPENDER DEL PAISAJE ENERGETICO DEL PLEGADO…
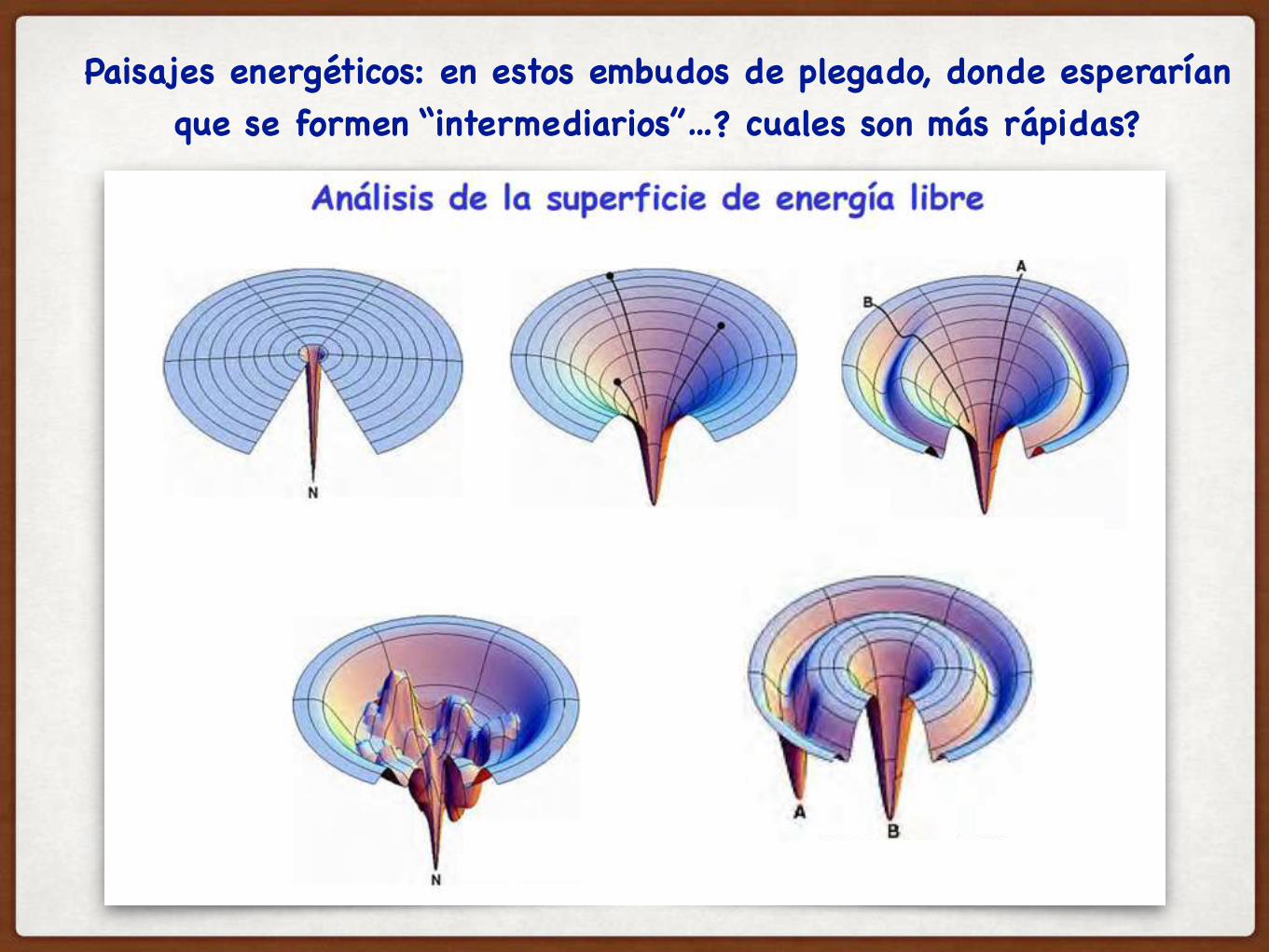
Paisajes energéticos: en estos embudos de plegado, donde esperarían que se formen “intermediarios”…? cuales son más rápidas?
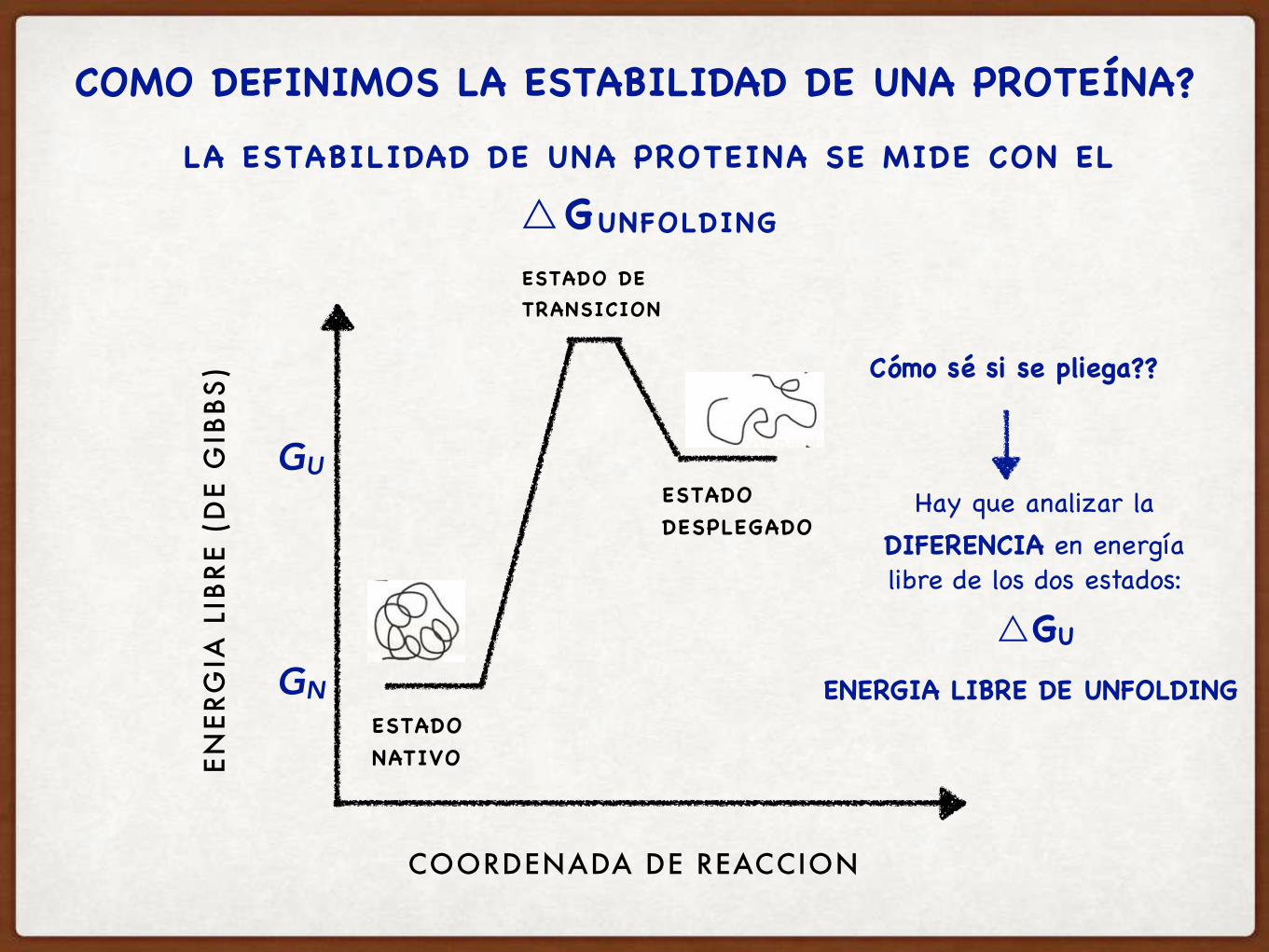
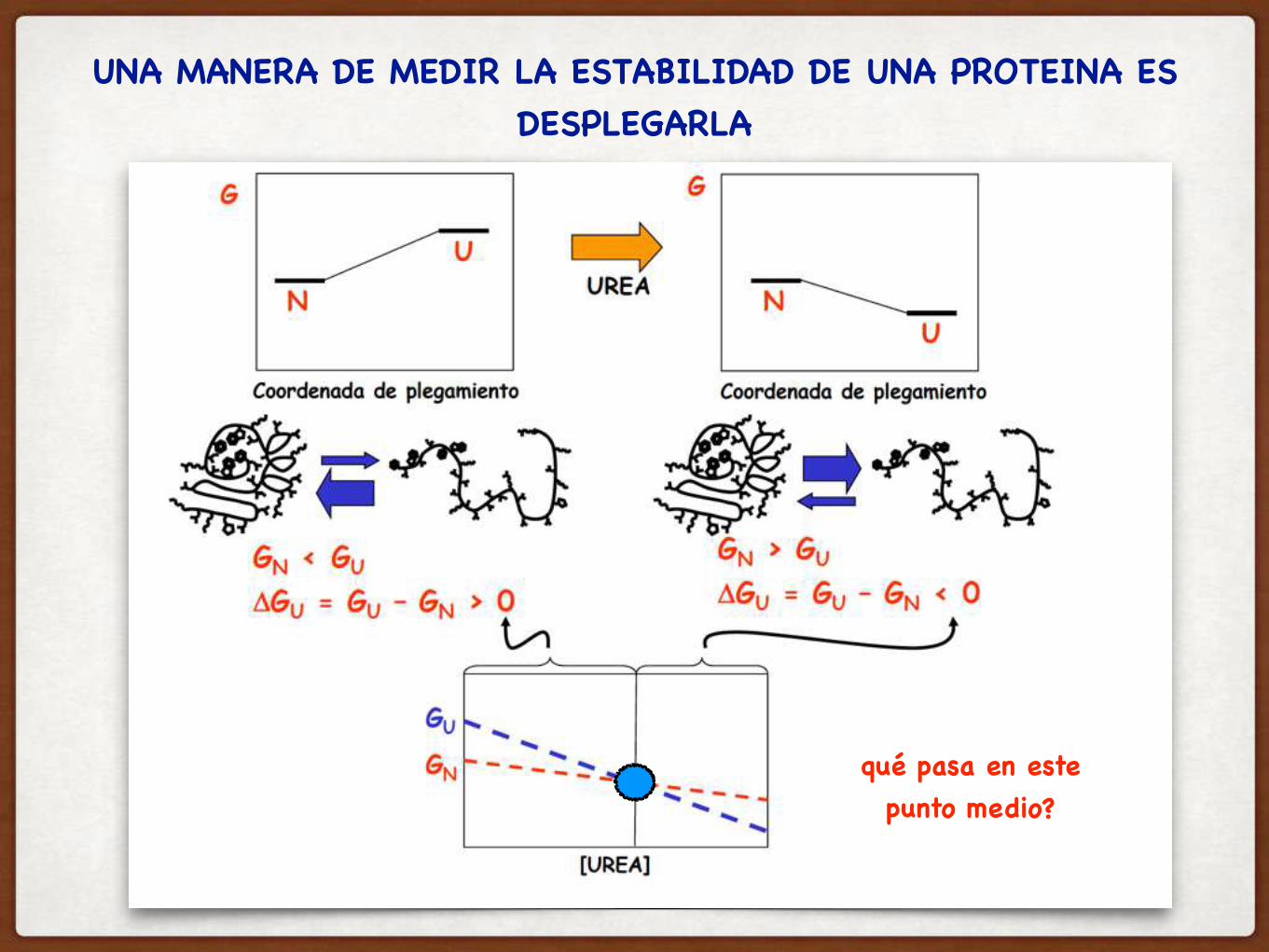
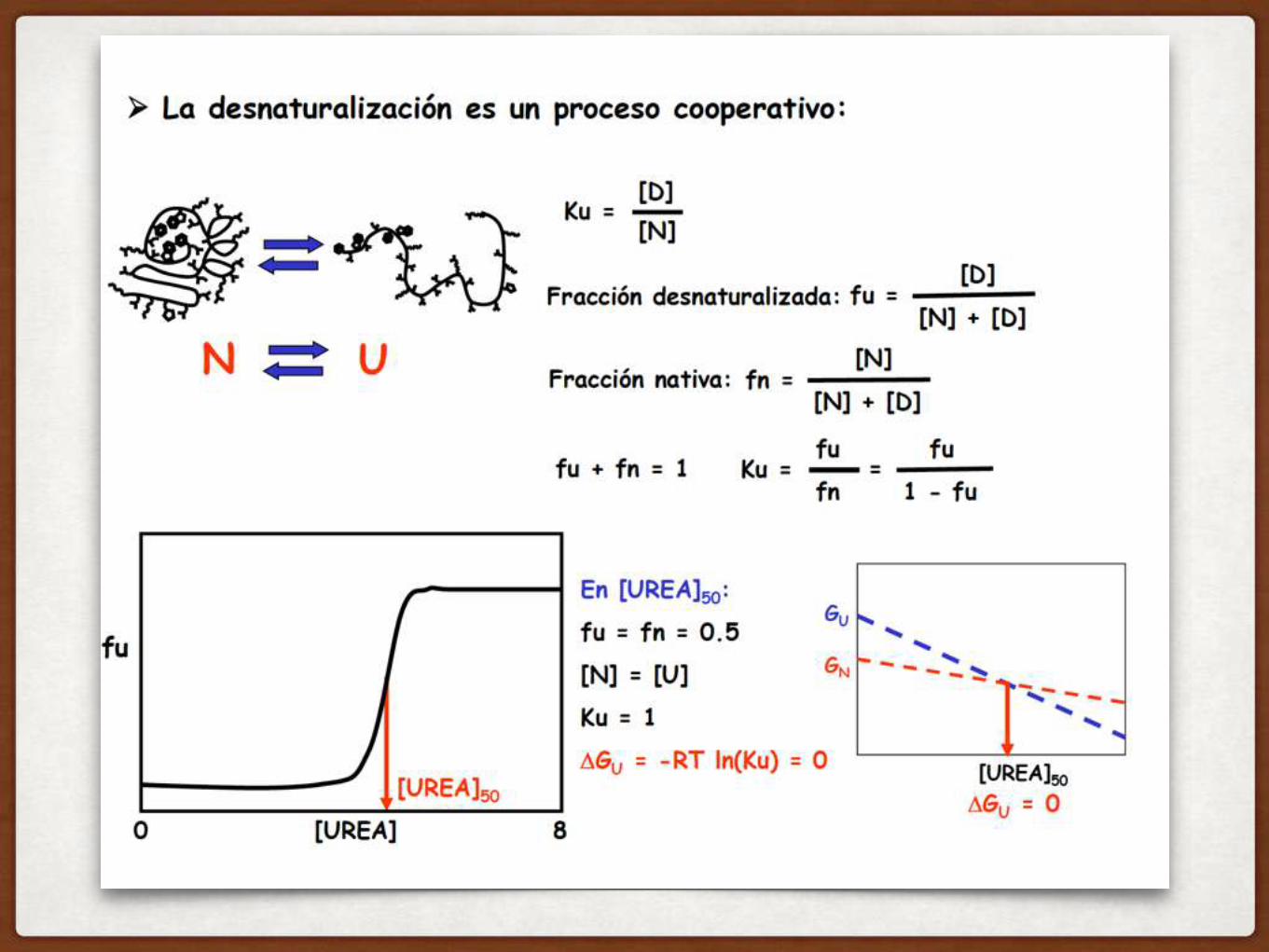
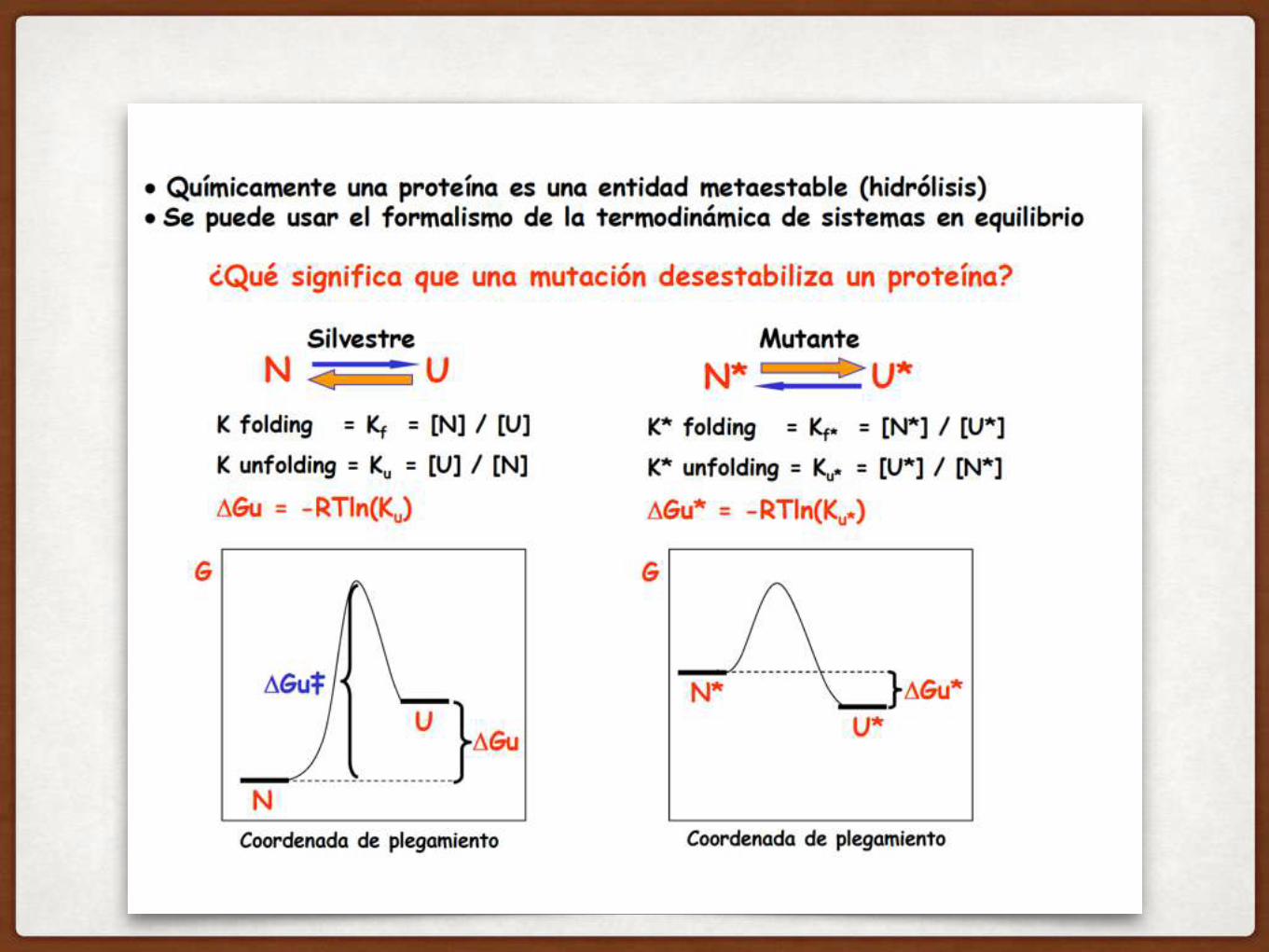
COMO DEFINIMOS LA ESTABILIDAD DE UNA PROTEÍNA?
LA ESTABILIDAD DE UNA PROTEINA SE MIDE CON EL
△GUNFOLDING
COORDENADA DE REACCION
ENER
GIA
LIB
RE
(DE
GIB
BS
)
ESTADO DESPLEGADO
ESTADO NATIVO
ESTADO DE TRANSICION
GU
GN
Hay que analizar la DIFERENCIA en energía libre de los dos estados:
△GU
Cómo sé si se pliega??
ENERGIA LIBRE DE UNFOLDING
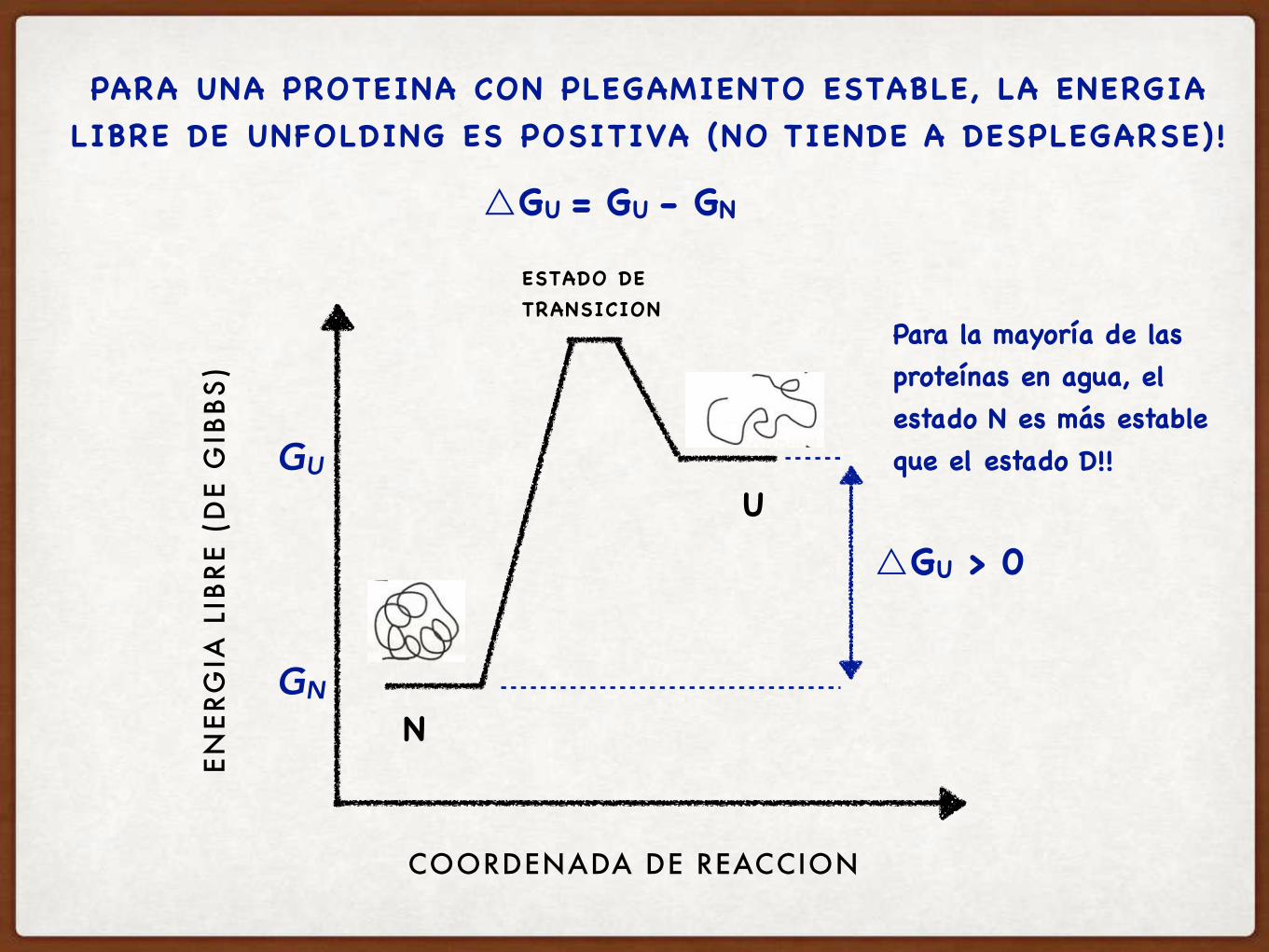
Para la mayoría de las proteínas en agua, el estado N es más estable que el estado D!!
PARA UNA PROTEINA CON PLEGAMIENTO ESTABLE, LA ENERGIA LIBRE DE UNFOLDING ES POSITIVA (NO TIENDE A DESPLEGARSE)!
△GU = GU - GN
△GU > 0
COORDENADA DE REACCION
ENER
GIA
LIB
RE
(DE
GIB
BS
)
N
ESTADO DE TRANSICION
GU
GN
U
COORDENADA DE REACCION
ENER
GIA
GU
GN
ESTADO DESPLEGADO
ESTADO NATIVO
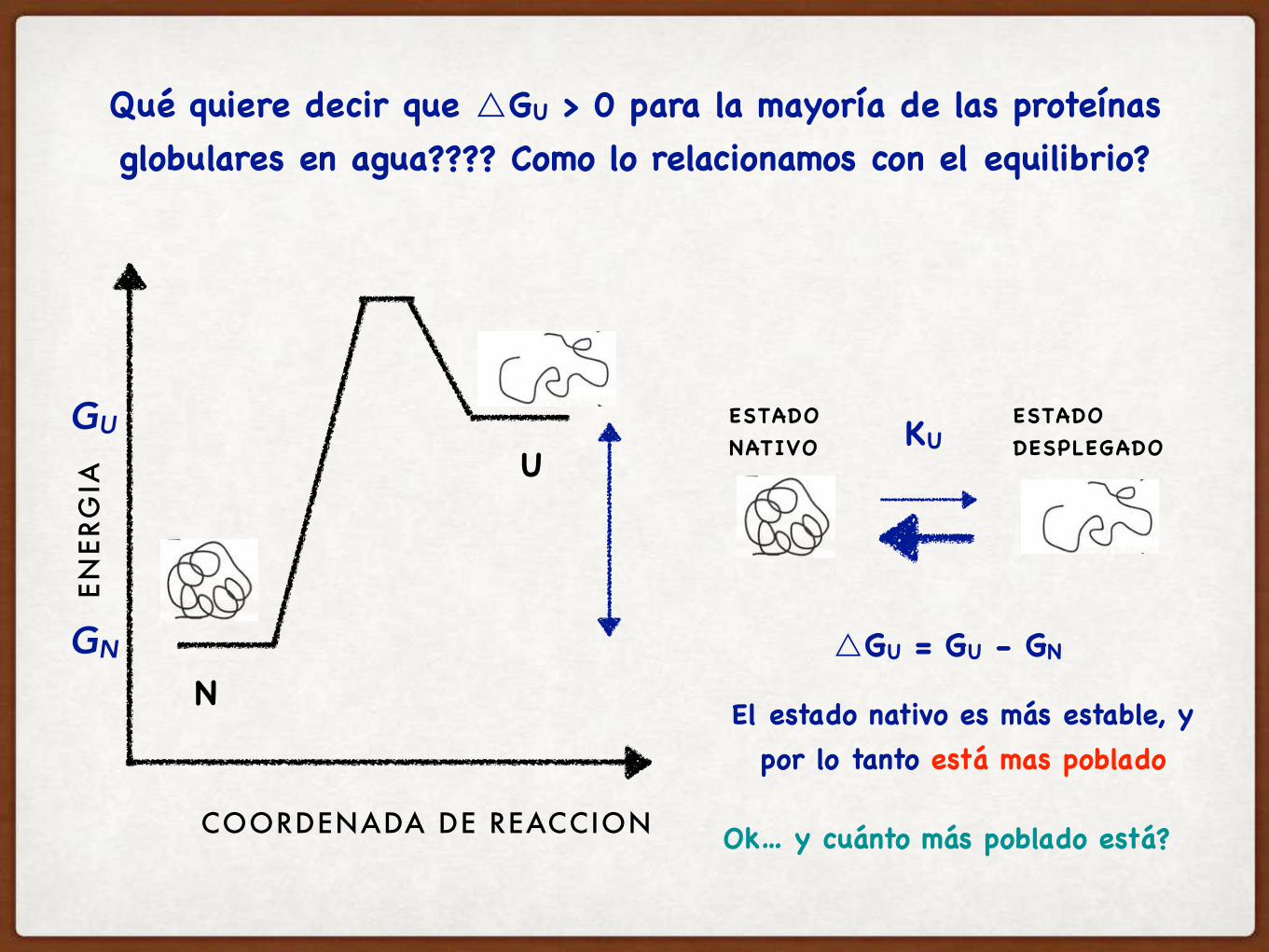
Qué quiere decir que △GU > 0 para la mayoría de las proteínas globulares en agua???? Como lo relacionamos con el equilibrio?
△GU = GU - GN
El estado nativo es más estable, y por lo tanto está mas poblado
Ok… y cuánto más poblado está?
N
UKU
ENER
GIA
N
U
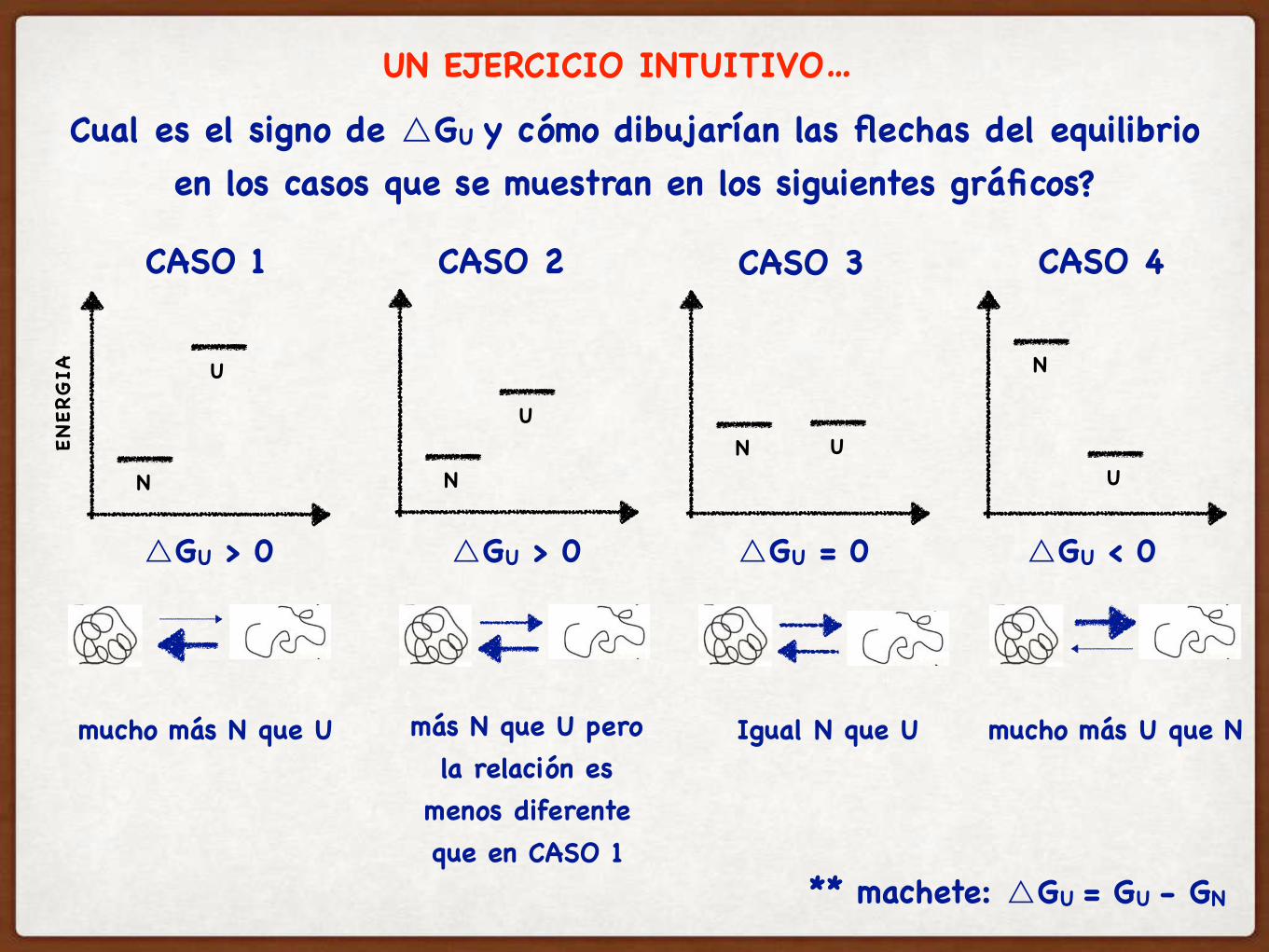
Cual es el signo de △GU y cómo dibujarían las flechas del equilibrio en los casos que se muestran en los siguientes gráficos?
CASO 1 CASO 2 CASO 3 CASO 4
N
UN U
N
U
mucho más N que U
△GU > 0
más N que U pero la relación es
menos diferente que en CASO 1
△GU > 0
Igual N que U
△GU = 0
mucho más U que N
△GU < 0
UN EJERCICIO INTUITIVO…
** machete: △GU = GU - GN

Cómo puedo calcular cuán poblado está cada estado??? Cómo se relaciona con el equilibrio que vemos acá??
ESTADO DESPLEGADO
ESTADO NATIVO
hay que definir la constante de equilibrio de esta reacción
KU
△GU = -RT ln (KU)
RT
Constante de los gasesTemperatura (K)
Además, la constante KU y △GU están directamente relacionados…cómo?
KU = [U]/[N] = fU/fN
Fracción de proteína desplegada
Fracción de proteína nativa

Volvamos al ejemplo y veamos cómo dan los números: calculemos KU para estas proporciones fU y fN
ENER
GIA
N
U
CASO 1 CASO 2 CASO 3 CASO 4
N
UN U
N
U
fN = 0.9fU = 0.1
KU = 0.1/0.9
KU = 0.111
fN = 0.7fU = 0.3
fN = 0.5fU = 0.5
fN = 0.1fU = 0.9
** machete: KU = fU/fN
KU = 0.3/0.7
KU = 0.43
△GU > 0 △GU > 0 △GU = 0 △GU < 0
KU = 0.5/0.5
KU = 1
KU = 0.9/0.1
KU = 9
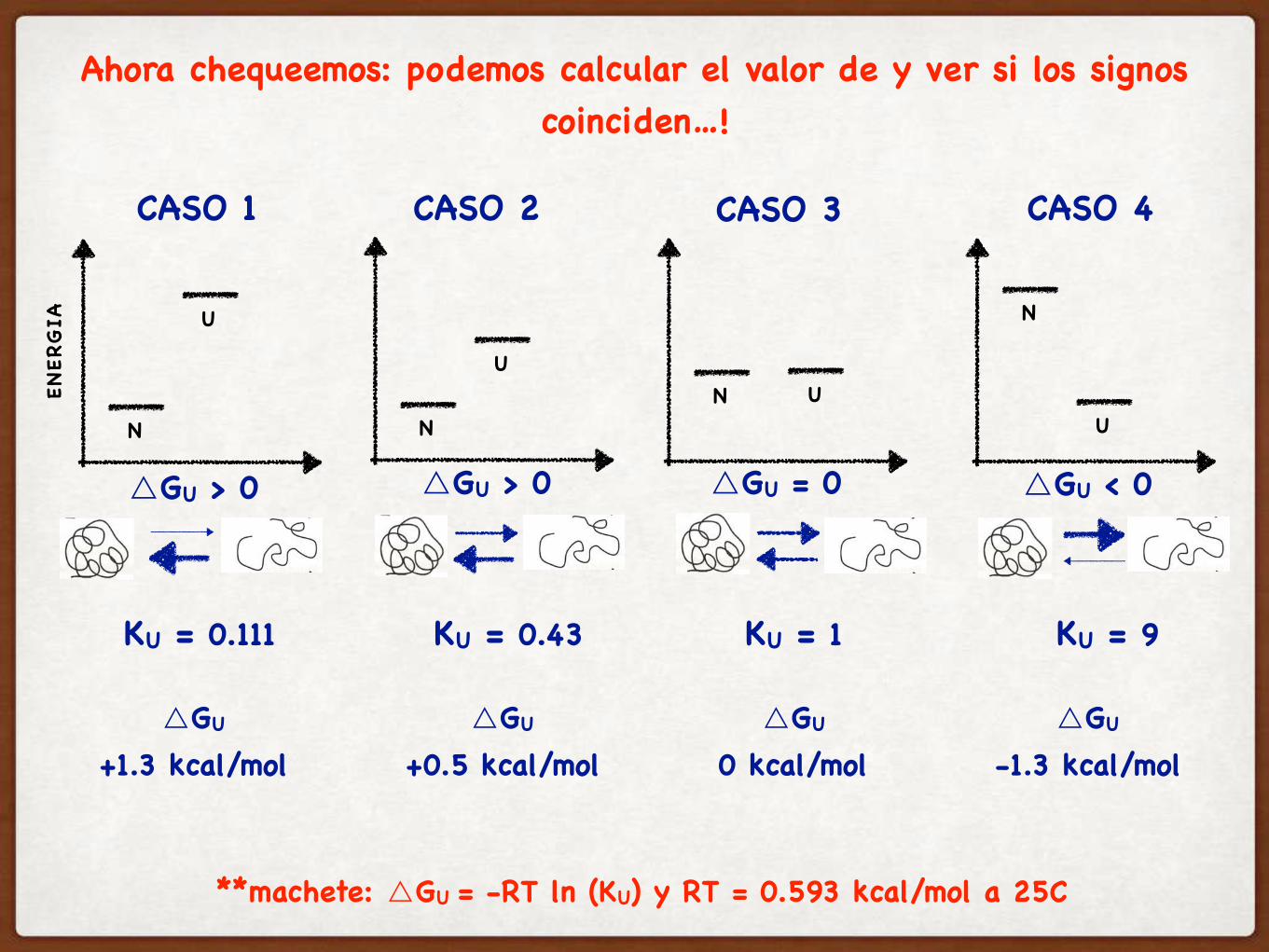
Ahora chequeemos: podemos calcular el valor de y ver si los signos coinciden…!
ENER
GIA
N
U
CASO 1 CASO 2 CASO 3 CASO 4
N
UN U
N
U
△GU > 0 △GU > 0 △GU = 0 △GU < 0
KU = 0.111 KU = 0.43 KU = 1 KU = 9
**machete: △GU = -RT ln (KU) y RT = 0.593 kcal/mol a 25C
△GU
+1.3 kcal/mol△GU
+0.5 kcal/mol△GU
0 kcal/mol△GU
-1.3 kcal/mol

LAS ESTABILIDADES DE LAS PROTEINAS “PLEGADAS” EN QUE RANGO ESTAN ???
Los rangos observados son ~ △GU de +5 a +15 kcal/mol
ENTONCES, CUANTA PROTEINA PLEGADA HAY??
en nuestro ejemplo, 90% proteína nativa correspondía a △GU = + 1.3kcal/mol
KU = e(-RT/△GU)
fN = 1/(1+ KU)
△GU = -RT ln (KU)
KU = fU/fN

LAS ESTABILIDADES DE LAS PROTEINAS “PLEGADAS” EN QUE RANGO ESTAN ???
Los rangos observados son ~ △GU de +5 a +15 kcal/mol
△GU = +5 kcal/mol
fN = 0,9999999999896 △GU = +15 kcal/mol
fN = 0,999782
ENTONCES, CUANTA PROTEINA PLEGADA HAY??
ESTADO DESPLEGADO
ESTADO NATIVO
EN AGUA, CASI TODA LA PROTEINA ESTA PLEGADA !!! SIN EMBARGO, NO ES MUY DIFICIL DESESTABILIZAR EL ESTADO PLEGADO…
el equilibrio esta MUY desplazado hacia la izquierda
en nuestro ejemplo, 90% proteína nativa correspondía a △GU = + 1.3kcal/mol
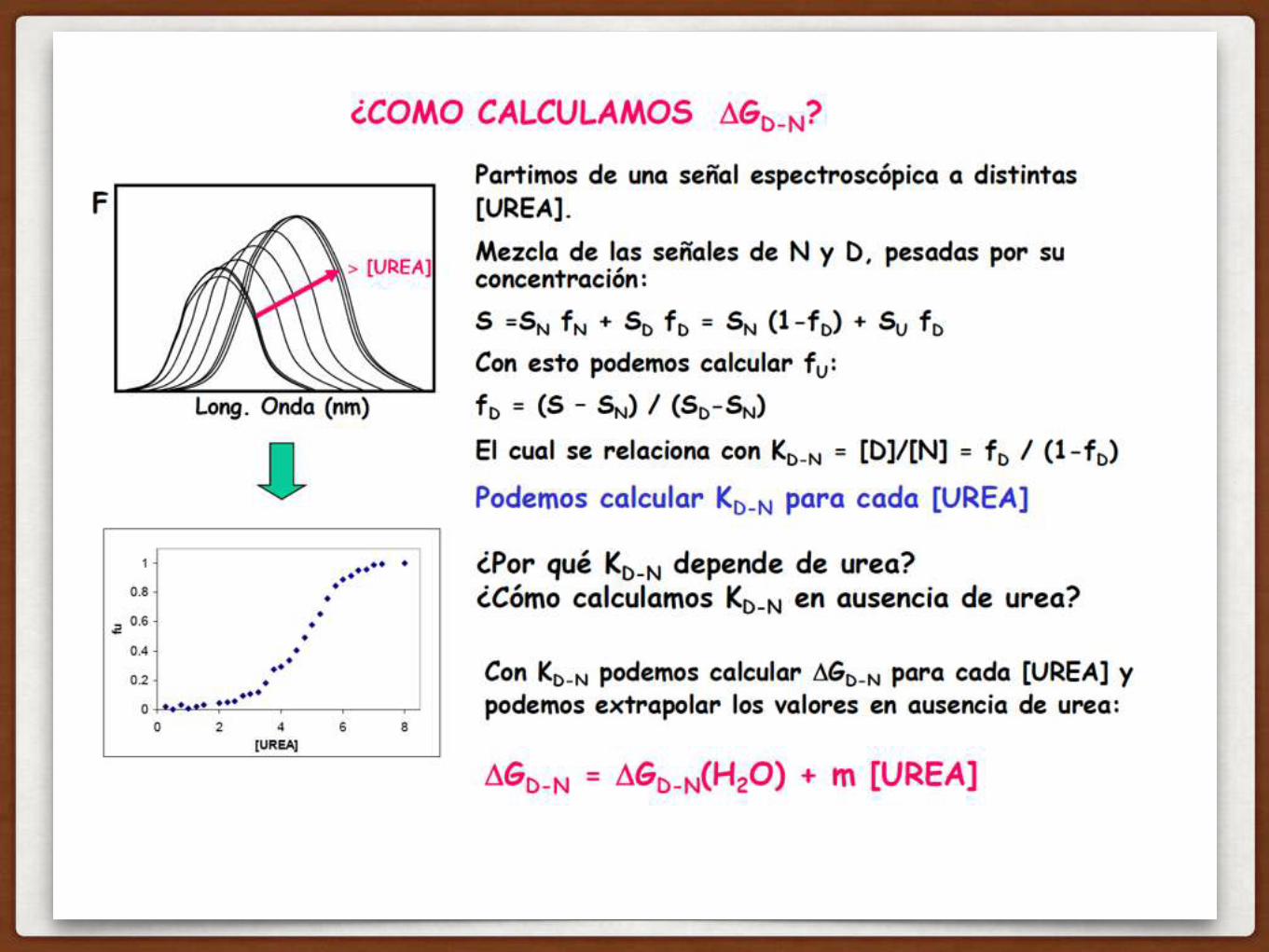
UNA MANERA DE MEDIR LA ESTABILIDAD DE UNA PROTEINA ES DESPLEGARLA
qué pasa en este punto medio?
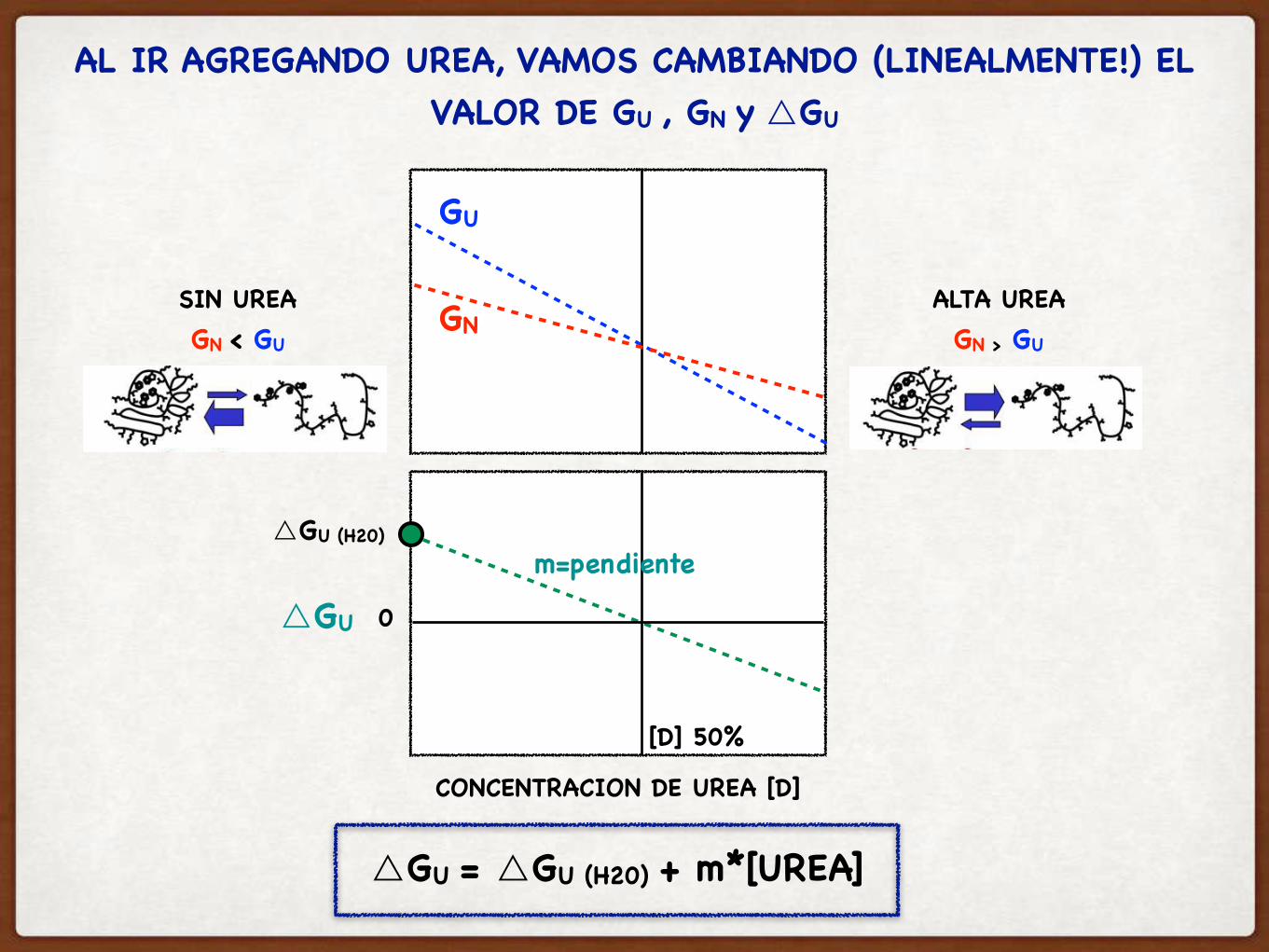
AL IR AGREGANDO UREA, VAMOS CAMBIANDO (LINEALMENTE!) EL VALOR DE GU , GN y △GU
CONCENTRACION DE UREA [D]
SIN UREAGN < GU
ALTA UREAGN > GU
GN
GU
△GU 0
[D] 50%
△GU = △GU (H20) + m*[UREA]
△GU (H20)m=pendiente
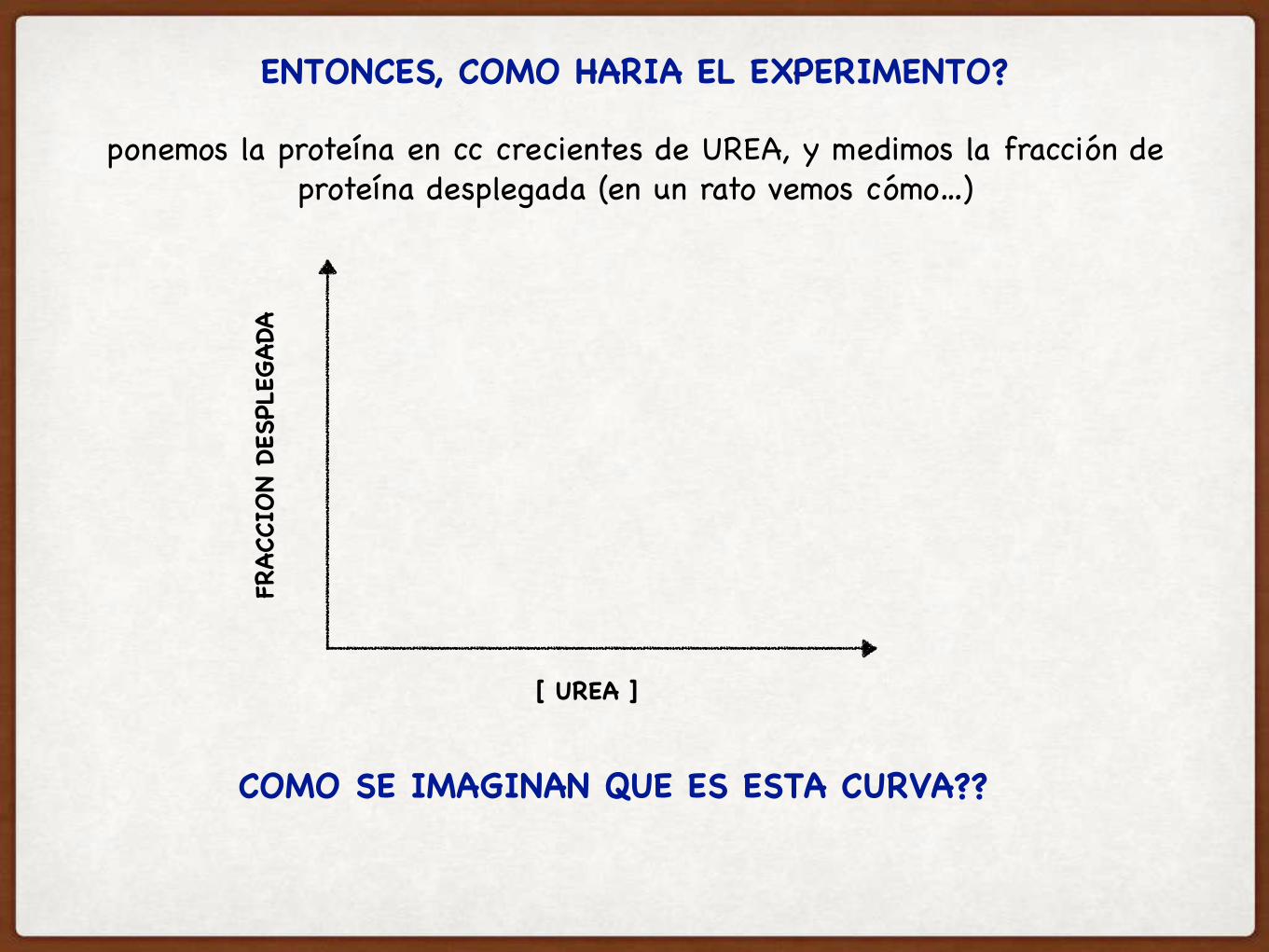
COMO SE IMAGINAN QUE ES ESTA CURVA??
FRAC
CION
DESP
LEGA
DA
[ UREA ]
ENTONCES, COMO HARIA EL EXPERIMENTO?
ponemos la proteína en cc crecientes de UREA, y medimos la fracción de proteína desplegada (en un rato vemos cómo…)

ES UNA CURVA ALTAMENTE COOPERATIVA: SE ROMPEN MUCHAS INTERACCIONES SIMULTANEAMENTE
la pendiente de la transición se relaciona con cuantos residuos se despliegan
FRAC
CION
DESP
LEGA
DA
[ UREA ]
ENTONCES, COMO HARIA EL EXPERIMENTO?

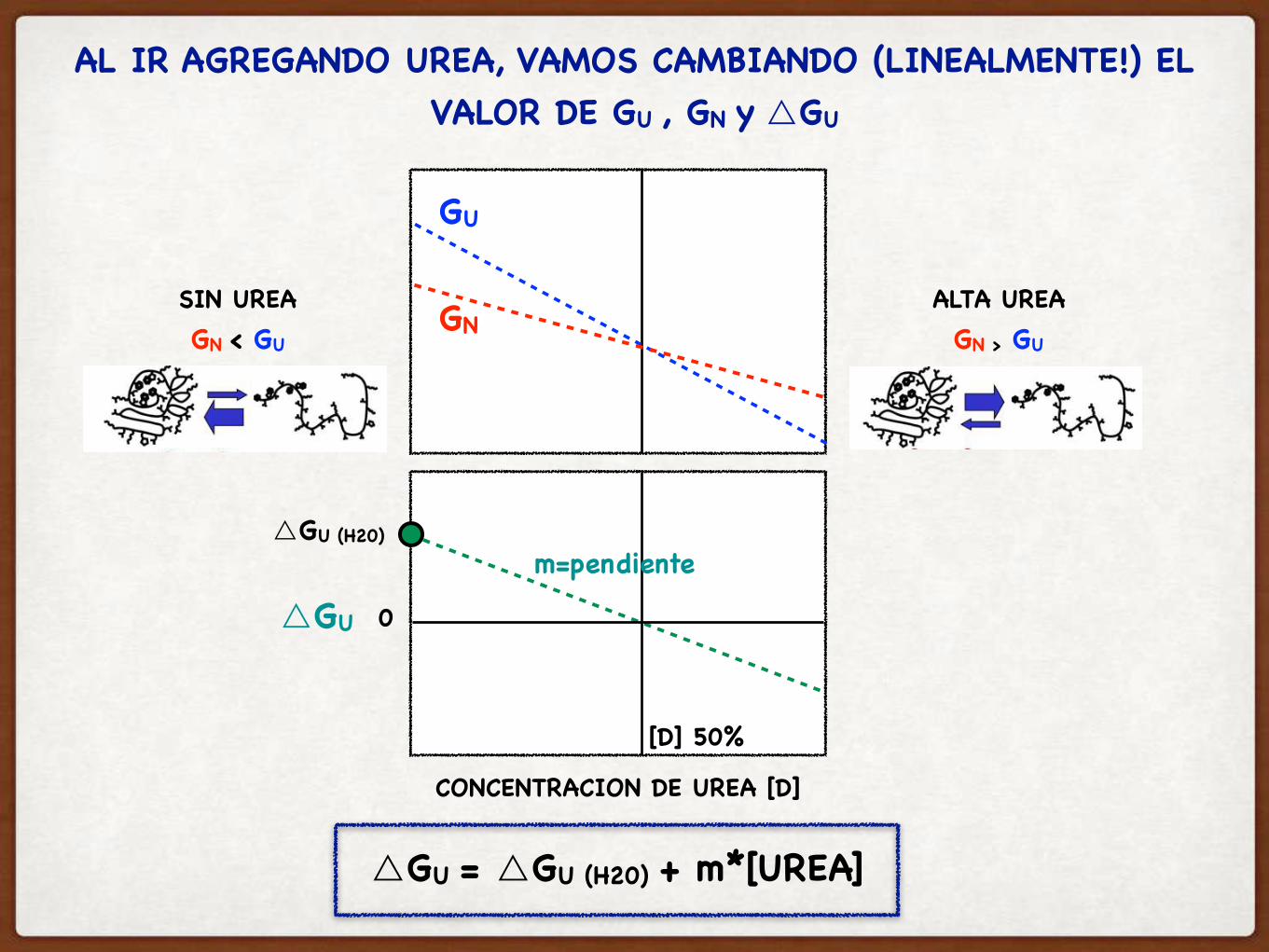
AL IR AGREGANDO UREA, VAMOS CAMBIANDO (LINEALMENTE!) EL VALOR DE GU , GN y △GU
CONCENTRACION DE UREA [D]
SIN UREAGN < GU
ALTA UREAGN > GU
GN
GU
△GU 0
[D] 50%
△GU = △GU (H20) + m*[UREA]
△GU (H20)m=pendiente

COMO MEDIMOS EL CONTENIDO DE ESTRUCTURA DE UNA PROTEINA?
ESTRUCTURA SECUNDARIA
ESTRUCTURA TERCIARIA
ESTRUCTURA CUATERNARIA
• DICROISMO CIRCULAR *• ESPECTROSCOPIA RAMAN• ESPECTROSCOPIA INFRARROJA• RESONANCIA MAGNETICA NUCLEAR *
• DICROISMO CIRCULAR *• EXCLUSION MOLECULAR (SEC)• FLUORESCENCIA INTRINSECA *
• EXCLUSION MOLECULAR (SEC)• CRISTALOGRAFIA DE RAYOS X *• DISPERSION DE LUZ LASER

METODOS PARA EL ANALISIS DE ESTRUCTURA SECUNDARIA
• DICROISMO CIRCULAR *• ESPECTROSCOPIA RAMAN• ESPECTROSCOPIA INFRARROJA• RESONANCIA MAGNETICA NUCLEAR *
TODOS LOS METODOS MIDEN DE DIFERENTE MANERA LA CONFIGURACION DEL ENLACE AMIDA…SE ACUERDAN DE RAMACHANDRAN?
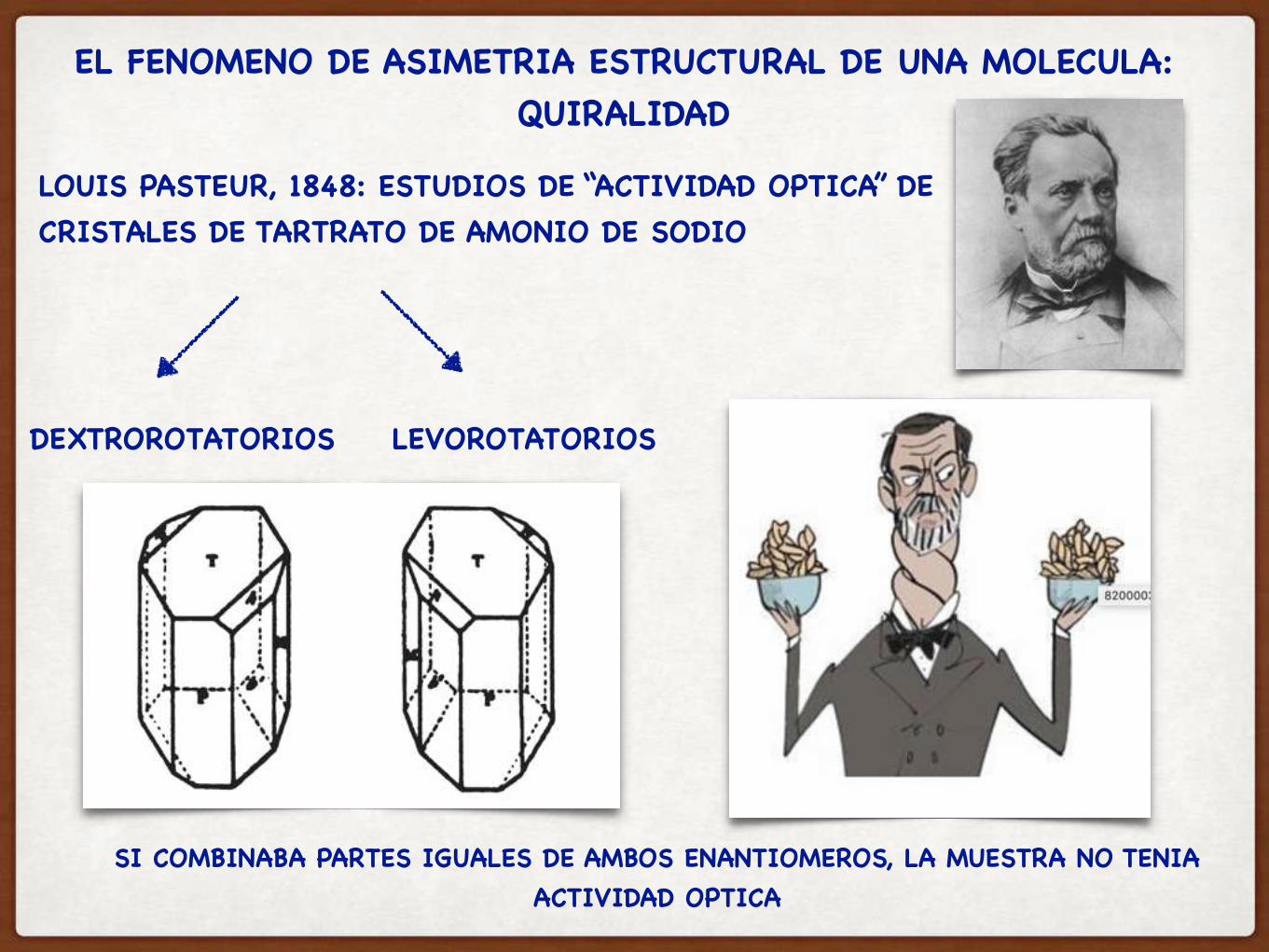
EL FENOMENO DE ASIMETRIA ESTRUCTURAL DE UNA MOLECULA: QUIRALIDAD
LOUIS PASTEUR, 1848: ESTUDIOS DE “ACTIVIDAD OPTICA” DE CRISTALES DE TARTRATO DE AMONIO DE SODIO
DEXTROROTATORIOS LEVOROTATORIOS
SI COMBINABA PARTES IGUALES DE AMBOS ENANTIOMEROS, LA MUESTRA NO TENIA ACTIVIDAD OPTICA
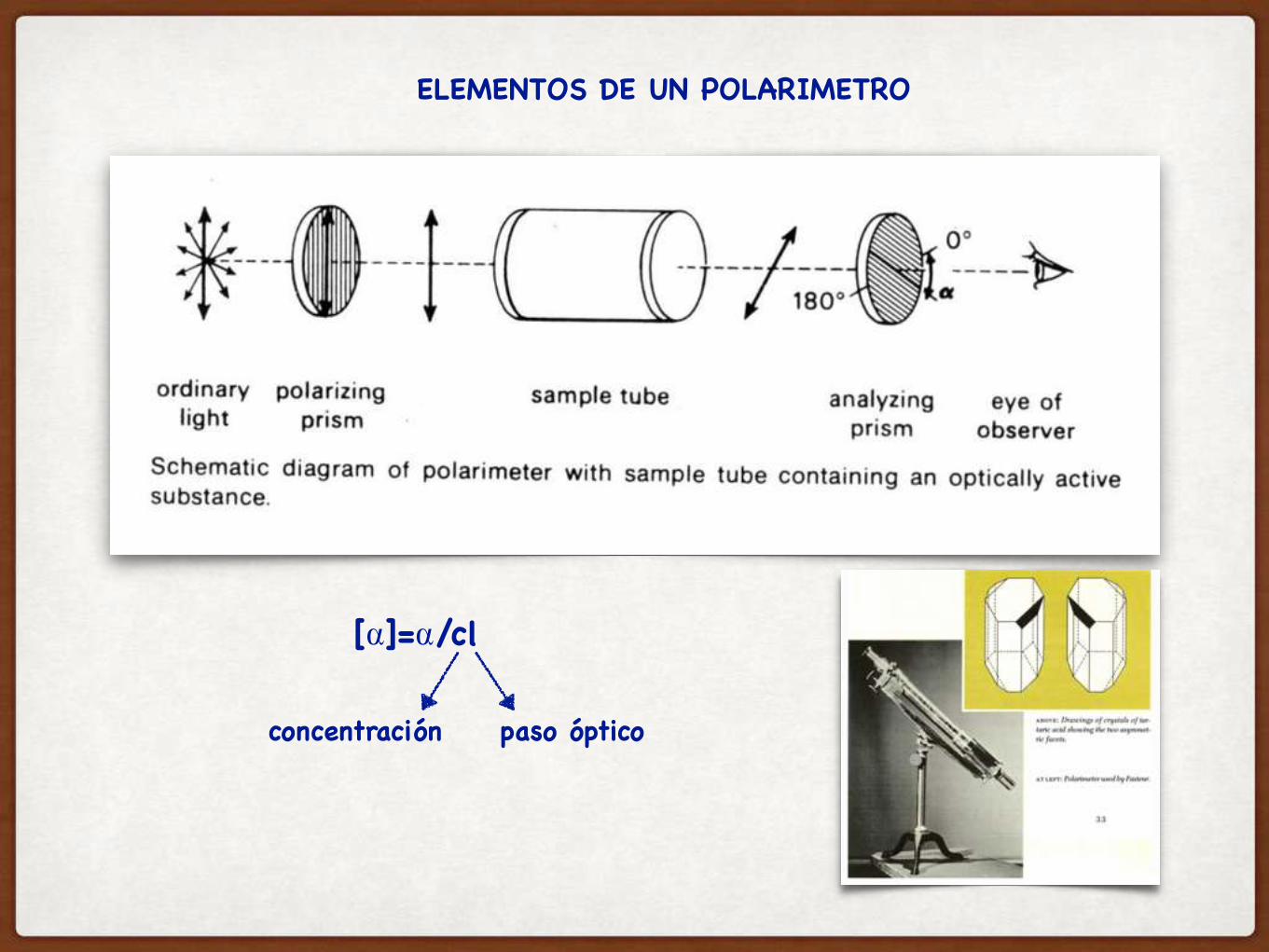
ELEMENTOS DE UN POLARIMETRO
[⍺]=⍺/cl
concentración paso óptico
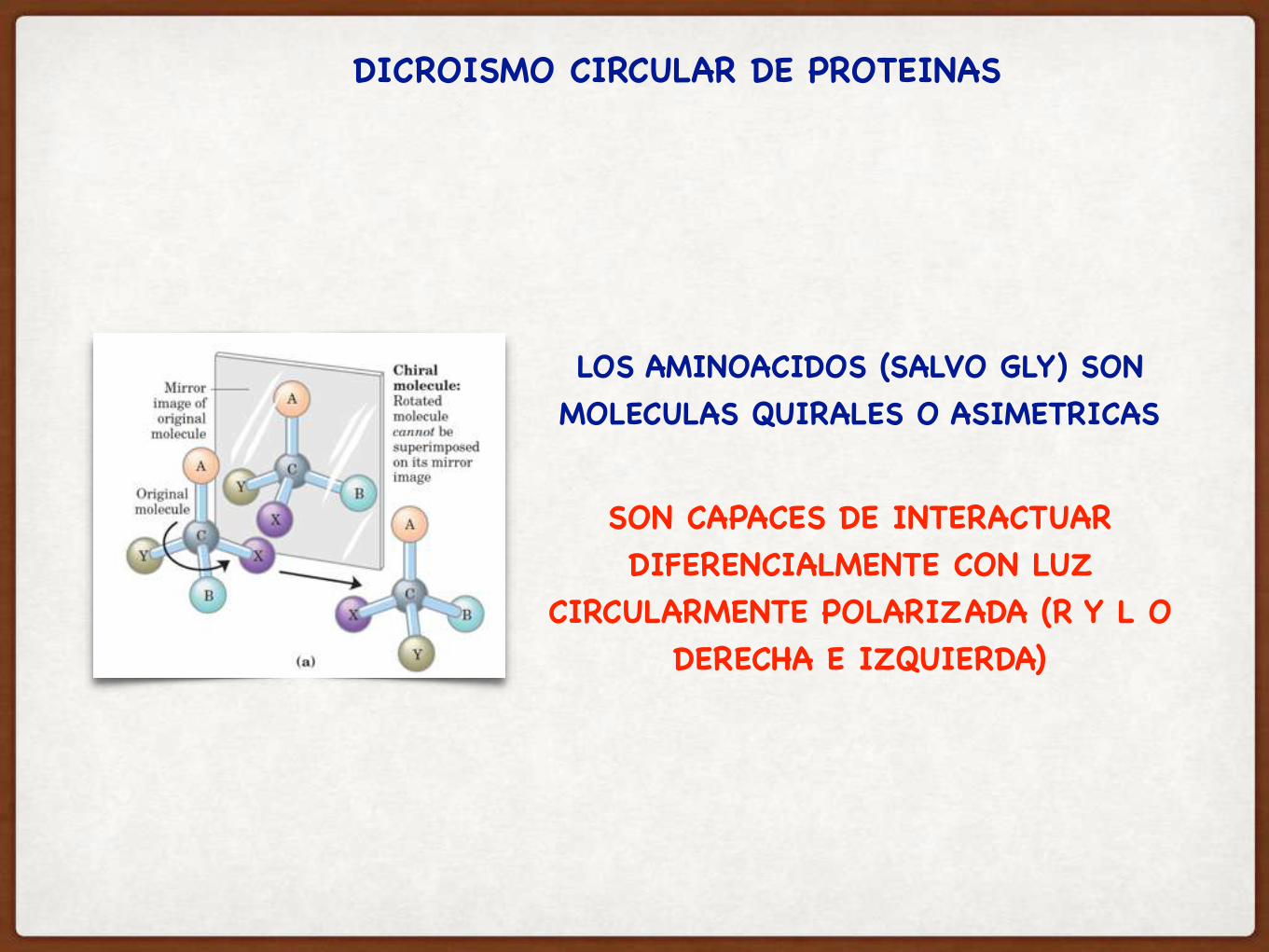
LOS AMINOACIDOS (SALVO GLY) SON MOLECULAS QUIRALES O ASIMETRICAS
DICROISMO CIRCULAR DE PROTEINAS
SON CAPACES DE INTERACTUAR DIFERENCIALMENTE CON LUZ
CIRCULARMENTE POLARIZADA (R Y L O DERECHA E IZQUIERDA)
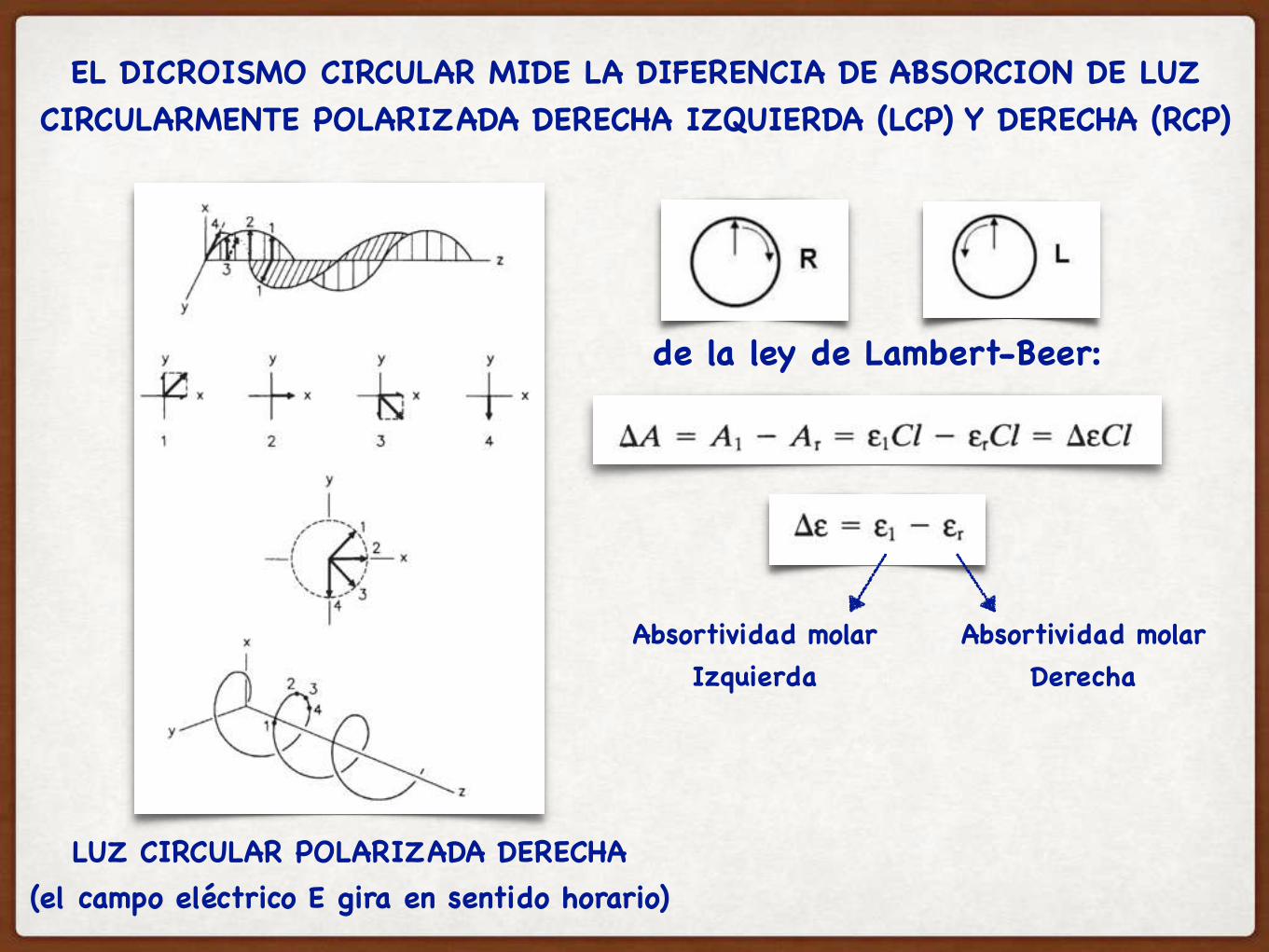
EL DICROISMO CIRCULAR MIDE LA DIFERENCIA DE ABSORCION DE LUZ CIRCULARMENTE POLARIZADA DERECHA IZQUIERDA (LCP) Y DERECHA (RCP)
de la ley de Lambert-Beer:
LUZ CIRCULAR POLARIZADA DERECHA(el campo eléctrico E gira en sentido horario)
Absortividad molar Derecha
Absortividad molar Izquierda
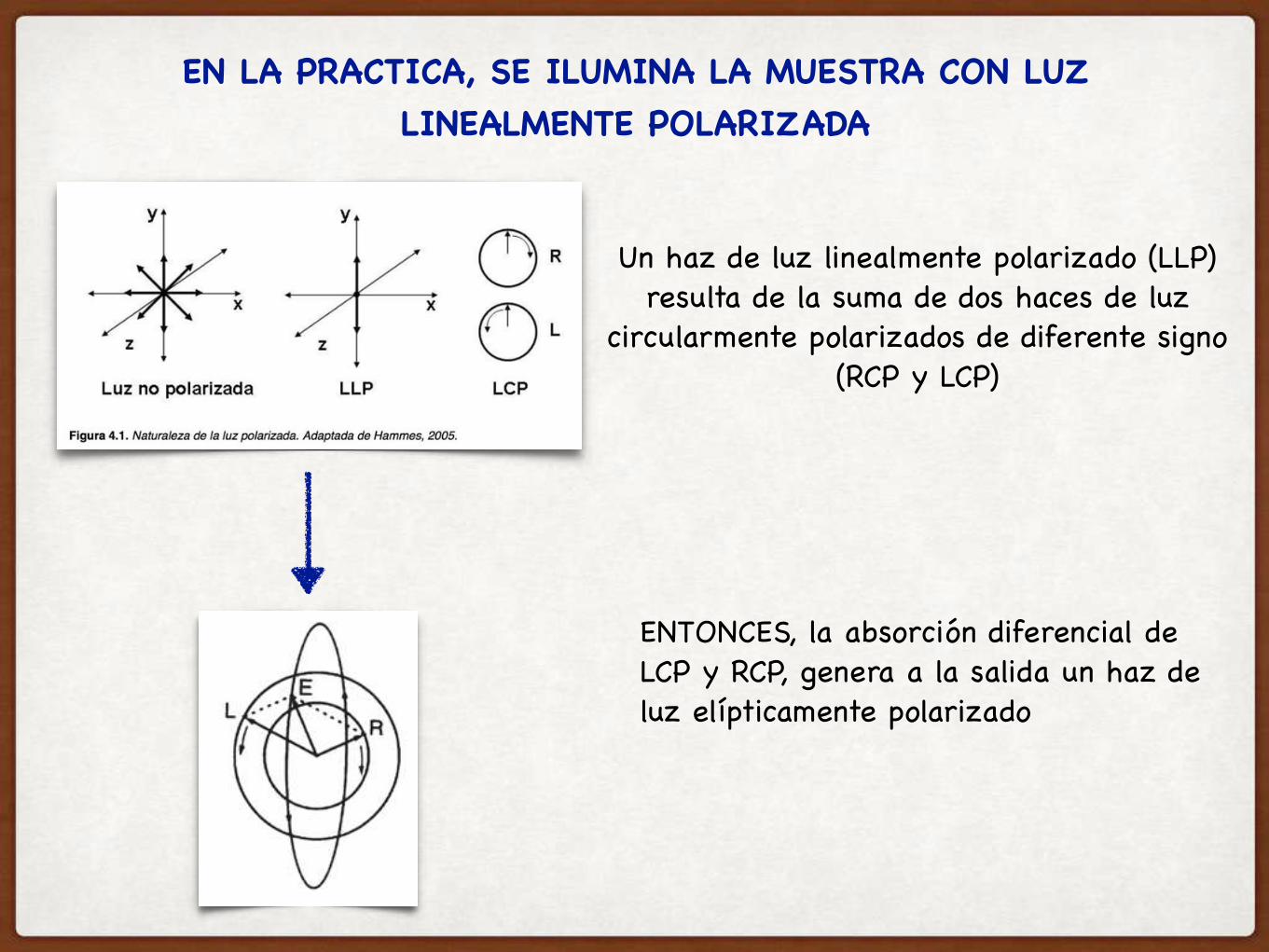
EN LA PRACTICA, SE ILUMINA LA MUESTRA CON LUZ LINEALMENTE POLARIZADA
Un haz de luz linealmente polarizado (LLP) resulta de la suma de dos haces de luz
circularmente polarizados de diferente signo (RCP y LCP)
ENTONCES, la absorción diferencial de LCP y RCP, genera a la salida un haz de luz elípticamente polarizado

Elipticidad
Elipticidad molar
EL CD SE EXPRESA EN UNIDADES DE ELIPTICIDAD
El eje mayor de la elipse corresponde a EL+ER (b)
El eje menor de la elipse corresponde a EL-ER (o)
para ángulos pequeños, podemos aproximar:
Definimos la elipticidad por el ángulo tita
tan = CO/CA = o/b
unidades: deg.cm2.dmol-1
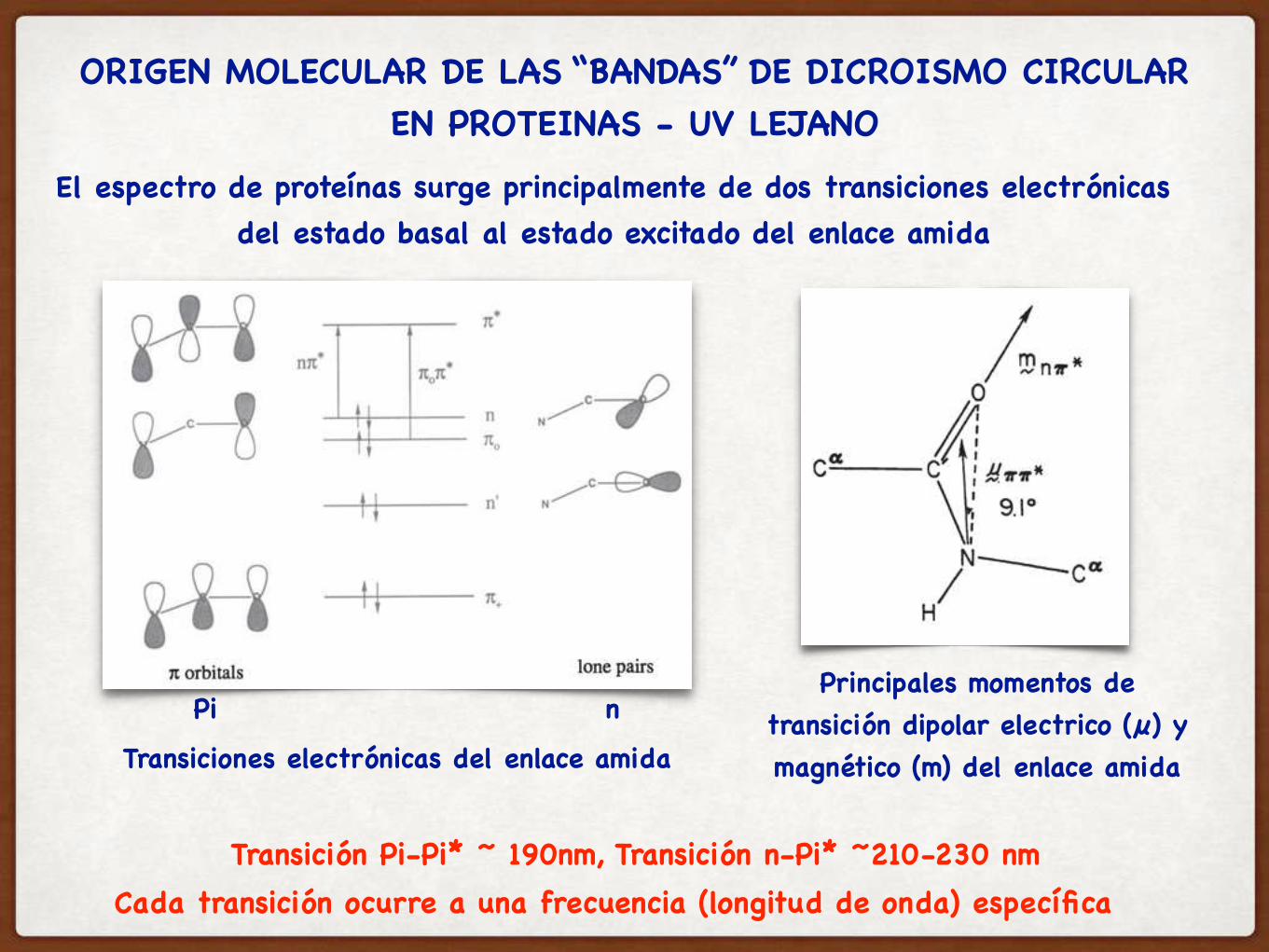
ORIGEN MOLECULAR DE LAS “BANDAS” DE DICROISMO CIRCULAR EN PROTEINAS - UV LEJANO
El espectro de proteínas surge principalmente de dos transiciones electrónicas del estado basal al estado excitado del enlace amida
Principales momentos de transición dipolar electrico (µ) y magnético (m) del enlace amida
Cada transición ocurre a una frecuencia (longitud de onda) específica
Transiciones electrónicas del enlace amida
Transición Pi-Pi* ~ 190nm, Transición n-Pi* ~210-230 nm
Pi n
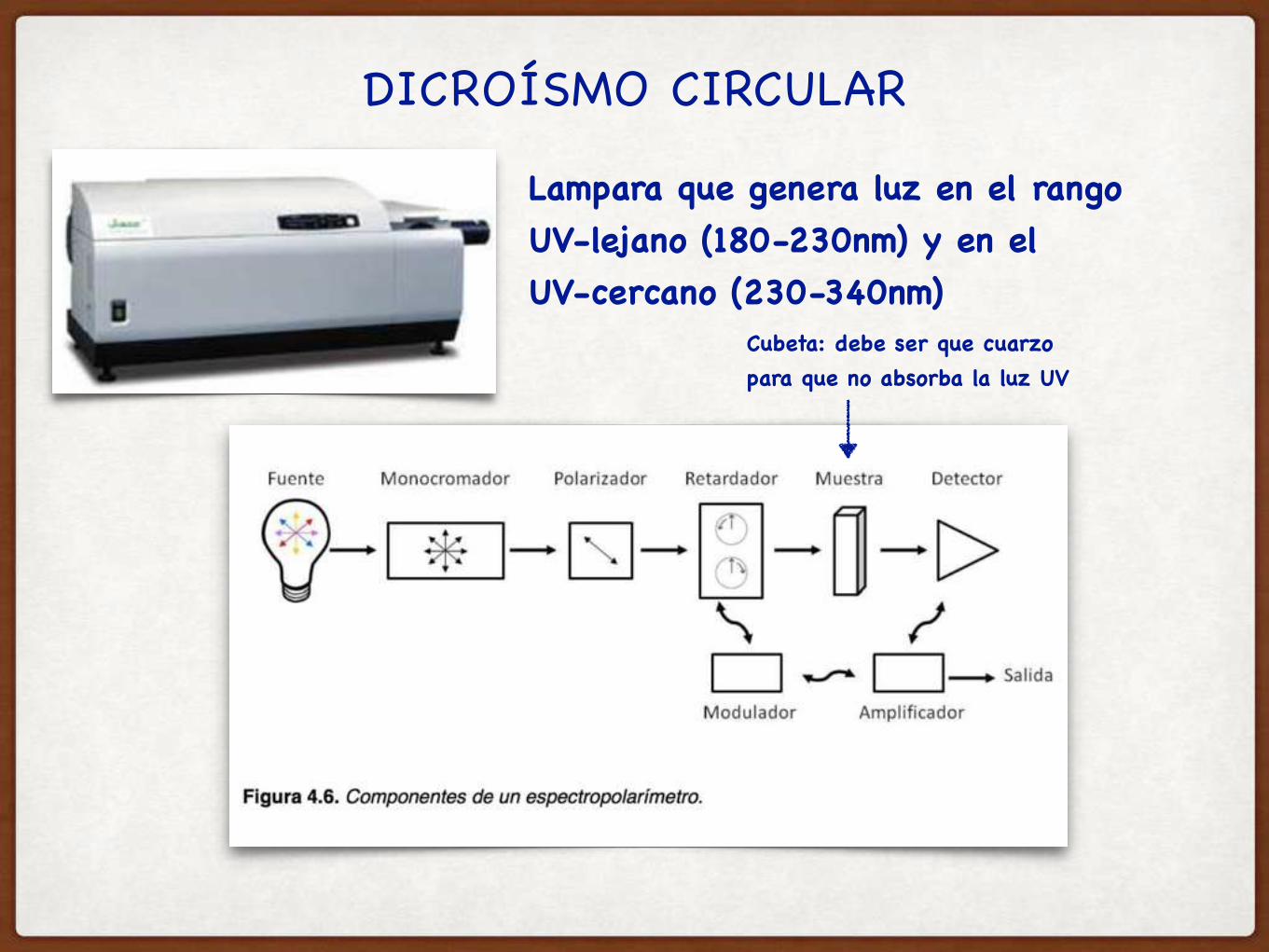
DICROÍSMO CIRCULAR
Lampara que genera luz en el rango UV-lejano (180-230nm) y en el UV-cercano (230-340nm)
Cubeta: debe ser que cuarzo para que no absorba la luz UV
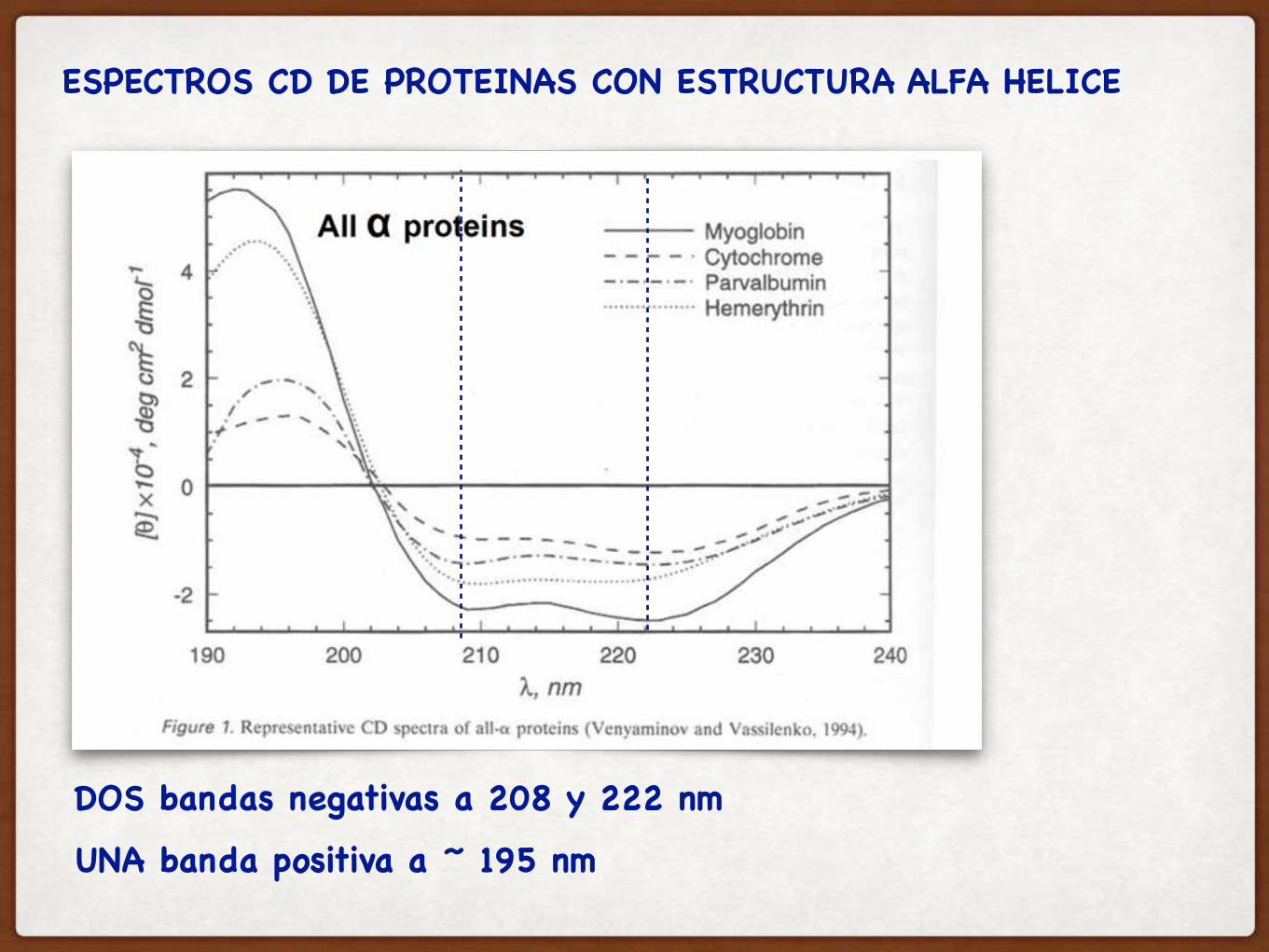
ESPECTROS CD DE PROTEINAS CON ESTRUCTURA ALFA HELICE
DOS bandas negativas a 208 y 222 nm
UNA banda positiva a ~ 195 nm
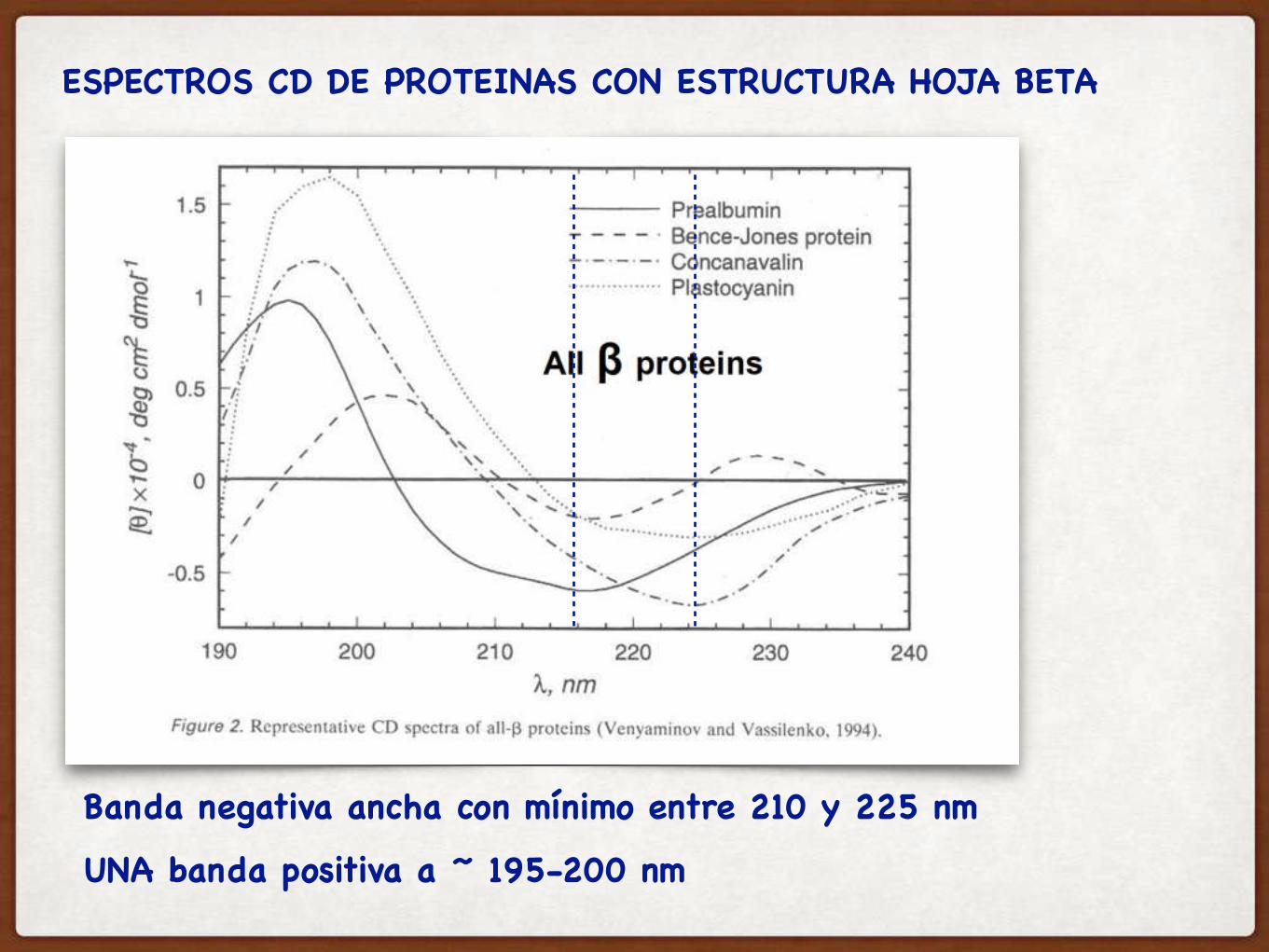
Banda negativa ancha con mínimo entre 210 y 225 nm
UNA banda positiva a ~ 195-200 nm
ESPECTROS CD DE PROTEINAS CON ESTRUCTURA HOJA BETA
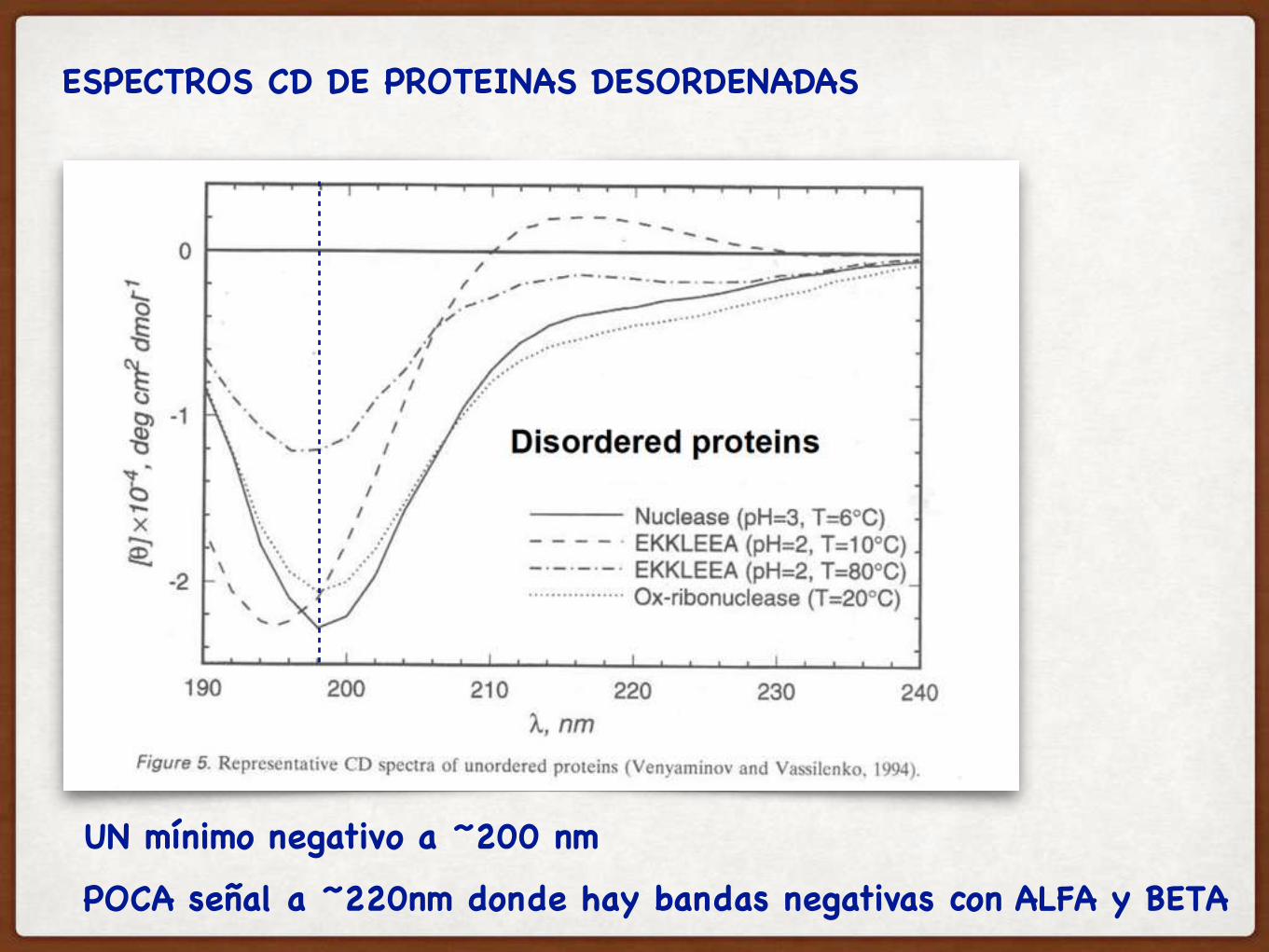
ESPECTROS CD DE PROTEINAS DESORDENADAS
UN mínimo negativo a ~200 nm
POCA señal a ~220nm donde hay bandas negativas con ALFA y BETA
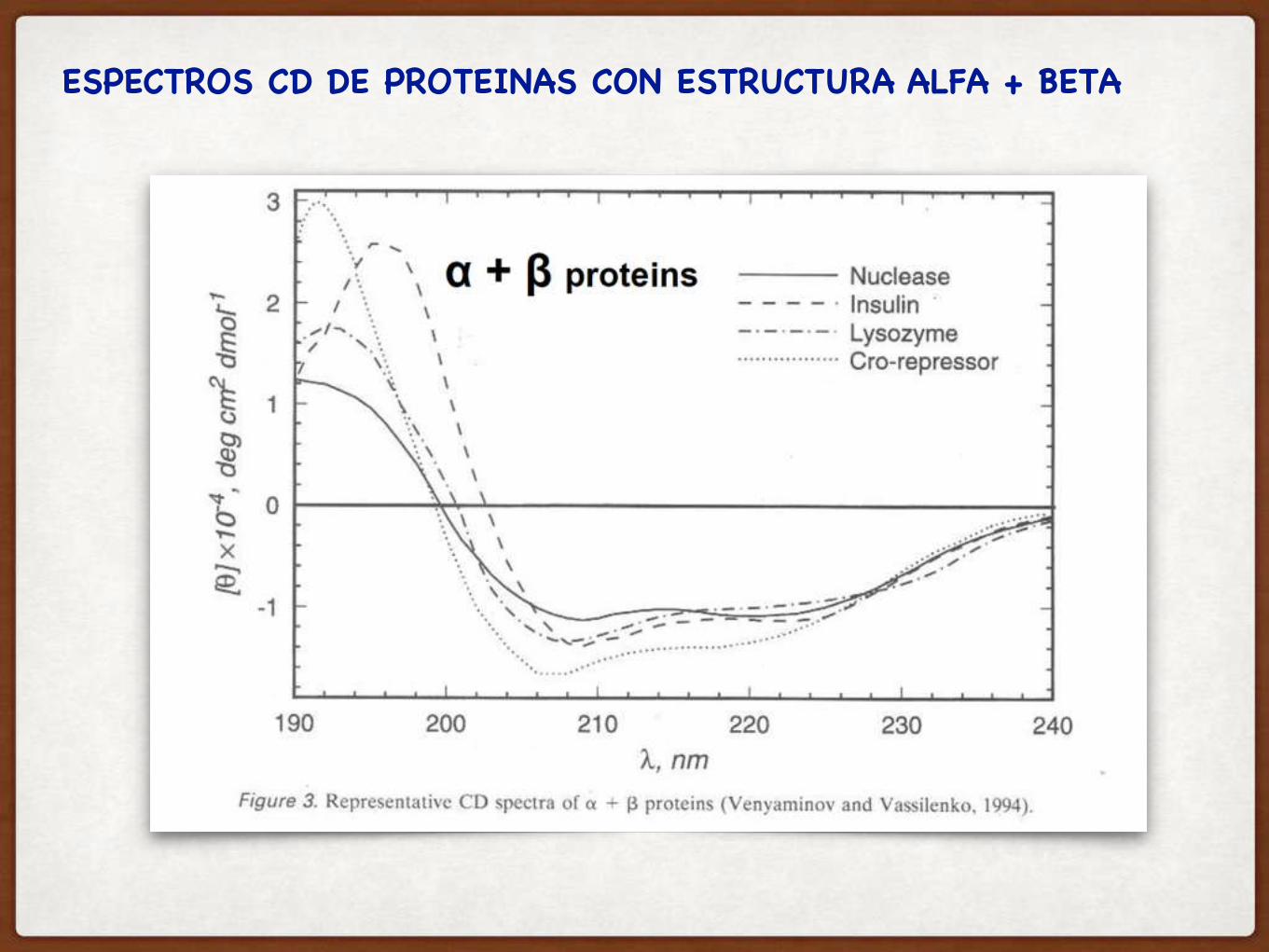
ESPECTROS CD DE PROTEINAS CON ESTRUCTURA ALFA + BETA
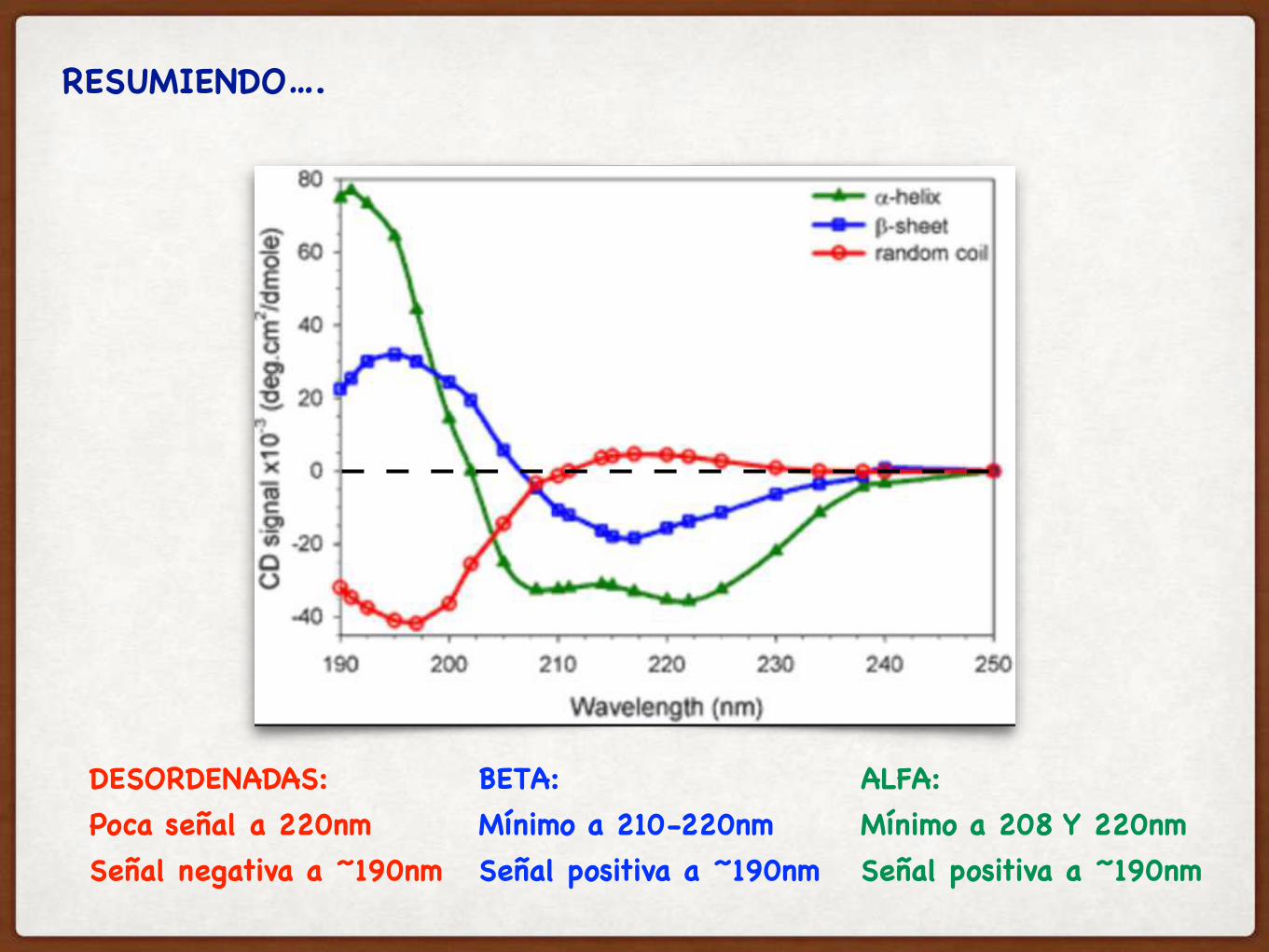
RESUMIENDO….
DESORDENADAS: Poca señal a 220nmSeñal negativa a ~190nm
BETA: Mínimo a 210-220nmSeñal positiva a ~190nm
ALFA: Mínimo a 208 Y 220nmSeñal positiva a ~190nm
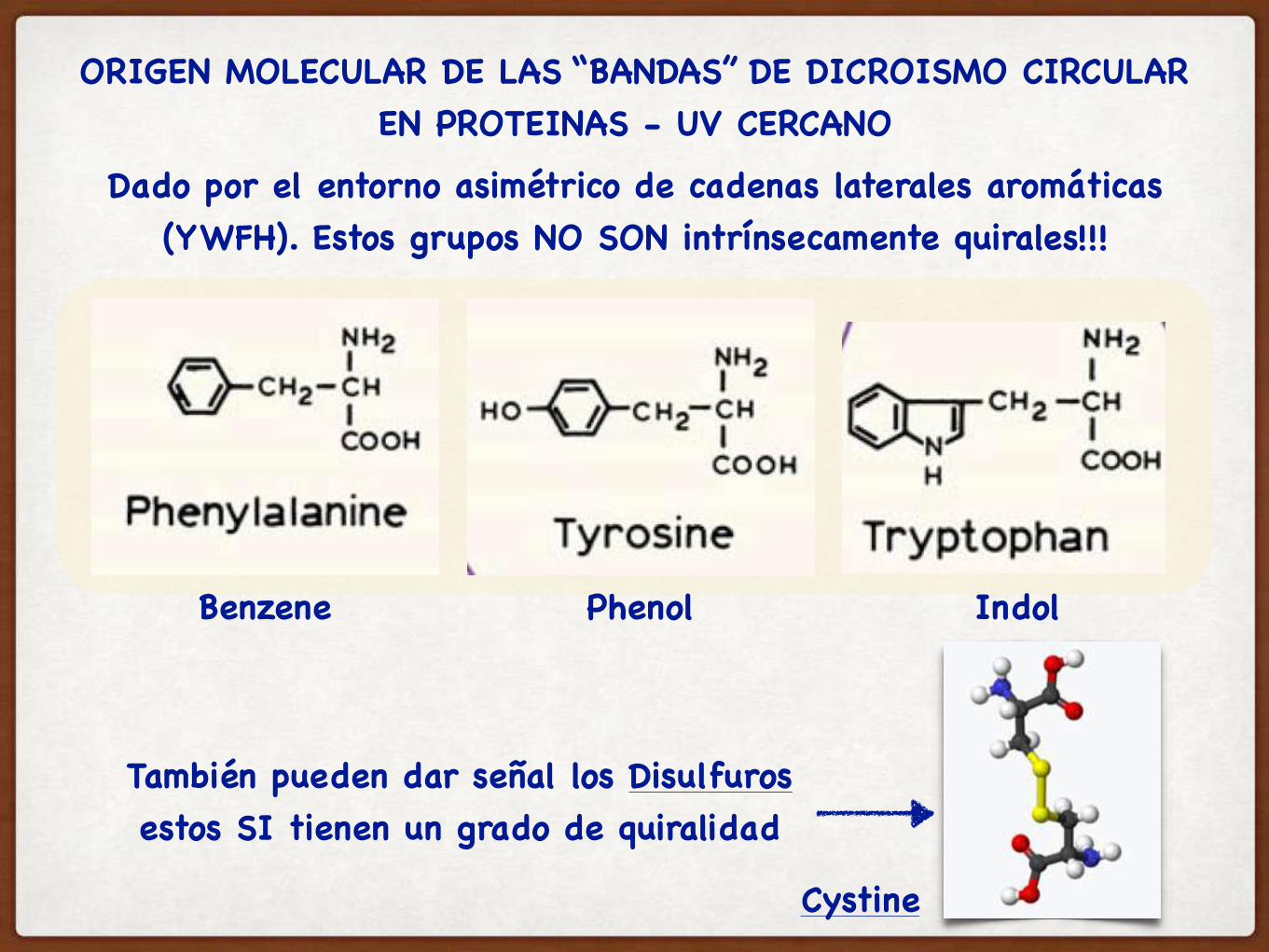
ORIGEN MOLECULAR DE LAS “BANDAS” DE DICROISMO CIRCULAR EN PROTEINAS - UV CERCANO
Dado por el entorno asimétrico de cadenas laterales aromáticas (YWFH). Estos grupos NO SON intrínsecamente quirales!!!
Benzene Phenol Indol
También pueden dar señal los Disulfurosestos SI tienen un grado de quiralidad
Cystine
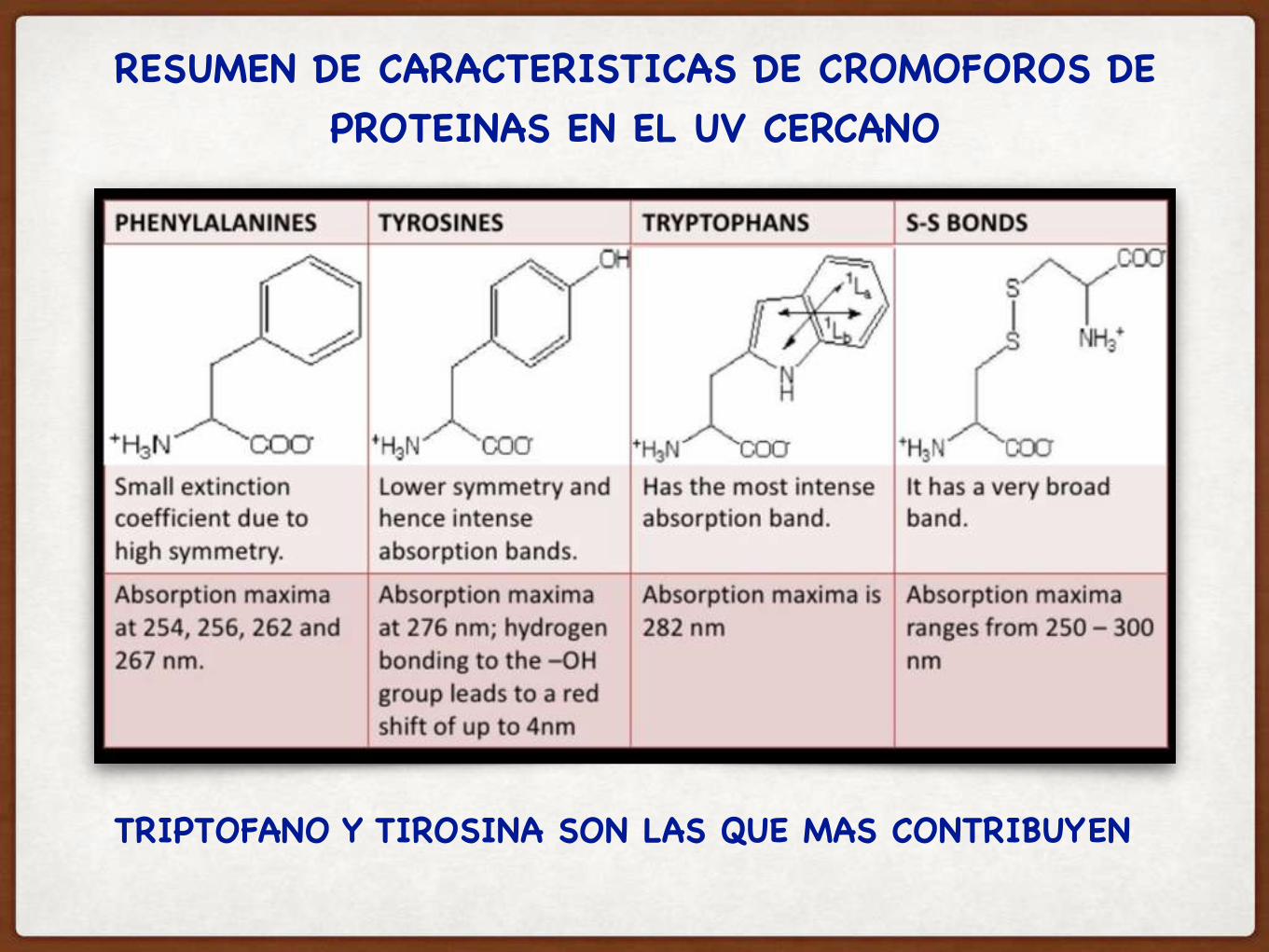
RESUMEN DE CARACTERISTICAS DE CROMOFOROS DE PROTEINAS EN EL UV CERCANO
TRIPTOFANO Y TIROSINA SON LAS QUE MAS CONTRIBUYEN
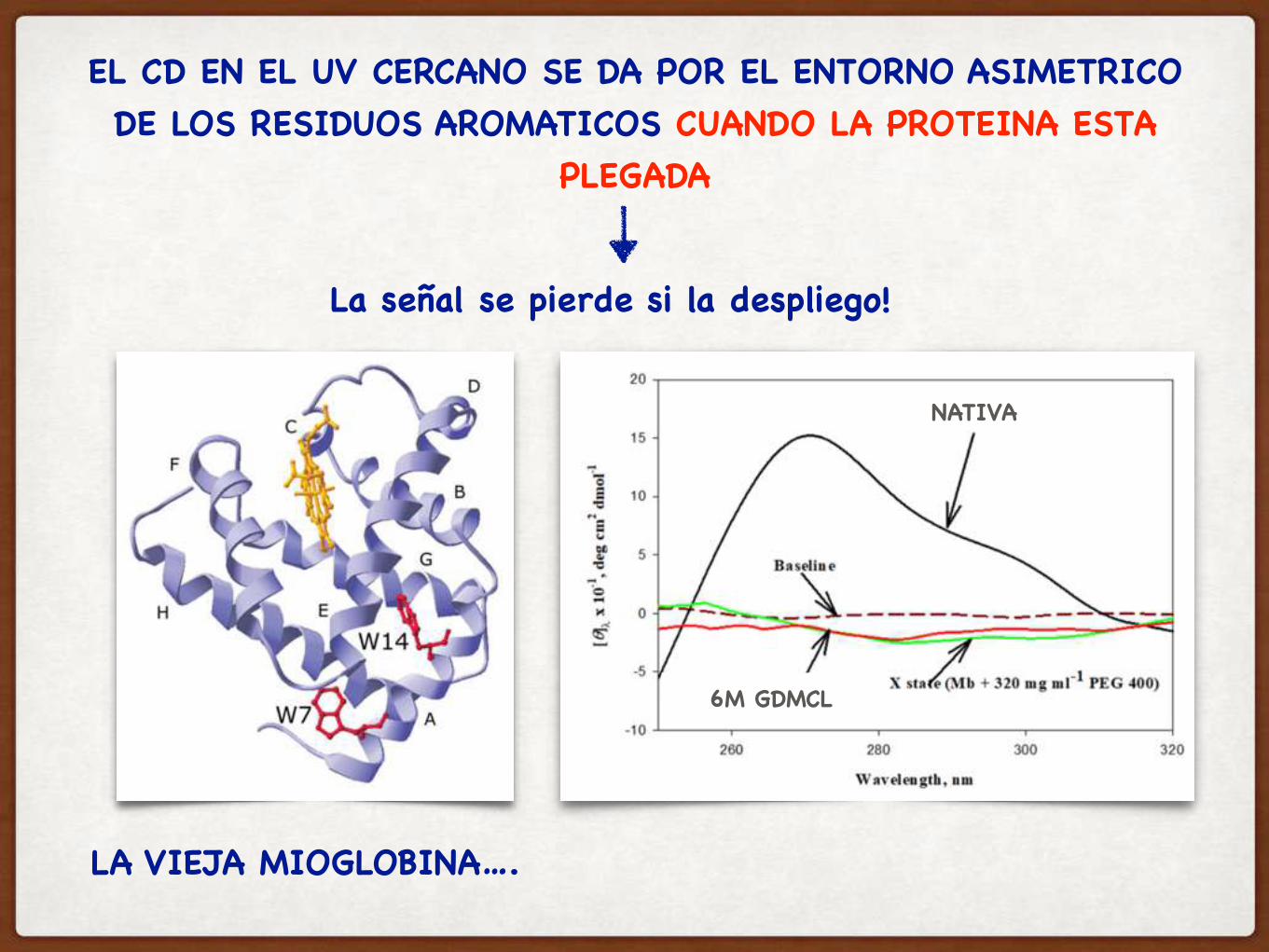
EL CD EN EL UV CERCANO SE DA POR EL ENTORNO ASIMETRICO DE LOS RESIDUOS AROMATICOS CUANDO LA PROTEINA ESTA
PLEGADA
LA VIEJA MIOGLOBINA….
NATIVA
6M GDMCL
La señal se pierde si la despliego!
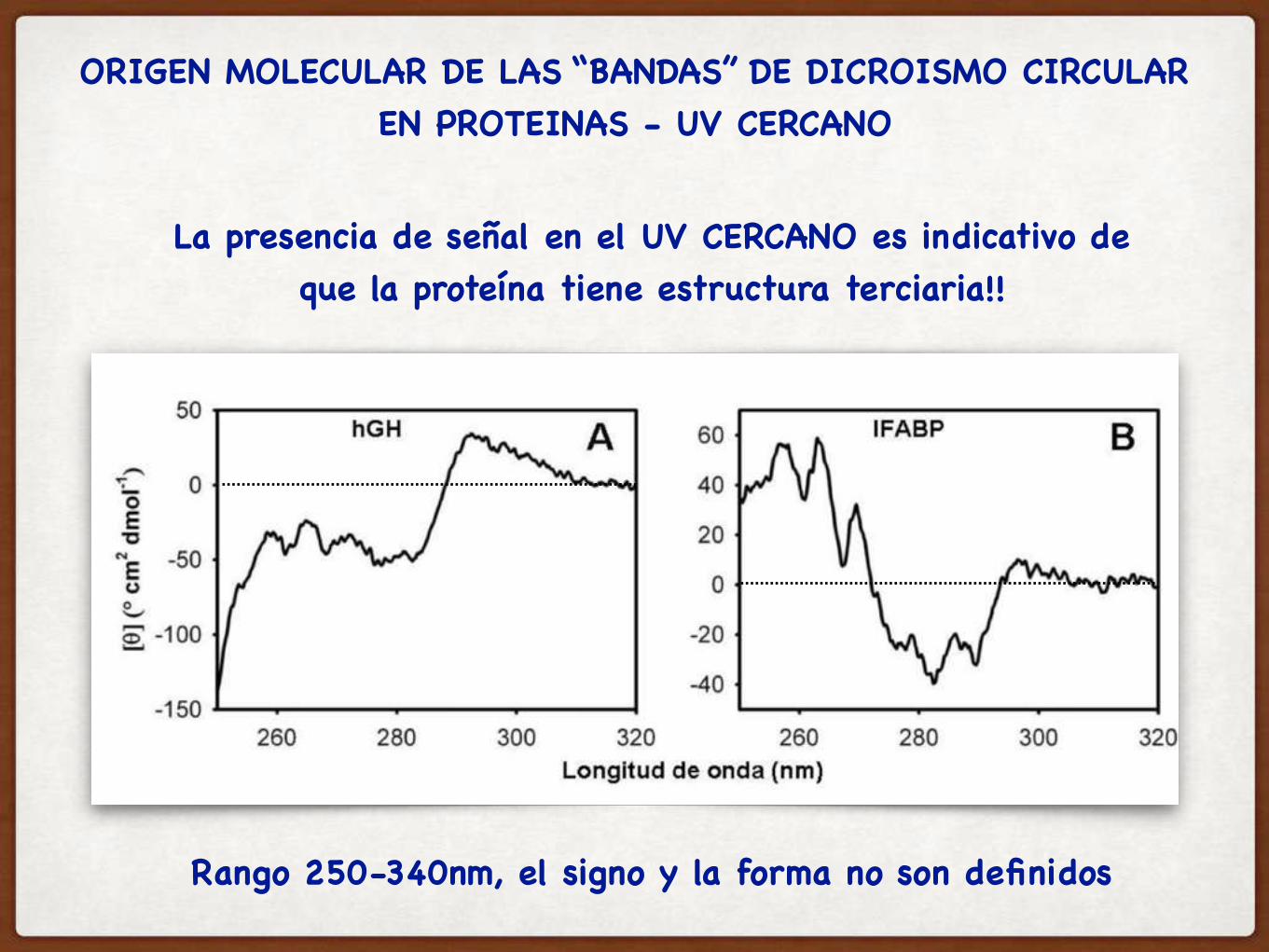
ORIGEN MOLECULAR DE LAS “BANDAS” DE DICROISMO CIRCULAR EN PROTEINAS - UV CERCANO
Rango 250-340nm, el signo y la forma no son definidos
La presencia de señal en el UV CERCANO es indicativo deque la proteína tiene estructura terciaria!!
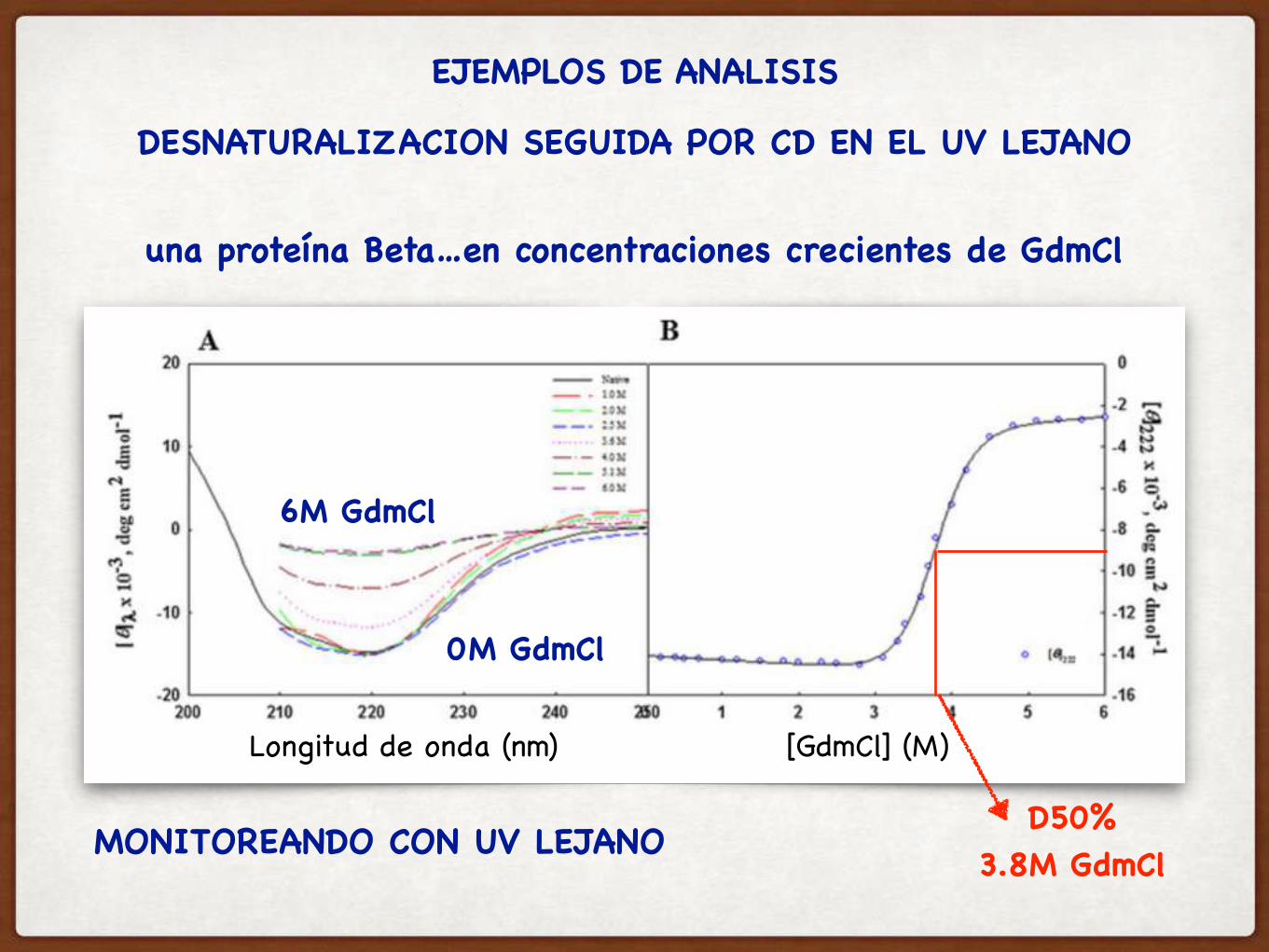
EJEMPLOS DE ANALISIS
DESNATURALIZACION SEGUIDA POR CD EN EL UV LEJANO
una proteína Beta…en concentraciones crecientes de GdmCl
0M GdmCl
6M GdmCl
Longitud de onda (nm) [GdmCl] (M)
D50%3.8M GdmClMONITOREANDO CON UV LEJANO

FLUORESCENCIA INTRINSECA DE PROTEINAS
EL FENOMENO DE FLUORESCENCIA
Es la emisión de luz por una sustancia al decaer del estado
excitado (S1) al basal (So)
característico de gr aromáticos
“DIAGRAMA DE JABLONSKI”
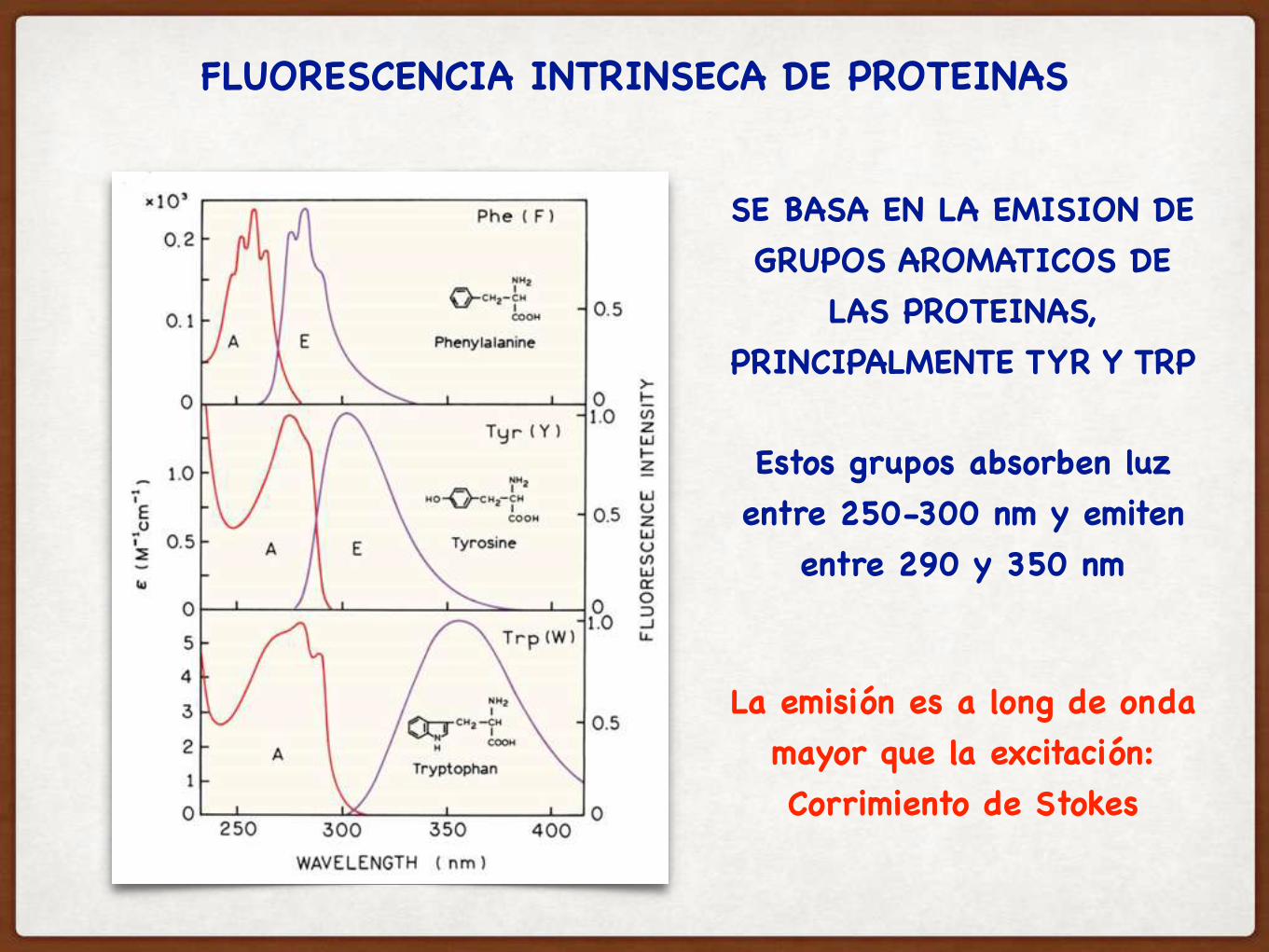
FLUORESCENCIA INTRINSECA DE PROTEINAS
SE BASA EN LA EMISION DE GRUPOS AROMATICOS DE
LAS PROTEINAS, PRINCIPALMENTE TYR Y TRP
Estos grupos absorben luz entre 250-300 nm y emiten
entre 290 y 350 nm
La emisión es a long de onda mayor que la excitación:Corrimiento de Stokes
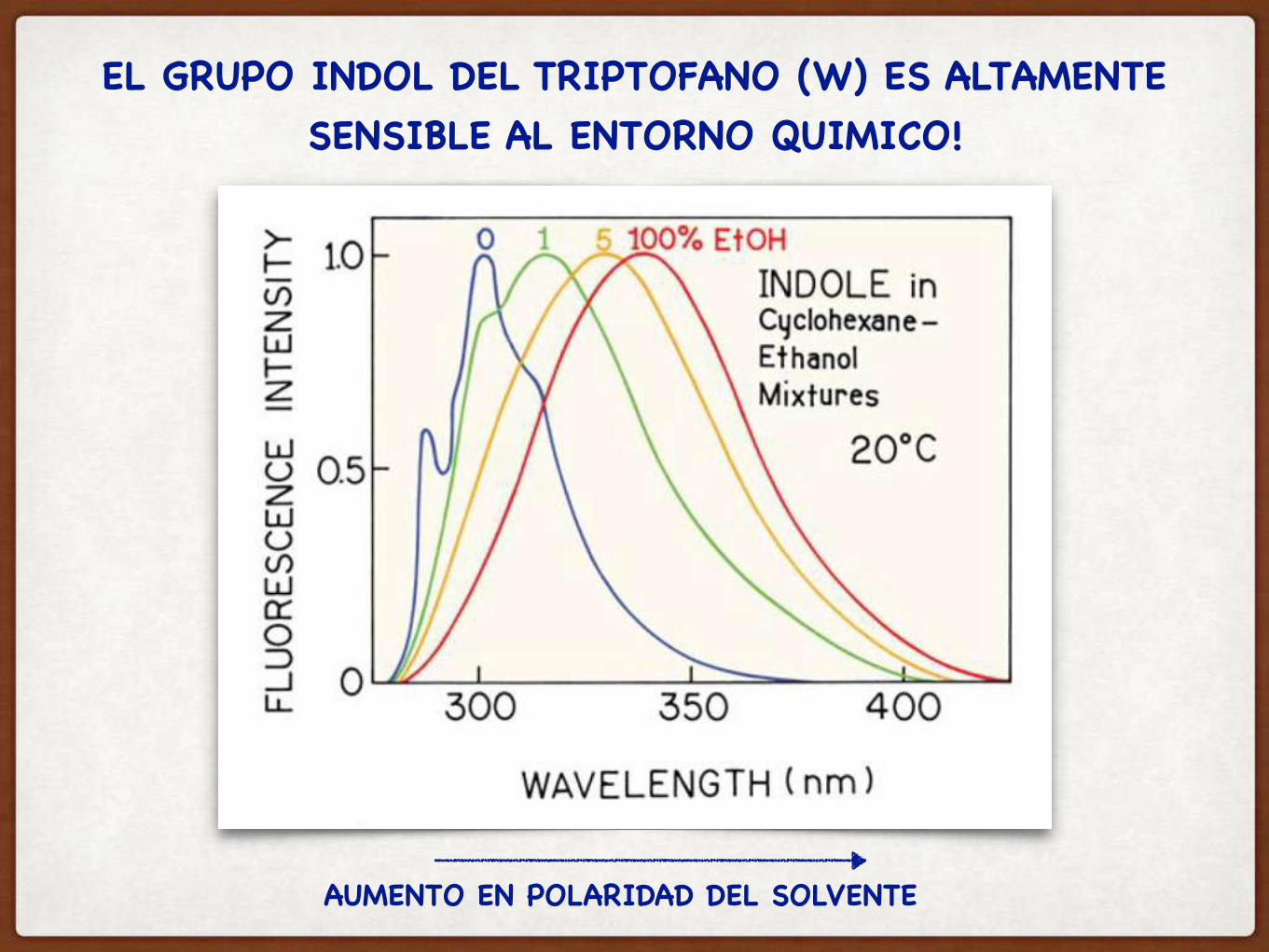
EL GRUPO INDOL DEL TRIPTOFANO (W) ES ALTAMENTE SENSIBLE AL ENTORNO QUIMICO!
AUMENTO EN POLARIDAD DEL SOLVENTE
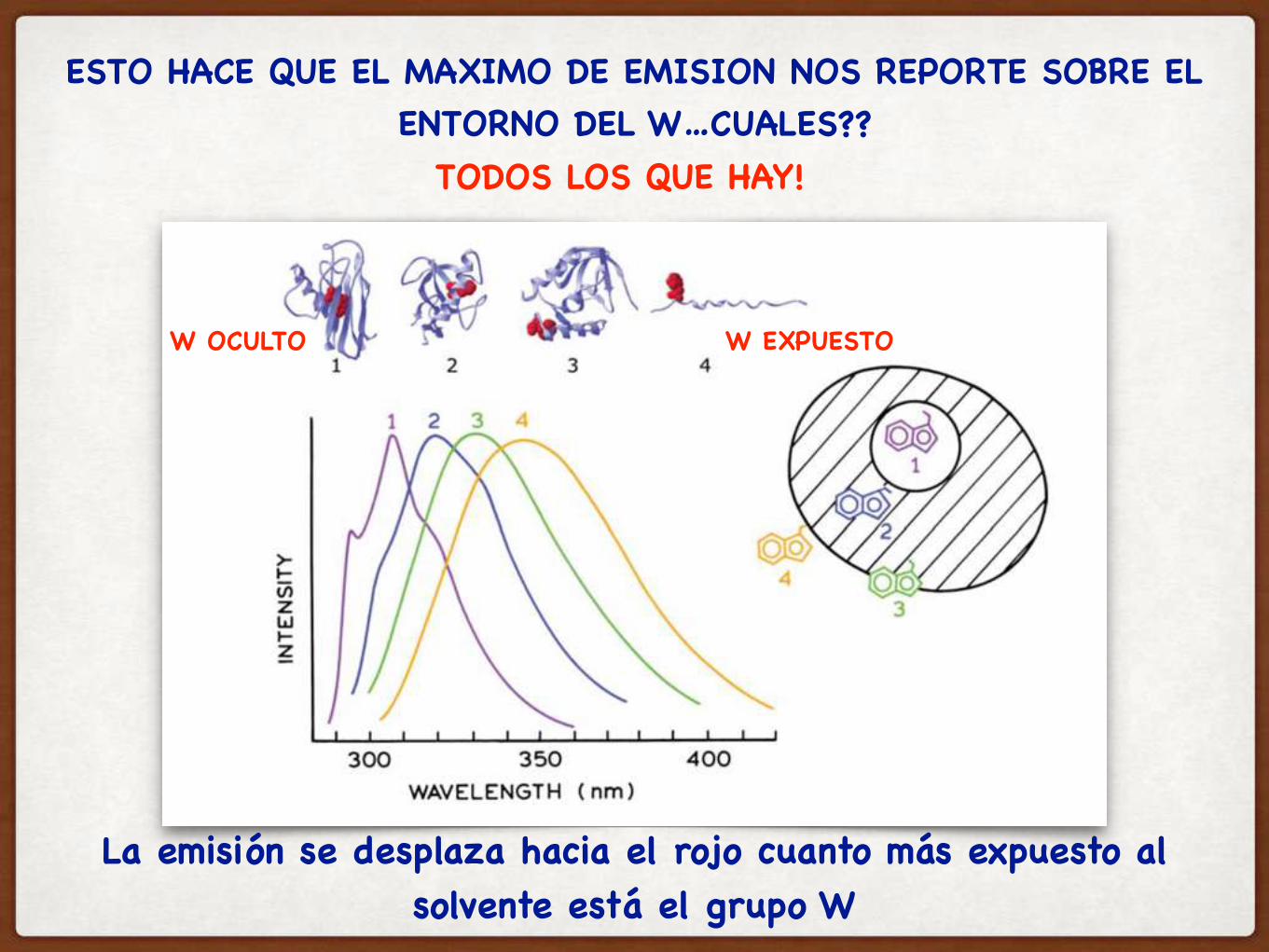
ESTO HACE QUE EL MAXIMO DE EMISION NOS REPORTE SOBRE EL ENTORNO DEL W…CUALES??
La emisión se desplaza hacia el rojo cuanto más expuesto al solvente está el grupo W
TODOS LOS QUE HAY!
W OCULTO W EXPUESTO
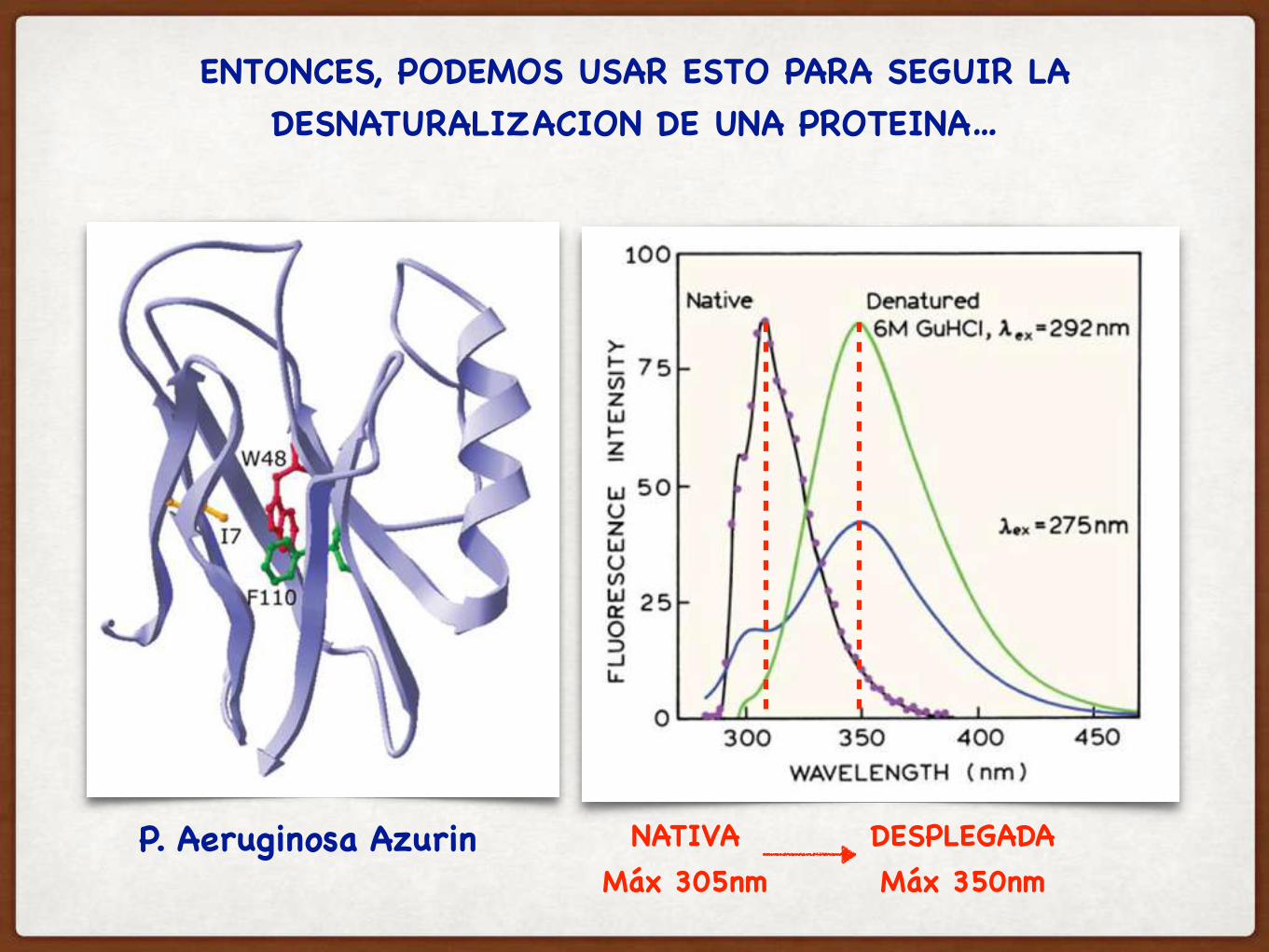
ENTONCES, PODEMOS USAR ESTO PARA SEGUIR LA DESNATURALIZACION DE UNA PROTEINA…
P. Aeruginosa Azurin NATIVAMáx 305nm
DESPLEGADAMáx 350nm
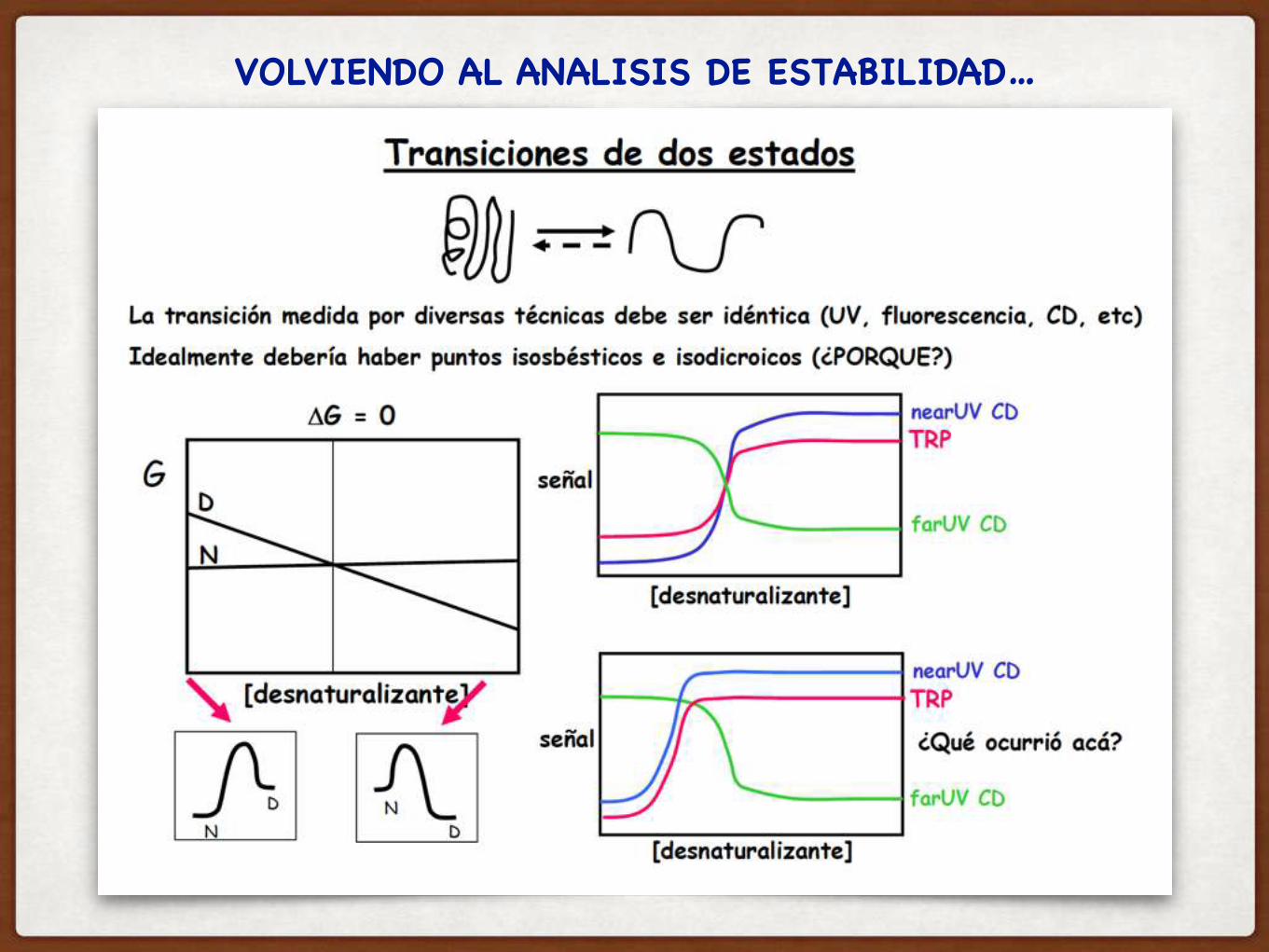
VOLVIENDO AL ANALISIS DE ESTABILIDAD…
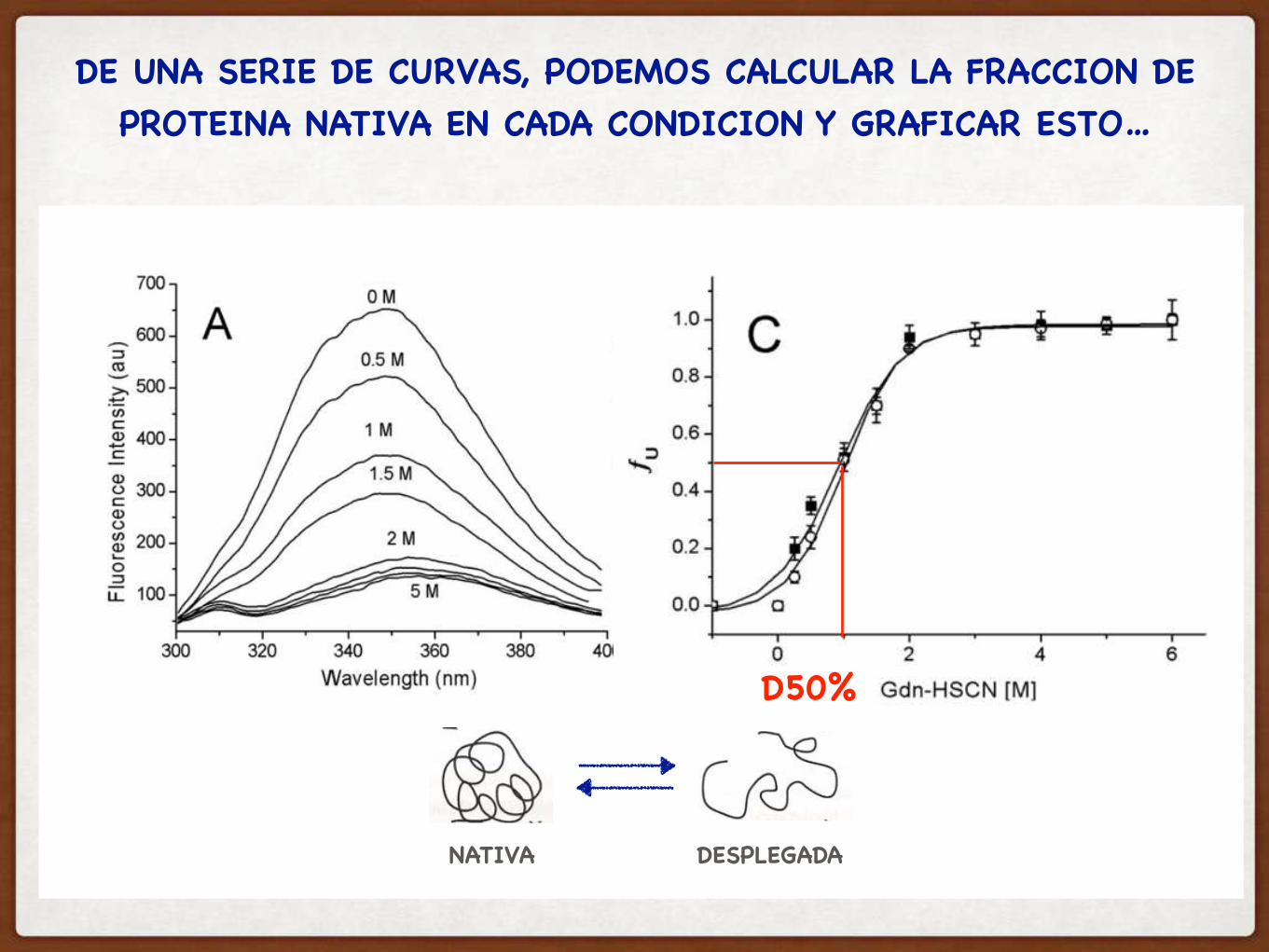
DE UNA SERIE DE CURVAS, PODEMOS CALCULAR LA FRACCION DE PROTEINA NATIVA EN CADA CONDICION Y GRAFICAR ESTO…
D50%
NATIVA DESPLEGADA

Y CUANDO LAS TRANSICION MEDIDA POR DIFERENTES TECNICAS ES DIFERENTE, COMO LO INTERPRETO??
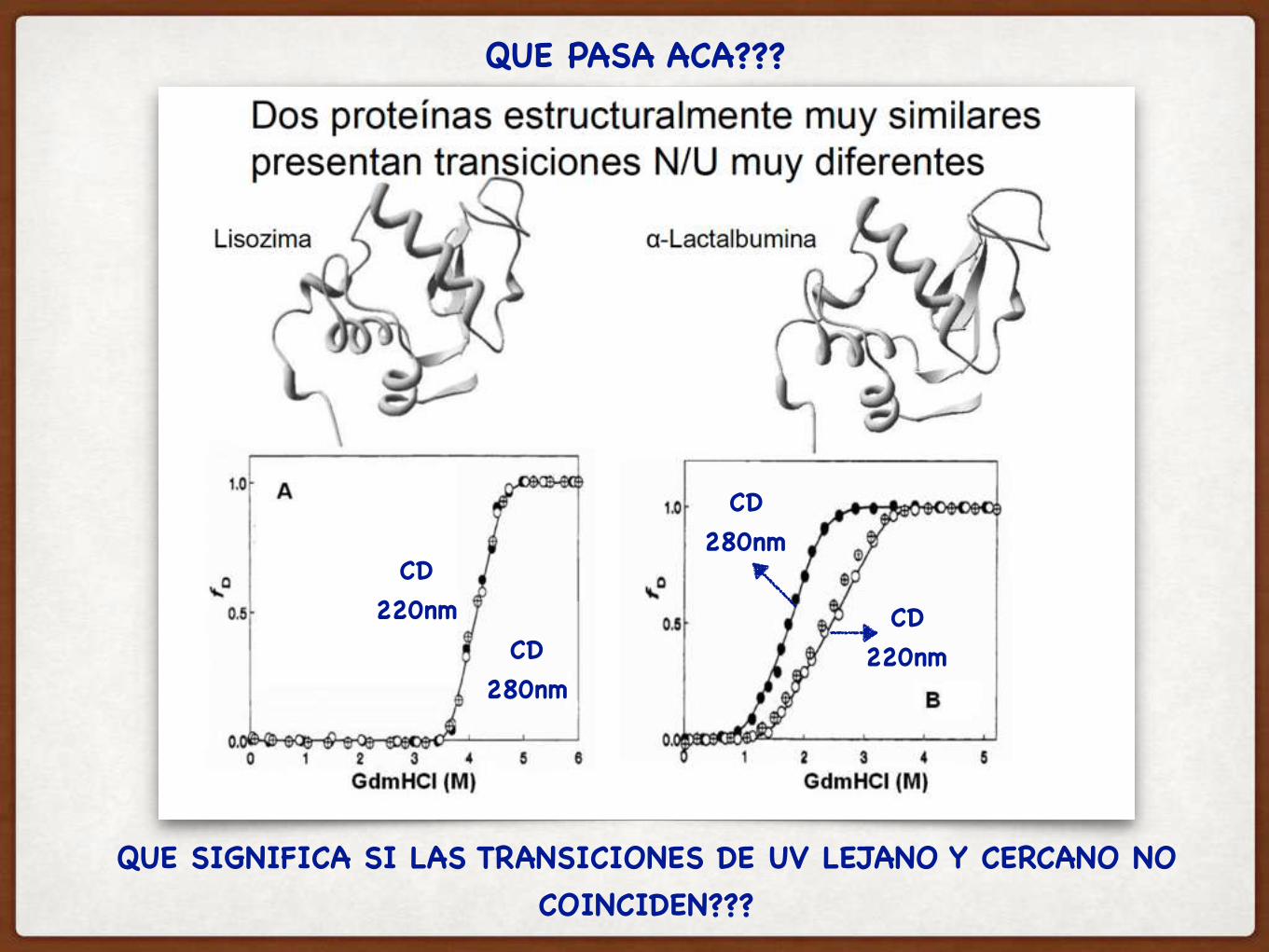
CD220nm
QUE PASA ACA???
QUE SIGNIFICA SI LAS TRANSICIONES DE UV LEJANO Y CERCANO NO COINCIDEN???
CD280nm
CD220nm
CD280nm
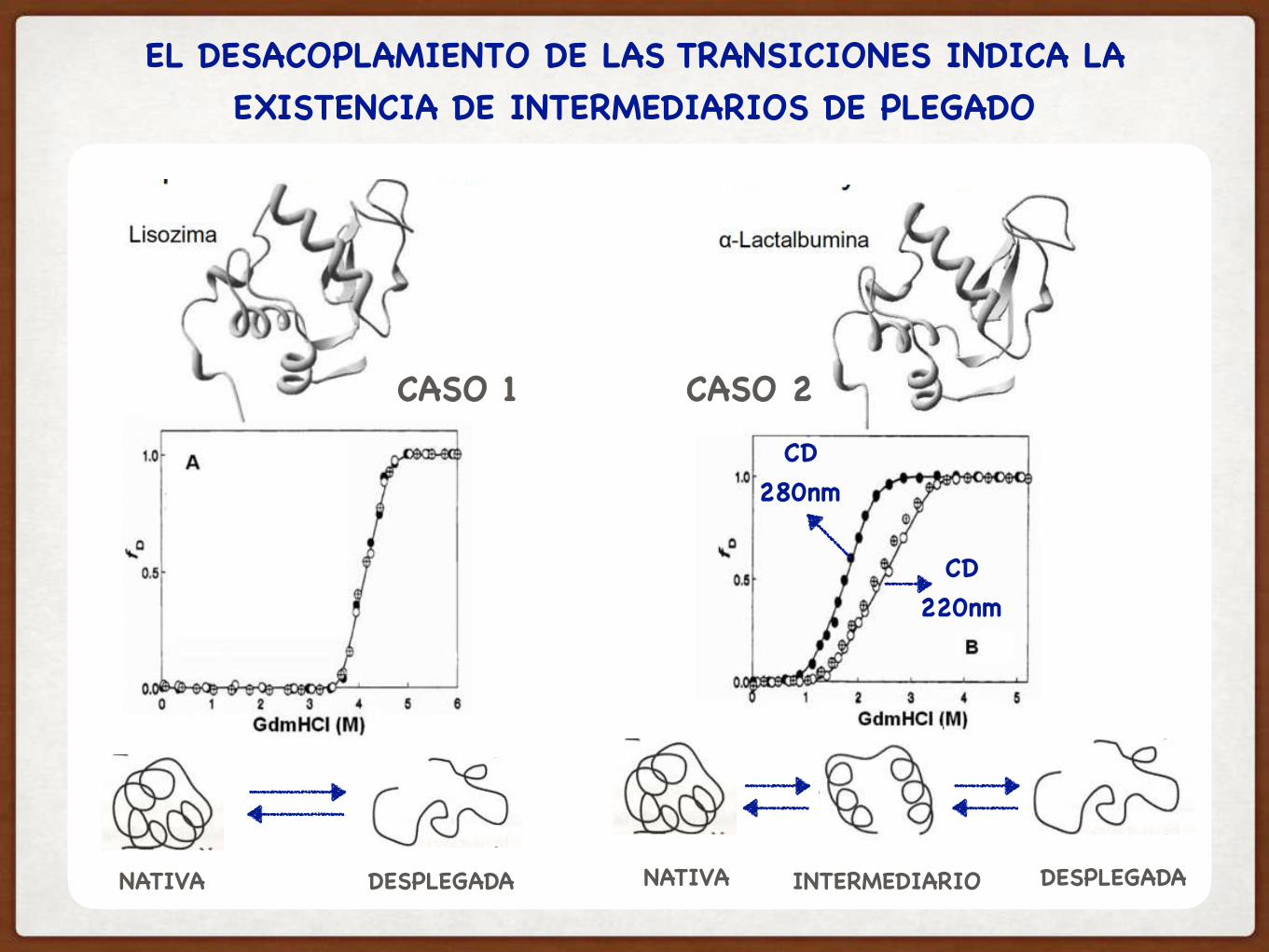
NATIVA DESPLEGADA NATIVA DESPLEGADAINTERMEDIARIO
EL DESACOPLAMIENTO DE LAS TRANSICIONES INDICA LA EXISTENCIA DE INTERMEDIARIOS DE PLEGADO
CASO 1 CASO 2
CD220nm
CD280nm
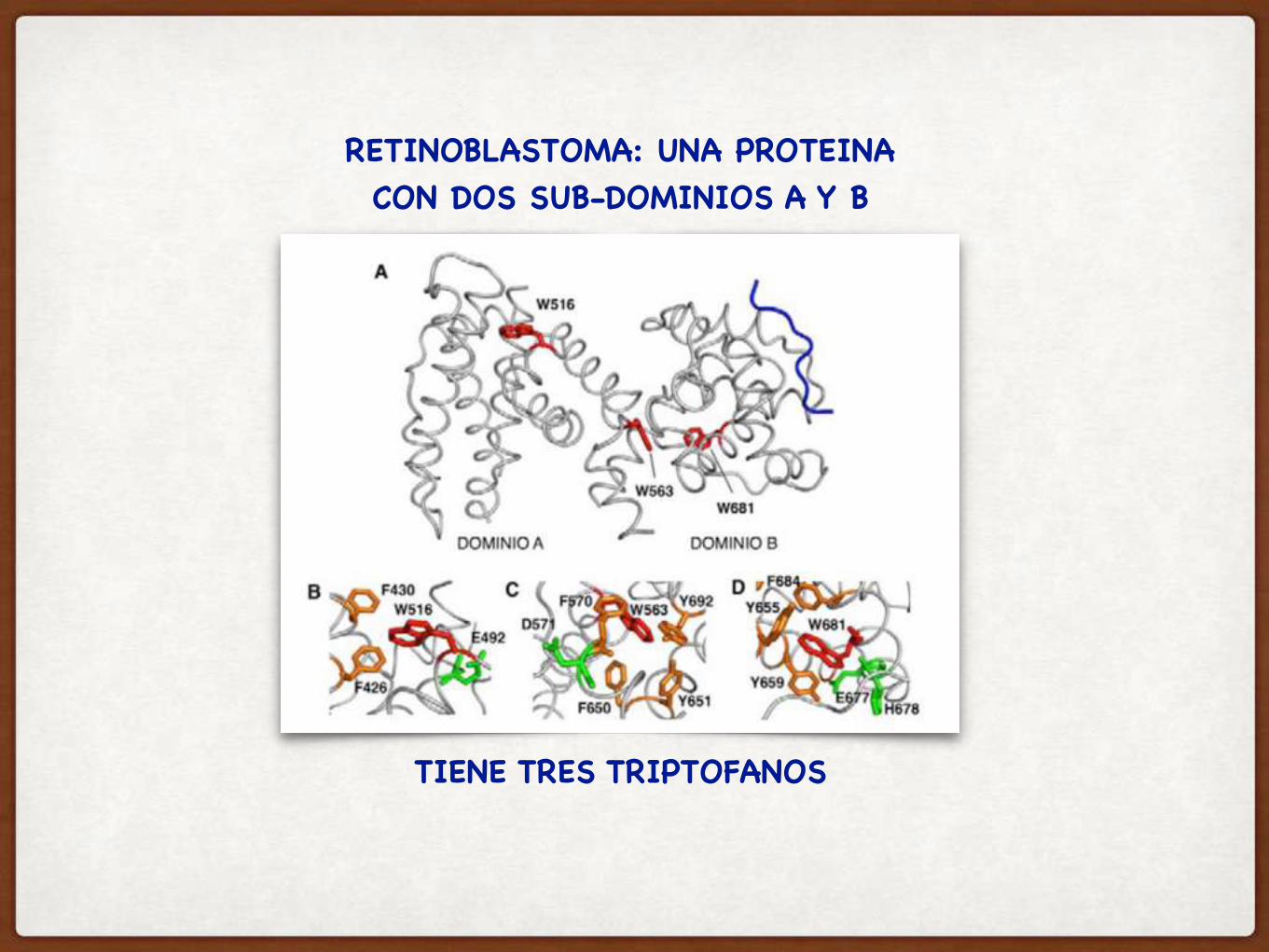
RETINOBLASTOMA: UNA PROTEINA CON DOS SUB-DOMINIOS A Y B
TIENE TRES TRIPTOFANOS
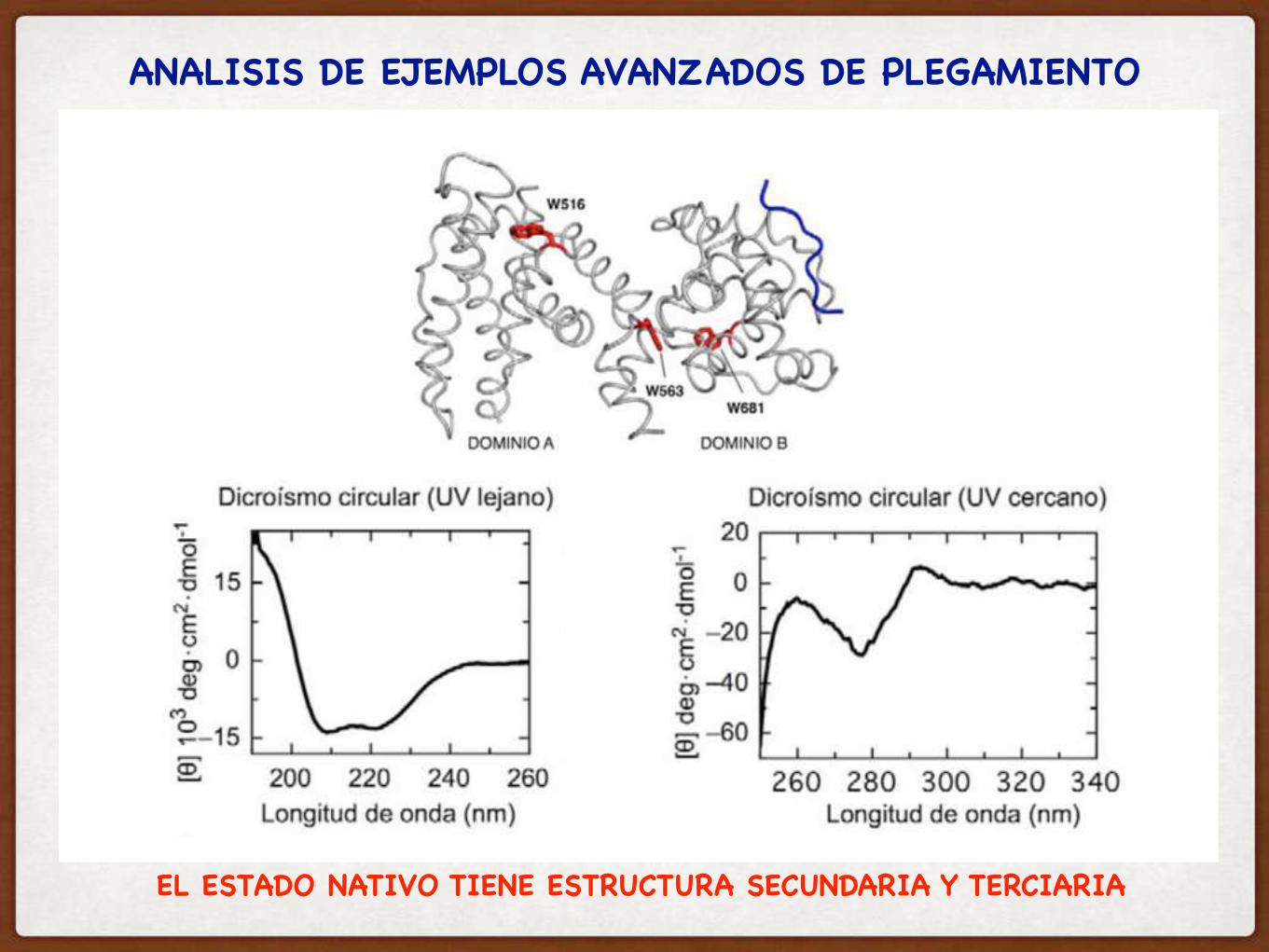
ANALISIS DE EJEMPLOS AVANZADOS DE PLEGAMIENTO
EL ESTADO NATIVO TIENE ESTRUCTURA SECUNDARIA Y TERCIARIA
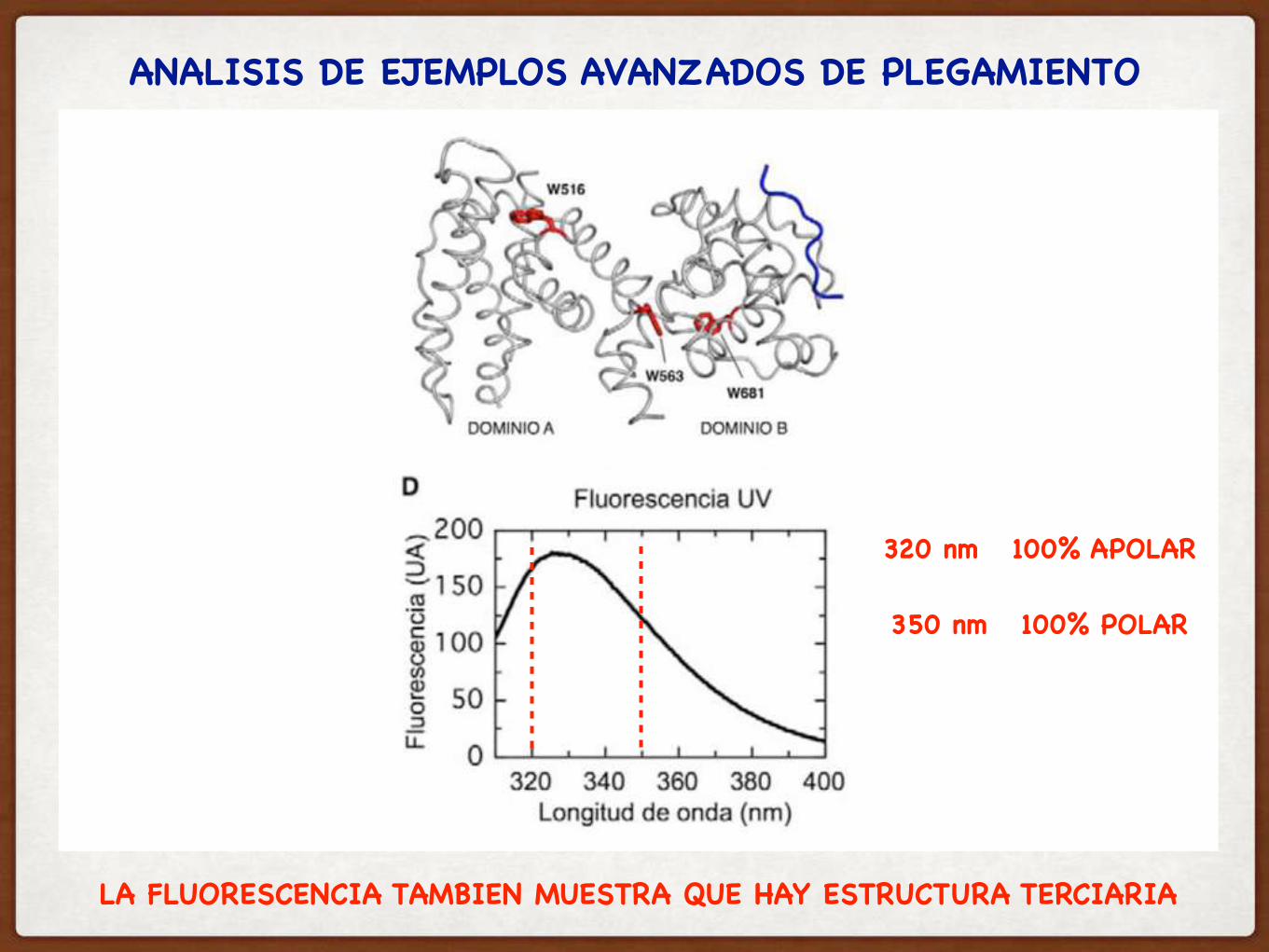
ANALISIS DE EJEMPLOS AVANZADOS DE PLEGAMIENTO
320 nm 100% APOLAR
350 nm 100% POLAR
LA FLUORESCENCIA TAMBIEN MUESTRA QUE HAY ESTRUCTURA TERCIARIA
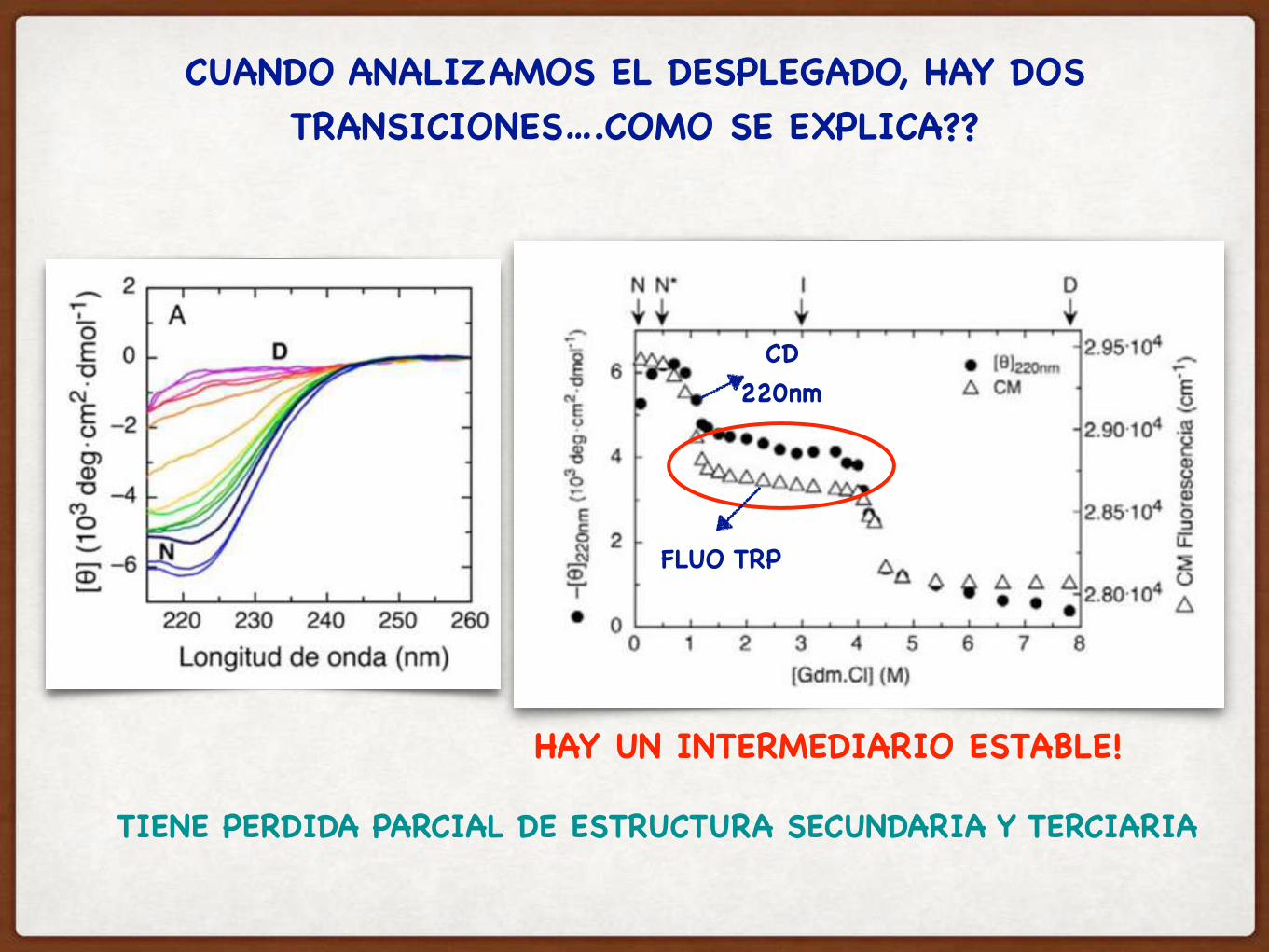
CUANDO ANALIZAMOS EL DESPLEGADO, HAY DOS TRANSICIONES….COMO SE EXPLICA??
HAY UN INTERMEDIARIO ESTABLE!
TIENE PERDIDA PARCIAL DE ESTRUCTURA SECUNDARIA Y TERCIARIA
CD220nm
FLUO TRP
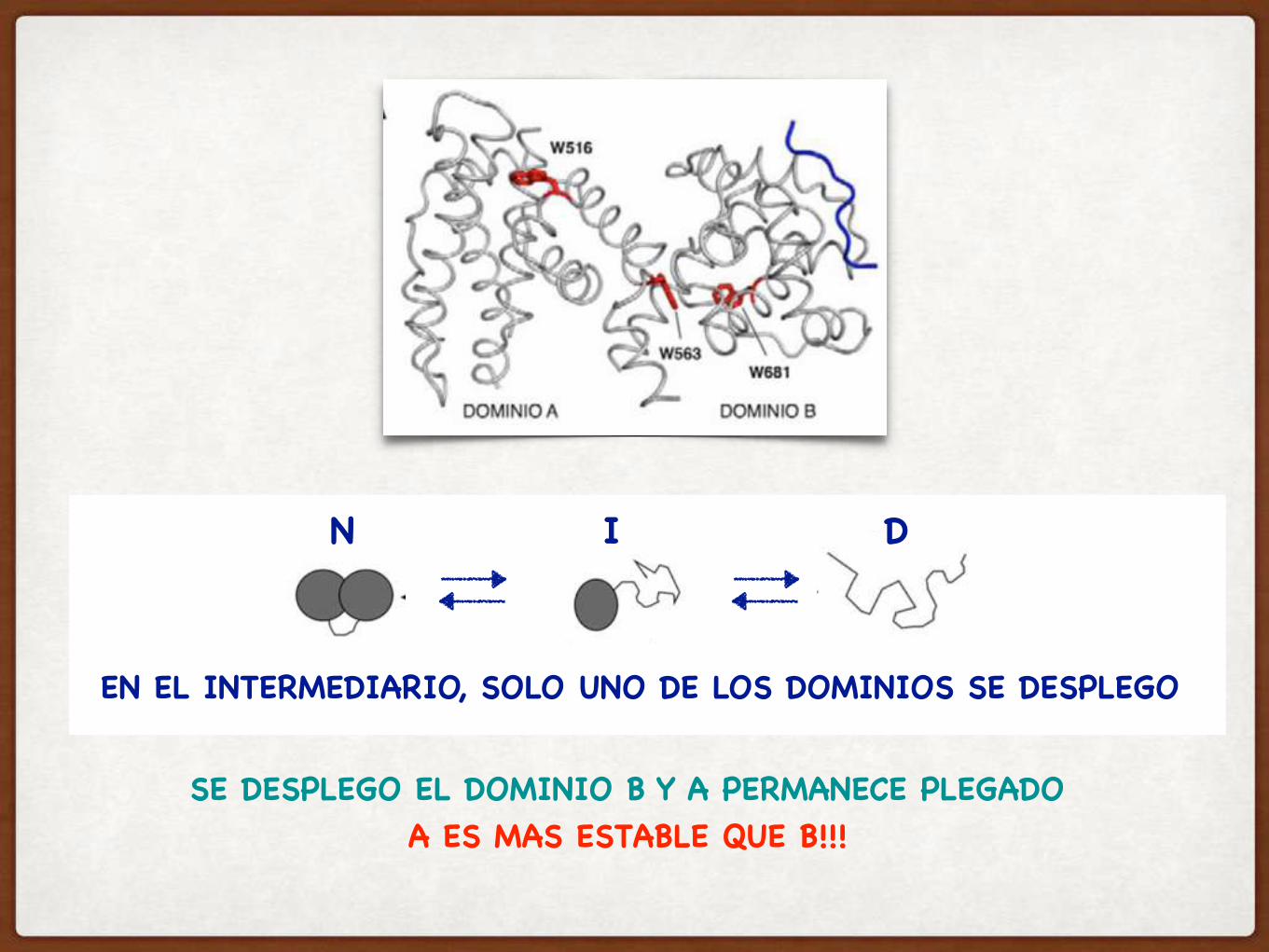
SE DESPLEGO EL DOMINIO B Y A PERMANECE PLEGADOA ES MAS ESTABLE QUE B!!!
N I D
EN EL INTERMEDIARIO, SOLO UNO DE LOS DOMINIOS SE DESPLEGO

ANALISIS DE EJEMPLOS AVANZADOS DE PLEGAMIENTOOLIGOMEROS
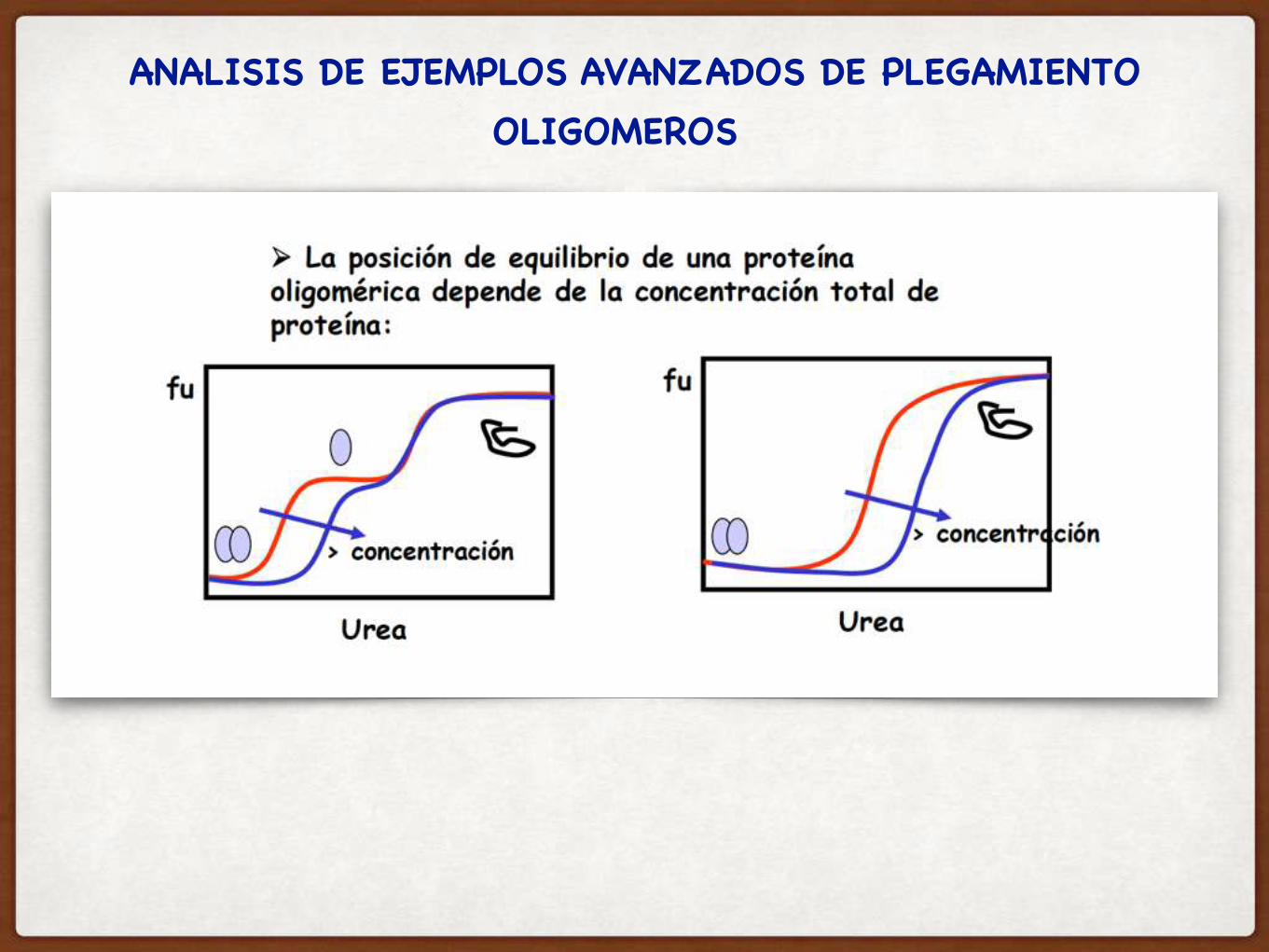
ANALISIS DE EJEMPLOS AVANZADOS DE PLEGAMIENTOOLIGOMEROS

ANALISIS DE EJEMPLOS AVANZADOS DE PLEGAMIENTO
INGENIERIA DE PROTEINAS Y MUTACIONES
FRAC
CION
DESP
LEGA
DA
[ UREA ]
Mutación estabilizante
Mutación desestabilizante
Aumento de estabilidad
D50 D50 D50


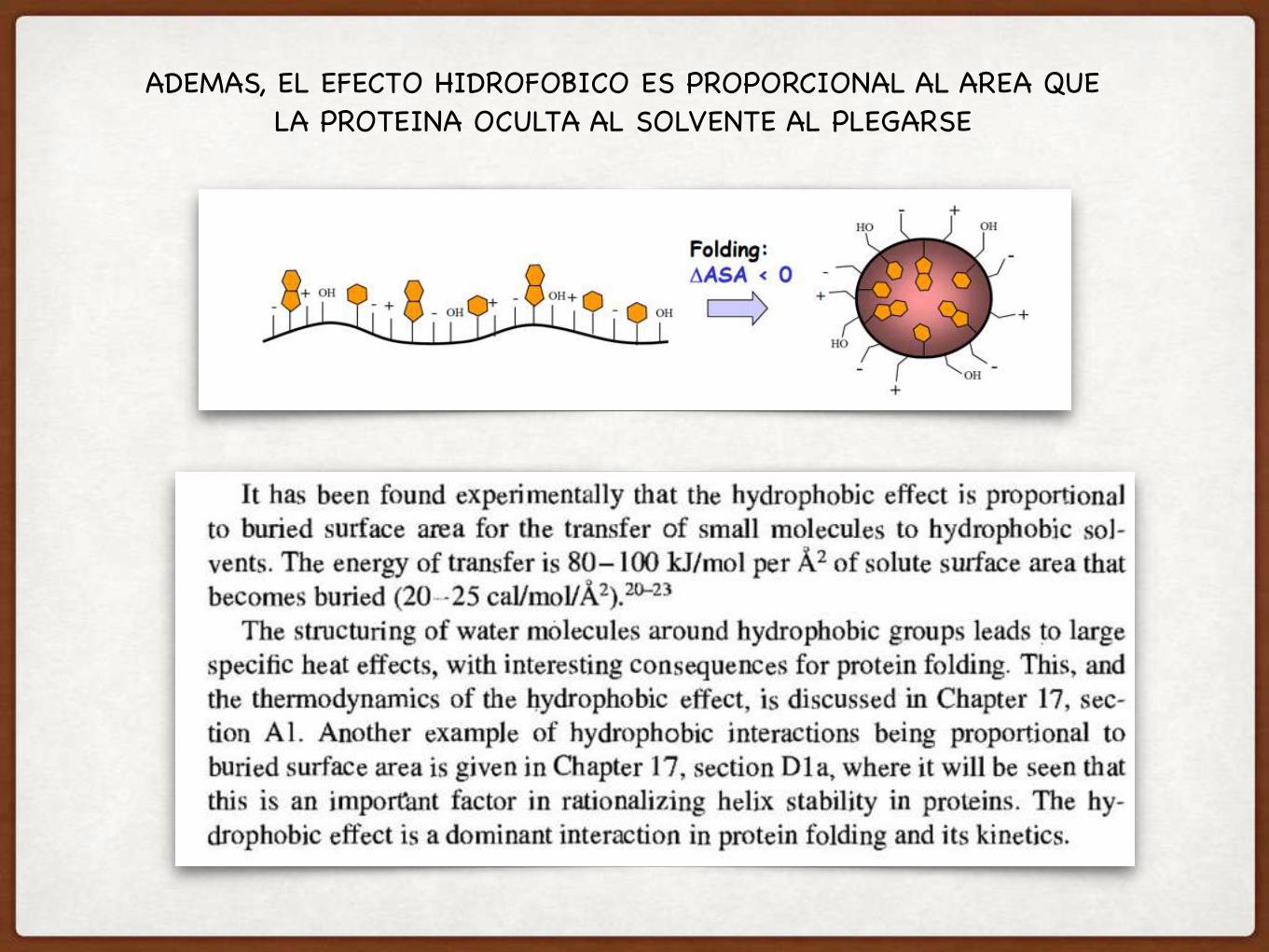
ADEMAS, EL EFECTO HIDROFOBICO ES PROPORCIONAL AL AREA QUE LA PROTEINA OCULTA AL SOLVENTE AL PLEGARSE






![2012_09_11 Clase 15 Pliegues y Plegamiento [Compatibility Mode]](https://img.pdfslide.tips/doc/110x75/5572110b497959fc0b8e3423/20120911-clase-15-pliegues-y-plegamiento-compatibility-mode.jpg)






















